Область изобретения
Настоящее изобретение относится к генам и способам высокопродуктивной экспрессии эукариотических и вирусных белков в эукариотических клетках.
Предпосылки изобретения
У прокариот экспрессия продуктов эукариотических генов иногда ограничена из-за наличия кодонов, которые редко используются у Е. coli. Экспрессия таких генов может усиливаться при систематической замене эндогенных кодонов на кодоны, чаще представленные в прокариотических генах с высокой продуктивностью экспрессии (Robinson et al., Nucleic Acids Res. 12:6663, 1984). Обычно полагали, что редкие кодоны создают пропуск в рибосоме, что приводит к повреждению полной образующейся полипептидной цепи и разобщению транскрипции и трансляции. Полагали, что пропуск в рибосоме ведет к обнажению 3'-конца мРНК для клеточных рибонуклеаз.
Сущность изобретения
Настоящее изобретение относится к синтетическому гену, кодирующему белок, обычно экспрессируемый в клетках млекопитающих или иных эукариотических клетках, в которых, по крайней мере, один непредпочтительный или менее предпочтительный кодон природного гена, кодирующий данный белок, заменен на предпочтительный кодон, кодирующий ту же аминокислоту.
Предпочтительные кодоны представляют собой А1а (gсс); Аrg (сgс); Asn (aac); Asp (gас); Cys (tgc); Gln (саg); Gly (ggc); His (cac); Ilе (atc); Leu (ctg); Lys (aag); Pro (ccc); Phe (ttc); Ser (agc); Thr (асc); Туг (tac); и Val (gtg). Менее предпочтительные кодоны представляют собой Gly (ggg); Ile (att); Leu (ctc); Ser (tcc); Val (gtc) и Arg (agg). Кодоны, которые не соответствуют описанию предпочтительных кодонов или менее предпочтительных кодонов, представляют собой непредпочтительные кодоны. Вообще, степень предпочтения отдельного кодона указывает на преобладание данного кодона в человеческих генах, экспрессируемых с высокой продуктивностью, которые указаны в Таблице 1 под заголовком “Высокопродуктивный”. Например, “atc” соответствует 77% кодона Ilе в высокопродуктивно экспрессируемых человеческих генах и представляет собой предпочтительный кодон Ilе; “att” соответствует 18% кодона Ilе в высокопродуктивно экспрессируемых человеческих генах и представляет собой менее предпочтительный кодон Ilе. Последовательность “ata” соответствует лишь 5% кодона Ilе в высокопродуктивно экспрессируемых человеческих генах как непредпочтительный кодон Ilе. Замена кодона другим кодоном, который преобладает в высокопродуктивно экспрессируемых человеческих генах, будет, как правило, повышать экспрессию данного гена в клетках млекопитающих. Соответственно, настоящее изобретение включает замену менее предпочтительного кодона на предпочтительный кодон, а также замену непредпочтительного кодона предпочтительным или менее предпочтительным кодоном.
Под “белком, обычно экспрессируемым в клетке млекопитающего”, подразумевается белок, который экспрессируется у млекопитающих в естественных условиях. Данный термин включает такие гены в геноме млекопитающих, как гены, кодирующие Фактор VIII, Фактор IX, интерлейкины и другие белки. Данный термин включает также гены, которые экспрессируются в клетках млекопитающих при болезненных состояниях, такие как онкогены, а также гены, которые кодируются вирусом (в том числе ретровирусом), которые экспрессируются в постинфекционных клетках млекопитающих. Под “белком, обычно экспрессируемым в эукариотических клетках”, подразумевается белок, который экспрессируется эукариотом в естественных условиях. Данный термин включает также гены, которые экспрессируются в клетках млекопитающих при болезненных состояниях.
В предпочтительных осуществлениях синтетические гены способны экспрессировать белок млекопитающего или эукариотический белок на уровне, который составляет, по меньшей мере, 110%, 150%, 200%, 500%, 1000%, 5000% или даже 10000% от уровня, который экспрессируется “природным” (или “нативным”) геном в in vitro культуральной системе клеток млекопитающих в идентичных условиях (т.е. тот же тип клеток, те же культуральные условия, тот же экспрессионный вектор).
Ниже описаны подходящие клеточные культуральные системы для измерения экспрессии данного синтетического гена и соответствующего природного гена. Другие подходящие экспрессионные системы, использующие клетки млекопитающих, хорошо известны специалистам в данной области техники и описаны, например, в указанных ниже ссылках на классические работы по молекулярной биологии. Векторы, подходящие для экспрессии данного синтетического и природного генов, описаны ниже и в ссылках на классические работы, также описанных ниже. Под “экспрессией” подразумевается экспрессия белка. Экспрессия может быть измерена с использованием антител, специфичных к интересующему белку. Такие антитела и методы измерения хорошо известны специалистам в данной области техники. Под “природным геном” и “нативным геном” подразумевается генная последовательность (включая природные аллельные варианты), которая кодирует данный белок в природе, т.е. нативная или природная кодирующая последовательность.
В других предпочтительных осуществлениях, по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% кодонов в природном гене представляют собой непредпочтительные кодоны.
В других предпочтительных осуществлениях, по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% непредпочтительных кодонов в данном природном гене заменены предпочтительными кодонами или менее предпочтительными кодонами.
В других предпочтительных осуществлениях, по меньшей мере, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% непредпочтительных кодонов в данном природном гене замещены предпочтительными кодонами.
В предпочтительном осуществлении белок представляет собой ретровирусный белок. В более предпочтительном осуществлении белок представляет собой лентивирусный белок. В еще более предпочтительном осуществлении белок представляет собой белок HIV. В других предпочтительных осуществлениях белок представляет собой gag, pol, env, gp 120 или gp l60. В других предпочтительных осуществлениях белок представляет собой человеческий белок. В более предпочтительных осуществлениях белок представляет собой человеческий Фактор VIII и белок с делецией В-области человеческого Фактора VIII. В другом предпочтительном осуществлении белок представляет собой зеленый флуоресцентный белок.
В различных предпочтительных осуществлениях, по меньшей мере, 30%, 40%, 50%, 60%, 70%, 80%, 90% и 95% кодонов синтетического гена представляют собой предпочтительные или менее предпочтительные кодоны.
В настоящем изобретении представлен также экспрессионный вектор, включающий синтетический ген.
Следующий аспект настоящего изобретения представляет клетку, несущую данный синтетический ген. В различных предпочтительных вариантах осуществления настоящего изобретения данная клетка является прокариотической клеткой, и данная клетка является клеткой млекопитающего.
В предпочтительных осуществлениях данный синтетический ген включает менее 50, менее 40, менее 30, менее 20, менее 10, менее 5 или не содержит “сg” - последовательностей.
В настоящем изобретении охарактеризован также способ получения синтетического гена, кодирующего белок, обычно экспрессируемый клеткой млекопитающего или иной эукариотической клеткой. Данный способ включает идентификацию не предпочтительных или менее предпочтительных кодонов в данном природном гене, кодирующем данный белок, и замену одного или нескольких данных непредпочтительных или менее предпочтительных кодонов на предпочтительный кодон, кодирующий ту же данную аминокислоту, что и замещаемый кодон.
В некоторых обстоятельствах (напр., чтобы дать возможность ввести сайт рестрикции) может оказаться желаемым заменить непредпочтительный кодон на менее предпочтительный кодон, а не на предпочтительный кодон.
Нет необходимости заменять все менее предпочтительные или непредпочтительные кодоны предпочтительными кодонами. Повышение экспрессии может быть достигнуто даже при неполной замене менее предпочтительных или непредпочтительных кодонов предпочтительными кодонами. В некоторых обстоятельствах может оказаться желаемым лишь частичная замена непредпочтительных кодонов предпочтительными или менее предпочтительными кодонами с целью получить промежуточный уровень экспрессии.
В других предпочтительных осуществлениях настоящее изобретение представляет векторы (включая экспрессионные векторы), включающие один или несколько синтетических генов.
Под “вектором” подразумевается молекула ДНК, полученная, например, из плазмиды, бактериофага или из вируса млекопитающего или насекомого, в которую могут быть вставлены или клонированы фрагменты ДНК. Вектор будет содержать один или несколько уникальных сайтов рестрикции и может обладать способностью к автономной репликации в определенном хозяине или организме-носителе, таком, в котором может быть воспроизведена клонируемая последовательность. Таким образом, под "экспрессионным вектором" подразумевается любой автономный элемент, способный управлять синтезом белка. Такие экспрессионные ДНК-векторы включают плазмиды и вирусы, вводимые в клетки млекопитающих.
В настоящем изобретении представлены также фрагменты синтетического гена, которые кодируют желаемую часть данного белка. Такие фрагменты синтетического гена аналогичны синтетическим генам настоящего изобретения за тем исключением, что они кодируют лишь часть данного белка. Такие генные фрагменты предпочтительно кодируют, как минимум, 50, 100, 150 или 500 смежных аминокислот данного белка.
При конструировании синтетических генов настоящего изобретения желательно избегать CpG-последовательностей, так как эти последовательности могут вызывать умолкание генов. Поэтому в предпочтительном варианте осуществления настоящего изобретения кодирующая область данных синтетических генов не включает последовательность “сg”.
Неоднозначность кодона, присутствующая в гене env gp120 HIV, присутствует также в генах gag и pol. Поэтому замена части непредпочтительных и менее предпочтительных кодонов, находящихся в этих генах, на предпочтительные кодоны должна произвести ген, способный к более высокопродуктивной экспрессии. Большая доля кодонов человеческих генов, кодирующих Фактор VIII и Фактор IX, является непредпочтительными кодонами или менее предпочтительными кодонами. Замена части данных кодонов на предпочтительные кодоны должна произвести гены, способные к более высокопродуктивной экспрессии в культуре клеток млекопитающих.
Синтетические гены настоящего изобретения могут быть интродуцированы в клетки живого организма. Например, векторы (вирусные или невирусные) могут использоваться для введения синтетического гена в клетки живого организма для генной терапии.
И наоборот, возможно желаемым окажется заменить предпочтительные кодоны в природном гене на менее предпочтительные кодоны в целях снижения экспрессии.
Ссылки на классические работы, описывающие общие принципы метода рекомбинантной ДНК, включают Watson et at., Molecular Biology of the Gene, Volumes I and II, the Benjamin/Cummings Publishing Company, Inc., publisher, Menio Park, CA (1987); Darnell et al., Molecular Cell Biology, Scientific American Books, Inc., Publisher, New York, N.Y. (1986); Old et al., Principles of Gene manipulation: An Introduction to Genetic Engineering, 2d edition. University of California Press, publisher, Berkeley, CA (1981); Maniatis et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Laboratory, publisher. Cold Spring Harbor, NY (1989); и Current Protocols in Molecular Biology, Ausubel et al., Wiley Press, New York, NY (1992).
Под “трансформированной клеткой” подразумевается клетка, в которую (или в предка которой) интродуцировали методами рекомбинантной ДНК выбранную молекулу ДНК, например синтетический ген.
Под “расположенной для экспрессии” подразумевается, что молекула ДНК, например синтетический ген, располагается по соседству с последовательностью ДНК, которая управляет транскрипцией и трансляцией данной последовательности (т.е. способствует образованию белка, кодируемого данным синтетическим геном).
На фиг.1 А - фиг.1D изображена последовательность синтетического гена gр120 и синтетического гена gр160, в которой кодоны были заменены на кодоны, обнаруженные в высокопродуктивно экспрессируемых человеческих генах.
На фиг.2 схематически изображен синтетический ген gр120 (HIV-1 MN). Заштрихованные части, отмеченные как v1-v5, изображают гипервариабельные области. Заполненный точками прямоугольник указывает сайт связывания с CD4. Показано небольшое количество сайтов рестрикции: Н (Hind3), Nh (Nhel), P (Pst1), Na (Nael), M (Mlu1), R (EcoR1), A (Age1) и No (Not1). Химически синтезированные фрагменты ДНК, которые служили в качестве матриц для ПЦР, показаны ниже последовательности gp120, наряду с локализацией праймеров, используемых для их амплификации.
Фиг.3 представляет собой фотографию результатов анализов неустойчивой трансфекции, используемых для измерения экспрессии gр120. Гель-электрофорез иммунопреципитированных супернатантов клеток 293Т, трансфицированных плазмидами, экспрессирующими gр120, кодируемого участком IIIB, выделенным из HIV-1 (gp120IIIB), участком MN, выделенным из HIV-1 (gp120mn), участком MN, выделенным из HIV-1, модифицированного путем замены эндогенного лидерного пептида на пептид из антигена CD5 (gp120mnCD5L), или химически синтезированным геном, кодирующего MN-вариант HIV-1, на CDSLeader человека (syngp120im). Супернатанты собирали после 12-часового периода мечения после 60 часов трансфекции и иммунопреципитации гибридным белком CD4:IgG1 и белок-А.сефарозой.
На фиг.4 графически изображены результаты анализов ELISA, используемых для измерения содержания белка в супернатантах временно трансфицированных клеток 293Т. Супернатанты клеток 293Т, трансфицированные плазмидами, экспрессирующими gp120, кодируемого участком IIIB из HIV-1 (gp120 IIIb), участком MN из HIV-1 (gp120mn), участком MN из HIV-1, модифицированного путем замены эндогенного лидерного пептида на пептид из CD5-антигена (gp120mn CD5L), или химически синтезированным геном, кодирующим MN-вариант HIV-1, на человеческий лидерный пептид CDS (syngp 120 mn), собирали через 4 суток и тестировали в gp120/CD4 ELISA. Содержание gр120 выражали в нг/мл.
Фиг.5 представляет собой фотографию геля, иллюстрирующую результаты анализа иммунопреципитации, используемого для измерения экспрессии нативного и синтетического gр120 в присутствии rev в транс и RRE в цис. В данном эксперименте клетки 293Т временно трансфицировали путем кальцийфосфатной копреципитации 10 мкг плазмиды, экспрессирующей: (А) синтетическую gр120МN-последовательность и RRE в цис, (В) часть gp120 HIV-1 IIIB, (С) часть gp120 HIV-1 IIIB и RRE в цис, все в присутствии или в отсутствие экспрессии rev. RRE-конструкты gp120IIIbRRE и syngp120mnRRE конструировали с использованием фрагмента RRE Eag/Hpa1, клонируемого с помощью ПЦР из провирусного клона HIV-1 НХВ2. Каждую экспрессионную плазмиду gp120 котрансфицировали с помощью 10 мкг плазмидной ДНК pCMVrev или CDM7. Через 60 часов после трансфекции собирали супернатанты, иммунопреципитировали с помощью гибридного белка CD4:IgG и белок-А агарозы, а затем разгоняли в 7%-ном восстанавливающем SDS-PAGE. Время экспонирования полученного геля удлиняли, чтобы дать возможность индуцировать демонстрацию gp120IIIbrre с помощью rev.
На фиг.5В представлен результат более короткого экспонирования аналогичного эксперимента, в котором syngp120mnrre котрансфицировали с или без pCMVrev.
На фиг.5С представлена схематичная диаграмма конструктов, используемых на фиг.5А.
Фиг.6 представляет собой сравнение последовательности гена ratTHY-1 дикого типа (wt) и синтетического гена ratTHY-1 (env), сконструированного путем химического синтеза и обладающего преобладающими кодонами, обнаруженными в гене HIV-1 env.
На фиг.7 представлена схематичная диаграмма синтетического гена ratTHY-1. Зачерненный прямоугольник обозначает сигнальный пептид. Заштрихованный прямоугольник обозначает последовательности предшественника, который управляет присоединением фосфатидилинозитолгликанового якоря. Уникальные сайты рестрикции, используемые для сборки конструктов THY-1, обозначены как Н (Hind3), M (Mlu1), S (Sacl) и No (Notl). Положения синтетических олигонуклеотидов, используемых для данного конструирования, показаны внизу данноой фигуры.
На фиг.8 представлено графическое изображение результатов проточно-цитометрического анализа. В этом эксперименте клетки 293Т временно трансфицировали экспрессионной плазмидой ratTHY-1 дикого типа (жирная линия), экспрессионной плазмидой ratTHY-1 с кодонами, кодирующими оболочку, (тонкая линия) или только вектором (пунктирная линия) путем кальцийфосфатной копреципитации. Клетки окрашивали анти-ratTHY-1 моноклональными антителами OХ7 с последующим поликлональным ФИТЦ-конъюгированным антителом против мышиного IgG через 3 дня после трансфекции.
Фиг.9А представляет собой фотографию геля, иллюстрирующую результаты анализа иммунопреципитации человеческих клеток 293Т, трансфицированных syngp120mn (A) или конструктом syngp120mn. rTHY-1env, который содержит ген rTHY-1env в 3'-нетранслируемой области гена syng120mn (В). Конструкцию syngp120mn.rTHY-1env получали путем вставки Not1-адаптера в затупленный Нind3-сайт плазмиды rTHY-1env. Затем Not1-фрагмент 0,5 т.п.н., содержащий ген rTHY-lenv клонировали в Not1-сайт плазмиды syngp120mn и проверяли на правильность ориентации. Супернатанты клеток, меченых 35S, собирали через 72 часа после трансфекции, преципитировали с помощью гибридного белка CD4:IgG и белка-А агарозы и разгоняли в 7%-ном восстанавливающем SDS-PAGE.
На фиг.9B представлена схематичная диаграмма конструктов, используемых в эксперименте, представленном на фиг.9А.
Фиг.10А представляет собой фотографию клеток COS, трансфицированных вектором, не испускающим лишь GFP-флуоресценцию.
Фиг.10В представляет собой фотографию клеток COS, трансфицированных экспрессионной плазмидой CDM7, кодирующей нативный GFP, сконструированный таким образом, чтобы включать консенсусную последовательность инициации трансляции.
Фиг.10С представляет собой фотографию клеток COS, трансфицированных экспрессионной плазмидой, содержащей те же фланкирующие последовательности, и консенсусом инициации, как и на фиг.10В, но содержащей генную последовательность с оптимальными кодонами.
Фиг.10D представляет собой фотографию клеток COS, трансфицированных экспрессионной плазмидой, такой же как и на Фигуре 10С, но несущей остаток Thr в положении 65 вместо Ser.
Фиг.11 изображает последовательность синтетического гена, кодирующего зеленые флуоресцентные белки (SEQ ID NO:40).
Фиг.12 изображает последовательность нативного гена человеческого Фактора VIII, не имеющего центрального В-домена (аминокислоты 760-1639 включительно) (SEQ ID NO:41).
Фиг.13 изображает последовательность синтетического гена человеческого Фактора VIII, не имеющего центрального В-домена (аминокислоты 760-1639 включительно) (SEQ ID NO:42).
Описание предпочтительных осуществлений
ПРИМЕР 1
Конструирование синтетического гена gp120, содержащего кодоны, обнаруженные в высокопродуктивно экспрессируемых человеческих генах.
Частотную таблицу кодона оболочечного предшественника LAV-субтипа HIV-1 получали с использованием программного обеспечения, разработанного The University of Wisconsin Genetics Computer Group. Полученные результаты, которые сведены в таблицу, сопоставлены в табл.1 с профилем использования кодонов коллекцией высокопродуктивно экспрессируемых человеческих генов. Для любой аминокислоты, кодируемой вырожденными кодонами, наиболее предпочтительный кодон высокопродуктивно экспрессируемых генов отличается от наиболее предпочитаемого кодона оболочечного предшественника HIV. Кроме того, простое правило описывает профиль предпочтительных - оболочечных кодонов, где оно применяется: предпочтительные кодоны максимизируют количество адениновых остатков в вирусной РНК. Во всех случаях подразумевается, что данный кодон, в котором третье положение представляет собой А, используется наиболее часто. В конкретном случае серина, три кодона одинаково обеспечивают один А-остаток в данной мРНК; вместе эти три кодона включают 85% сериновых кодонов, реально используемых в оболочечных транскриптах. Особенно яркий пример А-смещения обнаружен при выборе кодона для аргинина, в котором триплет AGA включает 88% аргининовых кодонов. Кроме данного преимущества А-остатков, отчетливое предпочтение среди вырожденных кодонов отдается уридину, третий остаток которого должен являться пиримидином. Наконец, несовместимость среди менее часто используемых вариантов может быть отнесена на счет наблюдения, что возникает динуклеотид CgG; поэтому третья позиция маловероятна для G всякий раз, когда вторая позиция представляет собой С, как в кодонах для аланина, пролина, серина и треонина; а триплеты CGX для аргинина почти совсем не используются.

Частоту встречаемости кодона вычисляли с использованием программы GCG созданной The University of Wisconsin Genetics Computes Group. Значения представляют собой процентную долю случаев, когда используется отдельный кодон. Частоты использования кодонов оболочечных генов из других участков вируса HIV-1 сравнимы и показывают аналогичное смещение.
С целью получения гена gр120, способного к высокопродуктивной экспрессии в клетках млекопитающих, сконструировали синтетический ген, кодирующий сегмент gр120 из HIV-1 (syngp120mn), на основе последовательности наиболее распространенного североамериканского субтипа, HIV-1 MN (Shaw et al., Science 226:1165, 1984; Gallo et al.,. Nature 321:119, 1986). В данном синтетическом гене gр120 почти все нативные кодоны были систематически заменены кодонами, более часто используемыми в высокопродуктивно экспрессируемых человеческих генах (фиг.1). Данный синтетический ген собирали из химически синтезированных олигонуклеотидов по 150-200 оснований в длину. Если химически синтезировали олигонуклеотиды, превышающие 120-150 оснований, процент полноразмерного продукта мог быть низким, а значительный избыток материала состоял из более коротких олигонуклеотидов. Так как эти более короткие фрагменты ингибируют процессы клонирования и ПЦР, может быть весьма затруднительно использовать олигонуклеотиды, превышающие определенную длину. Чтобы использовать сырой синтетический материал без предварительной очистки, перед клонированием одноцепочечные олигонуклеотидные пулы подвергали ПЦР-амплификации. Продукты ПЦР выделяли очисткой на агарозных гелях и использовали в качестве матриц для следующей стадии ПЦР. Два близких фрагмента могли быть коамплифицированы вследствие перекрывающихся последовательностей на конце каждого фрагмента. Данные фрагменты, которые обладали размером от 350 до 400 п.н., субклонировали в полученную из pCDM7 плазмиду, содержащую лидерную последовательность поверхностной молекулы CD5 с последующим полилинкером Nhe1/Pst1/Mlu1/EcoR1/BamH1. Каждый из ферментов рестрикции в этом полилинкере представляет сайт, который представлен либо на 5'-, либо на 3'-конце полученных в ходе ПЦР фрагментов. Поэтому последовательное субклонирование каждого из 4 длинных фрагментов позволяло собрать целый ген gp120. По каждому фрагменту субклонировали от трех до шести разных клонов и перед сборкой секвенировали. Схематичное изображение способа, примененного для конструирования синтетического gp120, показано на фиг.2. Последовательность синтетического гена gp120 (и синтетического гена gp160, созданного с использованием того же подхода) представлена на фиг.1.
Скорость мутаций была значительной. Наиболее часто возникавшие мутации представляли собой короткие (1 нуклеотид) и длинные (до 30 нуклеотидов) делеции. В некоторых случаях необходимо было заменить части в данной конкретной области на любые синтетические адаптеры или фрагменты из других субклонов без мутаций. Некоторые отклонения от точного наследования в сторону использования оптимальных кодонов осуществляли для обеспечения внесения сайтов рестрикции в полученный ген, чтобы облегчить замену различных сегментов (фиг.2). Данные уникальные сайты рестрикции вводили в ген приблизительно со 100 п.н. интервалами. Нативную лидерную последовательность HIV заменяли высокопродуктивным лидерным пептидом человеческого антигена CD5 для облегчения секреции (Aruffo et al., Cell 61:1303 1990). Плазмида, используемая для конструирования, представляет собой производное экспрессионного вектора млекопитающего pCDM7, транскрибирующего встроенный ген под контролем мощного человеческого предраннего промотора CMV.
Для сравнения кодирующих последовательностей gp120 дикого типа и синтетической синтетическую последовательность, кодирующую gp120, встраивали в экспрессионный вектор млекопитающего и тестировали в анализах неустойчивой трансфекции. В качестве контроля использовали различные нативные гены gp120 для исключения различий в уровнях экспрессии между различными вирусными участками и артефактов, индуцированных определенными лидерными последовательностями. Конструкт gp120 HIV IIIb, используемый в качестве контроля, получали путем ПЦР с использованием в качестве матрицы оболочечного фрагмента НХВ2 Sal1/Xho1 HIV-1. Для исключения мутаций, индуцируемых ПЦР, фрагмент Kpn1/Ear1, содержащий приблизительно 1,2 т.п.н. из данного гена, заменяли на соответствующую последовательность из провирусного клона. Конструкты gp120 дикого типа, используемые в качестве контролей, клонировали с помощью ПЦР из клеток С 8166, инфицированных HIV-1 MN (AIDS Repository, Rockville, MD) и экспрессировали gp120 либо с нативной оболочкой, либо с лидерной последовательностью CD5. Так как провирусные клоны в данном случае были не доступны, тестировали два клона каждого конструкта во избежание артефактов ПЦР. Для проведения полуколичественного анализа секретируемого gp120 в супернатантах клеток 293Т, временно трансфицированных кальцийфосфатной копреципитацией, иммунопреципитировали растворимым гибридным белком CD4: иммуноглобулин и протеин-А сефарозой.
Результаты данного анализа (фиг.3) показывают, что продукт данного синтетического гена экспрессируется с высокой продуктивностью в сравнении с уровнями нативных контролей gp120. Молекулярная масса синтетического гена gp120 была сравнима с таковой контрольных белков (фиг.3), лежит в диапазоне от 100 до 110 кДа. Несколько более быструю миграция может быть объяснена тем фактом, что в некоторых линиях опухолевых клеток, например 293Т, гликозилирование либо осуществляется не полностью, либо изменено до некоторой степени.
Для более точного сравнения уровней экспрессии белка gp120 осуществляли количественный анализ с использованием gp120-ELISA с CD4 в неподвижной фазе. Данный анализ свидетельствует (фиг.4), что данные ELISA сопоставимы с данными, полученными в ходе иммунопреципитации, с концентрацией gp120, составляющей 125 нг/мл для синтетического гена gp120 и меньше фонового порога (5 нг/мл) для всех нативных генов gp120. Поэтому экспрессия данного синтетического гена gp120, как минимум, на один порядок превышает наивысшее значение для генов дикого типа gp120. В эксперименте показано, как минимум, 25-кратное увеличение.
Роль rev в экспрессии gp120
Так как rev, по-видимому, оказывает свое действие на нескольких стадиях экспрессии вирусного транскрипта, была проверена возможная роль нетрансляционных эффектов в повышенной экспрессии синтетического гена gp120. Прежде всего, контролировали уровни цитоплазматической мРНК, чтобы исключить возможность, при которой элементы негативных сигналов вызывают повышенную деградацию мРНК или нуклеиновый захват, осуществляли изменение нуклеотидной последовательности. Цитоплазматическую РНК получали путем лизиса NP40 кратковременно трансфицированных клеток 293Т и последующим удалением ядер путем центрифугирования. Цитоплазматическую РНК затем получали из лизатов множественными фенольными экстракциями и осаждением, наносили на нитроцеллюлозу с использованием слот-блот-устройства и, наконец, гибридизовали с оболочечно-специфичным зондом.
Вкратце, цитоплазматическую мРНК клеток 293, трансфицированных CDM&, gp120IIIВ или syngp120, выделяли через 36 часов после трансфекции. Цитоплазматическую РНК клеток He1a, инфицированных вирусом коровьей оспы дикого типа или рекомбинантным вирусом, экспрессирующим gp120 IIIb или синтетический ген gp120, находящийся под контролем промотора 7, 5, выделяли через 16 часов после инфицирования. Равные количества наносили на нитроцеллюлозу с использованием слот-блот-устройства и гибридизовали со случайно меченым 1,5 т.п.н. фрагментами gp120IIIb и syngp120 или человеческим бета-актином. Уровни экспрессии РНК оценивали количественно путем сканирования гибридизованных мембран фосфорсодержащей меткой. Более подробно используемые методы описаны ниже.
Данный эксперимент продемонстрировал, что нет существенной разницы в содержании мРНК в клетках, трансфицированных нативным или синтетическим геном gp120. На самом деле, в некоторых экспериментах уровень цитоплазматической мРНК синтетического гена gp120 был даже ниже, чем нативного гена gp120.
Эти данные были подтверждены путем измерения экспрессии рекомбинантных вирусов коровьей оспы. Человеческие клетки 293 или клетки Hela, инфицированные вирусом коровьей оспы, экспрессировали gp120 IIIb дикого типа или syngp120mn при, как минимум, множественности инфицирования 10. Через 24 часа после заражения и иммунопреципитации с помощью гибридного белка CD4:иммуноглобулин и протеин-А сефарозы собирали супернатанты. Методики, используемые в данном эксперименте, описаны более подробно ниже.
Данный эксперимент показал, что повышенная экспрессия данного синтетического гена все еще наблюдалась, когда продукт эндогенного гена и продукт синтетического гена экспрессировались в рекомбинантах коровьей оспы под контролем сильного смешанного раннего и позднего 7,5 k-промотopa. Поскольку молекулы мРНК вируса коровьей оспы транскрибируются и транслируются в данной цитоплазме, повышенная экспрессия данного синтетического оболочечного гена в этом эксперименте не может приписываться его повышенному экспорту из ядра. Данный эксперимент повторяли в двух дополнительных типах человеческих клеток, клеточной линии злокачественной опухоли 293 и клеток HeLa. Как и в случае трансфицированных клеток 293Т, уровни мРНК были аналогичными в клетках 293, инфицированными любым рекомбинантным вирусом коровьей оспы.
Применение кодонов у лентивируса
Поскольку ясно, что применение кодонов оказывает существенное воздействие на экспрессию в клетках млекопитающих, изучали частоту встречаемости кодонов в генах белков оболочки других ретровирусов. В целом, в данном исследовании не обнаружили четкого примера кодонового предпочтения среди ретровирусов. Однако, когда вирусы из рода лентивирусов, к которому относится HIV-1, анализировались раздельно, использование кодоновой неоднозначности почти идентично тому, которое обнаружено для HIV-1.
Была получена таблица частоты встречаемости кодонов для оболочечных гликопротеинов разнообразных (преимущественно типа С) ретровирусов, за исключением лентивирусов, и создали таблицу сравнения частоты встречаемости кодона с помощью последовательностей белков оболочки четырех лентивирусов, отдаленно родственных HIV-1 (вирус артритного энцефалита коз, вирус инфекционной анемии лошадей, вирус иммунодефицита кошачьих и вирус висны) (табл.2).
Профиль кодонового применения у лентивирусов поразительно аналогичен таковому у HIV-1 во всех случаях, кроме одного, предпочтительный кодон для HIV-1 является тем же, что и предпочтительный кодон для других лентивирусов. Исключением является пролин, который кодируется с помощью ССТ для 41% не относящихся к HIV лентивирусных остатков белков оболочки и с помощью ССА для 40% остатков, ситуация, которая ясно отражает существенное предпочтение для триплета, заканчивающегося А.
Пример использования кодона для нелентивирусных белков оболочки не показывает аналогичного преимущества остатков А, а также не обладает такой же асимметрией в пользу третьего положения остатков С и G, как и при использовании кодона для продуктивно экспрессируемых человеческих генов. В целом, нелентивирусные ретровирусы используют различные кодоны в более равной степени, их профиль совпадает с менее продуктивной экспрессией человеческих генов.
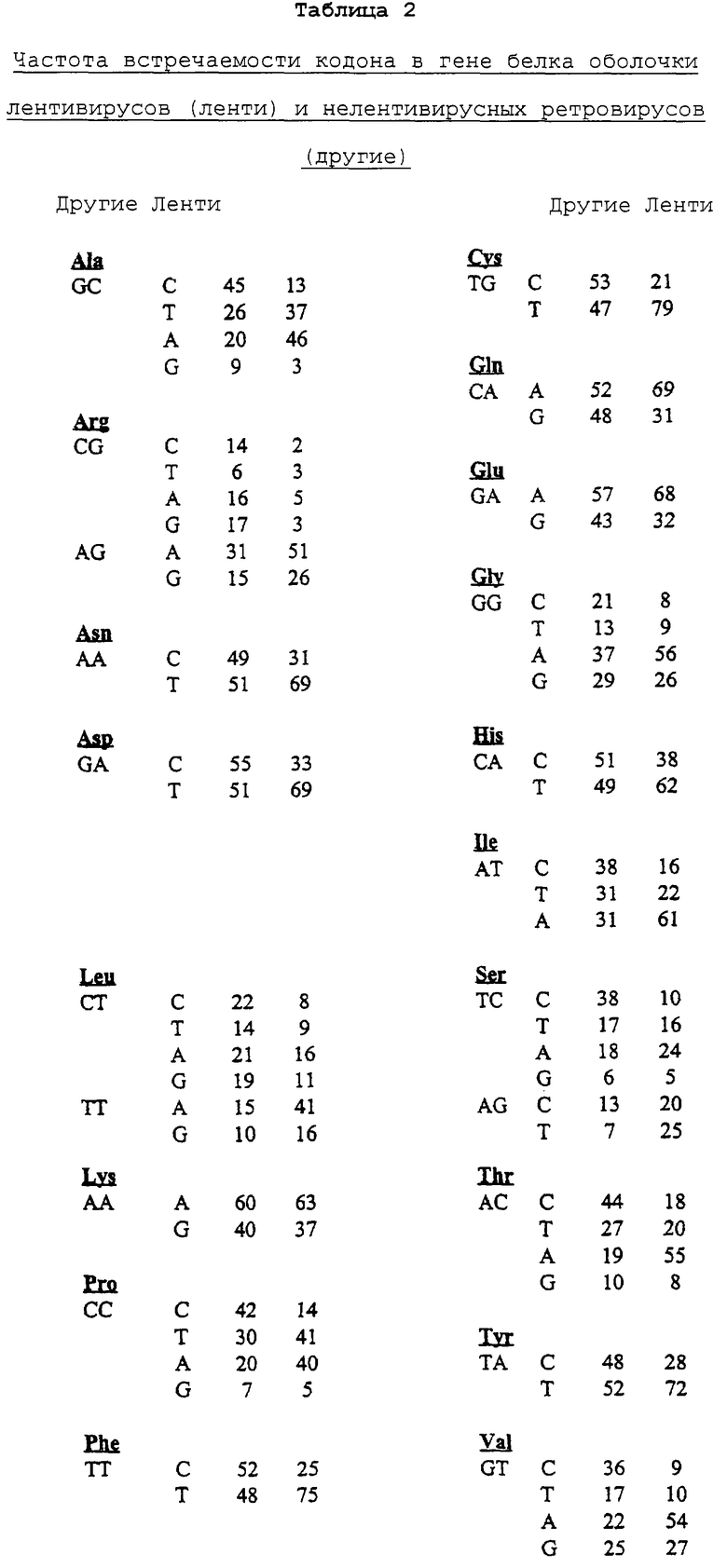
Частоту встречаемости кодона вычисляли с использованием программы GCG, разработанной The University of Wisconsin Genetics Computer Group. Числа соответствуют процентной доле использования отдельного кодона. Кодон, используемый нелентивирусными ретровирусами, составляли из последовательностей предшественника белка оболочки вируса лейкоза крупного рогатого скота, вируса лейкоза кошачьих, вируса Т-клеточного лейкоза человека I типа, лимфотропного вируса Т-клеток человека II типа; культуры, образующие очаг в клетках норки, вируса мышиного лейкоза (MuLV); культуры Раушера, образующие очаг в селезенке, культуры 10А1, амфотропной культуры 4070А и миелопролиферативной культуры вируса лейкоза, и из вируса лейкоза крыс, вируса саркомы человекообразных обезьян, вируса лейкоза Т-клеток человекообразных обезьян, лейкозогенного ретровируса Т1223/В, вируса лейкоза длинноруких обезьян. Таблицы частот встречаемости кодонов для лентивирусов, не относящихся к HIV, SIV составляли из последовательностей предшественника белка оболочки вируса артритного энцефалита коз, вируса инфекционной анемии лошадей, вируса иммунодефицита кошачьих и вируса висна.
Кроме преобладания кодонов, содержащих А, лентивирусные кодоны присоединяются к весьма представительному паттерну CpG в HIV, так, что третье положение для триплетов аланина, пролина, серина и треонина редко представляет собой G. Ретровирусные триплеты для белков оболочки демонстрируют сходство, но менее выражены представляемым CpG. Наиболее явное различие между лентивирусами и другими ретровирусами, в отношении преобладания CpG заключается в использовании варианта CGX в аргининовых триплетах, которые довольно часто представлены среди кодирующих последовательностей оболочки ретровируса, но почти никогда не присутствуют среди сопоставляемых лентивирусных последовательностей.
Различия в rev зависимости между нативным и синтетическим gp120
Чтобы установить, действительно ли регуляция с помощью rev связана с использованием кодона HIV-1, изучали влияние rev на нативный и синтетический ген. Так как регуляция с помощью rev нуждается в rev-связывающем сайте RRE в cis, были созданы конструкты, в которых этот связывающий сайт клонировали в 3'-нетранслируемую область данного нативного и данного синтетического гена. Эти плазмиды котрансфицировали с rev или с контрольной плазмидой в trans в клетки 193Т и проводили полуколичественный анализ уровня экспрессии gp120 в супернатантах путем иммунопреципитации. Методики, используемые в данном эксперименте, описаны более подробно ниже.
Как показано на фиг.5А и фиг.5В, rev положительно регулирует нативный ген gp120, но не оказывает влияния на экспрессию синтетического гена gp120. Следовательно, действие rev на субстрат, в котором отсутствует последовательность из эндогенных вирусных оболочечных последовательностей, не очевидно.
Экспрессия синтетического гена ratTHY-1 с кодонами для белков оболочки HIV
Вышеописанный эксперимент позволяет предположить, что фактически “последовательности оболочки” присутствуют для rev-регуляции. Для того чтобы проверить эту гипотезу, получали синтетический вариант гена, кодирующего небольшой, обычно продуктивно экспрессируемый белок клеточной поверхности, антиген ratTHY-1. Данный синтетический вариант гена ratTHY-1 предназначался для использования кодона, аналогичного HIV gp120. При создании этого синтетического гена последовательности AUUUA, которые связывают с нестабильностью мРНК, исключали. Кроме того, для упрощения обработки полученного гена (фиг.6) интродуцировали два сайта рестрикции. Данный синтетический ген с использованием кодонов для белков оболочки HIV (rTHY-lenv) конструировали с использованием трех 150-170-мерных олигонуклеотидов (фиг.7). В противоположность гену syngp120mn, продукты ПЦР клонировали сразу и осуществляли сборку в pUC12, a затем клонировали в pCDM7.
Уровни экспрессии нативного rTHY-1 и rTHY-1 с кодонами для белков оболочки HIV оценивали количественно с помощью иммунофлуоресценции временно трансфицированных клеток 293Т. На фиг.8 показано, что экспрессия нативного гена THY-1 почти на два порядка превышает уровень фона контрольных трансфицированных клеток (pCDM7). Напротив, экспрессия синтетического ratTHY-1 практически намного ниже, чем у указанного нативного гена (показано путем сдвига пика в направлении к обозначению канала).
Чтобы подтвердить, что негативные элементы последовательности, способствующие деградации мРНК, не были интродуцированы, получали конструкт, в котором ген rTHY-1 был клонирован по 3'-концу синтетического гена gp120 (фиг.9В). В данном эксперименте клетки 293Т трансфицировали геном syngp120mn или гибридным геном syngp120/ratTHY-1 env (syngp120mn. rTHY-1env). Экспрессию измеряли по методу иммунопреципитации с помощью гидридного белка CD4:IgG и протеин-А агарозы. Методы, используемые в данном эксперименте, более подробно описаны ниже.
Так как синтетический ген gp120 содержит стоп-кодон UAG, из данного транскрипта rTHY-1env не транслируется. Если негативные элементы, усиливающие деградацию, присутствуют в данной последовательности, содержание белка gp120, экспрессируемого из данного конструкта, должно быть снижено по сравнению с конструктом syngp120mn без rTHY-1env. На фиг.9А показано, что экспрессия обоих конструктов аналогична, свидетельствуя тем самым, что низкая экспрессия должна быть связана с трансляцией.
Rev-зависимая экспрессия синтетического гена ratTHY-1 с кодонами для белков оболочки
Чтобы выяснить действительно ли rev способен регулировать экспрессию гена ratTHY-1, содержащего кодоны env, создали конструкт с rev-связывающим сайтом на 3'-конце открытой рамки считывания rTHY-lenv. Для измерения реактивности rev конструкта ratTHY-1env, содержащего 3'-RRE, человеческие клетки 293Т котрансфицировали ratTHY-1envrre и либо CDM7, либо pCMVrev. Через 60 часов после трансфекции клетки отслаивали с помощью 1 мМ ЭДТА в ФБР и окрашивали с помощью 0Х7 анти-rTHY-1 мышиными моноклональными антителами и вторичными антителами, конъюгированными с ФИТЦ. Интенсивность флуоресценции измеряли с использованием цитофлуорометра EPICS XL. Более подробно эти методы описаны ниже.
В повторных экспериментах, когда rev был контрансфицирован геном rTHY-1env, обнаружили некоторое повышение экспрессии rTHY-1env. Для дальнейшего повышения чувствительности аналитической системы получали конструкт, экспрессирующий секретируемый вариант rTHY-lenv. Данный конструкт должен был давать более надежные данные, потому что накопленное количество белка в супернатанте отражает получение продукции белка в течение продолжительного периода, в противоположность белку, экспрессируемому на поверхности, что более точно отражает текущую скорость продукции. Ген, способный экспрессировать секретируемую форму, получали с помощью ПЦР с использованием прямых и обратных праймеров, путем отжига, соответственно 3' из эндогенной лидерной последовательности и 5' из мотива последовательности, необходимой для фосфатидилинозитолг-ликановом якоря. Продукт ПЦР клонировали в плазмиду, которая уже содержит лидерную последовательность CD5, создавая таким образом конструкт, в которой мембранный якорь был удален, а данную лидерную последовательность заменили гетерологичным (и, вероятно, более эффективным) лидерным пептидом.
Реактивность rev секретируемой формы ratTHY-1env измеряли путем иммунопреципитирования супернатантов человеческих клеток 293Т, котрансфицированных плазмидой, экспрессирующей секретируемую форму ratTHY-1env и RRE-последовательность в cis (rTHY-1envPI-rre) и CDM7 или pCMVrev. Конструкт rTHY-1envPI-RRE создавали с помощью ПЦР с использованием олигонуклеотида: cgcggggctagcgcaaa-gagtaataagtttaac (SEQ ID NO: 38) в качестве прямого праймера, с использованием олигонуклеотида: cgcggatcccttgtattttgtactaata (SEQ ID NO: 39) в качестве обратного праймера и синтетической конструкции rTHY-1env в качестве матрицы. После расщепления с помощью Nhe1 и Not1 полученный PCR-фрагмент клонировали в плазмиду, содержащую лидерную последовательность CD5 и последовательность RRE. Супернатанты клеток, меченных 35S, собирали через 72 часа после трансфекции, преципитировали мышиными моноклональными антителами 0Х7 против rTHY-1 и на сефарозе, конъюгированной с антителом против мышиного IgG, и разгоняли в 12%-ном восстанавливающем SDS-PAGE.
В этом эксперименте индукция rTHY-1 с помощью rev была намного более заметной и четкой, чем в вышеописанном эксперименте, и основательно подтверждала, что rev способен трансляционно регулировать транскрипты, которые супрессированы редко используемыми кодонами.
Rev-зависимая экспрессия гибридного белка rTHY-1env: иммуноглобулин
Чтобы проверить, действительно ли редко используемые кодоны должны быть представлены по всей целой кодирующей последовательности или действительно ли короткая область является достаточной для придания реактивности rev, был получен гибридный белок rTHY-1env: иммуноглобулин. В данной конструкции ген rTHY-1env (без последовательности мотива, ответственного за фосфатидилинозитолгликановый якорь) сцеплен с шарниром человеческого IgG1, доменами СН2 и СН3. Эту конструкцию получали с помощью якорной ПЦР с использованием праймеров с сайгами рестрикции Nhe1 и BamH1 и rTHY-1env в качестве матрицы. Полученный фрагмент ПЦР клонировали в плазмиду, содержащую лидерную последовательность из CD5-поверхностной молекулы и указанного шарнира, частей СН2 и СН3 человеческого иммуноглобулина IgG. Фрагмент Hind3/Eag1, содержащий вставку rTHY-1enveg1, клонировали затем в полученную pCDM7-плазмиду с последовательностью RRE.
Чтобы измерить ответ гибридного гена rTHY-1env/иммуноглобулин (rTHY-1enveg1rre) на rev, человеческие клетки 293Т котрансфицировали с помощью rTHY-1enveglrre и либо - pCDM7, либо - pCMVrev. Конструкт rTHY-1enveg1rre создавали с помощью якорной ПЦР с использованием прямого и обратного праймеров, соответственно с сайтами рестрикции Nhe1 и BamH1. Полученный фрагмент ПЦР клонировали в плазмиду, содержащую СВ5-лидер и шарнир человеческого IgG, домены СН2 и СH3. Супернатанты клеток, меченных 35S, собирали через 72 часа после трансфекции, преципитировали с помощью мышиного моноклонального антитела 0Х7 к rTHY-1 и антимышиный IgG-сефарозе и разгоняли в 12%-ном восстанавливающем SDS-PAGE. Используемые здесь методики подробно описаны ниже.
Как и в случае продукта гена rTHY-1envPI, данный гибридный белок rTHY-1env/иммуноглобулин выделяется в супернатант. Следовательно, данный ген должен отвечать на rev-индукцию. Однако в противоположность rTHY-1envPI котрансфекция rev в trans не индуцирует или индуцирует лишь незначительное повышение экспрессии rTHY-1enveg1.
Экспрессию гибридного белка rTHY-1: иммуноглобулин с помощью кодонов для белков оболочки нативного rTHY-1 или HIV измеряли путем иммунопреципитации. Вкратце, человеческие клетки 293Т, трансфицировали rTHY-1enveg1 (env-кодоны) или rTHY-1wteg1 (нативные кодоны). Конструкт rTHY-1wteg1 получали способом, аналогичным способу, используемому для конструирования rTHY-1enveg1, за тем исключением, что плазмида, содержащая нативный ген rTHY-1, использовалась в качестве матрицы. Супернатанты клеток, меченных 35S, собирали через 72 часа после трансфекции, преципитировали с помощью мышиного моноклонального антитела 0Х7 против rTHY-1 и анти-мышиной IgG-сефарозой и разгоняли в 12%-ном восстанавливающем SDS-PAGE. Указанные методики, используемые в данном эксперименте, более подробно описаны ниже.
Уровни экспрессии rTHY-1enveg1 были снижены по сравнению с аналогичным конструктом rTHY-1 дикого типа в виде гибридного партнера, но все еще были существенно выше, чем rTHY-1env. В соответствии с этим, обе части гибридного белка оказывают влияние на уровни экспрессии. Добавление rTHY-1env не ограничивает экспрессию на равную величину, как это видно для одного rTHY-1env. Следовательно, регуляция с помощью rev неэффективна, если экспрессия белка супрессирована не полностью.
Кодоновое предпочтение для генов белков оболочки HIV-1
Прямое сравнение частоты использования кодона для белка оболочки HIV и высокопродуктивно экспрессируемых человеческих генов обнаруживает четкое различие по всем двадцати аминокислотам. Простое измерение статистической значимости преимущества данного кодона обнаруживает, что среди девяти аминокислот с двукратной кодоновой вырожденностью, предпочтение отдано третьему остатку, который представляет собой А или U во всех девяти. Возможность того, что все девять, выбранные с одинаковой вероятностью, будут одними и теми же составляет, приблизительно, 0,004, и, следовательно, путем любого традиционного измерения выбор третьего остатка не может рассматриваться как случайный. Дополнительное доказательство асимметрии кодонового предпочтения обнаружено среди наиболее вырожденных кодонов, где можно наблюдать сильный отбор по триплетам, несущим аденин. Это контрастирует с паттерном для высокопродуктивно экспрессируемых генов, которые благоприятствуют кодонам, несущим С или, менее обычно, G в третьей позиции кодонов с трехкратной или более вырожденностью.
Систематическая замена нативных кодонов на кодоны высокопродуктивно экспрессируемых человеческих генов резко повышает экспрессию gp120. Количественный анализ с помощью ELISA показывает, что экспрессия данного синтетического гена была, как минимум, в 25 раз выше по сравнению с нативным gp120 после временной трансфекции в человеческих клетках 293. Концентрационные уровни в данном ELISA-эксперименте были значительно ниже. Так как ELISA используется для количественного анализа, который основан на связывании gp120 с CD4, обнаруживается лишь нативный, не денатурированный материал. Это может объяснять явно низкую экспрессию. Измерения уровней цитоплазматической мРНК демонстрируют, что разница в экспрессии белка обусловлена различиями в трансляции, а не стабильностью мРНК.
Ретровирусы обычно не проявляют подобного предпочтения в отношении А и Т, как обнаружено для HIV. Но если данное семейство разделить на две подгруппы, лентивирусы и нелентивирусные ретровирусы, аналогичное предпочтение для А и, менее часто, Т, определяется по третьему кодоновому положению для лентивирусов.
Таким образом, полученное доказательство дает основание полагать, что лентивирусы сохраняют характеристический паттерн кодонов для белков оболочки не вследствие исходного преимущества в обратной транскрипции или репликации таких остатков, а скорее в силу некоторой причины, присущей физиологии данного класса вирусов. Основное различие между лентивирусами и некомплексными ретровирусами заключается, как уже упоминалось, в дополнительных регуляторных и несущественных вспомогательных генах у лентивирусов. Таким образом, одним из простых объяснений ограничения экспрессии белков оболочки может заключаться в том, что на ней основан существенный регуляторный механизм одной из этих дополнительных молекул. Действительно, известно, что один из этих белков, rev, вероятнее всего обладает гомологией у всех лентивирусов. Поэтому кодон, используемый в вирусной мРНК, используется для создания класса транскриптов, которые восприимчивы к стимулирующему действию rev. Эта гипотеза была подтверждена с использованием стратегии, аналогичной вышеприведенной, однако на этот раз используемый кодон изменили в обратном направлении. Кодон, используемый в эффективно экспрессируемом клеточном гене, был заменен на более часто используемые кодоны в оболочке HIV. Предполагается, что уровни экспрессии были существенно ниже в сравнении с нативной молекулой почти на два порядка, когда анализировались путем иммунофлуоресценции поверхности экспрессируемой молекулы. Если rev коэкспрессировали в trans, a RRE-элемент присутствовал в cis, для поверхностной молекулы обнаруживалась лишь слабая индукция. Однако, если THY-1 экспрессировался в виде секретируемой молекулы, индукция с помощью rev была намного отчетливей, подтверждая вышеприведенную гипотезу. По-видимому, это можно объяснить накоплением секретируемого белка в супернатанте, который существенно преумножает эффект rev. Если rev, в целом, индуцирует лишь минорное повышение содержания поверхностных молекул, индукция белков оболочки HIV с помощью rev не может иметь результатом избыточное увеличение поверхности, но скорее повышение уровня внутриклеточного gp160. В настоящее время нет полной ясности почему это должно случиться.
Чтобы проверить, действительно ли небольшие субтотальные элементы гена достаточны для ограничения экспрессии и вызывают ее, были получены rev-зависимые гибридные белки rТНY1еnv: иммуноглобулин, в которых только около одной трети всего гена использует кодоны для белков оболочки. Экспрессионные уровни данной конструкции находились на промежуточном уровне, свидетельствуя, что элемент негативной последовательности rTHY-1env не является доминантным в пределах иммуноглобулиновой части. Данный гибридный белок не был или только слегка был rev-респонсивным, свидетельствуя, что только полностью супрессированные гены могут быть rev-реактивными.
Другой характеристической особенностью, которая была обнаружена в таблицах частоты встречаемости кодона, является поразительно малая представительность CpG триплетов. В сравнительном исследовании использования кодонов у Е. coli дрожжей, дрозофилы и приматов было показано, что для многочисленных анализируемых генов приматов, по меньшей мере, 8 используемых кодонов содержат все кодоны с динуклеотидной последовательностью CpG. Исключение кодонов, содержащих этот динуклеотидный мотив, было также обнаружено в последовательности других ретровирусов. По-видимому, вероятно, что причина меньшего представления триплетов, несущих CpG, отчасти создается вследствие исключения гена, замолкающего из-за метилирования CpG цитозинов. Ожидаемая численность динуклеотидов CpG в HIV, в целом, составляет почти одну пятую от ожидания на основе гетероциклической структуры. Это может указывать, что возможность высокопродуктивной экспрессии восстанавливается и что данный ген, в действительности, эффективно экспрессируется в определенный момент во время вирусного патогенеза.
Результаты, представленные здесь, ясно свидетельствуют, что кодоновое предпочтение оказывает сильный эффект на содержание белков и дает основание полагать, что рост цепи при трансляции контролирует экспрессию гена млекопитающего. Однако и другие факторы могут играть роль. Во-первых, избыток субмаксимально нагруженных молекул мРНК в эукариотических клетках указывает, что инициация является нормой, лимитирующей трансляцию, по крайней мере, в некоторых случаях, иначе все транскрипты были бы покрыты рибосомами. Кроме того, если бы останавливающиеся рибосомы и последующая деградация мРНК были таким механизмом, супрессия с помощью редких кодонов могла бы быть, вероятно, необратима при любом регуляторном механизме, подобно механизму, представленному здесь. Возможное объяснение влияния инициации и элонгации на трансляционную активность заключается в том, что скорость инициации или избыток рибосом контролируется, частично, сигналами, распределенными по всей данной РНК, так что лентивирусные кодоны склоняют РНК к накоплению пула из скудно инициированных молекул РНК. Однако данное ограничение не обязательно является кинетическим; например, выбор кодонов мог бы повлиять на вероятность того, что данный трансляционный продукт, однажды инициированный, должным образом укомплектовывается полностью. При данном механизме избыток менее благоприятных кодонов приводил бы к существенной кумулятивной вероятности неудачи в комплектации образующейся полипептидной цепи. Тогда бы изолированная РНК повышала бы скорость инициации под действием rev. Поскольку адениновые остатки находятся в избытке в rev-респонсивных транскриптах, это означало бы, что метилирование аденина РНК опосредует данную супрессию трансляции.
Детализация методик
В вышеописанных экспериментах использовались нижеследующие методики.
Анализ последовательности
Анализ последовательности осуществляли в соответствии с программным обеспечением, разработанным The University of Wisconsin Genetics Computer Group.
Конструирование плазмид
Конструирование плазмид создавали с помощью нижеследующих методов. Векторы и инсерцированную ДНК расщепляли при концентрации 0,5 мкг/10 мкл в надлежащем рестрикционном буфере в течение 1-4 часов (общий реакционный объем, приблизительно, 30 мкл). Расщепленный вектор обрабатывали с помощью 10% (о/о) щелочной фосфатазы кишечника теленка (1 мкг/мл) в течение 30 минут перед гелевым электрофорезом. Гидролизаты вектора и вставки (каждый 5-10 мкл) разгоняли в 1,5% геле легкоплавкой агарозы с ТАЕ-буфером. Кусочки геля, содержащие нужные полосы, переносили в 1,5 мл реакционную пробирку, расплавляли при 65°С и сразу вносили для лигирования, не удаляя агарозу. Лигаты обычно получали в общем объеме 25 мкл в 1х низкосолевом буфере с 1х лигирующими добавками 200-400 Eg лигазы, 1 мкл вектора и 4 мкл вставки. При необходимости, 5'-выступающие концы заполняли путем добавления 1/10 объема из 250 мкМ dNTPs и 2-5 Е полимеразы Кленова для инактивирования нагреванием или экстракции гидролизатов фенолом и инкубировали в течение 20 мин при комнатной температуре. При необходимости, 3'-выступающие концы заполняли путем добавления 1/10 объема из 2,5 мМ дНТФ и 5-10 Eg ДНК-полимеразы T4, чтобы инактивировать нагреванием или экстрагировать гидролизаты фенолом, с последующей инкубацией при 37°С в течение 30 мин. В этих реакциях использовали нижеследующие буферы: 10х низкосолевой буфер (60 мМ трис-HC1, рН 7,5, 60 мМ MgCl2, 50 мМ NaCl, 4 мг/мл BSA, 70 мМ (β-меркаптоэтанола, 0,02% NaN3); 10х среднесолевой буфер (60 мМ трис-НСl, рН 7,5, 60 мМ MgCl2, 50 мМ NaCl, 4 мг/мл BSA, 70 мМ β-меркаптоэтанола, 0,02% NаN3); 10х высокосолевой буфер (60 мМ трис-HCl, рН 7,5, 60 мМ MgCl2, 50 мМ NaCl, 4 мг/мл BSA, 70 мМ β-меркаптоэтанола, 0,02% NаN3); 10х лигирующие добавки (1 мМ АТФ, 20 мМ ДТТ, 1 мг/мл BSA, 10 мМ спермидина); 50х ТАЕ (2 М трис-ацетат, 50 мМ ЭДТА).
Олигонуклеотидный синтез и выделение очисткой
Олигонуклеотиды получали на синтезаторе Milligen 8750 (Millipore). Колонки элюировали с помощью 1 мл 30%-ного аммония гидроксида, а элюированные олигонуклеотиды деблокировали при 55°С в течение 6-12 часов. После деблокирования 150 мкл олигонуклеотида преципитировали с помощью 10х объема ненасыщенного н-бутанола в 1,5 мл реакционных пробирках, с последующим центрифугированием при 15000 об/мин в микроцентрифуге. Осадок промывали с помощью 70%-ного этанола и ресуспендировали в 50 мкл H2O. Полученную концентрацию определяли путем измерения оптической плотности при 260 нм для разведения 1:333 (1 OD260=30 мкг/мл).
Для конструирования синтетического гена gp120 использовали нижеследующие олигонуклеотиды (все последовательности, приведенные в данном тексте, находятся в 5'-3'-ориентации).
Олигонуклеотид 1 прямой (Nhel): сдс ggg cta gсс асе gаg ааg ctg (SEQ ID NO: 1).
Олигонуклеотид 1: асc gag aag ctg tgg gtg acc gtg tac tac ggc gtg ccc gtg tgg aag ag ag gcc асc асc асc ctg ttc tgc gcc agc gac gcc aag gcg tac gac acc gag gtg cac aac gtg tgg gcc ace cag gcg tgc gtg ccc acc gac ccc aac ccc cag gag gtg gag ctc gtg aac gtg acc gag aac ttc aac at (SEQ ID NO:2).
Олигонуклеотид 1 обратный: сса сса tgt tgt tct tcc аса tgt tga agt tct с (SEQ ID NO:3).
Олигонуклеотид 2 прямой: gac cga gaa ctt caa cat gtg gaa gaa caa cat (SEQ ID NO:4).
Олигонуклеотид 2: tgg aag aac aac atg gtg gag cag atg cat gag gac atc atc agc ctg tgg gac cag ag ctg aag ccc tgc gtg aag ctg acc cc ctg tgc gtg acc tg aac tgc acc gac ctg agg aac acc acc aac acc aac ac agc acc gcc aac aac aac agc aac agc gag ggc acc atc aag ggc ggc gag atg (SEQ ID NO:5).
Олигонуклеотид 2 обратный (Psti): gtt gaa gct gca gtt ctt cat ctc gcc gcc ctt (SEQ ID NO:6).
Олигонуклеотид 3 прямой (Pstl): gaa gaa ctg cag ctt caa cat cac cac cag с (SEQ ID NO:7).
Олигонуклеотид 3: aac atc асе асе agc atc cgc gac aag atg cag aag gag tac gcc ctg ctg tac aag ctg gat atc gtg agc atc gac aac gac agc acc agc tac cgc ctg atc tcc tgc aас асc agc gtg atc асc саg gсс tgc ccc aag atc agc ttc gag ccc atc ccc atc cac tac tgc gсс ccc gсс ggc ttc gсс (SEQ ID NO:8).
Олигонуклеотид 3 обратный: gaa ctt ctt gtc ggc ggc gaa gcc ggc ggg (SEQ ID NO:9).
Олигонуклеотид 4 прямой: gcg ccc ccg ccg gct tcg cca tcc tga agt gca acg аса аgа agt tc (SEQ ID NO:10).
Олигонуклеотид 4: gcc gac aag aag ttc agc ggc aag ggc agc tgc aag aac gtg agc acc gtg cag tgc acc cac ggc atc cgg ccg gtg gtg agc acc cag ctc ctg ctg aac ggc agc ctg gcc gag gag gag gtg gtg atc cgc agc gag aac ttc acc gac aac gcc aag acc atc atc gtg cac ctg aat gag agc gtg cag atc (SEQ ID NO:11)
Олигонуклеотид 4 обратный (Mlu1): agt tgg gac gcg tgc agt tga tct gca cgc tct с (SEQ ID NO:12).
Олигонуклеотид 5 прямой (Mlu1): gag agc gtg cag atc aac tgc acg cgt ccc (SEQ ID NO:13).
Олигонуклеотид 5; aac tgc acg cgt ccc aac tac aac aag cgc aag cgc atc cac atc ggc ccc ggg cgc gcc ttc tac acc acc aag aac atc atc ggc acc atc ctc cag gcc cac tgc aac atc tct aga (SEQ ID NO:14).
Олигонуклеотид 5 обратный: gtc gtt cca ctt ggc tct aga gat gtt gca (SEQ ID NO:15).
Олигонуклеотид 6 прямой: gca аса tct cta gag cca agt gga acg ac (SEQ ID NO:16).
Олигонуклеотид 6: gcc aag tgg aac gac acc ctg cgc cag atc gtg agc aag ctg aag gag cag ttc aag aac aag acc atc gtg ttc ас сад agc agc ggc ggc gac ccc gag atc gtg atg cac agc ttc aac tgc ggc ggc (SEQ ID NO:17).
Олигонуклеотид 6 обратный (EcoR1): gca gta gaa gaa ttc gcc gcc gca gtt gc (SEQ ID NO:18).
Олигонуклеотид 7 прямой (EcoR1): tca act gcg gcg gcg aat tct tct act gc (SEQ ID NO:19).
Олигонуклеотид 7: ggc gaa ttc ttc tac tgc aac acc agc ccc ctg ttc aac agc acc tgg aac ggc aac aac acc tgg aac aac acc acc ggc agc aac aac aat att acc ctc cag tgc aag atc aag cag atc atc aac atg tgg cag gag gtg ggc aag gcc atg tac gcc ccc ccc atc gag ggc cag atc cgg tgc agc agc (SEQ ID NO:20)
Олигонуклеотид 7 обратный: gca gac cgg tga tgt tgc tgc tgc acc gga tct ggc cct с (SEQ ID NO:21).
Олигонуклеотид 8 прямой: cga ggg cca gat ccg gtg cag cag caa cat cac cgg tct g (SEQ ID NO:22).
Олигонуклеотид 8: aac atc асc ggt ctg ctg ctg acc cgc gac ggc ggc aag gac acc gac acc aac gac acc gac atc ttc cgc ccc ggc ggc ggc gac atg cgc gac aac tgg aga tct gag ctg tac aag tac aag gtg gtg acg atc gag ccc ctg ggc gtg gcc ccc acc aag gcc aag cgc cgc gtg gtg cag cgc gag aag cgc (SEQ ID NO:23).
Олигонуклеотид 8 обратный (Not1): cgc ggg cgg ccg ctt tag cgc ttc tcg cgc tgc асе ас (SEQ ID NO:24).
Для конструирования гена ratTHY-lenv использовались нижеследующие олигонуклеотиды.
олигонуклеотид 1 прямой (Bam.H1/Hind.3): cgc ggg gga tcc aag ctt acc atg att cca gta ata agt (SEQ ID NO:25);
олигонуклеотид 1: atg aat cca gta ata agt ata аса tta tta tta agt gta tta caa atg agt aga gga caa aga gta ata agt tta аса gca tct tta gta aat caa aat ttg aga tta gat tgt aga cat gaa aat aat аса aat ttg cca ata caa cat gaa ttt tca tta acg (SEQ ID NO:26);
олигонуклеотид 1 обратный (EcoR1/Mlu1): cgc ggg gaa ttc acg cgt taa tga aaa ttc atg ttg (SEQ ID NO:27);
олигонуклеотид 2 прямой (BamH1/Mlu1): cgc gga tcc acg cgt gaa aaa aaa aaa cat (SEQ ID NO:28);
олигонуклеотид 2: cgt gaa aaa aaa aaa cat gta tta agt gga аса tta gga gta cca gaa cat аса tat aga agt aga gta aat ttg ttt agt gat aga ttc ata aaa gta tta аса tta gca aat ttt аса аса aaa gat gaa gga gat tat atg tgt gag (SEQ ID NO:29);
олигонуклеотид 2 обратный (EcoR1/Sac1): cgc gaa ttc gag ctc аса cat ata atc tcc (SEQ ID NO:30);
олигонуклеотид 3 прямой (BamH1/Sac1): cgc gga tcc gag ctc aga gta agt gga caa (SEQ ID NO:31);
олигонуклеотид 3: ctc aga gta agt gga caa aat cca аса agt agt aat aaa аса ata aat gta ata aga gat aaa tta gta aaa tgt ga gga ata agt tta tta gta caa aat аса agt tgg tta tta tta tta tta tta agt tta agt ttt tta caa gca аса gat ttt ata agt tta tga (SEQ ID NO:32);
олигонуклеотид 3 обратный (EcoR1/Not1): cgc gaa ttc gcg gcc gct tca taa act tat aaa ate (SEQ ID NO:33).
Полимеразная цепная реакция
Вкратце, перекрывающиеся 15-25-мерные олигонуклеотиды, отожженные по обоим концам, использовали для амплификации длинных олигонуклеотидов в полимеразной цепной реакции (ПЦР). Обычными условиями ПЦР были: 35 циклов, 55°С температура отжига, 0,2 с время удлинения. Продукты ПЦР выделяли очисткой в геле, экстрагировали фенолом и использовали в последующей ПЦР для получения более длинных фрагментов, состоящих из двух небольших смежных фрагментов. Эти более длинные фрагменты клонировали в CDM7-произведенную плазмиду, содержащую лидерную последовательность из СВ5-поверхностной молекулы с последующим полилинкером Nhe1/Pst1/Mlu1/EcoR1/BamH1.
В этих реакциях использовались нижеследующие растворы: 10х PCR-буфер (500 мМ КС1, 100 мМ трис-HCl, рН 7,5, 8 мМ MgCl2, 2 мМ каждого дНТФ). Данный конечный буфер дополняли 10% ДМСО для точности воспроизведения Taq-полимеразы.
Получение ДНК в небольшом количестве
Трансформированные бактерии выращивали в 3-х мл LB-культуры в течение более чем 6 часов или в течение ночи. Приблизительно 1,5 мл каждой культуры наливали в 1,5 мл-овые микроцентрифужные пробирки, крутили в течение 20 с для осаждения клеток и ресуспендировали в 200 мкл раствора I. Затем добавляли 400 мкл раствора II и 300 мкл раствора III. Эти микроцентрифужные пробирки закрывали колпачками, перемешивали и крутили в течение > 30 с. Супернатанты переносили в чистые пробирки и однократно экстрагировали фенолом. ДНК осаждали, заполняя эти пробирки изопропанолом, перемешивали и откручивали в микроцентрифуге в течение > 2 мин. Полученный осадок промывали 70%-ным этанолом и ресуспендировали в 50 мкл dH20, содержащем 10 мкл РНКазы А. В этих операциях использовались нижеследующая среда и растворы: LB-среда (1,0% NaCl, 0,5% дрожжевого экстракта, 1% триптона); раствор I (10 мМ ЭДТА, рН 8,0); раствор II (0,2 М NaOH, 1,0% SDS); раствор III (2,5 M KOAc, 2,5 М ледяной уксусной кислоты); фенол (рН доводили до 6,0, насыщенного ТЭ); ТЭ (10 мМ Трис-HCl, рН 7,5 мМ ЭДТА, рН 8,0).
Получение ДНК в большом количестве
1 л трансформированных бактерий выращивали в течение 24-36 часов (МС1061р3, трансформированные производными pCDM) или 12-16 часов (МС1061, трансформированные производными pUC) при 37°С либо в бактериальной среде М9 (производные pCDM) или LB (производные pUC). Бактерии центрифугировали в 1 л бутылках с использованием центрифуги Beckman J6 при 4200 об/мин, в течение 20 мин. Полученный осадок ресуспендировали в 40 мл раствора I. Затем добавляли 80 мл раствора II и 40 мл раствора III, и эти бутылки довольно энергично встряхивали до образования комков размером от 2-3 мм. Такую бутылку откручивали при 4200 об/мин в течение 5 мин, и полученный супернатант выливали через марлю в 250 мл-овую бутыль.
Наверх добавляли изопропанол и данную бутыль центрифугировали при 4200 об/мин в течение 10 мин.
Полученный осадок ресуспендировали в 4,1 мл раствора I и добавляли хлорид цезия до 4,5 г, 0,3 мл бромида этидия (10 мг/мл) и 0,1 мл 1%-ного раствора Тритон Х100. Пробирки центрифугировали в высокоскоростной центрифуге Beckman J2 при 10000 об/мин в течение 5 мин. Полученный супернатант переносили в ультрацентрифужные пробирки Beckman Quick Seal, которые затем силиконизировали и центрифугировали в ультрацентрифуге Beckman с использованием NVT90-poropa с фиксированным углом при 80000 об/мин в течение > 2,5 часов. Полученную полосу извлекали под видимым светом с использованием 1 мл шприца и иглы 20-го размера. К извлеченному материалу добавляли равный объем Н2О. ДНК однократно экстрагировали с помощью н.-бутанола, насыщенного 1 М хлоридом натрия с последующим добавлением равного объема 10 М ацетата аммония / 1 мМ ЭДТА. Полученный материал выливали в 13 мл-закрывающуюся пробирку, которую затем наполняли доверху абсолютным этанолом, перемешивали и центрифугировали в центрифуге Beckman J2 при 10000 об/мин в течение 10 мин. Полученный осадок промывали 70%-ным этанолом и ресуспендировали в 0,5-1 мл Н2О. Концентрацию полученной ДНК определяли путем измерения оптической плотности при 260 нм в разведении 1:200 (1 OD260=50 мкг/мл).
В этих операциях использовались нижеследующие среда и буферы: бактериальная среда М9 (10 г солей М9, 10 г казаминовых кислот (гидролизованных), 10 мл добавок М9, 7,5 мкг/мл тетрациклина (500 мкл исходного раствора (15 мг/мл)), 12,5 мкг/мл ампициллина (125 мкл исходного раствора (10 мг/мл) ); добавки М9 (10 мМ CaCl2, 100 мМ MgSO4, 200 мкг/мл тиамина, 70% глицерин); LB-среда (1,0% NaCl, 0,5% дрожжевого экстракта, 1% триптона); раствор I (10 мМ ЭДТА, рН 8,0); раствор II (0,2 М NaOH, 1,0% ДДС); раствор III (2,5 М КОАс, 2,5 М НОАс).
Секвенирование
Синтетические гены секвенировали по дидезоксинуклеотидному методу Сэнгера. Вкратце, 20-50 мкг двухцепочечной плазмидной ДНК денатурировали в 0,5 М NaOH в течение 5 мин. Затем эту ДНК осаждали с помощью 1/10 объема ацетата натрия (рН 5,2) и 2 объемами этанола и центрифугировали в течение 5 мин. Полученный осадок промывали 70%-ным этанолом и ресуспендировали при концентрации 1 мкг/мкл. Реакцию ренатурации осуществляли с помощью 4 мкг матричной ДНК и 40 нг праймера в 1х ренатурационном буфере в конечном объеме 10 мкл. Данную реакцию нагревали до 65°С и медленно охлаждали до 37°С.
Для каждой реакции в отдельную трубку вносили 1 мкл 0,1 М DTT, 2 мкл смеси для мечения, 0,75 мкл dH2O, 1 мкл [35S] дАТФ (10 мкКи) и 0,25 мкл Sequenase™ (12 Е/мл). 5 мкл этой смеси вносили в каждую пробирку отжигаемой прай-мер-матрицы и инкубировали в течение 5 мин при комнатной температуре. Для каждой реакции мечения добавляли 2,5 мкл, каждой, из 4-х терминирующей смеси в планшет Terasaki и предварительно нагревали при 37°С. В конце данного инкубационного периода в каждую из 4-х терминирующих смесей вносили 3,5 мкл смеси для мечения. Через 5 мин в каждую реакцию добавляли 4 мкл стоп-раствора и этот планшет Terasaki инкубировали при 80°С в течение 10 мин в термостате. Полученные реакции секвенирования электрофорезировали в 5%-ном денатурирующем полиакриламидном геле. Акриламидный раствор готовили путем добавления 200 мл 10х ТБЭ-буфера и 975 мл dH2О со 100 г акриламид:метиленбисакриламида (29:1). 5%-й полиакриламидный гель с 46% мочевины и 1х ТБЭ готовили путем смешивания 38 мл акриламидного раствора и 28 г мочевины. Полимеризацию инициировали добавлением 400 мкл 10%-ного аммония персульфата и 60 мкл ТЭМЭД. Гели формировали с использованием стеклянных пластин и гребенки и разгоняли в 1х ТБЭ-буфере при 60-100 W в течение 2-4 часов (в зависимости от области прочтения). Гели переносили на бумагу Watman для блоттирования, сушили при 80°С в течение почти 1 часа и экспонировали на рентгеновскую пленку при комнатной температуре. Время обычного экспонирования составляло 12 часов. В этих операциях использовались нижеследующие растворы: 5х ренатурационный буфер (200 мМ Трис-HCl, рН 7,5, 100 мМ MgCl2, 250 мМ NaCl); смесь для мечения (7,5 мкМ каждого из дЦТФ, дГТФ и дТТФ); смесь для терминации (80 мкМ каждого из дНТФ, 50 мМ NaCl, 8 мкМ дНТФ (каждого); стоп-раствор (95% формамид, 20 мМ ЭДТА, 0,05% бромфенолового синего, 0,05% ксиленцианола); 5х ТБЭ (0,9 М Трис-борат, 20 мМ ЭДТА); Полиакриламидный раствор (96,7 г полиакриламида, 3,3 г бисакриламида, 200 мл 1х ТБЭ, 957 мл dH2O).
Выделение РНК
Цитоплазматическую РНК выделяли из клеток 293Т, трансфицированных с помощью кальций-фосфата, через 36 часов после трансфекции и из Hela-клеток, зараженных оспой, через 16 часов после заражения, практически так, как описано у Gilman. (Gilman Preparation of cytoplasmic RNA from tissue culture cells. In Current Protocols in Molecular Biology, Ausubel et al., eds., Wiley & Sons, New York, 1992). Вкратце, клетки лизировали в 400 мкл лизирующего буфера, ядра центрифугировали, a SDS и протеиназу К добавляли соответственно в концентрации 0,2% и 0,2 мг/мл. Полученные цитоплазматические экстракты инкубировали при 37°С в течение 20 мин, дважды экстрагировали фенол/хлороформом и осаждали. Полученную РНК растворяли в 100 мкл буфера I и инкубировали при 37°С в течение 20 мин. Данную реакцию останавливали добавлением 25 мкл стоп-буфера и вновь осаждали.
В этой операции использовались нижеследующие растворы: Буфер для лизиса (TRUSTEE, содержащий 50 мМ Триса рН 8,0, 100 мМ NaCI, 5 мМ MgCl2, 0,5% NP40); Буфер I (TRUSTEE-буфер с 10 мМ MgCl2, 1 мМ ДТТ, 0,05 Е/мкл РНКазного ингибитора из плаценты, 0,1 Eg/мкл РНКазы, свободной от ДНКазы I); Стоп-буфер (50 мМ ЭДТА 1,5 М NaOAc 1,0% ДДС).
Слот-блот-анализ
Для слот-блот-анализа растворяли 10 мкг цитоплазматической РНК в 50 мкл dH2O, к которой приливали 150 мкл SSC/18% формальдегида. Эту солюбилизированную РНК инкубировали затем при 65°С в течение 15 мин и наносили пятнами в слот-блот-аппарат. Для гибридизации использовались радиоактивно меченые пробы 1,5 т.п.н. фрагменты gp120IIIb и syngp120mn. Каждый из этих двух фрагментов случайно метили в 50 мкл реакции с 10 мкл 5х буфера для мечения олигонуклеотидов, 8 мкл 2,5 мг/мл BSA, 4 мкл [α32P] - дЦТФ (20 мкКи/мкл; 6000 Ки/ммоль) и 5 Eg фрагмента Кленова. После 1-3 часов инкубации при 37°С добавляли 100 мкл TRUSTEE, а неинкорпорированный [α32]-дЦТФ удаляли с использованием spin-колонки G50. Активность измеряли на бета-счетчике Beckman и одинаковые специфические активности использовали для гибридизации. Мембраны прегибридизовали в течение 2-х часов и гибридизовали в течение 12-24 часов при 42°С с пробой 0,5×106 на мл гибридизующей жидкости. Полученную мембрану дважды промывали (5 мин) промывочным буфером I при комнатной температуре, в течение одного часа в промывочном буфере II при 65°С и затем экспонировали на рентгеновскую пленку. Аналогичные результаты были получены с использованием 1,1 т.п.н. фрагмента Not1/Sfi1 из pCDM7, содержащего 3 нетранслируемые области. Контрольные гибридизации осуществляли одновременно со случайно меченой пробой человеческого бета-актина. Экспрессию РНК оценивали количественно путем сканирования гибридизованных нитроцеллюлозных мембран с помощью фосфоиндикатора Magnetic Dynamics.
В этой операции использовались нижеследующие растворы: 5х буфер для меченья олигонуклеотидов (250 мМ Трис-HC1, рН 8,0, 25 мМ MgCl2, 5 мМ β-меркаптоэтанола, 2 мМ dATP, 2 мМ dGTP, мМ dTTP, 1 M Hepes, рН 6,6, 1 мг/мл гексануклеотидов [dNTP]6); гибридизационный раствор (0,05 M натрийфосфат, 250 мМ NaCl, 7% SDS, 1 мМ ЭДТА, 5% декстран-сульфат, 50% формамид, 100 мкг/мл денатурированной ДНК спермы лосося); промывочный буфер I (2x SSC, 0,1% SDS); промывочный буфер II (0,5х SSC, 0,1% SDS); 20х SSC (3 М NaCl, 0,3 М Nа3-цитрат, рН, доведенный до 7,0).
Рекомбинация коровьей оспы
Использовали модификацию рекомбинации коровьей оспы по методу, описанному Romeo и Seed (Romeo and Seed, Cell, 64:1037, 1991). Вкратце, CVl-клетки, на 70-90% слитые в монослой, заражали 1-3 мкл штаммом коровьей оспы дикого типа WR (2x 108 б. о. е./мл) в течение 1 часа в культуральной среде, не содержащей сыворотки теленка. Через 24 часа эти клетки трансфицировали кальций-фосфатом с 25 мкг TKG плазмидной ДНК на чашку. После дополнительных 24-48 часов эти клетки соскребали с данной чашки, осаждали центрифугированием и ресуспендировали в объеме 1 мл. После 3-х циклов замораживания/оттаивания добавляли трипсин до 0,05 мг/мл и лизаты инкубировали в течение 20 мин. Для заражения в небольших чашках (6 см) использовали серию разведений 10, 1 и 0,1 мкл лизата CV1-клеток, которые предварительно обрабатывали с помощью 12,5 мкг/мл микофеноловой кислоты, 0,25 мг/мл ксантина и 1,36 мг/мл гипоксантина в течение 6 часов. Зараженные клетки культивировали в течение 2-3 дней и затем окрашивали с помощью моноклонального антитела NEA9301 к gp120, а щелочную фосфатазу конъюгировали со вторичным антителом. Клетки инкубировали с 0,33 мг/мл NBT и 0,16 мг/мл BCIP в АР-буфере и, наконец, насыщали 1%-ной агарозой в PBS. Отбирали позитивные бляшки и ресуспендировали в 100 мкл Триса рН 9,0. Полученную бляшку очищали и очистку повторяли. Для получения штаммов с высоким титром инфицирование медленно повышали. Наконец, одну большую чашку Hela-клеток инфицировали половиной вируса из прежнего цикла. Инфицированные клетки разъединяли в 3 мл PBS, лизировали в Daunce-гомогенизаторе и осветляли от большого дебриса путем центрифугирования. Рекомбинантные штаммы VPE-8 вируса коровьей оспы сортировали благодаря созданию AIDS-хранилища, Rockville, MD и экспрессировали HIV-1 IIIB gp120 под 7,5 смешанным ранним/поздним промотором (Earl с соавт., J. Virol., 65:31, 1991). Во всех экспериментах с рекомбинантной коровьей оспой клетки инфицировали при множественности заражения, как минимум, 10.
В этой операции использовались нижеследующие растворы:
АР-буфер (100 мМ Трис-HCl, рН 9,5, 100 мМ NaCl, 5 мМ MgCl2).
Культура клеток
Линии клеток CV1 и Cos7 карциномы почек обезьяны, линия 293 Т карциномы почек человека и линия клеток Hela карциномы шейки матки человека получали из Американской Коллекции Типовых культур и сохраняли в IMDM с добавками. Их содержали в 10 см-овых чашках для тканевых культур и обычно расслаивали 1:5-1:20 каждые 3-4 дня. Для этой операции использовалась нижеследующая среда: IMDM с добавками (90%-ная модифицированная Iscove среда Дюльбекко с добавлением 10% телячьей сыворотки, железа, инактивированной нагреванием в течение 30 мин при 56°С, 0,3 мг/мл L-глутамина, 25 мкг/мл гентамицина, 0,5 мМ β-меркаптоэтанола (рН доводили 5 М NaOH, 0,5 мл).
Трансфекция
Кальций-фосфатную трансфекцию клеток 293Т осуществляли путем медленного добавления и при встряхивании 10 мкг плазмидной ДНК в 250 мкл 0,25 М СаСl2 до того же объема 2х HEBS-буфером во время встряхивания. После инкубации в течение 10-30 мин при комнатной температуре указанный преципитат ДНК вносили в небольшую чашку с клетками, образовавшими монослой на 50-70%. В контрансфекционных экспериментах с rev клетки трансфицировали с помощью 10 мкг gp120IIIb, gp120IIIbrre, syngp120mnrre или rTHY-lenveglrre и 10 мкг pCMVrev или CDM7 плазмидной ДНК.
В этой операции использовались нижеследующие растворы:
2х HEBS-буфер (280 мМ NaCl, 10 мМ КС1, 1,5 мМ стерильно фильтрованные); 0,25 мМ CaCl2 (автоклавированный).
Иммунопреципитация
Через 48-60 часов меняли среду и клетки инкубировали еще 12 часов в Cys/Met-свободной среде, содержащей 200 мкКи 35S-трансметки. Собирали супернатанты и для удаления дебриса центрифугировали их в течение 15 мин при 3000 об/мин. Затем добавляли протеазные ингибиторы лейпептин, апротинин и PMSF в количестве соответственно 2,5 мкг/мл, 50 мкг/мл, 100 мкг/мл, инкубировали 1 мл супернатанта с 10 мкл белок-А сефарозой, упакованной в чистом виде (rTHY-lenveglrre), или с белок-А сефарозой и 3 мкг очищенного слитого белка СD4/иммуноглобулин (созданный у Behring) (все gp120-конструкции) при 4°С в течение 12 часов на роторной качалке. Затем гранулы, конъюгированные с белком А, промывали 5 раз в течение 5-15 мин каждый раз. После окончательной промывки добавляли 10 мкл буфера для образца, образцы кипятили в течение 3-х мин и наносили на 7%-ный (все gp120-конструкции) или 10%-ный (rTHY-lenveg1rre) полиакриламидные гели с SDS (ТРИС-буфер, рН 8,8 для разделяющего геля, ТРИС-буфер, рН 8,6 для концентрирующего геля, ТРИС-глициновый буфер для электрофореза, Maniatis с соавт., см. выше, 1989). Гели фиксировали в 10%-ной уксусной кислоте и 10%-ном метаноле, инкубировали для усиления в течение 20 мин, высушивали и экспонировали в течение 12 часов.
В этой операции использовались нижеследующие буферы и растворы: промывочный буфер (100 мМ Триса, рН 7,5, 150 мМ NaCl, 5 мМ CaCl2, 1% NP-40); буфер для электрофореза (5х) (125 мМ Триса, 1,25 М глицина, 0,5% SDS); буфер для образца (10% глицерина, 4% SDS, 4% β-меркаптоэтанола, 0,02% бромфенолового синего).
Иммунофлуоресценция
Клетки 293Т трансфицировали путем кальций-фосфатной копреципитации и через 3 дня анализировали поверхностную экспрессию THY-1. После отслаивания с помощью ЭДТА/PBS клетки окрашивали моноклональным антителом оХ-7 в разведении 1:250 при 4°С в течение 20 мин, промывали с помощью PBS и затем инкубировали с разведенной 1:500 антисывороткой, конъюгированных с FITC, антител козы к антииммуноглобулину мыши. Клетки вновь промывали, ресуспендировали в 0,5 мл фиксирующего раствора и анализировали в цитофлуориметре EPICS XL (Coulter).
В этой операции использовались нижеследующие растворы: PBS (137 мМ NaCl, 2,7 мМ КС1, 4,3 мМ Na2HPO4, 1/4 мМ КН2РО4, рН, доведенный до 7,4); фиксирующий раствор (2% формальдегид в PBS).
ELISA
Концентрацию gp120 в культуральных супернатантах определяли с использованием планшетов ELISA, покрытых CD4 и козьей анти-gp120-антисывороткой в растворимой фазе. Супернатанты клеток 293Т, трансфицированные кальций-фосфатом, собирали через 4 дня, центрифугировали при 3000 об/мин в течение 10 мин для удаления дебриса и инкубировали в течение 12 часов при 4°С на планшетах. После 6 промывок с помощью PBS в течение 2-х часов добавляли 100 мкл козьей анти-gp120-антисыворотки, разведенной 1:200. Планшеты вновь промывали и инкубировали в течение 2-х часов с разведенной 1:1000 антисывороткой, конъюгированных с пероксидазой кроличьих антител к анти-IgG козы. Затем эти планшеты промывали и инкубировали в течение 30 мин со 100 мкл раствора субстрата, содержащего 2 мг/мл о-фенилендиамина в натрийцитратном буфере. Под конец данную реакцию останавливали с помощью 100 мкл 4 М серной кислоты.
Планшеты прочитывали при 490 нм с помощью спектрофотометра Coulter для микропланшетов. Очищенный рекомбинантный gp120IIb использовали в качестве контроля. В этой методике использовались нижеследующие буферы и растворы: промывочный буфер (0,1% NP40 в PBS); субстратный раствор (2 мг/мл о-фенилендиамина в натрийцитратном буфере).
ПРИМЕР 2
Синтетический ген зеленого флуоресцентного белка
Эффективность кодонового замещения по gp120 дает основание полагать, что замена непредпочтительных кодонов менее предпочтительными кодонами или предпочтительными кодонами (и замена менее предпочтительных кодонов предпочтительными кодонами) будет повышать экспрессию в клетках млекопитающих других белков, напр., других эукариотических белков.
Зеленый флуоресцентный белок (GFP) из медузы Aequorea victoria (Ward, Photochem. Photobiol. 4; 1, 1979; Prasher с соавт.. Gene 111:229, 1992; Cody с соавт., Biochem. 32:1212, 1993) привлек к себе внимание недавно за возможность его использования в качестве маркера или репортера для трансфекции и изучения рода (Chalfie с соавт.. Science 263:802, 1994).
Анализ таблицы кодонового использования, сконструированной из нативной кодирующей последовательности GFP, показывает, что GFP-кодоны предпочитают А или U в третьей позиции. Смещение в данном случае благоприятствует А меньше, чем смещению gp120, но является существенным. Был создан синтетический ген, в котором естественная GFP-последовательность была воссоздана биотехнологически во многом тем же способом, что и для gp120 (фиг.11; SEQ ID NO:40). Кроме того, инициацию трансляции последовательности GFP заменили последовательностями, корреспондирующими с инициации трансляции консенсуса. Полученная экспрессия результирующего белка была противоположна той, которая присуща последовательности дикого типа, подобно биотехнологической, чтобы нести оптимизированный консенсус инициации трансляции (фиг.10В и фиг.10С). Кроме того, изучали (фиг.10D) эффект включения мутации Ser 65→Thr, которая, как сообщалось, улучшает эффективность возбуждения GFP при 490 нм и поэтому предпочтительна для флуоресцентной микроскопии (Heim с соавт.. Nature 373:663, 1995). Биотехнологический кодон существенно повышает эффективность экспрессии (совместная процентная доля клеток очевидно позитивна для трансфекции), а сочетание мутации Ser 65→Thr и кодоновой оптимизации приводит к кодированию в сегменте ДНК превосходно видимого маркерного белка млекопитающих (ФИГ.10D).
Вышеописанную последовательность, кодирующую синтетический зеленый флуоресцентный белок, создавали аналогичным способом, что и gp120, из шести фрагментов, приблизительно, из 120 п.н. каждый, с использованием метода сборки, который зависит от способности ферментов рестрикции BsaI и BbsI расщеплять, за исключением их узнающей последовательности. Синтезировали длинные олигонуклеотиды, которые содержат части кодирующей последовательности GFP, вставленной во фланкирующие последовательности, кодирующие EcoRI и BsaI на одном конце и BamHI и BbsI - на другом конце. Таким образом, каждый олигонуклеотид обладает конфигурацией EcoRI/BsaI/GFP-фрагмент/BbsI/BamHI. Концы сайта рестрикции, образованные с помощью сайтов BsaI и BbsI, предназначались для получения сочетаемых концов, которые могли бы использоваться для соединения смежных GFP-фрагментов. Каждый из этих сочетаемых концов проектировался в качестве уникального и не самокомплементарного. Примерные синтетические фрагменты ДНК амплифицировали путем PCR, инсерцировали между EcoRI и BamHI в pUC9 и секвенировали. Затем интактную кодирующую последовательность собирали для лигирования шести фрагментов с использованием инсерцируемых фрагментов, полученных с помощью BsaI и BbsI. Две из шести плазмид, полученных в результате лигирования, имели отношение к вставке правильного размера, а одна содержала требуемую последовательность полной длины. Мутацию Ser 65→Thr осуществляли с помощью мутагенеза на основе PCR с использованием праймера, который перекрывает уникальный BssSI-сайт в данном синтетическом GFP.
Кодоновая оптимизация как метод улучшенной экспрессии в клетках млекопитающих
Представленные здесь данные дают основание полагать, что кодирующая биотехнологическая последовательность, созданная повторно, может обладать общей применяемостью для улучшения экспрессии эукариотических генов, млекопитающих и не принадлежащих млекопитающим, в клетках млекопитающих. Результаты, полученные здесь для трех не связанных родством белков: HIV gp120, поверхностный антиген Thy-1 крысиных клеток и зеленый флуоресцентный белок из Aequorea victoria и человеческий Фактор VIII (см. ниже) дают основание полагать, что кодоновая оптимизация может оказаться плодотворным подходом для улучшения экспрессии в клетках млекопитающих целого ряда эукариотических генов.
ПРИМЕР III
Создание кодон-оптимизированной экспрессии гена человеческого Фактора VIII, не имеющего центрального В-домена
Был создан синтетический ген, который кодирует зрелый человеческий Фактор VIII, охватывающий (остатки 779-1658, в том числе предшественника), не имеющий аминокислотных остатков 760-1639. Данный синтетический ген создавали путем выбора кодонов, соответствующих кодонам, которые благоприятствуют эффективно экспрессируемым человеческим генам. Некоторое отклонение от точного присоединения к предпочтительному паттерну остатка было сделано, чтобы позволить сайгам расщепления уникальным ферментом рестрикции быть интродуцированным по всей длине данного гена, чтобы облегчить будущие манипуляции. Для получения данного синтетического гена данную последовательность разделяли затем на 28 фрагментов по 150 пар оснований и на 29-й сегмент из 161 пары оснований.
Синтетический ген, экспрессирующий человеческий Фактор VIII, не имеющий центрального В-домена, конструировали следующим образом.
Синтезировали матричные олигонуклеотиды из двадцати девяти пар (см. ниже). 5'-матричные олигонуклеотиды имели в длину 105 оснований, а 3'-матричные олигонуклеотиды имели в длину 104 основания (за исключением последнего 3'-олигонуклеотида, который имел в длину 125 остатков). Матричные олигонуклеотиды создавали таким образом, чтобы каждая отжигаемая пара, состояла из одного 5'-олигонуклеотида и одного 3'-олигонуклеотида, создавая 19 парнуклеотидные двухцепочечные области.
Для того чтобы содействовать PCR и последующим операциям, 5'-концы данных олигонуклеотидных пар проектировали инвариантными в пределах первых 18 остатков, с учетом характерной пары PCR-праймеров, используемых для амплификации, и с учетом PCR-условий, используемых для всех пар. Первые 18 остатков каждого 5'-члена данной матричной пары представляют собой сgс gaa ttc gga аgа ссс (SEQ ID NO:110), а первые 18 остатков каждого 3'-члена данной матричной пары представляют собой ggg gat cct cac gtc tca (SEQ ID NO:43).
Пары олигонуклеотидов отжигали, а затем удлиняли и амплифицировали с помощью PCR в реакционной смеси следующим образом: матрицы, каждую, отжигали в 200 мкг/мл PCR-буфера (10 мМ Трис-HCl, 1,5 мМ MgCl2, 50 мМ КСl, 100 мкг/мл желатина, рН 8.3). PCR-реакции содержали 2 нг отожженных матричных олигонуклеотидов, 0,15 мкг каждого из двух 18-мерных праймеров (описаны ниже), 200 мкМ каждого из дезоксинуклеозидтрифосфатов, 10%, по объему, DMSO и PCR-буфер, поставляемый Boehringer Mannheim Biochemicals, в конечном объеме 50 мкл. После добавления Тaq-полимеразы (2,5 единицы, 0,5 мкл; Boehringer Mannheim Biochemicals) амплификацию осуществляли на амплификаторе Perkin-Elmer в течение 25 циклов (94°С в течение 30 с, 55°С в течение 30 с, и 72°С в течение 30 с). Заключительный цикл осуществляли с последующим 10-минутным удлинением при 72°С.
Полученные амплифицированные фрагменты обрабатывали с помощью EcoRI и BamHI (расщепление фрагментов по 5'- и 3'-концам) и лигировали с pUC9-производным, разрезанным с помощью EcoRI и BamHI.
Индивидуальные клоны секвенировали и идентифицировали коллекцию плазмид, корреспондирующих с полной требуемой последовательностью. Затем клоны объединяли путем мультифрагментного лигирования, учитывая преимущество сайтов рестрикции на 3'-концах PCR-праймеров, непосредственно примыкающих к данной амплифицированной последовательности. 5'-PCR-праймеp, содержащий BbsI-сайт, и 3'-PCR-праймер, содержащий BsmBI-сайт, располагали таким образом, чтобы расщепление соответствующими ферментами происходило до первого нуклеотида амплифицируемой части и левыми 4-м основанием 5'-концевого выступа, создаваемого первыми 4-мя основаниями данной амплифицируемой части. Одновременная обработка с помощью BbsI и BsmBI высвобождает таким образом данную амплифицированную часть с уникальным 4-м основанием 5'-концевого выступа на каждом конце, которая не содержит никакой праймерной последовательности. Обычно эти выступы не были самокомплементарны, учитывая мультифрагментные лигирующие реакции, которые образуют требуемый продукт с высокой эффективностью. Уникальная часть из первых 28 амплифицированных олигонуклеотидных пар была при этом 154 п.н. и после обработки, каждая, приводила к возникновению 150 п.н. фрагмента с уникальными концами. Первый и последний фрагменты не изменялись данным способом, однако, поскольку они обладали иными сайтами рестрикции, созданными в них для облегчения инсерции собранной последовательности в надлежащий экспрессионный вектор млекопитающего.
Практически процесс сборки протекал следующим образом.
Сборка синтетического гена Фактор VIII
Стадия 1: 29 фрагментов собраны с образованием 10 фрагментов.
29 пар олигонуклеотидов, которые образовали сегменты 1-29 посредством спаривания оснований, описаны ниже.
Плазмиды, несущие сегменты 1, 5, 9, 12, 16, 20, 24 и 27 были обработаны с помощью EcoRI и BsmBI и были выделены 170 п.н. фрагменты; плазмиды, несущие сегменты 2, 3, 6, 7, 10, 13, 17, 18, 21, 25 и 28 были обработаны с помощью BbsI и BsmBI и были выделены 170 п.н. фрагменты; и плазмиды, несущие сегменты 4, 8, 11, 14, 19, 22, 26 и 29 были обработаны с помощью EcoRI и BbsI и был выделен 2440 п.н. векторный фрагмент. Фрагменты, несущие сегменты 5, 6, 7 и 8, были лигированы с образованием сегмента "В"; фрагменты, несущие сегменты 9, 10 и 11, были лигированы с образованием сегмента "С"; фрагменты, несущие сегменты 12, 13 и 14, были лигированы с образованием сегмента "D"; фрагменты, несущие сегменты 16, 17, 18 и 19, были лигированы с образованием сегмента "F"; фрагменты, несущие сегменты 20, 21 и 22, были лигированы образованием сегмента "G"; фрагменты, несущие сегменты 24, 25 и 26, были лигированы с образованием сегмента "I"; и фрагменты, несущие сегменты 27, 28 и 29, были лигированы с образованием сегмента "J".
Стадия 2: Сборка 10 полученных фрагментов на стадии 1 в три фрагмента
Плазмиды, несущие сегменты "А", "D" и "G" были обработаны с помощью EcoRI и BsmBI, плазмиды, несущие сегменты В, 15, 23 и I, были обработаны с помощью Bbsl и BsmBI, a плазмиды, несущие сегменты С, F и J, были обработаны с помощью EcoRI и Bbsl. Фрагменты, несущие сегменты А, В и С, были лигированы с образованием сегмента "К"; фрагменты, несущие сегменты D, 15 и F, были лигированы с образованием сегмента "О"; и фрагменты, несущие сегменты G, 23, I и J, были лигированы с образованием сегмента "Р".
Стадия 3: Сборка трех окончательных кусков
Плазмиду, несущую сегмент К, обрабатывали с помощью EcoRI и BsmBI, плазмиду, несущую сегмент О, обрабатывали с помощью BbsI и BsraBI, и плазмиду, несущую сегмент Р, обрабатывали с помощью EcoRI и BbsI. Полученные три фрагмента лигировали с образованием сегментов.
Стадия 4: Инсерцирование полученного синтетического гена в экспрессионный вектор млекопитающего
Плазмиду, несущую сегмент S, обрабатывали с помощью NheI и NotI и инсерцировали между свитами NheI и NotI плазмиды CD51Neg1 с образованием плазмиды cd51sf8b-.
Секвенирование и коррекция синтетического гена Фактора VIII
После сборки указанного синтетического гена обнаружено, что в нем имелись два нежелательных остатка, закодированных в данной последовательности. Один из них представлял собой Arg-остаток на 749 позиции, который представлен в GenBank-последовательности, исходно введенный из Genen-tech, но не в последовательности, приведенной Genentech в литературе. Другой представлял собой Ala-остаток на 146 позиции, которая должна была быть Pro. Данная мутация на неидентифицированной стадии возникла после секвенирования 29 составных фрагментов. Мутацию Pro749Arg откорректировали путем инкорпорирования требуемого изменения в PCR-праймере (ctg ctt ctg acg cgt gct ggg gtg gcg gga gtt; SEQ ID NO:44), которое включает Mlul-сайт в позиции 2335 в нижеприведенной последовательности (последовательность сегмента от Hindi II до NotI) и амплификации между этим праймером и праймером (ctg ctg aaa gtc tcc agc tgc; SEQ ID NO:44) от 5' до сайта SgrAI в позиции 2225. Затем фрагмент SgrAI-MluI инсерцировали в экспрессионный вектор в родственные сайты данного вектора и полученное исправленное изменение последовательности проверили путем секвенирования.
Мутация Рго146А1а была исправлена путем инкорпорирования требуемого изменения последовательности в олигонуклеотид (ggс аgg tgc tta аgg аgа асg gсс cta tgg cca; SEQ ID NO:46), несущей сайт AflII по остатку 504, и амплификации фрагмента, полученного из PCR-реакции между этим олигонуклеотидом и праймером, обладающим последовательностью ctg tgt tct tca tac gcg tct ggg gct cct cgg ggc (SEQ ID NO:109), разрезанием полученного PCR-фрагмеыта с помощью AflII и AvrII по остатку 989, инсерцированием исправленного фрагмента в экспрессионный вектор и подтверждением данной конструкции путем секвенирования.
Конструирование эквивалентного нативного гена, экспрессирующего человеческий Фактор VIII, не имеющего центрального В-домена
Экспрессионную плазмиду с делегированным В-доменом эквивалентного Фактора VIII, обладающего нативной кодоновой последовательностью, конструировали путем интродуцирования NheI на 5'-конце зрелой кодирующей последовательности с использованием праймера сgс саа ggg cta gсс gсс асc аgа аgа tac tac ctg ggt (SEQ ID NO:47), амплификации между этим праймером и праймером att cgt agt tgg ggt tcc tct gga xag (соответствующий остаткам 1067-1093 последовательности, показанной ниже), разрезания с помощью NheI и AflII (остаток 245 в нижеприведенной последовательности) и инсерцирования данного полученного фрагмента в надлежащую расщепленную плазмиду, несущую нативный Фактор VIII. Делецию В-домена осуществляли с помощью метода перекрывающейся PCR с использованием ctg tat ttg atg aga асе g (соответствующие остаткам 1813-1831, ниже) и саа gас tgg tgg tgg ggt ggc att aaa ttg ctt t (SEQ ID NO:48) (комплементарные ниже остаткам 2342-2372) по 5'-концевому перекрыванию, и aat gcc асc сса сса gtc ttg aaa cgc ca (SEQ ID NO:49) (2352-2380 в последовательности ниже) и cat ctg gat att gca ggg ag (SEQ ID NO:50) (3145-3164). Затем полученные продукты из двух индивидуальных PCR-реакций смешивали и реамплифицировали путем использования наиболее удаленных от центра праймеров, получая фрагмент, расщепляемый по Asp718 (изошизомер Kpnl, 1837 в последовательности ниже) и PflMI (3100 в последовательности ниже) и инсерцировали соответствующую расщепленную экспрессионную плазмиду, несущую нативный Фактор VIII.
Полная последовательность (SEQ ID NO:41) нативного гена человеческого фактора VIII, делегированного по центральной В-области, представлена на фиг.12. Полная последовательность (SEQ ID NO:42) синтетического гена Фактора VIII, делегированного по центральной В-области, представлена на фиг.13.
Получение и анализ экспрессионных плазмид
Два независимых плазмидных изолята из природного и четыре независимых изолята из синтетического Фактора VIII экспрессионной плазмиды размножали в бактериях, а их ДНК получали путем центрифугирования в растворе с градиентом плавучей плотности CsCl с последующей фенольной экстракцией.
Анализ супернатантов СОЗ-клеток, трансфицированных данными плазмидами, показал, что синтетический ген дает увеличение Фактора VIII, приблизительно, в четыре раза больше, чем природный ген.
Затем СОS-клетки трансфицировали 5 мкг каждой конструкции фактора VIII в 6 см чашке Петри с использованием DEAE-декстранового метода. Через 72 часа после трансфекции в каждую чашку добавляли 4 мл свежей среды, содержащей 10% телячьей сыворотки, 12-ю часами позже из каждой чашки брали образец среды. Образцы тестировали с помощью ELISA с использованием мышиного моноклонального антитела к антителам легкой цепи человеческого фактора VIII и конъюгированных с пероксидазой поликлональных козьих антител к антителам человеческого фактора VIII. Выделенный очисткой человеческий плазменный фактор VIII использовали в качестве стандарта. Клетки, трансфицированные с помощью синтетической генной конструкции Фактора VIII, экспрессировали 138±20,2 нг/мл (эквивалентных нг/мл неделетированного Фактора VIII) Фактора VIII (n=4), а клетки, тарсфицированные с помощью природного гена Фактора VIII, экспрессировали 33,5±0,7 нг/мл (эквивалетные нг/мл неделетированнного Фактора VIII) Фактора VIII (n=2).
Для конструирования синтетического гена Фактора VIII использовались нижеследующие олигонуклеотидные матрицы.















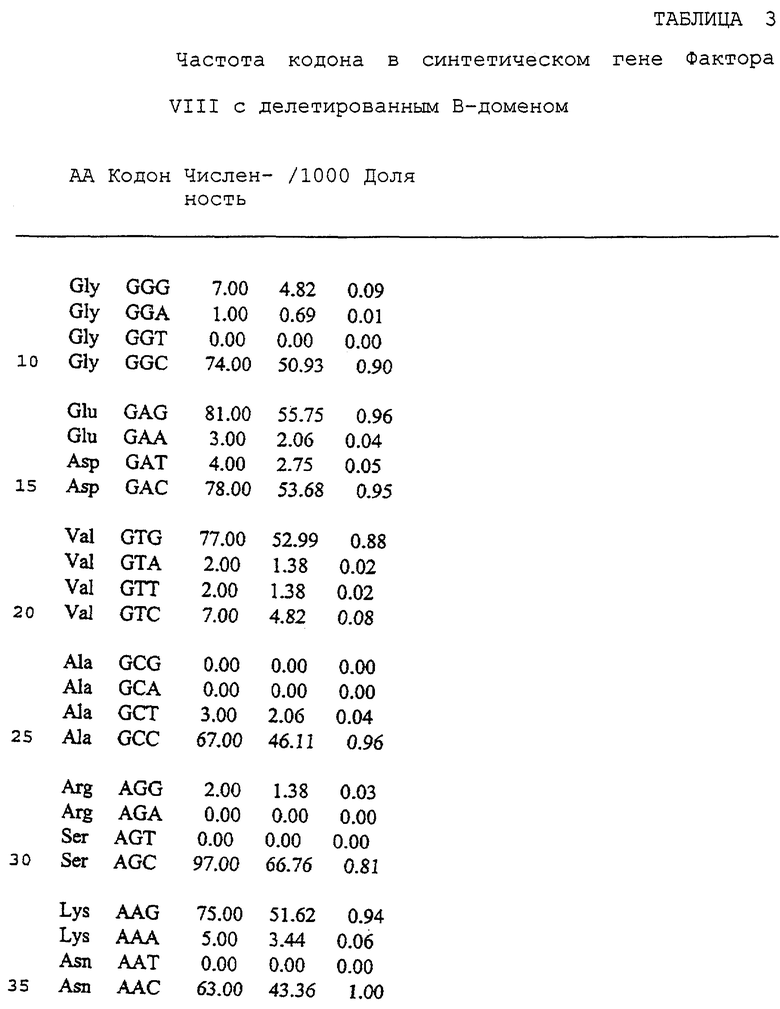
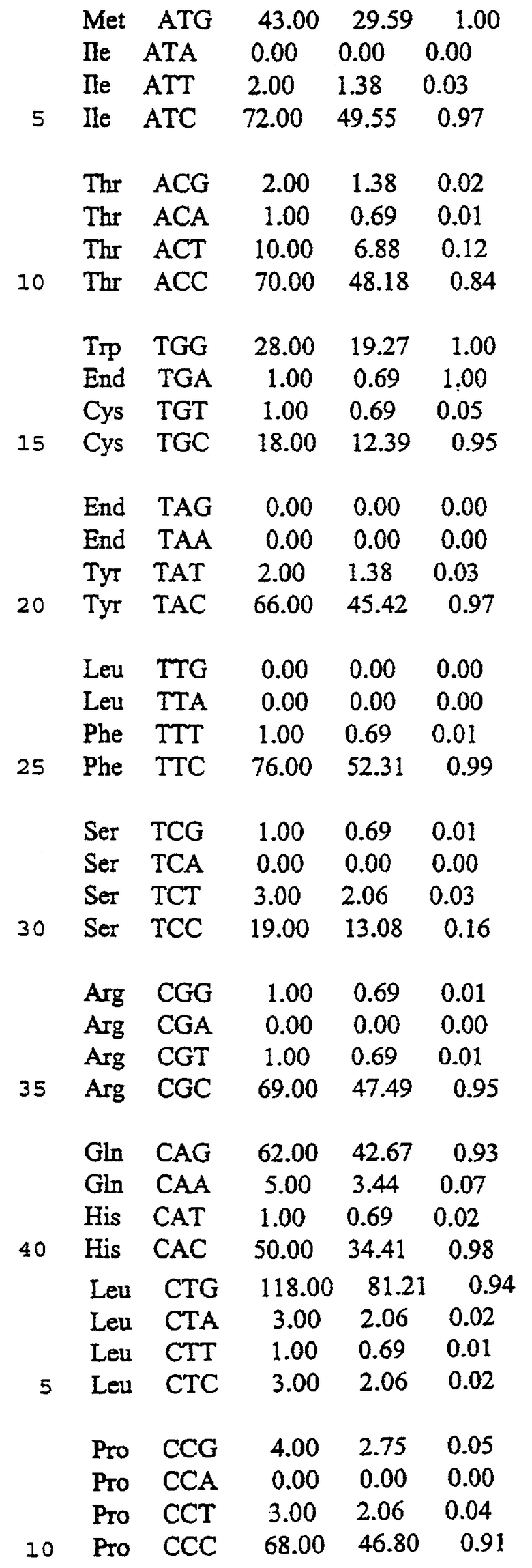
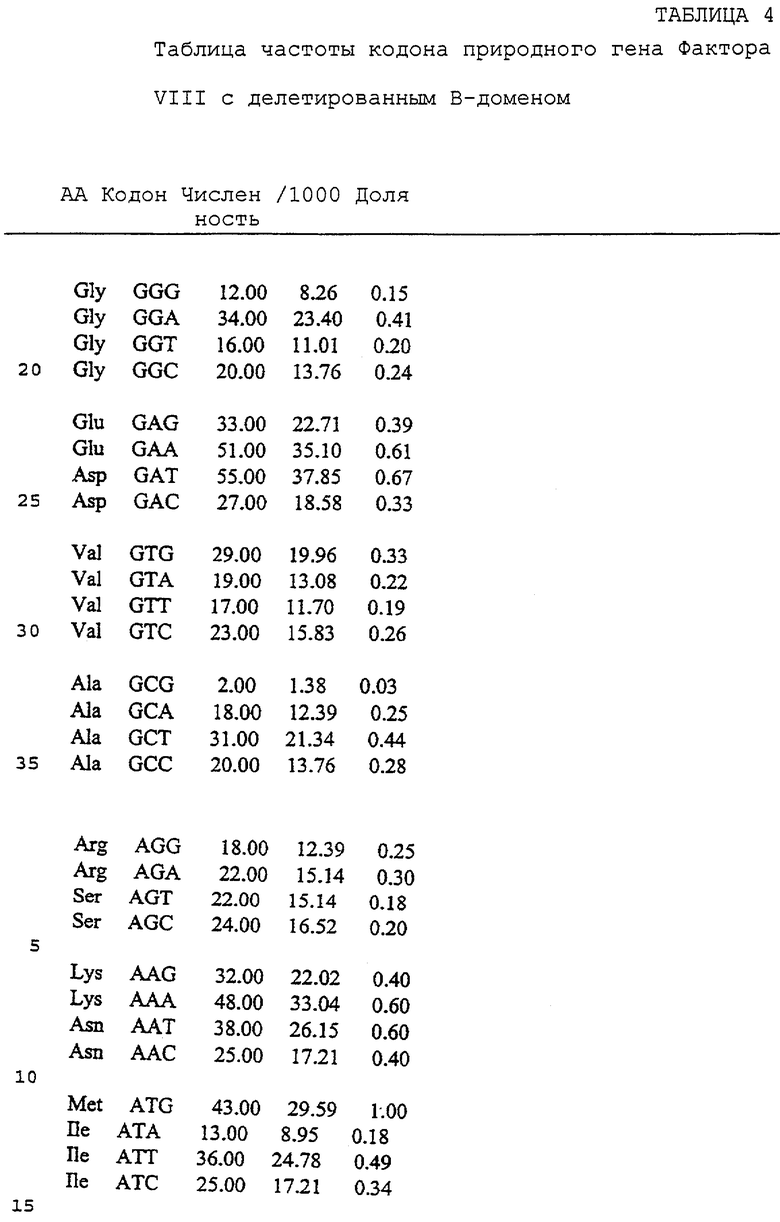
Использование
Синтетические гены настоящего изобретения пригодны для экспрессии белка, обычно экспрессируемого в клетках млекопитающих в клеточной культуре (например, для промышленного получения человеческих белков, таких как hGH, ТРА, Фактор VIII и Фактор IХ). Синтетические гены настоящего изобретения пригодны также для генной терапии. Например, синтетический ген, кодирующий выбранный белок, может быть интродуцирован в клетку, которая может экспрессировать белок для создания клетки, которая может быть введена пациенту, нуждающемуся в данном белке. Такие методики генной терапии, основанные на клетке, хорошо известны специалистам в данной области техники, смотрите, напр., Anderson с соавт., U.S. Patent No. 5399349; Mulligan and Wilson, U.S. Patent No. 5460959.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения линии гуманизированных мышей, содержащих инсерцию 3974insT в гене mGrin2a (mice glutamate [NMDA] receptor subunit epsilon-1), приводящую к преждевременному прекращению трансляции белка grin2a | 2021 |
|
RU2764650C1 |
| Искусственные гены, кодирующие белки-иммуногены EV.CTL и EV.Th, рекомбинантные плазмидные ДНК pEV.CTL и pEV.Th, обеспечивающие экспрессию искусственных генов, и искусственные Т-клеточные полиэпитопные белки-иммуногены EV.CTL и EV.Th, содержащие эпитопы антигенов вируса Эбола, используемые для создания вакцины против вируса Эбола | 2018 |
|
RU2713723C1 |
| СПОСОБ ПОЛУЧЕНИЯ ХИМИЧЕСКИХ ПРОДУКТОВ ТОНКОГО СИНТЕЗА С ПОМОЩЬЮ CORYNEBACTERIUM, СЕКРЕТИРУЮЩЕЙ МОДИФИЦИРОВАННЫЕ α-1,6-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2763317C2 |
| АНТИТЕЛО ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2781148C2 |
| АНТИТЕЛО К CD73 ЧЕЛОВЕКА | 2017 |
|
RU2754058C2 |
| Кодон-оптимизированная последовательность нуклеотидов, кодирующая hAIPL1, и её содержащий экспрессионный вектор | 2021 |
|
RU2785621C1 |
| АНТИТЕЛО ПРОТИВ С5 И СПОСОБ ПРЕДУПРЕЖДЕНИЯ И ЛЕЧЕНИЯ ОБУСЛОВЛЕННЫХ КОМПЛЕМЕНТОМ ЗАБОЛЕВАНИЙ | 2014 |
|
RU2663349C2 |
| АНТИТЕЛО ПРОТИВ Fn14 ЧЕЛОВЕКА | 2019 |
|
RU2787044C2 |
| Способ получения линии гуманизированных мышей, трансгенных по hACE2 | 2020 |
|
RU2757114C1 |
| Последовательность нуклеотидов, кодирующая β-N-ацетилгексозаминидазу Aggregatibacter actinomycetemcomitans, и генетические конструкции, её содержащие | 2021 |
|
RU2789544C1 |



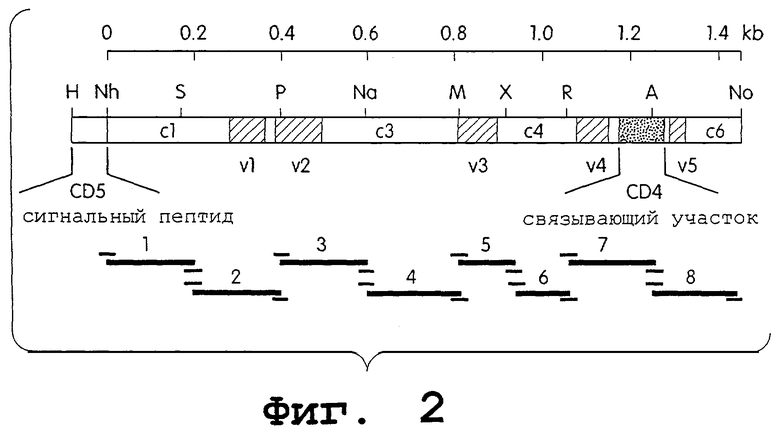
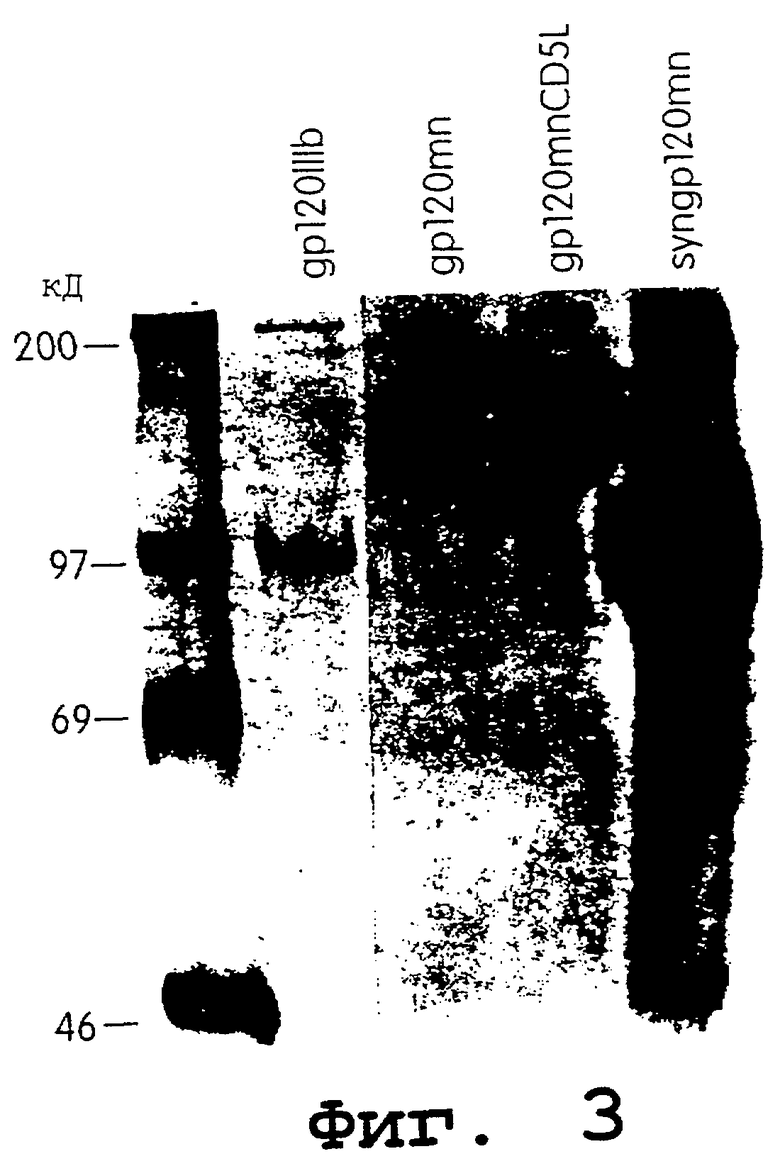
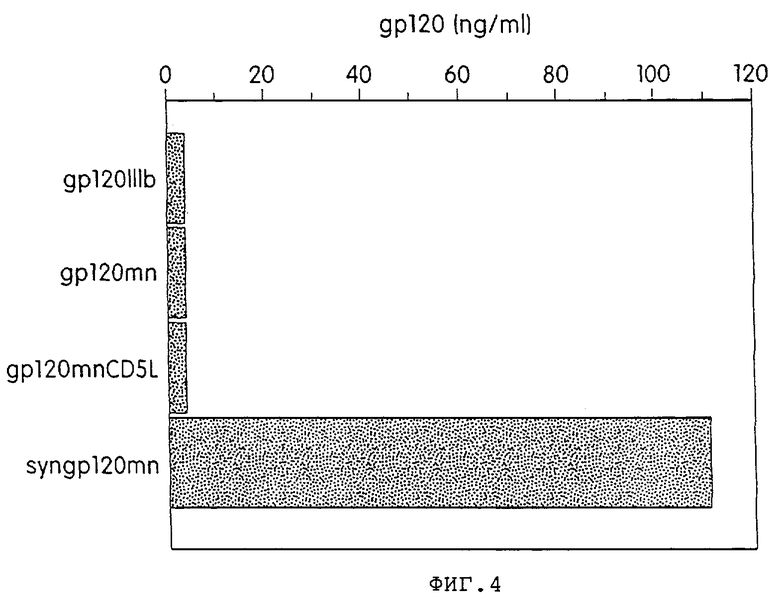
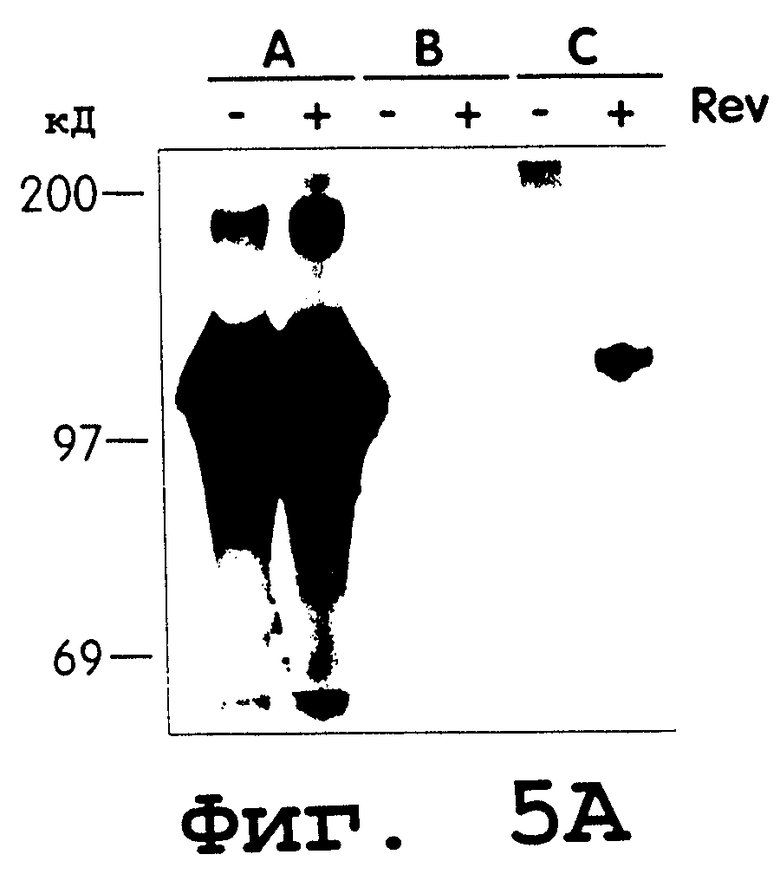
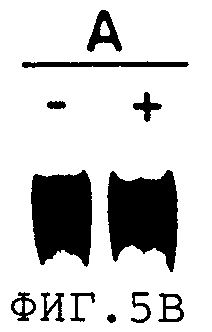
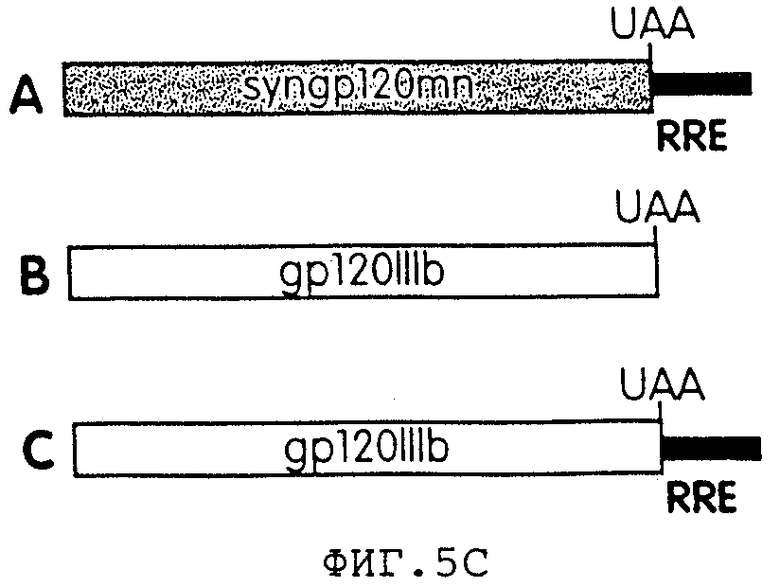

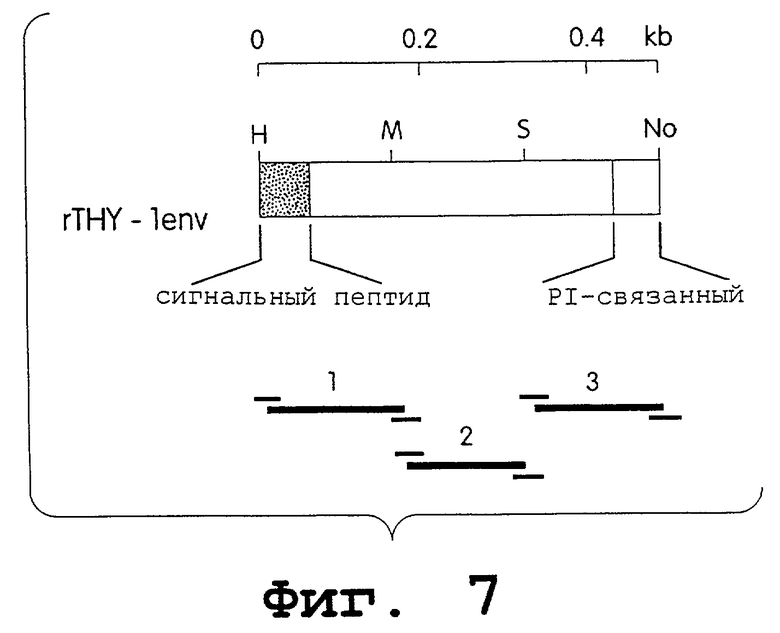
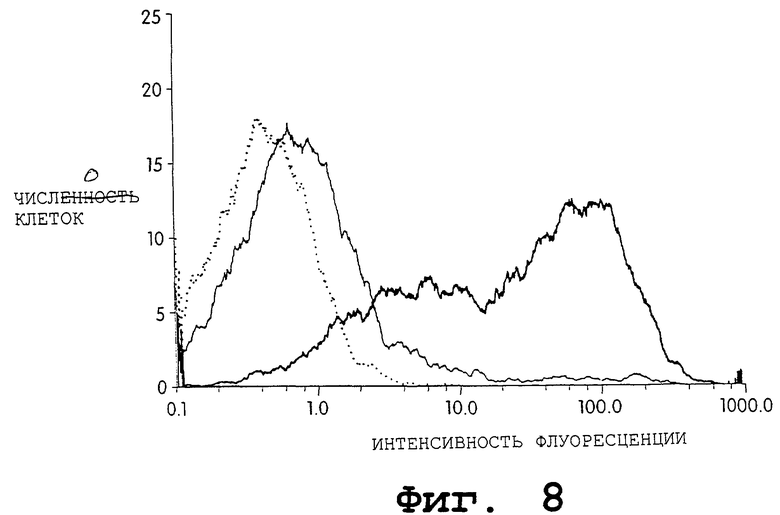

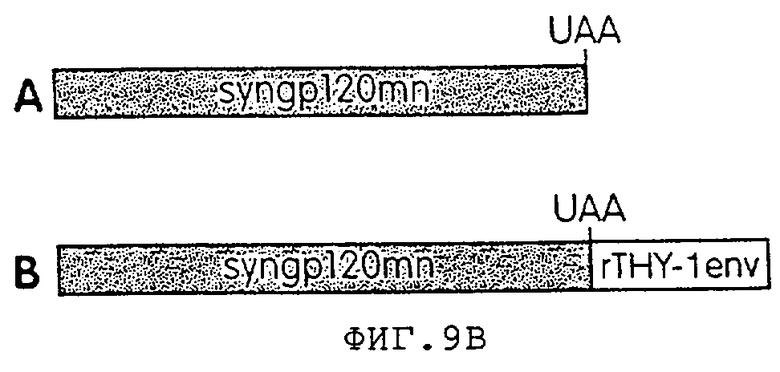
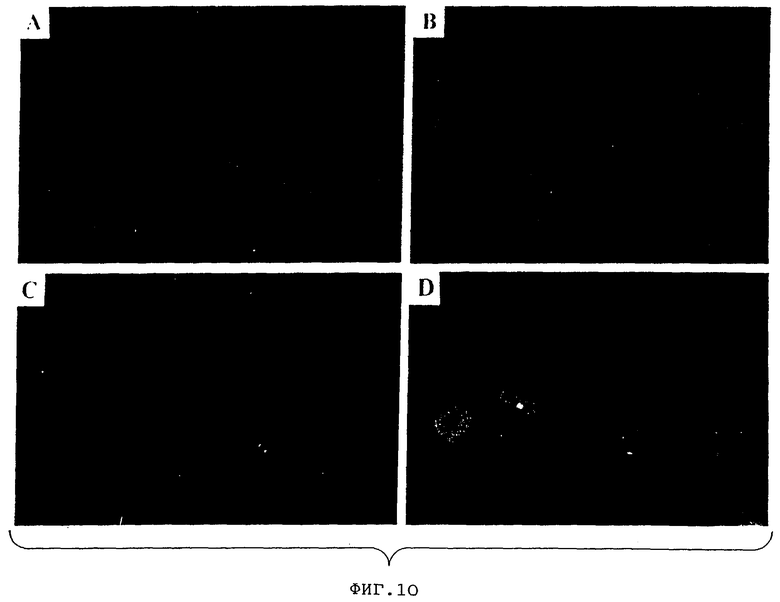

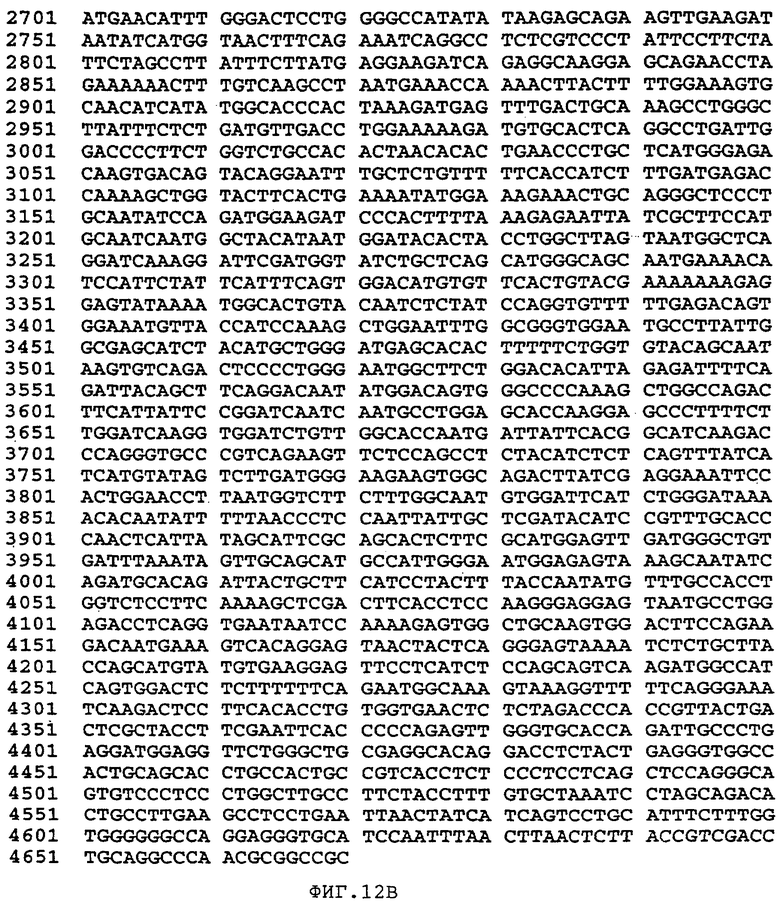
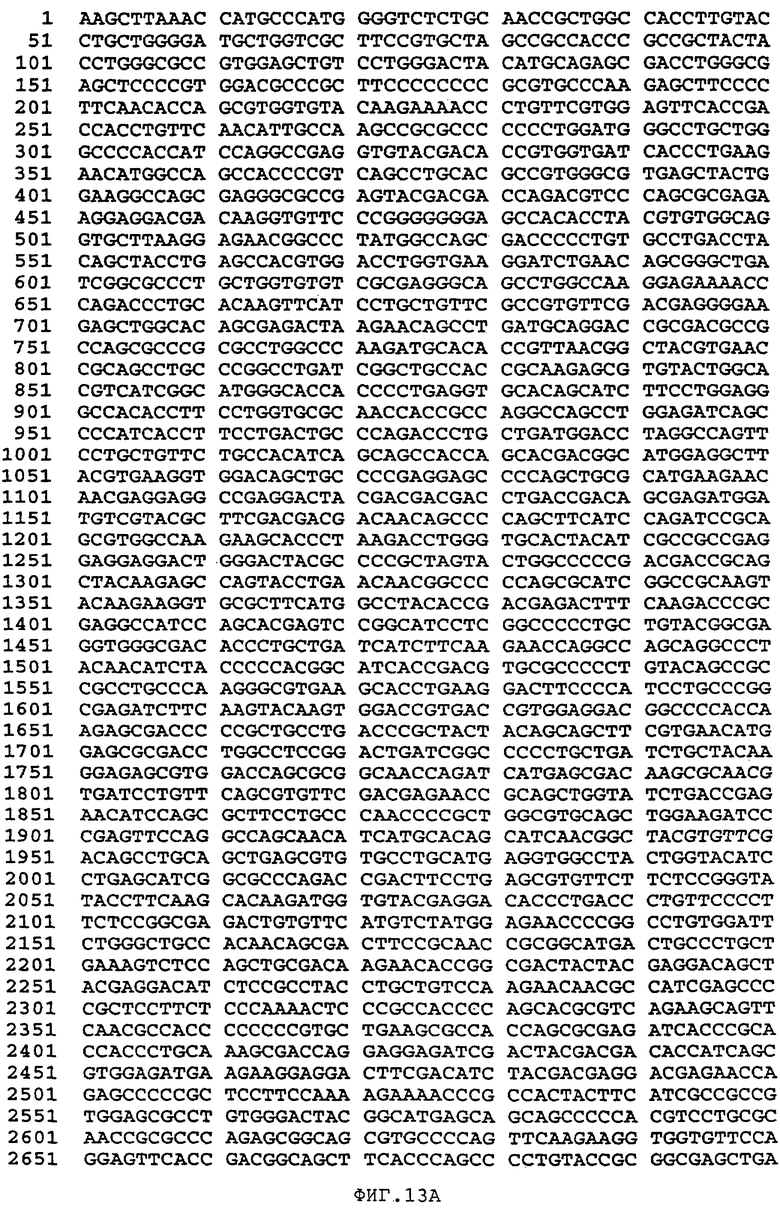
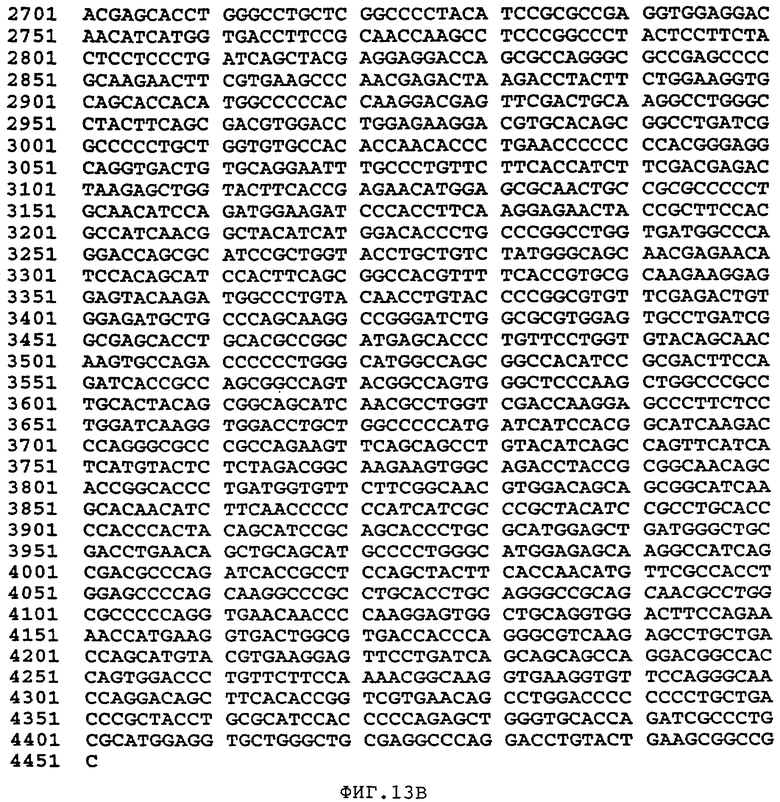
Изобретение относится к генной инженерии. Предложен новый синтетический ген, кодирующий зеленый флуоресцентный белок. Предложенный ген характеризуется заменами непредпочтительных или менее предпочтительных кодонов на предпочтительные кодоны. При этом предпочтительные кодоны представляют собой группу gcc, cgc, aac, gac, tgc, cag, ggc, cac, atc, ctg, aag, ccc, ttc, agc, acc, tac и gtg, а менее предпочтительные представляют собой ggg, att, ctc, tcc, gtc и agg, а непредпочтительные кодоны представляют собой оставшиеся кодоны. Новый синтетический ген характеризуется способностью экспрессировать зеленый флуоресцентный белок на уровне 110-1000% от уровня экспрессии природного гена. Также предложен способ получения синтетического гена зеленого флуоресцентного белка. Способ предусматривает определение непредпочтительных или менее предпочтительных кодонов в природном гене и замену одного или нескольких кодонов на предпочтительные кодоны, кодирующих ту же аминокислоту, что и заменяемые кодоны. Предложен также экспрессирующий вектор, содержащий синтетический ген, и способ получения культуры клеток млекопитающего, несущих синтетический ген, предусматривающий трансформирование клеток экспрессирующим вектором. Предложенное изобретение может быть использовано для промышленного получения необходимых белков, а также для генной терапии. 4 н. и 11 з.п. ф-лы, 4 табл., 21 ил.
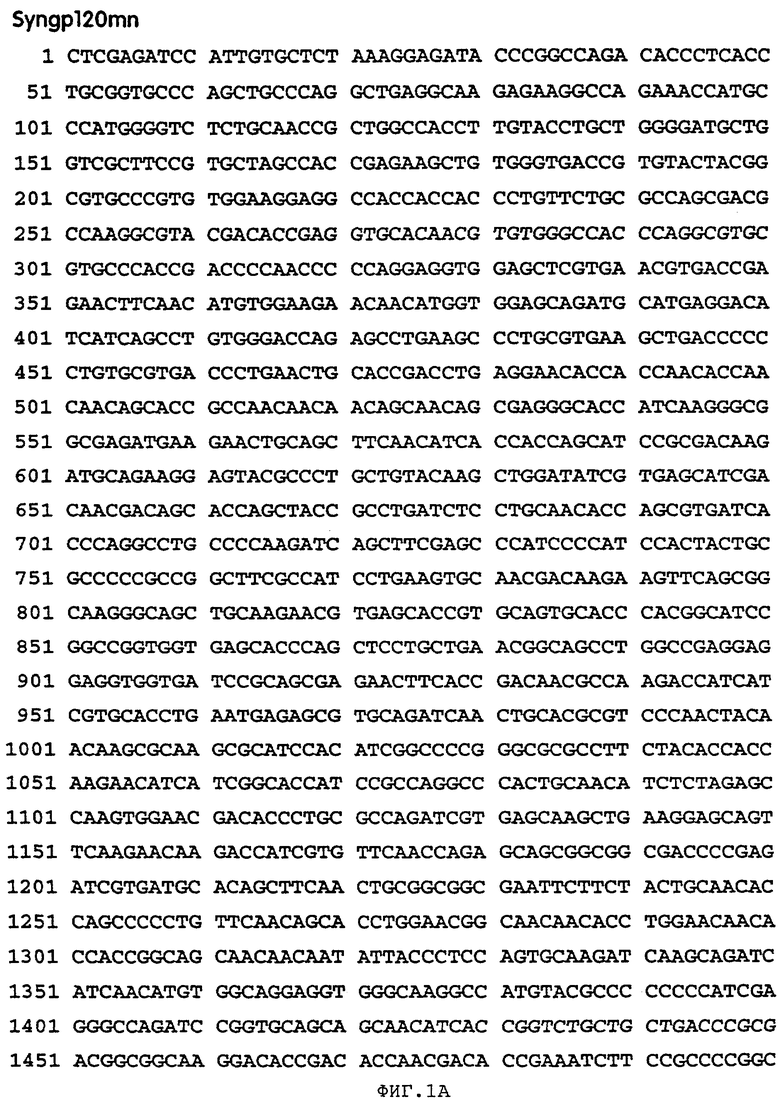
| WO 9609378, 28.03.1996 | |||
| SEETHARAM et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Biochem | |||
| Biophys Res | |||
| Comm | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |

