Данное изобретение относится к растению ячменя с пониженной активностью фермента синтазы крахмала II (SSII), приводящей к крахмалу, имеющему пониженное содержание амилопектина. Кроме того, данное изобретение относится к крахмалу и зерну, а также к пищевым продуктам, полученным из них.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Одним из открытий в науке о питании является тот факт, что устойчивый крахмал имеет важное значение для здоровья кишечника, в частности, для здоровья толстого кишечника. Благоприятные эффекты устойчивого крахмала являются результатом обеспечения питания толстого кишечника, где кишечная микрофлора получает источник энергии, который подвергается ферментации с образованием среди прочего короткоцепочечных жирных кислот. Эти короткоцепочечные жирные кислоты обеспечивают питательные вещества для колоноцитов, усиливают захват некоторых питательных веществ через толстый кишечник и способствуют физиологической активности ободочной кишки. Как правило, если не обеспечивать устойчивые крахмалы или другую диетическую клетчатку, ободочная кишка относительно неактивна в метаболизме.
В последние годы существует направление поиска разработки устойчивых крахмалов из различных источников, направленного на здоровье кишечника. Соответственно, высокоамилозные крахмалы обнаружены в некоторых хлебных злаках, таких как кукуруза, для использования в пищевых продуктах в качестве средств, способствующих здоровью кишечника.
Физическая структура крахмала может оказывать важное воздействие на питательные и технологические свойства крахмала для пищевых продуктов. Некоторые характеристики можно рассматривать в качестве показателя структуры крахмала, включая распределение длин цепей амилопектина, степень кристалличности и наличие форм кристалличности, таких как V-комплексная форма кристалличности крахмала. Формы с такими характеристиками можно также рассматривать в качестве показателя питательных или технологических свойств пищевых продуктов, содержащих эти крахмалы. Таким образом, малая длина цепи амилопектина может быть показателем низкой кристалличности и слабой желатинизации, и кроме того считают, что она коррелирует с пониженной ретроградацией амилопектина. Кроме того, считают, что распределение длин более коротких цепей амилопектина отражает органолептические свойства пищевых продуктов, в которые этот крахмал включен в значительных количествах. Пониженная кристалличность крахмала может также быть показателем пониженной температуры желатинизации крахмала, и, кроме того, считают, что она связана с улучшенными органолептическими свойствами. Наличие V-комплексной кристалличности или иначе связанного с крахмалом липида будет увеличивать уровень устойчивого крахмала и, следовательно, диетической клетчатки.
Линии ячменя, имеющие крахмалы с высоким содержанием амилозы, были идентифицированы ранее. Результатом этого являлись только относительно небольшие повышения содержания амилозы до максимального значения примерно 45% от суммарного крахмала, как например в сорте ячменя, известном как High Amylose Glacier (AC38). Хотя крахмалы с повышенным содержанием амилозы этого типа полезны, все же предпочтителен крахмал с более высоким содержанием амилозы, и выращивают некоторые другие виды хлебных злаков, чтобы получить крахмалы с более высоким содержанием амилозы при уровнях 90-процентильного диапазона. Они очень устойчивы к ферментативному гидролизу и приносят большую пользу для здоровья.
При разработке высокоамилозных крахмалов существует проблема, поскольку известные высокоамилозные крахмалы также имеют высокую температуру желатинизации. Температура желатинизации отражает энергию измельчения, необходимую для обработки таких пищевых продуктов. Следовательно, для обработки зерна или муки для производства пищевых продуктов из таких зерен или крахмалов обычно необходимы более высокие температуры. Поэтому, как правило, продукты, имеющие высокоамилозные крахмалы, являются более дорогостоящими. Подобным образом, с точки зрения потребителя, для приготовления этих произведенных пищевых продуктов или для приготовления пищи из муки, имеющей высокоамилозные крахмалы, могут потребоваться более длительные периоды времени и более высокие температуры. Следовательно, в обеспечении высокоамилозных крахмалов в пищевых продуктах существует значительное неудобство.
Другим питательным компонентом хлебных злаков и, в частности, ячменя являются β-глюканы. β-Глюканы состоят из глюкозных единиц, связанных β(1-4) и/или β(1-3) гликозидными связями, и также не разрушаются человеческими пищеварительными ферментами, что делает их подходящими в качестве источника диетической клетчатки. β-Глюканы могут частично подвергаться ферментативному гидролизу эндогенными бактериями ободочной кишки, в процессе ферментации которых образуются короткоцепочечные жирные кислоты (преимущественно ацетат, пропионат и бутират), которые полезны для клеток слизистой оболочки, выстилающих кишечник и ободочную кишку (Sakata and Engelhard Comp. Biochem. Physiol. 74a: 459-462 (1983)).
Кроме того, поглощение β-глюкана имеет эффект повышения выделения желчных кислот, приводящего к снижению суммарных сывороточных холестерина и липопротеинов низкой плотности (ЛПНП) с уменьшением риска ишемической болезни сердца. Подобным образом, β-глюканы действуют посредством ослабления сдвигов концентрации глюкозы в крови, возникающих после приема пищи. Считают, что оба этих эффекта основаны на повышении вязкости содержимого желудка и кишечника.
Состав пищевых продуктов, содержащих крахмалы, и тесные взаимодействия этих крахмалов с другими питательными или иными компонентами могут оказывать значительное воздействие на питательную ценность этих пищевых продуктов или на функциональные характеристики этих компонентов в приготовлении или структуре этих пищевых продуктов.
Хотя модифицированные крахмалы или β-глюканы, например, можно использовать в пищевых продуктах, которые обеспечивают функциональность, в норме не обеспечиваемую немодифицированными источниками, такая обработка имеет тенденцию либо изменять другие важные компоненты, либо является нежелательной вследствие процессов, вовлеченных в модификацию. Следовательно, предпочтительно предложить источники составных частей, которые можно использовать в немодифицированной форме в пищевых продуктах.
Сорт ячменя МК6827 доступен от Barley Germplasma Collection (USDA-ARS National Small Grain Germplasma Research Facility Aberdeen, Idaho 831290 USA). Зерно МК6827 является сморщенным и имеет сильно окрашенную шелуху и продолговатую форму, и в руках авторов изобретения это зерно очень трудно поддается обработке, включая то, что оно очень устойчиво к измельчению. Свойства зерна МК6827 ранее не были охарактеризованы, природа мутации не установлена, и его не считают подходящим для производства пищевых продуктов.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Данное изобретение является результатом выделения и характеристики SSII мутанта растений ячменя, зерно которого, как обнаружено, содержит крахмал, который имеет пониженное содержание амилопектина и, следовательно, высокие относительные уровни амилозы, и поэтому имеет повышенные уровни диетической клетчатки.
Зерно этого мутанта и зерно от скрещивания в некоторые генетические фоны, кроме того, имеет повышенный уровень β-глюкана. Сочетание повышенного уровня β-глюкана и устойчивого крахмала, вносящее вклад в высокий уровень диетической клетчатки, авторы изобретения считают уникальным для настоящего изобретения.
Кроме того, по меньшей мере в некоторых генетических фонах обнаружено, что зерно от таких мутантов содержит крахмал, который имеет высокие относительные уровни амилозы, а также имеет низкие температуры желатинизации. Низкие характеристики набухания такого крахмала во время и после желатинизации также имеют преимущества при некоторых применениях технологий диетических и пищевых продуктов.
Кроме того, обнаружено, что зерно от таких мутантов содержит крахмал, который имеет высокие относительные уровни амилозы, причем обнаруженные уровни амилозы выше 50% от содержания крахмала, что представляет собой уровень, который никогда ранее не обнаруживали в немодифицированном крахмале, полученном из ячменя.
Крахмал мутантов и линий обратного скрещивания, имеющих происхождение от этих мутантов (до той степени, до которой эти линии обратного скрещивания протестированы), ведет себя как устойчивый крахмал с измененной структурой, на которую указывают специфичные физические характеристики, включая одну или более чем одну из группы, включающей в себя наличие высокого относительного содержания амилозы, физическую недоступность вследствие наличия высокого содержания β-глюкана, измененную морфологию гранул и наличие связанного с крахмалом липида, и на эту измененную структуру также указывает характеристика, выбранная из одной или более чем одной из группы, включающей в себя низкую кристалличность, пониженное распределение длин цепей амилопектина и наличие заметного связанного с крахмалом липида.
Кроме того, зерно, полученное от этих мутантных растений ячменя, можно легко использовать в процедурах технологии пищевых продуктов.
Настоящее изобретение в одном аспекте относится к крахмалу, полученному из зерна растения ячменя, имеющего пониженный уровень активности SSII, причем указанные крахмальные гранулы имеют высокое содержание амилозы вследствие пониженного содержания амилопектина.
Настоящее изобретение в другом, более широком аспекте относится к зерну, полезному для производства пищевых продуктов, полученному из растения ячменя, имеющего пониженный уровень активности SSII, причем крахмал указанного зерна имеет высокое содержание амилозы вследствие пониженного содержания амилопектина.
Настоящее изобретение еще в одном, более широком аспекте относится к растению ячменя с пониженным уровнем активности SSII, причем указанное растение ячменя способно производить зерно, причем крахмал указанного зерна имеет высокое содержание амилозы вследствие пониженного содержания амилопектина, причем указанное зерно является подходящим для производства пищевых продуктов.
Альтернативно, настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей белок ячменя SSII, причем указанная нуклеиновая кислота способна к гибридизации в жестких условиях с SEQ ID NO 1, или клетке, несущей реплицируемый рекомбинантный вектор, несущий указанную молекулу нуклеиновой кислоты. Еще в одном варианте данное изобретение относится к выделенной молекуле нуклеиновой кислоты, способной к специфичной гибридизации с SEQ ID NO 1.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Для лучшего понимания данное изобретение будет описано далее со ссылкой на ряд примеров.
Фиг.1. Анализ распределения молекулярных размеров крахмала, как определено путем ВЭЖХ-разделения крахмала в 90% ДМСО. (а) Himalaya, (б) АС38, (в) 342, (г) 292.
Фиг.2. Фотографии, показывающие морфологию зерна мутантной и родительских линий, (a) Himalaya, (б) АС38, (в) 292, (г) Waxiro, (д) 342, (е) Tantangara, (ж) МК6827, (з) Sloop. Размеры зерна в длину (L), ширину (W) и толщину (Т) проиллюстрированы на панели (а).
Фиг.3. Анализ распределения длин цепей различных крахмалов мутанта и дикого типа с использованием FACE. (а) Нормализованное распределение длин цепей, (б) сравнение распределений длин цепей с помощью диаграммы разностей. Образцами являлись 342  , 292
, 292  , Tantangara (s), AC38
, Tantangara (s), AC38  , МК6827
, МК6827  и Himalaya (+).
и Himalaya (+).
Фиг.4. RVA-анализ образцов крахмала ячменя. Образцами являлись Himalaya , Namoi (Δ), AC38 (О), 342 (▿), 292  и МК6827 . Температурный профиль, используемый на протяжении профиля, указан непрерывной линией.
и МК6827 . Температурный профиль, используемый на протяжении профиля, указан непрерывной линией.
Фиг.5. Данные дифракции рентгеновских лучей для мутантной линии и линии дикого типа.
Фиг.6. Микрофотографии в сканирующем электронном микроскопе выделенных крахмалов ячменя, (a) Himalaya, (б) Waxiro, (в) AC38, (г) 292, (д) 342, (е) МК6827.
Фиг.7. Локусы на хромосоме 7Н ячменя, показывающие близость локусов nud1 и sex6. Диаграмма согласно GrainGenes (http://wheat.pw.usda.gov/) Barley morphological genes, 7H map, author; Franckowiak JD).
Фиг.8. Зависимости между размерами семян и распределением длин цепей крахмала для двойных гаплоидных линий 292×Tantangara. Линии, обозначенные (+), дали картину ПЦР Himalaya, а линии, обозначенные (о), дали результат ПЦР 292. Панель (А), отношение длины семени к толщине, нанесенное на график против процента цепей крахмала с DP от 6 до 11; панель (Б), масса семени, нанесенная на график против процента цепей крахмала с DP от 6 до 11.
Фиг.9. Последовательность кДНК SSII ячменя (SEQ ID NO 1) из сорта Himalaya.
Фиг.10. Структура генов SSII из (1) Т.tauschii (диплоидная пшеница), (2) сорта ячменя Morex. Линиями жирного шрифта представлены экзоны, а тонкими линиями - интроны. Прямая линия под каждым примером указывает участок последовательностей генов. Пунктирной линией показан участок гена SSII ячменя, от интрона 7, который не был секвенирован, но был определен ПЦР-анализом примерно как 3 т.п.о. в длину.
Фиг.11. Сравнения предсказанных кДНК SSII из МК6827 (SEQ ID NO 2), Morex (SEQ ID NO 3) и 292 (SEQ ID NO 4) и последовательности кДНК Himalaya (SEQ ID NO 1). Предсказанные последовательности получили путем идентификации участков геномных последовательностей, присутствующих в кДНК SSII Himalaya. Стартовый кодон ATG и стоп-кодон дикого типа указаны, как и дополнительные стоп-кодоны, присутствующие в МК6827 (#) и 292 (&), соответственно.
Фиг.12. Сравнение аминокислотных последовательностей, имеющих происхождение от генов, кодирующих SSII, из линий ячменя 292 (SEQ ID NO 7), Morex (SEQ ID NO 5), MK6827 (SEQ ID NO 8), Himalaya (SEQ ID NO 6). Дополнительные стоп-кодоны в 292 и MK6827 указаны символами (&) и (#), соответственно.
Фиг.13. Положение мутаций в MK6827 (SEQ ID NO 2) и 292 (SEQ ID NO 4) в гене SSII ячменя.
Фиг.14. Разработка и использование ПЦР-анализа для мутации 292. (а) Схематическое изображение участка SSII из Himalaya, амплифицированного с помощью праймеров ZLSS2P4 и ZLBSSIIP5, (б) изображение участка, амплифицированного из гена SSII из 292 с использованием ZLSS2P4 и ZLBSSIIP5, показывающее отсутствие одного сайта NIaIV, (в) агарозный гель-электрофорез продуктов ферментативного гидролиза NIaIV из ячменя; Дорожка М; лэддер маркеров ДНК, дорожка 1: МК6827, дорожка 2: Himalaya, дорожка 3: Tantangara, дорожка 4: 292, дорожка 5: 342.
Фиг.15. ДСН-ПААГ электрофорез (электрофорез в полиакриламидном геле с додецилсульфатом натрия) белков крахмальной гранулы. Панель (А) 8% акриламид (37,5:1 акрил/бис) ДСН-ПААГ гель, подвергнутый электроблоттингу и зондированию с антителом SSII, полученным против очищенного связанного с гранулами белка SSII из пшеницы. (Б) 12,5% акриламид (30:0,135 акрил/бис), окрашенный серебром. Миграция стандартов молекулярных масс определенной массы (единицами являются кД) указаны с каждой стороны фигуры.
Фиг.16. Схематическое изображение конструкций ДНК, предназначенных для негативной регуляции экспрессии SSII после стабильной трансформации ячменя. (1) Ген SSII от нуклеотида 1 до 2972 (см. последовательность на Фиг.9) встроен между промотором и терминатором в смысловой ориентации. (2) Ген SSII встроен между промотором и терминатором в антисмысловой ориентации от нуклеотида 2972 до 1 (см. последовательность на Фиг.9). (3) Двойная конструкция, в которой интрон 3 гена SSII ячменя (между нуклеотидами 1559 и 2851) геномной последовательности SSII Morex встроен между экзонами 2 и 3 из кДНК SSII ячменя из Himalaya (нуклеотиды 363-1157 на Фиг.9).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Гликемический Индекс. Это сравнение воздействия тестируемых пищевых продуктов, таких как белый хлеб или глюкоза, на сдвиги концентрации глюкозы в крови. Гликемический Индекс представляет собой меру вероятного эффекта пищи, связанного с концентрацией глюкозы в сыворотке после приема пищи и потребностью в инсулине для гомеостаза глюкозы в крови.
Устойчивый крахмал. Сумма крахмала и продуктов ферментативного гидролиза крахмала, не всасывающихся в тонком кишечнике здоровых людей, но поступающих в толстый кишечник. Таким образом, устойчивый крахмал исключает продукты, перевариваемые и всасывающиеся в тонком кишечнике.
Устойчивые крахмалы можно классифицировать на четыре группы.
RS1 физически недоступный крахмал. Примеры этой формы крахмала образуются там, где крахмал заключен внутри белкового или подобного матрикса, либо внутри растительной клеточной стенки, либо она может образоваться в связи с частичным измельчением зерна или бобовых после охлаждения.
RS2 устойчивые гранулы. Это, как правило, сырые крахмалы, такие как те, которые образуются из сырого картофеля или зеленых бананов, некоторых бобовых и высокоамилозных крахмалов.
RS3 ретроградированные крахмалы. Эти крахмалы образуются путем термической/влажной обработки крахмала или крахмальных пищевых продуктов, такие как встречаются в вареном и охлажденном картофеле, хлебе и кукурузных хлопьях.
RS4 химически модифицированные. Эти крахмалы образуются вследствие химических модификаций, таких как замещение или поперечное сшивание. Эту форму крахмала часто используют в обработанных пищевых продуктах.
Диетическая клетчатка. В данном описании это сумма углеводов или продуктов ферментативного гидролиза углеводов, которые не всасываются в тонком кишечнике здоровых людей, но поступают в толстый кишечник. Они включают в себя устойчивый крахмал, β-глюкан и другие растворимые и нерастворимые углеводные полимеры. Следует включать ту часть углеводов, которая по меньшей мере частично подвержена ферментации резидентной микрофлорой в толстом кишечнике.
Желатинизация представляет собой разрушение (прерывание) молекулярного порядка в пределах крахмальной гранулы с сопутствующими и необратимыми изменениями свойств, такими как гранулярное набухание, плавление кристаллита, потеря двойного лучепреломления, развитие вязкости и солюбилизация крахмала.
Данное изобретение возникло в результате выделения и характеристики SSII мутантных растений ячменя, зерно которых, как обнаружено, содержит крахмал, который имеет пониженное содержание амилопектина и, следовательно, высокие относительные уровни амилозы, и поэтому имеет повышенные уровни диетической клетчатки.
Обнаружено, что такие мутанты обладают рядом весьма желательных характеристик, и показано, что скрещивания в различные другие генетические фоны сохраняют по меньшей мере некоторые из этих характеристик.
Зерно мутанта и зерно от скрещиваний в некоторые генетические фоны, кроме того, имеют повышенный уровень β-глюкана. Сочетание повышенного уровня β-глюкана и высокого уровня диетической клетчатки авторы изобретения считают уникальным для настоящего изобретения.
Кроме того, обнаружено, что по меньшей мере в некоторых генетических фонах зерно от таких мутантов содержит крахмал, который имеет высокие относительные уровни амилозы, а также имеет низкие температуры желатинизации. Характеристики желатинизации, относящиеся к набуханию, такого крахмала также имеют преимущество, представляющее собой слабое набухание, которое является полезным при некоторых применениях в технологии диетических и пищевых продуктов.
Кроме того, обнаружено, что зерно от таких мутантов содержит крахмал, который имеет высокие относительные уровни амилозы, причем обнаруженные уровни амилозы выше 50% от содержания крахмала, что представляет собой уровень, который ранее никогда не обнаруживали в немодифицированном крахмале, полученном из ячменя.
Крахмал мутантов и линий обратного скрещивания до той степени, до которой они протестированы, ведет себя как устойчивый крахмал с измененной структурой, на которую указывают специфичные физические характеристики, включая одну или более чем одну из группы, включающей в себя наличие высокого относительного содержания амилозы, физическую недоступность вследствие наличия высокого содержания β-глюкана, измененную морфологию гранул и наличие связанного с крахмалом липида, и на эту измененную структуру также указывает характеристика, выбранная из одной или более чем одной из группы, включающей в себя низкую кристалличность, пониженное распределение длин цепей амилопектина и наличие заметного связанного с крахмалом липида.
Кроме того, по этой причине зерно, полученное от этих мутантных растений ячменя, можно легко использовать в процедурах технологии пищевых продуктов.
Зерно от таких мутантов в одной форме предпочтительно содержит крахмал, который имеет высокие относительные уровни диетической клетчатки, более конкретно амилозы, а также повышенный уровень β-глюкана. Сочетание повышенного уровня β-глюкана и высокого уровня амилозы авторы изобретения считают уникальным для настоящего изобретения, и оно обеспечивает уникальный источник сочетания β-глюкана и устойчивого крахмала, которое не требует, по меньшей мере в более широких аспектах изобретения, смешивания β-глюкана и растворимой диетической клетчатки вместе или модификации этих составных частей.
Насколько известно авторам изобретения растение ячменя по настоящему изобретению является впервые открытой зерновой культурой ячменя, обладающей повышенными относительными уровнями диетической клетчатки в форме устойчивого крахмала, имеющего повышенный уровень амилозы, который также имеет повышенные уровни β-глюкана, которые находятся на более высокой точке типичных уровней β-глюкана или которые превышают этот уровень. Зерна, которые имеют еще более высокое содержание β-глюкана, представляют собой фенотип waxy и, следовательно, обладают низкими уровнями амилозы.
Известно, что существует широкое разнообразие уровней β-глюкана в ячмене в диапазоне от примерно 4% до примерно 18% масс/масс ячменя, но более типично от 4% до примерно 8% (Izydorcyk et at. (2000) Journal of Agricultural and Food Chemistry 48, 982-989; Zheng et al. (2000) Cereal Chemistry 77, 140-144; Elfverson et al. (1999) Cereal Chemistry 76, 434-438; Andersson et al. (1999) Journal of the Science of Foods and Agriculture 79, 979-986; Oscarsson et al. (1996) J. Cereal Science 24, 161-170; Fastnaught et al. (1996) Crop Science 36, 941-946). Разработаны улучшенные линии ячменя, например Prowashonupana, которые имеют от примерно 15% до примерно 18% масс/масс β-глюкана, но обладает фенотипом waxy. Она имеется в продаже под названием Sustagrain™ (ConAgra™ Specially Grain Products Company, Omaha, Neb. USA).
Уровни β-глюкана, рассматриваемые данным изобретением, могут зависеть от генетического фона, при котором ферментативная активность синтеза амилопектина понижена. Однако предполагают, что понижение активности синтеза амилопектина будет обладать эффектом повышения относительного уровня диетической клетчатки, которая частично принимает форму амилозы, и в то же время повышения уровня β-глюкана. Одним из объяснений сопутствующего повышения β-глюкана при повышенных относительных уровнях амилозы является то, что такое повышение может быть результатом концентрационного эффекта уменьшенного эндосперма и может далее повышаться посредством переключения углерода от синтеза крахмала на синтез β-глюкана.
Таким образом, зерно растения ячменя предпочтительно имеет содержание β-глюкана, которое выше 6% от суммарной массы нешелушенного зерна, или более предпочтительно выше 7%, и наиболее предпочтительно выше 8%, однако, уровни β-глюкана у мутанта waxy измерили вплоть до уровней от 15 до 18%, и в настоящем изобретении можно рассматривать столь же высокие или более высокие уровни, чем эти.
Во второй предпочтительной форме зерно растения ячменя обладает пониженной температурой желатинизации (как измерено с помощью дифференциальной сканирующей калориметрии) в дополнение к относительно высокому содержанию амилозы. По данным, представленным для ячменя, служащего примером, эта пониженная температура желатинизации не только снижена по сравнению с крахмалом, продуцируемым ячменем с несколько повышенным содержанием амилозы, но также по сравнению с крахмалом, продуцируемым ячменем, имеющим крахмал, у которого нормальные уровни амилозы. Таким образом, хотя в данном изобретении рассматривают пониженные температуры желатинизации относительно соответствующего высокоамилозного крахмала, можно также рассматривать температуру желатинизации, пониженную относительно таковой у крахмала с нормальными уровнями амилозы.
Кроме того, в генетических фонах, проверенных таким образом, крахмал также характеризуется набуханием в избытке нагретой воды, которое является более слабым, чем набухание других протестированных крахмалов.
В третьей предпочтительной форме крахмал имеет уровни амилозы выше 50% от содержания крахмала, что является уровнем, который никогда ранее не обнаруживали в немодифицированном крахмале, полученном из ячменя.
Крахмал настоящего растения ячменя имеет высокое относительное содержание амилозы, более высокое, чем можно ожидать для мутации в гене SSII или другом гене синтазы крахмала. Так, у пшеницы мутанты по SSII приводят в результате к относительным уровням амилозы примерно 35% от крахмала. Содержание амилозы у крахмала можно считать повышенным, когда это содержание значительно выше 25%, или которое присутствует в зерне нормального ячменя, и, таким образом, может быть выше чем примерно 30% масс/масс от суммарного крахмала. Известные растения ячменя, которые считают высокоамилозными, имеют содержание 35-45%. Однако в настоящем изобретении предложен ячмень с содержанием амилозы, которое выше 50%, что является уровнем, который никогда ранее не обнаруживали в немодифицированном крахмале, полученном из ячменя.
Относительное содержание амилозы может быть выше 60% и, более предпочтительно, даже выше 70%. Может быть желательно иметь даже более высокие уровни и, следовательно, было возможно достичь даже более высоких уровней у других растений с помощью скрещивания с одиночными мутациями, причем такие уровни достигают 90%. Следовательно, данное изобретение может охватывать уровни амилозы, которые выше 80% или выше 90%.
В четвертой предпочтительной форме крахмал также имеет измененную структуру, которая приводит к устойчивому крахмалу. Это может являться результатом высокого содержания амилозы. Устойчивый крахмал может также образоваться, поскольку β-глюкан присутствует в повышенных уровнях и, вероятно, оказывает защитные воздействия вследствие связывания β-глюкана с крахмальной гранулой, причем прочность контакта потенциально обеспечивает защитный эффект для крахмала, чтобы посредством этого обеспечить устойчивость, которую можно охарактеризовать как RS1 форму, которая до некоторой степени недоступна ферментативному гидролизу. Подобным образом, наличие связи крахмал-липид, как измерено на основании V-комплексной кристалличности, также вероятно вносит вклад в уровень устойчивого крахмала. В этом случае устойчивость, по-видимому, является результатом физической недоступности крахмала благодаря наличию липида, и, соответственно, ее можно рассматривать как RS1 крахмал. Известно, что ретроградированный крахмал, который принимает V-комплексную конфигурацию, является высоко устойчивым к ферментативному гидролизу и соответственно ожидают, что амилопектин, который образует часть V-комплексной кристаллической структуры, также будет устойчив к ферментативному гидролизу. Крахмал растения ячменя, служащего примером, может быть устойчивым к ферментативному гидролизу вследствие структуры крахмальной гранулы и, соответственно, может иметь RS2 крахмал. Каждая из этих характеристик может присутствовать отдельно, либо в виде двух или более чем двух этих характеристик в сочетании.
Повышенный уровень диетической клетчатки может, по меньшей мере частично, принимать форму устойчивого крахмала, который можно охарактеризовать высоким содержанием амилозы крахмальных гранул, как объяснено выше.
Относительное содержание амилозы может быть выше 60% и более предпочтительно выше 70%. Может быть желательно иметь даже более высокие уровни, и поэтому было возможно достичь даже более высоких уровней в других растениях с помощью скрещивания с одиночными мутациями, и такие уровни достигают 90%. Таким образом, данное изобретение может охватывать уровни амилозы выше 80% или выше 90%.
Может быть желательно, чтобы растение ячменя дополнительно экспрессировало измененный уровень активности одного или более чем одного фермента синтеза амилозы или других ферментов для еще большего повышения относительного уровня амилозы. Таким образом, это растение ячменя может нести другую мутацию, которая дополнительно уменьшает или изменяет биосинтез амилопектина, либо мутацию или генетический фон, которые повышают биосинтез амилозы. Например, это растение ячменя может проявлять генотип amylose extender, как например растение ячменя, несущее мутацию amo 1. Примером такого растения является сорт, известный как АС38 (также известный как High Amylose Glacier).
Следует понимать, что относительный уровень амилозы, который обсуждали, составляет уровень по отношению к суммарному содержанию крахмала, и поэтому остальная часть крахмала может представлять собой преимущественно промежуточный тип крахмала, либо она может представлять собой преимущественно амилопектин, либо смесь обоих типов. В проанализированном ячмене повышенный уровень амилозы является результатом пониженных уровней амилопектина, и, соответственно, относительный уровень амилозы не является результатом повышенного синтеза амилозы.
Известно, что β-глюкан обладает эффектом замедления ферментативного гидролиза в тонком кишечнике просто в результате его присутствия вместе с другим пищевым компонентом. Подобным образом, известно, что устойчивые молекулы, которые имеют близкое соседнее положение с крахмальными гранулами, помогают маскировать крахмал и вносят вклад в его устойчивость, делая его физически недоступным. Повышенные уровни амилозы и других форм крахмала, которые могут образоваться в результате связи с липидом, будут, следовательно, дополнительно повышены в результате присутствия и физического соседнего положения с крахмальными гранулами. Следовательно, обеспечено значительное усиление эффектов устойчивого крахмала, а также обеспечение других полезных эффектов, являющихся результатом высоких уровней β-глюкана.
Кроме того, известно, что существует ответ на дозу в отношении полезных эффектов устойчивого крахмала и β-глюкана. Следовательно, предполагают, что повышенный уровень β-глюкана вместе с повышенными уровнями устойчивого крахмала будут обеспечивать повышенную пользу для здоровья.
Сочетание уровней β-глюкана и устойчивого крахмала по меньшей мере предпочтительных вариантов данного изобретения не было обнаружено ранее и, несомненно, ни из одного источника без некоторой степени модификации или очистки, и поэтому формы настоящего изобретения обеспечивают единственный практический источник этой пользы.
Другой предпочтительный аспект крахмала состоит в том, что, несмотря на высокое относительное содержание амилозы, он также имеет низкую температуру желатинизации, как измерено с помощью дифференциальной сканирующей калориметрии. Это находится в противоречии с общепринятыми сведениями относительно того факта, что высокоамилозные крахмалы склонны иметь повышенную температуру желатинизации, которая вводит ограничения на способ, при котором можно использовать высокоамилозные крахмалы. На основании данных, представленных для ячменя, служащего примером, эта пониженная температура желатинизации не только снижена по сравнению с крахмалом, продуцируемым линиями с несколько повышенным содержанием амилозы, но также по сравнению с крахмалом, продуцируемым ячменем, имеющим крахмал, у которого нормальные уровни амилозы. Таким образом, хотя в предпочтительном аспекте данного изобретения рассматривают пониженные температуры желатинизации относительно соответствующего высокоамилозного крахмала, можно также рассматривать температуру желатинизации, пониженную относительно таковой у крахмала с нормальными уровнями амилозы. Для высокоамилозных крахмалов аспекты обработки, требующие высоких температур, следовательно, неизбежно требуют высокого потребления энергии, что является дорогостоящим и может разрушить функциональность других пищевых компонентов. Подобным образом, с точки зрения непосредственного потребителя, пищевые продукты высокоамилозного крахмала могут быть менее удобны по причине высокой температуры или более длительного времени, необходимого для приготовления. Поэтому, например, в данном предпочтительном аспекте изобретения теперь возможно предложить продукт, такой как продукт в виде лапши, требующий добавления кипящей или нагретой воды в сосуд, такой как чашка, и не требующий нагревания в течение длительного периода времени, и в то же время обеспечивающий доставку устойчивых крахмалов и других составных частей, имеющих питательную ценность, в толстый кишечник.
Главным эффектом низких температур желатинизации этих крахмалов являются более низкие температурные требования и, следовательно, энергия измельчения этой пищи. Следствием также является то, что, если, как типично может происходить при некоторой обработке пищи, смешивание происходит при комнатной температуре, а затем эту смесь нагревают, более низкая температура желатинизации также уменьшает время, необходимое для достижения желатинизации. Кроме того, при диапазоне температур ниже температуры полной желатинизации нормального крахмала, будет происходить более полная желатинизация крахмала по настоящему изобретению, чем нормального крахмала.
Одна из мер способности к желатинизации отражена в термических свойствах на основании измерения с помощью ДСК (дифференциальной сканирующей калориметрии). Начало первого пика (пик желатинизации) ДСК может находиться при температуре менее 53°С, более предпочтительно при температуре менее 50°С и наиболее предпочтительно при температуре менее чем примерно 47°С. Начало первого пика можно рассматривать как начало желатинизации. Крахмал, полученный из зерна ячменя, может иметь первый пик при температуре менее чем примерно 60°С, более предпочтительно при температуре менее чем 55°С, и наиболее предпочтительно при температуре менее чем 52°С. ΔН (Энтальпия) первого пика может быть менее чем примерно 3,5, более предпочтительно менее чем примерно 1,0 и наиболее предпочтительно менее чем примерно 0,5.
Другим открытием в отношении желатинизации видов муки, содержащих крахмалы по данному изобретению, является то, что они проявляют пониженное набухание. Объем набухания типично измеряют путем смешивания крахмала или муки с избытком воды и нагревания до повышенных температур, типично выше 90°С. Затем образец собирают путем центрифугирования, и объем набухания выражают в виде массы осажденного материала, разделенной на сухую массу образца. Обнаружено, что объемы набухания муки из крахмалов waxy и нормальных ячменей выше чем примерно 5,5. Объемы набухания муки, полученной из зерна, которое представляет собой высокоамилозное зерно (АС38), составляют примерно 3,75. Тогда как из зерна исследованных мутантов и гибридов они составляют менее чем 3,2, предпочтительно менее чем 3,0, но, как правило, выше чем примерно 2.
Эта характеристика желатинизации, относящаяся к низкому набуханию, особенно полезна, если желательно увеличить содержание крахмала пищевого препарата, в частности, гидратированного пищевого препарата. В настоящем случае может быть желательно увеличить содержание диетической клетчатки золя или другого жидкого препарата, где иначе существовало бы ограничение на доставку этого пищевого препарата.
Данная характеристика в сочетании с пониженной температурой желатинизации, проявляемой настоящим крахмалом, обеспечивает перспективу значительного усиления питательной пользы пищевых продуктов, где имеется потребность в быстром приготовлении, таких как растворимые супы и лапши быстрого приготовления.
Постулировано, что эффекты температуры желатинизации являются результатом измененной структуры амилопектина в эндосперме зерна, и одним из измерений этой структуры является распределение длин цепей (степеней полимеризации) молекул крахмала после удаления разветвления цепи изоамилазой. Анализ длин цепей амилопектинового содержимого крахмала мутантов SSII, служащих примером, показал, что при удалении разветвления цепи они имеют распределение длин цепей в диапазоне от 5 до 60, что короче, чем распределение крахмала, которое дают немутантные линии после удаления разветвления цепи. Крахмал с меньшими длинами цепей будет также иметь соразмерное повышение частоты разветвления. Таким образом, этот крахмал может также иметь распределение более коротких длин цепей амилопектина. Доля цепей крахмала, которые имеют степень полимеризации, которая попадает в диапазон от 6 до 11 остатков, может быть более 25%, более предпочтительно более 30%, и наиболее предпочтительно более 35%. Доля цепей крахмала, которые имеют степень полимеризации, которая попадает в диапазон 12-30 остатков, может быть менее 65%, более предпочтительно менее 60%, и наиболее предпочтительно менее чем примерно 55%. Доля цепей крахмала, которые имеют степень полимеризации, которая попадает в диапазон 31-60 остатков, может быть менее 10%, более предпочтительно менее 8%, но также предпочтительно более чем примерно 5%, и наиболее предпочтительно более чем примерно 6%. Скорее чем взятые индивидуально, комбинации долей трех диапазонов длин цепей можно рассматривать в качестве показателя того, что крахмал представляет собой крахмал того типа, который соответствует настоящему изобретению.
Понижение в распределении длин цепей, вероятно, способствует низким температурам желатинизации. Также считают, что уменьшенная длина цепи улучшает органолептические свойства крахмала, в частности, вкусовое ощущение, тем самым, возможно, способствуя однородности продукта. Кроме того, постулировано, что уменьшенная длина цепи амилопектина может снижать степень разрушения амилопектина, которое оказывает воздействие на качество пищи, например, его считают важным при черствении хлеба.
Кроме того, показано, что структура крахмала в крахмале, служащем примером, отличается тем, что степень кристалличности снижена по сравнению с нормальным крахмалом, выделенным из ячменя. При сочетании с пониженным распределением длин цепей амилопектина сниженная гранулярная кристалличность может указывать на то, что температура желатинизации будет ниже. Считают также, что сниженная кристалличность крахмала связана с улучшенными органолептическими свойствами и, как и меньшая длина цепи амилопектина, способствует более однородному вкусовому ощущению. Таким образом, крахмал может дополнительно проявлять сниженную кристалличность в результате пониженных уровней активности одного или более чем одного фермента синтеза амилопектина. Доля крахмала, проявляющего кристалличность, может быть менее чем примерно 20%, и предпочтительно менее чем примерно 15%.
Дополнительной мерой свойств настоящего крахмала является измерение вязкости. При использовании Rapid Visco Analyser (быстрого анализатора вязкости) обнаружено, что пиковая вязкость крахмала по данному изобретению значительно отличается от таковой у нормального и waxy крахмалов и высокоамилозных крахмалов, полученных из ячменя. Эти измерения проводили на муке из цельного зерна, однако, свойства крахмала в данных измерениях будут преобладать. Нормальный и waxy крахмалы имеют пиковую вязкость от примерно 900 до примерно 500 единиц RVA, известный высокоамилозный крахмал имеет пиковую вязкость более чем 200, тогда как крахмал растений ячменя по настоящему изобретению имеет пиковую вязкость менее чем 100, при большинстве менее чем примерно 50, в некоторых растениях столь низкую, как примерно 10 единиц RVA. Специалисту в данной области техники следует понимать, что упомянутые параметры представляют собой эмпирические единицы, и упомянутые результаты предназначены для указания относительной характеристики этих крахмалов в приборах RVA или подобных приборах, таких как амилограф.
Кроме сниженной кристалличности, объясненной выше, настоящий крахмал можно охарактеризовать наличием V-комплексной формы крахмала. Авторы изобретения считают, что впервые именно эту форму крахмала продемонстрировали в заметных количествах в крахмальных гранулах хлебного злака. Эту форму крахмала обычно связывают с ретроградированным крахмалом, в частности, если имеется контакт с липидами, В случае настоящего изобретения постулировано, что структура крахмала дает возможность образования тесной взаимосвязи между растительными липидами и крахмалом, которое приводит в результате к V-комплексной структуре. Считают, что данная форма крахмала может обладать пользой для здоровья, поскольку она обладает пониженной усваиваемостью и, следовательно, может вносить вклад в устойчивый крахмал.
Другие формы структуры могут также являться результатом взаимодействия липид-крахмал и включают в себя некристаллические комплексы липид-крахмал. Таким образом, можно также сказать, что данное изобретение относится к растению ячменя, показывающему заметные количества комплексов крахмал-липид в крахмальном содержимом эндосперма его зерна, являющиеся результатом пониженных уровней активности одного или более чем одного фермента синтеза амилопектина. Крахмалы, которые содержат комплексы крахмал-липид, включая те, которые проявляют V-комплексную структуру, также обычно устойчивы к ферментативному гидролизу и, следовательно, вносят вклад в уровни диетической клетчатки. Предпочтительно доля кристаллического крахмала, проявляющего форму кристалличности, характерную для комплекса крахмал-липид, выше чем примерно 50%, и более предпочтительно выше чем примерно 80%.
Крахмал, кроме наличия V-комплексной формы крахмала, может также не проявлять заметных количеств А-комплексных форм крахмала. Отсутствие А-комплекса можно рассматривать как показатель наличия крахмала по данному изобретению.
Обнаружено также, что температура пастирования крахмалов и продукта, полученных из зерна по данному изобретению, значительно повышена. Температуры пастирования известных крахмалов составляют менее чем 70°С, как для нормального, так и для высокоамилозного крахмалов. Крахмалы по настоящему изобретению, однако, предпочтительно проявляют температуры пастирования выше чем примерно 75°С, или более предпочтительно выше чем примерно 80°С. Следует отметить, что эти характеристики являются эмпирическими, и их можно рассматривать как относящиеся к такому измерению других крахмалов.
Обнаружено, что крахмал растения ячменя, служащего примером, имеет значительные количества диетической клетчатки и устойчивого крахмала, причем преимущественно это повышение является, по меньшей мере частично, результатом высокого относительного уровня амилозы, однако, вклад в диетическую клетчатку может также являться результатом комплексов крахмал/липид, включая V-комплекс, либо тесной связи амилозы или амилопектина с β-глюканом. Подобным образом, просто повышенный уровень β-глюкана может также вносить значительный вклад в повышение уровня диетической клетчатки.
Повышенные относительные уровни амилозы в эндосперме растения ячменя, служащего примером, по всей вероятности, являются результатом измененного продуцирования амилопектина в результате снижения уровня активности фермента SSII.
Можно ожидать, что мутации в гене, кодирующем этот фермент, проявляются в повышенном содержании амилозы и/или в снижении уровня амилопектина. В тех случаях, когда снижен только синтез амилопектина, крахмал проявляет повышенный относительный уровень амилозы.
Пониженной активности фермента синтеза амилопектина можно достичь в результате подходящих мутаций в пределах соответствующего гена или регуляторных последовательностей этого гена. Степень, до которой этот ген ингибирован, будет до некоторой степени определять характеристики получаемого крахмала. Мутации, служащие примером, по данному изобретению, представляющие собой мутации SSII ячменя, представляют собой мутанты усечения, и известно, что они оказывают значительное воздействие на природу крахмала, однако, измененная структура амилопектина также будет результатом мутанта с просачивающимся исходным фенотипом, который достаточно снижает активность фермента синтеза амилопектина, чтобы обеспечить интересующую характеристику крахмала или зерна ячменя. Другие хромосомные перестройки могут быть также эффективны, и они могут включать в себя делеции, инверсии, дупликации или точечные мутации.
Такие мутации можно вводить в желаемые генетические фоны либо путем мутагенеза интересующих сортов, но, что более надежно, путем скрещивания мутанта с растением желаемого генетического фона и проведения подходящего числа обратных скрещиваний для вытеснения исходно нежелательного родительского генетического фона. Выделения мутаций можно достичь с помощью отбора растений, подвергнутых мутагенезу.
В качестве альтернативы общепринятым способам можно предпринять молекулярно-биологический подход. Последовательность SSII представлена в данном описании. Векторы, несущие желаемые мутации и селектируемый маркер, можно вводить в культуры тканей растений или в подходящие растительные системы, такие как протопласты. Растения, где мутация интегрирована в хромосому с замещением существующего аллеля дикого типа, можно подвергать отбору с помощью, например, использования подходящего зонда, представляющего собой нуклеиновую кислоту, специфичную для этой мутации, и наблюдения фенотипа. Способы трансформации однодольных растений, таких как ячмень, и регенерации растений из протопластов или незрелых растительных эмбрионов, хорошо известны в данной области техники, смотри, например, Канадскую патентную заявку 2092588 Nehra, Австралийскую патентную заявку №61781/94 National Research Council of Canada, Австралийский патент №667939 Japan Tobacco Inc., Международную патентную заявку PCT/US97/10621 Monsanto Company, патент США 5589617, а другие способы изложены в WO99/14314.
Можно также предпринять другие известные подходы к изменению активности фермента синтеза амилопектина, иные чем использование мутаций. Так, например, это можно сделать путем экспрессии подходящих антисмысловых молекул, которые препятствуют транскрипции или процессингу гена или генов, кодирующих фермент синтеза амилопектина. Они могут быть основаны на последовательности ДНК, описанной здесь для гена SSII ячменя. Это могут быть антисмысловые последовательности для структурных генов или для последовательностей, которые осуществляют контроль над экспрессией гена или событием сплайсинга. На эти последовательности ссылки приведены выше. Способы разработки аинтисмысловых последовательностей хорошо известны в данной области техники, и их примеры можно найти, например, в патенте США 5190131, ЕР 0467349 A1, ЕР 0223399 A1 и в ЕР 0240208, которые включены здесь путем ссылки в том смысле, что в них приведены способы осуществления антисмысловых методик. Способы введения и сохранения таких последовательностей в растениях также опубликованы и известны.
Вариантом антисмысловой методики является использование рибозимов. Рибозимы представляют собой молекулы РНК с ферментативной функцией, которая может расщеплять другие молекулы РНК в специфичных сайтах, определенных антисмысловой последовательностью. Расщепление РНК блокирует экспрессию гена-мишени. Ссылка дана на ЕР 0321201 и WO 97/45545.
Другим молекулярно-биологическим подходом, который также можно использовать, является косупрессия. Механизм косупрессии не полностью понятен, но он включает в себя внесение в растение экстра-копии гена в нормальной ориентации. В некоторых случаях эта дополнительная копия гена препятствует экспрессии гена-мишени растения. На способы осуществления технологии косупрессии сделана ссылка на WO 97/20936 и ЕР 0465572.
Следующим способом, который можно применять, используя последовательности ДНК, является генная супрессия, опосредованная дуплексной или двунитевой РНК. При данном способе используют ДНК, которая направляет синтез двунитевого продукта РНК. Присутствие этой двунитевой молекулы запускает ответ из защитной системы растения, который разрушает и двунитевую РНК, и также РНК, происходящую от гена-мишени растения, эффективно снижая или элиминируя активность этого гена-мишени. На способы применения этой методики сделана ссылка на Австралийский патент 99/292514-А и WO 99/53050.
Следует понимать, что данное изобретение вероятно является результатом понижения уровней активности двух или более чем двух из указанных выше генов при использовании молекулярно-биологического подхода.
Одним из важных продуктов, который можно рассматривать, в частности, как результат высокого содержания амилозы и высокого содержания β-глюкана, является низкокалорийный продукт с пониженным гликемическим индексом. Низкокалорийный продукт может быть основан на включении муки, полученной из измельченного зерна. Однако может быть желательно сначала переработать это зерно в перловую крупу, удаляя, возможно, 10% или 20% масс/масс зерна, тем самым удаляя алейроновый слой, и при большем измельчении удаляя также зародыш. Эффект стадии переработки ячменя в перловую крупу состоит в снижении содержания липида и, следовательно, в снижении калорийности пищевого продукта. Такие пищевые продукты будут обладать эффектом насыщения, улучшения здоровья толстого кишечника, снижения сывороточной концентрации глюкозы и липида после приема пищи, а также обеспечения низкокалорийного пищевого продукта. Использование продукта в виде перловой крупы приведет в результате к уменьшению питательной ценности, обеспечиваемой алейроновым слоем и зародышем. Мука, полученная из продукта в виде перловой крупы, вероятно, имеет улучшенный внешний вид, поскольку продукт, полученный таким способом, имеет тенденцию к белому цвету.
Аспекты данного изобретения также являются результатом комбинации алейронового слоя и зародыша в сочетании с высокими уровнями диетической клетчатки. Конкретно это является результатом несколько более высоких относительных уровней алейрона или зародыша, имеющихся в зерне, служащем примером. Во-первых, ячмень имеет значительно более высокий алейроновый слой, чем другие имеющиеся в продаже хлебные злаки, в результате наличия трехклеточного алейронового слоя. Во-вторых, зерно ячменя, служащее примером, также является сморщенным, что означает, что эндосперм присутствует в сниженных количествах, следствием чего является то, что алейроновый слой и зародыш присутствуют в повышенных относительных количествах. Таким образом, ячмень имеет относительно высокий уровень некоторых полезных элементов или витаминов в сочетании с системой доставки устойчивого крахмала, и такие элементы включают в себя двухвалентные катионы, такие как биологически доступный Са++, и витамины, такие как фолат, или антиоксиданты, такие как токоферолы и токотриенолы. Таким образом, установлено, что кальций обеспечивает материал для роста и образования кости и другой кальцинированной ткани и снижает риск остеопороза более поздних периодов жизни. Обнаружено, что фолиевая кислота обладает защитным эффектом против пороков нервной трубки при разумном употреблении и снижает риск сердечно-сосудистого заболевания, тем самым усиливая эффекты сочетания устойчивого крахмала и β-глюкана. Также считают, что фолиевая кислота обладает эффектом снижения риска некоторых видов рака. Токоферол и токотриенолы приносят пользу в качестве антиоксидантов, и считают, что они снижают риск рака и сердечного заболевания, а также они обладают эффектом снижения нежелательных эффектов окисления компонентов пищи, таких как жирные кислоты, которые могут привести к прогорклости. В то время как эти компоненты данной предпочтительной формы зерна ячменя или продукты, полученные из них, удобно помещены в одном зерне. Одной из конкретных форм измельченного продукта может быть такая форма, где алейроновый слой включен в этот измельченный продукт. Конкретный процесс измельчения можно осуществить так, чтобы увеличить количество алейронового слоя в измельченном продукте. На такой способ сделана ссылка на Fenech et al. ((1999), J. Nutr, 129: 1114-1119). Таким образом, любой продукт, полученный из зерна, измельченного или иначе обработанного для включения алейронового слоя и зародыша, будет иметь дополнительную питательную пользу, не требуя добавления этих элементов из отдельных источников.
Следует понимать, что растение ячменя по настоящему изобретению предпочтительно представляет собой растение, имеющее зерно, которое является полезным для получения пищевых продуктов и, в частности, для промышленного получения пищевых продуктов. Такое получение может включать в себя получение муки или другого продукта, который может представлять собой ингредиент в промышленном получении пищевых продуктов. Более низким уровнем пригодности может быть содержание крахмала выше чем примерно 12%, или, возможно, выше чем примерно 15%. Или, подобным образом, он может включать в себя способность к измельчению зерна; таким образом, хотя ячмень в виде перловой крупы можно производить из большинства форм зерна, некоторые конфигурации зерна особенно устойчивы к измельчению. Другой характеристикой, которая может влиять на разнообразие продукции промышленно применимого зерна, является обесцвечивание произведенного продукта. Таким образом, если шелуха или другая часть зерна проявляет значительное окрашивание, например пурпурное, оно будет проявляться через весь продукт и ограничивать его промышленные применения только до таких областей применения, как компонент хлеба, содержащего окрашенные цельные или дробленые зерна. Как правило, также удобнее, чтобы растения ячменя были беспокровным, поскольку наличие шелухи на зернах ячменя вводит значительные трудности при обработке зерна. Другим аспектом, который может придать растению ячменя более высокую ценность, является аспект на основе экстракции крахмала из зерна, причем более высокие степени экстракции более полезны. Форма зерна также является еще одним признаком, который может влиять на промышленную пригодность растения, таким образом, форма зерна может влиять на легкость или что-либо другое, с помощью чего это зерно можно измельчить, так, например, зерно растения ячменя МК6827 имеет необычно очень продолговатую морфологию зерна, которая затрудняет его измельчение и обработку. Удобной характеристикой этой продолговатой формы и применимости является отношение двух морфологических характеристик - длины зерна к толщине зерна (отношение L/T). Это отношение часто обусловлено природой крахмала. Авторами изобретения обнаружено, что МК6827 имеет отношение L/T больше 6. Растения ячменя, подвергнутые такому отбору, несущие мутантный ген SSII, имеют отношение L/T, находящееся в диапазоне от примерно 4 до примерно 5, хотя ожидают, что его можно расширить даже до большего диапазона, чтобы они все же были полезны, возможно, менее чем примерно 5,8 или по меньшей мере 5,5.
Желаемый генетический фон будет включать в себя соображения промышленного выхода и другие характеристики. Такие характеристики могут включать в себя, желательно ли иметь озимый или яровой тип ячменя, агрономическую характеристику, устойчивость к болезням и устойчивость к абиотическому стрессу. В Австралии может быть желательно скрещивать с сортами ячменя, такими как Sloop, Schooner, Chebec, Franklin, Arapiles, Tantangara, Galleon, Gairdner или Picolla. Представленные примеры являются конкретными для Австралийского промышленного района, а для других районов выращивания могут быть подходящими другие сорта.
Более наполненное зерно может быть желательным в отношении достижения более высоких выходов и некоторых преимуществ, которые можно достичь данным изобретением, таких как производство крахмала с высокими уровнями амилозы или варианта крахмала с измененными распределениями длин цепей. Другие аспекты данного изобретения могут, однако, быть лучше достигнуты с помощью зерна, которое является менее наполненным. Таким образом, доля алейронового слоя или зародыша в отношении к крахмалу может быть выше в менее наполненном зерне, тем самым обеспечивая ячменную муку или другой продукт, который имеет больше полезных составных частей алейронового слоя. Продукт с высоким алейроновым слоем может, таким образом, иметь более высокое содержание некоторых витаминов, таких как фолат, или более высокое содержание некоторых минералов, таких как кальций, и это в сочетании с более высокими уровнями устойчивого крахмала и/или более высокими уровнями β-глюкана может обеспечить синергические эффекты, такие как обеспечение усиленной абсорбции минералов в толстом кишечнике.
Чтобы максимизировать количество амилозы, для растения ячменя также может быть желательно иметь другие фенотипические характеристики в дополнение к пониженной активности одного или более чем одного фермента синтеза амилопектина. Генетический фон может, следовательно, включать в себя дополнительно высокоамилозный фенотип, например, мутацию amo 1 в АС38 (причинный ген неизвестен) и мутацию waxy (обнаруженную, например, в сорте Waxiro). Дополнительно может быть желательно получать двойные мутации у других доступных мутантов ячменя со сморщенными эндоспермами, где причинный ген неизвестен.
В следующем аспекте данное изобретение относится к зерну, полученному из растения ячменя, ссылаясь на данное описание.
Следует также понимать, что в объем данного изобретения входит обработанное зерно, включая измельченное, размолотое, дробленое, в виде перловой крупы или зерновых хлопьев, либо продукт, полученный из обработанного или цельного зерна растения ячменя, на которое ссылаются выше, включая муку. Эти продукты затем можно использовать в различных пищевых продуктах, например в мучных продуктах, таких как хлеба, пирожные, бисквиты и тому подобное, либо в пищевых добавках, таких как загустители, либо для получения солодовых или других ячменных напитков, лапши и супов быстрого приготовления.
Альтернативно, в объем данного изобретения входит крахмал, выделенный из зерна растения ячменя, на которое ссылаются выше. Крахмал можно выделить с помощью известных методик.
Следует понимать, что одним из преимуществ настоящего изобретения является то, что в нем предложен один или более чем один продукт, который обладает особенной питательной ценностью, и, кроме того, это сделано без необходимости модифицировать крахмал или другие составные части зерна ячменя.
Однако может быть желательно создавать модификации крахмала, β-глюкана или другой составной части зерна, и в объем изобретения входит такая модифицированная составная часть.
Способы модификации известны и включают в себя экстракцию крахмала или β-глюкана, либо другой составной части общепринятыми способами и модификацию крахмалов до получения желаемой устойчивой формы.
Таким образом, крахмал или β-глюкан можно модифицировать либо однократно, либо многократно посредством использования обработки, выбранной из группы, включающей в себя, но не ограниченной ими, нагревание и/или увлажнение, физическая обработка (например, дробление на шаровой мельнице), ферментативный (при использовании, например, α- или β-амилазы, пуллаланазы или тому подобного), химический гидролиз (влажный или сухой при использовании жидких или газообразных реагентов), окисление, поперечное связывание с дифункциональными реагентами (например, триметафосфатом натрия, оксихлоридом фосфора) или карбоксиметилирование.
Содержание диетической клетчатки зерна ячменя, служащего примером, не является исключительно результатом повышенного относительного содержания амилозы в эндосперме. Одной из основных причин является то, что β-глюкан присутствует в повышенных уровнях и вносит значительный вклад в уровень диетической клетчатки. Вероятно, также имеются защитные эффекты вследствие связывания β-глюкана с крахмальной гранулой, причем прочность этой связи потенциально обеспечивает защитный эффект для крахмала, чтобы посредством этого обеспечить устойчивость, которую можно охарактеризовать как RS1 форму, которая до некоторой степени недоступна ферментативному гидролизу. Подобным образом наличие связи крахмал-липид, как измерено на основании V-комплексной кристалличности, также, вероятно, вносит вклад в уровень устойчивого углевода. В данном случае устойчивость, вероятно, возникает вследствие физической недоступности в связи с присутствием липида и, соответственно, ее можно рассматривать как RS1 крахмал. Таким образом, известно, что ретроградированный крахмал, который принимает V-комплексную конфигурацию, высоко устойчив к ферментативному гидролизу и, соответственно, ожидают, что амилопектин, который образует часть крахмальной гранулы, имеющей V-комплексную кристаллическую структуру, будет обладать повышенной устойчивостью к ферментативному гидролизу. В-третьих, крахмал растения ячменя, служащего примером, может быть устойчив к ферментативному гидролизу вследствие структуры крахмальной гранулы и, соответственно, может иметь RS2 крахмал.
Следует понимать, что, хотя различные указания даны как на аспекты настоящего изобретения, данное изобретение может относиться к комбинациям двух или более чем двух аспектов настоящего изобретения.
ПРИМЕР 1
Предпосылки
Синтез крахмала в эндосперме высших растений осуществляется рядом ферментов, которые катализируют четыре ключевые стадии. Во-первых, АДФ-глюкозопирофосфорилаза активирует мономерный предшественник крахмала посредством синтеза АДФ-глюкозы из G-1-P и АТФ. Во-вторых, активированный донор гликозила, АДФ-глюкоза, переносится к невосстанавливающему концу ранее существовавшей α-1-4 связи синтазами крахмала. В-третьих, ферменты разветвления крахмала вводят точки разветвления посредством расщепления участка α-1,4-связанного глюкана с последующим переносом отщепленной цепи на акцепторную цепь с образованием новой α-1,6 связи. Наконец, генетические исследования показывают, что ферменты удаления разветвления цепи крахмала существенны для синтеза нормальных количеств крахмала у высших растений, однако, механизм, посредством которого действуют ферменты удаления разветвления цепи, не решен (Myers et at., 2000).
Хотя ясно, что по меньшей мере эти четыре активности необходимы для синтеза нормальной крахмальной гранулы у высших растений, в эндосперме высших растений обнаружены множественные изоформы каждой из этих четырех активностей, и для индивидуальных изоформ предположили конкретные роли на основании мутационного анализа (Wang et al., 1998, Buleon et al., 1998) или посредством модификации уровней генной экспрессии при использовании трансгенных подходов (Abel et al., 1996, Jobling et al., 1999, Scwall et al., 2000). Однако точные вклады каждой изоформы каждой активности в биосинтез крахмала до сих пор неизвестны, и неизвестно, различаются ли значительно эти вклады между видами. В эндосперме хлебных злаков присутствуют две изоформы АДФ-глюкозопирофосфорилазы, одна форма внутри амилопласта и одна форма в цитоплазме (Denyer et al., 1996, Thorbjornsen et al., 1996). Каждая форма состоит из двух типов субъединиц. Мутанты shrunken (sh2) и brittle (bt2) кукурузы означают повреждения в большой и малой субъединицах, соответственно (Girouz and Hannah, 1994). В эндосперме хлебных злаков обнаружены четыре класса синтазы крахмала, изоформа, исключительно локализованная внутри крахмальной гранулы, гранулосвязанная синтаза крахмала (GBSS, granule-bounded starch synthase), две формы, которые распределены между гранулой и растворимой фракцией (SSI, Li et al., 1999a, SSII, Li et al., 1999b) и четвертая форма, которая полностью локализована в растворимой фракции, SSIII (Сао et al., 2000, Li et al., 1999b, Li et al., 2000). Показано, что GBSS существенна для синтеза амилозы (Shure et al., 1983), и показано, что мутации в SSII и SSIII изменяют структуру амилопектина (Gao et al., 1998, Craig et al., 1998). Мутации, определяющие роль активности SSI, не описаны.
В эндосперме хлебных злаков экспрессируются три формы разветвляющего фермента, разветвляющий фермент I (BEI, branching enzyme I), разветвляющий фермент IIa (BEIIa) и разветвляющий фермент IIb (BEIIb) (Hedman and Boyer, 1982, Boyer and Preiss, 1978, Mizuno et al., 1992, Sun et al., 1997). Показано, что у кукурузы и риса высокоамилозные фенотипы являются результатом повреждений в гене BEIIb (Boyer and Preiss, 1981, Mizuno et al., 1993). У этих мутантов содержание амилозы значительно повышено, а частота разветвлений остаточного амилопектина снижена. Кроме того, имеется значительный пул вещества, которое определяют как "промежуточное" между амилозой и амилопектином (Boyer et al., 1980, Takeda et al., 1993). Мутации, определяющие роли BEIIa и BEI, еще нужно описать, несмотря на то, что у картофеля негативная регуляция одного BEI оказывает минимальные воздействия на структуру крахмала (Filpse et al., 1996). Однако у картофеля комбинация негативной регуляции BEII и BEI дает намного более высокое содержание амилозы, чем негативная регуляция одного BEII (Schwall et al., 2000). Два типа ферментов, удаляющих разветвление цепи, присутствуют у высших растений и определены на основании их субстратной специфичности, ферменты, удаляющие разветвление цепи, типа изоамилазы и ферменты, удаляющие разветвление цепи, типа пуллуланазы (Myers et al., 2000). Мутации Sugary-1 у кукурузы и риса связаны с недостаточностью обоих ферментов, удаляющих разветвление цепи (James et al., 1995, Kubo et al., 1999), однако, причинная мутация картирована в той же локализации, что и ген фермента, удаляющего разветвление цепи, типа изоамилазы. У мутанта Chlamydomonas sta-7 (Mouille et al., 1996), аналога мутации sugary-1 кукурузы, имеется негативная регуляция одной изоамилазной активности.
Известное варьирование структуры крахмала ячменя ограничено по сравнению с варьированием, доступным у кукурузы. Большинство высоко охарактеризованных мутаций представляют собой waxy и высокоамилозную мутацию, идентифицированную как АС38. Двойные мутанты также сконструированы и проанализированы (Schondelmaier et al., 1992, Fujita et al., 1999). Сообщают о широком диапазоне характеристик варьирования структуры и свойств крахмала (Czuchajowska et al., 1992; Schondelmaier et al., 1992; Vasanthan and Bhatty, 1995; Morrison et al., 1984; Gerring and DeHaas, 1974; Bankes et al., 1971; Persson and Christerson, 1997; Vasanthan and Bhatty, 1998; Czuchajowska et al., 1998; Song and Jane, 2000; Andreev et al., 1999; Yoshimoto et al., 2000), а также свойствах зерна (Swantson, 1992, Ahokas, 1979; Oscarsson et al., 1997; Oscarsson et al., 1998; Andersson et al., 1999; Elfverson et al., 1999; Bhatty, 1999; Zheng et al., 2000; Izydorczyk et al., 2000; Andersson et al., 2000), и исследована полезность этих мутантов в опытах кормления животных (Xue et al., 1996; Newman et al., 1978; Calvert et al., 1976; Wilson et al., 1975; Sundberg et al., 1998; Bergh et al., 1999), продуктов питания человека (Swantson et al., 1995; Fastnaught et al., 1996; Persson et al., 1996; Pomeranz et al., 1972) и питания человека (Pomeranz 1992; Granfeldt et al., 1994; Oscarsson et al., 1996; Akerberg et al., 1998).
В настоящем примере авторы изобретения выделили новый класс высокоамилозного мутанта из ячменя. Эти мутантные линии имеют содержание амилозы (65-70%), выше чем известно у хорошо охарактеризованного мутанта High Amylose Glacier (AC38) (45-48%) (Walker et al., 1968), и имеют крахмал со структурой амилопектина, в которой имеется повышение частоты разветвлений крахмала, что противоположно сниженной частоте разветвлений, связанной с мутантом amylose extender у кукурузы (Takeda et al., 1993).
Характеристики зерна и крахмала настоящего мутанта подробно исследованы, и картирована причинная мутация. Выделенные мутации аллельны известной ранее мутации shrunken у ячменя, sex6, и показано, что причинная мутация локализована внутри гена синтазы крахмала II. Эффекты этой мутации проливают новый свет на процесс биосинтеза крахмала и иллюстрируют, как мутации в специфичных генах могут иметь различные воздействия на структуру крахмала от вида к виду.
Материалы и методы
Мутагенез и отбор
Сорт беспокровного ячменя "Himalaya" подвергали мутагенезу, используя азид натрия, согласно Zwar and Chandler (1995). Селекцию вариантов с измененной морфологией зерна проводили согласно Green et al. (1997). Идентифицировали и поддерживали всего 75 линий с фенотипами сморщенного эндосперма согласно Green et al. (1997).
Выделение крахмала
Крахмал выделяли из зерна ячменя, используя способ Schulman et al. (1991).
Методы определения амилозы
Определение отношения амилоза/амилопектин по методу ВЭЖХ для разделения крахмалов с неразветвленными цепями и по методу связывания йода проводили, как описано Batey and Curtin (1996). Анализ отношения амилоза/амилопектин путем анализа крахмалов с разветвленными цепями проводили согласно Case et al. (1998).
Измерение содержания крахмала
Крахмал определяли, используя набор для анализа суммарного крахмала, поставляемый Megazyme (Bray, Co Wicklow, Republic of Ireland).
Содержание белка
Азот определяли методом Къельдаля, и содержание белка вычисляли, используя фактор 5,7.
Уровни β-глюкана
β-Глюкан определяли, используя набор, поставляемый Megazyme (Bray, Со Wicklow, Republic of Ireland).
Распределение длин цепей крахмала
Крахмалы подвергали удалению разветвления цепи, и распределения длин цепей анализировали, используя электрофорез углеводов с помощью флуорофора (FACE, fluorophore assisted carbohydrate electrophoresis), используя капиллярный электрофорез согласно Morell et al. (1998).
ДСК
Желатинизацию измеряли в дифференциальном сканирующем калориметре Pyris 1 (Perkin Elmer, Norwalk CT, USA). Крахмал смешивали с водой в соотношении 2 части воды: 1 часть крахмала, и эту смесь (40-50 мг, точно взвешенную) помещали в чашу из нержавеющей стали и герметично закрывали. Образец сканировали при 10°С в течение минуты от 20°С до 140°С с пустой чашей из нержавеющей стали в качестве сравнения. Температуры желатинизации и энтальпию определяли, используя программное обеспечение Pyris.
RVA-анализ
Вязкость измеряли на Rapid-Visco-Analyser (RVA, Newport Scientific Pty Ltd, Warriewood, Sydney), используя условия, как описано Batey et al., 1997, для непросеянной муки. Чтобы ингибировать α-амилазы, во все анализы включали нитрат серебра в концентрации 12 мМ. Измеряемыми параметрами были пиковая вязкость (максимальная вязкость горячей пасты), сила удерживания, конечная вязкость и температура пастирования. Кроме того, вычисляли разложение (пиковая вязкость минус сила удерживания) и задержку (конечная вязкость минус сила удерживания).
Набухание муки
Объем набухания муки определяли согласно способу Konik-Rose et al. (2001).
Данные рентгеновского анализа
Данные дифракции рентгеновских лучей собирали, используя стандартные методики (Buleon et al., 1998).
Сканирующая электронная микроскопия
Сканирующую электронную микроскопию проводили на оборудовании Joel JSM 35C. Очищенные крахмалы покрывали распылением золота и сканировали при 15 кВ при комнатной температуре.
Получение двойных гаплоидов
Двойные гаплоиды получали из растений F1, имеющих происхождение от скрещиваний между 292 и Hordeum vulgare cv Tantangara и между 342 и Н.vulgare cv Tantangara Dr. P.Davies, Waite Institute, Adelaide, Australia.
Анализ сцепления
Данные генетического сцепления вычисляли, используя MapManager.
Конструирование библиотеки кДНК ячменя
Пять мг полиА+мРНК из тканей эндосперма ячменя на 10, 12 и 15 сутки после опыления использовали для синтеза кДНК согласно протоколам (Life Technology). Для синтеза первой нити кДНК использовали праймер Notl-(dT) 18 (Pharmacia Biotech). Двунитевые кДНК лигировали с адаптером Sa/I-XhoI (Stratagene) и клонировали к плечам Sa/I-NotI ZipLox (Life Technology) после ферментативного гидролиза кДНК с использованием NotI с последующим фракционированием по размеру (колонка SizeStep 400 spun от Pharmacia Biotech). Лигированные кДНК упаковывали с упаковочным экстрактом Gigapack III Gold (Stratagene). Титр библиотеки, тестированный штаммом Е.coli Y1090(ZL), составлял 2×106 б.о.е. (бляшкообразующих единиц).
Клонирование специфичных участков кДНК синтазы крахмала II ячменя с использованием ПЦР
Клон кДНК wSSIIp1 использовали для скрининга библиотеки кДНК ячменя. Этот клон кДНК wSSIIp1 получили с помощью ПЦР, используя праймеры ssIIa (TGTTGAGGTTCCATGGCACGTTC SEQ ID NO 9) и ssIIb (AGTCGTTCTGCCGTATGATGTCG SEQ ID NO 10), амплифицируя участок между положениями нуклеотидов 1435 и 1835 wSSIIA (GenBank номер по каталогу AF 155217).
Амплификацию проводили, используя термосеквенатор FTS-1 (Corbett, Australia) в течение 1 цикла при 95°С по 2 минуты; 35 циклов при 95°С по 30 секунд, при 60°С по 1 минуте, при 72°С по 2 минуты и 1 цикла при 25°С в течение 1 минуты. Фрагмент wSSIIp1 клонировали в вектор pGEM-T (Promega).
Скрининг библиотеки кДНК ячменя
Скрининг библиотеки кДНК, сконструированной из РНК из эндосперма ячменя cv Himalaya, проводили фрагментом кДНК 347 п.н., wSSIIp1, при таких условиях гибридизации, которые описаны ранее (Rahman et al., 1998). Гибридизацию проводили в 50% формамиде, 6х SSPE, 0,5% ДСН, 5х растворе Денхардта и 1,7 мкг/мл ДНК спермы лосося при 42°С в течение 16 ч, затем отмывали 3х 2х SSC, содержащим 0,1% ДСН, при 65°С в течение 1 часа на отмывку.
Скрининг геномной библиотеки ячменя
Геномную библиотеку ячменя (ячмень cv Morex) конструировали и подвергали скринингу по существу так, как описано в Gubler et al (2000), используя кДНК SSII ячменя в качестве зонда.
Секвенирование геномных клонов
Ген SSII Morex субклонировали в плазмидные вектора и секвенировали. Гены 292 и МК6827 секвенировали с помощью ПЦР-амплификации перекрывающихся участков этого гена, используя праймеры, сконструированные на основе последовательности Morex. Фрагменты ПЦР либо секвенировали непосредственно, либо субклонировали и секвенировали из плазмид.
Идентификация экспрессируемых участков
Районы геномных последовательностей 292 и МК6827, предположительно присутствующие в кДНК, определили в сравнении с последовательностью кДНК Himalaya и геномной последовательностью Morex.
ПЦР-анализ мутации G на А в гене SSII
Сконструировали праймеры ПЦР, которые амплифицируют участок, содержащий транзицию G на А, идентифицированную в 292. Последовательности праймеров представляют собой: ZLSS2P4 (CCTGGAACACTTCAGACTGTACG SEQ ID NO 11) и ZLBSSII5 (CTTCAGGGAGAAGTTGGTGTAGC SEQ ID NO 12). Амплификацию проводили, используя термосеквенатор FTS-1 (Corbett, Australia) в течение 1 цикла при 95°С в течение 2 минут; 35 циклов при 95°С по 30 секунд, при 60°С по 1 минуте, при 72°С по 2 минуты и 1 цикла при 25°С в течение 1 минуты.
Анализ белков эндосперма ячменя в ДСН-ПААГ электрофорезе
Крахмал получали из развивающегося и зрелого эндосперма ячменя и пшеницы, и поверхностные белки удаляли с помощью протеиназы К, как описано (Rahman et al., 1995). Белки крахмальной гранулы экстрагировали из 20 мг крахмала (сухая масса), используя 0,5 мл экстрагирующего буфера, содержащего 50 мМ Трис рН 6,8, 10% ДСН и 10% 2-меркаптоэтанол. После желатинизации кипячением в течение 10 мин и сбора крахмала центрифугированием 15 микролитров супернатанта наносили на каждую дорожку.
Получение двойных гаплоидов
Двойные гаплоиды получали из растений F1, имеющих происхождение от скрещиваний между 292 и Hordeum vulgare cv Tantangara и между 342 и Н.vulgare cv Tantangara Dr. P.Davies, Waite Institute, Adelaide, Australia.
Стратегия обратных скрещиваний
Проводили скрещивания между 292 и Hordeum vutgare cv Sloop для получения посевного материала F1. Растения, выращенные из посевного материала F1, подвергали самоопылению с получением популяции посевного материала F2. Растения, выращенные из этого посевного материала F2, тестировали, используя ПЦР-анализ, и растения, гомозиготные по мутации 292, подвергали обратному скрещиванию со Sloop (BC1). Растения F1, полученные в результате BC1, снова тестировали с помощью ПЦР, и растения, гетерозиготные по мутации 292, отбирали и подвергали обратному скрещиванию со Sloop (BC2). Растения F1, полученные в результате ВС2, снова анализировали с помощью ПЦР и отбирали растения, гетерозиготные по мутации 292. Эти растения либо подвергали самоопылению с получением популяции BC2F2, либо снова скрещивали со Sloop (BC3). Растения F1, полученные в результате BC3, снова анализировали с помощью ПЦР и отбирали растения, гетерозиготные по мутации 292. Эти растения подвергали самоопылению с получением популяции BC3F2. Растения, выращенные из этого посевного материала, тестировали с помощью ПЦР, и растения, гомозиготные по мутации 292, отбирали для получения поколения одного семени и размножения семян.
Результаты
Отбор мутантов
Об идентификации ряда мутантов у сорта ячменя "Himalaya" без шелухи или с беспокровными семенами, индуцированных обработкой азидом натрия, ранее сообщали Zwar and Chandler (1995). Группа из 75 мутантов со сморщенным зерном идентифицирована авторами изобретения, и амилозное содержание крахмала из этого сморщенного семени определили с помощью ВЭЖХ (Фиг.1). Обнаружено, что две линии, 292 и 342, имели содержание амилозы 71 и 62,5%, соответственно (Табл. 1). Содержание амилозы 292 и 342 было существенно выше, чем в линии АС38, ранее хорошо охарактеризованной (47% амилозы, см. Табл. 1). В данном исследовании определили генетическую основу нового высокоамилозного фенотипа, проявляемого 292 и 342, и описали воздействия причинной мутации на структуру и функциональность зерна и крахмала.
Характеристики зерна
Размер и морфология зерна
Отмечены влияния мутации на массу и морфологию зерна (Табл. 2). Масса зерна снижена с 51 мг для родительской линии Himalaya до 32 мг для 292 и 35 мг для 342. У этих мутантов сохранились длина и ширина как у дикого типа, но в сравнении зерна являются расплющенными (от 2,82 мм средней толщины у Himalaya до 1,58 и 1,75 мм у 292 и 342, соответственно) и имеют по существу незаполненный центральный участок. На Фиг.2 показаны фотографии зерна мутанта и дикого типа. Размеры зерна измеряли традиционным путем, длину зерна (L), ширину зерна в самой широкой точке (W) и толщину (Т), как указано на Фиг.2. Отношение длины (L) к толщине (Т) является полезным диагностическим признаком для этой мутации при значениях >3,5, типично обнаруженных для семян, несущих мутации 292 или 342, и значениях <3,5 для немутантных растений ячменя.
Состав зерна:
Содержание крахмала мутантных линий понижено от 49,0% для Himalaya до 17,7 и 21,9% для 292 и 342, соответственно (см. Табл. 1). Вычитание массы крахмала из общей массы зерна с получением суммарного некрахмального содержимого зерна показало, что потеря содержания крахмала происходит за счет потери массы зерна при массах некрахмального содержимого 26,0, 26,3 и 27,3 мг для Himalaya, 292 и 342, соответственно.
Содержание белка 292 и 342 повышено относительно родительской линии Himalaya (Табл. 1), однако, этот эффект имеет место за счет потери крахмала из зерна и не связан с каким-либо повышением синтеза белка на зерновку.
Уровни β-глюкана мутантов 292 и 342 также повышены и являются более высокими, чем ожидалось бы в результате эффекта снижения содержания крахмала (Табл. 1). В обоих случаях содержание β-глюкана повышено примерно на 20% на зерновку, что, возможно, представляет собой переключение небольшой части входящего углерода от синтеза крахмала на синтез β-глюкана.
Состав и функциональность крахмала
Содержание амилозы и амилопектина
Содержание амилозы определяли, используя две методики, во-первых, вытеснительную по размеру ВЭЖХ в 90% (об/об) ДМСО и, во-вторых, показатель синего окрашивания йодом. Содержание амилозы, определенное каждым методом, было аналогичным, а данные ВЭЖХ представлены в Табл. 1.
На основании данных по массе зерна и содержанию амилозы для мутантных линий и линий дикого типа можно произвести вычисления количества запасаемой амилозы на зерно. Этот анализ показывает, что имеется снижение количества амилозы на зерно от 6,2 мг/зерновка у Himalaya до 4,0 мг/зерновка у 292 и 4,8 мг/зерновка у 342. Напротив, имеется резкое снижение синтеза амилопектина на зерновку от 18,7 мг у Himalaya до 1,6 мг у 292 и 2,9 мг у 342.
Распределение длин цепи
Распределение длин цепи крахмала после удаления разветвления цепи изоамилазой проводили, используя электрофорез углеводов с помощью флуорофора (FACE). Распределение длин цепи мутантов 292 и 342 и Himalaya показано на Фиг.3а. На Фиг.3б показан график разностей, на котором нормализованные распределения длин цепей для мутантов 292 и 342 вычитают из нормализованного распределения Himalaya. Проценты длин цепей из DP 6-11, DP 12-30 и DP 31-65 вычислены и представлены в Табл. 3. Имеется заметный сдвиг в распределении длин цепей у мутантов 292 и 342, такой что на участке из DP 6-11 процент цепей более высок по сравнению с DP 12-30.
Дифференциальная сканирующая калориметрия
Температуру желатинизации мутантов исследовали, используя дифференциальную сканирующую калориметрию, и данные представлены в Табл. 4. Как 292, так и 342 дают крахмалы, которые имеют значительно более низкие температуры желатинизации, чем крахмалы Himalaya, в отношении начальной, пиковой и конечной температур для пика желатинизации. Энтальпия для пика желатинизации для мутантов 292 и 342 также резко снижена по сравнению с диким типом. Начальная температура пика амилоза/липид также снижена для мутантов 292 и 342, однако, энтальпия повышена, что соответствует повышенному содержанию амилозы у мутантов.
Вязкость крахмала по RVA
RVA-анализ образцов непросеянной ячменной муки проводили, чтобы изучить их вязкость пастирования. Предшествующие исследования показали, что анализ образцов непросеянной муки имеет строгую корреляцию с анализом выделенных крахмалов (Batey et al., 1997). Этот анализ показал, что существуют большие различия между исследованными генотипами ячменя (см. табл. 5 и фиг.4). Два сорта ячменя, содержащих крахмал дикого типа, Himalaya и Namoi, показали типичные профили RVA, в которых наблюдался выступающий пик вязкости с последующим снижением вязкости до силы удерживания с последующим повышением вязкости по мере снижения температуры до конечной вязкости. Как обычно наблюдают для крахмалов ячменя, конечные вязкости для крахмалов дикого типа были эквивалентны пиковым вязкостям или ниже их (Табл. 5). У АС38 получили выдающийся пик вязкости, однако, в связи с повышенным содержанием амилозы у этой линии полученная конечная вязкость была выше, чем пиковая вязкость. Однако у 292, 342 и МК6827 получили сильно отличающийся профиль. У других крахмалов ячменя не было получено заметного начального повышения вязкости, соответствующего пиковой вязкости, и, следовательно, нельзя было вычислить никакого показателя разрушения. Значения для пиковой вязкости, данные в Табл. 5 для 292, 342 и МК6827, представляли собой вязкости, зарегистрированные в момент времени пиковой вязкости для Himalaya. У 292, 342 и МК6827 вязкость повышалась на протяжении всего анализа с достижением конечной вязкости, сравнимой с другими образцами непросеянной муки. При нормализации на основании содержания крахмала крахмалы 292 и 342 имели очень высокие конечные вязкости (см. Табл. 5).
Объем набухания представляет собой метод измерения свойств муки и крахмала, с помощью которого исследуют поведение этого материала при воздействии нагревания и избытка воды. Повышенное поглощение воды измеряют с помощью взвешивания образца перед смешиванием и после смешивания этого образца с водой при определенных температурах и последующего сбора желатинизированного материала. Этот анализ показал, что контрольные образцы, Himalaya и Tantangara, набухают в 6-8 раз от их сухой массы, напротив, 292 и 342 набухают только до 2-3 раз от их сухой массы (Табл. 9).
Кристалличность
Структуру крахмалов далее исследовали с помощью рентгеновской кристаллографии (см. Табл. 6 и Фиг.5). Himalaya показал ожидаемую картину для крахмала хлебных злаков, имеющего преимущественно кристалличность "А" типа, и как АС38, так и Waxiro показали очень сходные картины дифракции рентгеновских лучей, хотя уровни кристалличности были ниже для АС38 и выше для Waxiro. Для мутантов 292 и 342 картина дифракции рентгеновских лучей была сдвинута до смеси V и В картины. Кроме сдвига картины дифракции, количество кристалличности было резко снижено у мутантов 292 и 342 до 9 и 12%, соответственно. Этот результат соответствует низкому содержанию амилопектина крахмалов 292 и 342.
Морфология гранулы
Морфологию крахмальной гранулы исследовали, используя сканирующую электронную микроскопию (Фиг.6). Размер и форма гранул из Himalaya (Фиг.6, панель а), ячменя waxy (Waxiro, Фиг.6, панель б) и АС38 (Фиг.6, панель в) соответствовали ранее опубликованным наблюдениям крахмальных гранул у нормальных линий ячменя. Морфология крахмальных гранул "А" типа у мутантных линий 292 (Фиг.6, панель г), 342 (Фиг.6, панель д) и МК6827 (Фиг.6, панель е) явно изменена, так как гранулы имеют изогнутую поверхность по сравнению с гладкой двояковыпуклой формой гранул нормальных ячменей.
Диетическая клетчатка
Анализ диетической клетчатки был проведен согласно методике АОАС и показал, что увеличение диетической клетчатки наблюдалось в 292 и 342, и что такое увеличение диетической клетчатки было вероятнее следствием увеличения нерастворимой диетической клетчатки, чем растворимой диетической клетчатки (Табл. 1), в соответствии с компонентами диетической клетчатки, которые представляют собой устойчивый крахмал и β-глюкан. Следует отметить, что эта мера диетической клетчатки является химически определяемой мерой, которая совершенно отличается от физиологической меры, релевантной с точки зрения питания.
Генетическая основа мутации
Сегрегационное отношение
Скрещивание мутации с сортами ячменя, не проявляющими фенотип сморщенного эндосперма 292 или 342, показало, что эта мутация представляет собой рецессивную мутацию прямого направления, проявляющую отношение 3 нормальных: 1 сморщенное семя F2 популяций ауткроссинга и отношение 1 нормальное: 1 сморщенное семя двойной гаплоидной популяции, развившейся после одного ауткроссинга (см. Табл. 6). Нормальное семя определено как семя с отношением L/T<3,5, сморщенное семя определено как семя с отношением L/T>3,5.
Аллельная природа мутантов
Путем анализа потомства от скрещиваний 292 и 342 показали, что мутации 292 и 342 являются аллельными. Весь посевной материал F1, имеющий происхождение от реципрокных скрещиваний, проявлял фенотипы массы зерна и морфологии зерна в пределах диапазона размеров и форм, наблюдаемых для родительских линий 292 и 342 и вне диапазона размера и формы семени, обнаруженного для родительской линии Himalaya. Кроме того, весь посевной материал F2, имеющий происхождение от растений F1 292х342, проявлял типичный фенотип сморщенного семени мутантов 292 и 342.
Анализ морфологии зерна и характеристик крахмала серии мутантов со сморщенным зерном, доступных от Barley Germplasm Collection (USDA-ARS, National Small Grains Germplasm Research Facility, Aberdeen, Idaho 83210, USA), позволил предположить, что линия МК6827 (BGS31, также называемая GSHO 2476), несущая мутацию sex6, проявляла ряд характеристик зерна и крахмала, высоко подобный мутациям 292 и 342. Провели скрещивания между 292 и МК6827, и все зерно F1 показало типичный фенотип 292 в отношении фенотипа массы зерна и сморщенного семени. Весь посевной материал F2, имеющий происхождение от растений F1 292×МК6827, проявлял фенотип сморщенного эндосперма с соотношениями L/T>4. Напротив, посевной материал F2 от скрещивания между 292 и имеющимся в продаже сортом ячменя Sloop давал бимодальное распределение, показывая отношение сегрегации 3:1 между сморщенным и наполненным семенем (Табл. 6). Весь посевной материал F1, имеющий происхождение от скрещиваний 292 и 5 других линий с фенотипами сморщенного эндосперма (BGS 380, сморщенный эндосперм 4, 7HL (Jarvi et al., 1975); BGS 381, сморщенный эндосперм 5, 7HS (Jarvi et al, 1975); BGS 382, sex 1, 6HL (Eslick and Ries, 1976); BGS 396, сморщенный эндосперм 6, 3HL (Ramage and Eslick, 1975); BGS 397, сморщенный эндосперм 7, не картирован (Ramage and Eslick, 1975), давал зерно с морфологией наполненного семени. На этом основании мутации 292, 342 и МК6827 считают аллельными, и на основе ранее опубликованных локализаций на карте для локуса sex6 мутации 292 и 342 вероятно предсказаны на карте в коротком плече хромосомы 7Н ячменя примерно 4 сМ от центромера (Netsvetaev, 1990, Netsvetaev and Krestinkov, 1993, Biyashev et al., 1986, Netsvetaev, 1992).
Анализ сцепления
От скрещивания между 292 и имеющегося в продаже сорта пивоваренного ячменя, Tantangara, получили двойную гаплоидную популяцию, которая содержала 90 линий-потомков (Табл. 8).
Эти линии оценивали по морфологии семени (наполненное против сморщенного семени), распределению длин цепей путем FACE (процент цепей с DP 6-11), оболочке семени (беспокровное или покрытое шелухой) и по ПЦР-маркеру (см. ниже). Эти данные представлены в Табл. 8. В данной популяции признак сморщенного семени и FACE-распределение 292 точно косегрегировали, как можно было ожидать, если измененные размер и форма зерна являлись следствием измененного отложения крахмала. Косегрегация признаков проиллюстрирована на Фиг.8. На панели А показана взаимосвязь между длиной цепи крахмала (проиллюстрированной на основании процента цепей между DP 6-11) и отношением длины к толщине. Пустыми кружками показаны линии, которые имеют ПЦР-маркер для мутации 292, крестиками показаны линии, которые несут ПЦР-маркер дикого типа. Существует четкое определение между этими двумя группами линий. На панели Б Фиг.8 показана взаимосвязь между длиной цепи крахмала и массой семени, и показано, что масса семени является менее диагностической для этой мутации, чем отношение длины к толщине.
У ячменя наличие или отсутствие шелухи контролирует локус nud, локализованный на хромосоме 7Н, и поскольку Tantangara представляет собой ячмень, покрытый шелухой, а 292 представляет собой беспокровный тип, этот признак можно оценить в двойном гаплоидном потомстве. Анализ сцепления между признаком беспокровный/покрытый шелухой и данными FACE показал, что в данном скрещивании мутация 292 картирована в пределах 16,3 сМ от локуса nud. Эта локализация совпадает с предшествующими данными картирования для аллельной мутации sex6 (Netsvetaev, 1990, Netsvetaev and Krestinkov, 1993, Biyashev et al., 1986, Netsvetaev, 1992).
Идентификация причинного гена
Продемонстрировано, что ген nud локализован на хромосоме ячменя 7Н (Фиг.8, Fedak et al., 1972). У пшеницы три синтазы крахмала (GBSS, SSI и SSII) и фермент типа изоамилазы, удаляющий разветвление цепи (S.Rahman, личное сообщение), локализованы на коротком плече хромосомы 7, гомологичной хромосомы (Yamamori and Endo, 1996, Li et al., 1999a, Li et al., 1999b, Li et al., 2000). Тесное сцепление с локусом nud позволило предположить, что наиболее вероятным геном-кандидатом является ген SSII. Ген SSII пшеницы клонирован на уровне кДНК (Li et al., 1999b; Genbank, номер по каталогу AF155217) и на геномном уровне (Li et al., личное сообщение), и кДНК ячменя выделяли и клонировали (Фиг.9). Секвенирование геномных последовательностей SSII ячменя и пшеницы показало, что эти гены обладают очень сходными экзон/интронными структурами, однако, длины интронных участков различаются между последовательностями (Фиг.10). Сравнение геномной последовательности Morex и последовательности кДНК из Himalaya (Фиг.9) приводит к идентификации выведенных последовательностей кДНК из Morex, 292 и МК2827.
Мутант транзиции G на А обнаружен в гене SSII из 292 в положении, которое соответствует 1829 выравнивания, показанного на Фиг.11. Эта мутация вводит стоп-кодон в открытую рамку считывания SSII 292 (Фиг.12). Анализ последовательностей Tantangara и Himalaya показал, что оба гена дикого типа идентичны в данном участке, и как 292, так и 342 содержат одну и ту же мутацию транзиции G на А. Введенный стоп-кодон должен укорачивать продукт этого гена таким образом, что полноразмерный С-концевой каталитический домен гена синтазы крахмала II не будет транслироваться, и, следовательно, существует высокая вероятность того, что активность SSII полностью прекращена данной мутацией.
Транзиция G на А также присутствовала в МК6827 в положении 242 выравнивания, показанного на Фиг.11, и последовательности кДНК Himalaya на Фиг.9. Эта мутация также вводит стоп-кодон в открытую рамку считывания SSII 292 (Фиг.12) и должна предотвращать трансляцию более 90% гена SSII, прекращая активность SSII, кодируемую данным геном.
Мутация транзиции G на А у 292 прервала сайт рестрикции (NIaIV) в гене SSII ячменя. Локализация этого диагностического сайта NIalV показана на Фиг.14, панелях (а) и (б). На Фиг.14в показан электрофорез в агарозном геле продуктов ферментативного гидролиза NIalV из ячменя, показывающий, что диагностическая картина для мутации 292 находится в 292 и 342, но не в МК6827, Himalaya или Tantangara.
ПЦР-маркер для транзиции G на А оценивали у 90 линий двойной гаплоидной популяции 292×Tantangara и обнаружили, что она точно косегрегирует с фенотипами сморщенного семени и распределения длин цепей по FACE, указывая на то, что мутация 292 полностью сцеплена с данным фенотипом крахмала, и что существует высокая вероятность того, что эта мутация является причинной мутацией, лежащей в основе фенотипа 292. На Фиг.14г показан анализ линий от двойной гаплоидной популяции 292×Tantangara.
Биохимическое доказательство потери активности SSII
Состав ферментов биосинтеза крахмала в мутантных и нормальных линиях ячменя исследовали, используя ряд методик гель-электрофореза. Анализ растворимой фракции развивающегося эндосперма продемонстрировал, что все линии содержали BEI, BEIIa, BEIIb, SSI и SSIII, и что содержание этих изоформ BE и SS, соответственно, было по существу не измененным. Однако анализ крахмальной гранулы показал, что несколько полос отсутствовали. Во-первых, анализ путем электрофореза в ДСН-ПААГ (Фиг.16, панель Б) показал, что полоса 90 кД, которая присутствовала в Himalaya, Tantangara и АС38, отсутствовала в 292, 342 или МК6827. С помощью иммуноблоттинга было показано, что эта полоса содержит SSII (Фиг.16, панель Б, а также BEIIa и BEIIb. Обнаружение того факта, что BEIIa и BEIIb присутствуют в растворимой фракции, но не в крахмальной грануле, указывает на то, что у мутантов 292, 342 и МК6827 вероятнее существует изменение в распределении этих ферментов, чем мутация, прекращающая экспрессию. Напротив, не обнаружено никакого доказательства экспрессии SSII ни в растворимой, ни в гранулярной фракции (Фиг.16, панели А и Б), что совпадает с генетическим доказательством непосредственного сцепления мутации SSII с наблюдаемыми фенотипами у 292, 342 и МК6827.
Селекция линий, несущих мутацию 292
Для переноса мутации 292 в альтернативные генотипы ячменя использованы две стратегии.
В первом примере получили двойные гаплоидные линии от скрещивания между 292 и Tantangara. Данные для покрытия семени, массы семени, отношения L:T, распределения длин цепей и статуса ДНК-маркера SSII представлены в Табл. 8. Более полный анализ состава этих линий представлен в Табл. 9, включая RVA-анализ, содержание β-глюкана и объем набухания муки. Эти данные показывают, что линии, несущие мутацию 292, имеют значительно отличающиеся параметры RVA (примером чего является отношение пиковой/конечной вязкости), более высокое содержание β-глюкана и измененные объемы набухания муки.
Во втором примере мутацию переносили путем проведения двух обратных скрещиваний из 292 с сортом, имеющим нормальные свойства крахмала (cv Sloop). Для анализа собирали посевной материал F2 от трех растений F1 обратного скрещивания 2. Этот посевной материал F2 разделяли на категории семян с отношением LT>3,5 и с отношением LT<3,5. Распределение семян между этими классами соответствовало ожиданиям для одиночного рецессивного гена. Данные по объемам набухания муки для этих категорий семян, полученных от каждого растения, представлены на Фиг.10 и показывают, что признак набухания крахмала четко переносится посредством процесса скрещивания в линии со средним содержанием генетического фона Sloop 75%.
Обсуждение
Авторы изобретения описывают выделение новых мутантов, 292 и 342, у ячменя, которые имеют фенотип сморщенного эндосперма. Анализ состава зерна показывает, что сморщенный фенотип является следствием значительного снижения содержания крахмала, а анализ состава крахмала показывает, что это снижение проявляется в виде высокоамилозного фенотипа, который является результатом снижения синтеза амилопектина.
Мутанты 292 и 342 обладают уникальным сочетанием свойств зерна и крахмала в содержании как повышенных уровней β-глюкана, так и устойчивого крахмала. Уровни β-глюкана этих линий увеличены примерно на 15% выше того уровня, который ожидают вследствие эффекта пониженного содержания крахмала, что позволяет предположить, что углерод, неспособный превращаться в крахмал, переключается на синтез β-глюкана. Определения уровней диетической клетчатки демонстрируют, что зерно от мутантов обладает повышенными уровнями диетической клетчатки, и что это повышение является следствием повышения нерастворимой диетической клетчатки.
Данное сочетание свойств указывает на то, что эти мутанты могут иметь весьма представляющий интерес потенциал в качестве компонентов диеты человека. Во-первых, повышенные уровни β-глюкана предполагают, что эти линии могут быть полезны при снижении холестерина посредством хорошо установленного действия β-глюкана в снижении уровней холестерина. Во-вторых, наличие устойчивого крахмала указывает на то, что эти линии могут быть полезны вследствие перспективы здоровья кишечника благодаря хорошо установленной способности устойчивых крахмалов способствовать ферментации в ободочной кишке (Topping et al., 1997, Topping, 1999). В-третьих, состав зерна указывает на то, что эти линии будут обладать низкоэнергетической плотностью и что они могут медленно подвергаться ферментативному гидролизу, указывая на то, что они могут способствовать изготовлению пищевых продуктов с пониженным гликемическим индексом.
Свойства крахмала линий, служащих примером, также являются уникальными в том, что они объединяют высокоамилозный крахмал, который к тому же имеет низкую температуру желатинизации. Это противоречит высокоамилозным мутациям, являющимся результатом мутаций в разветвляющем цепь ферменте IIb, при которых температура желатинизации типично повышается, как например при мутации amylose extender у кукурузы (Ng et at., 1997, Katz et al., 1993, Krueger et al., 1987, Fuwa et al., 1999). Хотя содержание амилозы 292 сравнимо с линиями amylose extender, структура амилопектинового компонента крахмалов резко отличается (Wang et al., 1993). У 292 и 342 распределение длин цепей амилопектина сдвинуто в направлении более низкой степени полимеризации, тогда как у amylose extender распределение длин цепей сдвинуто в направлении повышенной степени полимеризации. Это позволяет предположить, что амилопектин, а не содержание амилозы, является первичным определяющим фактором температуры желатинизации, и что данный эффект опосредован силой взаимодействия между внешними цепями молекулы амилопектина. Подобные эффекты были отмечены для ряда крахмалов Jane et al., 1999.
Данные по вязкости на основании RVA-анализа указывают на то, что крахмал из мутантных по SSII линий заметно отличается от нормальных ячменей и АС38. Мутанты ячменя по SSII по существу не имеют пика вязкости, типично наблюдаемого при резком повышении температуры до 95°С в начале RVA-температурного профиля. Вместо этого у этих мутантов вязкость повышается монотонно до конечной вязкости, если она имеется. Эти данные находятся в соответствии с низким содержанием амилопектина гранул, низким уровнем амилопектиновой кристалличности в грануле и низкими температурой и энтальпией желатинизации, наблюдаемыми в дифференциальном сканирующем калориметре. Высокая конечная вязкость достигается, как только амилоза высвобождена из гранулы посредством нагревания в избытке воды и перемешивания. Эти RVA-характеристики являются уникальными для крахмала хлебных злаков и обеспечивают новый источник крахмала для пищевых продуктов и промышленных применений, где необходима низкая вязкость пастирования и все еще высокая конечная вязкость.
Наблюдения, сделанные по температуре желатинизации в ДСК, отражены в результатах исследований дифракции рентгеновских лучей. Гранулы 292 и 342 имеют пониженные уровни кристалличности и сдвиги кристаллической формы от типа А, типичного для крахмалов хлебных злаков, до смеси типов V и В. Тип V типичен для амилозы и отражает амилозный компонент крахмала, образующего комплекс с жирными кислотами, тогда как форма В имеет происхождение от амилопектина и преимущественно отражает содержание остаточного амилопектина крахмала (Buleon et al., 1998).
Анализ генетической основы мутаций 292 и 342 демонстрирует, что эти мутации представляют собой простые рецессивные мутации, которые дают типичные Менделевские отношения в экспериментах по ауткроссингу. Исследования скрещивания показали, что 292 и 342 являются аллельными. Дальнейший анализ взаимодействия между 292 и другими мутациями сморщенного эндосперма в экспериментах по скрещиванию продемонстрировал, что мутации 292/342 также являются аллельными с мутацией Sex6 в линии МК6827. Эта мутация картирована ранее, и показано, что она локализована в пределах 3 сМ от центромера короткого плеча хромосомы 7Н (Netsvetaev, 1990, Netsvetaev and Krestinkov, 1993, Biyashev et al., 1986, Netsvetaev, 1992).
Была создана двойная гаплоидная популяция между покрытым шелухой ячменем Tantangara и беспокровным мутантом 292, и мутация сморщенного эндосперма картирована на коротком плече хромосомы 7HS в пределах 16 сМ от гена nud в локализации, совпадающей с локализацией на карте мутации Sex6.
Локализация гена в участке, прилежащем к центромеру, на коротком плече хромосомы 7HS показывает, что причинная мутация (sex6) находится в гене, отличном от мутации, которая вызывает высокоамилозный фенотип у АС38 (amo 1), которая картирована на хромосоме 1Н (Schondelmeier et al., 1992). Эта локализация на карте позволила предположить, что одним из кандидатов для гена, прерванного при мутации sex6/292, является синтаза крахмала II, о котором известно, что у пшеницы он локализован в том же участке хромосомы (Yamamori and Endo, 1996, Li et al., 1999b). Анализ последовательности мутантов 292 и 342 показал, что в этом гене имела место мутация транзиции G на А, которая могла вызвать укорочение этого гена, так что С-концевой участок, содержащий активный сайт фермента, не должен транслироваться, что преимущественно приводит к синтезу полностью неактивного белка. Кроме того, секвенирование гена SSII из МК6827 показало мутацию транзиции G на А в положении 242, которая также должна была вызвать укорочение этого гена. Этот результат подтверждает аллельную природу мутаций 292 и МК6827.
Идентификация мутаций в гене SSII привела к разработке ПЦР-маркера, диагностического для мутации в 292. Этот ПЦР-маркер оценивали через 91 потомок популяции 292×Tantangara, и показано, что он на 100% косегрегирует с фенотипом сморщенного эндосперма и фенотипом пониженного распределения длин цепей крахмала. Обнаружение аллельных мутаций в генах SSII из ячменей различных генетических фонов (292 и МК6827), которые вызывают подобные фенотипы, и точное сцепление этой мутации с фенотипом сморщенного зерна дает высокую степень достоверности того, что эти мутации, присутствующие в генах SSII 292, 342 и МК6827, являются причинными мутациями, приводящими к признаку сморщенного эндосперма.
Фенотип, наблюдаемый здесь для мутации SSII у ячменя, в некоторых отношениях подобен фенотипам мутаций SSII у других растений, однако, мутации SSII не вызывают настолько высокое содержание амилозы, как мутации, обнаруженные в 292/342. Мутации SSII известны у гороха (rug5, Craig et al., 1998) и Chlamydomonas (Fontaine et al., 1993) и приводят к амилопектинам с пониженными распределениями длин цепей, как наблюдали здесь. Имеются также данные, позволяющие предположить, что мутация Shrunken-2 у кукурузы является результатом мутации гена SSII, хотя это еще следует окончательно продемонстрировать (Harn et al., 1998, Knight et al., 1998). У кукурузы мутация Shrunken-2 приводит к образованию крахмалов, имеющих пониженные температуры желатинизации (Campbell et al., 1994). У пшеницы Yamamori разработал линию тройных нуллисомиков, в которой отсутствует белок Sgp-1 (Yamamori, 1998), который, как показано Li et al (Li et al., 1999b), является продуктом гена SSII. В пшенице содержание амилозы повышено примерно до 35%, и наблюдаются аномальные крахмальные гранулы, измененная кристалличность и измененная температура желатинизации (Yamamori, 1998). Различия в свойствах между мутантами SSII ячменя и мутантами SSII из других видов являются совершенно неожиданными.
Показано, что мутация SSII способна к переносу посредством скрещивания из одного генетического фона в другой и дает диагностические морфологию и состав зерна, типичные для исходных 292, 342 и МК6827. В таблице (9) данные параметров от двойных гаплоидных линий 292 х Tantangara по отношению L/T, содержанию β-глюкана, распределению длин цепей, RVA и объему набухания муки демонстрируют, что линии, несущие мутацию 292, проявляют фенотипы, типичные для родителя 292. В следующем подтверждении сегрегация посевного материала от потомства от самоопыления второго обратного скрещивания 292 на Sloop показала отношение сегрегации, совпадающее с сегрегацией 3:1, для нормальных (74 семени с отношением L/T<3,5) и сморщенных фенотипов (21 семя с отношением L/T>3,5).
Доступность последовательности гена SSII и систем трансформации ячменя обеспечивает средства, необходимые для выключения гена SSII при использовании методик генной супрессии, с целью получения фенотипа, сравнимого с обнаруженным при мутациях SSII. Недавно разработанной высокоэффективной стратегией является получение шпилечной конструкции, предназначенной для получения двунитевой РНК, которая должна подавлять эндогенную активность SSII. Хотя мутации полного выключения, аналогичные мутациям, описанным здесь, будут представлять интерес, использование конструкций ДНК с различными промоторами и выделение трансгенов с различными уровнями экспрессии шпилечной конструкции, должны дать возможность оценить воздействие титрования экспрессии этого гена от нормальных уровней до полностью выключенных уровней.
Было показано, что эти мутации способны к переносу из 292 в альтернативные генетические фоны ячменя, в то же время сохраняя существенные признаки исходной мутации 292. В Таблицах 9 и 10 представлены фенотипические данные для двойного гаплоидного потомства 292×Tantangara и посевного материала от второго обратного скрещивания с Sloop, и они указывают на то, что эти фенотипы переносятся посредством процесса скрещивания.
б% от суммарного содержания крахмала
н.о. не определяли
б% распределения олигосахаридов, выраженных на молярной основе
** Значение, зарегистрированное в момент пиковой вязкости для Himalaya
*** Значение было ниже 0
н.о. не определяли
б двойное гаплоидное потомство
в в каждом случае, χ2(0,05), df=1=3,84, следовательно, популяция 292×Sloop соответствует сегрегации 3:1, а двойная гаплоидная популяция 292×Tantangara соответствует сегрегации 1:1
б фенотип шелухи: N - беспокровное; Н - с шелухой
в отношение L/T: отношение длины к толщине
д содержание амилозы определено по значению синего окрашивания йода
е оценка ПЦР: 292 - реакция ПЦР дает полосу, которая дает полосы 169 п.о. плюс 103 п.о. после ферментативного гидролиза NIaIV; Wt - реакция ПЦР дает полосу, которая дает полосы 111 п.о., 103 п.о. и 57 п.о. после ферментативного гидролиза NlaIV.
ПРИМЕР 2
Разработка и конструирование векторов
Участки гена SSII ячменя (как определено на Фиг.15) клонировали в вектора для трансформации. Получили три конструкции для каждого гена-мишени в целях стратегии генной супрессии, (1) смысловая косупрессия, (3) антисмысловая и (3) опосредованная дуплексом супрессия.
На Фиг.16 проиллюстрирована конфигурация последовательностей в конструкциях ДНК, предназначенных для подавления экспрессии эндогенного гена-мишени. Промотор может быть выбран либо из эндосперм-специфичных (таких как промотор глютенина высокой молекулярной массы, High Molecular Weight Glutenin, промотор SSI пшеницы, промотор BEII пшеницы), либо из промоторов, не специфичных для эндосперма (таких как убиквитин или 35S). Конструкция может также содержать другие элементы, которые усиливают транскрипцию, такие как nos 3 элемент OCS. Проиллюстрированные участки ДНК следует включить в векторы, содержащие подходящие последовательности селектируемых маркерных генов и другие элементы, или в векторы, которые котрансформируют с векторами, содержащими эти последовательности.
Трансформация хлебных злаков
Способы трансформации ячменя (Tingay et al., 1997; Wan et al., 1994), овса (Somers et al., 1992, 1994; Gless et al., 1998; Zhang et al., 1999, Cho et al., 1999) и ржи (Castillo et al., 1994; Pena et al., 1984) описаны, и их можно использовать для переноса конструкций ДНК, получая трансгенные растения.
Анализ трансгенных растений
Идентификацию трансгенных растений осуществляют посредством идентификации ДНК конструкции ДНК с помощью ПЦР или гибридизации по Саузерну. Уровни экспрессии индивидуальных генов биосинтеза крахмала ячменя измеряют как на уровне мРНК, так и на уровне белка с помощью стандартных методик, таких как гибридизация по Нозерну и Вестерн-блоттинг, соответственно. Содержание и состав зерна и крахмала измеряют, используя стандартные методики, такие как методики, приведенные в Примере 1.
Ссылки
Abel et al., (1996). The Plant Journal 10, 981-991.
Ahokas, (1979). Barley genetics Newsletter 9, 7-9.
Akerberg et al., (1998). Journal of Cereal Science 28, 71-80.
Andersson et al., (2000). Cereal Chemistry 77, 463-467.
Andersson et al., (1999). Journal of the science of food and agriculture 79, 979-986.
Andreev et al., (1999). StarchStarke 51, 422-429.
Banks et al., (1971). Starch 23, 12-15.
Batey and Curtin (1997). Starch 48, 338-344.
Batey et al., (1997). Cereal Chemistry 74, 497-501.
Bergh et al., (1999). Animal Feed Science and Technology 78, 215-226.
Bhatty (1999). Cereal Chemistry 76, 589-599.
Biyashev et al., (1986). Soviet Genetics 22, 296-303.
Boyer and Preiss, (1978). Carbohydrate research 61, 321-334.
Boyer and Preiss, (1981). Plant Physiology 67, 1141-1145.
Boye et al., (1980). Starch 32, 217-222.
Buleon et al., (1998). International Journal of Biological Macromolecules 23, 85-112.
Calvert et al., (1976). Nutrition Reports. International 14, 55-61.
Campbell et al., (1994). Cereal Chemistry 71, 464-468.
Cao et al., (2000). Achives. of Biochemistry and Biophysics. 373, 135-146.
Case et al., (1998). Journal of Cereal Science 27, 301-314.
Castillo, et al., (1994). Biotechnology 12, 1366-1371.
Cho, et al. (1999). Plant Science (Limerick) 148, 9-17.
Craig et al. (1998). Plant Cell 10, 413-426.
Czuchajowska et al. (1998). Cereal Chemistry 75, 747-754.
Czuchajowska et al. (1992). Cereal Chemistry 69, 413-418.
Denyere et al. (1996). Plant Physiology 112, 779-785.
Elfverson et al. (1999). Cereal Chemistry 76, 434-438.
Eslick and Ries, (1976). Barley Genetics Newsletter 6, 21-22.
Fastnaught et al. (1996). Crop Science 36, 941-946.
Fedak et al. (1972). Canadian Journal of Genetics and Cytology 14, 949-957.
Filpse et al. (1996). Planta 198, 340.
Fontaine et al. (1993). Journal of Biological Chemistry 268, 16223-16230.
Fujita et al. (1999). Breeding Science 49, 217-219.
Fuwa et al. (1999). Starch/Starke 51, 147-151.
Gao et al. (1998). Plant Cell 10, 399-412.
Giroux and Hannah (1994). Molecular and General Genetics 243, 400-408.
Gless et al. (1998). Journal of Plant Physiology 152, 151-157.
Goering and DeHaas, (1974). Cereal Chemistry 51, 573-578.
Granfeldt et al. (1994). American Journal of Clinical Nutrition 59, 1075-1082.
Green et al. (1997). Plant Physiology 114, 203-212.
Gubler et al. (2000). Plant Physiology 122, 1457.
Hedman and Boyer, (1982). Biochemical Genetics 20, 483-492.
Izydorczyk et al. (2000). Journal of Agricultural and Food Chemistry 48, 982-989.
James et al. (1995). Plant Cell 7, 417-429.
Jane et al. (1999). Cereal Chemistry 76, 629-637.
Jarvi and Eslick, R.F. (1975). Crop Science 15, 363-366.
Jobling et al. (1999). Plant Journal 18, 163-171.
Katz et al. (1993). Carbohydrate polymers 21, 133-136.
Knight et al. (1998). Plant Journal 14, 613-622.
Konik-Rose et al. (2001). Sfarch 53, 14-20.
Krueger et al. (1987). Cereal Chemistry 64, 187-190.
Kubo et al. (1999). Plant Physiology 121, 399-409.
Li et al. (1999). Plant Physiology 120, 1147-1155.
Li et al. (2000). Plant Physiology 123, 613-624.
Li et al. (1999). Theoretical and Applied Genetics 98, 1208-1216.
Mizuno et al. (1993). Journal of Biological Chemistry 268, 19084-19091.
Mizuno et al. (1992). Journal of Biochemistry 112, 643-651.
Morell et al. (1997). Plant Physiology 113, 201-208.
Morell et al. (1998). Electrophoresis 19, 2603-2611.
Morrison et al. (1984). Journal of Cereal Science 2, 257-271.
Mouille et al. (1996). The Plant Cell 8, 1353-1366.
Myers et al. (2000). Plant Physiology 122, 989-997.
Netsvetaev (1990). Nauchno-Tekh. Bull'VSGI, Odessa. № 175, 31-35.
Netsvetaev (1992). Cytology and Genetics (Kiev) 26, 26-30.
Netsvetaev and Krestinkov (1993). Barley Genetics Newsletter 22, 44-45.
Newman et al. (1978). Journal of Animal Science 47, 448-455.
Ng et al. (1997). Cereal Chemistry 74, 288-288.
Oscarsson et al. (1998). Journal of the science of food and agriculture 78, 359-366.
Oscarsson et al. (1996). Journal of Cereal Science 24, 161-170.
Oscarsson et al. (1997). Journal of Cereal Science 26, 259-264.
Pena et al. (1987). Nature, UK 325, 274-276.
Persson and Christerson, (1997). Sveriges. Utsadesforenings. Tidskrift. 107, 141-153.
Persson et al. (1996). Sveriges. Utsadesforenings. Tidskrift. 106, 79-86.
Pomeranz, (1992). European Journal of Clinical Nutrition 46, S63-S68.
Pomeranz et al. (1972). Cereal Chemistry 49, 629-635.
Rahman et al. (1995). Australian Journal of Plant Physiology 22, 793-803.
Ramage and Eslick (1975). Barley Genetics Newsletters 5, 114.
Ramage and Eslick, (1975). Barley Genetics Newsletter 6, 115.
Schondelmaier et al. (1992). Plant Breeding 109, 274-280.
Shulmann and Kammiovirta, (1991). Starch 43, 387-389.
Schwall et al. (2000). Nature Biotechnology 18, 551-554.
Shure et al. (1983). Cell 35, 225-233.
Somers et al. (1992). Fertile, transgenic oat plants. Bio/technology 10, 1589-1594.
Somers et al. (1994). Genetic engineering of oat. 37-46.
Song and Jane (2000). Carbohydrate Polymers 41, 365-377.
Sun et al. (1997). The New Phytologist 137, 215-215.
Sundberg et al. (1998). Journal of the Science of Food and Agriculture 76, 457-463.
Swanston (1992). Barley Genetics Newsletter 22, 66-67.
Swanston et al. (1995). Journal of Cereal Science 22, 265-273.
Takeda et al. (1993). Carbohydrate Research 246, 273-281.
Thorbjornsen et al. (1996). Plant Journal 10, 243-250.
Tingay et al. (1997). Plant Journal 11, 1369-1376.
Topping (1999). Asia Pacific Journal of Clinical Nutrition 8, S22-S26.
Topping et al. (1997). The Journal of Nutrition 127, 615-615.
Vasanthan and Bhatty (1995). Cereal Chemistry 72, 379-384.
Vasanthan and Bhatty (1998). Starch-Starke 50, 286-291.
Walker and Merritt (1968). Nature 221, 482-484.
Wan and Lemaux (1994). Plant Physiology 104, 37-48.
Wang et al. (1998). Journal of Experimental Botany 49, 481-502.
Wang et al. (1993). Cereal Chemistry 70, 521-525.
Wilson and McNab (1975). British Poultry Science 16, 497-504.
Xue et al. (1996). Cereal Chemistry 73, 588-592.
Yamamori (1998) (Anon.), pp.300-302. University Extension Press, University of Saskatchewan, Saskatoon.
Yamamori and Endo (1996). Theoretical and Applied Genetics 93, 275-281.
Yoshimoto et al. (2000). Cereal Chemistry 77, 279-285.
Zhang et al. (1999). Plant Celt Reports. 18, 959-966.
Zheng et al. (2000). Cereal Chemistry 77, 140-144.
Zwar and Chandler (1995). Planta 197, 39-48.












| название | год | авторы | номер документа |
|---|---|---|---|
| ЯЧМЕНЬ С ИЗМЕНЕННОЙ АКТИВНОСТЬЮ ВЕТВЯЩЕГО ФЕРМЕНТА И КРАХМАЛ И КРАХМАЛСОДЕРЖАЩИЕ ПРОДУКТЫ С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ АМИЛОЗЫ | 2003 |
|
RU2303870C2 |
| ПШЕНИЦА С МОДИФИЦИРОВАННОЙ АКТИВНОСТЬЮ ВЕТВЯЩЕГО ФЕРМЕНТА, А ТАКЖЕ ПОЛУЧЕННЫЕ ИЗ НЕЕ КРАХМАЛ И КРАХМАЛСОДЕРЖАЩИЕ ПРОДУКТЫ | 2004 |
|
RU2377303C2 |
| ВЫСОКОАМИЛОЗНАЯ ПШЕНИЦА-III | 2017 |
|
RU2811010C2 |
| Высокоамилозная пшеница | 2011 |
|
RU2619636C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ КРАХМАЛА С НОВЫМИ ФУНКЦИЯМИ | 2016 |
|
RU2745568C2 |
| СПОСОБ ПОВЫШЕНИЯ КАЧЕСТВА ЗЕРНА КУКУРУЗЫ | 1991 |
|
RU2083088C1 |
| ТИПЫ ВОСКОВИДНОГО ПШЕНИЧНОГО КРАХМАЛА, СОДЕРЖАЩИЕ В ГРАНУЛЕ ВОСКОВИДНЫЕ БЕЛКИ | 1997 |
|
RU2245617C9 |
| ИНКАПСУЛИРОВАНИЕ ПОЛИПЕПТИДОВ В МАТРИЦУ КРАХМАЛА | 1997 |
|
RU2268301C9 |
| СШИТЫЙ ВЫСОКОАМИЛОЗНЫЙ КРАХМАЛ ДЛЯ ИСПОЛЬЗОВАНИЯ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ С РЕГУЛИРУЕМЫМ ВЫСВОБОЖДЕНИЕМ И СПОСОБЫ ЕГО ПОЛУЧЕНИЯ | 2001 |
|
RU2274443C2 |
| АУТОПРОЦЕССИРУЮЩИЕСЯ РАСТЕНИЯ И ЧАСТИ РАСТЕНИЙ | 2002 |
|
RU2312144C2 |
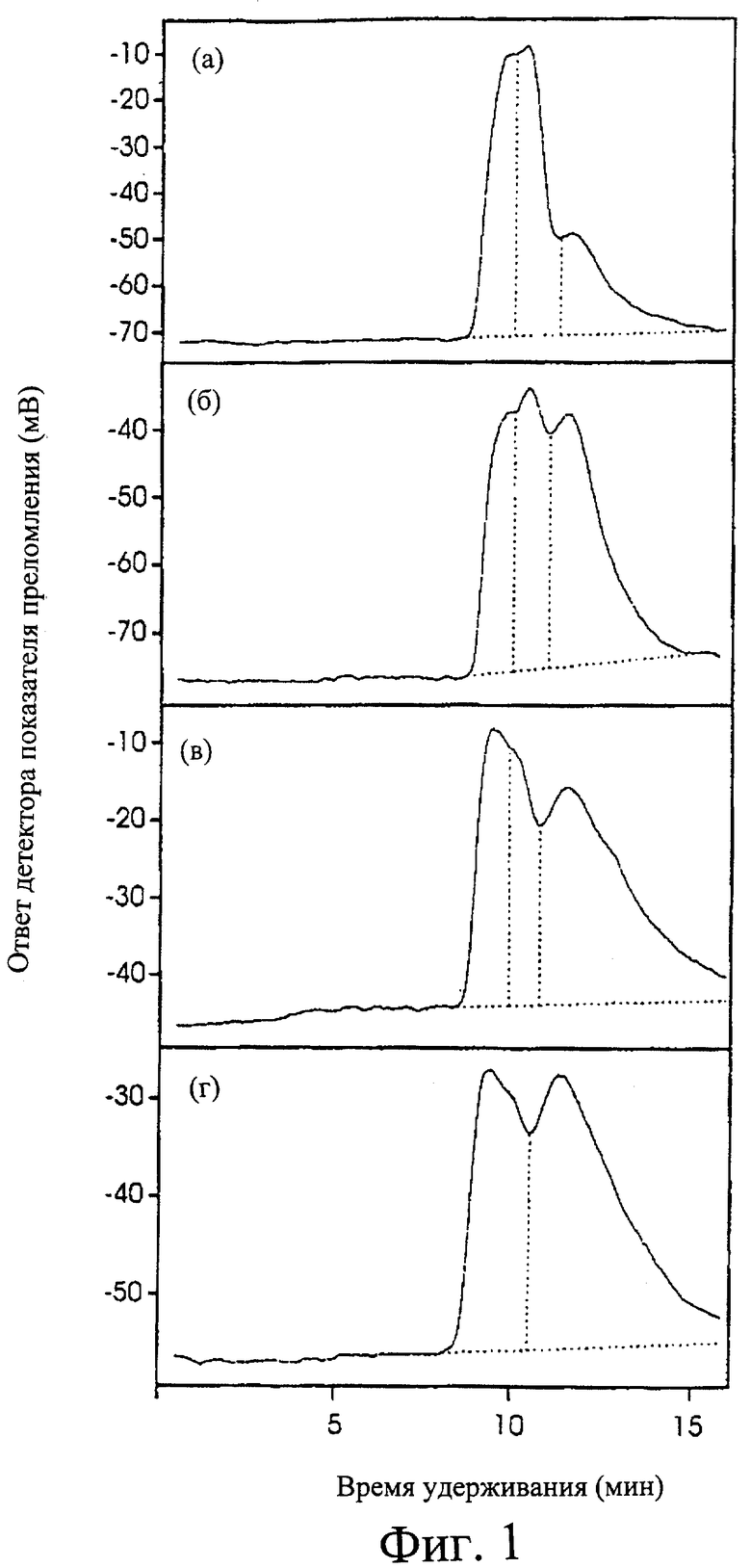
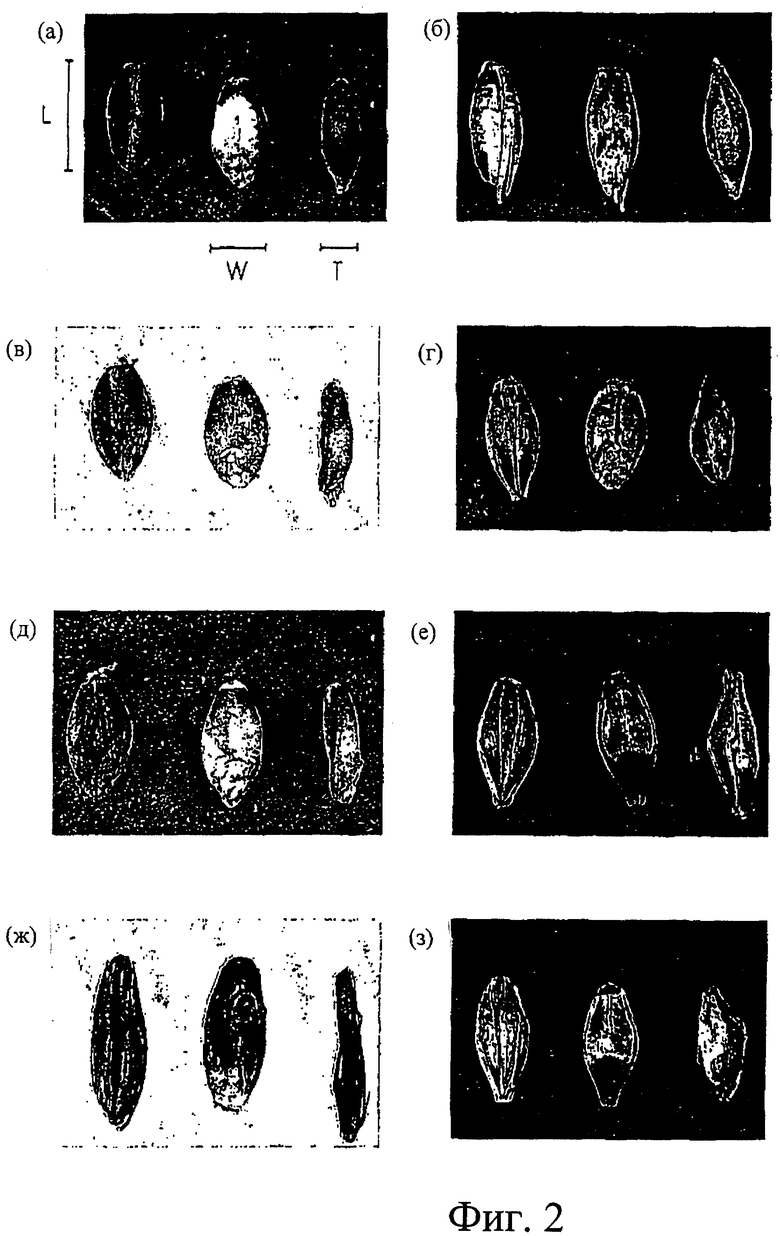
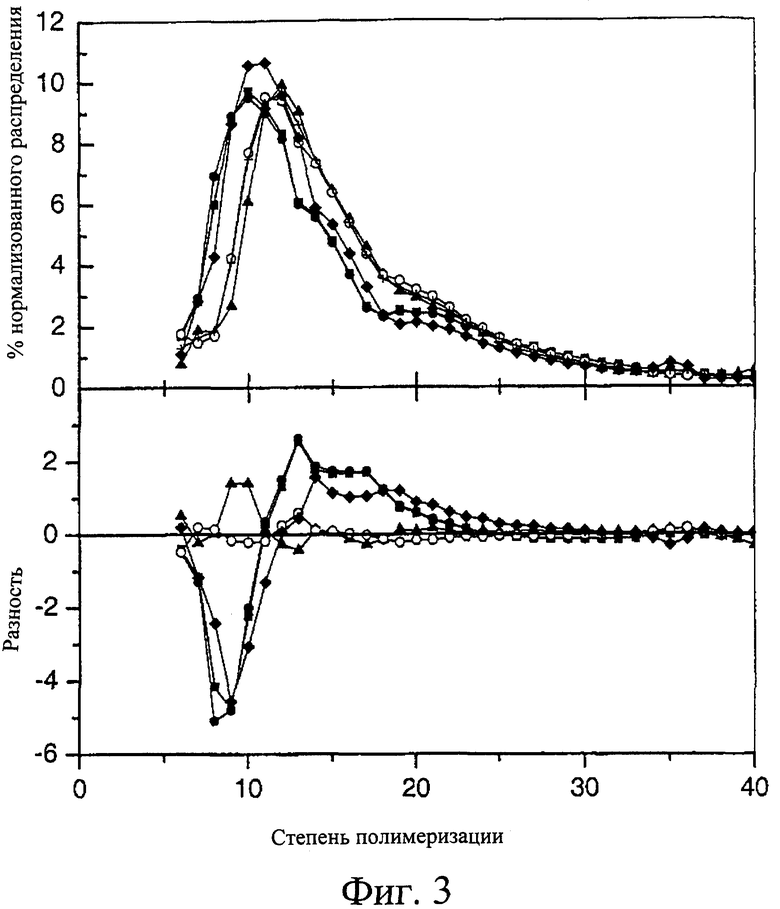
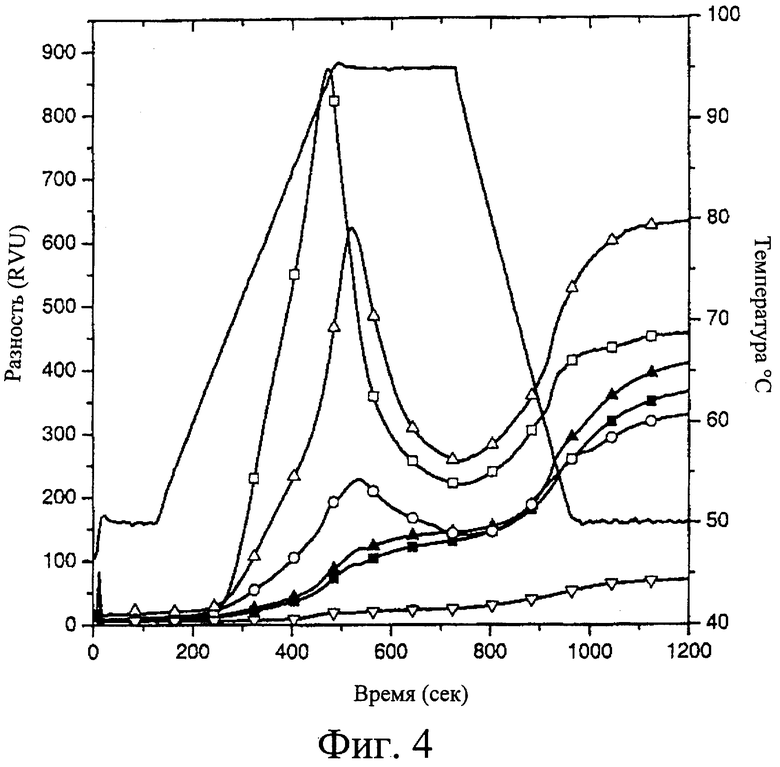
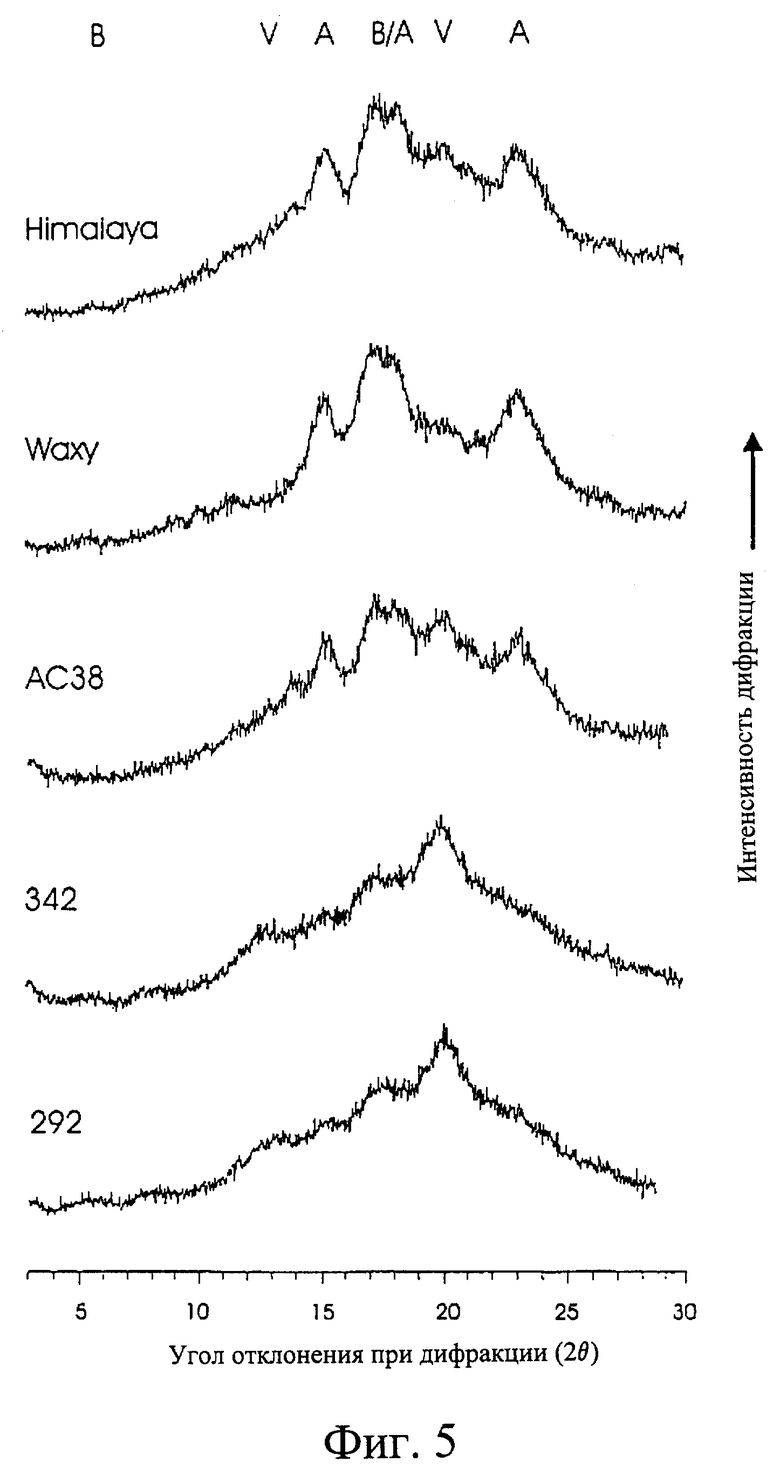


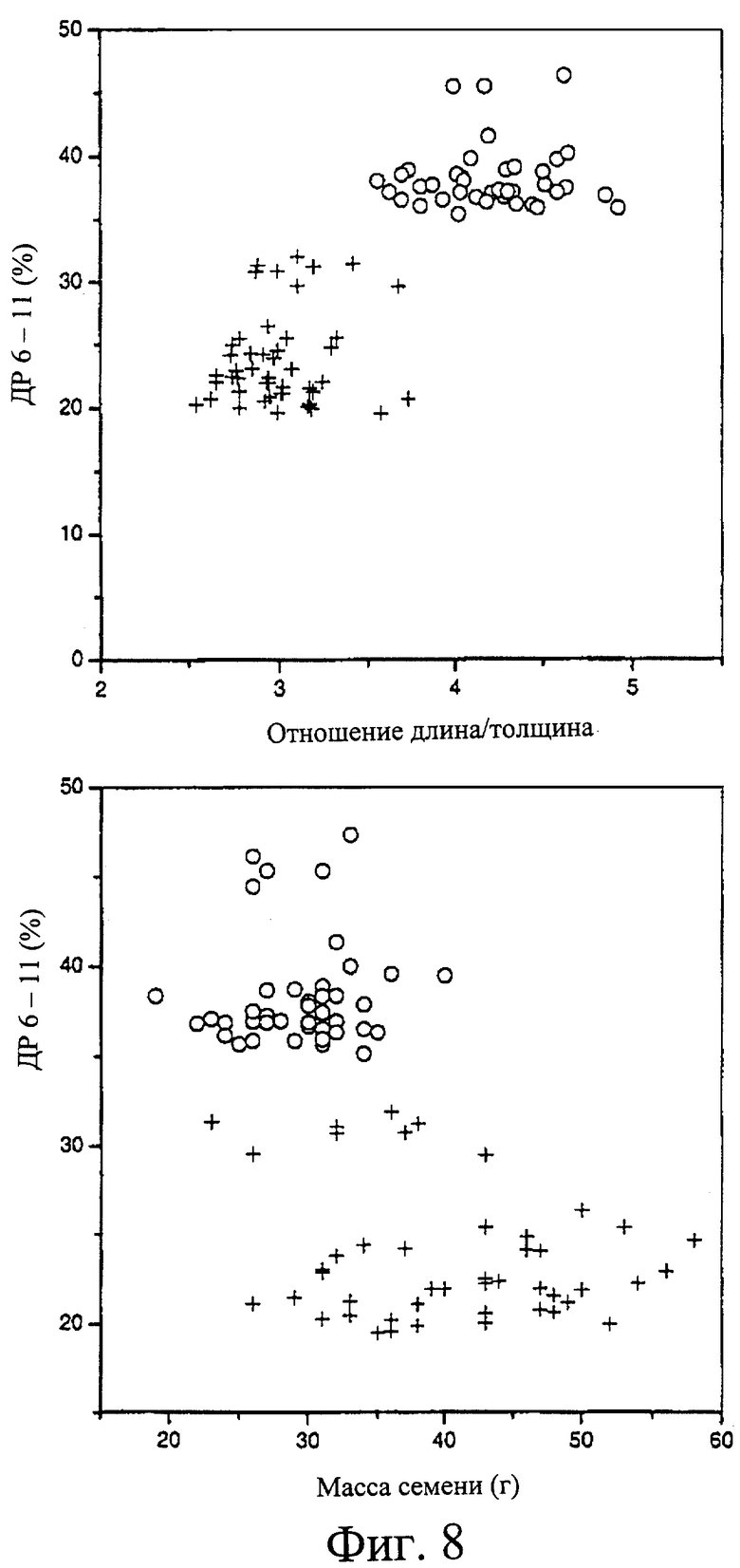
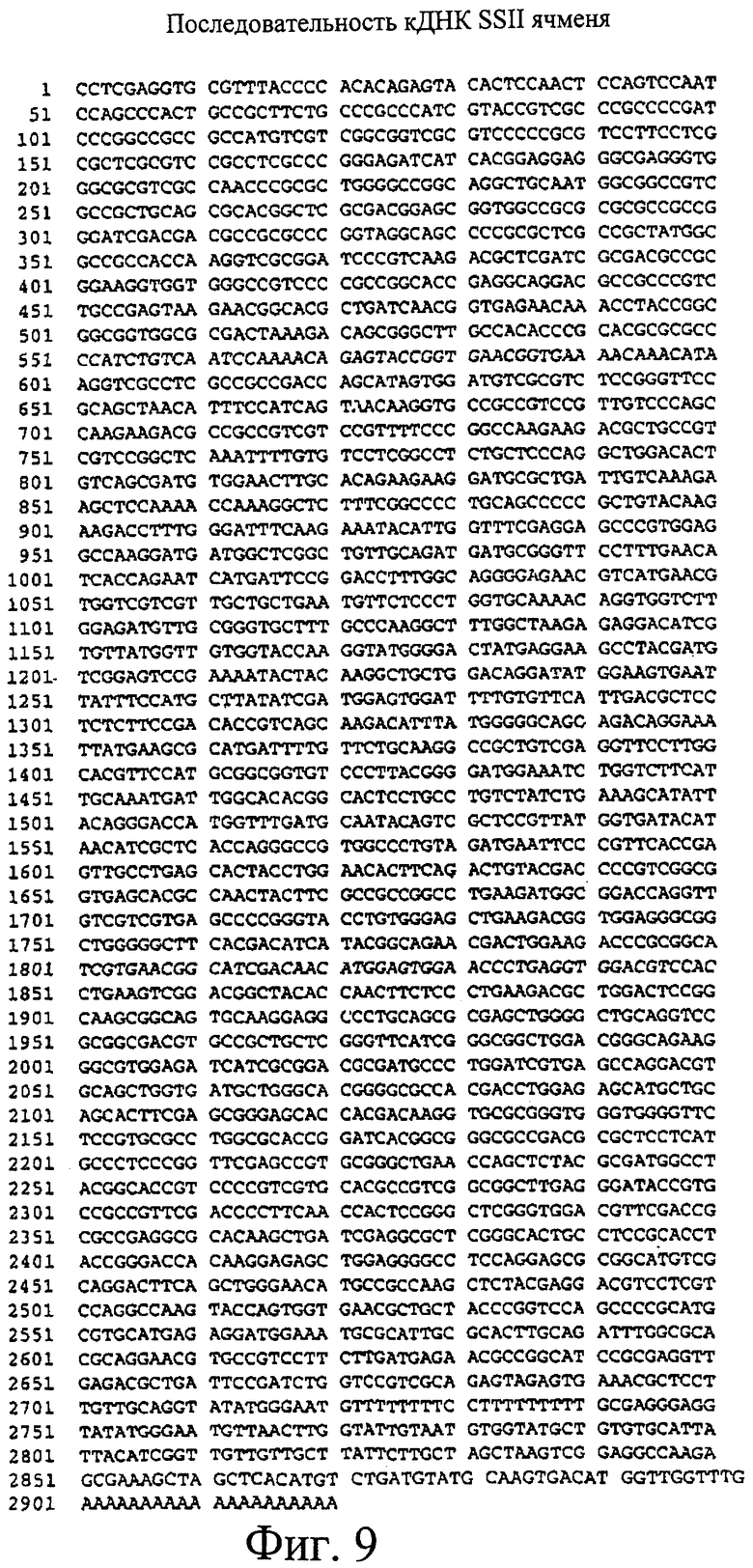
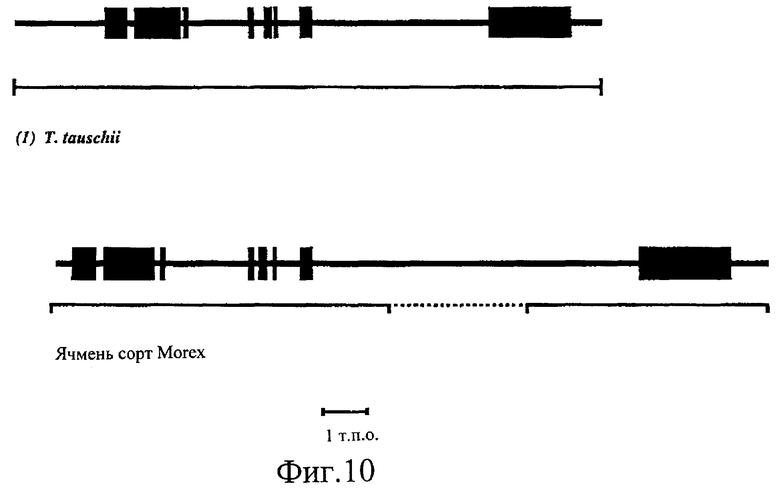
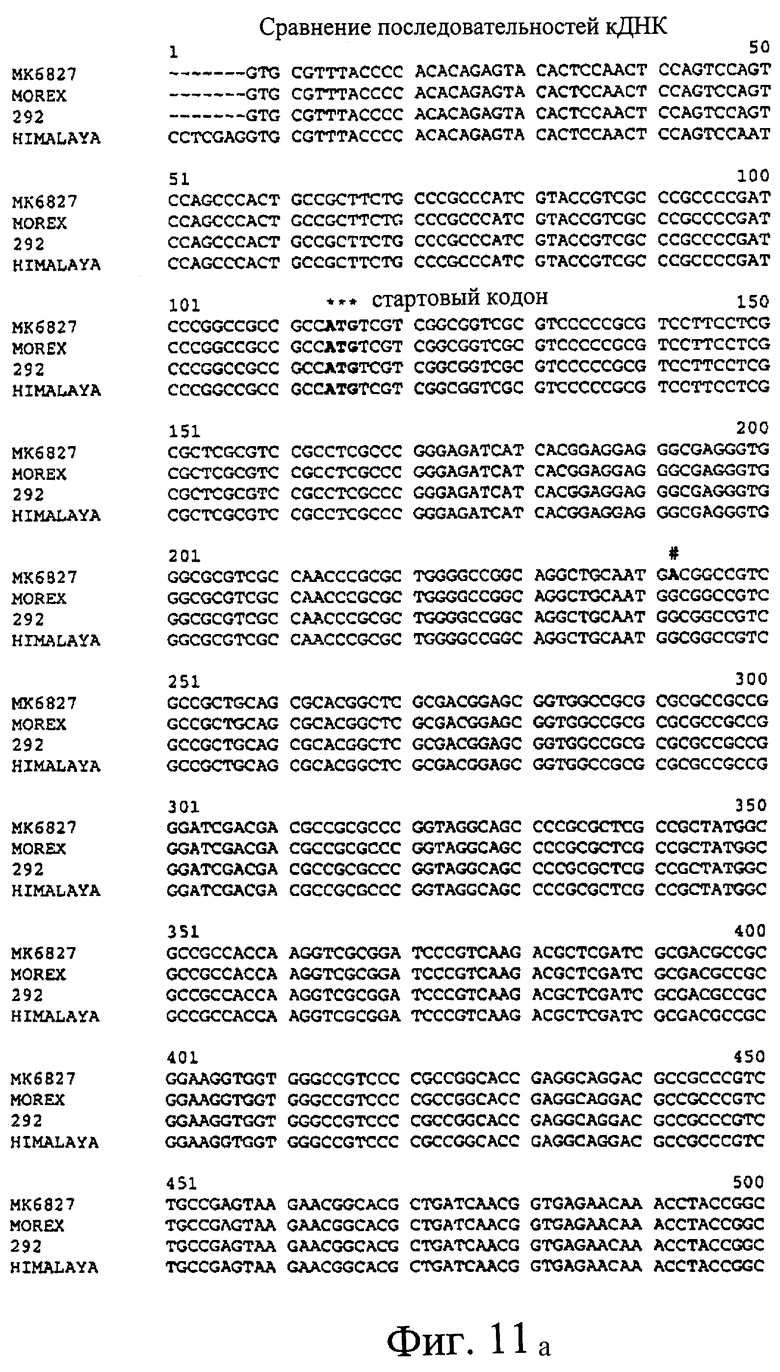



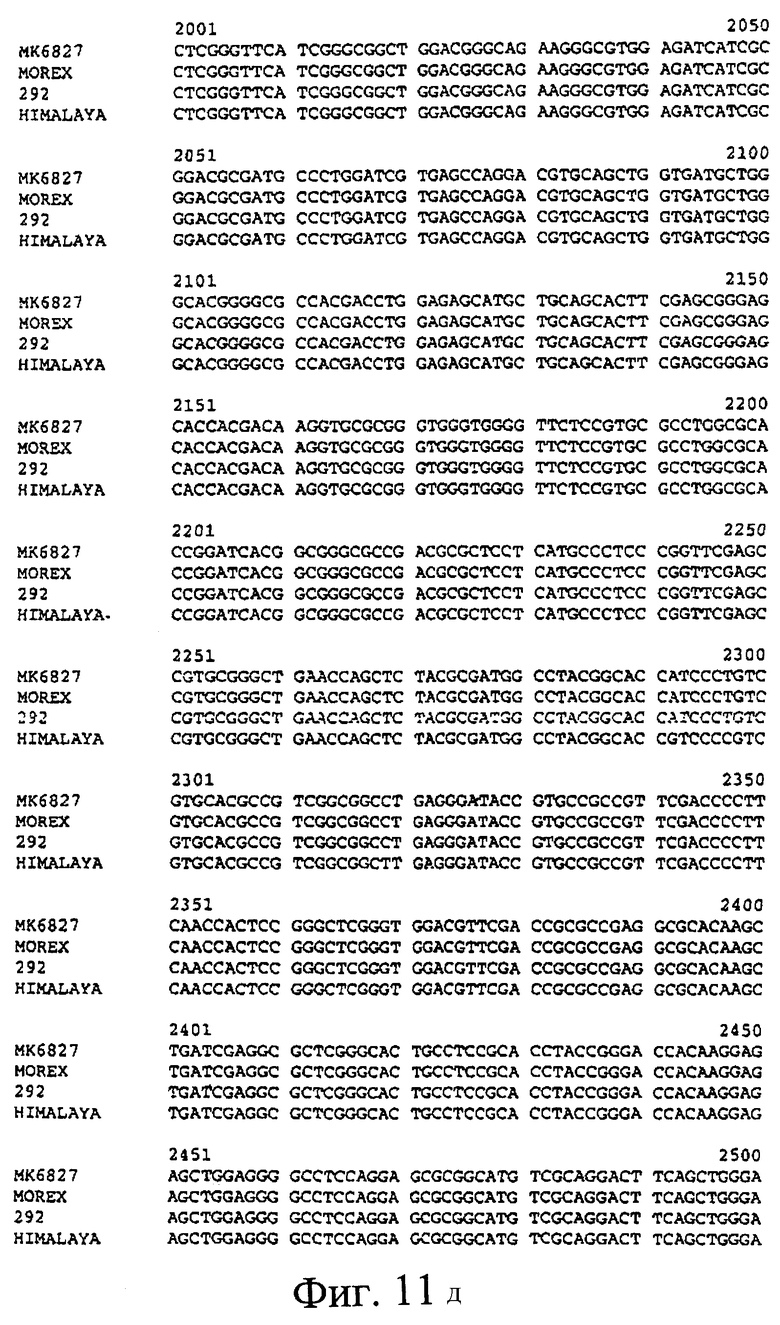
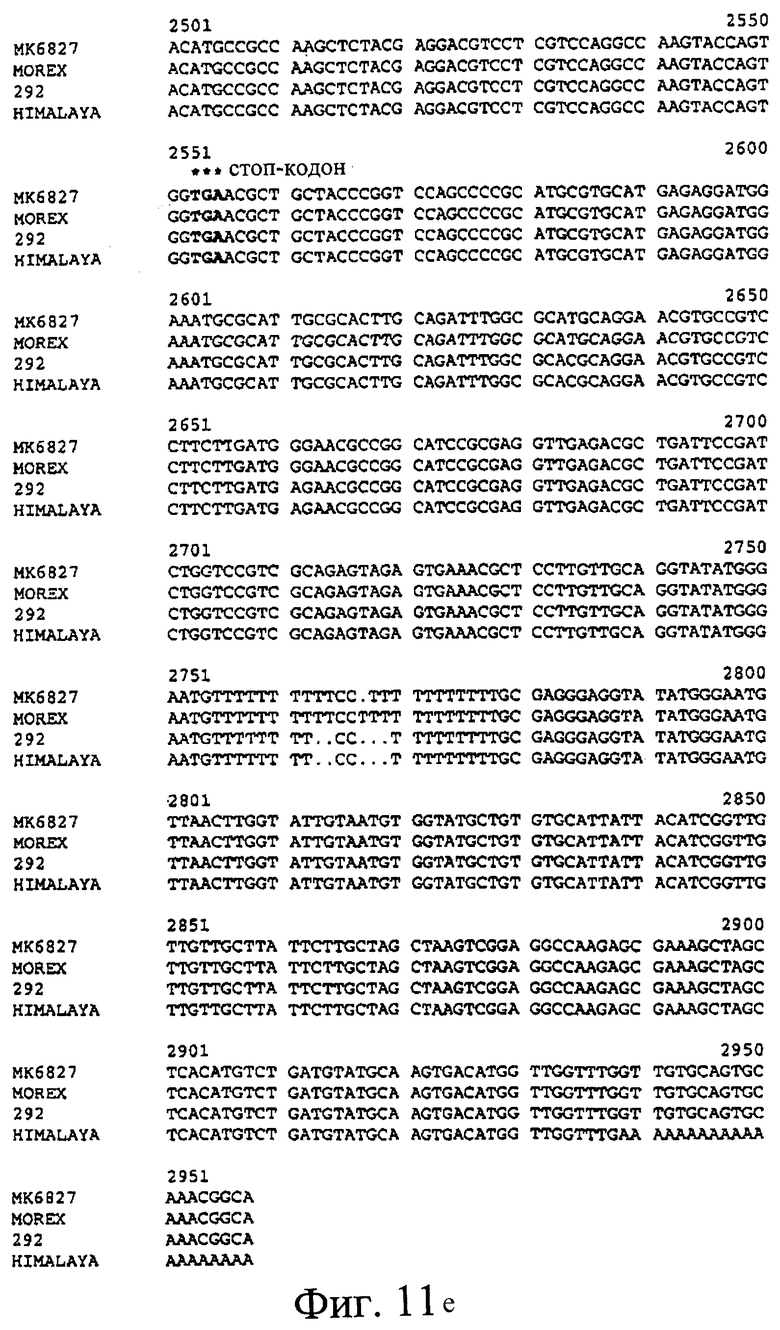


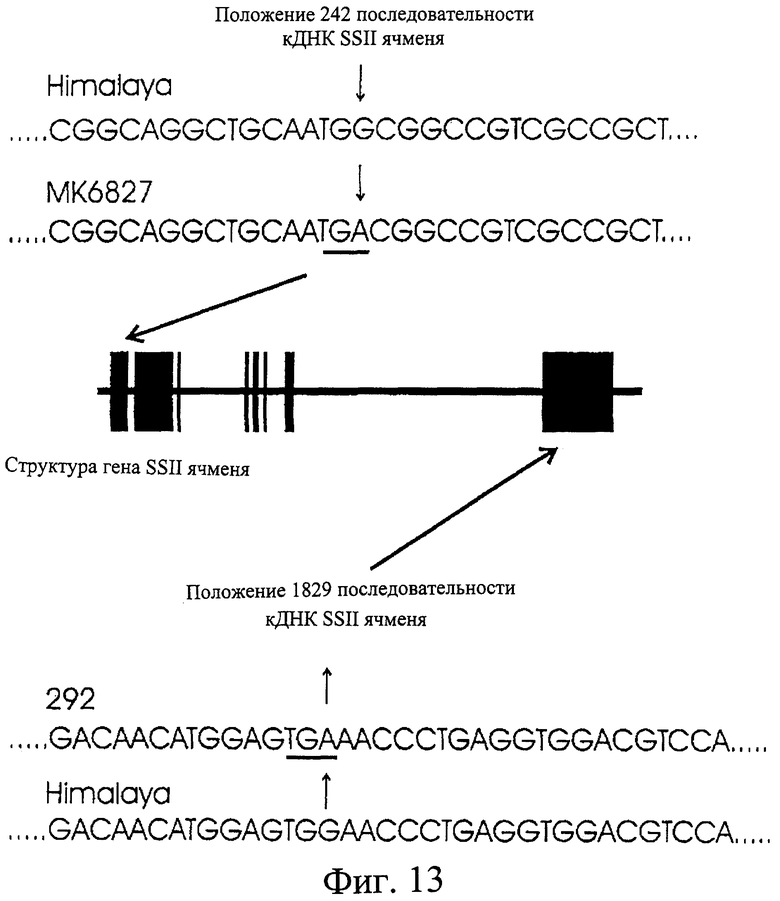
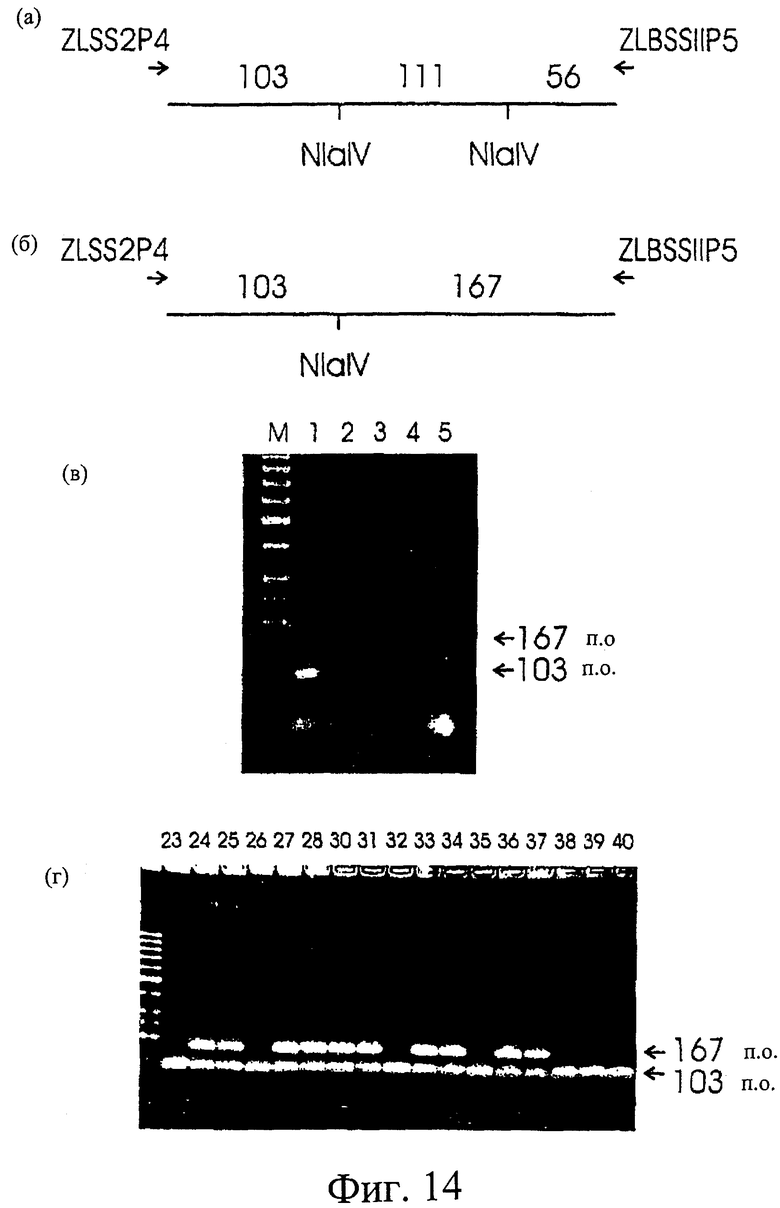
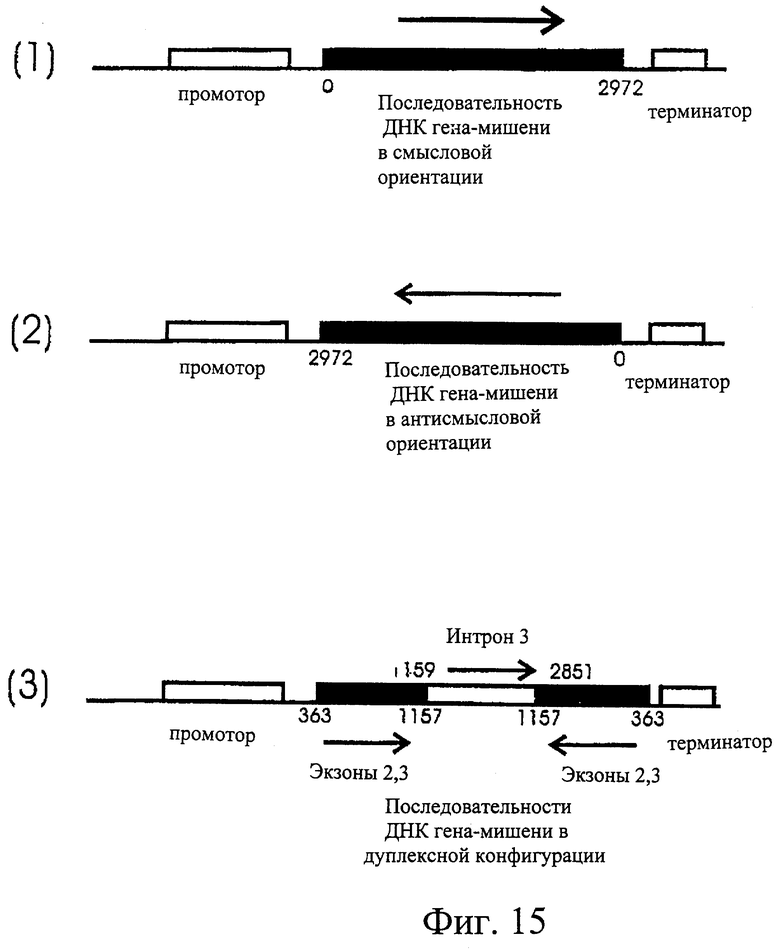
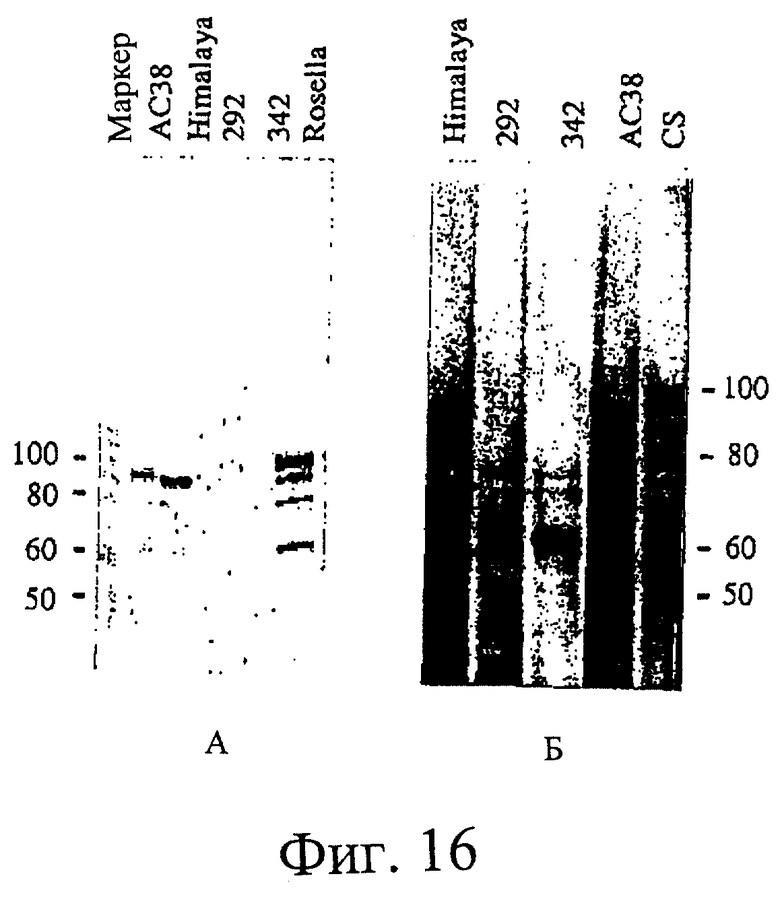
Изобретение относится к биотехнологии и пищевой промышленности. Растения ячменя подвергают мутации в гене SSII, которые понижают уровень активности SSII. Зерно такого ячменя имеет структуру крахмала с пониженным содержанием амилопектина и, в результате этого, с относительно высоким содержанием амилозы. Кроме того, это зерно может иметь относительно высокое содержание β-глюкана. Крахмал характеризуется пониженной вязкостью желатинизации и низкой кристалличностью, а также наличием высоких уровней связанного с липидом крахмала V-формы его кристалличности. 9 н. и 24 з.п. ф-лы, 39 ил., 10 табл.
ДНК, которая направляет синтез антисмысловой молекулы, которая препятствует транскрипции или процессингу гена SSII,
SSII-специфичного рибозима,
экстра-копии гена SSII для косупрессии гена SSII и
ДНК, которая направляет синтез двунитевого продукта РНК, опосредующего супрессию гена SSII.
а) доля цепей, имеющих длину в диапазоне от 6 до 11 остатков, составляет по меньшей мере 25%,
б) доля цепей, имеющих длину в диапазоне от 12 до 30 остатков, составляет менее 65%, и
в) доля цепей, имеющих длину в диапазоне от 31 до 60 остатков, составляет менее 10%.
а) вводят генетическую вариацию в родительское растение ячменя, которая представляет собой либо мутацию в гене SSII, либо трансген, которые понижают уровень активности SSII,
б) получают посевной материал от этого родительского растения ячменя и
в) осуществляют отбор семян, полученных в результате стадии (б), для тех семян, которые имеют пониженный уровень активности SSII, и
г) выращивают растение ячменя из семян со стадии (в).
а) измельчают зерно по любому из пп.1-8 и
б) экстрагируют крахмал из этого измельченного зерна.
| PLANT PHYSIOLOGY, 1999, vol.120, р.1147-55 | |||
| PLANT PHYSIOLOGY, 2000, vol.122, p.255-63 | |||
| PLANT CELL, 1998, vol.10, р.413-26 | |||
| GENOME, 2000, vol.43, р.768-75 | |||
| RU 96124069 А, 10.02.1999. |

