1. Область техники, к которой относится изобретение
Изобретение касается способов секвенирования или анализа нуклеиновых кислот, а более конкретно - способов анализа триплексных и дуплексных комплексов, полученных в результате гибридизации нуклеиновых кислот, с использованием измерений интенсивности флуоресценции.
2. Уровень техники
Флуоресцентные красители используют для обнаружения и количественного анализа нуклеиновых кислот уже в течение десятилетий. В своих наиболее общих формах анализы, основанные на определении интенсивности флуоресценции, как правило, включают осуществление контакта мишени с содержащим флуорофор зондом, полное удаление несвязанного зонда из связанного зонда и обнаружение флуоресценции в промытом образце. Гомогенный анализ имеет преимущество перед такими основными методиками анализов в том, что он не требует стадии промывки или обеспечения нежидкой подложки.
Например, в патентах США №5538848 (Livak et al.) и 4220450 (Maggio) описываются гомогенные основанные на флуоресценции анализы нуклеотидных последовательностей с использованием олигонуклеотидных зондов в растворе. Однако эти патенты требуют использования гасящей добавки в комбинации с репортинговым (reporting) агентом, для того, чтобы можно было различить сигналы, генерируемые гибридизированными зондами и негибридизированными зондами. Способ, описываемый Livak et al. требует также использования ферментов. Гасящие добавки и ферменты усложняют и удорожают эти способы.
В патенте США №5332659 (Kidwell) описывается способ обнаружения нуклеотидных последовательностей в растворе с помощью зондов, включающих, по меньшей мере, две флуорофорные группы. Флуорофоры следует выбирать так, чтобы они могли, находясь достаточно близко друг к другу, осуществлять электронное взаимодействие друг с другом, в результате чего изменяется длина волны, в зависимости от их спектров. Негибридизированные зонды гораздо более гибкие, чем зонды, гибридизированные с последовательностью мишени, и поэтому гораздо более вероятно, что две флуорофорных группы на каждом зонде будут расположены близко друг к другу в случае, если зонд не гибридизирован, чем в том случае, когда зонд гибридизирован. Таким образом, изменение в длинах испускаемых волн, коррелирующее со свободным зондом, можно отслеживать, и оно является показателем количества свободного зонда в образце.
В патенте США №5846729 (Wu et al.) также описываются гомогенные основанные на флуоресценции анализы для гибридизации нуклеиновых кислот.
В некоторых анализах используют интеркалирующие (встраивающиеся) флуорофоры для обнаружения гибридизации нуклеиновых кислот, что основано на способности таких флуорофоров встраиваться между цепями нуклеиновой кислоты в гибридизационном комплексе.
Например, патент США №5824557 (Burke et al.) описывает способ и набор для обнаружения и подсчета молекул нуклеиновой кислоты. Предпочтительный вариант осуществления этого способа основан на встраивании красителя в спираль двухцепочечной нуклеиновой кислоты или одноцепочечной нуклеиновой кислоты. После встраивания краситель флуоресцирует, и интенсивность флуоресценции является прямым показателем количества нуклеиновой кислоты, присутствующего в образце. Хотя способ Burke et al. претендует на полезность для измерения количества нуклеиновой кислоты в образце, но неспецифичное связывание между интеркалятором и нуклеиновой кислотой, на котором основан этот способ, делает его непригодным для определения специфичного связывания, особенно в условиях, когда присутствуют дуплексы нуклеиновой кислоты, не являющейся мишенью.
В патенте США №5814447 (Ishiguro et al.) описывается способ, претендующий на преимущество перед анализами, основанными на неспецифичном взаимодействии между интеркалирующими агентами и дуплексами нуклеиновых кислот, такими, как описываемый Burke et al., а также более ранним анализом, описанным Ishiguro et al. в японском патенте №237000/1993. Эти более ранние разработки включали добавку интеркалирующего флуорохрома, обладающего тенденцией к проявлению повышенной интенсивности флуоресценции после встраивания его в образец в растворе перед тем, как специфичная область нуклеиновой кислоты-мишени была амплифицирована с помощью ПЦР, и измерение интенсивности флуоресценции реакционного раствора в заданные интервалы времени для обнаружения и количественного определения нуклеиновой кислоты-мишени перед амплификацией. В патенте №`447 сделана попытка улучшить более ранние разработки путем обеспечения анализа, имеющего повышенную специфичность и отличающегося тем, что зонд представляет собой одноцепочечный олигонуклеотид, меченый интеркалирующим флуорохромом, который подлежит встраиванию в часть комплементарного связывания между нуклеиновой кислотой-мишенью и зондом, представляющим собой одноцепочечный олигонуклеотид.
Кроме вышеупомянутых разработок, с помощью которых определяют интенсивность флуоресценции, в некоторых разработках предлагается использовать преимущества анализов флуоресцентной поляризации. Однако анализы, основанные на поляризации, имеют существенные недостатки. Степень изменения поляризации в зависимости от связывания может быть непредсказуемой, а интерпретация данных, не согласующихся с теоретически ожидаемыми, может потребовать больше усилий, чем это желательно для аналитического метода, особенно в том случае, если этот метод должен быть автоматизирован.
Несмотря на вышеупомянутые разработки, в данной области техники продолжает существовать необходимость в простом, высокочувствительном, эффективном и быстром способе анализа взаимодействия между нуклеиновыми кислотами и/или аналогами нуклеиновых кислот.
Все цитированные здесь публикации включены в настоящее описание в качестве ссылок во всей их полноте.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение касается способа анализа связывания, включающего:
обеспечение мишени, включающей, по меньшей мере, одну последовательность нуклеиновой кислоты;
обеспечение зонда, включающего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты, не полностью комплементарную, по меньшей мере, части мишени;
обеспечение интеркалирующего агента, причем любой из указанных зондов или интеркалирующего агента включает флуорофор;
добавление зонда, мишени и интеркалирующего агента к гибридизационной среде для получения испытуемого образца;
облучение испытуемого образца возбуждающим излучением, чтобы заставить флуорофор испускать флуоресцентное излучение;
определение интенсивности флуоресцентного излучения, причем эта интенсивность является прямым показателем аффинности связывания между зондом и мишенью;
определение на основании указанной интенсивности степени ошибочного спаривания между зондом и мишенью.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Изобретение далее описывается в соответствии с нижеследующими чертежами, в которых одинаковые номера ссылок обозначают одинаковые элементы, и в которых фигуры 1А, 1В, 2, ЗА, 3В, 4, 5, 6А, 6В, 7А, 7В, 8А и 8В представляют собой сводные графики максимальных показателей интенсивности флуоресценции в зависимости от температуры для каждого проанализированного образца.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Изобретение касается быстрого, чувствительного, безвредного для окружающей среды и безопасного способа анализа связывания между мишенью и зондом, в котором мишень включает последовательность нуклеиновой кислоты или последовательность аналога нуклеиновой кислоты и зонд включает последовательность нуклеиновой кислоты или последовательность аналога нуклеиновой кислоты.
В отличие от некоторых анализов по прототипу настоящее изобретение позволяет не только обнаружить наличие гибридизации, но также обеспечивает получение качественной и количественной информации, касающейся природы гибридизации между зондом и мишенью. Таким образом, изобретение позволяет практическому работнику отличить друг от друга правильное спаривание, ошибочное спаривание одной пары оснований, ошибочное спаривание двух пар оснований, ошибочное спаривание трех пар оснований, делецию одной пары оснований, делецию двух пар оснований и делецию трех пар оснований.
Варианты осуществления изобретения включают калибровку интенсивности флуоресценции, измеренной для смеси первого зонда и мишени, по сравнению с показателями интенсивности флуоресценции, полученными для других зондов, объединенных с той же самой мишенью и тем же самым интеркалирующим агентом, причем каждый из других зондов отличается от первого зонда, по меньшей мере, одним основанием. Интенсивность флуоресценции, определяемая в способе по настоящему изобретению, увеличивается с увеличением аффинности связывания между зондом и мишенью. Настоящий способ не требует измерения поляризации флуоресценции в отличие от способов флуоресцентной анизотропии.
Можно построить калибровочную кривую, в которой интенсивность является функцией аффинности связывания между мишенью и зондом. Поскольку аффинность связывания между мишенью и множеством различных зондов варьирует в зависимости от количества ошибочно спаренных оснований, то природа ошибочного спаривания (A-G против А-С против T-G против Т-С и т.д.), места расположения ошибочного спаривания (спариваний) в гибридизационном комплексе и т.п., то анализ по настоящему изобретению можно использовать для секвенирования мишени.
Изобретение позволяет количественно определить аффинность связывания между зондом и мишенью. Такая информация может быть ценной для ряда применений, включая конструирование антисмысловых лекарств с оптимизированными характеристиками связывания.
В отличие от способов в уровне техники анализ по настоящему изобретению предпочтительно является гомогенным. Этот анализ можно проводить без отделения комплекса зонд-мишень от свободных зонда и мишени перед определением интенсивности флуоресценции. Анализ не требует стадии разделения в геле, что позволяет значительно повысить производительность тестирования. Количественные анализы просты и точны. Кроме того, анализ предпочтительно проводят в гомогенном растворе, что исключает требование отделять связанные комплексы от несвязанных зондов либо путем фильтрации с многочисленными стадиями промывки, либо путем гель-электрофореза. Вследствие этого настоящий анализ связывания экономит массу времени и средств и легко поддается автоматизации. Кроме того, он позволяет быстро определять переменные связывания, такие как буфер, рН, ионная концентрация, температура, время инкубации, относительные концентрации последовательностей зонда и мишени, концентрация интеркалятора, длина последовательностей мишени, длина последовательностей зонда и требования к возможным кофакторам.
В следующих предпочтительных воплощениях способ анализа по настоящему изобретению может проводиться в растворе в лунке, на непроницаемой поверхности или на микрочипе. Использование твердых подложек, таких как нитроцеллюлозные или нейлоновые мембраны (см., к примеру, Sambrook J., Fritsch E.E., and Maniatis Т. (1989) Molecular Cloning, A Laboratory Manual., 2nd Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York)), кварцевые пластины, стеклянные пластины, покрытое полиакриламидом стекла, полипропилен, полистирол, силикон и т.д. без ограничения, и собственно биочипов (к примеру, см. обзор L.J.Kricka, S. Surrey, P. Fortina. Multispot Array-Technologies (2001) Encyclopedia of Life Sciences, 2001 Macmillan Publisers Ltd., Nature Publishing Group) является ставшим уже рутинным подходом при проведении гибридизации. Специалисту в данной области понятно, что интеграция способа анализа гибридизации настоящего изобретения с форматом, например, микротитровальных планшетов или микрочипов (биочипов) может быть легко осуществлена и будет открывать широкие возможности для автоматизации/роботизации заявленного способа и применения его в качестве способа высокопроизводительного скрининга.
Кроме того, анализ по настоящему изобретению предпочтительно проводят без использования сигнальной гасящей добавки на мишени или на зонде.
Предпочтительные варианты осуществления изобретения специфически обнаруживают триплексную гибридизацию между зондом и двухцепочечной мишенью, и таким образом исключается необходимость в денатурации мишени. Хотя известно, что ДНК-зонды образуют триплексы с определенными классами мишеней (см., например, Egholm et al., 365 Nature 566 (1993) и Tomac et al., 118 J. Am. Chem. Soc. 5544 (1996)), авторы настоящего изобретения обнаружили, что они могут специфично анализировать триплексы, образованные между зондами, представляющими собой одноцепочечную нуклеиновую кислоту (например, одноцепочечную ДНК - ssDNA - или РНК) и мишенями, представляющими собой двухцепочечную нуклеиновую кислоту (например, двухцепочечную ДНК - dsDNA).
Подходящие для использования по настоящему изобретению зонды включают, например, ssDNA, РНК и PNA и другие аналоги нуклеиновых кислот, имеющие незаряженные остовы. Предпочтительными являются последовательности зондов, имеющие любую длину от 8 до 20 оснований, поскольку это интервал, внутри которого находятся самые мелкие единственные в своем роде последовательности ДНК прокариотов и эукариотов. Зонды длиной от 12 до 18 оснований особенно предпочтительны, поскольку это длина самых мелких единственных в своем роде последовательностей генома человека. В вариантах осуществления изобретения предпочтительны зонды, имеющие от 6 до 30 оснований, причем наиболее предпочтительны зонды, имеющие 15 оснований. Однако можно использовать множество более коротких зондов для обнаружения нуклеотидных последовательностей, заключающих в себе множество не являющихся единственными в своем роде последовательностей мишеней, которые объединены таким образом, что позволяют идентифицировать единственную в своем роде нуклеотидную последовательность.
Изобретение не требует использования радиоактивных зондов, которые являются опасными, требующими длительного времени, сложными в применении и требуют постоянной регенерации. Зонды по настоящему изобретению предпочтительно безопасны в применении и стабильны в течение нескольких лет. Поэтому зонды можно изготавливать или заказывать в больших количествах и хранить.
Авторы настоящего изобретения также обнаружили, что зонды, представляющие собой параллельную PNA, ведут себя лучше, чем обычно синтезируемые зонды, представляющие собой антипараллельную PNA. Когда последовательности нуклеиновых кислот мишени были более чем в три раза длиннее, чем последовательности PNA-зондов, то зонды, представляющие собой параллельную и антипараллельную PNA, были одинаково эффективны для того, чтобы отличать правильное спаренные гибридизационные комплексы нуклеиновых кислот от комплексов, содержащих различные ошибочные спаривания одной пары оснований, двух пар оснований или трех пар оснований. Однако в случае, если последовательности ДНК-мишени были такой же длины, как и последовательности PNA-зондов (т.е. длиной в 15 нуклеотидов), то зонды, представляющие собой параллельную PNA, были гораздо более предпочтительны, чем зонды, представляющие собой антипараллельную PNA, из-за большей величины различий в наблюдаемых показателях интенсивности флуоресценции между правильно спаренными комплексами и комплексами с ошибочным спариванием одной или двух пар оснований.
Хотя точный механизм, благодаря которому зонды, представляющие собой параллельную PNA, предпочтительны в таких условиях, неизвестен, но эти наблюдения подтверждают, что ошибочно спаренные гибридизационные комплексы параллельная PNA: ДНК менее стабильны, чем аналогичные ошибочно спаренные гибриды антипараллельная PNA:ДНК, что приводит к меньшему встраиванию представляющего собой нуклеиновую кислоту интеркалятора и, следовательно, наблюдаются более низкие значения интенсивности флуоресценции.
Предпочтительно чтобы зонд и мишень были немечеными, но в альтернативных вариантах осуществления изобретения имеется интеркалирующий агент, ковалентно связанный с зондом. В таких вариантах осуществления изобретения интеркалирующий агент предпочтительно связывается с зондом на любом конце.
В других вариантах осуществления изобретения интеркалирующий агент не связан ковалентно с зондом, хотя он может встраиваться между зондом и мишенью во время анализа, входя в смысловую связь с зондом, нековалентным образом.
Предпочтительные для использования по настоящему изобретению интеркалирующие агенты включают, например, YOYO-1, ТОТО-1, этидийбромид, гомодимер-1 этидия, гомодимер-2 этидия и акридин. Как правило, интеркалирующий агент представляет собой группу, которая способна встраиваться между цепями дуплексного и/или триплексного комплекса нуклеиновых кислот. В предпочтительных вариантах осуществления изобретения иктеркалирующий агент (или его компонент) является по существу не флуоресцентным в отсутствие нуклеиновых кислот, и флуоресцирует, когда он встраивается и возбуждается излучением с подходящей длиной волны, проявляя усиление флуоресценции в 100 до 10000 раз, когда он встраивается в дуплексный или триплексный комплекс нуклеиновых кислот.
В альтернативных вариантах осуществления изобретения интеркалирующий агент может выбывать сдвиг длины волны флуоресценции после встраивания и возбуждения излучением с подходящей длиной волны. Точная длина волны флуоресценции может зависеть от структуры нуклеиновой кислоты, в которую осуществляется встраивание, например, ДНК против РНК, дуплекс против триплекса и т.п. Особый случай изменения длины волны флуоресценции в результате электронного взаимодействия двух флуорофоров (резонансный перенос энергии), находящихся на одном и том же зонде, раскрыт в патенте US 5332659. Кроме того, YOYO-1, интеркалировавший в одноцепочечную нуклеиновую кислоту, имеет максимум флуоресценции при большей длине волны, чем YOYO-1, интеркалировавший в дуплексную или триплексную нуклеиновую кислоту, так что сдвиг максимума флуоресценции в коротковолновую сторону свидетельствует о событии, связанном с образованием дуплекса или триплекса. Примерами сдвига максимума испускания флуоресценции при образовании дуплекса служат результаты, представленные на фиг. 2А и 3А, а примерами сдвига максимума испускания флуоресценции при образовании локального триплекса при использовании конъюгированного основания служат результаты, приведенные на фиг.5А и 5В из патента US 6656692.
В качестве неограничивающего примера интеркалирующего агента, длина волны флуоресценции которого зависит от природы нуклеиновой кислоты, в которую он встроен, можно привести акридин, который имеет разные максимумы флуоресценции в зависимости от того, интеркалирован ли он в ДНК (зеленая флуоресценция с λmax 525 нм) или в РНК (красная флуоресценция с λmax около 650 нм) (Handbook of Fluorescent Probes and Research Chemicals, by R. P. Haugland, 6th Edition, Molecular Probes, 1996, p.145).
Длину волны возбуждения выбирают (обычным опытным путем и/или на основании обычных знаний) так, чтобы она соответствовала максимуму возбуждения для используемого флуорофора, и предпочтительно она составляет от 200 до 1000 нм. Предпочтительно выбирают интеркалирующие агенты, имеющие длину испускаемых волн от 200 до 1000 нм. В предпочтительных вариантах осуществления изобретения используют аргоновый ионный лазер для облучения флуорофора светом, имеющим длину волн в диапазоне от 400 до 540 нм, а флуоресцентную эмиссию обнаруживают в диапазоне от 500 до 750 нм.
Анализ по настоящему изобретению можно осуществлять в широком интервале температур, например, от 5 до 85°С. Некоторые анализы по прототипу требуют повышенных температур, что повышает стоимость анализа и увеличивает его длительность. С другой стороны, анализ по настоящему изобретению можно проводить при комнатной или более низкой температуре (например, при температуре ниже 25°С).
Надежность данного изобретения не зависит от содержания гуанина и цитозина (GC)в мишени. Поскольку пары оснований G-C образуют три водородные связи, а пары оснований А-Т образуют только две водородные связи, то последовательности мишени и зонда с более высоким содержанием G и С более стабильны, обладая более высокими температурами плавления. Вследствие этого ошибочные спаривания пар оснований, которые увеличивают содержание GC в гибридизированной области зонда и мишени по сравнению с тем, который присутствует в правильно спаренных гибридах, могут привести к ослаблению связывания, связанному с ошибочно спаренным зондом. Гибридизационные комплексы, содержащие все возможные ошибочные спаривания пар оснований между зондом и мишенью, оказались более нестабильными, чем правильно спаренные гибриды, что всегда приводит к более низким значениям интенсивности флуоресценции, чем у полностью комплементарных гибридов.
Анализ по настоящему изобретению обладает чрезвычайно высокой чувствительностью, поэтому нет необходимости осуществлять амплификацию мишени с помощью ПЦР. Например, можно проанализировать испытуемый образец, имеющий объем около 20 микролитров, содержащий около 10 фемтомоль мишени и около 10 фемтомоль зонда. Варианты осуществления настоящего изобретения обладают достаточно высокой чувствительностью, чтобы анализировать мишени при концентрации в 5×10-9 М, предпочтительно при концентрации не более, чем 5×10-10 М. Варианты осуществления настоящего изобретения обладают достаточно высокой чувствительностью, чтобы использовать зонды в концентрации 5×10-9 М, предпочтительно в концентрации не более чем 5×10-10 М. Само собой разумеется, что вышеуказанные значения не означают, что с помощью данного способа нельзя обнаружить более высокие концентрации.
В качестве гибридизационной среды можно использовать любую обычную среду, пригодную для сохранения нуклеотидов. См., например, Sambrook et al., "Molecular cloning: A Lab Manual", Vol.2 (1989). Например, жидкая среда может включать нуклеотиды, воду, буферы и стандартные концентрации солей.
Гибридизация между комплементарными основаниями происходит при широком разнообразии условий, в т.ч. при различных значениях температуры, концентраций солей, электростатической прочности и при различных составах буферов. Примеры таких условий и способов их применения известны в данной области техники.
Предпочтительно чтобы гибридизационные комплексы образовывались при температуре от около 15°С до около 25°С, в течение от около 1 минуты до около 5 минут. Более длительное время реакции не требуется, но инкубация в течение нескольких часов не оказывает отрицательного влияния на гибридизационные комплексы.
Если в реакционную смесь включен подходящий интеркалирующий агент (как описано выше), то в подавляющем большинстве случаев другие облегчающие гибридизацию реагенты не требуются. Однако можно облегчить гибридизацию в растворе с помощью некоторых реагентов. Предпочтительные примеры таких реагентов включают одноцепочечные связывающие белки, такие как белок Rec А, белок 32 гена Т4, одноцепочечный связывающий белок Е. coli, большие и малые белки связывания бороздок нуклеиновых кислот, дивалентные ионы, поливалентные ионы, виологен (viologen) и интеркалирующие вещества, такие как этидийбромид, актиномицин D, псорален и ангелицин. Такие облегчающие гибридизацию реагенты могут оказаться полезными в экстремальных условиях работы, например, при ненормальных уровнях рН или чрезвычайно высоких температурах.
Анализ по настоящему изобретению можно использовать, например, для идентификации доступных областей в складчатых последовательностях нуклеотидов, для определения количества ошибочно спаренных пар оснований в гибридизационном комплексе, а также для картирования геномов.
Изобретение далее иллюстрируется более подробно со ссылками на нижеследующие примеры, однако следует иметь в виду, что настоящее изобретение ими не ограничивается.
ПРИМЕРЫ
Пример 1
Смысловые и антисмысловые последовательности мишени, представляющей собой 50-членную ssDNA, происходящие из экзона 10 гена муковисцидоза человека (Nature 380, 207 (1996)), синтезировали на ДНК-синтезаторе (Expedite 8909, PerSeptive Biosystems) и очищали с помощью ВЭЖХ. Эквимолярные количества комплементарных олигонуклеотидов денатурировали при 95°С в течение 10 минут и давали им постепенно отжечься, пока температура понижалась до 21°С в течение 1,5 часов. Олигонуклеотиды dsDNA растворяли в ddH2O (бидистиллированной воде) в концентрации 1 пмоль/мкл.
Смысловая цепь мишени, представляющей собой ДНК дикого типа, имела нижеследующую последовательность (SEQ ID NO:1): 5′-TGG CAC CAT TAA AGA AAA TAT CAT CTT TGG TGT TTC СТА TGA TGA ATA TA-3′.
Антисмысловая цепь мишени, представляющей собой ДНК дикого типа, имела нижеследующую последовательность (SEQ ID NO:1): 5′ -TAT ATT CAT CAT AGG AAA CAC CAA AGA TGA TAT TTT CTT TAA TGG TGC CA-3′.
Получили последовательность мишени, представляющей собой 50-членную мутантную dsDNA, идентичную мишени, представляющей собой ДНК дикого типа (SEQ ID NO:1), за исключением мутации одной пары оснований (подчеркнуто) в позиции 507 аминокислоты, в которой последовательность дикого типа CAT была изменена на CGT.
Смысловая цепь этой 50-членной последовательности мишени имела следующую последовательность (SEQ ID NO:2): 5′-TGG CAC CAT TAA AGA AAA TAT CGT CTT TGG TGT TTC СТА TGA TGA ATA TA-3′.
Антисмысловая цепь этой 50-членной последовательности мишени имела следующую последовательность (SEQ ID NO:2): 5′-ТАТ АТТ CAT CAT AGG AAA САС САА AGA CGA TAT TTT СТТ ТАА TGG TGC СА-3′.
Получили последовательность мишени, представляющей собой 50-членную мутантную dsDNA, идентичную ДНК дикого типа мишени (SEQ ID NO:1), за исключением мутации двух последовательно расположенных пар оснований (подчеркнуто) в позициях 506 и 507 аминокислот, в которых последовательность дикого типа CAT была изменена на ACT.
Смысловая цепь этой 50-членной последовательности мишени имела следующую последовательность (SEQ ID NO:3): 5′-TGG CAC CAT ТАА AGA AAA TAT ACT CTT TGG TGT TTC СТА TGA TGA ATA TA-3′.
Антисмысловая цепь этой последовательности мишени имела следующую последовательность (SEQ ID NO:3): 5′-ТАТ АТТ CAT CAT AGG AAA CAC САА AGA GTA TAT TTT CTT TAA TGG TGC CA-3′.
Получили последовательность мишени, представляющей собой 50-членную мутантную dsDNA, идентичную ДНК дикого типа мишени (SEQ ID NO:1), за исключением мутации трех последовательно расположенных пар оснований (подчеркнуто) в позициях 506 и 507 аминокислот, в которых последовательность дикого типа CAT была изменена на ACG.
Смысловая цепь этой последовательности мишени имела следующую последовательность (SEQ ID NO:4): 5′-TGG CAC CAT TAA AGA AAA TAT ACG CTT TGG TGT TTC CTA TGA TGA ATA TA-3′.
Антисмысловая цепь этой последовательности мишени имела следующую последовательность (SEQ ID NO:4): 5′-TAT ATT CAT CAT AGG AAA CAC CAA AGCGTA TAT TTT CTT TAA TGG TGC CA-3′.
Получили последовательность мишени, представляющей собой 47-членную мутантную dsDNA, идентичную ДНК дикого типа мишени (SEQ ID NO:1), за исключением делеции трех последовательно расположенных пар оснований (указано многоточием) в позициях 507 и 508 аминокислот, в которых последовательность дикого типа CTT была утрачена.
Смысловая цепь этой 47-членной последовательности мишени имела следующую последовательность (SEQ ID NO:5): 5′-TGG CAC CAT TAA AGA AAA TAT CAT... TGG TGT TTC СТА TGA TGA ATA TA-3′.
Антисмысловая цепь этой 47-членной последовательности мишени имела следующую последовательность (SEQ ID NO:5): 5′-ТАТ ATT CAT CAT AGG AAA CAC CA... A TGA TAT TTT CTT TAA TGG TGC CA-3′.
PNA-зонды, использованные в примерах, синтезировали, очищали с помощью ВЭЖХ и подтверждали с помощью масс-спектроскопии в Commonwealth Biotechnologies, Inc. (Richmond, VA, USA). PNA-зонды сначала растворяли в 0,1% TFA (трифторуксусной кислоте) до концентрации 10 мг/мл, а затем разводили до 1 мг/мл путем добавления ddH2O. Окончательные маточные растворы PNA готовили в ddH2O в концентрации 1 пмоль/мкл.
Зонд №1 представлял собой 15-членную антипараллельную PNA и был сконструированный так, чтобы он был полностью комплементарным 15-нуклеотидному сегменту смысловой цепи 50-членной последовательности мишени, представлявшей собой ДНК дикого типа (SEQ ID NO:1), перекрывая позиции аминокислот от 505 до 510 (Nature 380, 207 (1996)). Зонд имел следующую структуру (SEQ ID NO: 6): 5′-H-CAC CAA AGA TGA TAT-Lys-CONH2-3′.
Зонд №2 представлял собой 15-членную PNA, с последовательностью, идентичной последовательности зонда №1, но в отличие от него, имел параллельную ориентацию вместо антипараллельной ориентации. Зонд имел следующую структуру (SEQ ID NO:7):
5′-Н-ТАТ AGT AGA AAC CAC-Lys-CONH2-3′.
Каждая гибридизационная реакционная смесь (80 мкл) содержала следующее: 4 пмоль dsDNA-мишени, 4 пмоль PNA-зонда, 0,5×ТВЕ и 500 нМ ДНК-интеркалятора YOYO-1 (Molecular Probes, Eugene, OR, USA). Реакционные смеси инкубировали при 95°С в течение 5-10 минут, чтобы произошла денатурация, а затем держали при 80°С до начала анализа. Образцы помещали в кварцевую кювету, облучали лучом аргонового ионного лазера с длиной волны 488 нм и периодически замеряли флуоресцентную эмиссию по мере того, как температура понижалась со временем. Совпадающие по времени измерения температуры производили с помощью контролируемого компьютером датчика температуры, помещенного непосредственно в каждый образец. Максимальная интенсивность флуоресценции наблюдалась при длине волны в 540 нм, что указывало на встраивание YOYO-1 в гибриды PNA:ДНК. Значения максимальной интенсивности флуоресценции показывали на графиках как функцию температуры для каждого проанализированного образца.
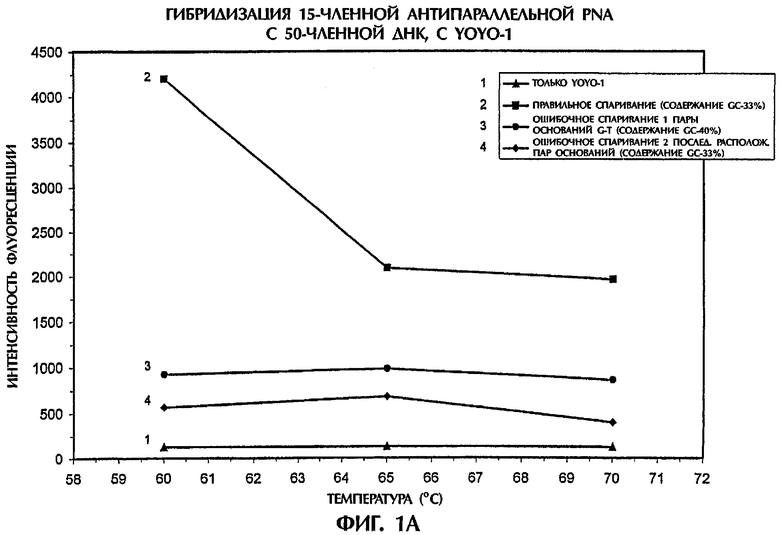
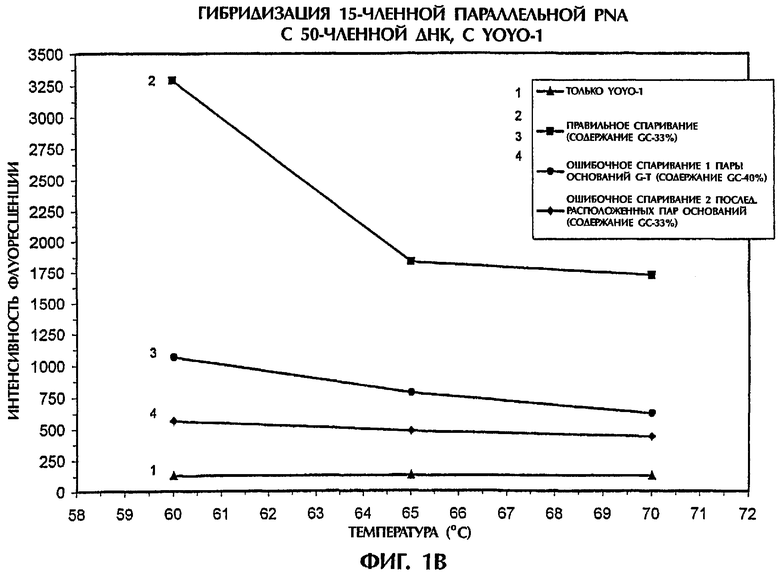
Значения интенсивности флуоресценции, наблюдавшиеся в отсутствие ДНК или PNA (присутствовал только YOYO-1), или когда последовательность дикого типа (SEQ ID NO:1), мутантная последовательность (SEQ ID NO:2) или мутантная последовательность (SEQ ID NO:3) реагировали с зондом №1, представляющим собой антипараллельную PNA, или с зондом №2, представляющим собой параллельную PNA, показаны на фиг.1А и 1В, соответственно. Гибриды ssDNA:PNA, состоящие из полностью комплементарных последовательностей (SEQ ID NO:1 + зонд №1) давали возможность максимального встраивания YOYO-1, что приводило к самым высоким значениям интенсивности флуоресценции, которые увеличивались по мере понижения температуры (фиг.1А). Значения интенсивности флуоресценции для гибридов ssDNA:PNA с ошибочным спариванием одной пары оснований (SEQ ID NO:2 + зонд №1) и гибридов ssDNA:PNA с ошибочным спариванием двух пар оснований (SEQ ID NO:3 + зонд №1) были соответственно на 81% и 89% ниже, чем для гибрида ssDNA:PNA с правильным спариванием, при температуре 60°С (фиг.1А). Аналогично, когда зонд №2, представляющий собой параллельную PNA, гибридизировали с последовательностями ДНК-мишени, гибриды ssDNA:PNA с ошибочным спариванием одной и двух пар оснований демонстрировали интенсивность флуоресценции, которая была соответственно на 70% и 86% ниже, чем у полностью комплементарного гибрида ssDNA:PNA (SEQ ID NO:1 + зонд №2), при температуре 60°С (фиг.1В). По мере увеличения степени ошибочного спаривания между зондом и мишенью уровень встраивания YOYO-1 уменьшался, а следовательно, понижался уровень интенсивности флуоресценции. Эта взаимосвязь выдерживалась не зависимо от того, использовали ли зонд, представляющий собой антипараллельную или параллельную PNA.
Интересно, что когда последовательности ДНК-мишени были такой же самой длины, что и последовательности PNA-зонда (т.е. длиной в 15 нуклеотидов), то зонды, представляющие собой параллельную PNA, были гораздо более предпочтительны, чем зонды, представляющие собой антипараллельную PNA, из-за бóльших различий в наблюдаемых значениях интенсивности флуоресценции между правильно спаренными комплексами и комплексами с ошибочно спаренными одной или двумя парами оснований (данные не показаны).
Хотя точный механизм, благодаря которому зонды, представляющие собой параллельную PNA, предпочтительны в таких условиях, неизвестен, но эти наблюдения подтверждают, что ошибочно спаренные гибридизационные комплексы параллельная PNA:ДНК менее стабильны, чем аналогичные ошибочно спаренные гибриды антипараллельная PNA:ДНК, что приводит к тому, что меньше встраивается представляющий собой нуклеиновую кислоту интеркалятор и, следовательно, наблюдаются более низкие значения интенсивности флуоресценции.
Пример 2
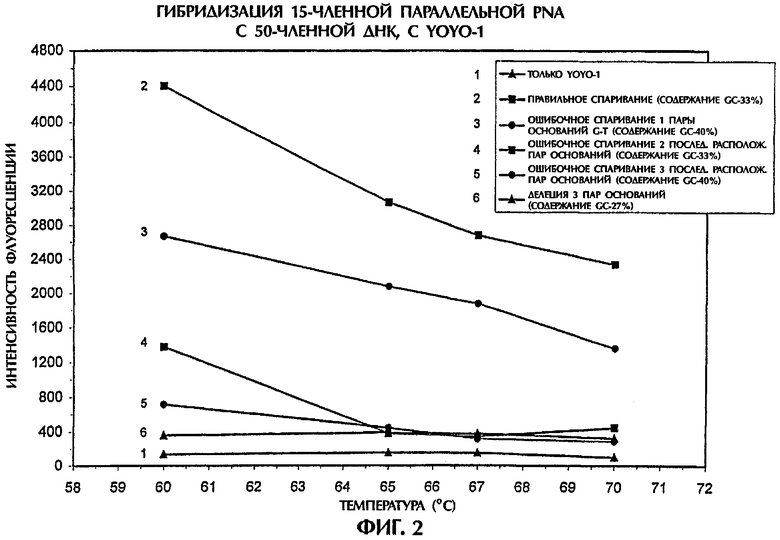
Фиг.2 демонстрирует, что анализ гибридизации по настоящему изобретению может также выявить различия между гибридами ssDNA:PNA с правильным спариванием и гибридами, содержащими 1, 2 или 3 ошибочно спаренных пар оснований, а также делеции 3 пар оснований, если используется зонд, представляющий собой параллельную PNA. Условия анализа были идентичными описанным в примере 1. Значения интенсивности флуоресценции для гибрида ssDNA:PNA с ошибочным спариванием одной пары оснований (SEQ ID NO:2 + зонд №2), с ошибочным спариванием двух последовательно расположенных пар оснований (SEQ ID NO:3 + зонд №2), с ошибочным спариванием трех последовательно расположенных пар оснований (SEQ ID NO:4 + зонд №2) и с делецией 3 пар оснований (SEQ ID NO:5 + зонд №2) были соответственно на 41%, 71%, 87% и 95% ниже, чем у правильно спаренного гибрида ssDNA:PNA (SEQ ID NO:1 + зонд №2), при температуре 60°С. По мере понижения температуры от 70°С до 60°С степень различий между правильно спаренными гибридами и гибридами с разным числом ошибочно спаренных пар оснований увеличивалась. Как и в примере 1, увеличение степени ошибочного спаривания между зондом и мишенью приводило к прогрессивно снижающимся уровням интенсивности флуоресценции. Кроме того, такой характер флуоресценции сохранялся даже в том случае, когда мутантные последовательности имели содержание GC выше, чем в последовательностях дикого типа (фиг.2).
Когда использовали зонд, представляющий собой параллельную PNA, то ошибочное спаривание двух расположенных не подряд пар оснований (в которых две области ошибочного спаривания величиной в 1 пару оснований каждая была разделены тремя парами оснований) привело к немного более низким значениям интенсивности флуоресценции, чем при ошибочном спаривании двух расположенных подряд пар оснований (данные не показаны). Аналогично ошибочное спаривание трех расположенных не подряд пар оснований (в которых каждая из трех областей ошибочного спаривания величиной в 1 пару оснований была отделена тремя парами оснований) привело к несколько более низким значениям интенсивности флуоресценции, чем для ошибочного спаривания трех расположенных подряд пар оснований (данные не показаны). Эти результаты подтверждают, что расположенные не подряд ошибочно спаренные основания в большей степени нарушают образование гибридов ssDNA:PNA, чем последовательно расположенные ошибочно спаренные пары оснований.
Пример 3
Последовательность SEQ ID NO:8 представляла собой 15-членную последовательность dsDNA-мишени, происходящую из SEQ ID NO:1, сконструированную так, чтобы она была полностью комплементарной зонду №1. Последовательности от SEQ ID NO:9 до SEQ ID NO:17 представляли собой 15-членные последовательности мишени, представляющей собой мутантную dsDNA, идентичные последовательности дикого типа SEQ ID NO:8, за исключением мутации одной пары оснований (подчеркнуто). Смысловые и антисмысловые 15-мерные последовательности ssDNA синтезировали, очищали и отжигали, как описано выше. Олигонуклеотиды dsDNA растворяли в ddH2O в концентрации 1 пмоль /мкл.
Последовательность для смысловой цепи мишени, представляющей собой ДНК дикого типа (SEQ ID NO:8) была следующей: 5′-АТА ТСА ТСТ TTG GTG-3′.
Последовательность для антисмысловой цепи мишени, представляющей собой ДНК дикого типа (SEQ ID NO:8) была следующей: 5′- САС САА AGA TGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:9) была следующей: 5′-ATA TCT TCT TTG GTG-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:9) была следующей: 5′- САС САА AGA AGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:10) была следующей: 5′-АТА ТСА TCT TTC GTG-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:10) была следующей: 5′-САС GAA AGA TGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO: 11) была следующей: 5′-ATA ТСА TGT TTG GTG-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:11) была следующей: 5′-САС САА АСА TGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO: 12) была следующей: 5′м-ATA ТСА TCT ATG GTG-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:12) была следующей: 5′-САС CAT AGA TGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:13) была следующей: 5′- АТА ТСА TCT CTG СТС-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:13) была следующей: 5′-CAC CAG AGA TGA TAT-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:14) была следующей: 5′-АТА ТСА ТСТ GTG GTG-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:14) была следующей: 5′-САС САС AGA TGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:15) была следующей: 5′- АТА TCG ТСТ TTG GTG-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:15) была следующей: 5′-САС САА AGA CGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO: 16) была следующей: 5′- АТА ТСА ТТТ TTG СТС-3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:16) была следующей: 5′-САС САА ААА TGA ТАТ-3′.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:17) была следующей: 5′- АТА ТСА ТСТ ТТТ GTG -3′.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:17) была следующей: 5′-CAC ААА AGA TGA ТАТ-3′.
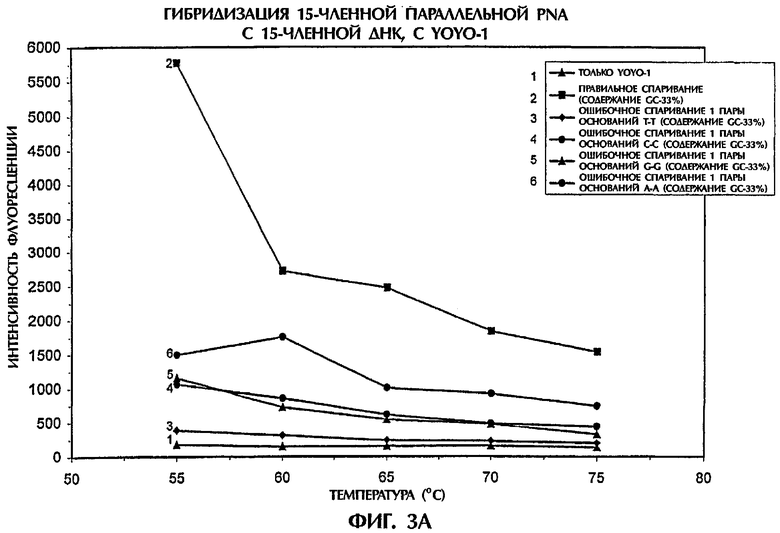
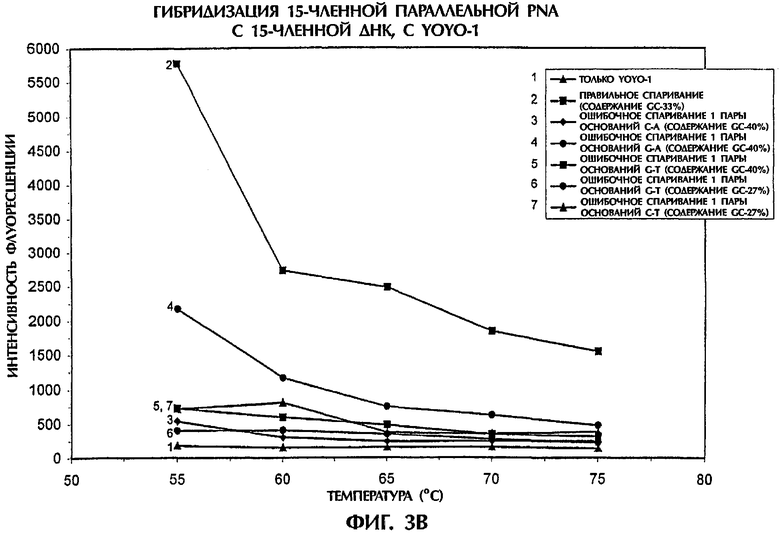
Затем испытывали специфичность анализа гибридизации путем проведения реакции зонда №2, представляющего собой параллельную PNA, с мишенью, представляющей собой 15-членную dsDNA дикого типа (SEQ ID NO:8) и с различными мишенями, представляющими собой 15-членную dsDNA с мутацией 1 пары оснований (от SEQ ID NO:9 до SEQ ID NO:17), что дает возможность получить все возможные типы ошибочного спаривания 1 пары оснований (фиг.3). Условия анализа были идентичными описанным в примере 1.
Самые высокие значения интенсивности флуоресценции получили для гибридов ssDNA:PNA, состоящих из полностью комплементарных последовательностей, при всех испытывавшихся температурах. Флуоресценция увеличивалась по мере понижения температуры. У всех гибридов ssDNA:PNA, у которых получены ошибочные спаривания одной пары оснований Т-Т, С-С, G-G, А-А, С-А, G-A, G-T и С-Т, значения интенсивности флуоресценции были ниже, чем наблюдавшиеся для правильно спаренных гибридов ssDNA:PNA (Фиг.3А и 3В). Значения интенсивности флуоресценции для ошибочных спаривании 1 пары оснований были на 64% до 96% ниже и на 57% до 95% ниже, чем наблюдавшиеся для правильного спаривания при температурах 55°С и 75°С, соответственно. Изменчивость в интенсивности флуоресценции, наблюдавшаяся между различными ошибочными спариваниями 1 пары оснований, больше зависела от ошибочного спаривания конкретной пары оснований, чем от изменения в процентном содержании GC в мутантных последовательностях, когда использовали зонд, представлявший собой параллельную PNA (фиг.3). Когда в подобном эксперименте испытывали зонд №1, представлявший собой антипараллельную PNA, то различия в интенсивности флуоресценции, наблюдавшиеся между правильным спариванием и различными ошибочными спариваниями 1 пары оснований, были менее резкими, чем в случае использования зонда №2, представлявшего собой параллельную PNA и, по-видимому, они зависели от температуры (данные не показаны). Анализ дал наилучшие результаты при температурах между 40°С и 60°С, когда использовали зонд, представлявший собой антипараллельную PNA. Поэтому зонд, представляющий собой параллельную PNA, является предпочтительным в тех случаях, когда последовательности ДНК-мишени имеют такую же самую длину, что и последовательности PNA-зонда.
Результаты, представленные на фиг.3, подтвердили надежность анализа гибридизации для идентификации всех возможных ошибочных спаривании 1 пары оснований с высокой точностью.
Пример 4
Анализы гибридизации, описанные в примерах от 1 до 3, осуществляли после денатурации последовательностей dsDNA-мишени и измеряли образование гибридов ssDNA:PNA при температурах выше точки плавления (Тm) dsDNA-мишеней. Данный пример демонстрирует надежность анализа по настоящему изобретению для осуществления дифференциации между правильными спариваниями и ошибочными спариваниями пар оснований, без необходимости в проведении предварительной денатурации.
Гибридизационная реакционная смесь (120 мкл) содержала следующее: 6 пмоль dsDNA-мишени, 6 пмоль зонда №2, представляющего собой параллельную PNA, 0,5×ТВЕ и 500 нМ ДНК-интеркалятора YOYO-1. Хотя объем реакционной смеси был немного больше, чем в примерах от 1 до 3, но количества ДНК, PNA и YOYO-1 корректировали так, чтобы поддерживать постоянную концентрацию образцов. Идентичные результаты были получены, когда объем реакции составлял 40 мкл, 80 мкл или 120 мкл. Реакционные смеси инкубировали при комнатной температуре (21°С) в течение 5 минут, помещали в кварцевую кювету, облучали лучом аргонового ионного лазера с длиной волны 488 нм и периодически замеряли флуоресцентную эмиссию по мере того, как температура повышалась со временем, в нагреваемой камере. Совпадающие по времени измерения температуры образцов производили с помощью контролируемого компьютером датчика температуры, помещенного непосредственно в каждый образец. Максимальные значения интенсивности флуоресценции показывали на графиках как функцию температуры для каждого проанализированного образца.
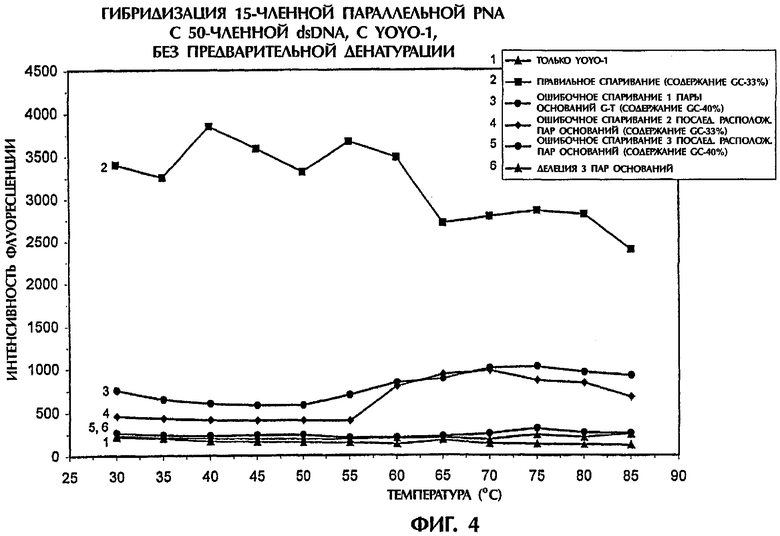
Из фиг.4 видно, что даже без предварительной денатурации были достигнуты самые высокие уровни интенсивности флуоресценции, когда проводили реакцию последовательности мишени, представляющей собой 50-членную dsDNA (SEQ ID NO:1), с зондом №2, представляющим собой 15-членную параллельную PNA, при температурах от 30°С до 85°С. При температуре ниже 65°С, которая является температурой плавления 50-членной dsDNA дикого типа, образовывались триплексы dsDNA:PNA. Когда температуру поднимали выше 65°С, триплексные структуры преобразовывались в дуплексы ssDNA:PNA. Ясно, что YOYO-1 был способен эффективно встраиваться как в триплексные, так и в дуплексные структуры. Вследствие этого значения интенсивности флуоресценции для триплекса dsDNA:PNA с ошибочно спаренной одной парой оснований (SEQ ID NO:2 + зонд №2), триплекса с ошибочно спаренными двумя последовательно расположенными парами оснований (SEQ ID NO:3 + зонд №2), триплекса с ошибочно спаренными тремя последовательно расположенными парами оснований (SEQ ID NO:4 + зонд №2) и для триплекса с делецией трех пар оснований (SEQ ID NO:5 + зонд №2), были ниже на 83%, 93%, 99% и 99,5%, соответственно, чем для правильно спаренного триплекса dsDNA:PNA (SEQ ID NO:1 + зонд №2), при температуре 30°С. Когда температура повышалась от 30°С до 85°С, степень различий между правильным спариванием и ошибочным спариванием пар оснований уменьшалась. При температуре 85°С значения интенсивности флуоресценции для гибридов с ошибочным спариванием 1 пары оснований, с ошибочным спариванием 2 пар оснований, с ошибочным спариванием 3 пар оснований и с делецией 3 пар оснований были ниже на 65%, 76%, 94% и 95% соответственно, чем наблюдавшиеся для полностью комплементарных последовательностей. Таким образом, анализ гибридизации по настоящему изобретению дает возможность отличать друг от друга последовательности дикого типа и последовательности, содержащие мутации или делеции 1 пары оснований, 2 пар оснований или 3 пар оснований, без предварительной денатурации последовательностей.
Пример 5
Зонд №3 представлял собой 15-членную ssDNA и был идентичен с точки зрения последовательности и ориентации зонду №1, представляющему собой 15-членную антипараллельную PNA (SEQ ID NO:6). Этот зонд имел следующую структуру:
5′-САС САА AGA TGA TAT - 3′.
Затем специфичность анализа гибридизации исследовали с помощью проведения реакции ssDNA-зонда №3 с 50-членными последовательностями мишени, представляющей собой дикого типа и мутантную dsDNA, без предварительной денатурации. Условия анализа были идентичными описанным в примере 4.
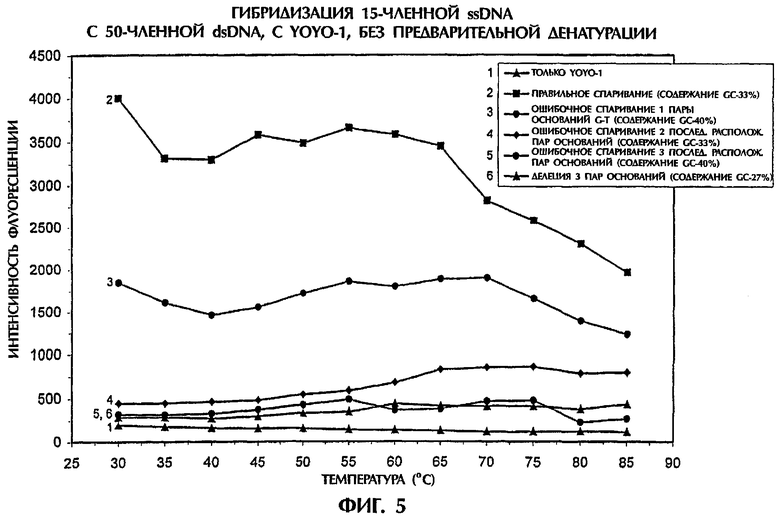
При стимулирующем воздействии ДНК-интеркалятора YOYO-1 триплексы dsDNA:ssDNA образовывались при температурах от 30°С до 65°С. Правильно спаренные ДНК-триплексы, состоящие из SEQ ID NO:1 + зонда №3, демонстрировали самые высокие значения интенсивности флуоресценции (фиг.5). В противоположность этому не полностью комплементарные комбинации зонда и мишени, несущие ошибочное спаривание 1 пары оснований (SEQ ID NO:2 + зонд №3), ошибочное спаривание последовательно расположенных 2 пар оснований (SEQ ID NO:3 + зонд №3), ошибочное спаривание последовательно расположенных 3 пар оснований (SEQ ID NO:4 + зонд №3) и делецию 3 пар оснований (SEQ ID NO:5 + зонд №3), демонстрировали значения интенсивности флуоресценции, которые были меньше на 57%, 94%, 97% и 98% при 30°С и на 47%, 79%, 92% и 91% при 65°С соответственно, чем наблюдавшиеся у правильно спаренных последовательностей (фиг.5). При повышении температуры выше 65°С степень различий между правильным спариванием и ошибочным спариванием пар оснований уменьшалась, что указывало на постепенное разрушение структур ДНК-триплексов. При 85°С значения интенсивности флуоресценции, полученные для ошибочного спаривания 1 пары оснований, ошибочного спаривания 2 пар оснований, ошибочного спаривания 3 пар оснований и делеции 3 пар оснований были на 40%, 63%, 92% и 83% ниже, чем наблюдавшиеся при правильном спаривании (фиг.5). Присутствие YOYO-1 позволяло использовать ssDNA-зонд вместо PNA-зонда, для дифференциации между полностью комплементарными последовательностями и последовательностями, содержавшими ошибочное спаривание 1 пары оснований, 2 пар оснований и 3 пар основания или делеции, без необходимости в предварительной денатурации.
Пример 6
Для того чтобы убедиться, что анализ гибридизации с помощью ssDNA-зондов и dsDNA-мишеней, осуществляемый без предварительной денатурации, применим к ДНК зондам и мишеням, имеющим резко различающееся содержание GC (гуанина и цитозина) (а следовательно, разные температуры плавления), синтезировали новые 15-членные последовательности ssDNA-зондов и 50-членные последовательности dsDNA-мишеней, очищали их и отжигали, как описано выше. Как ssDNA-зонды, так и dsDNA-мишени растворяли в ddH2O в концентрации 1 пмоль/мкл.
Последовательность SEQ ID NO:18 представляла собой 50-членную последовательность dsDNA-мишени - модифицированную SEQ ID NO:1, в которой процентное содержание GC было изменено с 30% на 52%.
Последовательность для смысловой цепи мишени, представлявшей собой ДНК дикого типа (SEQ ID NO:18), была следующей: 5′-GAG CAC CAT GAC AGA CAC TGT CAT CTC TGG TGT GTC СТА CGA TGA CTC TG-3′.
Последовательность для антисмысловой цепи мишени, представлявшей собой ДНК дикого типа (SEQ ID NO:18), была следующей: 5′- CAG AGT CAT CGT AGG АСА CAC CAG AGA TGA CAG TGT CTG TCA TGG TGC TC - 3′.
Последовательность SEQ ID NO:19 представляла собой 50-членную мутантную последовательность dsDNA-мишени, идентичную SEQ ID NO:18, за исключением мутации одной пары оснований (подчеркнуто), в которой последовательность CAT была изменена на CGT.
Последовательность для смысловой цепи мутантной последовательности SEQ ID NO:19 была следующей: 5′- GAG CAC CAT GAC AGA CAC TGT CGT CTC TGG TGT GTC СТА CGA TGA CTC TG -3′.
Последовательность для антисмысловой цепи мутантной последовательности SEQ ID NO:19 была следующей: 5′- CAG AGT CAT CGT AGG АСА CAC CAG AGA CGA CAG TGT CTG TCA TGG TGC TC-3′.
Последовательность SEQ ID NO:20 представляла собой 50-членную мутантную последовательность dsDNA-мишени, идентичную SEQ ID NO:18, за исключением мутации двух последовательно расположенных пар оснований (подчеркнуто), в которой последовательность CAT была изменена на ACT.
Последовательность для смысловой цепи мутантной последовательности SEQ ID NO:20 была следующей: 5′- GAG CAC CAT GAC AGA CAC TGT ACT CTC TGG TGT GTC СТА CGA TGA CTC TG -3′.
Последовательность для антисмысловой цепи мутантной последовательности SEQ ID NO:20 была следующей: 5′- CAG AGT CAT CGT AGG АСА CAC CAG AGA GTA CAG TGT CTG TCA TGG TGC TC-3′.
Последовательность SEQ ID NO:21 представляла собой 50-членную последовательность dsDNA-мишени, модифицированную из SEQ ID NO:1, в которой процентное содержание GC было изменено с 30% на 72%.
Последовательность для смысловой цепи мишени, представлявшей собой ДНК дикого типа (SEQ ID NO:21), была следующей: 5′- GAG CAC CCT CCC AGG САС GGT CGT CCC TGG TGC GAC CTC CGA CGA GCG TG-3′.
Последовательность для антисмысловой цепи мишени, представлявшей собой ДНК дикого типа (SEQ ID NO:21), была следующей: 5′- САС GCT CGT CGG AGG TCG CAC CAG GGA CGA CCG TGC CTG GGA GGG TGC ТС -3′.
Последовательность SEQ ID NO:22 представляла собой 50-членную мутантную последовательность dsDNA-мишени, идентичную SEQ ID NO: 21, за исключением мутации одной пары оснований (подчеркнуто), в которой последовательность CGT была изменена на CAT.
Последовательность для смысловой цепи мутантной последовательности SEQ ID NO 22: была следующей: 5′-GAG CAC CCT CCC AGG CAC GGT CAT CCC TGG TGC GAC CTC CGA CGA GCG TG -3′.
Последовательность для антисмысловой цепи мутантной последовательности SEQ ID NO:22 была следующей: 5′-САС GCT CGT CGG AGG TCG CAC CAG GGA TGA CCG TGC CTG GGA GGG TGC ТС -3′.
Последовательность SEQ ID NO:23 представляла собой 50-членную мутантную последовательность dsDNA-мишени, идентичную SEQ ID NO:21, за исключением мутации двух последовательно расположенных пар оснований (подчеркнуто), в которой последовательность CGT была изменена на АТТ.
Последовательность для смысловой цепи мутантной последовательности SEQ ID NO:23 была следующей: 5′-GAG CAC CCT ССС AGG CAC GGT АТТ ССС TGG TGC GAC CTC CGA CGA GCG TG-3′.
Последовательность для антисмысловой цепи мутантной последовательности SEQ ID NO: 23 была следующей: 5′- CAC GCT CGT CGG AGG TCG CAC CAG GGA ATA CCG TGC CTG GGA GGG TGC ТС-3′.
Зонд №4 представлял собой 15-членный ssDNA-зонд и был сконструирован полностью комплементарным 15-нуклеотидному сегменту смысловой цепи 50-членной последовательности мишени, представляющей собой ДНК дикого типа (SEQ ID NO:18). Зонд имел нижеследующую структуру (SEQ ID NO:24):
5′- CAC CAG AGA TGA CAG -3′.
Зонд №5 представлял собой 15-членный ssDNA-зонд и был сконструирован полностью комплементарным 15-нуклеотидному сегменту смысловой цепи 50-членной последовательности мишени, представляющей собой ДНК дикого типа (SEQ ID NO:21). Зонд имел нижеследующую структуру (SEQ ID NO:25):
5′- CAC CAG GGA CGA CCG -3′.
Условия анализа гибридизации были идентичными описанным в примере 4.
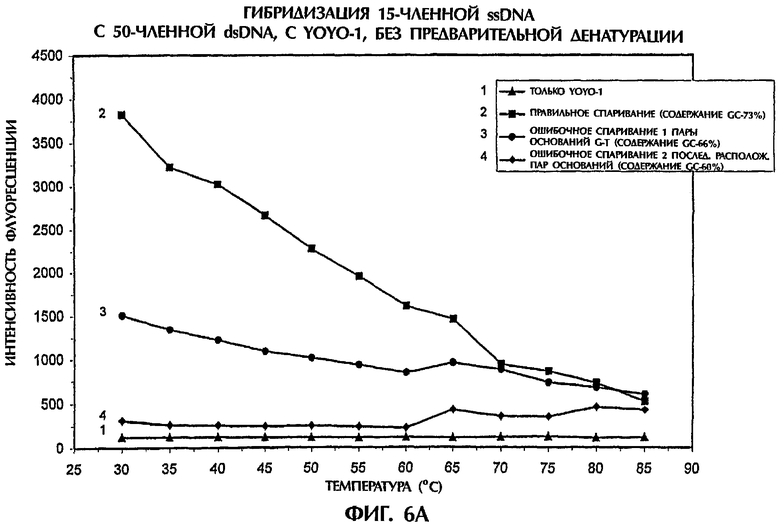
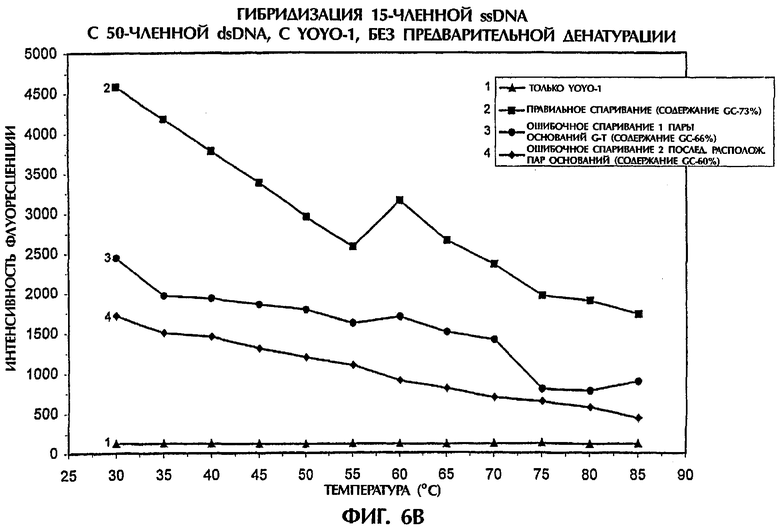
Когда ssDNA-зонд №4 (с содержанием GC 53%) гибридизировали с мишенью, представлявшей собой 50-членную dsDNA дикого типа (SEQ ID NO: 18) и с мишенями, представлявшими собой мутантные dsDNA (SEQ ID NO:19 и SEQ ID NO:20), триплексы dsDNA:ssDNA образовывались при низких температурах в условиях без денатурации (фиг.6А). В то время как правильно спаренные ДНК-триплексы достигали наивысших значений интенсивности флуоресценции, не полностью комплементарные триплексы с ошибочным спариванием 1 пары оснований (SEQ ID NO:19 + зонд №4) и с ошибочным спариванием 2 последовательно расположенных пар оснований (SEQ ID NO:20 + зонд №4) демонстрировали значения интенсивности флуоресценции, которые были соответственно на 63% и 95% ниже, чем полученные для правильно спаренных последовательностей при 30°С (фиг.6А). С повышением температуры происходило постепенное разрушение структуры ДНК-триплексов, что приводило к уменьшенным значениям интенсивности флуоресценции и к меньшим различиям между правильным спариванием и ошибочным спариванием пар оснований. При достижении 85°С наблюдались очень небольшие различия между правильно спаренными последовательностями и последовательностями, содержащими неправильно спаренные пары оснований (фиг.6А).
Аналогично в присутствии YOYO-1 образовывались триплексы dsDNA:ssDNA, когда проводили реакции ssDNA-зонда №5 (содержащего 73% GC) с соответствующей мишенью, представляющей собой 50-членную dsDNA дикого типа (SEQ ID NO:21) и с мишенями, представляющими собой мутантную dsDNA (SEQ ID NO:22 и SEQ ID NO:23). Значения интенсивности флуоресценции для ДНК-триплекса с ошибочно спаренной 1 парой оснований (SEQ ID NO: 22 + зонд №5) и для ДНК-триплекса с ошибочно спаренными последовательно расположенными 2 парами оснований (SEQ ID NO:23 + зонд №5) были соответственно на 48% и 64% ниже, чем полученные для правильно спаренных последовательностей при 30°С (фиг.6В). Флуоресценция всех образцов уменьшалась, когда температура повышалась от 30°С до 85°С, что указывало на уменьшение встраивания YOYO-1 и разрушение ДНК-триплексов.
Независимо от % содержания GC в ssDNA-зондах и dsDNA-мишенях YOYO-1 был способен стимулировать образование ДНК-триплексов в условиях без денатурации, что позволяло точно отличить полностью комплементарные последовательности от последовательностей, содержащих мутации 1 или 2 пар оснований.
Пример 7
Анализы гибридизации, описанные в примерах 5 и 6, доказали надежность способа по настоящему изобретению для того, чтобы отличать друг от друга последовательности ДНК дикого типа и последовательности, содержащие неправильные спаривания или делеции пар оснований, без необходимости проводить предварительную денатурацию, с использованием ssDNA-зондов. С помощью этих анализов измеряют образование ДНК-триплексов при температуре, которая ниже точки плавления dsDNA-мишеней. Кроме того, оптимальная дифференциация между последовательностями дикого типа и мутированными последовательностями наблюдалась при 30°С, что являлось наименьшей измеренной температурой. Для того чтобы выяснить, не будут ли температуры ниже 30°С еще более благоприятными для анализа, провели реакцию 50-членного ssDNA-зонда №3 с мишенью, представляющей собой 50-членную dsDNA дикого типа (SEQ ID NO:1) или с мишенью, представляющей собой 50-членную мутантную dsDNA (SEQ ID NO:2), при температурах между 5°С и 30°С.
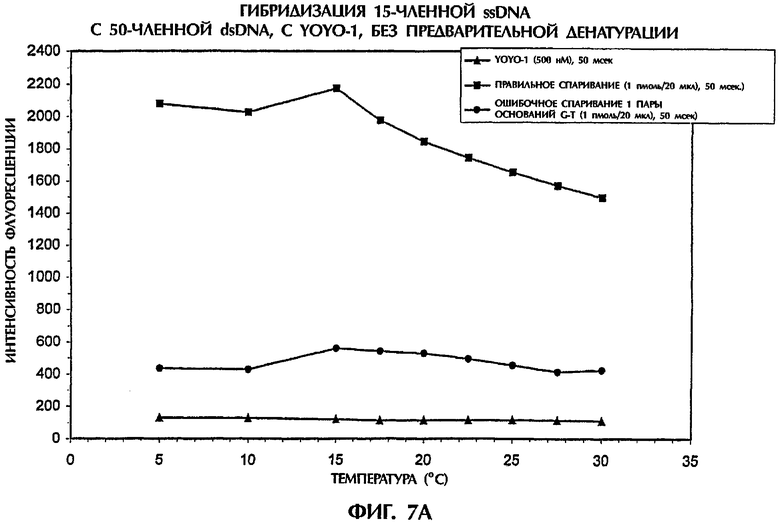
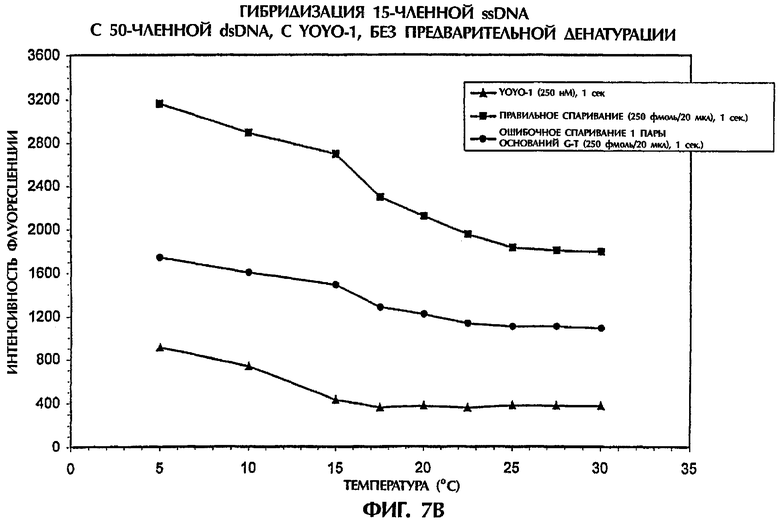
На фиг.7А показаны результаты реакции 2 пмоль dsDNA-мишени с 2 пмоль ssDNA-зонда №3 в 40 мкл реакционной смеси, содержащей 0,5 × ТВЕ и 500 нМ YOYO-1. На фиг.7В показаны результаты реакции 500 фмоль (т.е. 10-15 моль) dsDNA-мишени с 500 фмоль ssDNA-зонда №3 в 40 мкл реакционной смеси, содержащей 0,5 × ТВЕ и 250 нМ YOYO-1. Реакционные смеси инкубировали при комнатной температуре (21°С) в течение 5 минут, затем облучали лучом аргонового ионного лазера при 30°С, как это делали в примерах от 4 до 6. Затем кюветы с образцами помещали на лед. Когда температура каждого образца достигала 2°С (что определяли с помощью температурного датчика, помещенного непосредственно в каждый образец), образцы переносили в измерительную камеру и периодически замеряли флуоресцентную эмиссию по мере того, как температура со временем повышалась с 5°С до 30°С. Максимальные значения интенсивности флуоресценции выражали в виде графика в качестве функции температуры для каждого проанализированного образца.
Оптимальные различия между полностью комплементарными ДНК-триплексами (SEQ ID NO:1 + зонд №3) и ДНК-триплексами, содержащими 1 пару ошибочно спаренных оснований (SEQ ID NO:2 + зонд №3), наблюдали при 5°С при обеих испытывавшихся концентрациях ДНК (фиг.7А и 7В). Однако разница в значениях интенсивности флуоресценции между полностью и не полностью комплементарными ДНК-триплексами резко не менялась при повышении температуры от 5°C до 30°С. При концентрации в 1 пмоль/20 мкл как зонда, так и мишени, ДНК-триплекс с 1 ошибочно спаренной парой оснований демонстрировал показатели интенсивности флуоресценции, которые были на 84% и 77% ниже при 5°С и 30°С соответственно, чем полученные для правильно спаренных последовательностей (фиг.7А). Аналогично при в четыре раза более низкой концентрации зонда и мишени наблюдалась разница в интенсивности флуоресценции, составлявшая 63% и 50% при 5°С и 30°С соответственно, между правильно спаренными и ошибочно спаренными ДНК-триплексами (фиг.7В). Интенсивность флуоресценции всех образцов, измеренная при 30°С до и после охлаждения до 5°С, была очень близкой (данные не показаны). Хотя максимальные различия наблюдались при 5°С, но для удобства анализ гибридизации без применения денатурации можно надежно осуществлять при комнатной температуре.
Пример 8
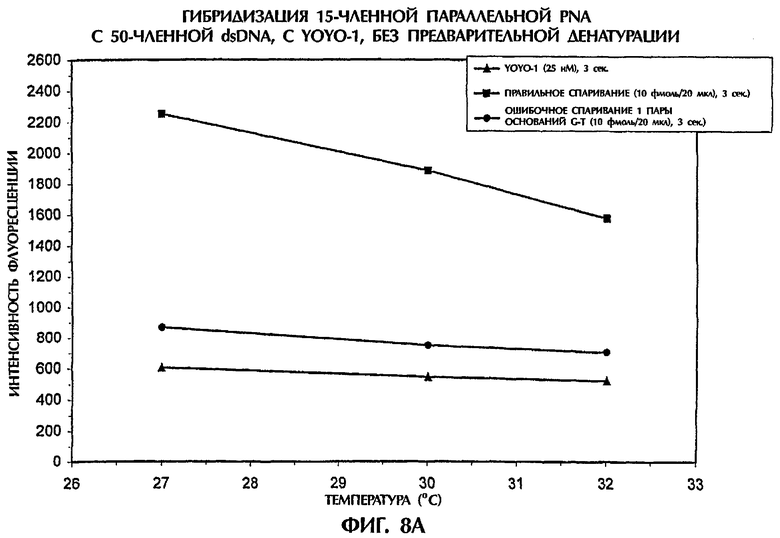
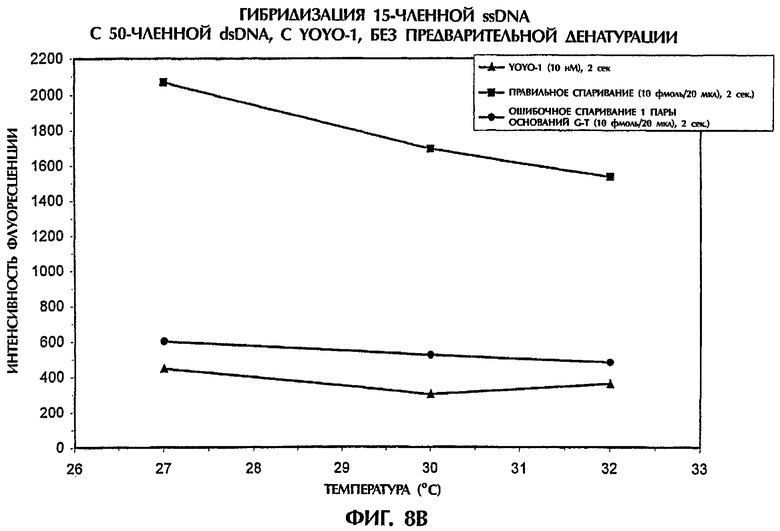
Чувствительность анализа гибридизации испытывали путем проведения реакции понижающихся концентраций зонда №2, представляющего собой параллельную PNA, или ssDNA-зонда №3 с понижающимися концентрациями мишеней, представляющих собой 50-членную dsDNA дикого типа или мутантную dsDNA (SEQ ID NO:1 и SEQ ID NO:2 соответственно), без денатурации, при температурах между 27°С и 32°С (фиг.8).
Гибридизационная реакционная смесь (40 мкл) содержала следующее: от 20 фмоль до 2 пмоль dsDNA-мишени, от 20 фмоль до 2 пмоль ssDNA-зонда №3 или зонда №2, представляющего собой параллельную PNA, 0,5 × ТВЕ и от 2,5 нМ до 500 нМ YOYO-1. В каждом образце последовательности мишени и последовательности зонда поддерживали в идентичных концентрациях. Условия анализа были идентичными описанным в примере 4.
Когда 1 пмоль/20 мкл зонда №2, представляющего собой параллельную PNA, или ssDNA-зонда №3 гибридизировали с 1 пмоль/20 мкл последовательностей dsDNA-мишени при нормальной концентрации YOYO-1 в 500 нМ, то dsDNA:PNA или dsDNA:ssDNA-триплексы, получаемые при ошибочном спаривании 1 пары оснований, демонстрировали значения интенсивности флуоресценции, которые были на 93% до 94% и на 56% до 54% ниже соответственно, чем наблюдавшиеся для правильно спаренных триплексов, при температуре от 27°С до 32°С (данные не показаны).
При концентрации в 10 фмоль/20 мкл как зонда, так и мишени (т.е. в 100 раз меньшей концентрации), гибрид dsDNA:PNA достигал значений интенсивности флуоресценции, которые были на 85%-83% ниже, чем полученные для полностью комплементарных триплексов, при температуре от 27°С до 32°С, при использовании 25 нМ YOYO-1 (фиг.8А). Аналогично, когда проводили реакцию 10 фмоль/20 мкл ssDNA-зонда №3 с 10 фмоль/20 мкл последовательностей dsDNA-мишени в присутствии 10 нм YOYO-1, значения интенсивности флуоресценции, получаемые при ошибочном спаривании 1 пары оснований, были на 90% до 84% ниже, чем для правильно спаренных ДНК-триплексов, при температуре от 27°С до 32°С (фиг.8В), что доказывает очень высокую чувствительность анализа гибридизации даже при очень низких концентрациях зонда и мишени.
Широкий диапазон концентраций YOYO-1 давал близкие результаты при каждой испытанной концентрации зонда и мишени. Когда гибридизировали 10 фмоль/20 мкл как зонда, так и мишени, оптимальные концентрации YOYO-1 составляли от 25 нМ до 2,5 нМ (для зонда, представлявшего собой параллельную PNA) и от 10 нМ до 2,5 нМ (для ssDNA-зонда), при этом различия в интенсивности флуоресценции между правильно спаренными и ошибочно спаренными последовательностями составляли от 90% до 71% (данные не показаны). Взятые вместе, эти результаты подтверждают очень высокую чувствительность и надежность не требующего денатурации анализа гибридизации для выявления различий между последовательностями дикого типа и последовательностями, содержащими различные мутации пар оснований.
Хотя настоящее изобретение описано подробно со ссылками на конкретные примеры его осуществления, но специалисту в данной области техники понятно, что могут быть осуществлены различные изменения и модификации данного изобретения, не изменяющие его существа и объема.








Изобретение относится к области биохимии и может быть использовано для гомогенного анализа гибридизации нуклеиновых кислот. Предложено два варианта способа анализа связывания мишени, содержащей полинуклеотидную последовательность. В соответствии с указанными вариантами мишень гибридизуют с зондом, включающим последовательность нуклеиновой кислоты или ее аналога, в присутствии интеркалирующего флуорофора. Образец, полученный при гибридизации, облучают возбуждающим излучением. В результате такой обработки флуорофор, содержащийся в образце, испускает флуоресцентное излучение. По интенсивности указанного излучения определяют степень ошибочного спаривания между зондом и мишенью или степень соответствия зонда и мишени. Использование изобретения позволяет упростить анализ взаимодействия между нуклеиновыми кислотами и/или аналогами нуклеиновых кислот, а также повысить чувствительность и скорость указанного процесса. 2 н. и 27 з.п. ф-лы, 13 ил.
обеспечение указанной мишени;
обеспечение указанного зонда;
обеспечение интеркалирующего агента, включающего флуорофор;
получение испытуемого образца путем добавления указанных зонда, мишени и интеркалирующего агента к гибридизационной среде с последующей инкубацией в условиях, подходящих для гибридизации;
облучение испытуемого образца возбуждающим излучением, чтобы заставить флуорофор испускать флуоресцентное излучение;
определение интенсивности флуоресцентного излучения;
определение на основании указанной интенсивности степени ошибочного спаривания между зондом и мишенью путем ее калибровки относительно интенсивностей, демонстрируемых образцами, полученными гибридизацией указанной мишени с другими зондами, различающимися между собой, по меньшей мере, одним основанием, причем указанный способ осуществляют при температуре, выбираемой из интервала от 5 до 85°С.
| СПОСОБ КОНТРОЛЯ СОСТОЯНИЯ ГОРНЫХ ВЫРАБОТОК | 2007 |
|
RU2333359C1 |
| WO 9745539 A, 04.12.1997 | |||
| WO 9501370 А, 12.01.1995. | |||

