ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к диизопропилфторфосфатазе (далее называемой ДФФаза, классификация ферментов ЕС 3.1.8.2) из Loligo vulgaris, к последовательности нуклеотидных оснований, кодирующей фермент, а также к применению фермента и способу его получения в трансформированных клетках.
УРОВЕНЬ ТЕХНИКИ
Термин ДФФаза широко используется при описании класса ферментов, которые способны гидролизовать диизопропилфторфосфат (ДФФ) и другие фосфорорганические соединения со сходной структурой. Группу этих фосфорорганических соединений можно свести к соединениям со следующей базисной формулой:
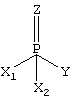
Здесь Z обозначает кислород или серу, Y обозначает группу, которая благодаря связи H-Y может обладать кислотными свойствами. В частности, это группы ангидридов кислот (депротонированные кислотные группы) (в частности, F-или СN-группы) или сложноэфирные группировки, такие как тиоэфирная, энолэфирная или п-нитрофенилэфирная группы. Группами X1 и Х2 могут быть неразветвленные или разветвленные, или циклические, содержащие от 1 до 15 атомов углерода, алкокси-, алкил-, алкиламино- или диалкиламиногруппы. Многие соединения такого рода используются или ранее использовались как инсектициды. Прочие соединения попадают под понятие так называемых нервно-паралитических газов. К нервно-паралитическим газам из этого класса соединений относятся, в частности ДФФ, табун, зоман, зарин, этилзарин и циклозарин.
Ликвидация мировых запасов этих высокотоксичных соединений представляет собой возрастающую проблему. Поэтому необходимо найти не оказывающие неблагоприятного влияния на окружающую среду способы, чтобы очистить (деконтаминировать) от таких соединений уже отравленные зоны окружающей среды.
До сих пор важнейшим способом уничтожения больших количеств этих веществ является сжигание их при высоких температурах.
Поэтому, например, патент US-A-4666696 посвящен разрушению нервно-паралитических газов и других ингибиторов холинэстеразы путем восстановления их расплавленным алюминием.
Другие попытки решения этой проблемы заключаются, например, в применении физических способов. Так, например, патент US-A-5550311 описывает термическое разложение токсичных соединений (нервно-паралитических газов) в струе воды. Другая попытка описана в патенте US-A-5648591, при которой химическое оружие, например зарин, разрушают путем активации в механических мельницах. Все эти способы уже обнаружили серьезные недостатки, связанные с их неэкономичностью и ограниченным спектром применений. Так, например, во всех этих способах существует необходимость использовать аппараты, соответствующие определенным токсичным веществам, в результате чего, например, использование их в полевых условиях затруднено, если вообще возможно.
Биологическое уничтожение запасов химического оружия и деконтаминация с помощью микроорганизмов или ферментов, которые можно производить в больших количествах, могли бы обеспечить упрощение или более эффективную попытку решения этой проблемы.
Поэтому от описанных выше способов деконтаминации отличается настоящее изобретение, предпосылки которого основаны на исследованиях Aldridge 1953 года. Aldridge наблюдал, что разнообразные ткани различных организмов содержат ферменты, которые гидролизуют параоксон, ингибитор холинэстеразы. Более поздние исследования 1966 года Hoskin и сотрудников показали, что в кальмаре Loligo pealii содержится расщепляющий ДФФ фермент. До сегодняшнего дня продолжались многочисленные исследования в этой области, однако, до сих пор все же не удавалось осуществить определение последовательности аминокислотных остатков или получение рекомбинантного белка. Исследованный Hoskin фермент не удалось точно охарактеризовать с точки зрения его молекулярной массы. В то время, как Hoskin исходил из молекулярной массы 26600 Да, другие группы исследователей, например Kopes-Smyth et al. (1993), считали вероятным, что, в случае белка с молекулярной массой 26,6 кДа, речь идет всего лишь о фрагменте очень нестабильного белка с молекулярной массой 42 кДа. Короткая последовательность аминокислотных остатков пептидного фрагмента этой ДФФазы была опубликована в 1993 г. Ward и Deschamps. Описанный там отрезок последовательности не обнаруживается в заявляемой аминокислотной последовательности согласно настоящему изобретению, из чего можно сделать вывод, что ДФФаза из Loligo pealii явно отличается от ДФФазы согласно настоящему изобретению.
Кроме Loligo peatii, сходные белки были идентифицированы в самых разнообразных организмах. Так, в том числе, их обнаружили у термофильных или галофильных бактерий, у Е. соli, Proteus vulgaris, Saccharomyces cerevisiae, Pseudomonas diminuta, Flavobacterium и у эукариотического одноклеточного организма Tetrahymena thermophila, а также у насекомых и беспозвоночных. Также удалось обнаружить их у млекопитающих, таких как мыши, крысы, кролики, свиньи, и у человека. Несмотря на усилия многих рабочих групп, которые занимаются ферментами, которые по своей специфичности напоминают ДФФазу из Loligo vulgaris согласно настоящему изобретению, ни одной из этих групп не удалось полностью описать фермент согласно настоящему изобретению в отношении последовательности аминокислотных остатков и лежащей в ее основе последовательности нуклеотидных оснований, трансформировать соответствующую последовательность нуклеотидных оснований при помощи вектора в клетку-реципиент и таким образом обеспечить возможность промышленного производства указанного фермента с исключительной чистотой. Хотя DierI (1955, диссертация в Университете имени Иоганна Вольфганга Гете, Франкфурт-на-Майне) в рамках его научной работы удалось расшифровать часть последовательности ДФФазы из Loligo vulgaris, характеристики последовательности аминокислотных остатков или последовательности нуклеотидных оснований нельзя заимствовать из этой работы. Представленная Dierl частичная информация является неполной, а доработка представленных результатов невозможна уже потому, что генетический банк кДНК, лежавший в основе работы, не является общедоступным.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Одна из задач настоящего изобретения состоит в том, чтобы предоставить в распоряжение полный и способный функционировать ДФФ-расщепляющий фермент из Loligo vulgaris и кодирующую фермент последовательность нуклеотидных оснований, для того чтобы обеспечить возможность промышленного производства с использованием генной инженерии. Другой задачей было получить ДФФазу, которую можно выделить в промышленных масштабах без потери стабильности посредством фракционирования на колонках с сульфатом аммония, и оптимум активности которой находится при нейтральном значении рН, равном примерно 7,5, и комнатной температуре (примерно 25°С). За счет этого, в конечном итоге, должна была быть решена задача осуществления не наносящего ущерба окружающей среде, энергосберегающего уничтожения нервно-паралитических газов и инсектицидов. Затем следовало решить задачу получения стабильной при хранении и устойчивой к растворителям ДФФазы. Так, например, концентрированный раствор должен был храниться в течение длительного времени при 4°С без существенного снижения активности. Также на активность фермента не должны были влиять различные растворители или водные среды, содержащие растворители (например, 10%-ный водный раствор этилового спирта). Далее нужно было решить задачу получения ДФФазы, которая активна в различных буферных системах. Получение такой ДФФазы должно было, как следствие, обеспечить возможность деконтаминации, детоксикации, а также обнаружения ингибиторов ацетилхолинэстеразы, которые являются субстратами ДФФазы. Получение такого фермента должно не только обеспечить промышленное уничтожение соответствующих нервно-паралитических газов и инсектицидов, но и дать возможность деконтаминировать загрязненные естественные среды (почвы, водоемы и т.д.). Также таким путем возможно клонирование в растениях. Далее возможно применение фермента для производства лекарственных средств для детоксикации или лечения человека и животных. При этом фермент мог бы применяться, например, местно в форме крема для кожи, парентерально в форме раствора для инфузий или ингаляций, или даже перорально.
Вышеперечисленные задачи были решены путем получения ДФФазы с последовательностью аминокислотных остатков приведены в конце.
Заявленная диизопропилфторфосфатаза при этом охватывает также последовательности аминокислотных остатков, которые можно получить путем делеции, вставки и/или замещения одной или нескольких аминокислот из вышеуказанной последовательности, если активность ДФФазы сохраняется. Это, разумеется, включает и укорочение с амино- и/или карбокситерминальной стороны.
В основе этой последовательности аминокислотных остатков согласно настоящему изобретению лежит ДНК-последовательность согласно настоящему изобретению, которая включает ДНК-последовательность, которая кодирует ДФФазу из Loligo vulgaris и которая в предпочтительной форме осуществления включает следующую последовательность нуклеотидных оснований.
В результате изменений последовательности нуклеотидных оснований можно изменить, например, субстратную специфичность, растворимость и/или стабильность фермента в соответствии с возникающими проблемными ситуациями.
Еще одна задача настоящего изобретения состоит в том, чтобы обеспечить вектор, который содержит, как минимум, одну копию ДНК-последовательности согласно настоящему изобретению. Этот вектор может быть любым прокариотическим или эукариотическим вектором, на котором находится ДНК-последовательность согласно настоящему изобретению, предпочтительно под контролем сигнала экспрессии. К сигналам экспрессии относятся, в частности, промоторы, операторы и усилители. Промоторами могут быть, например, Т7-промотор или lac-, tac-, lacUV5- или РL-промоторы. Подходящими прокариотическими векторами являются, например, хромосомные векторы, такие как бактериофаги (например, бактериофаг λ) и экстрахромосомные векторы, такие как плазмиды, причем при этом предпочтительны циркулярные плазмидные векторы. Вектор согласно настоящему изобретению, с другой стороны, может быть и эукариотическим вектором, например дрожжевым вектором или вектором, подходящим для более высокоразвитых клеток, как, например, плазмидным вектором или вирусным вектором. Векторы вышеуказанных видов известны специалистам в области молекулярной биологии, так что нет необходимости рассматривать этот вопрос подробнее. В предпочтительной форме осуществления изобретения в качестве векторов такого рода могут служить так называемые векторы секреции, которые осуществляют экспрессию белка слияния, состоящего из ДФФазы и сигнального белка, причем сигнальный белок отвечает за то, что белок пройдет через клеточную мембрану и при секреции будет точно отделен от ДФФазы при помощи сигнальных пептидов. В качестве сигнальных белков могут рассматриваться, например, реlВ-, α- или РН01-сигнальные белки. Соединение ДФФазы с сигнальными пептидами такого рода предпочтительно для того, чтобы воспрепятствовать образованию “инклюзионных тел”, то есть агрегатов ДФФазы внутри клетки, и вывести большие количества белка в периплазматическое пространство или межклеточную среду.
Настоящее изобретение далее относится к клетке, которая трансформирована при помощи ДНК-последовательности согласно настоящему изобретению или вектора согласно настоящему изобретению. Эта клетка может быть, например, прокариотической клеткой, предпочтительно грам-отрицательной прокариотической клеткой, особенно предпочтительно - эубактериальной клеткой. В одной из форм осуществления изобретения эта клетка является клеткой Е. соli. Трансформация прокариотических клеток при помощи экзогенных последовательностей нуклеиновых кислот известна специалистам в области молекулярной биологии. Клетка согласно настоящему изобретению может, однако, быть и эукариотической клеткой, такой как грибковая клетка (например, дрожжевая), животная или растительная клетка. Предпочтительными эукариотическими системами экспрессии являются, например, Pichia pastoris или Saccharomyces cerevisiae. Трансформация или трансфекция эукариотических клеток при помощи экзогенных последовательностей нуклеиновых кислот известна специалистам в области молекулярной биологии, и поэтому ее не нужно разъяснять здесь более подробно.
В результате получения вышеназванных белков, ДНК-последовательностей, векторов и трансформированных клеток будет возможно промышленное производство фермента с ДФФазной активностью. За счет этого впервые открывается возможность простым, экономичным и благоприятным для окружающей среды способом разрушить большие количества веществ, ингибирующих ацетилхолинэстеразу.
При этом ДФФазу можно получать при помощи технологии получения рекомбинантных ДНК как составную часть экстракта из организма-хозяина или в выделенном и очищенном виде, например, путем экспрессии в Е. coli. Применение согласно настоящему изобретению такой ДФФазы состоит в разрушении ингибиторов ацетилхолинэстеразы, содержащих P-F-связи или соответствующие базисной формуле P-Y-связи. Предпочтительно, чтобы при этом очищенная и изолированная ДФФаза помещалась в ферментативный реактор, например, для промышленного расщепления содержащих P-Y-связи (в соответствии с базисной формулой) ингибиторов ацетилхолинэстеразы. Иммобилизация ферментов в реакторе такого рода известна специалистам, и поэтому ее не нужно обсуждать здесь подробно. Кроме того, допустима полимеризация ДФФазы с пенопластами, например пенополиуретаном.
Еще одна форма осуществления применения - это, например, применение ДФФазы в интактном микроорганизме, который можно использовать, например, для деконтаминации почв в природных условиях, предпочтительно при этом используются трансформированные почвенные бактерии.
Еще одно применение согласно настоящему изобретению состоит в том, чтобы ДФФазу согласно настоящему изобретению включить в пену, причем пена выступает в качестве носителя и/или смачивающего агента. Такой пеной может быть пена поверхностно-активного вещества, которая, в частности, может быть использована для деконтаминации почв, поверхностей, ценных приборов или тому подобного. В некоторых случаях можно использовать также простое обрызгивание, а также нанесение в виде аэрозоля. Использование в форме пены тогда не является настоятельно необходимым.
Возможно также применение ДФФазы в технологиях для стационарной работы, в которых используются, например, реакторы, а также применение ДФФазы в мобильных устройствах при деконтаминации приборов или больших площадей открытого грунта. Другие формы осуществления применения согласно настоящему изобретению могут состоять в детоксикации загрязненных водоемов или питьевой воды. Иммобилизация фермента для применения его в качестве стационарной фазы в реакторе может осуществляться, например, за счет ковалентного присоединения фермента к твердому носителю.
К кодирующей кДНК может быть присоединен "His-Tag", который дает возможность получения модифицированной ДФФазы. Эта модифицированная ДФФаза способна присоединяться к никель-NТА-модифицированному материалу-носителю. Если в случае никель-NТА-модифицированного материала-носителя речь идет о материале-наполнителе для разделительной колонки, то этот способ может служить для очистки ДФФазы.
Далее можно импрегнировать ферментом текстильные изделия посредством ковалентного или нековалентного связывания, чтобы они служили в качестве защитной одежды.
Применение согласно настоящему изобретению может состоять также в новых способах обнаружения, например биосенсорных. При этом ДФФаза служит рецептором, который иммобилизован на “датчике”. Если ДФФаза расщепляет вещество, подлежащее выявлению, то есть вышеописанные субстраты, то биологический сигнал преобразуется в соответствующий измеряемый электрический сигнал, который усиливается электронным компонентом. Выходной сигнал обычно зависит от количества и/или вида анализируемого вещества. При этом в качестве рецептора может использоваться ДФФаза в чистом выделенном виде или в виде синтезирующих ее клеток. В качестве “датчика” могут рассматриваться известные специалистам в области биосенсорики преобразующие компоненты.
Еще одно применение согласно настоящему изобретению состоит в приготовлении лекарственных средств, которые в качестве активного вещества содержат ДФФазу и поэтому могут способствовать детоксикации или лечению людей или животных, отравленных ингибитором ацетилхолинэстеразы, содержащим P-Y-связи. При этом, наряду с местным применением, рассматриваются также парентеральное или пероральное введения.
Настоящее изобретение относится также к способу получения ДФФазы, при котором ее продуцирует клетка согласно настоящему изобретению. Этим данный способ отличается от полного химического синтеза ДФФазы. При способе получения ДФФазы согласно настоящему изобретению при помощи трансформированной клетки, трансформации при помощи вектора экспресии предшествуют следующие этапы работы:
(1) Выделение мРНК из головных ганглиев Loligo vulgaris, извлекаемых сразу же после умерщвления животного, причем необходимо немедленно подвергнуть ганглии глубокой заморозке в жидком азоте.
(2) Очистка мРНК посредством аффинной хроматографии на колонке с олиго-(dТ)-целлюлозой,
(3) Трансляция мРНК в кДНК,
(4) Клонирование кДНК в λ-фагах,
(5) Выявление λ-фагов, содержащих генетическую информацию ДФФазы, с помощью дегенерирующих зондов с использованием полимеразной цепной реакции (ПЦР),
(6) Амплификация генетической информации с помощью полимеразной цепной реакции,
(7) Перенос генетической информации в вектор экспрессии.
При этом способе приходилось преодолевать различные трудности, которые не смог бы преодолеть специалист в области генной инженерии с помощью имеющихся в его распоряжении стандартных методик. Так, например, ранее не было очевидно, но оказалось настоятельно необходимым, чтобы выделение мРНК Loligo vulgaris производилось из головных ганглиев только что умерщвленного животного, причем головные ганглии сразу же после их выделения необходимо помещать в жидкий азот. Если не поступать таким образом, шансы выделить мРНК “полной длины” близки к нулю. Малое число молекул мРНК, кодирующей ДФФазу, создает настоятельную необходимость использовать методику полимеразной цепной реакции. Для выявления генетической информации в банке кДНК необходимо было разработать новые высокоспецифичные зонды, которые должны были на длинных отрезках совпадать с кДНК Loligo vulgaris. Синтез этих зондов был затруднен тем, что на момент изобретения не были расшифрованы длинные белковые последовательности ДФФазы из Loligo vulgaris, которые могли бы помочь синтезировать высокоспецифичные зонды. Только с использованием информации о последовательности, представленной в настоящем изобретении, специалист в области молекулярной биологии может синтезировать зонды такого рода простым способом.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Нижеследующий пример должен служить для наглядной иллюстрации изобретения.
Примеры
1. Выделение головных ганглиев Loligo vulgaris и последующая экспрессия рекомбинантных белков в Escherichia coli
(1) Для выделения головных ганглиев голову кальмара отделяют от мешка с внутренностями и удаляют щупальца. После этого эпидермис легко можно поднять, чтобы произвести разрез, который идет от ротовой полости до затылка. Обнажают головную капсулу и глаз. После удаления всех нервных стволов извлекают ткань мозга. Сразу же после выделения ткань мозга переносят в жидкий азот и до получения препарата ДНК хранят при - 196°С.
Всю стеклянную посуду сушат в течение 4 часов при 250°С. Все растворы, насколько это возможно, обрабатывают диэтилпирокарбонатом. Ткань мозга переносят в заполненную жидким азотом ступку и растирают до получения гомогенной пасты. Затем ткань переносят в цитратный буфер, который содержит тиоизоцианат гуанидиния, β-меркаптоэтанол и N-лауроилсаркозин. Для растворения еще существующих мельчайших групп клеток полученную суспензию клеток гомогенизируют в стеклянной пробирке. Перед проведением ультрацентрифугирования еще присутствующие частицы ткани удаляют из гомогенизата путем центрифугирования. Затем для отделения РНК от остальных составных частей клетки производят ультрацентрифугирование в градиенте хлорида цезия. Осадок после центрифугирования промывают этиловым спиртом и после растворения в воде многократно экстрагируют фенолом/хлороформом. Для контроля качества и количества мРНК проводят обычные анализы.
(2) Обогащение мРНК осуществляют с помощью аффинной хроматографии на олиго-(dТ)-целлюлозе. Для этого после заполнения колонки для аффинной хроматографии общую РНК нагревают, а затем охлаждают в ванне со льдом. После растворения вторичных структур общую РНК при высокой концентрации солей переносят на колонку с олиго-(dТ)-целлюлозой. При этом поли-(А+)-РНК специфически связывается с материалом-носителем. Чтобы повысить выход мРНК, предпочтительно собрать элюат, повторно денатурировать и вновь пропустить через колонку. Колонку с присоединенной к ней поли-(А+)-РНК промывают заряженным и промывочным буферами. С заряженной олиго-(dТ)-целлюлозы поли-(А+)-РНК элюируется раствором с меньшей концентрацией соли и определяется спектрофотометрически в соответствующих фракциях.
(3) Для синтеза первой цепи кДНК используют олиго-(dТ)-праймер, который, кроме необходимой для связывания поли-(dТ)-последовательности, содержит Xhol-отрезок и последовательность "GAGA". После образования гибрида с помощью обратной транскриптазы синтезируется цепь ДНК, комплементарная мРНК. При проведении этой реакции из-за низкой активности РНазы Н для повышения эффективности процесса и меньшего ингибирования поли-(А-)-РНК используют обратную транскриптазу из вируса лейкемии мышей Moloney (M-Mul VRT). Для защиты от рестрикционного расщепления фермента при синтезе первой цепи используют смесь dАТФ, dГТФ, dТТФ и 5-метил-d-ЦТФ. После синтеза для первых циклов репликации используют бактериальный штамм Escherichia coli PKL-F’, который несет генетические маркеры тсrА- и тсrВ-.
После синтеза первой цепи кДНК синтезируют вторую цепь по методу Gubler и Hoffman. Вначале при помощи эндорибонуклеазы РНазы Н отщепляют РНК от двойных цепей РНК:ДНК. При этом образуются олигорибонуклеотиды с 5’-фосфат- и 3’-гидроксильными концами, которые распознаются ДНК-полимеразой I и используются как стартеры синтеза ДНК.
Чтобы клонировать полученную двойную цепь ДНК в подходящий вектор, на первом этапе выступающие концы ДНК-цепей дополняют при помощи Т4 ДНК-полимеразы. Затем с помощью Т4 ДНК-лигазы концы кДНК соединяют с адаптерами EcoR I. При этом используют смесь фосфорилированного 9-мерного и дефосфорилированного 13-мерного олигодезоксирибонуклеотидов. После тепловой инактивации лигазы выступающие 5’-концы фосфорилируют с помощью Т4 полинуклеотидкиназы и выравнивают двойную цепь ДНК с помощью рестрицирующего фермента XhoI. В результате использования двух различных участков рестрикции на концах кДНК возможно направленное клонирование в подходящий вектор.
(4) Полученную кДНК клонируют в векторные ветви λ-бактериофагов. Путем упаковки in vitro в предварительно подготовленные головки фагов и инкубации с различными клеточными экстрактами из мутантных фагов получают жизнеспособные бактериофаги. Создают два банка кДНК с различным числом рекомбинантных векторных молекул, которые служат исходными материалами для дальнейших работ.
(5) Для проверки обоих банков кДНК на содержание генетической информации диизопропилфторфосфатазы на основании ранее известной частичной информации о последовательности аминокислот в белке - с помощью дополнительных методик - выделяют олигодезоксирибонуклеотиды, которые должны иметь наибольшее из возможных соответствие кДНК последовательности белка. С использованием полимеразной цепной реакции из одного олигодезоксирибонуклеотида со специфической последовательностью -последовательность 5’-TTC-CAA-TTC-CCI-AAT-GGI-ATT-GCT-GT-3’ - при очень жестких условиях получают продукты. В ПЦР-экспериментах для идентификации диизопропилфторфосфатазы из Loligo vulgaris использовались два олигодезоксирибонуклеотида. Первый олигодезоксирибонуклеотид состоит из последовательности λ-бактериофага, которая примыкает к внедренной последовательности кДНК. Второй олигодезоксирибонуклеотид, который в нескольких положениях содержит инозин, является производным установленной последовательности аминокислот белка и поэтому специфичен для диизопропилфторфосфатазы из Loligo vulgaris.
В банках кДНК можно обнаружить продукт с длиной 550 пар оснований. После выделения и очистки продукта полимеразной цепной реакции можно прочитать содержащуюся в ДНК информацию в опытах по определению последовательности нуклеотидных оснований. Путем применения олигодезоксирибонуклеотидов, которые являются производными от уже известной последовательности ДНК, можно осуществить определение последовательности нуклеотидных оснований в обеих ДНК-цепях кДНК-вставки. Путем сравнения результатов, полученных при определении последовательности нуклеотидных оснований в ДНК, с ранее определенными последовательностями аминокислотных остатков в белках можно доказать, что в случае фрагмента с 550 парами оснований речь идет о частичной последовательности искомой диизопропилфторфосфатазы из Lo/igo vulgaris.
Исходя из этой частичной последовательности фермента можно определить всю последовательность кДНК диизопропилфторфосфатазы с общей длиной в 1210 пар оснований. Эта ДНК-последовательность характеризуется областью на 5’-конце длиной примерно в 210 пар оснований, которая не содержит кодирующей ДНК-последовательности. Открытый для чтения участок состоит из 942 пар оснований и кодирует белок с 314 аминокислотами, который имеет молекулярную массу, примерно равную 35 кДа.
(6) Для создания системы экспрессии открытый для чтения участок фермента амплифицируется при помощи фланкирующего олигодезоксирибонуклеотида в полимеразной цепной реакции. Дополнительно к информации о специфической последовательности белка, олигодезоксирибонуклеотиды содержат участки рестрикции, которые упрощают клонирование в систему экспрессии. К 5’-концу информационного участка присоединен отрезок Nсо I, к 3’-концу ДНК - отрезок Hind III. Продукт полимеразной цепной реакции обрабатывают Pfu-Taq-полимеразой, очищают на агарозном геле, выделяют и помещают в разомкнутый вектор pCR-Script SK(+).
(7) После выбора плазмид, которые содержат искомую генетическую информацию, ее вырезают при помощи Nсо I/Hind III и переносят в вектор экспрессии с trс-промоторной системой. В результате экспрессии в этой системе можно получить 100-200 мг рекомбинантной и биологически активной диизопропилфторфосфатазы на один литр среды.
2. Исследование ферментативной активности рекомбинантной ДФФазы
Способность полученной описанным выше способом рекомбинантной ДФФазы гидролизовать диизопропилфторфосфат (т.е ее ферментативная активность) определялась путем титрования выделяемых в результате гидролиза протонов раствором NaOH с использованием радиометра "рН Stat" согласно методикам, описанным в документе F.C.G.Hoskin, A.H.Roush, Science 215 (1982) 1255-1257.
В результате проведенных исследований было установлено, что активность рекомбинантной ДФФазы, выраженная в единицах (U) на миллиграмм фермента составляет 213±10 U/мг, где единица активности U соответствует 1 мкмоль гидролизованного диизопропилфторфосфата (ДФФ) в минуту.
3. Получение мутантной и химически модифицированной ДФФазы
Химическая модификация звеньев ДФФазы, имеющей описанную выше последовательность, осуществлялась с использованием методов, описанных в следующих источниках информации: G.L.Ellman, Arch. Biochem. Biophys., 82, 70-77 (1959); E.W.Miles, Modification of histidyl residues in proteins by diethylpyrocarbonate, Methods in Enzymol, 47, 431-442 (1977); J.F.Riordan, Biochemistry 12(20): 3915-3923 (1973); Т.Okuyama, К.Satake, J.Biochem. (Tokyo) 47, 454-466 (1960); J.F.Riordan, Mol. Cell. Biochem. 26, 71-92 (1979); D.G.Hoare, D.E.Koshland Jr., A Procedure for the Selective Modification of Carboxyl Groups in Protein. J.Am.Chem.Soc. 88, 2057-2058 (1966).
Указанная модификация осуществлялась с использованием групп-селективных реагентов путем инкубирования фермента с большим избытком реагента. В качестве групп-селективных реагентов использовались следующие:
Целевая аминокислота Реагент
Cys дитиобиснитробензоат
Cys иодоацетамид
Туr N-ацетилимидазол
Gly/Asp 1-этил-3-(3-иметиламинопро
пил)карбодиимид
Arg бутандион
Lys тринитробензолсульфокислота
His диэтилпирокарбонат
His диазобензолсульфокислота
His метиленовый синий
Избыток реагента составлял от 35 до 600 моль реагента на 1 моль белка. Мутантные белки были сконструированы из ДФФазы, имеющей описанную выше последовательность, путем сайт-направленного мутагенеза с использованием методик Quick-Change Protocol, Stratagene, La Jolla, CA, США. Таким образом были получены, в частности, мутантные белки H181N, H219N, H224N, H248N, H287N, в которых аминокислотные остатки His были заменены на Asp, а также E37Q, W244Y, W244H, W244F, W244L, S271A, E21Q и D229N.
4. Исследование ферментативной активности мутантной и химически модифицированной ДФФазы
Оценивалась относительная активность модифицированных и мутантных белков по сравнению с активностью ДФФазы дикого типа, имеющей описанную выше последовательность. Исследования ферментативной активности полученной мутантной и химически модифицированной ДФФазы осуществляли аналогично описанному выше для рекомбинантной ДФФазы дикого типа, а также с использованием ион-селективного фторидного электрода.
В результате проведенных исследований было установлено, что даже при модификации нескольких тирозиновых, цистеиновых, аргининовых, лизиновых, глутаминовых и аспартатных звеньев существенного снижения ДФФазной активности белка по сравнению с ДФФазой дикого типа не происходит, причем в ряде случаев ферментативная активность белка существенно повышается. Небольшое снижение активности мутантных белков может объясняться изменениями структуры белка, возникающей в результате замены гистидиновых звеньев на аспарагиновые. Заметное снижение активности, которое наблюдалось при химической модификации гистидиновых звеньев диазобензолсульфокислотой и метиленовым синим могло быть следствием либо селективной модификации активного сайта, либо конформационных изменений, возникающих в результате химической модификации звеньев.
5. Исследование субстратной специфичности ДФФазы
Субстратная специфичность рекомбинантной ДФФазы, а также мутантной и химически модифицированной ДФФазы, полученной описанными выше способами, исследовалась в отношении субстратов, представляющих собой соответствующие приведенной выше базисной формуле фосфорорганические соединения, являющиеся ингибиторами ацетилхолинэстеразы, в частности зарин, ДФФ, табун. Исследования осуществлялись в соответствии со стандартами НАТО.
Указанные субстраты наносились на покрытые полиуретаном металлические пластины размером 10×10 см. После этого на пластины наносились (распылялись) исследуемые вещества. После инкубирования в течение 10-15 минут следов субстрата на поверхности пластин не было обнаружено. При этом в результате указанных исследований было установлено, что некоторые из мутантных белков ДФФазы обладают активностью, превышающей активность ДФФазы дикого типа в 4-15 раз.
Учитывая механизм реакции гидролиза соединений, описываемых приведенной выше базисной формулой, ДФФазой согласно настоящему изобретению, который включает расщепление фосфорангидридной связи указанных соединений, на основании полученных результатов анализа специфической активности фермента можно сделать вывод о том, что:
- ДФФаза в соответствии с изобретением обладает способностью гидролизовать (разрушать) ингибиторы ацетилхолинэстеразы, содержащие фосфорангидридную связь и соответствующие приведенной выше базисной формуле;
- ДФФаза в соответствии с изобретением пригодна для детоксикации (деконтаминации) нервно-паралитических агентов G-типа, соответствующих указанной базисной формуле.
Изобретение относится к области энзимологии, в частности к получению ферментов с помощью технологии рекомбинантных ДНК, и может быть использовано в медицине и при разработке способов охраны окружающей среды. Из головных ганглиев кальмара Loligo vulgaris выделена суммарная мРНК и получена библиотека кДНК, путем скрининга которой клонирован фрагмент ДНК, кодирующий фермент диизопропилфторфосфатазу (ДФФазу). В результате экспрессии клонированного фрагмента в E.coli получена и охарактеризована рекомбинантная ДФФаза, а также ее мутантные и химически модифицированные формы, которые могут быть использованы в способах, предусматривающих разрушение ингибиторов ацетилхолинэстеразы. 4 с. и 11 з.п. ф-лы.
или последовательностью, которую можно получить из нее путем делеции, вставки и/или замещения одной или нескольких аминокислот, и которая обладает активностью, присущей исходной диизопропилфторфосфатазе.
ATGGAAATTC CAGTTATCGA ACCTCTTTTC ACAAAAGTGA CCGAAGATAT ACCAGGTGGA 60
GAGGGTCCCG TTTTTGACAA AAATGGCGAT TTTTATATCG TGGCCCCCGA AGTTGAAGTT 120
AACGGAAAAC CGGCGGGAGA AATTCTACGA ATCGATTTGA AAACAGGAAA GAAAACTGTG 180
ATCTGCAAAC CAGAAGTTAA TGGTTATGGA GGAATTCCTG CTGGCTGCCA ATGTGATCGA 240
GATGCCAACC AGCTGTTTGT GGCCGACATG AGACTCGGCT TGTTGGTCGT GCAAACTGAT 300
GGGACCTTTG AAGAGATTGC CAAAAAAGAC TCTGAAGGTA GAAGAATGCA GGGATGCAAT 360
GATTGCGCAT TTGATTATGA AGGTAACTTG TGGATCACTG CACCAGCTGG GGAAGTCGCA 420
CCTGCAGACT ACACCCGTTC AATGCAGGAA AAATTTGGCA GTATTTACTG CTTCACAACA 480
GATGGTCAAA TGATTCAAGT GGATACTGCT TTCCAGTTTC CAAATGGTAT TGCTGTTCGT 540
CACATGAACG ATGGCCGTCC TTACCAACTA ATTGTGGCTG AAACTCCAAC CAAGAAACTC 600
TGGAGTTATG ATATCAAAGG TCCAGCAAAG ATTGAAAACA AGAAAGTGTG GGGTCACATC 660
CCAGGTACTC ATGAAGGTGG TGCTGATGGA ATGGATTTTG ATGAAGACAA TAACCTTTTG 720
GTAGCCAACT GGGGGAGCTC ACACATCGAA GTGTTCGGCC CAGATGGGGG ACAGCCTAAA 780
ATGAGAATCC GTTGCCCATT TGAAAAACCC AGCAACTTGC ATTTCAAGCC CCAGACCAAA 840
ACCATTTTTG TCACGGAACA CGAGAACAAT GCTGTCTGGA AGTTTGAATG GCAAAGAAAT 900
GGCAAAAAAC AGTATTGTGA GACGTTAAAA TTTGGAATAT TT 942.
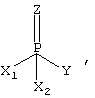
где Z обозначает кислород или серу;
Y - группу ангидрида кислоты (депротонированную кислотную группу), в частности F- или CN-группу, или сложноэфирную группу, в частности тиоэфирную, энолэфирную или п-нитрофенилэфирную группу;
Х1 и Х2 обозначают неразветвленную, разветвленную или циклическую, содержащую от 1 до 15 атомов углерода, алкокси-, алкил-, арил-, алкиламино- или диалкиламиногруппу, причем Х1 и Х2 являются одинаковыми или различными,
отличающийся тем, что включает этап взаимодействия указанных ингибиторов ацетилхолинэстеразы с ДФФазой по п.1.
Конвенционный приоритет по пп.1-15 установлен от 27.02.1998 в соответствии с заявкой 19808192.8, поданной в Патентное ведомство Германии.
| US 4666696, 19.05.1987 | |||
| US 5550311, 27.08.1996 | |||
| US 5648591, 15.07.1997. |

