Область изобретения
Настоящее изобретение относится к изоцитратдегидрогеназе, которая катализирует продуцирование НАДФН, необходимого для биосинтеза липидов, включая жирные кислоты, сквален и холестерин, и к ее применению в лечении болезней обмена веществ, включая ожирение, гиперлипидемию и жировую инфильтрацию печени. Также, настоящее изобретение относится к гену изоцитратдегидрогеназы, слитым генным конструкциям, содержащим этот ген, клеткам-трансфектантам, несущим эти гены в своем геноме, и к трансгенным животным, способным к непрерывной экспрессии изоцитратдегидрогеназы на протяжении всей их жизни.
Предшествующий уровень техники
Принимая участие в цикле ТКК (трикарбоновых кислот), изоцитратдегидрогеназа катализирует окислительное декарбоксилирование лимонной кислоты в α-кетоглутарат с сопутствующим продуцированием НАДН или НАДФН.
У высших животных изоферменты изоцитратдегидрогеназы можно разделить на три класса в соответствии с их кофакторами и локализацией в клетке: митохондриальная НАД+-зависимая изоцитратдегидрогеназа (далее называемая «IDH»), митохондриальная НАДФ+-зависимая изоцитратдегидрогеназа (далее называемая «IDPm») и цитоплазматическая НАДФ+-зависимая изоцитратдегидрогеназа (далее называемая «IDPc»). Предполагают, что из этих изоцитратных изоферментов IDH играет основную роль в окислительном декарбоксилировании изоцитрата в цикле трикарбоновых кислот (ТКК) с сопутствующим продуцированием α-кетоглутарата и НАДН. НАДН используется для генерации энергии посредством системы переноса электронов, а α-кетоглутарат представляет собой метаболит, используемый в синтезе аминокислот, таких как глутаминовая кислота, глутамин, аргинин и пролин, а также других биологических продуктов. Активность IDH регулируется как контрольная точка цикла ТКК. Следовательно, IDH является ключевым ферментом для регуляции не только цикла ТКК, но также энергетического метаболизма, биосинтеза белка и метаболизма азота, поскольку метаболиты цикла ТКК принимают участие в этих процессах метаболизма.
IDH находится в исследовании с момента ее выделения из дрожжей и свиней. Дрожжевая IDH представляет собой аллостерически регулируемый фермент, который существует в виде октамера, состоящего из двух неидентичных субъединиц IDH1 и IDH2, имеющих высокую гомологию друг с другом. IDH1 играет роль в регуляции ферментативной активности, тогда как IDH2 ответственна за каталитическую активность (Keys, D.А. & McAlister-Henn, L., J.Bacteriol., 172, 4280-4287, 1990). Разбитая на три субъединицы (α, β и γ субъединицы), свиная IDH также существует в виде октамера (2 (α 2β γ)) в активной форме.
Хотя обнаружено, что IDPm и IDPc структурно состоят из двух частей, однако их функции не известны. Хотя оба фермента имеют молекулярную массу примерно 45 кДа при высокой гомологии, они были идентифицированы как разные независимые белки, что проанализировали с помощью экспериментов с иммунологическими реакциями, включающими использование поликлональных антител (Plaut, G.W.E. et al., Biochem. Biophys. Acta, 760, 300-308, 1983; Fantania, H.R. et al., FEBS, 322, 245-248, 1993). В частности, IDPm и IDPc высокотканеспецифичны. В тканях сердечной мышцы, например, более 90% суммарной НАДФ+-зависимой изоцитратдегидрогеназы находится в митохондриях, а оставшиеся 10% обнаружены в цитоплазме. Напротив, сообщают, что 3% суммарной НАДФ+-зависимой изоцитратдегидрогеназы тканей печени находится в митохондриях, тогда как оставшиеся 97% находятся в цитоплазме (Plaut, G.W.E., Current Topics in Cell Regulation, 2, 1-27, 1983).
Как упомянуто выше, изоферменты изоцитратдегидрогеназы охарактеризованы в отношении их некоторых структурных характеристик, но не в отношении функций. В частности, исследования точных механизмов IDPm и IDPc не обнаружены до публикации недавних сообщений, в которых только сделано предположение, что IDPm катализирует обратную реакцию цикла ТКК по превращению α-кетоглутарата через изоцитрат в цитрат, который связан с трикарбоксилатпереносящей системой для доставки ацетил-КоА, предшественника для биосинтеза жирных кислот и холестерина, с сопутствующим превращением цитрата в оксалоацетат с повышением уровней фосфоенолпирувата в цитоплазме, стимулируя таким образом гликонеогенез (Des Rosiers, С. et al., J. Biol. Chem., 269, 27179-27182, 1994; Fernandez, С.A. et al., J. Biol. Chem., 270, 10037-10042, 1995).
Важность IDPm в гликонеогенезе предполагается вследствие катализа ею обратной реакции цикла ТКК. Напротив, ни одно из сообщений об IDPc не касается ее метаболических функций. Известно, что IDPc экспрессируется в больших количествах в яичнике и молочной железе. Из ферментов, продуцирующих НАДФН, существующих в печени крыс, количественно проанализировали, что IDPc продуцирует НАДФН в количествах, больших, чем это делают важные ферменты пентозофосфатного пути, то есть глюкозо-6-фосфатдегидрогеназа - для превращения глюкозо-6-фосфата в 6-фосфоглюконо-δ-лактон и НАДФН, 6-фосфоглюконатдегидрогеназа - для превращения 6-фосфоглюконата в рибулозо-5-фосфат и НАДФН и цитоплазматическая малатдегидрогеназа - для превращения малата в пируват и НАДФН, в 16, 8 и 18 раз соответственно (Veech, R.L. Et al., Biochem. J., 115, 609-619, 1969).
В цитоплазме различные ферменты, вовлеченные в пути метаболизма жирных кислот, холестерина и гормонов, требуют большого количества НАДФН для своей каталитической активности. Поэтому считают, что ферменты, продуцирующие НАДФ, такие как глюкозо-6-фосфатдегидрогеназа и малатдегидрогеназа, играют важную роль в поставке НАДФН в цитоплазму. Однако в свете способности IDPc продуцировать цитоплазматический НАДФН, ожидается, что она в большей степени ответственна за регуляцию поставки НАДФН. В конечном счете, предполагают, что IDPc играет критическую роль в биосинтезе жирных кислот и холестерина. Среди синтаз жирных кислот, вовлеченных в биосинтез жирных кислот, β-кетоацил-[ацил-переносящий белок]-редуктазе и еноил-АПБ-редуктазе для катализа необходим НАДФН в качестве кофактора. В биосинтезе холестерина большое количество НАДФН необходимо для реакций, катализируемых гидроксиметил-глютарил-КоА-редуктазой и скваленсинтетазой, и для конечной 19-стадийной реакции получения холестерина из ланостерина. Соответственно, регуляция активности IDPc, функции которой состоят в поставке большей части НАДФН, необходимого в клетке, очень важна для регуляции биосинтеза жирных кислот и их производных, липидов, сквалена, а также холестерина и его производных.
У высших животных отложение липидов происходит в следующем порядке. Когда имеется избыток источников энергии, дифференциация жировых клеток ускоряется, приводя в результате к увеличению числа и размера белых жировых тканей с сопутствующим отложением липидов. В свою очередь, белая жировая ткань дает возможность активной экспрессии гена ob, что приводит к повышению уровня лептина в организме. В ответ на это изменяется гормональное действие головного мозга в направлении снижения аппетита. В то же время избыточные калории потребляются для поддержания температуры тела при использовании белков-разобщителей (UCP, uncoupler proteins). В белой жировой ткани активируется экспрессия генов, которые кодируют основные факторы транскрипции для пролиферации жировых клеток, такие как активируемый пролифератором пероксисом рецептор γ (peroxysome proliferator-activated receptor γ, PPARγ), C/EBPα и ADD1/SREBP1. Таким образом, происходит стимуляция дифференциации жировых клеток и отложения липидов, и избыток энергии организма запасается в форме липидов, так что энергия организма сбалансирована (Hu, Е. et al., Proc. Natl. Acad. Sci. USA, 92, 9856-9860, 1995; Keller, H. et al., Proc. Natl. Acad. Sci. USA, 20, 9856-9860, 1993; Freytag S.О. et al., Genes Dev., 8, 1654-1663, 1994; Tontonoz, P. et al., Mol. Cell. Bid., 13, 4753-4759, 1993; Spiegelman, В.М., Cell, 87, 377-389, 1996). Примеры лигандов, необходимых для активации PPARγ, основного фактора транскрипции для дифференциации жировых клеток, включают полиненасыщенные жирные кислоты, такие как линолевая кислота, докозагексановая кислота (DHA) и арахидоновая кислота (Krey, G. et al., Mol. Endocrinol., 11, 779-791, 1997; Yu et al., J. Biol. Chem., 270, 23975-23983, 1995). Также известно, что простагландин J2 служит в качестве лиганда этого основного фактора транскрипции (Forman В.М. et al., Cell, 83, 803-812, 1995; Kliewer S. A. et al., Cell, 83, 813-819, 1995).
На основании данного обзора можно сделать вывод о высокой вероятности того, что IDPc может быть непосредственно вовлечен в регуляцию биосинтеза различных жирных кислот, холестерина и гормонов благодаря его способности продуцировать НАДФН. Можно также предполагать, что IDPc играет ключевую роль в ожирении и жировой инфильтрации печени, стимулируя продуцирование активирующих лигандов для PPARγ, таких как полиненасыщенные жирные кислоты и арахидоновая кислота, для запуска каскада экспрессии различных генов, связанных с дифференциацией жировых клеток. Кроме того, необходимость большого количества НАДФН для биосинтеза холестерина предлагает возможность того, что искусственная регуляция внутриклеточных уровней IDPc и продукта его реакции НАДФН может дать средство регуляции биосинтеза холестерина.
Краткое изложение сущности изобретения
Ведущее к настоящему изобретению интенсивное и тщательное исследование механизма биосинтеза липидов путем молекулярно-биологических и биохимических экспериментов с использованием трансфицированных клеток животных и трансгенных мышей, осуществленное авторами настоящего изобретения, привело в результате к открытию того, что внутриклеточные уровни IDPc и продукта реакции с его участием НАДФН оказывают бесспорное влияние не только на скорость дифференциации жировых клеток и отложение липидов в жировых клетках, но и на биосинтез липидов и холестерина.
Следовательно, в задачу настоящего изобретения входит предложение фермента изоцитратдегидрогеназы для продуцирования НАДФН и ее гена.
В задачу настоящего изобретения также входит предложение слитой генной конструкции, которая содержит ген, кодирующий изоцитратдегидрогеназу, клетки-трансфектанта, которая несет этот ген в своем геноме, и трансгенного животного, которое может экспрессировать этот ген непрерывно на протяжении всей своей жизни.
В задачу настоящего изобретения также входит предложение применения изоцитратдегидрогеназы и ее гена в лечении и профилактике ожирения, гиперлипидемии и жировой инфильтрации печени или в биосинтезе липидов.
Краткое описание графических материалов
На фиг.1 представлены схематические диаграммы, показывающие структуры базового вектора LNCX (вверху), рекомбинантного вектора, в который встроен ген IDPc в смысловой ориентации для повышения экспрессии гена IDPc в жировых клетках NIH3T3 F442A (посередине), и рекомбинантного вектора, в который ген IDPc встроен в антисмысловой ориентации для снижения экспрессии гена IDPc в жировых клетках NIH3T3 F442A (внизу).
На фиг.2а представлены оптические фотографии, показывающие окрашенные Oil-Red-О жировые клетки, дифференцировавшие из нормальных NIH3T3 F442A (слева), клетки-трансфектанты FS1 с повышенной экспрессией гена IDPc (в центре) и клетки-трансфектанты FAS1 со сниженной экспрессией гена IDPc (справа) на чашках (верхняя панель) и частично при 200-кратном увеличении (нижняя панель).
На фиг.2б представлены оптические фотографии, показывающие отложение липидов в жировых клетках в зависимости от дозы НАДФН.
Фиг.3 представляет собой схему, иллюстрирующую конструирование рекомбинантного вектора экспрессии для использования при создании трансгенного животного, в которого встроена кДНК IDPc в прямом направлении от промотора гена РЕРСК (фосфоенолпируваткарбоксикиназы) крысиного происхождения.
На фиг.4 представлены фотографии, показывающие сравнение размера тела и отложения в виде жирового тела в придатке яичка между потомством F1 от трансгенных мышей по настоящему изобретению и нормальными мышами.
На фиг.5 представлены авторадиографии, показывающие повышение уровня экспрессии генов, показательных для ожирения, в жировой ткани трансгенных мышей по настоящему изобретению по сравнению с нормальными мышами.
Фиг.6а представляет собой гистограмму сравнения массы тела трансгенных мышей F1 с массой тела нормальных мышей.
Фиг.6б представляет собой гистограмму сравнения массы печени трансгенных мышей F1 с массой печени нормальных мышей.
Фиг.6в представляет собой гистограмму сравнения активности IDPc и уровня IDPc в крови у трансгенных мышей F1 с этими же параметрами у нормальных мышей.
Фиг.6г представляет собой гистограмму сравнения [НАДФН]/[НАДФН+НАДФ+] у трансгенных мышей F1 с этими же параметрами у нормальных мышей.
Фиг.6д представляет собой гистограмму сравнения массы жирового тела придатка яичка у трансгенных мышей F1 с этим же параметром у нормальных мышей.
Фиг.6е представляет собой гистограмму сравнения уровней триглицеридов и холестерина в крови у трансгенных мышей F1 с этими же уровнями у нормальных мышей.
Фиг.6ж представляет собой гистограмму сравнения уровней триглицеридов и холестерина в печени у трансгенных мышей F1 с этими же параметрами у нормальных мышей.
Фиг.6з представляет собой гистограмму сравнения уровней лептина в крови у трансгенных мышей F1 с этими же параметрами у нормальных мышей.
На фиг.7а представлены фотографии, показывающие ткани печени трансгенных мышей по настоящему изобретению и контрольных мышей.
На фиг.7б представлены фотографии, показывающие жировую клетку трансгенных мышей по настоящему изобретению и контрольных мышей.
Фиг.8а представляет собой график, иллюстрирующий ингибиторную активность щавелево-яблочной кислоты в отношении активности изоцитратдегидрогеназы.
Фиг.8б представляет собой график, иллюстрирующий ингибиторную активность метилизоцитрата в отношении активности изоцитратдегидрогеназы.
На фиг.9 представлены оптические фотографии, показывающие окрашенные Oil-Red-О жировые клетки, дифференцировавшие из клеток NIH3T3 F442A, обработанных ингибиторами изоцитратдегидрогеназы (слева), обработанные щавелево-яблочной кислотой (в центре) и метилизоцитратом (справа) при 100-кратном увеличении (верхний ряд) и 200-кратном увеличении (нижний ряд).
Фиг.10а представляет собой гистограмму, иллюстрирующую сравнение масс печени и жирового тела придатка яичка у крыс, которым вводили ингибитор изоцитратдегидрогеназы по настоящему изобретению, с этими параметрами у крыс, которым не вводили ингибиторы.
Фиг.10б представляет собой гистограмму, иллюстрирующую сравнение уровней триглицеридов и холестерина в крови у крыс, которым вводили ингибитор изоцитратдегидрогеназы по настоящему изобретению, с таковыми параметрами у крыс, которым не вводили ингибитор.
Фиг.10в представляет собой гистограмму, иллюстрирующую сравнение уровня ЛПВП (липопротеинов высокой плотности) в крови у крыс, которым вводили ингибитор изоцитратдегидрогеназы по настоящему изобретению, с таковыми параметрами у крыс, которым не вводили ингибитор.
Подробное описание изобретения
В одном аспекте изобретение относится к ферменту изоцитратдегидрогеназе, который катализирует продуцирование НАДФН, необходимого для биосинтеза жирных кислот и холестерина и для отложения липидов, а также к гену, кодирующему изоцитратдегидрогеназу.
Полезной для настоящего изобретения является IDPc, выделенная из мышей. Ген IDPc мышиного происхождения по настоящему изобретению, соответствующий последовательности №3 в перечне последовательностей, имеет открытую рамку считывания (ОРС) размером 1245 п.н., с 3'-нетранслируемым районом (НТР), в котором имеется последовательность оснований ААТААА, предполагаемый поли-А-сигнал. Белок IDPc, кодируемый геном IDPc, состоит из 414 аминокислот, последовательность №4 в перечне последовательностей, с молекулярной массой 46575 Да. Сравнительный анализ аминокислотных последовательностей IDPc из различных видов показывает, что мышиная IDPc по изобретению имеет гомологию 97,8% с крысиной IDPc, 68,5% с бычьей IDPm и 64,4% с дрожжевой IDPc. В частности, аминокислотная последовательность 412-414 мышиной IDPc идентична последовательности-мишени пероксисомы, о которой известно, что она вовлечена в биосинтез и расщепление жирных кислот и холестерина. Следовательно, это позволяет предположить высокую вероятность того, что IDPc перемещается к пероксисомам и там принимает участие в синтезе жирных кислот и холестерина.
Кроме IDPc, IDPm, ген, имеющий последовательность оснований, схожую с последовательностью оснований гена IDPc, также используется для продуцирования НАДФН, необходимого для биосинтеза жирных кислот и холестерина и для отложения липидов в соответствии с настоящим изобретением.
В другом аспекте настоящее изобретение относится к слитой генной конструкции, содержащей ген, к новому клеточному штамму, в котором закреплен этот ген, и к трансгенному животному, которое экспрессирует этот ген непрерывно на протяжении всей своей жизни.
С этой целью, во-первых, интересующий ген встроен в вектор экспрессии млекопитающего таким образом, чтобы этот ген транскрибировался в смысловом направлении или в антисмысловом направлении.
В этом отношении в качестве носителя гена предпочтительно используют ретровирусные векторы экспрессии, причем наиболее предпочтителен ретровирусный вектор pLNCX. pLNCX, который имеет происхождение от MMLV (вирус лейкемии мышей Moloney), имеет промотор CMV (цитомегаловируса) для экспрессии экзогенных генов в клетках млекопитающих и ген неомицина в качестве селективного маркера вместе с последовательностью LTP (длинный концевой повтор, long terminal repeat), представляющей собой фактор идентификации ретровирусных векторов.
Среди слитых генных конструкций, полученных таким образом, те, которые транскрибируются в смысловом направлении с помощью промотора CMV, используют для усиления экспрессии IDPc, тогда как конструкции, которые предназначены для антисмысловой транскрипции, используют для подавления экспрессии. Полученные в результате рекомбинантные векторы вводят в клетки NIH3T3 L1, являющиеся видом предшественников жировых клеток. Среди клеток-трансфектантов, которые идентифицированы как интегрировавшие ген IDPc в свои геномы, клетки, в которых ген IDPc встроен в смысловом направлении, обозначили как FS1 и депонировали в Корейской Коллекции Типовых Культур Корейского Научно-Исследовательского Института Биологических наук и Биотехнологии (KRIBB) под номером КСТС 0861 ВР 6 сентября 2000 г. С другой стороны, клеточные штаммы, в которых ген IDPc встроен в антисмысловом направлении, обозначили как FAS1.
Определили, что по сравнению с мышиными клетками NIH3T3 L1, в которые введен только вектор pLNCX (контроль), ферментативная активность примерно в 2 раза выше в мышиных клетках-трансфектантах NIH3T3 L1, в которых ген IDPc встроен в смысловом направлении (FS1), но примерно в 0,4 раза ниже в мышиных клетках-трансфектантах NIH3T3 L1, в которых ген IDPc встроен в антисмысловом направлении (FAS1).
Действие IDPc на биосинтез жирных кислот можно количественно определить с помощью Oil-Red-O, красителя, специфичного в отношении липидов, который наносят на жировые клетки, которые дифференцировали из клеток-трансфектантов после обработки инсулином. В результате обнаружили, что продуцирование липидов происходит более активно в клетке-трансфектанте FS1 с повышенной экспрессией гена IDPc, чем в контрольной клетке. С другой стороны, обнаружили небольшое по сравнению с контрольными клетками отложение липидов в клетках-трансфектантах FAS1 со сниженной экспрессией гена IDPc (см. фиг.2а). При дифференциации в жировые клетки в присутствии НАДФН, который является продуктом ферментативной реакции с участием IDPc, клетки-трансфектанты, в которые введен только pLNCX, то есть контрольные клетки, показывают более высокие скорости дифференциации и большие внутриклеточные отложения липидов по мере повышения концентрации НАДФН (см. фиг.2б). Эти результаты указывают на то, что IDPc, ее ген или продукт ферментативной реакции с ее участием, НАДФН, играют ключевую роль в определении внутриклеточных липидных отложений.
Далее, чтобы исследовать активность изоцитратдегидрогеназы, слитую генную конструкцию использовали для получения трансгенного животного, в геноме которого закреплен ген IDPc.
1. Получение слитой генной конструкции
Для постоянной экспрессии гена IDPc необходима интеграция этого гена в геном животного. С этой целью, во-первых, необходимо сконструировать рекомбинантный вектор, который может экспрессировать интересующий ген в млекопитающих. В полученной в результате слитой генной конструкции экспрессия интересующего гена регулируется подходящим геном-промотором. Термин «слитая генная конструкция», как его используют здесь, означает функциональную совокупность генов для использования в трансформации определенных организмов, которая содержит по существу по меньшей мере один структурный ген и по меньшей мере один цис-действующий регуляторный элемент для контроля экспрессии этого структурного гена.
Как правило, цис-действующий регуляторный элемент может быть промотором, энхансером, интроном, 5'-НТР (нетранслируемый район) и 3'-НТР. В слитой генной конструкции цис-действующий регуляторный элемент может быть расположен в любом месте на расстоянии 10 т.п.н. или менее от 5'-фланкирующего района, 3'-фланкирующего района, 5'-конца или 3'-конца структурного гена или внутри этого структурного гена (в случае интрона). Кроме структурного гена и цис-действующего регуляторного элемента, слитая генная конструкция дополнительно содержит различные компоненты, включая сигнал полиаденилирования для повышения скоростей транскрипции или трансляции, последовательность связывания рибосом, интрон и т. д. Дополнительно к этому, слитую генную конструкцию можно обеспечить последовательностью оснований для повышения эффективности встраивания интересующего гена в геном или в определенные сайты и маркерным геном для идентификации встраивания.
Промотор для слитой генной конструкции, который следует использовать при получении трансгенного животного, включает промотор CMV или области, регулирующие экспрессию для генов, экспрессирующихся в белых жировых тканях, таких как гены, кодирующие липопротеинлипазу (LPL), адипсин, белок адипоцитов 2 (аР2) и IDPc. В предпочтительном воплощении настоящего изобретения используют промотор крысиного происхождения для гена цитозольной фосфоенолпируваткарбоксикиназы (РЕРСК), который экспрессируется как в печени, так и в белых жировых тканях.
Более подробно, получение трансгенного животного, у которого происходит постоянная экспрессия IDPc, начинается с крысиного гена цитозольной РЕРСК. Из этого гена получают последовательность 2,2 т.п.н. в обратном направлении от 5'-конца, содержащую промотор. В прямом направлении от этой последовательности встраивают кДНК мышиной IDPc в смысловой ориентации, чтобы получить слитую генную конструкцию, которую обозначили как pPEPCKIDPc. Существует два вида генов РЕРСК: один кодирует цитозольный фермент, а другой кодирует митохондриальный фермент. В настоящем изобретении использовали последовательность, находящуюся в обратном направлении от 5-конца гена, кодирующего цитозольный РЕРСК (далее называемый «РЕРСК-С»). В печени, кишечнике и почках ткани, где ген РЕРСК-С экспрессируется под контролем этого промотора, определяют в зависимости от регуляторных районов, имеющихся в последовательности, находящейся в обратном направлении от 5'-конца. Последовательность 2,2 т.п.н. в обратном направлении от 5'-конца гена РЕРСК-С, используемая в настоящем изобретении, содержит генную последовательность около 987 нуклеотидов (nt), которая известна как регуляторный район, необходимый для эффективной экспрессии в белой жировой ткани (Hanson, R.W. Annu. Tev. Biochem., 66, 581-611, 1997).
Для получения трансгенных животных полезны мыши, но и любое животное, если его можно сделать трансгенным, полезно в настоящем изобретении, поскольку IDPc представляет собой фермент, экспрессирующийся у всех высших животных.
2. Получение эмбриона
Одной из наиболее важных стадий в получении трансгенного животного является введение слитой генной конструкции в эмбрион. Это введение осуществляют с помощью микроинъекционной системы. При микроинъекции слитой генной конструкции в эмбрион предпочтительно используют автоматическую микроинъекционную систему, которая способна автоматически регулировать количества ДНК до предела 4 пл, поскольку по доле успешных попыток она превосходит традиционные ручные микроинъекционные системы. Мышиный эмбрион, который содержит слитую генную конструкцию IDPc, депонирован в Корейской Коллекции Типовых Культур Корейского Научно-исследовательского института Биологических наук и Биотехнологии (KRIBB) под номером КСТС 0874 ВР 4 ноября 2000 г.
3. Получение трансгенного животного
Далее эмбрион, содержащий слитую генную конструкцию, имплантируют суррогатной матери для получения трансгенного животного. По изобретению имплантацию эмбриона суррогатной матери для удобства проводили на одноклеточной стадии эмбриона, а не на двухклеточной стадии. Немедленно после микроинъекции слитой генной конструкции эмбрион на одноклеточной стадии имплантируют в маточную трубу суррогатной матери, чтобы сократить различные процессы, необходимые для культивирования эмбриона до двухклеточной стадии. Для имплантации в маточную трубу на двухклеточной стадии, например, необходимо культивировать эмбрион в течение одних дополнительных суток в инкубаторе. Чтобы имплантировать эмбрион на двухклеточной стадии в воронку маточной трубы этот эмбрион необходимо вводить глубоко в маточную трубу либо необходимо прокалывать маточную трубу с помощью иглы. Однако имплантацию эмбриона на одноклеточной стадии суррогатной матери можно проводить в условиях, подобных обычным условиям для мышиных эмбрионов, хотя местом имплантации является воронка маточной трубы.
Используя полученное таким образом трансгенное животное, исследовали активность IDPc in vivo в отношении следующих показателей:
1. Увеличение жирового тела придатка яичка
Через 23 недели после рождения гетерозиготные трансгенные мыши F1 выросли больше, чем контрольные мыши. При анатомировании определили, что у гетерозиготных трансгенных мышей F1 значительно увеличен размер жирового тела придатка яичка, причем масса тела была в 14 раз выше, чем у контрольных мышей. Кроме того, при разделке на волокна у трансгенных мышей на спинах наблюдали кожные тучные клетки большего размера и в большем количестве по сравнению с контрольными мышами. При полном удалении кожи живота у трансгенных мышей наблюдали значительное увеличение жирового тела придатка яичка (см. фиг.4).
2. Скорость экспрессии гена, показательного для ожирения
Проводили исследование, оказывает ли экспрессия гена IDPc влияние на экспрессию генов, показательных для ожирения. В жировом теле придатка яичка мышей, трансгенных по гену IDPc, обнаруживали экспрессию введенного рекомбинантного гена IDPc параллельно с экспрессией их эндогенного гена IDPc, что демонстрировало, что суммарная экспрессия IDPc повышена. Кроме того, обнаружили повышение экспрессии генов, показательных для ожирения, таких как гены, кодирующие белок адипоцитов 2 (аР2), адипсин, липопротеинлипазу (LPL), лептин, фактор некроза опухоли α (TNF-α) и активируемый пролифератором пероксисом рецептор γ (PPARγ), обо всех из которых известно, что они проявляют повышенную экспрессию, способствуя дифференциации тучных клеток (Hwang, С.S. et al., Ann. Rev. Cell Dev. Biol., 13, 231-259, 1997; Lemberger, Т. et al., Annu. Rev. Cell Dev. Bid., 12, 225-362, 1996; Spiegelman, В.М. et al., Cell, 87, 377-389, 1996).
В свете недавнего сообщения (Spiegelman, В.М. et al., Cell, 87, 377-389, 1996), показавшего, что PPARγ служит в качестве основного фактора транскрипции как при дифференциации тучных клеток, так и при биосинтезе липидов, повышение экспрессии генов, показательных для ожирения, таких как гены, кодирующие аР2, адипсин, LPL, лептин и/или TNF-α, у мышей, трансгенных по IDPc, является результатом повышения экспрессии гена PPARγ. Следовательно, можно сделать вывод, что повышение активности IDPc, связанное с активной экспрессией гена IDPc, и сопутствующее повышение уровня НАДФН, вызывает сильное повышение экспрессии гена PPARγ, который необходим как для дифференциации жировых клеток, так и для биосинтеза липидов (см. фиг.5). В свою очередь, с учетом сообщения (Kirn, J.В. et al., Proc. Natl. Acad. Sci. USA. 95, 4333-4337, 1998), в котором раскрыто, что повышение уровня лиганда, необходимого для активации PPARγ, стимулирует экспрессию самого гена PPARγ, эти результаты приводят к выводу, что повышение активности IDPc и уровня НАДФН, который является продуктом этого фермента, сначала стимулирует продуцирование полиненасыщенных жирных кислот, которые служат в качестве лигандов, способных индуцировать активацию PPARγ, и затем, в ответ на это, индуцирует экспрессию самого гена PPARγ, таким образом повышая уровень экспрессии генов, показательных для дифференциации, таких как гены, кодирующие ар2, адипсин, LPL и лептин, которые вовлечены в дифференциацию различных жировых клеток и, в конечном счете, вызывают ожирение и жировую инфильтрацию печени.
3. Идентификация отложения липидов в организме
Обнаружили, что трансгенные мыши проявляют активность IDPc в 2,7 и 1,4 раза выше соответственно в печени и в жировом теле придатка яичка по сравнению с активностью IDPc в этих органах у контрольных мышей (см. фиг.6в). Таким образом, отношение НАДФН к суммарному пулу НАДФ ([НАДФН]/[НАДФ+]+[НАДФН]) повышается при повышении ферментативной активности IDPc (см. фиг.6г). Масса трансгенных мышей была увеличена на 35% или более по сравнению с массой нормальных мышей (см. фиг.6а), при этом более значительно было увеличено жировое тело придатка яичка у трансгенных мышей по сравнению с контрольными мышами (см. фиг.6д). Изменений в массе печени, тем не менее, не обнаружили (см. фиг.6б).
Кроме того, измерили, что уровни триглицеридов и суммарного холестерина в крови трансгенных мышей были в 1,8 и 2,4 раза выше, чем эти показатели у контрольных мышей (см. фиг.6е). Как и в крови, уровни как триглицеридов, так и холестерина в печени у трансгенных мышей были повышены (см. фиг.6ж). Уровень лептина, белка, продуцируемого главным образом тучными клетками, в крови определили как в два раза более высокий у трансгенных мышей, чем у контрольных мышей (см. фиг.6з).
Кроме того, наблюдали, что трансгенные мыши имеют печень, в которой отложилось большее количество жиров по сравнению с контрольными мышами. Другое значительное увеличение у трансгенных мышей по сравнению с контрольными обнаружили в размере жировых клеток в жировом теле придатка яичка (см. фиг.7а и 7б).
Как объясняется выше, прирост массы трансгенных мышей, которые проявляют более активную экспрессию гена IDPc, объясняется повышением количества жира в организме, и различные показательные для ожирения гены более активно экспрессируются в жировых тканях трансгенных мышей, чем в таковых у контрольных мышей. Например, наблюдалось значительное повышение уровня PPARγ, фактора транскрипции, активирующего транскрипцию генов, которые кодируют ферменты, ответственные за биосинтез липидов. Следовательно, повышение экспрессии гена IDPc сначала приводит к продуцированию больших количеств НАДФН, который необходим для биосинтеза жирных кислот, и, таким образом, дает возможность этому избытку липидных производных индуцировать активацию и экспрессию гена PPARγ, который, в свою очередь, активирует экспрессию генов, показательных для ожирения, и, в конце концов, вызывает ожирение у трансгенных по IDPc мышей. Между тем, повышение экспрессии гена и активности IDPc, а также уровня НАДФН в клетках увеличивает активность ферментов, которые вовлечены в биосинтез холестерина, а также ферментов, активирующих биосинтез липопротеинов через повышение уровней липидов, с получением больших количеств соединений холестерина, в которых холестерин связан с липопротеинами.
В следующем аспекте настоящее изобретение относится к способу отбора веществ, ингибирующих ферментативную активность и экспрессию гена изоцитратдегидрогеназы, и, таким образом, эффективных для лечения болезней обмена веществ, таких как ожирение, гиперлипидемия и жировая инфильтрация печени.
На основании ферментативных функций, раскрытых выше, в настоящем изобретении предложено лечение болезней обмена веществ, вызванных повышением уровней жира in vivo, таких как ожирение, гиперлипидемия и жировая инфильтрация печени, при котором используют тот факт, что повышение ферментативной активности и экспрессии гена изоцитратдегидрогеназы способствует биосинтезу НАДФН, активирующего в свою очередь PPARγ и повышающего in vivo уровни жирных кислот, сквалена и холестерина.
В этой связи весьма полезна спектрофотометрия. Более подробно, сначала спектрофотометр устанавливают на нулевую оптическую плотность при 340 нм, используя смесь реакционного буфера десятикратной концентрации и трижды дистиллированной воды. После этого прозрачную кювету, содержащую 10х буфер, тестируемый образец и трижды дистиллированную воду, устанавливают в спектрофотометр, добавляют в эту кювету изоцитратдегидрогеназу и проводят измерение изменения оптической плотности при 340 нм во времени.
Поскольку оптическая плотность быстрее снижается в кювете, содержащей более высокую концентрацию образца, ингибирующего активность фермента, анализ спектрофотометрических данных позволяет производить отбор ингибиторов изоцитратдегидрогеназы.
Пять образцов, то есть никотиновую кислоту, никотинамид, бупропион, метилизолимонную кислоту и щавелево-яблочную кислоту, тестировали на ингибиторную активность в отношении изоцитратдегидрогеназы. В первых двух образцах не обнаружили ингибиторной активности, тогда как бупропион проявлял небольшой ингибиторный эффект. Напротив, установили, что метилизолимонная кислота и щавелево-яблочная кислота обладают явной ингибиторной активностью в отношении активности изоцитратдегидрогеназы.
В следующем аспекте настоящее изобретение относится к применению НАДФН в стимуляции биосинтеза липидов, холестерина и сквалена и активации PPARγ, учитывающему то первое открытие согласно настоящему изобретению, что искусственное повышение уровня НАДФН в клетках вызывает значительное ожирение и гиперлипидемию и повышает уровень триглицеридов в клетках.
ПРИМЕРЫ
Лучшего понимания настоящего изобретения можно достичь в свете следующих примеров, которые приведены для иллюстрации, но не предназначены для ограничения настоящего изобретения.
ПРИМЕР 1: Выделение и секвенирование гена IDPc
1-1. Выделение кДНК мышиной IDPc
Зонд для идентификации кДНК IDPc мыши получили, используя кДНК IDPc крысы, о которой недавно сообщали (Jennings et al., J. Biol. Chem., 169, 21328-23134, 1994). Смысловой праймер, последовательность №1 в перечне последовательностей, синтезировали на основе 532-550 нуклеотидов гена IDPc крысы, а антисмысловой праймер, последовательность №2 в перечне последовательностей, - на основании нуклеотидов 1263-1245. Отдельно мРНК, выделенную из печени крысы, превращали в кДНК с помощью обратной транскриптазы. Используя праймеры, ПЦР начинали с предварительной денатурации при 94°С в течение 4 мин и проводили 25 циклов денатурации при 94°С по 1 мин, отжига при 50°С в течение 1 мин и удлинения при 72°С в течение 2 мин, за которым следовало удлинение при 72°С в течение дополнительных 10 мин, причем в качестве матрицы использовали 100 нг библиотеки кДНК. В результате амплифицировали последовательность ДНК 0,8 т.п.н. Этот продукт ПЦР клонировали в pCR II (Invitrogen Co.). Вставки клонов секвенировали для идентификации гена IDPc крысы.
Молекулярное клонирование кДНК IDPc мыши начинали с гибридизации в бляшках библиотеки кДНК мышиных клеток NIH3T3 (Stratagene) с геном IDPc крысы, меченым [α-32P]dCTP-, в качестве зонда. Все процессы гибридизации и отмывки проводили при 65°С. Для первичного скрининга фаг библиотеки кДНК при 5×104 БОЕ (бляшкообразующих единицах) смешивали с 3×108 клеток Е.coli XL1-blue при 37°С в течение 15 мин. После того, как эту смесь фага и хозяина хорошо перемешали с 7 мл мягкой агарозной среды (0,7% агароза, 1% триптон, 0,5% дрожжевой экстракт, 1% NaCl), ее наливали в чашку на 150 мм с агарозой TYM-Ap (1% триптон, 0,5% дрожжевой экстракт, 1% NaCl, 1,5% агар, 10 мМ MgSO4, ампициллин 50 мкг/мл), давали отвердеть и инкубировали при 37°С в течение 12 часов. После образования зон фаговых бляшек чашку хранили при 4°С в течение 1 часа, а затем фаги переносили на нитроцеллюлозную мембрану. Для использования при гибридизации в бляшках нитроцеллюлозная мембрана была последовательно намочена в дистиллированной воде и 1М NaCl и высушена на фильтровальной бумаге 3 MM. Чашку с покрытым фагом мягким агаром накрывали этой нитроцеллюлозной мембраной, а затем еще одной - для дупликации. Эти мембраны-дупликаты погружали в денатурирующий буфер (0,5 М NaOH, 0,5 М NaCl) на 5 мин для денатурации фагов из лизата клеток хозяина, и фаговую ДНК и помещали в нейтрализующий буфер (0,5 М Трис-Cl, рН 8,0, 0,5 М NaCl) на 5 мин, за чем, наконец, следовало высушивание на фильтровальной бумаге 3 MM.
Запеканием при 80°С в течение 2 часов иммобилизировали фаговую ДНК на мембране, которую затем промывали 6×SSC (1×SSC; 0,15 М NaCl, 0,015 М цитрат натрия, рН 7,0), содержащим 1% додецилсульфат натрия (ДСН), с последующей предварительной гибридизацией в течение 2 часов при 65°С. Используя меченый [α-32P]dCTP-ген IDPc крысы в качестве зонда, гибридизацию проводили в растворе 6хSSC, содержащем 5× раствор Денхардта (0,1% фиколл, 0,1% поливинилпирролидон, 0,1% БСА), 100 мкг/мл ДНК спермы лосося и 0,01% ДСН. После завершения гибридизации мембрану держали в непосредственном контакте с рентгеновской пленкой в течение 12 часов. Полученная в результате авторадиограмма дала возможность выделения отдельных бляшек.
Шесть клонов ДНК, выделенных из независимых фагов, подвергали рестрикции EcoR1 и Саузерн-блоттингу с использованием меченого [α-32P]dCTP-гена IDPc крысы в качестве зонда. Идентифицировали, что эти 6 фаговых клонов имели фрагменты с размерами в диапазоне от 1,9 до 2,2 т.п.н. Из них отобрали самый большой фрагмент кДНК для выделения фрагмента кДНК IDPc мыши, после чего его субклонировали в векторе pGEM7 (+) (Promega).
1-2: Секвенирование кДНК IDPc мыши
кДНК-ген IDPc мыши, выделенный в примере 1-1, анализировали на последовательность оснований с помощью набора Sequenase, версия 2.0 (United States Biochemicals). На основании полученной последовательности оснований определили аминокислотную последовательность. Для поиска похожих аминокислотных последовательностей и сравнения на предмет гомологии использовали базу данных GeneBank.
На основании данных анализа последовательности оснований ДНК обнаружили, что кДНК IDPc мыши имеет ОРС (открытую рамку считывания) размером 1,245 п.н., последовательность №3 в перечне последовательностей, с последовательностью ААТААА, которую считают сигналом поли(А)+, находящейся в 3'-НТР (нетранслируемом районе). Аминокислотная последовательность, выведенная из последовательности оснований гена IDPc, описана как состоящая из 414 остатков и имеющая молекулярную массу 46,515 Да, как показано в SEQ No.4.
Сравнительный анализ IDPc мыши и белков-изоцитратдегидрогеназ, имеющих происхождение из других видов, показывает, что IDPc мыши по настоящему изобретению имеет гомологию 97,8% с IDPc крысы, 68,5% с бычьей IDPm и 64,4% с дрожжевой IDPc.
Обнаружено, что в аминокислотной последовательности IDPc мыши последовательность 412-414 нт (нуклеотиды) идентична последовательности-мишени пероксисом. Известно, что пероксисома вовлечена в биосинтез и расщепление жирных кислот и холестерина. Эти результаты позволяют предположить высокую вероятность того, что IDPc перемещается к пероксисомам и там принимает участие в синтезе жирных кислот и холестерина.
ПРИМЕР 2: Конструирование клеточных линий, трансформированных генами IDPc
2-1: Конструирование рекомбинантного ретровирусного вектора для экспрессии генов IDPc
Для экспрессии гена IDPc в клетках полученную в примере 1 кДНК IDPc субклонировали в ретровирусном векторе pLNCX (Miller, A.D. and Rosman, G.Т., Biotechniques, 7, 980-990, 1989) в смысловой и антисмысловой ориентации.
После того, как кДНК IDPc подвергали рестрикции по обоим концам с помощью Clal, рестрикт ДНК лигировали в ретровирусный вектор. Эту рекомбинантную плазмиду вводили в Е.coli DH5α и амплифицировали путем культивирования этого микроорганизма. С помощью гидролитического разложения ферментами рестрикции идентифицировали ориентацию вставок в клонах. В полученных рекомбинантных векторных конструкциях экспрессия смысловой или антисмысловой кДНК IDPc направлялась промотором цитомегаловируса, как показано на фиг.1. Рекомбинантные векторы, в которых кДНК IDPc была встроена в смысловых направлениях, как определили с помощью гидролитического разложения ферментами рестрикции, использовали для усиления экспрессии интересующего гена, тогда как рекомбинантный вектор, в котором кДНК IDPc находилась в антисмысловом направлении, использовали для ограничения экспрессии этого гена. В качестве контроля использовали ретровирусный вектор pLNCX, в котором не был закреплен такой ген. Рекомбинантные векторы, полученные таким образом, вводили в клеточную линию мышей NIH3T3 L1 - предшественников жировых клеток. В качестве маркера для идентификации этой трансфекции в эту клетку также одновременно вводили вектор pLNCX, содержащий кДНК GFP (зеленого флуоресцентного белка, green fluorescent protein).
2-2: Конструирование клеточных линий-трансфектантов
Трансфекцию рекомбинантных векторов в клетки NIH3T3 L1 осуществляли с помощью ретровирусных систем упаковки, используя клетки BOSC23. В связи с этим клетки BOSC23 инокулировали при плотности 2×106 клеток/мл в DMEM (среда Игла, модифицированная Дульбекко) с добавкой 10% ФБС (фетальная бычья сыворотка), а затем поддерживали в DMEM, содержащей 25 мкМ хлорохин и 10% ФБС, перед использованием в трансфекции. Трансфекцию осуществляли кальций-фосфатным методом (Pear, W.S. et al., Proc. Natl. Acad. Sci. USA, 90, 8392-8396, 1993). После смешивания с 2×HBS (20 мМ NaCl, 1,5 мМ Na2HPO4, 50 мМ HEPES, pH 7,1) раствор, содержащий 10 мкг рекомбинантной векторной ДНК и 0,25 М CaCl2, равномерно добавляли в чашку, на которой росли клетки BOSC23, и инкубировали в CO2-инкубаторе. После 10 часов инкубации клеткам обеспечивали свежую DMEM с добавкой 10% ФБС и культивировали в течение 24 часов. Только среду центрифугировали при 1200 об/мин, и супернатант фильтровали через 45 мкм фильтр. К фильтрату, который содержал только рекомбинантный ретровирусный вектор, добавляли полибрен (Sigma) до концентрации 4 мкг/мл.
При подготовке к трансфекции ретровирусным рекомбинантным вектором клетки NIH3T3 L1 инокулировали при плотности 5×105 клеток/мл и культивировали в DMEM с добавкой 10% ФБС. Когда число клеток увеличивалось на 50%, среду удаляли и ретровирусные частицы, отделенные от упаковывающих клеток, добавляли к NIH3T3 L1. После 5 часов инкубации к клеткам добавляли свежую среду и культивировали в течение 2 суток.
Подсчитывали количество клеток NIH3T3 L1, инфицированных рекомбинантными ретровирионами, с последующим распределением этих клеток на аликвоты с плотностью 50 клеток на лунку в 96-луночных планшетах, которые содержали DMEM с добавленным G-418 (Gibco BRL), концентрация 400 мкг/мл. В то время как среду с G-418 меняли через сутки, клетки NIH3T3 L1 в каждой лунке повторно разделяли и культивировали для отбора первых трансформантов NIH3T3 L1. Для вторичного скрининга проводили ПЦР геномной ДНК, чтобы подтвердить встраивание кДНК IDPc в геном.
Среди клеток, которые в конце концов идентифицировали как содержащие ген IDPc в своем геноме, трансформанты, в которых ген IDPc был встроен в смысловом направлении и в антисмысловом направлении, обозначили соответственно как FS1 и FAS1. Клеточная линия FS1, в геноме которой ген IDPc закреплен в смысловом направлении, депонирована Корейской Коллекцией Типовых Культур (КСТС) Корейского Научно-исследовательского института Биологических наук и Биотехнологии (KRIBB) под номером КСТС 0861 ВР 6 сентября 2000 г.
ПРИМЕР 3: Ферментативная активность IDPc и IDPm в клетках-трансфектантах
Для определения ферментативной активности IDPc в клетках-трансфектантах NIH3T3 L1 цитоплазму отделяли от клеток и определяли концентрацию белка, используя анализ по Брэдфорду. Сначала 3×107 клеток/мл дважды промывали 1×ФСБ (фосфатно-солевой буферный раствор) и подвергали лизису сахарозным буфером (0,32 М сахароза, 0,01 М Трис-Cl, рН 7,4). Клеточный лизат центрифугировали при 1000×g для удаления клеточного дебриса, а затем при 15000×g для осаждения митохондрий. К отделенному супернатанту, содержащему цитоплазматическую фракцию, добавляли ФСБ, содержащий 0,1% Тритон Х-100, в количестве 1/10 суммарного объема раствора с последующим количественным определением с помощью анализа по Брэдфорду. Ферментативную активность IDPc определяли путем измерения изменения количества продуцируемого НАДФН в буфере (50 мМ MOPS, рН 7,2, 35,5 мМ триэтаноламин, рН 7,2, 2 мМ НАДФ+, 2 мМ MgCl2, 5 мМ изоцитрат и ротенон 1 мкг/мл), поддерживаемом при 25°С. Используя спектрофотометр, измеряли оптическую плотность при 340 нм в течение 2 мин, чтобы определить количество НАДФН, продуцируемого IDPc, содержащейся в цитоплазматическом белке, и таким образом определить ферментативную активность IDPc. Для количественного определения этого фермента количество, которое может продуцировать 1 мкМ НАДФН за 1 мин, считали как 1 единицу.
По сравнению с контрольными клетками, в которые вводили только вектор LNCX, в клетках-трансфектантах FS1, в которых IDPc был введен в смысловом направлении, ферментативная активность IDPc была повышена примерно в 2 раза, тогда как она была снижена примерно в 0,4 раза в клетках-трансфектантах FAS1, в которых ген IDPc был встроен в антисмысловом направлении.
ПРИМЕР 4: Синтез жирной кислоты в линии клеток-трансфектантов в соответствии с активностью IDPc
Чтобы исследовать влияние IDPc на синтез жирных кислот, линии клеток-трансфектантов культивировали в DMEM, обогащенной 10% ФБС и содержащей 5 мкг/мл инсулина, 0,5 мМ 3-изобутил-1-метилксантин (IBMX, Sigma), 1 мкМ дексаметазон (DEX) и пенициллин-стрептомицин (Gibco BRL), каждого по 50000 единиц, в течение двух суток для увеличения количества клеток до плотности примерно 3×104 клеток/см2. После этого клетки дополнительно культивировали в течение 12 суток в DMEM, не содержащей IBMX и DEX, причем среду обновляли через сутки. Культивирование клеток осуществляли в CO2-инкубаторе при 37°С во влажной атмосфере и 5% CO2.
После культивирования клетки обрабатывали Oil-Red-O, который специфично окрашивает жир, чтобы наблюдать жировые отложения, образовавшиеся в жировых клетках. В связи с этим среду истощали, после чего к клеткам добавляли 10 мл какодилатного буфера (90 мМ какодилат, рН 7,2, 2% формальдегид, 2,5% глутаральдегид, 0,025% CaCl2, 5% сахароза), которым затем давали стоять при 4°С в течение 1 часа. После удаления буфера к клеткам добавляли 5 мл Oil-Red-O в 40% изопропаноле и медленно перемешивали в течение 1 часа с последующей промывкой 40% изопропанолом.
Со ссылкой на фиг.2а представлены наблюдения жировых клеток, окрашенных Oil-Red-O. Как показано на фиг.2, клетки-трансфектанты FS1, в которых экспрессия гена IDPc повышена, продуцировали жиры в большем количестве, чем контрольные клетки. С другой стороны, клетки-трансфектанты FAS1 со сниженной экспрессией гена IDPc показали значительно уменьшенные жировые отложения относительно контрольных клеток. Фотографии, сделанные при 200х увеличении, дополнительно показывают различия в размере жировых клеток среди этих групп клеток. Эти наблюдения показывают, что увеличение или снижение экспрессии гена и уровня IDPc, а также уровня НАДФН, метаболического продукта этого фермента, оказывают значительное влияние на увеличение или снижение клеточных жировых отложений.
ПРИМЕР 5: Изменение дифференциации жировых клеток и отложения жира внутри клеток в соответствии с концентрацией НАДФН
Действие НАДФН, метаболического продукта IDPc, на отложение клеточных жиров, количественно определяли с помощью Oil-Red-O, красителя, специфичного в отношении липидов. При культивировании контрольных клеток NIH3T3 L1, в которые был введен только вектор pLNCX, в тех же условиях, что и в примере 4, добавляли НАДФН в количествах 0 мкМ, 25 мкМ и 50 мкМ в среду для дифференциации этих клеток в жировые клетки. После дифференциации клетки окрашивали раствором Oil-Red-O для визуализации жировых отложений, образовавшихся внутри клеток. Визуализированные результаты показаны на фиг.2б. Как видно на фиг.2б, клетки аккумулировали большие количества жиров в присутствии внешнего НАДФН, чем в отсутствие внешнего НАДФН. Кроме того, более обширное отложение жиров наблюдали при подаче внешнего НАДФН в больших количествах. Следовательно, установили, что НАДФН, который может быть получен как продукт реакции с участием не только изоцитратных изоферментов IDPc и IDPm, но также и глюкозо-6-фосфатдегидрогеназы, 6-фосфоглюконатдегидрогеназы и малатдегидрогеназы, оказывает прямое положительное влияние на дифференциацию жировых клеток и сопутствующее отложение жиров, даже когда его добавляют искусственно.
ПРИМЕР 6: Идентификация активности IDPc in vivo с использованием трансгенного животного, содержащего ген IDPc в своем геноме
6-1: Создание трансгенной мыши
6-1-1: Получение слитой генной конструкции для микроинъекции
Для использования в тканеспецифичной постоянной экспрессии гена IDPc в печени и в жировых клетках последовательность 2,2 т.п.н. в обратном направлении от 5'-конца, содержащую промотор, из гена цитозольного РЕРСК крыс амплифицировали с помощью ДНК-полимеразы PfuTurbo (Stratagene), используя пару-праймеров - последовательности №№5 и 6 в перечне последовательностей. Продукт ПЦР подвергали ферментативному гидролизу Bg/II и Smal, а затем I-Pop-I, и обрабатывали нуклеазой фасоли золотистой для получения тупых концов, один из которых подвергали рестрикции Bg/II. Этот продукт ферментативного гидролиза ДНК встраивали в вектор экспрессии млекопитающих pCl-neo, содержащий промотор CMV. С этим вектором лигировали кДНК IDPc, которая была дважды подвергнута гидролизу Xhol и Sa/I. Полученный в результате рекомбинантный вектор, в котором ген IDPc экспрессировался под контролем последовательности РЕРСК, находящейся в обратном направлении от 5'-конца, обозначили pPEPCKIDPc. Процесс рекомбинации проиллюстрирован на фиг.3.
6-1-2: Получение слитой генной конструкции для микроинъекции
Слитую генную конструкцию, содержащую IDPc мыши (примерно 10 мкг), полученную в примере 6-1-1, подвергали двойному ферментативному гидролизу ферментами рестрикции Bg/II и NsiI, после чего раствор гидролизата разделяли на 0,7% агарозном геле, чтобы выделить фрагмент ДНК 4,9 т.п.н., содержащий интересующий ген. Вырезанную часть геля, содержащую этот фрагмент ДНК, обрабатывали смесью фенола и CIAA (хлороформ: изоамиловый спирт = 24:1 об/об) в объемном соотношении 1:1. Верхнюю часть центрифугировали при 12000 об./мин в течение 3 мин. К полученному супернатанту добавляли равный объем диэтилового эфира с последующим центрифугированием при 10000 об./мин в течение 5 сек. После удаления верхней эфирной части к нижней части, содержащей ДНК, добавляли два объема абсолютного этанола для осаждения ДНК. Осадок ДНК хорошо тщательно разводили в растворе для микроинъекции (10 мМ Трис рН 7,4, 0,1 мМ ЭДТА) в концентрации 2-10 мкг/2,4 мл. Полученный в результате раствор подвергали диализу против раствора для микроинъекции при 4°С в течение 24 часов. Для микроинъекции контролировали, чтобы ДНК находилась в концентрации 2-4 нг/мкл в сумме и хранилась при -20°С до использования.
6-1-3: Получение эмбриона
Потомкам мышей FVB/N, которые продуцируют много легко собираемых яйцеклеток и эмбрионы которых претерпевают небольшие повреждения при микроинъекции, вводили внутрибрюшинно гонадотропин сыворотки жеребой кобылы (PMSG, pregnant mare's serum gonadotropin) и человеческий хорионгонадотропин (hCG, human chorionic gonadotropin) в дозе 5 ME, каждый, для индукции суперовуляции. Через 20 часов после инъекции PMSG и hCG ампулу маточной трубы мыши разрывали, чтобы выделить скопление клеточной массы, которую затем освобождали от скоплений клеток обработкой гиалуронидазой (300 мкг/мл) в течение 3 мин. Из них отбирали эмбрионы на одноклеточной стадии, на которой наблюдали два эукариона на клетку, для использования в микроинъекции.
При мониторинге ядерной мембраны под инвертированным микроскопом, снабженным контрастными линзами дифференциальной интерференции (DIC) Normarski, проводили микроинъекцию слитой генной конструкции в отобранные эмбрионы с помощью микроманипулятора. После завершения микроинъекции выжившие эмбрионы культивировали в среде М16 при 37°С в атмосфере 5% CO2 в СО2-инкубаторе. Полученный в результате эмбрион мыши, несущий слитую генную конструкцию, содержащую ген IDPc, депонирован в Корейской Коллекции Типовых Культур Корейского Научно-исследовательского Института Биологических наук и Биотехнологии (KRIBB) под номером КСТС 0874ВР 5 ноября 2000 г.
6-1-4: Имплантация эмбриона мыши и идентификация трансгенных мышей
В качестве реципиентов использовали потомство мышей FVB/N благодаря большой площади их матки и высокой способности к размножению и вскармливанию. Вазэктомию проводили самцам мышей, которым давали возможность спариваться с самками на стадии течки. На следующее утро самок мышей, у которых наблюдали влагалищные пробки, отбирали в качестве окончательных реципиентов. Используя ножницы, производили надрез подкожного слоя около маточной трубы реципиентов длиной 1 см, а затем делали надрез в мышечном слое с последующей имплантацией эмбриона из примера 6-1-3 в лежащую напротив маточную трубу, обнаженную таким образом.
Потомство от этих реципиентов исследовали с помощью ПЦР на вставку микроинъецированной слитой генной конструкции ДНК IDPc мыши, то есть pPEPCKIDPc, в их геном. С этой целью у мышей 2- или 3-недельного возраста, рожденных реципиентом, отрезали кусочек хвоста длиной 2-3 см и погружали в 700 мкл буфера для лизиса (50 мМ Трис-Cl, рН 8,0, 100 мМ ЭДТА, 100 мМ NaCl, 1% ДСН) на 15-18 часов при 55°С в присутствии 35 мкл протеиназы К (10 мг/мл) при перемешивании. После гидролиза РНК 20 мкл РНКазы (13 мкг/мл) к этому гидролизованному раствору добавляли равный объем фенола, а затем центрифугировали. Фенольную экстракцию повторяли на супернатанте еще два или три раза. К конечному супернатанту добавляли два объема абсолютного этанола для осаждения ДНК. Полученную таким образом мышиную геномную ДНК (1 мкг) в качестве матрицы частично амплифицировали в ПЦР, используя 3'-фланкирующий район промотора CMV в качестве смыслового праймера Р1 (последовательность №7) и 5'-фланкирующий район гена IDPc в качестве антисмыслового праймера Р2 (последовательность №8). ПЦР начинали с денатурации при 95°С в течение 5 мин с последующими 30 циклами денатурации при 95°С в течение 1 мин, отжига при 51°С в течение 1 мин и удлинения при 72°С в течение 1,5 мин. Раствор ПЦР распределяли на 1,5% агарозном геле для отбора мыши, у которой представлена полоса ДНК 0,5 т.п.н., позволяющая идентифицировать ее как трансгенную мышь. У потомства от скрещивания отобранных трансгенных самцов мышей с самками мышей FVB/N дикого типа отрезали хвосты и анализировали на трансгенность тем же способом для отбора трансгенных мышей, которые унаследовали рекомбинантный ген IDPc в зародышевой линии. После того, как потомство трансгенной мыши идентифицировали с помощью ПЦР как имеющее рекомбинантный ген, это трансгенное потомство поддерживали в гетерозиготной линии F2. Обнаружили, что, подобно гетерозиготным трансгенным мышам F1, у этих гетерозиготных трансгенных мышей F2 проявлялись ожирение, гиперлипидемия и жировая инфильтрация печени.
Для управлениями этим видом трансгенных мышей у мышей 2-недельного возраста частично обрезали хвосты, получали геномную ДНК из этих сегментов хвостов и анализировали ее на вставку экзогенного интересующего гена. Как только эти мыши были идентифицированы как трансгенные, их разделяли по полу и маркировали по ушам.
6-2: Прирост массы жировой ткани и активности IDPc у трансгенного животного
6-2-1: Увеличение жирового тела придатка яичка у трансгенного животного
Трансгенных мышей и нормальных мышей, и те и другие были в возрасте 28 недель и потомством одних и тех же родителей, умерщвляли путем отделения их позвоночного столба, после чего их экзодермис частично иссекали ножницами, придерживая пинцетом. После полного отшелушивания экзодермиса иссекали эндодермис, чтобы обнажить жировые тела придатков яичек, которые затем сравнивали по размеру с жировыми телами придатков яичек у мышей FVB/N дикого типа. После этого суммарное количество жировой ткани брали как от трансгенных мышей, так и от мышей дикого типа, чтобы сравнить суммарные массы между ними. Вскоре после измерения массы отделенные суммарные жировые ткани фиксировали в формалине и быстро охлаждали. Используя микротом, эти замороженные жировые ткани нарезали при -20°С на ломтики, кусочки толщиной 10 мкм, которые окрашивали гематоксилином и эозином для визуализации под микроскопом.
Результаты наблюдения под микроскопом представлены на фиг.4. Через 26 недель после рождения, как видно на фиг.4а, гетерозиготные трансгенные мыши F1 вырастали больше, чем контрольные мыши. После снятия кожи наблюдали, что трансгенные мыши имели большие кожные тучные клетки в большем количестве на своих спинах по сравнению с контрольными мышами, как показано на фиг.4б. Кроме того, определяли, что у трансгенных мышей были значительно большие жировые тела придатков яичек. При дальнейшем анатомировании установили, что у гетерозиготных трансгенных мышей F1 были значительно большие жировые тела придатков яичек по сравнению с контрольными мышами, как показано на фиг.4в. После полного удаления кожи живота наблюдали значительное увеличение массы белой жировой ткани у трансгенных мышей по сравнению с контрольными мышами, как видно на фиг.4г. Между трансгенными мышами и нормальными мышами невооруженным глазом не было заметно различий в размере и цвете печени.
6-2-2: Изменение уровня экспрессии гена, показательного для ожирения, в соответствии с экспрессией гена IDPc у трансгенных мышей
Чтобы исследовать, оказывает ли экспрессия гена IDPc влияние на экспрессию генов, показательных для ожирения, каждый показательный для ожирения ген количественно измеряли в отношении уровня его экспрессии следующим образом.
Подробно, 1 г жирового тела придатка яичка, взятого от каждой из трансгенных по IDPc и нормальных мышей, добавляли в 9 мл лизогенного раствора (4 М гуанидинтиоцианат, 25 мМ цитрат натрия, рН 7,0, 0,5% саркозил, 0,72% (3-меркаптоэтанол) и гомогенизировали, используя гомогенизатор. После охлаждения в течение 2-3 мин во льду к гомогенату добавляли смесь 1 мл раствора для экстракции (2 М ацетат натрия, рН 4,0), 10 мл насыщенного DEPC-обработанной водой фенола, 2 мл смеси хлороформ - изоамиловый спирт (24:1) и оставляли стоять на льду в течение 15 мин. После того, как этот раствор центрифугировали при 3000×g при 4°С в течение 15 мин, супернатант снова экстрагировали смесью фенол/хлороформ. К экстракту добавляли равный объем изопропанола с последующим центрифугированием в течение 15 мин с получением осадка РНК. После растворения в 36% водном растворе формальдегида РНК разделяли на 1% агарозном геле, содержащем формальдегид в количестве 6,7%, в электрическом поле. После электрофореза разделенные полосы РНК переносили на нейлоновую мембрану в 20× растворе SSC, высушивали и фиксировали поперечным связыванием УФ светом. Нейлоновую мембрану промывали в течение 5 мин в 6× растворе SSC, а затем подвергали предварительной гибридизации при 42°С в течение 2 часов в подходящем количестве гибридизационного раствора (50% формамид, 6× SSC, 5× раствор Денхардта, 1,2% ДСН, 10 мкг/мл ДНК спермы лосося). Чтобы использовать в качестве зондов для Нозерн-блоттинга, различные кДНК, показательные для ожирения (аР2, адипсина, LPL (липопротеинлипазы), лептина, фактора некроза опухоли α [TNFα] и PPARγ) метили [α-32P]dCTP. Используя эти радиоактивно меченые зонды, проводили гибридизацию в течение 12 часов. После завершения гибридизации нейлоновую мембрану промывали при 65°С 6× раствором SSC, содержащим 0,1% ДСН, в течение 30 мин, а затем 2× SSC, содержащим 0,1% ДСН, в течение 20 мин и дополнительно промывали таким же образом, как указано выше, по меньшей мере еще один раз. Наконец нейлоновую мембрану дважды промывали 0,2× раствором SSC при комнатной температуре. Авторадиограмма, полученная экспонированием мембраны с рентгеновской пленкой при -70°С, дала возможность идентификации мРНК, транскрибируемых с генов, показательных для ожирения, в жировом теле придатка яичка.
На фиг.5 результаты гибридизации показаны на авторадиографах. Как видно на авторадиографах, обнаружена экспрессия рекомбинантного гена IDPc, введенного в трансгенных мышей, параллельно с экспрессией эндогенного гена IDPc, что показывает, что суммарная активность IDPc была повышенной. Кроме того, обнаружили увеличение экспрессии генов, показательных для ожирения, таких как гены, кодирующие аР2, адипсин, LPL, лептин, TNF-α и активируемый пролифератором пероксисом рецептор γ (PPARγ), о которых известно, что все они проявляют повышенную экспрессию, при этом способствуя дифференциации тучных клеток. Таким образом, показано, что повышенная активность IDPc в результате повышения экспрессии гена IDPc стимулирует экспрессию всех генов, показательных для ожирения. Как продемонстрировано на фиг.2б, эти результаты указывают на то, что НАДФН вовлечен в экспрессию генов, показательных для ожирения.
В свете недавних сообщений, описывающих механизм положительной обратной связи PPARγ, при котором активация PPARγ стимулирует далее экспрессию его гена (Kim, J.В. et al., Proc. Natl. Acad. Sci. USA, 95, 4333-4337, 1998), а также сообщений, касающихся функции производных полиненасыщенных жирных кислот и липидов в качестве лигандов для активации PPARγ, вышеуказанные результаты можно интерпретировать как означающие, что повышенный клеточный уровень НАДФН, связанный с повышением клеточной активности IDPc, стимулирует активность синтаз жирных кислот, которым обязательно необходим НАДФН для их ферментативных реакций, повышая таким образом уровень жирных кислот в клетках. Кроме того, в свете сообщений, показывающих, что повышенные в результате достаточного обеспечения НАДФН клеточные уровни производных жирных кислот приводят к активации PPARγ, который служит в качестве основного фактора транскрипции для стимуляции не только экспрессии генов, ответственных за синтез жирных кислот, необходимых для дифференциации тучных клеток, но также и генов, кодирующих белки, вовлеченные в биосинтез холестерина, повышение экспрессии генов, показательных для ожирения, таких как гены, кодирующие ар2, адипсин, LPL, лептин и TNFα, считают следствием повышенной экспрессии гена PPARγ. Следовательно, можно сделать вывод, что повышение активности IDPc вследствие активной экспрессии гена IDPc и сопутствующее повышение уровня НАДФН направлены на резкое повышение экспрессии гена PPARγ, который необходим как для дифференциации жировых клеток, так и для биосинтеза липидов.
6-2-3: Прирост массы и повышение внутриклеточного уровня липидов и холестерина в соответствии с экспрессией IDPc у трансгенного животного и его биохимическое исследование
Через 26 недель после рождения мышей F1, трансгенных по ожирению, и нормальных мышей сравнивали на внутриклеточное отложение липидов.
1. Измерение массы тела
Контейнер, подходящий для мыши, ставили на весы, которые затем устанавливали на ноль, после чего мышь осторожно помещали в этот контейнер. Поскольку числовые показания весов менялись, когда мышь двигалась, за массу тела этой мыши брали значение, определенное, когда мышь не двигалась.
2. Определение ферментативной активности IDPc
Печени и жировые ткани, взятые от трансгенных мышей и нормальных мышей, гомогенизировали в буфере (0,32 М сахароза, 0,01 М Трис-Claim, рН 7,4) и центрифугировали при 3000×g в течение 15 мин. Затем супернатант повторно центрифугировали при 10000×g в течение 15 мин. В виде супернатанта получили чистую цитозольную фракцию. Ферментативную активность IDPc определяли с помощью измерения изменения количества продуцированного НАДФН в буфере (50 мМ MOPS (3-(N-морфолино)пропансульфоновая кислота), рН 7,2, 35,5 мМ триэтаноламин, рН 7,2, 2 мМ НАДФ+, 2 мМ MgCl2, 5 мМ изоцитрат и 1 мкг/мл ротенон), поддерживаемом при 25°С. Используя спектрофотометр, измеряли оптическую плотность при 340 нм в течение 2 мин, чтобы определить количество НАДФН, продуцированного IDPc, содержащейся в цитоплазматическом белке, определяя таким образом ферментативную активность IDPc. Для количественного определения этого фермента количество, которое продуцировало 1 мкМ НАДФН за 1 мин, брали за 1 единицу.
3. Измерение [НАДФ]/[НАДФН+НАДФ+]
Количественное определение НАДФН основано на том принципе, что НАДФН взаимодействует с МТТ (3-[4,5-диметилтиазол-2-ил]-2,5-дифенилтетразолий) с сопутствующим изменением цвета. Готовили два цитозольных экстракта, содержащих 100 мкг белков: один был предварительно обработан реакцией при 60°С в течение 30 мин и охлаждением до 0°С, чтобы разложить весь НАДФ+ для измерения количества раннее существующего НАДФН, [НАДФН] (образец 1); а второй хранили при 0°С без предварительной обработки для измерения суммарного пула НАДФ, [НАДФН+НАДФ+] (образец 2). Каждый из этих двух образцов добавляли в реакционный раствор (0,1 М Трис-HCl-буфер, рН 8,0, 5 мМ ЭДТА, 2 мМ феназинэтосульфата, 0,5 мМ МТТ), к которому затем добавляли 1,3 единицы глюкозо-6-фосфатдегидрогеназы и инкубировали при 37°С в течение 5 мин для превращения в НАДФН всего НАДФ+, имеющегося в реакционном растворе. Для этого субстрат для фермента, глюкозо-6-фосфат, добавляли в количестве 1 мМ к каждому образцу (образцы 1 и 2), но не добавляли к контролю. После завершения реакции измеряли изменение оптической плотности при 570 нм после взаимодействия с МТТ для количественного определения НАДФН. Поскольку изменения оптической плотности, обнаруженные в образцах 1 и 2, приписывали [НАДФН] и [НАДФН+НАДФ+] соответственно, можно было определить отношение НАДФН к суммарному пулу НАДФ, т.е. ([НАДФН]/[НАДФ+]+[НАДФН]).
4. Количественное определение уровня триглицеридов и суммарного холестерина в крови
Для измерения уровня триглицеридов и суммарного холестерина в крови использовали аналитические наборы, выпускаемые Asan Pharmaceutics Co. Ltd. Образцы крови, взятые от трансгенных мышей и нормальных мышей, обрабатывали антикоагулянтом и центрифугировали с получением сыворотки. 10 мкл каждой сыворотки смешивали с 1,5 мл набора для анализа триглицеридов [раствор липопротеиназы 10800 ЕД, глицеролкиназы 5,4 ЕД, пероксидазы 135000 ЕД и L-α-глицерофосфатоксидазы 160 ЕД в 72 мл буфера с N,N-бис(2-гидроксиэтил)-2-аминометансульфоновой кислотой] или 1,5 мл набора для ферментативного анализа холестерина (смесь раствора ферментов (холестеролэстеразы 20,5 единиц Кимбрела/л, холестеролоксидазы 10,7 единиц Кимбрела/л и гидроксида натрия 1,81 г/л) и буфера (монофосфат калия 13,6 г/л, фенол 1,88 г/л) в соотношении 1:1) и инкубировали при 37°С в течение 5 мин для реакции. В устройстве для прочтения микропланшетов проводили измерение оптической плотности при 500 нм для холестерина и при 540 нм - для триглицеридов, чтобы количественно определить уровни триглицеридов и суммарного холестерина в крови. В связи с количественным определением триглицеридов и суммарного холестерина использовали стандартную кривую, которую получили с помощью применения вышеописанной методики к различным известным концентрациям стандартного раствора.
5. Количественное определение уровня триглицеридов и суммарного холестерина в печени
От трансгенных мышей и от нормальных мышей брали заданное количество печени и гомогенизировали в 1 мл CIAA (хлороформ : изоамиловый спирт = 2:1 об/об). 100 мкл этого гомогената растворяли в 200 мкл чистого этилового спирта и смешивали с 500 мкл каждого из аналитических наборов для триглицеридов и для холестерина, выпускаемых Asan Pharmaceutics Co. Ltd., содержащих 0,5% Тритон Х-100 и 3 мМ холат натрия, с последующей инкубацией при 37°С в течение 10 мин. После добавления 800 мкл воды каждый образец измеряли на уровни триглицеридов и суммарного холестерина в печени с помощью устройства для прочтения микропланшетов тем же способом, как описано выше. Подобным же образом вышеописанную методику применяли к различным известным концентрациям стандартного раствора для получения стандартной кривой, которую использовали для количественного определения уровней триглицеридов и холестерина в печени.
6. Количественное определение уровня лептина в крови
После коагуляции в течение ночи при 2-8°С кровь, взятую от трансгенных мышей и нормальных мышей, центрифугировали при 53000×g в течение 20 мин, чтобы отделить сыворотку. Эту сыворотку замораживали при -20°С до использования для измерения концентрации лептина. Уровень лептина в крови определяли с помощью набора для ELISA (набор для колориметрического определения мышиного лептина [OB], R&D Systems) согласно инструкциям изготовителя. Сначала 50 мкл сыворотки смешивали с равным объемом смеси 2,5 н. уксусной кислоты/10 М мочевины и полученной смеси давали стоять при комнатной температуре в течение 10 мин. Для нейтрализации к этой смеси добавляли 50 мкл 2,7 н. NaOH/1 M HEPES. Перед измерением концентрации полученную в результате смесь образца разбавляли в соотнощении 1/20 калибровочным разбавителем (RD5-3). В каждую лунку 96-луночных планшетов добавляли 50 мкл аналитического разбавителя (RD1W) с последующим добавлением 50 мкл образца сыворотки или 50 мкл эталонного материала. Полученный в результате раствор в каждой лунке полностью перемешивали в течение 1 мин и инкубировали при комнатной температуре в течение 2 часов для проведения реакции. После полного удаления жидкости каждую лунку промывали 4-5 раз отмывочным буфером. Остаток в каждой лунке подвергали взаимодействию со 100 мкл конъюгата лептина мыши при комнатной температуре в течение 2 часов и снова промывали 4-5 раз. В пределах 30 мин уровни лептина в крови количественно определяли путем измерения оптической плотности при 450 нм в устройстве для прочтения микропланшетов.
Результаты измерений, полученные в примере 6-2-3, анализировали и представили в виде графиков на фиг.6. После достижения 26 недель измерили, что трансгенные (Тг) мыши, как видно на фиг.6а, имели массу на 23-35% больше, чем нормальные мыши (не-Тг). Кроме того, обнаружили, что трансгенные мыши проявляют в 1,4 и 2,7 раз более высокую активность IDPc в печени и жировых тканях, соответственно, чем нормальные мыши, как показано на фиг.6в. На фиг.6д приведено сравнение отношений концентрации НАДФН к суммарному пулу НАДФ, ([НАДФН]/[НАДФ+]+[НАДФН]), в печени и в жировой ткани между трансгенными мышами и нормальными мышами. Отношения концентраций в печени и жировой ткани трансгенных мышей были выше в 1,2 и в 1,3 раза соответственно, чем таковые у нормальных мышей. Отмечено большое увеличение массы жирового тела придатков яичек. Измерили, что трансгенные мыши имеют жировое тело придатков яичек с массой примерно в 13,6 раз больше, чем у нормальных мышей, как показано на фиг.6д. Однако не обнаружено значительного различия в массе печени нормальных мышей и трансгенных мышей, как проиллюстрировано на фиг.6б. Что касается триглицеридов и суммарного холестерина, их уровни в крови были выше у трансгенных мышей в 1,8 и 2,4 раза соответственно по сравнению с таковыми у нормальных мышей, как показано на фиг.6е. Подобным образом, уровни триглицеридов и суммарного холестерина в печени трансгенных мышей были в 3,4 и 2,8 раза выше соответственно таковых параметров у нормальных мышей, как показано на фиг.6ж. На фиг.6з показано, что уровни в крови лептина, белка, продуцируемого, в основном, тучными клетками, у трансгенных мышей выше примерно в 2 раза по сравнению с нормальными мышами.
Кроме того, печень и жировые ткани, взятые от мышей F1, трансгенных по гену ожирения, и нормальных мышей наблюдали под микроскопом. Для удобства наблюдения ткани резали на срезы. Идентифицировали, что печень трансгенных мышей представляла собой печень с жировой инфильтрацией, которая аккумулировала больше жира, чем печень нормальных мышей, как видно на фиг.7а. Наблюдали также, что жировые клетки трансгенных мышей имеют в пять раз больший размер по сравнению с таковыми у нормальных мышей (фиг.7б).
Как описано выше, прирост массы у трансгенных мышей, в которых активно экспрессируется ген IDPc, является результатом накопления жира в организме. Кроме того, уровни в клетках различных белков, показательных для ожирения, включая PPARγ, который представляет собой фактор транскрипции, стимулирующий экспрессию генов, кодирующих ферменты, вовлеченные в метаболизм липидов и холестерина, значительно повышены в жировой ткани трансгенных мышей по сравнению с нормальными мышами. Следовательно, повышение экспрессии гена IDPc в первую очередь приводит к продуцированию больших количеств НАДФН, который необходим для биосинтеза жирных кислот и дает возможность полученному в результате обилию производных липидов индуцировать активацию и генную экспрессию PPARγ, который, в свою очередь, активирует экспрессию генов, показательных для ожирения, и, в конце концов, вызывает ожирение у мышей, трансгенных по IDPc.
ПРИМЕР 7: Отбор ингибиторов изоцитратдегидрогеназы
С целью отбора веществ, способных регулировать активность изоцитратдегидрогеназы, осуществляли следующую методику.
Аналитический буфер (50 мМ MOPS, pH 7,2, 35,5 мМ триэтаноламин, рН 7,2, 2 мМ НАДФ+, 2 мМ MgCl2, 5 мМ изоцитрат, ротенон 1 мкг/мл) и тестируемые образцы - все готовили в 10× концентрации. Ферментативные реакции, необходимые для отбора, проводили в конечном объеме 1 мл при 25°С в кварцевой кювете. Для использования концентрированные тестируемые образцы разбавляли трижды дистиллированной водой. Аналитический буфер, ингибиторы фермента и изоцитратдегидрогеназу, все в 10× концентрации, держали при низкой температуре, то есть во льду.
Спектрофотометр сначала устанавливали на ноль при 340 нм, используя смесь 10× аналитического буфера и трижды дистиллированной воды. В кварцевую кювету добавляли 100 мкл образца вместе со 100 мкл 10х аналитического буфера и смешивали с 600 мкл трижды дистиллированной воды. После завершения установки на ноль кювету с образцом устанавливали в спектрофотометре, а затем добавляли 200 мкл изоцитратдегидрогеназы и перемешивали этот раствор с помощью пипетки. Проводили мониторинг изменения оптической плотности при 340 нм во времени. В принципе, оптическая плотность уменьшается быстрее в кювете, содержащей более высокую концентрацию образца, ингибирующего активность фермента. На основании этого принципа количественное определение по данным спектрофотометрии позволяет производить отбор ингибиторов изоцитратдегидрогеназы.
Проводили исследование ингибиторной активности в отношении изоцитратдегидрогеназы пяти образцов, то есть никотиновой кислоты, никотинамида, бупропиона, метилизолимонной кислоты и щавелево-яблочной кислоты. Обнаружили, что первые два образца не проявляют ингибиторную активность, тогда как у бупропиона обнаружили только небольшую ингибиторную активность. Напротив, установили, что щавелево-яблочная кислота (фиг.8а) и метилизолимонная кислота (фиг.8б) обладают сильной активностью ингибирования изоцитратдегидрогеназы.
ПРИМЕР 8: Влияние ингибитора изоцитратдегидрогеназы на предупреждение ожирения
8-1: Предупреждение отложения жира в 3T3-L1 ингибитором изоцитратдегидрогеназы
После обработки ингибиторами изоцитратдегидрогеназы метилизолимонной кислотой и щавелево-яблочной кислотой клетки 3T3-L1, которые остались недифференцировавшими, исследовали на их дифференциацию в жировые клетки. Влияние ингибиторов изоцитратдегидрогеназы на синтез жирных кислот можно определить путем визуализации, используя Oil-Red-O, специфичный к жирам краситель. Результаты представлены на фиг.9, на которой показаны жировые отложения, окрашенные Oil-Red-O, в клетках, не обработанных ингибиторами изоцитратдегидрогеназы (слева), обработанных щавелево-яблочной кислотой (посередине) и обработанных метилизолимонной кислотой (справа), на фотографиях при 100-кратном увеличении (верхний ряд) и 200-кратном увеличении (нижний ряд). Как видно на этих оптических фотографиях, ингибиторы IDPc играют важную роль в метаболизме липидов in vivo, уменьшая жировые отложения в клетках.
8-2: Ограничивающее действие ингибитора изоцитратдегидрогеназы на ожирение у крыс
Через 26 недель после рождения крыс сравнивали на отложение липидов в клетках, когда их не обрабатывали ингибиторами изоцитратдегидрогеназы и когда их обрабатывали метилизолимонной кислотой.
1. Измерение массы тела
Контейнер, подходящий для крысы, ставили на весы, которые затем устанавливали на ноль, после чего крысу осторожно помещали в этот контейнер. Поскольку числовые показания весов менялись, когда крыса двигалась, за массу тела крысы принимали значение, определенное, когда эта крыса не двигалась.
2. Определение ферментативной активности IDPc
Печени и жировую ткань, взятые от крыс, которым вводили ингибитор, и от крыс, которым не вводили ингибитор, гомогенизировали в буфере (0,32 М сахароза, 0,01 М Трис-Claim, рН 7,4) и центрифугировали при 3000×g в течение 10 мин. Супернатант снова центрифугировали при 10000×g в течение 15 мин. Чистую цитозольную фракцию получили в виде супернатанта. Ферментативную активность IDPc определяли путем определения изменения количества продуцированного НАДФН в буфере (50 мМ MOPS, рН 7,2, 35,5 мМ триэтаноламин, рН 7,2, 2 мМ НАДФ+, 2 мМ MgCl2, 5 мМ изоцитрат и ротенон 1 мкг/мл), поддерживаемом при 25°С. Используя спектрофотометр, измеряли оптическую плотность при 340 нм в течение 2 мин, чтобы определить количество НАДФН, продуцируемого с участием IDPc, содержащейся в цитоплазматическом белке, определяя таким образом ферментативную активность IDPc. Для количественного определения этого фермента количество, которое продуцирует 1 мкМ НАДФН за 1 мин, принимали за 1 единицу. Количественное определение белка проводили анализом по Брэдфорду.
3. Количественное определение уровней триглицеридов и суммарного холестерина в крови
Для измерения уровней триглицеридов и суммарного холестерина в крови использовали аналитические наборы, выпускаемые Asan Pharmaceutics Co. Ltd. После обработки антикоагулянтом кровь, взятую у крыс, которым вводили ингибитор, и у крыс, которым не вводили ингибитор, центрифугировали с получением сыворотки. 10 мкл каждой сыворотки смешивали с 1,5 мл набора для анализа триглицеридов [раствор липопротеиназы 10800 ЕД, глицеролкиназы 5,4 ЕД, пероксидазы 135000 ЕД и L-α-глицерофосфатоксидазы 160 ЕД в 72 мл буфера с N,N-бис(2-гидроксиэтил)-2-аминометансульфоновой кислотой] или 1,5 мл набора для ферментативного анализа холестерина (смесь раствора ферментов (холестеролэстеразы 20,5 единиц Кимбрела/л, холестеролоксидазы 10,7 единиц Кимбрела/л и гидроксида натрия 1,81 г/л) и буфера (монофосфат калия 13,6 г/л, фенол 1,88 г/л) в соотношении 1:1) и инкубировали при 37°С в течение 5 мин для осуществления реакции. В устройстве для прочтения микропланшетов производили измерения оптической плотности при 500 нм для холестерина и при 540 нм для триглицеридов, чтобы количественно определить уровни триглицеридов и суммарного холестерина в крови. В связи с количественным определением триглицеридов и холестерина использовали стандартную кривую, которую получили с применением вышеописанной методики к разным известным концентрациям стандартного раствора.
4. Количественное определение уровней триглицеридов и суммарного холестерина в печени
От крыс, которым вводили ингибитор, и от крыс, которым не вводили ингибитор, брали заданное количество печени и гомогенизировали в 1 мл CIAA (хлороформ : изоамиловый спирт = 2:1 об/об). 100 мкл этого гомогената растворяли в 200 мкл чистого этилового спирта и смешивали с 500 мкл каждого из аналитических наборов для триглицерида и для холестерина, выпускаемых Asan Pharmaceutics Co. Ltd., содержащих 0,5% Тритон Х-100 и 3 мМ холат натрия, с последующей инкубацией при 37°С в течение 10 мин. После добавления 800 мкл воды каждый образец измеряли на уровни триглицеридов и суммарного холестерина в печени с помощью устройства для прочтения микропланшетов тем же способом, как описано выше. Подобным же образом вышеописанную методику применяли к разным известным концентрациям стандартного раствора для получения стандартной кривой, которую использовали для количественного определения уровней триглицеридов и холестерина в печени.
Результаты измерений, полученные в примере 8-2, анализировали и представили с виде графиков на фиг.10. Не было обнаружено значительного различия в массе тела крыс, которым вводили ингибитор, и крыс, которым не вводили ингибитор, где и те, и другие были в возрасте 10 недель. Как видно на фиг.10а, измерили, что у крыс, которым вводили ингибитор, масса жирового тела придатков яичек была примерно на 12% ниже по сравнению с крысами, которым не вводили ингибитор, при этом у нормальных крыс и у крыс, которым вводили ингибитор, отсутствовало различие в массе печени. Что касается триглицеридов и суммарного холестерина, их уровни в крови были ниже у крыс, которым вводили ингибитор, на 20% и 11% соответственно по сравнению с таковыми у крыс, которым не вводили ингибитор, как показано на фиг.10б. Атеросклеротический индекс также был в 10 раз ниже. С другой стороны, установили, что уровень липопротеинов высокой плотности (ЛПВП), которые транспортируют холестерин из тканей в печень по пути «обратного» транспорта холестерина и таким образом способствуют расщеплению и выделению холестерина, был на 11% более высокий у крыс, которым вводили ингибитор, по сравнению с крысами, которым не вводили ингибитор, как показано на фиг.10в.
Как описано выше, у крыс, которым вводили ингибитор IDPc, снижен уровень жировых тел придатков яичек, а также уровень триглицеридов и суммарного холестерина в крови. Кроме того, у крыс, которым вводили ингибитор IDPc, обнаружили снижение атеросклеротического индекса. Взятые вместе, эти результаты указывают на то, что ингибиторы IDPc оказывают отрицательное действие на ожирение у животных и снижают вероятность возникновения атеросклероза. Кроме того, установили, что ингибиторы IDPc ограничивают гиперхолестеринемию, поскольку они повышают клеточный уровень ЛПВП, которые вовлечены в расщепление и выделение холестерина.
В настоящем изобретении раскрыто, что прирост массы трансгенных мышей, у которых активно экспрессируется ген IDPc, является результатом аккумуляции жиров в организме и что уровни в клетках различных белков, показательных для ожирения, включая PPARγ, который представляет собой фактор транскрипции, стимулирующий экспрессию генов, кодирующих ферменты, вовлеченные в метаболизм липидов и холестерина, значительно повышены в жировой ткани у трансгенных мышей по сравнению с таковыми у нормальных мышей. Следовательно, увеличение экспрессии гена IDPc в первую очередь приводит к продуцированию больших количеств НАДФН, который необходим для биосинтеза жирных кислот и дает возможность полученному в результате обилию липидных производных индуцировать активацию и экспрессию гена PPARγ, который, в свою очередь, активирует экспрессию генов, показательных для ожирения, и в конце концов вызывает ожирение у мышей, трансгенных по IDPc.
На основании этих результатов исследований продемонстрировано, что ингибиторы IDPc полезны при лечении болезней обмена веществ. То есть, когда крысам вводят ингибиторы изоцитратдегидрогеназы, у них ограничивают ожирение и, кроме того, у них выявляют небольшие жировые тела придатков яичек, низкие уровни триглицеридов и суммарного холестерина и низкие атеросклеротические индексы. Следовательно, ингибиторы изоцитратдегидрогеназы могут подавлять индуцированный ожирением атеросклероз. Также, повышение уровня ЛПВП, связанное в ингибиторами IDPc, показывает, что снижение уровня суммарного холестерина после введения ингибиторов IDPc является следствием повышения уровня ЛПВП.
Промышленная применимость
В соответствии с настоящим изобретением установлено, что, как описано выше, что экспрессия гена IDPc и сопутствующее повышение уровня IDPc вызывает повышение уровня НАДФН в клетках, который, в свою очередь, вызывает отложение липидов в жировых клетках, приводя к ожирению и к жировой инфильтрации печени. Одновременно также обнаружено, что уровни триглицеридов и суммарный холестерин в крови повышены в результате экспрессии гена IDPc. С другой стороны, снижение клеточного уровня НАДФН в результате подавления экспрессии гена IDPc и сопутствующего внутриклеточного снижения уровней IDPc имеет эффект ингибирования отложения липидов в жировых клетках. Следовательно, ген IDPc, IDPc и НАДФН можно применять для синтеза липидов, включая жирные кислоты, сквален, DHA и т.д., и для активации PPARγ. Кроме того, поскольку мыши, трансгенные по гену IDPc, по настоящему изобретению явно проявляют симптомы ожирения, гиперлипидемии и жировой инфильтрации печени, продуцирующую НАДФН изоцитратдегидрогеназу, включая IDPc, и ее гены можно непосредственно применять для идентификации веществ, которые подавляют ожирение и жировую инфильтрацию печени, а также веществ, являющихся ингибиторами биосинтеза триглицеридов и холестерина. Далее, используя преимущество подавляющего или ингибирующего действия ингибиторов изоцитратдегидрогеназы на ожирение и биосинтез глицеридов и холестерина, можно разработать фармацевтически эффективные материалы для профилактики и лечения ожирения, гиперлипидемии и жировой инфильтрации печени.
Настоящее изобретение описано иллюстративно и следует понимать, что используемую здесь терминологию применяют для описания, но не для ограничения. Возможны многие модификации и варианты настоящего изобретения в свете описанных выше методик. Следовательно, следует понимать, что данное изобретение на практике можно применять в пределах объема прилагаемой формулы изобретения иначе, чем уже конкретно описано в данном описании изобретения.








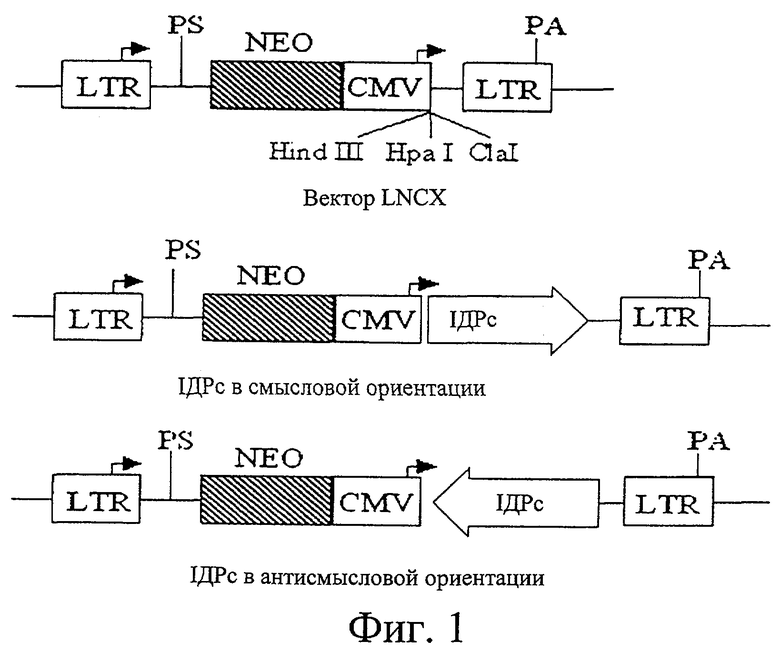
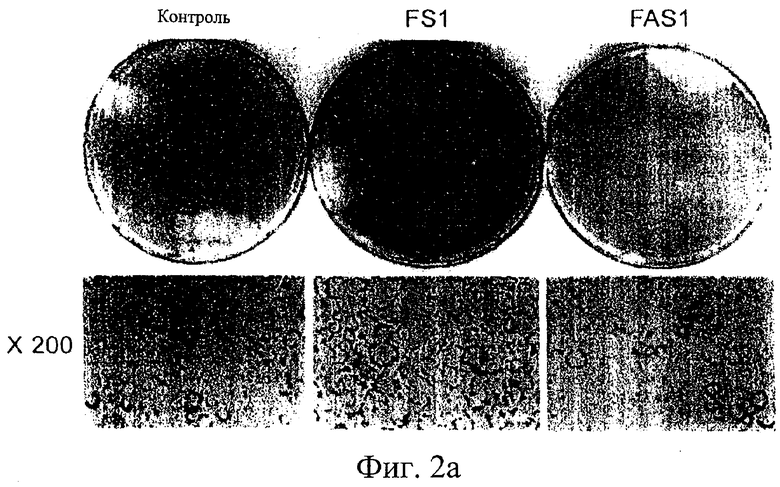
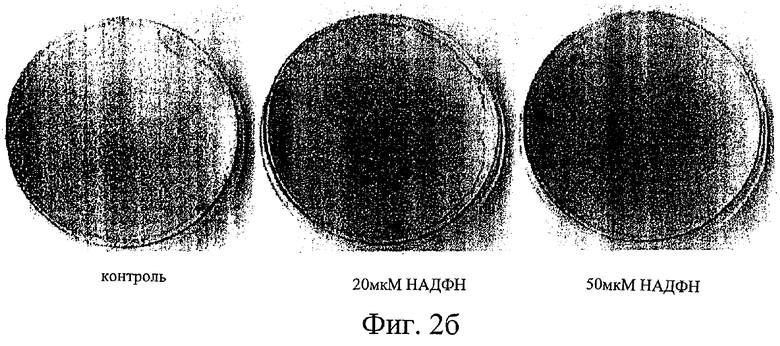
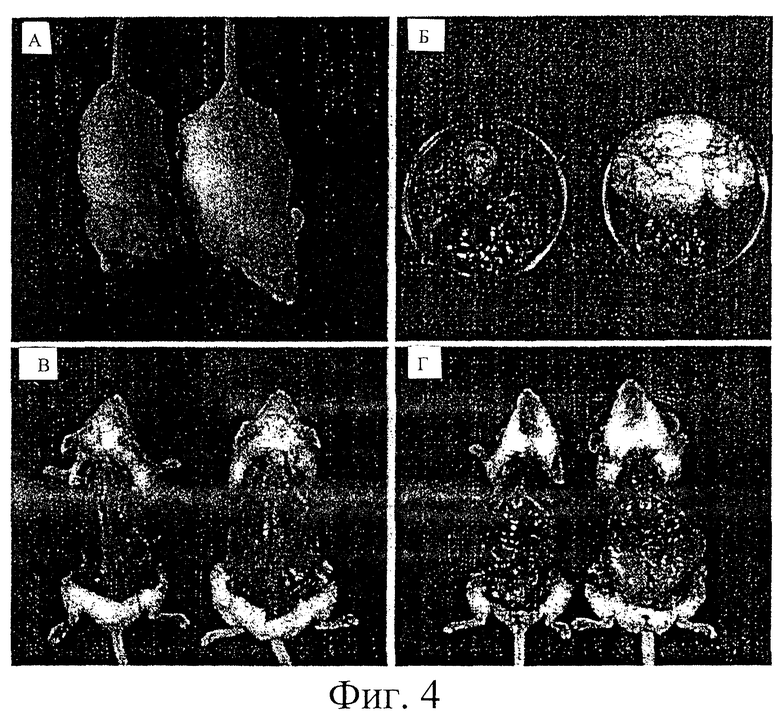
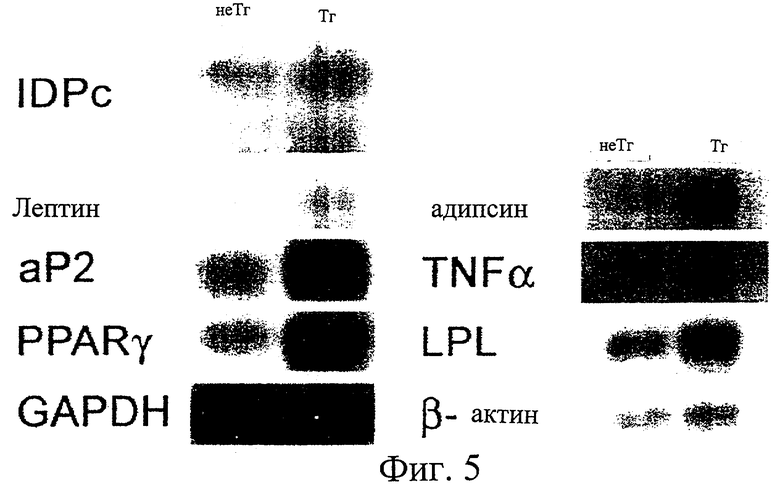
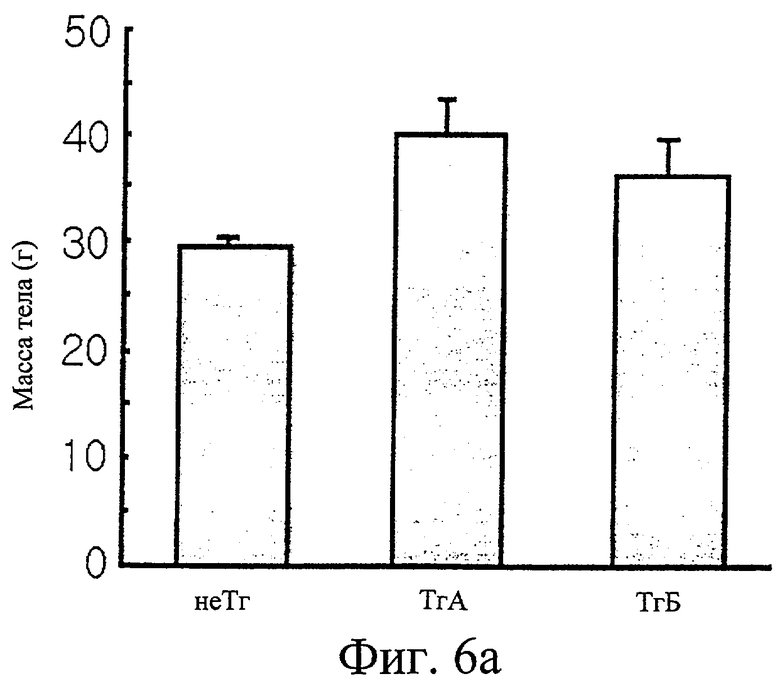
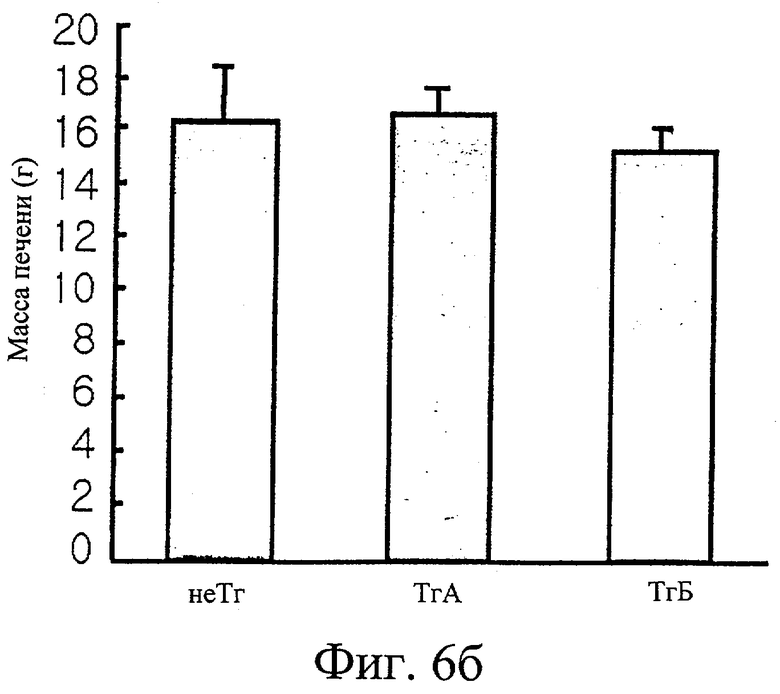
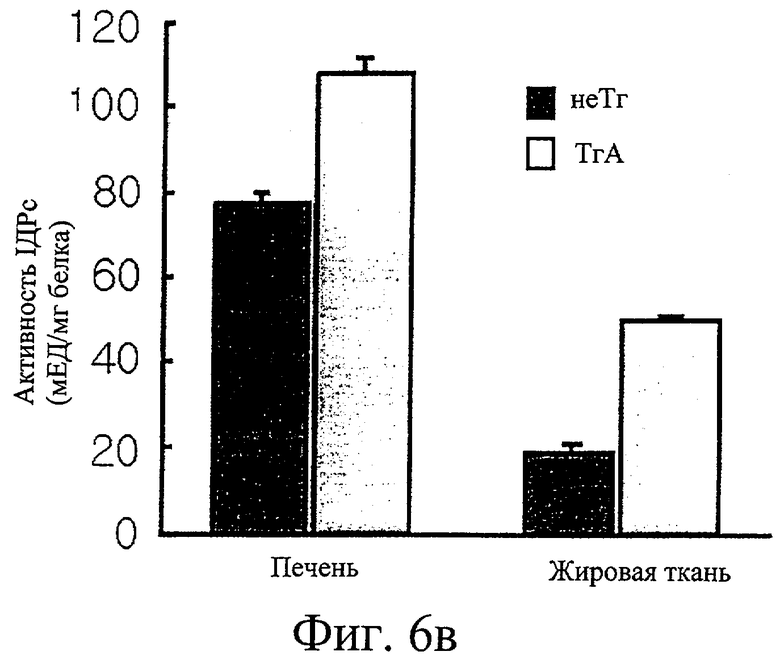
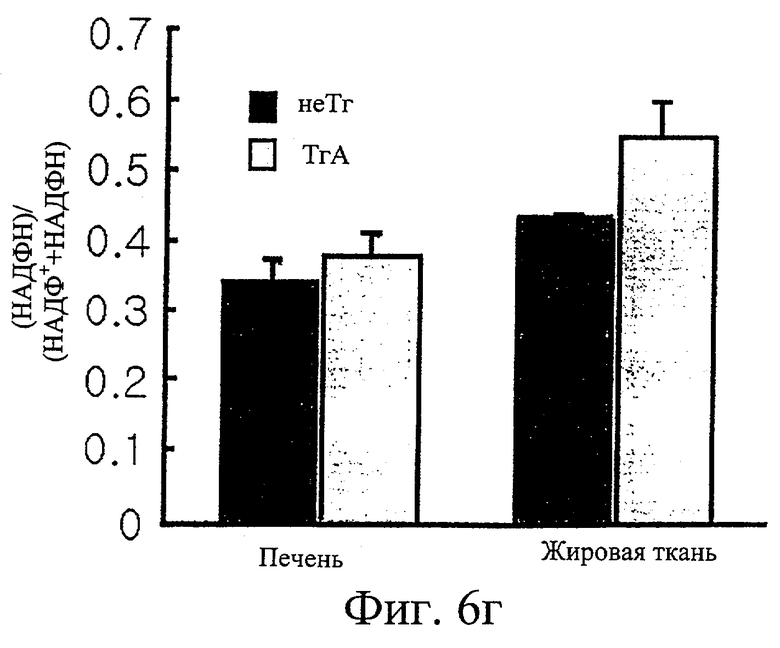
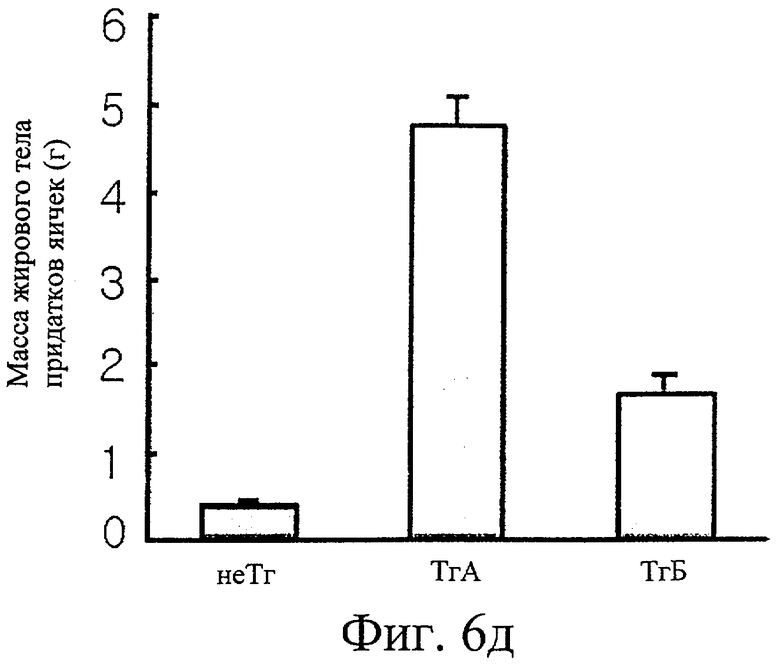
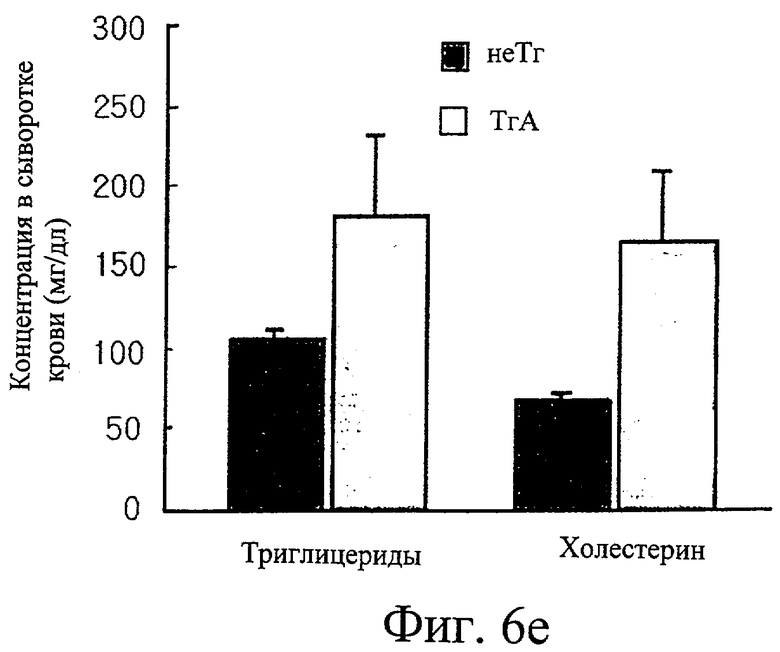
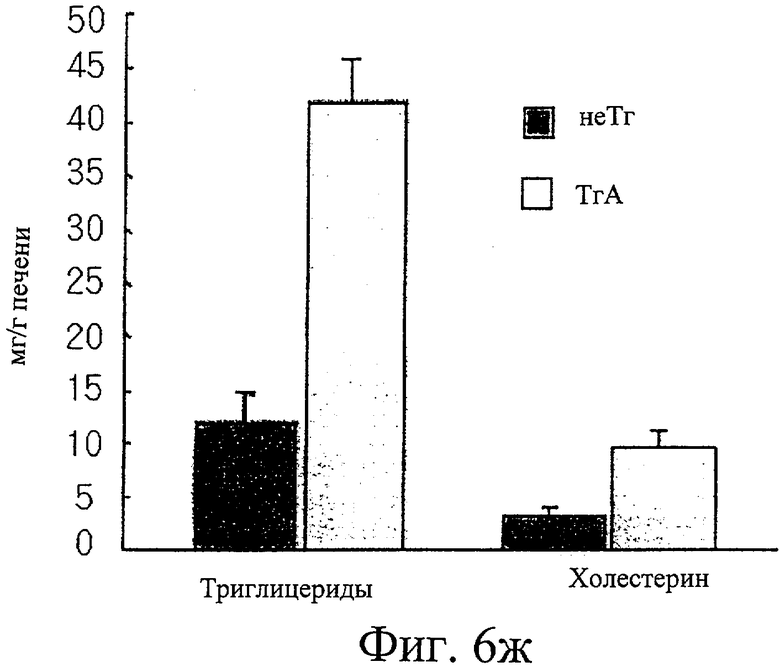
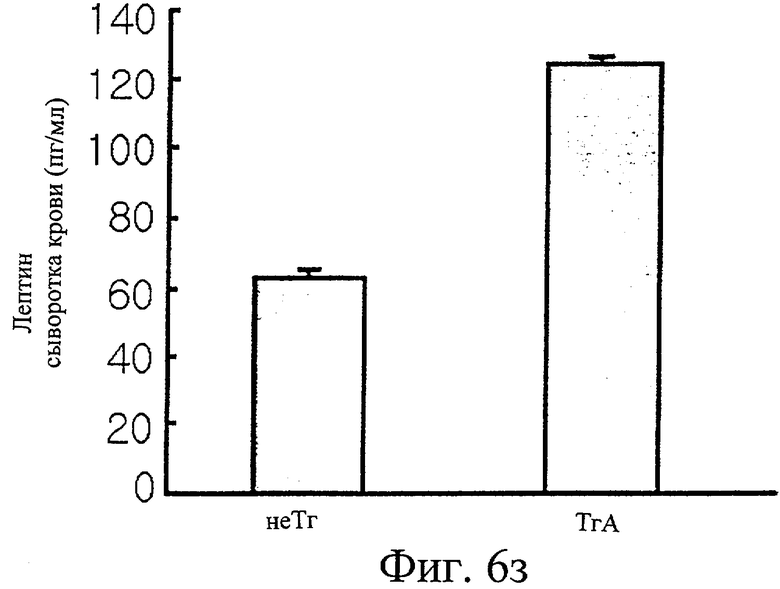

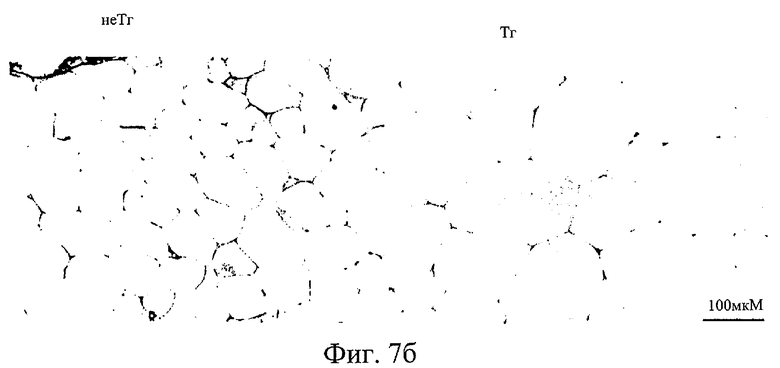
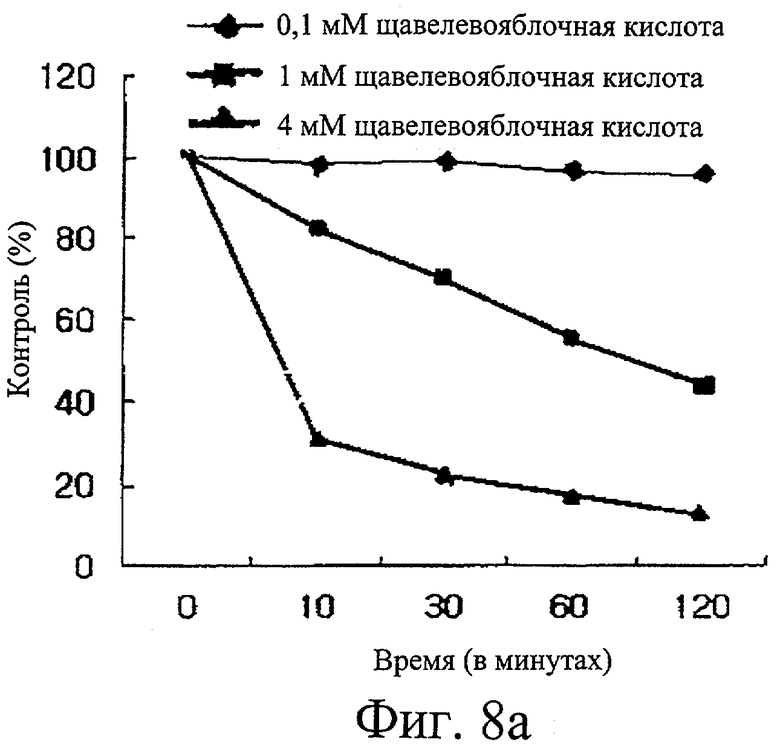
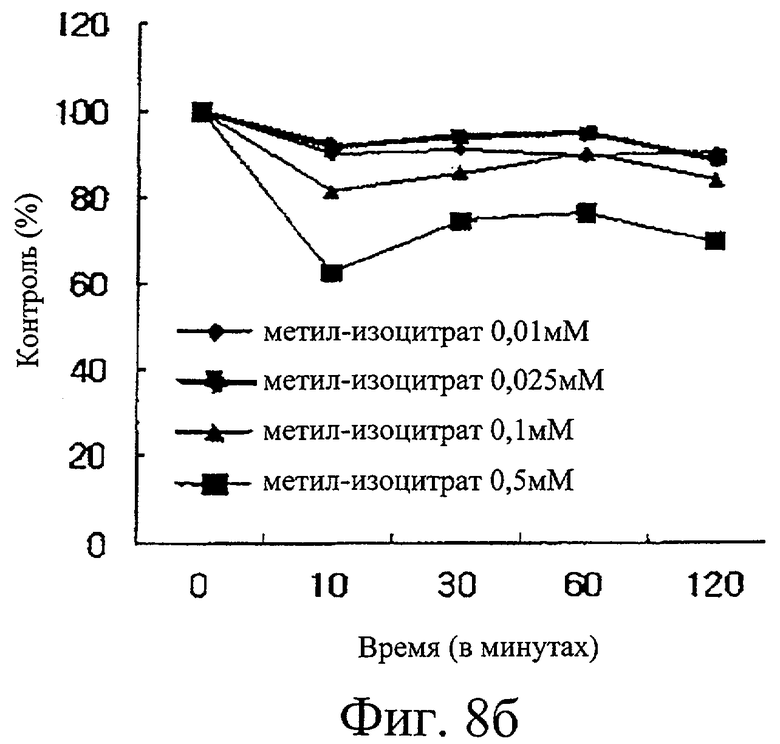
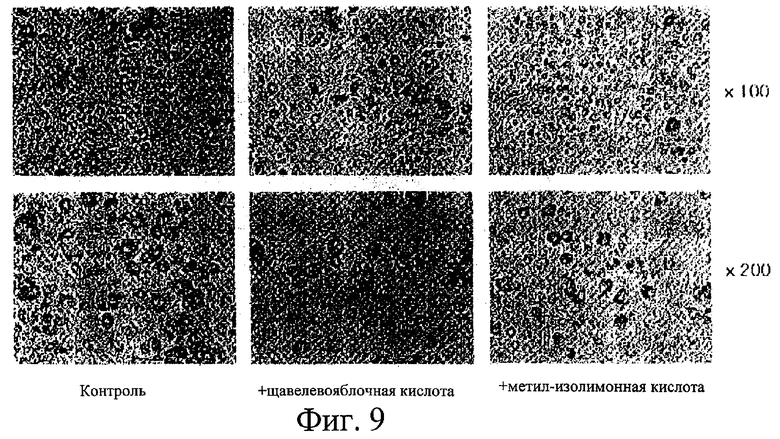
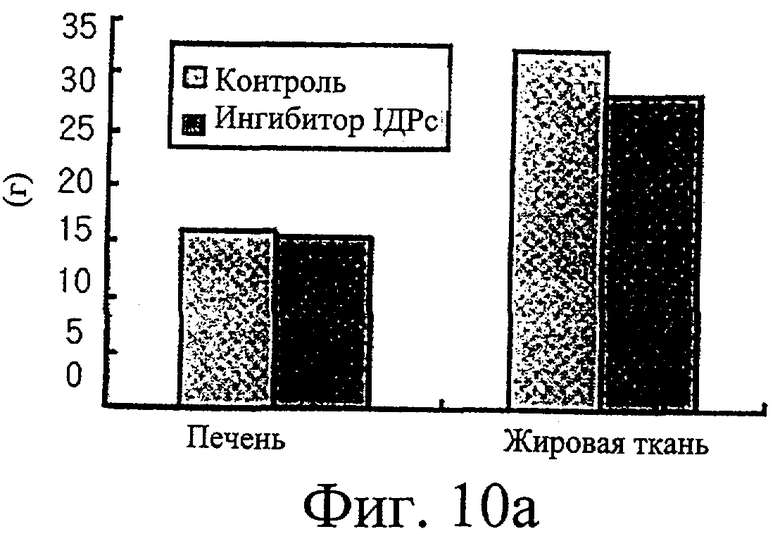
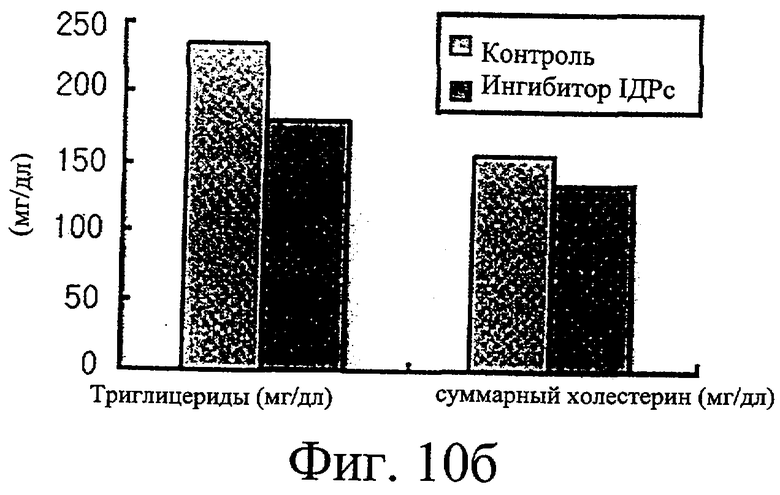
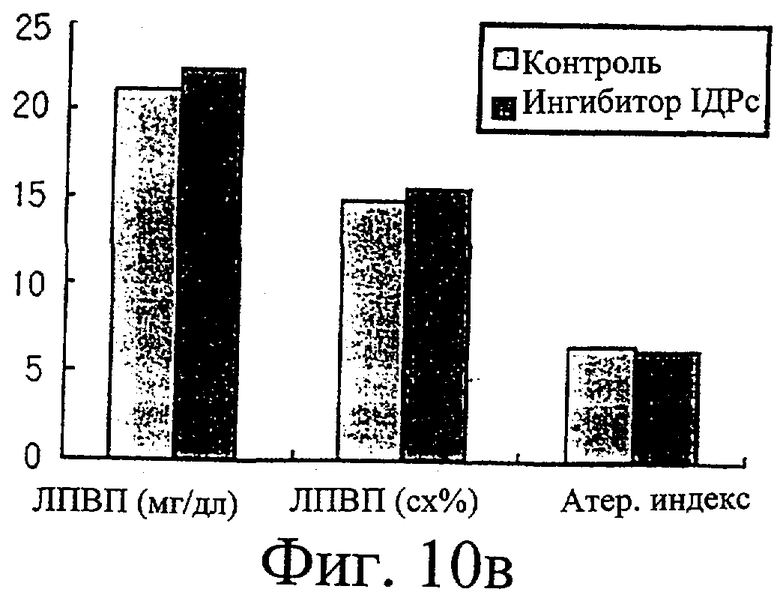
Изобретение относится к медицине и касается цитозольной изоцитратдегидрогеназы, ее гена и их использования в лечении ожирения, гиперлипидемии и жировой инфильтрации печени. Изобретение включает способ увеличения активности активируемого пролифератором пероксисом рецептора (PPAR), при котором вводят цитозольную НАДФ+-зависимую изоцитратдегидрогеназу (IDPc), ген IDPc или НАДФН, а также способ отбора антисмысловой молекулы, которая ингибирует отложение жиров или продуцирование триглицеридов и холестерина, при котором используют ген IDPc, а также способ лечения болезней обмена веществ, таких как ожирение, гиперлипидемия или жировая инфильтрация печени, при котором вводят антисмысловую молекулу гена IDPc или такой ингибитор IDPc, как щавелевояблочная кислота или метилизолимонная кислота. Изобретение позволяет снизить клеточный уровень НАДФ в результате подавления экспрессии гена IDPc, что приводит к ингибированию отложения липидов в клетках. 4 н.п. ф-лы, 10 ил.
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Felicia Gaskin, Clayton R.B | |||
| An interstain difference in cholesterol synthesis in vitro in mice, dependent upon a difference in endogenous NADPH-generating capacity // J | |||
| of Lipid Research, | |||

