Изобретение относится к нейротрофным факторам и в особенности к цилиарному нейротрофному фактору (CNTF), колирующей его ДНК и способу получения рекомбинантного CNTF.
Сильнейшие нарушения, психические и физические, происходят при гибели нервных или глиальных клеток в нервной системе. Гибель нервных или глиальных клеток может вызываться нейродегенеративными заболеваниями, такими, как старческий склероз мозга и рассеянный склероз, ишемией, вызванной инсультом, травматическим повреждением, естественными процессами старения.
Нейтротрофные факторы - класс молекул, которые способствуют выживанию и функциональной активности нервных и глиальных клеток. Существуют основания для предположения, что нейротрофные факторы предотвращают нарушения деятельности в условиях, перечисленных выше Appel, 1981, Ann. Neurology 10:499.
Лучше всех охарактеризован фактор роста нерва (NGF).
Эксперименты на животных демонстрируют, что NGF предотвращает смерть холинчувствительных нервных клеток диэнцефалона после травмы и что NGF способен предотвращать потерю познавательной способности, которая случается с возрастом. Hefti и Weiner, 1986, Ann. Neurology 20:275, Fisher et al, 1987, Nature, 329: 65. Эти результаты предполагают потенциальную клиническую полезность этого фактора для людей в лечении утраты познавательной способности, наблюдаемой при болезни, травме или старении.
Сложность использования нейротрофных факторов заключается в их специфичности только к тем субпопуляциям нервных клеток, которые обладают соответствующими мембранными рецепторами. У большинства нервных клеток тела NGF рецепторы отсутствуют. Следовательно, чрезвычайно важно было выявить новые нейротрофные факторы, которые смогли бы обеспечить выживание других типов нервных и глиальных клеток.
Новые нейротрофные факторы были найдены по их способности поддерживать выживание в культуре нервных клеток, которые не являются чувствительными к NGF. Так были обнаружены факторы, которые способствуют выживаемости цилиарных ганглиозных моторных нейронов, которые инвертируют скелетные и гладкие мышцы (цилиарные ганглиозные нервные клетки принадлежат парасимпатической нервной системе и их выживаемость не поддерживается NGF).
Присутствие факторов, которые облегчают выживаемость цилиарных ганглиозных клеток, отмечено для множества тканей и видов. Эти цилиарные ганглиозные нефротрофные активности имеют следующие сходные химические и биологические свойства: (1) данная активность присутствует в высокой концентрации в седалищных нервах, (2) данная активность выдерживает присутствие ионного детергента SDS и редуцирующих агентов бета-меркаптоэтанола (BME) или дитиотрейтола (DTT) при электрофорезе в полиакриламидном геле и (3) на таких гелях эта активность мигрирует с кажущейся молекулярной массой между 24-28 кДа. Collins, 1985, Developmental, Bioligy, 109:255-258, Monthorpe et al/, 1986, Brain Pesearch, 367-282:286.
На основе этих сходных свойств предположили, что за цилиарную ганглиозную нейротрофную активность ответственны одни и те же близкородственные молекулы, обычно называемые "цилиарный нейротрофный фактор" или "CNTF".
Без достаточного количества данных нельзя утверждать, что белки, ответственные за эти активности, являются идентичными, поэтому CNTF обычно различают по происхождению (ткань и вид). Так, если вид, взятый в качестве источника - кролик, номенклатура - CNTF кроличьего седалищного нерва (кроличий SN-CNTF)/
SN-CNTF, по-видимому, находится в периферических нервах высочайших концентрациях. Он освобождается из клеток в нервы при травме. SN-SNTF поддерживает выживание и рост всех изученных нервных клеток периферической нервной системы, включая сенсорные, симпатические и парасимпатические нервные клетки. Таким образом, SN-CNTF оказывает действие на более широкий спектр нервных клеток, чем NGF. Как было недавно показано, крысиный SN-CMTF регулирует формирование специфических типов глиальных клеток в центральной нервной системе (Hughes et al., 1988, Nature 335:70).
Предполагается, что CNTF седалищного нерва - компонент ответа нервной системы на травму. SN-CNTF, освобождаемый из клеток в поврежденный нерв, как можно было ожидать, способствует выживанию и восстановлению роста поврежденных нервных клеток и регулирует функциональную активность глиальных клеток, необходимую для регенерации. Эти соображения показывают, что SN-CNTF может иметь терапевтическое значение при повреждении нервной системы, вызванном болезнью или травмой.
Несмотря на расширяющийся научный интерес к SN-CNTF, трудность выделения значительных количеств из натуральных источников и недоступность SN-CNTF человека затрудняют попытки четко продемонстрировать его значение в поддержании жизнеспособности нервных клеток. Предыдущие попытки очистить крысиный SN-CNTF привели к 800-кратному обогащению по сравнению с грубым экстрактом нервных клеток в отношении специфической активности Manthorpe et al., 1986, Brain Research 367:282-286.
Однако восьмиcоткратное увеличение специфической активности было недостаточно, чтобы получить белок одного вида. Термин "белки одного вида" используют здесь и в прилагаемой формуле исследования для обозначения полипептидов с одной с той же аминокислотной последовательностью во всех активных сайтах. Другими словами, если оперативная часть аминокислотной последовательности - одна и та же у двух или более полипептидов, они считаются здесь "белками одного вида", даже если имеется минорная гетерогенность в отношении длины или заряда.
Цель изобретения - получение SN-CNTF, очищенного до значительно большей степени, чем когда-нибудь достигнутая ранее, так, чтобы получить белок одного вида.
Еще одна цель изобретения - создание зондов для скрининга кДНК и геномной библиотек с целью клонирования животного и человеческого генов, кодирующих SN-CNTF.
Другая цель изобретения - получение последовательности ДНК для животного и человеческого CNTF.
Еще одна цель изобретения - обеспечение рекомбинантных экспрессионных систем, в которых нуклеотидная последовательность для человеческого или животного CNTF могла бы быть экспрессирована с получением человеческого или животного CNTF.
Эти и другие цели достигаются благодаря методу очистки SN-CNTF до такой степени, что специфическая активность увеличивается больше, чем в 25000 раз при сравнении экстракта и очищенного SN-CNTF, возможности анализа очищенного белка, получению на этой основе для скрининга кДНК и геномной библиотек SN-CNTF, клонированию и определению нуклеотидных последовательностей кроличьего и человеческого CNTF и использованию их в подходящей рекомбинантной экспрессионной системе с получением биологически активного SN-CNTF.
Следует понимать, что как предшествующее общее описание, так и последующее детальное описание, являются только примерными и не ограничивают данное изобретение. Сопровождающие рисунки, которые объединены и составляют одну из частей спецификации, иллюстрируют различные воплощения изобретения и вместе с этим описанием служат, чтобы объяснить принципы изобретения.
На фиг. 1 изображены примерные результаты хроматографии на колонке Моно Р; на фиг. 2 - примерное распределение нейротрофной активности в элюате каждого из семи участков, вырезанных из SDS-ПААГ геля после электрофореза; на фиг. 3 - примерные результаты обратнофазовой хроматографии; на фиг. 4 - примерные результаты прогона фракций на окрашенном серебром редуцирующем ДДС-ПААГ геле, эквивалентные активным и соседним с ними фракциям, показанным на фиг. 3; на фиг. 5 - профиль элюции пептидов после обработки эндопротеазой Asp-N и Lys-C; на фиг. 6 - профиль элюции пептидов после переваривания эндопротеазой Lys-C; на фиг. 7 - примерные результаты хроматографии фракций сульфата аммония на колонке фенил-Сефарозы HIC; на фиг. 8 - примерные результаты хроматографии на Mono P хроматосфокусированных на колонке алкил-Сефарозы FPLC-HIC фракций; на фиг. 9 - примерные результаты хроматографии препаративных SDS-ПЭГ фракций на C8 обратнофазовой HPLC колонке ((A) иллюстрирует результаты первоначальной процедуры очистки, (B) - результаты процедуры последовательной очистки после добавления еще двух HIC хроматографических стадий); на фиг. 10 - примерные результаты электрофореза SDS-ПААГ и Вестерн-анализа очищенного обратнофазовой хроматографией SN-CNTF (дорожка 1 в каждой из двух частей несет белки стандартного молекулярного веса (SIGMA SDS-7), дорожка 2 - очищенный SN-CNTF, (A) иллюстрирует результаты окраски серебром, (B) иллюстрирует результаты Вестерн-анализа с аффинно-очищенным антипептид-A- антителом); на фиг. 11 - нуклеотидная последовательность, кодирующая кроличий SN-CNTF (трансляция этой последовательности дает соответствующую аминокислотную последовательность, напечатанную внизу одиночным буквенным кодом, последовательности, которые подчеркнуты, подтверждены аминокислотной последовательностью, полученной при анализе белка SN-CNTF); на фиг. 12 - нуклеотидная и соответствующая ей аминокислотная последовательность (трехбуквенный код) для кодирующей последовательности человеческого SN-CNTF. Последовательности человека находятся между линиями. Там, где нуклеотидная или аминокислотная последовательности кролика отличаются от человеческих, они надписаны выше или ниже линий, соответственно; на фиг. 13 - конструкция вектора экспрессии рCMVXVPL 2; на фиг. 14 - методы, использованные для конструирования CNTF-syn 1/3 для экспрессии SN-CNTF (рисунок характерен, но не в том масштабе, см. пример 7 для экспериментальных деталей); на фиг. 15 отражены методы, использованные для конструирования CNTF-syn 1/3 для экспрессии CNTF (рисунок типичный, но не в масштабе, см. пример 7 для экспериментальных деталей); на фиг. 16 - синтетические нуклеотиды с 1 по 4, используемые в конструкциях CNTF-syn 1/3 и CNTF-syn 2/3; на фиг. 17 - синтетические нуклеотиды с 5 по 10, используемые в конструкции CNTF-syn 2/3; на фиг. 18 - некоторые характеристики бактериального экспрессионного вектора pT5T, содержащего инсерцию ДНК, обеспечивающую экспрессию CNTF (рисунок типичен, но не в масштабе, см. пример 7 для экспериментальных деталей); на фиг. 19 отражены определенные черты бактериального экспрессионного вектора pT3х1-2, содержащего инсерцию ДНК для экспрессии CNTF (рисунок типичен, но не в масштабе, см. пример 7 для экспериментальных деталей); на фиг. 20 - редуцирующий SDS-полиакриламидный гель, в котором были разогнаны и окрашены Coomassie Brilliant Blue экстракты клеток, трансформированных различными экспрессионными конструкциями; на фиг. 21 - редуцирующий SDS-ПААГ, в котором были разогнаны и выявлены иммуноблоттом с аффинно-очищенным анти-CNTF пептид А-антителом экстракты клеток, трансформированных различными экспрессионными конструкциями; на фиг. 22 отражен биоанализ последовательных разведений супернатантов из бактериальных клеток, экспрессирующих pT5T:CNTF-syn 1/3 или pT3x1-2: CNTF-syn 2/3 ( вставка отражает биоанализ экстрагированных кусочков из редуцирующего SDS-ПАА-геля рТ5Т:CNTF-syn 1/3 супернатанта в районе геля непосредственно выше и ниже 24 кДа).
Хотя изобретение касается выделения CNTF из седалищного нерва кроликов, оно пригодно для выделения и очистки этого белка из других источников.
Вкратце, одно предпочтительное воплощение предлагаемого метода включает использование в качестве сырья растертого в порошок материала седалищного нерва кролика. Грубый экстракт из этого материала центрифугируют, супернатант окисляют, а полученный преципитат отбрасывают. Потом титруют супернатант NaOH и полученный осадок снова удаляют центрифугированием.
После pH-преципитаций к супернатанту добавляют насыщенный раствор сульфата аммония и удаляют преципитат центрифугированием. При дальнейшем добавлении сульфата аммония к супернатанту происходит преципитация белковой фракции, содержащей большинство SN-CNTF активности.
Вышеописанный препарат затем наносят на хроматофокусирующую FPLC колонку MonoP. Колоночные фракции собирают и анализируют pH и CNTF-активность. Фракции (указанные на фиг. 1 стрелкой с вертикальной чертой) с пиком SN-CNTF активности затем дополнительно обрабатывают, как это будет обсуждено в деталях ниже.
Сфокусированные фракции из множества прогонов через колонку MonoP наносят для электрофореза на SDS-полиакриламидный гель. Район геля, соответствующий молекулярным весам с 22 по 27 кДа, разрезают по всей толщине геля на многочисленные полоски и элюируют электрофоретически. Элюированные белки собирают и фракцию с наибольшей активностью в дальнейшем очищают, используя обратнофазовую HPLC. Этот процесс более детально описан в примерах, которые следуют.
Целесообразно проводить дополнительные стадии, которые включаются в процедуру очистки для того, чтобы обеспечить удобную обработку материала на ранних стадиях. В предпочтительном воплощении между фракционированием сульфатом аммония и хроматофокусированием вставляют хроматографию на гидрофобном носителе (фенил-Сефарозе), а гидрофобную хроматографию на колонке FPLC алкил-Сефарозы вставляют между хроматофокусированием и препаративным ЛДС-ПААГ (пример 1).
Разработанный метод очистки привел к получению SN-CNTF в очищенной форме (с более чем 25000-кратным увеличением специфической активности по сравнению с грубым экстрактом).
Увеличенная степень очистки облегчает определение аминокислотной последовательности SN-CNTF. В соответствии с изобретением определена аминокислотная последовательность, размер которой оказался достаточным, чтобы создать олигонуклеотидные пробы, которые облегчают скрининг к ДНК и геномной библиотек для клонирования животного и человеческого генов, кодирующих SN-CNTF.
Как будет обсуждаться более подробно ниже, эти гены в свою очередь сделали возможным производство в большом объеме (1) животного SN-CNTF, подходящего для изучения его способности восстанавливать животные модели повреждения нервной системы и человеческого (3) SN-CNTF, пригодного для включения в фармацевтические составы, полезные в лечении повреждений нервной системы человека.
Такие пептиды можно использовать для приготовления антител. Антитела к синтетическим пептидам получены и показано, что они реагируют с очищенным CNTF при Вестерн блот анализе (фиг. 10).
Из вышеприведенной работы конечная цель - клонировать и экспрессировать ген SN-CNTF человека для того, чтобы приготовить материал, подходящий для использования в фармацевтических препаратах для человека. Как только геномная последовательность известна, гены, кодирующие SN-CNTF, могут затем быть экспрессированы в животных или бактериальных клетках. Поскольку последовательность гена, кодирующего CNTF кролика и человека, была определена, в раскрытой здесь технологии рекомбинатной ДНК для производства CNTF могли использоваться не только природные, но и синтетические последовательности ДНК, кодирующие SN-CNTF.
Метод получения рекомбинантного CNTF включает
а) приготовление последовательности ДНК, способной обеспечить в хозяйской клетке производство белка, имеющего CNTF-активность;
б) клонирование последовательности ДНК в вектор, способный к введению, репликации и экспрессии последовательности ДНК в хозяйской клетке;
в) перенос вектора, содержащего чужеродную последовательность ДНК и необходимые элементы в клетку хозяина, способную экспрессировать эту последовательность ДНК, кодирующую CNTF;
г) культивирование клеток хозяина при условиях, подходящих для амплификации вектора и экспрессии CNTF;
д) сбор CNTF и
e) создание условий для образования полипептидом третичной структуры, благодаря которой он обладает CNTF-активностью.
В одном из воплощений изобретения (частично или полностью) синтетическая последовательность ДНК, использованная для получения рекомбинантного CNTF, может содержать иные нуклеотиды, чем природная последовательность ДНК, о при этом будет все еще кодировать полипептид с той же первичной структурой, что и CNTF, кодируемый CNTF генами животных или человека.
В другом воплощении изобретения природная последовательность ДНК, кодирующая CNTF, модифицируется, чтобы усилить экспрессию в организме хозяина или клетке. Такие модификации могут включать следующее:
а) если нативная последовательность ДНК-геномная ДНК, то с целью последующей экспрессии в бактериальной системе может использоваться удаление интронов;
б) изменение нуклеотидной последовательности путем введения последовательностей, узнающихся различными ферментами рестрикции, для простоты последующих стадий лигирования, клонирования и мутагенеза;
в) изменение последовательности нуклеиновых кислот путем использования кодонов, предпочитаемых организмом хозяина, используемым для получения рекомбинантного белка;
г) соединение нуклеотидных последовательностей с оперативными элементами, необходимыми для поддержания и экспрессии ДНК в организме хозяина или клетке.
Подготовленная таким образом последовательность вставляется в экспрессионный вектор, способный к существованию в организме хозяина и направлению экспрессии CNTF.
Векторы E.coli
Векторы, предлагаемые для использования в изобретении, включают любые векторы, в которые, как обсуждалось выше, последовательность ДНК может быть введена вместе с любыми предпочтительными или требующимися оперативными элементами, и полученный вектор может затем быть последовательно введен в клетку хозяина и быть реплицирован в такой клетке. Предпочтительные векторы - те, чьи рестрикционные сайты хорошо известны и которые содержат оперативные элементы, предпочтительные или требуемые для транскрипции последовательности ДНК. Однако определенные приложения изобретения касаются и не полностью изученных в настоящее время векторов, содержащих одну или более описанных последовательностей ДНК. В частности, предпочтительно, чтобы все эти векторы имели некоторые или все следующие характеристики: (1) обладали минимальным количеством последовательностей организма-хозяина, (2) стабильно поддерживались и размножались в желаемом хозяине, (3) были способны присутствовать в большом числе копий в желаемом хозяине, (4) обладали регуляторным промотором, расположенным так, чтобы способствовать транскрипции интересующего гена, (5) имели по меньшей мере одну маркерную последовательность ДНК, кодирующую селективный признак, представленный в части плазмиды, отделенной от той, куда может быть инсерцирована последовательность ДНК, и (6) ДНК последовательности, способные к терминированию транскрипции.
В различных предпочтительных воплощениях изобретения эти клонирующие векторы, содержащие и способные экспрессировать последовательности ДНК изобретения, содержат различные оперативные элементы. Эти "оперативные элементы", как здесь обсуждается, включают по меньшей мере один промотор, по меньшей мере одну последовательность Шайн-Дельгарно и инициирующий кодон и по меньшей мере один терминаторный кодон. Желательно, чтобы эти "оперативные элементы" также содержали по крайней мере один из следующих: по меньшей мере один оператор, по меньшей мере одну лидерную последовательность для секреции белков, по меньшей мере один ген для регуляторного белка и также другие последовательности ДНК, необходимые или предпочтительные для свойственной им транскрипции и последующей трансляции векторной ДНК.
Определенные из этих оперативных элементов могут быть представлены в каждом из предпочтительных векторов изобретения. Любые другие дополнительные оперативные элементы, которые, возможно, потребуются, можно добавить к этим векторам, используя методы, известные всем, имеющим самые обычные знания в этой области, особенно в свете данного руководства.
Например, нужные последовательности ДНК лигируют в подходящие векторы, как описано у Maniatis и др., Molecular Cloning, Cold Spring Harbor Laboratories (1984), приводимой здесь в виде ссылки.
Векторы, подходящие для использования в микроорганизмах, других, чем E. coli, также рассматриваются в рамках этого изобретения. Такие векторы описаны в таблице. В дополнение к этому определенные предпочтительные векторы обсуждаются ниже.
б) Векторы Pseudomonas
Некоторые векторные плазмиды, которые автономно реплицируются в широком спектре Грамм-негативных бактерий, являются предпочтительными для использования как клонирующих челночных векторов в хозяевах с геномом Pseudomonas. Определенные из них описаны Tait, R.C., Close T.J. Lundguist, R.C., Hagiya, M. Rodriguez, R. L., и Kado, C.I. B.Biotechnology, May, 1983, pp. 269-275, Panopoulos, N. J. в Genetic Engineering in the Plant Sciences, Praeger Publishers, New York, pp. 163-185 (1981); и Sakagucki, K. в Current Toric in Microbiology and Immunology 96:31-45 (1982), каждый из которых специально объединен здесь в ссылках. Одна особенно предпочтительная конструкция обычно содержит плазмиду RSF1010 и ее производные, как описано Bagdasarian, M., Bagdasarian, M. M. , Coleman, S., и Timmis, K.N. в Plasmids of Medical, Environmental and Commercial Importance, Timmis, K.N и Puhler, A. eds, Elsevier/Noth Holland Biomedical Press (1979), специально внесенные здесь в ссылки. Преимущество RSF 1010 в том, что она относительно маленькая, высококопийная, которая легко трансформируется и стабильно поддерживается, как в E.coli, так и в видах Pseudomonas. В этой системе предпочтительно использовать Tac экспрессионную систему, как описано для Esherichia, поскольку оказалось, что trp промотор E. coli охотно узнается PHK-полимеразой Pseudomonas (Sakagucki, K. в Curent Toris in Microbiology and Immunology 96:31-45 (1982) и Gray, G.L., Mckeown, K.A., Jones, A.J.S. Seeburg, P.H., и Heyneker, H.L. в Biotechnology, Feb. 1984, pp. 161-165).
В тех случаях, где рестрикционные минус штаммы хозяина видов Pseudomonas не имеются в наличии, трансформационная эффективность плазмидных конструкций, выделенных из E.coli, невысокая. Следовательно, желателен пассаж клонирующего вектора Pseudomonas перед трансформацией желаемого хозяина через r-m+ штамм других видов (Bagdasarian, M., и др. Plasmids of Medical Environmental and Commercial Importance, pp. 411-422, Timmis и Puhler eds., Elsevier/North Holland Biomedical Press (1979).
в) Векторы Bacillus
Предпочтительная экспрессионная система с хозяином рода Bacillus включает использование плазмиды pUB-110 в качестве клонирующего челночного вектора. Как в других векторных системах, в Bacillus возможно экспрессировать CNTF изобретения внутриклеточно или в виде секреторного белка. Настоящее приложение включает обе системы. Челночные векторы, которые реплицируются, как в Bacillus, так и в E.coli, имеются в распоряжении для конструирования и тестирования различных генов, как описано Dubnau, D., Gryczan, T., Contente S., и Shivakumar, A.G., Genetic Engineering, Vol. 2, Setlow и Hollander eds. , Plenum Press, New York, pp. 115-131) (1980), специально внесенных здесь в виде ссылки. Для экспрессии и секреции CNTF в B. subtilis предпочтительно присоединяют к району ДНК, кодирующему белок, сигнальную последовательность альфа-амилазы. Для синтеза внутриклеточного CNTF последовательность ДНК можно присоединить с сохранением рамки к сайту связывания рибосом альфа-амилазной лидерной последовательности.
Транскрипция любой из этих конструкций предпочтительно направляется альфа-амилазным промотором или его производным. Это производное содержит сайт узнавания РНК-полимеразы нативного альфа-амилазного промотора, но также присоединяет и район lac оператора. Сходные гибридные промоторы, сконструированные из промотора гена пенициллиназы и оператора lac, как было показано, функционируют в хозяине Bacillus в регуляторной манере, как установлено Yansura, D. G. и Henner в Genetics and Biotechnology of Bacilli, Gansean A.T и Hoch, J.A., eds., Academic Press, pp. 249-263 (1984), специально внесенные в ссылки. Ген lac I E. coli может также быть включен в плазмиду для внесения регуляции.
г) Векторы Clostridium
Одна из предпочтительных конструкций для экспрессии в Clostridium - плазмида pSU12, описанная Squires, C.H. и др. в J. Bacteriol., 159: 465-471 (1984), трансформированная в C. pertringens методом Heefner, D.L. др., как описано в J. Bacteriol. 159: 460-464 (1984). Транскрипция направляется промотором гена устойчивости к тетрациклину. Трансляция определяется наличием последовательности Шайн-Дальгарно того же гена tetr.
д) Дрожжевые векторы
Сохранение чужеродной ДНК, введенной в дрожжи, может осуществляться несколькими способами, как описано Botstein, D. и Davis, R.W., в The Molecular Biology of the Yeast Saccharomyces, Cold Spring Harbor Laboratory, Strathern, Jones and Braach, eds, pp. 607-636 (1982), специально внесенные здесь в ссылки. Одна из предпочтительных экспрессионных систем для использования Saccharomyces в качестве хозяйского организма и гена CNTF 2 μм плазмида. Преимущества ее включают относительно высокое число копий и стабильность при введении в cir' штаммы. Эти векторы также могут включать начало репликации и по меньшей мере один маркер устойчивости к антибиотику из pBR322, чтобы разрешить репликацию и селекцию в E. coli. Двухмикронную последовательность и дрожжевой ген LEU2 обеспечивают использование в LEU2 дефектных мутантах дрожжей.
Если планируется, что рекомбинантный CNTF будет в конце концов экспрессироваться в дрожжах, предпочтительно, чтобы клонирующий вектор сначала был введен в Escherichia coli, где вектору позволили бы реплицироваться, после чего вектор трансформировали бы в дрожжи для окончательной экспрессии CNTF.
е) Клетки млекопитающих
В качестве последовательностей для экспрессии CNTF в клетках млекопитающих служат кДНК и геномная ДНК, кодирующие SN-CNTF.
Должна иметься последовательность, которая будет эффективно связываться рибосомами, как описано Kozak, в Nucleic Acids Resеarch 15: 8125-8132 (1987), специально вынесенным здесь в ссылки. Рестрикционный фрагмент ДНК, несущий последовательность ДНК, кодирующую CNTF, можно вставить в экспрессионный вектор, который уже имеет транскрипционный промотор и энхансер, как описано у Guarente, L. в Cell 52: 303-305 (1988) и Kadonada, J.T. и др. в Cell 51: 1079-1090 (1987). Промотор может быть регуляторным, как в плазмиде (Pharmacia Cat. N 27450601), если конструктивная экспрессия ингибитора вредна для роста клеток. Вектор должен иметь полный сигнал полиаденилирования, как описано у Ausubel, F.M. и др. в Current Protocols in Molecular Biology, Wiley (1987), чтобы мРНК, транскрибируемая с этого вектора, правильно процессировалась. Кроме того, вектору следует иметь начало репликации и по меньшей мере один маркер устойчивости к антибиотику из pBR 322, чтобы разрешить репликацию и селекцию в E. coli.
Для того, чтобы селектировать стабильную клеточную линию, которая продуцирует SN-CNTF, экспрессионный вектор может нести ген селектируемого маркера, такой, как маркер лекарственной устойчивости, или нести комплементарный ген для дефектной клеточной линии, например, ген дигидрофолятредуктазы (dhfr) для трансформации dhfr - клеточной линии, как описано у Ausubel и др. supra. Плазмида, несущая селектирующий маркер, может быть котрансформирована вместе с экспрессионным вектором.
Клетки хозяина/Трансформация
Вектор, полученный описанным образом, вводят в подходящую клетку-хозяина. Эти хозяйские клетки могут быть клетками микроорганизмов или клетками млекопитающих.
а) Микроорганизмы
Считается, что может быть выбран любой микроорганизм, имеющий способность принимать экзогенную ДНК и экспрессировать ее гены и прилежащие оперативные элементы. После того, как организм хозяина выбран, в хозяйский организм вводят вектор, используя обычные методы. Примеры таких методов можно найти в Advanced Bacterial Genetics by R.W.Davis et al., Cold Spring Harbor Press, Cold Spring Harbor, New York (1980). Предпочтительно, чтобы трансформация происходила при низкой температуре, так как температурная регуляция обсуждается как средство регулирования генной экспрессии через использование оперативных элементов, как указано выше. В другом воплощении, если в вектор введены осмотические регуляторы, изменение болевых концентраций во время трансформации требует предусмотреть подходящий контроль в чужеродных генах.
Предпочтительно, чтобы микроорганизм-хозяин был факультативный аэроб или анаэроб. Среди дрожжей целесообразно использовать Saccharomyces и особенно Saccharomyces cerevisiae. Среди бактерий предпочтительно являются роды Bacillus, Escherichia и Pseudomonas, особенно Bacillus subtilis и Escherichia coli. Другие хозяйские клетки перечислены в таблице.
б) Клетки млекопитающих
Вектор может быть введен в культивируемые клетки млекопитающих несколькими технологиями, такими, как кальцийфосфат: ДНК-копреципитация, электропорация или слияние протопластов. Предпочтительный метод - копреципитация с фосфатом кальция, как описано Ausubel и др., supra.
Существует много стабильных клеточных типов, которые легко трансформируются и способны транскрибировать, процессировать и транслировать последовательность ДНК и продуцировать белок SN-CNTP. Однако клеточные типы могут быть вариабельными по отношению к гликозилированию белков и пост-трансляционным модификациям аминокислотных остатков (или любым другим). Таким образом, идеальные клеточные типы - те, что продуцируют рекомбинантный CNTP, идентичный природной молекуле.
Клетки хозяина культивируют в условиях, подходящих для экспрессии CNTF. Эти условия в основном специфичны для клетки хозяина и легко определяются специалистом средней квалификации в этой области в свете опубликованной литературы, касающейся условий роста для таких клеток и рекомендаций, содержащихся здесь. Например, Bergey's Manual of Determinative Bacteriology 8th Ed. , Williams & Wilkins Company, Baltimore, Maryland содержат информацию об условиях культивирования бактерий. Подобная информация по культивированию дрожжей и клеток млекопитающих может быть получена из работы Pollack, R. Mammalian Cell Culture, Cold Spring Habor Laboratories (1975), специально внесенной здесь в виде ссылки.
В одном из воплощений клетки выращивают до высокой плотности в присутствии подходящих регуляторных условий, которые ингибируют экспрессию последовательности ДНК. Когда достигнута оптимальная клеточная плотность, условия среды изменяют до подходящих для экспрессии последовательности ДНК.
Рекомбинантный SN-CNTF может быть очищен после экспрессии в клетке хозяина или организма.
В предпочтительном воплощении изобретения CNTF присутствует в его биологически активном состоянии уже при извлечении бактериальных культур. В альтернативном предпочтительном воплощении создают условия, в которых экспрессированный CNTF принимают свою активную структуру (на соответствующей стадии процесса очистки).
Для очистки рекомбинантного белка используют некоторые комбинации следующих стадий: анион-обменную хроматографию (Mono-Q, Mono-S и/или DEAE-Сефарозу), гельпроникающую хроматографию (Сефароза), хроматофокусирование (Mono-P) и хроматографию, оснащенную на гидрофобном взаимодействии (окти- и/или фенил-Сефароза).
Следующие примеры приводятся для иллюстрации определенных предпочтительных воплощений изобретения и не ограничивают его. Все ссылки, данные в этих примерах, специально внесены здесь.
Пример 1. Приготовление белка.
Материалы
Седалищные нервы взрослых кроликов получены от Pel- Freez Biologicals, Rogers, Arkansas. Сульфат аммония (высокой чистоты) был закуплен у Schwartz /Mann Biotech. Cleveland ohio. Фенилметилсульфонилфторид (PMSF) эпсилон-аминокапроновая кислота, бензамидин, пепстатин, дитиотреитол (ДТТ), поли- L орнитин (Р3655) и 3-/4,5-диметилтиазол-2 yl) 2,5-дифенилтетразолия бромид (МТТ) получены от Sigma Chemical Co., St. Louis, Missouri. _→ MonoP хроматофокусирующие FPLC колонки получены из Pharmacia, Inc., Pisataway, New Jersey. C8 обратнофазовые HPLC колонки получены от Synchrom, Inc. Lafayette, Indiana. Ацетонитрил закуплен у J.T. Baker Chemicak Co., Phillipsburg, New Jersey. Трифтороуксусная кислота получена от Pierce Chemicals, Rockford, Illinoice. Эндопротеазы Asp-N и Lys-c получены от Boehringer Mannerheim Biochemicals, Indianapolis, Indiana. Фетальная телячья сыворотка закуплена у Hyclone Laboratories, Logan, Utah. Культурные среды и солевые растворы получены от Irvine Scientific, Santa Ana, California. Культуральные чашки получены от Costar, Cambridge, Massachusetts. Патоген-свободные фертильные эмбриональные яйца курицы для сугубо практического использования получены от Spafas, Roanoke, Illinois.
б) Анализ SN-CNTF
Культуры основных куриных эмбриональных цилиарных ганглиев готовили, как описано ранее (Collins, 1978, Develop. Biol. 65:50; Manthorpe et al., 1986, Develop. Brain Res. 25:191. Цилиарные ганглии удаляли из фертильных, патоген-свободных куриных яиц, которые инкубировали в течение 9 - 10 дней при 38oC во влажной атмосфере. Ганглии подвергали химическому разделению при обработке сначала сбалансированным солевым раствором Хенкса без двухвалентных катионов, содержащих 10 мМ HEPES буфер, pH 7,2 10 мин при 37oC, а затем обрабатывали раствором 0,125% бактотрипсина 1:250 (Difco, Detroit, Michigan) в сбалансированном солевом растворе Хенкса, модифицированного, как описано выше, 12 мин при 37oC. Трапсинизацию останавливали добавлением фетальной телячьей сыворотки до конечной концентрации 10%.
После этой обработки ганглии переносили в раствор, состоящий из модифицированной Дульбекко среды Игла без бикарбоната, содержащей 10%-ную фетальную телячью сыворотку и 10 мМ HEPES, pH 7,2 и механически разделяли пропусканием 10 раз через пастеровскую пипетку, конец которой был гладко обожжен и вытянут до диаметра, позволяющего за 2 с наполнить пипетку.
Разделенные ганглии помещали в чашку (для тканевых культур диаметром 100 мм) с культуральной средой (см. выше), дополненной 10%-ной фетальной телячьей сывороткой, 4 мМ глутамина, 60 мг/л пенициллина-G, 25 мМ HEPES, pH 7,2 (40 разделенных ганглиев на чашку) на 3 ч. Такая процедура выполнялась для того, чтобы разделить ненейронные клетки, которые прилипали к чашке, и нервные клетки, которые не прилипали. После трех часов неприлипшие нервные клетки собирали центрифугированием, ресуспендировали в культуральной среде и высевали по 50 мкл на лунку в половину лунок 96-луночных микротитровальных планшетов для культуральной среды (с плотностью примерно 1500 нервных клеток на лунку).
Микротитровальные лунки предварительно обрабатывали раствором полиорнитина (1 мг/мл) в 10 мМ борате натрия, pH 8,4 в течение ночи при 4oC, отмывали дистиллированной водой и высушивали на воздухе.
10 мкл последовательных разведений образца добавляли в каждую лунку и планшеты инкубировали 20 ч при 37oC во влажной атмосфере, содержащей 7,5% CO2, с целью анализа нейротрофной активности. Через 18 ч в каждую лунку добавляли 15 мкм раствора тетразолиевой краски МТТ (1,5 мкг/мл) в среде Дульбекко без бикарбоната, содержащей 10 мМ HEPES, pH 7,2, и культуры помещали обратно в инкубатор (37oC) на 4 ч. Затем добавляли 75 мкл раствора (6,7 мл 12М HCl на 1 л изопропанола) и содержимое каждой лунки гомогенизировали 30 раз, чтобы разрушить клетки и суспендировать краситель. Затем определяли поглощение при 570 нм по сравнению с 690 нм поправкой для каждой лунки, используя автоматический считыватель микротитровальных планшетов (Dynatech, Chantilly, Virginia). Поглощение содержимого лунок, которые не содержали какого-либо нейротрофного агента (негативные контроли), вычитали из поглощения образец-содержащих лунок. Окончательное поглощение пропорционально количеству живых клеток в каждой лунке, которые трактуют как нервные клетки, способные превращать краситель. Количество единиц нейротрофной активности определяли как эквивалентное разделение, которое дает 50% максимальной выживаемости нервных клеток. Активность в трофических единицах на 1 мл получена делением общего количества единиц активности на объем анализа (60 мкл).
Удельная активность определялась делением общей активности трофических единиц на количество белка, присутствующего в образце.
в) Очистка SN-CNTF
В конце каждой из следующих стадий препарат либо использовался немедленно, либо оставлялся при -70oC не более чем на одну неделю до использования.
Стадия 1. Приготовление грубого экстракта.
100 г (сырой вес) кроличьего седалищного нерва (около 300 нервов) оттаивали и растирали, используя политронный роторный гомогенизатор (Kinematica, Switzerland) 1 мин в 10 объемах (вес/об) воды, содержащей 10 мМ ЭДТА, 1 мМ эпсилон, аминокапроновую кислоту, 1 мМ бензамидин и 0,1 мМ PMSF, и центрифугировали при 140000 g 30 мин при 4oC. Супернатант фильтровали через стеклянный фильтр, чтобы удалить флотирующие липиды.
Стадия 2. Обработка кислотой и аммонием
Стадии центрифугирования, на которые ссылаются ниже, выполнялись при 17000 g 20 мин и все операции выполнялись при 4oC, если не оговорено особо. Грубый экстракт центрифугировали. Супернатант закисляли до pH 3,65 н. HCl и полученный преципитат удаляли центрифугированием. Супернатант титровали до pH 6,3 1 н. NaOH и полученный проципитат снова удаляли центрифугированием. К указанному выше супернатанту добавляли раствор сульфата аммония до 30% насыщения и преципитат удаляли центрифугированием. Дальнейшее добавление сульфата аммония к супернатанту для достижения 60% насыщения приводило к преципитации белковой фракции, содержащей большую часть SN-CNTF активности. Преципитат растворяли в 20 мМ хлористого натрия, pH 6,7, содержащего 1 мМ ЭДТА, 0,1 мМ PMSF и 0,1 мкМ пепстатин, чтобы довести концентрацию до 8 - 13 мг/мл.
Стадия 3. Хроматофокусирование
Вышеуказанный препарат диализовали в течение ночи против в 500 раз большего объема 10 мМ фосфата натрия, pH 6,7 с однократной сменой буфера и центрифугировали при 1400000 g 30 мин. Супернатант пропускали через 0,22 мкм (диаметр пор) нейлоновый фильтр и наносили в 3 приема (по 2 мл каждый раз) на MonoР хроматофокусирующую FPLC колонку (объемом 4 мл), уравновешенную 25 мМ трис-HCl буфером, pH 5,8. Колонку промывали тем же буфером до возвращения поглощения элюата при 280 нм к базовой линии. Образец затем хроматографировали с полибуфером, pH 4,0 (1 - 10 разведение РВ74 от Pharmacia).
Колоночные фракции собирали и анализировали pH и CNTF активность. Фиг. 1 показывает результаты хроматографии на MonoP в виде оптической плотности (О. Д.) при 280 нм. Наложенные изображения - pH и SN-CNTF активности, измеренные в каждой фракции. Фракции, отмеченные стрелкой с вертикальной чертой (с пиком SN-CNTF активности около pH 5) собирали и обрабатывали твердым сульфатом аммония до достижения 95% насыщения, а осадок собирали центрифугированием, ресуспендировали в растворе насыщенного сульфата аммония и снова центрифугировали, чтобы удалить полибуфер. Преципитат растворяли в достаточном количестве 10 мМ натрий фосфатного буфера, pH 6,7, чтобы достичь белковой концентрации 3 - 5 мг/л (упоминается, как "собранная фракция"). Типично 1 л первоначального грубого экстракта требует 8 отдельных прогонов на колонке.
Стадия 4. Препаративный электрофорез в геле с додецилсульфатом натрия (SDS)
Собранные фракции из множества прогонов сквозь MonoP колонку объединяли и диализовали против 100-кратного объема 10 мМ натрий фосфатного буфера, pH 6,7 в течение 8 ч с одной сменой буфера, затем прогоняли в 15%-ном редуцирующем ДДС полиакриламидном геле в соответствии с методом Лэмли, 1970. Каждый разрешающий гель имеет размеры 0,3 см в толщину, 14 см в высоту, 11,5 см в ширину. 5,5 мг белка наносили на каждый гель. Электрофорез проводился при 15oC и 40 мА/гель до тех пор, пока стандарт молекулярного веса (окрашенный) 20 кДа не достигал конца разрешающего геля.
Чтобы обнаружить кривизну индивидуальных белковых полос вдоль ширины пластины геля, гель покрывают листом нитроцеллюлозы (0,45 мкм размер пор в форме рулона, полученный от Millipore Corporation, Beadford, Massachusetts), предварительно смоченным в воде, 2 листами смоченной и 2 листами сухой хроматографической бумаги (3ММ Chr, полученная от Whatman, Hillsboro, Oregon), стеклянной чашкой и 500 мл стеклянной колбой для веса. Через 30 - 45 мин очертания геля отмечали на нитроцеллюлозной бумаге, используя водонерастворимый маркер. Бумагу отмывали 3 раза 10 мМ трис-HCl буфером, pH 8,0, содержащим 0,15 NaCl и 0,3% NP - 40 детергент, и затем окрашивали 15 - 30 мин 1: 1000 разведенным Kohinuor Rapidograph Ink (имеется в магазинах канцелярских принадлежностей) (в вышеуказанном буфере).
Первоначальный гель помещали на стеклянную чашку и выравнивали его контуры по окрашенной нитроцеллюлозной бумаге, под стеклом. Район геля, соответствующий молекулярным весам между 22 - 27 локализовали отметкой, в соответствии с окрашенными стандартами молекулярного веса (BRL, Bethesda, MD), прогнанными в узких дорожках на обоих концах каждого геля. Этот район вырезали по всей ширине геля в виде семи 2,5 мм параллельных кусочков, используя кривизну полос, обнаруженную окрашенной нитроцеллюлозной бумагой. Каждый отдельный кусочек геля разрезали на меньшие кусочки (2,5•2 мм) и белки элюировали электрофоретически в течение 6 ч в 1:1 разведении буфера Лэмли, используя электрофоретический концентратор (ISCO Lincoln, Nebraska). Элюированные белки собираются в объеме 0,2 мл. Фиг.2 представляет план распределения нейротрофной активности в элюате из каждого из семи кусочков (отмеченных в порядке уменьшения молекулярного веса). Фракция с самой высокой активностью (полоска d) в дальнейшем очищается с использованием обратно-фазовой HPLC.
Стадия 5. Обратно-фазовая HPLC.
Дитиотреитол (ДТТ) и 10%-ная трифторуксусная кислота (TFA) были добавлены к элюатам геля до достижения конечной концентрации 2 и 0,3% соответственно. Образец фильтровали через 0,22 мкм нейлоновый фильтр, загружали на C8 обратнофазовую HPLC колонку и элюировали H2O/0,1% TFA:ацетонитрил/0,1% TFA - градиент. Фракции собирали в силиконизированные пробирки, содержащие 5 мкл 0,4% Твина 20. Анализировали нейротрофную активность аликвот каждой фракции. Белковую концентрацию определяли поглощением при 215 нм и совмещали с распределением нейротрофной активности. Фракции с пиком SN-CNTF активности (фракции 37-40, фиг.3) собирали для секвенирования, как описано в примере 2. В отдельном препарате фракции, соседние с включающими пик CNTF активности, эквивалентные фракциям 36-44 на фиг.3, также анализировали на окрашиваемом серебром редуцирующем ДДС-ПААГ (фиг.4).
Выполняли также дополнительные хроматографические стадии. Эти стадии подтверждали чистоту вышеполученных препаратов CNTF.
Две дополнительные хроматографические стадии основаны на принципе хроматографии по гидрофобным взаимодействиям (HIC). Первая HIC стадия - условная хроматографическая процедура, вставленная после стадии 2.
Растворенный материал после осаждения сульфатом аммония дополнительно разбавляют 10 мМ натрий фосфатным буфером, pH 6,7 (Буфер А) до ионной силы (измеренной прибором проводимости), равной ионной силе буфера В, содержащего 0,3 М сульфат аммония и 5% изопропанол (Буфера А). Затем добавляют изопропанол к разбавленному образцу до конечной концентрации 5% и смесь наносят на колонку фенил-Сефарозы (Pharmaccia, Inc., Piscatanay, New Jersey), уравновешенную буфером А. Не более чем 3 мг образца белка наносят на 1 мл объема колонки. 1 л грубого экстракта седалищного нерва дает 50 мл вновь растворенного осадка, осажденного сульфатом аммония, который затем разбавляют до 70-100 мл, как выше, и наносят на 110 мл фенил-сефарозную колонку. С колонки элюируют последовательно, начиная с 3 объемов буфера В, затем, используя 2 объема буфера В, содержащего 50% этиленгликоль (Буфер С), промывают 5 объемами воды 18 мл фракций.
Фиг. 7 показывает результаты каждого такого хроматографического прогона. Профиль элюированных белков непрерывно измерялся при 280 нм (толстая линия). О.Д. - контуром совмещен профиль элюированной биоактивности SN-CNTF в каждой фракции (линия, связывающая x's), измеренной в анализе выживания цилиарных ганглиев. SN-CNTF биоактивность выходит из колонки во время элюции буфером C (указано стрелкой с вертикальной чертой на фиг.7). Колоночные фракции, содержащие большую часть биоактивности, собирали и концентрировали, диализуя под давлением и используя мембрану Amicon JM-10 (Amicon Division, W.R.Grace & Co., Danvers, MA) до приблизительно 1/10 первоначального объема, который обычно приводил к конечной концентрации белков 2,5-3,0 мг/мл. Концентрат диализовали 6 ч против 3 смен 55-кратного объема В. Диализованный материал пропускали через 0,2 мкм (диаметр пор) фильтр Acrodise (Gelman Scinces, Inc., Ann Arbor, MI) и загружали множественными нанесениями (по 2 мл) на хроматофокусирующую колонку, как описано выше.
Без этой HIC - колоночной стадии 1 л грубого экстракта седалищного нерва требовал 8 отдельных прогонов через MonoP хроматофокусирующую колонку, а с добавлением HIC - колоночной стадии 1 л грубого экстракта можно было бы обработать в одном хроматофокусирующем прогоне.
Вторая HIC стадия введена после оригинальной стадии 3: хроматофокусированная на MonoP. К каждому 1 мл хроматофокусируемого материала (3-5 мг/мл белка) добавляли 2 мл 50 мМ фосфатного буфера, pH 6,7, содержащего 15 М сульфат аммония (буфер Д). Смесь затем пропускали через 0,2 мкм (диаметр пор) фильтр Acrodise и нагружали (множественными нанесениями по 2 мл каждое) на алкил-Сефарозную колонку HR 10/10 (FPLC(Pharmacia), уравновешенную буфером Д. Колонку промывали буфером Д до возвращения поглощения элюента (О.Д.) при 280 нм к базовой линии. Затем колонку обрабатывали для элюции 60 мл линейным градиентом буфер Д-буфер Е (50 мМ фосфатный буфер, pH 6,7) и собирали 1 мл фракции.
Фиг. 8 иллюстрирует результаты одного такого FPLC-HIC колоночного прогона. Непрерывная линия представляет профиль элюированного белка (О.Д.280). Совмещенные с ним линии - участок SN-CNTF активности в каждой градиентной фракции. Фракции, содержащие биоактивность (указанные стрелкой с вертикальной чертой на рис.8), собирали и концентрировали в Центрикон-10 концентр. (Amicon) до 0,5 мл. Образец разбавляли добавлением 2 мл буфера В до верхнего предела и вновь концентрировали центрифугированием до конечного объема 0,5 мл. Разбавление и повторное концентрирование повторяли еще 2 раза и окончательно сконцентрированный образец прогоняли в редуцирующем SDS-15%-ном полиакриламидном препаративном геле, как описано выше, исключая то, что предшествующий диализ не обязателен.
Фиг. 9 сравнивает окончательную стадию очистки на обратнофазовой HPLC в первоначальной процедуре очистки (верхняя часть) и в процедуре очистки после добавления двух стадий HIC (нижняя часть). Каждая часть показывает профиль элюированных белков (толстая линия О.Д.280) и совмещенную SN-СNTF активность (связывающая линия x's). Как видно из рисунка, в образце, проведенном через обратнофазовую HPLC, гораздо меньше примесного белка. Приготовленный оригинальной процедурой, описанной выше, белок очищен до гомогенности. Преимущество новой процедуры очистки в том, что 8 л первоначального материала можно обрабатывать так же удобно как 1 л, используя оригинальную методику.
Пример 2. Секвенирование очищенного нейротрофного фактора.
Фракции с пиком SN-CNTF активности (37-40, фиг.3) собирали и концентрировали до 50 мкл под вакуумной испаряющей центрифугой. Концентрированный образец содержал 0,14% Твин 20. Его разбавляли 1%-ным бикарбонатом аммония до конечного объема 350 мкл и обрабатывали эндопротеазой Asp-N или эндопротеазой Zys-C в течение ночи при 37oC. Смесь концентрировали до приблизительно 50-100 мкл на вакуумной испаряющей центрифуге и загружали по 1 мл образца петлей на узкую Aquapore RР-300 C8 обратнофазовую HPLS колонку (Broronbee Labs), 2,1•220 мм, элюировали H2O/0,1% TFA ацетонитрил/0,1% TFA градиент. Пептидсодержащие фракции собирали вручную в эппендорфовские пробирки, основываясь на поглощении при 215 нм. Фиг.5 показывает профиль элюированных пептидов после переваривания эндопротеазой Asp-N (по поглощению при 215 нм). Фиг.6 показывает профиль элюированных пептидов после переваривания эндопротеазой Lys-C - после редуцирования и карбоксиметилирования. Аминокислотную последовательность белков определяли, используя Applied Biosystems газовофазовый белковый секвенатор.
Дополнительная аминокислотная последовательность была получена с расщепляющими ферментами трипсином и эндопротеазой Glu-C(Boehringer Mannheim Biochemicals, Indianapolis, Inc.). Эта дополнительная белковая последовательность позволила некоторые из аминокислотных последовательностей, указанных выше, объединить в большие пептиды (по перекрыванию аминокислот). Новые аминокислотные последовательности и те, что объединены с предыдущими последовательностями, даны внизу:
H-S-A-L-T-P-H-R-R-E
L-A-R-K-I-R-S-D-L-T-A-L-T-E-S-Y-V-K-H-Q-G-L-N-K-N-I-N-L-D-S-V-D-G-V-P-M-A
D-Q-Q-V-H-F-T-P-A-E-G
D-G-L-F-E-K-K-L-W-G-L-K-V-L-Q-E-L-S-H-W-T-V
D-L-R-V-I
Пример 3. Приготовление антител к нейротрофному фактору.
Антитела, которые реагируют с очищенным кроличьим SN-CNTF, полезны при скрининге библиотек для получения гена, который кодирует кроличий SN-CNTF. В дополнение антитела, которые нейтрализуют его биологическую активность, используют в интактных животных для того, чтобы определить биологическую роль этого нейротрофного фактора.
Для того, чтобы приготовить такие антитела, синтезируют соответствующие районам последовательности SN-CNTF, используя Applied Biosystems автоматический белковый сектезатор. Такие синтетические пептиды ковалентно связывают с белком-носителем гемоцианином. Конъюгированный пептид инъецируют морским свинкам с полным адъювантом Френда, большими дозами через 3 и 6 недель - с неполным адъювантом. Сывороточные образцы отбирают из каждой свинки и используют в Вестерн-блоте против очищенного SN-CNTF для того, чтобы определить, реагирует ли антитело в сыворотке с очищенным белком. Позитивные в Вестерн-анализе образцы в дальнейшем тестируются на способность нейтрализовать нейротрофную активность в биоанализе, используемом для очистки. Серапозитивные Вестерн- и нейтрализационном анализе в дальнейшем очищают следующим образом: (1) сыворотку адсорбируют белком-носителем для того, чтобы удалить антитела, полученные против этого белка, затем сыворотку повторно тестируют вышеуказанным анализом, (2) фракцию IgG антител очищают от сыворотки стандартными методами и повторно тестируют вышеуказанным анализом. Обе эти стадии обеспечат поликлональное антитело, чистота которого достаточна, чтобы быть использованным для скринирования экспрессионных библиотек при клонировании мРНК и гена SN-CNTF.
Антитела продуцировались в кроликах к синтетическому пептиду "А", соответствующему части аминокислотной последовательности кроличьего SN-CNTF, данной в примере 2 ( E-S-J-V-K-H-Q-G-L-N-K-N). Ниже в этом примере даны методы. Аффинно-очищенные антитела против синтетического пептида А (антипептид-А-антитела) готовили проведением антисыворотки иммунизированных кроликов через аффинную колонку, несущую ковалентно-связанный синтетический пептид А и затем элюировали связавшиеся антитела. Нефракционированная иммунная антисывортка давала титр Ca•105 в ELISA анализе (при использовании пептида А для покрывания лунок и сыворотки в конечном разведении 1:50 для Вестерн-блот-анализа). Аффинно-очищенное антипептид А антитело, приготовленное, как описано выше, использовали в Вестерн-блот-анализе в конечной концентрации 80 мг/мл.
Как антипептид А - антисыворотка, так и аффинно-очищенные антитела, взаимодействуют с очищенным кроличьим SN-CNTF. Преиммунная сыворотка не взаимодействовала с SN-CNTF при этих условиях. Аликвоты фракций пика CNTF на конечной стадии очистки обратнофазовой HPLC (Фракция # 46, фиг.9, часть В) прогоняли на двух отдельных дорожках, сопровождаемых дорожками, содержащими белки-маркеры молекулярного веса. Гель разрезали на две части, каждая из которых содержала одну дорожку очищенного CNTF и соседнюю белков-маркеров. Один из этих кусочков окрашивали серебром, чтобы локализовать белки Bio-Rad Laboratories, Richmond, CA), а другой исследовали Вестерн-блот-анализом (Towbin и др., 1979, Proc. Natl. Acad. Sci., USA. 76: 4350) на белки, которые реагируют с аффинно-очищенными антипептид А-антителами.
Левая часть дорожек на фиг. 10 показывает, что обе эти полосы узнаются и окрашиваются аффинно-очищенными антипептид А-антителами. Это узнавание специфично, так как неродственные маркерные белки на самой левой дорожке справа не узнаются антипептид А-антителами, хотя присутствуют в высоких концентрациях, как показано на левых окрашенных серебром дорожках (фиг. 10). Преиммунная сыворотка из того же самого кролика также не узнает две полосы очищенного СNTF. Эти результаты показывают, что существуют по меньшей мере две различные CNTF, которые отличаются на Ca 5000 Да по молекулярному весу на редуцирующем SDS-ПААГ.
Чтобы приготовить антипептид А-антитела, синтетический пептид А конъюгировали с гемоцианином (KLH), чтобы усилить его антигенность. Для конъюгации 1 мг пептида А и 1 мг KLH (Calbiochem, La jolla, CA) в 50%-ном глицероле растворяли в 0,5 мл PBS (20 мМ натрий фосфатный буфер, pH 7,4, содержащий 0,15 М NaCl). 10%-ный глютаральдегид добавляли по каплям при перемешивании до конечной концентрации 1% и оставляли реакцию стоять при комнатной температуре в течение ночи при перемешивании, затем разбавляли до 5 мл PBS. Конъюгационную смесь эмульгировали с полным адъювантом Френда 1:2 и инъецировали под кутикулу во многие дорзальные места (в спину подкожно) двум новозеландским белым кроликам (100 мкг пептида А на кролика. Три недели спустя каждый кролик получал дополнительно дозу в 50 мкг конъюгированного пептида А с неполным адъювантом Френда. С этого времени подобные поддерживающие инъекции проводились с 2-недельными интервалами до тех пор, пока антисыворотка не давала титр по меньшей мере 100000 в ELISA анализе (Tainer и др. 1984, Nature 312. 127) при использовании покрытых пептидов лунок. Сыворотку готовили из крови, отобранной из ушной вены 5 недель спустя после первой инъекции и через каждые две недели после. Сыворотку хранили при -70oC.
Чтобы приготовить пептид-аффинную колонку, пептид А ковалентно присоединяли к матриксу хроматографической колонки следующим образом: к 3 мг пептида А, растворенного в 0,4 мл BBS, содержащего 4 М гуанидин гидрохлорид, добавляли 4,5 мл 0,1 М NaHCO3, pH 8,0 и 0,5 М NaCl. Один грамм лиофилизированной активированной СН Сефарозы 4В (Pharmacia) промывали, и она набухала в 200 мл 1 мМ HCl и непосредственно переносилась в раствор пептида А. Смесь качали в течение ночи при 4oC. Затем гель осаждали в центрифуге и супернатант сохраняли для определения количества пептида А, который остался связанным с матриксом. 15 мл 0,1 М трис-буфера, pH 8,0, добавляли к осадку геля и инкубировали при комнатной температуре 2 ч, чтобы блокировать непрореагировавшие спаривающиеся группы матрикса геля. Затем гель помещали в колонку (3 мл) и промывали три раза следующей последовательностью буферов: (1) 10 объемов 0,1 М ацетатного буфера, pH 4,0, содержащего 0,5 М NaCl, (2) 0,1 М трис-буфера, pH 8,0, содержащего 0,5 М NaCl. В конце концов колонку уравновешивали PBNS, содержащим 0,02% азид натрия. Различие в концентрации свободных аминных групп определяли в растворе первоначального пептида А и в супернатанте после конъюгации, используя флюоресцамин (Chen и др., 1978, Arch. Biochem. Biophys, 189-241; Nowicki, 1979. Anal. Letters 12: 1019). Этот анализ показал, что 92-95% пептида конъюгировало с матриксом сефарозного геля.
Перед аффинной очисткой антипептид А антитела 8 мл сыворотки иммунизированного кролика диализовали в течение ночи против 2 л PBS. Пептид А-сефарозную колонку промывали последовательно 10 объемами каждого из следующих растворов: 0,1 М глицин-HCl, pH 2,5, PBS 0,1 М триэтиламин, pH 11,5; PBS. Диализованную сыворотку пропускают через колонку три раза, чтобы добиться полного связывания антипептид А-антител. Колонку промывают 20 объемами PBS, затем элюируют последовательно 4 объемами каждого из следующих растворов: 0,1 М глицин-HCl, pH 2,5; PBS; 0,1 М триэтиламин, pH 11,5; затем PBS, 1 мл фракции собирают. Элюаты из глицинового и триэтиламинового промывов нейтрализуют сразу же 1 М трис, pH 9 и 7 соответственно и аликвоты анализируют на антипептид А-антитело ELISA анализом, используя покрытые пептидом А-лунки. Фракции с самым высоким титром (обычно в пределах 3 объемов начала глициновой и триэтиламиновой элюций) собирали и диализовали против PBS. После удаления части материала быстрым центрифугированием аффинно-очищенный антипептид А-антительный супернатант сохраняют при -70oC.
Пример 4. Клонирование гена
Учитывая возможность того, что кроличья и человеческая последовательности SN-CNTF неидентичны, целесообразным представляется сначала получить клоны гена кролика через гибридизацию с синтетическими олигонуклеотидами, основанными на белковой последовательности, а затем использовать клоны кролика как гибридизационные клоны в скрининге гена человека.
А. SN-СNTF ген
Чтобы получить геномную последовательность кролика, кодирующую SN-СNTF геномную библиотеку кролика (Clontech) переводили в бактериальный штамм E. coli nm 58 и выявляли при этом приблизительно 1000000 рекомбинантных клонов. Районы белковой последовательности кроличьего SN-СNTF, которые могут быть представлены самым незначительным числом кодонов, обратно транслируют и синтезируют соответствующие дегенеративные олигонуклеотидные пробы. Кроличьи олигонуклеотиды метят киназами в соответствии со стандартным протоколом Maniatis и др. (1982, Molecular Cloning, A Laboratory manual, cold Spring Harbor Laboratory). ДНК киназа получена от US Biochemical Corp. и гамма-меченый АТФ получен от ICN. Олигонуклеотиды метят со специфической активностью по меньшей мере 1000000 стр. на пикомоль.
При рассеве геномной библиотеки приблизительно 1 миллион бляшек переносятся на нитроцеллюлозные фильтры-дубликаты. Затем фильтры обрабатывают в соответствии с методами Manuatis и др. (1982, ibid) и гибридизируют течение ночи с радиоактивно-меченой олигонуклеотидной пробой. Гибридизационная смесь включает 6 • SSCр, 2х Денхард, 0,05% пирофосфат натрия, 1 мМ ЭДТА, 0,1% SDS, 100 мкг дрожжевой тРНК (Sigma), pH 8,0. Температура гибридизации должна быть на несколько градусов ниже измеренной (вычисленной) Tm олигонуклеотида. Клоны, которые гибридизуются с несколькими пробами, основанными на разных районах последовательности белка, рассеивают до отдельных колоний, и районы гибридизации секвенируют методом дидезокси-обрыва. (Sanger и др., 1977, Proc. Natl. Acad Sci 74: 5463), используя секвеназу (US Biochеmicals Corp.) для идентификации клонов, которые кодируют SN-CNTF белковую последовательность.
Б. SN-CNTF мРНК последовательности
Тотальную клеточную РНК получают из кроличьих и человеческих седалищных нервов. Ткань гомогенизируют в гуанидин тиоционат/бета-меркаптоэтанол растворе и РНК очищают седиментацией в градиенте хлористого цезия (Glison и др., 1974, Biochemistry 13: 2633).
Полиаденилированную РНК отбирают хроматографией на олиго (дТ) целлюлозе (Avid и Leder, 1972, Proc. Natl. Acad. Sci 69: 408). Количественный РНК блот гибридизационный анализ можно выполнять с "антисмысловыми" олигонуклеотидными пробами, чтобы оценить превалирование SN-CNTF последовательности в каждом РНК-препарате и этим оценить количество независимых клонов, необходимых для скрининга (по меньшей мере с 99% вероятностью) получающихся клонов CNTF. Достаточное количество двухцепочечной комплементарной ДНК можно синтезировать, как описано Gubler и Hoffman, 1983, Gene 25-263 и вставить в вектор лямбда gem (Promega Biontech) в соответствии с Palazzolo и Meyerowitz, 1987, Gene 52: 197.
Клоны, кодирующие кроличий SN-CNTF, идентифицируют гибридизацией рекомбинантных фаговых бляшек, как описано выше. Идентификацию этих клонов проверяют определением нуклеотидных последовательностей для того, чтобы установить соответствие с полностью известной белковой последовательностью. Скрининг с кДНК-пробами человеческого седалищного нерва (Feinberg и vogelstein, 1983, Anal Biochem 132: 6), представляющий собой более надежную процедуру для детекции перекрестной гибридизации, чем использование меньших олигонуклеотидов, использовали для скрининга кроличьих кДНК библиотек.
Полимерная цепная реакция (PCR) (Saiki и др., 1988 Science 239: 487) использовалась для амплификации фрагментов ДНК, соответствующих последовательностям аминокислот кроличьего CNTF. Такие фрагменты ДНК амплифицировали из человеческой и кроличьей геномной ДНК и кДНК седалищного нерва кролика и симпатического ганглия. Амплифицированные фрагменты ДНК субклонировали и секвенировали, используя стандартные технологии (Maniatis и др., 1982, Molecular Cloning: A Laboratory manual, Cold Spring Harbor, New York). Фрагменты ДНК, полученные PCR, также использовались как пробы для скрининга кДНК-библиотеки седалищного нерва кролика и геномной ДНК-библиотеки человека. Позитивные клоны кДНК-кролика и позитивные геномные клоны человека выделяли и частично секвенировали. Последовательность открытой рамки, соответствующая кодирующей (эквивалентной мРНК) последовательности для CNTF кролика или человека, подтвердила последовательность фрагментов ДНК кодирующего района, полученных с использованием PCR. Результирующие кодирующие последовательности для SN-CNTF кролика и человека даны на фиг. 11 и 12 соответственно.
Части аминокислотной последовательности, полученные из кроличьего SN-CNTF, обратно транслировали в дегенеративные олигонуклеотиды #'S 1, 13, 7 и 12 и им комплементарные #'S 3, 14, 8 и 17. Аминокислотная последовательность (сверху) и локализация и нумерация соответствующих смыслового и антисмыслового вырожденных олигонуклеотидов (внизу) дана ниже
Y-V-K-H-Q-G-L-N-K-N-I-N-L-D-S-V-D-G-V-P-M-A
(5') ****1****** ****13***** *****7*****(3')
(3') ****3****** ****14***** *****8*****(5')
K-L-W-G-L-K-V-L-Q-E-L-S
(5') ****12*****(3')
Нуклеотидная последовательность смысловой версии каждого из вырожденных олигонуклеотидов дана ниже (где N соответствует любому нуклеотиду)
(3') ****17*****(5')
#1 5'-TA(T/C) GTN AA (A/G) CA(T/C) CA(A/G) GG-3'
#13 5'-AA(T/C) AA(A/G) AA(T/C) AT(A/T/C) AA (T/C) (C/T)T-3'
#7 5'-GA(T/C) GGN GTN CCN ATG GC-3'
#12A 5'-AA(A/C) TT(A/G) TGG GGN TT(A/C) AA-3'
#12B 5'-AA(A/G) TT(A/G) TGG GGN CTN AA-3'
#12C 5'-AA(A/C) CTN TGG GGN TT(A/C) AA-3'
#12D 5'-AA(A/G) CTN TGG GGN CTN AA-3'
Отдельные полимеразные цепные реакции выполнялись при использовании либо человеческой, либо кроличьей геномной ДНК в качестве образца и олигонуклеотидов # 'S1 и 8 или #'S1 и 17 в качестве праймеров для того, чтобы амплифицировать соответствующие районы генов CNTF человека и кролика. Саузерн-блоты этих продуктов реакции (обработанных олигонуклеотидом # 13) обнаружили существование меченых полос размером 66 пн (#'S1 и 8) и размером 366 пн (#S1 и 17).
Те же PCR-реакции, описанные выше, проводили, используя кДНК, приготовленное из либо мРНК кроличьего седалищного нерва, либо мРНК кроличьего симпатического ганглия. РНК готовили из кроличьих седалищных нервов или симпатических ганглиев и пропускали через олиго-дТ колонку, чтобы собрать матричную РНК (мРНК), как описано выше. Комплементарную ДНК (кДНК) готовили с помощью обратной транскриптазы, используя мРНК как образец и олиго-дТ как праймер. При проведении PCR использовали как праймер, либо кДНК, либо олигонуклеотиды # 'S1 и 8 или #'S1 и 17 и амплифицировали те фрагменты, которые имели ту же последовательность, что и амплифицированные с геномной ДНК кролика (фиг. 11). Это указывает на то, что в кодирующем районе гена CMTF между олигонуклеотидами #'S1 и 17 не существует интронов.
Чтобы получить больше кодирующей последовательности (эквивалентной матричной РНК) CNTF кролика, использовали дополнительную стратегию: двухцепочечную кДНК готовили, используя мРНК кроличьего седалищного нерва, как образец и олиго-дТ Not I линкерный адаптор в качестве праймера. Впоследствии добавляли EcoR I/XmnI линкерный адаптор (5'-ААТТЦГААЦЦПЦТТЦГ-3') к 5'-концу двухцепочечной кДНК лигированием по тупым концам (Maniatis, et al., ibid). Полимеразную цепную реакцию выполняли, используя такую кДНК в качестве образца и олигонуклеотиды #8 и EcoR I/XmnI линкер-адаптер, как праймеры. Саузерн блот-продуктов реакции (обработанных меченым олигонуклеотидом #13) обнаружил существование меченой полосы размером приблизительно 200 пн.
Чтобы получить кДНК клоны кроличьего CNTF, кДНК библиотеку готовили из поли(А)+мРНК седалищного нерва кролика методом, описанным выше, за исключением того, что использовали лямбда-gt-вектор (Stratigene) лямбда-gem 2. Приблизительно 4 • 106 бляшек такой библиотеки скринировали, используя пробу, приготовленную случайным мечением субклона M13 PCR фрагмента, полученного использованием кДНК кроличьего симпатического ганглия в качестве образца и олигонуклеотида #8 и EcoR I/XmnI- линкера адаптора как праймеров (см. выше). Отдельной первичной позитивной считалась бляшка, очищенная третичным скринингом. При переваривании EcoR I ДНК этого клона давала три фрагмента в дополнение к фрагментам лямбда: 2,0, 1,5 и 0,6 тпн длиной. Саузерн блот-анализом 1,5 тпн фрагмента показано, что он гибридизуется с другими CNTF - специфическими олигонуклеотидами и фрагментами, указанными выше. Последовательность ДНК этого 1,5 тпн и кДНК фрагмента показала, что он соответствует полной кодирующей последовательности кроличьего SN-CNTF (фиг. 11).
Чтобы получить клоны геномной ДНК для человеческого CNTF, скринировали приблизительно 3 • 106 бляшек геномной ДНК-библиотеки человека в векторе лямбда EMBL 3, используя пробу, приготовленную случайным мечением M13 субклона PCR фрагмента, полученного из геномной ДНК человека в качестве матрицы и олигонуклеотидов #1 и #17 в качестве праймеров (см. выше). Шесть позитивных бляшек, очищенных последовательным скринингом, гибридизировались с дополнительными CNTF-специфическими олигонуклеотидами и PCR-фрагментами, 0,6 тпн Bam HI рестрикционный фрагмент из одного из этих субклонов, гибридизирующихся с олигом #13, был субклонирован в разрезанную Bam HI H13 mp 19 и секвенирован.
Последовательности ДНК-фрагментов, полученных PCR из кроличьего кДНК клона, комбинировали, основываясь на районах перекрывающейся последовательности, чтобы получить кодирующую последовательность (эквивалентную мРНК) для кролика, представленную на фиг. 11. Последовательности ДНК-фрагментов, полученных PCR с человеческой геномной ДНК и с геномных клонов человека, соединяли, основываясь на районах перекрывающейся последовательности, чтобы получить кодирующую последовательность для человеческого SN-CNTF, представленную на фиг. 12. Нуклеотидные последовательности CNTF кролика и человека являются приблизительно на 89% идентичными (фиг. 12), указывая на то, что последовательности кролика и человека происходят из гомологичных генов, кодирующих CNTF. Как показано на фиг. 11, части нуклеотидной последовательности для CNTF кролика подтверждаются аминокислотными последовательностями, полученными из очищенного SN-CNTF, и на которые ссылались в предыдущих примерах.
Полимеразную цепную реакцию выполняли, используя матрицы и праймеры, описанные выше. Программа реакций была следующая: денатурирующий цикл, 1 мин при 95oC, цикл отжига, 1,5 мин при 40oC и цикл наращивания цепи, 4 мин при 72oC. Реакцию проводили за 30 циклов. Продукты реакции наносили на 2% агарозные гели и переносили на мембраны Leta-Bind (Bio-Rad. Richmond, CA) для Саузерн-анализа. Для того, чтобы идентифицировать амплифицированные порции кодирующей последовательности CNTF, Саузерн-блоты обрабатывали меченым олигонуклеотидом #13, о котором известно, что он лежит между олигонуклеотидами, используемыми для праймирования реакции в последовательности белка CNTF. Меченые полосы, полученные после Саузерн-блота, вырезали из первоначального геля и готовили для клонирования достраиванием концов фрагментом Кленова ДНК-полимеразы (New England Biolabs, Beverly, MA) в присутствии всех четырех дНТФ и обработкой ДНК-полинуклеотидкиназой T4 (US Biochemical Corp., Cleveland, OH) и АТФ. Подходящие кусочки ДНК затем субклонировали в вектор M13mp10, разрезанный SmaI (дефосфорилированный, коммерческий препарат от Amersham Corp. , Arlington Heights, IL). Рекомбинантные фаги, содержащие интересующий фрагмент, идентифицировали процедурой скрининга Benton и Davis (1977, Science 196: 180), используя меченый олигонуклеотид #13 в качестве пробы. Эти рекомбинантные клоны выращивали до получения достаточного для секвенирования количества одноцепочечной ДНК и затем секвенировали методом дидезокси-обрыва цепи (Sanger и др., ibid).
Когда гибридизацию проводили длительно, используемые меченые пробы ДНК включали 5 • SSCP, 2х раствор Денхардта, 2 мМ ЭДТА, 0,05%-ный пирофосфат натрия, 0,1%-ный додецил сульфат натрия (SDS), 250 мкг/мл ДНК спермы сельди (неспецифический конкурент за связывание) pH 8,0. Гибридизацию проводили при 65oC и блоты или фильтры отмывали при 65oC в 0,1 • SSCP и 0,1% SDS. При коротких сроках гибридизации олигонуклеотидные пробы состояли из 6 • SSCP 2х раствора Денхардта, 2мМ ЭДТА, 0,05% пирофосфата натрия, 0,1% SDS, 100 мкг/мл тРНК дрожжей (неспецифический конкурент за связывание), pH 8,0, температуру гибридизации и условия отмывки блотов и фильтров приспосабливали индивидуально в зависимости от ГЦ-содержания каждого олигонуклеотида (Maniatis и др., ibid).
Пример 5. Экспрессия генов, кодирующих SN-CNTF в клетках животных.
Экспрессия SN-CNTF в культивируемых клетках животных требует проведения следующих стадий:
а) Конструирование экспрессионного вектора;
б) Выбор линии клеток хозяина;
в) Введение экспрессионного вектора в клетки хозяина;
г) Манипуляции с рекомбинантными клетками-хозяевами, чтобы увеличить уровни экспрессии SN-CNTF.
а) SN-CNTF экспрессионные векторы, предназначенные для использования в животных клетках, могут быть нескольких типов, включая строго конститутивные экспрессионные конструкции, индуцибельные генные конструкции, так же, как предназначенные для определенных клеточных типов. Во всех случаях промоторы и другие регуляторные районы гена, такие, как энхансеры (индуцибельные или нет) и сигналы полиаденилирования, помещаются в подходящее положение по отношению к последовательности кДНК в газовых плазмидных векторах. Два примера таких конструкций следуют далее. (1) Конструкцию, использующую район строго конститутивного промотора, делают, используя сигналы генетического контроля SV40 в таком расположении, как в плазмиде pSV2CAT, как описано Gormah и др., в Mol. Cel. Biol. 2: 1044-1051, 1982, специально вынесенных здесь в виде ссылки. Этой плазмидой следует манипулировать таким образом, чтобы заменить кодирующую последовательность хлорамфенилколацетилтрансферазы (САТ) SN-CNTF кДНК, используя стандартные молекулярно биологические технологии (Maniatis и др. , supra). (2) Индуцибельную генетическую конструкцию делают, используя плазмидную РМК, которая содержит промоторный район металлотионеина мыши (МТ-1) (Brinster и др., Cell 27: 228 - 231, 1981).
Эту плазмиду можно использовать как начальный материал и ей следует манипулировать, чтобы получить металл-индуцируемую генную конструкцию.
б) Можно использовать различные клеточные линии, чтобы экспрессировать SN-CNTF, работая с векторами, описанными выше для продуцирования активного белка. Две потенциальные клеточные линии, которые хорошо охарактеризованы по их способности усиливать экспрессию чужеродного гена - мышиная линия Ztk- и линия клеток яичников китайского хомячка (CHO) dhfr-, однако экспрессия SN-CNTF не лимитируется этим выбором.
Линии животных клеток, которые могут также использоваться для экспрессии, включает клетки почек мартышки COS-7, которые полезны для транзитной экспрессии, и человеческие эмбриональные клетки почек 293.
в) Векторную ДНК следует вводить в эти клеточные линии, используя любую из известных технологий. Метод, используемый здесь, включает технологию кальцийфосфатной ДНК-преципитации, описанную S.Z. Graham и A.S. van der Eb (Virology 52: 456-467, 1973), в которой экспрессионный вектор, кодирующий SN-CNTF, соосаждается со вторым экспрессионным вектором, кодирующим селектируемый маркер. В случае Ztk - клеточной трансфекции в качестве селектируемого маркера выступает ген тимидинкиназы и селекцию проводят, как описано Wigler и др., в Cell 16; L 777-785, 1979, в случае CHOdhfr-клеток селектируемый маркер-дигидрофолятредуктаза (DHFR) с селекцией, как описано Ringold и др. в J. Mol. Appl. Genet. 1: 165-175, 1981.
г) Клетки, которые экспрессируют генные конструкции, выращивают при условиях, которые будут увеличивать уровни продукции SN-CNTF. Клетки, несущие конструкции с металлотионеиновым промотором, можно выращивать в присутствии тяжелых металлов, таких, как кадмий, который будет приводить к 5-кратному увеличению использования MT-1 промотора (Mayo и др., Cell 29:99-108) и соответственно, к сравнимому увеличению в уровнях продукции белка SN-CNTF. Клетки, содержащие SN-CNTF экспрессионные векторы (либо SV40 - либо, MT-1-производные), вместе с DHFR-экспрессионным вектором можно подвергать генной амплификации по протоколу, описанному Ringold и др. в J. Mol. Appl. Genet. 1. 165-175, 1981, используя метотрексат, конкурентный антагонист DHFR. Это приводит к большему числу копий генов DHFR, присутствующих в этих клетках, и одновременно увеличению копий генов SN-CNTF, которые в свою очередь могут приводить к большему количеству SN-CNTF белка, продуцируемому клетками.
Дополнительный экспрессионный вектор pCMVXVPL2 использовали, чтобы экспрессировать кодирующую последовательность кроличьего CNTF за короткое время в COS-7 клетках. Этот плазмидный вектор содержит ранний промотор и энхансер, цитомегаловируса (CMV) (Boshart и др. CM 41:521-530, 1985).
Эту плазмиду можно сконструировать, как показано на рис. 13. Сигнал полиаденилирования поставляется SV40 последовательностью и координатами карты 2589-2452, см. Reddy и др., Science 200: 494-502, 1978. Начало репликации SV40 включается в эту плазмиду для облегчения ее использования в COS-клетках для быстрых экспрессионных анализов.
Кроличий SN-CNTF быстро экспрессировался в COS-7 клетках следующим образом: 1,5 тпн EcoRI фрагмент кДНК клона кроличьего седалищного нерва, содержащего полный кодирующий район для кроличьего SN-CNTF (пример 4), субклонировали в рестрицированный EcoRI вектор pCMVXPL2. Отобрали одиночный клон, который после переваривания SacI и Bam HI дает рестрикционные фрагменты, соответствующие по размеру предсказанным для инсерции в вектор фрагмента 1,5 тпн в правильной ориентации. Плазмидную ДНК из этой конструкции готовили методом щелочного лизиса, за которым следовало центрифугирование в градиенте плотности CsCl (Maniatis и др., ibid). Эту ДНК вводили в COS-7 клетки методом Sompayrac and Dama (Proc. Natl. Acad. Sci, USA 78: 7575-7578, 1981). Как контроль эквивалентные COS клеточные культуры трансфецировали плазмидной векторной ДНК без инсерции.
Через 48 ч после трансфекции ночную среду и осадок клеток собирали. Осадок клеток экстрагировали быстрым разрушением ультразвуком во льду в 20 мМ фосфате натрия, pH 6,7, содержащем 1 мМ ЭДТА, 0,1 мМ PMSF и 0,1 мкМ пепсатин. Серии разведений как клеточного экстракта, так и ночной среды из каждой культуры, анализировали на активность в анализе выживания цилиарных ганглиев.
Клеточные экстракты из культур, трансфецированных вектором, содержащим кДНК фрагмент CNTF, имели титр Ca 15000 Tu/мл в биоанализе и приблизительно 50 нг/мл CNTF, как определено Вестерн-блот-анализом. Ни клеточные экстракты из культур, трансфецированных одним вектором, ни среда из любых культур не показывали какую-либо детектируемую биоактивность CNTF белка или наличие его в Вестерн-блот-анализе. Этот результат ясно демонстрирует, что CNTF кДНК, которую мы клонировали, кодирует белок с биоактивностью подлинного SN-CNTF.
Пример 6. Выделение SN-CNTF из рекомбинантных животных клеток.
Так как SN-CNTF синтезируется хозяйскими клетками в виде, подобном натуральному материалу, методы, описанные выше для очистки природного белка, применимы для очистки и характеристики рекомбинантной формы. Для квалифицированных в этой области очевидно, что различные модификации и вариации могут быть допущены в процессах и продуктах изобретения в зависимости от ситуации.
Пример 7. Продукция рекомбинантного человеческого CNTF.
В одном из воплощений изобретения для производства рекомбинантного человеческого CNTF использовались бактерии Escherichia coli. При этом были применены два альтернативных метода для конструирования ДНК, которую надо экспрессировать, и два различных экспрессионных вектора. Все эти вариантные системы экспрессии продуцируют растворимый биологически активный белок CNTF с высоким выходом в бактериальном клеточном экстракте. Методы получения этих продуцирующих систем описаны ниже. Следовательно, положение характерной особенности какой-нибудь цифры, данное в скобках, например 233, обозначает номер пары в кодирующей последовательности CNTF человека (фиг. 12), с которой начинается какая-либо особенность или черта, по ходу транскрипции от A (1) в инициаторном кодоне ATG.
1. Приготовление ДНК для экспрессии CNTF.
Стратегия I для конструирования 5' конца (фиг. 14):
Клон геномной ДНК человека для CNTF в фаге лямбда EMBL3 из примера 4 переваривали рестрикционными ферментами Sal I и Hind III и 4,3 тпн фрагмент очищали электрофорезом в геле. Он содержит кодирующую последовательность CNTF выше Hind III сайта (233). Этот 4,3-тпн фрагмент также содержит одиночный, приблизительно 1,3-тпн интрон (114-115) в кодирующей последовательности. Чтобы разрешить экспрессию в бактериальных клетках, этот интрон удаляли сайт-направленным мутагенезом in vitro, используя синтетический олигонуклеотид, как описано G.A. Mc Clary, F. Whitney, G. Geiselsoder (1989) Biotechnigues 7:282-289.
А. Делеция интрона сайт-направленным мутагенезом с использованием фагмидного вектора и генетической селекции.
Сайт-направленный мутагенез для делеции 1,3-тпн-интрона осуществляли в 4,3-тпн Sal I/Hind III фрагменте ДНК, субклонированном в фагмидный вектор, Bluescript SK M 13 (-) (Strategene). Этот вектор выбрали, так как он может принимать Sal I/Hind III инсерцию большого размера (4,3 тпн). Фагмидные векторы - плазмидные векторы, содержащие межгенный район, бактериофага f1, который позволяет реплицироваться как одноцепочечная ДНК. В дополнение фагмидные векторы имеют целый ряд преимуществ над одноцепочечными векторами бактериофага M13. Так как фагмиды меньше половины размера векторов M13, которые предпочитают инсерции размером меньше, чем 2,3 тпн, большие инсерции могут проще субклонироваться в фагмиды, а возможность спонтанных делеций уменьшается. Другое преимущество фагмид - то, что инсерции можно секветировать прямо из двухцепочечной суперспирализованной ДНК, что облегчает их характеристику.
Мутагенез осуществляли, используя Muta-Gene в vitro Mutagenesis Kit et Bio Rad. Клетка-хозяин для приготовления матрицы для мутагенеза - штамм E. coli CS 236 (Генотип: dut, ung, thi, relA1, pCS 105 [Cap] CS236 несет F'-фактор, селектирующийся на хлорамфеникол и позволяющий "спасать" одноцепочечную фагмидную ДНК с помощью подходящего хелпернего фага R 408, использованного в настоящей работе. "Спасенная" одноцепочечная фагмидная ДНК претерпевает частичное замещение на урацил, обусловленное dut (дУТФаза) и Ung (урацил-гликозилаза)-мутациями в CS 236. ДНК-затравка с замененными урацилами, используемая для мутагенеза, селективно разрушается при трансформации в клетки хозяев, которые содержат ung локус дикого типа, в данном случае DH5L , таким образом позволяя предпочтительную репликацию новосинтезированной мутантной ДНК.
Для выполнения мутагенеза очищенный в геле 4,3 тпн-Sal I/Hind III фрагмент лигировали в переваренный и очищенный в геле Sal I/Hind III SKM 13 (-). Лигированную ДНК вводили в клетки штамма CS 236, сделанные компетентными по методу Hanahan, как описано в J. Mol. Biol., 166:557 (1983). Трансформанты селектировали на чашках, содержащих 50 мкг/мл ампициллина (для отбора фагмид) и 30 мкг/мл хлорамфеникола (для отбора на сохранение F'-фактора). Трансформантов проверяли на присутствие правильной инсерции рестрикционным анализом ДНК. Трансформант, несущий правильную инсерцию (pSHM-D19), использовали в последующем эксперименте по мутагенезу.
Одноцепочечный образец из pSHM-D19 "спасали", используя фаг R 408, как хелперный. Штамм CS 236, содержащий pSHM-D19, выращивали в бульоне Бурия, содержащем ампициллин (50 мкг/мл) и хлорамфеникол (30 мкг/мл) до A600 примерно 0,3. Клетки инфицировали R 408 хелперным фагом при множественности инфекции 20, затем трясли при 37oC 8 - 14 ч. Из "спасенных" фагмид экстрагировали одноцепочечный образец.
Сайт-направленный (интрон делетирующий) мутагенез выполняли, используя 72-пн олигонуклеотид (олигонуклеотид 1 на фиг. 16). Олигонуклеотид 1 имеет основания 1 - 30, комплементарные кодирующей цепи сразу ниже /3'/ и основания 31 - 71, комплементарные кодирующей цепи сразу выше /5'/ интрона, который надо делетировать из CNTF геномной ДНК. Для использования в реакциях мутагенеза олигонуклеотид фосфорилировали, используя полинуклеотидкиназу T4. Реакции мутагенеза, используя Rio Rad Muta Genekit, выполняли в соответствии со спецификациями производителя, за исключением того, что ДНК после реакций мутагенеза использовали, чтобы трансформировать штамм E.coli DH5L. Делеционных мутантов характеризовали рестрикционным картированием ДНК и ДНК-сиквенсом двухцепочечной ДНК из мутантов с подходящими рестрикционными картами. Правильно делетированный, лишенный интрона мутант, в Bluescript обозначен pMCN-21.
Б. Реконструкция 5'-конца гена CNTF для экспрессии.
5'-конец CNTF кодирующей последовательности реконструировали для того, чтобы сделать определенные замены, которые не изменяют кодируемую аминокислотную последовательность, но которые предположительно увеличивают эффективность экспрессии в бактериях. Частично перекрывающиеся, комплементарные олигонуклеотиды 2 и 3 (фиг. 16) синтезировали, очищали в геле и отжигали друг с другом. Олигонуклеотид 2 кодирует аминокислоты, присутствующие в человеческом CNTF выше Nhe I сайта (66). Кодирующая последовательность олигонуклеотида 2 содержит несколько оснований, которые отличаются от присутствующих в человеческом гене (сравните фиг. 12 и 16). Эти замены были сделаны либо чтобы изменить человеческие кодоны на те, которые используются предпочтительно в E. coli (в соответствии с de-Rocr Kastelein в From Gene to Protein : Steps Dictating the Maximal Level of Gene Expression (1986) Davis и Reznikoff, eds. pp. 225 - 283, Butterworths, N. Y.), или чтобы создать Bgl II сайт (фиг. 16), вставленный в последовательность для простоты последующих генетических манипуляций. Олигонуклеотид 2 также кодирует трансляционный линкер на 5' конце (фиг. 16), облегчающий трансляцию. Отожженные олигонуклеотиды 2 и 3 имеют Bam HI - выступающий конец на 5' и Nhe I выступающий конец на 3' для облегчения последующего лигирования и клонирования (фиг. 14 и 16).
Рестрикционный анализ последовательности ДНК гена CNTF человека выявил уникальный сайт Nhe I (фиг. 12). Поэтому олигонуклеотиды 2 и 3 были сделаны с Nhe I выступающим концом; это позволяло им быть объединенными с оставшимся 3'-фрагментом гена после переваривания Nhe I.
В. Присоединение олигонуклеотидов 2 и 3 к кодирующим последовательностям с удаленным интроном.
Олигонуклеотиды 2 и 3, содержащие достроенные N-концы CNTF гена, отжигали друг с другом и лигировали с разрезанной Nhe pMCN-2a. Лигированную ДНК затем переваривали Bam H I и Hind III, чтобы освободить фрагмент ДНК, обозначенный CNTF-Syn I, который содержит последовательности ДНК, подходящие для экспрессии в E. coli и кодирующие человеческий CNTF выше Hind III сайте (233).
Стратегия 2 для конструирования 5'-конца (фиг.15):
Осуществляли альтернативную стратегию, в которой интрон удаляли сайт-направленным мутагенезом, но готовили более полную синтетическую последовательность ДНК, кодирующую CNTF выше Hind III сайта (233), но без интрона. Чтобы сделать эту синтетическую конструкцию ДНК, были синтезированы олигонуклеотиды с 5 по 10, фиг.17. Эти олигонуклеотиды предназначены, чтобы сформировать частично перекрывающиеся двухцепочечные пары: 5, и 6, 7 и 8, 9 и 10. Каждая двухцепочечная пара содержит одноцепочечные выступающие концы, предназначенные для правильного лигирования их вместе в порядке (5 и 6)-(7 и 8)-(9 и 10). Олигонуклеотиды очищали в геле, отжигали и лигировали вместе в отдельный 261 пн двухцепочечный ДНК-олигонуклеотид, названный CNTF-Syn 2. Эта синтетическая ДНК также содержала: (1) измененные кодоны, которые учитывают предпочтительное использование кодонов в E. coli (ср. фиг. 12 и 17), (2) 5' трансляционный линкер, использованный выше (фиг. 16 и 17) и (3) 5' Bam H I выступающий и 3' Hind III выступающий концы, чтобы облегчить лигирование и клонирование (фиг.17).
Приготовление 3'-конца для экспрессии.
Конструкция (фиг. 14 или 15) клон человеческой геномной ДНК CNTF в фаге лямбда EMBL3 из примера 4 разрезали ферментом рестрикции Hind III, и 2, 1 тпн-фрагмент, содержащий кодирующую последовательность Hild III сайта, очищали в геле. Этот 2,1 тпн фрагмент клонировали в разрезанную Hild III плазмиду pFMBL8 (Donti и др., 1983, Nucleic Acidc Res 11: 1645). Spe I -сайт (613) вставляли с 2,1 тпн-инсерцию ДНК олигонуклеотид-направленным мутагенезом 13 нуклеотидами ниже стоп кодона, завершающего CNTF последовательность, используя синтетический олигонуклеотид 4 (фиг.16). Мутантную плазмиду разрезали Hind III и Spl I, чтобы высвободить нижележащий кодирующий фрагмент, который очищали в геле и назвали CNTF - Syn 3 (содержащий кодирующие последовательности для человеческого CNTF ниже Hind III сайта (233)).
Приготовление полной экспрессионной конструкции.
CNT-Syn I лигировали с CNTF-Cyn 3, чтобы получить CNTF-Syn 1/3 (фиг.14). CNTF-Syn 2 лигировали с CNTF-Syn 3, чтобы получить CNTF-Syn 2/3 (фиг.15). Получали два альтернативных фрагмента ДНК, каждый из которых кодирует человеческий CNTF и имеет подходящий модификации последовательности ДНК для усиления эффективной экспрессии в E. coli. Эти фрагменты ДНК затем экспрессировали в E. coli после субклонирования в: (A) бактериальный экспрессионный вектор PT5T, основанный на промоторе фага 17 или (Б) бактериальный экспрессионный вектор, pT3xI-2, основанный на гибридном промоторе лактозного триптофанового оперона ('Tac').
2. Экспрессия CNTF с использованием экспрессионного вектора, основанный на системе "Т7 промотора", характеристики данного вектора приведены на фиг. 17.
А. Описание pT5T.
Основанный на T7 промоторе экспрессионный вектор pT5T является в значительной степени тем же, что и pJU 1003, описанный Squires, et.al., J. Biol. Chem. (1988) 263: 16297-16302, за исключением того, что здесь короче размер ДНК между уникальным Bgl III сайтом 5' у T7 промотора и Cla I сайтом в гене устойчивости к тетрациклину. Последовательность этой ДНК следующая:
ATCGATGATA AGCTGCAAA CATGAGAATT GAGCTCCCCG GAGATCCTTA
GCGAAGCTA
Cla I
AGGATTTTTT TTAGATCT
Bgl II
Б. Конструкция полного экспрессионного вектора.
Очищенный в геле вектор линеаризовали Bam H I и Spe I ферментами рестрикции. CNTF - Syn 1/3 смешивали с лианизированным вектором и лигировали, чтобы сформировать экспрессионную конструкцию pT5T : CNTF - Syn 1/3 (см. фиг. 14 с общей схемой векторной конструкции).
В. Экспрессия рекомбинантного человеческого CNTF в E. coli: pT5T: CNTF - Syn 1/3 трансформировали в штамм E. coli BL21 (DE3) для экспрессии. Этот штамм, описанный у Studier и Moffat, J. Mol. Biol. (1986) 189: 113 - 130, содержит ген РНК-полимеразы T7 под контролем индуцируемого I PTG lac - промотора на невырезающемся лизогенном бактериофаге лямбда. Из 10 скринированных трансформантов два глона, как было обнаружено, экспрессируют IPTG индуцируемый белок, мигрирующий с молекулярным весом, соответствующим по величине CNTF (≈ 24 кДа). Эти два клона обозначены, как pT5T: CNTF - Syn1/3-5a и 5c. ДНК секвенирования pT5T : CNTF - Syn 1/3 - 5a и 5c подтвердило, что данные последовательности рекомбинантов правильные.
Высокий уровень экспрессии рекомбинантного CNTF достигали выращиванием клеток в бульоне Лурия с 15 мкг/мл тетрациклином до клеточной плотности, соответствующей A600 0,5-0,8. Клетки выращивали в течение 1,5-4 ч либо без IpTG ("неиндуцированные"), либо IPTG добавляли до конечной концентрации 1,0 мМ ("индуцированные").
Г. Анализ экспрессированного белка электрофорезом в SDS полиакриламидном геле с последующим окрашиванием красителем Кумассии или иммуноблоттингом.
Клетки собирали быстрым центрифугированием и растворяли непосредственно в буфере для образцов SDS - полиакриламидного геля (0,025% бромфеноловый синий, 11% глицерин, 1% β - мерраптоэтанол, 2% SDS, 0,0625 M трисHCl, pH 6,8 (и кипятили 2 мин) (фиг. 20 и 21). В клетках, трансформированных pT5T : CNTF - Syn 1,3-5a и индуцированных IPTG, в течение 2 ч (дорожка 5, фиг.20) наблюдается интенсивно окрашенная красителем Кумассии полоса, идущая в положении, ожидаемом для SN-CNTF (24 кДа). Если клетки выращивали без IPTG (дорожка 4, фиг. 20), наблюдали значительно меньшую интенсивность этой полосы, как и ожидалось для белка, экспрессия которого находится под контролем lac оперона. Клетки, трансформированные вектором pT5T без инсерции CNTF, не показывали такую полосу ни в индуцированном (дорожка 3, фиг.20), ни в неиндуцированном (дорожка 2, фиг.20) варианте. Дорожка 2 включает стандарты молекулярного веса.
Идентичный SDS - полиакриламидный гель переносили на нитроцеллюлозу или делали иммуноблотинг с аффинно-очищенными антителами к CNTF пептиду A (E-S-Y-V-K-H-Q-G-L-N-K-N). В клетках, трансформированных pT5T : CNTF - Syh 1,3-5a и индуцированных IPTG в течение 2 ч (дорожка 5, фиг. 21), наблюдается плотная полоса, узнаваемая афинно-очищенными антителами к CNTF пептиду A и идущая в позиции, ожидаемой для CNTF (34 кДа). Если клетки выращивали без IPTG (дорожка 4, фиг.21), наблюдали гораздо менее интенсивную полосу в этом положении. Клетки, трансформированные вектором pT5T без инсерции CNTF, не показывали этой полосы ни в индуцированном (дорожка 3, фиг.20), ни в неиндуцированном виде (дорожка 2, фиг.20).
Дорожка 1 содержит стандарты молекулярного веса.
В дополнение клетки, трансформированные pT5T : CNTF - Syn 1, 3-5 и индуцированные IPTG в течение 2 ч, разрушали прохождением три раза через клеточный компрессор Френча. Аликвоту грубого клеточного лизата разделяли на фракции супернатанта и осадка центрифугированием при 20000 rрm в JA-20 роторе (Beckman) 15 мин. Аликвоты грубого лизата, супернатанта и осадка, представляющих то же количество начальной клеточной суспензии, также прогоняли на том же SDS полиакриламидном геле, переносили на нитроцеллюлозу и делали иммуноблотинг с афинно-очищенными антипептид A антителами. Супернатант лизата (дорожка 8, фиг. 21) содержал гораздо более иммунореактивный CNTF, чем осадок лизата (дорожка 9, фиг.21). Уровень супернатанта CNTF был сравним с нефракционированным лизатом (дорожка 7, фиг.21).
Д. Биоактивность экспрессированного CNTF.
Клетки, собранные быстрым центрифугированием, ресуспендировали в 20 мМ трис-HCl, pH 8,2 в 1/35 начального объема клеточной суспензии, разрушали пассированием через клеточный компрессор Френча три раза и грубый клеточный лизат разделяли на фракции супернатанта и осадка центрифугированием при 20000 rpm в SA-20 роторе (Beckman) 15 мин. Последовательные разведения фракции клеточного супернатанта анализировали на их способность усиливать выживаемость цилиарных нервных клеток ганглия (как описано в примере 1). Супернатант демонстрировал значительную биоактивность при разведении 1:1000000 (фиг. 22). Специфическая активность рекомбинантного человеческого SN-CNTF, как было оценено на основании биоактивности и количества CNTF белка, рассчитанных из иммуноблотов, составляет приблизительно 275 TU/нг. Эта специфическая активность почти вдвое превосходит специфическую активность очищенного CNTF белка, указывая, что рекомбинантный CNTF биологически активен в бактериальном клеточном экстрате. Лизаты клеток, трансформированных pT5T, без инсерции CNTF, не демонстрировали детектируемой биоактивности.
Супернатант также прогоняли электрофорезом в 15%-ном полиакриламидном редуцирующем SDS-геле и разрезали на (1 мм кусочки ширины), которые экстрагировали в течение ночи в культуральной клеточной среде при 4oC со встряхиванием и анализировали на активность, как описано в примере 1. Вставка на фиг. 22 иллюстрирует, что пик биоактивности выявляется во фракциях, соответствующих 24 кДа (как это и ожидалось для CNTF).
E. Аминокислотная последовательность экспрессированного CNTF.
Район вокруг 24 кДа (аналогичный на фиг. 20) вырезали из SDS полиакриламидного геля, но не окрашивали Кумасси. Этот материал секвенировали деградацией по Эдману в белковом секвенаторе Applied Biosystems (как описано в примере 2). Была получена нижеследующая аминокислотная последовательность: AFTEHSPLTPHRRDL[? ]S... Значок вопроса соответствует C в человеческой последовательности (фиг. 12), которая не может быть определена этим методом. Эта последовательность соответствует ожидаемой для человеческого CNTF (фиг. 12) и обеспечивает дополнительное доказательство, что является правильно экспрессированным. Это также указывает на то, что аминотерминальный метионин удаляется во время экспрессии, оставляя аланин, как аминотерминальную аминокислоту в экспрессированном белке.
Вышеприведенные результаты демонстрируют, что иммунологически перекрестно-реактивный CNTF экспрессировался на высоком уровне в биологически активной форме, большая часть которой растворима после лизиса бактериальных клеток.
3. Экспрессия CNTF с использованием экспрессионного вектора, основанного на гибридном промоторе 'TAC' (см. фиг. 19 по характеристикам вектора).
A. Описание pT3XI-2 (модификации pKK223-3).
Начальной плазмидой для этой конструкции служила плазмида pKK 223-3, закупленная у Pharmacia. Плазмида pKK223-3 несет часть гена тетрациклической устойчивости. Этот нефункциональный ген заменяли полным геном устойчивости к тетрациклину pBR 322. Плазмиду pKK 223-3 переваривали полностью Sph I и частично Bam H I, 4,4 тпн фрагмент очищали в геле и комбинировали с синтетическим адаптором со следующей последовательностью:
5' GATCTAGAATTGTCATGTTTGACAGCTTATCAT 3'
3' ATCTTAACAGTACAAACTGTCGAATAGTAGC 5'
и 539 пн фрагментом ДНК из Cla I / Sph I рестрицированного гена тетрациклиновой устойчивости pBR (PL Biochemicals, 27-4891-01). Результирующую плазмиду обозначили pCJ1.
Затем XhoI линкер, закупленный у New England Biolals, инсерцировали в Pvu II сайт плазмиды pCS1, чтобы получить плазмиду pCSX-1. Эта инсерция нарушала rop ген, который контролирует копийность плазмиды. EcoR I Фрагмент, содержащий lac I ген, очищали из плазмиды pMC9(Colos и др., Proc. Natl. Acad. Sci USA (1983), 80:3015-19), а затем вставляли в Xho I сайт с Xho I - EcoR I адаптерами, имеющими последовательность
5' TCGAGTCTAGA 3'
3' CAGATCTTTAA 5'
Последовательность полилинкера между EcoR I и Pst I сайтами в плазмиде pKK223-3 была затем заменена полилинкерной последовательностью, указанной ниже
5' AATTCCCGGG TACCAGATCT GAGCTCACTA GTCTGCA 3'
3' GGGCCC ATGGTCTAGA CTCGAGTGAT CAG 5'
Полученный плазмидный вектор обозначили pCSXI-1.
В конце концов, ген тетрациклиновой устойчивости был заменен сходным геном, который имел сайты для ферментов рестрикции Hind III, Bam H I и Sal I, нарушенные мутагенезом с бисульфитом. Следующая процедура использовалась, чтобы заставить мутировать ген тетрациклиновой устойчивости pBR322. Плазмиду pBR322 разрезали Hind III, а затем подвергали мутагенезу с бисульфитом натрия (Shortle и Nathans, Proc., Natl. Acad. Sci. USA (1978) 5: 2170-2174). Мутантную ДНК лигировали, чтобы образовать циклическую ДНК, затем разрезали Hind III, чтобы линеаризовать любую плазмиду, избежавшую мутагенеза. E.coli SM 109 (Janisch-Perron и др., Gene (1985) 33: 103-119) трансформировали этой плазмидой и затем высевали на селективные среды. Выделяли плазмиды из тетрациклин-устойчивых колоний и проверяли на отсутствие Hind III сайта в гене устойчивости к тетрациклину. Успешно мутировавшую плазмиду обозначили pT1. Аналогичная процедура использовалась для введения мутации в BamH1 сайт pT1 и получения плазмиды pT2. Плазмиду pT2 в свою очередь подвергали мутагенезу, чтобы удалить Sal I сайт, и получали плазмиду pT3•Cla I/Bsm I - фрагмент pT3, несущий мутантный ген тетрациклиновой устойчивости, выделили и использовали, чтобы заменить гомологичный фрагмент pCJXI - 1 для создания pT3XI - 2. Мутантный ген тетрациклиновой устойчивости все еще кодировал функциональный белок.
B. Создание pT3XI - 2 - ⊘ 10TC3FGF syn (приготовление tac - промоторного вектора для CNTF).
Первоначально был синтезирован ген основного фактора роста фибробластов (в FGF). Этот ген кодирует последовательность, описанную Sommer и др., (1987, Biochem. Biophys. Res. Commun. 141.67), но использует кодоны, являющиеся предпочтительными в генах с высоким уровнем экспрессии в E.coli. Структура этого гена такова, что (см. Squires и др. 1988, ibid) обеспечивает эффективную инициацию трансляции.
FGF синтетический ген сначала вставили в M13mp18 между EcoR I и Hind III сайтами и секвенировали. Структура этого гена приведена далее полностью.
Главные особенности гена отмечены особо.
Затем его изолировали рестрикцией BamHI и Hind III и инсерцией в Bam H I / Hind III разрезанный pSU 1003/ Squires и др., 1988, ibid; с по pSH 1001003-Syn FGF. Эту плазмиду переварили xbaI и Hind III, и xba I/Hind III фрагмент, несущий FGF ген, выделили. Этот фрагмент лигировали в рT3XI-2, расщепленную EcoR I и Hind III, используя EcOR I-xba I линкер
pATT TCC ACA ACG GTT TCC, CT3';
3'GG TGC CAA AGG GAG ATCp5'.
Новую плазмиду обозначили pT3XI - 2 - ⊘ 10TC3 FGF syn.
Г. Инсерция CNTF экспресионных конструкций в Tac промоторный вектор.
pT3XI - 2 - ⊘ 10TC3 FGF syn расщепляли BamH I и Spl I которые проводили к линеаризации 7,4 тпн экспрессионного вектора и высвобождению примерно 0,5 тпн FGF фрагмента ДНК. В отдельных реакциях CNTFsyn 1/3 и CNTF syn 2/3 лигировали с очищенным в геле Bam H I/Sp lI расщепленном ДНК фрагментом вектора с образованием плазмид pT3XI - 2: CNTF - syn 1/3 и pT3XI - 2 : syn 2/3 соответственно.
Д. Экспрессия в E. coli pT3XI - 2 : CNTF - syn 1/3 трансформировали в фаг-устойчивый К-штамм E.coli SM 07, 14 трансформантов выращивали и анализировали на экспрессию CNTF электрофорезов в SDS - полиакриламидном теле с окрашиванием Coomassie Brilliant Blue 4 трансформанта демонстрировали интенсивно окрашенную полосу в приблизительной позиции CNTF. Высокий уровень экспрессии рекомбинантного CNTF достигался выращиванием клеток в бульоне Лурия с 15 мкг/мл тетрациклином до клеточной плотности, соответствующей A600 0,8. Добавляли IpTG до конечной концентрации 1,0 мМ и клеткам позволяли расти в течение 2 ч. Все четыре трансформанта демонстрировали почти одинаковую плотность CNTF как на Кумасси-окрашенном, так и на геле с иммуноблотингом. Кажущийся уровень CNTF экспрессии в этих трансформантах (по данным гелей) был около 1/4 по сравнению с демонстрируемым pT5T:CNTF - syn 4/3 - 5a, растущим, как и предыдущий, с тетрациклином. 7 мл этой клеточной суспензии добавляли на 50 мл бульона Лурия в большие флаконы и выращивали при 27oC со встряхиванием до A600 приблизительно 0,5. Затем добавляли IPTG в конечной концентрации 0,5 мМ, чтобы индуцировать экспрессию CNTF и продолжали выращивать до достижения A600 приблизительно 1,2, что обычно занимает 4 - 6 ч. Клетки собирали центрифугированием на SA-20 роторе при 7000 rpm 5 мин при 4oC и отмывали еще раз центрифугированием в 50 мМ фосфатном буфере, pH 8,0 (Буфер A) при 4oC. Супернатант удаляли и осадок клеток замораживали при -20oC как пасту.
Клеточную пасту суспендировали в буфере Б (буфер А, содержащий 1 мМ ЭГТА и 1 мМ ЭДТА) (по 0,5 г пасты на 1 мл буфера) при 4oC и проводили три раза через клеточный компрессор Френча, чтобы разрушить бактерии. Полиэтиленимин (PEI) добавляли до конечной концентрации 0,25% и встряхивали 30 мин при 4oC. Преципитат удаляли центрифугированием в течение 15 мин на самой высокой скорости в Mirofuge. Эта стадия обычно приводит к уменьшению содержания нуклеиновой кислоты, как измерено по соотношению A260/A280, от 25 до менее чем 5%.
Образец затем наносили на колонку Q-Сефарозы (Pharmacia), уравновешенной буфером Б. CNTF является основным компонентом лизата разрушенных клеток, поэтому хроматография может сопровождаться анализом колоночных фракций на SDS - полиакриламидном геле. Главная, Кумасси-окрашенная полоса SN-CNTF, приблизительно 24 кДа. Используя этот анализ, CNTF смывают с Q-Сефарозной колонки Буфером Б.
Фракции, содержащие основную массу CNTF белка, собирают, диализуют против Буфера С (5 мМ фосфатный буфер, pH 8,0, содержащий 1 мМ ЭГТА и 1 мМ ЭДТА) и применяют в колонке Q - Сефарозы, уравновешенной Буфером C. Колонку промывают Буфером C до тех пор, пока A280 не возвращается к базовой линии, указывая, что несвязавшиеся белки отмыты с колонки. В Буфере C CNTF связывается с Q - Сефарозной колонкой, а затем его элюируют 0 - 0,1 М NaCl градиентом в Буфере C. CNTF выходит из колонки при приблизительно 40 мМ NaCl и с более чем 90%-ной чистотой, судя по окрашенным Кумасси-SDS-полиакриламидным гелем фракции пика CNTF.
Последовательность гена в FGF приведена в конце текста.
Источники информации
1. Backman, K., Ptashne, M. and Gilbert, W. Proc. Natl. Acad. Sci. USA 73, 4174-4178 (1976).
2. de Boer, H.A., Comstock, L.J., and Vasser, M. Proc. Natl. Acad. Sci. USA 80, 21-25 (1983).
3. Shimatake, H. and Rosenberg, M. Nature 292, 128-132 (1981).
4. Derom, C., Gheysen, D. and Fiers, W. Gene 17, 45-54 (1982).
5. Hallewell, R.A. and Emtage, S. Gene 9, 27-47 (1980).
6. Brosius, J., Dull, T.J., Sleeter, D.D. and Noller, H.F. J. Mol. Biol. 148 107-127 (1981).
7. Normanly, J., Ogden, R.C., Horvath, S.J. and Abelson, J. Nature 321, 213-219 (1986).
8. Belasco, J.G., Nilsson, G., von Gabain, A. and Cohen, S.N. Cell 46, 245-251 (1986).
9. Schmeissner, U., McKenney, K., Rosenberg M. and Court, D. J. Mol. Biol. 176. 39-53 (1984).
10. Mott, J. E. , Galloway, J. L. and Platt, T. EMBO J. 4, 1887-1891 (1985).
11. Koshland, D. and Botstein, D. Cell 20. 749-760 (1980).
12. Morva, N.R., Kakamura, K. and Inouye, M. J. Mol. Biol. 143, 317-328 (1980).
13. Surin, B. P. , Jans, D.A., Fimmel, A.L., Shaw, D.C., Cox, G.B. and Rosenberg, H. J. Bacteriol. 157, 772-778 (1984).
14. Sutcliffe, J.G. Proc. Natl. Acad. Sci. USA 75, 3737-3741 (1978).
15. Peden, K.W.C. Gene 22, 277-280 (1983).
16. Alton, N.K. and Vapnek, D. Nature 282, 864-869 (1979).
17. Yang, M., Galizzi, A., and Henner, D. Nuc. Acids Res. 11(2), 237-248 (1983).
18. Wong, S.-L., Price, C.W., Goldfarb, D.S., and Doi, R.H. Proc. Natl. Acad. Sci. USA 81, 1184-1188 (1984).
19. Wang, P.-Z., and Doi, R. H. J. Biol. Chem. 251, 8619-8625, (1984).
20. Lin, C. -K. , Quinn, L.A. Rodriguez, R.L. J. Cell Biochem. Suppi. (9B), p. 198 (1985).
21. Vasantha, N., Thompson, L.D., Rhodes, C., Banner, C., Nagle, J., and Filpula, D. J. Bact. 159(3), 881-819 (1984).
22. Palva, l., Sarvas, M., Lehtovaara, P., Sibazkov, M., and Kaariainen, L. Proc. Natl. Acad. Sci. USA 79, 5582-5586 (1982).
23. Wong. S.-L., Pricee, C.W., Goldfarb, D.S., and Doi, R.H. Proc. Natl. Acad. Sci. USA 81, 1184-1188 (1984).
24. Sullivan, M.A., Yasbin, R.E., and Young, F.E. Gene 29, 21-46 (1984).
25. Vasantha, N., Thompson, L.D., Rhodes, C., Banner, C. Nagle, J., and Filula, D. J. Bact. 159(3), 811-819 (1984).
26. Yansura, D.G. and Henner, D.J. PNAS 81, 439-443 (1984).
27. Gray, G.L., McKeown, K.A., Jones, A.J.S., Seeburg, P.H. and Heyneker, H.L. Biotechnology, 161-165 (1984).
28. Lory, S., and Tai, P.C. Gene 22, 95-101 (1983).
29. Liu, P.V.J. Infect. Dis. 130 (suppl), 594-599 (1974).
30. Wood, D.G., Hollinger, M.F., nd Tindol, M.B.J. Bact. 145, 1448-1451 (1981).
31. St. John, T.P. and Davis, R.W. J. Mol. Biol. 152, 285-315 (1981).
32. Hopper, J.E., and Rowe, L.B. J. Biol. Chem. 253, 7566-7569 (1978).
33. Denis, C. L. , Ferguson, J. and Young, E.T. J. Biol. Chem. 258, 1165-1171 (1983).
34. Lutsdorf, L. and Megnet, R. Archs. Biochem. Biophys. 126, 933-944 (1968).
35. Meyhack, B. , Bajwa, N., Rudolph, H. and Hinnen, A. EMBO. J. 6., 675-680 (1982).
36. Watson, M.E. Nucleic Acid Research 12, 5145- 5164 (1984).
37. Gerband, C. and Guerineau, M. Curr. Genet, 1, 219-228 (1980).
38. Hinnen, A. , Hicks, J.B. and Fink, G.R. Proc. Natl. Acad. Sci. USA 75, 1929-1933 (1978).
39. Jabbar, M.A., Sivasubramanian, N. and Nayak, D.P. Proc. Natl. Acad. Sci. USA 82, 2019-2023 (1985).




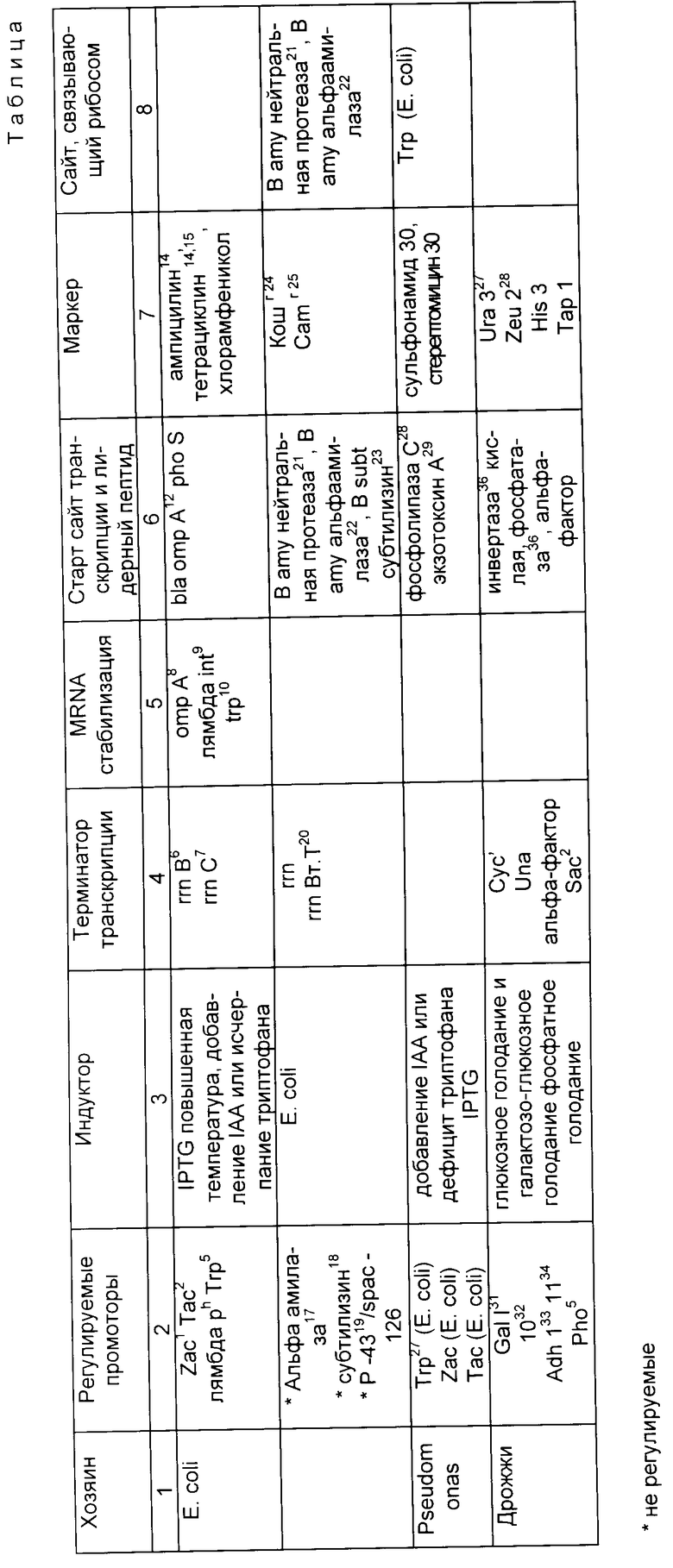
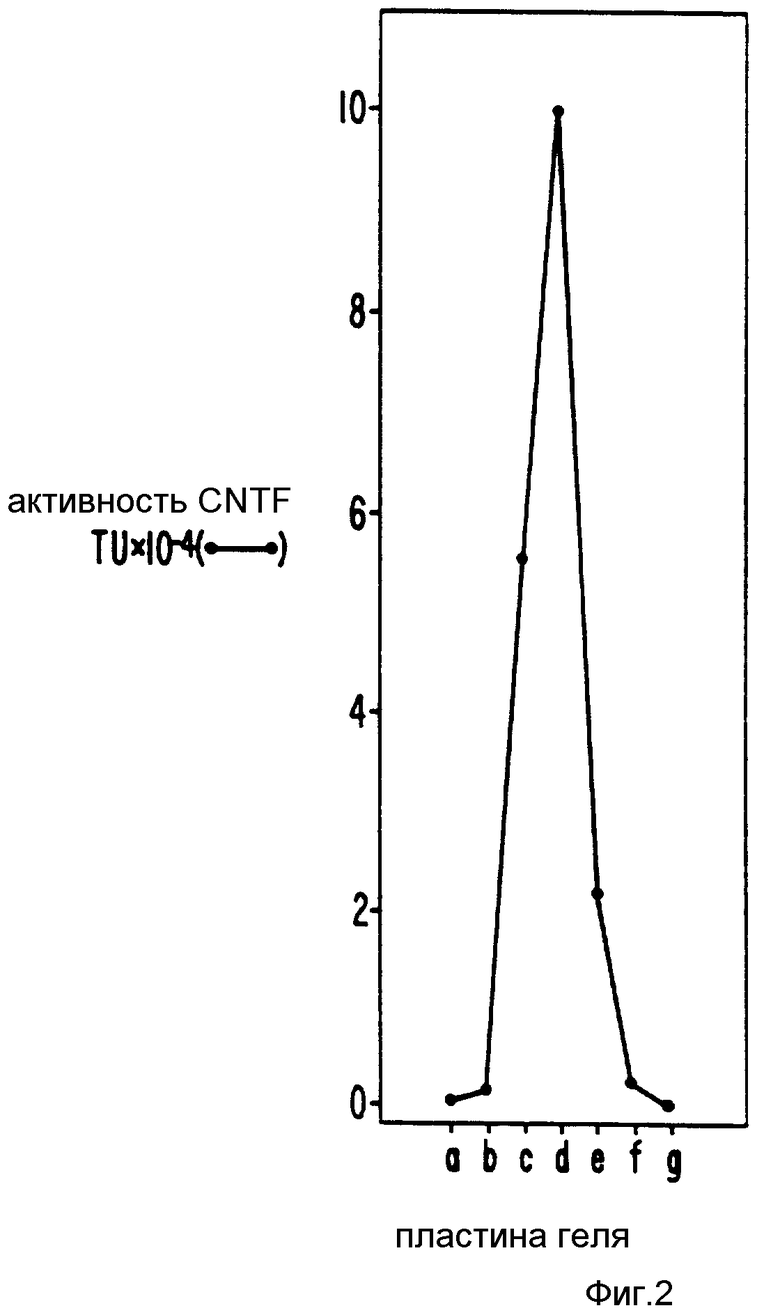
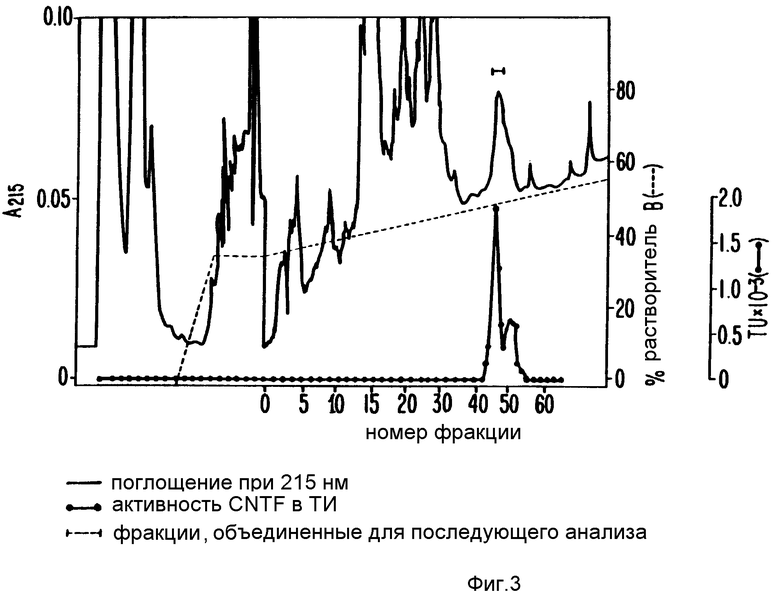

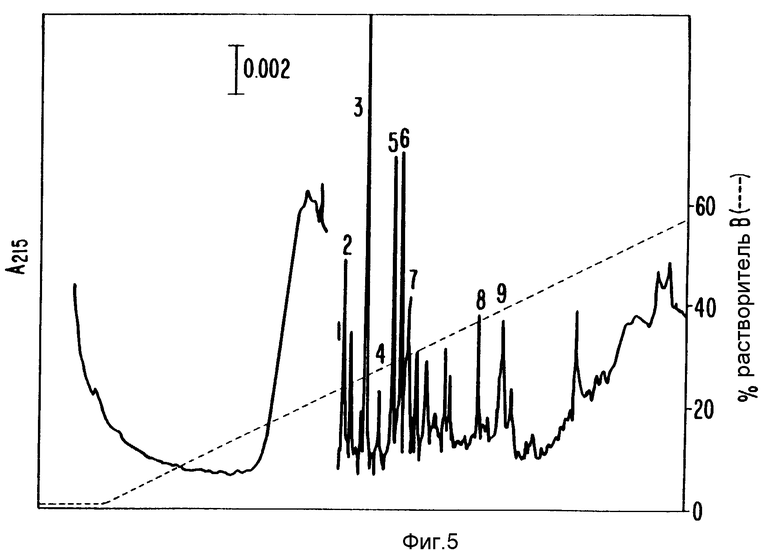
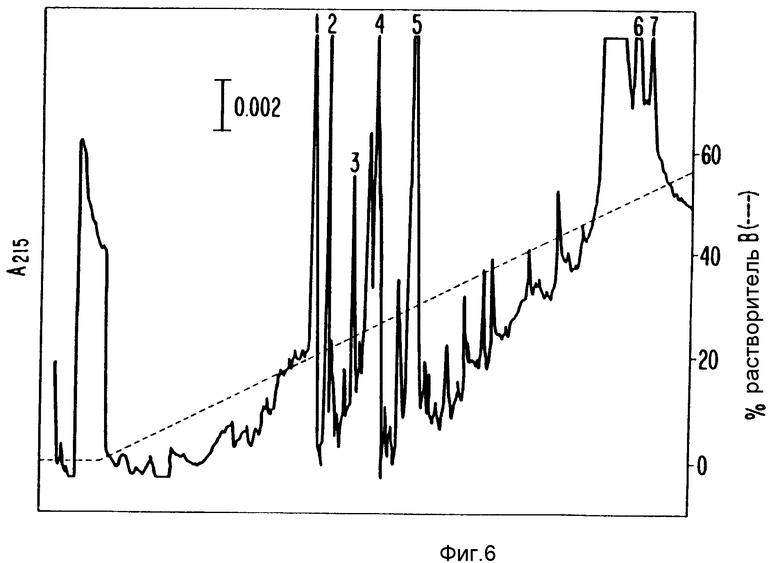
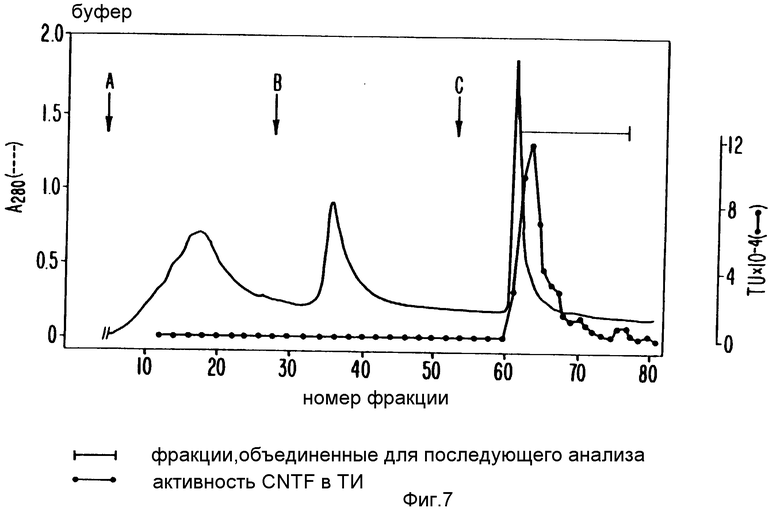
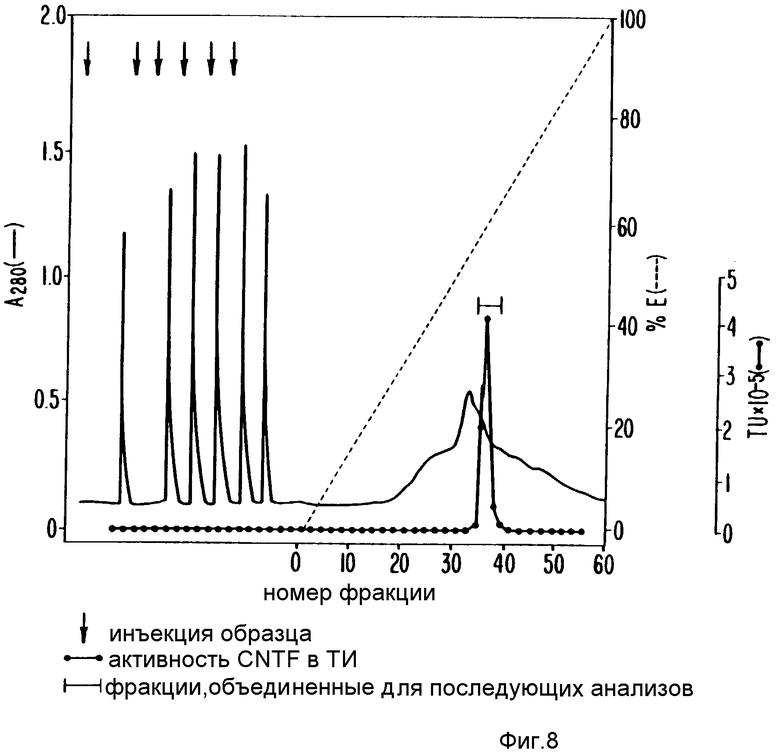
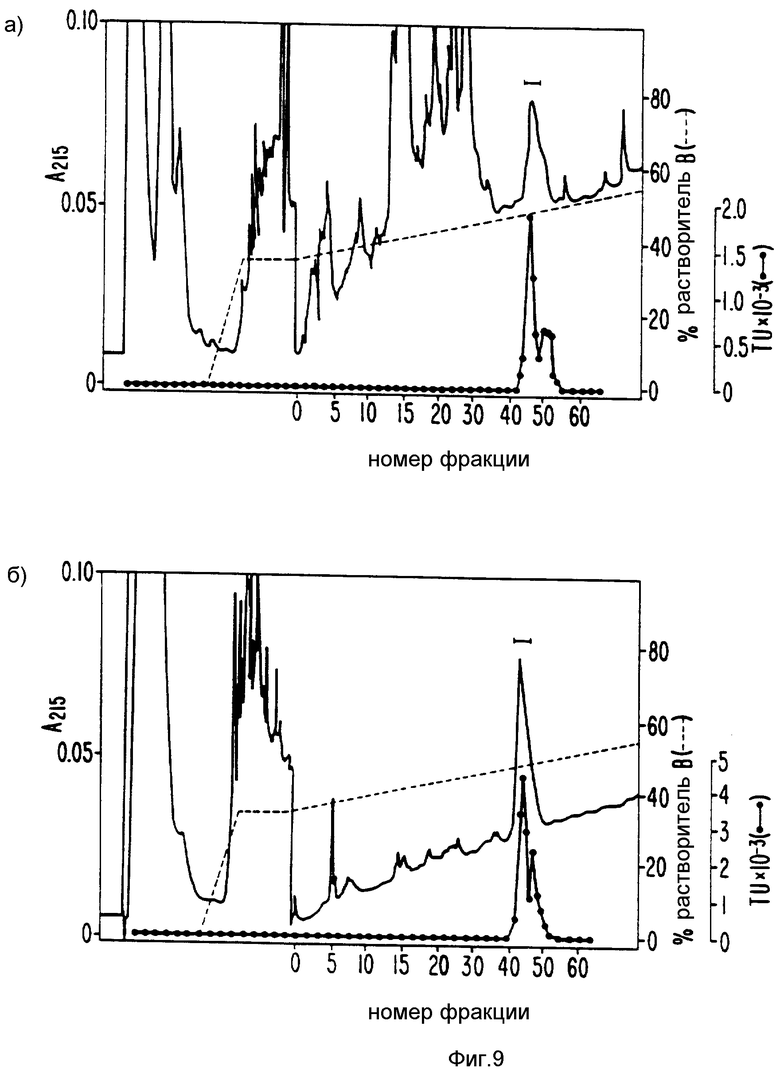
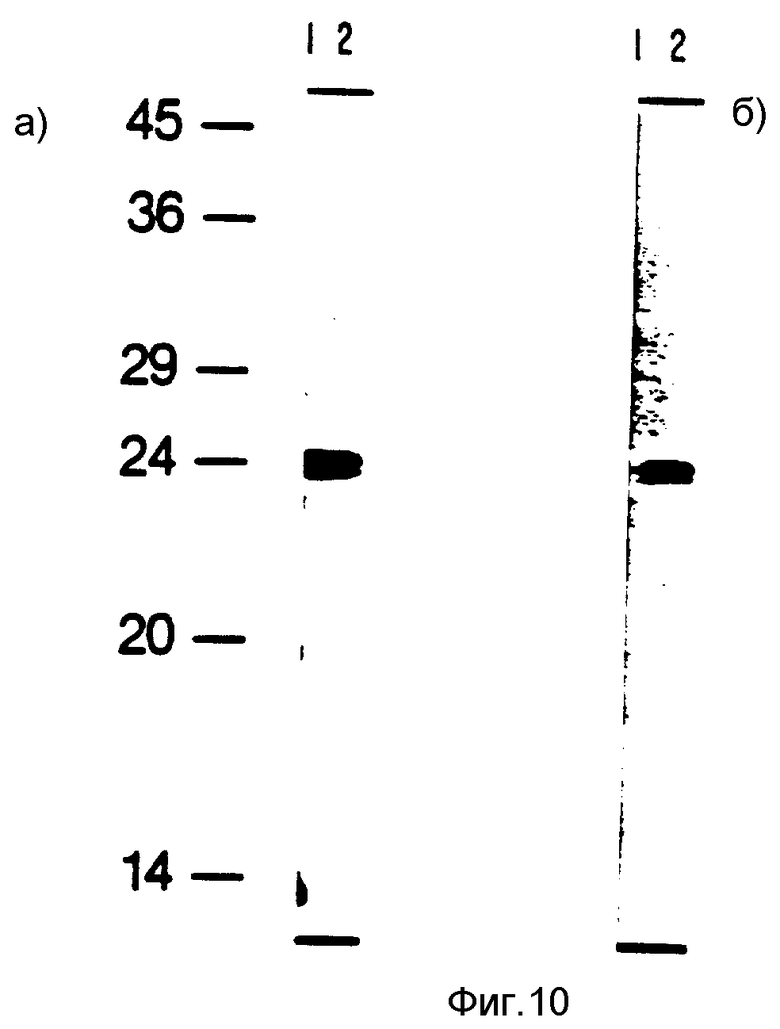


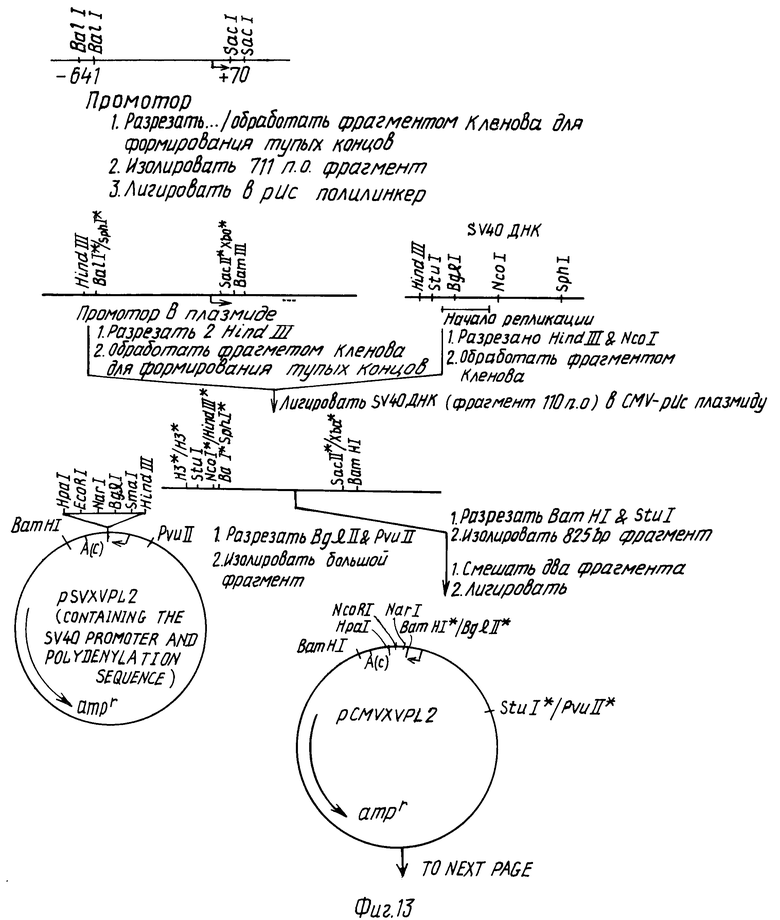
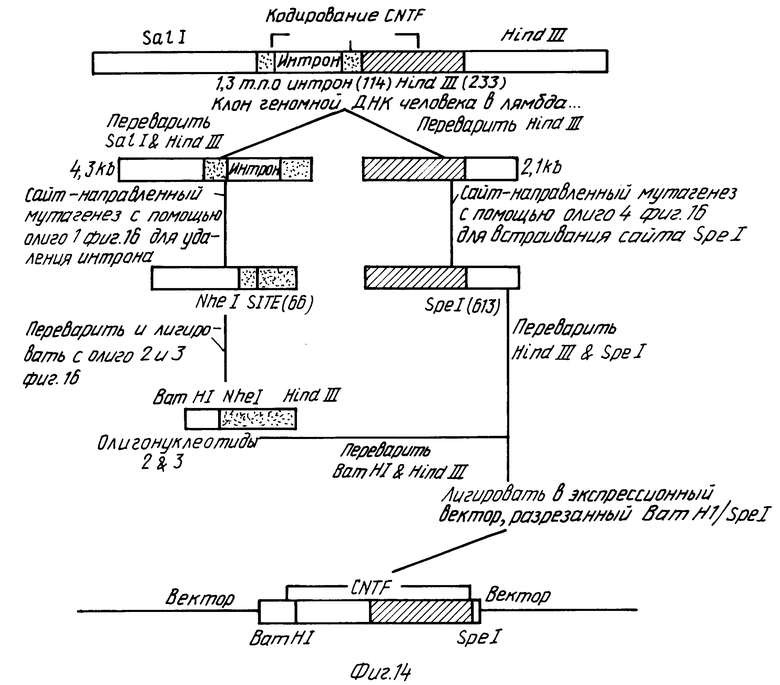
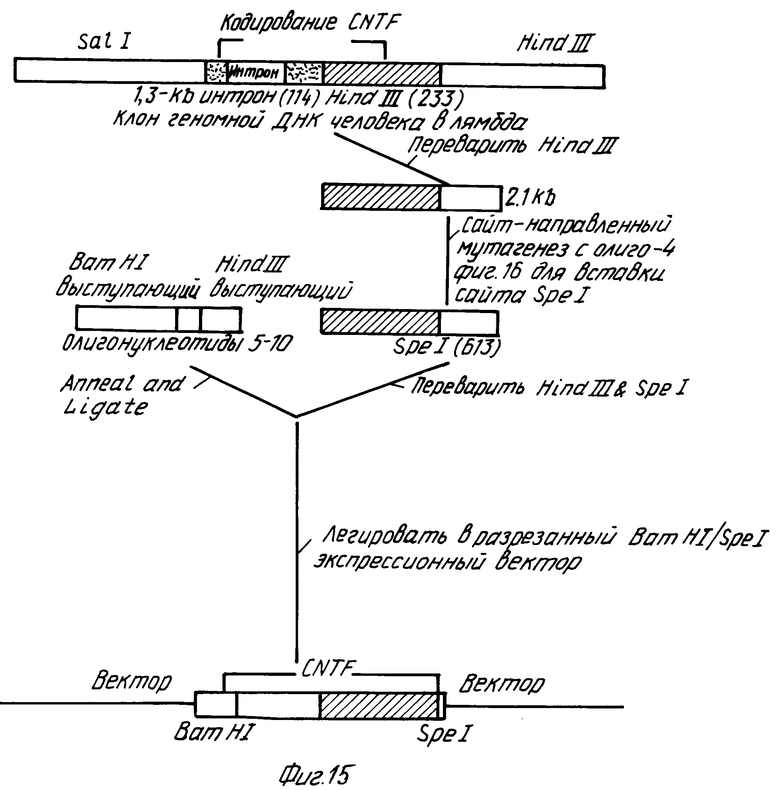


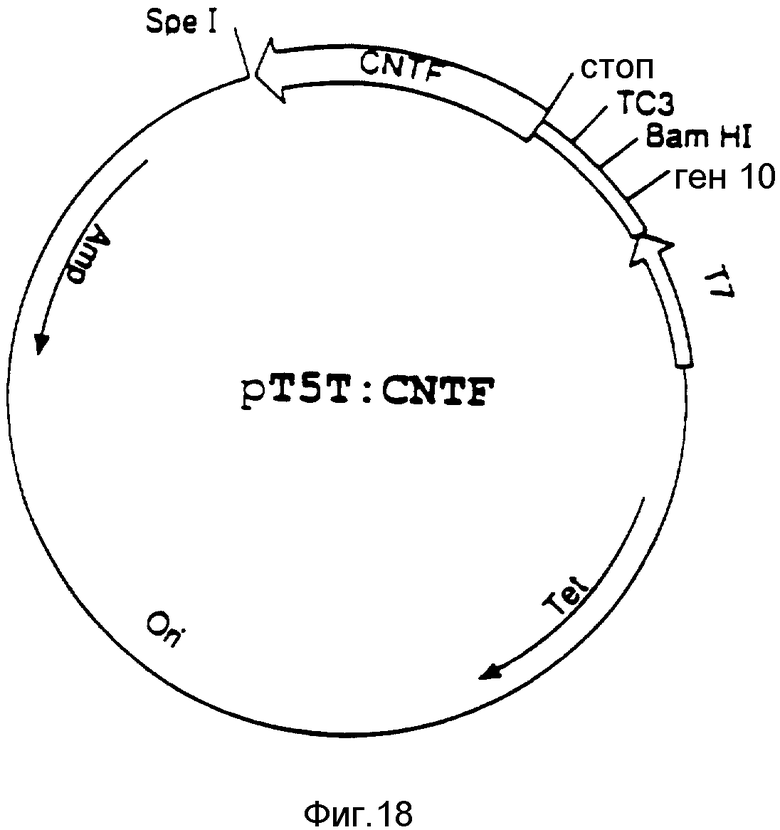
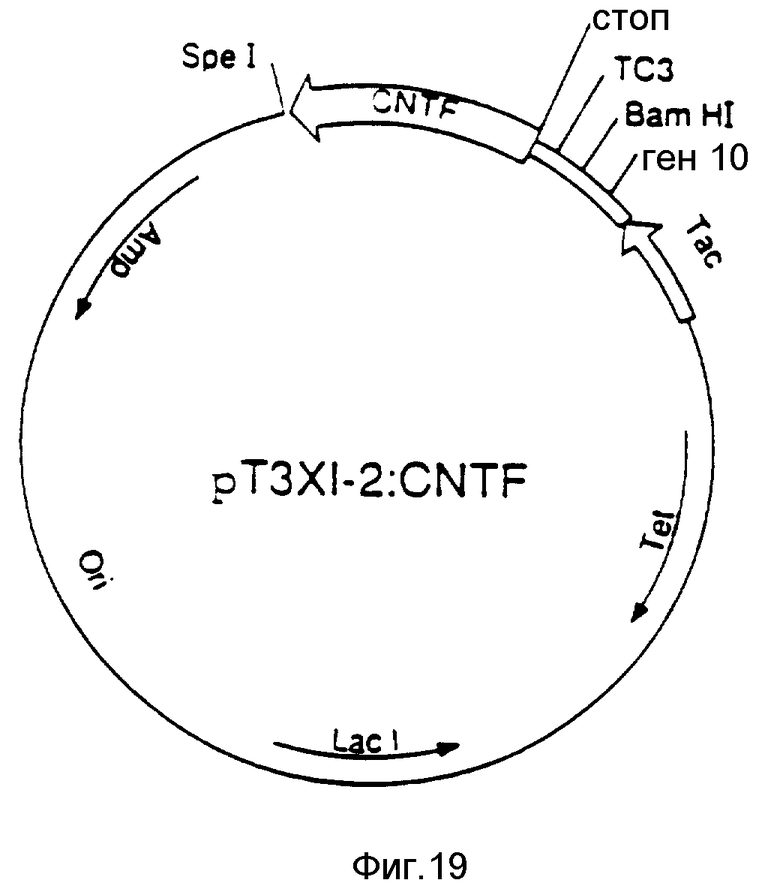


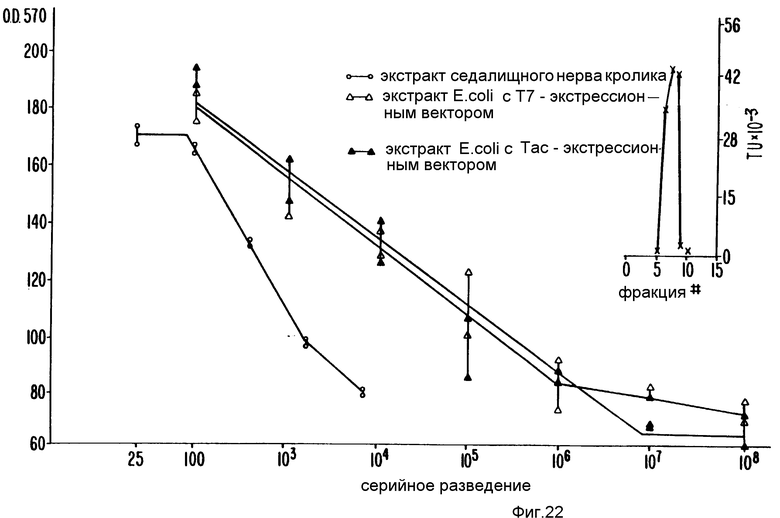
Клонирован и секвенирован фрагмент геномной ДНК из седалищного нерва человека, кодирующий цилиарный нейротрофный фактор (SN-CNTF). Получена последовательность ДНК, пригодная для экспрессии SN-CNTF человека в E.coli и сконструированы рекомбинантные векторы экспрессии, включающие названную последовательность. После трансформации указанными векторами подходящего штамма E.coli, отбора и культивирования трансформантов из бактериальных клеток выделили активную форму рекомбинантного SN-CNTF человека с мол. массой 24 кДа и N-концевой аминокислотной последовательностью, совпадающей с последовательностью нативного белка. 3 с. и 2 з.п. ф-лы, 1 табл, 22 ил.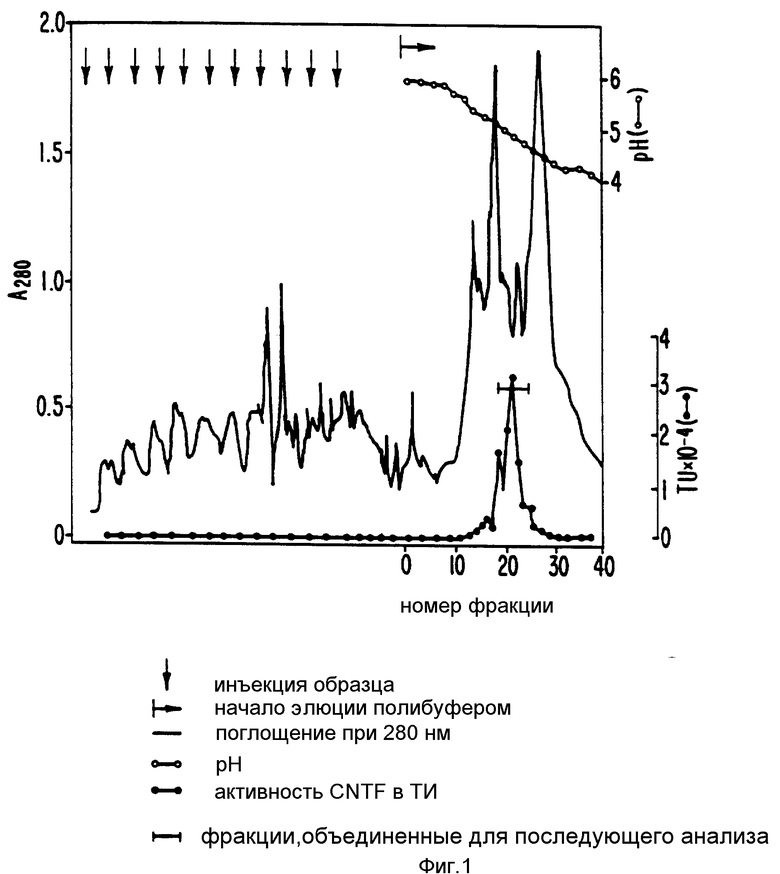
| Аналоги в патентной и научно-технической литературе не обнаружены. |
