Изобретение относится к области генной и белковой инженерии, а именно к новым белкам, мутантным Са2+-регулируемым фотопротеинам, которые проявляют отличительные свойства по сравнению с фотопротеином дикого типа, кДНК, кодирующие данные белки, и может быть использовано в исследованиях in vivo, в частности, для определения потоков кальция в клетках. Основное применение Са2+-регулируемых фотопротеинов обусловлено самой природой реакции - излучение света происходит в ответ на появление ионов кальция, поэтому наибольшее применение Са2+-регулируемые фотопротеины нашли в качестве биолюминесцентных индикаторов внутриклеточного кальция в различных типах клеток. Биолюминесценция инициируется ионами кальция и возникает вследствие окислительного декарбоксилирования связанного с белком субстрата - целентеразина. В результате образуются продукт реакции, целентерамид, в возбужденном состоянии и СО2. Переход целентерамида из возбужденного состояния в основное сопровождается излучением света. Обелин дикого типа генерирует «голубую» биолюминесценцию. Для мониторинга содержания внутриклеточного кальция сразу в разных частях клетки было бы полезно изменить длину волны испускания, так чтобы возможно было разделить сигналы, идущие от разных органелл. Фотопротеины диких типов позволяют измерять концентрацию Са2+ в диапазоне концентраций 10-7-10-4 М. Разное содержание свободного кальция в разных органеллах (в ЭПР, который является Са2+ депо, и в митохондриях максимальная концентрация свободного кальция очень высока и достигает 0.1-1 мМ, а в цитозоле, наоборот, низкая 10-8-10-7 М) диктует необходимость применения фотопротеинов с пониженной и повышенной чувствительностью к кальцию.
Известны мутанты акворина и обелина со сдвинутым спектром излучения [US 7345160, C12Q 1/66, publ. March 18, 2008], где сдвиг, кроме того, достигался использованием в качестве субстрата модифицированных аналогов целентеразина, что приводило к излучению в более длинноволновой части спектра.
Известен ряд мутантов акворина с пониженной [Kendall J.M. et al, Biochem. Biophys. Res. Commun., 1992] и повышенной чувствительностью к кальцию [Tricoire L. et al, PNAS, 2006, (прототип)].
Недостатком мутантов акворина и акворина дикого типа является влияние ионов магния на чувствительность к ионам кальция. Это существенно, поскольку ионы магния способны связываться в сайтах связывания кальция и изменять чувствительность к кальцию, а в клетках содержание магния достаточно велико (1-3 мМ).
Техническим результатом изобретения является получение на основе фотопротеина репортера внутриклеточного кальция, обладающего одновременно пониженной чувствительностью к кальцию и «синей биолюминесценцией», и репортера с повышенной чувствительностью к кальцию и «зеленой» биолюминесценцией.
Технический результат достигается тем, что в мутантном фотопротеине для определения внутриклеточного кальция одновременно в разных органеллах клетки, обладающем светоизлучающей функцией и характеризующемся аминокислотной последовательностью кальций-регулируемого фотопротеина дикого типа, в которой природные остатки фенилаланина, относящиеся к консервативным областям названной последовательности и соответствующие остатку 119 фотопротеина обелина Obelia longissima (или остатку 120 фотопротеина акворина Aequorea victoria, или остатку 122 фотопротеина клитина Clytia gregaria), а также остатку 88 обелина Obelia longissima (или остатку 89 фотопротеина акворина Aequorea victoria, или остатку 91 фотопротеина клитина Clytia gregaria), заменены другими остатками, отличными от фенилаланина, замены которых приводят к тому, что чувствительность кальций-регулируемого фотопротеина к ионам кальция уменьшается по сравнению с фотопротеином дикого типа и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа. Аминокислотный остаток, соответствующий остатку 119 в обелине Obelia longissima, является триптофаном, а аминокислотный остаток, соответствующий остатку 88 обелина Obelia longissima, представляет собой тирозин.
Технический результат достигается также тем, что мутантном фотопротеине для определения внутриклеточного кальция одновременно в разных органеллах клетки, обладающем светоизлучающей функцией и характеризующемся аминокислотной последовательностью кальций-регулируемого фотопротеина дикого типа, в которой природный остаток изолейцина, являющийся консервативной аминокислотой и соответствующий остатку 144 фотопротеина обелина Obelia longissima (или остатку 145 фотопротеина акворина Aequorea victoria, или остатку 147 фотопротеина клитина Clytia gregaria), заменен другим аминокислотным остатком, отличным от изолейцина, таким образом, что увеличивается чувствительность кальций-регулируемого фотопротеина к ионам кальция и изменяется длина волны излучения по сравнению с фотопротеином дикого типа. В мутантном фотопротеине гидрофобный аминокислотный остаток в положении 144 фотопротеина обелина Obelia longissima заменен на гидрофильный аминокислотный остаток. Гидрофильная аминокислота, обеспечивающая увеличение чувствительности кальций-регулируемого фотопротеина к ионам кальция и изменение длины волны испускаемого света, представляет собой гистидин.
Технический результат также обеспечивается ДНК, в которой кодоны, соответствующие аминокислотным остаткам в положениях 119 и 88, заменены кодонами, определяющими аминокислоты, отличные от фенилаланина, таким образом, что чувствительность кальций-регулируемого фотопротеина к ионам кальция уменьшается и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа. ДНК имеет нуклеотидную последовательность, описанную в Seq. №1, где нуклеотиды в положениях 262-264 образуют кодон, кодирующий аминокислоту, обеспечивающую смещение максимума излучения в более коротковолновую область спектра, и нуклеотиды в положениях 355-357 образуют кодон, кодирующий аминокислоту, обеспечивающую уменьшение чувствительности фотопротеина к ионам кальция. ДНК, в которой нуклеотиды в положениях 262-264 кодируют тирозин и нуклеотиды в положениях 355-357 кодируют триптофан.
Технический результат также обеспечивается ДНК, в которой кодон, соответствующий аминокислотному остатку в положении 144, заменен кодоном, определяющим аминокислоту, отличную от изолейцина, таким образом, что чувствительность кальций-регулируемого фотопротеина к ионам кальция увеличивается и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа. ДНК имеет нуклеотидную последовательность, описанную в Seq. №3, где нуклеотиды в положениях 430-432 образуют кодон, кодирующий аминокислоту, обеспечивающую смещение максимума излучения в более длинноволновую область спектра и увеличение чувствительности фотопротеина к ионам кальция. ДНК, в которой нуклеотиды в положениях 430-432 кодируют гистидин.
Вектор для экспрессии белка, в клетках млекопитающих, представляет собой плазмиду pCMV/myc/mito (Invitrogene), в которую лигирована ДНК, характеризующаяся последовательностью Seq. №1, продукт экспрессии которой будет локализован в митохондриях и излучать «синий» свет.
Вектор для экспрессии белка в клетках млекопитающих, представляющий собой плазмиду pcDNA3.1, в которую лигирована ДНК, характеризующаяся Seq. №3, продукт экспрессии которой будет локализован в цитоплазме клеток и при появлении ионов кальция (при стимуле) излучать «зеленый» свет. Плазмиды, содержащие гены мутантных фотопротеинов в таких формах, которые способны экспрессировать белки изобретения конститутивно, при введении в клетку-хозяина.
Согласно изобретению предусматриваются клетки млекопитающих линии СНО, способные экспрессировать мутантные фотопротеины изобретения, а также способ анализа, в котором внутриклеточное содержание кальция измеряется одновременно в двух частях клетки.
В заявляемом изобретении посредством сайт-направленного мутагенеза получены белки, одна молекула которых содержит одновременно замены, приводящие к изменению двух свойств фотопротеина: чувствительности к ионам кальция и спектра биолюминесценции. При этом не требуется дополнительного использования аналогов субстрата. И, в отличие от прототипа, кроме измененной чувствительности к кальцию, мутантные фотопротеины настоящего изобретения обладают измененными спектральными характеристиками излучения так, что максимумы биолюминесценции двух мутантных фотопротеинов отстоят друг от друга на спектральной шкале длин волн на 50 нм. Также в отличие от прототипа фотопротеины по изобретению имеют кальциевую чувствительность, которая не зависит от присутствия ионов магния.
Приобретаемые мутантными фотопротеинами новые свойства позволяют эффективно использовать их в качестве репортерных агентов при изучении потоков кальция в клетках, поскольку имея различную чувствительность к Са2+ позволяют измерять концентрацию ионов в компартментах клетки с разным содержанием кальция, а разные максимумы излучения позволяют проводить измерения одновременно.
Перечисленные выше отличительные от прототипа признаки позволяют сделать вывод о соответствии заявляемых технических решений критерию «новизна».
Признаки, отличающие заявляемые технические решения от прототипа, не выявлены в других технических решениях и, следовательно, обеспечивают заявляемому решению соответствие критерию «изобретательский уровень».
Описание чертежей
На фигуре 1 представлены последовательности олигонуклеотидов, использованные в качестве праймеров для осуществления мутаций сайт-направленным мутагенезом.
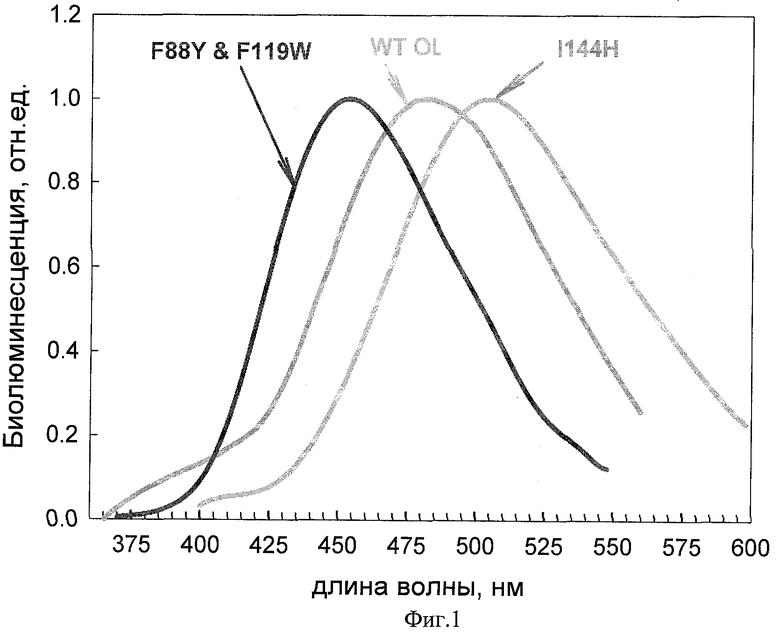
На фигуре 2 представлен график, иллюстрирующий спектры биолюминесценции обелина дикого типа WT и мутантов обелина с двойной заменой: фенилаланин в 88-м положении на тирозин и в 119-м положении на триптофан (F88Y&F119W) и с заменой изолейцина в положении 144 аминокислотной последовательности обелина на гистидин (I144H).
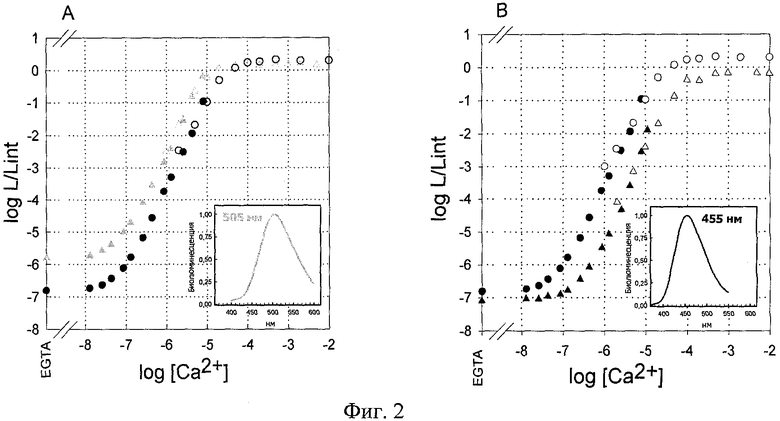
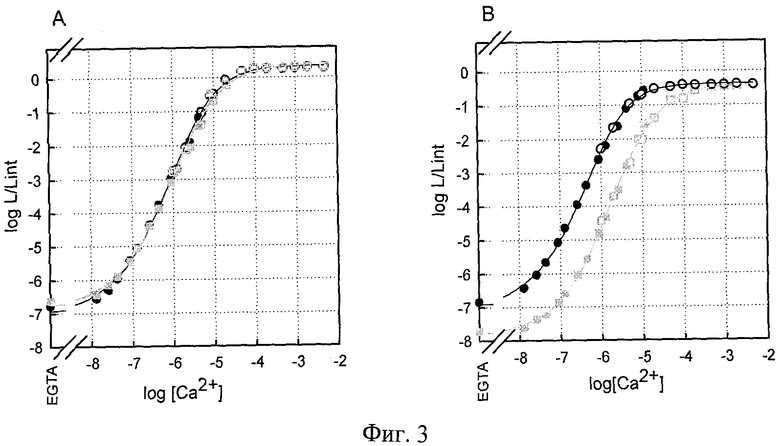
На фигуре 3 показаны графики зависимости интенсивности люминесценции от концентрации Са2+ (А) мутанта обелина OL I144H, (В) мутанта обелина OL F88Y&F119W. Обозначения: Δ - люминесценция мутантов, ○ - люминесценция обелина WT. Заполненные символы - концентрация кальция задавалась с помощью Са2+ - EGTA буферов. Открытые символы - концентрация кальция задавалась с помощью разведения раствора CaCl2. Интенсивность светового сигнала (L) выражена в единицах L/Lint, где Lint - суммарное количество квантов, излученное в ходе реакции, определенное с помощью кинетических измерений в тех же условиях в том же образце белка. Температура измерения 20°С. В правом нижнем углу графиков приведены спектры биолюминесценции соответствующих мутантных белков, интенсивность люминесценции выражена в относительных единицах, полученных при нормировании спектров на Lmax. Рядом с кривой цифрами обозначена длина волны, при которой наблюдается максимум биолюминесценции (λmax).
На фигуре 4 представлены графики зависимости интенсивности люминесценции мутантного обелина OL F88Y (А) и акворина Aeq N33D (В) от концентрации Са2+. Обозначения: ○ - люминесценция мутантов в отсутствие Mg2+; □ - люминесценция мутантов в присутствии 1 мМ Mg2+. Остальные обозначения аналогичны фиг.3.
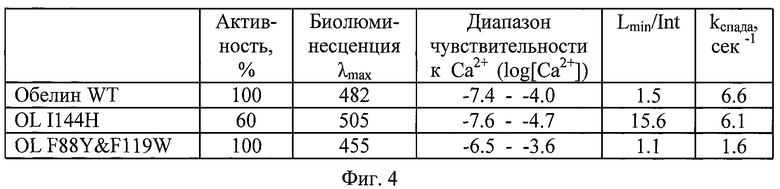
На фигуре 5 представлена таблица, в которой указаны основные характеристики биолюминесценции обелина WT и его мутантных вариантов: OL I144H и OL F88Y&F119W. Приведены характеристики биолюминесценции в растворе при рН 7.0. Биолюминесцентная активность выражена в процентных долях по отношению к удельной специфической активности обелина дикого типа (рекомбинантного). Параметр Lmin/Int обозначает интенсивность Са2+-независимой люминесценции, нормированную на суммарный световой выход. Са2+-независимая люминесценция (самопроизвольное излучение) является показателем стабильности фотопротеинов. kспада - характеризует скорость спада биолюминесцентного сигнала.
Последовательности, представленные в конце описания, описывают последовательности ДНК и аминокислот следующим образом:
Последовательность №1: представляет ДНК последовательность, кодирующую мутантный фотопротеин настоящего изобретения, в которой кодоны дикого типа Obelia longissima в положениях 262-264 и 355-357 подверглись мутациям, для тирозина основание 263 в результате мутации представлено А, для триптофана основания 356 и 357 в результате мутации представлены Т.
Последовательность №2: представляет аминокислотную последовательность фотопротеина настоящего изобретения, в которой аминокислота фенилаланин в положениях 88 и 119 Obelia longissima заменены на тирозин и триптофан соответственно.
Последовательность №3: представляет ДНК последовательность, кодирующую другой мутантный фотопротеин настоящего изобретения, в которой мутирован кодон дикого типа Obelia longissima в положении 430-432 таким образом, что основания в положении 430 и 431 представлены С и А.
Последовательность №4: представляет аминокислотную последовательность другого фотопротеина настоящего изобретения, в которой аминокислота изолейпин заменена на гистидин.
Пример 1
Получение мутантных фотопротеинов с измененной чувствительностью к кальцию и спектрами излучения.
Все генно-инженерные манипуляции, если не оговорено особо, были выполнены с помощью методов, хорошо известных для специалистов в данной области.
Сайт-направленный мутагенез для получения мутантных форм обелина проводился методом ПЦР с использованием набора для направленного на сайт мутагенеза QuikChange® Site-Directed Mutagenesis Kit (Stratagene, США), следуя руководству к набору. В качестве матрицы использовали плазмиду, содержащую ген обелина дикого типа pET19-OL8. Праймеры для введения единичных аминокислотных замен были синтезированы в ООО «Синтол» (Россия). Последовательности олигонуклеотидных праймеров и их комплементарных партнеров представлены на фиг.1. Все полученные мутации были подтверждены секвенированием полученной ДНК. Нуклеотидную последовательность ДНК определяли на системе для секвенирования Amerlight (Amersham, Великобритания) с использованием Themo Sequenase™ Cy™5 Dye Terminator Kit (Amersham, Великобритания). Для каждого комплекта из 4-х реакций (один образец) готовили реакционную смесь комбинацией следующих компонентов в одной пробирке: 1 мкг ДНК-матрицы (0.5 мкг/мкл), 4 пмоль сиквенсового праймера (20 мкМ), реакционного буфера, термосеквеназы (10 U/µ1), 2 мкл A, G, С или Т-смеси. Программа амплификации включала 31 цикл со следующими параметрами: 95°С 30 сек, следующие 30 циклов (95°С 30 сек, 56°С 30 сек, 72°С 1 мин 20 сек). Полученные образцы переосаждали этанолом с ацетатом аммония и глицерином, осадок получали центрифугированием в течение 15 мин при 16000 g, высушивали на воздухе и растворяли в 8 мкл стоп-раствора каждый. Затем ДНК денатурировали нагреванием в течение 3 минут при 72°С и помещали в лед до нанесения на гель. В качестве электрофорезного буфера использовали ТВЕ×0.5. На гель наносили по 1/2 объема реакции. Разделение осуществляли согласно стандартной программы ALFWin Control (Amerlight, Великобритания) при следующих условиях: 1500 В, 25 мА, 750 мин. Данные обрабатывали с помощью программы ALF Express.
Белки экспрессировали в клетках Е.соli и получали в высокоочищенном виде по методу, разработанному для обелина дикого типа (WT). Клеточную пасту суспендировали в пятикратном объеме 0,02 М Трис-HCl, рН 7.0 и разрушали озвучиванием при помощи ультразвукового дезинтегратора UD-20 (Techpan, Польша) 5-кратно по 20 сек при охлаждении льдом. Полученную смесь центрифугировали (14000 g, 2.9 мин). Осадок, состоящий в основном из телец включения, промывали, ресуспендируя последовательно растворами 0,02 М Трис-HCl, рН 7.0, содержащими 0,9% NaCl, 0,1% Тритон Х-100, 5 мМ CaCl2. Тельца включения растворяли в 2,5-кратном объеме 6 М мочевины, содержащей 20 мМ Трис-HCl и 5 мМ CaCl2 (40°С, в течение ночи), затем снова центрифугировали. Супернатант хроматографировали на колонке, заполненной DEAE-Sepharose FF (2×3,5 см) (хроматографическая система FPLC, Pharmacia, Швеция). Элюцию веществ с колонки проводили буфером, содержащим 6 М мочевину, 5 мМ CaCl2 0,02 М Трис-HCl рН 7.0 в градиенте концентраций NaOAc от 0 до 0,5 М. Скорость элюции составляла 2 мл/мин.
Чистоту конечных препаратов рекомбинантных апобелков, полученных после выделения, оценивали с помощью электрофореза в 12,5%-ном ПААГ в денатурирующих условиях (в присутствии 0.1%-ного SDS) по методу Лэммли.
Активные фотопротеины получали инкубацией апобелков с 10-кратным молярным избытком субстрата - целентеразина в буфере, содержащем 20 мМ Tris-HCl, рН 7.0, 5 мМ ЭДТА, 5 мМ дитиотреитола при 4°С в течение как минимум 4-х часов.
Активированные белки отделяли от алобелков ионообменной хроматографией на колонке Mono Q 5/5 с помощью хроматографической системы FPLC, Pharmacia, Швеция. Элюцию белков с колонки проводили в 20 мМ Tris-HCl, pH 7, 5 мМ ЭДТА градиентом NaCl (0-0,35 М).
Пример 2
Определение чувствительности фотопротеинов к ионам кальция
Определение чувствительности фотопротеинов к Ca2+ основано на том, что низкие концентрации Ca2+ задаются с помощью кальциевых буферных растворов (Ca2+-EGTA буферов), а высокие - простым разбавлением раствора CaCl2. Кальциевые буферные растворы готовили из двух стоковых растворов путем реципрокного разбавления: один из них содержал эквимолярное количество Ca2+ и хелатора - EGTA, другой точно такое же количество хелатора, но без Ca2+. Для определения концентрации кальция в готовых растворах использовали равновесную константу диссоциации хелатора, которая зависит от pH, ионной силы и температуры раствора. Все используемые растворы и посуда требуют тщательной обработки от следов ионов Ca2+ и EGTA. С этой целью все используемые растворы пропускали через колонку с сорбентом Chelex-100, связывающим следовые количества ионов кальция (Sigma). Для приготовления растворов использовали деионизованную воду (18.2 МОм). Во время измерений температура как растворов, так и измерительного кюветного блока поддерживалась на постоянном уровне (20°С).
Белковые растворы также очищались от следов ионов Ca2+ и EGTA посредством гель-фильтрации на колонке: 1.5×6.5 см D-Salt Dextran Desalting column (Pierce) в буфере, включающем 150 мМ KCl, 5 мМ PIPES, pH 7.0. Фракции, содержащие белок, идентифицировали по биолюминесцентной активности, при этом во избежание контаминации EGTA для определения зависимости биолюминесцентной активности от концентрации Ca2+ собирали только самые первые выходящие с колонки белковые фракции. Измерение светового сигнала проводили впрыскиванием 10 мкл таким образом подготовленного раствора фотопротеина в 1 мл Ca2+-раствора. Интенсивность светового сигнала (L) была выражена в единицах L/Lint, где Lint - суммарное количество квантов, излученное в ходе реакции, определенное с помощью кинетических измерений в тех же условиях в том же образце белка. Люминометр имел три фильтра разной оптической плотности, ослабляющих излучение, что позволило записать сигналы, отличающиеся по величине на 8 порядков.
Для определения чувствительности фотопротеинов к Са2+ в присутствии 1 мМ Mg2+, обессоленный белковый образец выдерживали в течение 1 часа с 1 мМ MgCl2. Все Са2+-растворы также содержали 1 мМ Mg2+.
В результате изучения зависимости интенсивности люминесценции от концентрации Са2+ были определены следующие показатели:
а) диапазон чувствительности к Са2+;
б) Lmin/Lmax - отношение нижней и верхней границ диапазона люминесцентного ответа, характеризующее ширину диапазона люминесцентного ответа;
в) Lmin/Int - интенсивность Са2+-независимой люминесценции, нормированная на суммарный световой выход.
Зависимость интенсивности люминесценции мутантных фотопротеинов от концентрации Са2+ представлена как функция логарифма нормированной люминесценции (log(L/Lint)) от логарифма концентрации Са2+ (log[Ca2+]). Диапазон чувствительности к Са2+ у мутантного фотопротеина I144H расположен в интервале -7.6 - -4.7 и сдвинут в сторону более низких концентраций Са2+ по сравнению с фотопротеином дикого типа, что означает увеличение чувствительности мутантов к Са2+. Напротив, замены F88Y&F119W приводят к смещению диапазона чувствительности к Са2+ в сторону более высоких концентраций иона, и диапазон составляет -6.5 - -3.6 (на порядок меньше по сравнению с обелином дикого типа). Данные мутантные фотопротеины выбраны как наиболее перспективные из ряда запланированных и исследованных точечных замен в последовательности обелина, включающих: С51А, F72V, V118F, E41G, D133E&I132V, F119W, L160D, F88W, F88Y, F88H, F88R, W92F, W92H, W92E, W92K, W92R, W92F&H22E, Н22Е, H22N, M25F, M25Y, М25Н, М25К, M25R, I144W, I144H, I144Q, Y138F, Y138W. Для мутантного фотопротеина F88Y&F119W величина Lmin/Lmax составляет почти 10-7, для мутанта I144H - 10-6. Сужение ширины диапазона люминесцентного ответа в случае последнего объясняется возрастанием интенсивности люминесценции в отсутствие Са2+ (возрастанием нижней границы диапазона) Lmin/Int.
Ионы магния (Mg2+) обладают меньшей аффинностью к кальций-связывающим сайтам фотопротеинов, чем ионы кальция (Са2+). Но их присутствие может отразиться на чувствительности к Са2+. Присутствие Mg2+ в физиологической концентрации (1 мМ) никак не влияет на чувствительность мутантных фотопротеинов настоящего изобретения к Са2+. Это выражается в том, что Са2+-кривые в отсутствие ионов магния и в присутствии 1 мМ Mg2+ идентичны. Отсутствие влияния ионов магния на чувствительность к ионам кальция свойственно и для исходного белка - обелина дикого типа (WT). По этому признаку обелин отличается от другого фотопротеина - акворина, несмотря на то, что оба белка близкородственны и структурные отличия между ними минимальны. Мы сконструировали 2 мутанта акворина: первый с заменой 33-го аспарагина на аспарагиновую кислоту в 1-ом Са2+-связывающем сайте акворина (N33D), второй - с заменой фенилаланина в 156-м положении на серин, находящегося в гидрофобной области белковой глобулы (F156S). Определили чувствительность полученных мутантов акворина к кальцию. Оказалось, что оба фотопротеина обладают повышенной чувствительностью к Са2+ по сравнению с акворином дикого типа, что характеризуется смещением диапазона чувствительности к Са2+ в сторону более низких концентраций Са2+. Добавление 1 мМ Mg2+ во все используемые в эксперименте растворы радикально сдвигает Са2+-кривые в сторону более высоких концентраций иона. На фигуре 4 представлены примеры влияния присутствия ионов магния на чувствительность к кальцию обелинового мутанта и акворинового мутанта.
Пример 3
Основные биолюминесцентные свойства фотопротеинов
Относительную удельную биолюминесцентную активность (относительно обелина дикого типа) мутантов определяли с помощью планшетного люминометра Luminoskan vl.30 (Labsystems, Финляндия). Для этого в лунки непрозрачного планшета вносили по 50 мкл раствора фотопротеина в 20 мМ Tris-HCl-буфере, рН 7.0, содержащем 5 мМ EDTA и 0,2 М NaCl. Реакцию запускали впрыскиванием 50 мкл 100 мМ Tris-HCl-буфера, рН 8.8, содержащего 100 мМ CaCl2, в каждую лунку планшета. Время сканирования сигнала - от 5 сек до 1 мин в зависимости от скорости люминесцентной реакции каждого из белков.
Биолюминесцентная активность мутантного фотопротеина F88Y&F119W идентична активности фотопротеина дикого типа (WT), активность мутантного фотопротеина I144H ниже на 40% (фиг.5).
Кальций-независимая люминесценция была измерена в белковых образцах (0,5-2,0 мг/мл), содержащих 2 мМ EDTA, помещенных в кювету люминометра.
Уровень кальций-независимой люминесценции, характеризующий стабильность фотопротеина, как комплекса белок-субстрат, у «двойного» мутанта такие же, как и у обелина дикого типа, а у «одинарного» в 10 раз выше, но при использовании этого белка для исследований in vivo, это не существенно, поскольку комплекс белок-субстрат образуется при проникновении гидрофобного субстрата в клетку и не требует его долгосрочного хранения.
Спектры биолюминесценции были измерены на спектрофлуориметре AMINCO (Thermo Spectronic, США). Все спектры излучения были корректированы на чувствительность ФЭУ к различным длинам волн с помощью программного обеспечения прибора. Спектры биолюминесценции измеряли в белковых растворах, содержащих 1 мМ EDTA, 50 мМ Бис-Трис пропан, рН 7.0, при впрыскивании раствора CaCl2 в том же буфере. Концентрация свободного кальция в реакционной смеси составляла около 0.5 µM (концентрация была рассчитана с помощью компьютерной программы Maxichelator) для того, чтобы обеспечить постоянный уровень свечения во время записи спектра излучения. Измерения были выполнены при комнатной температуре.
Обелин дикого типа генерирует голубую биолюминесценцию с максимумом 482 нм и плечом в районе 400 нм. Максимум биолюминесценции мутантного фотопротеина F88Y&F119W расположен в «синей» части спектра и равен 455 нм. Максимум биолюминесценции мутантного фотопротеина I144H расположен в «зеленой» части спектра и равен 505 нм. Спектры обоих фотопротеинов изобретения мономодальны и не содержат других пиков.
Константа спада биолюминесцентного сигнала была рассчитана по кривой спада, которая была записана при впрыске насыщающей концентрации Ca2+ в образец, находящийся в кювете люминометра, по формуле kспада=ln(I1/I2)/Δt (сек).
Кинетика биолюминесцентной реакции мутантов характеризуется более медленным спадом интенсивности по сравнению с обелином дикого типа. Константа спада биолюминесцентного сигнала для F88Y&F119W составляет 1.6 сек-1, что в 4 раза меньше, чем у обелина WT, для I144H константа составляет 6.1, что также меньше значения константы обелина дикого типа. Медленное свечение - еще одно преимущество фотопротеина как внутриклеточного репортерного белка, поскольку упрощает регистрацию светового потока.
Пример 4
Получение плазмид, содержащих ДНК настоящего изобретения
Плазмиды pCMV/myc/mito и pcDNA3.1(+) получены от hwtrogene Corporation, USA. Олигонуклеотидные праймеры были синтезированы ЗАО «Биосан», Россия.
Праймер, содержащий сайт рестрикции для нуклеазы KpnI для конструирования вектора, содержащего ген мутантного фотопротеина OL I144H, на основе pcDNA3.1(+) с помощью ПЦР характеризуется последовательностью:
olig №1 - 5'-TAACTTTAAGAAGGAGAGGTACCATGGCT-3'.
Праймер, содержащий сайт рестрикции для нуклеазы PstI, для конструирования вектора, содержащего ген мутантного фотопротеина OL F88Y&F119W, на основе pCMV/myc/mito с помощью ПЦР, имеет следующую последовательность:
olig №2 - 5'-CTTTAAGAAGGAGATATCTGCAGGCTTCAAAATACGCA-3'.
Праймер для синтеза сайта рестрикции для нуклеазы XhoI, комплементарный концевой части последовательности обелина представлен:
olig №3 - 5'-TACTCGAGATTAGGGAACTCCGTTGC-3'.
Для получения pCMV-OL88/119 вставку, содержащую кодирующую последовательность мутантного фотопротеина OL F88Y&F119W (Seq. 1) с концевьми сайтами рестрикции PstI и XhoI, получали на основе pET19b-OL88/119 путем ПЦР с праймерами olig №2 и olig №3. Рестрикцию фрагмента и вектора проводили с помощью PstI и XhoI (СибЭнзим, Россия), после чего препаративно очищали с помощью электрофореза в агарозном геле и лигировали.
Для получения pcDNA3.1(+)-OL144 вставку, содержащую кодирующую последовательность мутантного фотопротеина OL I144H (Seq. 3) с концевыми сайтами рестрикции KpnI и XhoI, получали на основе pET19b-OL144 путем ПЦР с праймерами olig №1 и olig №3. Рестрикцию фрагмента и вектора проводили с помощью KpnI и XhoI (СибЭнзим, Россия), после чего препаративно очищали с помощью электрофореза в агарозном геле и лигировали.
Полученные конструкции были проверены секвенированием ДНК. Трансформацию плазмид в клетки СНО (клетки яичников китайского хомяка) проводили с помощью трансфицирующего агента FreeStyle MAX (Invitrogene). Селективный отбор клонов, содержащих плазмиду, проводили на среде с неомицином.
Получены новые мутантные формы фотопротеина обелина, характеризующиеся измененной чувствительностью к кальцию и одновременно измененной длиной волны испускаемого света. Новые фотопротеины отличаются от исходного варианта точечными заменами в аминокислотной последовательности. Получены два репортера, которые можно использовать в качестве внутриклеточных индикаторов кальция. Синий и зеленый цвет биолюминесценции, а также высокая и низкая чувствительность к кальцию позволяет изучать изменение концентрации кальция одновременно (с помощью двухволновой детекции) в двух клеточных компартментах с разным содержанием Са2+, например, в цитозоле, где [Са2+] в состоянии покоя составляет 0.05-0.1 мкМ, повышаясь до 10 мкМ при различных стимулах, и митохондриях, где [Са2+] достигает сотен мкМ. Данный способ определения содержания внутриклеточного кальция применим для изучения процессов в клетке, опосредованных действием ионов кальция, для определения функции рецепторов, лигандов, ионных каналов, для изучения воздействий на клетки различных веществ: лекарств, токсинов и др.
Seq 1.

Seq 2.

Seq 3.
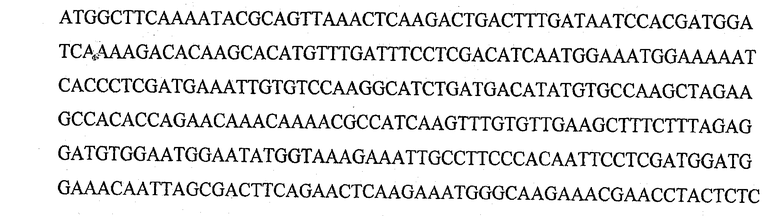
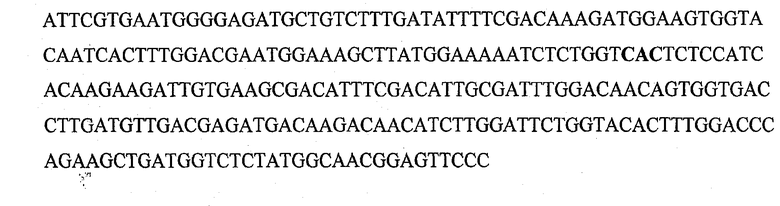
Seq 4.


Изобретение относится к биотехнологии и представляет собой мутантный фотопротеин для определения внутриклеточного кальция одновременно в разных органеллах клетки, обладающий светоизлучающей функцией и характеризующийся аминокислотной последовательностью кальций-регулируемого фотопротеина дикого типа, в которой природные остатки фенилаланина, относящиеся к консервативным областям названной последовательности и соответствующие остатку 119 фотопротеина обелина Obelia longissima, а также остатку 88 обелина Obelia longissima заменены другими остатками, отличными от фенилаланина, замены которых приводят к тому, что чувствительность кальций-регулируемого фотопротеина к ионам кальция уменьшается по сравнению с фотопротеином дикого типа и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа. Изобретение относится также к еще одному мутантному протеину, имеющему замену в положении 144 фотопротеина обелина Obelia longissima. Изобретение относится также к ДНК, кодирующим указанные выше белки, а также к векторам для экспрессии белков. Изобретение позволяет получить новые формы фотопротеина с измененной чувствительностью к кальцию и измененной длиной волны испускаемого света. 6 н. и 7 з.п. ф-лы, 5 ил.
1. Мутантный фотопротеин для определения внутриклеточного кальция одновременно в разных органеллах клетки, обладающий светоизлучающей функцией, и характеризующийся аминокислотной последовательностью кальций-регулируемого фотопротеина дикого типа, в которой природные остатки фенилаланина, относящиеся к консервативным областям названной последовательности и соответствующие остатку 119 фотопротеина обелина Obelia longissima (или остатку 120 фотопротеина акворина Aequorea victoria, или остатку 122 фотопротеина клитина Clytia gregaria), а также остатку 88 обелина Obelia longissima (или остатку 89 фотопротеина акворина Aequorea victoria, или остатку 91 фотопротеина клитина Clytia gregaria) заменены другими остатками, отличными от фенилаланина, замены которых приводят к тому, что чувствительность кальций-регулируемого фотопротеина к ионам кальция уменьшается по сравнению с фотопротеином дикого типа и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа.
2. Мутантный фотопротеин по п.1, характеризующийся аминокислотной последовательностью, описанной в Seq. №2, отличающийся тем, что аминокислотный остаток, соответствующий остатку 119 в обелине Obelia longissima, является триптофаном, а аминокислотный остаток, соответствующий остатку 88 обелина Obelia longissima, представляет собой тирозин.
3. Мутантный фотопротеин для определения внутриклеточного кальция одновременно в разных органеллах клетки, обладающий светоизлучающей функцией и характеризующийся аминокислотной последовательностью кальций-регулируемого фотопротеина дикого типа, в которой природный остаток изолейцина, являющийся консервативной аминокислотой и соответствующий остатку 144 фотопротеина обелина Obelia longissima (или остатку 145 фотопротеина акворина Aequorea victoria, или остатку 147 фотопротеина клитина Clytia gregaria) заменен другим аминокислотным остатком, отличным от изолейцина, таким образом, что увеличивается чувствительность кальций-регулируемого фотопротеина к ионам кальция и изменяется длина волны излучения по сравнению с фотопротеином дикого типа.
4. Мутантный фотопротеин по п.3, отличающийся тем, что гидрофобный аминокислотный остаток в положении 144 фотопротеина обелина Obelia longissima заменен на гидрофильный аминокислотный остаток.
5. Мутантный фотопротеин по п.4, характеризующийся аминокислотной последовательностью, описанной в Seq. №4, где гидрофильная аминокислота, обеспечивающая увеличение чувствительности кальций-регулируемого фотопротеина к ионам кальция и изменение длины волны испускаемого света, представляет собой гистидин.
6. ДНК, кодирующая мутантный фотопротеин по п.2, характеризующаяся тем, что ее полинуклеотидная последовательность представляет собой последовательность фотопротеина обелина дикого типа, в которой кодоны, соответствующие аминокислотным остаткам в положениях 119 и 88, заменены кодонами, определяющими аминокислоты, отличные от фенилаланина, таким образом, что чувствительность кальций-регулируемого фотопротеина к ионам кальция уменьшается и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа.
7. ДНК по п.6, отличающаяся тем, что имеет нуклеотидную последовательность, описанную в Seq. №1, где нуклеотиды в положениях 262-264 образуют кодон, кодирующий аминокислоту, обеспечивающую смещение максимума излучения в более коротковолновую область спектра, и нуклеотиды в положениях 355-357 образуют кодон, кодирующий аминокислоту, обеспечивающую уменьшение чувствительности фотопротеина к ионам кальция.
8. ДНК по п.7, где нуклеотиды в положениях 262-264 кодируют тирозин и нуклеотиды в положениях 355-357 кодируют триптофан.
9. ДНК, кодирующая мутантный фотопротеин по п.5, характеризующаяся тем, что ее полинуклеотидная последовательность представляет собой последовательность фотопротеина обелина дикого типа, в которой кодон, соответствующий аминокислотному остатку в положении 144, заменен кодоном, определяющим аминокислоту, отличную от изолейцина, таким образом, что чувствительность кальций-регулируемого фотопротеина к ионам кальция увеличивается и изменяется длина волны испускаемого света по сравнению с фотопротеином дикого типа.
10. ДНК по п.9, отличающаяся тем, что имеет нуклеотидную последовательность, описанную в Seq. №3, где нуклеотиды в положениях 430-432 образуют кодон, кодирующий аминокислоту, обеспечивающую смещение максимума излучения в более длинноволновую область спектра и увеличение чувствительности фотопротеина к ионам кальция.
11. ДНК по п.10, где нуклеотиды в положениях 430-432 кодируют гистидин.
12. Вектор для экспрессии белка, охарактеризованного в любом из пп.1 и 2, в клетках млекопитающих, представляющий собой плазмиду pCMV/myc/mito, в которую лигирована ДНК, охарактеризованная в любом из пп.6-8.
13. Вектор для экспрессии белка, охарактеризованного в любом из пп.3-5, в клетках млекопитающих, представляющий собой плазмиду pcDNA3.1, в которую лигирована ДНК, охарактеризованная в любом из пп.9-11.
| TRICOIRE L et al | |||
| Calcium dependence of aequorin bioluminescence dissected by random mutagenesis | |||
| Proc Natl Acad Sci USA | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| KENDALL J.M | |||
| et al | |||
| Targeting aequorin to the endoplasmic reticulum of living cells | |||
| Biochem Biophys Res Commun | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| MARKOVA S.V | |||
| et al | |||
| Obelin from the bioluminescent | |||

