ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к выделенным полинуклеотидам, кодирующим дельта-8-десатуразу, к дельта-8-десатуразам, кодируемым выделенными полинуклеотидами, векторам экспрессии, содержащим выделенные полинуклеотиды, клеткам-хозяевам, содержащим векторы экспрессии, и к способам получения дельта-8-десатураз и полиненасыщенных жирных кислот.
УРОВЕНЬ ТЕХНИКИ
Полиненасыщенные жирные кислоты (PUFA) участвуют в правильном функционировании всех живых организмов. Например, PUFA являются важными составляющими плазматической мембраны клетки, в составе которой они находятся в виде фосфолипидов. PUFA необходимы для нормального развития мозга у детей, а также для образования и восстановления тканей у взрослых млекопитающих.
В биологическом синтезе PUFA участвуют несколько типов ферментов, прежде всего, десатуразы и элонгазы (см. ФИГ.1). Десатуразы катализируют внесение участков ненасыщения (например, двойных связей) между атомами углерода в алкильной цепи жирной кислоты в субстрате. Элонгазы катализируют присоединение единицы, состоящей из двух атомов углерода, к субстрату жирной кислоты. Например, линолевую кислоту (LA, 18:2n-6) получают из олеиновой кислоты (OA, 18:1n-9) с помощью Δ12-десатуразы. Эйкозадиеновую кислоту (EDA, 20:2n-6) получают из LA с помощью Δ9-элонгазы. Дихомо-γ-линоленовую кислоту (DGLA, 20:3n-6) получают из EDA с помощью Δ8-десатуразы (см. ФИГ.1). Арахидоновую кислоту (ARA, 20:4n-6) получают из DGLA с помощью Δ5-десатуразы (см. ФИГ.1).
В данной области техники известно несколько важных PUFA с длинной цепью. Например, одной из наиболее важных PUFA с длинной цепью является эйкозапентаеновая кислота (EPA). EPA содержится в грибах и жирах морских продуктов. Второй важной PUFA с длинной цепью является докозагексаеновая кислота (DHA). DHA часто содержится в рыбьем жире, а также ее можно получить путем выделения из ткани мозга млекопитающих. Третьей важной PUFA с длинной цепью является ARA. ARA содержится в мицелиальных грибах, а также ее можно получить путем выделения из тканей млекопитающих, включая ткани печени и надпочечников.
ARA, EPA и/или DHA можно получить как с помощью альтернативной цепочки превращений Δ8-десатуразы/Δ9-элонгазы, так и с помощью традиционной цепочки превращений Δ6-десатуразы (см. ФИГ.1). Ранее были идентифицированы элонгазы, обладающие активностью в отношении жирных кислот в качестве субстрата в классической цепочке превращений Δ6 для образования PUFA с длинной цепью, в частности, ARA, EPA и DHA. В ходе традиционной цепочки превращений Δ6-десатуразы, направленной на превращение LA в DGLA и альфа-линоленовой кислоты (ALA) в ω3-эйкозатетраеновую кислоту (ω3-ETA), используют Δ6-десатуразный фермент для превращения LA в гамма-линоленовую кислоту (GLA) и ALA в стеаридоновую кислоту (SDA), и C18-элонгазный фермент для превращения GLA в DGLA и SDA в ω3-ETA. Тем не менее, в некоторых случаях более предпочтительной может являться альтернативная цепочка превращений Δ8-десатуразы/Δ9-элонгазы по сравнению с традиционной цепочкой превращений Δ6-десатуразы. Например, если в ходе получения DGLA, ARA, ω3-ETA, EPA, ω3-докозапентаеновой кислоты (DPA) и/или DHA является нежелательным образование остаточных промежуточных продуктов омега-6 или омега-3 жирных кислот, например, GLA или SDA, можно применять альтернативный путь Δ8-десатуразы/Δ9-элонгазы вместо традиционного пути Δ6-десатуразы для того, чтобы избежать образования GLA и SDA. Возможность применения Δ8-десатураз в указанной цепочке превращений обусловлена способностью десатурировать молекулу жирной кислоты в положении между восьмым и девятым атомами углерода (нумерация осуществляется от карбоксильного конца молекулы) и их способностью, например, катализировать превращение ω6-эйкозадиеновой кислоты (EDA) в DGLA и/или ω3-эйкозатриеновой кислоты (ω3-ETrA) в ω3-ETA. Следовательно, в данной области техники существует потребность в новых источниках Δ8-десатураз, которые можно применять для получения PUFA с длинной цепью.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном аспекте настоящее изобретение относится к выделенной нуклеотидной кислоте или ее фрагменту, содержащей последовательность нуклеотидов, кодирующую полипептид, обладающий десатуразной активностью, или комплементарной указанной последовательности, причем аминокислотная последовательность полипептида по меньшей мере на 55% идентична аминокислотной последовательности, содержащей SEQ ID NO:29. Выделенная нуклеиновая кислота или ее фрагмент кодирует функционально активный фермент Δ8-десатуразу, использующий в качестве субстрата ω6-эйкозадиеновую кислоту или ω3-эйкозатриеновую кислоту. Указанную выделенную последовательность нуклеиновой кислоты можно получать из Emiliana huxleyi, предпочтительно, из Emiliana huxleyi CCMP 378.
В другом аспекте настоящее изобретение относится к выделенной нуклеотидной последовательности или ее фрагменту, содержащей или комплементарной по меньшей мере 55% нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO:28 и SEQ ID NO:30. Выделенная нуклеотидная последовательность или ее фрагмент кодирует функционально активный фермент десатуразу, использующий в качестве субстрата ω6-эйкозадиеновую кислоту или ω3-эйкозатриеновую кислоту. Выделенная нуклеотидная последовательность может содержать последовательность, представленную в SEQ ID NO:28. В качестве альтернативного варианта, выделенная нуклеотидная последовательность может содержать последовательность, представленную в SEQ ID NO:30. Указанную выделенную нуклеотидную последовательность можно получить из Emiliana huxleyi, предпочтительно, Emiliana huxleyi CCMP 378.
В другом аспекте настоящее изобретение относится к вектору экспрессии. Указанный вектор экспрессии согласно настоящему изобретению содержит нуклеотидную последовательность, функционально связанную с регуляторной последовательностью, причем нуклеотидная последовательность содержит или комплементарна по меньшей мере 55% нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO:28 и SEQ ID NO:30.
В еще одном дополнительном аспекте настоящее изобретение относится к клетке-хозяину, содержащей описанный выше вектор экспрессии. Клетка-хозяин может представлять собой клетку эукариот. В частности, клетку эукариот выбирают из группы, состоящей из: клетки млекопитающего, клетки насекомого, клетки растения и клетки гриба. Примеры применяемых клеток грибов включают клетки грибов, выбранные из группы, состоящей из: Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Kluyveromyces spp., Hansenula spp., Aspergillus spp., Penicillium spp., Neurospora spp., Trichoderma spp. и Pichia spp. Примеры применяемых клеток растений включают клетки растений, выбранные из группы, состоящей из: клеток растений сои, видов рода Brassica, сафлора, подсолнечника, кукурузы, хлопка и льна.
В еще одном аспекте настоящее изобретение относится к клетке растения, зерну растения, растению или ткани растения, содержащим описанный выше вектор экспрессии, при этом экспрессия нуклеотидной последовательности вектора приводит к образованию клеткой растения, зерном растения растением или тканью растения по меньшей мере одной полиненасыщенной жирной кислоты. Полиненасыщенная жирная кислота, образуемая указанным вектором экспрессии, выбрана из группы, состоящей из: арахидоновой кислоты (ARA), эйкозапентаеновой кислоты (EPA), докозагексаеновой кислоты (DHA), дихомо-гамма-линоленовой кислоты (DGLA) или ω3-эйкозатетраеновой кислоты (ω3-ETA), а также комбинаций указанных кислот.
В еще одном аспекте настоящее изобретение относится к одному или более растительных масел или жирных кислот, образуемых клеткой растения, зерном растения, растением или тканью растения, описанными выше.
В еще одном аспекте настоящее изобретение относится к очищенному полипептиду, кодируемому нуклеотидной последовательностью, содержащей или комплементарной по меньшей мере 55% нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30.
В еще одном аспекте настоящее изобретение относится к очищенному полипептиду, десатурирующему полиненасыщенную жирную кислоту, длина углеродной цепи которой составляет 20 атомов углерода (C20-PUFA), используемую в качестве субстрата, в положении между 8 и 9 атомами углерода, и при этом аминокислотная последовательность полипептида по меньшей мере на 55% идентична аминокислотной последовательности, содержащей SEQ ID NO:29.
В еще одном варианте реализации настоящее изобретение относится к очищенному полипептиду, содержащему аминокислотную последовательность, представленную в SEQ ID NO:29.
В еще одном варианте реализации настоящее изобретение относится к способу получения фермента Δ8-десатуразы. Указанный способ включает этапы:
a) выделения нуклеотидной последовательности, содержащей или комплементарной по меньшей мере 55% нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30;
b) конструирования вектора экспрессии, содержащего выделенную нуклеотидную последовательность согласно этапу a), функционально связанную с регуляторной последовательностью; и
c) введения вектора экспрессии в клетку-хозяина на период времени и в условиях, достаточных для образования фермента Δ8-десатуразы.
В способе, описанном выше, клетка-хозяин представляет собой клетку эукариот. В частности, указанную клетку эукариот выбирают из группы, состоящей из: клетки млекопитающего, клетки насекомого, клетки растения и клетки гриба. Примеры применяемых клеток грибов включают клетки грибов, выбранные из группы, состоящей из: Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Kluyveromyces spp., Hansenula spp., Aspergillus spp., Penicillium spp., Neurospora spp., Trichoderma spp. и Pichia spp. Примеры применяемых клеток растений включают клетки растений, выбранные из группы, состоящей из: клеток растений сои, видов рода Brassica, сафлора, подсолнечника, кукурузы, хлопка и льна.
В еще одном варианте реализации настоящее изобретение относится к способу получения полиненасыщенной жирной кислоты, включающий этапы:
a) выделения нуклеотидной последовательности, содержащей или комплементарной по меньшей мере 55% нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30;
b) конструирования вектора экспрессии, содержащего выделенную нуклеотидную последовательность согласно этапу a), функционально связанную с регуляторной последовательностью;
c) введения вектора экспрессии в клетку-хозяина на период времени и в условиях, достаточных для образования фермента Δ8-десатуразы; и
d) приведения во взаимодействие фермента Δ8-десатуразы с субстратом, выбранным из группы, состоящей из: ω6-эйкозадиеновой кислоты, ω3-эйкозатриеновой кислоты, а также комбинаций указанных кислот, для превращения субстрата в получаемую полиненасыщенную жирную кислоту.
В способе, описанном выше, получаемая полиненасыщенная жирная кислота представляет собой дихомо-гамма-линоленовую кислоту (DGLA), ω3-эйкозатетраеновую кислоту (ω3-ETA) или любые комбинации указанных кислот.
Также, способ, описанный выше, дополнительно может включать этап:
приведения во взаимодействие получаемой полиненасыщенной жирной кислоты с по меньшей мере одной дополнительной десатуразой или элонгазой, для превращения получаемой полиненасыщенной жирной кислоты в другую или дополнительную полиненасыщенную жирную кислоту. Получаемая в результате полиненасыщенная жирная кислота представляет собой арахидоновую кислоту (ARA), эйкозапентаеновую кислоту (EPA), докозапентаеновую кислоту (DPA) или докозагексаеновую кислоту (DHA) или любую комбинацию указанных кислот.
В еще одном аспекте настоящее изобретение относится к способу получения полиненасыщенной жирной кислоты в клетке-хозяине, включающий этапы:
a) выделения нуклеотидной последовательности, содержащей или комплементарной по меньшей мере 55% нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30;
b) конструирования вектора экспрессии, содержащего выделенную нуклеотидную последовательность согласно этапу a), функционально связанную с регуляторной последовательностью;
c) введения вектора экспрессии согласно этапу b) и по меньшей мере одной дополнительной рекомбинантной ДНК конструкции, содержащей выделенную нуклеотидную последовательность, функционально связанную с по меньшей мере одной регуляторной последовательностью, кодирующую дельта-9 элонгазу, в клетку-хозяина;
d) приведения во взаимодействие экспрессированного фермента Δ8-десатуразы и дельта-9 элонгазы с субстратом, выбранным из группы, состоящей из: линолевой кислоты (LA), альфа-линоленовой кислоты (ALA) и комбинаций указанных кислот, для превращения субстрата в получаемую полиненасыщенную жирную кислоту.
В описанном выше способе, получаемая полиненасыщенная жирная кислота представляет собой дихомо-гамма-линоленовую кислоту (DGLA) или ω3-эйкозатетраеновую кислоту (ω3-ETA) или любую комбинацию указанных кислот.
описанный выше способ дополнительно может включать этап:
приведения во взаимодействие полиненасыщенной жирной кислоты с по меньшей мере одной дополнительной десатуразой или элонгазой для превращения получаемой полиненасыщенной жирной кислоты в другую или дополнительную полиненасыщенную аминокислоту. Получаемая полиненасыщенная жирная кислота представляет собой арахидоновую кислоту (ARA), эйкозапентаеновую кислоту (EPA), докозапентаеновую кислоту (DPA), докозагексаеновую кислоту (DHA) или любую комбинацию указанных кислот.
В способе, описанном выше, клетка-хозяин представляет собой клетку эукариот. В частности, клетку эукариот выбирают из группы, состоящей из: клетки млекопитающего, клетки насекомого, клетки растения и клетки гриба. Примеры применяемых клеток грибов включают клетки грибов, выбранные из группы, состоящей из: Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Kluyveromyces spp., Hansenula spp., Aspergillus spp., Penicillium spp., Neurospora spp., Trichoderma spp. и Pichia spp. Примеры применяемых клеток растений включают клетки растений, выбранные из группы, состоящей из: клеток растений семейства бобовых, видов рода Brassica, сафлора, подсолнечника, кукурузы, хлопка и льна.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На ФИГ.1 показан путь биологического синтеза жирной кислоты и роль Δ8-десатуразы в указанном пути.
На ФИГ.2A и 2B показано выравнивание аминокислотной последовательности, кодируемой ED3-8 (SEQ ID NO:29) с известными Δ8-десатуразами Pavlova lutheri CCMP 459 (SEQ ID NO:2), Pavlova salina (SEQ ID NO:3), Euglena gracialis (SEQ ID NO:1) и Perkinsus (SEQ ID NO:4). Идентичные остатки выделены, консервативные гистидиновые боксы подчеркнуты, консервативный участок в домене цитохрома b5 подчеркнут (двойной чертой).
На ФИГ.3A показаны аминокислотные последовательности Δ8-десатуразы Euglena gracialis (# доступа AF139720, SEQ ID NO:1).
На ФИГ.3B показаны аминокислотные последовательности Δ8-десатуразы Pavlova lutheri CCMP 459 (SEQ ID NO:2).
На ФИГ.4A показаны аминокислотные последовательности Δ8-десатуразы Pavlova salina (# доступа DQ995518, SEQ ID NO:3).
На ФИГ.4B показаны аминокислотные последовательности Δ8-десатуразы Perkinsus marinus (# доступа DQ508730, SEQ ID NO:4).
На ФИГ.4C показаны аминокислотные последовательности Δ8-десатуразы Acanthamoenba castellani (# доступа CS608483, SEQ ID NO:5).
На ФИГ.5 показана последовательность ДНК (SEQ ID NO:11) клона ED3-8, полученного в соответствии со способом, описанным в Примере 2.
ФИГ.6 показана расшифрованная аминокислотная последовательность (SEQ ID NO:12) клона ED3-8 полученного в соответствии со способом, описанным в Примере 2.
ФИГ.7A показана последовательность ДНК (SEQ ID NO:15) клона PK15 полученного в соответствии со способом, описанным в Примере 2.
ФИГ.7B показана аминокислотная последовательность (SEQ ID NO:16) клона PK15, полученного в соответствии со способом, описанным в Примере 2.
ФИГ.8A показана последовательность ДНК (SEQ ID NO:24) предполагаемого 3′-конца клона клона ED3-8, полученного в соответствии со способом, описанным в Примере 2.
ФИГ.8B показана аминокислотная последовательность (SEQ ID NOS:25 и Δ4-46, соответственно, в порядке появления) предполагаемого 3′-конца клона ED3-8, полученного в соответствии со способом, описанным в Примере 2.
На ФИГ.9 показана последовательность гена 1254 пары азотистых оснований предполагаемой Δ8-десатуразы Emiliana huxleyi CCMP 378 (SEQ ID NO:28).
На ФИГ.10 показан белок, содержащий в составе 417 аминокислот (SEQ ID NO:29), кодируемый последовательностью гена, содержащего 1254 пары азотистых оснований, (SEQ ID NO:28) предполагаемой Δ8-десатуразы Emiliana huxleyi CCMP 378.
На ФИГ.11 показана последовательность гена, оптимизированная относительно кодонов, предполагаемой Δ8-десатуразы Emiliana huxleyi CCMP 378 (SEQ ID NO:30), обозначенная 'ED3-8-EP2-5-SC'. ED3-8-EP2-5-SC обладает 66.98% идентичности с исходной последовательностью гена ED3-8 (SEQ ID NO:28; ФИГ.9). Ни одна из модификаций в составе гена, оптимизированного относительно кодонов, не изменяет аминокислотной последовательности кодируемого белка (SEQ ID NO:29; ФИГ.10).
На ФИГ.12 показана последовательность гена Δ9-элонгазы, полученная из Isochrysis galbana (IsoD9) (№ доступа CQ831422, SEQ ID NO:31).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к нуклеотидным последовательностям (например, нуклеотидным последовательностям гена) и транслированным аминокислотным последовательностям гена Δ8-десатуразы Emiliana sp; например, Emiliana huxley, в частности, Emiliana huxley CCMP 378. Далее, в объем настоящего изобретения включены области применения гена и фермента, кодируемого указанным геном. Например, нуклеотид и соответствующий фермент можно применять для получения полиненасыщенных жирных кислот, например, DGLA, ARA, EPA, ω3-ETA DPA и DHA или любых комбинаций указанных кислот, которые можно включать в состав фармацевтических композиций, пищевых составов и других ценных продуктов.
A. Определения
Формы единственного числа, используемые в настоящем описании, включают также и множественное число обозначаемых объектов, если в контексте четко не указано обратного. Для перечисления в пределах областей числовых значений, приведенных в настоящем описании, предусматривается каждое промежуточное значение с одинаковой степенью точности. Например, в интервале 6-9, помимо чисел 6 и 9, также предусматриваются 7 и 8, а для интервала 6.0-7.0, явно предусматриваются числа 6.0, 6.1, 6.2, 6.3, 6.4, 6.5, 6.6, 6.7, 6.8, 6.9 и 7.0.
a) Виды рода Brassica
Фраза ″виды рода Brassica″, используемая в настоящем описании, относится к любому из растений: Brassica juncea, Brassica napus, Brassica carinata, Brassica oleracea, Brassica nigra и Brassica campestris.
b) Химерная конструкция
Термин ″химерная конструкция″, используемая в настоящем описании, относится к комбинации молекул нуклеиновых кислот, не встречающемуся в естественном виде в природе. Соответственно, химерная конструкция может содержать регуляторные последовательности и кодирующие последовательности, полученные из различных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного источника, но расположенные в порядке, отличающемся от встречающегося в естественном виде в природе.
c) Кодирующая последовательность
Термин ″кодирующая последовательность″, используемый в настоящем описании, относится к последовательности ДНК, кодирующей определенную аминокислотную последовательность. Термин ″регуляторные последовательности″ относится к нуклеотидным последовательностям, расположенным в 3′-5′ направлении (5′ некодирующие последовательности), в составе кодирующей последовательности или в 5′-3′ направлении (3′ некодирующие последовательности) от кодирующей последовательности, влияющим на транскрипцию, процессинг и стабильность РНК, или на трансляцию соответствующей кодирующей последовательности. Регуляторные последовательности могут включать, но не ограничиваются укаанными: промоторы, лидерные последовательности трансляции, интроны и последовательности распознавания полиаденилирования.
d) Оптимизированный относительно кодона
Термин ″оптимизированный относительно кодона″, употребляемый в отношении гена или молекулы нуклеиновой кислоты, относится к гену или молекуле нуклеиновой кислоты, частота использования кодонов которых приспособлена для воспроизведения предпочтительной частоты использования кодонов в клетке-хозяине.
e) Комплементарность
Термин ″Комплементарность″, использцемый в настоящем описании, относится к степени соответствия между двумя сегментами ДНК. Комплементарность определяют путем оценки способности кодирующей нити одного сегмента ДНК гибридизироваться с некодирующей нитью другого сегмента ДНК, в соответствующих условиях, с образованием двойной спирали. В двойной спирали, аденин находится в составе одной нити, тимин находится в составе другой нити. Также, в тех положениях, в которых в составе одной нити находится гуанин, в составе другой находится цитозин. Чем больше степень соответствия между нуклеотидными последовательностями двух сегментов ДНК, тем выше способность образовывать гибридные дуплексы указанными двумя сегментами ДНК.
f) Кодируемый, гибридизация и жесткие условия
Термин ″кодируемый″, используемый в настоящем описании, относится к последовательности нуклеиновой кислоты, кодирующей последовательность полипептида, причем последовательность полипептида или ее фрагмент содержит аминокислотную последовательность, состоящую по меньшей мере из трех последовательных аминокислот, более предпочтительно - по меньшей мере 8 последовательных аминокислот, еще более предпочтительно - 15 последовательных аминокислот из состава полипептида, кодируемого последовательностью нуклеиновой кислоты.
Также в объем настоящего изобретения входит выделенная нуклеотидная последовательность, кодирующая фермент, обладающий десатуразной активностью в отношении PUFA, гибридизуемая в жестких условиях с нуклеиновой кислотой, нуклеотидная последовательность которой содержит или комплементарна нуклеотидной последовательности, содержащей SEQ ID NO:28 или SEQ ID NO:30 (См., ФИГ.9 и 11). Молекула нуклеиновой кислоты способна ″гибридизоваться″ с другой молекулой нуклеиновой кислоты, если молекула нуклеиновой кислоты в одноцепочечной форме может соединяться с дугой молекулой нуклеиновой кислоты в соответствующих температурных условиях и при соответствующей ионной силе (См., Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition (1989), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.)). Температурные условия и ионная сила определяют ″жесткость″ гибридизации. Для ″гибридизации″ необходимо, чтобы две нуклеиновые кислоты содержали комплементарные последовательности. Тем не менее, в зависимости от жесткости условий гибридизации, могут возникать несоответствия в основаниях. Соответствующая жесткость условий гибридизации нуклеиновых кислот зависит от длины нуклеиновых кислот и степени комплементарности. Указанные параметры хорошо известны специалистам в данной области техники. В частности, чем больше степень сходства или гомологичности двух нуклеотидных последовательностей, тем большая величина Tm соответствует гибридам нуклеиновых кислот, содержащих указанные последовательности. Для гибридов, длина которых составляет более 100 нуклеотидов, были получены уравнения для расчета Tm (См., Sambrook et al., выше). Для гибридизации с более короткой нуклеиновой кислотой, положение несоответствий имеет большее значение, и длина олигонуклеотида определяет его специфичность (См., Sambrook et al., выше).
Обычно, жесткими условиями считаются такие условия, в которых концентрация солей составляет менее приблизительно 1.5 M ионов Na, обычно приблизительно от 0.01 до 1.0 M ионов Na (или других солей) при pH от 7.0 до 8.3 и температуре по меньшей мере приблизительно 30°C для коротких зондов (например, от 10 до 50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов (например, длина которых составляет более 50 нуклеотидов). Жесткие условия также можно создать путем добавления дестабилизирующих агентов, например, формамида. Пример минимально жестких условий включает гибридизацию с применением буферного раствора с 30-35% формамидом, 1 M NaCl, 1% SDS (додецилсульфат натрия) при 37°C, и отмывкой в 1× - 2×SSC (20×SSC=3.0 M NaCl/0.3 M тринатриевым цитратом) при 50-55°C. Пример умеренно жестких условий включает гибридизацию в 40-45% формамиде, 1 M NaCl, 1% SDS при 37°C, и отмывкой в 0.5× to 1×SSC при 55-60°C. Пример очень жестких условий включает гибридизацию в 50% формамиде, 1 M NaCl, 1% SDS при 37°C, и отмывкой в 0.1×SSC при 60-65°C.
g) Экзон
Термин ″экзон″, используемый в настоящем описании, относится к участку последовательности гена, подвергаемому транскрипции и обнаруживаемому в составе зрелой информационной РНК, полученной на основе гена, но не обязательно являющемуся частью последовательности, кодирующей конечный продукт гена.
h) Экспрессия, антисмысловое ингибирование и ко-супрессия
Термин ″экспрессия″, используемый в настоящем описании, относится к образованию функционально активного конечного продукта. Экспрессия гена включает транскрипцию гена и трансляцию мРНК в предшественник зрелого белка.
Выражение ″антисмысловое ингибирование″, используемое в настоящем описании, относится к образованию антисмысловых транскриптов РНК, способных подавлять экспрессию целевого белка.
Термин ″ко-супрессия″, используемый в настоящем описании, относится к образованию смысловых транскриптов РНК, способных подавлять экспрессию идентичных или, по существу, аналогичных посторонних или эндогенных генов (См. патент США №5,231,020).
i) Фрагмент или субфрагмент, являющийся функционально эквивалентным
Термины ″фрагмент или субфрагмент, являющийся функционально эквивалентным″ и ″функционально активный фрагмент или субфрагмент″, используемые взаимозаменяемо в настоящем описании, относятся к фрагменту или субпоследовательности молекулы выделенной нуклеиновой кислоты, сохраняющим способность влиять на экспрессию гена или проявлять определенный фенотип, независимо от того, кодирует ли указанный фрагмент или субфрагмент активный фермент. Например, фрагмент или субфрагмент можно применять для разработки химерных конструкций, предназначенных для получения целевого фенотипа трансформированного растения. Химерные конструкции можно разрабатывать с целью применения для ко-супрессии или антисмыслового ингибирования путем присоединения фрагмента или субфрагмента нуклеиновой кислоты, независимо от того, кодирует ли он активный фермент, в соответствующем положении относительно промоторной последовательности растения.
j) Ген, нативный ген и трансген
Термин ″ген″, используемый в настоящем описании, относится к молекуле нуклеиновой кислоты, экспрессирующей определенный белок, включая регуляторные последовательности, предшествующие (5′ не-кодирующие последовательности) и следующие за (3′ не-кодирующие последовательности) кодирующей последовательностью.
Выражение ″нативный ген″, используемое в настоящем описании, относится к гену, обнаруживаемому в природе, вместе с характерными для него регуляторными последовательностями.
Термин ″трансген″, используемый в настоящем описании, относится к гену, введенному в состав генома с помощью процедуры трансформации.
k) Виды рода Gossypium
Выражение ″виды рода Gossypium″, употребляемое в настоящем описании, относится к любому из растений Gossypium arboreum, Gossypium barbadense, Gossypium herbaceum, Gossypium hirsutum, Gossypium hirsutum var hirsutum, Gossypium hirsutum var marie - galante, Gossypium lapideum, Gossypium sturtianum, Gossypium thuberi, Gossypium thurberi, Gossypium tomentosum или Gossypium tormentosum.
l) Гомология
Термины ″гомология″, ″гомологичный″, ″по существу, аналогичный″ и ″по существу, соответствующий″, используются взаимозаменяемо в настоящем описании и относятся к молекулам нуклеиновых кислот, изменения в одном или более нуклеотидных оснований в составе которых не влияют на способность молекулы нуклеиновой кислоты опосредовать экспрессию гена или влиять на проявление определенного фенотипа. Указанные термины также относятся к таким модификациям молекул нуклеиновых кислот согласно настоящему изобретению, как делеция или введение одного или более нуклеотидов, по существу, не влияющим на функциональные свойства получаемой молекулы нуклеиновой кислоты по сравнению с исходной молекулой, не содержащей изменений. Следовательно, для специалиста в данной области техники будет очевидно, что в объем настоящего изобретения входит более одной примерной определенной последовательности.
m) Клетка-хозяин
Выражение ″клетка-хозяин″, используемое в настоящем описании, обозначает клетку, содержащую выделенную последовательность нуклеиновой кислоты или ее фрагмент, согласно настоящему изобретению. Клетки-хозяева могут представлять собой клетки прокариот (например, Escherichia coli, цианобактерии и Bacillus subtilis) или клетки эукариот (например, клетки грибов, насекомых, растений или млекопитающих).
Примерами клеток грибов, подходящих для применения, являются Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Kluyveromyces spp., Hansenula spp., Aspergillus spp., Penicillium spp., Neurospora spp., Trichoderma spp. и Pichia spp. Особо предпочтительными клетками грибов являются клетки Saccharomyces cerevisiae.
Клетки растений могут представлять собой клетки однодольных или двудольных растений. Особо предпочтительными клетками растений являются клетки Glycine max (например, соя), видов рода Brassica, Carthamus tinctorius L. (например, сафлор), Helianthus annuus (например, подсолнечника), Zea mays (например, кукруза), видов рода Gossypium и Linum usitatissimum (например, лен).
n) Идентичность, идентичность последовательностей и процент идентичности последовательностей (% идентичности)
Термины ″идентичность″ или ″идентичность последовательностей″, используемые взаимозаменяемо в настоящем описании в контексте нуклеотидных последовательностей или последовательностей полипептидов, относятся к одинаковым азотистым основаниями или аминокислотным остаткам в составе двух последовательностей, при выравнивании на максимальное соответствие в определенном окне сравнения. Следовательно, идентичность определяется степенью подобия, соответствия или эквивалентности одинаковых цепей (кодирующих или не кодирующих) двух сегментов ДНК или полипептидов.
″Процент идентичности последовательностей″ или ″% идентичности″ вычисляют в ходе сравнения двух оптимально выровненных последовательностей на определенном участке, путем определения числа положений, в которых в обеих последовательностях находится идентичное основание, для получения числа соответствующих положений, деления числа указанных положений на общее число положений в сравниваемом участке и умножения результата на 100. Оптимальное выравнивание последовательностей можно осуществлять с помощью алгоритма Smith & Waterman, Appl. Math. 2:482 (1981), алгоритма Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), способа Pearson & Lipman, Proc. Natl. Acad. Sci. (USA) 85:2444 (1988) и с помощью компьютерных программ, применяющих соответствующие алгоритмы (например, Higgins et al., CABIOS. 5L151-153 (1989)), FASTDB (Intelligenetics), BLAST (National Center for Biomedical Information; Altschul et al., Nucleic Acids Research 25:3389-3402 (1997)), PILEUP (Genetics Computer Group, Madison, Wis.) or GAP, BESTFIT, FASTA and TFASTA (Wisconsin Genetics Software Package Release 7.0, Genetics Computer Group, Madison, Wis.). (См. патент США №5,912,120). Пригодные примеры процента идентичности последовательностей включают, но не ограничиваются указанными, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Указанные величины процента идентичности можно определять с помощью любой из программ, описанных в настоящей заявке.
о) Прямо или опосредованно
Термин ″опосредованно″, используемый в настоящем описании в отношении применения гена и соответствующего ему фермента для получения полиненасыщенных жирных кислот, обозначает превращение первой кислоты во вторую кислоту (то есть, наличие промежуточного продукта пути) с помощью первого фермента (например, LA превращают в EDA, например, с помощью Δ9-элонгазы), после чего вторую кислоту превращают в третью кислоту с помощью второго фермента (например, EDA превращают в DGLA например, с помощью Δ8-десатуразы).
Термин ″прямо″, используемый в настоящем описании в отношении применения гена и соответствующего ему фермента для получения полиненасыщенных жирных кислот, обозначает непосредственное превращение ферментом первой кислоты во вторую кислоту, при этом вторую кислоту впоследствии применяют в составе композиции (например, превращение LA в EDA например, с помощью Δ9-элонгазы или ω3-ETra в ω3-ETA например, с помощью Δ8-десатуразы).
p) Интрон
Термин ″интрон″, используемый в настоящем описании, относится к промежуточной последовательности в составе гена, не кодирующей часть последовательности белка. Следовательно, указанные последовательности транскрибируются в РНК, но после этого исключаются и не транслируются. Термин также применяется для обозначения исключенных последовательностей РНК.
q) Выделенный
Термин ″выделенный (выделенная)″, используемый в настоящем описании, относится к молекуле нуклеиновой кислоты (ДНК или РНК) или белку и его участку, обладающему биологической активностью, извлеченной из естественного окружения или источника, с помощью стандартных методик, известных в данной области техники (например, из бактерии, водоросли, гриба, растений, позвоночных, млекопитающих, и т.д.). Выделенные молекулы нуклеиновых кислот или белки являются, по существу, или главным образом свободными от компонентов, в нормальных условиях сопутствующих или взаимодействующих с молекулами нуклеиновых кислот или белков в их естественном окружении.
r) Фрагмент выделенной нуклеиновой кислоты или последовательность выделенной нуклеиновой кислоты
Выражения ″фрагмент выделенной нуклеиновой кислоты″ или "последовательность выделенной нуклеиновой кислоты", используемые в настоящем описании, относятся к одно- или двухцепочечному полимеру РНК или ДНК, возможно, содержащему синтезированные, не природные или измененные азотистые основания. Фрагмент выделенной нуклеиновой кислоты в форме полимера ДНК может включать один или более сегментов кДНК, геномной ДНК или синтезированной ДНК. (″Фрагмент″ определенного полинуклеотида относится к последовательности полинуклеотида, включающей непрерывную последовательность, длина которой составляет по меньшей мере приблизительно 10 последовательных нуклеотидов по меньшей мере приблизительно 15 последовательных нуклеотидов по меньшей мере приблизительно 20 последовательных нуклеотидов, и т.д., идентичной или комплементарной участку указанной нуклеотидной последовательности.) Для нуклеотидов (обычно находящихся в форме их 5′-монофосфатов) использованы их следующие однобуквенные обозначения: ″A″ для аденилата или дезоксиаденилата (для РНК или ДНК, соответственно), ″C″ для цитидилата или дезоксицитидилата, ″G″ для гуанилата или дезоксигуанилата, ″U″ для уридилата, ″T″ для дезокситимидилата, ″R″ для пуринов (A или G), ″Y″ для пиримидинов (C или T), ″K″ для G или T, ″H″ для A или C или T, ″I″ для инозина и ″N″ для любого нуклеотида.
s) Зрелый и предшественник
Термин ″зрелый″, используемый в настоящем описании в отношении термина ″белок″, относится к полипептиду, прошедшему пост-трансляционные модификации; то есть, к белку, из состава которого удалили все пре- или пропептиды, присутствующие в первичном продукте трансляции.
Термин ″предшественник″, используемый в настоящем описании в отношении термина ″белок″, относится к первичному продукту трансляции мРНК; то есть, в составе которого еще присутствуют пре- и пропептиды. Пре- и пропептиды могут являться сигналами внутриклеточной локализации, но не ограничиваются указанными.
t) 3′ He-кодирующие последовательности
Выражение ″3′-некодирующие последовательности″, используемое в настоящем описании, относится к последовательностям ДНК, расположенным в 5′-3′ направлении от кодирующей последовательности и включающим последовательности распознавания полиаденилирования и другие последовательности, кодирующие сигналы регуляции, способные влиять на процессинг мРНК или на экспрессию гена. Сигнал полиаденилирования часто характеризуется влиянием на внесение путей полиадениловой кислоты в 3′-конец предшественника мРНК. Применение различных 3′-некодирующих последовательностей описано у Ingelbrecht et al., (1989) Plant Cell 1:671-680.
u) He-природный, не встречающийся в природе
Выражения ″не природный″ или ″не встречающийся в природе″, используемые в настоящем описании, относятся к объекту, полученному искусственным путем, не соответствующему обычно встречающемуся в природе.
v) Функционально связанный
Выражение ″функционально связанный″, используемое в настоящем описании, относится к объединению последовательностей нуклеиновых кислот в одной молекуле нуклеиновой кислоты таким образом, что активность одной последовательности регулируется другой. Например, промотор функционально связан с кодирующей последовательностью, если он способен регулировать экспрессию указанной кодирующей последовательности (то есть, транскрипция кодирующей последовательности находится под контролем промотора). Кодирующая последовательность может быть функционально связана с регуляторными последовательностями в кодирующей или не кодирующей ориентации. В другом примере, комплементарные участки РНК согласно настоящему изобретению можно функционально связывать, прямо или опосредованно, с 5′-концом целевой мРНК или 3′-концом целевой мРНК, или вводить ее в пределы целевой мРНК, или первый комплементарный участок можно присоединять к 5′-концу, а участок, комплементарный ему - к 3′-концу целевой мРНК.
w) Растение
Термин ″растение″, используемый в настоящем описании, относится к целым растениям, органам растений, тканям растений, семенам, клеткам растений, семян и предшественникам всего перечисленного. Клетки растений включают, без ограничений, клетки семян, суспензии культур, эмбрионов, участков меристемы, клетки каллюса, листьев, корней, побегов, гаметофитов, спорофитов, пыльцу и микроспоры.
x) Полимеразная цепная реакция или ПЦР
Выражения ″полимеразная цепная реакция″ или ″ПЦР″, используемые в настоящем описании, относятся к методике синтеза большого количества специфичных сегментов ДНК, состоящей из серий повторяющихся циклов (Perkin Elmer Cetus Instruments, Norwalk, Conn.). Обычно, двухцепочечную ДНК разрушают при нагревании, при низкой температуре присоединяют два праймера, комплементарные 3′-границам сегмента - мишени, и затем последовательности удлиняют при средних значениях температуры. Один повтор трех приведенных последовательных этапов обозначают как цикл.
ПЦР представляет собой эффективную методику, применяемую для амплификации ДНК в миллионах повторов, путем повторения репликации матрицы, за короткий период времени (Mullis et al, Cold Spring Harbor Symp. Quant. Biol. 51:263-273 (1986); Erlich et al, заявка на европейский патент 50,424; заявка на европейский патент 84,796; заявка на европейский патент 258,017, заявка на европейский патент 237,362; Mullis, заявка на европейский патент 201,184, Mullis et al, патент США №4,683,202; Erlich, патент США №4,582,788; и Saiki et al, патент США №4,683,194). В ходе процесса применяют набор специфичных олигонуклеотидов, синтезированных in vitro, для начала синтеза ДНК. Структура праймеров зависит от целевых анализируемых последовательностей ДНК. Методику осуществляют в ходе большого количества циклов (обычно 20-50) плавления матрицы при высокой температуре, позволяющих праймерам присоединится к комплементарным последовательностям матрицы с последующей репликацией матрицы с помощью ДНК-полимеразы. Продукты реакции ПЦР анализируют путем разделения в агарозном геле с последующей окраской бромидом этидия и визуализацией с помощью УФ просвечивания. В качестве альтернативного варианта, в реакции ПЦР можно вносить радиоактивные дНТФ для включения меток в продукты. В данном случае продукты ПЦР визуализируют путем приведения геля в контакт с рентгеновской пленкой. Дополнительным преимуществом внесения радиоактивных меток в продукты ПЦР является возможность количественной оценки уровней отдельных продуктов амплификации.
y) Промотор и энхансер
Термин ″промотор″, используемый в настоящем описании, относится к последовательности ДНК, способной контролировать экспрессию кодирующей последовательности или функционально активной РНК. Последовательность промотора состоит из проксимальных и более удаленных в 3′-5′ направлении элементов, последние элементы часто обозначают как энхансеры.
Термин ″энхансер″, используемый в настоящем описании, относится к последовательности ДНК, способной стимулировать активность промотора и возможно, являющейся естественным элементом промотора или гетерологичным элементом, введенным для повышения уровня тканеспецифичности промотора. Промоторная последовательность также может быть расположена внутри транскрибируемых участков генов и/или 5′-3′ направлении относительно транскрибируемых генов. Промоторы могут целиком происходить из нативного гена или состоять из различных элементов, полученных из различных промоторов, встречающихся в природе, или даже включать различные синтезированные сегменты ДНК. Специалистам в данной области техники следует понимать, что различные промоторы могут контролировать экспрессию гена в различных типах тканей или клеток, или на разных стадиях их развития или в зависимости от различных условий окружающей среды. Промоторы, стимулирующие экспрессию гена в большинстве типов клеток в большинстве случаев, обычно обозначают как ″конститутивные промоторы″. Постоянно обнаруживаются новые промоторы различных типов, пригодные для применения в клетках растений; несколько примеров можно найти в компиляции, составленной Okamuro and Goldberg, (1989) Biochemistry of Plants 15:1-82. Далее обнаруживается, что, поскольку в большинстве случаев не были окончательно установлены точные границы регуляторных последовательностей, некоторые разновидности молекул ДНК могут обладать идентичной промоторной активностью.
z) Рекомбинантный
Термин ″рекомбинантный″, используемый в настоящем описании, относится к искусственному сочетанию двух сегментов последовательности, обычно расположенных отдельно, например, путем химического синтеза или манипуляций с выделенными сегментами нуклеиновых кислот с помощью методик генной инженерии.
aa) Рекомбинантная конструкция, экспрессионная конструкция и рекомбинантная экспрессионная конструкция
Выражения ″рекомбинантная конструкция″, ″экспрессионная конструкция″ и ″рекомбинантная экспрессионная конструкция″ используются взаимозаменяемо в настоящем описании и относятся к функциональной единице генетического материала, которую можно включать в состав генома клетки с помощью стандартных методик, хорошо известных специалисту в данной области техники. Указанную конструкцию можно применять отдельно или совместно с вектором. В случае применения вектора, его выбирают в зависитмости от способа, применяемого для трансформации растений-хозяев, как хорошо известно специалисту в данной области техники. Например, можно применять плазмидный вектор. Специалист в данной области техники хорошо осведомлен о генетических элементах, присутствие которых в составе вектора необходимо для успешной трансформации, селекции и размножения клеток-хозяев, содержащих молекулу любой из выделенных нуклеиновых кислот согласно настоящему изобретению. Также для специалиста в данной области техники будет понятно, что различные независимые события трансформации приведут к различным уровням и характеру экспрессии (Jones et al., (1985) EMBO J. 4:2411-2418; De Almeida et al., (1989) Mol. Gen. Genetics 218:78-86), и, следовательно, следует рассмотреть большое количество событий для получения линий, представляющих целевой уровень и характер экспрессии. Подобное скрининговое исследование можно провести путем анализа ДНК с помощью Саузерн-блоттинга, анализа экспрессии мРНК с помощью Нозерн-блоттинга, анализа экспрессии белка с помощью Вестерн-блоттинга или анализа фенотипа.
bb) Транскрипт РНК, информационная РНК, кДНК, функционально активная РНК и эндогенная РНК
Выражение ″транскрипт РНК″, используемое в настоящем описании, относится к продукту, образующемуся в результате транскрипции последовательности ДНК, катализируемой РНК-полимеразой. Если транскрипт РНК представляет собой абсолютно комплементарную копию последовательности ДНК, его обозначают как первичный транскрипт, или он может представлять собой последовательность РНК, полученную в результате посттранскрипционного процессинга первичного транскрипта и обозначаться как зрелая РНК.
Выражение ″информационная РНК (мРНК)″, используемое в настоящем описании, относится к РНК, не содержащей в составе интронов и способной транслироваться клеткой в белок.
Термин ″кДНК″, используемый в настоящем описании, относится к ДНК, являющейся комплементарной и синтезированной на основе матрицы мРНК с помощью фермента обратной транскриптазы. Молекула кДНК может быть одноцепочечной или ее можно преобразовать в двухцепочечную форму с помощью молекулы полимеразы I Кленова. ″Смысловая (кодирующая)″ РНК относится к траскрипту ДНК, включающему мРНК и способному транслироваться в белок в клетке или in vitro. ″Антисмысловая (некодирующая) РНК″ относится к транскрипту РНК, являющемуся комплементарным целиком первичному транскрипту-мишени или его части, и блокирующему экспрессию гена-мишени (патент США №5,107,065). Антисмысловая РНК может быть комплементарна любому участку определенного транскрипта гена, то есть, 5′ некодирующей последовательности, 3′ некодирующей последовательности, интронам или кодирующей последовательности.
Выражение ″функционально активная ДНК″, используемое в настоящем описании, относится к антисмысловой РНК, рибозимной РНК или РНК другого типа, еще не прошедшей трансляцию, но воздействующей на процессы в клетке.
Термины ″комплементарная цепь″ и ″обратно комплементарная цепь″ используются взаимозаменяемо в настоящем описании по отношению к транскриптам мРНК, и обозначают антисмысловую РНК транскрипта.
Выражение ″эндогенная РНК″, используемое в настоящем описании, относится к любой молекуле РНК, кодируемой любой последовательностью нуклеиновой кислоты, присутствующей в геноме хозяина до трансформации с помощью рекомбинантной конструкции согласно настоящему изобретению, как встречающейся, так и не встречающейся в природе, то есть, внесенной путем рекомбинации, мутагенеза, и т.д..
cc) Сходство
Термин ″сходство″, используемый в настоящем описании в отношении ″сходства″ между двумя аминокислотными последовательностями, белками или полипептидами, относится к группам идентичных, а также консервативных аминокислотных остатков, в составе обеих последовательностей. Чем выше степень сходства между двумя аминокислотными последовательностями, тем выше соответствие, подобие или эквивалентность двух последовательностей.
dd) Устойчивая трансформация, временная трансформация и трансформация
Выражение ″устойчивая трансформация″, используемое в настоящем описании, относится к переносу молекулы нуклеиновой кислоты в геном организма-хозяина, включая как ядерный геном, так и геном, присутствующий в органеллах, приводящему к генетически устойчивому наследованию.
Напротив, выражение ″временная трансформация″, используемое в настоящем описании, относится к переносу молекулы нуклеиновой кислоты в ядро или ДНК-содержащую органеллу организма-хозяина, приводящему к экспрессии гена без внедрения или устойчивого наследования. Организмы-хозяева, содержащие трансформированные молекулы нуклеиновых кислот, обозначают как ″трансгенные″ организмы. Предпочтительным способом трансформации клеток риса, кукурузы и других однодольных является применение технологии трансформации с помощью ускоренных частиц или ″генной пушки″ (Klein et al., (1987) Nature (London) 327:70-73; патент США №4,945,050) или способ, включающий применение агробактерий с соответствующей Ti плазмидой, несущей трансген (Ishida Y. et al., (1996) Nature Biotech. 14:745-750).
Термин ″трансформация″, используемый в настоящем описании, относится как к устойчивой трансформации, так и к временной трансформации.
ee) Лидерная последовательность трансляции
Выражение ″лидерная последовательность трансляции″, используемое в настоящем описании, относится к последовательности ДНК, расположенной между промоторной последовательностью гена и кодирующей последовательностью. Лидерная последовательность трансляции присутствует в полностью процессированной мРНК в 3′-5′ направлении от последовательности начала трансляции. Лидерная последовательность трансляции может влиять на процессинг первичного транскрипта в мРНК, устойчивость мРНК или точность трансляции. Примеры лидерных последовательностей трансляции известны (Turner, R. and Foster, G.D. (1995) Molecular Biotechnology 3:225).
Все патенты, патентные публикации и приоритетные документы, цитированные в настоящем описании, полностью включены в настоящую заявку посредством ссылки.
B. Ген Δ8-десатуразы и фермент, кодируемый указанным геном
Фермент, кодируемый геном Δ8-десатуразы, согласно настоящему изобретению, необходим для образования полиненасыщенных жирных кислот (PUFA), содержащих в составе, по меньшей мере, два участка ненасыщенности (двойные связи), полная длина которых составляет 20 атомов углерода или более. В частности, фермент согласно настоящему изобретению является функционально активным (например, обладает активностью Δ8-десатуразы), что означает, что он вносит двойную связь между атомом углерода номер 8 (C8) и атомом углерода номер 9 (C9) в PUFA, длина цепи которой составляет, по меньшей мере, 20 атомов углерода, и которая уже содержит двойные связи в Δ9, Δ12 и/или Δ15 положениях. Как показано на ФИГ.1, фермент, кодируемый геном Δ8-десатуразы, согласно настоящему изобретению, образует PUFA, длина цепей которых составляет 20 атомов углерода или более, в ходе альтернативного пути Δ8-десатуразы/Δ9-элонгазы. Субстраты, ω6-эйкозадиеновая кислота, ω3-эйкозатриеновая кислота или обе кислоты, ω6-эйкозадиеновая и ω3-эйкозатриеновая, используются Δ8-десатуразой согласно настоящему изобретению, в ходе указанного пути.
Ген Δ8-десатуразы согласно настоящему изобретению, был выделен из генома Emiliana sp., точнее, Emiliana huxleyi, в частности, Emiliana huxleyi CCMP 378. Нуклеотидная последовательность выделенного гена Δ8-десатуразы из генома Emiliana huxleyi CCMP 378 показана на Фигуре 9 и в SEQ ID NO:28. Выделенная нуклеотидная последовательность предполагаемой нуклеотидной последовательности, оптимизированная относительно кодонов, показана на Фигуре 11 и в SEQ ID NO:30. Выделенная или очищенная аминокислотная последовательность, кодируемая SEQ ID NO:28 и SEQ ID NO:30, показана на Фигуре 10 и в SEQ ID NO:29.
Химическое превращение LA в DGLA и ALA в ω3-ETA с помощью фермента Δ9-элонгазы и фермента Δ8-десатуразы называют альтернативным путем Δ8-десатуразы/Δ9-элонгазы. Классический путь Δ6 для превращения LA в DGLA и ALA в ω3-ETA использует фермент Δ6-десатуразу для преобразования LA в GLA и ALA в SDA, и ген Δ6-элонгазы для преобразования GLA в DGLA и SDA в ω3-ETA, соответственно. В обоих путях, образование ARA или EPA затем катализируется, например, Δ5-десатуразой. DHA, например, можно получать путем преобразования EPA в ω3-докозапентаеновую кислоту (DPA), и ω3-докозапентаеновую кислоту в DHA, с использованием, например, C20-элонгазы и Δ4-десатуразы, соответственно.
Хотя, например, DGLA, ARA, ω3-ETrA, ω3-ETA, EPA, DPA и/или DHA можно получать как с помощью альтернативного пути Δ8-десатуразы/Δ9-элонгазы, так и с помощью классического пути Δ6, в некоторых случаях альтернативный путь Δ8-десатуразы/Δ9-элонгазы может являться более предпочтительным, чем классический путь Δ6. Например, если в ходе получения DGLA, ARA, ω3-ETrA, ω3-ETA, EPA, DPA и/или DHA, образование побочных промежуточных продуктов омега-6 или омега-3 жирных кислот, например GLA или SDA, является нежелательным, вместо классического пути Δ6 можно применять альтернативный путь Δ8-десатуразы/Δ9-элонгазы, для того, чтобы избежать образования GLA и SDA.
Как было упомянуто выше, Δ8-десатураза является необходимым ферментом пути Δ8-десатуразы/Δ9-элонгазы. EPA, например, не может быть синтезирована в ходе альтернативного пути Δ8-десатуразы/Δ9-элонгазы без гена Δ8-десатуразы и фермента, кодируемого им. Как показано на ФИГ.1, выделенный фермент Δ8-десатураза согласно настоящему изобретению превращает, например, EDA в DGLA и ω3-ETrA в ω3-ETA. Затем образование ω3-ETA из ω3-ETrA, и EPA из ω3-EPA катализируется, например, Δ8-десатуразой и Δ5-десатуразой, соответственно. В результате применения альтернативного пути Δ8-десатуразы/Δ9-элонгазы удается избежать образования жирных кислот GLA и SDA.
Также согласно настоящему изобретению предложены выделенные или очищенные нуклеотидные последовательности (и соответствующие белки, кодируемые ими), последовательности которых содержат или комплементарны, по меньшей мере, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% нуклеотидов в последовательности (то есть, обладают идентичностью с последовательностью), приведенной в SEQ ID NO:28 (выделенная нуклеотидная последовательность Δ8-десатуразы из генома Emiliana huxleyi CCMP 378) или SEQ ID NO:30 (выделенная нуклеотидная последовательность Emiliana huxleyi CCMP 378, оптимизированная относительно ко донов). Указанные последовательности могут происходить из организма человека или из других организмов (например, С.elegans или мыши).
Также в объем настоящего изобретения входят фрагменты и производные, содержащие или состоящие из нуклеотидной последовательности SEQ ID NO:28 или SEQ ID NO:30. Фрагменты, полученные на основе SEQ ID NO:28 или SEQ ID NO:30, могут содержать в составе от 10 до 1250 нуклеотидов, от 10 до 1000 нуклеотидов, от 10 до 750 нуклеотидов, от 10 до 500 нуклеотидов, от 10 до 250 нуклеотидов, от 10 до приблизительно 100 нуклеотидов или от 10 до приблизительно 50 нуклеотидов или от 15 до 40 нуклеотидов. В одном аспекте, фрагменты SEQ ID NO:28 и SEQ ID NO:30 кодируют полипептид, обладающий Δ8-десатуразной активностью. В другом аспекте, фрагменты SEQ ID NO:28 и SEQ ID NO:30 можно применять в качестве праймеров и зондов. Способы создания праймеров и зондов хорошо известны специалистам в данной области техники. Длина указанных праймеров и зондов может составлять от 10 до 50 нуклеотидов, предпочтительно от 15 до 40 нуклеотидов.
Также согласно настоящему изобретению предусмотрены варианты нуклеотидной последовательности SEQ ID NO:28 и SEQ ID NO:30. Указанные варианты могут содержать вставку, замену или делецию одной или более пар оснований, и предусматривается, что указанные вставки, замены или делеции не затрагивают ни одного из трех (3) высоко консервативных ″гистидиновых боксов″ в цитохром b5 - подобном участке, расположенном на 5′- конце SEQ ID NO:29 (См., ФИГ.2). Участки ″гистидиновых боксов″ и цитохром b5 - подобный участок более подробно описаны в настоящей заявке в связи с вариантами последовательности, приведенной в SEQ ID NO:29. Примеры вариантов нуклеотидных последовательностей, входящих в объем настоящего изобретения, приведены ниже, в Таблице A.
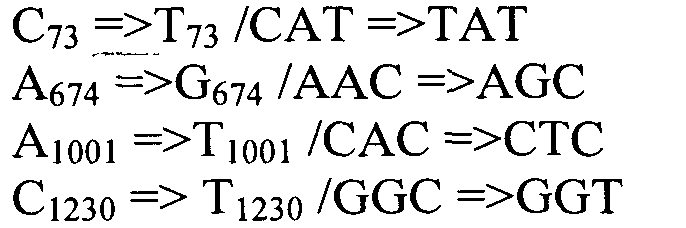


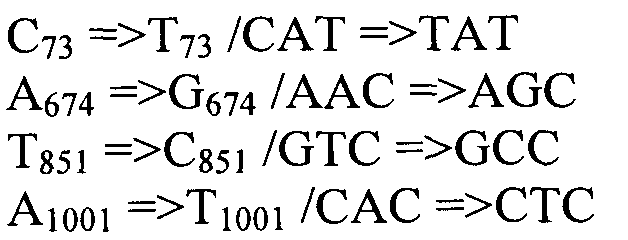
Также в объем настоящего изобретения входят нуклеотидные последовательности, полученные из других источников, и обладающие описанной выше комплементарностью или соответствием последовательности, приведенной в SEQ ID NO:28 или SEQ ID NO:30. Функциональные аналоги последовательности, приведенной в SEQ ID NO:28 или SEQ ID NO:30 (то есть, последовательности, включающие Δ8-десатуразу) также входят в объем настоящего изобретения.
Также в объем настоящего изобретения входят нуклеотидные последовательности или их фрагменты, обладающие Δ8-десатуразной активностью, причем аминокислотная последовательность указанного полипептида по меньшей мере на 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:29. Указанные последовательности могут происходить как из организма человека, так и из других источников (например, C.elegans или мыши).
Также настоящее изобретение обеспечивает выделенный и/или очищенный полипептид, десатурирующийполиненасыщенную жирную кислоту посредством внесения дополнительной двойной связи в положении между 8 и 9 атомами углерода (что означает, что указанный полипептид обладает Δ8-десатуразной активностью) в составе жирной кислоты, содержащей по меньшей мере 20 атомов углерода участок ненасыщенности у атома углерода в 9 положении, и по меньшей мере на 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% сходна с или идентична аминокислотной последовательности SEQ ID NO:29 (показанной на ФИГ.10). В частности, настоящее изобретение обеспечивает выделенный полипептид, содержащий аминокислотную последовательность, приведенную в SEQ ID NO:29.
Также настоящее изобретение обеспечивает фрагменты полипептида, содержащего последовательность, приведенную в SEQ ID NO:29. Длина указанных фрагментов может оставлять 10-400 последовательных аминокислот, 10-300 последовательных аминокислот, 10-200 последовательных аминокислот, 10-100 последовательных аминокислот, 10-50 последовательных аминокислот, 10-40 последовательных аминокислот или 10-30 последовательных аминокислот, 10-20 аминокислот. Указанные фрагменты можно применять, например, в качестве иммуногенов для получения антител. Также, указанные фрагменты можно применять в качестве специфичных участников связывания для одного или более иммунологических анализов.
Также настоящее изобретение обеспечивает варианты полипептида, содержащего последовательность, приведенную в SEQ ID NO:29. Указанные варианты могут содержать вставку, замену или делецию одной или более пар оснований, и предусматривается, что указанные вставки, замены или делеции не затрагивают ни одного из трех (3) высоко консервативных ″гистидиновых боксов″ в цитохром b5 - подобном участке, расположенном на 5′-конце SEQ ID NO:29 (См., ФИГ.2). Гистидиновые боксы расположены в 155-160 (HDYLH (SEQ ID NO:32)), 197-201 (HNTHH (SEQ ID NO:33)) и 355-359 (QTEHH (SEQ ID NO:34)) аминокислотных положениях SEQ ID NO:29 (См., ФИГ.2). Цитохром b5 - подобный участок на 5′-конце содержит консервативный мотив связывания гема HPGG (38-41 аминокислотные положения SEQ ID NO:29) (См., ФИГ.2). Цитохром b5 - подобный участок встречается в составе нескольких ферментов десатураз, связанных с мембраной ′передним концом′, например, Δ6-Δ5- и Δ4-десатуразы, участвующие в образовании PUFA с длинными цепями (См., Napier J A et al. (2003) Prostaglandins Leukot Essent Fatty Acids. 68:135-43). Считается, что цитохром b5 функционирует в качестве доноров электронов для указанным ферментов в ходе реакции десатурации, и нарушения в указанной области могут приводить к потере или изменению ферментативной активности (См., Sayanova O et al (1999) Plant Physiol. 121:641-646; Guillou H. et al (2004) J Lipid Res. 45:32-40). Примеры вариантов нуклеотидных последовательностей, входящих в объем настоящего изобретения, приведены ниже, в Таблице B.
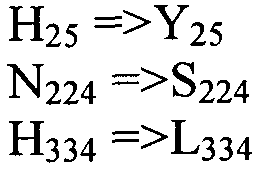
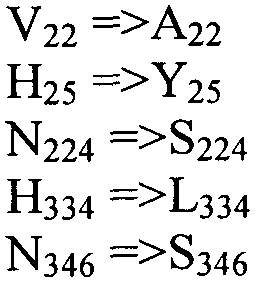
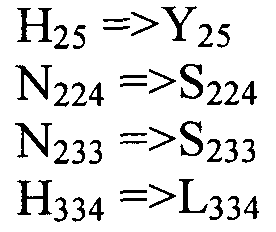
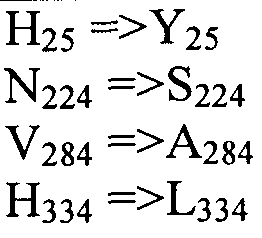
C. Получение фермента Δ8-десатуразы
После выделения и/или очистки последовательности нуклеиновой кислоты (например, гена), кодирующей фермент Δ8-десатуразу, ее можно вносить в клетку-хозяина, как прокариотическую, так и эукариотическую, с помощью вектора или конструкции. Вектор, например, бактериофаг, космида или плазмида, может содержать нуклеотидную последовательность, кодирующую фермент Δ8-десатуразу, а также любую регуляторную последовательность (например, промотор), являющиеся функционально активными в клетке-хозяине и способными стимулировать экспрессию фермента Δ8-десатуразы, кодируемой нуклеотидной последовательностью. Регуляторная последовательность находится в функциональном взаимодействии или функционально связана с нуклеотидной последовательностью. (Регуляторную последовательность называют ″функционально связанной″ с кодирующей последовательностью, если регуляторная последовательность влияет на транскрипцию или экспрессию кодирующей последовательности.) Примеры соответствующих промоторов включают, но не ограничиваются указанными: промоторы генов, кодирующих алко гольд егидрогеназу, глицеральдегид-3-фосфатдегидрогеназ, фосфоглюкоизомеразу, фосфоглицераткиназу, кислотную фосфатазу, T7, TPI, лактазу, металлотионеин, предранний белок цитомегаловируса, кислый белок сыворотки, глюкоамилазу, и промоторы, активирующиеся в присутствии галактозы, например, GAL1 и GAL10. Также, в состав вектора можно включать нуклеотидные последовательности, кодирующие другие белки, ферменты (например, Δ9-элонгазу), олигосахариды, липиды, и так далее, а также другие регуляторные последовательности, например, сигнал полиаденилирования (например, сигнал поли-A антигена SV-40T, овальбумин или гормон роста крупного рогатого скота). Выбор последовательностей для включения в состав конструкции зависит от целевых продуктов экспрессии, а также от природы клетки-хозяина.
Как было упомянуто выше, после конструирования вектора его можно ввести в целевую клетку-хозяина с помощью способов, известных среднему специалисту в данной области техники, включая, например, способ трансфекции, трансформации и электропорации (См., Molecular Cloning: A Laboratory Manual, 2 nd ed., Vol.1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press (1989)). После этого клетку-хозяина культивируют в соответствующих условиях, обеспечивающих экспрессию генов, ведущую к образованию целевой PUFA, которую затем восстанавливают и очищают с помощью рутинных методик, известных в данной области техники.
Примеры соответствующих клеток прокариот включают, но не ограничиваются указанными: клетки бактерий, например, Escherichia coli, Bacillus subtilis, а также клетки цианобактерий, например, Spirulina spp. (то есть, сине-зеленых водорослей). Клетки эукариот включают, но не ограничиваются указанными: клетки млекопитающих, клетки насекомых, клетки растений или клетки грибов. Клетки грибов включают, но не ограничиваются, указанными: Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Aspergillus spp., Penicillium spp., Neurospora spp., Kluyveromyces spp., Hansenula spp., Trichoderma spp. или Pichia spp. В частности, клетка гриба может являться клеткой дрожжей, включая, но не ограничиваясь указанными: Saccharomyces spp., Candida spp., Hansenula spp. и Pichia spp. Клетка дрожжей также может являться клетокой Saccharomyces cerevisiae. Клетки растений включают, но не ограничиваются указанными: клетки Glycine max (например, сои), видов рода Brassica, Carthamus tinctorius L. (например, сафлора), Helianthus annuus (например, подсолнечника), Zea mays (например, кукурузы), видов рода Gossypium и Linum usitatissimum (например, льна).
Экспрессию клетки-хозяина можно осуществлять во временной или устойчивой форме. Временную экспрессию можно осуществлять с помощью внесенных конструкций, содержащих сигналы экспрессии, функционально активные в клетке-хозяине, но указанные конструкции не реплицируются и редко интегрируются в клетку-хозяин, или в не пролиферирующей клетке. Также временную экспрессию можно осуществлять путем стимулирования активности регулируемого промотора, функционально связанного с целевым геном, хотя указанные индуцируемые системы часто проявляют низкий основной уровень экспрессии. Устойчивая экспрессия обеспечивается путем внесения конструкции, способной интегрироваться в геном клетки-хозяина, или автономно реплицирующейся в клетке-хозяине. Селекцию по признаку устойчивой экспрессии целевого гена можно проводить с помощью селективного маркера, находящегося в составе или трансфицируемого вместе с экспрессионной конструкцией, с последующей селекцией клеток, экспрессирующих маркер. Если устойчивую экспрессию осуществляют путем интегрирования, сайт интегрирования конструкции в геноме хозяина может быть случайным, или можно осуществлять направленное интегрирование путем применения конструкций, содержащих области гомологии с геномом хозяина, достаточные для направленной рекомбинации с локусом генома хозяина. Если конструкции направляют на эндогенный локус, все или некоторые участки, регулирующие транскрипцию и трансляцию, могут быть получены из эндогенного локуса.
Также экспрессию фермента Δ8-десатуразы и, в конечном итоге, целевой (целевых) PUFA, можно проводить с помощью трансгенного млекопитающего. В частности, после создания описанной выше конструкции, ее можно вносить в пронуклеус зародыша. После этого зародыш можно имплантировать в самку-реципиента. В качестве альтернативного варианта можно применять метод переноса ядра (Schnieke et al., Science 278:2130-2133 (1997)). После этого обеспечивают прохождение процессов созревания и рождения (См., например, патент США №5,750,176 и патент США №5,700,671). Молоко, ткань или другие жидкие образцы, полученные от потомства, будут содержать повышенные уровни PUFA относительно нормальных уровней содержания в организме не трансгенного животного. Можно наблюдать последующие поколения для выявления образования измененных или повышенных уровней PUFA и, следовательно, включения гена, кодирующего целевой фермент десатуразу, в их геномы. В качестве хозяев можно использовать таких млекопитающих, как, например, мышь, крыса, кролик, свинья, коза, овца, лошадь и корова. Тем не менее, можно использовать любых других млекопитающих, при условии, что существует возможность введения ДНК, кодирующей целевой фермент, в геном указанных млекопитающих.
Для экспрессии полипептида Δ8-десатуразы, функционально активные участки инициации и терминации транскрипции и трансляции функционально связывают с ДНК, кодирующей полипептид десатуразы. Участки инициации и терминации транскрипции и трансляции получают из различных общедоступных источников, включая экспрессируемую ДНК, гены, для которых известна или предполагается способность к экспрессии в целевой системе, векторы экспрессии, химический синтез или получение из эндогенного локуса в клетке-хозяине. В некоторых случаях существует возможность экспрессии в ткани и/или части растения, в частности, если ткань или часть растения созревают рано, например, семена, листья, фрукты, цветы, корни и т.д.. Экспрессию можно осуществлять в указанной части растения путем применения определенной регуляторной последовательности, например, описанной в патентах США №5,463,174, 4,943,674, 5,106,739, 5,175,095, 5,420,034, 5,188,958 и 5,589,379.
В качестве альтернативного варианта, экспрессируемый белок может представлять собой фермент, образующий продукт, который можно вводить, как прямо, так и с помощью дальнейших модификаций, в жидкую составляющую, получаемую из растения-хозяина. Экспрессия гена Δ8-десатуразы или антисмысловых транскриптов Δ8-десатуразы способна изменять уровни определенных PUFA или их производных, обнаруживаемых в частях и/или тканях растения.
Участок, кодирующий полипептид Δ8-десатуразы, можно экспрессировать как отдельно, так и вместе с другими генами (например, с геном, кодирующим Δ9-элонгазу, геном, кодирующим Δ5-десатуразу, геном, кодирующим Δ17-десатуразу, геном, кодирующим Δ5-элонгазу и/или геном, кодирующим Δ4-десатуразу), для получения тканей и/или частей растения с высоким содержанием целевых PUFA, или сочетание PUFA в составе которых имеет большее сходство с грудным молоком человека (См., WO 95/24494). Участок терминации можно получать из 3′-участка гена, из которого получали участок инициации, или другого гена. Известно об удовлетворительном функционировании большого количества участков терминации в различных хозяевах, принадлежащих к одному и тому же или к различным родам и видам. Обычно участок терминации выбирают скорее из соображений удобства, чем на основании какого-либо определенного свойства.
Как было упомянуто выше, растение (например, Glycine max или Brassica napus (canola)) или ткань растения также можно использовать в качестве хозяина или клетки-хозяина, соответственно, для экспрессии фермента Δ8-десатуразы, который, в свою очередь, можно использовать, например, для получения полиненасыщенных жирных кислот. В частности, целевые PUFA можно экспрессировать в семени. Способы выделения масел из семян известны в данной области техники. Следовательно, в дополнение к получению источника PUFA, на компоненты масла из семян можно воздействовать путем экспрессии гена Δ8-десатуразы, а также генов элонгазы (например, Δ9-элонгазы, Δ5-элонгазы, и т.д.) и генов других десатураз (например, Δ5- десатуразы, Δ17-десатуразы, Δ4-десатуразы, и т.д.), для получения масел из семян, которые можно включать в состав пищевых композиций, фармацевтических композиций, еды для животных и косметических средств. Кроме того, вектор, содержащий последовательность ДНК, кодирующую Δ8-десатуразу, функционально связанную с промотором, вносят в ткань растения или растение, на период времени и в условиях, достаточных для экспрессии гена Δ8-десатуразы. Вектор также может включать один или более генов, кодирующих другие ферменты, например, Δ4-десатуразу, Δ5-десатуразу, Δ6-десатуразу, Δ10-десатуразу, Δ12-десатуразу, Δ15-десатуразу, Δ17-десатуразу, Δ19-десатуразу, Δ9-элонгазу, Δ6-элонгазу и/или Δ5-элонгазу. Ткань растения или растение может обеспечивать соответствующий субстрат для действия ферментов, или в ткань растения, клетку растения или растения можно вносить вектор, кодирующий ферменты, обеспечивающие указанный субстрат. Также, субстрат можно наносить на ткани растения, в которых осуществляется экспрессия соответствующих ферментов. С помощью различных приведенных методик можно производить PUFA путем применения клетки растения, ткани растения или растения. Следует заметить, что в объем настоящего изобретения также входит трансгенное растение, содержащее описанный выше вектор, экспрессия нуклеотидной последовательности вектора в котором приводит к образованию полиненасыщенной жирной кислоты, например, в семенах трансгенного растения.
Воспроизведение, развитие и культивирование растений, полученных из трансформантов одного протопласта растения или из различных трансформированных эксплантатов, хорошо известно в данной области техники (Weissbach and Weissbach, In: Methods for Plant Molecular Biology, (Eds.), Academic Press, Inc. San Diego, Calif., (1988)). Указанное воспроизведение и процесс роста в обычном случае включают этап селекции трансформированных клеток, культивирование указанных отдельных клеток в ходе обычных стадий эмбрионального развития до стадии проростка, имеющего корни. Трансгенные зародыши и семена воспроизводятся сходным образом. Полученные в результате зародыши, имеющие корни, сажают на соответствующую среду для роста растений, например, в почву.
Процессы развития или воспроизведения растений, содержащих посторонний, экзогенный ген, кодирующий целевой белок, хорошо известны в данной области техники. Предпочтительно, растения проходят самоопыление для получения гомозиготных трансгенных растений. Другим путем является перекрестное опыление выросших из семян растений из агрономически значимых линий с помощью пыльцы, полученной от вторичной генерации растений. И наоборот, пыльцу, полученную от растений указанных агрономически значимых линий, применяют для опыления растений вторичной генерации. Трансгенное растение согласно настоящему изобретению, содержащее целевой полипептид, культивируют с помощью способов, известных специалисту в данной области техники.
Существует множество способов воспроизведения растений из ткани растения. Конкретный способ воспроизведения будет зависеть от исходной ткани растения и определенных видов растений, предназначенных для воспроизведения.
Способы трансформации двудольных, в первую очередь, путем применения Agrobacterium tumefaciens, и получения трансгенных растений, были опубликованы на примере хлопка (патент США №5,004,863, патент США №5,159,135, патент США №5,518,908); соевых (патент США №5,569,834, патент США №5,416,011, McCabe et. al., Bio/Technology 6:923 (1988), Christou et al., Plant Physiol. 87:671-674 (1988)); Brassica (патент США №5,463,174); арахиса (Cheng et al., Plant Cell Rep. 15:653-657 (1996), McKently et al., Plant Cell Rep. 14:699-703 (1995)); папайи; и гороха (Grant et al., Plant Cell Rep. 15:254-258, (1995)).
Также описана трансформация однодольных с помощью метода электропорации, бомбардирования частицами и с помощью агробактерий. Трансформацию и воспроизведение растений удалось осуществить на аспарагусе (Bytebier et al., Proc. Natl. Acad. Sci. (USA) 84:5354, (1987)); ячмене (Wan and Lemaux, Plant Physiol 104:37 (1994)); Zea mays (Rhodes et al., Science 240:204 (1988), Gordon-Kamm et al., Plant Cell 2:603-618 (1990), Fromm et al., Bio/Technology 8:833 (1990), Koziel et al., Bio/Technology 11:194, (1993), Armstrong et al., Crop Science 35:550-557 (1995)); овсе (Somers et al., Bio/Technology 10:15 89 (1992)); еже сборной (Horn et al. Plant Cell Rep. 7:469 (1988)); рисе (Toriyama et al, TheorAppl. Genet. 205:34, (1986); Part et al., Plant Mol. Biol. 32:1135-1148, (1996); Abedinia et al., Aust. J. Plant Physiol. 24:133-141 (1997); Zhang and Wu, Theor. Appl. Genet. 76:835 (1988); Zhang et al. Plant Cell Rep. 7:379. (1988); Battraw and Hall, Plant Sci. 86:191-202 (1992); Christou et al., Bio/Technology 9:957 (1991)); ржи (De la Pena et al., Nature 325:274 (1987)); сахарном тростнике (Bower and Birch, Plant J. 2:409 (1992)); tall fescue (Wang et al., Bio/Technology 10:691 (1992)) и пшенице (Vasil et al., Bio/Technology 10:667 (1992); патент США №5,631,152).
Были разработаны методы анализа экспрессии гена на основе временной экспрессии клонированных конструкций нуклеиновых кислот, применяющие введение молекул нуклеиновых кислот в клетки растений с помощью обработки полиэтиленликолем, электропорации или бомбардирования частицами (Marcotte et al., Nature 335:454-457 (1988); Marcotte et al., Plant Cell 1:523-532 (1989); McCarty et al., Cell 66:895-905 (1991); Hattori et al., Genes Dev. 6:609-618 (1992); Goff et al., EMBO J. 9:2517-2522 (1990)).
Системы с временной экспрессией можно применять для функционального разрезания генных конструкций (См., в общих чертах, Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor Press (1995)). Следует понимать, что любую из молекул нуклеиновых кислот согласно настоящему изобретению можно постоянно или временно вводить в клетку растения в сочетании с другими генетическими элементами, например, с векторами, промоторами, энхансерами и т.д..
В дополнении к описанному выше способу, практикующим специалистам хорошо известны стандартные источники материала, описывающие определенные условия и способы для конструирования, манипулирования и выделения макромолекул (например, молекул ДНК, плазмид и т.д.), создания рекомбинантных организмов и скрининга и выделения клонов (См., Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press (1989); Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor Press (1995); Birren et al., Genome Analysis: Detecting Genes, 1, Cold Spring Harbor, N.Y. (1998); Birren et al., Genome Analysis: Analyzing DNA, 2, Cold Spring Harbor, N.Y. (1998); Plant Molecular Biology: A Laboratory Manual, eds. Clark. Springer, N.Y. (1997)).
Как следует из описанного выше, в объем настоящего изобретения также входят способы получения фермента Δ8-десатуразы. Указанные способы включают этапы: 1) выделения нуклеотидной последовательности, содержащей, или по меньшей мере на 55% комплементарной нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30; 2) конструирования вектора экспрессии, содержащего указанную нуклеотидную последовательность; и 3) введения указанного вектора в клетку-хозяина на период времени и в условиях, достаточных для образования фермента Δ8-десатуразы.
Также в объем настоящего изобретения входят способы получения полиненасыщенных жирных кислот. В одном аспекте, способ включает: 1) выделение нуклеотидной последовательности, содержащей, или по меньшей мере на 55% комплементарной нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30; 2) конструирование вектора экспрессии, содержащего нуклеотидную последовательность из этапа 1), функционально связанную с регуляторной последовательностью; 3) введение указанного вектора в клетку-хозяина на период времени и в условиях, достаточных для образования фермента Δ8-десатуразы; и 4) приведение экспрессированного фермента Δ8-десатуразы во взаимодействие с субстратом, выбранным из группы, состоящей из: ω6-эйкозадиеновой кислоты, ω3-эйкозатриеновой кислоты или как ω6-эйкозадиеновой кислоты, так и ω3-эйкозатриеновой кислоты, для превращения субстрата в первую получаемую полиненасыщенную жирную кислоту. Примерами полиненасыщенных жирных кислот, которые можно получать на первом этапе с помощью указанного способа, являются DGLA, ω3-ETA или как DGLA, так и ω3-ETA. Далее, способ включает этап(ы) приведения во взаимодействие первой полученной полиненасыщенной жирной кислоты по меньшей мере с одной десатуразой или по меньшей мере с одной элонгазой и, возможно, повтор этапа (а именно, приведения во взаимодействие полиненасыщенной жирной кислоты, полученной на втором или следующем этапе, с десатуразой или элонгазой (которые могут быть теми же десатуразой и элонгазой, которые применяли на предыдущем этапе, или любыми другими)) для превращения первой полученной полиненасыщенной жирной кислоты (например, DGLA и/или ω3-ETA) во вторую или следующую полиненасыщенную жирную кислоту (например, третью, четвертую, пятую, шестую, и т.д.). Указанный этап можно повторять любое необходимое количество раз, для получения целевой полиненасыщенной жирной кислоты. Например, если первой получаемой полиненасыщенной жирной кислотойявляется DGLA, способ далее может включать приведение DGLA во взаимодействие с Δ5-десатуразой с образованием ARA (второй получаемой полиненасыщенной жирной кислоты, образующейся в качестве второго продукта). Возможно последующее приведение ARA во взаимодействие с Δ17-десатуразой с образованием ЕРА (третьей получаемой полиненасыщенной жирной кислоты). Далее, возможно последующее приведение ETA во взаимодействие с Δ5-элонгазой с образованием DPA (четвертой получаемой полиненасыщенной жирной кислоты). Далее, возможно последующее приведение DPA во взаимодействие с Δ4-десатуразой с образованием DHA (пятой получаемой полиненасыщенной жирной кислоты).
В другом аспекте, способ включает: 1) выделение нуклеотидной последовательности, содержащей, или по меньшей мере на 55% комплементарной нуклеотидной последовательности, выбранной из группы, состоящей из: SEQ ID NO:28 и SEQ ID NO:30; 2) конструирование вектора экспрессии, содержащего нуклеотидную последовательность согласно этапу 1), функционально связанную с регуляторной последовательностью; 3) введение вектора согласно этапу 2) и по меньшей мере одной дополнительной рекомбинантной сконструированной ДНК, содержащей выделенную нуклеотидную последовательность, функционально связанную по меньшей мере с одной регуляторной последовательностью, кодирующей Δ9-элонгазу (См., например, патентную публикацию США №2008/0214667, в которой описывают выделенную нуклеотидную последовательность, кодирующую Δ9-элонгазу) в клетку-хозяина; и 4) приведение экспрессированного фермента Δ8-десатуразы и Δ9-элонгазы во взаимодействие с субстратами, выбранными из группы, состоящей из: LA, ALA или LA и ALA, для превращения субстрата в первую полиненасыщенную жирную кислоту. Примерами полиненасыщенных жирных кислот, которые можно получать на первом этапе с помощью указанного способа, являются DGLA, ω3-ETA или как DGLA, так и ω3-ETA. Указанный способ может дополнительно включать этап(ы) приведения во взаимодействие первой полученной полиненасыщенной жирной кислоты по меньшей мере с одной десатуразой или по меньшей мере с одной элонгазой и, возможно, повтор этапа (а именно, приведения во взаимодействие второй или последующей полученной полиненасыщенной жирной кислоты с десатуразой или элонгазой (которые могут быть теми же десатуразой и элонгазой, которые применяли на предыдущем этапе, или любыми другими)) для превращения первой полученной полиненасыщенной жирной кислоты, (например, DGLA и/или ω3-ETA) во вторую или следующую (например, третью, четвертую, пятую, шестую, и т.д.), получаемую полиненасыщенную жирную кислоту. Указанный этап можно повторять любое необходимое количество раз, для получения в целевой полиненасыщенной жирной кислоты. Например, если первой получаемой полиненасыщенной жирной кислотой является DGLA, способ далее может включать приведение DGLA во взаимодействие с Δ5-десатуразой с образованием ARA (второй получаемой полиненасыщенной жирной кислоты). Возможно последующее приведение ARA во взаимодействие с Δ17-десатуразой с образованием EPA (третьей получаемой полиненасыщенной жирной кислоты). Далее, возможно последующее приведение EPA во взаимодействие с Δ5-элонгазой с образованием DPA (четвертой получаемой полиненасыщенной жирной кислоты). Далее, возможно последующее приведение DPA во взаимодействие с Δ4-десатуразой с образованием DHA (пятой получаемой полиненасыщенной жирной кислоты).
Следовательно, согласно описанному выше, Δ8-десатуразу согласно настоящему изобретению можно применять для получения полиненасыщенных жирных кислот, которые, в свою очередь, могут быть полезны для определенных целей, или для получения других полиненасыщенных жирных кислот.
D. Применение гена Δ8-десатуразы
Как было упомянуто выше, выделенный ген Δ8-десатуразы и кодируемый им фермент Δ8-десатураза, имеют множество областей применения. Например, ген и соответствующий ему фермент можно прямо или опосредованно применять для получения полиненасыщенных жирных кислот, например, Δ8-десатуразу можно применять для получения DGLA, ARA, EPA, ω3-ETrA, ω3-ETA, DPA и/или DHA. Указанные полиненасыщенные жирные кислоты (то есть, кислоты, прямо или опосредованно полученные благодаря активности фермента Δ8-десатуразы) можно включать в состав, например, пищевых композиций, фармацевтических композиций, косметических средств и кормов для животных, все перечисленные применения включены в объем настоящего изобретения. Указанные области применения подробно описаны ниже.
E. Питательные композиции
Настоящее изобретение обеспечивает питательные композиции. Указанные композиции, в контексте настоящего изобретения, включают любую пищу или препарат, предназначенные для применения человеком, включая энтеральное или паренетеральное применение, которые, при поступлении в организм, (a) служат для питания или построения тканей или запаса энергии и/или (b) поддерживают, восстанавливают или сохраняют соответствующий пищевой статус или метаболическую функцию.
Питательная композиция согласно настоящему изобретению включает по меньшей мере одно масло или кислоту, полученные при прямом или опосредованном применении гена Δ8-десатуразы, описанного в настоящей заявке, в соответствии с настоящим изобретением, и находящееся либо в твердой, либо в жидкой форме. Также, композиция может включать пищевые макронутриенты, витамины и минералы в количествах, соответствующих определенной области применения. Количество указанных составляющих будет изменяться в зависимости от того, предназначена ли композиция для применения здоровыми младенцами в нормальном состоянии, детьми или взрослыми, имеющими особые потребности, например, связанные с определенным состоянием метаболизма (например, с нарушениями метаболизма).
Примеры макронутриентов, подходящих для включения в состав композиции, включают, но не ограничиваются указанными: пищевые жиры, углеводы и белки. Примеры указанных пищевых жиров включают, но не ограничиваются указанными: кокосовое масло, соевое масло и моно- и диглицериды. Примеры указанных углеводов включают, но не ограничиваются указанными: глюкозу, пищевую лактозу и гидролизованный крахмал. Также, примеры белков, которые можно применять в составе пищевой композиции согласно настоящему изобретению, включают, но не ограничиваются указанными: белки сои, сыворотку, подвергнутую элекродиализу, обезжиренное молоко, подвергнутое электродиализу, сыворотку молока или гидролизаты указанных белков.
В отношении витаминов и минералов, в состав питательных композиций согласно настоящему изобретению можно добавлять следующие: кальций, фосфор, калий, натрий, хлор, магний, марганец, железо, медь, цинк, селен, йод, а также витамины A, E, D, C и витамины комплекса B. Также можно добавлять другие подобные витамины и минералы.
Компоненты, применяемые в составе пищевых композиций согласно настоящему изобретению, должны находится в очищенной или частично очищенной форме. Под частично очищенным понимают вещество, полученное путем очистки природного материала или путем синтеза.
Примеры пищевых композиций согласно настоящему изобретению включают, но не ограничиваются указанными: смеси для младенцев, диетические добавки, пищевые заменители и композиции для восстановления водного баланса. Пищевые композиции, представляющие особый интерес, включают, но не ограничиваются указанными: композиции, применяемые для энтерального и парентерального кормления младенцев, специализированные смеси для младенцев, пищевые добавки для пожилых людей и пищевые добавки для лиц, страдающих заболеваниями желудочно-кишечного тракта и/или нарушением всасывания.
Пищевую композицию согласно настоящему изобретению также можно добавлять к пище в случаях, когда не требуется дополнение диеты. Например, композицию можно добавлять к любой пище, включая, но не ограничиваясь указанными: маргарины, модифицированные масла, сыры, молоко, йогурт, шоколад, сладости, закуски, масла для салатов, кулинарные масла, кулинарные жиры, мясо, рыбу и напитки.
В предпочтительном варианте реализации настоящего изобретения, пищевая композиция представляет собой пищевой продукт, предназначенный для энтерального применения, более предпочтительно, пищевой продукт, предназначенный для энтерального применения взрослыми или детьми. Указанную композицию можно вводить взрослым или детям, испытывающим стресс или имеющим особые потребности в связи с хроническими или острыми болезненными состояниями. Композиция может включать, в дополнение к полиненасыщенным жирным кислотам, произведенным в соответствии с настоящим изобретением, макронутриенты, витамины и минералы, согласно описанному выше. Макронутриенты могут содержаться в количествах, эквивалентных количествам, присутствующим в молоке человека или в пересчете на содержащуюся энергию, то есть, в пересчете на содержащиеся калории.
Способы создания жидких или твердых пищевых смесей, предназначенных для энтерального и парентерального введения, хорошо известны в данной области техники.
Например, состав, предназначенный для энтерального введения, можно стерилизовать и впоследствии применять как готовый для потребления (RTF) или хранить в форме концентрированной жидкости или порошка. Порошок можно изготавливать путем сушки распылением состава, приготовленного согласно описанному выше, и восстановления его путем регидратирования концентрата. Пищевые составы, предназначенные для применениями взрослыми и детьми, хорошо известны в данной области техники и коммерчески доступны (например, Симилак®, Эншур®, Джевити® и Алиментум®, Ross Products Division, Abbott Laboratories, Columbus, Ohio). К любому из указанных составов можно добавлять масло или кислоту, полученные в соответствии с настоящим изобретением.
Энергетическая плотность пищевых композиций согласно настоящему изобретению, находящихся в жидкой форме, может варьировать приблизительно от 0.6 ккал приблизительно до 3 ккал на мл. Пищевые добавки, составленные в твердой или порошкообразной форме, могут содержать приблизительно от 1.2 до более 9 ккал на грамм, предпочтительно, приблизительно от 3 приблизительно до 7 ккал/г. В общем случае, осмолярность жидкого продукта должна составлять менее 700 миллиосмоль, более предпочтительно, менее 660 миллиосмоль.
Пищевой состав может включать макронутриенты, витамины и минералы, согласно описанному выше, в дополнение к PUFA, полученным в соответствии с настоящим изобретением. Присутствие указанных дополнительных составляющих помогает удовлетворять минимальную ежедневную потребность в указанных элементах. В дополнение к введению PUFA, также может быть желательным добавление в состав композиции цинка, меди, фолиевой кислоты и антиоксидантов. Считают, что указанные вещества поддерживают иммунную систему, подвергнутую стрессу, и, следовательно, их применение приносит дальнейшую пользу лицу, применяющему композицию. Указанные элементы также можно добавлять в состав фармацевтической композиции.
В более предпочтительном варианте реализации, пищевая композиция содержит, в дополнение к антиоксидантам и по меньшей мере одной PUFA, источник углеводов, при этом по меньшей мере 5 процентов углевода по массе представляет собой неусваиваемый олигосахарид. В более предпочтительном варианте реализации, пищевая композиция дополнительно содержит белок, таурин и карнитин.
Как было упомянуто выше, PUFA или их производные, полученные в соответствии с настоящим изобретением, можно включать в состав диетических заменителей или добавок, в частности, детских смесей, для пациентов, находящихся на внутривенном кормлении, или для предотвращения или лечения истощения или других нарушений или болезненных состояний. В качестве дополнительной информации, следует заметить, что содержание жирных кислот в грудном молоке человека составляет приблизительно от 0.15% приблизительно до 0.36% DHA, приблизительно от 0.03% приблизительно до 0.13% EPA, приблизительно от 0.30% приблизительно до 0.88% ARA, приблизительно от 0.22% приблизительно до 0.67% DGLA и приблизительно от 0.27% приблизительно до 1.04% GLA. Следовательно, такие жирные кислоты, как ARA, EPA и/или DHA, полученные в соответствии с настоящим изобретением, можно применять для изменения, например, композиции детских смесей для улучшенного воссоздания составляющей PUFA в грудном молоке человека или для изменения состава PUFA, обычно присутствующего в молоке млекопитающих, не относящихся к роду человек. В частности, композиция, предназначенная для применения в фармакологии и в качестве пищевой добавки, предпочтительно будет содержать одну или более из ARA, EPA, DGLA, и DHA. Более предпочтительно, масло будет включать приблизительно от 0.3 до 30% ARA и приблизительно от 0.2 до 30% DGLA.
В объем настоящего изобретения входят пищевые композиции, предназначенные для парентерального введения, содержащие приблизительно от 2 приблизительно до 30 процентов по массе жирных кислот в пересчете на триглицериды. Возможно также включение других жирорастворимых витаминов, например, витаминов A, D, E и L-карнитина. При необходимости, можно добавлять такие консерванты, как альфа-токоферол, в количестве, составляющем приблизительно 0.1% по массе.
В дополнение следует заметить, что отношение содержания ARA и DGLA можно варьировать в зависимости от определенного конечного применения. При оформлении композиции в виде заменителя грудного молока, в композиции, включающей одну или более из ARA, DGLA и GLA, отношение будет составлять приблизительно от 1:19:30 приблизительно до 6:1:0.2, соответственно. Например, отношение содержания ARA:DGLA:GLA в грудном молоке животных может варьировать от 1:19:30 до 6:1:0.2, что включает промежуточные значения отношения, предпочтительно составляющие приблизительно 1:1:1, 1:2:1, 1:1:4. При совместном получении в клетке-хозяине, для точного контроля отношений содержания PUFA можно применять регулирование отношения и процента превращения субстрата-предшественника, например, EDA и DGLA, в ARA. Например, для получения отношения содержания ARA и DGLA, составляющего приблизительно 1:19, можно применять отношение превращения DGLA в ARA, составляющее приблизительно от 5% до 10%, тогда как для получения отношения содержания ARA и DGLA, составляющего приблизительно 6:1, можно применять отношение превращения приблизительно от 75% до 80%. Следовательно, как в системе культуры клеток, так и в организме животного-хозяина, для модулирования уровней и отношений PUFA можно применять регуляцию экспрессии десатуразы, а также экспрессии элонгаз (не ограничивающим примером является Δ9 элонгаза) и других десатураз по времени, активности и специфичности. PUFA/кислоты, производимые в соответствии с настоящим изобретением (например, ARA и EPA), впоследствии можно комбинировать с другими PUFA/кислотами (например, с DGLA) в желаемых концентрациях и отношениях.
Также, PUFA, полученные в соответствии с настоящим изобретением, или клетки-хозяева, содержащие их, можно применять в качестве пищевых добавок для животных для изменения составляющей жирных кислот молока или ткани животных на более благоприятное для потребления животными или человеком.
Примеры некоторых пищевых добавок, смесей для детского питания, пищевых заменителей и других пищевых растворов, в которых можно применять полиненасыщенные жирные кислоты, полученные в соответствии с настоящим изобретением, описаны ниже.
I. СМЕСИ ДЛЯ ДЕТСКОГО ПИТАНИЯ
A. Соевая смесь Изомил®, содержащая железо:
Применение: В качестве напитка для новорожденных, детей и взрослых, страдающих от аллергии или чувствительности к коровьему молоку. Питание для пациентов с нарушениями, при которых следует избегать употребление лактозы: включая лактозная недостаточность, непереносимость лактозы и галактоземию.
Описание:
- Изолят соевого белка, применяемый для предотвращения проявления симптомов аллергии или чувствительности к белку коровьего молока.
- Отсутствие в составе лактозы, для предотвращения диареи, связанной с употреблением лактозы.
- Низкая осмолярность (200 мосмоль/кг воды).
- Парные углеводы (кукурузный сироп и сахароза), предназначенные для максимального повышения всасывания и снижения до минимума риска развития нарушения всасывания.
Состав: 43.2% сухая кукурузная патока, 14.6% изолят белка сои, 11.5% высокомасличное масло сафлора, 10.3% сахар (сахароза), 8.4% соевое масло, 8.1% кокосовое масло: менее 2%: фосфат кальция, цитрат калия, хлорид калия, хлорид магния, хлорид натрия, аскорбиновая кислота, холихлорид, L-метионин, таурин, аскорбилпальмитат, сульфат железа, м-инозитол, смешанные токоферолы, сульфат цинка, d-альфа-токоферил ацетат, L-карнитин, никотинамид, пантотенат кальция, сульфат меди, тиаминхлорид гидрохлорид, витамин A-пальмитат, рибофлавин, пиридоксина гидрохлорид, фолиевая кислота, йодид калия, гидроксид калия, филлохинон, биотин, селенат натрия, бета-каротинин, витамин D3 и цианокобаламин.
В. Соевая смесь Изомил® DF, применяемая при диарее:
Применение: Для диетотерапии при диарее новорожденных и детей 1-3 лет.
Описание:
- Первая детская смесь, содержащая пищевую клетчатку, полученную на основе соевой клетчатки, предназначенная для терапии диареи.
- Клинически доказана эффективность для снижения продолжительности жидкого, водянистого стула как при легкой, так и при тяжелой диарее новорожденных.
- Отсутствие в составе лактозы, для предотвращения диареи, связанной с употреблением лактозы.
- Низкая осмолярность (240 мосмоль/кг воды) для снижения риска осмотической диареи.
Состав: 85.7% вода, 4.8% кукурузная патока, 2.6% сахар (сахароза), 2.1% соевое масло, 2.0% изолят соевого белка, 1.4% кокосовое масло, 0.77% соевая клетчатка, цитрат кальция, цитрат калия, фосфат кальция, фосфат калия, хлорид калия, моно- и диглицериды, соевый лецитин, хлорид магния, каррагенан, аскорбиновая кислота, L-метионин, хлорид натрия, холинхлорид, таурин, сульфат железа, м-инозитол, d-альфа-токоферил ацетат, сульфат цинка, L-карнитин, никотинамид, пантотенат кальция, сульфат меди, витамин A-пальмитат, тиаминхлорид гидрохлорид, рибофлавин, пиридоксина гидрохлорид, фолиевая кислота, сульфат марганца, йодид калия, филлохинон, биотин, селенит натрия, витамин D3 и цианокобаламин.
C. Соевая смесь Изомил® Эдванс®, содержащая железо:
Применение: В качестве напитка для новорожденных, детей и взрослых, страдающих от аллергии или чувствительности к коровьему молоку. Питание для пациентов с нарушениями, при которых следует избегать употребления лактозы: включая лактозную недостаточность, непереносимость лактозы и галактоземию.
Описание:
- Содержит DHA и ARA, два питательных вещества, входящие в состав грудного молока, имеющие значение для умственного развития и развития зрительной системы.
- Изолят соевого белка, применяемый для предотвращения проявления симптомов аллергии или чувствительности к белку коровьего молока.
- Отсутствие в составе лактозы для предотвращения диареи, связанной с употреблением лактозы.
- Низкая осмолярность (200 мосмоль/кг воды).
- Парные углеводы (кукурузный сироп и сахароза), предназначенные для максимального повышения всасывания и снижения до минимума риска развития нарушения всасывания.
Состав: 43.2% сухая кукурузная патока, 14.6% изолят соевого белка, 11.5% высокомасличное масло сафлора, 10.3% сахар (сахароза), 8.4% соевое масло, 7.7% кокосовое масло, масло С.cohnii, масло M.alpina. Фосфат кальция, цитрат калия, хлорид калия, хлорид магния, хлорид натрия, аскорбиновая кислота, холинхлорид, L-метионин, таурин, аскорбилпальмитат, сульфат железа, м-инозитол, смешанные токоферолы, сульфат цинка, d-альфа-токоферилацетат, L-карнитин, никотинамид, пантотенат кальция, сульфат меди, тиаминхлорид гидрохлорид, витамин A-пальмитат, рибофлавин, пиридоксина гидрохлорид, фолиевая кислота, йодид калия, гидроксид калия, филлохинон, биотин, селенат натрия, бета-каротинин, витамин D3 и цианокобаламин.
D. Готовая к применению соевая смесь Изомил® Эдванс® 20, содержащая железо, 20 кал/жидкую унцию:
Применение: в случаях, когда желательным является соевое питание.
Состав: 85.9% вода, 6.7% кукурузная патока, 1.9% изолят соевого белка, 1.4% высокомасличное масло сафлора, 1.3% сахар (сахароза), 1.1% соевое масло, 1.0% кокосовое масло, масло С.cohnii, масло M.alpina, цитрат кальция, фосфат кальция, цитрат калия, хлорид калия, моно- и диглицериды, соевый лецитин, каррагенан, аскорбиновая кислота, L-метионин, хлорид магния, фосфат калия, хлорид натрия, холинхлорид, таурин, сульфат железа, м-инозитол, d-альфа-токоферилацетат, сульфат цинка, L-карнитин, никотинамид, пантотенат кальция, сульфат меди, витамин A-пальмитат, тиаминхлорид гидрохлорид, рибофлавин, пиридоксина гидрохлорид, фолиевая кислота, сульфат марганца, йодид калия, филлохинон, биотин, селенит натрия, витамин D3 и цианокобаламин.
E. Смесь для младенцев Симилак®:
Применение: В случаях, когда требуется смесь для младенцев: если принято решение о прекращении грудного вскармливания до наступления возраста 1 год, если существует потребность в добавке, помимо грудного вскармливания, или если грудное вскармливание не подходит в качестве регулярного кормления. Порошок, концентрированная жидкость и формы, готовые к употреблению.
Состав: Вода, обезжиренное молоко, лактоза, высокомасличное масло сафлора, соевое масло, кокосовое масло, концентрат белка сыворотки, цитрат калия, карбонат кальция, аскорбиновая кислота, соевый лецитин, моноглицериды, каррагенан, хлорид калия, хлорид магния, сульфат железа, холинхлорид, холинбитартрат, таурин, м-инозитол, сульфат цинка, никотинамид, d-альфа-токоферилацетат, пантотенат кальция, l-карнитин, витамин A-пальмитат, рибофлавин, сульфат меди, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, фолиевая кислота, сульфат марганца, филлохинон, биотин, бета-каротин, селенит натрия, витамин D3, цианокобаламин, фосфат кальция, фосфат калия, хлорид натрия, гидроксид калия и нуклеотиды (аденозин-5′-монофосфат, цитидин-5′-монофосфат, динатрийгуанозин-5′-монофосфат, динатрийуридин-5′-монофосфат).
F. Смесь для младенцев Симилак® Эдванс®, содержащая железо:
Применение: Для применения в качестве добавки или альтернативы грудному вскармливанию. Порошок, концентрированная жидкость и формы, готовые к употреблению.
Состав: Вода, обезжиренное молоко, лактоза, высокомасличное масло сафлора, соевое масло, кокосовое масло, концентрат белка сыворотки, масло C.cohnii oil, масло M.alpina, цитрат калия, карбонат кальция, аскорбиновая кислота, соевый лецитин, моноглицериды, каррагенан, хлорид калия, хлорид магния, сульфат железа, холинхлорид, холинбитартрат, таурин, м-инозитол, сульфат цинка, никотинамид, d-альфа-токоферилацетат, пантотенат кальция, l-карнитин, витамин A-пальмитат, рибофлавин, сульфат меди, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, фолиевая кислота, сульфат марганца, филлохинон, биотин, бета-каротин, селенит натрия, витамин D3, цианокобаламин, фосфат кальция, фосфат калия, хлорид натрия, гидроксид калия и нуклеотиды (аденозин-5′-монофосфат, цитидин-5′-монофосфат, динатрийгуанозин-5′-монофосфат, динатрийуридин-5′-монофосфат).
G. Смесь для младенцев Симилак® Неошур® Эдванс®, содержащая железо:
Применение: Специализированная смесь для применения в таких случаях, как недоношенность.
Описание:
- Жировая смесь, характеризующаяся хорошим всасыванием, содержащая 25% дополнительных триглицеридов со средней длиной цепи (МСТ).
- Повышенное содержание белка, витаминов и минералов на 100 ккал по сравнению со стандартной смесью.
- Повышенное содержание кальция и фосфора по сравнению со стандартной смесью.
Состав: обезжиренное молоко, сухая кукурузная патока, лактоза, соевое масло, высокомасличное масло сафлора, концентрат белка сыворотки, триглицериды со средней длиной цепи, кокосовое масло, масло C.cohnii, масло M.alpina, цитрат калия, фосфат кальция, м-инозитол, аскорбиновая кислота, хлорид магния, карбонат кальция, таурин, сульфат железа, холинбитартрат, холинхлорид, аскорбилпальмитат, L-карнитин, хлорид калия, хлорид натрия, сульфат цинка, смешанные токоферолы, d-альфа-токоферилацетат, цитрат натрия, никотинамид, фосфат калия, пантотенат кальция, сульфат меди, витамин A-пальмитат, тиаминхлорида гидрохлорид, рибофлавин, пиридоксина гидрохлорид, бета-каротин, фолиевая кислота, сульфат марганца, филлохинон, биотин, селенит натрия, витамин D3, цианокобаламин и нуклеотиды (аденозин-5′-монофосфат, цитидин-5′-монофосфат, динатрийгуанозин-5′-монофосфат, динатрийуридин-5′-монофосфат).
H. Готовый к применению обогатитель молока человека Симилак с низким содержанием железа, 24 кал/жидкую унцию:
Применение: Разработано для смешивания с молоком человека или в качестве альтернативы молоку человека для младенцев, родившихся с низкой массой тела.
Состав: Вода, обезжиренное молоко, сухая кукурузная патока, лактоза, триглицериды со средней длиной цепи, концентрат белка сыворотки, соевое масло, кокосовое масло, масло C.cohnii, масло M.alpina, фосфат кальция, цитрат калия, аскорбиновая кислота, карбонат кальция, хлорид магния, соевый лецитин, моно- и диглицериды, м-инозитол, цитрат натрия, каррагенан, холинбитартрат, таурин, холинхлорид, никотинамид, d-альфа-токоферилацетат, L-карнитин, сульфат цинка, хлорид калия, двухосновный фосфат калия, пантотенат кальция, сульфат железа, сульфат меди, рибофлавин, витамин A-пальмитат, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, биотин, фолиевая кислота, бета-каротин, сульфат марганца, филлохинон, витамин D3, селенит натрия, цианокобаламин и нуклеотиды (аденозин-5′-монофосфат, цитидин-5′-монофосфат, динатрийгуанозин-5′-монофосфат, динатрийуридин-5′-монофосфат).
Различные PUFA согласно настоящему изобретению можно замещать и/или добавлять в смеси для новорожденных, описанные выше, и в другие смеси для новорожденных, известные специалистам в данной области техники.
II. ПИЩЕВЫЕ СМЕСИ
A. Эншур®
Применение: Питательная смесь ЭНШУР со вкусом сливок является источником полноценного, сбалансированного питания при применении в качестве добавки между приемами пищи или во время приема пищи, или в качестве единственного источника пищи при временном применении. Применение ЭНШУРА может быть полезно людям, подверженным риску, связанному с питанием, испытывающим патологическую потерю веса, проходящим восстановление после болезни или хирургического вмешательства, или в ходе модифицированных или бесшлаковых диет. Для орального применения. Для временного применения в качестве единственного источника питания. Розничный продукт для дополнительного перорального питания.
Состав: Вода, сахар (сахароза), кукурузный мальтодекстрин, изолят белка молока, соевое масло, кукурузное масло, каноловое масло, концентрат белка сои, цитрат калия, натуральный и искусственный ароматизатор, фосфат магния, цитрат натрия, соевый лецитин, фосфат кальция, хлорид магния, соль (хлорид натрия), холинхлорид, каррагенан, аскорбиновая кислота, dl-альфа-токоферилацетат, сульфат железа, сульфат цинка, никотинамид, пантотенат кальция, сульфат марганца, сульфат меди, витамин A-пальмитат, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, рибофлавин, фолиевая кислота, хлорид хрома, биотин, молибдат натрия, селенат натрия, филлохинон, йодид калия, витамин D3 и цианокобаламин.
B. Эншур® с высоким содержанием белка:
Применение: ЭНШУР С ВЫСОКИМ СОДЕРЖАНИЕМ БЕЛКА полезен для людей, испытывающих потребность в дополнительном белке и питании в диете. ЭНШУР С ВЫСОКИМ СОДЕРЖАНИЕМ БЕЛКА подходит для людей, проходящих восстановление после общего хирургического вмешательства или перелома бедренной или другой кости, и является хорошим источником питания для лиц, подверженных риску появления пролежневых язв. Для дополнительного перорального питания.
Состав: Вода, сахар (сахароза), кукурузный мальтодекстрин, казеинаты кальция и натрия, соевое масло, изолят белка сои, кукурузное масло, цитрат калия, каноловое масло, фосфат кальция, цитрат натрия, хлорид магния, фосфат магния, искусственный ароматизатор, соль (хлорид натрия), соевый лецитин, холинхлорид, аскорбиновая килсота, каррагенан, сульфат цинка, dl-альфа-токоферилацетат, сульфат железа, геллановая камедь, никотинамид, пантотенат кальция, сульфат марганца, сульфат меди, витамин A-пальмитат, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, рибофлавин, фолиевая кислота, хлорид хрома, биотин, молибдат натрия, йодид калия, селенат натрия, филлохинон, витамин D3 и цианокобаламин.
C. Эншур Плюс®
Применение: ЭНШУР ПЛЮС является источником полноценного, сбалансированного питания, обеспечивающего организм концентрированными калориями и белком, что помогает пациентам набрать или поддерживать здоровую массу тела. Его можно применять одновременно с приемом пищи или между приемами пищи, или в качестве заменителя пищи. Для временного питания в качестве единственного источника пищи. Для пациентов, имеющих ограничения в потреблении жидкости или нуждающихся в ограничении объемов поступающей пищи.
Описание:
- 650 мг омега-3 жирной кислоты ALA (40% 1.6 г RDI) для поддержания здоровья сердца.
Превосходный источник 24 незаменимых витаминов и минералов.
- Источник антиоксидантов, селена и витаминов C и E для укрепления иммунной системы.
- Низкий уровень холестерола.
- Кошерность.
- Без содержания глутена.
- Без содержания лактозы.
Состав: Ваниль: вода, кукурузная патока, мальтодекстрин (кукурузный), кукурузное масло, казеинаты натрия и кальция, сахар (сахароза), изолят белка сои, хлорид магния, цитрат калия, трехосновный фосфат кальция, соевый лецитин, естественный и искусственный ароматизатор, цитрат натрия, хлорид калия, холинхлорид, аскорбиновая кислота, каррагенан, сульфат цинка, сульфат железа, альфа-токоферилацетат, никотинамид, пантотенат кальция, сульфат марганца, сульфат меди, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, рибофлавин, витамин A-пальмитат, фолиевая кислота, биотин, хлорид хрома, молибдат натрия, йодид калия, селенит натрия, филлохинон, цианокобаламин и витамин D3.
D. Эншур® в порошке:
Применение: ЭНШУР® В ПОРОШКЕ (ресуспендируемый в воде) является полноценным, сбалансированным питанием для применения в качестве добавки одновременно с приемом пищи или между приемами пищи. Он может быть полезен людям, придерживающимся модифицированной диеты, подверженным риску, связанному с питанием, испытывающим патологическую потерю веса, проходящим восстановление после болезни или хирургического вмешательства или находящимся на бесшлаковой диете.
Описание:
- Удобство в применении, легкость смешивания
- Слабое образование осадка
- Без содержания лактозы и глутена
Состав: кукурузная патока, кукурузный мальтодекстрин, сахар (сахароза), кукурузное масло, казеинаты натрия и кальция, изолят белка сои, искусственный ароматизатор, цитрат калия, хлорид магния, цитрат натрия, фосфат кальция, хлорид калия, соевый лецитин, аскорбиновая кислота, холинхлорид, сульфат цинка, dl-альфа-токоферилацетат, никотинамид, сульфат железа, пантотенат кальция, сульфат марганца, сульфат меди, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, рибофлавин, витамин A-пальмитат, фолиевая кислота, биотин, хлорид хрома, молибдат натрия, йодид калия, селенат натрия, филлохинон, цианокобаламин и витамин D3.
E. Эншур® в форме пудинга
Применение: ЭНШУР В ФОРМЕ ПУДИНГА представляет собой питательную альтернативу другим закускам и десертам. Он обеспечивает полноценное сбалансированное питание в приятной, легкой для употребления форме. Он подходит для лиц с пониженной массой тела или страдающих истощением или находящихся на диете с ограничением в потреблении жидкости или ограничением объемов поступающей пищи. Для людей, находящихся на диете с модифицированием консистенции (например, употребляющих мягкую пищу, пюре или исключительно жидкую пищу). Для людей с затруднениями при проглатывании. Для дополнительного перорального питания.
Описание:
- Хороший источник 24 незаменимых витаминов и минералов.
- Удобство - не требует замораживания.
- Без содержания глутена.
- Содержит 1 г фруктоолигосахаридов (FOS) в одной порции (FOS представляют собой пребиотики, стимулирующие рост полезных бактерий в кишечнике).
Состав:
Ваниль: вода, сахар (сахароза), модифицированный кукурузный крахмал, частично гидрогенизированное соевое масло, концентрат белка молока, обезжиренное молоко, фруктоолигосахариды, сульфат магния, фосфат калия, фосфат натрия, стеароиллактилат натрия, искусственный ароматизатор, аскорбат натрия, сульфат цинка, dl-альфа-токоферилацетат, сульфат железа, никотинамид, сульфат марганца, пантотенат кальция, FD&C Желтый #5 и #6, сульфат меди, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, витамин A-пальмитат, рибофлавин, фолиевая кислота, хлорид хрома, биотин, молибдат натрия, йодид калия, селенат натрия, филлохинон, витамин D3 и цианокобаламин.
F. Эншур® с клетчаткой:
Применение: ЭНШУР С КЛЕТЧАТКОЙ является источником полноценного, сбалансированного питания для людей, которым может быть полезна диета с повышенным содержанием клетчатки и питательных веществ. Смесь пищевых волокон с FOS, пребиотиком, способствует поддержанию нормального функционирования пищеварительного тракта. ЭНШУР С КЛЕТЧАТКОЙ подходит для людей, не нуждающихся в бесшлаковой диете. Его можно вводить перорально или через трубку. ЭНШУР С КЛЕТЧАТКОЙ может быть полезен для людей, находящихся на модифицированной диете, подверженных нутриционному риску, страдающих патологической потерей веса или проходящих восстановление после болезни или хирургического вмешательства. Для перорального приема. Для временного употребления в качестве единственного источника пищи.
Описание:
- Включает 1 г FOS/8 жидких унций FOS клетчатки (неусвояемый углевод), способствующей осуществлению естественных защитных реакций в кишечнике.
- Превосходный источник 24 незаменимых витаминов и минералов.
- Обеспечивает 2.8 г общей пищевой клетчатки на порцию 8 жидких унций.
- Без содержания лактозы и глутена.
Состав:
Ваниль: вода; кукурузный мальтодекстрин, сахар (сахароза), казеинаты натрия и кальция, соевое масло, изолят белка сои, кукурузное масло, овсяная клетчатка, фруктоолигосахриды, каноловое масло, соевая клетчатка, фосфат кальция, хлорид магния, цитрат калия, целлюлозный гель, соевый лецитин, фосфат калия, цитрат натрия, натуральный и искусственный ароматизатор, холинхлорид, фосфат магния, аскорбиновая кислота, карбоксиметилцеллюлоза, хлорид калия, каррагенан, сульфат железа, dl-альфа-токоферилацетат, сульфат цинка, никотинамид, сульфат марганца, пантотенат кальция, сульфат меди, витамин A-пальмитат, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, рибофлавин, фолиевая кислота, хлорид хрома, биотин, молибдат натрия, йодид калия, селенат натрия, филлохинон, витамин D3 и цианокобаламин.
Различные пищевые добавки, описанные выше, а также другие, известные специалистам в данной области техники, можно заменять или дополнять PUFA, произведенными в соответствии с настоящим изобретением.
G. Диетический продукт Оксепа™
Способность продукта Оксепа™ модулировать воспалительные реакции у тяжелобольных, находящихся на искусственной вентиляции, пациентов, клинически доказана. Он подходит пациентам с сепсисом, ССВР (синдромом системной воспалительной реакции), ОПЛ (острым повреждением легких) или ОРДС (острым респираторным дистресс-синдромом). Для введения с помощью трубки. Для употребления в качестве единственного источника питания.
Распределение калорий: Распределение калорий в составе продукта оксепа приведено в таблице C.
Состав: вода, казеинаты кальция и натрия, сахар (сахароза), каноловое масло, триглицериды со средней длиной цепи, сардиновое масло, масло бурачника, хлорид магния, фосфат кальция, соевый лецитин, цитрат калия, цитрат натрия, аскорбиновая кислота, фосфат калия, натуральный и искусственный ароматизатор, холинхлорид, таурин, d-альфа-токоферилацетат, L-карнитин, соль (хлорид натрия), геллановая камедь, сульфат цинка, сульфат железа, никотинамид, пантотенат кальция, сульфат марганца, сульфат меди, тиаминхлорида гидрохлорид, пиридоксина гидрохлорид, рибофлавин, бета-каротин, витамин A-пальмитат, фолиевая кислота, хлорид хрома, биотин, молибдат натрия, йодид калия, селенат натрия, филлохинон, витамин D3 и цианокобаламин.
Различные жирные кислоты в составе пищевого продукта Оксепа™ можно заменять и/или дополнять PUFA, полученными в соответствии с настоящим изобретением.
F. Фармацевтические композиции
Также в объем настоящего изобретения входит фармацевтическая композиция, содержащая в составе одну или более кислот и/или образующихся в результате масел, полученных с применением гена Δ8-десатуразы, описанного в настоящей заявке, в соответствии со способами, описанными в настоящей заявке. В частности, указанная фармацевтическая композиция может включать одну или более кислот и/или масел, а также общепринятый, хорошо известный, не токсичный фармацевтически приемлемый носитель, вспомогательное вещетво или наполнитель, например, фосфатный буферный солевой раствор, воду, этанол, многоатомные спирты, растительные масла, увлажняющие агенты или эмульсию, например, эмульсию вода в масле. Композиция может существовать как в жидкой, так и в твердой форме. Например, композицию можно предоставлять в форме таблетки, капсулы, жидкости или порошка, предназначенных для приема внутрь, мази или крема, предназначенных для приема внутрь или местного применения. Соответствующую текучесть можно обеспечивать, например, с помощью поддержания требуемого размера частиц в случае дисперсий, а также путем применения поверхностно-активных веществ. Также может быть желательным включение в состав композиции изотонических агентов, например, Сахаров, хлорида натрия и им подобных. Помимо указанных инертных разбавителей, композиция может включать вспомогательные вещества, например, увлажняющие агенты, эмульгирующие и суспендирующие агенты, подслащивающие агенты, вкусовые агенты и ароматизирующие агенты.
Суспензии, в дополнение к действующим веществам, также могут содержать суспендирующие агенты, например, этоксилированные изостеариловые спирты, сорбитоловые и сорбитановые эфиры полиоксиэтилена, микрокристаллическую целлюлозу, метагидроксид алюминия, бентонит, агар-агар и трагакант или смеси указанных веществ.
Твердые единицы дозировки, например, таблетки и капсулы, можно изготавливать с помощью технологий, известных в данной области техники. Например, PUFA, полученные в соответствии с настоящим изобретением, можно получать в виде таблеток, с такими традиционными наполнителями, как лактоза, сахароза и кукурузный крахмал, в сочетании с такими связывающими веществами, как гуммиарабик, кукурузный крахмал или желатин, агентами, улучшающими распадаемость, например, картофельный крахмал или альгиновая кислота, и лубрикантами, например, стеариновая кислота или стеарат магния. Капсулы можно изготавливать путем помещения указанных наполнителей в желатиновую капсулу вместе с антиоксидантами и соответствующей (соответствующими) PUFA. Включаемые в состав антиоксиданты и PUFA должны соответствовать рекомендациям, приведенным выше.
Для внутривенного введения, PUFA, полученные в соответствии с настоящим изобретением, или их производные, можно включать в коммерческие составы, например, Интралипид™. В обычном случае содержание жирных кислот в плазме крови составляет от 6.64 до 9.46% ARA, от 1.45 до 3.11% DGLA и от 0.02 до 0.08% GLA. Указанные PUFA или их предшественники в метаболических реакциях можно вводить отдельно или в сочетании с другими PUFA для достижения нормального содержания жирных кислот в организме пациента. При желании, отдельные составляющие лекарственных форм можно предоставлять по отдельности, в форме набора, для однократного или многократного применения. Обычная дозировка определенной жирной кислоты составляет от 0.1 мг до 20 г (до 100 г) ежедневно, предпочтительно от 10 мг до 1, 2, 5 или 10 г ежедневно.
Возможные способы введения фармацевтических композиций согласно настоящему изобретению включают, например, энтеральный (например, пероральный и ректальный) и парентеральный. Например, жидкий состав можно вводить, например, пероральным или ректальным путем. Также, гомогенную смесь можно полностью диспергировать в воде, смешанной в стерильных условиях с физиологически приемлемыми разбавителями, консервантами, буферами или пропеллентами, для образования спрея или средства для ингаляции.
Способ введения будет зависеть, разумеется, от целевого эффекта. Например, если композицию применяют для лечения шелушения, сухости или старения кожи, для лечения повреждения или ожога кожи или для лечения кожи или волос, поврежденных в результате заболевания или болезненного состояния, возможно местное применение.
Дозировку композиции, вводимую пациенту, должен определять специалист в данной области техники, и она будет зависеть от различных факторов, например, масса тела пациента, возраст пациента, иммунный статус пациента и т.д..
В отношении формы, композицию можно предоставлять, например, в виде раствора, дисперсии, эмульсии или стерильного порошка, восстанавливаемого перед введением.
Также в объем настоящего изобретения входит лечение различных нарушений путем применения фармацевтических и/или пищевых композиций, описанных в настоящей заявке. В частности, композиции согласно настоящему изобретению можно применять для лечения рестиноза, развившегося в результате пластики сосудов. Далее, с помощью композиций согласно настоящему изобретению также можно лечить симптомы воспаления, ревматоидного артрита, астмы и псориаза. Также доказано, что PUFA могут принимать участие в метаболизме кальция; следовательно, композиции согласно настоящем у изобретению можно применять, например, для лечения или предотвращения развития остеопороза или камней, образующихся в почках или мочевыводящих путях.
Также, композиции согласно настоящему изобретению можно применять для лечения рака. Было показано изменение состава жирных кислот в злокачественных клетках. Доказано, что добавление жирных кислот замедляет их рост, вызывает смерть клеток и повышает их чувствительность к химиотерапевтическим агентам. Более того, композиции согласно настоящему изобретению могут быть полезны для лечения истощения, связанного с раком.
Композиции согласно настоящему изобретению можно применять для лечения диабетов (см. патент США №4,826,877 и Horrobin D F et al., (1993) Am. J. Clin. Nutr. Vol.57 (Suppl.) 732S-737S). В организме животных, страдающих диабетом, были показаны изменения в метаболизме и составе жирных кислот.
Далее, композиции согласно настоящему изобретению, содержащие PUFA, полученные прямо или опосредованно путем применения фермента Δ8-десатуразы, также можно применять для лечения экземы, для снижения кровяного давления и для повышения экзаменационных оценок по математике. Также, композиции согласно настоящему изобретению можно применять для ингибирования агрегации тромбоцитов, индукции расширения просвета кровеносных сосудов, снижения уровней холестерола, подавления пролиферации в гладкомышечной и фиброзной ткани стенок сосудов (Brenner et al., (1976) Adv. Exp. Med. Biol. Vol.83, p.85-101), уменьшения или предотвращения кровотечений в желудочно-кишечном тракте и других подобных эффектов применения нестероидных и противовоспалительных препаратов (См. патент США №4,666,701), предотвращения или лечения эндометриоза и предменструального синдрома (См. патент США №4,758,592), а также для лечения миалгического энцефаломиелита и синдрома хронической усталости после перенесения вирусных инфекций (См. патент США №5,116,871).
Дальнейшие способы применения композиций согласно настоящему изобретению включают применение их для лечения СПИДа, множественного склероза и воспалительных заболеваний кожи, а также для общего поддержания здоровья организма.
Также, композицию согласно настоящему изобретению можно применять в косметических целях. Указанную композицию можно добавлять в уже существующие фармацевтические композиции, для составления смеси или в качестве основы для композиции.
G. Способы применения в ветеринарии
Следует заметить, что описанные выше фармацевтические и пищевые композиции можно применять в отношении животных (то есть, домашних и не домашних), также как и в отношении людей, поскольку животные испытывают многие потребности и состояния, сходные с таковыми для людей. Например, масла или кислоты согласно настоящему изобретению можно применять в пищевых добавках для животных или для водной культуры, заменителях пищи для животных, витаминах для животных или в составе мазей местного применения для животных.
Настоящее изобретение можно пояснить с помощью следующих примеров, не ограничивающих объем настоящего изобретения.
Пример 1
Разработка вырожденных олигонуклеотидов для выделения Δ8-десатуразы из штамма Emiliana huxleyi CCMP 378 и составления библиотеки кДНК
В ходе анализа состава жирных кислот морской водоросли выявили присутствие значительного количества докозагексаеновой кислоты (DHA, 22:6 n-3) (40% по массе от общего содержания липидов) в штамме Emiliana huxleyi CCMP 378 (См., таблица 1). Также, в указном организме представлены промежуточные продукты альтернативного пути 'А8-десатуразы-Δ9-элонгазы' (См., Фиг 1), что указывает на активность указанного пути в указанном организме. Следовательно, предполагают, что указанный организм содержит активную Δ9-элонгазу, способную к превращению линолевой кислоты (LA, 18:2n-6) в эйкозадиеновую кислоту (EDA, 20:2n-6) или альфа-линоленовой кислоты (ALA, 18:3, n-3) в эйкозатриеновую кислоту (ETrA, 20:3n-3), а также активную Δ8-десатуразу, превращающую эйкозадиеновую кислоту (EDA, 20:2n-6) в дихомо-гамма-линоленовую кислоту (DGLA, 20:3n-6) или ω3- эйкозатриеновую кислоту (ω3-EtrA, 20:3n-3) в ω3-эйкозатетраеновую кислоту (ω3-ETA, 20:4n-3) (См., Фигуру 1).
Целью данного исследования было выделение прогнозируемого полноразмерного гена Δ8-десатуразы из штамма Emiliana huxleyi CCMP 378 и проверка его активности путем экспрессии в Saccharomyces cerevisiae. Для этого создавали библиотеку нормализованных кДНК для Emiliana huxleyi CCMP 378. Массу клеток Emiliana huxleyi CCMP 378 получали в Национальном Центре морского фитопланктона Provasoli-Guillard (CCMP-Bigelow Laboratories, West Boothbay, Me.). Общую РНК очищали с помощью набора Qiagen RNeasy Maxi (Qiagen, Valencia, Calif.) в соответствии с инструкциями производителя. Вкратце, замороженную массу клеток разрушали в жидком азоте с помощью ступки и пестика, суспендировали в буфере RLT buffer (Qiagen RNeasy Plant Mini kit) и пропускали через QiaShredder. РНК очищали с помощью макси-колонок RNeasy в соответствии с инструкциями производителя.
Библиотеки первичных и нормализованных кДНК для Emiliana huxleyi CCMP 378 создавали с помощью Agencourt Biosciences (Waltham, Mass.), с применением запатентованной ими технологии. Agencourt применяет несколько нестандартных и запатентованных этапов в ходе первого витка, который, в конечном итоге, обеспечивает повышение эффективности на 25-30% по сравнению с обычно применяемыми методиками. В ходе осуществления запатентованного способа, проводят обратную транскрипцию РНК в одноцепочечную ДНК при условиях, применяемых для уменьшения или устранения посторонних событий прайминга. Сочетание приведенного процесса со специализированной программой циклирования повышает количество полноразмерных клонов. В ходе второго витка синтеза, отбирают клоны кДНК, размер которых превышает 1.2 т.н., для снижения уровня клонирования небольших, усеченных молекул кДНК. Размер вставок для библиотеки крупных вставок составлял >4 п.н., для отбора клонов со вставками большего размера. После проведения отбора по размеру, концы кДНК зачищают, и кДНК расщепляют с помощью редкощепящего фермента. ″Редкощепящий″ фермент рестрикции, сайт для которого вводят в клоны в ходе этапа праймирования ДНК, затем применяют для создания клонов для направленного клонирования в вектор pAGEN. Вероятность расщепления клонов кДНК ″редкощепящим″ ферментом рестрикции в 20 раз меньше, вследствие чего образуется значительно больше полноразмерных клонов по сравнению с другими способами создания библиотеки кДНК, в ходе которых применяют стандартные ферменты рестрикции, вносящие разрезы в случайных интервалах в пределах клона. Итогом является вставка на 5′-тупом конце и ″липкий″ 3′-конец, образующийся в результате применения редкощепящего фермента. Благодаря описанному способу, не требуется дополнительного адаптерного лигирования для обеспечения направленного клонирования. Это улучшает общую эффективность процесса клонирования. Вектор специально конструируют для направленного клонирования без применения 5′-адаптеров, что обеспечивает дальнейшее повышение эффективности трансформации благодаря снижению количества манипуляций с кДНК в ходе клонирования. После завершения создания библиотеки первичных кДНК, ее исследуют на количество независимых клонов, процент рекомбинантных клонов и средний размер вставки.
Процесс нормализации начинают с разделения стандартной библиотеки на две популяции. Первую популяцию линеаризируют и транскрибируют с кДНК в РНК, при включении биотинилированных нуклеотидов. Из второй популяции создают одноцепочечные ДНК плазмиды путем образования фагмид. Двухцепочечную ДНК в лизате клеток расщепляют с помощью ДНКазы I. Таким образом устраняют контаминацию ДНК плазмид из препарата одноцепочечной ДНК.
После этого две популяции смешивают, и любые перепредставленные клоны из плазмид одноцепочечной ДНК вступают в реакцию гибридизации с их партнерами в составе популяции биотинированой РНК. Agencourt применяет олиго-dT и праймер удлинения для предварительного блокирования участка поли-A до гибридизации. Таким образом предотвращают гибридизацию клона РНК поли-A и клона поли-U. С помощью способа экстракции, применяющего стрептавидин/фенол, удаляют все биотинилированные гибридные пары и линеаризованные биотинилированные молекулы РНК, следовательно, остаются одноцепочечные, перепредставленные плазмиды ДНК. С помощью олигонуклеотида, гибридизующегося только с клонами, содержащими вставку, синтез ДНК праймируют для воссоздания двухцепочечных клонов ДНК. После этого клоны трансформируют в бактерии для создания итоговой библиотеки нормализованных кДНК. Путем применения приведенного протокола, была создана библиотека нормализованных кДНК с титром, составлявшим 2.6×107 KOE/мл, с общим количеством полученных колоний, составлявшим 1.8×108, и средним размеров вставки, составлявшим 1.05 т.п.н..
Для выделения кандидатов, похожих на ген Δ8-десатуразы, из библиотеки, разрабатывали вырожденные олигонуклеотиды (то есть, праймеры), кодирующие консервативные аминокислотные мотивы, присутствующие в составе известных Δ8-десатураз. После этого указанные праймеры применяли в ходе реакции ПЦР для определения фрагментов ДНК, содержащих указанные консервативные участки в составе прогнозируемой Δ8-десатуразы.
Для выравнивания и разработки праймеров применяли известные аминокислотные последовательности Δ8-десатуразы следующих организмов: Euglena gracialis (№ доступа AF 139720, SEQ ID NO:1; Фигура 3A), Pavlova lutheri CCMP 459 (WO 2007/127381A2, SEQ ID NO:2; Фигура 3B), Pavlova salina ((№ доступа DQ995518, SEQ ID NO:3; Фигура 4A), Perkinsus marinus ((№ доступа DQ508730, SEQ ID NO:4; Фигура 4B) и Acanthamoenba castellani ((№ доступа CS608483, SEQ ID NO:5; Фигура 4C).
Применяемые вырожденные праймеры представляли собой следующие:
Белковый мотив 1: (SEQ ID NO:38):
NH3 - R D A T D/E A/Q F E/M S/V Y/M H-COOH
Праймер RO 1714 (Прямой) (SEQ ID NO:6):
5′-CGC GAC GCG ACG GAS SMG TTC RWG KYK WWS CAC-3′
Приведенный праймер содержал последовательность консервативного мотива в прогнозируемом участке цитохрома b5.
Белковый мотив 2: (SEQ ID NO:39):
NH3 - G W L A/S H D Y/I L/S H H-COOH
Праймер RO 1715 (Прямой) (SEQ ID NO:7):
5′-GGC TGG CTT KCK CAC GAC WWC YYG CAT CAC-3′
Приведенный праймер содержал последовательность консервативного мотива '1 гистидинового бокса
Белковый мотив 3: (SEQ ID NO:40):
NH 3 - W K/R A/L R H N T/A H H-COOH
Праймер RO 1716 (Прямой) (SEQ ID NO:8)
5′-TGG MRS SYG CGC CAT AAC RCG CAC CAC GTG KSC AGC AAC-3′
Приведенный праймер содержал последовательность консервативного мотива '2 гистидинового бокса'
Белковый мотив 4: (SEQ ID NO:41):
NH 3 - F A/G T A/G I/V V V F A T H Y-COOH
Праймер RO 1717 (Обратный) (SEQ ID NO:9)
5′-ATA GTG GGT TGC AAA GAC AAC SAY SSC CGT CSC GAA-3′
Белковый мотив 5: (SEQ ID NO:42):
NH 3 -Q I/T H H L F P T/M M P-COOH
Праймер RO 1718 (Обратный) (SEQ ID NO:10)
5′-GGG CAT SRT GGG GAA GAG GTG ATG CTC GRT CTG-3′
Приведенный праймер содержал последовательность консервативного мотива '3 гистидинового бокса'
Стандартное определение MixBase для синтеза олигонуклеотидов представляло собой: K=G,T; R=A,G; S=C,G; M=A,C; W=A,C; Y=C,T; B=C,G,T; H=A,C,T; V=A,C,G; D=A,T,C; X=A,C,G,T.
Пример 2
Выделение прогнозируемого гена Δ8-десатуразы Emiliana huxleyi CCMP 378
Для выделения гена Δ8-десатуразы Emiliana, для реакций ПЦР применяли различные перестановки и сочетания описанных выше вырожденных олигонуклеотидов (См., Пример 1). Амплификацию в ходе ПЦР проводили в объеме, составлявшем 50 мкл, содержащих: 2 мкл плазмидной ДНК, выделенной из библиотеки нормализованных кДНК, в качестве затравки, 1 × буфер ПЦР минус MgCl 2 (20 мМ Tris-HCl, pH 8.4, 50 мМ KCl), 1.5 мМ MgSO 4, 200 мкм каждого из дНТФ, 2 пмоль каждого праймера и платиновую ДНК Taq-полимеразу (Invitrogen). Амплификацию осуществляли следующим путем: начальная денатурация при 94°C/3 мин, с последующими 35 циклами следующего (94°C/30 секунд; 55°C/30 секунд; 72°C/1 минута), итоговое удлинение при 72°C/5 минут, и реакцию останавливали при 4°C. Реакцию ПЦР очищали с помощью набора Qiagen MinElute Reaction Cleanup (Qiagen Valencia, Calif.), и реакцию разделяли в 0.8% агарозном геле. Бэнды соответствующего размера (на основе известных Δ8-десатураз) очищали в геле с применением набора QiaQuick Gel Extraction (Qiagen), и указанные фрагменты ДНК клонировали в вектор клонирования TOPO-TA (Invitrogen, Carlsbad, Calif.), в соответствии с протоколом производителя. Рекомбинантные плазмиды трансформировали в суперкомпетентные клетки TOP10 (Invitrogen, Carlsbad, Calif.), и клона секвенировали. Единственным сочетанием праймеров среди различных исследованных наборов праймеров, образовавшим фрагменты ДНК со сходством последовательностей с известными десатуразами Δ8-десатураз было RO 1715 и RO 1717.
Следовательно, был выделен один клон, для которого была показана гомология последовательностей с ранее идентифицированными Δ8-десатуразами. Длина указанного клона (ED3-8) составляла 531 т.п.н., и расшифрованная аминокислотная последовательность, полученная из него, обладала 62% идентичности аминокислотной последовательности Δ8-десатуразы Pavlova lutheri CCMP 459 (См., WO 2007/127381A2, SEQ ID NO:2; Фигура 3В) как самый высокий процент совместимости в исследовании BLAST. ДНК и расшифрованная аминокислотная последовательность указанного клона приведены (SEQ ID NOS:11 и 12; Фигуры 5 и 6, соответственно).
Для выделения 5′-конца фрагмента ED3, проводили ПЦР амплификацию с применением плазмидной ДНК, очищенной из библиотеки кДНК, в качестве матрицы, и олигонуклеотиды (праймеры): RO 1720 (SEQ ID NO:13) (5′-GAT CAC CGG GCT GTT GCG CAC GAA G-3′) и RO 899 (SEQ ID NO:14) (5′-AGCGGATAACAATTTCACACAGGAAACAGC-3′). Праймер RO 1720 создавали на основе фрагмента ED3-8 прогнозируемой Δ8-десатуразы а праймер RO 899 соответствовал последовательности из вектора pAGEN, применявшегося для создания библиотеки кДНК. Амплификацию проводили с применением 10 пмоль каждого праймера, 1 мкл матрицы ДНК, 1.5 мкл 50 мМ MgSO4, 1 × буфера ПЦР (Qiagen), 0.5 × раствора энхансера (итоговая концентрация), 1 мкл 10 мМ дНТФ и 0.5 мкл платиновой ДНК Taq-полимеразы(Qiagen) в итоговом объеме, составлявшем 50 мкл, в соответствии с инструкциями производителя. Образцы подвергали денатурации при начальном значении температуры, составлявшем 94°C, в течение 2 минут, затем проводили 35 циклов при следующих режимах: 94°C в течение 45 секунд, 55°C в течение 30 секунд, 68°C в течение 1 минуты. Итоговый цикл удлинения проводили при 68°С в течение 7 минут, после чего реакцию останавливали при 4°C. Фрагменты, полученные в ходе ПЦР, разделяли в 0.8% агарозном геле и очищали в геле с применением набора Qiagen Mini-elute Gel Extraction. Фрагменты ДНК клонировали в вектор клонирования TOPO-TA (Invitrogen). Рекомбинантные плазмиды трансформировали в суперкомпетентные клетки TOP10 (Invitrogen), и клоны секвенировали.
Клон PK15 содержал вставку 692 т.п.н. (SEQ ID NO:15; Фигура 7A), в которой установили содержание 5'-конца прогнозируемого гена Δ8-десатуразы на основе гомологии аминокислотных последовательностей с известными Δ8-десатуразами, и наличие стартового кодона 'ATG' 'Met'. Кодируемая аминокислотная последовательность указанного клона PK15, содержащего 5′-конец прогнозируемой Δ8-десатуразы, приведена (SEQ ID NO:16; Фигура 7B).
Для выделения 3′-конца указанной Δ8-десатуразы Emiliana huxleyi, применяли следующие праймеры:
Прямые праймеры RO 1719 (SEQ ID NO:17):
5′-GTA CCA GTG GCT GCT GCT GAC GAT G-3′)
или
Прямые праймеры RO 1724 (SEQ ID NO:18):
5′-CTG GCG CTT CGA GTC GAT GCA GTA CCT-3′
или
Прямые праймеры RO 1727 (SEQ ID NO:19):
5′-CTT CGT GCG CAA CAG CCC GGT GAT C-3′
и
Обратный праймер RO 898 (SEQ ID NO:20):
5′-CCCAGTCACGACGTTGTAAAACGACGGCCAG-3′
RO 1719, RO 1724 и RO 1727 были разработаны на основе последовательности фрагмента ED3-8, идентифицированного ранее. RO 898 был разработан на основе последовательности вектора pAGEN, применявшегося для конструирования библиотеки кДНК, амплифицировавшего в ходе ПЦР 3′-конец Δ8-десатуразы из библиотеки кДНК. Применяли условия реакции ПЦР, аналогичные описанным для выделения 5′-конца фрагмента ED3-8. Тем не менее, ни один из созданным подобным образом продуктов ПЦР не содержал 3′-конец ED3-8 прогнозируемой Δ8-десатуразы.
Также проводили реакцию ПЦР с помощью платинового фермента высокой точности Taq High Fidelity (HF) (Invitrogen) в соответствии с инструкциями производителя с применением 1×HF буфера ПЦР (итоговая концентрация), предоставляемого в наборе, 10 пмоль каждого праймера, 1 мкл ДНК матрицы, 1.5 мкл 50 мМ MgSO4, 1 мкл 10 мМ дНТФ м 0.5 мкл платиновой ДНК Taq HF-полимеразы (Qiagen) в итоговом объеме, составлявшем 50 мкл. Образцы подвергали денатурации при начальном значении температуры, составлявшем 94°C, в течение 2 минут, затем проводили 30 циклов при следующих режимах: 94°C в течение 30 секунд, 55°C в течение 30 секунд, 68°C в течение 2 минут. Итоговый цикл удлинения проводили при 68°C в течение 7 минут, после чего реакцию останавливали при 4′C. Тем не менее, ни один из созданным подобным образом продуктов ПЦР не содержал 3′-конец ED3-8 прогнозируемой Δ8-десатуразы.
Итоговую ПЦР амплификацию с помощью праймеров RO 1727 и RO 898, в сочетании с ДНК-полимеразой AccuPrime pfx (Invitrogen), проводили в соответствии с протоколом производителя. Реакция ПЦР содержала 2 мкл матрицы ДНК из библиотеки кДНК, 1 × итоговую концентрацию буфера AccuPrime Pfx, 20 пмоль каждого праймера и 1 мкл ДНК-полимеразы AccuPrime pfx в мкл общей реакции. Образцы подвергали денатурации при начальном значении температуры, составлявшем 95°C, в течение 2 минут, затем проводили 30 циклов при следующих режимах: 95°С в течение 15 секунд, 55°C в течение 30 секунд, 68°С в течение 1 минуты. Реакцию останавливали при 4°C. Тем не менее, ни один из созданным подобным образом продуктов ПЦР не содержал 3′-конец ED3-8 прогнозируемой Δ8-десатуразы.
После этого применяли новый способ выделения 3′-конца прогнозируемого гена Δ8-десатуразы, проводили ED3-8. RACE (быстрая амплификации концов кДНК) с применением набора GeneRace (Invitrogen) для выделения 3′-конца. Для создания первой цепи кДНК применяли 5 мкг общей ДНК, выделенной из Emiliana, в сочетании с праймером GeneRacer Oligo dT (5′-GCT GTC AAC GAT ACG CTA CGT AAC GGC ATG АСА GTG (T) 24-3′ (SEQ ID NO:43)), в соответствии с рекомендациями производителя.
ПЦР амплификацию готовых кДНК, описание создания которых с помощью RACE приведено выше, проводили с применением 30 пмоль праймера GeneRacer 3′ (5′-GCT GTC AAC GAT ACG CTA CGT AAC G-3′(SEQ ID NO:22)) и 10 пмоль RO 1724 прямого праймера (SEQ ID NO:18), обладающих специфичностью по отношению к прогнозируемой Δ8-десатуразе (ED3-8), в соответствии с рекомендациями производителя. Реакция ПЦР включала 2 мкл матрицы кДНК, 1×HF буфера ПЦР (Invitrogen), 1 мкл 10 мМ дНТФ, 2 мкм 50 мМ MgSO4, 0.5 мкм платиновой ДНК Taq HF-полимеразы, в сочетании с указанными выше праймерами, в общем объеме реакции, составлявшем 50 мкл. ПЦР амплификацию проводили следующим способом: начальная денатурация при 94°C/2 минуты; 5 циклов денатурации при 94°C/30 секунд; удлинение при 72°C/1 минута; 5 циклов денатурации при at 94°C/30 секунд; удлинение при 70°C/1 минуту; 20 циклов денатурации при 94°C/30 секунд; удлинение при 65°C/30 секунд; итоговое удлинение при 65°C/10 минут; остановка реакции при 4°C. В ходе анализа продуктов ПЦР выявили полосы очень небольшой интенсивности, возможно, по причине низкого количественного содержания гена в пуле кДНК. Вследствие этого проводили гнездовую реакцию ПЦР с применением 1 мкл реакции ПЦР, получение которой описано выше, в качестве матрицы, в сочетании с 10 пмоль праймера RO 1719 (SEQ ID NO:17), обладающего специфичностью по отношению к гену, и 10 пмоль гнездового праймера GeneRacer 3′ (5′-CGC TAC GTA ACG GCA TGA CAG TG-3′ (SEQ ID NO:23)). Ход реакции ПЦР был аналогичен описанному для основной реакции с готовой матрицей кДНК (выше). Применяли следующие условия амплификации: денатурация при 94°C/2 минуты; 25 циклов денатурации при 94°C/30 секунд, отжиг при 65°C/30 секунд, удлинение при 68°C/1 минуту; итоговое удлинение при 68°C/10 минут, и реакцию останавливали при 4°C. В ходе анализа фрагментов, полученных в реакции ПЦР, в 0.8% агарозном геле, выявили наличие четких бэндов. Их очищали в геле с помощью набора Qiagen Mini-elute Gel Extraction. Концы фрагментов ДНК заплавляли с помощью полимеразы T4, и образованные фрагменты с тупыми концами клонировали в вектор клонирования ТОРО-Blunt cloning (Invitrogen). Рекомбинантные плазмиды трансформировали в суперкомпетентные клетки TOP 10 (Invitrogen), и клоны секвенировали. В ходе анализа последовательностей выявили вставку, длина которой составляла 589 т.п.н. (SEQ ID NO:24; Фигура 8A), кодируемая последовательность которой (SEQ ID NOS: 25 и Δ4-46; Фигура 8B) содержала прогнозируемый 3′-конец ED3-8, на основе гомологии аминокислотных последовательностей с известными Δ8-десатуразами и наличии стоп-кодона 'TAG' и поли-A хвоста.
Полноразмерный ген прогнозируемой Δ8-десатуразы, ED3-8, выделили с помощью реакции ПЦР с применением следующих праймеров:
RO 1736 (SEQ ID NO:26):
(Прямой, содержащий стартовый кодон ATG (выделен жирным шрифтом) и сайт клонирования EcoRI (подчеркнут)
5′-AAA GAA TTC ATG GGC AAG GGC GGC AAC GCG AAC C-3′
RO 1737 (SEQ ID NO:27):
(Обратный, содержащий стоп-кодон TGA (выделен жирным шрифтом) и сайт клонирования HindIII (подчеркнут)
5′-AAA AAG CTT CTA GTG CGG CAT CTC TGC CCA CTC G-3′
Матрицы, применявшиеся для проведения реакции ПЦР, включали либо готовую кДНК RACE, либо ДНК, выделенную из библиотеки нормализованных кДНК Emiliana, в соответствии со следующим:
Амплификацию проводили следующим способом: начальная денатурация при 95°C/2 минуты; 30 циклов (денатурация при 95°C/15 секунд; отжиг при 55°C/30 секунд; удлинение при 68°C/1.5 минуты); итоговое удлинение при 68°C/4 минуты; реакцию останавливали при 4°C. В результате реакции ПЦР получали один бэнд, длина которого составляла ~1254 т.п.н., который клонировали в вектор TOPO-Blunt (Invitrogen) в соответствии с протоколом производителя. В результате применения обеих матриц образовывался бэнд ДНК одинакового размера. В ходе секвенирования продукта ПЦР (ED3-8-EP2-5), полученного путем применения библиотеки кДНК в качестве матриц, выявили последовательность полноразмерного гена прогнозируемой Δ8-десатуразы Emiliana huxleyi CCMP 378, размер которой составлял 1254 т.п.н. (SEQ ID NO:28; Фигура 9), колирующую белок, включающий 417 аминокислот (SEQ ID NO:29; Фигура 10). Указанный ген обозначили как ED3-8, и применяли его для исследования экспрессии.
Помимо ED3-8, в ходе секвенирования обнаружили дополнительные варианты клонов, обладающие некоторыми различиями последовательностей в отдельных участках полноразмерного гена (См., Таблица 2). Возможно, указанные вариации вызваны мутациями, в норме происходящими в ходе ПЦР амплификации, по причине низкой специфичности применявшейся полимеразы. Также клоны получали как в результате применения библиотеки кДНК в качестве матрицы (обозначенные как ED3-8-EP-X), так и при применении в качестве матрицы готовым кДНК RACE (обозначенные как ED3-8-ER-X). Указанные гены также изучали на предмет активности Δ8-десатуразы, в соответствии с описанием, приведенным ниже, для исходного клона ED3-8.
Нуклеотидную последовательность, кодирующую Δ8-десатуразу ED3-8, клонировали в вектор клонирования pUC57, обозначенный как pRSP61. Указанный вектор был помещен в Американскую коллекцию типовых культур (ATCC), 10801 University Boulevard, Manassas, Va. 20110 в соответствии с условиями Будапештского договора, 9 сентября, 2008, и ему было присвоено патентное цифровое обозначение ATCC PTA-9477.
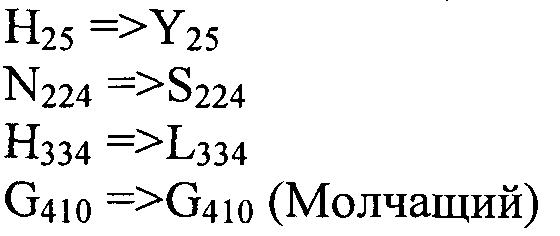
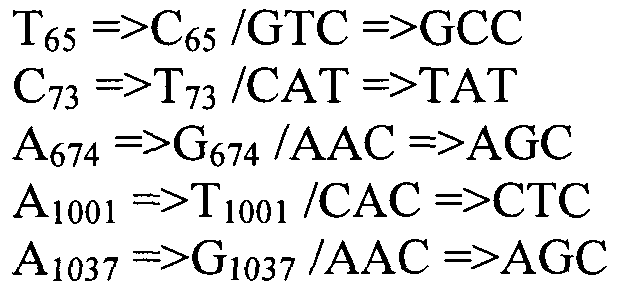
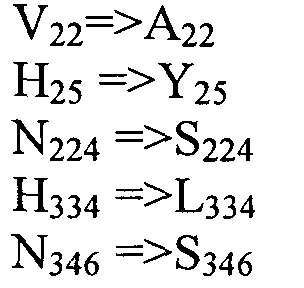
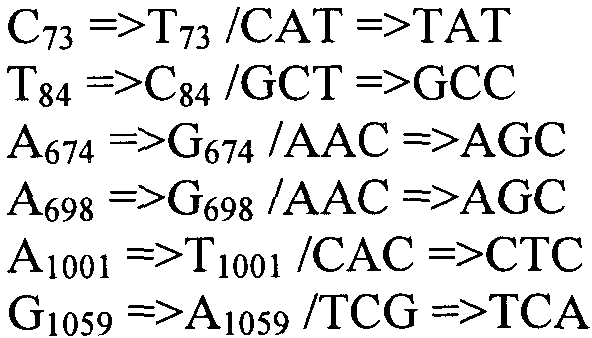

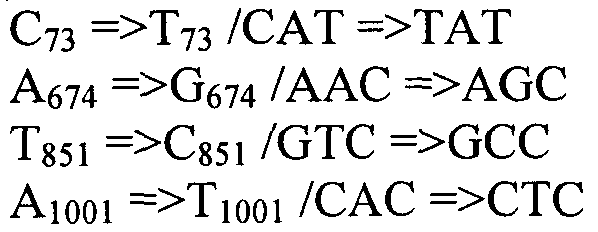
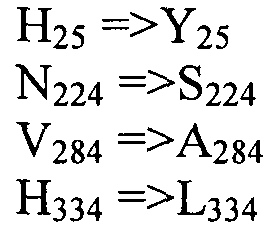
В ходе анализа бластов выявили аминокислотную последовательность, кодируемую полноразмерным геном ED3-8 (SEQ ID NO:29), обладающую высокой гомологией с последовательностью известных Δ8-десатураз. Указанные последовательности включали последовательности Δ8-десатураз Pavlova lutheri CCMP 459 ((SEQ ID NO:2; Фигура 3B), Pavlova salina (SEQ ID NO 3; Фигура 4A), Perkinsus marinus (SEQ ID NO:4; Фигура 4В) и Euglena gracialis (SEQ ID NO:1; Фигура 3A). Указанный кодируемый белок обладал самой высокой идентичностью аминокислотной последовательности (52.3%)аминокислотной последовательности Δ8-десатуразы Pavlova lutheri CCMP 459. Он также включал три консервативных 'гистидиновых бокса', обнаруживаемые в составе всех известных связанных с мембраной десатураз (Okuley, et al. (1994) The Plant Cell 6:147-158; Pereira S L et al (2003) Prostaglandins Leukot Essent Fatty Acids. 68:97-106), для которых показана значительная ферментативная активность (Sayanova O et al. (2001) J Exp Bot. 52:1581-1585; Sayanova O et al. (2000) Biochem Soc Trans. 28:636-638). Консервативные гистидиновые боксы в составе белка, кодируемого ED3-8, находились в 155-160 аминокислотных положениях (HDYLH (SEQ ID NO:32)), 197-201 (HNTHH (SEQ ID NO:33)) и 355-359 (QTEHH (SEQ ID NO:34)) (См., Фигура 2). Указанная последовательность также включала область, подобную цитохрому b5, на 5′-конце, с консервативным мотивом связывания гема HPGG (38-41 аминокислотных положениях) (См., Фигура 2). Общее содержание G+C в указанном гене составляло ~65%.
Пример 3
Описание ферментативной активности прогнозируемой Δ8-десатуразы, кодируемой геном ED3-8
Ген ED3-8, кодирующий прогнозируемую Δ8-десатуразу, клонировали в области сайтов EcoRI/HindIII дрожжевого вектора экспрессии, pYX242 (Novagen) для создания клона pRSP60, который затем трансформировали в компетентный штамм Saccharomyces cerevisiae SC334. Трансформацию дрожжей осуществляли с помощью набора для трансформации дрожжей Alkali-Cation (QBioGene) в соответствии с условиями, определенными производителем. Трансформанты отбирали по признаку ауксотрофности по отношению к лейцину, на среде без добавления лейцина (DOB [-Leu]).
Для определения специфичной десатуразной активности фермента, кодируемого ED3-8, трансформанты выращивали в присутствии 50 мкм специфичных субстратов жирных кислот (приведенных ниже), и для определения специфичности субстрата применяли превращение в определенный продукт:
Для изучения активности Δ8-десатуразы:
Эйкозадиеновая кислота (EDA, 20:2n-6)  дихомо-гамма-линоленовая кислота (DGLA, 20:3n-6)
дихомо-гамма-линоленовая кислота (DGLA, 20:3n-6)
Эйкозатриеновая кислота (ETrA, 20:3n-3) ω3-эйкозатетраеновая кислота (ω3-ЕТА, 20:4n-3)
Для изучения активности Δ6-десатуразы:
Линолевая кислота (18:2n-6) гамма-линоленовая кислота (GLA, 18:3n-6)
Альфа-линоленовая кислота (18:3n-3) стеаридоновая кислота (SDA, 18:4n-3)
Для изучения активности Δ5-десатуразы:
Дихомо-гамма-линоленовая кислота (20:3n-6) арахидоновая кислота (ARA, 20:4n-6)
ω3-эйкозатетраеновая кислота (ω3-ЕТА, 20:4n-3) эйкозапентаеновая кислота (ЕРА, 20:5n-3)
Для изучения активности Δ4-десатуразы:
ω6-адреновая кислота (ADA, 22:4n-6) ω6-докозапентаеновая кислота (ω6-DPA, 22:5n-6)
ω3-докозапентаеновая кислота (ω3-DPA, 22:5n-3) докозагексаеновая кислота (DHA, 22:6n-3)
Штамм, применявшийся в качестве отрицательного контроля, включал вектор pYX242, экспрессированный в S.cerevisiae 334.
Трансформированные колонии, выделенные из селективной среды DOB [Leu], выращивали в течение ночи в 10 мл жидкой питательной среды YPD при 30°C, при активном перемешивании. После этого 5 мл ночной культуры добавляли к 45 мл селективной среды (DOB [Leu]), содержащей 50 или 25 нМ (итоговая концентрация) различных субстратов жирных кислот (в соответствии с указаниями), и образованную смесь подвергали активному перемешиванию (250 об.) в течение 48-72 часов (согласно указаниям) при 24°C.
Для полной экстракции липидов, клетки дрожжей откручивали при 2000 об/15 минут, и добавляли 0.5 мл воды, образцы перемешивали на вортексе, после чего добавляли 10 мл метанола при легком покручивании. После этого добавляли 20 мл хлороформа, образцы перемешивали на вортексе в течение 1 минуты на большой скорости и оставляли на 2 часа при комнатной температуре. После этого к образцам добавляли 6 мл солевого раствора с последующим центрифугированием при 2200 об в течение 10 минут. Верхний слой хлороформа удаляли в чистую сухую пробирку, объем которой составлял 30 мл, и хлороформ выпаривали досуха при 40°C под потоком азота. После полного выпаривания растворителей, в каждую пробирку добавляли по 2 мл хлороформа, и получали производные из образцов.
Для получения метиловых эфиров жирных кислот (FAME) в качестве производных липидов, в каждую пробирку вносили по 100 мкл тригептадеканоина в качестве внутреннего стандарта (17.216 мкг/100 мкл). Хлороформ выпаривали досуха под азотом при 40°C, добавляли 2 мл трехфтористого бора в 14% метаноле, после чего добавляли 2 капли (~50 мкл) толуола. Каждую пробирку промывали под азотом, и нагревали в течение 15 минут при 95°C. После охлаждения пробирок, добавляли 2 мл солевого раствора, и липиды экстрагировали с помощью 4 гексана при интенсивном перемешивании на вортексе в течение 1 минуты. После этого экстракт в гексане переносили в в чистую сухую пробирку с завинчивающейся крышкой, объем которой составлял 20 мл, добавляли 5 мл ди-H2O и перемешивали образцы на вортексе, после чего центрифугировали при 1500 об/мин в течение 4 минут. После этого переносили отмытый гескан в пробирку для реагентов, объем которой составлял 20 мл. гексан выпаривали досуха, и каждый образец восстанавливали с помощью 0.5 мл свежего гексана. Итоговую восстановленную реакцию в гексане перемешивали на вортексе для диспергирования липидов. Весь объем образца помещали в пробирки для автодозатора GC, и 4 мкл использовали для анализа. Калибровку GC проводили с помощью NuChek Std. 461.
Процент превращения субстрата в продукт вычисляли с помощью следующей формулы:
В Таблице 3 приведена ферментативная активность белка, кодируемого ED3-8, полученная на основе процента превращения вносимого субстрата. Клон pRSP60, содержавший ген ED3-8 Emiliana, преобразовывал 1.68% субстрата EDA (20:2n-6) в DGLA (20:3n-6), и 0.58% субстрата ETrA (20:3n-3) в ETA (20:4n-3). Данные указывают на то, что ген ED3 кодирует Δ8-десатуразу, способную распознавать субстраты как n-6, так и n-3, преимущественно, субстрат n-6, EDA. При применении только вектора в качестве контроля не обнаружили фоновой активности (не специфичного преобразования субстрата) (См. Таблицу 2). Фермент, кодируемый ED3-8, не обладает активностью в отношении каких-либо других исследованных субстратов (данные не приведены), что указывает на отсутствие у него Δ6-, Δ5- или Δ4-десатуразной активности.
Также, другие варианты белка, кодируемого ED3-8 (См. Таблицу 2) экспрессировали в S.cerevisiae, при аналогичных условиях культивирования (См. Таблицу 4). Результаты указывают на то, что варианты обладают более низкой Δ8-десатуразной активностью или не обладают ей, в сравнении с исходным белком, кодируемым ED3-8 (См. Таблицу 4). Указанные минорные замены в составе аминокислотной последовательности ED3-8 могут влиять на ферментативную активность, в зависимости от их расположения. Возможно, указанные замены влияют на каталитические центры фермента или на стабильность фермента, что приводит к снижению активности. Поскольку кристаллическая структура указанных мембранно-связанных десатураз еще не была расшифрована, невозможно предсказать каждый участок фермента, имеющий значение для ферментативной активности. Хорошо известно, что участки 'гистидиновых боксов' и участок цитохрома b5 принимают значительное участие в активности фермента. Тем не менее, ни один из указанных вариантов ED3-8 не содержал замен в участках 'гистидиновых боксов' и цитохрома b5 (См. Таблицу 2). Следовательно, определяли дополнительные участки в указанной Δ8-десатуразе, кодируемой ED3-8, имеющие значение для ферментативной активности.
20:3n-6)
Поскольку Δ8-десатуразная активность ED3-8 была ниже при анализируемых условиях культивирования, условия культивирования изменяли для определения возможности повышения активности каким-либо способом, путем изменения количества вносимого субстрата или изменением температуры и времени экспрессии. Соответственно, культуру дрожжей, трансформированных pRSP60. Выращивали в присутствии 25 мкм субстрата при 24°С в течение 48 часов или в присутствии 50 мкм или 25 мкм субстрата при 20°C в течение 72 часов. Данные, приведенные в Таблице 5, указывают на то, что изменения условий культивирования способны повышать экспрессию в клетках дрожжей. Процент преобразования субстрата в продукт повышался с ~1.7% до ~4.5%.
Пример 4
Оптимизация ED3-8 относительно кодонов и экспрессия белка, кодируемого им, в Saccharomyces cerevisiae
Поскольку содержание G+C в составе ED3-8 достигает высоких значений (~65%), это может быть причиной относительно низкой Δ8-десатуразной активности, проявлявшейся в ходе экспрессии в клетках дрожжей. Соответственно, частоту использования кодона ED3-8, Δ8-десатуразы Emiliana huxleyi CCMP 378 (SEQ ID NO:28; Фигура 9), оптимизировали для экспрессии в Saccharomyces cerevisiae. Характер частоты использования кодона Saccharomyces cerevisiae определяли в базе данных частоты использования кодона (См., Nakamura, Y., Gojobori, T. and Ikemura, T. (2000) Nucl. Acids Res. 28, 292), и полученные данные применяли для секвенирования гена ED3-8 с помощью программы Vector NTI (Invitrogen). Общий размер модифицированного участка для выравнивания частоты использования кодона с частотой использования кодона у Saccharomyces cerevisiae, составлял 412 т.п.н., в составе кодирующего участка, размер которого оставлял 1254 т.п.н. (~33%). В дополнение, удаляли внутренний сайт для HindIII, для усиления клонирования гена в сайт HindIII сайта множественного клонирования различных векторов экспрессии. Следовательно, общий размер модифицированного участка в составе кодирующего участка размером 1254 т.п.н., составлял 414 т.п.н.. Идентичность новой последовательности, оптимизированной относительно кодонов, и исходной последовательностью гена ED3-8, составляла 66.98%. Ни одна из модификаций в составе указанного гена, оптимизированного относительно частоты использования кодона, не изменяла аминокислотную последовательность колируемого белка (SEQ ID NO:29; Фигура 10). Также, к 5′-концу кодона инициации трансляции ATG добавляли 'ААА', что считается способствующим усилению экспрессии в клетках дрожжей. Указанную последовательность обозначали как 'ED3-8-EP2-5-SC' (SEQ ID NO:30; Фигура 11). Также добавляли фланкирующие сайты расщепления для усиления клонирования в различные векторы экспрессии. Ген, обозначенный как 'ED3-8-EP2-5-SC', синтезировали в корпорации GenScript (Piscataway, N.J.) и клонировали в участок клонирования 'ТА' в составе вектора клонирования pUC57. После этого указанный ген субклонировали в сайт EcoRI/SpeI в составе дрожжевого вектора экспрессии pESC-Ura (Stratagene) для создания конструкции, обозначенной как pRSP62.
Клон, содержавший ген ED3-8-EP2-5-SC (SEQ ID NO:30), клонировали в вектор pESC-Ura, обозначенный как pRSP62, помещали на хранение в американскую коллекцию типовых культур (АТСС), 10801 University Boulevard, Manassas, Va. 20110, в соответствии с условиями Будапештского соглашения, 26 сентября, 2008, и соответствовал патентному номеру депонирования ATCC PTA-9532.
pRSP62 трансформировали в Saccharomyces cerevisiae в соответствии с протоколом, описанным в Примере 3, и трансформанты отбирали по признаку ауксотрофности по отношению к урацилу на среде без добавления урацила (DOB[-Ura]) (QBioGene). Трансформированные колонии, выделенные из селективной среды DOB[Ura], выращивали в течение ночи в 10 мл жидкой среды YPD при 30°C, при энергичном перемешивании. После этого 5 мл ночной культуры добавляли в 45 мл селективной среды (DOB[-Ura] без декстрозы) с добавлением 2% галактозы (итоговая концентрация) для индуцирования экспрессии pRSP62 и 50 мкМ (итоговая концентрация) различных субстратов жирных кислот (в соответствии с указанным). Культуры подвергали энергичному перемешиванию (250 об/мин) в течение 48 часов (в соответствии с указанным) при 24°C. Выделение и анализ общего содержания жирных кислот также проводили в соответствии с описанием, приведенным в Примере 3.
В Таблице 6 представлены данные ферментативной активности белка, кодируемого 'ED3-8-EP2-5-SC', в процентах преобразования внесенного субстрата. Δ8-десатуразная активность гена ED3-8, оптимизированного относительно частоты использования кодона, была значительно выше, чем у белка, кодируемого исходным ED3-8. Результаты составили 12.51% преобразования субстрата EDA (20:2n-6) в DGLA (20:3n-6) и 8.45% преобразования субстрата ETrA (20:3n-3) в ETA (20:4n-3). При применении только вектора pESC-Ura в качестве контроля не обнаружили фоновой активности (не специфичного преобразования субстрата) (См. Таблицу 6). Фермент, кодируемый ED3-8, оптимизированным относительно частоты использования ко дона, не обладает Δ6-, Δ5- или Δ4-десатуразной активностью в отношении каких-либо других исследованных субстратов (данные не приведены).
Пример 5
Ко-экспрессия гена Δ8-десатуразы 'ED3-8-EP2-5-SC', оптимизированного относительно частоты кодона, и гена Δ9-элонгазы Isochrysis, в клетках дрожжей
Осуществляли ко-экспресссию гена Δ8-десатуразы 'ED3-8-EP2-5-SC', оптимизированного относительно частоты кодона, с последовательностью нуклеиновой кислоты Δ9-элонгазы, полученной из Isochrysis galbana (IsoD9) (№ доступа CQ831422, SEQ ID NO:31; Фигура 12). Синтезированную конструкцию гена IsoD9 создавали в GenScript (Piscataway, N.J.) и клонировали в вектор клонирования pUC57. Указанный ген субклонировали в сайты EcoRJ/BamHI дрожжевого вектора экспрессии pYX242 и полученную конструкцию обозначали как pIsoD9.
Конструкции pRSP62 (ED3-8-EP2-5-SC в pESC-Ura) и pIsoD9 ко-трансформировали в штамм SC334 Saccharomyces cerevisiae в соответствии с протоколом, описанным в Примере 3. Отбор ко-трансформантов осуществляли по признаку ауксотрофности как по отношению как к лейцину, так и к урацилу (среда DOB[-Leu-Ura]). Трансформированные колонии выращивали в течение ночи в 10 мл жидкой среды YPD при 30°C при энергичном перемешивании. После этого 5 мл ночной культуры добавляли в 45 селективной среды (DOB[-Leu-Ura] без декстрозы + 2% галактозы), содержащей 50 мкМ (итоговая концентрация) LA (18:2n-6) или ALA (18:3n-3) (в соответствии с указанным), и энергично перемешивали (250 об/мин) в течение 48-72 часов (в соответствии с указанным) при 24°C или 20°C. Выделение и анализ общего содержания жирных кислот проводили в соответствии с описанием, приведенным в Примере 3.
Δ8-десатураза и Δ9-элонгаза способны к совместному действию по химическому превращению LA (18:2n-6) в DGLA (20:3n-3). Δ9-элонгаза (IsoD9) преобразует LA до EDA (20:2n-6), а Δ8-десатураза (фермент, кодируемый ED3-8-EP2-5-SC) преобразует EDA в DGLA. Данный факт указывает на то, что Л8-десатураза, кодируемая ED3-8, выделенным из Emiliana huxleyi, способна функционировать в ходе альтернативного пути (Δ8-десатуразы/Δ9-элонгазы), результатом которого является биологический синтез арахидоновой кислоты (ARA) или EPA/DHA. Соответственно, указанные ферменты можно применять в сочетании с дополнительными десатуразами и элонгазами (то есть, с Δ5-десатуразой, С20-элонгазой и Δ4-десатуразой) для получения ARA из LA или EPA/DHA из ALA в различных организмах-хозяевах.
Пример 6
Экспрессия в растениях
Ген Δ8-десатуразы 'ED3-8-EP2-5-SC', оптимизированный относительно частоты кодона, ко-экспрессировали с геном Δ9-элонгазы, полученным из Isochrysis galbana (IsoD9) (№ доступа CQ831422, SEQ ID NO:31; Фигура 12), а также с геном Δ9-элонгазы, полученным из Euglenoid deses Ehr. CCMP2916 (EugMO7ELO, SEQ ID No:35) (более полное описание приведено в публикации патентной заявки США сер. №12/505,293, поданной 17 июля, 2009, включенной в настоящую заявку посредством ссылки, в соответствующем объеме) в экспериментальных масличных растениях Arabidopsis.
Кодирующую последовательность EugMO7ELO (SEQ ID NO:35) амплифицировали в ходе ПЦР из плазмиды, содержащей соответствующий ген, со смысловыми и анти-смысловыми олигонуклеотидами: 5′-TATAGAATTCAAATGGACGTCGCGACTACGCTG-3′ (SEQ ID NO. 36) и 5′-TATTCTCGAGTTCTAGTCCACTTTCTTCTCATCCTTC-3′ (SEQ ID NO 37) (внесенные последовательности распознавания фермента подчеркнуты). Реакцию ПЦР проводили с применением полимеразы Phusion высокой точности (New England Biolabs). После расщепления ферментами рестрикции EcoRI и XhoI, продукт присоединяли по его 5′-концу к промотору глицина-1 соевых, обладающему специфичностью в отношении семян, а по его 3′-концу - к 3′-нетранслируемому участку глицина-1 в составе двойного вектора pBinGlyRed2, с получением плазмиды pEugMO7ELO. Регуляторные элементы глицина-1 были описаны ранее (Nielsen, N.С.et al. (1989) Characterization of the glycinin gene family in soybean. Plant Cell, 1, 313-328). Указанный вектор также содержит трансген Ds-Red, находящийся под контролем промотора вируса мозаики маниока, для отбора трансформированных семян с применением флуоресценции, и марке устойчивости к канамицину для селекции бактерий. Указанный вектор также содержит несколько уникальных сайтов расщепления ферментами (например, MluI), позволяющие осуществлять клонирование других трансгенов для экспрессии нескольких генов в растении-хозяине. В качестве контроля, в указанных экспериментах клонировали Δ9-ELO Isochrysis galbana (SEQ ID NO:31) как фрагмент EcoRI/XhoI, под контролем помотора глицина-1 в pBinGlyRed2 для создания плазмиды pIsoD9.
Кодирующую последовательность ED3-8-EP2-5-SC (SEQ ID NO:30) синтезировали с сайтами расщепления NotI, фланкировавшими открытую рамку считывания. Кодирующую последовательность ED3-8-EP2-5-SC клонировали во фрагмент NotI в соответствующих сайтах вектора pBCon, содержащего промотор, обладающий специфичностью в отношении семян, к α′-субъединице гена β-конглицинина соевых, и 3′-нетранслируемый участок гена фазеолина Phaseolus vulgaris. ED3-8-EP2-5-SC фланкировали на 5′-конце с помощью последовательности промотора β-конглицинина, и на 3′-конце - с помощью 3′-нетранслируемого участка фазеолина. Образованную кассету, содержавшую промотор, кодирующую последовательность ED3-8-EP2-5-SC и 3′-нетранслируемый участок, удаляли из состава вектора pBCon с помощью сайтов расщепления AscI, фланкировавших кассету. После этого кассету AscI клонировали в совместимый сайт MluI в составе pEugELO для создания плазмиды pEugMO7ELO-'ED3-8-EP2-5-SC', содержащей трансгены для ко-экспресии EugMO7ELO и десатуразы ED3-8-EP2-5-SC, обладающей специфичностью в отношении семян. Кассету AscI, содержавшую трансген для десатуразы ED3-8-EP2-5-SC, обладающий специфичностью в отношении семян, также клонировали в сайт MluI в составе pIsoD9 для создания pIsoD9-'ED3-8-EP2-5-SC'.
pEugMO7ELO-'ED3-8-EP2-5-SC' и pIsoD9-'ED3-8-EP2-5-SC' вводили в штамм Agrobacterium tumefaciens C58MP90 с помощью электропорации. После чего, агробактерии, обладавшие устойчивостью к канамицину, применяли для трансформации экотипа Col-0 Arabidopsis thaliana с помощью метода погружения цветка (Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method for Agrobacterium - mediated transformation of Arabidopsis thaliana. Plant J, 16, 735-743). После трансформации методом погружения цветка с применением агробактерии, растения культивировали при 22°C в течение 16 дней до достижения зрелости и высыхания. В ходе указанных экспериментов применяли fad3/fae1 мутант Arabidopsis, содержащий низкие уровни α-линолевой кислоты и очень длинные цепи жирных кислот (>C20), и при этом повышенные уровни линолевой кислоты, в масле семян (Cahoon, Е.В. et al (2006) Conjugated fatty acids accumulate to high levels in phospholipids of metabolically engineered soybean and Arabidopsis seeds. Phytochemistry, 67, 1166-1176). Указанная генетическая подоплека сближает характер составляющей жирных кислот масел семян, полученных из таких зерновых, как сафлор, и соевых с низким содержанием линолевой кислоты. Трансгенные семена, полученные от растений Arabidopsis, трансформированных с помощью Agrobacterium, определяли с помощью метода флуоресценции маркерного белка DsRed с применением известной методики (Pidkowich, M.S. et al (2007) Modulating seed beta-ketoacyl-acyl carrier protein synthase II level converts the composition of a temperate seed oil to that of a palm-like tropical oil. Proc Natl Acad Sci USA, 104, 4742-4747). Простые трансгенные и не трансгенные семена, применявшиеся в качестве контроля, подвергали прямой трансэтерификации содержащихся липидов, включая триацилглицеролы, путем применения реагента гидроксид атриметилсульфония (TMSH) (Cahoon, E.B. and Shanklin, J. (2000) Substrate-dependent mutant complementation to select fatty acid desaturase variants for metabolic engineering of plant seed oils. Proc Natl Acad Sci USA, 97, 12350-12355). Метиловые эфиры жирных кислот, полученные из простых семян, анализировали с помощью газовой хроматографии с детекцией ионизации в пламени с применением газового хроматографа Agilent 7890, снабженного колонкой INNOWax (длина 30 м × внутренний диаметр 0.25 мм), и при температуре в термостате, установленной со 185°C (выдерживание в течение 1 мин) до 230°C (выдерживание в течение 2 мин) при 7°C/мин. Метиловые эфиры входящих в состав жирных кислот определяли на основе их времени удерживания по сравнению с метиловыми эфирами известных жирных кислот из семян Arabidopsis thaliana Col-0 дикого типа и путем сравнения времени удерживания с показателями для метиловых эфиров жирных кислот, применявшихся в качестве стандартов.
В Таблице 8 приведены сочетания жирных кислот в простых семенах T1, полученных в результате пяти независимых событий трансформации растений, трансформированных pEugMO7ELO-'ED3-8-EP2-5-SC'. Также приведены сочетания жирных кислот в простых семенах T1, полученные в результате независимых событий трансформации растений pIsoD9-'ED3-8-EP2-5-SC' (Таблица 9). В семенах трансформантов pEugMO7ELO-'ED3-8-EP2-5-SC', в которых осуществляется ко-экспрессия элонгазы Euglena-MO7 и Δ8-десатуразы ED3-8, накапливалась преимущественно Δ8,11,14-эйкозатриеновая кислота (20:3Δ8,11,14) и меньшие количества Δ11,14-эйкозадиеновой кислоты. В указанных семенах, наиболее распространенной жирной кислотой являлась 20:3Δ8,11,14, и ее относительное количественное содержание варьировало в интервале от 28% до 37% от общего содержания жирных кислот. Относительное количественное содержание 20:2Δ11,14 в указанных семенах варьировало в интервале от 10.8% до 14.5% от общего содержания жирных кислот. Для сравнения, 20:3Δ8,11,14 и 20:2Δ11,4 не были определены или найдены даже в следовых количествах в семенах не трансформированных растений Arabidopsis fad3/fae1 (Таблица 10). В семенах, в которых осуществлялась ко-экспрессия Isochrysis galbana 49 ELO и Δ8-десатуразы ED3-8, содержание 20:3Δ8,11,14 составляло до 37% от общего содержания жирных кислот, а содержание 20:2Δ11,14 составляло до 6% от общего содержания жирных кислот (Таблица 9). В общем, указанный результат доказывает, что семена, богатые 18:2, в которых осуществляется ко-экспрессия элонгазы Euglena-MO7 и Δ8-десатуразы ED3-8 или 49 ELO Isochrysis galbana и Δ8-десатуразы ED3-8, способны к образованию значительных количеств 20:3Δ8,11,14, представляющей собой промежуточный продукт и предшественник в ходе биологического синтеза арахидоновой кислоты (20:4Δ5,8,11,14). Приведенные результаты также отражают эффективность ко-экспрессии 18:2-специфичной элонгазы, например, EugMO7ELO, и 20:2 Δ8-десатуразы, например, Δ8-десатуразы ED3-8, или осуществления важных этапов конструирования при проризводстве ARA или EPA/DHA, в семенах масличных растений. Целесообразной является ко-экспрессия указанных генов с d5-десатуразой, для достижения итоговой цели, заключающейся в производстве масел ARA или EPA в трансгенных семенах масличных растений.
Изобретение, наглядно описанное в настоящей заявке, можно успешно применять в отсутствие любого элемента или элементов, при любом ограничении или ограничениях, что не указано отдельно в настоящей заявке. Термины и выражения, использованные для описания, не являются ограничивающими, и их значение не исключает любые эквиваленты приведенных и описанных черт или их частей. Следует понимать, что в объем настоящего изобретения входят различные модификации, согласно формуле настоящего изобретения. Следовательно, следует понимать, что, хотя настоящее изобретение было описано, в частности, в предпочтительных вариантах его реализации, специалист в данной области техники может прибегать к внесению дополнительных признаков, модификаций и созданию вариаций понятий, описанных в настоящей заявке, и указанные вариации рассматриваются как входящие в объем настоящего изобретения, в соответствии с определенным в прилагаемой формуле изобретения.































Изобретение относится к биохимии и представляет собой выделенные полинуклеотиды, кодирующие Δ8-десатуразу. Изобретение также относится к самим Δ8-десатуразам, кодируемым выделенными полинуклеотидами, векторам экспрессии, содержащим выделенные полинуклеотиды, клеткам-хозяевам, содержащим векторы экспрессии, и к способам получения Δ8-десатураз и полиненасыщенных жирных кислот, выбранных из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ЕТА) и любых их сочетаний. Изобретение позволяет расширить ассортимент Δ8-десатураз, используемых для получения полиненасыщенных жирных кислот. 10 н. и 2 з.п. ф-лы, 12 ил., 14 табл., 6 пр.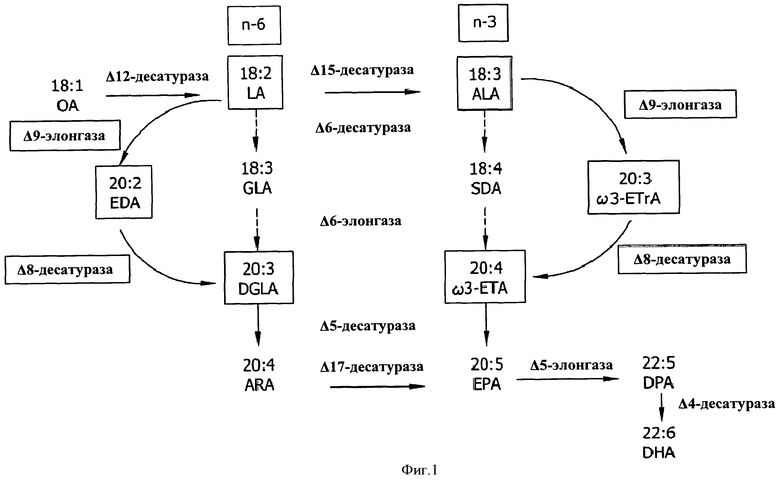
1. Выделенная нуклеиновая кислота, содержащая нуклеотидную последовательность, кодирующую полипептид, обладающий активностью Δ8-десатуразы, причем аминокислотная последовательность полипептида содержит аминокислотную последовательность SEQ ID NO:29.
2. Выделенная нуклеиновая кислота, содержащая нуклеотидную последовательность SEQ ID NO:28 или содержащая по меньшей мере 90% нуклеотидной последовательности SEQ ID NO:30, причем указанная нуклеиновая кислота кодирует полипептид, обладающий активностью Δ8-десатуразы.
3. Выделенная нуклеиновая кислота по п.2, отличающаяся тем, что указанная последовательность кодирует функционально активный фермент Δ8-десатуразу, использующий в качестве субстрата ω6-эйкозадиеновую кислоту или ω3-эйкозатриеновую кислоту.
4. Вектор экспрессии, включающий:
нуклеотидную последовательность, функционально связанную с регуляторной последовательностью, где нуклеотидная последовательность содержит SEQ ID NO:28 или содержит по меньшей мере 90% нуклеотидной последовательности SEQ ID NO:30, причем указанная нуклеиновая кислота кодирует полипептид, обладающий активностью Δ8-десатуразы.
5. Эукариотическая клетка для продуцирования полиненасыщенных жирных кислот, выбранных из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ETA) и любых их сочетаний, содержащая вектор по п.4.
6. Эукариотическая клетка по п.5, выбранная из группы, состоящей из клетки млекопитающего, клетки насекомого, клетки растения или клетки гриба.
7. Растение для продуцирования полиненасыщенных жирных кислот, выбранных из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ЕТА) и любых их сочетаний, содержащее вектор по п.4, причем в результате экспрессии нуклеотидной последовательности вектора в растении образуется по меньшей мере одна полиненасыщенная жирная кислота.
8. Очищенный полипептид, обладающий активностью Δ8-десатуразы, кодируемый нуклеиновой кислотой, содержащей нуклеотидную последовательность SEQ ID NO:28 или содержащей по меньшей мере 90% нуклеотидной последовательности SEQ ID NO:30.
9. Очищенный полипептид, обладающий активностью Δ8-десатуразы, десатурирующий субстрат в виде полиненасыщенной жирной кислоты с 20 атомами углерода в цепи (C20-PUFA) между 8 и 9 атомами углерода субстрата, причем указанный полипептид содержит аминокислотную последовательность SEQ ID NO:29.
10. Способ получения полипептида, обладающего активностью Δ8-десатуразы, включающий этапы:
a) выделения нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:28 или содержащую по меньшей мере 90% нуклеотидной последовательности SEQ ID NO:30, причем указанная нуклеиновая кислота кодирует полипептид, обладающий активностью Δ8-десатуразы,
b) конструирования вектора экспрессии, содержащего выделенную нуклеиновую кислоту согласно этапу a), функционально связанную с регуляторной последовательностью; и
c) введения вектора экспрессии в эукариотическую клетку на период времени и в условиях, достаточных для образования полипептида, обладающего активностью Δ8-десатуразы.
11. Способ получения полиненасыщенной жирной кислоты, выбранной из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ETA) и любых их сочетаний, включающий этапы:
a) выделения нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:28 или содержащую по меньшей мере 90% нуклеотидной последовательности SEQ ID NO:30, причем указанная нуклеиновая кислота кодирует полипептид, обладающий активностью Δ8-десатуразы;
b) конструирования вектора экспрессии, содержащего выделенную нуклеиновую кислоту согласно этапу a), функционально связанную с регуляторной последовательностью;
c) введения вектора экспрессии в эукариотическую клетку на период времени и в условиях, достаточных для образования полипептида, обладающего активностью Δ8-десатуразы; и
d) приведения во взаимодействие экспрессированного полипептида, обладающего активностью Δ8-десатуразы, с субстратом, выбранным из группы, состоящей из ω6-эйкозадиеновой кислоты, ω3-эйкозатриеновой кислоты и комбинаций указанных кислот, для превращения субстрата в конечную полиненасыщенную жирную кислоту, выбранную из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ETA) и любых их сочетаний.
12. Способ получения полиненасыщенной жирной кислоты, выбранной из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ETA) и любых их сочетаний в эукариотической клетке, включающий этапы:
a) выделения нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:28 или содержащую по меньшей мере 90% нуклеотидной последовательности SEQ ID NO:30, причем указанная нуклеиновая кислота кодирует полипептид, обладающий активностью Δ8-десатуразы;
b) конструирования вектора экспрессии, содержащего выделенную нуклеиновую кислоту согласно этапу a), функционально связанную с регуляторной последовательностью;
c) введения вектора экспрессии согласно этапу b) и по меньшей мере одной дополнительной рекомбинантной ДНК конструкции, содержащей выделенную нуклеотидную последовательность, функционально связанную с по меньшей мере одной регуляторной последовательностью, кодирующей дельта-9 элонгазу, в эукариотическую клетку;
d) приведения во взаимодействие экспрессированного полипептида, обладающего активностью Δ8-десатуразы и дельта-9 элонгазы с субстратами, выбранными из группы, состоящей из линолевой кислоты (LA), альфа-линоленовой кислоты (ALA) и комбинаций указанных кислот, для превращения субстрата в конечную полиненасыщенную жирную кислоту, выбранную из группы, состоящей из дигомо-гамма-линоленовой кислоты (DGLA), ω3-эйкозатетраеновой кислоты (ω3-ETA) и любых их сочетаний.
| WO 2007127381 A2, 08.11.2007 | |||
| US 20080138868 A1,12.06.2008 | |||
| ZHOU XR et.al.Isolation and characterization of genes from the marine microalga Pavlova salina encoding three front-end desaturases involved in docosahexaenoic acid biosynthesis, Phytochemistry | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| ГЕН ДЕСАТУРАЗЫ КИСЛОТЫ ЖИРНОГО РЯДА ИЗ РАСТЕНИЙ | 2000 |
|
RU2274657C2 |

