Область, к которой относится изобретение
Изобретение относится, в основном, к вакцинам, применяемым в ветеринарной практике, и, более конкретно, к живой рекомбинантной аттенуированной вакцине против заболеваний, вызываемых организмами, которые в норме инкапсулированы, причем капсула необходима для вирулентности, но не для иммунопротекции. Изобретение, в частности, относится к генно-инженерной вакцине на основе инфекционных агентов, лишенных капсулы.
Уровень техники
Вакцины представляют собой препараты, которые используются для предотвращения заболеваний животных и человека путем индукции иммунитета. Это достигается введением пациенту антигена, специфичного для конкретного заболевания, который, в свою очередь, заставляет иммунную систему пациента продуцировать большие количества антител. Наличие антител в крови пациента защищает его при дальнейших контактах с инфекционным агентом. Вакцины могут включать субъединицы агента или собственно агент, живой или убитый. Например, полиомиелит, обычно обозначаемый как “полио”, предотвращают введением или живой аттенуированной оральной полиовирусной вакцины (как правило, детям) или же введением убитой или инактивированной полиовирусной вакцины, которой чаще иммунизируют взрослых, поскольку для них выше риск заболеваемости полиомиелитом в результате иммунизации живой вакциной. Если используется живая вакцина, то вирулентность инфекционного агента должна быть каким-либо образом снижена (аттенуирована); в противном случае содержащийся в вакцине вирус будет вызывать заболевание вместо того, чтобы обеспечивать защиту от него.
Ряд заболеваний вызывается инкапсулированными бактериями, при этом капсула, представляющая собой клееподобный слой полисахарида или полипептида снаружи клеточной стенки бактерии, необходима для патогенеза. Плевропневмония свиней является одним из примеров такого рода заболеваний. Она вызывается бактерией Actinobacillus pleuropneumoniae, к факторам вирулентности которой относятся капсулярный полисахарид, эндотоксин и белковые экзотоксины. Плевропневмония свиней - это одно из основных респираторных заболеваний, приносящих значительный ущерб свиноводству во всем мире. Только в США ежегодный ущерб от этого заболевания оценивается в миллионы долларов.
В Патенте США 5429818, упоминаемом в качестве ссылки, Inzana описывает некапсулированные мутанты Actinobacillus pleuropneumoniae, которые невирулентны и способны обеспечивать превосходную защиту от вирулентных бактерий. Некапсулированные мутанты, описанные Inzana, были получены этилметансульфонатным мутагенезом. Недостатками подобных процедур являются нестабильность спонтанных или химически-индуцированных мутантов, а также тот факт, что природа мутаций остается неизвестной.
Краткое описание изобретения
Задача настоящего изобретения заключается в получении безопасной и эффективной, живой, аттенуированной рекомбинантной вакцины против плевропневмонии животных, в частности свиней.
Другой задачей настоящего изобретения является разработка генно-инженерного метода, в результате которого возбудитель плевропневмонии Actinobacillus pleuropneumoniae утрачивает капсулу, становясь при этом авирулентным.
Согласно настоящему изобретению был получен рекомбинантный, живой, аттенуированный штамм Actinobacillus pleuropneumoniae, который в результате генно-инженерных манипуляций утратил капсулу. Поскольку капсула необходима для вирулентности, но не для иммунопротекции, штамм полезен для использования в качестве вакцинного против плевропневмонии свиней. Вакцина была получена с использованием клонированного плазмидного вектора, неспособного реплицироваться в А.pleuropneumoniae. Были секвенированы гены А.pleuropneumoniae серотипа 5, ответственные за синтез и экспорт капсулы. В последовательности данных генов была произведена большая делеция и в сайт делении были клонированы гены устойчивости к канамицину и чувствительности к сахарозе, которые служили маркерными генами. Данный суицидный вектор был введен в вирулентный штамм А.pleuropneumoniae серотипа 5 с помощью электропорации с тем, чтобы получить гомологичную рекомбинацию между гомологичными областями хромосомы и плазмиды. Было получено четыре изолята, каждый из которых был лишен радужности, что свидетельствует об утрате капсулы. Для одного штамма утрата капсулы и делеция части капсулярных генов были подтверждены дот-блоттингом и блоттингом по Саузерну соответственно. Было также подтверждено присутствие маркерных генов в рекомбинантном штамме. Не было идентифицировано каких-либо других изменений фенотипических свойств, и маркерные гены не были обнаружены в других областях хромосомы. Рекомбинантный штамм, обозначенный J45-100, оказался очень чувствительным к сыворотке, имел пониженную вирулентность для свиней - (LD50 оказалась в 10 раз ниже, чем у исходного штамма) и должен обеспечивать защиту свиней от плевропневмонии.
Настоящее изобретение будет полезно для получения вакцин против инкапсулированных организмов, продуцирующих токсины или другие факторы вирулентности, при том, что капсула необходима для вирулентности, но не для иммунопротекции. Все, что необходимо осуществить в этом случае, - это клонировать гены, кодирующие у данного организма синтез капсулы, затем делегировать и заменить часть клонированного гена маркерным геном в суицидном векторе, затем ввести вектор в нужный организм и провести скрининг генетически модифицированных организмов, утративших капсулу.
Краткое описание чертежей
Задачи и преимущества изобретения будут более ясными из последующего изложения конкретных вариантов осуществления изобретения и чертежей, описание которых приведено ниже.
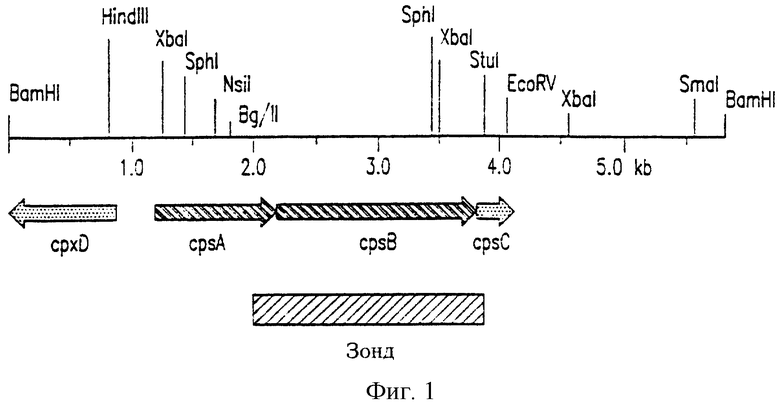
На фиг.1 представлена физическая карта клонированной в pCW-11Е ДНК из области, кодирующей синтез капсулы A.pleuropneumoniae J45. Указаны локализация и направление транскрипции двух полных открытых рамок считывания (ORFs) (cspA и cspB, темные стрелки), идентифицированных дидеоксисеквенированием. Указано также положение частичной третьей открытой рамки считывания (cspC). Указаны также локализация и направление транскрипции неполного гена экспорта капсулы cspD, локализованного в этом фрагменте ДНК. Приведен 2.1 кб BglII-StuI фрагмент pCW-11E, использованный в качестве ДНК-зонда в экспериментах, проиллюстрированных на фиг.2. Отмеченная точками стрелка обозначает неполные рамки считывания (ORFs).
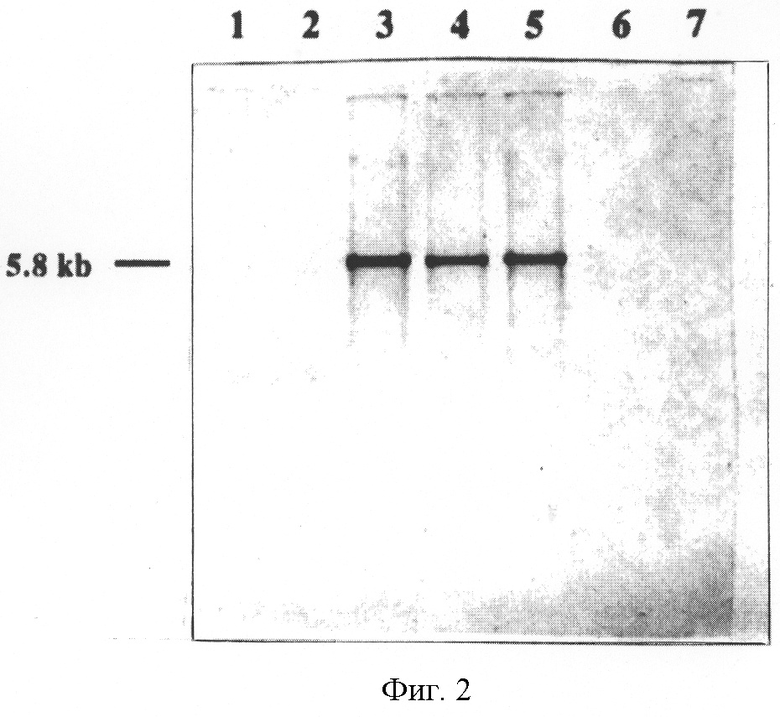
На фиг.2 приведены результаты Саузерн-блоттинга геномной ДНК А.pleuropneumoniae, гибридизованной с дигоксигенин-меченным 2.1 кб BglII-StuI фрагментом pCW-11E. ВamHI-рестрицированная геномная ДНК серотипа 1 штамма 4074 (полоса 1), серотипа 2 штамма 1536 (полоса 2), серотипа 5а штамма J45 (полоса 3), серотипа 5а штамма К17 (полоса 4), серотипа 5 штамма 178 (полоса 5), серотипа 7 штамма 29628 (полоса 6) и серотипа 9 штамма 13261 (полоса 7) были гибридизованы, как описано ниже. Указана молекулярная масса фрагментов (в кб) в полосах гибридизации.
На фиг.3а и 3b представлена нуклеотидная последовательность 3.2 кб HindIII-EcoRV фрагмента pCW-11E, содержащего серотип-специфическую ДНК Л. pleuropneumoniae J45 (SEQ.ID.NO.1). В данной последовательности предполагается наличие аминокислотных последовательностей двух полных открытых рамок считывания (ORFs): cspA (SEQ.ID.NO.2), cspB (SEQ.ID.NO.3) и N-концевой последовательности третьей неполной рамки считывания, cspC (SEQ.ID.NO.4), аббревиатуры которых указаны под нуклеотидной последовательностью. Предполагаемые сайты связывания рибосом каждой рамки считывания отмечены жирными буквами, отмечены также предполагаемые -10 и -35 промоторные последовательности “выше” cspA.
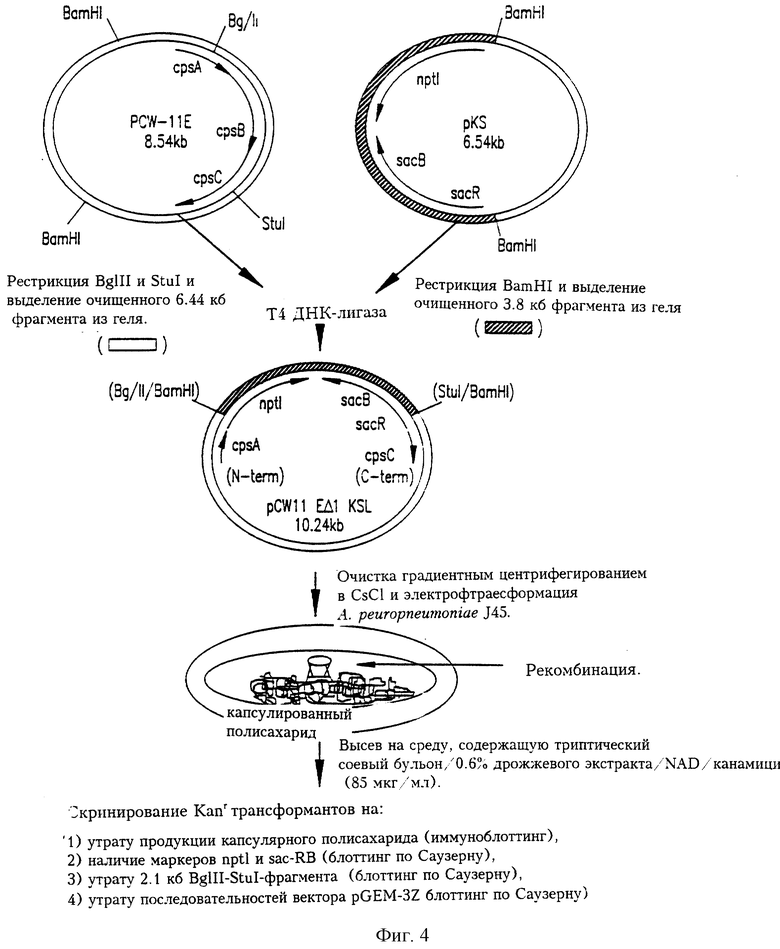
На фиг.4 представлены схема конструирования суицидного вектора, содержащего делению направляющей синтез капсулы ДНК, pCW11 ЕΔ1 KS1, и получения некапсулированных мутантов А. pleuropneumoniae J45 аллельным обменом. Плазмидный вектор pCW11 ЕΔ1 KS1 конструировали, рестрицируя pCW-11E BglII и StuI, затупляя концы и лигируя большой 6.4 кб фрагмент с 3.8 кб ВamHI-фрагментом pKS (также с затупленными концами), содержащим nptl-sacRB (Kanr SucS) кассету. Сайты рестрикции (в скобках) указывают на первоначальные концы фрагментов, встроенных в pCWI1 ЕΔ1 KS1. Вектор pCW11 ЕΔ1 KS1 был электротрансформирован в A.pleuropneumoniae и на среде, содержащей 85 мкг/мл канамицина, по утрате радужности были отобраны некапсулированные Каnr трансформанты.
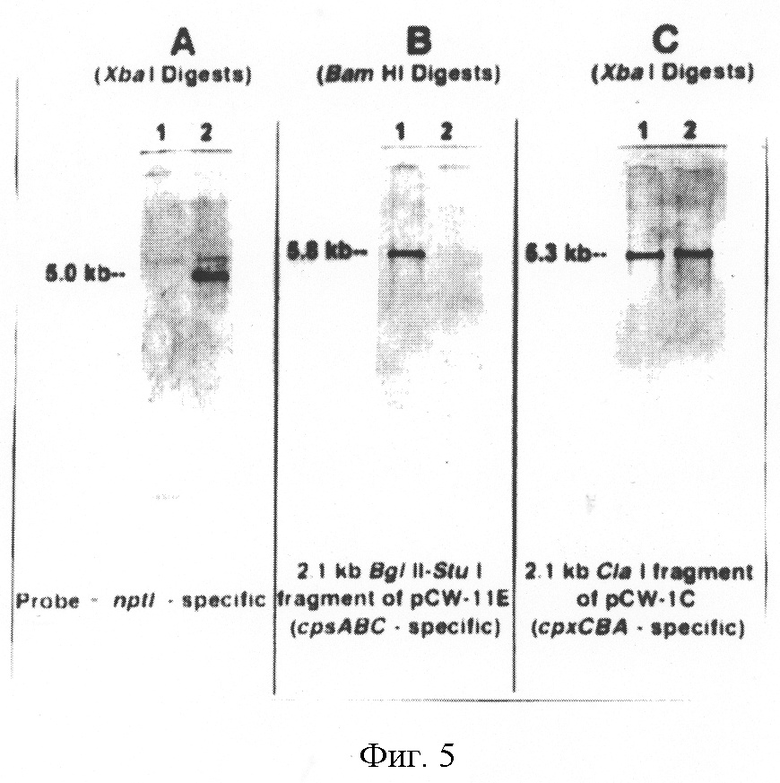
На фиг.5 приведены результаты Саузерн-блоттинга геномной ДНК, выделенной из A.pleuropneumoniae J45 (полоса 1) и J45-100 (полоса 2) после гибридизации с дигоксигенин-меченными зондами, специфичными к nptl или части локуса капсуляции A.pleuropneumoniae. Геномную ДНК A.pleuropneumoniae J45 (полоса 1) и J45-100 (полоса 2) рестрицировали ХbаI (панели А и С) или BamHI (панель В) и гибридизовали или с 1.24 кб PstT-фрагментом pKS (nptl-специфичный), панель А, или с 2.1 кб BglII-Stul-фрагментом pCW-11E (cpsABC-специфичный, см. фиг.1), панель В, или с 2.1 кб ClaI-фрагментом pCW-1C (срхСВА-специфичный, см. фиг.3.2), панель С.
На фиг.6 представлены результаты иммуноблоттинга колоний A.pleuropneumoniae J45 и J45-100 после реакции со свиной антисывороткой к капсулярному полисахариду. На нитроцеллюлозный фильтр было нанесено приблизительно 5×105 (полоса 1) или 5×104 (полоса 2) CFU (колониеобразующих единиц вируса) на ячейку. Нанесенные на фильтр бактерии лизировали хлороформом и фильтр инкубировали со свиной антисывороткой, содержащей антитела к капсулярному полисахариду серотипа 5а, но не к другим поверхностным антигенам A.pleuropneumoniae J45.
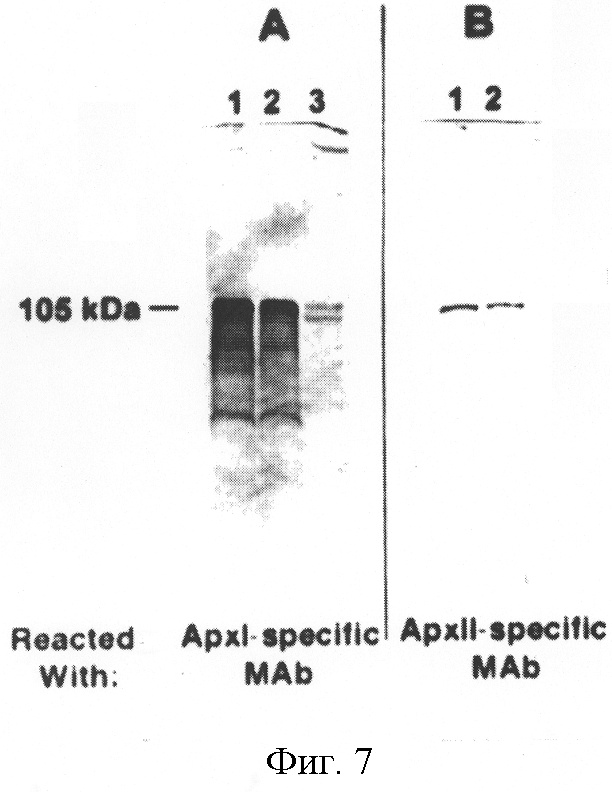
На фиг.7 представлены иммуноблоты концентрированных культуральных супернатантов A.pleuropneumoniae J45 (полоса 1) и J45-100 (полоса 2), содержащих преимущественно экзотоксины ApxI и ApxII. Панель А реагировала с ApxI-специфическими моноклональными антителами, а панель В - с ApxII-специфическими моноклональными антителами. В качестве отрицательного контроля в панель А был включен концентрированный культуральный супернатант A.pleuropneumoniae серотипа 2 штамма 1536 (полоса 3), поскольку данный серотип не синтезирует ApxI. Блот панели А обрабатывали ApxI-специфическими моноклональными антителами.
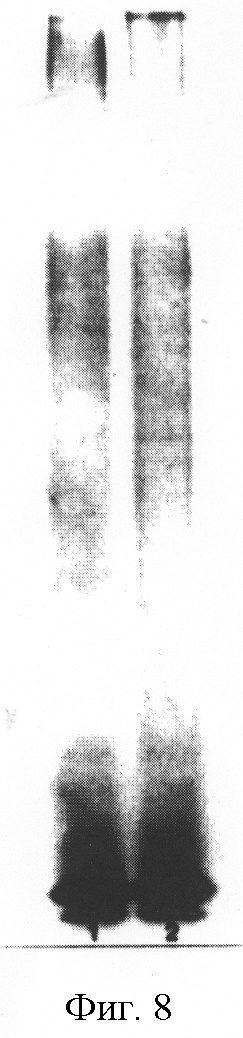
На фиг.8 представлены электрофоретические профили липополисахаридов (LPS), выделенных из A.pleuropneumoniae J45 (полоса 1) и рекомбинантного некапсулированного мутанта J45-100 (полоса 2). LPS подвергали электрофорезу в 15%-ном разделяющем геле и окрашивали аммонийным серебром.
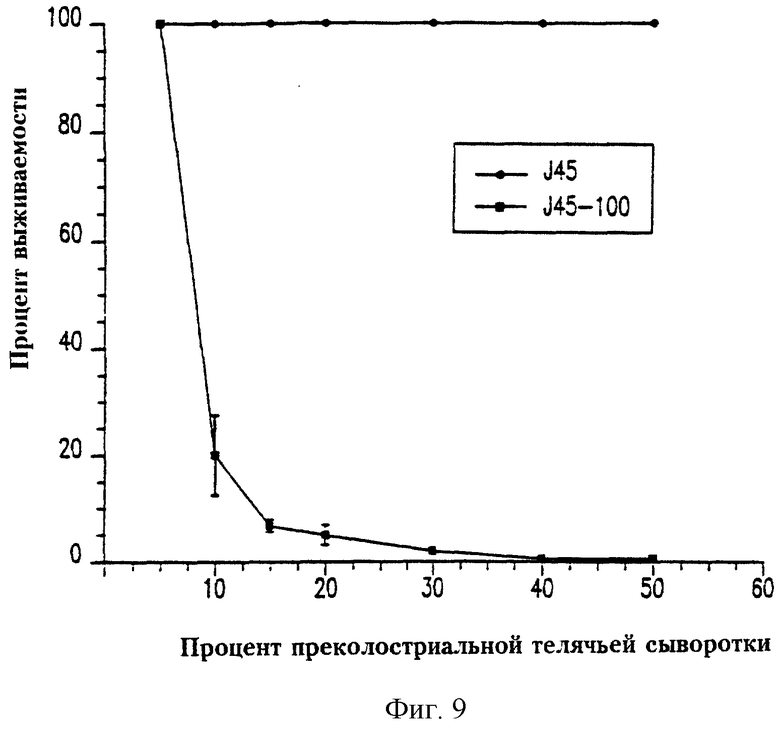
На фиг.9 отражена бактерицидная активность преколостриальной телячьей сыворотки для A. pleuropneumoniae J45 и J45-100. Процент жизнеспособных клеток для каждого штамма оценивали после 60 минут инкубации при 37°С. Каждая точка отражает среднее значение для трех независимых экспериментов, проведенных дважды. Представлено стандартное отклонение для каждого значения. Максимальный процент жизнеспособности, отмеченный для J45, составлял 100%, хотя эти значения были обычно выше, поскольку бактерии вырастали за время опыта. Значения, превышающие 100%, не учитывались, поскольку их нельзя было определить точно.
На фиг.10а и 10b представлена нуклеотидная последовательность 3.2 кб ХbаI-СlaI-фрагмента pCW-1C, кодирующего гены экспорта капсулы A.pleuropneumoniae J45 (SEQ.ID.No.5). Приведены предсказанные аминокислотные последовательности (SEQ.ID.Nos.6-8) белков, участвующих в экспорте капсулярного полисахарида А. pleuropneumoniae серотипа 5а.
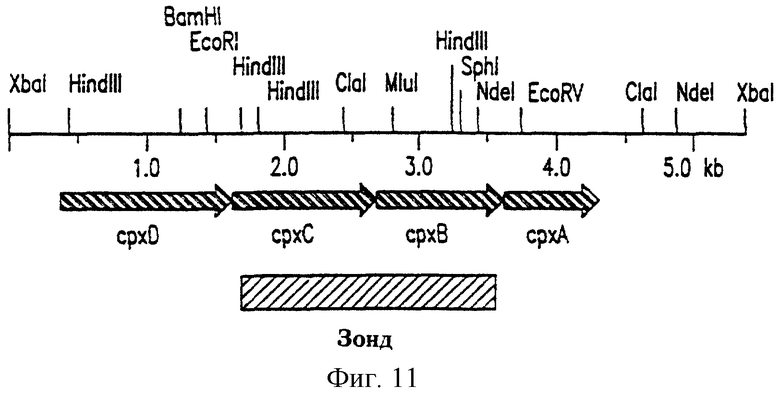
На фиг.11 приведена физическая карта ДНК pCW-1C из А.pleuropneumoniae J45.
Подробное описание предпочтительных вариантов осуществления изобретения
Изобретение относится к использованию живого, рекомбинантного, авирулентного штамма микроорганизма (например, бактерий или грибов), который в результате генно-инженерных манипуляций утратил капсулу, в качестве вакцины против вызываемого данным микроорганизмом заболевания. Изобретение будет полезно в случае таких заболеваний, когда капсула необходима для вирулентности, но не для иммунопротекции, и когда заболевание обусловлено токсинами или другими факторами вирулентности. В конкретном варианте осуществления изобретения был получен некапсулированный штамм Actinobacillus pleuropneumoniae, который может служить в качестве вакцинного против плевропневмонии свиней. Главной отличительной особенностью изобретения является генетическая модификация микроорганизма, которая, как в случае варианта осуществления при использовании Actinobacillus pleuropneumoniae, означает генерацию делеции дезоксирибонуклеиновой кислоты (ДНК) в той области, которая ответственна за синтез капсулы. Исключительно для целей иллюстрации изобретения описано получение трансформированного мутанта Actinobacillus pleuropneumoniae серотипа 5а, однако необходимо понимать, что согласно изложенному ниже способу могут быть получены и другие серотипы, которые могут оказаться полезными в качестве вакцин сами по себе или в сочетании с одним или несколькими рекомбинантными мутантами различных серотипов.
Охарактеризованный ниже штамм, а также другие штаммы некапсулированных токсигенных бактерий и других микроорганизмов, полученные согласно описанным ниже процедурам, могут представлять собой эффективные вакцины, поскольку они авирулентны, но продуцируют все антигены, необходимые для развития у хозяина защитного иммунного ответа. Вакцины могут вводиться различными способами, однако внутримышечные и подкожные инъекции предпочтительней. Преимущество данных живых вакцин заключается в том, что токсины, которые первоначально ответственны за развитие заболевания, а также другие компоненты, вырабатываемые только живыми организмами или in vivo, будут производиться в месте иммунизации, приводя к генерации у хозяина иммунного ответа, который защищает его от вызываемых токсинами повреждений. Поэтому заболевание (острое или хроническое) не развивается. Организмы не могут диссеминировать, поскольку без капсулы они крайне чувствительны к сыворотке и немедленно выводятся из кровотока или респираторного тракта. Кроме того, в случае живой вакцины клеточный иммунный ответ будет более высоким и защитный эффект более продолжительным, нежели в случае убитой вакцины.
Пример
Была идентифицировна и охарактеризована область ДНК, задействованная в биосинтезе капсулярного полисахарида у Actinobacillus pleuropneumoniae. Зонд, специфичный к cpxD гену, участвующему в экспорте капсулярного полисахарида у A.pleuropneumoniae серотипа 5а J45, был использован для идентификации и клонирования прилегающего 5.8 кб ВаmHI-фрагмента геномной ДНК J45. Результаты блоттинга по Саузерну показали, что часть этой области содержит ДНК, которая является серотип-специфической. Секвенироване ДНК выявило, что эта область содержит две полные открытые рамки считывания cpsA и cpsB и потенциальную неполную рамку считывания cpsC. CpsA и cpsB обладают слабой гомологией с гликозилтрансферазами, участвующими в биосинтезе липополисахарида Escfierichia coli и капсулярного полисахарида Haemophilus influenzae типа b соответственно. Для получения мутанта J45-100 деления размером 2.1 кб, покрывающая клонированные открытые рамки считывания cpsABC, была введена аллельной заменой в хромосому J45. Этот мутант не продуцирует внутриклеточного или внеклеточного капсулярного полисахарпда, что свидетельствует о том, что cpsA, cpsB и/или cpsC участвуют в биосинтезе капсулярного полисахарида А. pleuropneumoniae. Профили Арх-токсина и липополисахарида у J45-100 были идентичны таковым у капсулированного исходного штамма J45. Однако, J45-100 in vitro рос быстрее, чем J45. J45-100 был чувствителен к преколостриальной телячьей сыворотке, а J45 - нет. При интратрахеальном заражении свиней в дозе, равной трем LD50 штамма J45, J45-100 оказался невирулентным. При заражении свиней в дозе, равной шести LD50 штамма J45, J45-100 вызывал умеренные повреждения легких, но не гибель. Эти результаты свидетельствуют о том, что капсулярный полисахарид является главной детерминантой устойчивости к сыворотке и вирулентности у A.pleuropneumoniae.
Материалы и методы
Бактериальные штаммы, плазмиды и условия выращивания
Бактериальные штаммы и плазмиды, использованные в настоящем исследовании, приведены в Таблице 1. Для выделения геномной ДНК и бактерицидных тестов штаммы A.pleuropneumoniae выращивали при встряхивании при 37°С в экстракте мозга и сердца (Difco Laboratories, Detroit. Mich.), содержащем 5 мкг/мл никотинамидадениндинуклеотида (NAD) (Sigma Chemical Co., St.Lous, Mo.). Для электропорации штаммы A.pleuropneumoniae выращивали при встряхивании при 37°С в триптическом соевом бульоне (Difco Laboratories), содержащем 0.6% дрожжевого экстракта (Difco Laboratories) и 5 мкг/мл никотинамидадениндинуклеотида (NAD) (TSY-N). Для экспериментов по заражению свиней штаммы A.pleuropneumoniae выращивали при встряхивании при 37°С в бульоне Колумбия (Difco Laboratories), содержащем 5 мкг/мл NAD. Штаммы Escherichia coli выращивали на бульоне Лурии-Бертани (Sambrook et al., 1989) при рутинном культивировании или на Террифик бульоне (Tartof and Hobbes, 1987) для выделения плазмид. Для поддержания плазмид в Е. Coli в ростовую среду добавляли антибиотики в следующих концентрациях: ампициллин (Аmр) 100 мкг/мл и канамицин (Каn) 50 мкг/мл. Канамицин в концентрации 85 мкг/мл использовали для селекции рекомбинантных мутантов А.pleuropneumoniae.

Вычисление времени генерации
Время генерации в логарифмической фазе роста для штаммов А.pleuropneumoniae, выращиваемых на среде TSY-N, вычисляли по следующему уравнению: R=1/g, где R - средняя скорость роста бактерий, a g - время генерации популяции бактерий (Pelczar et al., 1993). Среднюю скорость роста, R, вычисляли по следующему уравнению: R=3.32(log10 N-log10 N0)/t, где t - истекшее время, N - число бактерий на момент t, а N0 -исходное число бактерий на момент t=0 (Pelczar et al., 1993)
Гибридизационный анализ ДНК
ДНК, переваренную эндонуклеазами рестрикции (приблизительно 5 мкг на полосу), подвергали электрофорезу в 0,7%-ном геле агарозы и переносили за счет капиллярных сил на найлоновые фильтры MagnaGraph (Micron Separations Inc., Westboro, Mass.) с использованием 20Х натрий-цитратного раствора (20Х SSC - это 3 М NaCl, 300 мМ цитрат натрия, рН 7), как описано ранее (Sambrook et al., 1989; Southern, 1975). ДНК ковалентно “пришивали” к найлоновым фильтрам ультрафиолетовым облучением с помощью UV Stratalinker (Stratagene, La Jolla, Calif.). Дигоксигенин-меченные зонды для гибридизации ДНК синтезировали методом выборочной затравки с использованием нерадиоактивной системы мечения и проявления Genius System (Boehringer Mannheim Corp, Indianapolis, Ind.) согласно инструкциям производителя. Гибридизацию ДНК проводили при 68°С в растворах, содержащих 5Х SSC. Фильтры отмывали и проявляли согласно инструкциям Genius System для колориметрического обнаружения.
Методы рекомбинантных ДНК и реагенты
Геномную ДНК из выращенных в жидкой среде клеток A.pleuropneumoniae выделяли по методу, описанному S.Spinola. Вкратце, бактерии ресуспендировали в 10 мМ Tris-EDTA (рН 8) и инкубировали с додецил-сульфатом натрия (0.66%) и РНКазой (100 мкг/мл) 1 час при 56°С. Смесь экстрагировали один раз забуференным фенолом и четыре раза забуференным фенол-хлороформом (Amresco, Inc., Solon, Ohio), геномную ДНК осаждали этанолом и ресуспендировали в 10 мМ Тris-1 мМ EDTA (рН 8). Плазмидную ДНК выделяли быстрым методом щелочного лизиса (Ish-Horowicz and Burke, 1981). Рестрикционные фрагменты, необходимые для клонирования и синтеза зондов, элюировали из агарозных гелей, как описано ранее (Zhen and Swank, 1993). Рестрикцию, электрофорез в гелях агарозы и лигирование ДНК проводили, как описано ранее (Sambrook et al., 1989). Концы рестрикционных фрагментов затупляли, достраивая 5’-вилки нуклеотидами (dNTPs) с использованием фрагмента Кленова ДНК-полимеразы 1, как описано ранее (Sambrook et al., 1989). Плазмидной ДНК трансформировали штаммы Е.соli при помощи электропорации (Dower et al.,1988) с использованием электропоратора ВТХ ЕСМ 600 (ВТХ, Inc., San Diego, Calif.).
Эндонуклеазы рестрикции и фрагмент Кленова ДНК-полимеразы 1 получали от Promega Corporation (Madison, Wis.). ДНК-лигаза Т4 была получена от Gibco BRL (Gaithersburg, Md.). Нуклеотиды (dNTPs) для достройки концов были получены от Boehringer Mannheim Corp. (Indianapolis, Ind.).
Секвенирование и анализ ДНК
Определяли нуклеотидную последовательность обеих цепей 2.7 кб XbaI-EcoRV-фрагмента ДНК pCW-11E при помощи дидезокси-метода терминации цепи (Sanger et al.,1977) с использованием версии 2.0 набора для секвенирования ДНК Sequenase (United States Biochemical Corp., Cleveland, Ohio) с α 35[S]dATP (DuPont/NEN Research Products, Boston, Mass.) Двуцепочечные матрицы ДНК были секвенированы с использованием специальных олигонуклеотидных праймеров (DNAgency, Inc., Malverne, Pa.) с тем, чтобы сделать возможным прочтение каждой цепи.
Полученные нуклеотидные последовательности были смешаны с нуклеотидной последовательностью 4.6 кб XbaI-ClaI-фрагмента pCW-1C, кодирующей расположенные “выше” структурные капсулярные гены (фиг.10), и были проанализированы с помощью программы DNASTAR (DNASTAR, Inc., Madison, Wis.). Поиск сходства последовательностей в базах данных EMBUGenBank/DDBJ был проведен с помощью программы BLAST (Altschul et al., 1990) в Национальном Центре Информации по Биотехнологии (Bethesda, Md.).
Консервативные области cap локуса Н.influenzae типа b, задействованные в экспорте капсулярного полисахарида, были использованы для идентификации, клонирования и характеристики части капсулярного локуса A.pleuropneumoniae серотипа 5а, участвующей в экспорте капсулярного полисахарида. Был проведен анализ Саузерн-блоттингом геномной ДНК A.pleuropneumoniae серотипа 5а штамма J45 с зондами, специфичными к непрерывным областям локуса капсуляции (capb) Н.influenzae типа b. Данные зонды не гибридизовались с геномной ДНК A. pleuropneumoniae в жестких условиях (68°С, 5xSSC), но гибридизовались в мягких и средних условиях (55°С, 5×SSC). 4.4 кб EcoRI-фрагмент локуса сарb Н.influenzae из плазмиды pSKHl, содержащей область 1 гена bехD, участвующего в экспорте капсулярного полисахарида, и две открытые рамки считывания области 2, участвующие в биосинтезе капсулярного полисахарида, гибридизовался с 1.2 кб HindIII и 5.3 кб ХbaI-фрагментами геномной ДНК J45. 9.0 кб FcoRI-фрагмент локуса capb H.influenzae из плазмиды pSKH2, содержащей область 1 генов bехСВА, участвующих в экспорте капсулярного полисахарида, некую неохарактеризованную область ДНК 3, общую для нескольких серотипов Н.influenzae, и некую область ДНК 2, участвующую в биосинтезе капсулярного полисахарида, гибридизовался с 1.5 кб HindIII, 5.3 кб XbaI и 2.4 кб XhoI-фрагментами геномной ДНК J45. Полученные данные свидетельствуют о том, что локусы капсулярных генов Н.influenzae типа b и A.pleuropneumoniae серотипа 5а имеют гомологичные области. Оба специфических зонда Н.influenzae capb содержали область ДНК 1, участвующую в экспорте капсулярного полисахарида, позволяя предположить, что 5.3 кб XbaI-фрагмент геномной ДНК J45, который гибридизовался с двумя зондами Н.influenzae capb, может содержать гены, кодирующие белки, которые участвуют в экспорте капсулярного полисахарида. A.pleuropneumoniae серотипа 5а. 5.3 кб ХbaI-фрагмент геномной ДНК J45, который гибридизовался с двумя зондами capb H.influenzae, был клонирован по сайту XbaI в плазмиду pGEM-3Z (в обеих ориентациях). Этот фрагмент был выбран из ХbaI-рестрицированных фрагментов геномной ДНК J45 в пределах от 4.8 до 6.0 кб, которые после электрофореза были элюированы из агарозного геля. Одна из полученных плазмид была обозначена pCW-1C. Был проведен Саузерн-блоттинг с целью определения, что bexD, bехС, bехВ и bехА H.influenzae типа b гибридизуются с прилегающими фрагментами pCW-1C в том же порядке (bехDСВА), в котором эти гены расположены у H.influenzae. Результаты свидетельствуют о том, что была успешно клонирована та область ДНК A.pleuropneumoniae серотипа 5а, которая необходима для экспорта капсулярного полисахарида, и что данная область организована аналогично локусу bex H.influenzae типа b.
Была определена нуклеотидная последовательность 4.6 кб XbaI-ClaI-рестрикционного фрагмента и 3.2 ХbaI-СlaI-рестрикцнонный фрагмент представлен на фиг.10а-b (SED ID NO 5). Четыре открытые рамки считывания (показаны на фиг.10а-b и 11), обозначенные как cpxDCBA (сокращение срх использовано для обозначения экспорта капсулярного полисахарида), были обнаружены на одной и той же цепи ДНК в непосредственной близости друг к другу. Инициирующий кодон AUG cpxC (SED ID NO.7) находился на 26 нуклеотидов “ниже” терминирующего кодона UAA cpxD (SED ID NO.6), тогда как инициирующий кодон AUG срхB (SED ID NO.8) перекрывал терминирующий кодон UAA cpxC (SED ID NO.7) и инициирующий кодон AUG срхА частично перекрывал терминирующий кодон UGA cpxB (SED ID NO.8). Консенсусные последовательности связывания рибосом Шайн-Дальгарно были обнаружены на 17 нуклеотидов “выше” каждого инициирующего кодона AUG и предположительно содержащие промотор последовательности, аналогичные консенсусным последовательностям E.coli σ70-10 (ТАТААТ) и -35 (TTGACA), были идентифицированы “выше” cpxD (SED ID NO.6). “Выше” срхА была идентифицирована палиндромная последовательность, которая может функционировать как rho-независимый сигнал терминации транскрипции (не показано). Генетическая организация дает основания полагать, что cpxDCBA транскрибируется в виде одной полицистронной мРНК.
Электротрансформация A.pleuropneumoniae
A.pleuropneumoniae выращивали до средней логрифмической фазы в TSY-N, осаждали центрифугированием при 7000×g и 4°С, а затем промывали четыре раза в охлажденном (4°С), стерилизованном фильтрованием буфере, содержащем 272 мМ маннитола, 2.43 мМ К2НРО4, 0.57 мМ KH2PO4 15% глицерина, рН 7.5. Этот буфер был модифицирован (содержит маннитол вместо сахарозы) по сравнению с ранее описанным буфером, использованным для отмывания клеток A. pleuropneumoniae перед электропорацией (Lalonde et al., 1989b). Затем клетки отмывали однократно в охлажденном, стерилизованном фильтрованием 15%-ном глицерине и ресуспендировали в 15%-ном глицерине до концентрации 1010 CFU/мл. Аликвоты этой суспензии (90 мкл) смешивали (в 1.5 мкл дистиллированной воды) с 1.5-2.0 мкг плазмидной ДНК, очищенной ультрацентрифугированием в градиенте плотности цезия (Sambrook et al., 1989), помещали в охлажденные 2-мм кюветы для электропорации (ВТХ, Inc.) и электропорировали с использованием электропоратора ВТХ ЕСМ 600 (ВТХ, Inc.) со следующими параметрами: вольтаж заряда 2.5 kV, a сопротивление R7 (246 Ом). Реально отпущенный импульс составлял 2.39 kV при продолжительности около 10.7 миллисекунд. После электропорации клетки “восстанавливали”, инкубируя при осторожном встряхивании 3.5 часа при 37°С в 1 мл TSY-N, содержащем 5 мМ MgCl2). Затем клетки высевали на TSY-N агар, содержащий 85 мкг канамицина на мл и инкубировали при 37°С.
Иммуноблоттинг
Для иммуноблоттинга колоний целые клетки A.pleuropneumoniae, выращенные в течение ночи на чашках с TSY-N агаром, собирали в фосфатный буферный раствор (PBS) и доводили концентрацию до 109 CFU/мл, что определяли спектрофотометрически. На нитроцеллюлозный фильтр (NitroBind; Micron Separations Inc.,) наносили приблизительно от 5×104 CFU до 5×105 CFU на ячейку с помощью аппарата Bio-Dot (Bio-Rad Laboratories, Richmond, Calif.). Фильтры помещали в хлороформ на 15 минут при комнатной температуре для лизиса бактериальных клеток на фильтре. Затем фильтр досуха подсушивали на воздухе и инкубировали 1 час при комнатной температуре в Трис-буфере, рН 7.5 (TBS), содержащем 2% снятого молока для блокирования неспецифических сайтов связывания на фильтре. Фильтр инкубировали 1 час при комнатной температуре с разведенной 1:2000 (2% молоко-TBS) адсорбированной свиной антисывороткой, которая содержала антитела к капсулярному полисахариду серотипа 5а, но не к другим поверхностным антигенам A.pleuropneumoniae. Эта антисыворотка, обогащенная антителами к капсулярному полисахариду, была получена истощением гипериммунной свиной антисыворотки к A.pleuropneumoniae K17 спонтанным некапсулированным мутантом, К17-С (Inzana and Mathison, 1987), как описано ранее (Inzana, 1995). Фильтр отмывали TBS, содержащим 0.05% Твин 20, а затем инкубировали 1 час при комнатной температуре с кроличьими антисвиными IgG в разведении 1:1000, конъюгированными с пероксидазой хрена (тяжелые и легкие цепи; Cappel, Durham, N.C.). Фильтр отмывали TBS и проявляли 4-хлор-1-нафтолом (Bio-Rad Laboratories) в TBS, содержащем 0.02% Н2О2.
Иммуноблоттинг концентрированных культуральных супернатантов A.pleuropneumoniae проводили, как описано ранее (Ма and Inzana, 1990). В частности, приблизительно 15 мкг общего белка культурального супернатанта разделяли в непрерывном SDS-PAGE (Laemmli, 1970) в 8%-ном разделяющем геле. Белки переносили на нитроцеллюлозный фильтр (NitroBind; Micron Separations Inc.,) по методу Towbin et al. (1979). Фильтр инкубировали в TBS, содержащем 2% бычьего сывороточного альбумина, для блокирования неспецифического связывания и разрезали на полоски. Полоски инкубировали в течение ночи при 4°С с моноклональными антителами, специфическими к токсину ApxII (Ma and Inzana, 1990), или с моноклональными антителами, специфическими к токсину ApxI (Devendish et al., 1989; Frey et al., 1992) и отмывали TBS. Блот, реагировавший с ApxII-специфическими моноклональными антителами, инкубировали с козьими антимышиными IgG в разведении 1:2000, конъюгированными с пероксидазой хрена (Cappel), отмывали TBS и проявляли, как описано ранее. Блот, реагировавший с ApxI-специфическими моноклональными антителами, инкубировали с козьими антимышиными IgG в разведении 1:2000, конъюгированными с щелочной фосфатазой, и проявляли, как описано ранее (Frey et al., 1992).
Экстракция и электрофорез LPS
Липополисахариды (LPS) экстрагировали из A.pleuropneumoniae с применением микрометода экстракции горячим фенолом, как описано ранее (Inzana, 1983). Очищенные LPS подвергали электрофорезу в 15%-ном разделяющем полиакриламидном геле, содержащем мочевину, как описано ранее (Inzana, 1988). Электрофоретические профили LPS визуалилизировали, окрашивая гель аммонийным серебром (Tsai and Frasch, 1982).
Тест на бактерицидность сыворотки
Определяли чуствительность A.pleuropneumonuie к бактерицидной активности преколостриальной телячьей сыворотки. Процент жизнеспособных бактерий в присутствии 5, 10, 15, 20, 30, 40 и 50% преколостриальной телячьей сыворотки оценивали после 60 минут инкубации при 37°С.
Изучение вирулентности
Поросят в возрасте от 7 до 9 недель получали из двух местных хозяйств, свободных от инфекции A.pleuropneumoniae, и распределяли на две группы случайным образом. Поросят различных групп содержали в различных загонах, что исключало прямой физический контакт между животными различных групп. Виварий Виргинского политехнического института и Университета штата функционирует в соответствии с требованиями Американской ассоциации по аккредитации лабораторий для работы с животными. Для экспериментов по заражению поросят штаммы A.pleuropneumoniae выращивали при встряхивании в бульоне Колумбия (Difco Laboratories), содержащем 5 мкг/мл NAD, осаждали при 7000×g и ресуспендировали до концентрации 109 CFU/мл в PBS. Поросят заражали интратрахеально, вводя 10 мл разведения данной суспензии. При этом животные находились под мягким седативным воздействием стреснила (Stresnil) (Pittman-Moore.Inc., Washington Crossingm N.J.). Поросят вскрывали как можно скорее после гибели или после эвтаназии, произведенной пентобарбиталом натрия. Повреждения легких оценивались ветеринарным патологоанатомом по следующим критериям: 0, ничем не примечательные легкие (нет заметных повреждений); 1+, 1-10% ткани легких повреждено каким-либо сочетанием закупорки, эдемы, геморрагий, уплотнений и/или плеврита; 2+, 11-49% ткани легкого повреждено; 3+, 50-74% ткани легкого повреждено; 4+, 75% или более ткани легкого повреждено. При вскрытии забирали образцы ткани легкого из правой краниально-дорсальной части каудальной доли и культивировали в экстраке мозга и сердца, содержащем NAD, для обнаружения A.pleuropneumoniae.
Результаты
Идентификация и клонирование серотип-специфической области ДНК A.pleuropneumoniae
Для идентификации и клонирования ДНК A.pleuropneumoniae J45, задействованной в биосинтезе капсулярного полисахарида, был проведен блоттинг по Саузерну с целью идентификации области ДНК, отстоящей “выше” (в 5’-направлении) от кластера генов cpxDCBA, участвующих в экспорте капсулярного полисахарида (фиг.10а-b и 11). Ожидалось, что расположенная “выше” область ДНК будет кодировать серотип-специфические гены, участвующие в биосинтезе капсулярного полисахарида, поскольку локус капсуляции cap у A.pleuropneumoniae должен быть организован аналогично локусам капсуляции у Haemophilus influenzae типа b и Neisseria meningitidis группы В. Геномную ДНК A.pleuropneumoniae J45, рестрицированную BamHI, гибридизовали с меченным дигоксигенином 1.2 кб BamHI-XbaI-фрагментом pCW-1C, содержащим часть гена cpxD. Данный cpxD-специфический зонд гибридизовался с единственным 5.8 кб BamHI-фрагментом геномной ДНК (не показано). Данный 5.8 кб BamHI-фрагмент клонировали по BamHI-сайту в pGEM-3Z, причем источником данного фрагмента служили фрагменты геномной ДНК A.pleuropneumoniae J45, рестрицированной BamHI, в пределах от 5.0 до 6.5 кб, которые после электрофоретического разделения были элюированы из геля. Полученная плазмида была обозначена pCW-11E и была получена ее рестрикционная карта (фиг.1). Часть ДНК-вставки в pCW-11Е (1.2 кб BamHI-ХbaI-фрагмент) перекрывалась с ДНК-вставкой в pCW-1C.
ВаmHI-рестрицированную геномную ДНК нескольких различных серотипов A.pleuropneumoniae гибридизовали с 2.1кб BglII-StuI-фрагментом pCW-11E (фиг.1) для определения серотип-специфичности данной области (фиг.2). 2.1кб ВglII-Stul-фрагмент гибридизовали с 5.8 кб BamHI-фрагментом геномной ДНК трех штаммов A.pleuropneumoniae серотипа 5а, но не с геномной ДНК серотипов 1, 2, 7 и 9 (фиг.2). Таким образом, ДНК A.pleuropneumoniae в pCW-11E содержала ДНК, специфичную для штаммов серотипа 5. Поскольку данная ДНК была серотип-специфической, весьма вероятно, что она участвует в синтезе капсулярного полисахарида.
Нуклеотидная последовательность и анализ серотип-специфической области ДНК A.pleuropneumoniae
Была определена нуклеотидная последовательность 2.7 кб Xbal-EcoRV-фрагмента pCW-11E. Данную нуклеотидную последовательность смешали с нуклеотидной последовательностью 4.6 кб Clal-ХbaI-фрагмента pCW-1C и проверили на наличие ранее не идентифицированных открытых рамок считывания (ORFs). Нуклеотидная последовательность 3.2 кб HindIII-ЕсоRV-фрагмента. pCW-11E содержала новые неидентифицированные ORFs, которые приведены на фиг.3. Две полные ORFs, обозначенные cpsA и cpsB (аббревиатура cps означает синтез капсулярного полисахарида), были идентифицированы “выше” и на противоположной цепи гена cpxD, участвующего в экспорте капсулярного полисахарида A.pleuropneumoniae (фиг.1 и 3). Инициирующий кодон AUG cpsB был расположен на 3 нуклеотида “ниже” от терминирующего кодона UAA cpsA. Инициирующий кодон AUG третьей потенциальной ORF, cpsC, был идентифицирован на 15 оснований “ниже” терминирующего кодона UAA cpsB. Консенсусные рибосом-связывающие последовательности Шайн-Дальгарно (Shine and Dalgarno, 1974) были идентифицированы на 13 оснований “выше” инициирующих кодонов AUG cpsA, cpsB и cpsC (фиг.3). Предполагаемый промотор, содержащий последовательность, сходную с консенсусными последовательностями Е.соli70-10 (ТАТААТ) и -35 (TTGACA) (Hawley and McClure, 1983), был идентифицирован “выше” cpsA (фиг.3). Близкое “соседство” cpsABC, а также обнаружение “выше” них предполагаемого промотора дают основания предполагать, что эти ORFs могут котранскрибироваться. Содержание G+C в области ДНК, кодирующей cpsABC, составляло 28%.
Предсказанные полипептиды cpsA и cpsB состояли из 321 (CpsA) и 526 (CpsB) аминокислот (фиг.3). Предсказанные молекулярные массы CpsA и CpsB составляли 36.9 и 61.7 килодальтон (кДа) соответственно. Гидропатические графики показали, что CpsA и CpsB являются относительно гидрофильными белками, что может свидетельствовать об их ассоциации с цитоплазматическим компартментом A.pleuropneumoniae (данные не приведены). Поиски с помощью программы BLAST (Altschul et al.,1990) в комбинированных неизбыточных базах данных нуклеотидных и белковых последовательностей в Национальном Центре Информации по Биотехнологии не выявили какой-либо существенной гомологии на нуклеотидном и аминокислотном уровнях между cpsABC и другими последовательностями, представленными в базах данных (данные не приведены). Однако слабая гомология (15% сходства) наблюдалась между CspA и белком Rfb E.coli, O-антиген гликозилтрансферазой, участвующей в биосинтезе LPS (Cheah and Manning, 1993). Низкая степень гомологии (около 14% сходства) была установлена между CspB и предсказанным белковым продуктом области 2 открытой рамки считывания 3 (ORF3) локуса капсуляции H.influenzae типа b. Предсказанный белок ORF3 участвует в биосинтезе полирибозилрибитолфосфата капсулярного полисахарида Н.influenzae типа b (Van Eldere et al., 1995). He было обнаружено какой-либо существенной гомологии между 83 N-концевыми аминокислотами CpsC и другими белками, последовательности которых содержатся в базах данных.
Получение канамицин-устойчивых трансформантов A.pleuropneumoniae серотипа 5а
A.pleuropneumoniae
На фиг.4 схематически представлена методика, использованная для получения рекомбинатных некапсулированных мутантов A.pleuropneumoniae J45 путем гомологичной рекомбинации и аллельного обмена. Исходно, для того, чтобы осуществить обмен капсулирующей ДНК A.pleuropneumoniae дикого типа на генетически-измененную капсулирующую ДНК A.pleuropneumoniae путем двойной гомологичной рекомбинации с кроссинговером, был сконструирован нереплицирующийся суицидный вектор pCW11EΔ1KS1. Вектор pCW11EΔ1KS1 получали, рестрицируя pCW-11E эндонуклеазами BglII и StuI для получения большой делеции в серотип-специфической капсулирующей ДНК A.pleuropneumoniae. Концы рестрицироваиной ДНК затупляли и большой 6.4 кб фрагмент дотировали с 3.8 кб BamHI-фрагментом pKS (также с затупленными концами), содержащим кассету nptl-sacR-sacB. Данная кассета содержит ген nptl Tn903, который обеспечивает A.pleuropneumoniae устойчивость к канамицину (Kanr) (Tascon et al., 1994), и последовательности sacRB, ответственные за чувствительность к сахарозе (Sucs) многих грамотрицательных бактерий (Gay et al., 1983; Ried and Collmer, 1987). Введенная в pCW11EΔ1KS1 делеция перекрывала cpsABC (фиг.1 и 4) и потому должна влиять на белковые продукты указанных открытых рамок считывания.
Вектор pCW11EΔ1KS1 нe реплицируется в A.pleuropneumoniae и потому функционирует как суицидный вектор. После электропорации вектора pCW11ЕΔ1КS1 в A.pleuropneumoniae J45 и выращивания клеток при 37°С в течение 2 дней было получено семь канамицин-устойчивых трансформантов. Четыре из этих канамицин-устойчивых трансформантов J45 не обладали радужностью при просмотре чашек в косом освещении, что давало основание считать их некапсулированными (данные не приведены). Состав среды, использованной для выращивания A.pleuropneumoniae перед электропорацией вектором pCW11EΔ1KS1, был очень важен, поскольку никогда не было получено некапсулированных канамицин-устойчивых трансформантов, если A.pleuropneumoniae выращивали на экстракте мозга и сердца с добавлением NAD.
Генотипическая характеристика канамицин-устойчивых трансформантов A.pleuropneumoniae
Первичный гибридизационный анализ колоний семи канамицин-устойчивых трансформантов показал, что четыре трансформанта, которые при визуальной инспекции оказались не капсулированными, гибридизовались с nptl-специфическим ДНК-зондом (1.24 кб PstI-фрагментом pKS), но не с зондами, специфичными к pGEM-3Z (I.I кб BglII-фрагмент pGEM-3Z) или серотип-специфическим 2.1 кб BglII-StuI-фрагментом pCW-11E (данные не приведены). Данные результаты свидетельствуют о том, что акт двойной рекомбинации произошел в каждом из этих четырех канамицин-устойчивых трансформантов. Наоборот, колонии трех других канамицин-устойчивых трансформантов гибридизовались с зондами, специфичными к гену npti, pGEM-3Z и 2.1 кб BglII-StuI-фрагменту pCW-11E, что свидетельствует об единичном кроссинговере и о том, что полный суицидный вектор pCW11EΔ1KS1 встроился в хромосому указанных трансформантов (данные не приведены). Результаты анализа блоттингом по Саузерну геномной ДНК, выделенной из четырех канамицин-устойчивых, потенциально некапсулированных трансформантов (с использованием описанных выше зондов), оказались идентичны, что указывает на то, что в каждом из этих трансформантов имела место одна и та же двойная рекомбинация. Один из этих трансформантов был случайным образом выбран для дальнейших исследований и обозначен J45-100.
Был проведен блоттинг по Саузерну геномной ДНК, выделенной из J45 и J45-100, с использованием ДНК-зондов, специфичных к генам nptl, к 2.1 кб ВglII-Stul-фрагменту pCW-11E и 2.1 кб ClaI-фрагменту pCW-1C (фиг.5). Nptl-специфичный ДНК-зонд гибридизовался с 5.0 кб фрагментом ХbaI-рестрицированной ДНК J45-100, но не ДНК J45, что подтверждает присутствие маркера npte в хромосоме J45-100 (фиг.5А). Гибридизация nptl-зонда с 5.0 кб ХbaI-фрагментом геномной ДНК J45-100 указывала на фрагмент того же размера в XbaI-рестрицированной ДНК суицидного вектора pCW11EΔ1KS1, использованного для получения J45-100. 2.1 кб BglII-StuI-фрагмент pCW-11E гибридизовался с 5.8 кб BamHI-рестрицированной геномной ДНК J45, но не ДНК J45-100, что подтверждает делецию данного фрагмента из J45-100 (фиг.5В). Зонд, специфичный к генам срхСВА, участвующим в экспорте капсулярного полисахарида (2.1 кб СlaI-фрагмент pCW-1C) гибридизовался с 5.3 кб СlaI-фрагментом как J45, так и J45-100 (фиг.5С). Данный результат свидетельствует о том, что часть локуса инкапсуляции A.pleuropneumoniae осталась незатронутой двойной рекомбинацией, произошедшей в соседних областях ДНК. Зонд, специфичный к pGEM-3Z, не гибридизовался с геномной ДНК как J45, так и J45-100, что подтверждает отсутствие ДНК вектора в геноме J45-100. В совокупности результаты гибридизации ДНК свидетельствуют о том, что желаемая двойная рекомбинация и аллельный обмен действительно произошли в J45-100.
Фенотипическая характеристика канамицин-устойчивого трансформанта A.pleuropneumoniae, J45-100
J45-100 оценивали на способность к образованию капсулярного полисахарида путем иммуноблоттинга колоний и латекс-агглютинации. Антисыворотка, содержащая антитела к капсулярному полисахариду A.pleuropneumoniae серотипа 5а, но не к другим компонентам бактериальной поверхности реагировала с J45, но не реагировала с J45-100 (фиг.6). Поскольку бактериальные колонии на нитроцеллюлозном фильтре лизировали хлороформом, эти результаты свидетельствуют о том, что J45-100 не продуцирует внутриклеточного или внеклеточного капсулярного полисахарида. Целые или разрушенные ультразвуком клетки J45-100 не агглютинировали латексных частиц, ковалентно конъюгированных с очищенными антителами к капсулярному полисахариду A. pleuropneumoniae серотипа 5а (Inzana, 1995), тогда как целые и разрушенные ультразвуком клетки J45 эффективно агглютинировали латексный реагент (данные не приведены). Эти результаты подтверждают тот факт, что делеция, произведенная в локусе cap A.pleuropneumoniae J45-100, приводит к утрате биосинтеза капсулярного полисахарида. Кроме того, эти результаты свидетельствуют о том, что некапсулированный мутант, J45-C, полученный путем метансульфонатного мутагенеза (Inzana et al., 1993a), продуцирует внутриклеточный, но не внеклеточный капсулярный полисахарид.
Сравнивали экспрессию токсина Арх и электрофоретические профили LPS J45 и J45-100 для того, чтобы определить, влияет ли на эти важные показатели вирулентности мутация, введенная в локус cap J45-100. Не было обнаружено различий между J45 и J45-100 в секреции 105 кДа токсинных белков ApxI и ApxII в культуральный супернатант (фиг.7). Кроме того, не обнаружено разницы в электрофоретических профилях LPS J45 и J45-100 (фиг.8).
Сравнивали ростовые характеристики J45 и J45-100 в TSY-N и чувствительность LPS J45 и J45-100 к бактерицидной активности преколостриальной телячьей сыворотки, чтобы определить воздействие утраты инкапсуляции на эти фенотипические показатели. Кривые роста J45 и J45-100 в TSY-N были сходы, но не идентичны (данные не приведены). Однако подсчет жизнеспособных колоний на чашках показал, что во время логарифмической фазы роста J45-100 рос быстрее (время генерации около 23 минут), чем исходный капсулированный штамм (время генерации около 28 минут) (данные не приведены). Присутствие от 10 до 50% преколостриальной телячьей сыворотки в качестве источника комплемента вызывало эффективную гибель рекомбинантного некапсулированного мутанта J45-100 в течение 60 минут, тогда как исходный капсулированный штамм, J45, оставался жизнеспособным (фиг.9).
Исследовали чувcтвительность J45-100 к сахарозе с тем, чтобы определить, могут ли последовательности sacRB функционировать как противоселективный маркер в A.pleuropneunoniae и впоследствии индуцировать эксцизию кассеты nptl-sacRB из хромосомы J45-100. Выращенные в жидкой среде клетки J45-100 с трудом росли при прямом высеве на чашки или после разведения с последующим высевом на чашки со средой TSY-N или средой Лурии-Бертани (с добавлением 5 мкг/мл NAD), содержащей 5 или 8% сахарозы. Присутствие последовательностей sacRB в хромосоме J45-100 было подтверждено блоттингом по Саузерну. Эти результаты свидетельствуют о том, что или маркер sacRB не экспрессируется в A.pleuropneumoniae, или продукт леван, образуемый sacRB левансахаразой в присутствии сахарозы, не является токсичным для J45-100.
Интратрахеальное заражение свиней рекомбинантным некапсулированным мутантом A.pleuropneumoniae, J45-100
Рекомбинантный некапсулированный мутант J45-100 не вызывал смертности у поросят при введении в дозах, в 3-6 раз превышающих (1.45×107 CFU и 2.95×10 CFU соответственно) 50%-ную летальную дозу (LD50) исходного капсулированного штамма J45 (5×106 CFU) (Inzana et al., 1993a) (Таблица 2). Наоборот, у всех трех поросят, получивших 6.5-кратную LD50 штамма J45, развились острые поражения легких и животные погибли (Таблица 2).

У пяти поросят, зараженных низкой дозой J45-100 (1.45×107 CFU), не проявилось каких-либо клинических симптомов, характерных для плевропневмонии свиней, и у них не развилось каких-либо поражений легких. Более того, из образцов ткани легких, взятых при вскрытии через четыре для после заражений, не удалось выделить в культуре A.pleuropneumoniae.
Двое из пяти поросят, зараженных высокой дозой J45-100 (2.95×107 CFU), оставались клинически нормальными, и при вскрытии у них не обнаружено поражений легких. У одного поросенка из этой группы, получившей высокую дозу J45-100, была отмечена умеренная одышка, а вскрытие показало некоторое уплотнение легких и слабые геморрагии (оценка поражения легких 1+). У двух остальных поросят из этой группы была отмечена легкая одышка, а вскрытие обнаружило признаки плеврита и уплотнения (оценка поражения легких 2+). A.pleuropneumoniae J45-100 был выделен в культуре только от двух поросят с наиболее острыми поражениями легких. Выделенные от этих поросят бактерии не агглютинировали латексные частицы, специфичные к серотипу 5а. Таким образом, выделенные бактерии оставались некапсулированными, что свидетельствует о том, что J45-100 не ревертирует к капсулированному фенотипу in vivo.
Были получены следующие данные по иммунизации свиней путем введения штамма J45-100 (рекомбинантный некапсулированный мутант). Шести свиньям дважды вводили стандартную дозу J45-100. Через 4 недели после первой прививки у всех свиней наблюдалось более чем четырехкратное усиление реакции на внеклеточные токсины Ар. Такой рост титра коррелируется с активностью защиты от введенного вещества. При интратрахеальном введении свиньям большой дозы гетерогенного штамма (серотип 2) все свиньи оставались живы через 24 часа после введения, кроме одной, которую пришлось усыпить из-за болей. Штаммы серотипа 2 выделяют токсины (Арх.III), которые не вырабатываются штаммом J45-100. Данные по состоянию легких колебались от 0% патологии до 55% - уплотнение и воспаление легочной массы (0%, 0%, 3%, 20%, 45% и 55%).
При том, что гены nptl (устойчивость к канамицину) и SacB/SacR (чувствительность к сахарозе) были клонированы в сайт делеции, данные гены использовались только как маркерные. Также могут быть использованы альтернативные маркерные гены. Может оказаться предпочтительным отказ от использования маркеров устойчивости к антибиотикам, таких как nptl, по соображениям охраны здоровья и безопасности или же из стремления использовать механизм модификации или инактивации антибиотикового маркера. К приемлемым “неантибиотиковым” маркерам может относиться устойчивость к ртути.
Полученный согласно настоящему изобретению штамм Actinobacillus pleuropneumoniae серотипа 5а продуцирует только два из трех токсинов Actinobacillus pleuropneumoniae. При том, что модифицированный штамм Actinobacillus pleuropneumoniae является протективным и иммуногенным, может оказаться полезным клонирование в сайт делеции третьего гена токсина RTX. Это можно осуществить, клонируя ген токсина RTX в кассету гена устойчивости к канамицину штамма J45-100, инактивируя тем самым ген устойчивости к канамицину.
Вакцина предпочтительно должна быть приготовлена в форме, аналогичной для других известных из уровня техники вакцин.
Предпочтительно, чтобы вакцина была расфасована в виде лиофилизированной смеси и могла включать один или несколько серотипов мутантных штаммов. Для сохранения жизнеспособности могут быть добавлены такие вещества, как бульон Колумбия, трегалоза, альбумин, глицерин или другие агенты. Лиофилизированную смесь остается только растворить в стерильной воде или буфере и ввести животному (внутримышечно, внутривенно, внутрибрюшинно, подкожно и т.д.). Может быть также приготовлена вакцина для других способов введения (например, орального, трансдермального, подъязычного и т.д) с использованием соответствующих смесей наполнителей (например, крахмалов, полисахаридов, масел, липосом, смол и т.д.).
Доза вакцины, вводимая животному, будет определяться такими факторами, как возраст и пол животного, а также способ введения. Во всех случаях достаточное количество живого, авирулентного, некапсулированного Actinobacillus pleuropneumoniae будет обеспечивать развитие иммунного ответа у вакцинированного животного. Успешные результаты были получены при двукратной иммунизации с интервалом 2-3 недели дозой 109 колониеобразующих единиц.
При том, что в настоящем описании приведены примеры предпочтительных вариантов осуществления изобретения, квалифицированному специалисту очевидно, что могут быть разработаны модификации изобретения, не выходящие за пределы его объема и охватываемые приведенной ниже формулой.




Изобретение относится к биотехнологии и касается вакцины, применяемой в ветеринарной практике. Вакцина против плевропневмонии свиней содержит генетически модифицированную некапсулированную бактерию Actinobacillus pleuropneumoniae, у которой удалены последовательности ДНК, кодирующие синтез капсулы. Описан также способ получения указанной вакцины. Способ получения вакцины включает идентификацию последовательностей ДНК капсульного синтеза кодирующих синтез капсулы Actinobacillus pleuropneumoniae и последующее делегирование указанных последовательностей ДНК капсульного синтеза с получением при этом некапсулированных мутантов этого организма. Полученная рекомбинантная вакцина является безопасной и эффективной, живой, аттенуированной. 2 с. и 2 з.п.ф-лы, 2 табл., 11 ил.
| US 5429818 А, 04.07.1995 | |||
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| БАКТЕРИЦИДНАЯ КОМПОЗИЦИЯ ДЛЯ КОНСЕРВИРОВАНИЯ ПИЩЕВЫХ ПРОДУКТОВ | 1990 |
|
RU2048151C1 |
| US 5441736 А, 15.08.1995. | |||

