Настоящее изобретение относится к экспрессирующему вектору для секреторного продуцирования человеческого интерферона альфа (hIFNα), содержащему полинуклеотид, кодирующий модифицированную сигнальную последовательность термостабильного энтеротоксина II E.coli, и hIFNα - кодирующий полинуклеотид, лигированный с его 3'-концом; к микроорганизму, трансформированному указанным экспрессирующим вектором; и к способу продуцирования hIFNα, не содержащего дополнительного метионинового остатка на своем N-конце и секретирующегося в периплазму клетки E.coli.
Предшествующий уровень техники
В 1957 году Иссак и Линденман сообщали, что у кур, инфицированных вирусом гриппа А, продуцировался фактор ингибирования репликации вируса, названный интерфероном (Issacs, К. & Lindenmann, J.Proc.R.Soc.Lond., В 147:258-267, 1957).
Человеческие интерфероны представляют собой белки цитокины, ингибирующие иммунный ответ in vivo или репликацию вируса, и классифицируются как интерферон альфа (IFNα), интерферон бета (IFNβ) и интерферон гамма (IFNγ) по типу продуцирующих их клеток (Kirchner, H. et al., Tex.Rep.Biol.Med., 41:89-93, 1981; Stanton, G. J. et al., Тех. Rep.Biol.Med. 41:84-88, 1981).
Хорошо известно, что указанные интерфероны, действуя вместе, обладают синергическим эффектом, который проявляется в антивирусной активности, противораковой активности, активации NK (клеток-киллеров) и в активности, направленной на ингибирование пролиферации клеток костного мозга (Klimpel, et al., J.Immunol. 129:76-78, 1982; Fleischmann, W.R. et al., J.Natl. Cancer. Inst. 65:863-966, 1980; Weigent, et al., Infect. Immun. 40:35-38, 1980). Кроме того, интерфероны действуют как факторы регуляции экспрессии, структуры и функции генов в данной клетке и обладают прямым антипролиферирующим действием.
IFNα продуцируются в случае, когда лейкоцит стимулируется В-клеточным митогеном, вирусом или раковыми клетками. В настоящее время известны гены, кодирующие более 20 видов интерферонов, каждый из которых включает 165 или 166 аминокислот.
IFNα, используемый для ранних клинических тестов, получают из лейкоцитарной пленки, стимулированной вирусом Сендай, и его чистота составляет лишь менее 1% (Cantell, К & Hirvonen, Тех. Rep. Biol. Med. 35:138-144, 1997).
В 1980 г. с помощью техники рекомбинантных ДНК стало возможным продуцировать большое количество IFNa, обладающего биофизической активностью (Goedell, D.V. et al., Nature, 287:411-416, 1980). Клинические тесты с использованием рекомбинантного hIFNα показали, что он является эффективным для лечения различных твердых раковых опухолей, а в частности рака мочевого пузыря, рака почек, ВИЧ-ассоциированной саркомы Капоши и т.п.(Torti, F.M. J.Clin. Oncol. 6:476-483, 1988; Vugrin, D. et al., Cancer. Treat. Rep. 69:817-820, 1985; Rios, A. et al., J. Clin. Oncol. 3:506-512, 1985). Он также является эффективным для лечения вируса гепатита С (Davis, G.G. et al., N.Engl. J.Med., 321:1501-1506, 1989) и границы его применения в качестве терапевтического агента расширяются день ото дня.
Результаты клонирования гена IFNα из лейкоцита показали, что IFNα кодируется группой, по крайней мере, из 10 различных генов. Это указывает на то, что ДНК-последовательности этих генов продуцируют не один вид белка, и что IFNα представляет собой смесь белков определенного подтипа, имеющих сходные структуры. Белки такого подтипа были названы IFNα-1, 2, 3 и т.п. (Nature, 290:20-26, 1981).
Среди нескольких типов интерферонов hIFNα, выделенный из человеческих лейкоцитов, имеет молекулярную массу 17500-21000 и очень высокую природную активность, составляющую примерно 2 × 108 ед/мг белка. Jn vivo, IFNα представляет собой белок, состоящий из 165 аминокислот. В случае, если 23-й аминокислотой является лизин, то такой интерферон обозначается IFNα-2a (SEQ ID No:1), а в случае, если 23-й аминокислотой является аргинин, то он обозначается IFNα-2b (SEQ ID No:2). Сначала hIFNα был продуцирован методом с использованием клеточной культуры. Однако этот метод оказался неподходящим для промышленного продуцирования из-за его низкой продуктивности, которая составляет примерно 250 мкг/л.
Для решения этой проблемы были разработаны способы выделения большого количества интерферона из микроорганизмов с использованием техники рекомбинантных ДНК, и эти способы используются до настоящего времени.
Наиболее широкое применение имеет способ с использованием E.coli, которая в соответствии со свойствами клеток E.coli продуцирует IFNα, состоящий из 166 или 167 аминокислот. Эти продукты содержат дополнительный метиониновый остаток, добавленный у N-конца благодаря кодону ATG, присутствующему в сайте инициирующего кодона. Однако сообщалось, что в случае человеческого гормона роста этот дополнительный метиониновый остаток может стимулировать нежелательный иммунный ответ (публикация патента ЕР № 256843).
Кроме того, большинство экспрессированных IFNα аккумулируется в цитоплазме в форме нерастворимых телец включения и должно быть превращено в активную форму в результате повторной укладки в процессе очистки. Поскольку этот способ повторной укладки является неэффективным, IFNα присутствует частично в восстановленной форме или образует межмолекулярное дисульфид-связывающее тельце или дефектное дисульфид-связывающее тельце. Удаление побочных продуктов, которые приводят к заметно низкому выходу, представляет определенные трудности. В частности, чрезвычайно трудно удалить такие нежелательные побочные продукты интерферона, как интерфероны с неправильной укладкой.
Недавно для решения вышеупомянутых проблем, связанных с продуцированием чужеродного белка в микробной клетке, был предпринят ряд попыток разработать способ, основанный на эффективной секреции растворимой формы целевого белка, не имеющего у своего N-конца дополнительного метионина.
В этом способе нужный белок экспрессируется в форме гибридного белка, несущего сигнальный пептид, присоединенный к его N-концу. При прохождении указанного гибридного белка через клеточную мембрану сигнальный пептид удаляется ферментом в E.coli, и нужный белок секретируется в нативной форме.
Способ секреторного продуцирования является более предпочтительным, чем способ микробного продуцирования, поскольку аминокислотная последовательность и высшая структура продуцированного белка, обычно, идентичны указанным последовательности и структуре белка дикого типа. Однако выход, который дает способ секреторного продуцирования, в большинстве случаев является очень низким из-за недостаточной эффективности как мембранного транспорта, так и последующей очистки. Это соответствует хорошо известному факту, что выход белка млекопитающего, продуцированного секреторным способом в прокариотах, гораздо ниже, чем выход прокариотического белка, продуцированного тем же способом в прокариотах. Поэтому были предприняты попытки разработать более эффективный способ секреторного продуцирования. Так, например, в публикации патента Кореи № 93/1387 описаны попытки крупномасштабного продуцирования IFNα с использованием сигнального пептида щелочной фосфатазы E.coli, однако его выход в культуральной среде при 109 МЕ/л (10 мкг/л культуральной среды) был очень низким. Поэтому разработка способа продуцирования растворимого IFNα, не содержащего на своем N-конце дополнительного метионинового остатка, с использованием микроорганизма для крупномасштабного производства, представляет огромный интерес.
Ранее авторами настоящего изобретения был продуцирован новый сигнальный пептид термостабильного энтеротоксина II E.coli (заявки на патент Кореи № 98-38061 и 99-27418) и было обнаружено, что этот новый секреторный сигнальный пептид может быть использован для крупномасштабного продуцирования природной формы IFNα. А именно авторами настоящего изобретения был сконструирован экспрессирующий вектор, содержащий ген, полученный путем лигирования IFNα-кодирующего гена, вместо гена, кодирующего энтеротоксин II, с модифицированным секреторным сигнальным пептидом E.coli, и был разработан способ секреторного продуцирования IFNα, обладающего природной биологической активностью, предусматривающий использование микробной секреторной системы путем культивирования микроорганизма, трансформированного указанным экспрессирующим вектором.
Краткое описание изобретения
В соответствии с этим целью настоящего изобретения является получение экспрессирующего вектора, который способен к секреторному продуцированию человеческого интерферона альфа (hIFNα).
Другой целью настоящего изобретения является получение микроорганизма, трансформированного указанным экспрессирующим вектором.
Еще одной целью настоящего изобретения является способ продуцирования растворимой формы hIFNα с использованием указанного микроорганизма, который не содержит дополнительного метионинового остатка, присоединенного к амино-концу.
Краткое описание графического материала
Вышеуказанные и другие цели и признаки настоящего изобретения будут очевидны из нижеследующего описания настоящего изобретения и прилагаемого графического материала, на котором соответственно проиллюстрированы:
Фиг.1: процедура конструирования вектора pT-IFNα-2a;
Фиг.2: процедура конструирования вектора pT14SIα-2a;
Фиг.3: процедура конструирования вектора pT14SSIα-2a;
Фиг.4: процедура конструирования вектора pT14OSSIα-2a-4T22Q;
Фиг.5а и 5b: результаты электрофореза в ДСН-ПААГ, которые подтверждают экспрессию IFNα-2a и чистоту IFNα-2a, экспрессированного из рекомбинантных клеточных линий, и результат Вестерн-блот-анализа, который подтверждает молекулярную массу экспрессированного IFNα-2b соответственно.
Подробное описание настоящего изобретения
В соответствии с одним из своих аспектов настоящее изобретение относится к экспрессирующему вектору для секреторного продуцирования hIFNα, содержащего полинуклеотид, кодирующий модифицированную сигнальную последовательность термостабильного энтеротоксина II (далее называемой “мутантом STII”) и полинуклеотид, кодирующий hIFNα, лигированный с его 3'-концом.
Полинуклеотидом, кодирующим hIFNα и используемым для конструирования экспрессирующего вектора настоящего изобретения, может быть любой из полинуклеотидов, кодирующих произвольные подтипы hIFNα, такие как природный hIFNα-2a (SEQ ID No:1), IFNα-2b (SEQ ID No:2), IFNα-1 и IFNα-3, и им может также быть рекомбинантный полипептид, который имеет модифицированную последовательность оснований, кодирующую любой из вышеуказанных подтипов IFNα.
Полинуклеотидом, кодирующим модифицированную сигнальную последовательность термостабильного энтеротоксина II E.coli настоящего изобретения, лигированную перед 5'-концом полинуклеотида, кодирующего hIFNα и используемого для секреторного продуцирования hIFNα, может быть полинуклеотид, кодирующий мутант, полученный путем замены одной или нескольких аминокислот сигнальной последовательности термостабильного энтеротоксина II E.coli, описанной в SEQ ID No:3, а предпочтительно одной или более из ее 4-й, 20-й и 22-й аминокислот другой(ими) аминокислотой(ами). Примерами таких полинуклеотидов являются полинуклеотиды, кодирующие мутанты, полученные путем следующих замен: 4-й аминокислоты треонином ([Thr4]STII); 4-й аминокислоты треонином и 22-й аминокислоты глутамином соответственно ([Thr4, Gln22]STII); 4-й аминокислоты треонином, 20-й аминокислоты валином и 22-й аминокислоты глутамином соответственно ([Thr4, Val20, Gln22]STII); и 4-й аминокислоты треонином и 20-й аминокислоты валином соответственно ([Thr4, Val20]STII) в сигнальной последовательности термостабильного энтеротоксина II E.coli (STII), описанной в SEQ ID No:3, а предпочтительными полинуклеотидными последовательностями являются SEQ ID No: 4, 5, 6 и 7. Однако известно, что вследствие вырожденности кодона могут существовать и некоторые другие полинуклеотиды, кодирующие мутанты настоящего изобретения, а в частности полинуклеотид, модифицированный путем введения предпочтительных кодонов Е.coli без каких-либо замен в аминокислотной последовательности, может быть использован для стимуляции скорости экспрессии IFNα.
Кроме того, экспрессирующий вектор настоящего изобретения может также включать последовательность Шайна-Дальгарно термостабильного энтеротоксина II E.coli (последовательность SD, SEQ ID No:8) или ее мутантную последовательность, лигированную перед 5'-концом полинуклеотида, кодирующего модифицированную сигнальную последовательность термостабильного энтеротоксина II. По сравнению с последовательностью дикого типа, которая имеет 7 оснований (TGATTTT), следующих за GAGG на 5'-конце последовательности SD термостабильного энтеротоксина II в E.coli, представленной в SEQ ID No:8, мутантная последовательность SD имеет укороченную последовательность из 6 или 5 оснований. Использование этого мутанта может приводить к увеличению скорости секреторной экспрессии IFNα. Однако, если указанная последовательность оснований становится короче на 4 основания, то скорость экспрессии заметно снижается. Конкретным примером предпочтительного мутанта, который может быть использован в настоящем изобретении, является мутантная последовательность SD термостабильного энтеротоксина II E.coli, имеющая нуклеотидную последовательность SEQ ID No:9.
Промотором, используемым для получения экспрессирующего вектора настоящего изобретения, может быть любой из промоторов, который способен экспрессировать гетерологичный белок в микроорганизме-хозяине. В частности, если гетерологичный белок экспрессируется в E.coli, то предпочтительным промотором является промотор lac, Tac и арабинозы.
Настоящее изобретение также относится к трансформированным микроорганизмам, которые могут быть получены путем трансформации таких штаммов E.coli, как BL21 (DЕ3) E.coli (Novagen, USA) или XL-1 blue E.coli (Novagen, USA) с использованием указанного экспрессируищего вектора. Примерами настоящего изобретения являются такие трансформированные микроорганизмы, как BL21(DE3)/pT14OSSIα-2а-4Т (“НМ 10603”) E.coli, BL21(DE3)/ pT14OSSIα-2a-4T22Q (“HM 10611”) E.coli, BL21 (DЕ3) /pT14OSSIα-2b-4T (“HM 10703”) E.coli, BL21(DЕ3)/pT14OSSIα-2b-4T22Q (“HM 10711”) E.coli.
Вышеуказанные трансформированные микроорганизмы были депонированы в Корейском центре культур микроорганизмов (КССМ) (Адрес: Yurium Bidg., 361-221, Hongje 1-dong, Seodaemun-gu, Seoul 120-091, Republic of Korea) 23 декабря 1999 г. под номерами доступа КССМ-10175, КССМ-10176, КССМ-10177 и КССМ-10178 соответственно, в рамках Будапештского договора, заключенного в соответствии с Международным соглашением о депонировании микроорганизмов для проведения патентной экспертизы.
В соответствии с другим своим аспектом настоящее изобретение также относится к способу продуцирования hIFNα, не содержащего дополнительного метионинового остатка у своего N-конца, путем его секреции в периплазму E.coli, где указанный способ предусматривает культивирование трансформированного микроорганизма в подходящих условиях культивирования, которые могут быть аналогичными стандартным условиям культивирования, используемым для трансформированных микроорганизмов.
hIFNα, продуцируемый секреторным способом настоящего изобретения, включает произвольные подтипы hIFNα, такие как IFNα-1, IFNα-3 и т.п., а также природные подтипы hIFNα-2a (SEQ ID No:1) и hIFNα-2b (SEQ ID No:2), состоящие из 165 аминокислот. Кроме того, способ настоящего изобретения может быть также использован для продуцирования любого другого интерферона, такого как hIFNβ и hIFNγ.
В соответствии со способом настоящего изобретения 80% или более IFNα, продуцируемого трансформантом E.coli настоящего изобретения, секретируется в периплазму с высокой степенью продуктивности, составляющей более 1 г/л. Продуцированный IFNa имеет ту же аминокислотную последовательность, что и природный IFNα, у N-конца которого отсутствует дополнительная аминокислота, присоединенная к его N-концу, и который обнаруживает биологическую активность, аналогичную активности природного IFNα.
Настоящее изобретение подробно проиллюстрировано нижеследующими примерами, которые не ограничивают его объема. Сравнительный пример: ген IFNα-2a и конструирование вектора, содержащего этот ген.
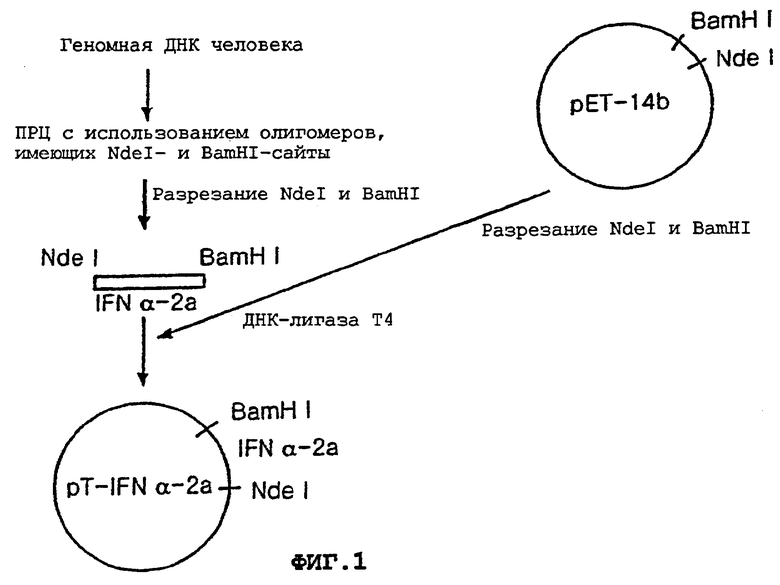
Ген, кодирующий hIFNα-2a, получали путем проведения ПЦР с использованием геномной ДНК человека в качестве матрицы и SEQ ID No: 10 и 11 в качестве праймеров. Праймер SEQ ID No:10 был сконструирован для получения рестрикционного NdeI-сайта (5'-CATATG-3'), расположенного выше от кодона, кодирующего первую аминокислоту (цистеин) природного hIFNα, а праймер SEQ ID No:11 был сконструирован для получения рестрикционного BamHI-сайта (5'-GGATCC-3'), расположенного ниже от кодона терминации природного hIFNα.
Амплифицированный ПЦР-продукт расщепляли ферментами NdeI и BamHI с получением ДНК-фрагмента, кодирующего hIFNα-2a. Этот ДНК-фрагмент встраивали в NdeI/BamHI-сайт вектора рЕТ-14b (Novagen, USA) с получением вектора pT-IFNα-2a.
На фиг.1 проиллюстрирована вышеописанная процедура конструирования вектора pT-IFNα-2a.
Сравнительный пример 1: Конструирование вектора, содержащего сигнальную последовательность энтеротоксина и гены IFNα-2a
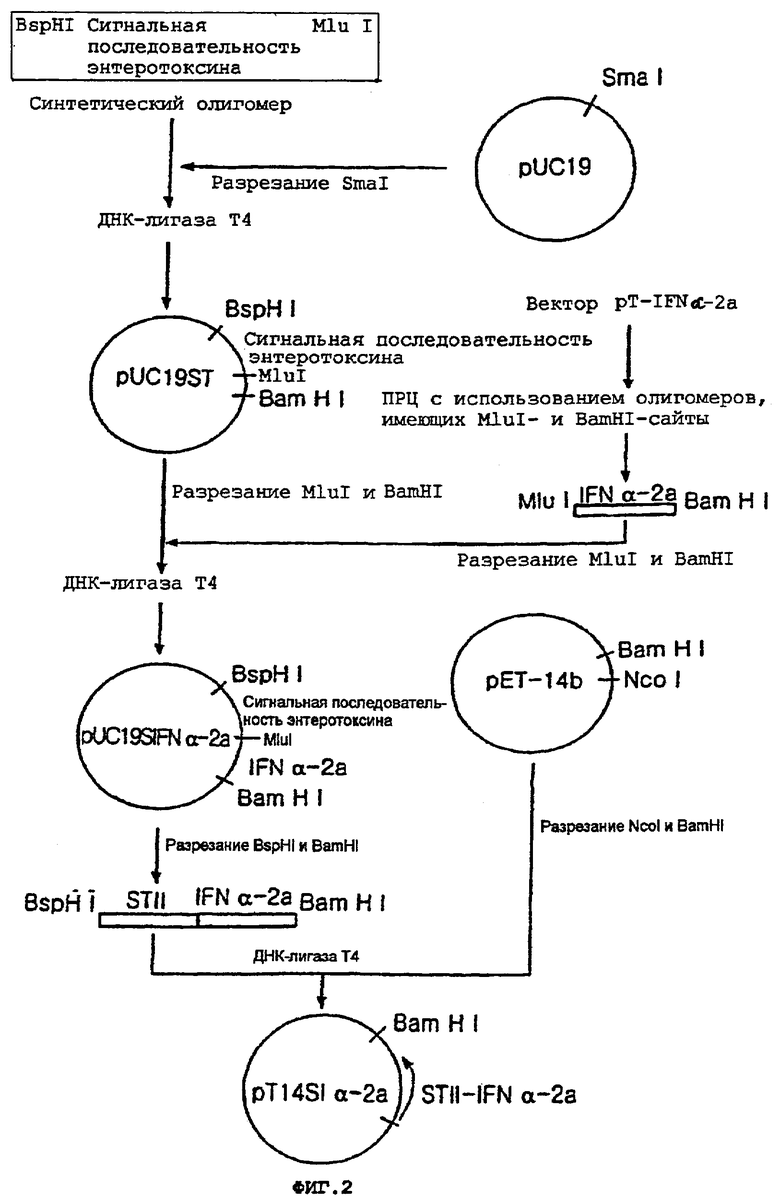
Для получения гена сигнальной последовательности энтеротоксина II E.coli, пару комплементарных олигонуклеотидов SEQ ID No:12 и 13 конструировали на основе ранее известной нуклеотидной последовательности сигнального пептида энтеротоксина II E.coli, и синтезировали с использованием ДНК-синтезатора (Model 380B, Applied Biosystem, USA). Вышеуказанные олигонуклеотиды конструировали для встраивания рестрикционного BspHI-сайта (сайта, комплементарного рестрикционному NdeI-сайту) выше кодона инициации энтеротоксина II E.coli и рестрикционного MluI-сайта, введенного посредством молчащей замены в другой конец. Оба олигонуклеотида отжигали при 95°С с получением затупленного по концам ДНК-фрагмента, имеющего нуклеотидную последовательность, кодирующую сигнальную последовательность энтеротоксина II E.coli. Вышеуказанный ДНК-фрагмент встраивали в SmaI-сайт вектора pUC19 (BioLabs, USA) с получением вектора pUC19ST.
Кроме того, вектор pT-IFNα-2a, содержащий ген IFNα-2a, полученный в сравнительном примере, подвергали ПЦР с использованием праймеров SEQ ID No:14 и 15 для лигирования сигнального пептида энтеротоксина с геном IFNα-2a. Праймер SEQ ID No:14 конструировали так, чтобы он соответствовал 5'-концу гена IFNα-2a, а праймер SEQ ID No: 15 конструировали так, чтобы рестрикционный BamHI-сайт (5'-GGATCC-3') был введен ниже кодона терминации гена. ДНК-фрагмент, содержащий указанный полинуклеотид, кодирующий природный IFNα-2a, амплифицировали посредством ПЦР с использованием вышеуказанных полинуклеотидных праймеров. Амплифицированный ДНК-фрагмент расщепляли ферментами MluI и BamHI с получением ДНК-фрагмента IFNα-2a, имеющего MluI/BamHI-концы.
При этом вектор pUC19ST, содержащий сигнальный пептид энтеротоксина, расщепляли ферментом MluI, а затем гидролизовали BamHI с получением фрагмента вектора, имеющего MluI/BamHI-концы. Указанный фрагмент вектора лигировали с ДНК-фрагментом IFNa-2a, в результате чего получали вектор pUC19SIFNα-2a.
Вектор pUC19SIFNα-2a расщепляли ферментами BspHI и BamHI с получением ДНК-фрагмента (564 п.н.). ДНК-фрагмент встраивали в NcoI/BamHI-участок вектора pET-14b (Novagen, USA) с получением вектора pT14SIα-2a.
На фиг.2 проиллюстрирована описанная выше процедура конструирования вектора pT14SIα-2a.
Затем штамм BL21(DE3) E.coli обрабатывали 70 мМ раствором хлорида кальция с получением компетентной E.coli, a затем в него добавляли вектор рТ14Iα-2а в 10 мМ Трис-буфере (рН 7,5). Трансформант E.coli, экспрессирующий IFNα-2a, отбирали стандартным методом, основанным на чувствительности трансформированного вектора к антибиотикам, и обозначали НМ 10600 E.coli.
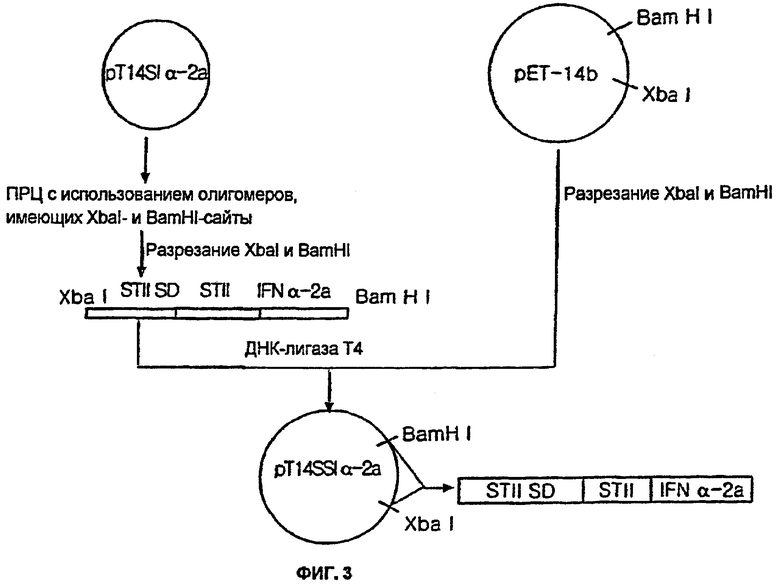
Кроме того, вектор pT14SIα-2a подвергали ПЦР с использованием праймеров SEQ ID No: 16 и 17 для амплификации ДНК-фрагмента, полученного посредством последовательного лигирования последовательности Шайна-Дальгарно энтеротоксина; сигнального пептида энтеротоксина; и гена IFNα-2a, а затем ДНК-фрагмент расщепляли ферментами XbaI и BamHI с получением вставки.
Полученную вставку лигировали в XbaI/BamHI-участок вектора pET-14b (Novagen, USA) для конструирования вектора pT14SSIα-2a.
На фиг.3 проиллюстрирована вышеописанная процедура конструирования вектора pT13SSIα-2a. Штамм BL21(DE3) E.coli (Stratagene, USA) трансформировали вектором pT14SSIα-2a с получением трансформанта, обозначенного НМ 10601 E.coli.
Сравнительный пример 2: Конструирование вектора, содержащего сигнальную последовательность энтеротоксина и гены IFNα-2b
23-й лизиновый кодон гена IFNα-2a в векторе pT14SSIα-2a заменяли кодоном аргинина посредством сайт-направленного мутагенеза (Papworth, С et al., Strategies, 9, 3, 1996) в целях конструирования экспрессирующего вектора, содержащего ген IFNα-2b. Вектор pT14SSIα-2a подвергали гибридизации с синтетическими олигонуклеотидами SEQ ID No:19 и 20, содержащими замененный кодон, с образованием гибридной молекулы и осуществляли амплификацию ДНК с использованием pfu-ДНК-полимеразы (Stratagene, USA) и четырех нуклеотидтрифосфатов (АТР, GTP, ТТР, СТР), которые удлиняют указанные олигонуклеотиды в направлении 5'→3'.
Последовательность интерферона α-2b

Амлифицированный ДНК-фрагмент выделяли и к нему добавляли рестриктирующий фермент DpnI для полного удаления нетрансформированных плазмид.
XL-1 Blue E.coli (Novagen, USA) трансформировали указанной модифицированной плазмидой. Определяли последовательность оснований ДНК, выделенной из трансформированных колоний, и таким образом получали плазмиду pT14SSIα-2b, которая содержала ген, кодирующий аргинин вместо 23-й аминоксилоты IFNα-2a, то есть лизина.
Затем BL21(DE3) E.coli трансформировали модифицированным вектором pT14SSIα-2b с получением трансформанта, обозначенного НМ 10701 E.coli, методом, описанным в сравнительном примере 1. Анализ N-концевой аминокислотной последовательности белка, продуцированного путем культивирования трансформанта, подтвердил, что этот трансформант экспрессировал IFNα-2b, имеющий нативную аминокислотную последовательность.
Пример 1: Конструирование вектора, содержащего мутант сигнального пептида энтеротоксина
(1) Конструирование вектора, содержащего [Thr4]STII
Для модификации конкретного аминокислотного остатка пептида сигнальной последовательности энтеротоксина получали вектор, содержащий полинуклеотид, кодирующий мутантную сигнальную последовательность энтеротоксина, с помощью сайт-направленного мутагенеза, описанного ниже.
Сначала вектор pT14SSIα-2a, полученный, как описано в сравнительном примере 1, подвергали ПЦР с использованием олигонуклеотидов SEQ ID No:22 и 23, в результате чего, в соответствии с процедурой сайт-направленного мутагенеза, описанной в сравнительном примере 2, получали модифицированную плазмиду, где 4-я аминокислота сигнальной последовательности энтеротоксина заменена треонином (Thr).

XL-1 Blue E.coli (Novagen, USA) трансформировали указанной модифицированной плазмидой. Определяли последовательность оснований ДНК, выделенной из трансформированных колоний, и таким образом получали плазмиду, которая содержала ген, кодирующий пептид сигнальной последовательности энтеротоксина, имеющий в 4-м аминокислотном положении Thr. Полученную таким образом плазмиду расщепляли ферментами XbaI и MluI, а затем встраивали в XbaI/MluI-участок вектора pT14SSIα-2a с получением вектора pT14SSIα-2a-4T.
Затем BL21(DE3) E.coli (Stratagene, USA) трансформировали вектором pT14SSIα-2a-4T с получением трансформанта E.coli, обозначенного HМ 10602 E.coli.
Вектор pT14SSIa-2a-4T конструировали с использованием pT14SSIα-2b, а затем в соответствии с методом, описанным выше, трансформировали в BL21(DE3) E.coli (Stratagene, USA) с получением трансформанта E.coli, обозначенного НМ 10702 E.coli.
(2) Конструирование вектора, содержащего [Thr4,Gln22]STII
Вектор pT14SSIα-2a-4T, полученный в стадии (1), подвергали ПЦР с использованием олигонуклеотидов SEQ ID No: 25 и 26, которые были сконструированы так, что в нем 22-я аминокислота сигнального пептида энтеротоксина, имеющего Thr в 4-м положении, была заменена на кодон Gln в соответствии с процедурой сайт-направленного мутагенеза в стадии (1), в результате чего получали модифицированную плазмиду.

Затем XL-1 Blue E.coli (Novagen, USA) трансформировали указанной модифицированной плазмидой. Определяли последовательность оснований ДНК, выделенной из трансформированных колоний, и таким образом получали плазмиду pT14SSIa-2α-4T22Q, которая содержала ген, кодирующий сигнальный пептид энтеротоксина, имеющий Thr и Gln в 4-м и 22-м аминокислотных положениях сигнальной последовательности энтеротоксина соответственно. Затем BL21(DE3) E. Coli (Stratagene, USA) трансформировали вектором pT14SSIα-2a-4T22Q в соответствии с методом, описанным в стадии (1), в результате чего получали трансформант E.coli, обозначенный НМ 10604 E.coli.
Для модификации последовательности Шайна-Дальгарно модифицированной сигнальной последовательности энтеротоксина с образованием последовательности SEQ ID No:9 векторы pT14SSIα-2a-4T и pT14SSIα-2a-4T22Q подвергали процедуре сайт-направленного мутагенеза, описанной в стадии (2) с использованием олигонуклеотидов SEQ ID No:27 и 28 с получением нужной модифицированной плазмиды.
XL-1 Blue E.coli (Novagen, USA) трансформировали указанной модифицированной плазмидой. Определяли последовательность оснований ДНК, выделенной из трансформированных колоний, и таким образом получали плазмиды pT14OSSIα-2a-4T и pT14OSSIα-2a-4T22Q, содержащие модифицированную последовательность Шайна-Дальгарно сигнальной последовательности энтеротоксина.
На фиг.4 проиллюстрирована вышеописанная процедура конструирования вектора pT14OSSIα-2a-4T22Q.
BL21(DE3) E.coli трансформировали векторами pT14OSSIα-2а-4Т и pT14OSSIα-2a-4T22Q соответственно с получением трансформантов, обозначенных НМ 10603 и НМ 10611 E.coli, которые были депонированы в Корейской коллекции культур микроорганизмов (КССМ) 23 декабря 1999 г. под номерами доступа КССМ-10175 и КССМ-10176 соответственно.
Кроме того, векторы pT14OSSIα-2b-4T и pT14OSSIα-2b-4T22Q получали в соответствии с процедурой, описанной выше для вектора pT14SSIα-2b, которые были использованы для трансформации BL21(DE3) E.coli с получением трансформантов, обозначенных НМ 10703 и НМ 10711 E.coli соответственно. Трансформанты НМ 10703 и НМ 10711 E.coli были депонированы в КССМ 23 декабря 1999 г. под номерами доступа КССМ-10177 и КССМ-10178 соответственно.
(3) Конструирование вектора, содержащего [Thr4,Val20, Gln22]STII
Для последующей замены 20-й аминокислоты пептида сигнальной последовательности энтеротоксина, имеющей Thr и Gln в 4-м и 22-м положениях, на кодон Val, векторы pT14OSSIα-2a-4T22Q и pT14OSSIα-2b-4T22Q, полученные на стадии (2), подвергали ПЦР с использованием олигонуклеотидов SEQ ID No: 29 и 30 методом сайт-направленного мутагенеза, описанного в стадии (2), в результате чего получали нужные модифицированные плазмиды, обозначенные pT14OSSIα-2a-4T20V22Q и pT14OSSIα-2b-4T20V22Q.
XL-1 Blue E.coli трансформировали указанными модифицированными плазмидами. Определяли последовательности оснований ДНК, выделенных из трансформированных колоний, и таким образом получали плазмиды pT14OSSIα-2a-4T20V22Q и pT14OSSIα-2b-4T20V22Q, которые содержали ген, имеющий кодоны Thr, Val и Gln вместо кодонов Asp в 4-м положении. Asp в 20-м положении и Tyr в 22-м положении соответственно. BL21(DE3) Е.coli трансформировали указанными плазмидами с получением трансформантов, обозначенных НМ 10612 и НМ 10712 E.coli соответственно.
Пример 2. Получение мутантной последовательности Шайна-Дальгарно для термостабильного энтеротоксина II
Для снижения числа оснований между сайтом связывания с рибосомой и инициирующим колоном ATG модифицированной сигнальной последовательности термостабильного энтеротоксина II E.coli внутри последовательности Шайна-Дальгарно термостабильного энтеротоксина II в полученном, как описано выше, экспрессирующем векторе конструировали модифицированную плазмиду в соответствии с процедурой сайт-направленного мутагенеза, описанной в сравнительном примере 2.
А именно для снижения числа оснований между сайтом связывания с рибосомой GAGG и инициирующим кодоном ATG с 7 до 5 вектор pT14OSSIα-2a-4T22Q, полученный, как описано в примере 1(2), подвергали сайт-направленному мутагенезу, описанному в сравнительном примере 2 с использованием олигонуклеотидов SEQ ID No: 31 и 32, в результате чего получали модифицированную плазмиду, обозначенную pT14NSSIα-2a-4T22Q. Кроме того, для снижения числа оснований между сайтом связывания с рибосомой GAGG и инициирующим кодоном ATG до 4, вектор pT14NSSIα-2a-4T22Q подвергали процедуре сайт-направленного мутагенеза, описанной в сравнительном примере 2 с использованием олигонуклеотидов SEQ ID No: 33 и 34, в результате чего получали модифицированную плазмиду, обозначенную pT14MSSIα-2a-4T22Q.
XL-1 Blue E.coli трансформировали указанными модифицированными плазмидами. Определяли последовательности оснований ДНК, выделенных из трансформированных колоний, и таким образом получали IFNα-экспрессирующие плазмиды pT14NSSIα-2a-4T22Q и pT14MSSIα-2a-4T22Q, которые соответственно содержали 5 и 4 основания между сайтом связывания с рибосомой GAGG и инициирующим кодоном ATG. BL21(DЕ3) E.coli трансформировали указанными экспрессирующими плазмидами с получением трансформантов, обозначенных НМ 10613 и НМ 10614 E.coli соответственно.
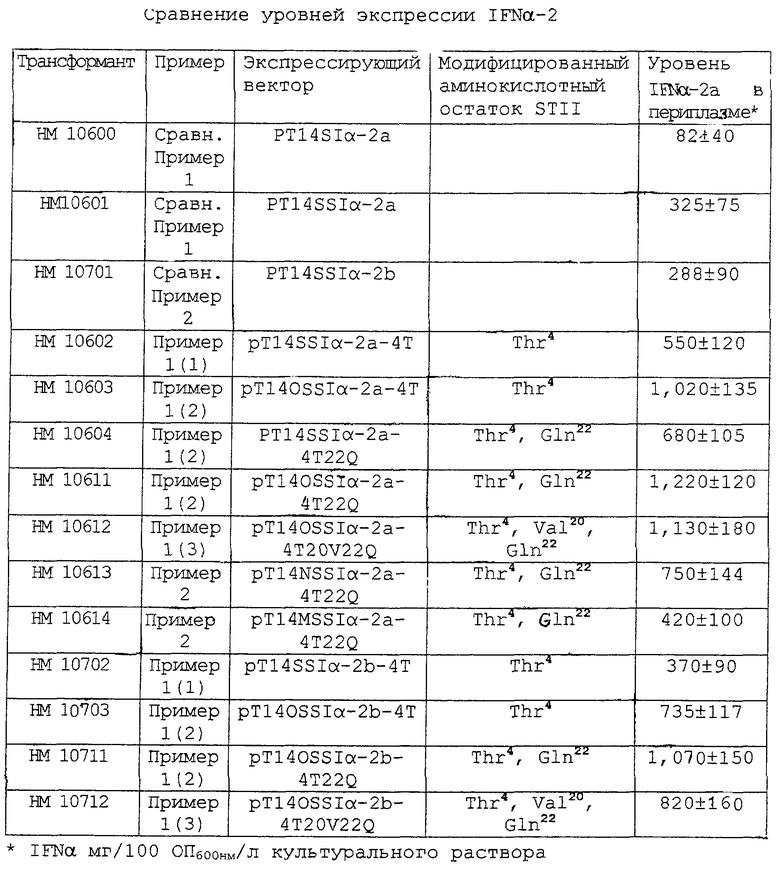
Пример 3. Сравнение уровней экспрессии IFNα-2
Трансформанты, полученные, как описано выше в Сравнительных примерах и в Примерах, культивировали в среде LB, а затем инкубировали в присутствии IPTG в течение 3 часов. Каждую из этих культур центрифугировали при 6000 об/мин в течение 20 минут для осаждения бактериальных клеток, и преципитат обрабатывали методом осмотического шока (Nossal, G.N., J.Biol.Chem., 241:3055, 1966), как описано ниже.
Преципитат суспендировали в 1/10 объема изотонического раствора (20% сахароза, 10 мМ буфер Трис-Cl, содержащий 1 мМ EDTA, рН 7,0). Полученную суспензию оставляли на 30 минут при комнатной температуре, а затем центрифугировали для сбора бактериальных клеток. Полученные клетки ресуспендировали в D.W. при 4°С для экстракции белков, присутствующих в периплазме клеток, и центрифугировали с получением супернатанта в виде периплазматического раствора. Уровень IFNα-2 в периплазматическом растворе анализировали методом ELISA (Kato, К. et al., J. Immunol., 116, 1554, 1976) с использованием антитела против IFNα-2 (R&D, USA), и этот уровень вычисляли как количество IFNα-2a, продуцируемого на 1 л культуры. Результаты представлены в таблице.
Пример 4. Последующая обработка и очистка
В соответствии с процедурой, описанной в примере 3, трансформант НМ 10611 E.coli, полученный, как описано в примере 1(2), культивировали в среде LB, и культуру центрифугировали при 6000 об/мин в течение 20 минут для сбора клеток. Из этих клеток получали периплазматический раствор методом осмотического шока.
Указанный периплазматический раствор доводили до рН 5,0-5,5, адсорбировали на колонке с S-сефарозой (Pharmacia Inc., Sweden), предварительно уравновешенной до рН 5,3, а затем колонку промывали 25 мМ NaCl. IFNα-2 элюировали путем последовательного добавления растворов, забуферных уксусной кислотой и содержащих 50 мМ, 100 мМ, 200 мМ и 1 М NaCl соответственно, и фракции, содержащие IFNα-2, собирали и объединяли.
Объединенные фракции подвергали хроматографии на колонке с сефарозой Blue (Pharmacia Inc., Sweden) и элюировали добавлением в колонку буферных растворов, содержащих более 2 М NaCl, с получением активной фракции.
Активную фракцию диализовали буфером, и, наконец, подвергали фракционированию на колонке с анионообменной смолой DEAE при рН 5,8 с получением IFNα-2a, имеющего чистоту более 99%. Кроме того, IFNα-2b выделяли из трансформанта НМ 10711 E.coli путем повторения вышеописанной процедуры.
Каждую из очищенных фракций IFNα-2a и IENα-2b подвергали электрофорезу в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ) для определения чистоты и приблизительной концентрации IFNα, а затем подвергали стандартному ELISA-методу, как описано в примере 3 для определения точной концентрации IFNα в периплазматическом растворе. Кроме того, с помощью анализа N-концевой аминокислотной последовательности было подтверждено, что IFNα-2a и IFNα-2b представляют собой природные типы, не содержащие дополнительного метионина.
Пример 5: Определение молекулярной массы IFNα-2a, продуцированного из рекомбинантных клеточных линий
Уровень экспрессии и молекулярные массы IFNα-2a и IFNα-2b, продуцированных из рекомбинантных клеточных линий, определяли с помощью электрофореза в ДСН-ПААГ и Вестерн-блоттинга.
Сначала периплазматическую фракцию трансформанта НМ 10611 E.coli, полученного, как описано в примере 4, и очищенный IFNα-2a, полученный из нее, подвергали электрофорезу в ДСН-ПААГ стандартным методом с использованием коммерчески доступного продукта IFNα-2a (3 × 106 МЕ/мл) в качестве контроля.
На фиг.5а представлены результаты электрофореза в ПААГ с ДСН, где дорожка 1 соответствует IFNα-2а-контролю; дорожка 2 соответствует периплазматической фракции трансформанта НМ 10611 E.coli; а дорожка 3 соответствует очищенному IFNα-2a. Как видно из фиг.5а, очищенный IFNa-2a имеет ту же молекулярную массу, что и природный IFNα-2a, и присутствует в периплазматической фракции трансформанта НМ 10611 E.coli на высоком уровне.
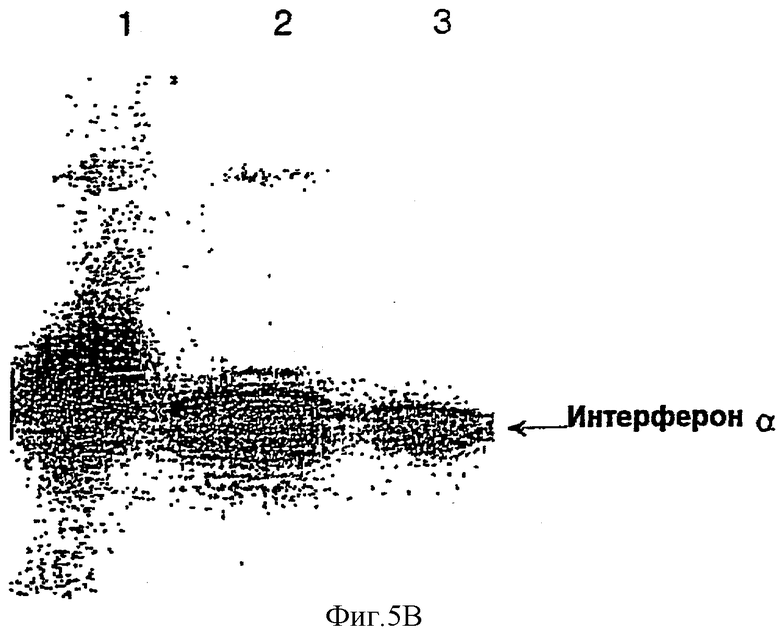
Кроме того, периплазматическую фракцию трансформанта НМ 10711 E.coli, очищенную фракцию, полученную в результате хроматографии периплазматического раствора на колонке с S-Сефарозой, и наконец, очищенный IFNα-2b подвергали электрофорезу в ПААГ с ДСН стандартным методом.
Нитроцеллюлозный фильтр (Bio-Rad Lab, USA) смачивали в буферном растворе для блоттинга (170 мМ глицин, 25 мМ Трис • НСl [рН 8], 20% метанола) и разделенные на геле белки переносили на нитроцеллюлозный фильтр в течение 3 часов с использованием набора для блоттинга. Фильтр выдерживали в 1% казеине в течение 1 часа и три раза промывали PBS, содержащим 0,05% Твина 20. Затем фильтр помещали в раствор кроличьего антитела против IFNα (Chemicon, # AB1434, USA), разведенного PBS, и оставляли на 2 часа при комнатной температуре для прохождения реакции. После проведения реакции фильтр 3 раза промывали раствором PBST для удаления непрореагировавшего антитела. Затем добавляли конъюгированное с пероксидазой хрена козье антитело против кроличьего IgG (Bio-Rad Lab, USA), разведенное в PBS, и подвергали реакции при комнатной температуре в течение 2 часов. Фильтр промывали PBST, и к нему добавляли раствор набора субстратов для пероксидазы (Bio-Rad Lab, USA) для развития цветной реакции. Результаты вышеописанного Вестерн-блоттинга приводятся на фиг.5b, где дорожка 1 соответствует периплазматической фракции трансформанта НМ 10711 E.coli; дорожка 2 соответствует фракции, очищенной хроматографией на колонке с S-сефарозой; а дорожка 3 соответствует конечному очищенному IFNα-2b.
Результаты этого примера подтверждают, что из рекомбинантных штаммов Е.coli настоящего изобретения экспрессируется большое количество растворимого IFNα.















Изобретение относится к области биотехнологии, конкретно к производству рекомбинантных интерферонов человека. Экспрессирующий вектор для секреторного продуцирования человеческого интерферона альфа (hIFNα) содержит полинуклеотид, кодирующий модифицированную сигнальную последовательность термостабильного энтеротоксина II E.coli, и полинуклеотид, кодирующий hIFNα, лигированный с его 3′-концом. Вектор используют в способе секреторного продуцирования человеческого интерферона путем трансформации им штамма Е.coli и культивирования указанного штамма. Способ позволяет секретировать растворимую форму активного hIFNα, не содержащего дополнительного метионинового остатка на своем N-конце, в периплазму клетки Е.coli. 8 с. и 5 з.п. ф-лы, 5 ил., 1 табл.
| US 5710027 А, 20.01.1998 | |||
| CHANG CN, et al | |||
| High-level secretion of human growth horman by Escherichia coli: Gene | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| SAEED AM, et al | |||
| Molecular homogeneity of heat-stable enterotoxins produced by bovine enterotoxigenic Escherichia coli | |||
| Infec | |||
| Immun, 1984, vol.45, no.1, р.242-247 | |||
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК РТТG КM2, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ИММУННОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА, ШТАММ ESCHERICHIA COLI T3G - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ИММУННОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУННОГО ИНТЕРФЕРОНА ЧЕЛОВЕКА | 1996 |
|
RU2097428C1 |
| МОДИФИЦИРОВАННЫЙ СИГНАЛЬНЫЙ ПЕПТИД ЭНТЕРОТОКСИНА - II E. COLI И МИКРООРГАНИЗМ, ЭКСПРЕССИРУЮЩИЙ БЕЛОК СЛИЯНИЯ УПОМЯНУТОГО ПЕПТИДА С ГЕТЕРОЛОГИЧНЫМ БЕЛКОМ | 1999 |
|
RU2198179C2 |
| УСТРОЙСТВО ДЛЯ ТРАНСПОРТИРОВАНИЯ ШТУЧНЫХ ГРУЗОВ | 0 |
|
SU177343A1 |

