ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка является продолжением заявки США № 15/090926, поданной 5 апреля 2016 года, описание которой включено в настоящее описание в качестве ссылки в полном объеме для всех целей.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к кукурузному крахмалу, полученному из растений и зерен, композициям и способам получения такого крахмала.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Существующий крахмал кукурузы с двойной мутацией waxy-sugary-2 демонстрирует очень хорошую стабильность при замораживании/размораживании. Однако вязкость существующего крахмала кукурузы с двойной мутацией waxy-sugary-2 являлась, по существу, более низкой, чем у коммерческого крахмала кукурузы waxy, такого как крахмал AMIOCA™. Кроме того, температура клейстеризации существующего крахмала кукурузы с двойной мутацией waxy-sugary-2 являлась, по существу, более низкой, чем у крахмала AMIOCA™. Таким образом, в этой области существует потребность в получении кукурузного крахмала с хорошей стабильностью при замораживании/размораживании, повышенной вязкостью и/или более высокой температурой клейстеризации.
Авторы настоящего изобретения разрабатывали новый кукурузный крахмал с более высокой вязкостью и/или более высокой температурой клейстеризации, чем у коммерческого кукурузного крахмала waxy, такого как крахмал AMIOCA™, посредством встраивания GBSSI низкой активности в существующую кукурузу с двойной мутацией waxy-sugary-2.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к крахмалу с новыми функциями, не обнаруживаемыми в природе. В некоторых вариантах осуществления крахмал имеет содержание амилозы от приблизительно 2% масс. до приблизительно 20% масс., определяемое с использованием набор для анализа амилозы/амилопектина, например, Megazyme®. В некоторых вариантах осуществления крахмал имеет температуру клейстеризации водного раствора крахмала, составляющую по меньшей мере на приблизительно 5% больше, чем температура клейстеризации контрольного крахмала, измеряемая с использованием Rapid Visco Analyzer (RVA, Newport Scientific, Sydney, Australia). В некоторых вариантах осуществления крахмал получают из растения кукурузы с двойной мутацией waxy-sugary 2, содержащего по меньшей мере один аллель wx-Stonor (wxS) в эндосперме. В некоторых вариантах осуществления растение кукурузы с двойной мутацией waxy-sugary 2 содержит рецессивный мутантный аллель su2.
В некоторых вариантах осуществления крахмал по настоящему изобретению получают из растений, не являющихся трансгенными (т.е. не-ГМО) по генотипу, продуцирующих описываемый и заявленный в формуле изобретения крахмал.
В некоторых вариантах осуществления содержание амилозы в крахмале составляет от приблизительно 8% масс. до приблизительно 15% масс.
В некоторых вариантах осуществления температура клейстеризации водного раствора составляет на приблизительно 5%-15% больше, чем у контрольного крахмала.
В некоторых вариантах осуществления крахмал дополнительно имеет изменение энтальпии кристаллизации. В некоторых вариантах осуществления изменение энтальпии кристаллизации составляет по меньшей мере на приблизительно 30% больше, чем изменение энтальпии кристаллизации контрольного крахмала, как измеряют с использованием дифференциального сканирующего калориметра (DSC). В некоторых вариантах осуществления изменение энтальпии кристаллизации составляет на приблизительно 30%-200% больше, чем изменение энтальпии кристаллизации контрольного крахмала, как измеряют с использованием DSC.
В некоторых вариантах осуществления крахмал дополнительно имеет распределение размера гранул крахмала, где распределение размера гранул крахмала включает по меньшей мере на приблизительно 40% меньше гранул размером менее 5 мкм по сравнению с долей гранул размером менее 5 мкм в контрольном крахмале, что наблюдали с использованием сканирующего электронного микроскопа.
В некоторых вариантах осуществления крахмал имеет гранулы, демонстрирующие повышенную целостность по сравнению с крахмалом, полученным из контрольного растения кукурузы, что наблюдали с использованием сканирующего электронного микроскопа.
В некоторых вариантах осуществления крахмал способен выдерживать по меньшей мере три цикла замораживания/размораживания. В некоторых вариантах осуществления крахмал способен выдерживать по меньшей мере от трех до по меньшей мере пяти циклов замораживания/размораживания.
В некоторых вариантах осуществления растение, продуцирующее крахмал по настоящему изобретению, получают посредством перекрестного опыления первого растения кукурузы, имеющего генотип wxwxsu2su2, и второго растения кукурузы, имеющего генотип wxSwxSsu2su2 или генотип wxSwxsu2su2. В некоторых вариантах осуществления эндосперм растения кукурузы waxy-sugary 2 имеет одну дозу, две дозы или три дозы аллеля wxS, и эндосперм имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2.
Настоящее изобретение также относится к способам получения крахмала с улучшенным профилем клейстеризации. В некоторых вариантах осуществления способы включают встраивание по меньшей мере одного wxS аллеля в эндосперм растения кукурузы waxy.
Настоящее изобретение также относится к новым растениям с двойной мутацией. В некоторых вариантах осуществления растения являются растениями кукурузы. В некоторых вариантах осуществления растения являются гомозиготами по рецессивному гену синтазы крахмала IIa (su2). В некоторых вариантах осуществления растения являются гомозиготами или гетерозиготами по мутантному гену гранулосвязанной синтазы крахмала I (GBSSI). В некоторых вариантах осуществления мутантный ген GBSSI имеет меньшую активность GBSSI, чем ген GBSSI дикого типа (Wx), но более высокую активность GBSSI, чем рецессивный ген GBSSI с потерей функции (wx).
В некоторых вариантах осуществления мутантный ген GBSSI имеет от приблизительно 8% до приблизительно 10% активности гена GBSSIа GBSSI дикого типа. В некоторых вариантах осуществления мутантный ген GBSSI представляет собой аллель wx-Stonor (wxS). В некоторых вариантах осуществления мутантный ген GBSSI представляет собой аллель wx-G, аллель wx-B5 или аллель wx-M.
В некоторых вариантах осуществления растение с двойной мутацией получают посредством перекрестного опыления двух родительских растений. В некоторых вариантах осуществления первое растение имеет генотип wxwxsu2su2. В некоторых вариантах осуществления второе растение имеет генотип wxSwxSsu2su2 или генотип wxSwxsu2su2.
В некоторых вариантах осуществления эндосперм растения имеет одну дозу, две дозы или три дозы аллеля wxS. В некоторых вариантах осуществления эндосперм растения имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2.
В некоторых вариантах осуществления крахмал, продуцируемый в эндосперме растения, имеет более высокую вязкость по сравнению с крахмалом, продуцируемым в эндосперме растения с двойной рецессивной мутацией waxy-sugary-2, имеющего генотип wxwxwxsu2su2su2.
В некоторых вариантах осуществления крахмал, продуцируемый в эндосперме растения, имеет более высокую температуру клейстеризации крахмала по сравнению с крахмалом, продуцируемым в эндосперме растения с двойной рецессивной мутацией waxy-sugary-2, имеющего генотип wxwxwxsu2su2su2.
Настоящее изобретение также относится к частям растения, растительным клеткам и культурам ткани растений. В некоторых вариантах осуществления частью растения является эндосперм, лист, цветок, семязачаток, пыльца, корневище или черенок. В некоторых вариантах осуществления частью растения является семя растения.
Настоящее изобретение также относится к крахмалу, экстрагируемому из растений, описываемых выше.
Настоящее изобретение также относится к способам получения крахмала. В некоторых вариантах осуществления способы включают получение крахмала из эндосперма растений по настоящему изобретению.
В некоторых вариантах осуществления способы получения крахмала включают выращивание растений по настоящему изобретению и сбор семян растений.
Настоящее изобретение также относится к способам получения семени. В некоторых вариантах осуществления способы включают скрещивание первого родительского растения со вторым родительским растением и сбор полученных семян. В некоторых вариантах осуществления указанное первое родительское растение и/или второе родительское растение является растением по настоящему изобретению. В некоторых вариантах осуществления способы включают самоопыление растения по настоящему изобретению и сбор полученных семян.
Настоящее изобретение также относится к способам вегетативного размножения растений. В некоторых вариантах осуществления способы включают сбор частей растения и регенерацию растения из указанных частей.
Настоящее изобретение также относится к способам получения растения, полученного из растений по настоящему изобретению. В некоторых вариантах осуществления способы включают самоопыление растения по настоящему изобретению, по меньшей мере, однократно для получения растения-потомка из растения. В некоторых вариантах осуществления способы включают скрещивание растения по настоящему изобретению со вторым растением для получения растения-потомка.
Настоящее изобретение также относится к способам получения трансгенного растения. В некоторых вариантах осуществления способы включают встраивание интересующего трансгена в растение по настоящему изобретению. Интересующий трансген придает растению один или несколько желаемых фенотипов.
Настоящее изобретение также относится к способам получения растения с желаемыми фенотипами, включающим редактирование гена для получения генетической последовательности в растении по настоящему изобретению. Генетическая последовательность придает растению один или несколько желаемых фенотипов.
Настоящее изобретение также относится к способам получения растения с желаемыми фенотипами, включающим встраивание одной или несколько мутаций в растение по настоящему изобретению для получения генетической последовательности. Генетическая последовательность придает растению один или несколько желаемых фенотипов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 изображены профили клейстеризации крахмала AMIOCA™, крахмала wxwxwxSU2su2su2 и крахмала из растений с двойной мутацией waxy-sugary 2 с нулем, одной, двумя или тремя дозами гена wxS, как измеряют с помощью Rapid Visco Analyzer (RVA) в течение цикла нагревания-охлаждения.
На фиг. 2 изображены свойства клейстеризации крахмала AMIOCA™, крахмала wxwxwxSU2su2su2 и крахмала из растений с двойной мутацией waxy-sugary 2 с нулем, одной, двумя или тремя дозами гена wxS, включая температуру клейстеризации, RVU пика, RVU удержания, конечные RVU, RVU потери вязкости и setback RVU.
На фиг. 3 изображено тестирование стабильности при замораживании/размораживании крахмала из растений с двойной мутацией waxy-sugary 2 с одной, двумя, тремя или нулем доз гена wxS, крахмала из семян Waxy Stonor, крахмала wxwxwxSU2su2su2 и крахмала AMIOCA™. Указанные выше крахмалы показаны рядами, и пояснительные изображения гелей крахмалов представлены после первого замораживания/размораживания (цикла 1), после третьего замораживания/размораживания (цикла 3) и после пятого замораживания/размораживания (цикл 5).
На фиг. 4 изображены термические свойства крахмала AMIOCA™, крахмала wxwxwxSU2su2su2 и крахмала из растений с двойной мутацией waxy-sugary 2 с нулем, одной, двумя или тремя дозами гена wxS.
На Фиг. 5 изображено распределение размера гранул крахмала AMIOCA™, крахмала wxwxwxSU2su2su2 и крахмала из растений с двойной мутацией waxy-sugary 2 с нулем, одной, двумя или тремя дозами гена wxS.
На фиг. 6 изображена морфология гранул крахмала из растений с двойной мутацией waxy-sugary 2 с одной, двумя, тремя или нулем доз гена wxS, крахмала wxwxwxSU2su2su2, крахмала AMIOCA™ и крахмал из семян Waxy Stonor. Пояснительные изображения для каждого из указанных выше крахмалов получены с использованием сканирующего электронного микроскопа.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
В рамках изобретения, глагол "содержат" в описании и формуле изобретения и его спряжения используют в его неограничивающем смысле для обозначения того, что элементы после этого слова включены, но элементы, не указанные конкретно, не исключены.
Изобретение относится к композициям и способам получения растения. В рамках изобретения, термин "растение" относится к любому живому организму Plantae (т.е. любому роду/виду в царстве Растений). В некоторых вариантах осуществления растение способно продуцировать крахмал, включая, в качестве неограничивающих примеров, кукурузу, картофель, пшеницу, тапиоку, рис, картофель и сорго. В некоторых вариантах осуществления растение является растением кукурузы.
Изобретение относится к частям растений. В рамках изобретения, термин "часть растения" относится к любой части растения, включая, в качестве неограничивающих примеров, эмбрион, эндосперм, побег, корень, стебель, семя, прилистник, лист, лепесток, цветок, семязачаток, прицветник, ветку, черешок, междоузлие, кору, опушение, отросток, ризом, вайю, лацинию, семязачаток, пыльцу, тычинку и т.п.
Изобретение относится к генам, содержащим выделенную (например, дикого типа, эндогенную для организма), химерную, рекомбинантную или синтетическую последовательность нуклеиновой кислоты. В рамках изобретения, термин "ген" относится к любому сегменту ДНК, ассоциированному с биологической функцией. Таким образом, гены включают, в качестве неограничивающих примеров, кодирующие последовательности и/или регуляторные последовательности, необходимые для их экспрессии. Гены также могут включать неэкспрессирующиеся сегменты ДНК, например, образующие последовательности распознавания для других белков. Гены можно получать из множества источников, включая клонирование из интересующего источника или синтез с помощью информации об известной или предсказанной последовательности, и они могут включать последовательности, сконструированные так, что они имеют желаемые параметры.
Изобретение относится к полинуклеотидам с заменой нуклеотидов по сравнению с референсной последовательностью дикого типа. В рамках изобретения, термин "замена нуклеотида" относится, например, к замене, делеции, инсерции нуклеотида или транспозону, как хорошо известно в этой области. Например, мутации включают изменения, приводящие к молчащим заменам, добавлениям, делециям или альтернативному сплайсингу, изменяющему или не изменяющему свойства или активности кодируемого белка или то, как белки получают. Замена нуклеотида также может являться инсерцией и/или вырезанием ретротранспозонного элемента в гене.
Изобретение относится к полипептидам с модификацией белка по сравнению с референсной последовательностью дикого типа. В рамках изобретения, термин "модификация белка" относится, например, к замене аминокислоты, модификации, делеции, инсерции аминокислоты или изменению активности белка, как хорошо известно в этой области.
Изобретение относится к полинуклеотидам и полипептидам, полученным из референсных последовательностей дикого типа. В рамках изобретения, термин "полученный из" относится к происхождению или источнику и может включать природные, рекомбинантные, неочищенные или очищенные молекулы. Нуклеиновая кислота или аминокислота, полученная из источника, может иметь все типы замен нуклеотидов или модификации белка, как определено где-либо в настоящем описании.
Изобретение относится к биологически активным вариантам или функциональным вариантам последовательностей нуклеиновой кислоты и полипептидных последовательностей по настоящему изобретению. В рамках изобретения, фраза "биологически активный вариант" или "функциональный вариант" в отношении белка относится к аминокислотной последовательности, измененной в одной или нескольких аминокислотах относительно референсной последовательности, все равно сохраняющей существенную биологическую активность референсной последовательности. Вариант может иметь "консервативные" замены, где замещенная аминокислота имеет схожие структурные или химические свойства, например, замену лейцина изолейцином. В следующей таблице приведены примеры консервативных аминокислотных замен. В некоторых вариантах осуществления вариант имеет одну или несколько замен аминокислот, где одна, или несколько, или все замены представляют собой кислую аминокислоту, такую как аспарагиновая кислота, аспарагин, глутаминовая кислота или глутамин.
Альтернативно, вариант может иметь "неконсервативные" замены, например, замену глицина триптофаном. Аналогичные минорные варианты также могут включать делецию или инсерцию аминокислот, или и то, и другое. Руководство по определению того, какие аминокислотные остатки можно подвергать замене, инсерции или делеции без устранения биологической или иммунологической активности, можно найти с использованием компьютерных программ, хорошо известных в этой области, например, программного обеспечения DNASTAR. В случае полинуклеотидов вариант содержит полинуклеотид, имеющий делеции (т.е. укорачивания) на 5'- и/или 3'-конце; делеции и/или добавления одного или нескольких нуклеотидов в одном или нескольких внутренних участках референсного полинуклеотида; и/или замену одного или нескольких нуклеотидов в одном или нескольких участках в референсном полинуклеотиде. В рамках изобретения, "референсный" полинуклеотид содержит нуклеотидную последовательность, получаемую способами, представленными в настоящем описании. Варианты полинуклеотидов также включают синтетически полученные полинуклеотиды, такие как получаемые, например, с использованием сайт-специфического мутагенеза, но все равно содержащие генетический регуляторный элемент. Как правило, варианты конкретного полинуклеотида или молекулы нуклеиновой кислоты, или полипептида по изобретению имеют по меньшей мере приблизительно 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 91,5%, 92%, 92,5%, 93%, 93,5%, 94%, 94,5%, 95%, 95,5%, 96%, 96,5%, 97%, 97,5%, 98%, 98,5%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или более идентичности последовательности по отношению к полинуклеотиду/полипептидам, как определяют с помощью программ и параметров для выравнивания последовательностей, как представлено где-либо в настоящем описании.
Варианты полинуклеотидов также включают последовательности, полученные способом мутагенеза и рекомбинации, такого как ДНК-перестановка. Стратегии такой ДНК-перестановки известны в этой области. См., например, Stemmer (1994) PNAS 91:10747-10751; Stemmer (1994) Nature 370:389-391; Crameri et al. (1997) Nature Biotech. 15:436-438; Moore et al. (1997) J. Mol. Biol. 272:336-347; Zhang et al. (1997) PNAS 94:4504-4509; Crameri et al. (1998) Nature 391:288-291; и патенты США №№ 5605793 и 5837458. В случае ПЦР-амплификации полинуклеотидов, представленных в настоящем описании, можно конструировать олигонуклеотидные праймеры для использования в реакциях ПЦР для амплификации соответствующих последовательностей ДНК из кДНК или геномной ДНК, выделенной из любого интересующего растения. Способы конструирования праймеров для ПЦР и ПЦР-клонирования, как правило, известны в этой области и описаны в Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2nd ed., Cold Spring Harbor Laboratory Press, Plainview, New York). See also Innis et al., eds. (1990) PCR Protocols: A Guide to Methods and Applications (Academic Press, New York); Innis and Gelfand, eds. (1995) PCR Strategies (Academic Press, New York); and Innis and Gelfand, eds. (1999) PCR Methods Manual (Academic Press, New York). Известные способы ПЦР включают, в качестве неограничивающих примеров, способы с использованием парных праймеров, праймеров для гнездовой ПЦР, отдельных специфичных праймеров, вырожденных праймеров, гено-специфичных праймеров, вектор-специфичных праймеров, частично несовпадающих праймеров и т.п.
Изобретение относится к трансгенным растениям. В рамках изобретения, термин "трансгенный" относится к клеткам, культурам клеток, организмам (например, растениям) и потомству, в который встраивали чужеродный или модифицированный ген одним из разных способов трансформации, где чужеродный или модифицированный ген происходит из одного вида или вида, иного, чем вид организма, в который встраивали чужеродный или модифицированный ген.
Ген Waxy: ген Waxy является геном гранулосвязанной синтазы крахмала I (GBSSI). Ген кодирует гранулосвязанную ADP-глюкоза-глюкозилтрансферазу крахмала (EC 2.4.1.242), отвечающую за биосинтез амилозы в триплоидном эндосперме развивающегося зерна и в гаплоидной пыльце и зародышевом мешке. "Wx" относится к аллелю дикого типа, и "wx" относится к нулевому, референсному мутантному аллелю. Термины "wxS" (также известный как wx-Stonor), "wx-G", "wx-B5" и "wx-M" относятся к промежуточным аллелям, имеющим инсерцию ретротранспозона в гене waxy, как описано в Wessler and Varagona (PNAS, 82:4177-4181, 1985) и Varagona et al. (The Plant Cell, 4:811-820, 1992), каждый из которых включен в настоящее описание в качестве ссылки в полном объеме. Эти промежуточные аллели приводят к меньшей активности белка waxy в растениях, имеющих аллели, чем активность белка waxy в растениях дикого типа. Ген waxy кукурузы кодирует белок, имеющий последовательность SEQ ID NO: 1.
wx-Stonor: аллель wx-Stonor в растении кукурузы образуется вследствие инсерции ретротранспозона в участке донора сплайсингового фрагмента интрона 5 гена waxy. Хотя инсерция не влияет на аппарат сплайсинга, она приводит к более низкому уровню ферментативной активности по сравнению с аллелем дикого типа (например, лишь приблизительно 9,5% активности GBSSI относительно дикого типа). В результате растения, имеющие аллель wx-Stonor, имеют более высокий уровень амилозы, чем то же растение, не имеющее аллель wx-Stonor.
Ген Sugary-2: ген Sugary-2 кодирует изоформу синтазы крахмала IIa (SSIIa, EC 2.4.1.21). "SU2" относится к аллелю дикого типа, и "su2" относится к рецессивному мутантному аллелю. Функция белка Sugary-2 в кукурузе описана в Zhang et al. (Plant Molecular Biology, 54(6):865-879, 2004), включенной в настоящее описание в качестве ссылки в полном объеме. Ген SU2 кукурузы дикого типа кодирует белок дикого типа, имеющий последовательность SEQ ID NO: 2.
Более подробно ген waxy и ген sugary-2 кукурузы описаны в патентах США №№ 4428972, 5954883, 6828474, 6960703 и 7678898, каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
Крахмал AMIOCA™: термин относится к кукурузному крахмалу AMIOCA™, производимому National Starch and Chemical Company, в настоящее время - Ingredion Incorporated.
Крахмал wxwxwxSU2su2su2: термин относится к крахмалу, выделенному из эндосперма кукурузы, имеющей генотип wxwxwxSU2su2su2.
Крахмал wxwxwxsu2su2su2: термин относится к крахмалу, выделенному из эндосперма кукурузы, имеющему генотип wxwxwxsu2su2su2, являющегося эндоспермом с двойной рецессивной мутацией waxy-sugary-2. Мутантный ген waxy способствует его хорошей стабильности при замораживании/размораживании, и мутантный ген sugary-2 приводит к короткой разветвленной цепи. Однако крахмал имеет низкую целостность гранул крахмала. Гранулы легко разламываются, и крахмал имеет низкую вязкость.
Крахмал MELOJEL®: термин относится к кукурузному крахмалу MELOJEL®, производимому National Starch and Chemical Company, в настоящее время - Ingredion Incorporated.
Контрольное растение кукурузы: термин относится к растению для надлежащей проверки в целях сравнения. В некоторых вариантах осуществления растение для проверки может являться растением дикого типа. В некоторых вариантах осуществления растение для проверки является растением, не имеющим промежуточный ген waxy, как представлено в настоящем описании. В некоторых вариантах осуществления растение для проверки является растением кукурузы, продуцирующим крахмал wxwxwxSU2su2su2 или крахмал wxwxwxsu2su2su2. Таким образом, в некоторых вариантах осуществления контрольное растение кукурузы является растением кукурузы, не имеющим аллель wx-S в эндосперме.
Контрольный крахмал: термин относится к контрольному крахмалу.
wxSwxwxsu2su2su2: термин относится к эндосперму кукурузы с одной дозой wxS.
Крахмал wxSwxwxsu2su2su2: термин относится к крахмалу, выделенному из эндосперма кукурузы с одной дозой wxS, эндосперма, имеющего генотип wxSwxwxsu2su2su2.
wxSwxSwxsu2su2su2: термин относится к эндосперму кукурузы с двумя дозами wxS.
wxSwxSwxsu2su2su2 крахмал: термин относится к крахмалу, выделенному из эндосперма кукурузы с двумя дозами wxS, эндосперма, имеющего генотип wxSwxSwxsu2su2su2.
wxSwxSwxSsu2su2su2: термин относится к эндосперму кукурузы с тремя дозами wxS.
wxSwxSwxSsu2su2su2 крахмал: термин относится к крахмалу, выделенному из эндосперма кукурузы с тремя дозами wxS, эндосперма, имеющего генотип wxSwxSwxSsu2su2su2.
В следующих примерах проиллюстрированы различные аспекты по изобретению. Разумеется, следует понимать, что примеры предназначены исключительно для иллюстрирования конкретных вариантов осуществления изобретения, а не ограничения объема изобретения.
Крахмал с новыми функциями
В одном из аспектов настоящее изобретение относится к крахмалу, полученному из растения кукурузы с двойной мутацией, содержащего промежуточный аллель GBSSI, имеющий меньшую активность GBSSI, чем ген GBSSI дикого типа (Wx), но более высокую активность GBSSI, чем рецессивный ген GBSSI с потерей функции (wx). В некоторых вариантах осуществления растение кукурузы с двойной мутацией с промежуточным аллелем GBSSI является растением кукурузы waxy-sugary 2, содержащим по меньшей мере один аллель wx-Stonor (wxS) в эндосперме. В некоторых вариантах осуществления растение кукурузы waxy-sugary 2 получают посредством перекрестного опыления первого растения кукурузы, имеющего генотип wxwxsu2su2, и второго растения кукурузы, имеющего генотип wxSwxSsu2su2 или генотип wxSwxsu2su2. В дополнительных вариантах осуществления эндосперм растения кукурузы waxy-sugary 2 имеет одну дозу, две дозы или три дозы аллеля wxS, и эндосперм имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2.
В некоторых вариантах осуществления крахмал имеет содержание амилозы от приблизительно 2% масс. до приблизительно 20% масс. при измерении с использованием набора для анализа амилозы/амилопектина Megazyme®, и температуру клейстеризации водного раствора крахмала, составляющую по меньшей мере приблизительно на 5% больше, чем температура клейстеризации контрольного крахмала, при измерении с использованием анализа RVA.
Содержание амилозы. Крахмал состоит из амилозы и амилопектина. Амилоза является спирополимером, состоящим из единиц α-D-глюкозы, соединенных α(1→4)-гликозидными связями. Амилопектин является сильноразветвленным полимером глюкозы, содержащим α(1→4)- и α(1→6)-гликозидные связи. Разветвление происходит с использованием α(1→6)-связей, расположенных приблизительно через каждые 24-30 единиц глюкозы. В некоторых вариантах осуществления содержание амилозы в крахмале можно измерять с использованием набора для анализа амилозы/амилопектина, такого как набор для анализа амилозы/амилопектины, производимый Megazyme®, как указано в примерах.
Наблюдали, что промежуточная мутация гена GBSSI в растении кукурузы с двойной мутацией повышает содержание амилозы в полученном из него крахмале относительно контрольного растения кукурузы с нулевым аллелем гена GBSSI. Например, наблюдали, что содержание амилозы крахмала, полученного из растения кукурузы waxy-sugary 2, содержащего по меньшей мере один аллель wx-Stonor (wxS) в эндосперме, повышается относительно растения кукурузы. В некоторых вариантах осуществления крахмал, полученный из растения кукурузы по меньшей мере с одним аллелем wx-Stonor (wxS) в эндосперме, имеет содержание амилозы от приблизительно 2% масс. до приблизительно 20% масс., от приблизительно 4% масс. до приблизительно 18% масс., от приблизительно 5% масс. до приблизительно 15% масс. или приблизительно 8% масс. до приблизительно 15% масс., включая все значения и поддиапазоны. В других вариантах осуществления содержание амилозы составляет приблизительно 5% масс., приблизительно 6% масс., приблизительно 7% масс., приблизительно 8% масс., приблизительно 9% масс., приблизительно 10% масс., приблизительно 11% масс., приблизительно 12% масс., приблизительно 13% масс., приблизительно 14% масс., приблизительно 15% масс., приблизительно 16% масс., приблизительно 17% масс., или приблизительно 18% масс., включая все значения между ними.
Не будучи связанными какой-либо теорией, ожидают, что повышение содержания амилозы будет приводить к снижению содержания амилопектина в крахмале и, таким образом, снижению общего количества единиц глюкозы и групп с водородными связями, доступными для взаимодействия с водной средой. Такое снижение водородных связей должно соответствовать общему снижению силы взаимодействий указанного крахмала с водной средой и снижению свойств клейстеризации крахмала. Однако наблюдали неожиданное улучшение профиля клейстеризации при повышении содержания амилозы в крахмале, представленном в настоящем описании. Не будучи связанными какой-либо теорией, этот неожиданный результат можно приписать повышению длины разветвленной цепи, наблюдаемому в растении кукурузы waxy-sugary 2, содержащем по меньшей мере один аллель wx-Stonor (wxS) в эндосперме. Более длинные разветвленные цепи в крахмале содержат больше единиц глюкозы, что повышает количество взаимодействий посредством водородных связей с водной средой по сравнению с более короткими разветвленными цепями.
Профиль клейстеризации крахмала можно измерять известными способами, такими как с использованием Rapid Visco Analyzer (RVA) (Newport Scientific, Sydney, Australia). Например, с помощью RVA можно измерять температуру клейстеризации (°C), пиковая вязкость (RVU), вязкость удержания (RVU), конечная вязкость (RVU), потери вязкости (RVU) и set back вязкость (RVU). В некоторых вариантах осуществления улучшенный профиль клейстеризации делает возможной желатинизацию, помимо прочего, при более низком эффективном количестве крахмала и/или при более высокой температуре клейстеризации по сравнению с контрольным крахмалом. Желатинизация крахмала относится к процессу разрыва межмолекулярных связей молекул крахмала в присутствие воды и повышенной температуры, что позволяет участкам водородных связей (например, гидроксильных групп) связывать больше воды. Для измерения профиля клейстеризации крахмала можно добавлять эффективное количество крахмала в водный раствор при нейтральном pH. Например, крахмал можно добавлять в водный раствор при 8% масс./масс. твердых веществ.
Температура клейстеризации. В рамках изобретения, температуру клейстеризации предполагают измерять, как указано в примерах. Температура клейстеризации крахмала является температурой, при которой происходит исходное набухание гранул крахмала при суспендировании в воде. В некоторых вариантах осуществления крахмал, представленный в настоящем описании, имеет температуру клейстеризации водного раствора, составляющую по меньшей мере на приблизительно 5% больше, чем температура клейстеризации контрольного крахмала, как измеряют с использованием анализа RVA. В некоторых вариантах осуществления температуру клейстеризации водного раствора повышают на от приблизительно 1% до приблизительно 100%, от приблизительно 1% до приблизительно 50%, от приблизительно 5% до приблизительно 50%, от приблизительно 5% до 40%, от приблизительно 5% до приблизительно 30%, от приблизительно 5% до приблизительно 25%, от приблизительно 5% до приблизительно 20% или от приблизительно 5% до приблизительно 14%, включая все значения и поддиапазоны. В конкретном варианте осуществления температура клейстеризации водного раствора составляет на от приблизительно 5% до приблизительно 14% больше, чем у контрольного растения кукурузы, такого как растение кукурузы с двойной мутацией waxy-sugary 2.
В других вариантах осуществления температуру клейстеризации водного раствора повышают по меньшей мере на приблизительно 2%, по меньшей мере на приблизительно 3%, по меньшей мере на приблизительно 4%, по меньшей мере на приблизительно 5%, по меньшей мере на приблизительно 6%, по меньшей мере на приблизительно 7%, по меньшей мере на приблизительно 8%, по меньшей мере на приблизительно 9%, по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 11%, по меньшей мере на приблизительно 12%, по меньшей мере на приблизительно 13%, по меньшей мере на приблизительно 14%, по меньшей мере на приблизительно 15%, по меньшей мере на приблизительно 15%, по меньшей мере на приблизительно 16%, по меньшей мере на приблизительно 17%, по меньшей мере на приблизительно 18%, по меньшей мере на приблизительно 19% или по меньшей мере на приблизительно 20% больше по сравнению с температурой клейстеризации контрольного крахмала, включая все значения между ними.
В других вариантах осуществления температура клейстеризации водного раствора повышают на от 1°C до приблизительно 20°C или от приблизительно 5°C до приблизительно 10°C, включая все значения и поддиапазоны. В других вариантах осуществления температуру клейстеризации водного раствора повышают по меньшей мере на приблизительно 4°C, по меньшей мере на приблизительно 5°C, по меньшей мере на приблизительно 6°C, по меньшей мере на приблизительно 7°C, по меньшей мере на приблизительно 8°C, по меньшей мере на приблизительно 9°C, по меньшей мере на приблизительно 10°C, по меньшей мере на приблизительно 11°C, по меньшей мере на приблизительно 12°C, по меньшей мере на приблизительно 13°C, по меньшей мере на приблизительно 14°C, или на приблизительно 15°C или 15°C по сравнению с температурой клейстеризации контрольного крахмала.
В других вариантах осуществления крахмал, представленный в настоящем описании, имеет температуру клейстеризации водного раствора по меньшей мере приблизительно 50°C, по меньшей мере приблизительно 55°C, по меньшей мере приблизительно 60°C, по меньшей мере приблизительно 65°C, по меньшей мере приблизительно 70°C, по меньшей мере приблизительно 75°C, по меньшей мере приблизительно 80°C, по меньшей мере приблизительно 85°C, по меньшей мере приблизительно 90°C, по меньшей мере приблизительно 95°C или по меньшей мере приблизительно 100°C. В конкретных вариантах осуществления крахмал, полученный из растения кукурузы waxy-sugary 2, содержащего по меньшей мере один аллель wx-Stonor (wxS) в эндосперме, имеет температуру клейстеризации водного раствора от приблизительно 55°C до приблизительно 68°C или от приблизительно 59°C до приблизительно 65°C, включая все значения и поддиапазоны. В других вариантах осуществления температура клейстеризации водного раствора составляет приблизительно 59°C, приблизительно 60°C, приблизительно 61°C, приблизительно 62°C, приблизительно 63°C, приблизительно 64°C, приблизительно 65°C, приблизительно 66°C, приблизительно 67°C или приблизительно 68°C, включая все значения между ними.
Изменение энтальпии кристаллизации. В рамках изобретения, изменение энтальпии кристаллизации предполагают измерять, как указано в примерах. Кристаллизация относится к процессу образования твердых кристаллов из раствора посредством осаждения. В некоторых вариантах осуществления наблюдали, что встраивание по меньшей мере одного аллеля wx-Stonor (wxS) в эндосперм растения кукурузы с двойной мутацией waxy-sugary 2 повышает изменение энтальпии кристаллизации относительно крахмала из контрольного растения кукурузы. Изменение энтальпии можно измерять с использованием, например, дифференциальной сканирующей калориметрии (DSC). В некоторых вариантах осуществления изменение энтальпии кристаллизации повышают на от приблизительно 10% до приблизительно 300%, от приблизительно 10% до приблизительно 200%, от приблизительно 10% до приблизительно 150%, от приблизительно 10% до приблизительно 100% или от приблизительно 10% до приблизительно 50%, включая все значения и поддиапазоны. В одном таком варианте осуществления изменение энтальпии кристаллизации составляет на от приблизительно 70% до приблизительно 200% больше, чем изменение энтальпии кристаллизации контрольного крахмала, как измеряют с использованием DSC.
В некоторых вариантах осуществления изменение энтальпии кристаллизации повышают по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 20%, по меньшей мере на приблизительно 30%, по меньшей мере на приблизительно 40%, по меньшей мере на приблизительно 50%, по меньшей мере на приблизительно 60%, по меньшей мере на приблизительно 70%, по меньшей мере на приблизительно 80%, по меньшей мере на приблизительно 90%, по меньшей мере на приблизительно 100%, по меньшей мере на приблизительно 110%, по меньшей мере на приблизительно 120%, по меньшей мере на приблизительно 130%, по меньшей мере на приблизительно 140%, по меньшей мере на приблизительно 150%, по меньшей мере на приблизительно 160%, по меньшей мере на приблизительно 170%, по меньшей мере на приблизительно 180%, по меньшей мере на приблизительно 190% или по меньшей мере на приблизительно 200%, включая все значения между ними.
В некоторых вариантах осуществления изменение энтальпии кристаллизации составляет от приблизительно 1 Дж/г до приблизительно 20 Дж/г, от приблизительно 5 Дж/г до приблизительно 15 Дж/г, от приблизительно 6 Дж/г до приблизительно 13 Дж/г, включая все значения и поддиапазоны. В других вариантах осуществления изменение энтальпии кристаллизации 8% масс./масс. составляет приблизительно 4 Дж/г, приблизительно 5 Дж/г, приблизительно 6 Дж/г, приблизительно 7 Дж/г, приблизительно 8 Дж/г, приблизительно 9 Дж/г, приблизительно 10 Дж/г, приблизительно 11 Дж/г, приблизительно 12 Дж/г, приблизительно 13 Дж/г, включая все значения между ними. В конкретном варианте осуществления растение кукурузы содержит один аллель wxS в эндосперме, и изменение энтальпии кристаллизации составляет по меньшей мере на приблизительно 150% больше, чем у крахмала, полученного из контрольного растения с генотипом эндосперма wxwxwxsu2su2su2. В другом конкретном варианте осуществления растение кукурузы содержит два аллеля wxS в эндосперме, и изменение энтальпии кристаллизации составляет по меньшей мере на приблизительно 30% больше, чем у крахмала, полученного из контрольного растения с генотипом эндосперма wxwxwxsu2su2su2. В другом конкретном варианте осуществления растение кукурузы содержит три аллеля wxS в эндосперме, и изменение энтальпии кристаллизации составляет по меньшей мере на приблизительно 75% больше, чем у крахмала, полученного из контрольного растения с генотипом эндосперма wxwxwxsu2su2su2.
Стабильность при замораживании/размораживании. В рамках изобретения, стабильность при замораживании/размораживании предполагают измерять, как указано в примерах. В некоторых вариантах осуществления крахмал, представленный в настоящем описании, проявляет желаемые характеристики при замораживании/размораживании, например, улучшенную стабильность при замораживании/размораживании. Стабильность при замораживании/размораживании можно измерять посредством добавления эффективного количества крахмала в воду для получения суспензии, которую затем варят и измеряют синерезис после циклов замораживания/охлаждения. В некоторых вариантах осуществления встраивание по меньшей мере одного аллеля wx-Stonor (wxS) в эндосперм растения кукурузы не снижает стабильность полученного из него крахмала при замораживании/размораживании по сравнению с контрольным крахмалом. В других вариантах осуществления встраивание по меньшей мере одного аллеля wx-Stonor (wxS) в эндосперм растения кукурузы улучшало стабильность крахмала при замораживании/размораживании. В одном таком варианте осуществления крахмал способен выдерживать всего по меньшей мере от трех до по меньшей мере пяти циклов замораживания/размораживания.
Распределение и целостность гранул. В рамках изобретения, распределение и целостность гранул предполагают измерять, как указано в примерах.
В некоторых вариантах осуществления наблюдали, что встраивание по меньшей мере одного аллеля wx-Stonor (wxS) в эндосперм растения кукурузы изменяет распределение размеров гранул крахмала относительно контрольного крахмала. В некоторых таких вариантах осуществления распределение размера гранул крахмала, представленного в настоящем описании, включает по меньшей мере на приблизительно 40% меньше гранул размером менее 5 мкм по сравнению с долей гранул размером менее 5 мкм в контрольном крахмале, что наблюдали с использованием сканирующего электронного микроскопа.
В рамках изобретения, целостность гранул относится к способности крахмала сохраняться в виде интактной гранулы после растворения в водной среде. В некоторых вариантах осуществления крахмал, представленный в настоящем описании, является гранулой, проявляющей повышенную целостность по сравнению с крахмалом, полученным из контрольного растения кукурузы, что наблюдали с использованием сканирующего электронного микроскопа.
Вязкость. В рамках изобретения, вязкость предполагают измерять, как указано в примерах. В некоторых вариантах осуществления крахмал, представленный в настоящем описании, имеет пиковую вязкость в водном растворе, составляющую по меньшей мере на приблизительно 10% больше, чем пиковая вязкость водного раствора контрольного крахмала, как измеряют с использованием анализа RVA. В некоторых вариантах осуществления пиковую вязкость водного раствора повышали на от приблизительно 10% до приблизительно 300%, от приблизительно 10% до приблизительно 250%, от приблизительно 10% до приблизительно 200%, от приблизительно 10% до приблизительно 150%, от приблизительно 10% до приблизительно 100% или от приблизительно 10% до приблизительно 50%, включая все значения и поддиапазоны. В некоторых вариантах осуществления пиковую вязкость водного раствора повышали по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 20%, по меньшей мере на приблизительно 30%, по меньшей мере на приблизительно 40%, по меньшей мере на приблизительно 50%, по меньшей мере на приблизительно 60%, по меньшей мере на приблизительно 70%, по меньшей мере на приблизительно 80%, по меньшей мере на приблизительно 90%, по меньшей мере на приблизительно 100%, по меньшей мере на приблизительно 110%, по меньшей мере на приблизительно 120%, по меньшей мере на приблизительно 130%, по меньшей мере на приблизительно 140%, по меньшей мере на приблизительно 150%, по меньшей мере на приблизительно 160%, по меньшей мере на приблизительно 170%, по меньшей мере на приблизительно 180%, по меньшей мере на приблизительно 190% или по меньшей мере на приблизительно 200%, включая все значения между ними.
В некоторых вариантах осуществления пиковая вязкость водного раствора крахмала при 8% масс./масс. твердых веществ составляет от приблизительно 100 RVU до приблизительно 500 RVU, от приблизительно 100 RVU до приблизительно 400 RVU, от приблизительно 100 RVU до приблизительно 300 RVU, от приблизительно 100 RVU до приблизительно 200 RVU или от приблизительно 100 RVU до приблизительно 150 RVU, включая все значения и поддиапазоны. В некоторых вариантах осуществления пиковая вязкость водного раствора составляет приблизительно 150 RVU, приблизительно 155 RVU, приблизительно 160 RVU, приблизительно 165 RVU, приблизительно 170 RVU, приблизительно 175 RVU, приблизительно 180 RVU, приблизительно 185 RVU, приблизительно 190 RVU, приблизительно 195 RVU, приблизительно 200 RVU, приблизительно 205 RVU, приблизительно 210 RVU, приблизительно 215 RVU, приблизительно 220 RVU, приблизительно 225 RVU, приблизительно 230 RVU, приблизительно 235 RVU, приблизительно 240 RVU, приблизительно 245 RVU, приблизительно 250 RVU, приблизительно 255 RVU, приблизительно 260 RVU, приблизительно 265 RVU, приблизительно 270 RVU, приблизительно 275 RVU, приблизительно 280 RVU, приблизительно 285 RVU, приблизительно 290 RVU, приблизительно 295 RVU, приблизительно 300 RVU, приблизительно 305 RVU, приблизительно 310 RVU, приблизительно 315 RVU, приблизительно 320 RVU, приблизительно 325 RVU, приблизительно 330 RVU, приблизительно 335 RVU, приблизительно 340 RVU, приблизительно 345 RVU, приблизительно 350 RVU, приблизительно 355 RVU, приблизительно 360 RVU, приблизительно 365 RVU, приблизительно 370 RVU, приблизительно 375 RVU, приблизительно 380 RVU, приблизительно 385 RVU, приблизительно 390 RVU, приблизительно 395 RVU или приблизительно 400 RVU, включая все значения между ними.
В конкретном варианте осуществления растение кукурузы waxy-sugary 2 содержит один аллель wx-Stonor (wxS) в эндосперме, и крахмал имеет пиковую вязкость на приблизительно 110% больше, чем у контрольного растения с генотипом эндосперма wxwxwxsu2su2su2. В другом конкретном варианте осуществления растение кукурузы waxy-sugary 2 содержит два аллеля wx-Stonor (wxS) в эндосперме, и крахмал имеет пиковую вязкость на приблизительно 30% больше, чем у контрольного растения с генотипом эндосперма wxwxwxsu2su2su2. В другом конкретном варианте осуществления растение кукурузы waxy-sugary 2 содержит три аллеля wx-Stonor (wxS) в эндосперме, и крахмал имеет пиковую вязкость на приблизительно 40% больше, чем у контрольного растения с генотипом эндосперма wxwxwxsu2su2su2. В другом конкретном варианте осуществления растение кукурузы waxy-sugary 2 содержит два аллеля wx-Stonor (wxS) в эндосперме, и крахмал имеет пиковую вязкость на приблизительно 20% больше, чем у контрольного растения с генотипом эндосперма wxwxwxSU2su2su2. В другом конкретном варианте осуществления растение кукурузы waxy-sugary 2 содержит два аллеля wx-Stonor (wxS) в эндосперме, и крахмал имеет пиковую вязкость, по меньшей мере на приблизительно 30% больше, чем у контрольного растения с генотипом эндосперма wxwxwxSU2su2su2.
Экстрагирование. Экстрагирование крахмала из зерна растений по настоящему изобретению можно осуществлять стандартным способом посредством мокрого размола или сухого размола, хорошо известных в этой области, но такие способы являются неограничивающими примерами. В одном типичном способе мокрого размола растение очищают сильными струями воздуха, с помощью сита и электромагнитов для удаления нежелательного материала. Затем его погружают в теплую воду, содержащую небольшое количество диоксида серы. Воду для замачивания откачивают и размягченные зерна пропускают через рафинаторы для их разделения. Зародыш удаляют и оставшуюся смесь измельчают, промывают и просеивают как суспензию. Крахмал отделяют от глютена посредством центрифугирования, а затем оставшийся суспендированный крахмал фильтруют, промывают, ресуспендируют и повторно фильтруют.
Экстрагирование муки или ее вариантов из растения можно осуществлять посредством сухого размола. В типичном способе, подходящем для настоящего описания, но не исключающем другие способы, зерно, полученное из растения по настоящему изобретению, сначала тщательно очищают и пропускают через лущильный аппарат, затем смягчают или кондиционируют и пропускают через дегерминатор. Сток из дегерминатора сушат, а затем охлаждают, пропускают через сепаратор для дробленой кукурузы и аспиратор, измельчают и, в конечном итоге, просеивают в соответствии с тем, являются ли желательными цельные или разделенные фракции.
Модификация. Следует понимать, что некоторые модификации указанных выше способов экстрагирования, таких как, например, с использованием температуры воды для замачивания ниже той, которую, как правило, используют, могут являться желательными и будут известны практикующему специалисту в области крахмала.
Крахмал по настоящему изобретению можно, при желании, модифицировать известными в этой области способами, такими как дериватизация для получения простых эфиров или сложных эфиров, таких как гидроксипропиловые простые эфиры, ацетаты, фосфаты, сукцинаты, например, октенилсукцинат, простые эфиры третичных и четвертичных аминов и т.д., или любыми другими способами модификации, которыми получают крахмал, имеющий характеристики, определенные в настоящем описании. Предпочтительными группами заместителей в настоящем описании являются гидроксипропильные, фосфатные или ацетатные группы.
В коммерческих целях одной из модификаций крахмала по настоящему изобретению является перекрестная сшивка для укрепления гранулы относительно условий хранения и обработки, часто встречающихся при производственных операциях, и для получения крахмала, способного обеспечивать желаемые реологические свойства в пищевых системах. Для этой цели можно использовать любое сшивающее средство, известное в этой области, включая, в качестве неограничивающих примеров, эпихлоргидрин, линейные ангидриды дикарбоновых кислот, лимонную кислоту, акролеин, оксихлорид фосфора, смешанные ангидриды адипиновой/уксусной кислот и тримефосфатные соли для пищевых систем и эпихлоргидрин, линейные ангидриды дикарбоновых кислот, лимонную кислоту, акролеин, оксихлорид фосфора, смешанные ангидриды адипиновой/уксусной кислот, тримефосфатные соли, формальдегид, цианурхлорид, диизоцианаты и дивинилсульфоны в непищевых системах. Реакцию перекрестной сшивки осуществляют известными в этой области способами, например, описываемыми в патентах США №№ 2328537 и 2801242. Способы модификации крахмалов описаны в главе "Starch and Its Modification" в M. W. Rutenberg, стр. от 22-26 до 22-47, Handbook of Water Soluble Gums and Resins, R. L. Davidson, Editor (McGraw-Hill, Inc., New York, N.Y. 1980).
Количество сшивающего средства, необходимое для получения подходящего продукта, будет варьироваться в зависимости, например, от типа используемого сшивающего средства, концентрации сшивающего средства, условий реакции и необходимости получения перекрестно сшитого крахмала, попадающего в желаемый диапазон вязкости. Практикующему специалисту в этой области будет понятно, какое количество можно использовать, т.к. оно хорошо известно в этой области. Как правило, это количество будет находиться в диапазоне от приблизительно 0,001% масс. крахмала до того количества, которое считается приемлемым для использования в пищевых продуктах.
Крахмалы по настоящему изобретению также можно модифицировать физически, например, посредством термического ингибирования, описываемого в WO 95/04082 (опубликованном 9 февраля 1995 года).
Обработка. Крахмалы также можно прежелатинизировать. Примеры способов получения пептизированных крахмалов описаны в патенте США № 4280851 (Pitchon, et al.), патенте США № 4465702 (Eastman, et al.), патенте США № 5037929 (Rajagopalan), патенте США № 5131953 (Kasica, et al.) и патенте США № 5149799 (Rubens). Общепринятые способы прежелатинизации крахмала хорошо известны специалистам в этой области и описаны в таких статьях, как Глава XXII-"Production and Use of Pregelatinized Starch", Starch: Chemistry and Technology, Vol. III--Industrial Aspects, R. L. Whistler and E. F. Paschall, Editors, Academic Press, New York 1967.
Крахмалы по настоящему изобретению можно очищать любым известным в этой области способом для удаления посторонних привкусов и цветов, присущих крахмалу или возникших при модификации крахмала. Способы очистки, предпочтительные для обработки крахмалов по настоящему изобретению, описаны в патенте США № 07/832838, зарегистрированном 7 февраля 1992 года на имя Kasica, et al. Способы промывки щелочью крахмалов, предназначенных для использования в гранулярной или прежелатинизированной форме, также применимы и описаны в семействе патентов, представленном патентом США № 5187272 (Bertalan, et al.).
Продукты переработки, полученные из крахмалов по настоящему изобретению, включая, в качестве неограничивающих примеров, крахмалы жидкой варки или крахмалы жидкой варки, полученные посредством окисления, ферментативной переработки, в частности, с помощью α-амилазы, кислотного гидролиза, высокой температуры и/или кислотной декстринизации, продукты термической обработки и/или фрагментации также применимы в настоящем описании.
Растения, продуцирующие крахмал
Настоящее изобретение относится к растениям, продуцирующим описываемый выше крахмал. Растения, продуцирующие крахмал, включают, в качестве неограничивающих примеров, кукурузу, картофель, пшеницу, тапиоку, рис, маниок и сорго.
Крахмал по настоящему изобретению можно экстрагировать из растений кукурузы, например, из эндосперма растений кукурузы. Растения кукурузы, продуцирующие крахмал по настоящему изобретению, являются растениями с двойной мутацией с генотипом waxy/sugary-2. Ген waxy локализуется на хромосоме 9 кукурузы, в то время как ген sugary-2 локализуется на хромосоме 6. (См. M. G. Nueffer, L. Jones, and M. Zuber, "The Mutants of Maize" (Crop Science Society of America, Madison, WI, 1968), pp. 72 and 73).
Растения по настоящему изобретению являются гомозиготами по рецессивному гену синтазы крахмала IIa (su2), и гомозиготами или гетерозиготами по мутантному гену гранулосвязанной синтазы крахмала I (GBSSI). Мутантный ген GBSSI имеет меньшую активность GBSSI, чем ген GBSSI (Wx) дикого типа, но более высокую активность GBSSI, чем рецессивный ген GBSSI (wx) с потерей функции. В рамках изобретения, такой мутантный ген waxy обозначают как промежуточный мутант waxy. В некоторых вариантах осуществления промежуточный мутант waxy имеет менее 30%, менее 25%, менее 20%, менее 15%, менее 10% или менее 5% экспрессии гена GBSSI дикого типа. В некоторых вариантах осуществления промежуточный мутант waxy имеет приблизительно 5%, приблизительно 6%, приблизительно 7%, приблизительно 8%, приблизительно 9%, приблизительно 10%, приблизительно 11%, приблизительно 12%, приблизительно 13%, приблизительно 14% или приблизительно 15% экспрессии гена GBSSI дикого типа.
Промежуточный ген GBSSI, имеющий меньшую активность GBSSI, чем ген GBSSI дикого типа, но более высокую активность GBSSI, чем рецессивный ген GBSSI с потерей функции, может быть уже известен в этой области, или его можно получать любым подходящим способом мутагенеза.
Мутантные аллели, такие как wxS (также известный как wx-Stonor), wx-G, wx-B5 и wx-M, имеют промежуточную активность GBSSI. Эти мутантные аллели являются результатом инсерции ретротранспозона в ген GBSSI, как описано в Wessler and Varagona (PNAS, 82:4177-4181, 1985) и Varagona et al. (The Plant Cell, 4:811-820, 1992). Эти промежуточные аллели приводят к образованию белка waxy, имеющего меньшую активность, чем белок waxy дикого типа.
Дополнительные мутантные аллели, имеющие промежуточную активность GBSSI, можно получать подходящими способами. Например, ретротранспозонный мутагенез можно использовать для получения популяции для скрининга растений кукурузы, имеющих мутантный ген GBSSI, имеющий промежуточную активность GBSSI. В некоторых вариантах осуществления wxS (также известный как wx-Stonor), wx-G, wx-B5 и wx-M можно использовать в качестве исходных материалов для получения дополнительных мутантных аллелей.
Другие способы мутагенеза, которые можно использовать для получения мутантных аллелей, имеющих промежуточную активность GBSSI, включают, в качестве неограничивающих примеров, химический мутагенез, радиационный мутагенез, транспозонный мутагенез, инсерционный мутагенез, мутагенез с использованием генных сигнатур в качестве меток, сайт-специфический мутагенез и природный мутагенез.
В некоторых вариантах осуществления для получения мутантных аллелей, имеющих промежуточную активность GBSSI, можно использовать антисмысловую РНК, рибозим, dsRNAi, РНК-интерференцию (РНКи). Технология антисмысловой РНК включает экспрессию или встраивание в клетку молекулы РНК (или производного РНК), комплементарной, или антисмысловой, последовательностям, обнаруживаемым в конкретной мРНК в клетке. Посредством связывания с мРНК, антисмысловая РНК может ингибировать трансляцию кодируемого продукта гена. Использование антисмысловой технологии для снижения или ингибирования экспрессии конкретных генов растений описано, например, в европейской патентной публикации № 271988, Smith et al., Nature, 334:724-726 (1988); Smith et. al., Plant Mol. Biol., 14:369-379 (1990)). Рибозим является РНК, имеющей каталитический домен и последовательность, комплементарную конкретной мРНК. Рибозим функционирует посредством связывания с мРНК (с помощью комплементарного домена рибозима), а затем расщепления (деградации) матричной РНК с использованием каталитического домена. РНК-интерференция (РНКи) является процессом специфичного в отношении последовательности, посттранскрипционного сайленсинга гена или транскрипционного сайленсинга гена у животных и растений, инициируемым двухцепочечной РНК (дцРНК), являющейся гомологичной в отношении последовательности гену, подвергаемому сайленсингу. Способ РНКи обсуждают, например, в Elibashir, et al., Methods Enzymol. 26:199 (2002); McManus & Sharp, Nature Rev. Genetics 3:737 (2002); PCT application WO 01/75164; Martinez et al., Cell 110:563 (2002); Elbashir et al., выше; Lagos-Quintana et al., Curr. Biol. 12:735 (2002); Tuschl et al., Nature Biotechnol. 20:446 (2002); Tuschl, Chembiochem. 2:239 (2001); Harborth et al., J. Cell Sci. 114:4557 (2001); et al., EMBO J. 20:6877 (2001); Lagos-Quintana et al., Science 294:8538 (2001); Hutvagner et al., loc cit, 834; Elbashir et al., Nature 411:494 (2001).
Кроме того, в объем по настоящему изобретению входит крахмал, полученный из мутантов, где мутантные аллели wx и/или su2 перемещают в другую часть генома растения посредством транслокации, инверсии или любыми другими способами хромосомной инженерии. Кроме того, крахмал, экстрагированный из растения, выращенного с искусственными мутациями и вариантами указанной выше генетической композиции, которую можно получать известными стандартными способами мутационной селекции, также применимы в настоящем описании.
Кроме того, в объем по настоящему изобретению входит крахмал, получаемый из растений, иных, чем растения кукурузы, при условии, что растения, продуцирующие крахмал, имеют мутантный ортолог гена GBSSI кукурузы, имеющий промежуточную активность GBSSI, и рецессивный мутантный ортолог гена кукурузы SSIIa. В некоторых вариантах осуществления растения могут являться пшеницей, тапиокой, рисом, маниоком, саго, картофелем или сорго.
В некоторых вариантах осуществления ортолог ген GBSSI в растениях, иных, чем кукуруза, кодирует белок, имеющий по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или более идентичности по отношению к SEQ ID NO: 1 и активность GBSSI. В некоторых вариантах осуществления ортолог имеет последовательность регистрационной записи в UniProt №№ O82627 (Antirrhinum majus (львиный зев)), P12615 (Avena sativa (овес)), Q42968 (Oryza glaberrima (африканский рис)), Q43784 (Manihot esculenta (маниок)), P0C585 (Oryza sativa (рис)), Q00775 (Solanum tuberosum (картофель)), P27736 (Triticum aestivum (пшеница)), Q43134 (Sorghum bicolor (сорго)), Q43092 (Pisum sativum (горох огородный)), P84766 (Aegilops tauschii subsp. strangulata (эгилопс)), Q0DEV5 (Oryza sativa subsp. japonica (рис)), Q42857 (Ipomoea batatas (батат), P84633 (Fagopyrum esculentum (гречиха посевная)), P84765 (Secale cereale (рожь)), Q9MAQ0 (Arabidopsis thaliana), P09842 (Hordeum vulgare (ячмень) или A2Y8X2 (Oryza sativa subsp. indica (рис)), каждая из которых включен в настоящее описание в качестве ссылки в полном объеме.
В некоторых вариантах осуществления ортолог гена SSIIa в растениях, иных, чем кукуруза, кодирует белок, имеющий по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или более идентичности по отношению к SEQ ID NO: 2 и активность SSIIa. В некоторых вариантах осуществления ортолог имеет последовательность регистрационной записи в UniProt №№ V6BPG5 (Triticum urartu (урартская пшеница)), V6BPN2 (Triticum monococcum subsp. Monococcum), C3W8L4 (Hordeum vulgare var. distichum (одомашненный ячмень)), E2GHR4 (Oryza sativa subsp. indica (рис)) или E2GHR6 (Oryza sativa subsp. japonica (рис)), каждая из которых включен в настоящее описание в качестве ссылки в полном объеме.
В некоторых вариантах осуществления для получения растений с двойной мутацией, имеющих промежуточный мутантный wx и рецессивный мутантный su2, можно осуществлять перекрестное опыление между растением, имеющим промежуточный мутантный wx, и растением, имеющим рецессивный мутантный su2.
Например, можно осуществлять перекрестное опыление между гомозиготным мутантом wxS (wxSwxSSU2SU2) или гетерозиготным мутантом wxS (wxSWxSU2SU2) и гомозиготным мутантом waxy su2 (wxwxsu2su2) или гетерозиготным мутантом waxy su2 (wxWxsu2su2 или wxwxSU2su2). После скрещивания для получения растений F2 можно использовать самоопыление поколения F1, имеющего генотип wxSwxSU2su2. Таким образом, можно выделять растения, имеющие нуль, одну, две или три дозы гена wxS в эндосперме и три дозы рецессивного гена su2 (например, эндосперм, имеющий генотип wxwxwxsu2su2su2, wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwSsu2su2su2). Это всего лишь один пример способа скрещивания, можно использовать другие схемы скрещивания, и они входят в объем настоящего изобретения.
В некоторых вариантах осуществления получение растений с эндоспермами, имеющими по меньшей мере одну дозу промежуточного аллеля waxy и рецессивного аллеля sugary-2, в полевых условиях осуществляют посредством скрещивания женских растений кукурузы с мужскими растениями кукурузы. Типичное расположение посадок представляет собой один ряд мужских растений на несколько рядов женских растений. Ряды женских растений обессултанивают или мужские растения делают стерильными различными другими способами, известными в этой области, такими как цитоплазматические или генетические средства.
Крахмал, используемый в настоящем изобретении, можно получать из инбредных линий или крахмал можно получать из гибридов, полученных из инбредных растений, содержащих двойную мутацию промежуточного waxy/рецессивного su2. Хотя в настоящем описании кукуруза является примером растения, служащего источником крахмала waxy, настоящее изобретение также применимо к другим видам растений, таким как, например, рис waxy, ячмень waxy и сорго waxy, при условии, что они имеют промежуточный ген waxy и рецессивный ген su2.
Способы скрещивания
Для получения растений по настоящему изобретению можно использовать подходящие способы скрещивания или селекции. Распространенные способы селекции, как правило, включают родословную селекцию, модифицированную родословную селекцию, массовую селекцию, рекуррентную селекцию и возвратное скрещивание.
Перекрестноопыляющиеся популяции. Улучшение перекрестноопыляющихся популяций таких сельскохозяйственных культур, как рожь, многие сорта кукурузы и сахарной свеклы, пастбищные травы, бобовые растения, такие как люцерна и клевер, и тропические древесные культуры, такие как какао, кокос, масличная пальма и некоторые каучуконосы, зависит, по существу, от изменения частот генов в сторону фиксации предпочтительных аллелей при одновременном поддержании высокой (но далекой от максимальной) степени гетерозиготности. Однородность в таких популяциях является невозможной, и сортовая чистота в перекрестноопыляющемся сорте является статистическим признаком популяции в целом, а не характеристикой отдельных растений. Таким образом, гетерогенность перекрестноопыляющихся популяции отличается от гомогенности (или фактически) инбредных линий, клонов и гибридов.
Способы улучшения популяции, естественно, разделены на две группы, те, которые основаны исключительно на фенотипической селекции, как правило, называемые массовой селекцией, и те, которые основаны на селекции с тестированием потомства. В межпопуляционном улучшении используют концепцию открытых скрещиваемых популяций; позволяющую генам попадать из одной популяции в другую. Растения в одной популяции (культивар, штамм, экотип или любой источник зародышевой плазмы) скрещиваются естественным образом (например, с помощью ветра) или руками человека или пчелами (как правила, Apis mellifera L. или Megachile rotundata F.) с растениями из других популяций. Селекцию используют для улучшения одной (или иногда обеих) популяции посредством выделения растений с желаемыми признаками из обоих источников.
В целом, существует два основных способа улучшения перекрестноопыляющихся популяций. Во-первых, есть ситуация, в которой популяцию изменяют целиком с помощью выбранного способа селекции. Результатом является улучшенная популяция, бесконечно размножающаяся посредством случайного скрещивания внутри самой себя в изоляции. Во-вторых, в случае синтетического сорта достигают того же конечного результата, что и при улучшении популяции, но он не размножается сам по себе; его необходимо восстанавливать из родительских линий или клонов. Эти способы скрещивания растений для улучшения перекрестноопыляющихся популяций хорошо известны специалистам в этой области и всеобъемлющие обзоры способов скрещивания, общепринято используемых для улучшения перекрестноопыляющихся растений, представлены в многочисленных текстах и статьях, включая: Allard, Principles of Plant Breeding, John Wiley & Sons, Inc. (1960); Simmonds, Principles of Crop Improvement, Longman Group Limited (1979); Hallauer and Miranda, Quantitative Genetics in Maize Breeding, Iowa State University Press (1981); и Jensen, Plant Breeding Methodology, John Wiley & Sons, Inc. (1988).
Массовая селекция. При массовой селекции желаемые отдельные растения выбирают, собирают и получают композицию семян без тестирования потомства для получения следующего поколения. Т.к. селекция основана исключительно на родителе по материнской линии, и нет контроля опыления, целью массовой селекции является повышение доли лучших генотипов в популяции.
Синтетическое получение. Синтетический сорт получают посредством скрещивания между собой ряда генотипов, выбранных по хорошей комбинационной способности среди всех возможных комбинаций гибридов, с последующим поддержанием сорта посредством перекрестного опыления. В принципе, нет различий в том, являются ли родители (более или менее инбредные) выращиваемыми из семян линиями, как у некоторых сортов сахарной свеклы и бобов (Vicia), или клонами, как у пастбищных трав, клевера и люцерны. Родителей выбирают по общей комбинационной способности, иногда посредством тестовых скрещиваний или топкроссов, более типично - посредством поликроссов. Родительские линии семян можно преднамеренно скрещивать (например, посредством самоопыления или скрещивания сибсов). Однако даже если родителей не скрещивают преднамеренно, селекция в пределах линий при поддержании линии будет обеспечивать то, что будет происходить некоторый инбридинг. Разумеется, клональные родители будут оставаться неизмененными и высокогетерозиготными.
То, может ли синтетический сорт сразу попадать из родительского семенного участка к фермеру, или сначала его необходимо подвергать одному или двум циклам размножения, зависит от семенной продуктивности и масштаба спроса на семена. На практике, травы и клевер, как правило, размножают однократно или двукратно и, таким образом, в значительной степени, удаляют из исходного синтетического сорта.
Хотя иногда используют массовую селекцию, тестирование потомства, как правило, является предпочтительным в случае поликроссов по причине простоты их эксплуатации и очевидной значимости для цели, а именно использования общей комбинационной способности в синтетическом сорте.
Широко варьируется количество родительских линий или клонов, входящих в синтетический сорт. На практике, количества родительских линий находятся в диапазоне от 10 до нескольких сотен при среднем значении 100-200. Ожидают, что универсальные синтетические сорта, полученные из 100 или более клонов, будут более стабильными при размножении семян, чем специализированные синтетические сорта.
Родословные сорта. Родословный сорт представляет собой лучший генотип, разработанный посредством селекции отдельных растений из расщепляющейся популяции после размножения и увеличения семян самоопыляющегося потомства и тщательного тестирования генотипа в течение нескольких поколений. Это способ перекрестного опыления, хорошо работающий в отношении естественным образом самоопыляющихся видов. Этот способ можно использовать в комбинации с массовой селекцией при разработке сорта. Варианты родословной и массовой селекции в комбинации являются наиболее распространенными способами получения сортом самоопыляющихся культур.
Гибриды. Гибрид является отдельным растением, полученным при скрещивании между родителями с различными генотипами. В настоящее время широко используют коммерческие гибриды многих сельскохозяйственных культур, включая кукурузу, сорго, сахарную свеклу, подсолнечник и брокколи. Гибриды можно получать рядом различных способов, включая скрещивание непосредственно двух родителей (простые гибриды), скрещивание простого гибрида с другим родителем (тройные гибриды) или скрещивание двух разных гибридов (гибриды, полученные посредством четырехлинейного или двойного скрещивания).
Родословная селекция. Родословное скрещивание, как правило, используют для улучшения самоопыляющихся сельскохозяйственных культур или инбредных линий перекрестноопыляющихся сельскохозяйственных культур. Двух родителей, обладающих благоприятными, дополняющими признаками, скрещивают для получения F1. Популяцию F2 получают посредством самоопыления одного или нескольких F1 или перекрестного скрещивания двух F1 (скрещивание сибсов). Также можно использовать способ дигаплоидного скрещивания или другие способы снижения плоидности. Селекцию лучших индивидуумов, как правило, начинают в популяции F2; затем, начиная с F3, выбирают лучших индивидуумов в лучших семействах. Повторные тестирования семейств или комбинации гибридов, включающих индивидуумов из этих семейств, часто проводят в поколении F4 для улучшения эффективности селекции на предмет признаков с низкой наследуемостью. На поздней стадии инбридинга (т.е. F6 и F7) лучшие линии или комбинации фенотипически схожих линий тестируют для потенциального выпуска новых культиваров. Аналогично, для разработки новых культиваров с помощью дигаплоидной системы необходима селекция культиваров с последующим тестированием в течение от двух до пяти лет на дублируемых опытных участках.
Возвратное скрещивание. Возвратное скрещивание используют для переноса генов для просто наследуемого, высоконаследуемого признака в желаемый гомозиготный культивар или инбредную линию, являющуюся рекуррентным родителем. Источник признака, подлежащего переносу, называют донорным родителем. Ожидают, что полученное растение будет иметь признаки рекуррентного родителя (например, культивара) и желаемый признак, перенесенный из донорного родителя. После исходного скрещивания индивидуумов, обладающих фенотипом донорного родителя, подвергают селекции и повторно скрещивают (подвергают возвратному скрещиванию) с рекуррентным родителем. Ожидают, что полученное растение будет иметь признаки рекуррентного родителя (например, культивара) и желаемый признак, перенесенный из донорного родителя.
Массовый сегрегационный анализ (BSA). BSA, также известный как массовый сегрегационный анализ, является способом, описываемым в Michelmore et al. (Michelmore et al., 1991, Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proceedings of the National Academy of Sciences, USA, 99:9828-9832) and Quarrie et al. (Quarrie et al., 1999, Journal of Experimental Botany, 50(337):1299-1306).
Для BSA интересующего признака родительские линии с различными конкретными фенотипами выбирают и скрещивают для получения F2, двойных гаплоидных или рекомбинантных инбредных популяций с использованием анализа QTL. Затем популяцию фенотипируют для идентификации отдельных растений или линий, имеющих высокую или низкую экспрессию признака. Получают две совокупности ДНК, одну из индивидуумов, имеющих один фенотип (например, резистентный к вирусу), и другую из индивидуумов, имеющих обратный фенотип (например, восприимчивый к вирусу), и анализируют частоту аллеля с использованием молекулярных маркеров. Необходимо лишь несколько индивидуумов в каждой совокупности (например, по 10 растений каждая), если маркеры являются доминантными (например, RAPD). Необходимо больше индивидуумов, если маркеры являются кодоминантными (например, RFLP). Маркеры, связанные с фенотипом, можно идентифицировать и использовать для скрещивания или QTL-картирования.
Поиск индуцированных локальных нарушений в геномах (TILLING®). Схемы скрещивания по настоящему изобретению могут включать скрещивания с культиварами растений, подвергнутыми TILLING®. TILLING® является способом молекулярной биологии, делающим возможной направленную идентификацию мутаций в конкретном гене. TILLING® был представлен в 2000 году с использованием модельного растения Arabidopsis thaliana. С тех пор TILLING® используют в качестве способа обратной генетики в других организмах, таких как данио-рерио, кукуруза, пшеница, рис, соя, томат и Brassica rapa var. nipposinica.
В способе комбинируют стандартный и эффективный способ мутагенеза с использованием химического мутагена (например, этилметансульфоната (EMS)) с чувствительным способом скрининга ДНК, с помощью которого идентифицируют однонуклеотидные мутации (также называемые точечными мутациями) в гене-мишени. EcoTILLING является способом, в котором используют способы TILLING® для поиска природных мутаций в индивидуумах, как правило, для популяционного генетического анализа (см. Comai, et al., 2003 The Plant Journal 37, 778-786; Gilchrist et al. 2006 Mol. Ecol. 15, 1367-1378; Mejlhede et al. 2006 Plant Breeding 125, 461-467; Nieto et al. 2007 BMC Plant Biology 7, 34-42, каждая из которых включена в настоящее описание в качестве ссылки для всех целей). DEcoTILLING является модификацией TILLING® и EcoTILLING, в котором используют недорогой способ идентификации фрагментов (Garvin et al., 2007, DEco-TILLING: An inexpensive method for SNP discovery that reduces ascertainment bias. Molecular Ecology Notes 7, 735-746).
Способ TILLING® основан на образовании гетеродуплексов, образующихся, когда множество аллелей (которые могут быть из гетерозиготы или совокупности множества гомозигот и гетерозигот) амплифицируют посредством ПЦР, нагревают, а затем медленно охлаждают. В области несовпадения двух цепей ДНК (индуцированной мутации при TILLING® или природной мутации или SNP при EcoTILLING) образуется "пузырь", который затем расщепляется нуклеазами, специфичными к одноцепочечной ДНК. Затем продукты разделяют по размеру с использованием нескольких разных платформ.
Более подробное описание способов и композиций для TILLING® можно найти в патенте США № 5994075, патентной заявке США № 2004/0053236 A1, WO 2005/055704 и WO 2005/048692, каждая из которых, таким образом, включена в настоящее описание в качестве ссылки для всех целей.
Таким образом, в некоторых вариантах осуществления способы скрещивания по настоящему изобретению включают скрещивание с одним или несколькими линиями растений TILLING® с одной или несколькими идентифицированными мутациями.
Мутационная селекция. Мутационная селекция является другим способом внесения новых признаков в растения. Мутации, которые возникают спонтанно, или которые индуцируют искусственно, могут являться применимыми источниками изменчивости для селекционера. Целью искусственного мутагенеза является повышение частоты мутации для желаемой характеристики. Частоты мутаций можно повышать множеством различных способов или с помощью средств мутагенеза, включая температуру, длительное хранение семян, условия культивирования ткани, радиацию (такую как рентгеновское излучение, гамма-излучение, нейтронное излучение, бета-излучение или ультрафиолетовое излучение), химические мутагены (таких как аналоги оснований, подобные 5-бром-урацилу), антибиотики, алкилирующие средства (такие как сернистые иприты, азотистые иприты, эпоксиды, этиленамины, сульфаты, сульфонаты, сульфоны или лактоны), азид, гидроксиламин, азотную кислоту или акридины. После получения желаемого признака посредством мутагенеза признак можно вносить в существующую зародышевую плазму общепринятыми способами скрещивания. Подробное описание мутационной селекции можно найти в W. R. Fehr, 1993, Principles of Cultivar Development, Macmillan Publishing Co. Для достижения генетической изменчивости и внесения новых признаков в сорта также необходимо использовать новые способы скрещивания, такие как способы, включающие использование нуклеазы с цинковыми пальцами или олигонуклеотид-направленный мутагенез.
В некоторых вариантах осуществления мутационная селекция включает однолокусную конверсию. Один локус может содержать несколько генов и/или трансгенов, таких как трансген для резистентности к заболеванию, который в том же экспрессирующем векторе также содержит трансген для резистентности к гербициду. Ген для резистентности к гербициду можно использовать в качестве селективного маркера и/или фенотипического признака. Однолокусная конверсия сайт-специфической системы встраивания делает возможным встраивание множества генов в конвертируемый локус. Однолокусная конверсия также делает возможным получение одного или нескольких сайт-специфических изменений в геноме растения. В некоторых вариантах осуществления однолокусную конверсию осуществляют посредством редактирования генома, также известного как редактирование генома с использованием сконструированных нуклеазы (GEEN). В некоторых вариантах осуществления редактирование генома включает использование одной или нескольких сконструированных нуклеаз. В некоторых вариантах осуществления сконструированные нуклеазы включают, в качестве неограничивающих примеров, нуклеазы с цинковыми пальцами (ZFN), подобные активаторам транскрипции эффекторные нуклеазы (TALEN), систему CRISPR/Cas и сконструированные мегануклеазы, реконструированные хоминг-эндонуклеазы. В некоторых вариантах осуществления с помощью однолокусной конверсии изменяют одну или несколько нуклеиновых кислот в геноме растения.
Двойные гаплоиды и дупликация хромосом. Одним из способов получения гомозиготных растений без потребности в скрещивании двух родительских линий с последующей длительной селекцией сегрегированного потомства и/или множествами возвратных скрещиваний является получение гаплоидов, а затем дупликация хромосом для получения удвоенных гаплоидов. Гаплоидные растения могут возникать спонтанно, или их можно индуцировать искусственно посредством химических обработок или посредством скрещивания растений с индукторными линиями (Seymour et al. 2012, PNAS vol 109, pg 4227-4232; Zhang et al., 2008 Plant Cell Rep. Dec 27(12) 1851-60). Получение гаплоидного потомства может происходить посредством множества механизмов, которые могут влиять на распределение хромосом при образовании гамет. Хромосомный набор гаплоидов иногда удваивается спонтанно с образованием гомозиготных удвоенных гаплоидов (DH). Иногда могут возникать миксоплоиды, являющиеся растениями, содержащими клетки, имеющие различные плоидности, и они могут представлять собой растения, подвергающиеся дупликации хромосом таким образом, что спонтанно образуются удвоенные гаплоидны ткани, органы, побеги, части цветов или растения. Другим распространенным способом является индуцирование образования двойных гаплоидных растений с использованием обработки для дупликации хромосом, такой как обработка колхицина (El-Hennawy et al., 2011 Vol 56, issue 2 pg 63-72; Doubled Haploid Production in Crop Plants 2003, опубликованная Maluszynski, ISBN 1-4020-1544-5). Получение двойных гаплоидных растений приводит к получению высокооднородных культиваров и является особенно желательным в качестве альтернативы полового инбридинга сельскохозяйственных культур с более длительными поколениями. При получении двойного гаплоидного потомства количество возможных комбинаций генов для наследуемых признаков является более управляемым. Таким образом, с помощью эффективной технологии двойных гаплоидов можно значительно снижать время и стоимость инбредной разработки и разработки культивара.
Слияние протопластов. В другом способе скрещивания растений также можно использовать слияние протопластов для переноса придающего признак геномного материала из донорного растения в растение-реципиент. Слияние протопластов является индуцируемым или спонтанным объединением, таким как соматическая гибридизация, между двумя или более протопластами (клетками, стенки которых удаляют с помощью ферментативной обработки) для получения отдельной двух- или многоядерной клетки. Слитая клетка, которую можно получать с использованием вида растения, который не может скрещиваться в природе, представляет собой ткань, культивируемую в гибридном растении, проявляющем желаемую комбинацию признаков.
Культура ткани. В рамках изобретения, термин "культура ткани" означает композицию, содержащую выделенные клетки одного или разных типов или скопление таких клеток, организованных в части растения. Примерами типов культур тканей являются протопласты, каллюс, скопления растительной ткани и растительные клетки, которые могут образовывать культуру ткани, являющуюся интактной в растениях или частях растений, таких как зародыши, пыльца, цветы, семена, листья, стебли, корни, кончики корней, пыльники, пестики, меристемные клетки, пазушные почки, завязи, семенная оболочка, эндосперм, гипокотили, семядоли и т.п. Способы получения и поддержания культуры растительной ткани хорошо известны в этой области. В качестве примера, культуру ткани, содержащую органы, используют для получения регенерированных растений. В патентах США №№ 5959185, 5973234 и 5977445 описаны конкретные способы, описания которых включены в настоящее описание в качестве ссылок.
Настоящее изобретение дополнительно проиллюстрировано с помощью следующих примеров и вариантов осуществления, которые не следует истолковывать в качестве ограничений. Содержание всех ссылок, патентов и опубликованных патентных заявок, процитированных на всем протяжении настоящей заявки, а также фигуры и список последовательностей включены в настоящее описание в качестве ссылки.
ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ
1. Крахмал, имеющий:
содержание амилозы от приблизительно 2% масс. до приблизительно 20% масс., как измеряют с использованием набора для анализа амилозы/амилопектина; и
температуру клейстеризации водного раствора крахмала, составляющую по меньшей мере на приблизительно 5% больше температуры клейстеризации контрольного крахмала, как измеряют с использованием анализа RVA,
где крахмал получают из растения кукурузы waxy-sugary 2, содержащего по меньшей мере один аллель wxS в эндосперме.
2. Крахмал по варианту осуществления 1, где содержание амилозы составляет от приблизительно 8% масс. до приблизительно 15% масс.
3. Крахмал по любому из вариантов осуществления 1 или 2, где температура клейстеризации водного раствора составляет на от приблизительно 5% до приблизительно 14% больше, чем у контрольного крахмала.
4. Крахмал по любому из вариантов осуществления 1-3, дополнительно имеющий изменение энтальпии кристаллизации по меньшей мере на приблизительно 30% больше, чем изменение энтальпии кристаллизации контрольного крахмала, как измеряют с использованием анализа RVA.
5. Крахмал по варианту осуществления 4, где изменение энтальпии кристаллизации составляет на от приблизительно 70% до приблизительно 200% больше, чем изменение энтальпии кристаллизации контрольного крахмала, как измеряют с использованием дифференциального сканирующего калориметра (DSC).
6. Крахмал по любому из вариантов осуществления 1-4, дополнительно имеющий распределение размера гранул крахмала, где распределение размера гранул крахмала включает по меньшей мере на приблизительно 40% меньше гранул размером менее 5 мкм по сравнению с долей гранул размером менее 5 мкм в контрольном крахмале, что наблюдают с использованием сканирующего электронного микроскопа.
7. Крахмал по любому из вариантов осуществления 1-6, где крахмал представляет собой гранулу, демонстрирующую повышенную целостность по сравнению с контрольным крахмалом, что наблюдают с использованием сканирующего электронного микроскопа.
8. Крахмал по любому из вариантов осуществления 1-7, где крахмал способен выдерживать по меньшей мере три цикла замораживания/размораживания.
9. Крахмал по варианту осуществления 8, где крахмал способен выдерживать от по меньшей мере трех до по меньшей мере пяти циклов замораживания/размораживания.
10. Крахмал по любому из вариантов осуществления 1-9, где растение кукурузы waxy-sugary 2 получают посредством перекрестного опыления первого растения кукурузы, имеющего генотип wxwxsu2su2, и второго растения кукурузы, имеющего генотип wxSwxSsu2su2 или генотип wxSwxsu2su2.
11. Крахмал по любому из вариантов осуществления 1-10, где эндосперм растения кукурузы waxy-sugary 2 имеет одну дозу, две дозы или три дозы аллеля wxS, и эндосперм имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2.
12. Способ улучшения по меньшей мере одной характеристики в профиле клейстеризации крахмала, включающий встраивание по меньшей мере одного аллеля wxS в эндосперм растения кукурузы waxy, где полученный крахмал, полученный из растения кукурузы waxy, содержащего по меньшей мере один аллель wxS в эндосперме, имеет температуру клейстеризации водного раствора крахмала, составляющую по меньшей мере на приблизительно 5% больше, чем температура клейстеризации контрольного крахмала, как измеряют с использованием анализа RVA, таким образом, улучшение по меньшей мере одной характеристики профиля клейстеризации крахмала по сравнению с контрольным крахмалом.
13. Растение кукурузы с двойной мутацией, где растение кукурузы является гомозиготой, рецессивной по гену синтазы крахмала IIa (su2), и гомозиготой или гетерозиготой по мутантному гену гранулосвязанной синтазы крахмала I (GBSSI), где мутантный ген GBSSI имеет меньшую активность GBSSI, чем ген GBSSI (Wx) дикого типа, но более высокую активность GBSSI, чем рецессивный ген GBSSI (wx) с потерей функции.
14. Растение кукурузы с двойной мутацией по варианту осуществления 13, где мутантный ген GBSSI представляет собой аллель wx-Stonor (wxS).
15. Растение кукурузы с двойной мутацией по варианту осуществления 14, где растение получают посредством перекрестного опыления первого растения кукурузы, имеющего генотип wxwxsu2su2, и второго растения кукурузы, имеющего генотип wxSwxSsu2su2 или генотип wxSwxsu2su2.
16. Растение кукурузы с двойной мутацией по варианту осуществления 15, где эндосперм растения кукурузы имеет одну дозу, две дозы или три дозы аллеля wxS, где эндосперм имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2.
17. Растение кукурузы с двойной мутацией по любому из вариантов осуществления 13-16, где крахмал, продуцируемый в эндосперме растения кукурузы, имеет более высокую температуру клейстеризации по сравнению с крахмалом, продуцируемым в эндосперме растения кукурузы с двойной рецессивной мутацией waxy sugary-2, имеющего генотип wxwxwxsu2su2su2.
18. Семя растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-17.
19. Часть растения, растительная клетка или культура ткани растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-18.
20. Часть растения по варианту осуществления 19, где часть растения выбрана из группы, состоящей из эндосперма, листа, цветка, семязачатка, пыльцы, корневища и черенка.
21. Способ получения крахмала, включающий получение крахмала из эндосперма растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-20.
22. Способ получения крахмал, включающий выращивание растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-21 и сбор семян из растения кукурузы с двойной мутацией.
23. Способ получения семян кукурузы, включающий скрещивание первого родительского растения кукурузы со вторым родительским растением кукурузы и сбор полученных семян кукурузы, где указанное первое родительское растение кукурузы и/или второе родительское растение кукурузы является растением кукурузы с двойной мутацией по любому из вариантов осуществления 13-22.
24. Способ получения семян кукурузы, включающий самоопыление растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-23 и сбор полученных семян кукурузы.
25. Способ вегетативного размножения растения кукурузы по любому из вариантов осуществления 13-24, включающий сбор растения по любому из вариантов осуществления 1-11 и регенерацию растения из указанной части.
26. Способ получения растения кукурузы, полученного из растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-17, включающий самоопыление растения с двойной мутацией по любому из вариантов осуществления 13-17, по меньшей мере, однократно для получения растения-потомка, полученного из растения с двойной мутацией по любому из вариантов осуществления 13-17.
27. Способ получения растения кукурузы, полученного из растения с двойной мутацией по любому из вариантов осуществления 1-11, включающий скрещивание растения с двойной мутацией по любому из вариантов осуществления 13-17 со вторым растением кукурузы для получения растения-потомка растения с двойной мутацией по любому из вариантов осуществления 13-17.
28. Способ получения растения кукурузы, включающий встраивание трансгена в растение кукурузы, где трансген придает желаемый фенотип растения с двойной мутацией по любому из вариантов осуществления 13-17.
29. Способ получения растения кукурузы, включающий редактирование гена для получения генетической последовательности для придания желаемого фенотипа растения с двойной мутацией по любому из вариантов осуществления 13-17.
30. Способ получения растения кукурузы, включающий встраивание мутации для получения генетической последовательности для придания желаемого фенотипа растения с двойной мутацией по любому из вариантов осуществления 13-17.
31. Крахмал, экстрагированный из растения кукурузы с двойной мутацией по любому из вариантов осуществления 13-17.
32. Крахмал, экстрагированный из растения кукурузы с двойной мутацией по варианту осуществления 15.
33. Крахмал, экстрагированный из эндосперма растения кукурузы с двойной мутацией по варианту осуществления 16.
ПРИМЕР 1
Получение зерна с 1, 2 или 3 дозами аллеля wxS
Из-за триплоидной природы эндосперма кукурузы в случае, когда два полярных ядра из родителя по материнской линии оплодотворяют одной мужской половой клеткой родителя по отцовской линии, возможны три генотипические комбинации. Использование растения wxSwxSsu2su2 в качестве женского растения, опыляемого мужским растением wxwxsu2su2, приводит к "дозе-2" или зерну wxSwxSwxsu2su2su2. Использование wxSwxSsu2su2 в качестве мужского растения, опыляющего женское растение wxwxsu2su2, приводит к "дозе-1" или зерну wxSwxwxsu2su2su2. Самоопыление wxSwxSsu2su2 приводит к "дозе-3" или зерну wxSwxSwxSsu2su2su2.
ПРИМЕР 2
Определение вязкости крахмала с использованием Rapid Visco Amylograph (RVA)
Вязкость кукурузного крахмала, полученного из растений, содержащих 0, 1, 2 и 3 дозы аллеля wxS в рецессивных мутантных растениях sugary-2, определяли с использованием Rapid Visco Amylograph (Newport Scientific, Eden Prairie, Minn.). Крахмал AMIOCA™ и крахмал wxwxwxSU2su2su2 включали в качестве контролей. Генотип эндоспермов, из которых выделяли крахмал, приведен в таблице ниже. Содержание амилозы измеряли с использованием набора для определения амилозы Megazyme®. Содержание амилозы в крахмале повышается с повышением активности GBSSI. Содержание амилозы в крахмале wxwxwxsu2su2su2 (7,4%), крахмале AMIOCA™ (7,1%) и крахмале wxwxwxSU2su2su2 (6,1%) являлось более низким, чем в крахмале Waxy Stonor (12,8%) и wxwxwxsu2su2su2 с 1-3 дозами гена wxS в эндосперме (8,2-14,6%). Содержание амилозы в крахмале wxwxwxsu2su2su2 (7,4%, 8,2%, 13,8%, и 14,6%) повышалось с повышением дозы wxS (0, 1, 2, и 3 доз wxS, соответственно). Результаты позволяют предполагать, что встраивание гена wxS в wxwxwxsu2su2su2 немного повышало содержание амилозы в крахмале.
Свойство клейстеризации кукурузного крахмала waxy анализировали с помощью Rapid Visco-Analyzer (RVA, Newport Scientific, Sydney, Australia) с использованием способа профилей "Standard 2". Суспензию (28,0 г), содержащую 8% крахмала (масс./масс., в пересчете на сухой крахмал, dsb) уравновешивали при 50°C в течение 1 мин, нагревали при 95°C со скоростью 6°C/мин, держали при 95°C в течение 5 мин, а затем охлаждали до 50°C со скоростью 6°C/мин. Скорость вращения лопатки составляла 160 об./мин., за исключением того, что в первые 10 сек использовали960 об./мин. Результаты представлены на фиг. 1 и фиг. 2. Существующий крахмал wxwxwxsu2su2su2 имел наименьшую температуру клейстеризации, пиковую вязкость и конечную вязкость. После встраивания гена wxS в растение wxwxwxsu2su2su2 образцы крахмала (Q3017a [1 wxS], Q8412a [2 wxS] и Q7738a [3 wxS]) демонстрировали повышенную пиковую вязкость по сравнению с существующим крахмалом wxwxwxsu2su2su2 (0 wxS). Однако лишь крахмал wxSwxwxsu2su2su2 с одной дозой гена wxS демонстрировал пиковую вязкость, гораздо более высокую, чем Waxy Stonor, крахмал AMIOCA™ и крахмал wxwxwxSU2su2su2.
Результаты свидетельствуют о том, что улучшение температуры клейстеризации сделает возможным экстрагирование крахмала способом мокрого размола.
ПРИМЕР 3
Стабильность крахмала при замораживании/размораживании
Кукурузный крахмал, полученный из растения, содержащего 0, 1, 2 и 3 доз аллеля wxS в рецессивных мутантных растениях sugary-2, тестировали на стабильность при замораживании/размораживании. Крахмал AMIOCA™ и крахмал wxwxwxSU2su2su2 включали в качестве контролей. Образцы крахмала с содержанием твердых веществ 8% варили на водяной бане при 95°C в течение 20 минут. Суспензию крахмала перемешивали стеклянной палочкой в течение трех минут с последующим непрерывным нагреванием в течение 17 минут при варке. Стабильность геля крахмала (8%, масс./масс.) при замораживании/размораживании при нейтральном pH анализировали с использованием циклического морозильника Caron в течение пяти циклов замораживания/размораживания. В каждом цикле образец замораживали в течение 6 часов и размораживали в течение 6 часов при 2 циклах/день. Образцы визуально осматривали на предмет синерезиса на поверхности и при нажатии, а также на предмет мутности и изменений желатинизации. Этот цикл замораживания/размораживания повторяли. Результаты представлены на фиг. 3. После первого цикла замораживания/размораживания только гель крахмала Waxy Stonor демонстрировал высокую мутность, что свидетельствует о высокой степени ретроградации крахмала. Мутность геля крахмала повышалась с повышением циклов замораживания/размораживания. На третьем цикле замораживания/размораживания гели крахмала AMIOCA™, крахмала wxwxwxSU2su2su2 и Waxy Stonor становились очень мутными, в то время как гели крахмала Q3017a (1 wxS), Q8412a (2 wxS), Q7738a (3 wxS) и существующего крахмала wxwxwxsu2su2su2 все равно не являлись мутными. После пяти циклов замораживания/размораживания Q3017a (1 wxS), Q8412a (2 wxS), Q7738a (3 wxS) и существующий крахмал wxwxwxsu2su2su2 становились немного мутными.
Результаты позволяют предполагать, что небольшое повышение амилозы в образцах крахмала имеет небольшой эффект в отношении стабильности при замораживании/размораживании. Крахмалы Q3017a (1 wxS), Q8412a (2 wxS), Q7738a (3 wxS) имели лучшую стабильность при замораживании/размораживании, чем крахмал wxwxwxSU2su2su2 и крахмал AMIOCA™.
ПРИМЕР 4
Дифференциальная сканирующая калориметрия (DSC)
Термические свойства кукурузного крахмала, полученного из растения, содержащего 0, 1, 2 и 3 доз аллеля wxS в рецессивных мутантных по sugary-2 растениях, определяли с использованием дифференциальной сканирующей калориметрии (DSC). Крахмал AMIOCA™ и крахмал wxwxwxSU2su2su2 включали в качестве контролей. С помощью дифференциальной сканирующей калориметрии (DSC) измеряют тепло, поглощаемое или отдаваемое образцом в контролируемой атмосфере при определенных температурах. С помощью DSC получают информацию об удельной теплоте и латентной теплоте образцов с повышением температуры, что свидетельствует об изменениях в аморфных и кристаллических структурах. Данные регистрируют в терминах теплоотдачи и представляют в Джоулях/грамм (Дж/г) (Cassel, 2002). При анализе крахмала собирали информацию о параметрах желатинизации крахмала, таких как начало пика, пиковая температура, конец пика и изменение энтальпии желатинизации. Результаты представлены на фиг. 4. Крахмал wxSwxwxsu2su2su2 (Q3017a), крахмал wxSwxSwxsu2su2su2 (Q8412a), крахмал wxSwxSwxSsu2su2su2 (Q7738a) и крахмал wxwxwxsu2su2su2 (0 wxS) демонстрировали более низкие температуры желатинизации, чем крахмал AMIOCA™, Waxy Stonor и крахмал wxwxwxSU2su2su2, что соответствует о лучшей стабильности при замораживании/размораживании крахмала, выделенного из эндосперма с кукурузой с одной, двумя и тремя дозами wxS. Крахмал wxSwxwxsu2su2su2 с одной дозой wxS (Q3017a) имеет немного более высокую температуру начала желатинизации, чем другие гибриды (Q8412a [2 wxS], Q7738a [3 wxS] и крахмал wxwxwxsu2su2su2), что свидетельствует о том, что Q3017a имел лучшие характеристики при мокром размоле, чем другие гибриды.
ПРИМЕР 5
Гранула крахмала
Средний размер частиц кукурузного крахмала waxy, полученного из растений, содержащих 1, 2, и 3 доз аллеля wxS в рецессивных мутантных по sugary-2 растениях, измеряли по рассеянию лазерного излучения. Крахмал AMIOCA™ и крахмал wxwxwxSU2su2su2 включали в качестве контролей. Результаты представлены на фиг. 5. Кроме того, на фиг. 6 представлены микрофотографии, полученные с помощью сканирующего электронного микроскопа, для демонстрации морфологии гранул. Средний размер гранул крахмала Q3017a (11,4 мкм), Q8412a (11,4 мкм) и Q7738a (10,9 мкм) был больше, чем у существующего крахмала wxwxwxsu2su2su2 (10,4 мкм), что позволяет предполагать, что встраивание гена wxS в Waxy VO повышало размер гранулы крахмала. Результат соответствовал изображениям, полученным с помощью SEM (фигура 5), что свидетельствует о том, что встраивание гена wxS в крахмал wxwxwxsu2su2su2 повышало целостность гранулы крахмала.
Если не указано иначе, все технические и научные термины в настоящем описании обладают тем же значением, которое, как правило, понятно специалисту в области, к которой принадлежит настоящее изобретение. Хотя любые способы и материалы, схожие или эквивалентные представленным в настоящем описании, можно использовать в практическом осуществлении или тестировании настоящего изобретении, предпочтительные способы и материалы представлены в настоящем описании. Все процитированные публикации, патенты и патентные публикации включены в настоящее описание в качестве ссылок в полном объеме для всех целей.
Публикации, обсуждаемые в настоящем описании, относятся исключительно к их описанию до даты подачи настоящей заявки. Ничто в настоящем описании не следует истолковывать как признание того, что настоящее изобретение не дает право датировать такую публикацию задним числом в силу предшествующего изобретения.
Хотя настоящее изобретение описано применительно к конкретным вариантам его осуществления, следует понимать, что можно его дополнительно модифицировать, и настоящая заявка предназначена для охвата любых вариантов, применения или адаптаций изобретения, главным образом, в соответствии с принципами изобретения и включая такие отклонения от настоящего описания, которые подпадают под известную или принятую практику в области, к которой относится изобретение, и которые можно применить к изложенным выше существенным признаками, и которые соответствуют объему прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| Высокоамилозная пшеница | 2011 |
|
RU2619636C2 |
| ЯЧМЕНЬ С ИЗМЕНЕННОЙ АКТИВНОСТЬЮ ВЕТВЯЩЕГО ФЕРМЕНТА И КРАХМАЛ И КРАХМАЛСОДЕРЖАЩИЕ ПРОДУКТЫ С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ АМИЛОЗЫ | 2003 |
|
RU2303870C2 |
| ЯЧМЕНЬ С ПОНИЖЕННОЙ АКТИВНОСТЬЮ СИНТАЗЫ КРАХМАЛА II (SSII) И КРАХМАЛСОДЕРЖАЩИЕ ПРОДУКТЫ С ПОНИЖЕННЫМ СОДЕРЖАНИЕМ АМИЛОПЕКТИНА | 2001 |
|
RU2268584C2 |
| ТИПЫ ВОСКОВИДНОГО ПШЕНИЧНОГО КРАХМАЛА, СОДЕРЖАЩИЕ В ГРАНУЛЕ ВОСКОВИДНЫЕ БЕЛКИ | 1997 |
|
RU2245617C9 |
| ВЫСОКОАМИЛОЗНАЯ ПШЕНИЦА-III | 2017 |
|
RU2811010C2 |
| ПШЕНИЦА С МОДИФИЦИРОВАННОЙ АКТИВНОСТЬЮ ВЕТВЯЩЕГО ФЕРМЕНТА, А ТАКЖЕ ПОЛУЧЕННЫЕ ИЗ НЕЕ КРАХМАЛ И КРАХМАЛСОДЕРЖАЩИЕ ПРОДУКТЫ | 2004 |
|
RU2377303C2 |
| ТЕРМИЧЕСКИ ОБРАТИМЫЙ ЖЕЛИРУЕМЫЙ КРАХМАЛ | 2017 |
|
RU2758089C2 |
| КУКУРУЗНЫЕ ПРОДУКТЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2012 |
|
RU2650764C2 |
| СПОСОБ ПОВЫШЕНИЯ КАЧЕСТВА ЗЕРНА КУКУРУЗЫ | 1991 |
|
RU2083088C1 |
| ГЕН-СПЕЦИФИЧЕСКИЙ АНАЛИЗ НА FLUORY2 В МАИСЕ ДЛЯ ИНТРОГРЕССИИ МУЧНИСТОГО ПРИЗНАКА (FL2) | 2013 |
|
RU2661110C2 |
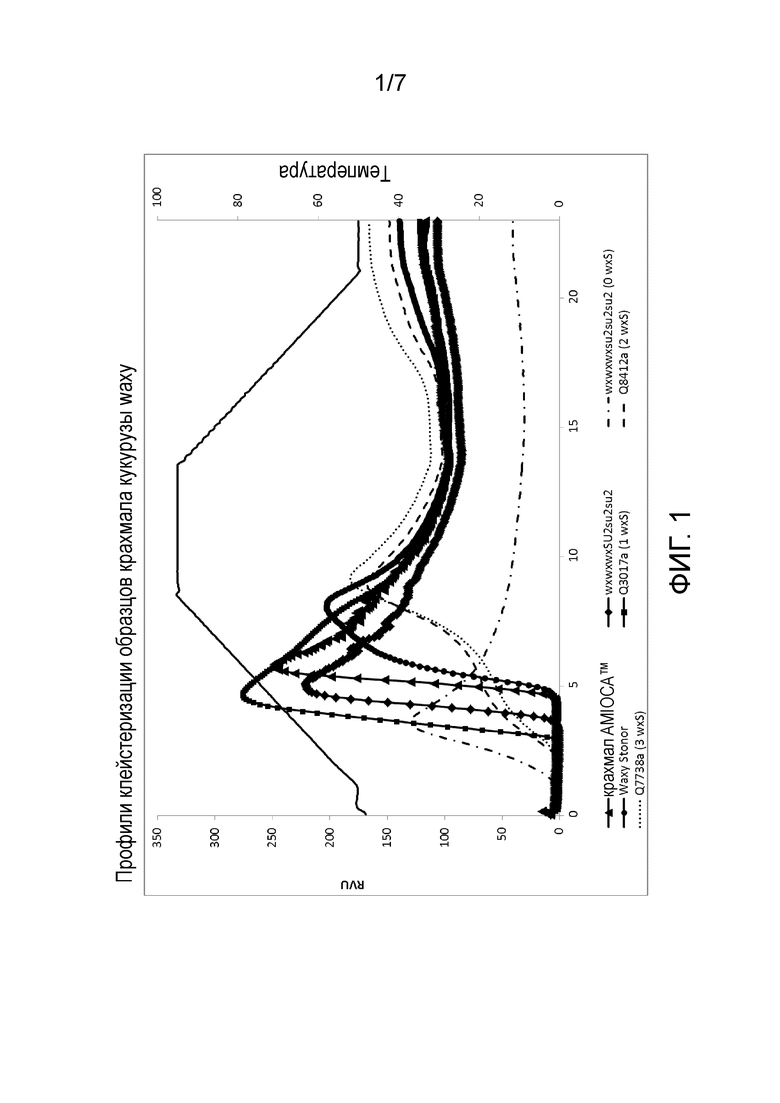
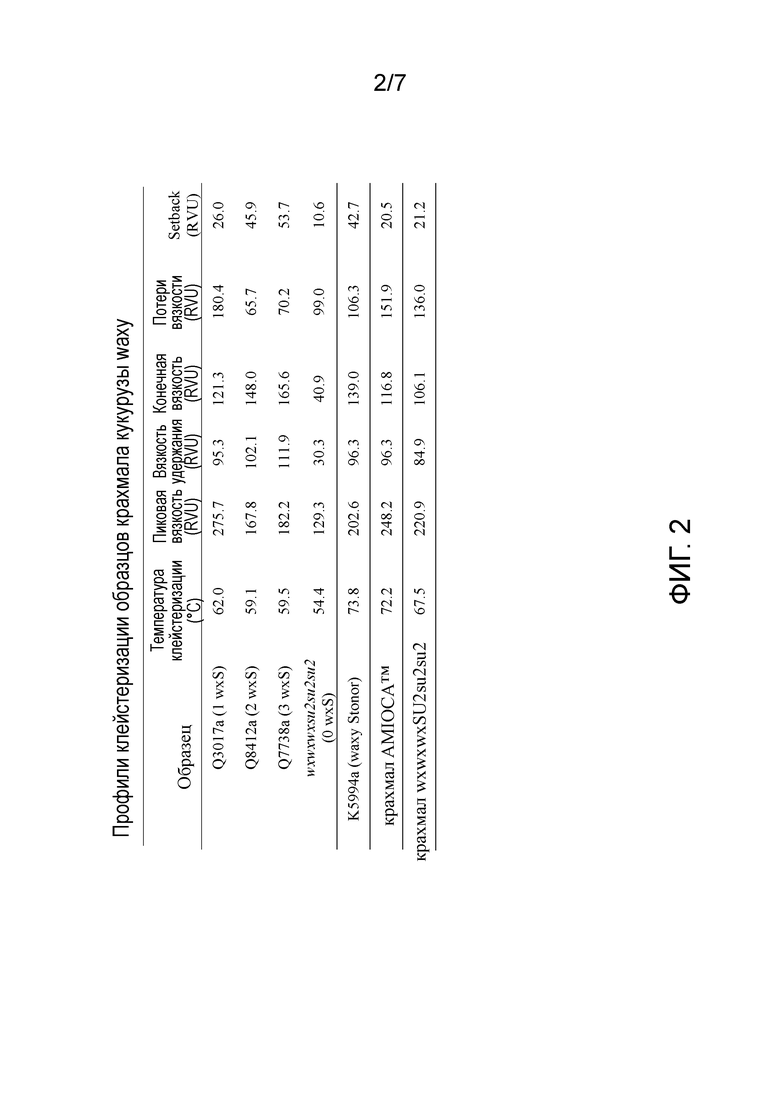

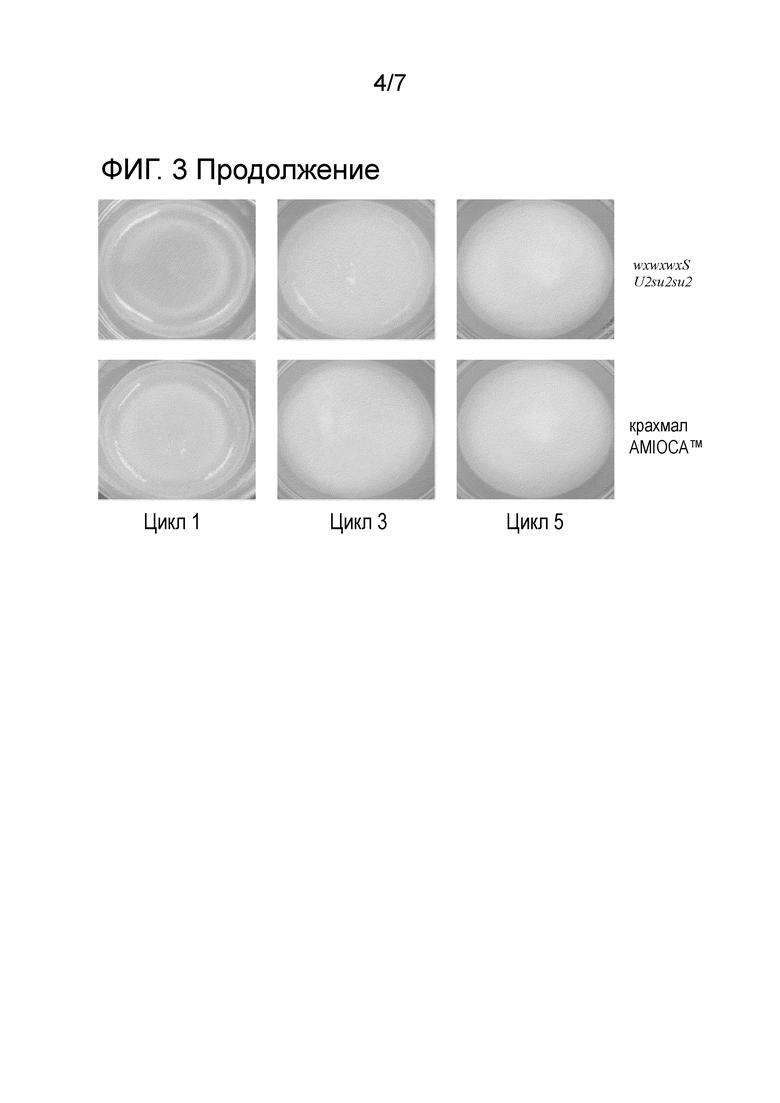
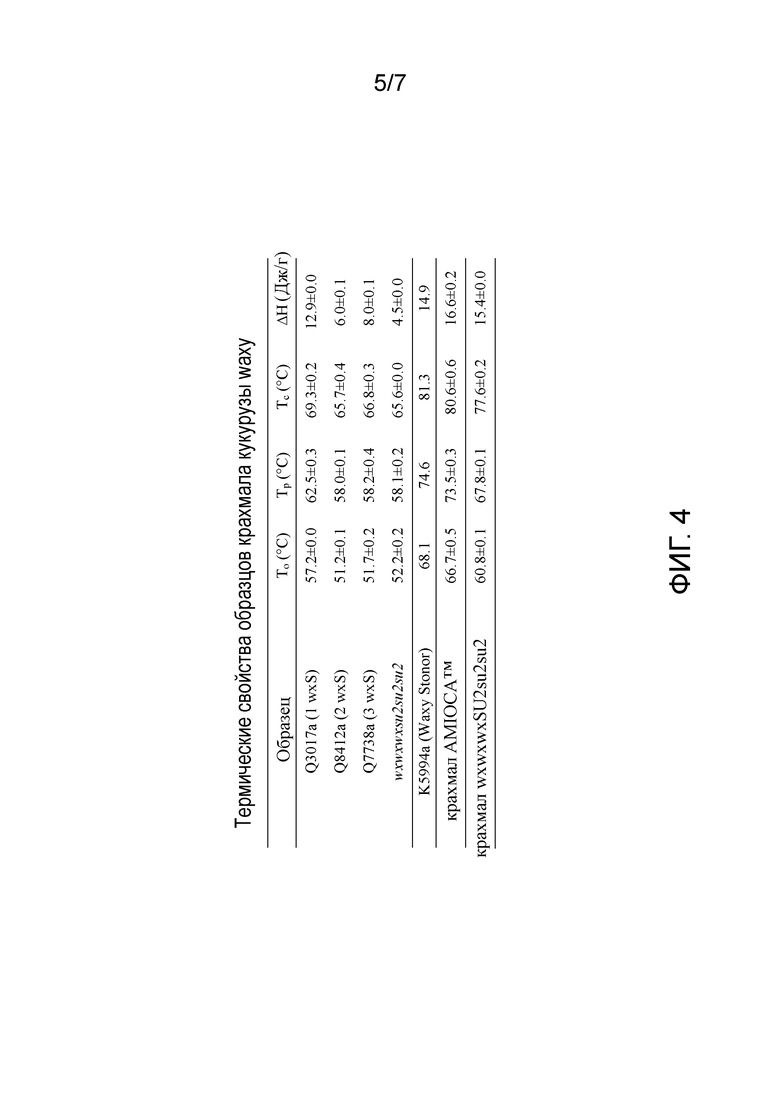
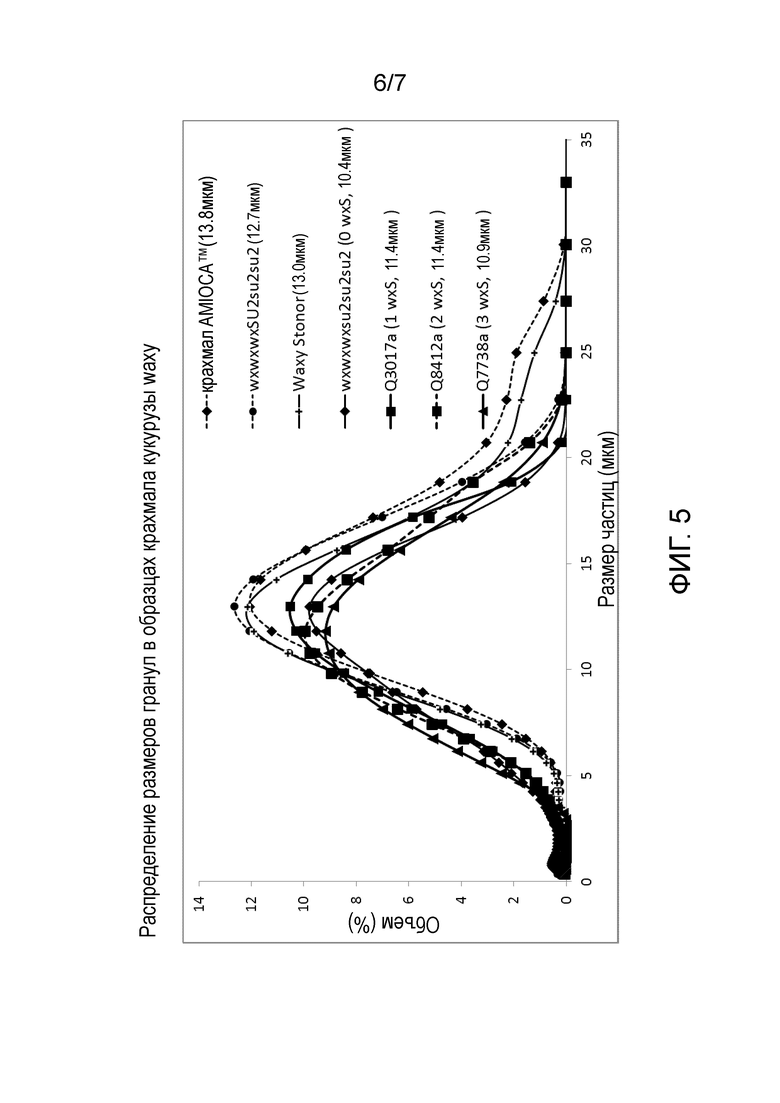

Изобретение относится к области биотехнологии. А именно к новому кукурузному крахмалу. Крахмал можно получать из недавно разработанной кукурузы с двойной мутацией waxy-sugary-2, имеющей низкую активность гранулосвязанной синтазы крахмала I (GBSSI), что приводит к низкому уровню амилозы. Крахмал из недавно разработанной кукурузы с двойной мутацией waxy-sugary-2 является стабильным при замораживании/размораживании и имеет высокую вязкость. По сравнению с крахмалом из существующей кукурузы с двойной мутацией waxy-sugary-2 крахмал из новой кукурузы с двойной мутацией waxy-sugary-2 демонстрировал, помимо прочего, улучшенный профиль клейстеризации, целостность гранул крахмала, более крупный размер гранул крахмала и более высокую вязкость. 8 з.п. ф-лы, 6 ил., 5 пр.
1. Крахмал, включающий:
содержание амилозы от 6,1% масс. до 14% масс. согласно измерению с использованием набора для анализа амилозы/амилопектина и
температуру клейстеризации водного раствора крахмала, составляющую от 59,1°C до 73,8°C, согласно измерению с использованием анализа RVA,
где крахмал получают из растения кукурузы waxy-sugary 2, имеющего одну дозу, две дозы или три дозы аллеля wxS, и эндосперм имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2;
причем контрольный крахмал представляет собой крахмал, полученный из контрольного растения кукурузы с генотипом эндосперма wxwxwxsu2su2su2.
2. Крахмал по п.1, дополнительно имеющий изменение энтальпии кристаллизации по меньшей мере на приблизительно 30% больше, чем изменение энтальпии кристаллизации контрольного крахмала, при измерении с использованием дифференциального сканирующего калориметра (DSC).
3. Крахмал по любому из пп. 1, 2, дополнительно имеющий распределение размера гранул крахмала, где распределение размера гранул крахмала включает по меньшей мере на приблизительно 40% меньше гранул размером менее 5 мкм по сравнению с долей гранул размером менее 5 мкм в контрольном крахмале, что наблюдают с использованием сканирующего электронного микроскопа.
4. Крахмал по любому из пп. 1-3, где крахмал представляет собой гранулу, демонстрирующую повышенную целостность по сравнению с контрольным крахмалом, что наблюдают с использованием сканирующего электронного микроскопа.
5. Крахмал по любому из пп. 1-4, где крахмал способен выдерживать по меньшей мере три цикла замораживания/размораживания.
6. Крахмал по любому из пп. 1-5, где растение кукурузы waxy-sugary 2 получают посредством перекрестного опыления первого растения кукурузы, имеющего генотип wxwxsu2su2, и второго растения кукурузы, имеющего генотип wxSwxSsu2su2 или генотип wxSwxsu2su2.
7. Крахмал по любому из пп. 1-6, экстрагированный из растения кукурузы с двойной мутацией, где растение кукурузы является гомозиготой, рецессивной по гену синтазы крахмала IIa (su2), и гомозиготой или гетерозиготой по мутантному гену гранулосвязанной синтазы крахмала I (GBSSI), где мутантный ген GBSSI имеет меньшую активность GBSSI, чем ген GBSSI (Wx) дикого типа, но более высокую активность GBSSI, чем рецессивный ген GBSSI (wx) с потерей функции,
где эндосперм растения кукурузы имеет одну дозу, две дозы или три дозы аллеля wxS и эндосперм имеет генотип wxSwxwxsu2su2su2, wxSwxSwxsu2su2su2 или wxSwxSwxSsu2su2su2.
8. Крахмал по п.7, где растение получают посредством перекрестного опыления первого растения кукурузы, имеющего генотип wxwxsu2su2, и второго растения кукурузы, имеющего генотип wxSwxSsu2su2 или генотип wxSwxsu2su2.
9. Крахмал по любому из пп. 7 или 8, где крахмал, продуцируемый в эндосперме растения кукурузы, имеет более высокую температуру клейстеризации по сравнению с крахмалом, продуцируемым в эндосперме растения кукурузы с двойной рецессивной мутацией waxy-sugary-2, имеющего генотип wxwxwxsu2su2su2.
| DANUPOL KETTHAISONG et al | |||
| Physicochemical and morphological properties of starch from fresh waxy corn kernels, J | |||
| Food Sci Tecnol, October 2015, 52 (10), p6529-6537, DOI 10.1007/s13197-015-1750-2 | |||
| AYOADE L | |||
| Relationship between α-amylase degradation and amylase/amylopectin content of maize starches, advances in Applied Science Research, 2013, 4 |

