Изобретение относится к биохимии, конкретно к новым биологически активным пептидам общей формулы Arg1-Trp2-Cys3-Val4-Tyr5-Ala6-Tyr7-Val8-Arg9-X10-Arg11-Gly12-Val13-Leu14-Val15-Arg16-Tyr17-Arg18-Arg19-Cys20-Trp21, где Х - Val (ареницин-1) или Ile (ареницин-2), обладающим антимикробным действием, которые могут найти применение в медицине.
В настоящее время клиническая медицина столкнулась с проблемой устойчивости патогенов к антибиотикам из имеющегося арсенала лекарственных средств, которые в ряде случаев уже не обеспечивают эффективного контроля над возбудителем при развитии хронических и рецидивирующих инфекционных заболеваний [Daw M.A., Drah A.M. 2001. Antibiotic resistance: prospects for the new millenium. J. Chemoter. 13: 587-594]. Ряд штаммов Mycobacterium tuberculosis, Enterococcus faecium и Pseudomonas aeroginosa все труднее подавляются конвенциальными антибиотиками. Возрастающая резистентность наблюдается также в отношении таких распространенных патогенов человека, как Staphylococcus aureus и Staphylococcus pneumoniae. Стремительный рост бактериальных инфекций, резистентных к так называемым "классическим" антимикробным средствам, диктует необходимость поиска новых терапевтических агентов, не имеющих структурных аналогий с антибиотиками, утратившими свою эффективность. Открытие антимикробных пептидов и выявление их многообразных биологических функций показывает, что эти природные антибиотики являются перспективными молекулами для создания новых терапевтических антимикробных средств.
Антимикробные пептиды представляют собой единственный новый структурный класс антибиотиков, открытых за последние 30 лет [Hancock R.E. 1997. Peptide antibiotics. Lancet 349:418-422].
Наиболее изученными из известных антимикробных пептидов являются дефенсины (от англ. "defence" - защита). Такое название эти пептидные антибиотики получили благодаря их свойству быстро убивать бактерии, грибки, простейших, оболочечные вирусы и обеспечивать завершенность процесса фагоцитоза. Дефенсины и структурно-родственные им пептидные антибиотики (протегрины, тахиплезины, полифемузины и др.) выделены в настоящее время из фагоцитов млекопитающих (человека, свиньи, коровы, кролика, крысы, морской свинки, обезьяны), птиц (кур, индеек), рыб, ракообразных, а также из амебоцитов, эпителиальных клеток и гемолимфы насекомых (мухи, пчелы, стрекозы).
Антимикробные пептиды выделены из различных водных организмов, включая моллюсков, ракообразных, асцидий и рыб. В настоящее время выявлено более 80 различных дефенсинов, экспрессируемых в лейкоцитах и эпителиальных клетках птиц и млекопитающих [Lehrer R.I., Ganz T. 2002. Defensins of vertebrate animals. Curr. Opin. Immunol. 14: 96-102].
Три различных семейства дефенсинов обнаружены у позвоночных: α-дефенсины, β-дефенсины и циклические θ-дефенсины [Tang Y.Q., Yuan J., Osapay G. et al. 1999. A cyclic antimicrobial peptide produced in primate leukocyte s by the ligation of two truncated α-defensins. Science 286: 498-502].
У человека выявлено по крайней мере 10 различных молекул дефенсинов: шесть α-дефенсинов и четыре β-дефенсина [Cole A.M., Waring A.J. 2002. The role of defensins in lung biology and therapy. Ann. J. Respir. Med. 1: 249-259]. Известные пептиды обладают способностью инактивировать широкий спектр микроорганизмов, а потому именно с их наличием в нейтрофилах связывают эффективность киллерной стадии фагоцитоза.
Все обнаруженные ранее антимикробные пептиды имеют ряд общих структурных (катионный заряд молекул и наличие четного числа остатков цистеина, образующих внутримолекулярные дисульфидные мостики) и функциональных (антимикробных, цитотоксических и мембранотропных) свойств. α-Дефенсины содержат 29-35 аминокислотных остатков, в том числе 6 остатков цистеина, образующих 3 дисульфидных связи (Cys1-Cys6, Cys2-Cys4, Cys3-Cys5) [Ganz Т., Selsted M.E., Szklarek D. et al. 1985. Defensins. Natural peptide antibiotics of human neutrolhils. J. Clin. Invest. 76: 1427-1435]. β-Дефенсины содержат до 45 аминокислотных остатков, в том числе 6 остатков цистеина, образующих 3 дисульфидных связи (Cys1-Cys5, Cys2-Cys4, Cys3-Cys6) [Huttner K.M., Bevins C.L. 1999. Antimicrobial peptides as mediators of epithelial host defense. Pediatr. Res. 45: 785-794]. Таким образом, дефенсины являются достаточно крупными пептидами со сложной пространственной организацией. Это создает ряд сложностей при получении этих пептидов для использования в качестве терапевтических или профилактических средств. Во-первых, химический синтез таких крупных пептидных молекул неизбежно приводит к низкому выходу продукта и, следовательно, к его высокой себестоимости. Во-вторых, в процессе фолдинга вновь синтезированных молекул дефенсинов могут возникать проблемы, связанные с неправильным замыканием дисульфидных связей и формированием биологически неактивных структур.
Альтернативой химическому синтезу является использование систем для биологической экспрессии генов дефенсинов. Некоторые дефенсины были экспрессированы в клеточных биореакторах [Porter E.M., Liu L., Oren A., Anton P.A., Ganz Т. 1997. Localization of human intestinal defensin 5 in Paneth cell granules. Infect. Immunol 65: 2389-2395. Valore E.V., Park C.H., Quayle A.J. et al. 1998. Human β-defensin-1: an antimicrobial peptide of urogenital tissue. J. Clin. Invest. 101: 1633-1642], однако пока еще не известны высокоэффективные продуценты, позволяющие получать дефенсины в фармацевтическом производстве.
Изобретение решает задачу расширения арсенала пептидов, обладающих антимикробным действием.
Поставленная задача решается за счет структуры пептидов ареницинов, выделенных из морского кольчатого червя Arenicola marina, имеющих следующие аминокислотные последовательности:

где Х=Val или Ile
Пептиды проявляют антимикробную активность в отношении грамположительной (Listeria monocytogenes) и грамотрицательной (Escherichia coli) бактерий, а также дрожжеподобного гриба Candida albicans.
Заявляемые пептиды ареницин-1 и ареницин-2 принадлежат к новому структурному семейству пептидов, содержащих 21 аминокислотный остаток и стабилизированных только одной дисульфидной связью, образующей макроцикл из 18 аминокислотных остатков. Эта структурная особенность дает большие преимущества при получении биологически активной субстанции или аналогов заявляемых пептидов вследствие снижения риска неправильного замыкания дисульфидных связей и некорректного фолдинга.
Пептиды содержат по 21 аминокислотному остатку, имеют общую формулу Arg1-Trp2-Cys3-Val4-Tyr5-Ala6-Tyr7-Val8-Arg9-X10-Arg11-Gly12-Val13-Leu14-Val15-Arg16-Tyr17-Arg18-Arg19-Cys20-Trp21, являются гомологами, различающимися одним аминокислотным остатком в положении 10 (X - Val у ареницина-1 и Х - Ile у ареницина-2). Каждый из заявляемых пептидов содержит по 2 остатка цистеина, образующих одну дисульфидную связь. Ареницин-1 и ареницин-2 имеют молекулярные массы 2758,3 Да и 2772,3 Да соответственно.
Пептиды были выделены из природного источника - морского кольчатого червя пескожила (Arenicola marina), который принадлежит к семейству Arenicolidae, отряду Drilomorpha, классу Polychaeta, типу Annelida. В качестве источника новых антимикробных пептидов использованы целомоциты пескожила, представленные двумя основными морфологически и физиологически отличающимися типами: элеоцитами, выполняющими трофическую функцию, и лейкоцитами, принимающими участие в фагоцитозе и других реакциях противоинфекционной защиты.
Заявляемые пептиды принадлежат к новому структурному классу пептидных антибиотиков и могут быть использованы для борьбы с возбудителями болезней грибкового и бактериального происхождения. Они образуют цикл, состоящий из 18 аминокислотных остатков, и содержат только одну дисульфидную связь. Эта структурная особенность значительно облегчает процесс получения биологически активных ареницинов химическим или биотехнологическим путем.
Изобретение иллюстрируют примеры.
Пример 1.
Выделение антимикробных пептидов из целомоцитов пескожила Arenicola marina
Пептиды выделяют из морского кольчатого червя (пескожила) Arenicola marina. Сначала был проведен скрининг кислотных экстрактов различных органов и тканей животного (кишечника, покровного эпителия, железистых образований в ротовой области, целомоцитов, подкисленной бесклеточной целомической жидкости) на наличие в них антимикробных пептидов. Тестирование показало наличие антимикробных пептидов только в целомоцитах пескожила. На первой стадии выделения из животных получают целомическую жидкость, из которой целомоциты отделяют центрифугированием при 100 g в течение 10 минут. Полученную суспензию целомоцитов замораживают. Второй стадией выделения является экстракция антимикробных пептидов из целомоцитов пескожила 10% уксусной кислотой. Целомоциты пескожила размораживают и полученную суспензию гомогенизируют в растворе уксусной кислоты (на 1 г клеток 10 мл 10% раствора СН3СООН). Гомогенат суспендируют на магнитной мешалке на холоду (+4°С) в течение 18-24 часов, затем центрифугируют 2 часа при 25000 g. Супернатант подвергают ультрафильтрации через фильтр Amicon YM-10. Ультрафильтрат концентрируют высушиванием в вакууме на центрифуге Speed Vac CS-18 и подвергают препаративному электрофорезу в полиакриламидном геле (акриламид-12,5%, бис-акриламид - 0,33%) в кислой буферной системе (концентрация уксусной кислоты в геле 5,6%) в присутствии мочевины (концентрация мочевины в геле - 5,3 М). В качестве электродного буфера используют 5%-ную уксусную кислоту. Элекрофорез проводят при силе тока 30 мА на аппарате для препаративного электрофореза 491 Prep Cell фирмы "BioRad". Аликвоты каждой фракции тестируют на антимикробную активность методом радиальной диффузии в агарозном геле.
Активные фракции после препаративного электрофореза подвергают лиофилизации и дальнейшему фракционированию с помощью обращенно-фазовой высокоэффективной жидкостной хроматографии на колонке Vydac С-18 (4,6×250 мм; размер пор 300Å; диаметр частиц 5 мкм) в линейном градиенте концентрации ацетонитрила от 0 до 60% (1% мин-1) в 0,1% трифторуксусной кислоте. Содержание пептидов в препаратах и их чистоту оценивают методом электрофореза в пластинах полиакриламидного геля в кислой буферной системе в присутствии мочевины. В качестве контрольного препарата используют смесь дефенсинов кролика (NP-1,2,3,4,5). В результате хроматографического разделения одной из активных фракций было выявлено два мажорных пика, каждый из которых по данным электрофоретического анализа представляет собой чистый пептид. Пептид I элюируют с колонки при 29%, а пептид II - при 31% концентрации ацетонитрила. Хроматограмма разделения смеси, содержащейся в активной фракции, приведена на чертеже А. Как видно из хроматограммы, в активной фракции наряду с искомыми пептидами содержится еще ряд компонентов, среди которых нами были выявлены пептиды с молекулярными массами 1159 Да, 1745 Да, 2007 Да, 2450 Да, 3627 Да, также обладающие антимикробной активностью. В настоящее время для двух их них, имеющих молекулярные массы 1745 Да и 2450 Да, установлены частичные первичные структуры, отличные от структур заявляемых пептидов.
Гомогенность очищенных пептидов оценивают с помощью электрофореза в полиакриламидном геле в двух системах - в щелочной буферной системе в присутствии додецилсульфата натрия (чертеж В) и в кислой буферной системе в присутствии мочевины (чертеж C). Анализ исходной смеси компонентов, содержащихся в активной фракции, методом электрофореза в кислой буферной системе показывает наличие целого ряда пептидов с иными, нежели у заявленных пептидов, физико-химическими характеристиками, в частности с иной электрофоретической подвижностью (чертеж C, трек 2). Выделенные нами пептиды (чертеж C, треки 3 и 4, соответственно) по своим кислотно-основным свойствам близки к дефенсину кролика NP-4 (смесь дефенсинов кролика приведена на чертеж C, трек 1).
Молекулярную массу оценивают с помощью диск-электрофореза в присутствии додецилсульфата натрия. В качестве стандарта используют стандартную смесь белков для определения молекулярной массы (чертеж В; Sigma, США). По результатам электрофоретического анализа было сделано заключение, что пептиды I и II (на чертеже В треки 1 и 2 соответственно) имеют молекулярные массы около 2,8 кДа.
Пример 2.
Определение антимикробного действия.
Антимикробную активность выделенных пептидов определяют методом радиальной диффузии пептидов в агарозном геле с тест-культурой микроорганизма, позволяющим оценивать микробицидную активность, расходуя минимальное количество препарата. Для определения антимикробной активности отдельных фракций полипептидов используют культуры следующих микроорганизмов: Escherichia coli, штамм ML-35p (грамотрицательная бактерия); Listeria monocytogenes, штамм EGD (грамположительная бактерия); Candida albicans (гриб класса аскомицетов). Антимикробное действие выделенных пептидов фиксируют через 24 часа по наличию зон ингибирования роста тест-культуры (таблица).
Показано, что пептиды I и II целомоцитов пескожила в количестве ˜1 мкг проявляют микробицидную активность в отношении трех микробов. Действие обоих пептидов из целомоцитов пескожила в отношении грамположительных и грамотрицательных бактерий, а также их противогрибковая активность практически одинаковы.
Антимикробное действие пептидов ареницинов
Пример 3.
Определение молекулярной массы ареницинов.
Для определения молекулярной массы ареницинов используют MALDI (matrix-assisted lazer desorption ionization) масс-спектрометрический анализ на приборе VISION 2000 (ThermoBioAnalysis, OK). В качестве матрицы используют 0,15М 2,5-дигидроксибензойную кислоту в смеси, содержащей 25% метанол и 0,1% трифторуксусную кислоту. Образец облучают УФ-лазером с длиной волны 337 нм и максимальной энергией 250 мкДж в импульсном режиме с частотой 3 нс. При масс-спектрометрическом исследовании первого пептида (мощность лазера - 45%, число лазерных ударов - 47) получен пик с m/z 2758,1, соответствующий молекулярному иону М+ нового антибиотика ареницина-1 (расчетная молекулярная масса ареницина-1 составляет 2758,32 Да). При масс-спектрометрическом исследовании второго пептида (мощность лазера - 45%, число лазерных ударов - 69) получен пик с m/z 2772,3, соответствующий молекулярному иону M+ нового антибиотика ареницина-2 (расчетная молекулярная масса ареницина-2 составляет 2772,35 Да).
Пример 4.
Качественное и количественное определение дисульфидных связей.
Одной из характеристик, положенных в основу современной классификации антимикробных пептидов, является наличие в их структуре внутримолекулярных дисульфидных связей, играющих важную роль в формировании и поддержании их пространственной структуры. Наличие дисульфидных связей в выделенных пептидах определяют методом окисления надмуравьиной кислотой. О присутствии дисульфидных связей свидетельствует изменение электрофоретической подвижности пептидов, обработанных надмуравьиной кислотой, по сравнению с контрольными пробами при проведении аналитического электрофореза в кислой буферной системе в присутствии мочевины. При электрофоретическом анализе в качестве контроля используют дефенсин нейтрофилов кролика NP-1, содержащий три дисульфидных связи. Тест показал, что пептиды из целомоцитов пескожила изменяют свою электрофоретическую подвижность в геле после обработки надмуравьиной кислотой. Это свидетельствует о наличии в структуре пептидов дисульфидных связей.
Определение числа остатков цистеина проводилось путем их химической модификации с помощью 4-винилпиридина. Ареницины подвергают денатурации в 6М гуанидинхлориде в буферном растворе, содержащем 0,25 М Tris-Cl (pH 8,5) и 1 мМ ЭДТА. Затем проводят восстановление дисульфидных связей 10-кратным молярным избытком дитиотреитола с последующим алкилированием 5-кратным молярным избытком 4-винилпиридина под аргоном при комнатной температуре в течение 2 часов с образованием стабильных и легко идентифицируемых производных цистеина - S-пиридинэтилцистеинов. Масс-спектрометрический анализ полученных производных выявил молекулярные ионы с m/z 2970,3 для пиридинэтилированного ареницина-1 и m/z 2984,7 для пиридинэтилированного ареницина-2, что соответствует возрастанию молекулярной массы каждого пептида на 212 Да после модификации. Это свидетельствует о наличии в каждом пептиде двух остатков цистеина, образующих одну дисульфидную связь.
Пример 5
Определение первичной структуры пептидного антибиотика.
Для определения N-концевой аминокислотной последовательности выделенных антибиотиков применяют автоматическое микросеквенирование на приборе Procise 491 cLC Protein Sequencing System (Applied Biosystems, США). Идентификацию фенилтиогидантоин-производных аминокислот проводят на анализаторе 120А РТН Analyzer (Applied Biosystems, США). В основе методики автоматического определения аминокислотной последовательности пептидов и белков лежит метод химической деградации полипептидной цепи по методу Эдмана, позволяющему последовательно отщеплять N-концевые аминокислотные остатки в виде фенилтиогидантоинов и идентифицировать отщепленные производные аминокислот методом обращенно-фазовой высокоэффективной жидкостной хроматографии. В результате автоматического микросеквенирования выделенных пептидов установлены их частичные N-концевые аминокислотные последовательности:
Ареницин-1
Arg1-Trp2-X3-Val4-Tyr5-Ala6-Tyr7-Val8-Arg9-Val10-Arg11-Gly12-Val13-Leu14-Val15-Arg16-Tyr17-Arg18-Arg19-X20-X21
Ареницин-2
Arg1-Trp2-X3-Val4-Tyr5-Ala6-Tyr7-Val8-Arg9-Ile10-Arg11-Gly12-Val13-Leu14-Val15-Arg16-Tyr17-Arg18-Arg19-X20-X21
В результате повторного автоматического микросеквенирования пиридинэтилированных производных выделенных пептидов были подтверждены их N-концевые аминокислотные последовательности и установлена локализация остатков цистеина:
Пиридинэтилированный ареницин-1
Arg1-Trp2-PECys3-Val4-Tyr5-Ala6-Tyr7-Val8-Arg9-Val10-Arg11-Gly12-Val13-Leu14-Val15-Argl6-Tyr17-Arg18-Arg19-PECys20-X21
Пиридинэтилированный ареницин-2
Arg1-Trp2-PECys3-Val4-Tyr5-Ala6-Tyr7-Val8-Arg9-Ile10-Arg11-Gly12-Val13-Leu14-Val15-Arg16-Tyr17-Arg18-Arg19-PECys20-Х21,
где PECys - пиридинэтилцистеин
Таким образом, установлена частичная (95%) аминокислотная последовательность за исключением последнего аминокислотного остатка для каждого из выделенных пептидов:
Ареницин-1

Ареницин-2

Для установления полной первичной структуры выделена суммарная РНК из целомоцитов Arenicola marina. Для выделения РНК используют колонки Spin (Promega, США). Измельченные в жидком азоте ткани пескожила обрабатывают лизирующим раствором, содержащим гуанидинтиоцианат и β-меркаптоэтанол, для подавления активности эндогенных рибонуклеаз. Нуклеиновые кислоты осаждают этанолом, а затем перерастворяют в лизирующем растворе и снова добавляют этанол. Образовавшуюся смесь наносят на Spin колонку, которую обрабатывают ДНКазой I для освобождения от примесей геномной ДНК. Колонку промывают буферным раствором для удаления солей и других примесей, а затем элюируют тотальную РНК водой. Очищенную РНК переосаждают этанолом и доводят ее концентрацию до 0,25 мкг/мкл.
Определение структуры генов, кодирующих ареницины, проводят путем обратной транскрипции и амплификации (RT-PCR с помощью набора реактивов SMART* RACE** cDNA Amplification Kit (Clontech, США; *SMART - Switching Mechanism at 5' end of RNA Transcript; "RACE - Rapid Amplification of cDNA Ends). RACE-PCR представляет собой метод быстрой амплификации концевых последовательностей кДНК с помощью полимеразной цепной реакции. Этот метод является быстрой и менее трудоемкой альтернативой скринингу кДНК-библиотек. SMART RACE обладает рядом преимуществ по сравнению с другими методиками, а именно: более высокий выход полноразмерных 5'-последовательностей кДНК, отсутствие необходимости синтеза второй цепи кДНК и дотирования адаптера. Для амплификации используют TaqAdv II-полимеразу, отличающуюся повышенной процессивностью, чувствительностью и точностью амплификации. Структура ген-специфических праймеров для амплификации была выбрана, исходя из известной аминокислотной последовательности каждого пептида. При выборе олигонуклеотидной последовательности учитывают частоту использования кодонов у Arenicola marina и филогенетически близких организмов. Электрофорез продуктов полимеразной цепной реакции (ПЦР), полученных с разными ген-специфическими праймерами, показывает, что преимущественно амплифицируется фрагмент размером около 600 н.п. Фрагмент элюируют из геля и используют в лигазной реакции с плазмидным вектором для клонирования продуктов ПЦР. Трансформация клеток Escherichia coli с последующим скринингом на среде с карбенициллином и IPTG/X-Gal позволяет получить ряд клонов, содержащих плазмиду со вставкой. Из них 6 были отобраны для выделения плазмидной ДНК и секвенирования.
В результате секвенирования по методу Сэнгера стала известна последовательность 3'-концевой области кДНК, на основании структуры которой были синтезированы праймеры для амплификации 5'-концевой области. Амплификация 5'-концевой области представляет собой более трудную задачу, поскольку обратная транскриптаза часто обрывает синтез, не доходя до конца матрицы, поэтому в реакции образуется смесь полноразмерных и укороченных цепей кДНК. Использование второго праймера, который играет роль продолжения РНК-матрицы, позволяет получить полноразмерную к ДНК. На следующем этапе специфически амплифицируют фрагменты только полноразмерных кДНК. Дальнейшая работа по клонированию и определению 5'-концевой последовательности проводилась по той же схеме, что и для 3'-концевой области. Таким образом, была определена полная первичная структура кДНК.
В результате анализа полноразмерной кДНК нами была определена полная первичная структура предшественника зрелого пептида размером 202 аминокислотных остатка; найдена граница между препро-частью и зрелым пептидом и предполагаемая сигнальная последовательность. Установленная нуклеотидная последовательность показывает, что обнаруженные антимикробные пептиды имеют следующие аминокислотные последовательности:
Ареницин-1

Ареницин-2
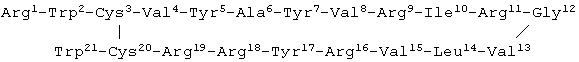
После анализа баз данных по аминокислотным последовательностям выявлено, что приведенные выше структуры не описаны ранее, а выделенные пептиды являются новыми антибиотиками. Заявляемые пептиды, названные ареницинами, не имеют структурного сходства с известными ранее представителями эндогенных пептидных антибиотиков и принадлежат к новому структурному типу антимикробных пептидов.
Подпись к чертежу.
А. Хроматограмма разделения активной фракции экстракта целомоцитов пескожила методом обращенно-фазовой высокоэффективной жидкостной хроматографии. Разделение на колонке Vydac C-18 (4,6×250 мм; размер пор 300 Å; диаметр частиц 5 мкм) в линейном градиенте концентрации ацетонитрила от 0 до 60% (1% мин-1) в 0,1% трифторуксусной кислоте.
В. Электрофорез ареницинов в полиакриламидном геле в щелочной буферной системе в присутствии додецилсульфата натрия. Трек 1 - ареницин-1, трек 2 - ареницин-2, трек 3 - стандартная смесь пептидов для определения молекулярной массы (Sigma, США).
С. Электрофорез активной фракции экстракта целомоцитов пескожила и выделенных ареницинов в кислой буферной системе в присутствии мочевины. Трек 1 - смесь дефенсинов кролика; трек 2 - исходная смесь пептидов, содержащихся в активной фракции целомоцитов пескожила; треки 3 - ареницин-1; трек 4 - ареницин-2.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АНТИМИКРОБНОГО ПЕПТИДА АРЕНИЦИНА | 2006 |
|
RU2316595C1 |
| Бета-шпилечный пептид, обладающий антимикробной активностью в отношении бактерий с множественной лекарственной устойчивостью | 2023 |
|
RU2812977C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pE-His8-TrxL-Ar2, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК, СОДЕРЖАЩИЙ АНТИМИКРОБНЫЙ ПЕПТИД АРЕНИЦИН МОРСКОГО КОЛЬЧАТОГО ЧЕРВЯ Arenicola marina, И ШТАММ Escherichia coli BL21(DE3)/pE-His8-TrxL-Ar2 - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА, СОДЕРЖАЩЕГО АРЕНИЦИН | 2006 |
|
RU2316590C1 |
| ПЕПТИД НИКОМИЦИН ИЗ МОРСКОГО КОЛЬЧАТОГО ЧЕРВЯ NICOMACHE MINOR, ОБЛАДАЮЩИЙ АНТИМИКРОБНЫМ И ПРОТИВООПУХОЛЕВЫМ ДЕЙСТВИЕМ. | 2019 |
|
RU2721273C1 |
| БЕТА-ШПИЛЕЧНЫЙ ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ | 2015 |
|
RU2624020C2 |
| ПЕПТИДЫ ЛАТАРЦИНЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2005 |
|
RU2302466C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pЕ-Trx-Lc-def, ШТАММ Escherichia coli ДЛЯ ЭКСПРЕССИИ АНТИМИКРОБНОГО ПЕПТИДА ДЕФЕНСИНА ЧЕЧЕВИЦЫ Lens culinaris И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ПЕПТИДА | 2010 |
|
RU2456345C1 |
| ПЕПТИДЫ, ПРОЯВЛЯЮЩИЕ АНТИМИКРОБНУЮ АКТИВНОСТЬ | 2005 |
|
RU2302425C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET-KSI-Buf2, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК, СОДЕРЖАЩИЙ АНТИМИКРОБНЫЙ ПЕПТИД БУФОРИН-2, ШТАММ Escherichia coli BL21(DE3)/pET-KSI-Buf2 - ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ АНТИМИКРОБНОГО ПЕПТИДА БУФОРИНА-2 | 2007 |
|
RU2347811C1 |
| ПЛАЗМИДНЫЙ ВЕКТОР pE-Trx-Aur, ШТАММ ESCHERICHIA COLI ДЛЯ ЭКСПРЕССИИ АНТИМИКРОБНОГО ПЕПТИДА АУРЕЛИНА И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ПЕПТИДА | 2009 |
|
RU2412999C1 |
Изобретение относится к области биохимии и может быть использовано в медицине. Из кольчатого червя Arenicola Marina выделены две формы пептида с широким спектром антимикробного действия. Указанные формы различаются одним аминокислотным остатком в положении 10. В положении 10 ареницин-1 имеет Val, а ареницин-2 - Ile. Применение изобретения позволяет расширить ассортимент антимикробных средств. 1 ил., 1 табл.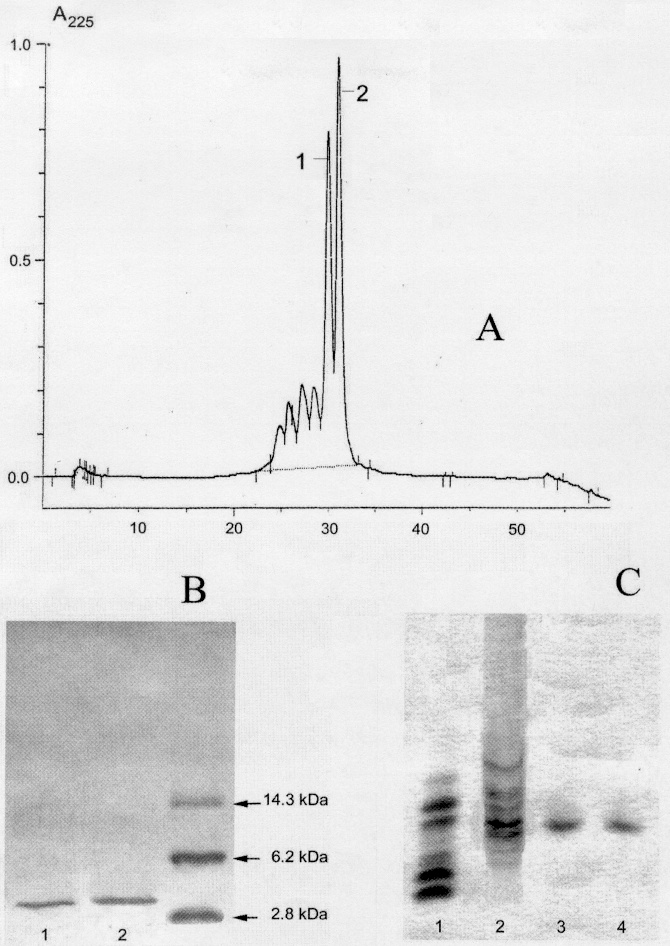
Пептид, обладающий антимикробным действием и имеющий следующую формулу:

где X=Val (ареницин-1) или X=Ile (ареницин-2).
| US 6337093, 08.01.2002 | |||
| Новые методы анализа аминокислот, пептидов и белков | |||
| Под ред | |||
| акад | |||
| Ю.А.Овчинникова | |||
| - М.: "МИР", 1974, с.50-80 | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |

