Область техники, к которой относится изобретение
Настоящее изобретение касается модифицированного сигнального пептида энтеротоксина-II E. coli, гена, кодирующего упомянутый пептид, вектора, включающего упомянутый ген, химеризованный на ген, кодирующий гетерологичный белок, микроорганизма, трансформированного упомянутым вектором, и способа получения гетерологичного белка с использованием упомянутого микроорганизма.
Предпосылки изобретения
Многие гетерологичные белки получают с применением преобразованных с помощью генной инженерии микроорганизмов-хозяев с использованием "внутриклеточного способа" или "секреторного способа".
Во "внутриклеточном способе" ген, кодирующий гетерологичный белок, экспрессируется и накапливается в цитоплазме микроорганизма. Хотя данный способ, как известно, дает относительно высокий выход гетерологичного белка, экспрессированный гетерологичный белок не находится в естественной активной форме из-за наличия остатка метионина на его N-конце. Более того, биологически неактивный гетерологичный белок, выработанный в таком способе, часто образует нерастворимые включенные тельца, которые должны быть подвергнуты солюбилизации и конверсии в натурализованную активную форму с помощью процесса рефолдинга.
Что касается "секреторного способа", то ген, кодирующий слитый белок, состоящий из сигнального пептида и гетерологичного белка, экспрессируется в цитоплазме микроорганизма, а затем слитый белок процессируется с участием сигнальной пептидазы этого микроорганизма с удалением сигнального пептида в ходе прохождения через цитоплазматическую мембрану. Процессированный белок секретируется в периплазматическое пространство между цитоплазматической (внутренней) мембраной и внешней мембраной данного микроорганизма. Однако известно, что данный способ дает существенно меньший выход гетерологичного белка по сравнению с "внутриклеточным способом". Следовательно, имеется необходимость в улучшении продуктивности "секреторного способа". В связи с этим, сообщалось, что точное и эффективное отщепление составляющей в виде сигнального пептида от экспрессированного слитого белка под действием сигнальной пептидазы является важным с точки зрения повышения выхода секретируемого гетерологичного белка (M. Akita et al., 1990, J. Biol. Chem., 265, 8164).
В целом, сигнальные пептиды подразделяют на две группы - гидрофильные сигнальные пептиды и гидрофобные сигнальные пептиды. Гидрофильный сигнальный пептид обычно состоит из 12-70 аминокислот. Типичный гидрофобный сигнальный пептид, например сигнальный пептид энтеротоксина-II E.coli, состоит из 13-30 аминокислот и составлен тремя участками: N-концевым гидрофильным сегментом, включающим одну или две основные аминокислоты, центральным гидрофобным сегментом, включающим примерно 10 основных аминокислот, и С-концевым гидрофильным сегментом, включающим аминокислоты с небольшими боковыми цепями.
Поскольку гетерологичный белок, экспрессированный в форме слитого белка с сигнальным пептидом, обычно быстро разрушается цитоплазматическими протеиназами, выход секретируемого гетерологичного белка уменьшается по мере снижения секреторной эффективности данного сигнального пептида. Следовательно, выход секретированных гетерологичных белков может быть повышен путем модифицирования сигнальной пептидной составляющей слитых белков, экспрессированных в микроорганизмах-хозяевах.
Ген гормона роста человека (hGH) состоит из 191 аминокислоты и имеет молекулярную массу 21500 Д. После того, как впервые очищенная форма hGH была выделена из гипофиза человека в 1956 году (Li & Papkoff, 1956, Science, 124, 1293), было проведено значительное число исследований hGH с целью оценки, например, влияния hGH на метаболизм в организме человека (J.С. Beck et al., 1957, Science, 125, 884) и активности hGH по подавлению гипофизарной карликовости (M. S. Raben, 1958, J. Clin. Endocrinol., 18, 901). Недавно было сообщено, что hGH также эффективен в лечении синдрома Тернера, остеопороза, ран и ожогов.
Поскольку количество hGH, получаемого из гипофиза человека, ограничено, были предприняты попытки выработать значительное количество hGH в генетически преобразованных клетках E.coli с применением "внутриклеточного способа" (D.V. Goeddel et al., 1979, Nature, 281, 544). Однако данный способ отягощен упоминавшейся выше проблемой выработки метионилированного hGH, который непригоден для применения в отношении человека. Дальнейшая попытка удалить метионин из состава метионилированного hGH с использованием дипептидиламинопептидазы-I привела к неприемлемо низкому выходу hGH.
Соответственно, была предпринята попытка секреторной выработки нативного hGH. Например, в европейских патентах 55942, 20147 и 114695 описаны способы экспрессии нативной формы hGH и выделения его с помощью секреции. Однако доступное для выделения количество выработанного в таких способах hGH оказывается минимально возможным.
Европейский патент 177343 описывает способ выработки hGH, который включает экспрессию гена, кодирующего слитый белок из hGH и сигнального пептида щелочной фосфатазы или энтеротоксина, в присутствии индуктора экспрессии - изопропилтио-β-D-галактозида (IPTG) и секрецию hGH в периплазму. Однако этот способ дает низкий выход hGH и требует использования дорогого экспрессионного индуктора - IPTG.
Следовательно, существует необходимость в разработке нового эффективного способа выработки hGH с высоким выходом.
Краткое содержание изобретения
Таким образом, целью настоящего изобретения является представление модифицированного сигнального пептида энтеротоксина-II E.coli, который может быть предпочтительно использован в "секреторном способе" выработки гетерологичного белка с целью повышения эффективности секреции.
Другой целью настоящего изобретения является представление гена, кодирующего упомянутый пептид.
Следующей целью настоящего изобретения является представление вектора, включающего упомянутый ген, химеризованный на ген, кодирующий гетерологичный пептид.
Еще одной целью настоящего изобретения является представление микроорганизма, трансформированного упомянутым вектором.
Следующей целью настоящего изобретения является представление способа получения гетерологичного белка с использованием упомянутого микроорганизма.
В соответствии с одним из аспектов настоящего изобретения представляется модифицированный сигнальный пептид энтеротоксина-II E.coli (обозначенный MST), отличающийся тем, что по крайней мере одна из аминокислот в положениях 2, 4, 5, 12, 20 и 22 сигнального пептида энтеротоксина-II E.coli, представленного следующей аминокислотной последовательностью (SEQ ID NO 1), заменена на другую аминокислоту при условии, что по крайней мере одна из аминокислот в положениях 2 и 4 MST является лизином:
Краткое описание чертежей
Названные выше цели и отличительные признаки настоящего изобретения станут ясны из нижеследующего описания предпочтительных вариантов, принимаемых вместе с прилагаемыми чертежами, в которых:
на фигуре 1 показана процедура конструирования вектора pT-hGH;
на фигуре 2 изображена процедура конструирования векторов рUC19ST и pUC19SH;
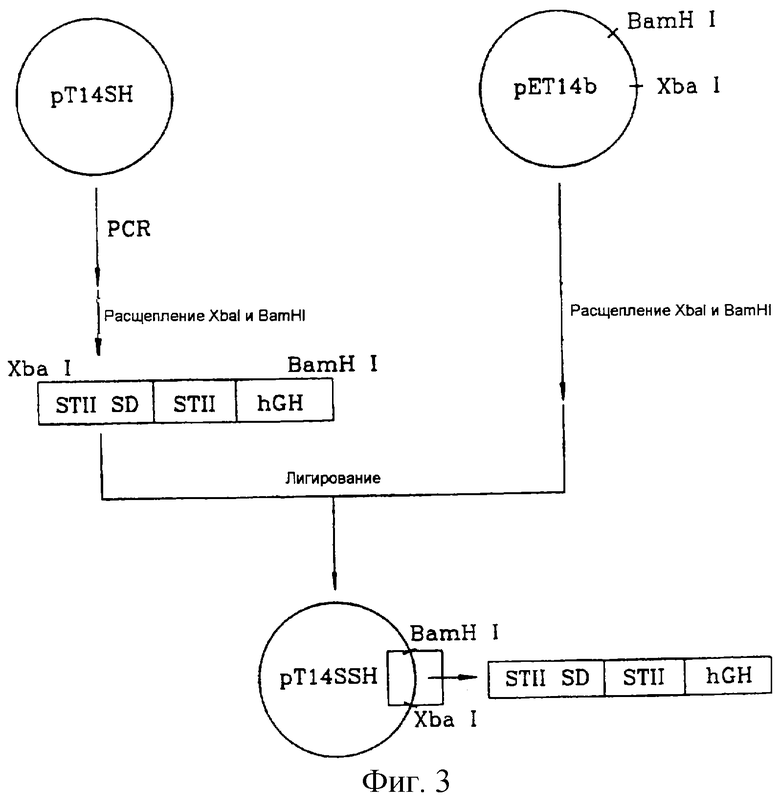
на фигуре 3 представлена процедура конструирования вектора pT14SSH;
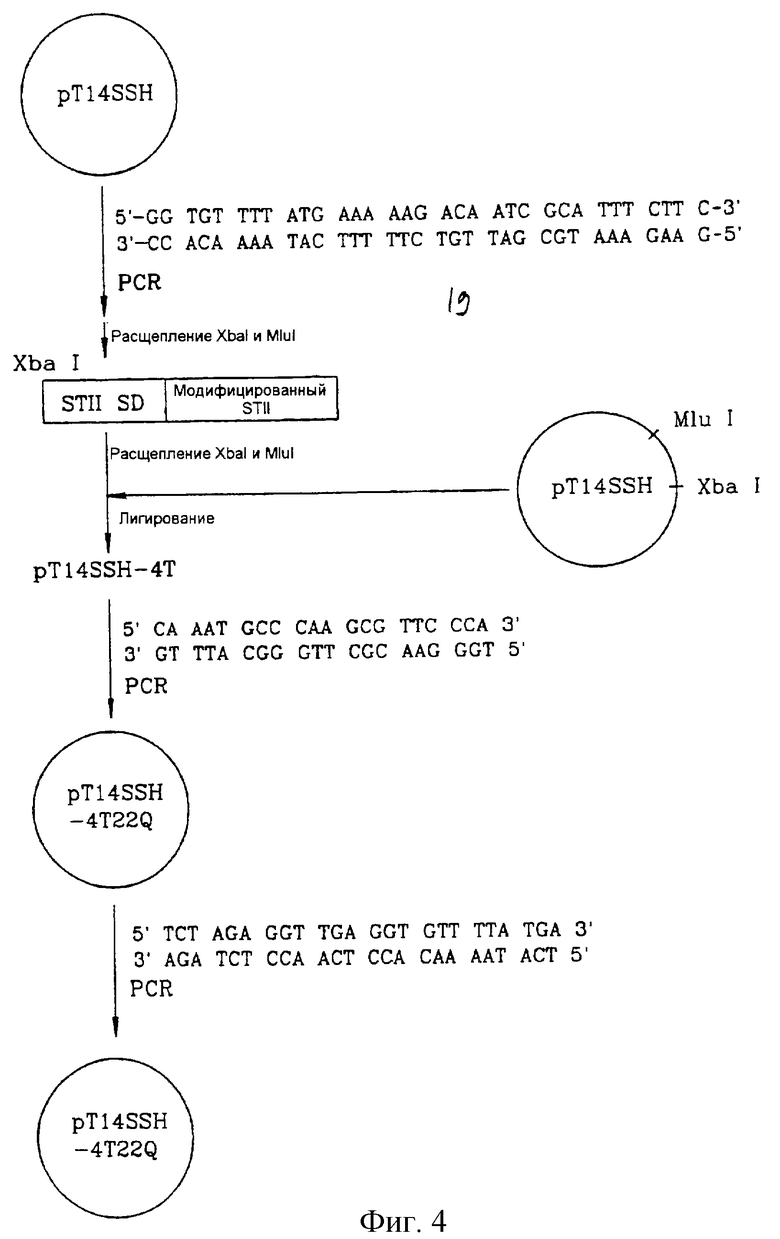
на фигуре 4 показана процедура конструирования вектора рT14S1SH;
на фигуре 5 воспроизведены результаты анализа очищенного hGH методом ДСН-ПААГ.
Подробное описание изобретения
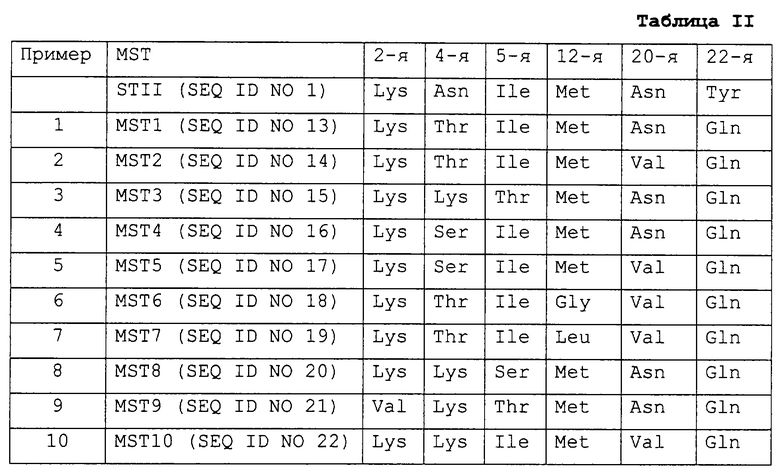
Среди модифицированных сигнальных пептидов энтеротоксина-II E. coli (MSTs) по настоящему изобретению предпочтительными являются те, в которых:
2-я аминокислота Lys не заменена;
4-я аминокислота Asn заменена на Ser, Thr, Lys или Gln;
5-я аминокислота Ilе не заменена или заменена на Thr или Ser;
12-я аминокислота Met не заменена или заменена на Аlа, Gly, Val, Leu или Ile;
20-я аминокислота Asn не заменена или заменена на Ilе, Phe, Ala или Val;
22-я аминокислота Туr не заменена или заменена на Gln, Asn, Ala или Lys.
Также предпочтительными являются те, в которых:
2-я аминокислота Lys заменена на любую другую аминокислоту;
4-я аминокислота Asn заменена на Lys;
5-я аминокислота Ilе заменена на Ser, Thr, Asn, Gln или Arg;
12-я аминокислота Met не заменена или заменена на Ala, Gly, Val, Leu или Ile;
20-я аминокислота Asn не заменена или заменена на Ilе, Phe, Ala или Val;
22-я аминокислота Туr не заменена или заменена на Gln, Asn, Ala или Lys.
Наиболее предпочтительными MST являются те, которые характеризуются одним из следующих наборов аминокислотных замен:
(a) 4-я Asn на Thr и 22-я Туr на Gln;
(b) 4-я Asn на Thr, 20-я Asn на Val и 22-я Туr на Gln;
(c) 4-я Asn на Lys, 5-я Ilе на Thr и 22-я Туr на Gln;
(d) 4-я Asn на Ser и 22-я Туr на Gln;
(e) 4-я Asn на Ser, 20-я Asn на Val и 22-я Туr на Gln;
(f) 4-я Asn на Thr, 12-я Met на Gly, 20-я Asn на Val и 22-я Туr на Gln;
(g) 4-я Asn на Thr, 12-я Met на Leu, 20-я Asn на Val и 22-я Туr на Gln;
(h) 4-я Asn на Lys, 5-я Ilе на Ser и 22-я Туr на Gln;
(i) 2-я Lys на Val, 4-я Asn на Lys, 5-я Ilе на Thr и 22-я Туr на Gln;
(j) 4-я Asn на Lys, 20-я Asn на Val и 22-я Туr на Gln.
MST по настоящему изобретению может кодироваться геном, включающим нуклеотидную последовательность, определенную по аминокислотной последовательности MST в соответствии с генетическим кодом. Известно, что по несколько разных кодонов, кодирующих одну и ту же аминокислоту, может существовать благодаря эффекту вырожденности генетического кода, а следовательно, MST по настоящему изобретению включает все нуклеотидные последовательности, установленные по аминокислотной последовательности MST. Предпочтительно ген MST может включать один или большее число предпочтительных кодонов E.coli.
Ген MST может быть получен путем мутирования одного или большего числа нуклеотидов нативного гена сигнального пептида энтеротоксина-II E.coli (обозначенного как ген STII) с применением сайт-специфичного (направленного) мутагенеза (С. Papworth et al., 1996, Strategies, 9, 3). Ген STII E.coli может быть выделен с помощью стандартного метода (Sambrook et al., 1989, "Molecular Cloning: A Laboratory Manual", 2d ed., Cold Spring Harbor Lab. ress). Кроме того, ген MST может быть синтезирован химическим путем.
MST по настоящему изобретению, в случае его химеризации на гетерологичный белок, обеспечивает высокоэффективную секрецию гетерологичного белка через цитоплазматическую мембрану микроорганизма, например E.coli. Соответственно, используя экспрессирующий вектор, который включает ген MST, химеризованный на ген, кодирующий гетерологичный белок, слитый белок из MST и гетерологичного белка (обозначенный как MST/гетерологичный белок) может быть преимущественно экспрессирован в цитоплазме E.coli, т.к. слитый белок эффективно процессируется за счет удаления составляющей MST, что приводит к быстрому выходу гетерологичного белка в периплазму E.coli. Таким образом, использование заявляемого MST приводит к значительно повышенному уровню выработки гетерологичного белка.
Химеризация гена MST и гена, кодирующего гетерологичный белок, может быть осуществлена в соответствии со стандартным методом лигирования (Sambrook et al., цит. выше).
Репрезентативные гетерологичные белки включают гормон роста человека (hGH), колония-стимулирующий фактор гранулоцитов (G-CSF), интерферон, интерлейкин, про-урокиназу, инсулин, фактор VIII свертывания крови, гирудин, супероксиддисмутазу и кальцитонин, однако это не ограничивает те гетерологичные белки, которые могут быть использованы в настоящем изобретении. Ген, кодирующий гетерологичный белок, может быть выделен с помощью стандартного метода, например путем скрининга библиотеки кДНК и ПЦР.
Экспрессирующий вектор по настоящему изобретению дополнительно может включать модифицированную последовательность Шайна-Дальгано энтеротоксина-II E. coli (модифицированная последовательность STII-SD) со следующей нуклеотидной последовательностью (SEQ ID NO 2), встроенной непосредственно перед старт-кодоном гена MST:
5'-GAGGTGTTTT-3'.
Модифицированная последовательность STII-SD составлена 4-нуклеотидной последовательностью STII-SD (GAGG) и 6-нуклеотидной Т-богатой последовательностью. Последовательность STII-SD в модифицированной последовательности STII-SD представляет собой очень мощный сайт связывания на рибосоме, который повышает уровень экспрессии в отсутствие индуктора экспрессии, например изопропилтио-β-D-галактозида (IPTG). Т-обогащенная последовательность модифицированной последовательности STII-SD играет роль в предотвращении образования вторичных структур транскрибированной с нее мРНК, что тем самым повышает эффективность экспрессии. Модифицированная последовательность STII-SD может быть получена стандартными методами (Sambrook et al., цит. выше), например с помощью химического синтеза. Кроме того, ген STII-SD, характеризующийся следующей нуклеотидной последовательностью (SEQ ID NO 3), может быть подвергнут направленному мутагенезу с целью получения модифицированной последовательности STII-SD:
5'-GCTCTAGAGGTTGAGGTGTTTTATGAAAAAGAATA-3'.
Модифицированная последовательность STII-SD может быть встроена впереди от старт-кодона ATG гена MST или может быть модифицирована последовательность STII-SD, предшествующая старт-кодону ATG гена MST.
Примерами экспрессирующих векторов по настоящему изобретению являются векторы pTG14S1SH-4T22Q, pT14S1SH-4T20V22Q, рTG14S1SH-4K5T22Q, pTG14S1SH-4S22Q, pTG14S1SH-4S20V22Q, pTG14S1SH-4T12G20V22Q, pTG14S1SH-4T12L20V22Q, pTG14SSH-4K5S22Q, pTG14SSH-2V4K5T22Q и pTG14SSH-4K20V22Q, которые получают в примерах 1-10, а предпочтительными векторами являются рT14S1SH-4T22Q и pT14S1SH-4T20V22Q.
Экспрессирующие векторы по настоящему изобретению могут быть внесены в микроорганизм, например E. coli, в соответствии со стандартным методом трансформации (Sambrook et al., цит. выше). Среди трансформированных микроорганизмов предпочтительными являются трансформанты E. coli HM10011 и НМ10012, которые были депонированы в Корейский центр культивируемых микроорганизмов (КССМ) (адрес: Dept. Food Engineering, Coll. Engineering, Yonsei Univ. , Sodaemungu, Seoul 120-749, Rep. Korea) 12 августа 1998 г. под депозитарными КССМ-10137 и КССМ-10138 соответственно в соответствии с положениями Будапештского Соглашения по международной сертификации депозитариев микроорганизмов для целей патентования.
Гетерологичный белок может быть получен путем культивирования микроорганизма-трансформанта с целью экспрессии гена, кодирующего химеру МSТ/гетерологичный белок, и секреции гетерологичного белка в периплазму и выделения гетерологичного белка из периплазмы. Микроорганизм-трансформант можно культивировать в соответствии со стандартным методом (Sambrook et al., цит. выше). Культуру микроорганизма можно отцентрифугировать или профильтровать с целью сбора микроорганизма, секретирующего гетерологичный белок. Трансформированный микроорганизм можно разрушить в соответствии со стандартным методом (F.M. Ausubel et al., 1989, "Current Protocols in Molecular Biology") с целью получения раствора периплазмы. Например, микроорганизм может быть разрушен в гипотоническом растворе, например в дистиллированной воде, за счет осмотического шока. Выделение гетерологичного белка из раствора периплазмы может быть осуществлено с помощью стандартного метода (Sambrook et al., цит. выше), например с помощью ион-обменной хроматографии, гель-фильтрационной хроматографии на колонках или иммунологической хроматографии на колонках. Например, hGH может быть очищен последовательно с помощью хроматографии на колонках с DEAE-сепарозой, хроматографии на колонках с фенилсепарозой и хроматографии на колонках Сефадекс G-100.
Гетерологичный белок, полученный в соответствии с настоящим изобретением, находится в нативной форме, не имеет остатка метионина на N-конце, и, следовательно, его можно использовать для различных практических целей.
Следующие примеры призваны дополнительно проиллюстрировать настоящее изобретение без ограничения его объема.
Пример получения 1. Скрининг кДНК гена гормона роста человека
Стадия 1. Конструирование библиотеки кДНК гипофиза человека
К 1 г гипофиза человека добавляли 10 мл раствора гуанидина (4 М гуанидинизоцианата, 50 мМ Трис-HCl, рН 7,5, 10 мМ ЭДТА и 5% 2-меркаптоэтанола) и гомогенизировали. Гомогенат центрифугировали при 10000 об/мин в течение 10 минут при 6oС. К надосадочной фракции добавляли 0,1 объема 2% саркозилового эфира (Sigma, США) и полученную смесь выдерживали при 65oС в течение 2 минут. Хлорид цезия добавляли к полученному в результате раствору до концентрации 0,1 г/мл и полученную смесь центрифугировали при 25000 об/мин в течение 16 часов с 9 мл буфера (5,7 М CsCl и 0,1 мМ ЭДТА) с получением РНК-преципитата. Преципитат растворяли в 3 мл суспензионного раствора (5 мМ ЭДТА, 0,5% саркозила и 5% меркаптоэтанола) и затем экстрагировали последовательно смесью фенола, хлороформа и изоамилового спирта (25:24:1 - об/об/об) и смесью хлороформа и изоамилового спирта (24: 1 - об/об). К объединенным экстрактам добавляли 0,1 объема 3 М ацетата натрия и 2,5 объема этанола, а полученную смесь центрифугировали с использованием стандартного метода (Sambrook et al. , цит. выше) с получением РНК-преципитата. РНК-преципитат растворяли в дистиллированной воде (a.d.) и выдерживали при 70oС в течение 10 минут. К этому раствору добавляли хлорид лития до концентрации 0,5 М и затем подвергали хроматографии с олиго-дТ-целлюлозой (тип 3: Collaboratory Research, США) с выделением полиаденилированной РНК в соответствии с методом Авива-Ледера (H. Aviv & P. Leder, 1972, J. Mol. Biol., 134, 743). Полученную таким образом поли(А)-РНК обрабатывали при 65oС в течение 5 минут, охлаждали до 0oС и немедленно добавляли 20 мкл 5 мМ dNTPs, 40 мкл 5х буферного раствора (0,25 М Трис-НСl - рН 8,3; 0,5 М КСl и 50 мМ MgCl2), 10 мкл 200 мМ дитиотреитола, 20 мкл 0,5 мг/мл олиго-дТ12-18 (Pharmacia Inc., Швеция), 80 мкл a. d., 10 мкл (10 ед.) RNAsin (Promega, США) и 20 мкл (20 ед.) обратной транскриптазы AMV (Life Science Inc., США). После реакции смеси при 42oС в течение 90 минут к реакционной смеси добавляли 5 мкл 0,5 М ЭДТА (рН 8,0) и 200 мкл Трис-забуференного фенола, перемешивали и центрифугировали при 10000 об/мин в течение 10 минут при комнатной температуре. Надосадочную фракцию дважды экстрагировали диэтиловым эфиром и объединенные экстракты смешивали с 20 мкл 3 М ацетата натрия и 1 мл 95%-ного этанола с осаждением одноцепочечных кДНК (оц-кДНК).
Для синтеза двухцепочечных кДНК (дц-кДНК) на матрице оц-кДНК преципитат оц-кДНК растворяли в 284 мкл a.d. и добавляли 40 мкл 5 мМ NTPs, 80 мкл 5х буферного раствора второй цепи (250 мМ Трис-НСl - рН 7,2; 450 мМ КСl, 15 мМ дитиотреитола, 15 мМ MgCl2 и 0,25 мг/мл бычьего сывороточного альбумина), 12 мкл 5 мМ β-NAD+, 2 мкл 3000 Ки/ммоль α-32Р-дЦТФ, 4 мкл (4 ед.) ДНК-лигазы E. coli и 10 мкл (100 ед.) ДНК-полимеразы I E.coli. После того, как реакционную смесь оставляли на 16 часов при 14oС, эту реакционную смесь подвергали фенольной экстракции и осаждению этанолом в соответствии с тем, что было описано выше, с получением преципитата дц-кДНК.
Для "затупления" концов дц-кДНК преципитат дц-кДНК растворяли в 42 мкл a. d. и добавляли к нему 5 мкл dNTPs, 16 мкл 5х буферного раствора второй цепи, 1 мкл 5 мМ β-NAD+, 4 мкл РНКазы-А (2 мкг/мл: Biolabs, США), 4 мкл (4 ед. ) РНКазы-Н, 2 мкл (20 ед.) ДНК-лигазы E.coli и 4 мкл (8 ед.) ДНК-полимеразы фага Т4, оставляя после этого реакционную смесь на 45 минут при 37oС. По завершении реакции реакционную смесь подвергали фенольной экстракции и осаждению этанолом в соответствии с описанным выше с получением преципитата дц-кДНК с "тупыми концами".
Для защиты рестрикционного EcoRI-сайта в составе дц-кДНК с помощью метилирования преципитат дц-кДНК с "тупыми концами" растворяли в 25 мкл a.d. и добавляли к нему 27 мкл 2х метилазного буфера (100 мМ NaCl, 100 мМ Трис-HCl - рН 8,0, и 1 мМ ЭДТА), 1 мкл 50х раствора SАМ (1 мг S-аденозилметионина в 0,14 мл ацетата натрия - рН 5,2) и 10 мкл (10 ед.) метилазы EcoRI (Biolabs, США). После того, как смесь оставляли реагировать в течение 2 часов при 37oС, реакционную смесь подвергали фенольной экстракции и осаждению этанолом в соответствии с описанным выше. Осажденную кДНК объединяли с EcoRI-линкером (Biolabs, США) и ДНК-лигазой Т4 и полученную смесь реагировали в течение 16 часов при 4oС с получением кДНК, лигированной на EcoRI-линкер.
Лигированную на EcoRI-линкер кДНК обрабатывали рестриктазой EcoRI и подвергали хроматографической очистке на колонке сефарозы CL-4B с целью удаления остаточных линкеров. Лигированную на EcoRI-линкер кДНК встраивали по EcoRI-сайту в состав λgt11 (Amersham, США). Полученный таким образом λgt11 подвергали упаковке in vitro с использованием набора для упаковки λ-фагов in vitro (Amersham, США) и полученной конструкцией трансфицировали клетки E.coli штамма Y1088 (АТСС 37195) с получением библиотеки кДНК гипофиза человека.
Стадия 2. Скрининг кДНК гена гормона роста человека
Для скрининга клонов гена гормона роста человека из библиотеки кДНК, сформированной на стадии 1, метод гибридизации бляшек осуществляли следующим образом.
Основываясь на опубликованной аминокислотной последовательности N-концевой части гормона роста человека (W.K. Liu et al., 1964, Biochem. Biophys. Acta, 93, 428; C.H. Li et al., 1966, J. Amer. Chem. Soc., 88, 2050), был выстроен и синтезирован 30-нуклеотидный фрагмент олигонуклеотидного зонда со смешанной последовательностью, представленный следующей нуклеотидной последовательностью:
Phe Pro Thr Ile Pro Leu Ser Arg (SEQ ID NO 4)
5'-TTCCCAACCATTCCCTTATCCAGG-3' (SEQ ID NO 5).
Первичную гибридизацию с бляшками проводили с использованием олигонуклеотидного зонда со смешанной последовательностью в соответствии с методом Бентона с соавт. (W.E. Benton et al., 1977, Science, 196, 180) с получением позитивных клонов. Эти клоны подвергали гибридизации с бляшками во второй и в третий раз с получением клона, включающего кДНК гена гормона роста человека.
Для подтверждения того, что этот клон включает ген гормона роста человека, клонированную фаговую ДНК расщепляли рестриктазой EcoRI и затем полученные ДНК-фрагменты подвергали Саузерн-блоттингу (Е. Southern, 1975, J. Mol. Biol., 98, 503) с использованием олигонуклеотидного зонда со смешанной последовательностью. После этого 650-нуклеотидный EcoRI-фрагмент, включающий ген гормона роста человека, встраивали по EcoRI-сайту вектора M12mp18 (Pharmacia, США) с получением вектора M13-hGH. Нуклеотидную последовательность гена гормона роста человека в векторе M13-hGH определяли с использованием метода Сэйнджера с терминацией цепи дидезоксирибонуклеотидами (F. Sanger et al., 1977, Proc. Natl. Acad. Sci. USA, 74, 5463-5467).
Пример получения 2. Получение гена, кодирующего зрелый гормон роста человека
Для формирования кДНК гена, кодирующего зрелый гормон роста человека, вектор M13-hGH, полученный на 2-й стадии примера получения 1, подвергали ПЦР с использованием следующих праймеров S1 и AS1. Прямой праймер S1 был сконструирован так, чтобы внести рестрикционный NdeI-сайт (5'-CATATG-3'), находящийся выше кодона 1-й аминокислоты (фенилаланин) зрелого гормона роста человека, а обратный праймер - так, чтобы внести рестрикционный BamHI-сайт (5'-GGATCC-3'), расположенный ниже его терминирующего кодона.
Прямой праймер S1 (SEQ ID NO 6):
5'-CCGCATATGTTCCCAACCATTCCC-3'.
Обратный праймер AS1 (SEQ ID NO 7):
5'-GCTGGATCCTAGAAGCCACAGCTGC-3'.
Амплифицированный ген гормона роста человека расщепляли рестриктазами NdeI и BamHI с целью получения гена, кодирующего зрелый гормон роста человека (обозначенный как ген hGH). Ген hGH встраивали по NdeI/BamHI-сегменту вектора pET14b (Novagen, США) с получением вектора pT-hGH.
На фиг.1 показана описанная выше процедура конструирования вектора pT-hGH.
Пример получения 3. Конструирование вектора, включающего ген, кодирующий слитый белок из сигнального пептида энтеротоксина-II E.coli и hGH
Стадия 1. Клонирование гена сигнального пептида энтеротоксина-II E.coli
Для получения гена сигнального пептида энтеротоксина-II E.coli следующую пару комплементарных олигонуклеотидов конструировали на основе нуклеотидной последовательности сигнального пептида энтеротоксина-II E.coli и синтезировали ее с помощью автоматического ДНК-синтезатора модели 380В (Applied Biosystem, США).
Олигонуклеотид смысловой цепи STII S1 (SEQ ID NO 8):
5'-TCATGAAAAAGAATATCGCATTTCTTCTTGCATCTATGTTCGTTTTTTCTATTGCTACAAATGCCTACGCGT-3'.
Олигонуклеотид антисмысловой цепи STII AS1 (SEQ ID NO 9):
5'-ACGCGTAGGCATTTGTAGCAATAGAAAAAACGAACATAGATGCAAGAAGAAATGCGATATTCTTTTTCATGA-3'.
Эти олигонуклеотиды были сконструированы так, чтобы включить рестрикционные NcoI- и BspHI-сайты выше старт-кодона энтеротоксина-II E.coli и рестрикционый MluI-сайт, внесенный путем несмысловой замены, в другом конце гена.
Оба олигонуклеотида отжигали при 95oС с целью получения двухцепочечных ДНК-фрагментов с "тупыми концами", характеризующихся нуклеотидной последовательностью, кодирующей сигнальный пептид энтеротоксина-II E. coli (ген. STII).
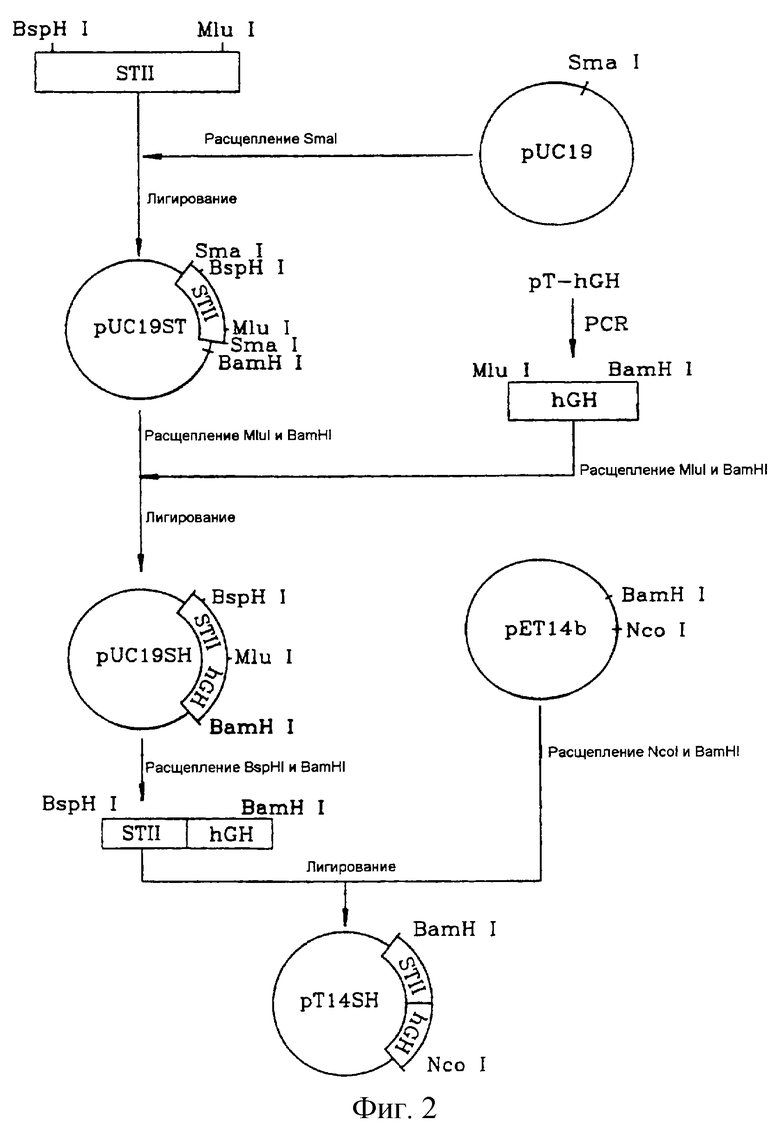
Ген STII был встроен по SmaI-сайту в состав вектора pUC19 (Biolabs, США) с получением вектора pUC19ST.
Стадия 2. Получение гена, кодирующего слитый белок STII/hGH
Для получения гена, кодирующего слитый белок STII/hGH, вектор pT-hGH, сконструированный в примере получения 2, подвергли ПЦР с использованием праймеров S2 и AS1, использованного в примере получения 2. Прямой праймер S2 был сконструирован так, чтобы включить рестрикционный MluI-сайт (5'-CATATG-3') выше кодона первой аминокислоты (фенилаланин) зрелого гормона роста человека.
Прямой праймер S2 (SEQ ID NO 10):
5'-GCGACGCGTTCCCAACCATTCCCTTATCC-3'.
Амплифицированные ДНК-фрагменты расщепляли рестриктазами MluI и BamHI и затем встраивали по MluI/BamHI-сегменту вектора pUC19ST, сконструированного на стадии 2. Сформированный таким путем вектор pUC19SH включал ген, кодирующий слитый белок STII/hGH (обозначен как ген STII-hGH).
На фиг.2 показана описанная выше процедура конструирования векторов pUC19ST и pUC19SH.
Стадия 3. Добавление к гену STII-hGH последовательности Шайна-Дальгано энтеротоксина-II E.coli
Вектор pUC19SH, полученный на 2-й стадии, расщепляли рестриктазами BspHI и BamHI с образованием 640-нуклеотидного фрагмента STII-hGH, который встраивали по NcoI/BamHI-сегменту вектора pET14b (Novagen, США) с получением вектора pT14SH.
Вектор pT14SH был подвергнут ПЦР с использованием праймеров S3 и AS3. Прямой праймер S3 сконструировали так, чтобы включить последовательность Шайна-Дальгано энтеротоксина-II E. coli (обозначена как последовательность STII-SD) и рестрикционный XbaI-сайт, а обратный праймер AS3 - так, чтобы внести рестрикционный BamHI-сайт, расположенный ниже терминирующего кодона зрелого hGH, с получением ДНК-фрагмента (STII-SD/STII-hGH), включающего слитый ген из STII-SD и STII-hGH.
Прямой праймер S3 (SEQ ID NO 11):
5'-GCTCTAGAGGTTGAGGTGATTTTATGAAAAAGAATA-3'.
Обратный праймер AS3 (SEQ ID NO 12):
5'-GGATGCCACGCTGGATCCTAGAAAGCCACAGCTGC-3'.
Фрагмент STII-SD/STII-hGH расщепляли рестриктазами XbaI и BamHI и затем встраивали по XbaI/BamHI-сегменту вектора pET14b (Novagen, США) с получением вектора pT14SSH. Клетки E.coli штамма BL21 (DE3) (Stratagene, США) трансформировали вектором pT14SSH с получением трансформанта, обозначенного как E.coli НМ10010.
На фиг. 3 представлена описанная выше процедура конструирования вектора pT14SSH.
Пример получения 4. Выработка hGH с использованием гена STII-hGH
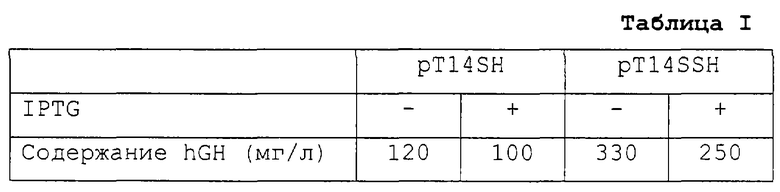
Для анализа влияния последовательности SD энтеротоксина-II E.coli на выработку hGH клетки E.coli BL21 (DE3), трансформированные вектором pT14SH, полученным на 3-й стадии примера получения 3, и клетки E.coli HM10010, полученные также на 3-й стадии примера получения 3, культивировали в присутствии и отсутствии индуктора экспрессии (IPTG) соответственно в среде LB (1% бактотриптона, 0,5% бактодрожжевого экстракта и 1% NaCl) при 37oС в течение 24 часов. Каждую из культур центрифугировали при 10000 об/мин в течение 10 минут для осаждения бактериальных клеток и полученный осадок суспендировали в 0,1 объема изотонического раствора (20% сахарозы, буферный раствор 10 мМ Трис-Cl, содержащий 1 мМ ЭДТА - рН 7,0). Суспензию оставляли при комнатной температуре на полчаса и затем центрифугировали при 10000 об/мин в течение 15 минут для сбора бактериальных клеток. Эти клетки ресуспендировали в a.d. при 4oС и центрифугировали при 12000 об/мин в течение 20 минут с получением надосадочной фракции в виде раствора периплазмы. Содержание hGH в растворе периплазмы оценивали с помощью метода ТИФА (K. Kato et al., 1976, J. Immunol., 116, 1554) с использованием антитела, специфичного в отношении hGH (Boehringer Mannheim), которое подсчитывали как количество hGH, выработанное в расчете на 1 л культуры. Полученные результаты показаны в табл.I.
Как можно видеть из табл.I, вектор pT14SSH, который включает последовательность STII-SD, вырабатывает более высокое количество hGH, причем даже в отсутствии индуктора экспрессии - IPTG.
Примеры 1-10
В примерах 1-10 описывается конструирование векторов, каждый из которых включает ген, кодирующий слитый белок MST/hGH по настоящему изобретению, причем MST соответствует модифицированному сигнальному пептиду энтеротоксина-II E. coli. Ген STII или последовательность STII-SD плазмиды pT14SSH, полученной на 3-й стадии примера получения 3, модифицировали в соответствии с методом направленного мутагенеза (C. Papworth et al., 1996, Strategies, 9, 3), который был осуществлен с помощью ПЦР этой плазмиды с использованием прямого праймера, характеризующегося модифицированной нуклеотидной последовательностью, и обратного праймера, характеризующегося нуклеотидной последовательностью, комплементарной прямому праймеру.
Модифицированные сигнальные пептиды энтеротоксина-II E.coli, полученные в примерах 1-10 - MSTs (от MST1 до MST10), - охарактеризованы в табл.II наряду с STII, а препаративные процедуры примеров 1-10 описаны ниже.
Пример 1. Конструирование вектора, включающего ген, кодирующий слитый белок MST1/hGH
Стадия 1
Вектор pT14SSH, полученный на 3-й стадии примера получения 3, подвергали ПЦР с использованием следующих комплементарных праймеров S4 и AS4, которые были сконструированы так, чтобы заменить 4-й кодон (АТТ) в составе STII на кодон треонина (АСА).
Прямой праймер S4 (SEQ ID NO 23):
5'-GGTGTTTTATGAAAAAGACAATCGCATTTCTTC-3'.
Обратный праймер AS4 (SEQ ID NO 24):
5'-GAAGAAATGCGATTGTCTTTTTCATAAAACACC-3'.
Полученный таким образом вектор расщепляли рестриктазами XbaI и MluI с получением 100-нуклеотидного XbaI/MluI-фрагмента, который был встроен по XbaI/MluI-сегменту вектора pT14SSH с получением вектора pT14SSH-4T. Вектор pT14SSH-4T включает ген, кодирующий модифицированный слитый белок STII/hGH, имеющий остаток Thr вместо 4-й аминокислоты STII.
Стадия 2
Вектор pT14SSH-4T был подвергнут ПЦР с использованием следующих комплементарных праймеров S5 и AS5, которые были сконструированы так, чтобы вместо 22-го кодона (ААТ) STII внести кодон глутамина (САА), с получением вектора pT14SSH-4T22Q.
Прямой праймер S5 (SEQ ID NO 25):
5'-CAAATGCCCAAGCGTTCCCA-3'.
Обратный праймер AS5 (SEQ ID NO 26):
5'-TGGGAACGCTTGGGCATTTG-3'.
Вектор pT14SSH-4T22Q включает ген, кодирующий слитый белок MSTl/hGH, у которого 4-я и 22-я аминокислоты в составе STII заменены на Thr и Gln соответственно.
Стадия 3
Вектор pT14SSH-4T22Q подвергали ПЦР с использованием следующих комплементарных праймеров S6 и AS6, имеющих шесть нуклеотидных последовательностей, показанных ниже, между последовательностью STII-SD 5'-CAGG-3' и инициирующим кодоном STII, с целью предотвращения образования вторичных структур соответствующей транскрибируемой мРНК.
Прямой праймер S6 (SEQ ID NO 27):
5'-TCTAGAGGTTGAGGTGTTTTATGA-3'.
Обратный праймер AS6 (SEQ ID NO 28):
5'-TCATAAAACACCTCAACCTCTAGA-3'.
Полученный таким образом вектор pT14S1SH-4T22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок MST1/hGH, в котором 4-я и 22-я аминокислоты в составе STII заменены на Thr и Gln соответственно.
На фиг. 4 показана описанная выше процедура конструирования вектора рТ14S1SН.
Клетки E.coli BL21 (DE3) были трансформированы вектором pT14S1SH-4T22Q с получением трансформанта, обозначенного E.coli HM10011, который был депонирован в Корейский центр культивируемых микроорганизмов (КССМ) 12 августа 1998 года под депозитарным номером КССМ-10137.
Пример 2. Конструирование вектора, включающего ген, кодирующий слитый белок MST2/hGH
Была повторена процедура 2-й стадии примера 1, за исключением использования следующих комплементарных праймеров S7 и AS7, которые были сконструированы так, чтобы внести кодоны валина и глутамина (GTT и САА) вместо 20-го и 22-го кодонов (ААТ и ТАТ) в составе STII соответственно, с получением вектора pT14SSH-4T20V22Q.
Прямой праймер S7 (SEQ ID NO 29):
5'-GTTTTTTCTATTGCTACAGTTGCCCAAGCGTTCCCAACCATTCCC-3'.
Обратный праймер AS7 (SEQ ID NO 30):
5'-GGGAATGGTTGGGAACGCTTGGGCAACTGTAGCAATAGAAAAAAC-3'.
Затем повторяли процедуру 3-й стадии примера 1, за исключением использования вектора pT14SSH-4T20V22Q, с получением вектора pT14S1SH-4T20V22Q. Вектор pT14S1SH-4V20V22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок МSТ3/hGH, в котором 4-я, 20-я и 22-я аминокислоты в составе STII заменены на Thr, Val и Gln соответственно.
Клетки E. coli BL21 (DE3) трансформировали вектором pT14S1SH-4T20V22Q с получением трансформанта, обозначенного как E.coli HM10012, который был депонирован в КССМ 12 августа 1998 года под депозитарным номером КССМ-10138.
Пример 3. Конструирование вектора, включающего ген, кодирующий слитый белок МSТ3/hGH
Повторяли процедуру 1-го этапа примера 1, за исключением того, что использовали следующие комплементарные праймеры S8 и AS8, которые конструировали так, чтобы внести кодоны лизина и треонина (AAG и АСА) вместо 4-го и 5-го кодонов (ААТ и АТС) в составе STII соответственно, с получением вектора pT14SSH-4К5Т.
Прямой праймер S8 (SEQ ID NO 31):
5'-GAGGTGTTTTATGAAAAAGAAGACAGCATTTCTTC-3'.
Обратный праймер AS8 (SEQ ID NO 32):
5'-GAAGAAATGCTGTCTTCTTTTTCATAAAACACCTC-3'.
Используя вектор pT14SSH-4K5T, повторяли процедуру 2-й стадии примера 1 с получением вектора pT14SSH-4K5T22Q.
Затем повторяли процедуру 3-й стадии примера 1 с использованием вектора pT14SSH-4K5T22Q с получением вектора рT14S1SH-4K5T22Q. Вектор pT14S1SH-4K5T22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок MST3/hGH, в котором 4-я, 5-я и 22-я аминокислоты в составе STII заменены на Lys, Thr и Gln соответственно.
Клетки E.coli BL21 (DE3) были трансформированы вектором pT14S1SH-4K5T22Q с получением трансформанта, обозначенного как E.coli HM10013.
Пример 4. Конструирование вектора, включающего ген, кодирующий слитый белок MST4/hGH
Повторяли процедуру 1-й стадии примера 1, за исключением того, что использовали следующие комплементарные праймеры S9 и AS9, которые конструировали так, чтобы внести кодон серина (ТСТ) вместо 4-го кодона (ДАТ) в составе STII, с получением вектора pT14SSH-4S.
Прямой праймер S9 (SEQ ID NO 33):
5'-GAGGTGTTTTATGAAAAAGTCTATCGCATTTCTTC-3'.
Обратный праймер AS9 (SEQ ID NO 34):
5'-GAAGAAATGCGATAGACTTTTTCATAAAACACCTC-3'.
Используя вектор pT14SSH-4S, повторяли процедуру 2-й стадии примера 1 с получением вектора pT14SSH-4S22Q.
Затем повторяли процедуру 3-й стадии примера 1 с использованием вектора pT14SSH-4S22Q с получением вектора pT14S1SH-4S22Q. Вектор pT14S1SH-4S22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок MST4/hGH, в котором 4-я и 22-я аминокислоты в составе STII заменены на Ser и Gln соответственно.
Клетки E. coli BL21 (DE3) трансформировали вектором pT14S1SH-4S22Q с получением трансформанта, обозначенного как E.coli HM10014.
Пример 5. Конструирование вектора, включающего ген, кодирующий слитый белок MST5/hGH
Повторяли процедуру 2-й стадии примера 1, за исключением того, что использовали вектор pT14SSH-4S, полученный в примере 4, и праймеры S7 и AS7, использовавшиеся в примере 2, с получением вектора p14SSH-4S20V22Q.
Затем повторяли процедуру 3-й стадии примера 1, используя вектор pT14SSH-4S20V22Q, с получением вектора pT14S1SH-4S20V22Q. Вектор pT14S1SH-4S20V22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок MST5/hGH, в котором 4-я, 20-я и 22-я аминокислоты в последовательности STII заменены на Ser, Val и Gln соответственно.
Клетки E. coli BL21 (DE3) трансформировали вектором pT14S1SH-4S20V22Q с получением трансформанта, обозначенного как E.coli HM10015.
Пример 6. Конструирование вектора, включающего ген, кодирующий слитый белок MST6/hGH
Вектор pT14SSH-4T20V22Q, полученный в примере 2, подвергали ПЦР с использованием следующих комплементарных праймеров S10 и AS10, которые были сконструированы так, чтобы заменить 12-й кодон (ATG) в составе STII на кодон глицина (GGT), с получением вектора pT14SSH-4T12G20V22Q.
Прямой праймер S10 (SEQ ID NO 35):
5'-GCATTTCTTCTTGCATCTGGTTTCGTTTTTTCTATTGC-3'.
Обратный праймер AS10 (SEQ ID NO 36):
5'-GCAATAGAAAAAACGAAACCAGATGCAAGAAGAAATGC-3'.
Затем повторяли процедуру 3-й стадии примера 1, используя вектор pT14SSH-4T12G20V22Q, с получением вектора рТ14S1SН-4T12G20V22Q. Вектор pT14S1SH-4T12G20V22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок МSТ6/hGH, в котором 4-я, 12-я, 20-я и 22-я аминокислоты в последовательности STII заменены на Thr, Gly, Val и Gln соответственно.
Клетки E.coli BL21 (DE3) трансформировали вектором pT14S1SH-4T12G20V22Q с получением трансформанта, обозначенного как E.coli HM10016.
Пример 7. Конструирование вектора, включающего ген, кодирующий слитый белок MST7/hGH
Вектор pT14SSH-4T12G20V22Q, полученный в примере 6, подвергали ПЦР с использованием следующих комплементарных праймеров S11 и AS11, которые были сконструированы так, чтобы внести кодон лейцина (СТТ) вместо 12-го кодона (GGT) в составе MST6, с получением вектора pT14SSH-4T12L20V22Q.
Прямой праймер S11 (SEQ ID NO 37):
5'-GCATTTCTTCTTGCATCTCTTTTCGTTTTTTCTATTGC-3'.
Обратный праймер AS11 (SEQ ID NO 38):
5'-GCAATAGAAAAAACGAAAAGAGATGCAAGAAGAAATGC-3'.
Затем повторяли процедуру 3-й стадии примера 1, используя вектор pT14SSH-4T12L20V22Q, с получением вектора pT14S1SH-4T12L20V22Q. Вектор pT14S1SH-4T12L20V22Q включает модифицированную последовательность STII-SD и ген, кодирующий слитый белок MST7/hGH, в котором 4-я, 12-я, 20-я и 22-я аминокислоты в последовательности STII заменены на Thr, Leu, Val и Gln соответственно.
Клетки E.coli BL21 (DE3) трансформировали вектором pT14S1SH-4T12L20V22Q с получением трансформанта, обозначенного как E.coli HM10017.
Пример 8. Конструирование вектора, включающего ген, кодирующий слитый белок MST8/hGH
Повторяли процедуру 1-й стадии примера 1, за исключением того, что использовали следующие комплементарные праймеры S12 и AS12, которые конструировали так, чтобы внести кодоны лизина и серина (AAG и ТСТ) вместо 4-го и 5-го кодонов (ААТ и АТС) в составе STII соответственно, с получением вектора pT14SSH-4K5S.
Прямой праймер S12 (SEQ ID NO 39):
5'-GAGGTGTTTTATGAAAAAGAAGTCTGCATTTCTTC-3'.
Обратный праймер AS12 (SEQ ID NO 40):
5'-GAAGAAATGCAGACTTCTTTTTCATAAAACACCTC-3'.
С использованием вектора pT14SSH-4K5S повторяли процедуру 2-й стадии примера 1 с получением вектора pT14SSH-4K5S22Q. Вектор pT14SSH-4K5S22Q включает ген, кодирующий слитый белок MST8/hGH, в котором 4-я, 5-я и 22-я аминокислоты последовательности STII заменены на Lys, Ser и Gln соответственно.
Клетки E.coli BL21 (DE3) трансформировали вектором pT14SSH-4K5S22Q с получением трансформанта, обозначенного как E.coli HM10018.
Пример 9. Конструирование вектора, включающего ген, кодирующий слитый белок MST9/hGH
Повторяли процедуру 1-й стадии примера 1, за исключением того, что использовали следующие комплементарные праймеры S13 и AS13, которые сконструировали так, чтобы заменить 2-й, 4-й и 5-й кодоны (ААА, ААТ и АТС) в составе STII на кодоны валина, лизина и треонина (GTT, AAG и АСА) соответственно, с получением вектора pT14SSH-2V4K5T.
Прямой праймер S13 (SEQ ID NO 41):
5'-GAGGTGTTTTATGGTTAAGAAGACAGCATTTCTTC-3'.
Обратный праймер AS13 (SEQ ID NO 42):
5'-GAAGAAATGCTGTCTTCTTAACCATAAAACACCTC-3'.
С использованием вектора pT14SSH-2V4K5T повторяли процедуру 2-й стадии примера 1 с получением вектора pT14SSH-2V4K5T22Q. Вектор pT14SSH-2V4K5T22Q включает ген, кодирующий слитый белок MST9/hGH, в котором 2-я, 4-я, 5-я и 22-я аминокислоты в составе STII заменены на Val, Lys, Thr и Gln соответственно.
Клетки E. coli BL21 (DE3) трансформировали вектором pT14SSH-2V4K5T22Q с получением трансформанта, обозначенного как E.coli HM10019.
Пример 10. Конструирование вектора, включающего ген, кодирующий слитый белок MST10/hGH
Повторяли процедуру 1-й стадии примера 1, за исключением того, что использовали следующие комплементарные праймеры S14 и AS14, которые были сконструированы таким образом, чтобы заменить 4-й кодон (ААТ) в STII на кодон лизина (AAG), с получением вектора pT14SSH-4K.
Прямой праймер S14 (SEQ ID NO 43):
5'-GAGGTGTTTTATGAAAAAGAAGATCGCATTTCTTC-3'.
Обратный праймер AS14 (SEQ ID NO 44):
5'-GAAGAAATGCGATCTTCTTTTTCATAAAACACCTC-3'.
С использованием вектора pT14SSH-4K повторяли процедуру 2-й стадии примера 1 с получением вектора pT14SSH-4K22Q.
Вектор pT14SSH-4K22Q подвергали ПЦР с использованием праймеров S7 и AS7, использовавшихся в примере 2, с получением вектора pT14SSH-4K20V22Q. Вектор pT14SSH-4K20V22Q включает ген, кодирующий слитый белок MST10/hGH, в котором 4-я, 20-я и 22-я аминокислоты в последовательности STII заменены на Lys, Val и Gln соответственно.
Клетки E. coli BL21 (DE3) трансформировали вектором pT14SSH-4K20V22Q с получением трансформанта, обозначенного как E.coli HM10020.
Пример 11. Выработка hGH с использованием гена MST/hGH
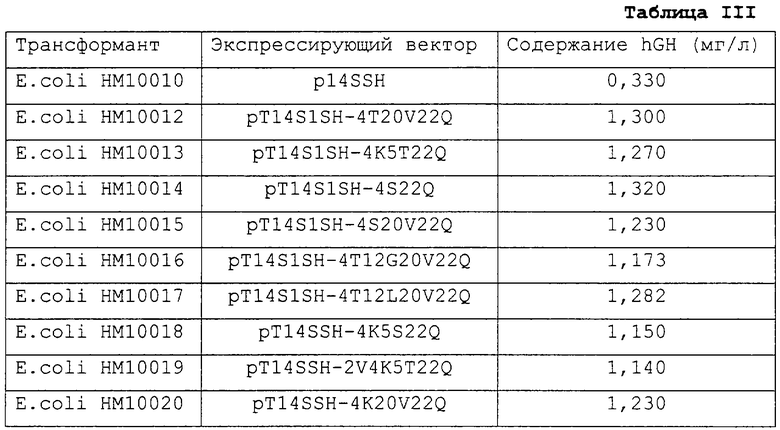
Для анализа влияния MST на выработку hGH процедуру примера получения 4 повторяли с использованием трансформантов (E.coli HM10011-HM10020), полученных в примерах 1-10, в отсутствие добавления IPTG. В качестве контроля использовали трансформант НМ10010, полученный на 3-й стадии примера получения 3. Уровень hGH определяли как количество hGH, выработанного в расчете на 1 л культуральной среды. Полученные результаты показаны в табл.III.
Как можно видеть из табл.III, каждый из векторов по настоящему изобретению, включающий ген MST, вырабатывает hGH с более высоким выходом по сравнению с контрольным вектором p14SSH, включающим нативный ген STII. Кроме того, среди векторов по настоящему изобретению более высокий выход hGH был в тех случаях, когда имелись модифицированные последовательности STII-SD, по сравнению с теми векторами, в которых имелась нативная последовательность STII-SD.
Пример 12. Очистка hGH
Трансформант E. coli HM10011, полученный в примере 1, культивировали в среде LB, а экспрессию гена MST/hGH индуцировали с использованием IPTG, после чего культуру центрифугировали при 6000 об/мин в течение 20 минут для сбора клеток. Раствор периплазмы приготавливали из этих клеток путем повторения процедуры из примера получения 4.
Раствор периплазмы доводили до рН 5,3-6,0, наносили на DEAE-сепарозную колонку (Pharmacia Inc., Швеция), предварительно уравновешенную до рН 5,8, и затем эту колонку промывали раствором 10 мМ NaCl. hGH элюировали с использованием буферных растворов, содержащих 20 мМ, 40 мМ и 80 мМ NaCl соответственно, собирая и объединяя фракции, содержащие hGH.
Объединенные фракции подвергали хроматографии на фенилсепарозной колонке (Pharmacia Inc., Швеция) с получением hGH, характеризующегося чистотой 99%, который далее очищали хроматографически на колонке Сефадекс G-100 (Pharmacia Inc., Швеция).
Очищенную фракцию hGH подвергали электрофорезу в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ) с целью определения чистоты и приблизительной концентрации hGH, a затем анализировали методом ТИФА с целью определения точной концентрации hGH в данной фракции.
На фиг. 5 воспроизведены результаты ДСН-ПААГ, где дорожка 1 показывает белки с маркерным размером; дорожка 2 - очищенный hGH. Как видно на фиг.5, высокий уровень чистого hGH получен при культивировании трансформанта по настоящему изобретению.
Кроме того, была определена N-концевая аминокислотная последовательность hGH и полученные результаты показывают, что hGH, выработанный в соответствии с настоящим изобретением, не включает метионина по N-концу.
Хотя настоящее изобретение было описано в отношении вышеназванных конкретных вариантов, должно быть понятно, что специалистом в данной области техники в настоящее изобретение могут быть внесены различные модификации и изменения, которые также попадают в объем настоящего изобретения в соответствии с определенным в прилагаемой формуле изобретения.

Модифицированный сигнальный пептид энтеротоксина-II E.coli отличается тем, что по крайней мере одна из аминокислот в положениях 2, 4, 5, 12, 20 и 22 сигнального пептида энтеротоксина-II E.coli с аминокислотной последовательностью Met Lys Lys Asn Ile Ala Phe Leu Leu Ala Ser Met Phe Val Phe Ser Ile Ala Thr Asn Ala Tyr Ala заменена на другую аминокислоту при условии, что по крайней мере одна из аминокислот во 2-м и 4-м положениях модифицированного пептида является лизином. Описаны также ген, кодирующий указанный пептид, а также плазмидный вектор, включающий этот ген. Описаны штаммы E.coli, трансформированные плазмидным вектором, для получения гормона роста человека, а также способ получения гормона роста человека в микроорганизме. 8 с. и 4 з.п.ф-лы, 5 ил. 3 табл.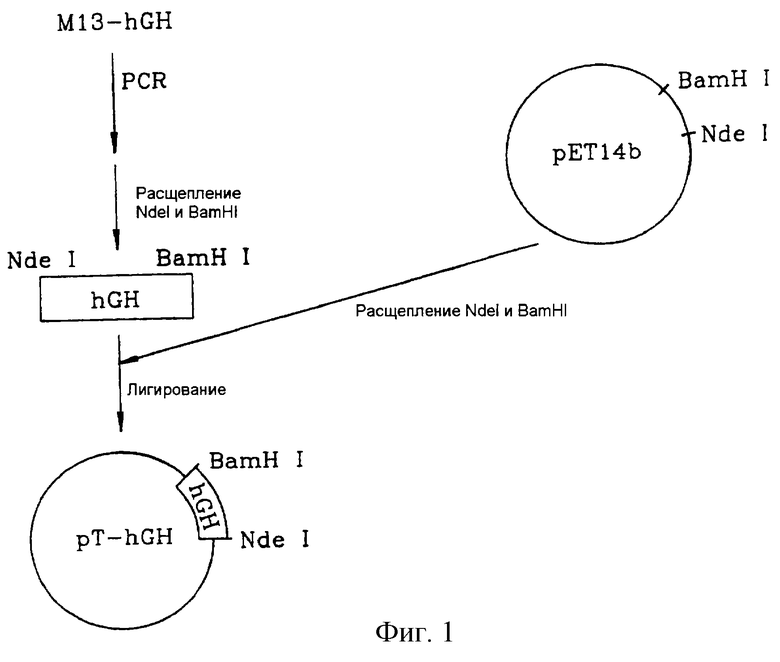

2. Модифицированный сигнальный пептид энтеротоксина-II E.coli no п.1, в котором
2-я аминокислота Lys не заменена;
4-я аминокислота Asn заменена на Ser, Thr, Lys или Gln;
5-я аминокислота Ilе не заменена или заменена на Thr или Ser;
12-я аминокислота Met не заменена или заменена на Ala, Gly, Val, Leu или Ilе;
20-я аминокислота Asn не заменена или заменена на Ilе, Phe, Ala или Val; и
22-я аминокислота Туr не заменена или заменена на Gln, Asn, Ala или Lys.
2-я аминокислота Lys заменена на любую другую аминокислоту;
4-я аминокислота Asn заменена на Lys;
5-я аминокислота Ilе заменена на Ser, Thr, Asn, Gln или Arg;
12-я аминокислота Met не заменена или заменена на Ala, Gly, Val, Leu или Ilе;
20-я аминокислота Asn не заменена или заменена на Ilе, Phe, Ala или Val; и
22-я аминокислота Туг не заменена или заменена на Gln, Asn, Ala или Lys.
(a) 4-я Asn на Thr и 22-я Туr на Gln;
(b) 4-я Asn на Thr, 20-я Asn на Val и 22-я Туr на Gln;
(c) 4-я Asn на Lys, 5-я Ilе на Thr и 22-я Туr на Gln;
(d) 4-я Asn на Ser и 22-я Туr на Gln;
(e) 4-я Asn на Ser, 20-я Asn на Val и 22-я Туr на Gln;
(f) 4-я Asn на Thr, 12-я Met на Gly, 20-я Asn на Val и 22-я Туr на Gln;
(g) 4-я Asn на Thr, 12-я Met на Leu, 20-я Asn на Val и 22-я Туr на Gln;
(h) 4-я Asn на Lys, 5-я Ilе на Ser и 22-я Туr на Gln;
(i) 2-я Lys на Val, 4-я Asn на Lys, 5-я Ilе на Thr и 22-я Туr на Gln;
(k) 4-я Asn на Lys, 20-я Asn на Val и 22-я Туr на Gln.
| УСТРОЙСТВО ДЛЯ ТРАНСПОРТИРОВАНИЯ ШТУЧНЫХ ГРУЗОВ | 0 |
|
SU177343A1 |
| RU 94030871 A1, 20.05.1997 | |||
| Устройство для наложения изоляционной ленты на замкнутые кольцевые сердечники | 1976 |
|
SU626448A1 |
| US 5308835 А, 03.05.1994 | |||
| Способ приготовления сернистого красителя защитного цвета | 1915 |
|
SU63A1 |

