Данное изобретение относится к способу идентификации локусов регуляции экспрессии генов в любом организме. Более конкретно, настоящее изобретение относится к применению транскриптов мРНК и изменчивости уровней транскриптов для идентификации локусов количественного признака, являющегося заданным признаком организма.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Система отбора заданных признаков у растений в программах селекции растений, как правило, основана на отборе одного или более фенотипических признаков. Однако многие важные агрономические признаки являются сложными, в значительной степени подверженными влиянию окружающей среды и находятся скорее под полигенным контролем, когда фенотипический признак регулируется множеством генов, нежели под контролем единственного локуса. В признаках под полигенным или мультигенным контролем вклад в представляющий интерес фенотип может вносить экспрессия аллелей во многих локусах.
Неоднократно показано, что многие гены не удается экспрессировать в ответ на эволюционные или внешние сигналы (окружающей среды). Понимание факторов, которые контролируют экспрессию этих генов, особенно важно для трансгенных организмов, например, при введении чужеродного гена с целью регуляции уровня их транскриптов в зависимости от эволюции, ткани или стресса. Такие исследования выявляют сложность и многоуровневую избыточность контроля экспрессии генов. Механизмы контроля экспрессии генов могут значительно варьироваться в зависимости от генов (см., например, Hirt. H. 1999 Trends Plant Sci. 4: 7-8). При такой совокупности внешних условий или условий эволюции гены, участвующие в данном процессе, не всегда реагируют одинаково и не накапливаются в клетках или тканях одного и того же типа, что указывает на различные механизмы контроля или различные сигнальные механизмы.
Используя современные методики, трудно классифицировать ключевые регуляторные гены, которые регулируют экспрессию интересующих нас генов среди целого каскада событий.
Гены с дифференциальной экспрессией идентифицированы такими методами, как дифференциальный скрининг библиотек кДНК, секвенирование генома в сочетании с поисками гомологии в банках генов, "нокаутирование" и комплементация генов, мутация гомеотических генов, высокопроизводительный скрининг EST с применением ряда генов с высокой плотностью. Эти методы, хотя они и являются трудоемкими также, иногда применяются для идентификации генов, участвующих в регуляции экспрессии генов, но они не предназначены специально для этой цели. Методы "нокаутирования" генов (мутантный вариант) для идентификации факторов, контролирующих экспрессию генов, применимы к малым видам геномов, таким как Arabidopsis. Такие методы также трудоемки, неспецифичны и часто приводят к недектируемым или летальным фенотипам. Комбинация таких методов как дифференциальная визуализация видов мРНК, субтрактивные или нормализованные библиотеки и совокупность (ряд) генов, может применяться для выделения регуляторных генов, однако эти методы не специфичны для анализа экспрессии одиночного гена и длительны. Кроме этого, в этих методах транскрипты регуляторных генов, представляющих интерес, должны присутствовать в образцах, применяемых для создания библиотек для идентификации транскриптов. Задачу можно решить, если взять множественные регуляторные гены с несинхронизированной экспрессией.
Локус количественного признака (QTL) представляет собой область генома, которая кодирует один или много белков, и это объясняет значительную долю вариабельности данного фенотипа, которая может быть под контролем полимерных генов. Как правило, для идентификации заданного QTL использовали один или более генетических маркеров. До настоящего времени в большинстве исследований QTL растительных видов рассматриваются явные (макроскопические) морфологические или агрономические фенотипы (например, урожайность, устойчивость к болезням или стрессу, время цветения и т.д.). Например, в Международной заявке WO 2000/18963 описаны растения сои, содержащие QTL, связанные с повышенной урожайностью, и методы отбора и селекции таких растений. Этот метод включает применение специфической маркерной нуклеиновой кислоты, способной гибридизоваться со второй нуклеиновой кислотой, которая на генетической карте относится к специфической области Glycine soja и которая связана с повышенной урожайностью. В патенте США QTL, ассоциируемый с BSR-резистентностью, можно применять для маркерной селекции растения. В WO 99/31964 описано применение маркерной нуклеиновой кислоты, которая, как правило, связана с набором 63 точно определенных локусов, для селекции растения. Идентифицируемый полиморфизм можно применять для фингерпринтинга ДНК и для картирования генов или QTL, связанных с устойчивостью к вредителям или болезням. В некоторых последних исследованиях QTL использовали соотношение между количественной изменчивостью специфических метаболических изменений, включая аккумуляцию метаболитов или изменения активности ферментов. Например, Byrn et al. (Byrn, P.F., McCulen, M.D., Snook, M.E., Musket, T.A., Theuri, J.M., Widstrom, N.W., Wiseman, B.R. and E.H.Coe. Proc. Natl. Acad. Sci. 93: 8820-8825) описывают картирование QTL, при этом считают для 58% вариантов концентрации майсина, флавона, действующего как фактор устойчивости хозяина-растения против совки хлопковой, для локуса, кодирующего активатор транскрипции для участков флавоноидного пути. Prioul et al. (Prioul, J.-L., Quarrie, S., Causse, M. and D.de Vienne. 1997. J. Exp. Bot. 48: 1151-1163; Prioul, J.-L., Pelleschi, S., 
 C., Causse, M., de Vienne, D. and A. Leonardi. 1999. J. Exp. Bot. 50: 1281-1288) использовали активности ферментов, уровни субстрата и продукта известного биосинтетического пути как количественные признаки в анализе QTL. Аналогично, Pelleschi et al. (Pelleschi, S., Guy, S., Kim, J.-Y., Pointe, C.,
C., Causse, M., de Vienne, D. and A. Leonardi. 1999. J. Exp. Bot. 50: 1281-1288) использовали активности ферментов, уровни субстрата и продукта известного биосинтетического пути как количественные признаки в анализе QTL. Аналогично, Pelleschi et al. (Pelleschi, S., Guy, S., Kim, J.-Y., Pointe, C.,  A., Barthes, L., Leonardi, A., and J.-L. Prioul. 1999. Plant Mol. biol. 39: 373-380) описывают применение инвертазной активности для идентификации генов-кандидатов на QTL, связанные с изменчивостью инвертазной активности в кукурузе. Damerval et al. (Damerval С., Maurice, A., Josse, J.M. and D.de Vienne. 1994. Genetics 137: 289-301) описывают применение пептидов на 2D-геле в качестве количественного признака для картирования QTL.
A., Barthes, L., Leonardi, A., and J.-L. Prioul. 1999. Plant Mol. biol. 39: 373-380) описывают применение инвертазной активности для идентификации генов-кандидатов на QTL, связанные с изменчивостью инвертазной активности в кукурузе. Damerval et al. (Damerval С., Maurice, A., Josse, J.M. and D.de Vienne. 1994. Genetics 137: 289-301) описывают применение пептидов на 2D-геле в качестве количественного признака для картирования QTL.
Ни один из вышеперечисленных методов не демонстрирует и не предполагает, что метод картирования QTL можно использовать для идентификации генов, участвующих в регуляции экспрессии одиночных генов или каскадов генов, и не предлагают использовать транскрипты мРНК в качестве фенотипического признака для картирования QTL.
Dumas et al. (Dumas p., Sun Y., Corbeil G., Tremblay S., Pausova Z., Kren V., Krenova D., Pravenec M., Hamet P., and J. Tremblay 2000, J. Hypertens 18:545-551) описывают применение мРНК в качестве фенотипического маркера для картирования QTL, связанного с экспрессией гена стресса у крысы. Сходный подход, с использованием экспрессии характерного гена для картирования QTL, связанного с кровяным давлением у крыс, был предложен Cicila and Lee (Cicila G.T. and S J Lee 1998 Hypertens Res 21: 289-296). Ни в одной из этих работ не предлагается метод QTL для идентификации генов, участвующих в регуляции одиночных генов или сложных путей регуляции (каскадов), ответственных за уровни контролируемого транскрипта. Помимо этого, нигде нет ни обсуждения картирования QTL экспрессии характерного гена в организмах, характеризующихся тем, что у них уровень плоидности выше, чем у диплоида, ни обсуждения идентифицирования QTL, связанных с экспрессией характерного гена в хозяевах неживотного происхождения.
Настоящее изобретение относится к идентификации геномных областей, участвующих в генетической регуляции одного или более интересующих генов, связанных с заданным признаком. С помощью анализа генома на основе картирования идентифицированы QTL локусы генов-регуляторов, связанных с характерной аккумуляцией транскриптов или генных продуктов.
Целью данного изобретения является преодолеть недостатки предыдущей техники.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к применению транскриптов мРНК и к изменяемости уровней транскриптов для идентификации локусов количественных признаков, относящихся к заданному признаку организма. Более конкретно, настоящее изобретение относится к способу идентификации локусов генов-регуляторов, которые контролируют экспрессию гена данного признака любого организма. Наконец, данное изобретение относится к способу идентификации генов и последовательностей генов, которые регулируют экспрессию генов и каскады-регуляторы, контролирующие экспрессивные признаки организма.
В соответствии с настоящим изобретением предлагается способ идентификации (опознавания) одной или более областей в геноме представляющего интерес организма, которые опосредуют экспрессию одного или более представляющих интерес генов, включающий:
i) определение первого представляющего интерес организма, характеризующегося тем, что интересующий организм проявляет измеримую реакцию на внешние раздражители или иным образом проявляет фенотип, связанный с дифференциальной генной экспрессией, ассоциированной с интересующим процессом;
ii) определение второго интересующего организма, характеризующегося тем, что второй интересующий организм:
а) не проявляет никакой реакции на раздражитель или проявляет не такую сильную реакцию на раздражитель, как первый интересующий организм;
б) проявляет фенотип, отличный от фенотипа первого интересующего организма, причем отличный фенотип ассоциирован с интересующим процессом;
в) проявляет фенотип, представляющий интерес, который сегрегирует в популяции, полученной в результате скрещивания с первым интересующим организмом;
г) или комбинация двух или более из а), б), в);
iii) скрещивание первого и второго интересующих организмов с целью получения популяции сегрегированного потомства;
iv) выделение РНК каждого сегрегированного потомства и количественное определение уровня генной экспрессии одного или более генов, представляющих интерес, причем один или более интересующих генов связаны с реакцией на внешний раздражитель или с интересующим процессом;
v) получение карты сцепления сегрегированного потомства с использованием одного или более маркеров;
vi) определение соотношения между одним или более маркером на карте сцепления и генной экспрессией одного или более интересующих генов и идентификация одного или более локусов количественных признаков (QTL).
vii) подтверждение того, что один или более генов локализованы в одном или более QTL.
Настоящее изобретение также включает способ, описанный выше и отличающийся тем, что после стадии скрещивания (стадия iii)) и перед стадией выделения РНК (стадия iv) сегрегированное потомство подвергают действию заданного внешнего раздражителя или характеризуют как находящееся на специфической стадии развития.
Способ по данному изобретению можно применять для идентификации одного или более QTL, соответствующих фактору транскрипции или любому фактору, контролирующему экспрессию одного или более генов, представляющих интерес.
Помимо этого, данное изобретение включает способ, описанный выше и отличающийся тем, что после стадии определения (стадия vi)) один или более генов, локализованных в одном или более QTL, выделяют и характеризуют.
Настоящее изобретение также охватывает способ идентификации регуляторного гена, который опосредует экспрессию интересующего гена, включающий:
i) определение одного или более Локусов Количественных Признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес;
ii) картирование одного или более генов-регуляторов;
iii) определение, имеется ли в одном или более QTL карта одного или более регуляторных генов;
iv) выделение одного или более регуляторных генов в одном или более QTL;
v) секвенирование одного или более регуляторных генов, выделенных на стадии iv).
В другом варианте изобретения способ идентификации регуляторного гена включает в себя:
(i) определение одного или более локусов количественных признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес; и
ii) выделение одного или более регуляторных генов в указанных одном или более QTL;
iii) секвенирование регуляторного гена.
Настоящее изобретение охватывает также способ идентификации регуляторной последовательности, которая опосредует экспрессию интересующего гена, включающий:
i) определение одного или более Локусов Количественных Признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес;
ii) картирование одного или более генов-регуляторов;
iii) определение, имеются ли в одном или более QTL карты одного или более регуляторных генов;
iv) выделение одного или более регуляторных генов в одном или более QTL;
v) секвенирование регуляторной последовательности одного или более регуляторных генов, выделенных на стадии iv).
Другой вариант способа идентификации регуляторной последовательности согласно настоящему изобретения включает в себя:
i) идентификацию одного или более локусов количественных признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес; и
ii) выделение одного или более регуляторных генов в указанных одном или более QTL;
iii) секвенирование регуляторных генов, включающих регуляторные последовательности.
Настоящий способ делает возможной идентификацию генов, имеющих общие локусы регуляции или локусы под тем же контролем (регуляцией). Это может иметь важное значение для многих заявляемых применений, таких, например, но без ограничения, как маркерная селекция, фенотипирование, картирование каскадов генов, регуляция генетического взаимодействия, анализ каскада генов и т.д.
Геномные области, ассоциированные с уровнем экспрессии генов, можно идентифицировать, используя QTL-картирование. Этот подход, использующий уровень генной экспрессии в качестве количественного признака, позволяет идентифицировать механизмы регуляции экспрессии генов, сужая область поиска таких генов-регуляторов и регуляторных последовательностей до конкретных (специфических) областей генома. Значительные QTL, идентифицируемые в данном описании, и неожиданно высокий уровень вариансы (дисперсии), который это объясняет, указывают на возможное применение QTL-анализа генной экспрессии для локализации важных регуляторных факторов, отвечающих за стимулируемую стрессом генную экспрессию. Сравнительный QTL-анализ полимерных генов проливает дополнительный свет на сложные пути регуляции генной экспрессии и позволяет по-другому классифицировать гены, имеющие общие регуляторные факторы.
Методы по данному изобретению применимы к представляющим интерес генам, экспрессия которых регулируется любыми факторами, использующими сегрегацию популяции для уровней экспрессии таких генов. Это делает возможным идентификацию геномных областей. Будучи идентифицированными, эти геномные области могут использоваться для характеристики данного генотипа без внешнего или эволюционного вмешательства, присущего методам, основанным на генной экспрессии.
С помощью идентификации одного или более QTL для количественных признаков, стимулируемых внешними раздражителями (например, но без ограничения, температурным стрессом) или связанных с данным процессом (например, но без ограничения, с определением выхода), методика по данному изобретению позволяет идентифицировать также один или более QTL, которые обычно связаны с анализируемым фенотипическим признаком и с экспрессией генов, имеющих большое адаптивное значение для данного признака. Регуляторные гены и регуляторные последовательности, обнаруженные в этих локусах, по-видимому, важны для улучшения количественного признака.
Следует отдавать себе отчет, что способ по данному изобретению применим для любого организма-хозяина, однако существуют некоторые организмы, - например, но без ограничения, одноклеточные организмы или растения, - которые имеют уникальные преимущества перед другими эукариотными системами в том, что касается применения способа по настоящему изобретению. Например, у некоторых видов растений поколение двойных галлоидов пыльцы, в которой каждый локус является гомозиготным, самовоспроизводство или обратное скрещивание с одной или другой родительской линией F1s с образованием сегрегированной F2 популяции или бэккросс-популяции, возможность неоднократного самоопыления отдельных растений F2 в течение нескольких поколений (F6-F10) с образованием ряда рекомбинантных инбредных линий, возможность каждого генотипа размножаться путем клонального размножения позволяют осуществлять множественный деструктивный анализ одного и того же генетического материала и дают возможность подвергать генетически идентичный материал действию различных внешних раздражителей или другим воздействиям. Помимо этого, эти типы клонального материала могут воспроизводиться в различных внешних условиях, когда можно изучать влияние взаимодействия генотипа, окружающей среды, или генотипа и окружающей среды, на фенотип.
Это краткое описание данного изобретение не обязательно описывает все необходимые отличительные признаки изобретения, но содержание данного изобретения может также отражаться в "субкомбинациях" описанных признаков.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Эти и другие отличительные признаки данного изобретения станут более наглядными из следующего описания, в котором имеются ссылки на прилагаемые фигуры, где:
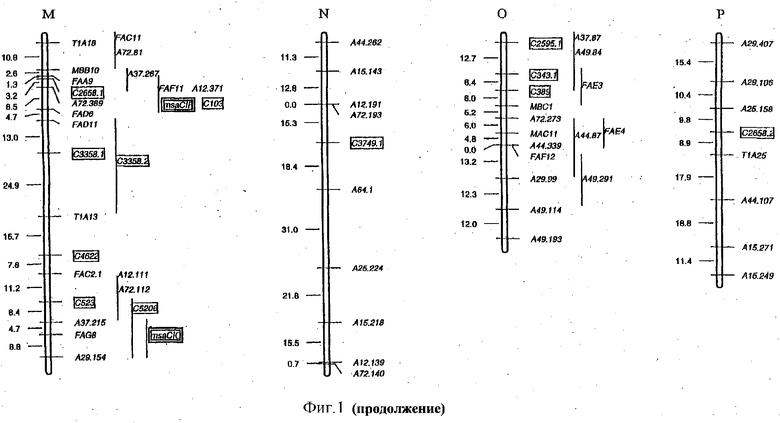
ФИГУРА 1 изображает карту сцепления диплоидной люцерны. Локусы перечисляются справа, а расстояния между рекомбинациями (см) перечисляются слева от каждой группы сцепления. Столбцы справа от групп сцепления указывают примерное положение локусов. Гены, регулируемые холодовой реакцией (msaCI), транскрибируется с двойной рамкой считывания, а гены, имеющие гомологию с генами с регуляторной функцией, - с рамкой считывания. Гены, являющиеся представителями мультигенного семейства, отличаются нумерацией, например, С3358.1 и С3358.3.
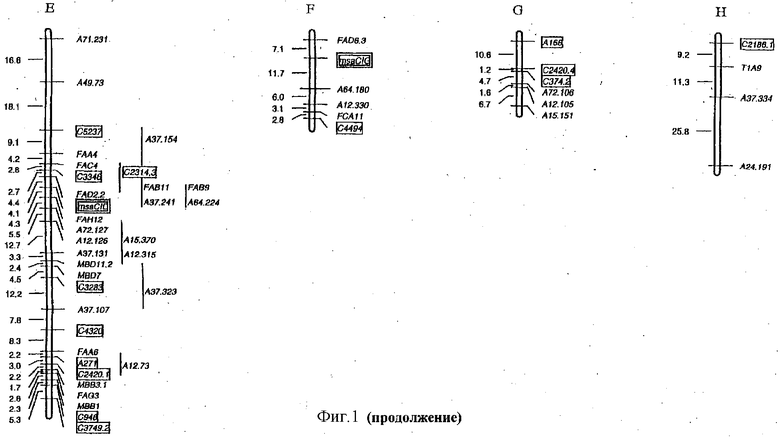
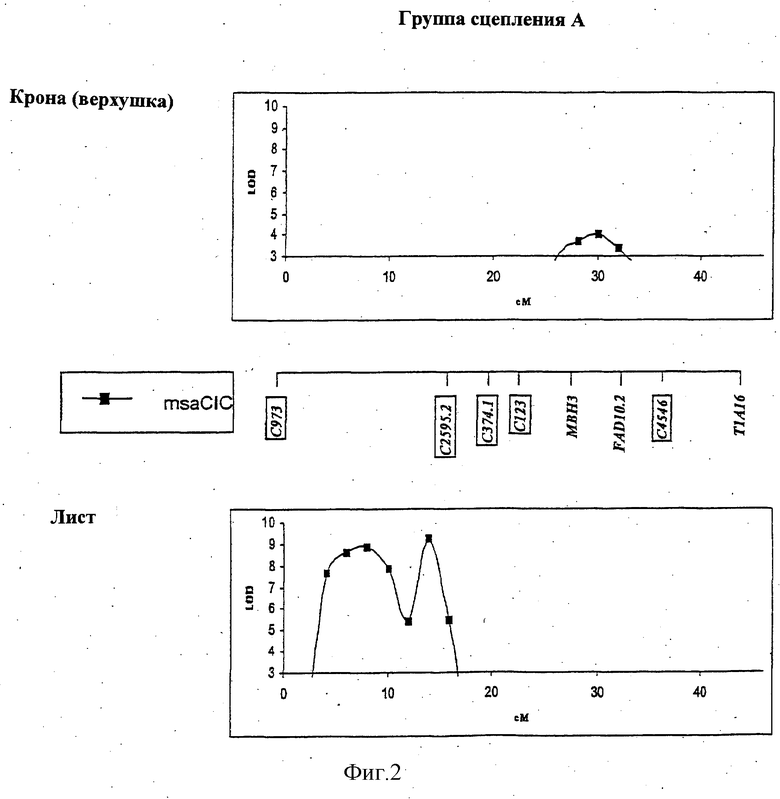
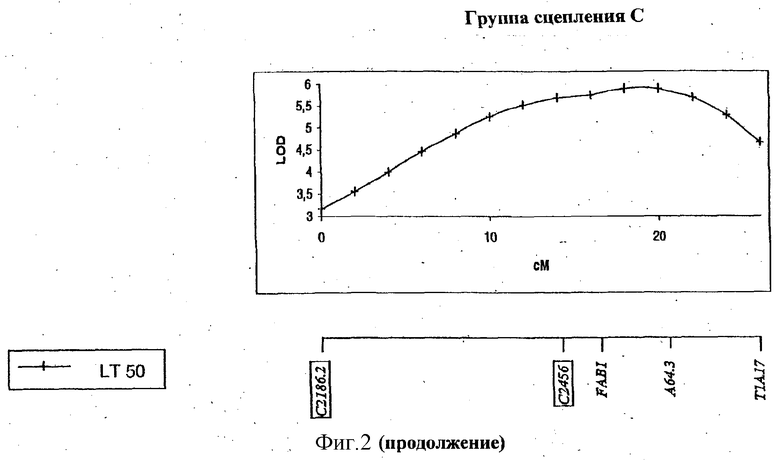
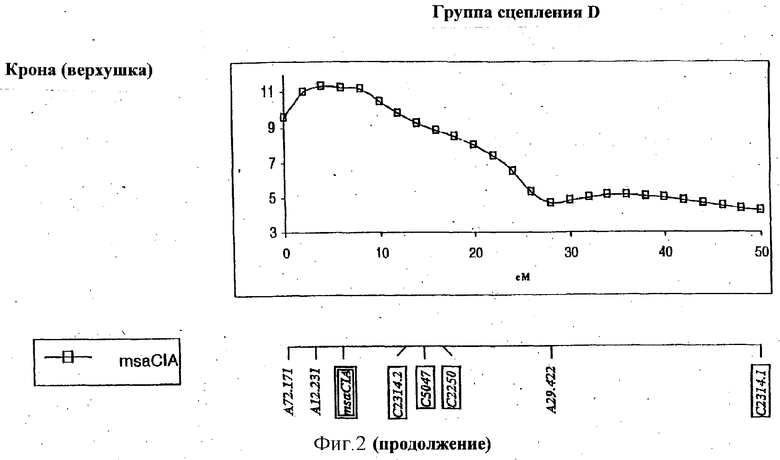
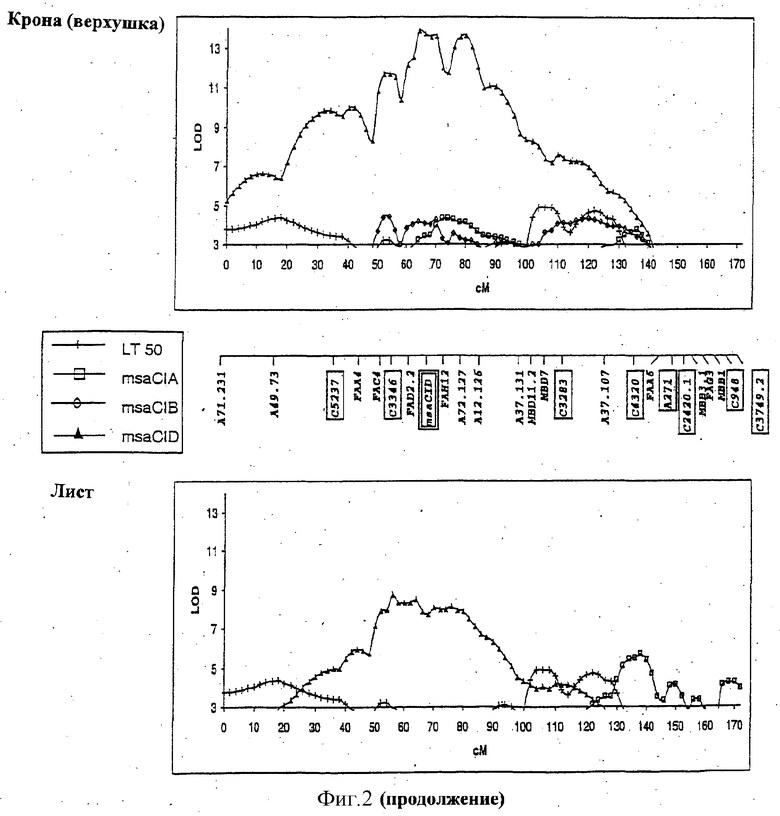
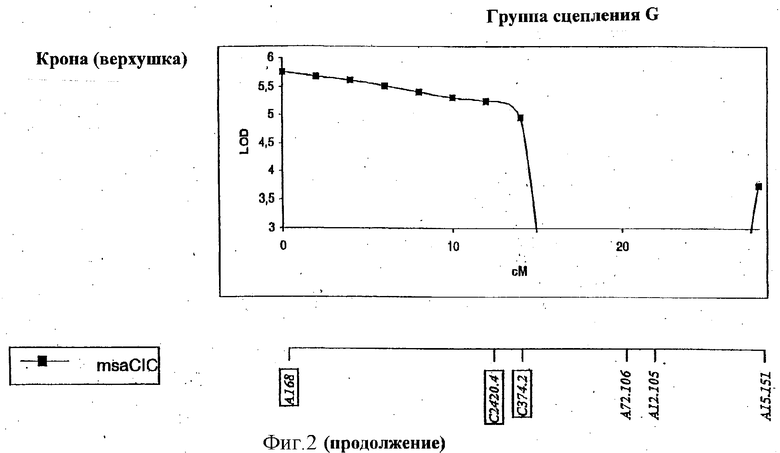
ФИГУРА 2 - положение и статистическая значимость (значения LOD) QTL для LT50 и экспрессии генов, регулируемых холодовой реакцией (msaCI). Гены msaCI транскрибируется с двойной рамкой считывания, а гены, имеющие гомологию с генами с регуляторной функцией - с рамкой считывания. Если QTL генной экспрессии обнаружены как в листьях, так и в кроне, график LOD-значений экспрессии в кроне дан вверху, а график LOD-значений экспрессии приводится внизу. LT50 QTL относятся к растению в целом и не зависят от типа ткани. Гены, являющиеся представителями мультигенного семейства, отличаются нумерацией, например С3358.1 и С3358.3.
ФИГУРА 3 представляет собой один пример анализа дот-блоттинга экспрессии интересующего гена, в данном случае гена msaCIA, в сегрегированном потомстве F2 диплоидной люцерны. Каждое пятно соответствует уникальному генотипу, за исключением последней колонки. Уровень транскриптов количественно определяют денситометрией и используют в качестве количественного признака для анализа QTL. Указано значение оптической плотности для каждого генотипа и контрольное.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНОГО ВАРИАНТА ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу идентификации регуляторных локусов, которые контролируют генную экспрессию в организме. Более конкретно, настоящее изобретение относится к применению транскриптов мРНК и изменчивости уровней транскриптов для идентификации локусов количественного признака, являющегося заданным признаком организма.
Нижеследующее описание представляет собой описание предпочтительного варианта изобретения только на примере и без ограничения в отношении сочетания отличительных признаков, необходимых для осуществления изобретения.
Локус количественных признаков (QTL) представляет собой область генома, которая объясняет значительную долю вариабельности данного фенотипа, контролируемого полимерными генами, например, но без ограничения, признаки, ассоциируемые с повышенной урожайностью, устойчивостью к морозам, к засухе и т.п. В этих областях локализован один или более генов, кодирующих факторы, которые оказывают значительное влияние на фенотип организма.
Как указано в данном описании, карта сцепления, построенная с применением маркера любого типа, например морфологических, маркеры на основе изозима или ДНК могут использоваться для локализации каждого интересующего гена на карте. Экспрессия отдельного интересующего гена рассматривается как количественный признак. Не желая связывать себя теорией, отмечаем, что если изменчивость экспрессии интересующего гена является результатом различий в самом гене, например, результатом различных аллельных форм гена, затем основной локус количественного признака следует локализовать близ местоположения структурного гена. Однако, как подчеркивается ниже, наблюдается, что варианса экспрессии интересующего гена высока и в значительной степени объясняется QTL, локализованными где-либо в геноме (см. Фигура 1). Следовательно, геномные области, локализованные на расстоянии от интересующего гена, являются контролирующими или регулирующими экспрессию интересующего гена. Эти геномные области можно идентифицировать с помощью карты сцепления, а затем методами, известными специалистам в данной области техники, определяют регуляторные гены, кодирующие факторы, которые контролируют экспрессию интересующего гена.
Под "геном, представляющим интерес", "интересующим геном" понимают один или более известных генов, которые могут применяться в качестве количественного признака, который можно охарактеризовать с помощью способа по настоящему изобретению, и могут включать области в обратном, 3'-5'-направлении (upstream), в прямом, 5'-3'-направлении (downstream), интроны и экзоны. Интересующий ген, как правило, но не всегда, экспрессирует в условиях, применяемых для определения влияния регуляторных генов на экспрессию или супрессию интересующего гена. Уровень экспрессии интересующего гена можно определить любыми методами, известными из уровня техники, например, нозерн-блоттингом, защитой от РНК-азы, матричным анализом, PCR и т.п. Интересующий ген или уровень его транскриптов представляют собой количественный признак, который используется для дополнительной идентификации одного или более QTL, ассоциированных с экспрессией интересующего гена. По способу согласно данному изобретению для анализа реакции на внешний раздражитель или другого интересующего процесса по данному описанию можно использовать один или более интересующих генов. Увеличивая число интересующих генов, используемых в данном анализе, можно идентифицировать один или более QTL, отвечающих за регуляцию некоторых интересующих генов, ассоциированных с ответом на внешний раздражитель или с другими процессами в растении, тем самым увеличивается значение QTL. Такой способ описан ниже и в Примерах.
Под "регуляторным геном", "геном регуляции" понимают ген, генный продукт которого прямо или косвенно влияет на экспрессию интересующего гена, и который включает кодирующую область, обратную (5', upstream) и прямую (3', downstream) некодирующие и регуляторные области и интроны. Как правило, регуляторный ген по-разному экспрессирует в ответ на раздражитель, например, но без ограничения, внешний раздражитель, такой как тепловой шок или температурный стресс. Примеры регуляторного гена включают, но без ограничения, ген, который кодирует фактор транскрипции, или ген, который кодирует другой белковый фактор, каким-либо образом регулирующий экспрессию другого гена. Продукт, кодируемый геном-регулятором, может непосредственно влиять на экспрессию интересующего гена, например, но никоим образом не ограничивая, связывая регуляторные области "вверх" (5', upstream) и "вниз" (3', downstream) от кодирующей области интересующего гена, или в интронах, как известно из уровня техники, и усиливая или ослабляя (заставляя "замолчать") экспрессию интересующего гена. Регуляторный ген может также кодировать продукт, который влияет либо на стабильность транскрипта интересующего гена, скорость или устойчивость трансляции, либо на транскрипт, синтезированный из интересующего гена, или как на событие трансляции, так и на пост-трансляционные события, относящиеся к экспрессии интересующего гена. Также предполагается, что регуляторный ген может влиять на дифференциальную экспрессию интересующего гена косвенно, опосредуя экспрессию одного или более вторичных генов, генные продукты которых могут затем действовать на экспрессию интересующего гена. Также ожидается, что один или более регуляторных генов может действовать как синергист, опосредуя дифференциальную экспрессию интересующего гена.
Способ по данному изобретению позволяет идентифицировать гены, имеющие общие регуляторные локусы или находящиеся под одним и тем же контролем. Это может иметь важное значение для применения во многих направлениях, например, но без ограничения, таких как селекция с помощью маркера, фенотипирование, картирование каскадов генов, регуляция генетического взаимодействия, анализ каскада генов или идентификация генов-регуляторов, оказывающих заметное влияние на количественные признаки.
Под "внешним раздражителем" ("раздражителем из внешней среды") понимают любой раздражитель (стимул), влияющий на и вызывающий измеримый ответ (измеримую реакцию) в интересующем организме. Например, но без намерения какого-либо ограничения, интересующий организм может представлять собой растение, часть растения, его ткань или клетку, а внешний раздражитель может представлять собой, например, но без ограничения, биотические стимулы, включая вирусное, бактериальное, грибковое заражение, заражение насекомым, нематодой или другими травоядными, или абиотические раздражители, например, но без ограничения, стресс, обусловленный охлаждением, замораживание, вода, засуха, осмос, тепло, окислитель или загрязнитель. Внешний раздражитель может также включать действие на интересующий организм, его орган, ткань или клетку минерального питания, света и эндогенных или экзогенных химических веществ.
Под "интересующим процессом" подразумевают любой процесс, который вызывает измеримый эффект в интересующем организме или его органе, ткани или клетке, например, но это ни в коем случае не ограничение, в растении. Интересующий процесс может включать, но без ограничения, эволюционный, химический или экологический контроль генной экспрессии во время, но без ограничения, развития растения, эмбриогенеза, развития цветов, семян, корней или листьев, органогенеза, или циркадный, ультрадный и другие внутренние ритмы. Другие интересующие процессы могут включать, но без ограничения, сбор урожая, транслокацию фотосинтана, сток и источник, индекс листовой поверхности, соотношение массы надземных и подземных органов растения, питательную ценность собранного материала, морфологию растения, скорость клеточного цикла, скорость клеточной дифференцировки, размер клетки, жизненный цикл растения, старение, созревание, покой, зарождение или геномные перестройки, например, вследствие активации транспозона. Однако следует понимать, что аналогичные или другие уникальные процессы в любом интересующем организме могут быть охарактеризованы представленными в данном описании способами.
Под "интересующим организмом" ("организмом, представляющим интерес") понимают любой организм, в котором нужно определить и охарактеризовать один или более QTL, например, но без ограничения, растение (водоросли, мхи, папоротники, покрытосемянные или голосемянные растения), животное, одноклеточный организм, бактерии, фитопланктон, дрожжи, грибы, а также клеточные и тканевые культуры этих организмов.
Способ по данному изобретению можно применять для идентификации уровня транскриптов гена, конкретное местоположение которого в геноме определяют по карте, и для того, чтобы определить, регулируется ли аккумуляция кодируемого транскрипта или белка генами, локализованными в локусе где-то в геноме. Применяя анализ генома на основе карты QTL (локусов количественных признаков), можно идентифицировать локусы, ассоциируемые с дифференциальной аккумуляцией транскриптов или генных продуктов. Следовательно, гены, обнаруженные в этих QTL, могут кодировать представителей мультигенных семейств под различным транскрипционным контролем (например, укрывание (депонирование) важных цис-регулирующих последовательностей, которые, в свою очередь, влияют на регуляцию транскрипции или действуют на "upstream" (вверх) или "downstream" (вниз) последовательности, требующиеся для усиленной трансляции или устойчивости мРНК), факторы транскрипции (включая энхансер и репрессоры), белки, участвующие в сигнальной трансдукции (например, протеинкиназы, протеинфосфатазы, белки 14-3-3 и т.д.), рецепторы внешних сигналов, белки, участвующие в регуляции уровней вторичных мессенджеров (Са2+, IP3, сАМР и т.д.), нуклеазы, участвующие в расщеплении транскриптов, протеазы и ингибиторы протеаз, новые факторы регуляции транскрипции.
Пример способа, применяемого для определения регуляторных генов, которые контролируют, опосредуют или влияют на экспрессию одного или более интересующих генов, например, но без ограничения, дифференциальную экспрессию в ответ на внешний раздражитель, или любое событие в интересующем организме, приводящее в результате к дифференциальной экспрессии гена, включает:
i) определение первого интересующего родительского организма, который проявляет реакцию на внешние стимулы или иным образом проявляет фенотип, обусловленный дифференциальной генной экспрессией, связанной с интересующим процессом, и второго интересующего родительского организма, у которого отсутствует или проявляется не так сильно реакция на раздражитель, или он проявляет отличный фенотип, обусловленный дифференциальной генной экспрессией, связанной с интересующим процессом, вне зависимости от того, обнаружен ли этот вариант в природе или получен мутацией гена, разрывом гена или в результате инсерции в гене. Желательно, но не обязательно, чтобы первый и второй родители по-разному отвечали на раздражитель, или чтобы они отличалась по дифференциальной экспрессии гена, обусловленной интересующим процессом, пока ответ на стимул или различие в дифференциальной генной экспрессии сегрегирует в потомстве;
ii) скрещивание первого и второго родителей с получением популяции сегрегированного потомства. Типы популяций могут включать, но без ограничения, F1 (от гетерозиготных родителей), семейства F2 или F3, рекомбинантные инбредные линии, двойные гаплоиды, линии обратного скрещивания (бэккросс-линии), линии тест-кросс и мегагаметофиты, расщепляющие голосемянные растения;
iii) если требуется, воздействие на поколение от этого скрещивания заданного внешнего раздражителя (стимула);
iv) выделение РНК из одного или более органов, тканей или типа клеток, например, в случае растения как интересующего организма РНК можно получать из тканей или клеток цветка, примордия, листа, ствола или корня;
v) измерение уровней транскрипта одного или более интересующих генов любым подходящим способом, такими как, например, но без ограничения, дот-блоттинг или нозерн-блоттинг, "array" анализ или количественная PCR (ПЦР), "array" анализ и количественное определение генной экспрессии для каждого потомства;
vi) получение генетической карты сцепления для сегрегированной популяции любым подходящим методом, например, но без ограничения, с применением маркеров на основе ДНК, таких как RFLP (полиморфизм длин рестрикционных фрагментов), AFLP (полиморфизм длин амплифицированных фрагментов), RAPD (статистически амплифицированная полиморфная ДНК), микросателлиты, IMP (интер MITE полиморфизм) или SNP (полиморфизм одиночного нуклеотида), маркеров белков или морфологических маркеров - для характеристики отдельных представителей картируемой популяции и рассмотрение данных, полученных одним или более из вышеуказанных методов, для анализа сцепления двух или многих факторов с целью создания карты сцепления;
vii) определение связи между маркерами на карте сцепления и экспрессией одного или более интересующих генов в ответ на внешний раздражитель, или связанной с интересующим процессом, с целью идентификации статистически значимых QTL. Такую связь можно определить любым методом, известным специалистам в данной области техники, например, но без ограничения, однофакторные дисперсионные анализы ANOVA, единичную регрессию, картирование в интервале (Interval mapping), картирование в сложном интервале (Composite interval mapping). Такой анализ можно осуществлять с применением MAPMAKER/QTL, MQTL, QTL Cartographer или другого подобного программного обеспечения. Вычисляют также статистическую значимость и процент дисперсии для каждого QTL.
Способ, представленный в данном описании, является эффективным для исчерпывающей идентификации многих, если не всех, регуляторных факторов (генов), участвующих в регуляции экспрессии интересующего гена, и он позволяет оценить относительный вклад каждого фактора в наблюдаемую изменчивость экспрессии интересующего гена, а также идентифицировать локализацию в геноме регуляторного гена и связанные маркеры, которые можно использовать в традиционном разведении растений. Помимо этого, "хозяин-переключатель" - регуляторные гены, влияющие на экспрессию многих генов, например, в каскаде, могут быть идентифицированы, когда одновременно анализируются полимерные гены, стимулированные данным внешним раздражителем или связанные с данным процессом.
Когда один или более QTL, которые в значительной степени связаны с экспрессией интересующего гена, идентифицированы, тогда каждый из этих локусов и связанные маркеры можно использовать либо непосредственно в качестве маркеров, например, но без ограничения, для целей размножения или скрининга, включая разведение растения, или для дополнительной характеристики с целью определения гена или генов, связанных с экспрессией интересующего гена, методами клонирования на основе картирования, известными специалисту в данной области техники. Например, один или более регуляторных генов можно картировать, чтобы определить, соответствует ли генетическая локализация этих генов QTL, регулирующим экспрессию мРНК интересующего гена. Подтверждение того, что такой соответствующий регуляторный ген влияет на экспрессию одного или более интересующих генов, можно получить с помощью обычно применяемых в технике методов, например, но без ограничения, методов генетической трансформации, генетической комплементации или нокаутирования, или с помощью сверхэкспрессии. Карту генетического связывания также можно использовать для выделения регуляторного гена, включая любые новые регуляторные гены, с помощью методов клонирования на основе карты, которые известны в технике, при этом маркеры, позиционированные в QTL, применяются для того, чтобы подойти к интересующему гену, используя контиги геномных клонов с большими инсерциями. Позиционное клонирование представляет собой такой метод, который можно использовать для выделения одного или более регуляторных генов, как описано в Martin et al. (Martin, G. В., Brommonschenkel, S.H., Chungwongse, J., Frary, A., Ganal, m. W., Spivey, R., Wu, Т., Earle, E.D. and S.D. Tanksley, 1993, Science 262: 1432-1436; вводится в данное описание в качестве ссылки), однако можно также использовать другие подходящие методы, такие, которые признаны специалистами в данной области техники. Опять же подтверждение того, что такой соответствующий регуляторный ген влияет на экспрессию одного или более интересующих генов, можно получить методами генетической трансформации и комплементации или нокаутирования, описанными ниже.
Следовательно, настоящее изобретение относится к идентифицированию областей в геноме растения, которое участвует в опосредовании экспрессии одного или более интересующих генов в ответ на внешний признак, или процесса, связанного с дифференциальной экспрессией гена в интересующем процессе. Данное изобретение также относится к характеристике идентифицируемых QTL для определения одного или более регуляторных генов и регуляторных последовательностей в этих локусах.
Способ по данному изобретению можно также использовать для идентификации локусов, соответствующих факторам (кодируем регуляторными генами), ассоциированным с контролем или опосредованием экспрессии интересующего гена в интересующем организме. Этот способ можно применять для идентификации регуляторных генов, которые контролируют и модулируют уровень транскриптов мРНК одного или более интересующих генов, вне зависимости от того, обнаружен ли этот вариант в природе или индуцирован мутацией гена, расщеплением гена или инсерцией, включая, например, но без ограничения, интересующие гены, которые регулируются факторами, вовлеченными в:
- биотические стрессы, например, но без ограничения, вирус, бактерии, грибки, насекомые или нематоды;
- абиотические стрессы, включая, но без ограничения, охлаждение, замораживание, воду, засуху, осмотические стрессы, тепло, соль, окислительные стрессы, загрязнители;
- генную экспрессию под эволюционным, химическим и экологическим контролем;
- эмбриогенез, например, но без ограничения, развитие ткани или органа, которое в случае растения может включать развитие семян, органогенез, развитие цветка;
- минеральное питание, включая как макро-, так и микроэлементы;
- свет (уровень освещенности и световые характеристики);
- циркадный, ультрадный и другие внутренние ритмы;
- химические индукторы, включая природные регуляторы роста, например, в случае, если интересующий организм представляет собой растение, ауксины, гиббереллины, АВА, цитокинины, этилен и их аналоги, синтетические гормоны, гербициды, салициловая кислота, жасминовая кислота;
или интересующие гены, оказывающие значительное влияние на биологические процессы, такие как:
- определение выхода, например, но без ограничения, у растений, определение собираемого урожая за счет влияния регуляторных генов на биомассу, завязывание семян (количество и размер), транслокацию фотосинтата, сток и источник, индекс листовой поверхности, соотношение массы подземных и надземных органов;
- питательная ценность биомассы, например, но без ограничения, собираемый материал и связанное с ним влияние на уровни качественных показателей, таких как кофакторы, витамины, белки, антиоксиданты, легко усваиваемые волокна;
- морфология, например, но без ограничения, в растениях, влияние регуляторных генов на окраску цветка (например, гены, участвующие в синтезе антоцианинов), высоту растения (например, гены, участвующие в синтезе гиббереллинов), длину междоузлия, место прикрепления листа;
- частота клеточного цикла, например, но без ограничения, циклины;
- степень клеточной дифференцировки, например, но без ограничения, белки гомеодомена;
- размер клетки; например, но без ограничения, в растениях, гены, участвующие в синтезе ауксинов, рецепторы ауксина, ауксин-индуцированные гены;
- жизненный цикл организма, например, за счет влияния регуляторных генов на старение, созревание, покой, прорастание;
- геномные перестройки, например, активация транспозона.
Помимо применения способа по настоящему изобретению для отнесения локусов, соответствующих фактору транскрипции или любым факторам, контролирующим экспрессию генов в интересующем организме, к различным заданным признакам, маркеры, локализованные в этих QTL, могут также применяться для практического улучшения сорта. Например, идентифицированные полиморфные формы, ассоциируемые с генетическими маркерами, локализованными в локусе (QTL), можно использовать: в методах диагностики и селекции на основе генома:
- для прослеживания потомства организма;
- определения гибридности организма;
- для идентификации изменчивости связанных фенотипических признаков, признаков экспрессии мРНК; или как фенотипических признаков, так и признаков экспрессии мРНК;
- в качестве генетических маркеров для конструирования генетических карт сцепления;
- для идентификации индивидуального потомства от скрещивания, при этом в потомстве имеется "вклад" родителя-донора, родителя-реципиента или обоих родителей - донора или реципиента;
- для выделения последовательности геномной ДНК около кодирующей ген или некодирующей последовательности ДНК, например, но без ограничения, последовательности промотора или регуляторной последовательности;
- в маркерной селекции, при клонировании с помощью карты, сертификации гибридов, для фингерпринтинга, генотипирования и аллель-специфического маркера;
- в качестве маркера интересующего организма.
Анализ с помощью QTL-карты позволяет также определить регуляторные локусы, ассоциированные с экспрессией интересующего гена в организме в целом, органе, ткани или на специфическом клеточном уровне, в ответ на внешний раздражитель, или фенотип, ассоциированный с интересующим процессом. Например, но это ни в коем случае не следует рассматривать как ограничение, возможные регуляторные гены, которые модулируют экспрессию мРНК, такие как (см. также Таблицу 2, Пример 2):
- Транскрипционные факторы (цинковые пальцы и белки АР2);
- Митоген-активированные протеинкиназы (ММК4, ММК3, ММК2, ММК1);
- Кальций-зависимые протеинкиназы;
- Серин/треонинпротеинкиназа;
- Са2+-транспортирующая АТФ-аза;
- ГТФ-связывающие белки;
- РНК-связывающие белки;
- Протеинфосфатазы (тип 2А и 2С);
- Са+2-связывающие белки, например, кальмодулин;
- Белки 14-3-3;
- Белки, активирующие ГТФ-азу;
- Белок аденилатциклазы;
- Фосфолипаза С;
- Липоксигеназа;
- Гистондеацетилаза;
- Рецепторная киназа; или
- Фосфатидилинозит-3-киназа,
можно оценить количественно, чтобы определить, не локализованы ли они на карте в областях генома (QTL), которые объясняют высокий уровень (соотношение) экспрессии генов, регулируемых холодовой реакцией, в растении, например, но без ограничения, в люцерне (Medicago falcata L).
Представленный способ можно использовать для пополнения высокопроизводительных аналитических исследований генома на основе элементов (чипов) ДНК и сетчатых матриц (array) или любой другой системы, способной измерять генную экспрессию. Используя сегрегированную популяцию, можно локализовать QTL, контролирующие уровень транскриптов генов, экспрессия которых связана с интересующими фенотипами (например, устойчивость к стрессу и болезни, биохимическая продукция, морфологические варианты и т.д.). Картирование предполагаемых генов (кандидатов) из коллекций EST устанавливает возможную колокализацию с важными QTL и позволяет идентифицировать аллельный полиморфизм и наличие аллель-специфических маркеров. Идентификация этих QTL и ассоциированных регуляторных генов можно использовать для методов геномной диагностики и селекции с целью оценки потенциала генной экспрессии в данных эволюционных и экологических (внешних) условиях и координации генной экспрессии в зависимости от эволюции, ткани и экологии.
Геномные области, связанные с уровнем экспрессии генов, например, но без ограничения, области, активируемые низкой температурой, можно идентифицировать методом QTL-картирования. Этот подход, использующий уровень генной экспрессии в качестве количественного признака, делает возможным определение механизмов контроля генной экспрессии за счет сужения области поиска таких регуляторных генов до специфических геномных областей. Последовательности ДНК, отвечающие за изменяемость генной экспрессии, по-видимому, также можно обнаружить в этих областях. Метод идентификации гена, представленный в данном описании, будет проще и быстрее, если станут более доступными последовательности генома в целом и рутинная интеграция все более увеличивающегося числа EST при нанесении на карты с высокой плотностью или за счет преимущества обширной колинеарности блоков генов в хромосоме родственных видов по сравнению с генетическим картированием.
Весьма значительные QTL, идентифицируемые в Примерах, приведенных в данном описании, и неожиданно высокий уровень дисперсии показателя, который они объясняют, указывает на возможность применения QTL-анализа генной экспрессии для локализации важных регулирующих факторов, ответственных за индуцированную стрессом генную экспрессию. Сравнительный QTL-анализ полимерных генов, индуцированных одинаковыми внешними раздражителями или участвующих в данном процессе, позволяет прояснить сложные пути регуляции генной экспрессии и по-новому сгруппировать гены с общими сходными факторами регуляции.
Способ по данному изобретению применим к генам, экспрессия которых регулируется любыми факторами, использующими сегрегацию популяции для уровней экспрессии таких генов. Это позволяет идентифицировать геномные области, которые можно использовать для того, чтобы охарактеризовать данный генотип без экологического (внешнего) или эволюционного влияния, присущего способам, основанным на экспрессии генов.
Для идентификации регуляторных генов, которые могут быть локализованы в одном или более QTL, которые объясняют значительную долю изменяемости экспрессии одного или более интересующих генов, получают зонды известных регуляторных генов, например, но без ограничения, вышеперечисленные регуляторные гены и регуляторные гены, приведенные в Таблице 2, Пример 2, и эти регуляторные гены или продукты их экспрессии наносят на карту сцепления. Эти зонды могут включать нуклеотидные фрагменты или полноразмерные гены для обнаружения ДНК или РНК, или антитела для обнаружения продуктов экспрессии регуляторного гена. Следует отдавать себе отчет, неизвестные регуляторные гены, локализованные в одном или более QTL, можно идентифицировать, используя методы, известные специалисту в данной области техники, например, но без ограничения, позиционное клонирование секвенированной ДНК, чтобы определить, имеется ли колокализация ДНК (карты) с одним или более идентифицированных QTL. Предпочтительно, локализацию предполагаемых регуляторных генов (кандидатов) в одном или более TL подтверждают, например, картированием областей "вверх" (3'-5') и "вниз" (5'-3') от каждого предполагаемого регуляторного гена на карте сцепления. Таким образом, только регуляторные гены, которые имеют 5' или 3' области, колокализующиеся с одними и теми же QTL, используют для дальнейшей характеристики.
Регуляторные гены, локализованные в одном или более QTL, ассоциирующемся с экспрессией одного или более интересующих генов, можно выделить и охарактеризовать, и их экспрессию можно модулировать в трансгенном растении, например, но без ограничения, в люцерне, для того, чтобы оценить их влияние на экспрессию интересующего гена и, если требуется, их влияние на ответ интересующего организма на экологический стресс. Такой анализ может включать, но без ограничения, комплементацию или "нокаутирование" гена, и активацию гена, исследования, в которые входит трансформация интересующего организма при участии смысловой или антисмысловой конструкции регуляторного гена или энхансерных последовательностей, соответственно. Интересующий организм, не отвечающий на внешний раздражитель или проявляющий слабую реакцию на него, или интересующий процесс, трансформируются при использовании смысловой конструкции регуляторного гена, тогда как интересующий организм, который проявляет заметную (сильную) реакцию или оценивается как позитивный для интересующего процесса, трансформируется при участии антисмысловой конструкции регуляторного гена или разрываются с помощью инсерции чужеродной ДНК, такой как Т-ДНК, до кодирующей области гена, или активируется при участии инсерции энхансерной последовательности 5' или 3' интересующего гена. Затем можно проводить мониторинг экспрессии одного или более интересующих генов и регуляторного гена, чтобы определить влияние регуляторного гена со смысловой, антисмысловой инсерцией, "нокаутированного" регуляторного гена или регуляторного гена с энхансерной инсерцией на реакцию организма на внешний раздражитель или интересующий процесс. Можно также определить влияние смысловой, антисмысловой, "нокаутирующей" или энхансерной инсерционной конструкции на физиологический ответ на экологический стресс.
Также ожидается, что регуляторный ген, локализованный в специфическом QTL или в группе QTL, можно использовать и в качестве маркера для одного или более QTL в программах по улучшению сорта (породы).
Таким образом, данное изобретение также относится к способу идентификации регуляторного гена, который опосредует экспрессию интересующего гена который включает:
i) создание карты сцепления интересующего организма;
ii) идентификацию одного или более QTL, которые объясняют значительную долю изменяемости экспрессии одного или более интересующих генов; и
iii) выделение и картирование одного или более регуляторных генов с целью определить, имеются ли карты регуляторного гена в одном или более из идентифицируемых QTL.
Настоящее изобретение относится также к опосредованию реакции интересующего организма на внешний (экологический) стресс или к определению характеристик in vivo влияния идентифицируемого и выделяемого регуляторного гена путем введения в интересующий организм регуляторного гена и, необязательно, определяя влияние регуляторного гена на экспрессию одного или более интересующих генов, или его влияние на реакцию интересующего организма на внешний раздражитель.
Настоящее изобретение далее иллюстрируется нижеприведенными примерами. Однако следует понимать, что эти примеры даны только с целью иллюстрации и никоим образом не должны ограничивать объем настоящего изобретения.
Пример 1: Идентификация QTL, ассоциируемых с экспрессией генов, регулируемой холодовой реакцией.
Общее описание экспериментального метода
Фенотипы экспрессии генов, регулируемых холодовой реакцией (генов msaCI), и LT50 используются в качестве количественных признаков, которые следует охарактеризовать в клональных побегах генотипов F2 диплоидной люцерны. Признаки (элементы, patterns) сегрегации фенотипов (гены экспрессии msaCI и LT50) применяют для поиска локусов количественных признаков (QTL) на карте сцепления генома диплоидной люцерны.
Растительный материал
Популяцию F1 создают перекрестным опылением диплоидных родителей, резко отличающихся по своей толерантности к холодовой реакции (М.falcata x M.sativa). Линию F1 получают перекрестным опылением между чувствительным к холоду генотипом культурной люцерны М.sativa, диплоидные растения (Bingham, E.T. and T.J.МсСоу. 1979. Crop Sci. 19: 97-100), в качестве женского родителя и выносливым генотипом диплоидных растений М.falcata cv. Anik (Pankiw,P. and Siemens, 1976. Can. J. Plant Sci. 56: 203-205) в качестве мужского родителя. Проводят эмаскуляцию цветков и вручную проводят опыление. Один случайно выбранный генотип Fi оплодотворяют, опыляя вручную, и потомство F2 от этого скрещивания используют для составления карты сцепления и QTL-анализа.
Условия выращивания
Черенки генотипов 117F2 разводят клонированием и перевивают в виде вставок в глубоких вегетационных сосудах после заложения корня. Клональные побеги выращивают затем в контролируемых внешних условиях при температурах 21°С и 17°С (день, ночь, соответственно), при 16-часовом световом периоде и освещенности с примерной плотностью светового потока 225 мкмол м-2 s-1.
Условия акклимации к холоду
Растения проходят акклимапию к холоду в течение двух недель в камере для выращивания при 2°С с 8-часовым световым периодом и освещенностью с плотностью светового потока 125 мкмол м-2 s-1.
Определение LT50
Клональные побеги каждого генотипа сегрегированной популяции и родительских генотипов проверяют с помощью теста на замерзание, описанного Castongway et al. (Castongway, Y., Nadeau, P., and S. Laberge 1993. Plant Cell Physiol. 34: 31-38). Растения помещают при комнатной температуре и постепенно (поэтапно) ее понижают, в перерывах отдельные растения реабилитируют и дают возможность снова расти в начальных условиях роста (21/17°С, дневная/ночная температуры, соответственно) в течение трех недель перед тем, как оценивать выживаемость растения. Температуру, при которой гибнет 50% растений (LT50), определяют по методу SAS™ Probit (Statistical Analysis System, Cary, NC). Устанавливают три копии неполноблочной конструкции с циклическими перестановками (12 блоков), чтобы статистически сравнить большое количество экотипов, применяемых для исследования.
Описание msaCI генов
Семь msaCI генов, выделенных с использованием библиотеки кДНК акклимированных к холоду крон тетраллоидной люцерны (М sativa cv Apica; Laberge, S, Castongway, Y. and  L.-P. 1993. Plant Physiol 101: 1411-1412; Castongway, Y., S.Laberge, P.Nadeau and L.-
L.-P. 1993. Plant Physiol 101: 1411-1412; Castongway, Y., S.Laberge, P.Nadeau and L.- 1997, p. 175-202. In B.D.McKersie and D.W.Brown (ed.). Biotechnology and the improvement of forage legumes. CAB International, Wallingford, UK), использованы в этом исследовании:
1997, p. 175-202. In B.D.McKersie and D.W.Brown (ed.). Biotechnology and the improvement of forage legumes. CAB International, Wallingford, UK), использованы в этом исследовании:
L.-P. 1993. Plant Physiol 101: 1411-1412.
** Monroy,A.F., Castongway, Y., Laberge, S., Sarhan, F., L.-P. and Dhindsa, R.S. 1993. Plant Physiol 102, 873-879.
*** Castongway, Y., Laberge, S., Nadeau, P. and L.-P. 1994. Plant Mol. Biol. 24, 799-804.
Выделение РНК
Примерно, 0,5 г (вес сырой ткани) листовой ткани каждого акклимированного к холоду генотипа пестиком в ступке измельчают в тонкий порошок в жидком N2 и экстрагируют обычными методами, как описано ранее (Castongway et al. 1994. Plant Mol. Biol. 24, 799-804). Тотальную РНК определяют количественно УФ-спектроскопией при 260 нм (Foumey, R.M., J.Miyakoshi, R.S.Day III and M.C.Paterson. 1988. Focus 10:1).
Количественный дот-блот анализ
Для каждого родительского генотипа и каждого генотипа сегрегированной популяции пять мкг тотальной РНК переносят в вакууме на мембрану из найлона (Hybond N+, Amersham Pharmacia Biotech, Oakvill, ON), используя устройство Bio-Dot™ (Bio-Rad, Mississauga, ON) и иммобилизуют, сшивая под действием УФ в течение 3 минут. Мембраны гибридизуют в течение ночи при 68°С в среде 2Х SSC, 0,25% (вес/объем) нежирного порошкового молока, 1% SDS с меченым [32P]-dCTP зондом, приготовленным из очищенных инсертов регулируемых холодовой реакцией генов в соответствии со стандартными протоколами (Sambrook, J., Maniatis, T. and Fritsch. 1989. Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York). Мембраны экспонируют с рентгеновской пленкой Kodak X-OmatAR5 X-ray при -80°С и уровень транскриптов количественно определяют денситометрическим анализом радиоавтографов, используя программное обеспечение OneD Scan™ (Scanalytics inc., Billerica, MA). См. на Фигуре 3 пример одного из таких дот-блотов, полученных при использовании в качестве зонда гена msaCIA. Средние уровни транскриптов всех образцов на каждой мембране используют для стандартизации гибридизационного сигнала через мембраны.
Карта сцепления
Генетическую карту сцепления строят на основе AFLP и RFLP. ДНК выделяют из 1-2 г ткани свежего листа 4-6 недельных клональных побегов. Применяют метод выделения, описанный Doyle and Doyle (Doyle, J.J., and Doyle, J.L. 1990. Isolation of plant DNA from fresh tissue. FOCUS 12, 13-15), за исключением того, что измельчают в отсутствие жидкого азота при комнатной температуре.
Для того чтобы обнаружить маркеры RFLP, ДНК, очищенную от 169 F2 генотипов, по отдельности расщепляют рестриктазами DraI и EcoRV. Каждый образец гидролизованной ДНК (6 мкг на генотип выделяют на горизонтальных гелях (0,9% агароза, 1 ХТАЕ) и разделенные фрагменты переносят на найлоновые мембраны (Hybond N+, Amersham Phannacia Biotech, Oakvill, ON) "капиллярным блоттингом". Для анализа одновременного разделения с множеством зондов готовят десять двойных копий (дубликатов) мембран. Мембраны гибридизуют в течение ночи при 65°С с зондами, мечеными [32P]-dCTP. Зонды ДНК состоят из случайно выбираемых кДНК и кДНК, отбираемых в соответствии с их гомологией регуляторным генам, как полученным с использованием библиотек на основе кДНК Medicago sativa, так и 7 msaCI кДНК, описанных выше. Инсерты кДНК сначала специфически амплифицируют с помощью PCR и метят методом случайных шестичленников в качестве затравки, используя набор QuickPrime™ Kit (Amersham Pharmacia Biotech, Oakvill, ON). Предгибридизацию, гибридизацию и мечение зонда осуществляют в соответствии со стандартными протоколами (Sambrook, J., Maniatis, Т. and Fritsch. 1989. Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York).
Для идентификации маркеров AFLP F2 ДНК расщепляют с помощью рестриктаз EcoRI и MseI, лигируют к адаптерам Есо- и Mse-специфичных праймеров, затем амплифицируют при участии предварительно отобранных Есо- и Mse PCR праймеров (Vos. P., Hogers, R., Bleeker, M., Reijans, M., van der Lee, Т., Homes, M., Frijters, A., Pelerman, J., Kuiper, M., and Zabeau, M. 1995, Nucleic Acids Res. 23, 4407-4414). Предварительно отобранные ампликоны разбавляют в 20 раз для того, чтобы использовать в качестве материала для селективной амплификации. Как Есо, так и Mse праймеры, применяемые для селективной амплификации, содержат 3 селективных нуклеотида на 3' конце. Праймер Есо метят красителями IRD, которые позволяют обнаруживать ампликоны с помощью инфракрасных лазеров автоматического секвенатора LI-COR (LI-COR, Inc. Lincoln, Neb, USA). Селективную амплификацию проводят согласно протоколу "touch down", описанному Vos et al. (1995, Nucleic Acids Res. 23, 4407-4414).
После селективной амплификации ампликоны разделяют на 25 см гелях, состоящих из 7% акриламида Long Ranger, 7M мочевины и 0,6 Х ТВЕ на автоматическом секвенаторе ДНК LI-COR модели 4000L.
Генетическую карту диплоидного скрещивания создают, используя данные сегрегации, полученные с помощью сегрегирующих маркеров 112 RFLP и 117AFLP. Косегрегационные анализы осуществляют с помощью программного обеспечения MAPMAKER/EXP v 3.0 (Lander,E.S., Green, P., Albertson, J., Barlow, A., Daly, M.J., Lincoln, S.E. and Newburg, L 1987. Genomics 1, 174-181), чтобы осуществить отнесение групп сцепления каждого маркера и локализацию каждого молекулярного маркера в каждой группе сцепления. Отдельные группы сцепления получают с применением анализа по двум факторам с минимальным числом LOD 15 и максимальным уровнем рекомбинации 30%. Затем для того, чтобы расположить локусы в группах сцепления, применяют многомерный анализ с пределом LOD 2.0. Полученная в результате карта (Фигура 1) покрывает, примерно, 1400сМ со средним маркерным интервалом около 8 сМ. Пятнадцать маркеров остаются несвязанными.
Идентификация QTL, ассоциируемых с экспрессией генов msaCI и морозостойкостью
QTL, участвующие в экспрессии генов msaCI, идентифицируют с использованием MAPMAKER/QTL v 1.1. (Lander, E.S., and Boststein, D. 1989, Genetics 121, 185-199). Карту сцепления сканируют на присутствие QTL в специфических областях генома, используя в качестве количественного признака экспрессию каждого гена. Определяют действие каждого QTL по отношению друг к другу и процент дисперсии экспрессии генов msaCI, объясняемый каждым QTL. Для идентификации QTL берут величину предела LOD 2.0.
Анализ регулируемой холодовой реакцией аккумуляции транскриптов для каждого из семи генов msaCI выявляет генотипическую вариабельность их экспрессии с разницей экспрессии между генотипами в некоторых случаях в 10-40 раз (например, см. Фигуру 3).
QTL, связанные с вариабельностью уровней транскриптов генов msaCI, наблюдаемой среди генотипов, обнаружены для всех 7 генов msaCI и для фенотипа LT50 (Таблица 1). Гены, которые играют важную регуляторную роль в экспрессии генов msaCI, обнаружены в этих QTL. Число QTL, ассоциируемых с экспрессией генов msaCI, варьируется от 1 (msaCIF) до 5 (msaCID). В большинстве случаев их LOD весьма значительны. Фенотипическое изменение, объясняемое этими QTL, варьируется от 10% до 60%. В большинстве случаев за увеличение экспрессии и морозоустойчивость, соответственно, ответственен аллель холодоустойчивого родителя M.falcata.
Идентифицированы три QTL, относящиеся к морозоустойчивости (LT50). Два из этих QTL локализованы в группе сцепления Е в геномной области, ассоциируемой также с экспрессией msaCIB и msaCID.
Обнаружены QTL, относящиеся к экспрессии генов msaCI, ассоциируемые с положением соответствующего структурного гена для msaCIA (см. группу сцепления D, Фигура 2), msaCID (см. группу сцепления Е, Фигура 2) и msaCIG (группа сцепления F, Фигура 2).
В случае msaCID локус RFLP совпадает с QTL, связываемым с экспрессией трех генов: самого гена msaCID, msaCIA и msaCIB (см. группу сцепления Е, Фигура 2). Это наводит на мысль, что либо msaCID является регуляторным геном, либо регуляторный ген локализован в его близости.
QTL для экспрессии в тканях кроны и листьев, которые локализованы в той же самой геномной области, наблюдаются для msaCIA (между 120-130 см. Фигура 2, группа сцепления Е) и msaCID (см. группу сцепления Е, Фигура 2). Иначе локусы, влияющие на ткани каждого типа, были бы различными. QTL, контролирующие экспрессию нескольких генов, локализованы в группах сцепления В и Е, что предполагает общий регуляторный механизм. Особенно поразительный пример представляет собой QTL в группе сцепления В, который оказывает влияние на экспрессию пяти генов msaCI в листовой ткани.


Пример 2: Выделение регуляторных генов и регуляторных последовательностей, ассоциированных с представляющими интерес QTL
Общее описание экспериментального метода
Обнаружение QTL, которые объясняют значительную долю изменяемости экспрессии генов msaCI, указывает на возможную локализацию регуляторных генов в этих локусах. Картирован ряд меток экспрессируемых последовательностей (EST) кодирующих генов акклимированной к холоду люцерны с известными функциями в регуляции генной экспрессии. Возможные регуляторные гены, локализованные в QTL, локализованных в QTL, ассоциированных с экспрессией генов msaCI, выделены и охарактеризованы, и изучена их экспрессия в трансгенной люцерне с целью оценить их действие на регулируемую холодовой реакцией генную экспрессию.
Описание возможных регуляторных генов
Ряд EST, проявляющих гомологию с генами с известньми регуляторными функциями, выделены из библиотеки λgt10 кДНК, созданной на основе мРНК, выделенной из крон тетраплоидной люцерны (М.sativa L. cv. Apica; R.Michaud, С.Richard, С.Willemot and H.Gasser 1983. Can. J. Plant Sci. 547-549):
Факторы транскрипции (цинковые пальцы и белки АР2)
Митоген-активированные протеинкиназы (ММК4, ММК3, ММК2, ММК1)
Кальций-зависимые протеинкиназы
Серин-треонинпротеинкиназа
Са2+транспортирующая АТФ-аза
GTP-связывающие протеиназы
РНК-связывающие протеиназы
Протеинфосфатазы (типа 2А и 2С)
Кальмодулин
14-3-3 белки
GTP-активирующие протеиназы
Белок аденилатциклазы
Фосфолипаза С
Липоксигеназа
Са2+-связывающие белки
Гистондеацетилаза
Рецепторная киназа
Фосфатидилинозит-3-киназа
Генетическое картирование возможных регуляторных генов
Зонды специфичных регуляторных генов, перечисленных в Таблице 2, получают PCR амплификацией, очищают на колонках Sephacril S-200 MicroSpin (Amersham Pharmacia Biotech, Oakvill, ON) и хранят при -20°С. Эти зонды применяют для Саузерн-гибридизации с рестриктированной ДНК F2 диплоидных генотипов потомства от скрещивания, описанного в Примере 1. Очищенные зонды метят радиоактивной меткой до высокой степени специфической активности методом использования случайных шестичленников в качестве затравки и гибридизуют с ДНК-блотами в соответствии со стандартными протоколами для RFLP, как описано в Примере 1 (Sambrook, J., Maniatis, T. and Fritsch. 1989. Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York). Регуляторные гены, являющиеся полиморфными и картированные, наряду с их функциями, предполагаемыми исходя из их гомологии с последовательностями в базе данных GENBANK/EMBL, представлены в Таблице 2.


Последовательности EST также применяются для создания праймеров для амплификации идентифицированных локусов в качестве STS (ДНК-маркирующих сайтов) ДНК сегрегирующей популяции. Если образующиеся являются мономорфными среди отдельных представителей популяции, PCR-продукты разрезают различными ферментами для обнаружения полиморфизма. Если полиморфизм не обнаруживают, продукты PCR, полученные от двух родителей, по отдельности секвенируют и ищут SNP (полиморфизмы одиночных нуклеотидов).
Предполагаемые регуляторные гены, описанные выше, картированы как RFLP в потомстве F2, как описано в Примере 1, чтобы оценить, локализованы ли они в QTL, ассоциируемых с изменением экспрессии генов msaCI, индуцируемой холодовой реакцией, как идентифицировано в Примере 1.
Все гены, перечисленные в Таблице 1 и выделенные в Таблице 2, локализованы в геномных областях, содержащих QTL, и, следовательно, они являются предполагаемыми регуляторными генами для экспрессии генов msaCI, для признака LT50 или их комбинации. Два из этих генов, С4494 и С2186, проявляют гомологию с геном CBF1 Arabidopsis и с геном SCOF-1 сои, соответственно, для которых продемонстрирована причастность к холодостойкости (Jaglo-Ottosen, K.R., Gilmour, S.J., Zarka, D.G., Schabenberger, О., and Thomashow, M.F. 1998 Arabidopsis CBF1 overexpression induces COR genes and freezing tolerance. Science 280: 104-106. Kim, J.C., Lee, S.H., Chsong, Y.H, Yoo, C.-M, Lee, S.I., Chun, H.J., Hong, J.C., Lee, S.Y., Lim, C.O., and Cho, M.J., 2001. A novel cold-inducible zinc-finger protein from soybean, SCOF-1, enhances cold tolerance in transgenic plants. The plant Journal 25: 247-257). C4494 (гомолог CBF1) локализован в QTL, ассоциируемых с экспрессией msaCIB в листьях (группа сцепления F, Фигура 2), в то время как С2186 (гомолог SCOF-1) отнесен на карте к области, содержащей QTL для LT50 (группа сцепления С, Фигура 2). Эти гены кодируют факторы транскрипции, индуцируемые холодовой реакцией, которые, как было недвусмысленно показано, вызывают конститутивную экспрессию генов COR (msaCI) в трансгенных растениях и повышают морозостойкость без акклимации.
Результаты также указывают на ряд интересных новых возможных генов, причастных к экспрессии под действием холодовой реакции и к экспрессии генов msaCI (Таблица 1, Фигура 2). Например, клон С2784, проявляющий гомологию с серин/треонинпротеинкиназой, картирован на группе сцепления В, где обнаружены QTL экспрессии msaCIA, msaCID, msaCIE и msaCIG в листьях. В клетках люцерны осмотический стресс приводит к быстрой активации двух протеинкиназ. Одна из этих киназ, по-видимому, представляет собой гомолог ASK1 серин/треонинкиназы 1 Arabidopsis (Munnik, T. and H.J.G. Meijer. 2001. FEBS Letters 498: 172-178). Подтверждение роли таких предполагаемых регуляторных генов может быть получено с применением описанной ниже методологии.
Секвенирование геномных клонов возможных регуляторных генов и возможных регуляторных последовательностей
EST предполагаемых регуляторных генов, которые картированы на одном или более QTL, ассоциируемых с изменением экспрессии генов msaCI, выделены при использовании геномной библиотеки, созданной на основе фага EMBL3 (Sambrook, J., Maniatis, T. and Fritsch. 1989. Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York) с применением ДНК двух родителей, идентифицированных выше. Каждый возможный регуляторный ген секвенируют во всей полноте, включая участки upstream и downstream кодирующей области методом терминации дидезоксинуклеотидной цепи (Sambrook, J., Maniatis, T. and Fritsch. 1989. Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York). Проведено сравнение аллельных форм каждого родителя для идентификации мутаций, отвечающих за QTL. Подтверждение локализации возможных регуляторных генов (кандидатов) проводят картированием upstream или downstream участков каждого регуляторного гена на карте сцепления, полученной выше. Только регуляторные гены, которые характеризуются наличием 5' или 3' областей, совпадающих на карте (co-mapping) с теми же самыми одним или более QTL, используются для дальнейшей характеристики в исследованиях коплементации или "нокаутирования" гена.
Изменяемая экспрессия предполагаемых генов (кандидатов) в трансгенных растениях (исследование комплементации и "нокаутирования" гена)
Трансформацию люцерны осуществляют с использованием Agrobacterium tumefaciens, как ранее описано  et al. ( R., S.Laberge, G.Allard, H.Khoudi, Y.Casonguay, J.Lapointe, R.Michaud and L.-
et al. ( R., S.Laberge, G.Allard, H.Khoudi, Y.Casonguay, J.Lapointe, R.Michaud and L.-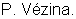 1994. Plant Cell Tissue Organ Cult 42: 129-140). Применяются два способа: 1) Конститутивная экспрессия предполагаемого гена (кандидата) с использованием слияния гена с конститутивным промотором, например, но без ограничения, с 35S; и индуцибельная экспрессия. Кодирующую область предполагаемого гена сливают в рамке считывания с полноразмерньм промотором 35S и полученную конструкцию клонируют в двойной вектор экспрессии pGA482. Полноразмерные предполагаемые регуляторные гены (кандидаты) (т.е. предполагаемые регуляторные гены, которые, с прилегающими 5' и 3' участками, картированы к одному или более QTL, идентифицированных выше) наряду, примерно, с 1 тыс. п.о. 5' и 3' участков некодирующих областей, клонируют в двойной вектор экспрессии pGA482. Конструкции, полученные в результате как конститутивной, так и индуцибельной экспрессии, в смысловой ориентации используют для трансформации растений, у которых отсутствует аллель в QTL, соответствующем экспрессии интересующего признака (например, но без ограничения, экспрессии гена msaCI), и конструкции в антисмысловой ориентации используют для трансформации растений, которые "укрывают" аллель в QTL. Затем выращивают трансгенные растения и подвергают их холодовой акклимации, как требуется для определения влияния введенного смыслового или антисмыслового гена как на экспрессию одного или более генов msaCI, так и на LT50, тем самым ген-кандидат оценивается как регуляторный фактор, контролирующий экспрессию мРНК генов msaCI в QTL.
1994. Plant Cell Tissue Organ Cult 42: 129-140). Применяются два способа: 1) Конститутивная экспрессия предполагаемого гена (кандидата) с использованием слияния гена с конститутивным промотором, например, но без ограничения, с 35S; и индуцибельная экспрессия. Кодирующую область предполагаемого гена сливают в рамке считывания с полноразмерньм промотором 35S и полученную конструкцию клонируют в двойной вектор экспрессии pGA482. Полноразмерные предполагаемые регуляторные гены (кандидаты) (т.е. предполагаемые регуляторные гены, которые, с прилегающими 5' и 3' участками, картированы к одному или более QTL, идентифицированных выше) наряду, примерно, с 1 тыс. п.о. 5' и 3' участков некодирующих областей, клонируют в двойной вектор экспрессии pGA482. Конструкции, полученные в результате как конститутивной, так и индуцибельной экспрессии, в смысловой ориентации используют для трансформации растений, у которых отсутствует аллель в QTL, соответствующем экспрессии интересующего признака (например, но без ограничения, экспрессии гена msaCI), и конструкции в антисмысловой ориентации используют для трансформации растений, которые "укрывают" аллель в QTL. Затем выращивают трансгенные растения и подвергают их холодовой акклимации, как требуется для определения влияния введенного смыслового или антисмыслового гена как на экспрессию одного или более генов msaCI, так и на LT50, тем самым ген-кандидат оценивается как регуляторный фактор, контролирующий экспрессию мРНК генов msaCI в QTL.
Все цитированные источники вводятся в данное описание в качестве ссылок.
Настоящее изобретение описано с привлечением предпочтительных вариантов изобретения. Однако, и это очевидно для специалистов в данной области техники, что можно осуществить много изменений и модификаций, не отступая от объема изобретения, описанного выше.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОТБОРА ЖИВОТНЫХ ПО ПРИЗНАКАМ, НАСЛЕДУЕМЫМ ПО МЕХАНИЗМУ РОДИТЕЛЬСКОГО ИМПРИНТИНГА | 1999 |
|
RU2262229C2 |
| МАРКЕРЫ, СЦЕПЛЕННЫЕ С УСТОЙЧИВОСТЬЮ РАСТЕНИЙ СОИ К SCN | 2011 |
|
RU2620973C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ НИЗКОГО СОДЕРЖАНИЯ ПАЛЬМИТИНОВОЙ КИСЛОТЫ В ПОДСОЛНЕЧНИКЕ (HELIANTHUS ANNUS) И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2670517C2 |
| ГЕНЕТИЧЕСКИЕ ЛОКУСЫ, СВЯЗАННЫЕ С УСТОЙЧИВОСТЬЮ К ПЫЛЬНОЙ ГОЛОВНЕ У МАИСА | 2009 |
|
RU2562864C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM2 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2717017C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С УСТОЙЧИВОСТЬЮ ПОДСОЛНЕЧНИКА К OROBANCHE | 2018 |
|
RU2776361C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM4 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2718584C2 |
| ГЕНЕТИЧЕСКИЙ ЛОКУС, АССОЦИИРОВАННЫЙ С ГНИЛЬЮ КОРНЯ И СТЕБЛЯ, ОБУСЛОВЛЕННОЙ PHYTOPHTHORA, У СОИ | 2016 |
|
RU2748688C2 |
| РАСТЕНИЕ, УСТОЙЧИВОЕ К НАСЕКОМЫМ | 2008 |
|
RU2522477C2 |
| РАСТЕНИЯ ТОМАТОВ, ИМЕЮЩИЕ БОЛЕЕ ВЫСОКИЕ УРОВНИ УСТОЙЧИВОСТИ К BOTRYTIS | 2005 |
|
RU2403709C2 |




Изобретение относится к генной инженерии и предназначено для идентификации одной или более областей в геноме организма. Благодаря построению карты сцепления для сегрегированного потомства скрещенных организмов можно идентифицировать один или более локусов количественных признаков. 5 н. и з.п. ф-лы, 3 ил., 2 табл.
i) идентификацию первого представляющего интерес организма, характеризующегося тем, что этот указанный первый интересующий организм проявляет измеримую реакцию на внешний раздражитель или иным образом проявляет фенотип, связанный с дифференциальной генной экспрессией, ассоциированной с интересующим процессом;
ii) идентификацию второго интересующего организма, характеризующегося тем, что указанный второй интересующий организм:
а) не проявляет никакой реакции на раздражитель или проявляет менее сильную реакцию на указанный раздражитель, чем первый указанный интересующий организм;
б) проявляет фенотип, отличный от фенотипа указанного первого интересующего организма, причем указанный отличный фенотип ассоциирован с указанным интересующим процессом;
в) проявляет фенотип, представляющий интерес, который сегрегирует в популяции, полученной в результате скрещивания с указанным первым интересующим организмом;
г) или комбинация двух или более из а), б), в);
iii) скрещивание указанных первого и второго интересующих организмов с целью получения популяции сегрегированного потомства;
iv) выделение РНК каждого указанного сегрегированного потомства и количественное определение уровня генной экспрессии одного или более генов, представляющих интерес, причем указанные один или более интересующие гены связаны с реакцией на внешний раздражитель или с указанным интересующим процессом;
v) получение карты сцепления указанного сегрегированного потомства с использованием одного или более маркеров;
vi) определение соотношения между указанными одним или более маркерами на указанной карте сцепления и указанной генной экспрессией указанных одного или более интересующих генов и идентификация одного или более локусов количественных признаков (QTL);
vii) подтверждение того, что один или более генов локализованы в одном или более QTL.
биотический стресс;
абиотический стресс;
изменение минерального питания;
изменение световых характеристик и уровня освещенности;
реакцию, вызываемую добавлением одного или более экзогенных химических веществ;
любой стресс, который вызывает изменение уровня эндогенных химических соединений в указанном растении;
эволюционный, химический или экологический контроль генной экспрессии во время эмбриогенеза;
органогенез;
старение;
созревание;
диапаузу;
прорастание;
циркадный, ультрадный и другие внутренние ритмы;
собираемый урожай;
завязывание семян;
транслокацию фотосинтата;
изменение емкости источника и стока в указанном растении;
индекс листовой поверхности;
изменение соотношения массы подземных и наземных органов растения;
изменение питательной ценности собранного материала;
изменение морфологии растения;
изменение скорости клеточного цикла;
изменение скорости клеточной дифференцировки;
изменение размера клеток;
изменение жизненного цикла растения и
геномные перестройки.
i) определение одного или более локусов количественных признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес;
ii) картирование одного или более регуляторных генов;
iii) определение, картированы ли один или более регуляторных генов с указанными одним или более QTL;
iv) выделение одного или более регуляторных генов в указанных одном или более QTL и
v) секвенирование одного или более регуляторных генов, выделенных на стадии iv).
i) определение одного или более локусов количественных признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес, и
ii) выделение одного или более регуляторных генов в указанных одном или более QTL;
iii) секвенирование регуляторного гена.
i) идентификацию одного или более локусов количественных признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес, и
ii) картирование одного или более регуляторных генов;
iii) определение, картированы ли один или более регуляторных генов с указанными одним или более QTL;
iv) выделение одного или более регуляторных генов в указанных одном или более QTL и
v) секвенирование одного или более регуляторных генов, включающих регуляторные последовательности указанных одного или более регуляторных генов, выделенных на стадии iv).
i) идентификацию одного или более локусов количественных признаков (QTL), которые объясняют значительную долю изменяемости экспрессии одного или более генов, представляющих интерес, и
ii) выделение одного или более регуляторных генов в указанных одном или более QTL;
iii) секвенирование регуляторных генов, включающих регуляторные последовательности.
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| WO 00/18963 А, 06.04.2000 | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| PLANT PHYSIOLOGY, v.108, № 3, 1995, р.1233-1239. | |||

