Изобретение относится к генетической инженерии и биотехнологии, конкретно к способам получения рекомбинантного иммунного интерферона человека медицинского назначения (далее γ-интерферон) из микробиологического продуцента.
Известна рекомбинантная плазмидная ДНК pINF-trp, кодирующая синтез g-интерферона [1] сконструированная на основе плазмиды pBR 322 с синтетическим участком связывания рибосом и триптофановым промотором. Однако продуктивность рекомбинатнтного штамма с плазмидой pINF-trp определена при культивировании в малом объеме и только в процентах от суммарного белка. Участок плазмидной ДНК, кодирующий g-интерферон, не содержит в своем составе терминатор транскрипции, что отрицательно сказывается на стабильности плазмиды и продуктивности рекомбинантного штамма. Отсутствуют данные о создании промышленного штамма-продуцента на основе этой плазмиды.
Известна плазмидная ДНК pLPPT, кодирующая синтез g-интерферона [2] созданная на основе плазмиды pBR322 с двумя промоторами trp и lpp генов и имеющая в качестве селективного маркера amp ген. Однако, данная плазмида не обеспечивает достаточно высокий уровень синтеза g-интерферона. Так, рекомбинантный штамм с плазмидой pLPPT способен синтезировать 100 мг g-интерферона на литр культуральной жидкости.
Известен штамм E. coli W3110 (pLPPT) продуцент g-интерферона [2] полученный трансформацией плазмиды pLPPT в прототрофный штамм E. coli W3110. Уровень синтеза в данном штамме недостаточно высок (100 мг g-интерферона на литр культуральной жидкости). Кроме того, для селективного поддержания этого штамма при культивировании используют ампициллин, являющийся сильным аллергеном, что создает определенные трудности при промышленном производстве g-интерферона.
Известен способ получения g-интерферона [2] заключающийся в глубинном культивировании штамма E. coli W3110 (pLPPT) в питательной среде в условиях аэрации и индукции синтеза g-интерферона путем добавления в определенный момент культивирования индолилакриловой кислоты и изопропилтиогалактозида. Для выделения и очистки g-интерферонаа используется ультразвуковое разрушение биомассы, очистка агрегированного g-интерферона от растворимых примесей, осаждение денатурированного g-интерферона, диализ разбавленного раствора белка.
Однако данный способ накладывает повышенные требования к выбору момента добавления индуктора для получения максимального выхода целевого продукта с единицы объема ферментера, что усложняет технологию процесса культивирования. Помимо этого, разрушение больших количеств биомассы ультразвуком и диализ разбавленного раствора белка создает определенные технологические трудности при крупномасштабном производстве g-интерферона.
Цель изобретения конструирование рекомбинантной плазмидной ДНК с геном g-интерферона, получение штамма-продуцента g-интерферона и создание способа получения g-интерферона.
Задача решается путем создания рекомбинантной плазмидной ДНК pTTg Km2, кодирующей синтез g-интерферона, размером 4440 пар оснований (п.о.), состоящей из следующих элементов:
HindIII-EcoRI фрагмент ДНК размером 220 п.о. включающий последовательность триптофаного промотора;
EcoRI-XbaI фрагмент ДНК размером 20 п.о. содержащий синтетический участок связывания рибосом;
5'GAATTCAGGAGGAATCTAGA 3'
3'CTTAAGTCCTCCTTAGATCT 5'
XbaI-BamHI фрагмент ДНК размером 450 п.о. кодирующий g-интерферон;
BamHI-ScaI фрагмент ДНК размером 850 п.о. плазмиды pKK233-3, содержащий терминатор транскрипции rrnBT1T2
PstI-PstI фрагмент ДНК размером 1500 п.о. плазмиды pUC4K, кодирующий устойчивость к канамицину (kan);
BglI-HindIII фрагмент ДНК размером 1400 п.о. плазмиды pUC19.
Штамм Escherichia coli T3g продуцент g-интерферона получен трансформацией клеток E. coli C600 Str (3) плазмидой pTTg Km2, хранится в Центральной коллекции микроорганизмов Государственного концерна "БИОПРЕПАРАТ" ЦКМК "Б" под N ЦКМ B-58g.
Характеристика штамма E. coli T3g
Культурально-морфологические признаки.
Морфологические признаки клетки прямые, палочковидной формы (0,5-0,7)•(2-2,5) мкм, подвижные, грамотрицательные, неспороносные.
Культуральные признаки клетки хорошо растут на простых питательных средах.
При росте на мясо-петонном агаре, питательном агаре "Дифко" колонии гладкие, круглые, выпуклые, блестящие, серые, края ровные, мутные.
При росте в жидких средах: мясо-пептонном бульоне, LB-бульоне образует ровную интенсивную муть.
Физиолого-биохимические признаки.
Клетки растут в пределах от 4 до 45oC при оптимуме pH 5,8-7,8.
Сбраживают до кислоты глюкозу, мальтозу, маннит, маннозу, глицерин.
В качестве источника азота используют как минеральные соли в аммонийной и нитратной форме, так и в органической форме в виде пептона, аминокислот.
Нитраты восстанавливают до нитритов.
Желатину не разжижают.
Индол не образуют.
Уреазная активность не обнаруживается.
Устойчивость к антибиотикам.
Проявляет устойчивость к стрептомицину и к канамицину, обусловленную наличием плазмиды pTTg Km2.
Способ получения g-итерферона предусматривает глубинное культивирование рекомбинантных бактерий E. coli в питательной среде в условиях аэрации, выделение агрегированного g-интерферона с последующей очисткой, причем в качестве рекомбинантного штамма используют E. coli T3g, а культивирование осуществляют на питательной среде с пониженным содержанием триптофана, величину pH уменьшают после прекращения удвоения клеток с 8-6 до 6-5, агрегированный g-интерферон отмывают фосфатным буфером, растворяют в концентрированном растворе мочевины с цетавлоном, далее либо примеси из раствора ренатурированного интерферона осаждают добавлением солей с двух- или трехзарядными анионами органических и неорганических кислот, такими как анионы серной, фосфорной, лимонной кислот в области величины pH от 7 до 9 с последующей хроматографией на анионообменнике или мочевинный раствор g-интерферона очищают с помощью высокоэффективной обращенно-фазной хроматографии, с последующей ренатурацией и хроматографией на анионообменнике.
Пример 1. Рекомбинантную плазмиду pTTg Km2 получают в несколько этапов:
1) выделение иРНК из индуцированных лимфоцитов человека, синтез и клонирование кДНК;
2) клонирование триптофанового проматора из генома E. coli;
3) конструирование рекомбинантной плазмиды pINF-trp8;
4) конструирование рекомбинантной плазмиды pTTgl;
5) конструирование рекомбинантной плазмиды pTTgKm2.
Этап 1. Выделение иРНК из индуцированных лимфоцитов человека, синтез и клонирование кДНК.
Индуцированные лимфоциты крови человека (1•1010 клеток) стафилококковым энтеротоксином B (0,04 мкг/мл) промываются на льду фосфатным буфером, приготовленным на физиологическом растворе (PBS 0,01 мМ натрия фосфорнокислого двузамещенного, 0,003 мМ калия фосфорнокислого однозамещенного, pH 7,2, 0,137 мМ натрия хлористого) и центрифугируют при 2000 g в течение 6 мин при температуре 4oC. Недостаточную жидкость удаляют, осадок клеток ресуспендируют в охлажденном во льду лизирующем буфере (0,14 мМ хлористого натрия, 1,5 мМ магния хлористого, 10 мМ Tris-HCl, pH 8,6, 0,5% NP-40). Суспензию перемешивают в течение 10 с и наслаивают на равный объем лизирующего буфера, содержащего 24% сахарозы (вес/объем) и NP-40 (1%). После центрифугирования при 10000 g 20 мин при температуре 4oC отбирают непрозрачный верхний (цитоплазматический) слой и добавляют равный объем буфера, имеющего состав 0,2 мМ Tris-HCl, pH 7,5, 25 мМ ЭДТА, 0,3 мМ натрия хлористого, 2% SDS. Белки удаляют двукратной экстракцией смеси фенол-хлороформ, РНК осаждают 2,5 объемами спирта этилового. Осадок промывают 75%-ным охлажденным этиловым спиртом и растворяют в минимальном объеме воды. иРНК используют для синтеза кДНК. Реакционная смесь для синтеза кДНК содержит 100 мкг/мл иРНК, 50 мМ dNTP, 50 Мм Tris-HCl, pH 8,3, 6 мМ хлористого магния, 80 мМ калия хлористого, 5 мкг/мл синтетического олигонуклеотида.
5'CCGGATCCTTACTGGGATGCTCTTCGACC3',
и 15 ед/мкг обратной транскриптазы вируса миелобластоза птиц. Синтез проводят при температуре 42oC в течение 1 ч. Реакционную смесь депротеинизируют фенолом, ДНК осаждают этиловым спиртом. кДНК амплифицируют in vitro, основанном на многократном повторении циклов денатурации ДНК при температуре 94oC, сплавлении с матрицей специфических синтетических олигонуклеотидных затравок при температуре 50oC и синтезе с них комплементарных цепей при температуре 72oC с помощью термостабильной ДНК-полимеразы, выделенной из Thermus aguaticus. Реакционная смесь содержит 10 мМ Tris-HCl, pH 8,3, 50 мМ хлористого калия, 1,5 мМ хлористого магния, 0,001% (w/v) желатина, 0,2 мМ dNTP, 5 мкг/мл синтетических олигонуклеотидов:
5'CCTCTAGATGCAGGACCCATAT3',
5'CCGGATCCTTACTGGGATGCTCTTCGACC3',
и 2,5 ед. термостабильной ДНК-полимеразы. Один цикл включает следующие стадии:
1) денатурация 94oC, 1,0 мин;
2) сплавление 50oC, 1,0 мин;
3) полимеризация 72oC, 1,0 мин.
Процесс амплификации состоит из 30 циклов. Реакционную смесь депротеинизируют фенолом, ДНК осаждают этиловым спиртом.
Затем проводят клонирование кДНК. Для этого 1 мкг ДНК плазмиды pUC19 и 1 мкг кДНК рестрицируют ферментами XbaI и BamHI в 50 мкл смеси: 10 мМ Tris-HCl, pH 7,5, 50 мМ хлористого натрия, 10 мМ хлористого магния, 1 мМ дитиотрейтола, по 5 ед. ферментов XbaI и BamHI. Реакцию проводят при температуре 37oC, 60 мин. Далее смесь прогревают 10 мин при температуре 70oC для инактивации ферментов, добавляют АТФ (1 мМ), дитиотрейтол (1 мМ), 10 ед. ДНК-лигазы фага T4. Лигирование проводят при температуре 14oC 12 ч, после чего лигатом трансформируют компетентные клетки штамма E. coli JM83, затем высевают их на Мак-Конки агар, содержащий ампициллина 100 мкг/мл. Через 12 ч (температура 37oC) белые рекомбинантные клоны отсевают, выделяют плазмидные ДНК, проводят рестрикционный анализ и определяют нуклеотидную последовательность. В результате получают плазмиду pXBl (фиг.1), размером 3140 п.о. в которой ген g-интерферона фланкирован сайтами XbaI и BamHI, кодоны аминокислот Cys-Tyr-Cys заменены на кодон ATG (Met) (фиг.5).
Этап 2. Кклонирование триптофанового промотора из генома E. coli.
Для этого выделяют тотальную ДНК E. coli. Ночную культуру штамма E. coli HB101 центрифугируют при 5000 g 10 мин, суспендируют в лизирующем буфере (50 мМ глюкозы, 25 мМ Tris-HCl, 10 мМ ЭДТА) до концентрации 1 мг/мл, выдерживают 30 мин при температуре 4oC, смешивают с равным объемом 2%-ного раствора додецил-сульфата натрия, добавляют 10 мкг/мл протеиназы K и ингибируют в течение 12 ч при температуре 37oC. Далее ДНК депротеинизируют фенолом, смесью фенола и хлороформа (1:1), хлороформом, осаждают этиловым спиртом, дважды промывают этиловым спиртом и растворяют в воде до концентрации 1 мг/мл.
Клонирование триптофанового промотора осуществляют с помощью реакции амплификации ДНК in vitro. Для этого готовят смесь: 10 мМ Tris-HCl, pH 8,3; 50 мМ хлористого калия; 1,5 мМ хлористого магния; 0,2 мМ dNTP; 1 мкг/мл ДНК E. coli; 0,002 мМ синтетических олигонуклеотидов
5'GGAAGCTTCTGGCGTCAGGCAGCCAT 3'
5'CCTGAATTCCCTTTTTACGTGAACTTGCG 3'
и 2,5 ед. термостабильной ДНК полимеразы. Один цикл включает следующие стадии:
1) денатурация 94oC, 1,0 мин;
2) сплавление 50oC, 1,0 мин;
3) полимеризация 72oC, 1,0 мин.
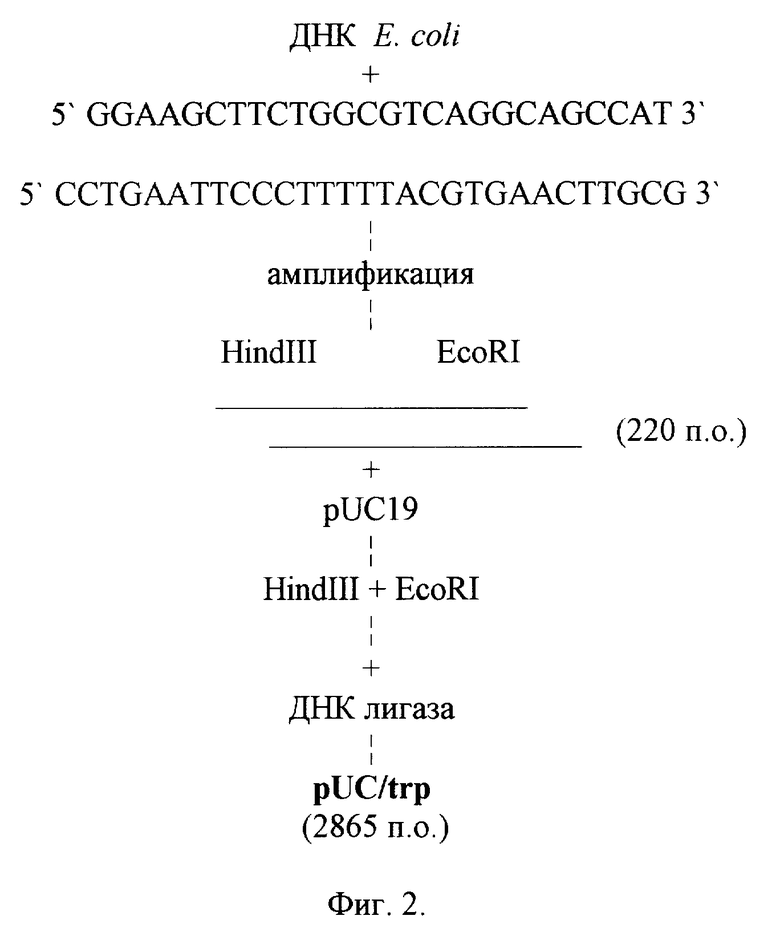
Процесс амплификации состоит из 30 циклов. Реакционную смесь депротеинизируют фенолом, ДНК осаждают этиловым спиртом и растворяют в минимальном объеме. Далее проводят клонирование амплифицированной ДНК, для чего готовят следующую реакционную смесь: 10 мМ Tris-HCl с pH 7,5, 50 мМ натрия хлористого, 10 мМ магния хлористого, 1 мМ дитиотрейтола, 1 мкг ДНК pUC19, 1 мкг амплифицированной ДНК, ферменты EcoRI и HindIII по 5 ед. Рестрикцию проводят при температуре 37oC 1,5 ч. Далее смесь прогревают при температуре 70oC 10 мин, добавляют дитиотрейтол (1 мМ), АТФ (1 мМ), 5 ед. ДНК лигазы фага T4. Реакцию лигирования проводят при температуре 14oC 12 ч, после чего лигатом трансформируют клетки штамма E. coli JM83 и трансформанты высевают на среду Мак-Конки, содержащую 100 мкг/см3 ампицилина. Через 12 ч (температура 37oC) белые рекомбинантные клоны отсеивают, выделяют плазмидные ДНК, проводят рестрикционный анализ и определяют нуклеотидную последовательность. В результате получают плазмиду pUC/trp размером 2856 п.о. (фиг.2), в которой ДНК триптофанового промотора фланкирована сайтами рестрикции HindIII и EcoRI.
Этап 3. Конструирование рекомбинантной плазмиды pINF-trp8.
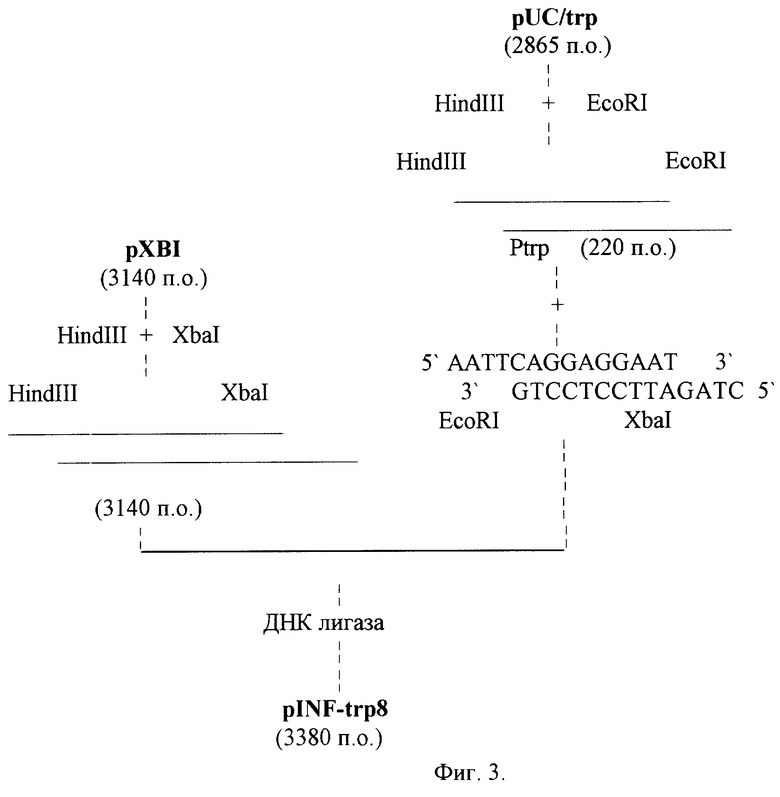
Для получения рекомбинантной плазмиды pINF-trp8 предпочтительно ДНК плазмиды pXBl обрабатывают ферментами HindIII и XbaI, выделяют и очищают фрагмент HindIII-EcoRI из плазмиды pUC/trp. 10 мкг плазмиды pXBl, содержащей ген g-интерферона, расщепляют смесью рестриктаз HindIII и XbaI, для чего готовят следующую реакционную смесь: 10 мМ Tris-HCl с pH 7,5, 50 мМ натрия хлористого, 1 мМ дитиотрейтола, 10 мкг ДНК pXBl, ферменты HindIII и XbaI по 5 ед. Рестрикцию проводят при температуре 37oC 1,5 ч. Далее смесь прогревают при температуре 70oC 10 мин. 10 мкг плазмиды pUC/trp, содержащей последовательность триптофанового промотора (220 п.о.), расщепляют смесью рестриктаз HindIII и EcoRI, для чего готовят следующую реакционную смесь: 10 мМ Tris-HCl с pH 7,5, 50 мМ натрия хлористого, 10 мМ магния хлористого, 1 мМ дитиотрейтола, 10 мкг ДНК pUC/trp, ферменты EcoRI и HindIII по 5 ед. Рестрикцию проводят при температуре 37oC 1,5 ч. Далее смесь прогревают при температуре 70oC 10 мин. Фрагмент ДНК длиной 220 п.о. отделяют электрофоретически в 1,2% -ном агарозном геле, элюируют, очищают хроматографией на мини-колонке NACS PREPAC (BRL). Далее проводят лигирование фрагментов ДНК (плазмиды pXBl, обработанной ферментами HindIII, XbaI и фрагмента ДНК, содержащего триптофановый промотор, фланкированный сайтами рестрикции HindIII, EcoRI) с синтетическим фрагментом ДНК размером 20 п.о. содержащий последовательность (SD) Шайн-Дельгарно и фланкированный сайтами рестрикции EcoRI и XbaI.
5' AATTCAGGAGGAAT 3'
3' GTCCTCCTTAGATC 5'.
Лигированную ДНК трансформируют в клетки штамма E. coli JM83. Трансформанты высевают на среду LB, содержащую 100 мкг/мл ампициллина. Через 12 ч (температура 37oC) рекомбинантные клоны отсевают, выделяют плазмидные ДНК, проводят рестрикционный анализ и определяют нуклеотидную последовательность. В результате получают плазмиду pINF-trp8 размером 3380 п.о. в которой ген g-интерферона находится под контролем триптофанового промотора (фиг.3).
Этап 4. Конструирование рекомбинантной плазмиды pTTgl.
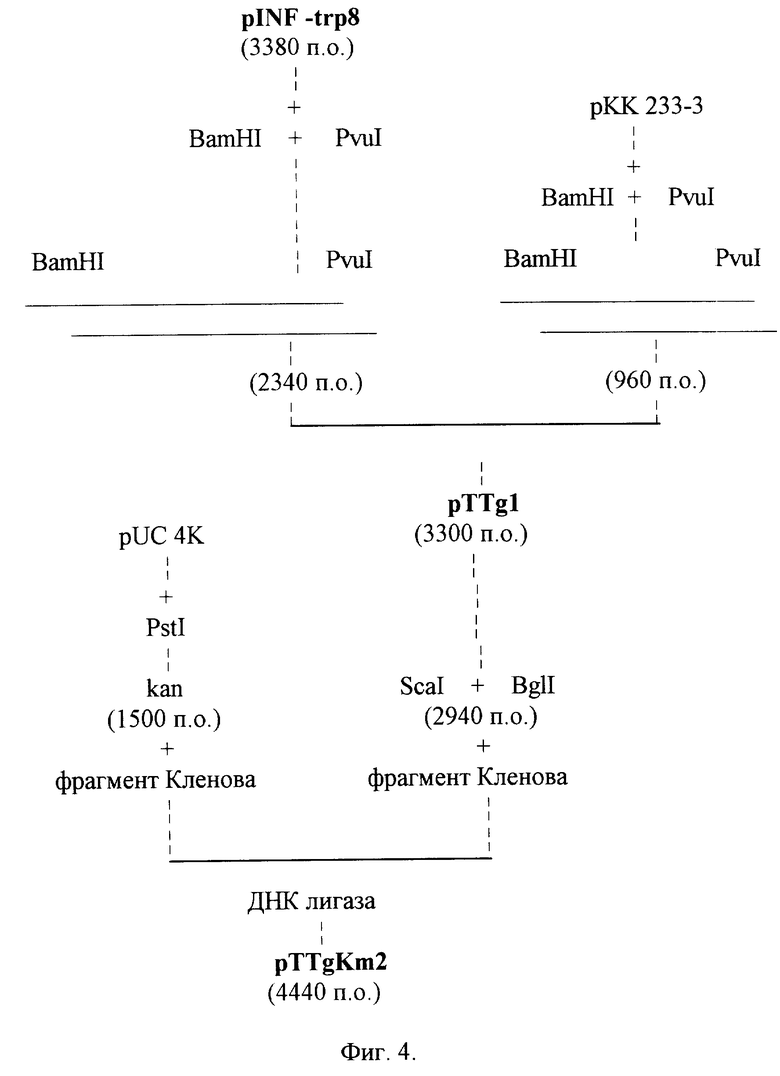
Для получения рекомбинантной плазмиды pTTgl предварительно выделяют и очищают следующие фрагменты ДНК: BamHI-PvuI фрагмент размером 2340 п.о. из плазмиды pINF-trp8 и BamHI-PvuI фрагмент размером 960 п.о. из плазмиды pKK233-3. 10 мкг плазмиды pINF-trp8, содержащей ген g-интерферона, расщепляют смесью рестриктаз BamHI, PvuI. Фрагмент ДНК размером 2340 п.о. отделяют электрофоретически в 0,8% -ном агарозном геле, элюируют, очищают хроматографией на мини-колонке NACS PREPAC (BRL). 10 мкг плазмиды pKK 233-3, содержащей последовательность "строгого" терминатора транскрипции rrnB T1T2, расщепляют смесью рестриктаз BamHI, PvuI. Фрагмент ДНК размером 960 п.о. отделяют электрофоретически в 1,2% -ном агарозном геле, элюируют, очищают хроматографией на мини-колонке NACS PREPAC (BRL).
Далее проводят лигирование фрагментов ДНК. Лигированную ДНК трансформируют в клетки штамма E. coli JM83. Трансформанты высевают на среду LB, содержащую 100 мкг/мл ампициллина. Через 12 ч (температура 37oC) рекомбинантные клоны отсевают, выделяют плазмидные ДНК, проводят рестрикционный анализ и определяют нуклеотидную последовательность. В результате получают плазмиду pTTGl размером 3300 п.о. в которой ген g-интерферона находится под контролем триптофанового промотора и терминатора транскрипции (фиг.4).
Этап 5. Конструирование рекомбинантной плазмиды pTTG Km2.
Для получения рекомбинантной плазмиды pTTg Km2 предварительно выделяют и очищают следующие фрагменты ДНК: Scal-BglI фрагмент размером 2940 п.о. из плазмиды pTTgl и PstI фрагмент размером 1500 п.о. из плазмиды pUC 4K. 10 мкг плазмиды pTTg Kml, содержащей ген g-интерферона, расщепляют смесью рестриктаз Scal, BglI. Фрагмент ДНК размером 2940 п.о. отделяют электрофоретически в 0,8%-ном агарозном геле, элюируют, очищают хроматографией на мини-колонке NACS PREPAC (BRL). 10 мкг плазмиды pUC 4K, содержащей последовательность гена устойчивости к канамицину, расщепляют рестриктазой PstI. Фрагмент ДНК размером 1500 п. о. отделяют электрофоретически в 1,2%-ном агарозном геле, элюируют, очищают хроматографией на мини-колонке NACS PREPAC (BRL). Далее фрагменты ДНК объединяют, обрабатывают фрагментом Кленова, как описано выше, и лигируют. Лигированную ДНК трансформируют в клетки штамма E. coli C600. Трансформанты высевают на среду LB, содержащую 20 мкг/мл канамицина. Через 12 ч (температура 37oC) рекомбинантные клоны отсевают, выделяют плазмидные ДНК, проводят рестрикционный анализ и определяют нуклеотидную последовательность. В результате получают плазмиду pTTg Km2 размером 4440 п.о. в которой ген g-интерферона находится под контролем триптофанового промотора и терминатора транскрипции (фиг.4-6).
На фиг.1 дана схема конструирования плазмиды pXB1; на фиг.2 схема конструирования плазмиды pUC/trp; на фиг.3 схема конструирования плазмиды pINF-trp8; на фиг. 4 схема конструирования плазмиды pTTG 1 и pTTg Km2; на фиг.5 нуклеотидная последовательность гена g-интерферона, аминокислотная последовательность белка g-интерферона и нуклеотидная последовательность олигонуклеотидов, использованных при конструировании плазмиды pTTg Km2 (последовательности олигонуклеотидов подчеркнуты); на фиг.6 рестрикционная карта плазмиды pTTg Km2.
Пример 2. Получение штамма Escherichia coli T3G продуцента g-интерферона
Для получения репициентного штамма E. coli C600 StrR берут 4•109 микробных клеток E. coli C600 (ATCC N23724) [3] выращенных в течение 18 ч при температуре 37oC на мясо-пептонном бульоне, высевают на LA-агар с 1000 мкг/мл стрепомицина и инкубируют при 37oC 48 ч. Появившиеся 15 колоний засевают на LA-агар с 1000 мкг/мл стрепомицина и выращивают при 37oC 20 ч. Отбирают один наиболее пышно растущий вариант и его рассевают до изолированных колоний на LA-агаризованной среде с 1000 мкг/мл стрептомицина и выращивают при 37oC в течение 18 ч. В качестве реципиентного штамма для трансформации используют отдельный клон.
Штамм-продуцент g-интерферона Escherichia coli T3g получают путем трансформации штамма E. coli C600 StrR рекомбинантной плазмидой pTTg Km2 с последующим отбором рекомбинантного клона на среде с канамицином при температуре 37oC и с определением активности g-интерферона в экстрактах клеток, выращенных до оптической плотности 4,0-6,0 в среде М9: 10% медицинского гидролизата казеина, 1% глюкозы, 20 мкг/мл канамицина, при температуре 37oC.
Активность g-интерферона определяют методом ингибирования цитопатического действия вируса везикулярного стоматита на диплоидные фибробласты человека и иммуноферментным методом. Содержание g-интерферона в 1 л культуры после 8 ч выращивания составляет 1,2•1010 ME активности.
Пример 3. Получение g-интерферона из штамма Escherichia coli T3g с использованием ионно-обменной хроматографии
этап 1) биосинтез g-интерферона;
этап 2) выделение и очистка g-интерферона;
этап 3) хроматографическая очистка g-интерферона.
Этап 1.
Приготавливают 10 л питательной среды для посевного ферментера марки ЗУ (ОКБ ТБМ, г. Кириши, Ленинградской области) емкостью 16 л следующего состава, г/л:
Гидролизат казеина 150
Экстракт пекарских дрожжей 5
Д-глюкоза 10
Сернокислый аммоний 2
Фосфорнокислый однозамещенный калий трехводный 4
Фосфорнокислый двухзамещенный натрий 12
Хлористый натрий 5
Сернокислый магний семиводный 0,25
Хлористый кальций 0,011
Канамицин 0,04
Дистиллированная вода Остальное
1 л этой среды разливают по 250 мл в 4 качалочные колбы и засевают по 1 мл жидкозаконсервированной культуры штамм-продуцента g-интерферона Escherichia coli T3g. колбы помещают на 24 ч в термостатируемую качалку при (24±0,5)oC. Полученным инокулятом засевают посевной ферментер. Температуру во время выращивания выдерживают 37,3oC, pH 7,3 pO2 60% Посевную культуру выращивают до оптической плотности 4 на длине волны 540 нм и переносят в ферментер БИОР-0,1 (ОКБ ТБМ г. Кириши) объемом 100 л, содержащий 60 л питательной среды следующего состава, г/л:
Гидролизат казеина 150
Д-глюкоза 10
Сернокислый аммоний 2
Фосфорнокислый однозамещенный калий трехводный 4
Фосфорнокислый двузамещенный натрий 12
Хлористый натрий 5
Сернокислый магний семиводный 0,25
Хлористый кальций 0,011
Канамицин 0,04
Дистиллированная вода Остальное
Температуру ферментера выдерживают 37oC, величину pH среды 7,1 до оптической плотности 4 на длине волны 540 нм и pH среды 5,8 до конца процесса культивирования. pO2 поддерживают 70% от насыщения до оптической плотности 4 и 40% до конца процесса. Накопление g-интерферона в виде "телец включений" контролируют с помощью фазово-контрастного микроскопа. Процесс культивирования завершают спустя 2 ч после прекращения удвоения клеток. Биомассу после культивирования осаждают центрифугированием на проточной центрифуге и хранят до использования при температуре минус 70oC.
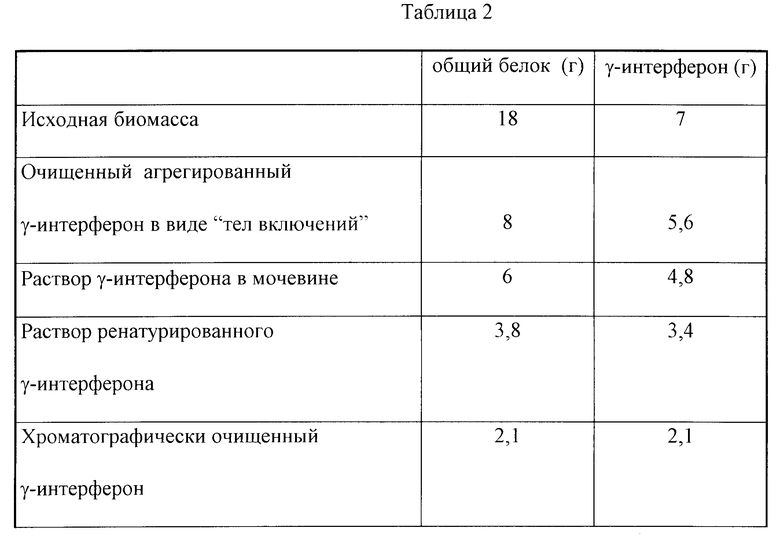
В табл.1 приведены данные об изменении оптической плотности и накопления g-интерферона в зависимости от времени культивирования. Из табл.1 видно, что в конце процесса культивирования концентрация g-интерферона по данным иммуноферментного метода достигает 1,1•1010 ME на 1 л культуры при плотности биомассы 7 г/л.
Величина выхода g-интерферона практически не меняется, если начало культивирования проводить при величине pH от 8 до 6, а конечную стадию культивирования проводить при величине pH от 6 до 5.
Этап 2. Выделение и очистка g-интерферона.
150 г замороженной при температуре минус 70oC биомассы E. coli T3g, продуцента g-интерферона ресуспендируют в 1500 мл буфера (50 мМ натрия фосфорнокислого двухзамещенного, 1 мМ ЭДТА, 0,4 М натрия хлористого) при температуре 8oC и центрифугируют в течение 20 мин на центрифуге Бэкман J2-21 при скорости 8000 об/мин на роторе JA-10. Получают отмытый осадок клеток. Осадок клеток повторно ресуспендируют в 450 мл того же буфера, добавляют 150 мг лизоцима и инкубируют с перемешиванием при температуре 8oC в течение 30 мин. Обработанную лизоцимом суспензию замораживают при минус 20oC в течение 3 ч и размораживают при температуре 25oC. Вязкость суспензии снижают добавлением ДНКазы 5 мг и магния сернокислого до концентрации 5 мМ и инкубацией в течение одного часа. После добавления равного объема 8% раствора Triton X-100 в буфере (50 мМ натрия фосфорнокислого двузамещенного, 1мМ ЭДТА, 0,4 М натрия хлористого) и инкубации в течение 1 ч суспензию ценрифугируют в течение 20 мин на центрифуге Бэкман J2-21 при скорости 8000 об/мин на роторе JA-10.
Полученный осадок ресуспендируют в 150 мл буфера (50 мМ натрия фосфорнокислого двузамещенного, 1 мМ ЭДТА, 0,4 натрия хлористого), добавляют 1250 мл буфера (50 мМ натрия фосфорнокислого трехзамещенного, 1мМ ЭДТА), и инкубируют с перемешиванием в течение 20 мин, после чего центрифугируют в течение 20 мин на центрифуге Бэкман J2-21 при скорости 8000 об/мин на роторе JA-10. Получают отмытый осадок агрегированного g-интерферона. Этот осадок ресуспендируют в 1400 мл 7 М раствора мочевины, pH 5,0 с добавлением 0,05% цетавлона при температуре 8oC и инкубируют в течение 2 ч. К суспензии добавляют натрий хлористый до концентрации 50 мМ, трис-HCl до 50 мМ, pH 8,0 и центрифугируют в течение 40 мин на центрифуге Бэкман J2-21 при скорости 10000 об/мин на роторе JA-10 при 4oC. Супернатант собирают, инкубируют при 4oC 18-20 ч и повторно центрифугируют в течение 20 мин на центрифуге J2-21 при скорости 10000 об/мин на роторе JA-10 при 4oC. Получают мочевинный раствор интерферона. Мочевинный раствор g-интерферона разбавляют в 10 раз водой и инкубируют при температуре 8oC в течение 4 ч. Затем добавляют ЭДТА pH 8,0 до концентрации 5 мМ, сульфат аммония до 0,2 М и инкубируют 20-24 ч. Выпавший осадок удаляют фильтрованием, а раствор g-интерферона концентрируют на установке с полыми волокнами в 10 раз и фракционируют сульфатом аммония. Фракцию белка, выпавшую при добавлении сульфата аммония до 300 г на 1 л раствора и инкубацией с перемешиванием при температуре 8oC в течение 2 ч, удаляют центрифугированием в течение 20 мин на центрифуге Бэкман A2-21 при скорости 8000 об/мин на роторе JA-10. Фракцию g-интерферона, осажденную при добавлении еще 200 г сульфата аммония на 1 л исходного раствора и инкубации с перемешиванием при температуре 8oC в течение 2 ч, собирают центрифугированием в течение 20 мин на центрифуге Бэкман J2-21 при скорости 8000 об/мин на роторе JA-10. Величину pH раствора в процессе сульфатного осаждения поддерживают в районе 7,5. Осадок растворяют в 1000 мл буфера (0,1 М аммония ацетата, pH 7,5). Получают ренатурированный раствор  -интерферона.
-интерферона.
Для осаждения примесей из ренатурированного раствора g-интерферона можно использовать двух-, трехразрядные анионы органических и неорганических кислот, такие как анионы серной, фосфорной, лимонной кислот в области величин pH от 7 до 9.
Этап 3. Хроматографическая очистка g-инерферона.
Полученный раствор белков наносят на хроматографическую колонку К100/45 "Pharmacia" (Швеция), содержащую 3000 мл КМ-52 целлюлозы, уравновешенную буфером 0,1 М аммония ацетата, о,1 М натрия хлористого, pH 7,5 и пропускают уравновешивающий буфер до достижения базовой линии контрольного оборудования для детекции белков. Фракцию g-интерферона элюируют пропусканием буфера: 0,1 М аммония ацетата, 0,18 М натрия хлористого, pH 7,5. Этот и все предыдущие растворы стерилизуют и очищают от пирогенов ультрафильтрацией, а весь процесс с этой стадии проводят в стерильных условиях при температуре 8oC. Скорость элюции на колонке 3000 мл/ч. Собирают фракции g-интерферона, не содержащие видимых примесей по данным электрофореза в 15% полиакриламидном геле с додецилсульфатом натрия при нанесении 30 мкг белка на лунку и окрашиванием геля красителем кумаси R250. В результате объединения фракций, содержащих электрофоротически чистый g-интерферон, получают 2000 мл раствора белка с концентрацией 1,05 мг/мл и удельной биологической активностью 1,5•107 МЕ/мг белка. Доля g-интерферона, находящегося в виде димерной формы при электрофорезе в 15% полиакриламидном геле с додецилсульфатом натрия, составляет не более 2% Отношение величины поглощения раствора белка при длине волны 280 нм к поглощению при 250 нм равно 2,6-2,7.
В табл.2 приведены данные о выходе белка g-интерферона на этапах: биосинтез (исходная биомасса), выделение и очистка (выделение тел включения, растворение тел включений, ренатурация) и хроматографическая очистка белка.
Пример 4. Очистка g-интерферона с использованием обращенно-фазной высокоэффективной хроматографии. Этап 1) биосинтез g-интерферона; этап 2) выделение и очистка g-интерферона; этап 3) очистка гамма-интерферона с помощью высокоэффективной обращенно-фазной хроматографии; этап 4) ренатурация и ионно-обменная хроматография g-интерферона.
Этап 1. Осуществляется, как в примере 3.
Этап 2. Выделение и очистка g-интерферона.
150 г замороженной при температуре минус 70oC биомассы E.coli T3q, продуцента g-интерферона ресуспендируют в 1500 мл буфера (50 мМ натрия фосфорнокислого двузамещенного, 1 мМ ЭДТА, 0,4 М натрия хлористого) при температуре 8oC и центрифугируют в течение 20 мин на центрифуге Бэкман J2-21 при скорости 8000 об/мин на роторе JA-10. Получают отмытый осадок клеток. Осадок клеток повторно ресуспендируют в 450 мл того же буфера, добавляют 150 мл лизоцима и инкубируют с перемешиванием при температуре 8oC в течение 30 мин. Обработанную лизоцимом суспензию замораживают при минут 20oC в течение 3 ч и размораживают при температуре 25oC. Вязкость суспензии снижают добавлением ДНКазы 5 мг магния сернокислого до концентрации 5 мМ инкубацией в течение одного часа. После добавления равного объема 8% раствора Triton X-100 в буфере (50 мМ натрия фосфорнокислого двузамещенного, 1 мМ ЭДТА, 0,4 М натрия хлористого) и инкубации в течение 1 ч суспензию центрифугируют в течение 20 мин на центрифуге Бэкман J2-21 при скорости 8000 об/мин на роторе JA-10.
Полученный осадок ресуспендируют в 150 мл буфера (50 мМ натрия фосфорнокислого двузамещенного, 1 мМ ЭДТА, 0,4 М натрия хлористого), добавляют 1250 мл буфера (50 мМ натрия фосфорнокислого трехзамещенного, 1 мМ ЭДТА) и инкубируют с перемешиванием в течение 20 мин, после чего центрифугируют в течение 20 мин на центрифуге Бэкмана J2-21 при скорости 8000 об/мин на роторе JA-10. Получают отмытый осадок агрегированного g-интерферона. Этот осадок ресуспендируют в 1400 мл 7 М раствора мочевины, pH 5,0 с добавлением 0,05% цетавлона при температуре 8oC и инкубируют в течение 2 ч. К суспензии добавляют натрий хлористый до концентрации 50 мМ, трис-HCl до 50 мМ, pH 8,0 и центрифугируют в течение 40 мин на центрифуге Бэкман J2-21 при скорости 10000 об/мин на роторе JA-10 при 4oC. Супернатант собирают, инкубируют при 4oC 18-20 ч и повторно центрифугируют в течение 20 мин на центрифуге J2-21 при скорости 10000 об/мин на роторе JA-10 при 4oC. Получают мочевинный раствор интерферона.
Этап 3. Очистка g-интерферона с помощью высокоэффективной обращенно-фазной хроматографии.
Мочевинный раствор g-интерфорона наносят на колонку размером 25x250, заполненную сорбентом Диасорб 130 С8/Т, 6 мкм, подключенную к установке для градиентной высокоэффективной жидкостной хроматографии фирмы Gilson, которая включает в себя два насоса серии 305, манометрический модуль 805, миксер 811 B, проточный денситометр 115. Колонку перед нанесением g-интерферона промывают 3 объемами 8 М раствора мочевины с pH 4,0. После нанесения препарата g-интерферона на колонку через нее пропускают 2 объема 8 М раствора мочевины с pH 4,0. Элюцию проводят линейным градиентом изопропиловый спирт 8 М раствор мочевины с pH 4,0 от 0 до 50% в течение 40 мин при температуре 25oC. g-интерферон элюируется при концентрации изопропилового спирта 30-34% Собирают фракции g-интерферона, не содержащие существенных примесей по данным электрофореза в 15% полиакриламидном геле с додецилсульфатом натрия при нанесении 30 мкг белка на лунку и окрашиванием геля красителем кумаси R250. В результате объединения фракций, содержащих электрофоретически чистый g-интерферон, получают 1200 мл раствора белка с концентрацией 2,4 мг/мл.
Этап 4. Ренатурация и ионно-обменная хроматография g-интерферона.
Величину pH мочевинного раствора g-интерферона доводят 2 М раствором Tris-base до 7,5, раствор разбавляют в 10 раз водой и инкубируют при температуре 8oC в течение 4 ч. Затем добавляют ЭДТА pH 7,5 до концентрации 5 мМ, аммоний ацетат до 0,1 М и инкубируют 20-24 ч. Выпавший осадок удаляют фильтрованием, а раствор g-интерферона концентрируют на установке с полыми волокнами в 10 раз. Получают ренатурированный раствор g-интерферона.
Полученный раствор белков наносят на хроматографическую колонку К100/45 "Pharmacia" (Швеция), содержащую 3000 мл КМ-52 целлюлозы, уравновешенную буфером 0,1 М аммония ацетата, 0,1 М натрия хлористого, pH 7,6 и пропускают уравновешивающий буфер до достижения базовой линии контрольного оборудования для детекции белков. Фракцию g-интерферона элюируют пропусканием буфера: 0,1 М аммония ацетата, 0,18 М натрия хлористого, pH 7,5. Этот и все предыдущие растворы стерилизуют и очищают от пирогенов ультрафильтрацией, а весь процесс с этой стадии проводят в стерильных условиях при температуре 8oC. Скорость элюции на колонке 3000 мл/ч. Собирают фракции g-интерферона, не содержащие видимых примесей по данным электрофореза в 15% полиакриламидном геле с додецилсульфатом натрия при нанесении 30 мкг белка на лунку и окрашиванием геля красителем кумаси R250. В результате объединения фракций, содержащих электрофоретически чистый гамма-интерферон, получают 2400 мл раствора белка с концентрацией 0,9 мг/мл и удельной биологической активностью 1,5•107 МЕ/ мл белка. Доля гамма-интерферона, находящегося в виде димерной формы при электрофорезе в 15%-ном полиакриламидном геле с додецилсульфатом натрия, составляет не более 0,1% Отношение величины поглощения раствора белка при длине волны 280 нм к поглощению при 250 нм равно 2,7.
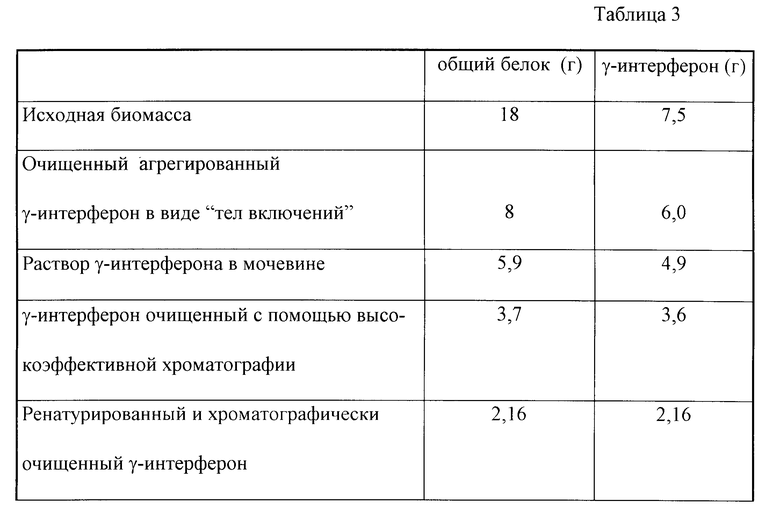
В табл. 3 приведены данные о выходе белка гамма-интерферона на этапах: биосинтез (исходная биомасса), выделение и очистка (выделение тел включения, растворение тел включений, высокоэффективная хроматография, ренатурация и хроматографическая очистка белка).
Таким образом, предлагаемая рекомбинантная плазмида, а также штамм, содержащий эту плазмиду продуцент рекомбинантного иммунного интерферона человека, позволяют значительно упростить процесс крупномасштабного получения гамма-интерферона при сохранении высокого уровня биосинтеза. Предлагаемый способ выделения и очистки позволяет получать биологический активный ремокбинантный иммунный интерферон человека не менее 98% чистоты и содержащий не более 2% гамма-интерферона, находящегося в виде димерной формы при электрофорезе в 15% полиакриламидном геле с додецилсульфатом натрия, а в случае использования высокоэффективной обращенно-фазной хроматографии не более 0,1% в объемах, достаточных для приготовления производственных партий лекарственных препаратов. Выход целевого продукта составляет не менее 30%


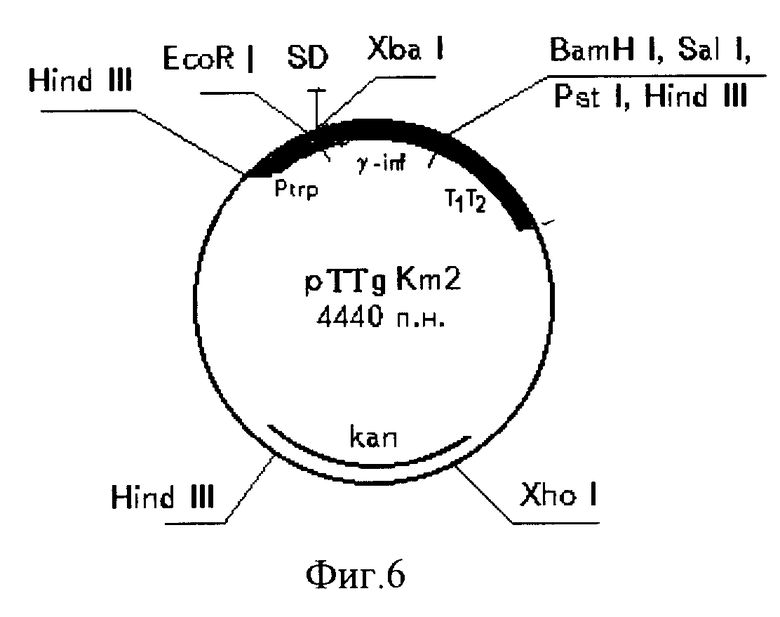
Сконструирована рекомбинантная ДНК pTTg Km2, кодирующая синтез γ-интерферона человека, содержащая триптофановый промотор из генома E. Coli, синтетическую последовательность участка связывания рибосом (последовательность Шайн-Дальгарно, фланкированная сайтами рестрикции EcoRI и xbaI), фрагмент кДНК, кодирующий γ-интерферон человека, терминатор транскрипции rrn BTT, ген устойчивости к канамицину в качестве генетического маркера и фрагмент векторной плазмиды pUC19. В результате трансформации реципиентного штамма E. coli C600 StrR рекомбинантной плазмидой pTTg Km2 получен штамм E. coli T3g - продуцент рекомбинантного γ-интерферона человека с продуктивностью до 1,2•1010 ME/л культуральной жидкости. Разработан способ получения рекомбинантного γ-интерферона человека, основанный на использовании созданного рекомбинантного штамма E. coli T3g и предусматривающий глубинное культивирование на питательной среде с пониженным содержанием триптофана, растворение агрегированного белка в концентрированном растворе мочевины с цетавлоном, ренатурацию и очистку от примесей путем обработки солью с двух- или трехвалентными анионами и последующей хроматографии раствора на анионообменнике. 3 с. и 1 з.п. ф-лы, 6 ил., 3 табл.
5' GAATCCAGGAGGAATCTAGA 3'
3' CTTAAGTCCTCCTTAGATCT 5',
Xba I Bam HI фрагмента ДНК размером 450 п.о. кодирующего иммунный интерферон человека с аминокислотной последовательностью, приведенной на фиг.5; Bam HI-Sca I фрагмента ДНК размером 850 п.о. плазмиды РКК 233-3, содержащего последовательность терминатора транскрипции rrn ВТТ; Pst I-Pst I фрагмента ДНК размером 1500 п.о. плазмиды pUC 4K, кодирующего устойчивость к канамицину (kan); Bgl I-Hind III фрагмента ДНК размером 1400 п.о. плазмиды pUC 19.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| SU, авторское свидетельство, 1433019, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Peret etal., Appl.Microbiol | |||
| Biotechnol., v | |||
| Способ сопряжения брусьев в срубах | 1921 |
|
SU33A1 |
| Способ получения снабженных окрашенными узорами формованных изделий из естественных или искусственных смол | 1925 |
|
SU429A1 |

