Изобретение относится к области генной и белковой инженерии и может быть использовано для детекции D-аминокислот в сложных образцах и в процессах ферментативного синтеза оптически активных веществ, альфа-кетокислот, цефалоспориновых антибиотиков и других органических соединений с использованием оксидаз.
Получены новые мутантные формы оксидазы D-аминокислот (DAAO), характеризующиеся субстратной специфичностью, отличной от субстратной специфичности фермента дикого типа, по отношению к ряду D-аминокислот и их производных. В качестве основы для получения новых мутантных форм DAAO использовалась последовательность кДНК, кодирующая аминокислотную последовательность оксидазы D-аминокислот из дрожжей Trigonopsis variabilis. Предложенные мутантные формы характеризуются тем, что природный аминокислотный остаток цистеина в положении, соответствующем положению 108 в аминокислотной последовательности оксидазы D-аминокислот из Trigonopsis variabilis, заменен на остаток аланина, или остаток фенилаланина, или остаток серина. Данные замены могут быть использованы в комбинации с другими мутациями, обеспечивающими приобретение оксидазами D-аминокислот новых или улучшенных свойств.
Настоящее изобретение относится к новым белкам, в частности мутантным оксидазам из дрожжей, которые обладают субстратной специфичностью, отличной от субстратной специфичности фермента дикого типа, по отношению к ряду D-аминокислот и их производных, к кДНК, кодирующей мутантные оксидазы D-аминокислот, к применению данных ферментов в аналитических тестах на содержание D-аминокислот в биологических и иных образцах, к наборам, включающим их, и в процессах биокаталитического получения оптически активных веществ, альфа-кетокислот и других органических соединений, окисления цефалоспорина С и его производных с использованием оксидаз.
ФАД-зависимая оксидаза D-аминокислот катализирует окислительное дезаминирование D-аминокислот до соответствующих α-кетокислот с образованием иона аммония и пероксида водорода:
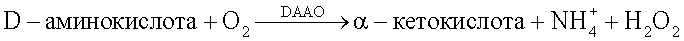
Исключительно высокую специфичность оксидазы D-аминокислот к D-формам аминокислот применяют для выделения L-изомеров и ферментативного синтеза α-кетокислот из дешевых рацемических смесей аминокислот. Это используют, в частности, при синтезе синтонов - исходных соединений для получения оптически чистых лекарственных препаратов. В качестве примера можно привести процесс получения Омапатрилата - ингибитора ангеотензинпревращающего фермента из рацемической смеси 6-гидроксинорлейцина с использованием оксидазы D-аминокислот из дрожжей Trigonopsis variabilis (Patel R.N. Enzymatic synthesis of chiral intermediates for Omapatrilat, an antihypertensive drug. Biomol. Eng., 2001, v.17, №6, pp.167-182). Также фермент активно применяется в аналитической биотехнологии для детекции D-аминокислот в биологических жидкостях, например спинно-мозговой жидкости человека (Wcislo M., Compagnone D., Trojanowicz M. Enantioselective screen-printed amperometric biosensor for the determination of D-amino acids. Bioelectrochem., 2006, v.71, pp.223-230,). Помимо этого, необратимость реакции, катализируемой этим ферментом, и специфическая активность оксидазы D-аминокислот позволяют ее использовать для превращения цефалоспорина С в 7-аминоцефалоспорановую кислоту, которая является исходным соединением при синтезе полусинтетических цефалоспориновых антибиотиков различных поколений (Pilone M.S., Buto S., Pollegioni L. A process for bioconversion of cephalosporin С by Rhodotorula gracilis D-amino acid oxidase. Biotechnol. Lett., 1995, v.17, №2, pp.199-204).
Оксидазы D-аминокислот широко распространены в природе и могут быть выделены из разнообразных микроорганизмов (бактерий, дрожжей, микроскопических грибов, водорослей), а также моллюсков, рыб, рептилий, птиц и млекопитающих. Подробный список известных источников оксидаз D-аминокислот можно найти в обзоре (Тишков В.И., Хороненкова С.В. Оксидаза D-аминокислот: структура, механизм действия и практическое применение. Биохимия, 2005, т.70, №1, с.51-67). Многие из генов, кодирующих данные ферменты, были клонированы и секвенированы, поэтому их можно получать с использованием технологии рекомбинантных ДНК. Последовательность кДНК, кодирующую оксидазу D-аминокислот, встраивают в генно-инженерный вектор (плазмида, фаг и т.д.), который используют для трансформации микроорганизмов, например бактерии E.coli или метилотрофные дрожжи Hansenula polymorpha, затем экспрессирующие желаемый ферментный продукт.
Основными недостатками известных природных и рекомбинантных оксидаз D-аминокислот дикого типа являются:
1) во-первых, неоптимальный спектр субстратной специфичности, что проявляется или в низкой каталитической активности с рядом природных и неприродных субстратов, или в неоптимальном соотношении активностей между различными D-аминокислотами. Например, с D-аланином большинство оксидаз D-аминокислот проявляют очень близкую или даже более высокую активность каталитической активностью, чем с D-серином. Поэтому такие ферменты не могут быть использованы в биосенсорах для селективного определения D-серина в сложных биологических объектах (например, в спинно-мозговой жидкости) в присутствии сравнимых концентраций D-аланина. В случае с цефалоспорином С каталитической активностью обладают только ферменты из дрожжей T.variabilis и Rhodotorula gracilis, причем в случае последнего фермента эта активность не очень высока (Pollegioni L., Caldinelli L., Molla G. et al. Catalytic properties of D-amino acid oxidase in cephalosporin С bioconversion: a comparison between proteins from different sources. Biotechnol. Prog., 2004, v.20, №2, pp.467-473).
2) вторым важным недостатком является невысокая температурная и операционная стабильность ферментов. Среди известных оксидаз D-аминокислот наиболее стабильным при высоких температурах является фермент из дрожжей T.variabilis (TvDAAO) (Тишков В.И., Хороненкова С.В. Оксидаза D-аминокислот: структура, механизм действия и практическое применение. Биохимия, 2005, т.70, №1, с.51-67).
Благодаря наилучшей среди оксидаз D-аминокислот каталитической активности с цефалоспорином С наиболее перспективной для практического применения в процессах синтеза цефалоспориновых антибиотиков в настоящее время является оксидаза D-аминокислот из T.variabilis. Однако для практического применения фермента необходимо улучшать его каталитические характеристики с цефалоспорином С и стабильность. Одним из подходов для решения поставленной задачи является мутагенез аминокислотных остатков целевого белка.
В настоящее время наибольшее количество экспериментов по мутагенезу оксидазы D-аминокислот выполнено для рекомбинантного фермента из дрожжей R.gracilis (см. обзор Pollegioni L., Piubelli L., Sacchi S., Pilone M.S., Molla G. Physiological functions of D-amino acid oxidases: from yeast to humans. Cell. Mol. Life Sci., 2007, v.64, №11, pp.1373-1394). Для фермента из T.variabilis подобных работ опубликовано всего 3. В двух работах с помощью метода направленного мутагенеза изучали роль консервативных остатков His324 и Asp206 в катализе и связывании ФАД (Lin L.L., Wang W.C., Ju S.S. The role of a conserved histidine residue, His324, in Trigonopsis variabilis D-amino acid oxidase. FEMS Microbiol. Lett., 1999, v.176, №2, pp.443-448; Ju S.S., Lin L.L., Wang W.C., Hsu W.H. A conserved aspartate is essential for FAD binding and catalysis in the D-amino acid oxidase from Trigonopsis variabilis. FEBS Lett., 1998, v.436, №1, pp.119-122). Замены по остаткам His324 и Asp206 сильно ухудшали кинетические свойства или приводили к полной потере активности. В третьей работе направленный мутагенез в молекуле TvDAAO шести остатков метионина на остаток лейцина показал, что эти замены повышают стабильность фермента против инактивации пероксидом водорода (Ju S.S., Lin L.L., Chien H.R., Hsu W.H. Substitution of the critical methionine residues in Trigonopsis variabilis D-amino acid oxidase with leucine enhances its resistance to hydrogen peroxide. FEMS Microbiol. Lett., 2000, v.186, №2, pp.215-219). Однако четыре мутации (Met104Leu, Met226Leu, Met245Leu и Met339Leu) приводили к резкому ухудшению кинетических свойств с D-аланином в качестве субстрата. В случае мутации Met209Leu ухудшилась константа Михаэлиса КМ (увеличение в 3,3 раза). В результате замены Met156Leu значение КМ возросло в 1,5 раза, однако за счет увеличения каталитической константы kcat в 3 раза общая каталитическая эффективность (отношение kcat/КМ) повысилась в два раза. Субстратная специфичность полученных мутантов с другими D-аминокислотами не исследовалась.
Методом химической модификации было показано, что модификация или окисление остатков цистеина в положении 298 и 108 приводит к полной или 75% потере активности соответственно (Schrader T., Andreesen J.R. Evidence for the functional importance of Cys298 in D-amino acid oxidase from Trigonopsis variabilis. Eur. J. Biochem., 1993, v.218, №2, pp.735-744 и Slavica A., Dib I., Nidetzky B. Single-site oxidation, cysteine 108 to cysteine sulfinic acid, in D-amino acid oxidase from Trigonopsis variabilis and its structural and functional consequences. Appl. Environ. MicrobioL, 2005, v.71, №12, pp.8061-8068).
Техническая задача, решаемая посредством предлагаемого технического решения, состоит в получении мутантных форм оксидазы D-аминокислот из Т.variabilis с субстратной специфичностью, отличной от субстратной специфичности фермента дикого.
Технический результат, получаемый при реализации предлагаемого технического решения, состоит в изменении субстратной специфичности полученных форм оксидазы D-аминокислот по отношению к ряду D-аминокислот и их производных.
Для достижения указанного технического результата предложено использовать рекомбинантную оксидазу D-аминокислот из Т.variabilis, характеризующуюся аминокислотной последовательностью, которая соответствует аминокислотной последовательности оксидазы D-аминокислот из T.variabilis дикого типа с заменой природного остатка цистеина в положении 108 или остатком аланина, или остатком фенилаланина, или остатком серина. Введение одного из трех перечисленных остатков обеспечивает изменение субстратной специфичности модифицированного фермента по отношению к ряду D-аминокислот и их производных. Кроме того, помимо замены природного остатка цистеина в положении 108 на остаток аланина, или остаток фенилаланина, или остаток серина аминокислотная последовательность мутантной оксидазы D-аминокислот из T.variabilis может содержать, по меньшей мере, одну дополнительную замену, обеспечивающую улучшение термостабильности, химической стабильности, изменение кинетических свойств, попарное или одновременное улучшение всех вышеуказанных свойств по сравнению с аминокислотной последовательностью оксидазы D-аминокислот из T.variabilis дикого типа.
В дальнейшем способ получения и преимущества полученных модифицированных оксидаз D-аминокислот будут рассмотрены с использованием примеров его реализации.
Пример 1.
Получение мутантной оксидазы D-аминокислот Trigonopsis variabilis с заменой Cys108Ala методом направленного мутагенеза.
Все генно-инженерные манипуляции, если не оговорено особо, осуществлены в соответствии с Манниатис Е., Фрич. Э., Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984, или в соответствии с инструкциями фирм-производителей приборов, наборов и других реагентов. Все эти методы хорошо известны специалистам в данной области.
Направленный мутагенез в гене daao из T.variabilis проводили с использованием полимеразной цепной реакции (ПЦР). В качестве матрицы для проведения направленного мутагенеза использовали плазмиду pKanDAAO, в которой ген оксидазы D-аминокислот находится под контролем сильного промотора РНК-полимеразы фага Т7. Плазмида pKanDAAO получена из коммерчески доступной плазмиды рЕТ33b (Novagen, США), в которую по сайтам рестрикции Ncol и BamHI встроен ген daao из T.variabilis (подробно получение этой плазмиды см. в Давыдова Е.Е., Тишков В.И. Клонирование гена оксидазы D-аминокислот из дрожжей Trigonopsis variabilis. Вестник Московского Университета, Серия 2. Химия, 2002, v.43, №6, с.353-355). Плазмида была выделена из клеток E.coli с использованием набора для выделения плазмид NucleoSpin® Plus согласно инструкции фирмы-производителя (Macherey-Nagel GmbH & Со. KG).
Для введения мутации в ген daao из T.variabilis с помощью ПЦР использовали праймеры на начало и конец гена:
DAOForl
5'-АТА ТАС CAT GGC ТАА ААТ CGT TGT TAT TGG TGC-3'
DAORev4
5'-GGA AGA GAG TTT CGA АСА AGG ACG AC-3',
а также прямой и обратный праймеры, несущие требуемую замену в триплете, кодирующем в гене daao остаток Cys в положении 108:
DAO_C108Afor
5'-G TCG GCC АТС GCT CAA CGC AAC CCC TGG TTC AA-3'
DAO_C108Arev
5'-GTT GCG TTG AGC GAT GGC CGA CAT GGC ACC TTC-3'.
Жирным шрифтом выделен триплет, обеспечивающий мутацию Cys108Ala.
Реакцию ПЦР проводили в тонкостенной пластиковой пробирке объемом 0,5 мл (Eppendorf, Германия). Для предотвращения испарения реакционной смеси и ее конденсации на крышке в пробирку добавляли 30 мкл минерального масла. Пробирку прогревали 5 мин при 95°С и затем проводили реакцию ПЦР по следующей программе: 1-я стадия - 95°С, 60 с; 2-стадия - 56°С, 60 с и 3-я стадия - 72°С, 2 мин, всего 25-35 циклов. После этого реакционную смесь выдерживали еще 10 мин при 72°С. Температуру на второй стадии выбирали на 3-5 градусов ниже Тm температуры плавления дуплексов, образуемых праймерами.
Реакционная смесь для проведения ПЦР имела следующий состав (таблица 1):
Условия проведения полимеразной цепной реакции (ПЦР).
На первом этапе проводили две полимеразные цепные реакции с использованием следующих пар праймеров: 1) DAOForl+DAO_C108Arev и 2) DAO_C108Afor+DAORev4. В качестве матрицы использовали плазмиду pKanDAAO. Полученные продукты ПЦР, фрагмент 1 и фрагмент 2, очищали с помощью специального набора, например QIAquick PCR Purification Kit (QIAGEN). Затем проводили третью объединяющую ПЦР с праймерами DAOForl и DAORev4, где в качестве ДНК-матрицы использовали полученные ранее фрагменты 1 и 2.
Продукт третьей ПЦР очищали с помощью препаративного электрофореза в агарозном геле с использованием предназначенных для этого наборов (например, NucleoSpin Extract (Macherey-Nagel GmbH & Co. KG)). Очищенный ПЦР-продукт обрабатывали рестриктазой Bsp19 в течение 2 часов при 37°С, затем инкубировали рестрикционную смесь для инактивации рестриктазы в течение 20 минут при 65°С и обрабатывали рестриктазой Bso31 в течение 2 часов при 55°С. В эксперименте были использованы рестриктазы и буфера фирмы Сибэнзим (Россия). Затем полученные фрагменты ДНК очищали от коротких фрагментов препаративным электрофорезом в агарозном геле с последующей экстракцией нужной полосы из агарозы с использованием специального набора, например NucleoSpin Extract (Macherey-Nagel GmbH & Co. KG) и лигировали с фрагментом исходной плазмиды pKanDAAO, из которой расщеплением по тем же сайтам рестрикции Bsp19 и Bso31 был удален фрагмент гена daao из T.variabilis дикого типа. Полученной после лигирования реакционной смесью трансформировали клетки Е.coli TG1 (генотип supE hsdΔ5 thi Δ (lac-proAB) F'[traD36 proAB+ lacIq lacZΔM15]) и выращивали в течение ночи при 37°С на твердой агаризованной среде LB, содержащей канамицин в концентрации 30 мкг/мл (плазмида pKanDAAO содержит ген устойчивости к канамицину). Выросшие колонии инокулировали в 6-8 мл среды 2YT, содержащей 30 мкг/мл канамицина, и растили при 37°С в течение 8-10 часов при аэрировании. Плазмидную ДНК выделяли из полученной культуры клеток с использованием предназначенного для этого набора, например QIAprep Spin Miniprep (QIAGEN). Для контроля введения требуемых мутаций проводили секвенирование плазмидной ДНК с помощью набора реактивов ABI PRISM® BigDye™ Terminator v.3.1 (Perkin Elmer Applied Biosystems), который основан на дидезокситерминационном методе [Sanger F., Nicklen S., Coulson A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA, 1977, v.74, №12, pp.5463-5467] с последующим анализом продуктов реакции на автоматическом секвенаторе ДНК ABI PRISM 3100-Avant. В результате из 5 выделенных плазмид все 5 содержали только требуемую замену Cys108Ala. Таким образом, был получен новый мутант TvDAAO Cys108Ala (плазмида pKanDAAO_C108A).
Для получения мутантного фермента клетки E.coli BL21 (DЕ3) pLysS Codon Plus (генотип Е.coli В F- ompT hsdS(rB- mB-) dcm+ Tetr gal λ(DE3)pLysS endA Hte [argU proL Camr) трансформировали плазмидой pKanDAAO_C108A и проводили их культивирование (см. ниже).
Пример 2. Получение мутантной оксидазы D-аминокислот T.variabilis с заменой Cys108Phe с использованием метода направленного мутагенеза.
Данный пример идентичен примеру 1 за исключением того, что в качестве прямого и обратного праймеров, несущих требуемую замену в положении 108 гена daao, использовали следующие дезоксирибоолигонуклеотиды:
DAO_C108Ffor
5'-G TCG GCC АТС ТТС САА CGC AAC CCC TGG ТТС АА-3'
DAO_C108Frev
5'-G GTT GCG TTG САА GAT GGC CGA CAT GGC АСС ТТС-3'.
Жирным шрифтом выделен триплет, обеспечивающий мутацию Cys108Phe.
Плазмида pKanDAAO с нуклеотидными заменами, обеспечивающими в гене фермента мутацию Cys108Phe, получила название pKanDAAO_C108F.
Для получения мутантного фермента клетки E.coli BL21 (DE3) pLysS Codon Plus (генотип Е.coli В F- ompT hsdS(rB- mB-) dcm+ Tetr gal λ(DE3)pLysS endA Hte [argU proL Camr]) трансформировали плазмидой pKanDAAO_C108F и проводили их культивирование (см. ниже).
Пример 3. Получение мутантной оксидазы D-аминокислот Trigonopsis variabilis с заменой Cys108Ser с использованием метода направленного мутагенеза.
Данный пример идентичен примеру 1 за исключением того, что в качестве прямого и обратного праймеров, несущих требуемую замену в положении 108 гена daao, были использованы следующие дезоксирибоолигонуклеотиды:
DAO_C108Sfor
5'-G TCG GCC АТС ТСТ САА CGC AAC CCC TGG ТТС АА-3'
DAO_C108Srev
5'-G GTT GCG TTG AGA GAT GGC CGA CAT GGC ACC TTC-3'.
Жирным шрифтом выделен триплет, обеспечивающий мутацию Cys108Ser.
Плазмида pKanDAAO с нуклеотидными заменами, обеспечивающими в гене фермента мутацию Cys108Ser, получила название pKanDAAO_C108S.
Для получения мутантного фермента клетки E.coli BL21 (DE3) pLysS Codon Plus (генотип Е.coli В F- ompT hsdS(rB- mB-) dcm+ Tetr gal λ(DE3)pLysS endA Hte [argU proL Camr) трансформировали плазмидой pKanDAAO_C108S и проводили их культивирование (см. ниже).
Пример 4. Изучение субстратной специфичности мутантных оксидаз D-аминокислот и фермента дикого типа.
Для точной оценки эффекта изменения субстратной специфичности оксидаз D-аминокислот за счет введения точечных замен Cys108Ala, Cys108Phe, Cys108Ser мутантные ферменты и оксидазу D-аминокислот дикого типа выделяли в высокоочищенном состоянии и определяли константы Михаэлиса и максимальные скорости взаимодействия ферментов с различными субстратами.
Методика получения и очистки была одинакова как для мутантных TvDAAO, так и для фермента дикого типа. Клетки Е.coli BL21 (DE3) pLysS Codon Plus, содержащие одну из плазмид (pKanDAAO, или pKanDAAO_C108A, или pKanDAAO_C108F, или pKanDAAO_C108S) и синтезирующие целевой фермент, получали культивированием единичной колонии в колбах объемом 1 л, содержащих 150 мл среды 2YT и 30 мкг/мл канамицина, при 37°С при непрерывном перемешивании (140 об/мин). По достижении поглощения среды на 600 нм величины А600 0,6-0,9 в колбы добавляли изопропил-бета-D-тиогалактозид (ИПТГ) до концентрации 0,1 мМ и культивировали в течение 24 часов при 25°С. Клетки собирали центрифугированием (7000 об/мин, 20 мин, 4°С) и ресуспендировали в 20 мМ Трис-HCl, рН 7,5 для получения 20% суспензии. Для предварительного разрушения клетки подвергали замораживанию-размораживанию и далее обрабатывали ультразвуком при охлаждении льдом, каждые 5 минут отбирая пробы для измерения активности фермента. По достижении постоянного значения активности полученный раствор центрифугировали (18000 об/мин, 30 мин, 4°С) для удаления осадка. Полученный супернатант нагревали в течение 10 мин при 45°С для инактивации каталазы и некоторых других белков, осадок удаляли центрифугированием (18000 об/мин, 30 мин, 4°С).
Ионообменную FPLC хроматографию проводили на установке фирмы Pharmacia (Швеция). Раствор фермента наносили на колонку Source 15Q объемом 40 мл (Pharmacia Biotech, Швеция), уравновешенную 20 мМ Трис-HCl, рН 7,5 (буфер А). После нанесения образца колонку промывали тем же буфером до нулевого поглощения на 280 нм выходящего из колонки раствора. Фермент элюировали с колонки со скоростью 10 мл/мин линейным градиентом 0-1М NaCl в буфере А. Собирали фракции, обладающие оксидазной активностью, объединяли их и концентрировали на мембране РМ10 в ячейке для концентрирования объемом 50 мл (мембрана и ячейка производства Amicon, США) до объема 5 мл. Полученный фермент наносили на колонку 2,5×100 см с Sephacryl S-200 (Pharmacia Biotech, Германия), уравновешенную 0,1 М фосфатным буфером, рН 8,0. Собирали фракции по 5 мл, определяли их поглощение на длине волны 280 нм A280 на спектрофотометре Shimadzu 1601 PC (Shimadzu GmBH, Германия) и измеряли величину ферментативной активности с помощью пероксидазной реакции (методику см. ниже). Отбирали фракции, имеющие постоянное отношение (активность/А280). Чистота полученных препаратов составляла не менее 95% согласно данным аналитического электрофореза в 12% полиакриламидном геле в присутствии додецилсульфата натрия. Электрофорез проводили на приборе BioRad (Англия) согласно инструкции фирмы-производителя.
Активность ферментов определяли по образованию пероксида водорода, для чего использовали пероксидазу из корней хрена (ПХ) фирмы Reanal (Венгрия) и ее субстрат АБТС (2,2'-азино-бис(3-этилбензотиазолин-6-сульфонат)) фирмы Sigma (США). В качестве субстратов для мутантной DAAO и фермента дикого типа использовали различные D-аминокислоты. В кювету спектрофотометра (рабочий объем 1 мл, оптический путь 1 см) добавляли раствор D-аминокислоты в 50 мМ натрий- или калий-фосфатном буфере (ФБ), 20 мкл раствора АБТС (16 мг/мл) в воде, 10 мкл раствора пероксидазы в 50 мМ ФБ (1000 Ед/мл) и насыщенный кислородом 50 мМ ФБ (рН 8,0) до общего объема 980 мкл. После термостатирования в течение 10 мин при 30°С в реакционную смесь добавляли 20 мкл фермента. Накопление продукта окисления АБТС регистрировали на длине волны 414 нм (ε414=36000 М-1 см-1).
Для определения величин максимальной скорости реакции и константы Михаэлиса оксидазы D-аминокислот дикого типа и мутантных ферментов концентрацию соответствующей D-аминокислоты варьировали в диапазоне 0,5-5 КM. Приблизительное значение КM определяли в отдельном эксперименте, измеряя скорость реакции при концентрациях D-аминокислоты 0,1, 0,5, 1,0, 5,0, 10 мМ. Значения активности при каждой концентрации измеряли не менее 3 раз, а на границах и в середине диапазона определение проводили 5 раз. Величины Vm и КM определяли из зависимости скорости реакции от концентрации D-аминокислоты с помощью нелинейной регрессии, используя программу "Origin Pro 7.0". Программа также позволяет оценить ошибки расчета параметров.
В таблице 2 приведены относительные значения констант Михаэлиса [(Kм)мут/(Kм)дт]·100%, максимальных скоростей ((kcat)мут/(kcat)дт*100%) и общей каталитической эффективности [(kcat/KM)мут/(kcat/KM)дт]·100% с различными субстратами для и мутантных TvDAAOCys108Phe, TvDAAO Cys108Ala и TvDAAOCys108Ser относительно таковых для оксидазы D-аминокислот дикого типа. В случае константы Михаэлиса уменьшение КM соответствует относительным величинам менее 100%, а увеличение kcat и kcat/КM соответствует относительным величинам более 100%.
Из таблицы следует, что введение замен Cys108Ala, Cys108Phe, Cys108Ser существенно изменило профиль субстратной специфичности полученных мутантных форм оксидазы D-аминокислот по сравнению с профилем субстратной специфичности фермента дикого типа. Например, все три мутантные формы перестали катализировать окисление D-аланина, который являлся "хорошим субстратом" (высокое значение отношения kcat/КM) для фермента дикого типа, т.е. полученные мутанты являются идеальными для использования в биосенсорах для селективного определения в клетках нервной системы D-серина в присутствии высоких концентраций эндогенного D-аланина. Природные оксидазы D- аминокислот с таким соотношением активностей с D-серин - D-аланин в литературе не описаны. В случае D-лейцина, наоборот, параметр каталитической эффективности kcat/KM в случае мутантных ферментов выше в 2,2-7,5 раз, чем таковой у TvDAAO дикого типа. В плане улучшения каталитических свойства наиболее эффективной оказалась замена Cys108Ala, которая обеспечила увеличение значения kcat/КM для 8 из 12 изученных D-аминокислот.
Таким образом, полученные мутантные формы оксидазы D-аминокислот обладают субстратной специфичностью, отличной от субстратной специфичности фермента дикого типа, по отношению к ряду D-аминокислот и их производных.
Пример 5. Каталитическая активность мутантных форм оксидаз D-аминокислот и фермента дикого типа с цефалоспорином С.
Для определения эффективности взаимодействия TvDAAO дикого типа и мутантных форм с цефалоспорином С в пробирке объемом 2,0 мл (Eppendorf, Германия) смешивали 5 мМ цефалоспорина С и предварительно насыщенный кислородом 0,1 М ФБ, рН 8,0. Реакцию запускали добавлением одинакового по массе количества мутантных TvDAAO или фермента дикого типа (10 мкг). Реакцию проводили при 30°С и непрерывном перемешивании открытой пробирки на шейкере (300 об/мин). Через определенные промежутки времени из реакционной смеси отбирали пробы объемом 50 мкл и анализировали с использованием HPLC системы (BioRad, США) на колонке 4,0×125 мм с носителем Силасорб С 18, 4 мкм. Элюция производилась в 20 мМ СН3СООNН4, 2% СН3СN, рН 4,9 со скоростью 0,5 мл/мин. Уменьшение концентрации субстрата цефалоспорина С в ходе реакции регистрировали спектрофотометрически по поглощению раствора на выходе из колонки на длине волны 260 нм.
Кинетика окисления цефалоспорина С с помощью полученных мутантных TvDAAO и фермента дикого типа представлена на чертеже. Во всех случаях в реакцию вводили одинаковые по массе количества фермента. Из приведенного рисунка хорошо видно, что мутантные TvDAAO с заменами Cys108Ala и Cys108Phe проявляют гораздо более высокую каталитическую активность в реакции окисления цефалоспорина С по сравнению с ферментом дикого типа. Например, период времени, за который степень превращения цефалоспорина С составляла 98%, в случае мутантной TvDAAO с заменой Cys108Ala был примерно в 4 раза меньше, чем в случае TvDAAO дикого типа.
Изобретение относится к области генной и белковой инженерии и может быть использовано для детекции D-аминокислот в сложных образцах и в процессах ферментативного синтеза оптически активных веществ, альфа-кетокислот, цефалоспориновых антибиотиков и других органических соединений с использованием оксидаз. Варианты мутантной оксидазы D-аминокислот отличаются от оксидазы D-аминокислот из дрожжей Trigonopsis variabilis аминокислотным остатком в положении 108 аминокислотной последовательности и обладают измененной субстратной специфичностью по сравнению с ферментом дикого типа. Применение изобретения позволяет варьировать ряд D-аминокислот, которые могут служить субстратом для оксидаз D-аминокислот в реакциях ферментативного окисления указанных соединений. 3 н.п. ф-лы, 1 ил., 2 табл.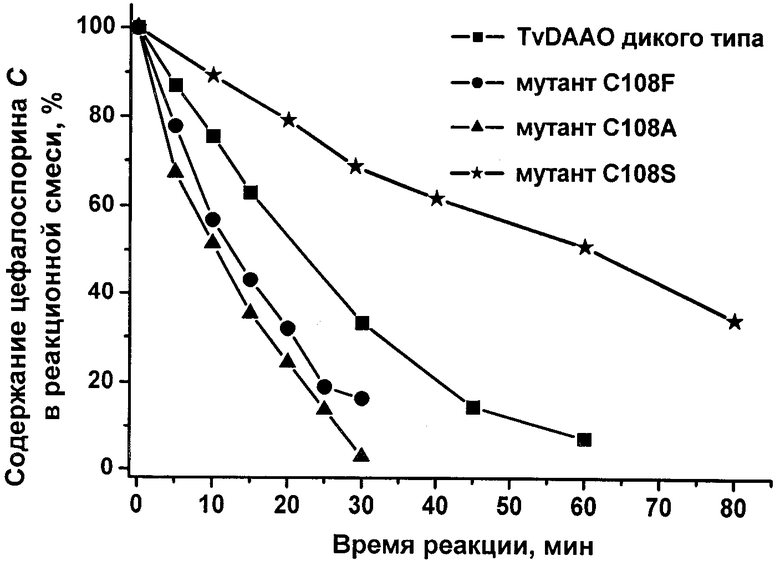
1. Мутантная оксидаза D-аминокислот с субстратной специфичностью, отличной от субстратной специфичности фермента дикого типа по отношению к ряду D-аминокислот и их производных, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности оксидазы D-аминокислот из Trigonopsis variabilis дикого типа, в которой остаток цистеина в положении 108 замещен остатком аланина.
2. Мутантная оксидаза D-аминокислот с субстратной специфичностью, отличной от субстратной специфичности фермента дикого типа по отношению к ряду D-аминокислот и их производных, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности оксидазы D-аминокислот из Trigonopsis variabilis дикого типа, в которой остаток цистеина в положении 108 замещен остатком фенилаланина.
3. Мутантная оксидаза D-аминокислот с субстратной специфичностью, отличной от субстратной специфичности фермента дикого типа по отношению к ряду D-аминокислот и их производных, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности оксидазы D-аминокислот из Trigonopsis variabilis дикого типа, в которой остаток цистеина в положении 108 замещен остатком серина.
| SLAVICA A., DIB I., NIDETZKY B., Single-site oxidation, cysteine 108 to cysteine sulfinic acid, in D-amino acid oxidase from Trigonopsis variabilis and its structural and functional consequences | |||
| Appl Environ Microbiol | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Устройство для измерения ускорений | 1990 |
|
SU1748073A1 |
| JU S.S, LIN L.L., CHIEN H.R., HSU W.H., Substitution of the critical methionine | |||

