Изобретение касается области генетической инженерии и биотехнологии. Более точно, изобретение касается нуклеотидной последовательности, полученной от патогенной бактерии Neisseria meningitidis нуклеотидная последовательность которой кодирует белок внешней мембраны указанной бактерии. Указанный белок клонируется и экспрессируется в хозяине - E. coli. Характеристики этого белка, а также его способность индуцировать иммунологически активные антитела (бактерицидные антитела) в своем естественном хозяине, позволяет использовать его для приготовления вакцин против патогенных штаммов указанного микроорганизма.
Грам-отрицательная бактерия N. meningitidis, по мировой статистике, вызывает каждый третий случай бактериального менингита. Она была впервые описана Антоном Вайксельбаумом в 1887 году (см. I. Devoe, 1982, Microbiol. Revs. 46: 162 - 190), и до сих пор единственным естественным хозяином этой бактерии остается человек.
В отношении метаболизма и серологической дифференциации этого микроорганизма в первой половине XX века были обнаружены некоторые существенные аспекты. Первые безуспешные попытки получения вакцинных препаратов основывались на капсулярном полисахариде. (E. Kabat и др. 1945, J. Exp. Med. 80: 299 - 307). Согласно химическому составу этого капсулярного полисахарида бактерия N. meningitidis серогруппирована на A, B, C, 29-E, H, I, K, J, W-135, X, Y или Z, и основной процент заболеваний вызывается группами A, C, Y, W-135 и B. Неинкапсулированные штаммы не связывают с этим заразным заболеванием.
С использованием различных способов очистки этих полисахаридов (E. Cotschlich и др. 1969, J. Exp. Med. 129: 1349 - 1365) было показано, что первые четыре полисахарида (ПС) являются хорошими иммуногенами и индукторами бактерицидных антител у человека (там же, 1367 - 1384). Присутствие этого вида антител ранее связывали с невосприимчивостью к инфекции (там же, 1307 - 1326). В настоящее время для серотипов A, C и W-135 хорошо изучены моно-, би- и тетравелентные вакцины (F. Ambrosch и др. 1983, Bulletin of the Who 61: 317 - 323; I. Vodopija и др. 1983, Infect. Immunol. 42: 599 - 604; M. Cadoz и др., 1985, Vaccine 3: 340 - 342; H. Peltola и др., 1985, Peliatrios 76: 91 - 96).
Эти вакцины разрешены для их применения на людях в различных странах (Centers for Desease Control, 1985, Morbid. Mortal. Weeklyheport 34: 255 - 259) и некоторые из них имеются в продаже от различных фирм и производителей (Connaught Laboratories, США; Smith Kline-RI, T Бельгия, Institute Merieux, Франция; Behringwerke Aktiengesellscchaft, Германия; Instituto Sieroterapico e vaccino genea Toscano "Sclavo", Италия, Swiss Serum and Vaccine Institute, Berne, Швейцария, и другие).
Однако традиционная вакцина против N. meningitidis серогруппы C не вызывает достаточных уровней бактерицидных антител у детей младше двух лет, которые являются основными жертвами этой болезни. Было продемонстрировано, что титр специфических антител против N. meningitidis у детей младше 4 лет после трех лет вакцинации сходен у вакцинированных и невакцинированных детей (H. Kayhty и др., 1980, J. of Infect. Dis. 142: 861 - 868). Так никакого иммунитета не было обнаружено против N. meningitidis после 8 лет вакцинации у молодых людей (N. Rautonen и др., 1986, J. of Immund, 137: 2670 - 2675).
Полисахарид, соответствующий N. meningitidis серогруппы B, является слабо иммуногенным (E. Cotschlich и др., 1969, J. Exp. Med. 129; 1349 - 1365) и вызывает слабую реакцию 1 М низкой специфичности (W. Zdlinger и др., 1979, J. Clin. Inoest. 63: 836 - 848). Имеется несколько теорий, касающихся этой проблемы, такие как перекрестная реактивность между B-полисахаридом и структурами мозга плода, антигенными структурами, модифицированными в растворе и чувствительностью к нейроаминидазе (C. Moreno и др., 1985, Infect. Immunol. 47: 527 - 533). Недавно была получена химическая модификация ПС "B", которая вызывала реакцию у хозяина (H. Jennings и др. 1988, пат. США N 4.727.136; F. Ashton и др. , 1989, Microb. Pathogen. 6: 455 - 458), однако безопасность этой вакцины для людей не была продемонстрирована.
Из-за отсутствия эффективной вакцины против N. meningitidis "B", и вследствие того, что опасность эндемии низка и ограничена, в основном, детьми, обязательная иммунизация полисахаридами не рекомендуется (C. Frash, 1989, Clin. Microbiol. Revs. 2: 134 - 138), за исключением эпидемических случаев.
Поскольку после Второй Мировой войны эта болезнь в большинстве случаев вызывалась N. meningitidis типа B, то вакцины против серогруппы B приобрели особое значение.
Компоненты внешней мембраны N. meningitidis включают фосфолипиды, липополисахариды (ЛПС или эндотоксины), пили-протеины и др. Различные иммунотипы ЛПС были описаны для N. meningitidis (W. Zollinger and R. Mandrell, 1977, Infect. Immun. 18: 424 - 433; C.M. Tsai и др., 1983 J. Bacteriol, 155: 498 - 504) и была исследована иммуногенность с использованием нетоксичных производных (H. Jennings и др., 1984, Infest. Immund. 43: 407 - 412), однако, их вариабельность (H. Schneider и др., 1984, Inf. Immunol. 45: 544 - 549) и пирогенность (когда ЛПС присоединен к липиду A) являются ограничивающими факторами до сих пор.
Пили-структуры, необходимые клеткам для их фиксации к слизистой оболочке носоглотки (D. Stephens и др., 1983, J. Infect. Dis., 148: 369 - 376), имеют антигенные различия у разных штаммов (J.Greenblatt и др., 1988, Infect. Immun. 56: 2356 - 2362) наряду с несколькими общими эпитопами (D. Stephens и др., 1988, The J. Infect Dis. 158: 332 - 343). В настоящее время имеются некоторые сомнения в отношении эффективности вакцины, основанной на этих структурах. Однако, было получено все же несколько таких типов вакцин с неизвестными результатами в отношении их применения на людях (C. Brinton, 1988, патент США N 4.769.240).
Недавно внимание переключилось на другие белки внешней мембраны этой бактерии. У этих белковых комплексов имеется множество иммунологических типов.
Штаммы N. meningitidis подразделяются на серотипы согласно присутствию специфических эпитопов в мажоритарном белке P1/P2 и на подтипы согласно другим эпитопам в белке P1 (C. Frasch и др., 1985, Rev. Infect. Dis. 7: 504-510).
Опубликовано несколько статей и патентных заявок, касающихся вакцин, основанных на "коктейле" из этих белков, с предварительным селективным удалением эндотоксинов с использованием биосовместимых детергентов. Иммуногенность этих коктейлей у животных и людей продемонстрировали следующие авторы: (W. Zollinger и др., 1979, J. Clin. Invest. 63: 836-848; C. Frasch и M. Peppler, 1982, Infect. Immun. 37: 271-280; E. Benverg и др., 1983, Infect. Immun. 40: 369-380, E. Kosenquist и др., 1983, NIPH Annals 6: 139-149; L. Wanq и C. Frasch, 1984, Infect. Immun. 46: 408-414; C. Moreno и др., 1985, Infect. Immun. 47: 527-533; E. Wedege и Froholm, 1986, Infect. Immun. 51: 571-578; C. Frasch и др., 1988, The J. Infect. Dis. 158: 710-718; M. Lifely и Z. Wang, 1988, Infect. Immun. 56: 3221-3227; J. Poolman и др., 1988, - в кн. J. Poolman и др. (изд.) "Gonococci and Meningococci" Kluwer Acad. Publ., Dordrecht, the Netherlands, стр. 159-165; E. Rosenquist и др., 1988, J. Clin. Microbiol. 26: 1543-1548), включая результаты массовых полевых исследований в Кейптауне, ЮАР, в 1981 (C. Frasch, 1985, Eur. J. Clin. Microbiol. 4: 533-536); в Икике, Чили, в 1987 г. (W. Zollinger, 1988, Proceedings of the Sixth International Pathogenic Neisseria Conference. Callaway Gardens Conference Center), и на Кубе в 1986 и 1988 годах (G. Sierra, 1988, Proceedings of the Sixth International Pathogenic Neisseria Conference. Callaway Gardens Conference Center). Однако, за исключением последнего случая, бактерицидные антитела, индуцируемые этими препаратами, были ограничены антителами к тем же самым (по серотипу) штаммам или родственным им.
На одну из таких вакцин указывается в патенте США N 4.601.903, которая ограничивается одним из типов N. meningitidis, вызывающих менингит (серотип 2), с широким диапазоном действия, однако у пациентов часто выделяли и другие серотипы, такие как серотипы 4 (Куба, от 1981 до 1983, H. Abdillahi и др. , 1988, Eur. J. Clin. Microbiol. Infect. Dis. 7: 293-296, Финляндия, от 1976 по 1987, H. Kayhty и др., 1989, Scand. J. Infect. Dis. 21: 527-535), 8 (Австралия от 1971 по 1980, F. Ashton и др., 1984, Can. J. Med. Biol. 30: 1289-1291) и 15 (Норвегия от 1982 по 1984, L. Froholm и др., 1985, Proceedings of the 4-th Int. Symp. on Path. Neisseria American Society for Microbiology; Чили с 1985 по 1987, S. Ruiz и др., 1988, Proceedings of the 6th Int. Path. Neisseria Conf. Callaway Gardens Conference Center), а также штаммы неидентифицированного серотипа (F. Ashton и др., 1980, Can. J. Microbiol. 26: 1480-1488, Австралия с 1971 по 1980, F. Ashton и др., 1984, Can. J. Med. Biol. 30: 1289-1291; Финляндия с 1976 по 1987, H. Kayhty и др., 1989, Scan. J. Infect. Dis. 21: 527-535).
Кубинская вакцина, полученная в 1988 году в "Сентро Насиональ де Биопрепарадос" (см. заявку EP N 301 992), оказалась весьма эффективной. Она основана на антигенном комплексе высокого молекулярного веса. Эта вакцина обладает широким диапазоном перекрестной реактивности с другими штаммами, продуцирует и поддерживает бактерицидные антитела в иммунизированном хозяине.
Однако методики получения этого типа вакцины начинаются с размножения в подходящей среде высокопатогенного микроорганизма, со связанным с этим биологическим риском непосредственной работы с бактерией. Кроме того, этот вид препаратов содержит липосахариды, являющиеся загрязнителями, которые, хотя и могут повысить эффективность вакцины, в то же время создают нежелательные побочные эффекты ввиду их сильной пирогенности. Также в разных партиях препарата нельзя контролировать изменения содержания минорных антигенных компонентов, которые образуют часть препарата, что затрудняет соблюдение важных параметров, касающихся реактогенности и иммуногенности.
По этой причине постоянно увеличивается интерес к идентификации нуклеотидных последовательностей, кодирующих высококонсервированные белки во всех штаммах, и еще более - к идентификации белков, индуцирующих бактерицидные антитела, которые являются общими для большинства патогенных N.meningitidis, чтобы получить вакцинные препараты с широким спектром защиты.
Имеется несколько белков высокого молекулярного веса, присутствующих в небольших количествах во внешней мембране N.meningitidis, когда этот микроорганизм выращивают в традиционной культурной среде, но дает сильную реакцию у инфицированных особей (J. Black и др., 1986, Infect. Immun. 54: 710-713; L. Aoun и др., 1988, Ann. Inst. Pasteur/Microbiol. 139: 203-212) и/или усиливают их реакцию при особых условиях культивирования (J. Van Putten и др., 1987, Antoine van Leenwenhock 53: 5557-5564; A.Schryvers и L.Morris, 1988. Mol. Microbiol., 2: 281-288, и Inf. Immun. 56: 1144-1149). Некоторые из этих белков являются высококонсервированными у различных штаммов, в частности у тех, которые отвечают за усвоение железа микроорганизмами, что делает их интересными кандидатами на приготовление вакцины (L.Mocca и др., 1988, Proceedings of the 6th Int. Path. Neisseria Conference Callaway Gardens Conference Center; C.Frash, 1989, Clin. Microbiol. Revs. 2: 134-138).
В дополнение к чистым белкам, полученным из микроорганизмов или штаммов соответствующих видов (например, белок размером 36 кДа, Т. Митцнер и С. Морзе, патент США N 4.681.761), было клонировано и экспрессировано несколько родственных генов. Среди этих белков есть следующие:
- протеаза IgAI (J.Koomey и S.Falkow, 1984, Inf. Imm. 43: 101-107);
- протеин P1 (A. Barlow и др., 1987, Inf. Imm. 55: 2734-2740, и 1989, Mol. Microbiol., 3: 131-139);
- протеин P5a (T.Kawula и др., 1988, Inf. Imm. 56: 380-386);
- протеин P5c (T.Olyhoek и M.Achtman, 1988, Proceedings of the 6th Int. Path. N. Conference Callaway Gardens Conf. Cent.);
- протеин P4 (K.Klugman и др., 1989, Inf. Imm. 57: 2066-2071);
- протеин P2 (K.Murakami и др., 1989, Inf. Imm. 57: 2318-2323) и от N. gonorrhoeae, которая кодирует белки с перекрестной реактивностью с их соответствующими белками от N.meningitidis;
- антиген H.8 (W.Black и J.G.Cannon, 1985, Inf. Imm. 47: 322-325);
- макромолекулярный комплекс (W.Tsai и C.Wilde, 1988, Proceedings oh the 6th Int. Path. Neisseria Conference. Callaway Gardens Conference Center);
- белок размером 37 кДа, репрессируемый в присутствии железа (S.Berish и др. , 1988, Proceedings of the 6th Int. Path. Neisseria Conference. Callaway Gardens Conference Center).
Использование этих белков в качестве активных вакцинирующих препаратов не сообщалось ранее, или же испытания на индуцируемые ими бактерицидные антитела дали отрицательный результат, как в случае мышиных моноклональных антител против H.8 (J.Woods и др., 1987, Infect. Immun., 55: 1927-1928).
До сего момента белок P1, находящийся во внешней мембране N.meningitidis, является одним из наиболее хорошо охарактеризованных и изученных антигенов. Этот белок не представляет варианбельности внутри одного штамма. Однако имеется более 17 типов белков P1 у Neisseria, которые имеют отличия в трех различных областях, что и явилось основанием для классифицирования Neisseria на разные подтипы. Этот белок является очень иммуногенным у людей (W. D. Zollinger и R.E.Mandrell, 1983, Med. Trop., 43: 143-147), вызывающим защитные антитела (E.Wedege и L.O.Froholm, 1986, Infect. Immun. 51: 571-578; K. Saukkonen и др., 1987, Micr. Path., 3, 261-267), что придает ему особое значение для приготовления вакцин.
Некоторые подтипы белков P1 клонировали в E.coli, исходя из геномных библиотек (А. К.Барлов и др., 1989, Мол. Микроб. 3: 131-139), или с использованием цепной реакции полимеразы (С. Бутчер, Седьмая Междунар. Конференц. по Нейсерии; Р. Зайд, патентная заявка WO 90/06696; Брайн Мак-Гиннесс и др., 1990, Ж. Эксп. Мед. 171: 1871-1882, М.С.Ж. Мейден и др., 7-я Межд. Конф. по Нейсерии, Берлин, 9-14 сент. 1990 и 1991, Молек. Микр. 3: 727, Дж. Сукер и др. , 7-я Межд. Конф. по Нейсерии, Берлин, 9-14 сент. 1990). Однако, до сих пор не имеется генетической конструкции, способной продуцировать этот белок с высокими уровнями экспрессии. Сообщалось лишь о низких уровнях экспрессии (Д. А. Вайт и др., 1990, Мол. Микроб. 4: 769-776) или о его экспрессии в Bacillus subtilis, лигированным с белком A внешней мембраны E.coli (omp A) (Е. Ваальштром и др., 7-я Межд. Конф. по Нейсерии, Берлин).
Можно подтвердить, что до сих пор не было выделено антигена, который был бы общим для всех типов и серогрупп N.meningitidis и был способен продуцировать бактерицидные антитела. По этой причине антиген такого типа, конъюгированный или лигированный к другим белкам или полисахаридам, представляющим иммунологический интерес, представлял бы собой кандидата для бивалентного вакцинного препарата.
Изобретение касается нуклеотидной последовательности, кодирующей белок, имеющий молекулярный вес около 64 кДа. Эта последовательность была обнаружена у всех серотипов N.meningitidis и всех исследованных серогруппах, что было проверено нуклеиновокислотной гибридизацией. Вестерн-блоттингом, дот-блоттингом и твердофазным иммуноферментным анализом (ELISA).
Технической задачей изобретения является идентификация нуклеотидной последовательности, которая кодирует высококонсервированный белок и является общей для большинства патогенных штаммов Neisseria (название P64к), с целью получения рекомбинантным методом белка, который имел бы высокую степень чистоты в коммерчески полезных количествах таким образом, чтобы его можно было применять в диагностических способах и как составную часть вакционного препарата широкого спектра защиты.
На уровне генетической информации (ДНК и РНК) изобретение обеспечивает рекомбинантный полинуклеотид, включающий нуклеотидную последовательность, кодирующую протеин P64к Neisseria meningitidis, причем указанный протеин P64к в основном имеет аминокислотную последовательность, показанную в SEQ ID NO: 1. В предпочтительном выполнении указанная нуклеотидная последовательность, кодирующая протеин P64к N.meningitidis, в основном состоит из нуклеотидной последовательности, показанной в SEQ ID NO:1. Далее рекомбинантный полинуклеотид может включать нуклеотидную последовательность клонирующего или экспрессионного вектора.
Изобретение также обеспечивает трансформированный микроорганизм, содержащий рекомбинантный полинуклеотид, как он определен выше, предпочтительно трансформированный микроорганизм, который способен экспрессировать протеин P64к N.meningitidis. В особенно предпочтительном выполнении изобретения трансформированным микроорганизмом является штамм E.coli, например штамм E.coli HB101, трансформированный экспрессионным вектором, содержащим нуклеотидную последовательность, кодирующую протеин P64к N.meningitidis, например экспрессионный вектор pM-6.
Изобретение также обеспечивает рекомбинантное белковое вещество, включающее аминокислотную последовательность, соответствующую аминокислотной последовательности по меньшей мере части протеина P64к N.meningitidis, причем указанный протеин P64к в основном имеет аминокислотную последовательность, показанную в SEQ ID NO:1. Указанное белковое вещество может в основном состоять из протеина P64к, или быть легированным протеином или белково-полисахаридным конъюгатом, включающим аминокислотную последовательность протеина P64к N.meningitidis.
Изобретение далее обеспечивает вакцинирующую композицию, включающую рекомбинантный протеин, как он определен выше, вместе с подходящим носителем, разбавителем или адъювантом. Частное выполнение изобретения обеспечивает вакцинную композицию, включающую липоамид-дегидрогеназу или ацетил-трансферазу, которая способна индуцировать антитела, которые могут связывать протеин P64к N. meningitidis вместе с подходящим носителем, разбавителем или адъювантом.
Дополнительно изобретение обеспечивает моноклональное антитело, созданное против рекомбинантного белкового вещества, как оно определено выше, или против липоамид-дегидрогеназы или ацетил-трансферазы, и способное связывать протеин P64к N.meningitidis.
Изобретение также обеспечивает способ получения протеина P64к, имеющего в основном аминокислотную последовательность, показанную в SEQ ID NO:1, который предусматривает стадии трансформирования микроорганизма экспрессионный вектором, содержащим нуклеотидную последовательность, кодирующую указанный протеин P64к, или указанный легированный протеин, культивирование трансформированного микроорганизма для получения экспрессии указанного протеина P64к, или указанного лигированного протеина, и выделение указанного продукта экспрессии.
Новым аспектом настоящего изобретения является ген, выделенный из N. meningitidis штамма B:4:P1.15, который назван M-6 и имеет в качестве основной своей характеристики стабильность в векторах E.coli. Этот ген не оказывает нежелательного воздействия на хозяина, позволяя получать выходы свыше 25% от общего белка (соотношение P64к от общего белка штамма-хозяина). С другой стороны, гибридизацией по Саузерну и Вестерн-болту (Е.Саузерн, 1975, J. Mol.Biol. 98: 503-527 и В.Брюнет, 1981, Анал. Биохем., 12: 195-203), было продемонстрировано, что протеин P64к присутствует у всех следующих штаммов N.meningitidis:
N.Meningitidis A N.Meningitidis B:9
N.Meningitidis B:1 N.Meningitidis B:11
N.Meningitidis B:2 N.Meningitidis B:15
N.Meningitidis B:4 N.Meningitidis B:4:P1.15
N.Meningitidis B:5 N.Meningitidis C
N.Meningitidis B:8 N.Meningitidis B:15:P1.16
N.Meningitidis N.Meningitidis B:15:P1.16(H44/76)
N.Meningitidis B: T(121/85)
N.Meningitidis B: T(71/86)
N.Meningitidis B: T(210/86), а также у
N.Mucsa, N.subflave и N.gonorrhoeae.
Этот белок не присутствует у N.cinerea, N.lactamica, N.sicca и N.flavescens, однако они не представляют интереса ввиду их непатогенности.
Этот протеин, имеющий молекулярный вес около 64 кДа, может быть локализован с помощью электронного микроскопа на внешней мембране N.meningitidis. Поэтому он является экспонированным антигеном, благоприятным для его использования в приготовлении вакцины. Этот протеин распознавался экспериментами по Вестерн-блот-иммуноидентификации в сыворотке выздоравливающих и вакцинированных традиционной кубинской вакциной "Ва-Менгок-БЦ" (Сентро Насиональ де Биопрепарадос, Гавана, Куба). Этот аспект гарантирует иммуногенность антигена и в то же время подтверждает его присутствие в белковой фракции высокого молекулярного веса, содержащейся в этой вакцине, которая отвечает за длительную иммунную реакцию на заболевание.
Другим новым аспектом изобретения является то, что этот протеин, являющийся объектом изобретения, продуцирует антитела с широким бактерицидным спектром (различные серогруппы, серотипы и подтипы), т.е. имеет характеристику, которая ранее не сообщалась ни для одного белка от N.meningitidis.
Высокие уровни этого протеина, полученные у E.coli, делают его важным кандидатом для усовершенствования иммуногенности, когда он экспрессируется как протеин, лигированный с другими протеинами. Также можно усилить экспрессию посредством передачи повышенной стабильности и совместимости в молекулярной структуре во время процессов транскрипции и трансляции. Будучи принадлежащим Neisseria, этот протеин может быть также легирован с другими протеинами Neisseria с целью получения вакцинных препаратов против этого микроорганизма с повышенной иммунногенностью. Эти лигированные протеины также являются объектами настоящего изобретения.
С другой стороны было неожиданного обнаружено, что ген M-6, полученный из геномной библиотекам штамма N.meningitidis B:4:P1.15, продемонстрировал значительную гомологию с последовательностями липоамид-дигидрогеназ и ацетил-трансфераз других микроорганизмов и высших организмов. Присутствие общих антигенных детерминантов позволяет использовать эти другие родственные белки в качестве иммуногенов, способных сообщить защиту посредством индуцирования бактерицидных антител, которые распознают антигенные детерминанты, общие с протеином Р64к. Поэтому в объем настоящего изобретения также включается использование этих липоамид-дегидрогеназ и ацетил-трансфераз (выделенных не от N. meningitidis) или их производных, таких как пептиды, фрагменты ферментной деградации конструкции, полученные легированием с другими белками или конъюгацией с протеинами, полисахаридами или липидами, или посредством инсерции в такие комплексы, как липосомы или везикулы и т.д., для целей вакцинирования.
Важным объектом настоящего изобретения является нуклеотидная последовательность, кодирующая ген М-6 (SEQ IDNO:1 в списке последовательностей), продуктом которой является протеин Р64к.
Этот ген был извлечен из генома штамма B385, выделенного на Кубе (N.meningitidis B: 4: P1.15), с помощью конструкции из геномной библиотеки в фаге EMBI 3.
Рекомбинантная ДНК, включающая ген М-6, представляет собой другой объект изобретения, который включает фаг лямбда, плазмиду pM-3 и экспрессионный вектор для экспрессии в бактериях.
В частности, для внутриклеточной экспрессии E.coli M-6 ген клонировали под триптофановым промотором с использованием его собственного сигнала терминации транскрипции и соединительного фрагмента между М-6 и клонирующим E. coli, который добавляет следующую нуклеотидную последовательность к концу 5: ATG CTAGAT AAA AGA (SEQ IDNO2).
N-терминал протеина Р64к, кодированного геном М-6, вставленным в плазмиду pM-6, который добавляет 5 аминокислот к N-терминалу исходного протеина.
Другим объектом настоящего изобретения являются микроорганизмы, полученные посредством трансформирования штамма E.coli HB 101 вектором pM-6, которые характеризуются высокими уровнями экспрессии протеина Р64к, хорошей жизнеспособностью и высокой стабильностью.
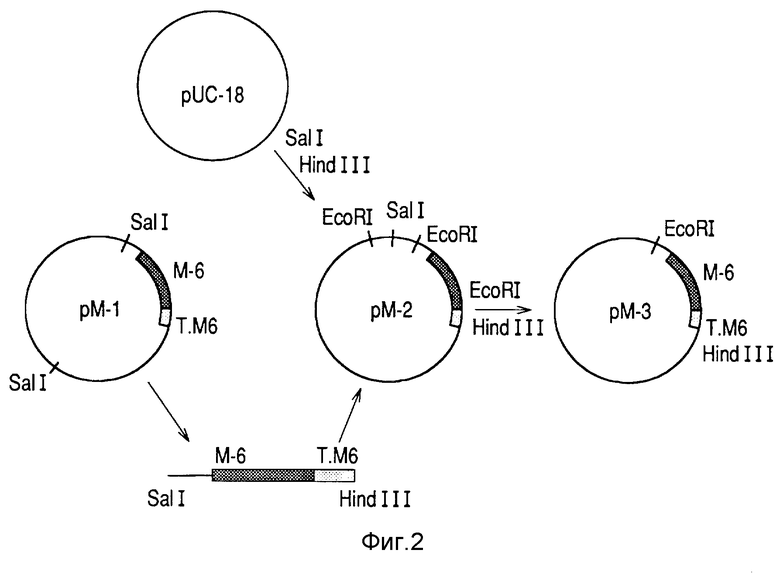
Трансформированный клон E. coli был обозначен HBM64 (см. фиг. 2), и представляет более высокие уровни экспрессии Р64к (на 25% по отношению к общему белку клетки) (фиг. 6).
Процедура, описанная в настоящем изобретении, благодаря уровням экспрессии этого продукта, позволяет достичь оптимальной чистоты для использования этого протеина для людей.
С другой стороны, антиген, полученный из выделенной последовательности, может быть очень полезен при получении различных типов потенциальных вакцинных препаратов, таких как бивалентные вакцины с широким спектром действия, например протеин-полисахаридные конъюгаты, лигированные протеины и т.д.
Примеры
Далее приводятся примеры для лучшей иллюстрации изобретения, которые, однако, не ограничивают его объемы.
Пример 1.
Для выделения геномной ДНК из N.meningitidis B:4:P1.15 клетки выращивали в среде Мюллера-Хинтона (OXOID, Лондон). Биомассу из 100 мл культуры ресуспендировали в 8 мл Трис (гидроксиметил-аминометан) 100 мМ, ЭДТК 1 мМ, pH 8. Клетки обрабатывали лизоцимом (10 мг/мл), с последующим добавлением 200 мкл самоперевариваемой проназы (20 мг/мл) и 1.1 мл 10% додецилсульфата натрия. Смесь инкубировали при 37oC в течение часа, а затем ее обрабатывали фенол-хлоформом (об/об), и остатки фенола удаляли с использованием 2-бутанола. Наконец, ДНК осаждали абсолютным спиртом, и РНК удаляли рибонуклеазой A (Сигма, Лондон).
ДНК размером около 60 кб/тыс.оснований/ подвергали частичному перевариванию ферментом SAU3A, с получением популяции фрагментов размером примерно 15 кб. Эту мажоритарную фракцию выделили и очистили посредством разделения на агарозном геле (Т. Маниатис и др., 1982, Молекулярное клонирование: Лабораторное руководство. Коулд Спринг Харбор Лаб.: (Нью-Йорк). Для создания геномной библиотеки в основном следовали процессу, описанному Маниатисом (см. там же). 4 мкг очищенной ДНК лигировали к 8 мкг ВамН1-переваренной плазмиде EMBL-3. Продукт лигирования был упакован и фаги, наконец, разместили (посеяли) на штамме E.coli C66P2.
Эту библиотеку скринировали посредством иммуноидентификации - (Р.Янг и Р. Дэвис, 1983, PNAS США 80:1194-1198) с использованием сыворотки кролика, полученной препаратом протеинов, принадлежащих внешней мембране штамма N. meningitidis B: 4: P1.15. Клоны анализировали вестерн-блотингом (Бурнет, 1981), и обнаруживали экспрессию протеина Р64к с молекулярным весом около 70 кДа. Полученный рекомбинатный фаг был обозначен 31. Вестерн-блотинг проводили также с использованием смеси сыворотки от выздоравливающих от менингококковой инфекции, свободной от антител от E.coli, с получением того же результата, что и с использованием сыворотки гипериммунизированных кроликов.
Этот эксперимент был повторен с использованием сыворотки от нескольких здоровых людей, и полученный сигнал был отрицательным против рекомбинантного протеина Р64к.
Пример 2.
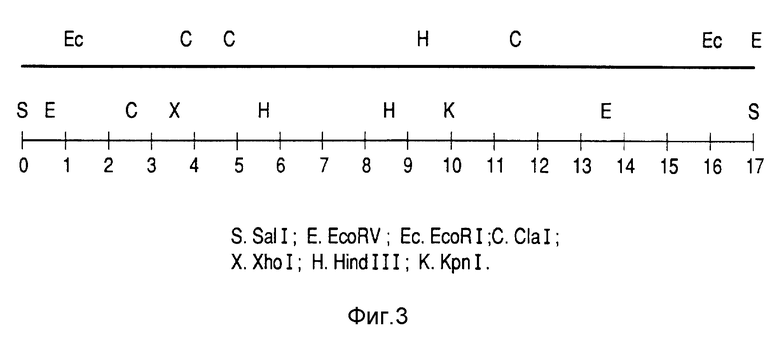
Для субклонирования в бактериях инсерт размером 17 кб, соответствующий выделенному из библиотеки фагу, клонировали в плазмиде pUC18 после отделения от плеч фага с использованием фермента SaI 1. Это привело к конструкции pM-1 (фиг. 2), которую подвергали рестрикционному анализу (фиг. 3).
Фрагмент SaI I-Hind III размером около 6 кб реклонировали в плазмиде pUC 18 и получили конструкцию pM-7 (фиг. 2). Чтобы получить более точное положение (локализацию) гена, кодирующего протеин P64к, осуществляли делеции с помощью ферментов Cla I, EcoR I и Hind II. Полный фрагмент гена M-6 был, наконец, локализован как EcoR I-Hind III инсерт, соответствующий конструкции pM-3 (фиг. 2)
Во всех конструкциях присутствие гена было подтверждено распознаванием этого протеина посредством иммуноидентификации колоний и вестерн-блоттингом с использованием сыворотки гипериммунизированных кроликов.
Последовательность инсерта в pM-3 была определена по способу Зангера (Ф. Зангер и др., 1977, PNAS США 74: 5463-5467).
Из полученной последовательности был получен дедукцией приблизительный молекулярный вес протеина, кодируемого этим геном.
С целью получения конструкции для высокой экспрессии протеина P64к плазмиду pM-3 (фиг. 2) выпрямили (линеаризовали) ферментом EcoR I и провели последовательные супрессии гена, инкубируя образцы с нуклеазами ExO III и SI.
Полученные фрагменты отделяли от остальных участков вектора pUC 18 посредством вырезания рестрикционным ферментом Hind III и клонировали лигированным к стабилизаторному фрагменту (EPA-0.416.673), с использованием Xbа-тупого адаптера для консервирования сайта Хbа I стабилизаторного гена:
5' C T A G A T A A A A G A 3' (SEQ ID NO:4)
3' T A T T T T T C T 5' (SEQ ID NO:5).
Конструкции, в которых лигированный фрагмент совпадал с рамкой считывания, отбирались посредством иммуноидентификации с использованием сыворотки гипериммунизированных кроликов.
Инсертные последовательности были установлены с использованием способа Зангера (Ф. Зангер и др., 1977, PNAS США 74: 5463-5467). Из полученных последовательностей был получен дедукцией средней молекулярный вес протеина, кодированного этим геном.
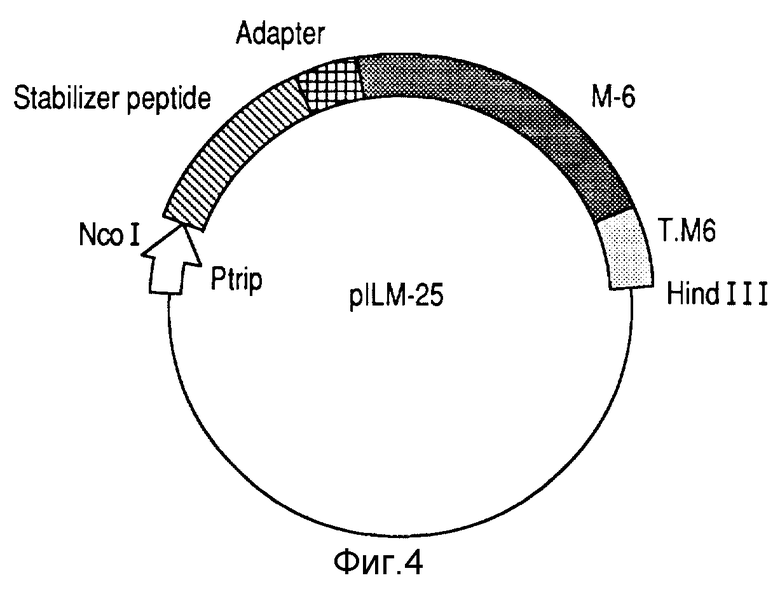
Область слияния между протеинами была локализована в генных последовательностях. В клоне pILM-25 (фиг. 4) АTГ гена, предопределенного последовательностью инсерта ДНК, выделенного из библиотеки, совпала с сайтом лигирования.
Фрагмент Nco I - Xba I, соответствующий стабилизирующей пептид-кодирующей последовательности, был удален делецией из pIIM-25 с получением нелигированного протеина, экспрессированного под триптофановым промотором с его природным терминатором из N.meningitidis B:4:PI.15 согласно конструкции pM-6 (фиг. 1).
Плазмиду pM-6 трансформировали в различных штаммах E.coli, таких как W 3110, JA-221, HB-101, LE-392 и MC-1061, и сравнивали экспрессию протеина p64к. Наилучшие результаты были получены со штаммами W3110, JA-221 и HB-101. Эти штаммы выбрали для масштабирования (оценки) ферментации, были получены уровни экспрессии до 25% общего клеточного белка.
Пример 3.
Для подтверждения правильной экспрессии клонированного гена H-терминальную область интактного протеина подвергали деградационному способу Эдмана (П. Эдман, 1950, "Акта Хем. Сканд., 4: 283-293). Эта методика позволила выяснить последовательность (первичную структуру) этой области молекул.
Протеин P64к был обессолен посредством гель-фильтрационной хроматографии (РД-10, фармация), элюирован водой и исследован мониторингом при длине волны 280 нм. Белковую фракцию концентрировали до 0.5 нМ/л. Один мкл этого раствора фильтровали через ПВДФ (поливинилиден-дифторид) (Миллипор)фильтр, предварительно активированный метанолом.
Проводили деградацию Эдмана с использованием Автоматического Секвенатора Кнауера, модель 810, в соединении с системой HPLC (высокопроизводительная жидкостная хроматография) так, чтобы обнаружить фенил-тиогидантоиновые производные аминокислот (PTH-аминокислоты). Стандартной процедурой секвенирования была процедура согласно рекомендациям производителя указанного оборудования. Разделение PTH-аминокислот осуществляли на колонке для обращенно-фазовой хроматографии C-18 (5 мкм) 250 мм х 2 мм (Мерцк), элюировали ацетонитрилом с градиентом (Б-буфер) в ацетате натрия (А-буфер), приготовленный по рекомендациям изготовителя, с потоком 200 мкл/мин при 42oC. PTH-аминокислоты обнаруживали при 269 нм.
Обработку данных и регистрирование осуществляли в автоматическом интеграторе Шимадзу, модель CP-6a, с использованием программы обработки данных посредством вычитания двух последовательных хроматограмм, для облегчения оценки деградационных циклов Эдмана. Идентификация последовательности получена посредством хроматографической оценки соответствующего проанализированного цикла и подтверждена хроматографией, полученной вычитанием, позволяющим определять 25 остатков.
Пример 4.
Для того, чтобы продемонстрировать, что протеин P64к распознается сывороткой пациентов, вакцинированных препаратом Ва-Менгок-БЦ (Сентро Насиональ де Препарадос, Гавана, Куба) осуществляли вестерн-блоттинг, со смесью 12 сывороток от взрослых (иммунизированных двумя дозами кубинской вакцины), которые были разведены в растворе, содержащем обезжиренное молоко (Oxoid, Лондон).
Эксперимент включал:
рекомбинантный протеин P64к, очищенный из штамма E.coli HB-101, трансформированного плазмидной pM-6;
супернатанат разорванных ультразвуком клеток нетрансформированного штамма E.coli HB-101;
реакция обнаруживалась посредством конъюгата протеин A-коллоидное золото.
Было показано, что протеин P64к распознается пулом сывороток.
Пример 5.
Бактерицидное испытание против B385 (B:4:PI.15) было проведено согласно процедуре, описанной Ларриком и др. (Сканд. Ж. Иммунол., 32, 1990, 121-128) с модификациями. Для этой цели была создана смесь:
а) суспензии бактерий, культивировавшихся при особых условиях (1-5 единиц образования колоний/1);
б) сбалансированный солевой раствор Гея;
в) сыворотка (3-4 недельного) кролика в качестве источника комплемента;
г) пул сывороток от мышей, иммунизированных против протеина P64к в Алюминий-Гидроксидном Геле (AHG) и инактивировали при 56oC в течение 30 мин. Иммунизацию мышей проводили согласно иммунизационной схеме - 3 дозы по 20 г. Пропорции, использованные в вышеупомянутой смеси, были 1:2:1:1 в общем объеме 125 л. Смесь инкубировали при 37oC в течение 1 часа и высевали в свежий Агар Мюллера-Хинтона (Оксоид, Лондон) с добавлением 5% сыворотки теленка (КубаВет, Гавана). Подсчет выживших колоний проводили через 18 часов инкубирования чашек в атмосфере 5% CO2 при 37oC.
Бактерицидный титр рассматривали как максимальное сывороточное разбавление, необходимое для вызывания 50% ингибирования клеточного роста, по отношению к той же самой смеси без тестовой сыворотки. Было обнаружено, что разбавление сыворотки 1:20 все еще поддерживает бактерицидное воздействие. В качестве отрицательного контроля (небактерицидного при разбавлении 1:2) использовали пул сывороток от мышей, иммунизированных AHG, и пул сывороток от мышей, иммунизированных рекомбинантной вакциной против гепатита B (Куба). Бактерицидный эффект был специфичным по отношению анти-P64к антител.
Пример 6.
Осуществляли бактерицидный тест против различных штаммов N.meningitidis с использованием:
1. Аммониево-сульфатного преципитата супернатанта, собранного из культуры клеток гибридомы, которые секретировали моноклональные антитела, специфичные против P64к (анти-P64к)/ анализируемый образец.
2. Аммониево-сульфатного преципитата супернатанта, собранного из гибридомных клеток, секретирующих моноклональные антитела, специфичные против PI. 15 протеина, присутствующего в штамме N. meningitidis B385 (анти-PI. 15)/положительный контроль системы. Максимальные исследованные разведения составляли всегда 1:16. В таблице указаны максимальные разведения, которые дали бактерицидный эффект согласно примеру 5 (см. в конце текста).
Как видно, моноклональные анти-P64к антитела обладают значительными бактерицидными титрами против различных серогрупп (A, B и C), серотипов (4, 14, 13, 15 и NT) и подтипов (7, 15, 16 и NT) бактерий.
Пример 7.
Лигированные протеин M-14 (P64к и PI.15)
Чтобы получить генетическую конструкцию для высокой экспрессии, которая содержала бы вариабельные эпитопы протеина PI.15 (протеина внешней мембраны N. meningitidis B:4:PI.15), лигированного к протеину P64к, клонировали ген, кодирующий протеин PI.15, с использованием цепной реакции полимеразы (PCR). Следующую последовательность (область)(см. I в конце текста), содержащую вариабельные иммунодетерминанты протеина PI. 15 вставили в MIuI сайт гена M-6, кодирующего P64к, после получения его тупых концов с помощью фрагмента Кленова из ДНК-полимеразы I (см. последовательность I в конце описания). Сайты для лигирования гена PI.15 с М-6 приведены в конце текста.
Полученный лигированный протеин (M-14) был экспрессирован в E.coli с использованием плазмидного вектора под триптофановым промотором до уровней свыше 10% общего клеточного белка. Этот протеин распознавался моноклональными антителами и антителами против PI.15 и P64к при вестерн-блоттинге.
Пример 8.
Конъюгация полисахарид/P64к.
Протеин P64к был конъюгирован с полисахаридом от Haemophilus influenzae с использованием метода редуктивного аминирования. Полисахарид Haemophilus influenzae (Полирибозил-рибитол-фосфат, PPP) очищали холодным фенольным методом, описанным Фрашем, 1990 (в: "Бактериальные вакцины", 1990, Алан Р.Лисс, Инк. , стр. 123-145). Конечное загрязнение PRP протеинами или нуклеиновыми кислотами было менее 1%. Этот полисахарид был деградирован с использованием метода Париха (Parikh и др., 1974, в: "Methods in EnzymoI. 34B: 77-102) с периодатом натрия в PPP (соотношение по весу 1:5), растворенном в 0.1 M ацетата натрия (pH 4.5). Инкубирование осуществляли в темноте в течение 30 минут при перемешивании. Излишек периодата удаляли посредством добавления рибитола. Соединения очень низкого молекулярного веса удаляли диализом (Медицелл Интернешнал Лтд. Мембрейн, Лондон). Полученные олигосахариды имели свободные альдегидные группы, способные к реакции с первичными аминами (напр. , лизиновые остатки в протеинах). Конъюгат получали смешиванием протеина и полисахарида в весовом соотношении 1:1 с добавлением цианоборгидрида натрия и подверганием смеси инкубированию, сначала 48 часов при 4oC, а затем при 37oC в течение 24 часов. Комплекс высокого молекулярного веса, который содержит полученный конъюгат в соотношении протеин/полисахарид 1:2.3, мог быть отделен от нереактивных загрязнителей посредством HPLC.
Пример 9.
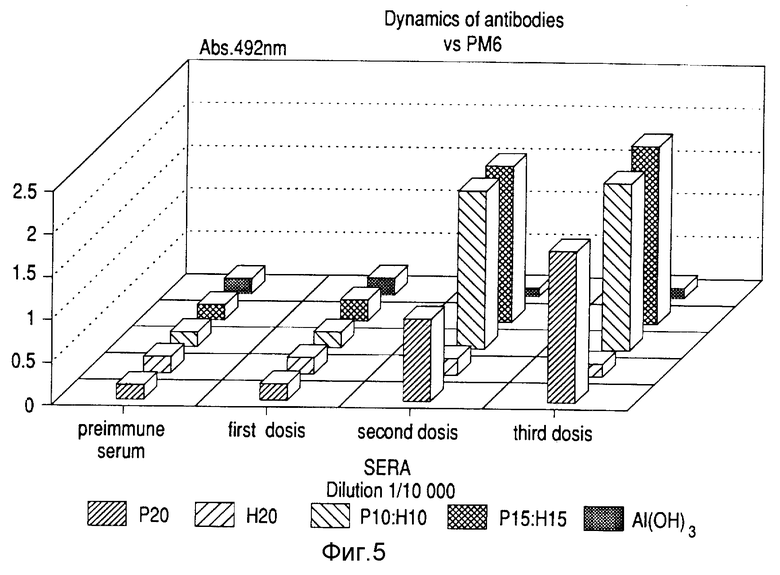
Бивалентный вакцинный препарат против вируса гепатита "B" и N.meningitidis. Для получения бивалентного вакционного препарата смешивали различные количества протеина Р64к и Поверхностного Антигена гепатита "B" (торговое наименование - Вакуна Рекомбинанте контра ла Гепатитис "B" "Хебер Биотек", Гавана, Куба. Эти антигены примешали к алюминий-гидроксидному гелю в количестве 2 мг на дозу и инокулировали BaI /с мышам с весом тела 20 г тремя дозами по 0,5 мл каждая.
Испытывали различные варианты:
1. Р64к 20 мкг (Р20).
2. HB9Ag 20 мкг (H20).
3. Р64к 10 мкг + HBsAg 10 мкг (P10:H10).
4. Р64к 15 мкг + HBsAg 5 мкг (P15:H5).
5. Плацебо (Al/OH/3).
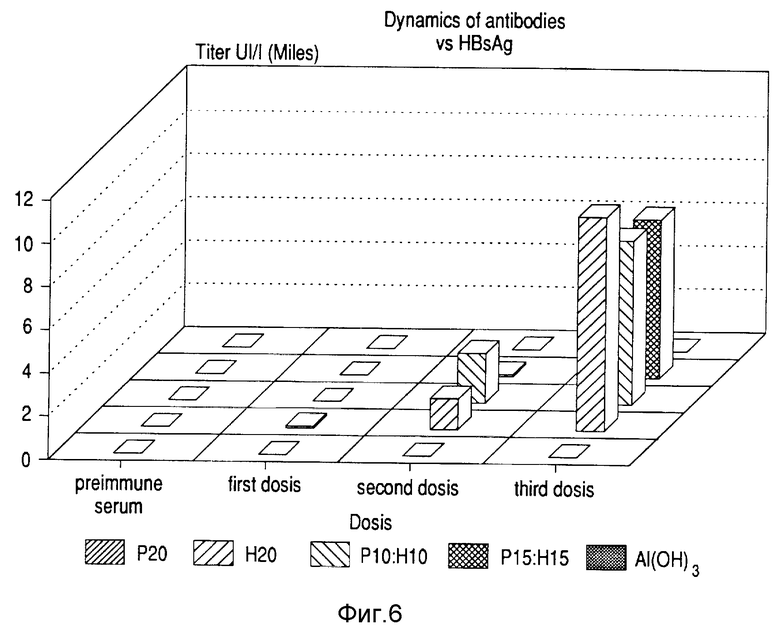
Через семь дней после иммунизации первыми дозами применяли вторую дозу. Третью дозу давали через 14 дней после второй. Семью днями позже брали кровь и отделяли сыворотку у каждого иммунизированного животного. Титры антител против Р64к протеина измеряли твердофазным ELISA с использованием Р64к в количестве 5 мг/мл для покрытия полистирольной чашки. Титры антител против HBsAg определяли посредством коммерческой ELISA (Органон Текника, Бокстел). Фиг. 5 показывает динамику реакции антител против протеина Р64к с использованием сыворотки, разведенной до 1/10 000. В реакцию против Р64к не вмешивался другой антиген. Фиг. 6 показывает титры против HBsAg после каждой дозы. Титры против этого белка не уменьшались в присутствии Р64к в препарате. Высокие титры получены против обоих антигенов в том же самом вакцинном препарате.
Пример 10.
Была создана компьютерная программа для поиска и обнаружения гомологий между Р64к и другими белками (EMBL - Европейская лаборатория молекулярной биологии - Дата-Бейс, т.е. информационный фонд EMBL). В результате поиска было обнаружено, что имеется гомология одного сегмента последовательно Р64к с сегментами последовательности N. gonorrhoeae. Было установлено, что эта последовательность является характеризующей как у N.gonorrhoeae, так и у N. Meningitidis (F.F. Correia, S. Jnouye и M. Jnouye, 1988, J. Biol. Chem. 263, N 25. 12194-12198).
Другая область с высокой гомологией была найдена у двух белков Пируват-Дегидрогеназного комплекса от штамма E.coli K:12:
а) ацетилтрансферазы от E.coli и белка Р64к внешней мембраны N.meningitidis.
Гомология существует между сегментом, включающий 100 аминокислот, повторяющимся в начале аминокислотной последовательности Ацетилтрансферазы ("Липоил-домен", включая "Липоил-связывающий сайт") (P.E. Stephens и др., 1983, Eur. J. Biochem 133, 481-489) и областью, расположенной в первых III аминокислотах Р64к (см. II в конце текста описания).
б) Липоамид-дегидрогеназа от E.coli и протеин Р64к внешней мембраны N. meningitidis.
Гомология существует между Липоамид-дегидрогеназой от E.coli (этот белок имеет 473 аминокислоты, P. E. Stephens и др., 1983, Eur, J. Biochem, 133, 481-489) и протеином Р64к, особенно в сегменте (см. III в конце текста), который представляет почти весь протеин, за исключением области с гомологией с "Липоил-доменом" Ацетилтрансферазы (см. III в конце описания), где: (-1-): Аденин-связывающий сайт (ГАД); (-2-): окислительно-восстановительная активная дисульфидная область; (-3): активный сайт гистидина.
Депонированные штаммы:
Клон E. coli HB101, содержащий плазмиду pM3 (pUC 18-плазмида, содержащая 4,1 кб ДНК-фрагмент N. meningitidis, штамма B:4:P1.15, клонированный между EcOR I и Hind III сайтами) был депонирован в августе 1991 в Баарне (Нидерланды), в Центральном Бюро культур грибков (CBS) и получил депозитарный номер CBS 4,8,5.91.
Список последовательностей.
Последовательность: N 1 (см. в конце текста).
Тип последовательности: нуклеотиды с соответствующими аминокислотами.
Длина последовательности: 1830 пар оснований (бр).
Количество нитей: одна.
Топология: линейная.
Тип молекулы: геномная ДНК.
Исходный организм-источник: N. meningitidis группы B.
Непосредственный экспертиментальный источник: штамм B:4:P.1:15, выделенный на Кубе.
Признаки: от 1 до 1830 бр - зрелый протеин.
Свойства: ген, кодирующий протеин Р64к из внешней мембраны N.meningitides.
Последовательность: N 2
Тип последовательности: нуклеотиды.
Длина последовательности: 15 оснований
ATGCTAGATA AAAGA (15).
Последовательность: N 3
Тип последовательности: аминокислоты.
Длина последовательности: 25 аминокислот.
Тип молекулы: N-терминал последовательности протеина Р64к из внешней мембраны
Последовательность: N 4
Тип последовательности: нуклеотиды.
Длина последовательности: 12 оснований
CTAGATAAAA GA (12)
Последовательность: N 5
Тип последовательности: нуклеотиды.
Длина последовательности: 8 оснований
TCTTTTAT (8)
Последовательность: N 6 (см. в конце текста).
Тип последовательности: аминокислоты.
Длина последовательности: 221 аминокислота.
Тип молекулы: последовательность, включающая вариабельные облаcти протеина Р64к.
Последовательность: N 7
Тип последовательности: аминокислоты.
Длина последовательности: 6 аминокислот.
Тип молекулы: последовательность, соответствующая сайту лигирования между N-терминалом Р64к и Р1.15
Последовательность: N 8
Тип последовательности: нуклеотиды.
Длина последовательности: 19 оснований.
Тип молекулы: последовательность, соответствующая лигированию N-терминала гена M-6 и гена P1.15
Последовательность: N 10
Тип последовательности: 18 оснований.
Тип молекулы: последовательность, соответствующая лигированию C-терминала от гена M-6 и гена P1.15
GAAGCCAACG CGATAGAA (18)
Последовательность: N 11 (см. в конце текста).
Тип последовательности: III аминокислот
Тип молекулы: Р64к N-терминал, включающий область гомологии с "липоил-связывающим сайтом" от Ацетилтрансферазы E.coli.
Последовательность: N 12 (см. в конце текста).
Тип последовательности: аминокислоты.
Длина последовательности: 112 аминокислот.
Тип молекулы: "липоил-связывающий сайт" от E.coli Ацетилтрансферазы.
Последовательность: N 13 (см. в конце текста).
Тип последовательности: аминокислоты.
Длина последовательности: 481 аминокислота.
Тип молекулы: фрагмент Р64к, включающий область гомологии с липоамид-дегидрогеназой от E.coli.
Последовательность: N 14 (см. в конце текста).
Тип последовательности: аминокислоты.
Длина последовательности: 472 аминокислоты.
Тип молекулы: сегмент E.coli липоамид-дегидрогеназы с гомологией с протеином Р64к.
| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛОК NMB1125 И ЕГО ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ | 2004 |
|
RU2336900C2 |
| БЕЛОК NMB0928 И ЕГО ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ | 2004 |
|
RU2335505C2 |
| ХИМЕРНЫЙ ГЕН CR3 И КОДИРУЕМЫЙ ИМ ХИМЕРНЫЙ БЕЛОК CR3 (ВАРИАНТЫ), ИНДУЦИРУЮЩИЙ ИММУННЫЙ ОТВЕТ ПРОТИВ ВИЧ-1 | 2002 |
|
RU2302461C2 |
| НАБОР ДЛЯ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ ПРОТИВ Neisseria meningitidis СЕРОЛОГИЧЕСКОЙ ГРУППЫ В | 2008 |
|
RU2498815C2 |
| ВАКЦИНЫ С ИСПОЛЬЗОВАНИЕМ ВЕЗИКУЛ НА ОСНОВЕ GNA 1870 ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ДЛЯ ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ NEISSERIA MENINGITIDIS | 2006 |
|
RU2432962C2 |
| АНТИГЕНЫ NEISSERIA MENINGITIDIS | 1999 |
|
RU2343159C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ ИНФЕКЦИИ, ВЫЗВАННОЙ БАКТЕРИЕЙ NEISSERIA | 2000 |
|
RU2244749C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТРАНСФЕРРИН-СВЯЗЫВАЮЩИЙ БЕЛОК И HSF ИЗ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ | 2003 |
|
RU2359696C2 |
| НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНУЮ РЕКОМБИНАНТНУЮ IgA1 ПРОТЕАЗУ NEISSERIA MENINGITIDIS СЕРОГРУППЫ B, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ НУКЛЕОТИДНУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩУЮ АКТИВНУЮ IgA1 ПРОТЕАЗУ, ШТАММ-ПРОДУЦЕНТ, СОДЕРЖАЩИЙ ПЛАЗМИДНУЮ ДНК, ПРОДУЦИРУЮЩИЙ ЗРЕЛУЮ ФОРМУ IgA1 ПРОТЕАЗЫ, РЕКОМБИНАНТНАЯ Ig ПРОТЕАЗА NEISSERIA MENINGITIDIS СЕРОГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОЙ ФОРМЫ IgA1 ПРОТЕАЗЫ, ОБЛАДАЮЩЕЙ ИММУНОГЕННЫМИ И ПРОТЕКТИВНЫМИ СВОЙСТВАМИ | 2011 |
|
RU2453599C1 |
















Изобретение касается метода выделения нуклеотидной последовательности, кодирующей протеин, имеющий мол.м. 64000 дальтон, который располагается на внешней мембране N. meningitidis так же, как и рекомбинантной ДНК, полученной на ее основе. Целью изобретения является идентификация нуклеотидной последовательности, кодирующей высококонсервативный протеин, производства этого протеина с высокой чистотой. Изобретение может быть использовано в способах диагностики и в изготовлении вакцин с широким спектром иммунозащиты. 6 с. и 5 з.п. ф-лы, 6 ил., 1 табл.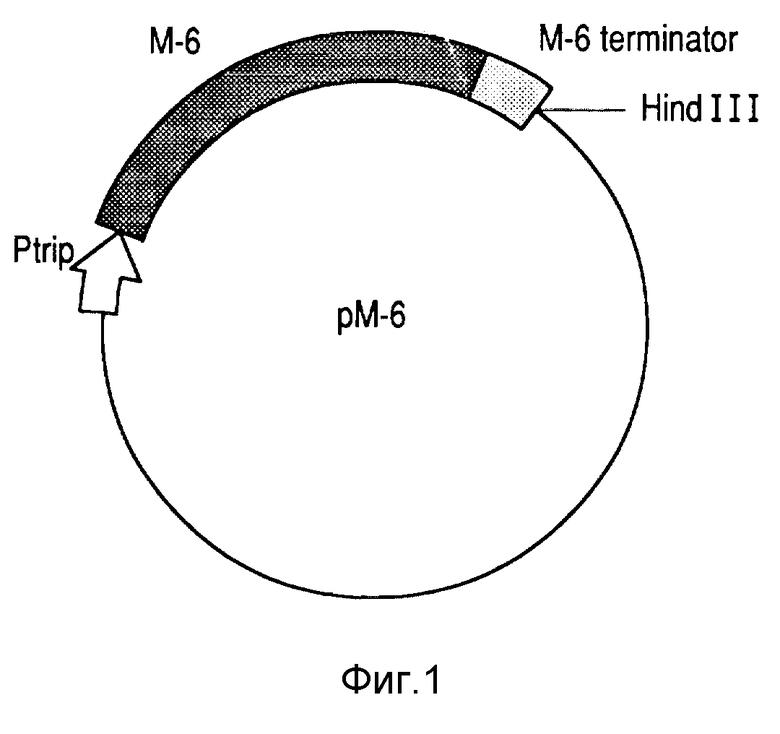
| EP 0301992, 05.06.89. |

