Это изобретение относится к областям генной и белковой инженерии, молекулярной и клеточной биотехнологии и представляет собой укороченную мутантную люциферазу из Metridia longa (далее - ML-Green), выявляющую улучшенные свойства в сравнении с последовательностью дикого типа, для использования в качестве генетически-кодируемого биолюминесцентного репортера для визуализации молекулярных процессов в живых клетках, мечения белков, клеток и организмов.
Целентеразин-зависимая люцифераза из Metridia longa (MLuc) излучает свет при окислении низкомолекулярного субстрата целентаразина в простой реакции пары энзим-субстрат и кислорода. Катализируемая реакция окислительного декарбоксилирования приводит к образованию продукта реакции, целентерамида в возбужденном состоянии и СО2. Переход целентерамида из возбужденного состояния в основное сопровождается излучением света. Длина волны излучаемого света зависит от состояния эмиттера целентерамида, что в свою очередь определяется и белковым окружением. Простота реакции, легкость и неинвазивность регистрации квантов, отсутствие токсичности для живой клетки, широкий динамический диапазон, сопоставимый с радиоизотопной меткой, а также проницаемость клеточных мембран для субстрата целентеразина позволяют широко использовать целентеразин-зависимые люциферазы как генетически-кодируемые репортеры молекулярных процессов в живых клетках и организмах.
Люцифераза дикого типа из Metridia longa и ее известные варианты, как и другие целентеразин-зависимые люциферазы, имеют «голубую биолюминесценцию» с максимумом около 485 нм (фиг.2), что не является оптимальным для использования репортера в живых тканях, интенсивно поглощающих голубой свет. Собираемый свет от репортера в живых тканях, а следовательно, и его чувствительность зависят от количества излученных фотонов, попадающих в так называемое «окно прозрачности» биологических тканей в районе 600-800 нм. Таким образом, получение мутантных форм люциферазы со смещенным в длинноволновую область спектром излучения увеличивает чувствительность репортера за счет увеличения количества регистрируемых фотонов, проходящих сквозь животную ткань. Также очень желательными качеством для биолюминесцентного репортера, используемого in vivo, является его термостабильность.
Известны биолюминесцентные репортеры со сдвинутыми в длинноволновую область спектрами на основе целентеразин-зависимых люцифераз: мутанты люциферазы из Renilla reniformis (RLuc, ~36 кДа) [Loening AM et al., Nat. Methods., 2007].
Их основные недостатки заключаются в низкой биолюминесцентной активности полученных производных люциферазы RLuc (не удалось существенно увеличить низкий квантовый выход, характерный для люциферазы из Renilla reniformis), а также в большом молекулярном весе RLuc.
Известны мутанты люциферазы из Gaussia princeps (GLuc, ~18.2 кДа) [US 20120034672, 435/189, C12N 9/02, C12N 1/19, Oct 2011: (прототип)], которые показывали лучшую чувствительность при использовании в живых клетках в сравнении с своими дикими прототипами.
Недостатки этих мутантов заключаются в том, что они никак не охарактеризованы с точки зрения термостабильности при 37°С, кроме того, имеют меньший сдвиг спектра излучения в длинноволновую область - максимум в 503 нм [уточнение в Kim SB et al, Anal Chem., 2011].
Основной задачей, на решение которой направлено заявляемое изобретение, является расширение арсенала маркировочных средств, используемых в молекулярной и клеточной биологии, медицинской диагностике и биотехнологии.
Техническим результатом изобретения является получение на основе люциферазы MLM4 из Metridia longa биолюминесцентного репортера ML-Green размером ~16 кДа с повышенной чувствительностью и термостабильностью.
Технический результат достигается укороченной мутантной люциферазой из Metridia longa для применения в качестве биолюминесцентного репортера в живых клетках, обладающей повышенной термостабильностью и сдвигом спектра излучения в длинноволновую область, что повышает чувствительность в живых клетках, и характеризующейся введением аминокислотных замен I69L, Н74Е, K125V, W139F в сравнении с люциферазой MLM4 из Metridia longa.
Технический результат достигается путем введения точечных аминокислотных замен в природную последовательность MLM4 варианта люциферазы из Metridia longa размером ~16 кДа обладающей светоизлучающей функцией, в положениях 69, 74, 125 и 139 (нумерация по полноразмерной последовательности, включающей отщепляемый природный сигнальный пептид для секреции - Seq.2), соответствующих высоконсервативным аминокислотным остаткам (фиг.1). При этом в природной последовательности в положении 69 изолейциновый остаток заменен на лейциновый (I69L), в положении 74 гистидиновый остаток заменен на остаток глутаминовой кислоты (Н74Е), в положении 125 остаток лизина заменен на валиновый, и в положении 139 остаток триптофана заменен на фенилаланиновый (W139F). ML-Green получена мутагенезом из укороченной люциферазы MLM4 из Metridia longa размером ~16 кДа, являющейся на сегодняшний день наименьшей из существующих люцифераз. Данный вариант люциферазы MLM4 был получен путем делеционного мутагенеза из последовательности дикого типа люциферазы MLuc164 из Metridia longa и отличается от исходной полноразмерной последовательности повышенной в несколько раз биолюминесцентной активностью. При этом основные физико-химических свойства сохраняются, за исключением существенного снижения термостабильности. Аминокислотные замены для получения ML-Green привели к существенному повышению термостабильности и сдвигу спектра излучения в длинноволновую область с новым максимумом около 510 нм.
В заявляемом изобретении посредством сайт-направленного мутагенеза получен белок люциферазы ML-Green, молекула которого содержит аминокислотные замены, приводящие к существенному повышению термостабильности (практически не теряется активность при 37°С в течение суток) и сдвигу спектра в длинноволновую область, что как по отдельности, так и в сочетании обуславливает общее увеличение чувствительности биолюминесцентного репортера для живых клеток и тканей. Основными отличиями предлагаемого биолюминесцентного репортера ML-Green ~16 кДа для использования в живых клетках и организмах в сравнении с выбранным прототипом на основе мутантов люциферазы из Gaussia princeps (GLuc, ~18.2 кДа) являются другая аминокислотная последовательность с наименьшим размером среди известных люцифераз с другими мутациями, а также больший сдвиг спектра излучения в длинноволновую область - максимум в 510 нм в сравнении с 503 нм [Kim SB et al, Anal Chem., 2011], а также наличие термостабильности при 37°С как минимум в течение суток.
Таким образом, перечисленные выше отличительные от прототипа признаки позволяют сделать вывод о соответствии заявляемого технического решения критерию «новизна».
Признаки, отличающие заявляемое техническое решение от прототипа, не выявлены в других технических решениях и, следовательно, обеспечивают заявляемому решению соответствие критерию «изобретательский уровень».
Сущность изобретения поясняется с помощью графических материалов.
На фиг.1. представлено сравнение выровненных последовательностей (элаймент): ML-Green для эукариотической и прокариотической экспрессии, а также прототипы ML-Green: люцифераза дикого типа MLuc164 из Metridia longa и ее укороченный вариант MLM4. Мутации для получения ML-Green выделены серым цветом и звездочками внизу.
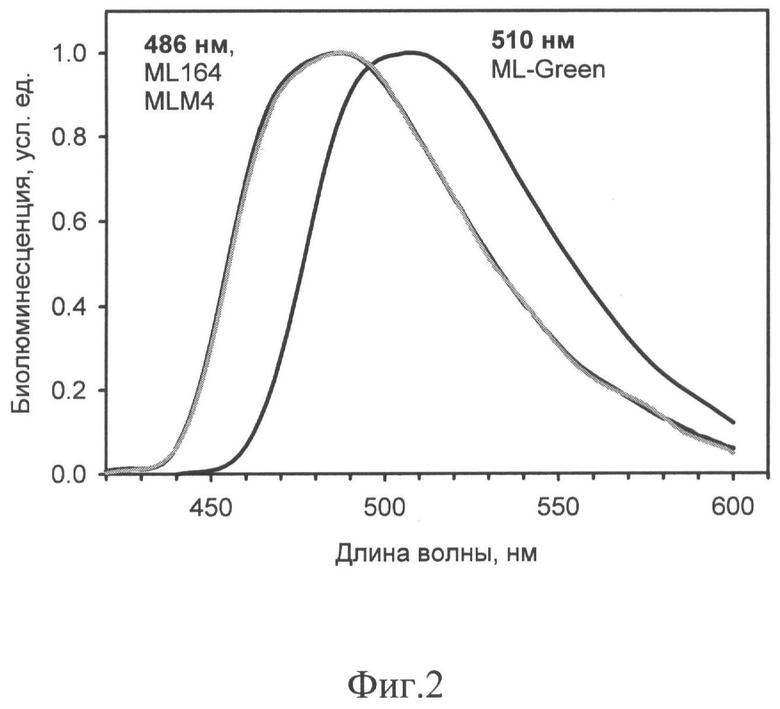
На фиг.2 представлены нормированные спектры биолюминесценции белка изобретения ML-Green в сравнении с природными последовательностями люциферазы дикого типа MLuc164 из Metridia longa и ее укороченного варианта (серая линия) MLM4.
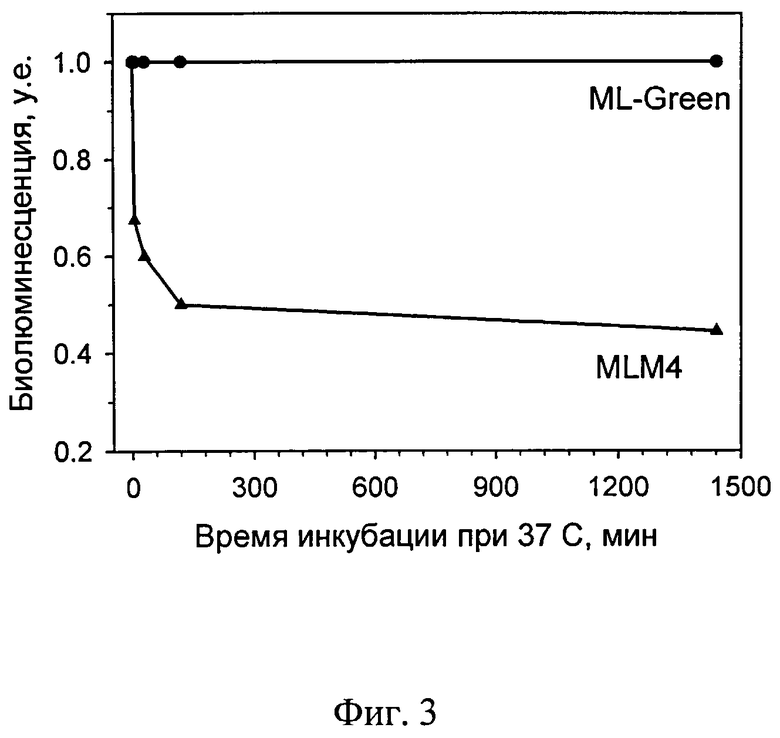
На фиг.3 представлена кинетика термоинактивации при 37°С мутантной люциферазы ML-Green (отмечено кружками) и ее прототипа - укороченной люциферазы MLM4 (кривая из треугольников).
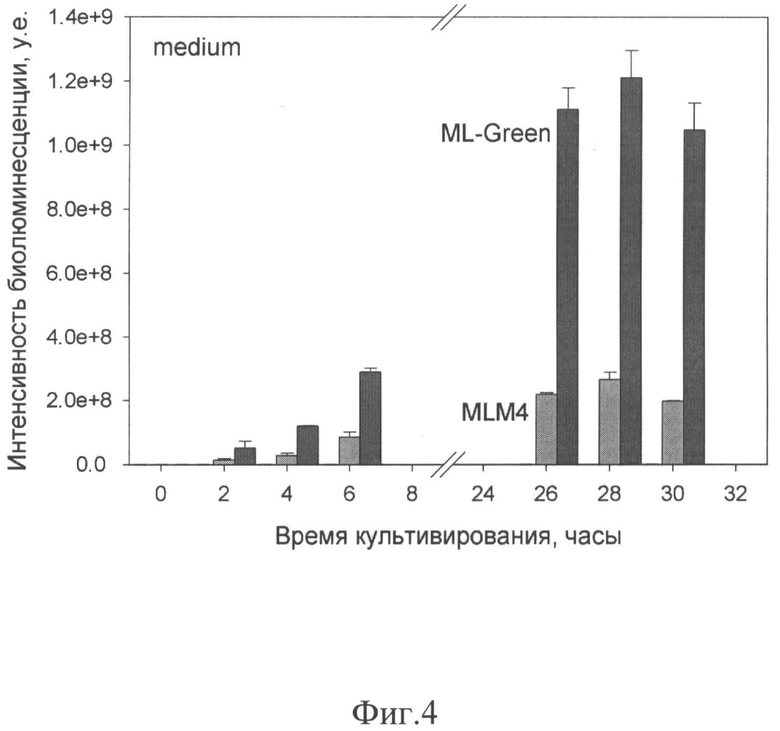
На фиг.4 представлен временной график биолюминесцентной активности в зависимости от времени роста клеток человека HEK293, синтезирующих белок изобретения ML-Green (черные столбики) и ее прототип - укороченную люциферазу MLM4 (серые столбики). Клетки, трансфецированные только векторной плазмидой, практически не выявляли биолюминесценции, поэтому этих данных не видно на графике. Каждая точка - результат трех независимых измерений по трем лункам с указанным средним квадратичным отклонением.
Описание представленных аминокислотных и ДНК последовательностей
Последовательность №1 представляет зрелую последовательность ML-Green без отщепляемого природного сигнального пептида для секреции. Такая же последовательность, дополненная метионином впереди, используется для бактериальной наработки люциферазы для определения биолюминесцентных свойств.
Последовательность №2 представляет полноразмерную последовательности ML-Green, включающую отщепляемый природный сигнальный пептид для секреции.
Последовательность №3 представляет ген, кодирующий полноразмерную последовательности ML-Green с отщепляемым природным сигнальным пептидом для секреции, включающий некодирующие области для успешной экспрессии в эукариотических клетках
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение мутантной люциферазы ML-Green (Seq. 1) с улучшенными репортерными свойствами
Генно-инженерные конструкции. Все генно-инженерные манипуляции, если не оговорено особо, были выполнены с помощью методов, хорошо известных для специалистов в данной области. Для получения мутантной люциферазы ML-Green с улучшенными свойствами в последовательности MLuc были выбраны консервативные аминокислотные остатки, предположительно участвующие в формировании активного центра, получен ряд мутантов и после характеризации многочисленных мутантов выбраны отдельные мутации, формирующие желаемые свойства. Сайт-направленный мутагенез для получения мутантной люциферазы ML-Green проводился методом ПЦР с использованием набора для направленного на сайт мутагенеза QuikChange® Site-Directed Mutagenesis Kit (Stratagene, США), следуя руководству к набору. В качестве матрицы использовали плазмиду, содержащую полноразмерный ген кДНК ген укороченной люциферазы MLM4 из Metridia longa размером ~16 кДа, являющуюся на сегодняшний день наименьшей из существующих люцифераз [Markova SV et al, Biochem. Biophys. Res. Commun., 2012]. Праймеры для мутагенеза, содержащие измененные кодоны для введения единичных аминокислотных замен, были синтезированы в ООО «Синтол» (Россия). 4 аминокислотных замены было получены путем последовательных раундов мутагенеза с одновременным введением не более двух мутаций. Все полученные мутации были подтверждены секвенированием полученной ДНК.
Для получения белка мутантной люциферазы ML-Green в достаточных количествах с целью характеризации свойств использовали наработку рекомбинантного белка в бактериальных клетках с экспрессионной плазмидой pET22b+ (Novagen). Последовательность, кодирующую зрелую мутантную люциферазу ML-Green (Seq. 1 - без первых 17 аминокислотных остатков сигнального пептида для секреции), амплифицировали в ходе ПЦР праймерами, в последовательности которой были введены сайты для клонирования NdeI и XhoI, а также стартовый метиониновый остаток в самое начало. Далее последовательность мутантной люциферазы ML-Green для бактериальной экспрессии была клонирована в соответствующие сайты pET22b+. Для наработки белка мутантной люциферазы ML-Green полученная плазмида была трансформированы в штамм Е.coli BL21-CodonPlus (DE3)-RIPL (Stratagene).
Получение препарата белка ML-Green для характвризации. Клетки Е.coli BL21 (DE3) Codon Plus (RIPL), содержащие экспрессионную плазмиду с ML-Green, наращивали в среде LB, содержащей ампициллин (200 мкг/мл), при 37°С до достижения оптической плотности OD600=0.8-1.0. Индукцию синтеза проводили добавлением ИПТГ до конечной концентрации 1 мМ, а затем культивировали с интенсивным перемешиванием в течение 2 часов при 37°С. При экспресиии в клетках Е.coli в экспрессионном векторе pET22b+ (Novagen), рекомбинантные белки MLuc164 и ее мутанты образуют нерастворимые тельца включения, из которых и проводили очистку мутантной люциферазы ML-Green последующим рефолдингом. Клеточный осадок суспендировали в пятикратном объеме буфера (20 мМ Трис-HCl, рН 8.8) и разрушали ультразвуком (1 мин × 6) при 0°С. После центрифугирования (5000 g, 15 мин) тельца включения 3-кратно последовательно промывали 0.9% NaCl, 20 мМ Трис-HCl, рН 8.8, и 2 М раствором мочевины с 0.1% Тритон Х-100. Чистоту телец включения контролировали с помощью электрофореза в ПААГ.
Тельца включения растворяли в буфере 6 М гуанидин-HCl, 1 мМ ЭДТА, 20 мМ Трис-HCl, рН 8.8. Для корректного фолдинга белка 1 мл экстракта по каплям добавляли к 50 мл буфера, содержащего 5 мМ цистеина, 0,5 мМ цистина, 50 мМ NaCl, 1 мМ ЭДТА, 20 мМ Трис-HCl, рН 8,8, и оставляли на 16 часов при 4°С. Полученный раствор концентрировали центрифугированием на ячейках Amicon-Ultra (Millipore) до объема 1 мл. 500 мкл наносили на 30-мл колонку Superdex-200 (GE Healthcare) и элюировали буфером (1 мМ ЭДТА, 0,5 М NaCl, 20 мт Трис-HCl, рН 7,5). Чистоту полученного белка контролировали с помощью электрофореза в ПААГ. Содержание целевого белка составило не менее 99%.
Мономерная фракция была использована для тестирования физико-химических свойств мутантов.
Пример 2. Основные биолюминесцентные свойства мутантной люциферазы ML-Green
Полученный препарат мутантной люциферазы ML-Green анализировали по спектрам биолюминесценции, удельной биолюминесцентной активности, термостабильности в сравнении с исходной укороченной MLM4 и полноразмерной MLuc164 люциферазами Metridia longa дикого типа.
Спектр биолюминесценции. Спектр биолюминесценции измеряли на спектрофлуориметре Сагу Eclipse (Varian Cary, Австралия) при температуре 17°С. Спектр записывали в растворе, содержащем 0.1 М NaCl, 50 мМ Трис-HCl, рН 7,5. Образцы выравнивали по концентрации белка, которая в кювете составляла 0.01 мг/мл. Биолюминесцентную реакцию запускали добавлением 10-кратного избытка целентаразина, спектры записывали при минимальном падении интенсивности сигнала, не требующего коррекции спектра на падение интенсивности света. Коррекция спектров на биолюминесценции на спектральную чувствительность ФЭУ была проведена с помощью программного обеспечения прибора.
Нормированные спектры всех трех люцифераз представлены на фиг.2, указанные максимумы излучения приведены также в Таблице 1. Как можно видеть из приведенных спектров, мутантная люцифераза ML-Green имеет существенный спектральный сдвиг в длинноволновую область в сравнении с оригинальными люциферазами: новый максимум лежит в области 510 нм.
Удельная биолюминесцентная активность. Удельную биолюминесцентную активность определяли, впрыскивая 5 мкл спиртового раствора целентеразина в кювету биолюминометра, содержащую 500 мкл раствора белка с конечной концентрацией 0.0006 мг/мл в 50 мМ Трис-HCl, рН 7.5, и 0.1 М NaCl. Измерения проводили при 10°С. Молярное соотношение белок:целентеразин было 1:1.5. Запись проводили до полного падения биолюминесцентного сигнала. Концентрацию белка определяли по методу Лоури.
Как можно видеть из таблицы, мутантная люцифераза ML-Green по изобретению сохраняет свою активность практически такой же как у исходной MLM4 люциферазы, что более чем в 4 раза выше по тотальному свету, чем у люциферазы дикого типа MLuc164.
Термостабильность. Для исследования термоинактивации белков при разных температурах, аликвоты, содержащие по 100 мкл раствора белка в 1 мМ ЭДТА, 0,5 М NaCl, 50 мМ Трис-HCl, рН 7,5, с концентрацией 0.03 мг/мл, инкубировали 5, 30, 120 и 1440 минут при температуре 37°С. При этом, в отличие от исходной MLM4 люциферазы, мутантная люцифераза ML-Green по изобретению была более термостабильна. Если потеря биолюминесцентной активности для исходной люциферазы MLM4 составила 50% за два часа, то для ML-Green при температуре 37°С наблюдалось стабильное сохранение биолюминесцентной активности в течение суток - фиг.3.
Пример 3. Применение мутантной люциферазы ML-Green как секретируемого биолюминесцентного репортера в клетках человека HEK293
Для тестирования мутантной люциферазы ML-Green как биолюминесцентного репортера могут быть использованы, например, клетки почки эмбриона человека линии HEK293, транзиентно трансфецированные, например, векторной плазмидой pcDNA3.1 (Invitrogene), в которую лигирована ДНК, характеризующаяся Seq.3 - полноразмерный кДНК ген мутантной люциферазы ML-Green. Плазмида pcDNA3.1 является вектором для экспрессии целевого белка в клетках млекопитающих под сильным конститутивным цитомегаловирусным промотором. В данном случае продукт экспрессии мутантная люцифераза ML-Green будет секретирована в культуральную среду.
Секретируемый репортер является очень ценным инструментом исследований, поскольку позволяет анализировать внутриклеточные события без разрушения клеток и тканей путем простого отбирания аликвот межклеточной среды. Полноразмерный ген (Seq.3), кодирующий мутантную люциферазу ML-Green (Seq.2) с отщепляемым природным сигнальным пептидом для секреции и включающий некодирующие области для экспрессии в эукариотических клетках, был полученный в ходе примера 1. Для эукариотической экспрессии полноразмерный ген (Seq.3), кодирующий мутантную люциферазу ML-Green, был синтезирован в ходе ПЦР со специфическими праймерами с введенными сайтами рестрикции KpnI и XhoI, затем вставлен по соответствующим сайтам в плазмиду pcDNA3.1.
Аналогичным образом была получена конструкция с исходным геном люциферазы MLM4 для сравнения. В этих конструкциях присутствовал природный сигнальный пептид для секреции Metridia люциферазы. Трансформацию плазмид в клетки HEK293 проводили с помощью трансфецирующего агента Lipofectamine 2000 (Invitrogen) согласно рекомендациям производителя. Трансфекцию ставили в 96-тилуночном формате, когда клетки достигали приблизительно 90% конфлюэнтности, множественные повторы были сделаны для усреднения результата.
Клетки HEK293, трансфецированные генетическими конструкциями pcDNA3.1-MLM4, pcDNA3.1-ML-Green и pcDNA3.1 как контроль, были проанализированы на биолюминесцентную активность в присутствии целентеразина, так же как и культуральная среда клеток в зависимости от времени культивирования. Клеточные экстракты были получены двукратным замораживанием-оттаиванием. Результаты представлены на графике зависимости величины биолюминесцентного сигнала от времени после трансфекции (Фиг.4). Как видно из приведенного графика, трансфецированные клетки секретировали исследуемые люциферазы непрерывно, вследствие чего биолюминесцентная активность непрерывно возрастала в культуральной среде, в то время как внутриклеточная люциферазная активность возрастала незначительно. В контрольных клетках, трансфецированных только векторной плазмидой, практически не наблюдалось никакой биолюминесцентной активности. Интенсивные биолюминесцентные сигналы в клетках, трансфецированных люциферазными генами, были получены уже через 2 часа после трансфекции, причем сигнал от мутантной люциферазы ML-Green возрастал в течение суток как минимум в 3 раза интенсивнее, чем от природной укороченной MLM4 люциферазы (фиг.4). Несомненно, термостабильность ML-Green обуславливает более высокие сигналы вследствие отсутствия термоинактивации накапливающихся молекул, следовательно, более высокую чувствительность ML-Green как биолюминесцентного репортера в живых клетках.
Мутантная люцифераза ML-Green с улучшенными свойствами для использования в живых клетках, представляет собой другую аминокислотную последовательность размером ~16 кДа, наименьшую из известных люцифераз обладающую термостабильностью при 37°С и сдвинутым спектром излучения в длинноволновую область (max ~510 нм). Малый размер является преимуществом для генетически-кодируемого репортера, приводя к снижению биосинтетической нагрузки на клетку и также, как правило, к улучшенному фолдингу рекомбинантного белка в чужеродном окружении.
Мутантная люцифераза ML-Green является новой биолюминесцентный репортерной молекулой для оценки молекулярных процессов в живых клетках, тканях и организмах, что расширяет арсенал репортеров в молекулярной биологии и биотехнологии, а также увеличивает чувствительность измерений.


Изобретение относится к биохимии и представляет собой укороченную мутантную люциферазу MLM4 из Metridia longa размером ~16 кДа с улучшенными свойствами для использования в качестве генетически-кодируемого биолюминесцентного репортера для визуализации молекулярных процессов в живых клетках. Аминокислотные замены I69L, H74E, K125V, W139F введены в последовательность укороченной люциферазы MLM4 из Metridia longa размером ~16 кДа. Изобретение позволяет получить люциферазу с повышенной термостабильностью и со сдвигом спектра в длинноволновую область, что обуславливает общее увеличение чувствительности биолюминесцентного репортера при использовании в живых клетках и организмах. 4 ил., 1 табл, 3 пр.
Укороченная мутантная люцифераза из Metridia longa для применения в качестве биолюминесцентного репортера в живых клетках, обладающая повышенной термостабильностью и сдвигом спектра излучения в длинноволновую область, что повышает чувствительность в живых клетках, и характеризующаяся введением аминокислотных замен I69L, Н74Е, K125V, W139F в укороченную люциферазу MLM4 из Metridia longa.
| US 20120034672 A1, 09.02.2012 | |||
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ЛЮЦИФЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ИЗОЛИРОВАННАЯ НУКЛЕИНОВАЯ КИСЛОТА, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, НАБОР ДЛЯ ИСПОЛЬЗОВАНИЯ В БИОЛЮМИНЕСЦЕНТНОМ АНАЛИЗЕ, АНАЛИТИЧЕСКИЙ ТЕСТ ДЛЯ ОПРЕДЕЛЕНИЯ ПРИСУТСТВИЯ В ОБРАЗЦЕ СоА | 2000 |
|
RU2251571C2 |
| RU 2009130108 A, 10.02.2011 | |||
| MARKOVA S.V | |||
| Cloning and expression of cDNA for a luciferase from the marine copepod Metridia longa | |||
| A novel secreted bioluminescent reporter enzyme | |||
| J Biol Chem | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

