Область техники, к которой относится изобретение
Данное изобретение относится к способам секвенирования или анализа нуклеиновых кислот, более конкретно к способам точного анализа триплексных и дуплексных комплексов гибридизации нуклеиновых кислот.
Уровень техники
В течение ряда лет было понятно, что биологические молекулы могут быть выделены и охарактеризованы посредством приложения к пробе электрического поля.
Электрофорез является, возможно, наиболее хорошо известным примером способа выделения и характеристики на основе влияния электрических полей на биологические молекулы. В гель-электрофорезе образуется однородный матрикс или гель, например полиакриламида, к которому прилагается электрическое поле. Смеси, нанесенные на одну сторону этого геля, мигрируют через гель в соответствии с их размером и взаимодействием с электрическим полем. Подвижность зависит от уникальных свойств вещества, таких как конформация, размер и заряд. Подвижность можно изменять изменением размеров пор геля, например образованием градиента концентрации или рН, или изменением состава буфера (рН, ДСН, DOC, глицина, соли). Одно- или двумерный гель-электрофорез являются довольно рутинными процедурами в большинстве исследовательских лабораторий. Вещества-мишени могут быть очищены пропусканием через гель и/или физической экстракцией из геля.
Относительно недавно разработанный способ, в котором электрическое поле прилагают к биологической пробе, описан в патенте США № 5824477, выданном Stanley. Патент Stanley описывает способ обнаружения присутствия или отсутствия заданной нуклеиновой кислоты в пробе. Этот способ предусматривает: (а) денатурацию двухцепочечной нуклеиновой кислоты посредством напряжения, прилагаемого на пробы в растворе при помощи электрода; (b) гибридизацию денатурированной нуклеиновой кислоты с олигонуклеотидным зондом для данной последовательности; и (с) определение, произошла ли гибридизаця. Патент Stanley описывает приложение электрического поля к подлежащей анализу пробе для ограниченной цели денатурации последовательности-мишени.
Более хорошо известный тип анализа гибридизации основан на применении флуоресцентно меченых агентов. В их наиболее основной форме анализы на основе интенсивности флуоресценции обычно предусматривали контактирование мишени с флуорофорсодержащим зондом, удаление любого несвязанного зонда из связанного зонда и детектирование флуоресценции в промытой пробе. Гомогенные анализы усовершенствуют такие основные анализы благодаря тому, что первые не требуют стадии промывания или обеспечения нежидкофазного носителя.
Некоторые анализы использовали интеркалирующиеся флуорофоры для обнаружения гибридизации нуклеиновых кислот на основе способности таких флуорофоров связываться между цепями нуклеиновой кислоты в гибридизационном комплексе.
Например, патент США № 5824557, выданный Burke et al., описывает способ и набор для обнаружения и количественного определения молекул нуклеиновых кислот. Предпочтительный вариант основан на интеркаляции (внедрении) красителя в двухцепочечную спираль нуклеиновой кислоты или одноцепочечную кислоту. Краситель флуоресцирует после интеркаляции, и эта интенсивность является прямой мерой количества нуклеиновой кислоты, присутствующей в пробе. Хотя способ Burke et al. имеет целью его применение для измерения количества нуклеиновой кислоты в пробе, неспецифическое связывание между интеркалятором и нуклеиновой кислотой, на котором основан этот способ, делает этот способ непрактичным для обнаружения специфического связывания, в частности, в условиях, когда присутствуют не являющиеся мишенями дуплексы нуклеиновых кислот.
Патент США № 5814447, выданный Ishiguro et al., описывает анализ, который имеет целью усовершенствование анализов, которые основаны на неспецифическом взаимодействии между интеркалирующими агентами и дуплексами нуклеиновых кислот, таких как Burke et al. и более ранний анализ, описанный Ishiguro et al. в опубликованной заявке на патент Японии № 237000/1993. Более ранняя разработка предусматривала добавление интеркалируюшего флуорохрома, имеющего тенденцию проявлять увеличенную интенсивность флуоресценции при интеркаляции в раствор пробы, перед тем как специфический район нуклеиновой кислоты-мишени амплифицировали при помощи ПЦР, и измерение интенсивности флуоресценции из реакционной смеси при заданных интервалах времени для детектирования и количественного определения нуклеиновой кислоты-мишени перед амплификацией. В патенте '447 предпринята попытка усовершенствования более ранней разработки обеспечением анализа, имеющего улучшенную специфичность, посредством того, что зонд является одноцепочечным олигонуклеотидом, меченым интеркалирующимся флуорохромом, который должен быть интеркалирован в часть комплементарного связывания между нуклеиновой кислотой-мишенью и одноцепочечным олигонуклеотидным зондом.
В ведущемся поиске на более чувствительные, точные и быстрые способы анализа одна исследовательская группа разработала тест, предусматривающий анализ действий электрического поля на интенсивность флуоресценции гибридизационных дуплексов нуклеиновых кислот. См. патентные заявки США с номерами 08/807901 и 08/870370, соответственно поданные 27 февраля 1997 года и 6 июня 1997 года. Эти исследователи показали, что интенсивность флуоресценции дуплекса с одной ошибочно спаренной парой оснований отличается от интенсивности флуоресценции точно спаренного дуплекса. Таким образом, эта заявка имеет целью описание способа для обнаружения нуклеотидной последовательности, в котором электрическое поле прилагают к жидкой среде перед стадией детектирования или одновременно со стадией детектирования и изменение интенсивности флуоресцентной эмиссии как функции электрического поля детектируют как указания, гибридизуется ли данный зонд с полностью комплементарной нуклеотидной последовательностью или неполностью комплементарной нуклеотидной последовательностью.
Несмотря на предыдущие разработки, в данной области продолжает существовать потребность в простом, высокочувствительном, эффективном и быстром способе для анализа взаимодействия между нуклеиновыми кислотами и/или аналогами нуклеиновых кислот.
Все цитируемые здесь ссылки включены здесь в качестве ссылки в их полном виде.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает способ анализа последовательность-специфической гибридизации, предусматривающий:
обеспечение мишени, содержащей, по меньшей мере, одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
приложение первого стимула к указанной тест-пробе для обеспечения первой стимулированной тест-пробы;
детектирование первого сигнала из указанной первой стимулированной тест-пробы, где указанный первый сигнал коррелирует с аффинностью связывания между указанным зондом и указанной мишенью;
калибрование указанного первого сигнала против контрольного сигнала, проявляемого контрольной пробой, содержащей, по меньшей мере, один контрольный зонд, комбинированный с указанной мишенью, где относительно указанной мишени каждый из указанного зонда и указанного, по меньшей мере, одного контрольного зонда является отличающимся членом, выбранным из группы, состоящей из точного спаривания, ошибочного спаривания одного основания, ошибочного спаривания двух оснований ошибочного спаривания трех оснований, делеции одного основания, делеции двух оснований и делеции трех оснований; и
определение на основании этого калибрования первого определения степени спаривания между указанным зондом и указанной мишенью;
приложение второго стимула к указанной первой стимулированной тест-пробе для обеспечения второй стимулированной тест-пробы; и
детектирование второго сигнала из указанной второй стимулированной тест-пробы, где указанный второй сигнал коррелирует с аффинностью связывания между указанным зондом и указанной мишенью;
определение на основании указанного детектирования указанного второго сигнала второго определения указанной степени спаривания между указанным зондом и указанной мишенью; и
сравнение указанного первого определения и указанного второго определения.
Обеспечен также другой способ анализа гибридизации, предусматривающий:
обеспечение мишени, содержащей, по меньшей мере, одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
измерение первого сигнала первого состояния указанной тест-пробы для обеспечения первичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанный первый сигнал коррелирует с гибридизацей между указанным зондом и указанной мишенью;
измерение второго сигнала второго состояния указанной тест-пробы для обеспечения вторичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанный второй сигнал коррелирует с гибридизацей между указанным зондом и указанной мишенью при условии, что, когда указанное первое состояние и указанное второе состояние являются одинаковыми, стимул прилагают к указанной тест-пробе после измерения указанного первого сигнала и перед измерением указанного второго сигнала, где указанный стимул значимо влияет на неполностью комплементарную гибридизацию между указанным зондом и указанной мишенью и не влияет значимо на полностью комплементарную гибридизацию между указанным зондом и указанной мишенью; и
сравнение указанного первичного определения и указанного вторичного определения для оценки, требует ли какое-либо несовпадение между ними повторного тестирования.
Кроме того, данное изобретение обеспечивает и другой способ анализа гибридизации, предусматривающий:
обеспечение мишени, содержащей, по меньшей мере, одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
измерение перед подачей электрического тока интенсивности флуоресценции указанной тест-пробы для обеспечения первичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанная интенсивность флуоресценции перед подачей электрического тока коррелирует с гибридизацией между указанным зондом и указанной мишенью;
приложение напряжения к указанной тест-пробе;
измерение после подачи электрического тока интенсивности флуоресценции указанной тест-пробы, во время или после указанного приложения напряжения, для обеспечения вторичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанная интенсивность флуоресценции после подачи электрического тока коррелирует с гибридизацией между указанным зондом и указанной мишенью; и
сравнение указанного первичного определения и указанного вторичного определения для оценки, требует ли какое-либо несовпадение между ними повторного тестирования.
Обеспечен также способ анализа последовательность-специфической гибридизации, предусматривающий:
обеспечение мишени, содержащей, по меньшей мере, одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
приложение электрического напряжения к указанной тест-пробе;
детектирование сигнала указанной тест-пробы во время или после указанного приложения указанного электрического напряжения, где указанный сигнал коррелирует с аффинностью связывания между указанным зондом и указанной мишенью;
калибрование указанного сигнала против контрольного сигнала, проявляемого контрольной пробой, содержащей, по меньшей мере, один конкретный зонд, комбинированный с указанной мишенью, где относительно указанной мишени каждый из указанного зонда и указанного, по меньшей мере, одного контрольного зонда является отличающимся членом, выбранным из группы, состоящей из точного спаривания, ошибочного спаривания одного основания, ошибочного спаривания двух оснований, ошибочного спаривания трех оснований, делеции одного основания, делеции двух оснований и делеции трех оснований; и
определение на основании этого калибрования степени спаривания между указанным зондом и указанной мишенью.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Данное изобретение будет описано вместе со следующими чертежами, в которых одинаковые цифры обозначают одинаковые элементы и в которых:
фиг.1А и 1В являются графиками тока как функции времени и комплементарности;
фиг.1С и 1D являются графиками тока как функции температуры и комплементарности;
фиг.2А, 2В, 2С, 3А и 3В являются графиками тока как функции температуры, комплементарности и дополнительных факторов;
фиг.4 является графиком тока как функции времени и комплементарности; и
фиг.5А, 5В, 5С и 6 являются спектрами интенсивности флуоресценции.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ
Данное изобретение обеспечивает быстрый, чувствительный, не приносящий вреда окружающей среде и безопасный способ для анализа связывания между мишенью и зондом, в котором мишень содержит последовательность нуклеиновой кислоты или последовательность аналога нуклеиновой кислоты и зонд содержит последовательность нуклеиновой кислоты или последовательность аналога нуклеиновой кислоты.
В противоположность некоторым анализам прежнего уровня техники, данное изобретение не только детектирует присутствие гибридизации, но также обеспечивает качественную и количественную информацию в отношении характера гибридизации между зондом и мишенью. Так, данное изобретение позволяет практикующему специалисту различать точное спаривание, ошибочное спаривание одной пары оснований, ошибочное спаривание двух пар оснований, ошибочное спаривание трех пар оснований, делецию одной пары оснований, делецию двух пар оснований и делецию трех пар оснований.
Варианты данного изобретения включают в себя калибрование измеренного сигнала (например, электрического тока и/или интенсивности флуоресценции) для первой смеси зонд-мишень против сигнала того же самого типа, проявляемого другими зондами, комбинированными с той же самой мишенью, где каждый из других зондов отличается от первого зонда, по меньшей мере, одним основанием.
В некоторых вариантах низкое напряжение подают к пробе перед измерением указанного сигнала или одновременно с измерением указанного сигнала. Обычно напряжение выбирают таким образом, что оно является достаточно высоким для дестабилизации дефектно спаренных партнеров гибридизации, но не настолько высоким, чтобы дестабилизировать точно спаренные партнеры гибридизации. В некоторых предпочтительных вариантах напряжение равно от приблизительно 1 В - до приблизительно 20 В.
Может быть получена калибровочная кривая, где величина измеренного сигнала (например, электрического тока и/или интенсивности флуоресценции) является функцией аффинности связывания между мишенью и зондом. Поскольку аффинность связывания между мишенью и множеством различных зондов меняется в зависимости от числа ошибочно спаренных оснований, характера ошибочного спаривания (A-G против А-С против T-G против Т-С, и т.д.), местоположения этого ошибочного спаривания (спаривании) в гибридизационном комплексе и т.д., данный анализ по изобретению может быть использован для секвенирования мишени.
Измеряемым сигналом может быть, например, электропроводность. В таких вариантах аффинность связывания между зондом и мишенью прямо коррелирует с величиной этого сигнала. То есть электропроводность увеличивается вместе со степенью спаривания между зондом и мишенью предпочтительно на протяжении диапазона, включающего 0-2 ошибочных спаривания и/или делеций, более предпочтительно на протяжении диапазона, включающего 0-3 ошибочных спаривания и/или делеций.
В других вариантах измеряемым сигналом может быть интенсивность флуоресценции флуорофора, включенного в тест-пробу. В таких вариантах аффинность связывания между зондом и мишенью может быть прямо или обратно коррелированной с интенсивностью в зависимости от гибридизации сигналов флуорофора через гашение сигнала или усиление сигнала. Так, интенсивность флуоресценции, генерируемой интеркалирующими (внедряющимися) агентами, является прямо коррелированной с аффинностью связывания зонд/мишень, тогда как интенсивность флуоресценции вариантов с использованием неинтеркалирующихся флуорофоров, ковалентно связанных с зондом, обратно коррелирует с аффинностью связывания зонд/мишень. Интенсивность флуоресценции увеличивается (или уменьшается для неинтеркаляторов) с увеличением степени спаривания между зондом и мишенью, предпочтительно на протяжении диапазона, включающего 0-2 ошибочных спаривания и/или делеции, более предпочтительно на протяжении диапазона, включающего 0-3 ошибочных спаривания и/или делеции.
Хотя авторы изобретения ранее описали преимущества анализов интенсивности флуоресценции для гибридизации (см. патентную заявку США № 09/468679, поданную 21 декабря 1999 года), приложение электрического поля к пробе, по-видимому, увеличивает разрешающую способность анализа, как показано в примере 6 ниже.
Кроме того, в особенно предпочтительном варианте данного изобретения анализ предусматривает измерение, по меньшей мере, двух сигналов пробы. Первый сигнал является предпочтительно интенсивностью флуоресценции, а второй сигнал выбран предпочтительно из нескольких измерений электропроводности (или наоборот).
В предпочтительных вариантах со множественными измерениями первый сигнал может быть одинаковым или отличающимся в сравнении со вторым сигналом. Когда первый и второй измеряемые сигналы являются одинаковыми, второй сигнал может быть откалиброван против первого сигнала и/или против того же самого контрольного сигнала (сигналов), используемых для калибрования первого сигнала. Кроме того, к тест-пробе подают изменяющий состояние стимул, после того как измеряют первый сигнал и перед тем как измеряют второй сигнал. Этот стимул предпочтительно является достаточным для значимого действия на дефектную комплементарную гибридизацию между зондом и мишенью и недостаточным для значимого действия на точную комплементарную гибридизацию между зондом и мишенью.
Например, в особенно предпочтительном варианте данного изобретения первым измеряемым сигналом является измеренная перед подачей электрического тока интенсивность флуоресценции (т.е. интенсивность, измеряемая перед подачей изменяющего состояние напряжения), а вторым измеряемым сигналом является измеренная после подачи электрического тока интенсивность флуоресценции (т.е. интенсивность, измеряемая во время или после изменяющего состояние напряжения, подаваемого к тест-пробе).
Дополнительные измерения в предыдущих вариантах увеличивают надежность данного анализа и делают возможным немедленное повторное тестирование сомнительных результатов. Несовместимые результаты, получаемые, по меньшей мере, в двух измерениях, обычно будут требовать повторного тестирования.
Данное изобретение делает возможным количественное определение аффинности связывания между зондом и мишенью. Такая информация может быть ценной для различных применений, в том числе конструирования антисмысловых лекарственных средств с оптимизированными характеристиками связывания.
В противоположность способам прежнего уровня техники анализ данного изобретения является предпочтительно гомогенным. Анализ может проводиться без отделения комплекса зонд-мишень от свободного зонда перед обнаружением величины измеряемого сигнала. Анализ не требует стадии разделения в геле, делая тем самым возможным большое увеличение производительности тестирования. Количественные анализы являются простыми и точными. В результате анализ связывания экономит массу времени и расходов и может быть легко автоматизирован. Кроме того, он позволяет быстро определять переменные связывания, такие как буфер, рН, концентрация ионов, температура, время инкубирования, относительные концентрации последовательностей зонда и мишени, концентрация интеркалятора, длина последовательностей-мишеней, длина последовательностей зондов и возможные требования кофакторов.
Данный анализ можно проводить, например, в растворе в лунке, на непроницаемой поверхности или на биочипе (наборе матриц).
Кроме того, анализ по данному изобретению предпочтительно проводят без обеспечения агента гашения сигнала на мишени или на зонде.
Предпочтительные варианты данного изобретения специфически детектируют триплексную гибридизацию между зондом и двухцепочечной мишенью, устраняя таким образом необходимость денатурации мишени. Хотя было известно, что ПНК-зонды образуют триплексы с определенными классами мишеней (см., например, Egholm et al., 365 Nature 566 (1993) и Tomas et al., 118 J. Am. Chem. Soc. 5544 (1996)), авторы изобретения были удивлены, что они были способны специфически анализировать триплексы, образуемые между одноцепочечными нуклеиновокислотными зондами (например, оцДНК (ssDNA) и РНК) и такими мишенями, как двухцепочечная нуклеиновая кислота (например, дцДНК (dsDNA)). Образование и/или стабилизация триплекса усиливается присутствием интеркалирующего агента в тестируемой пробе.
Подходящие зонды для применения в анализе данного изобретения включают в себя, например, оцДНК, РНК, ПНК и другие аналоги нуклеиновых кислот, имеющие незаряженные или частично заряженные скелеты. Хотя в некоторых вариантах предпочтительными являются антипараллельные зонды, ПНК-зонды также могут быть параллельными. Последовательности зондов, имеющие любую длину от 8 до 20 оснований, являются предпочтительными, так как эта длина находится в диапазоне, в котором находятся самые малые уникальные ДНК-последовательности прокариот и эукариот. Зонды из 12-18 оснований являются особенно предпочтительными, так как эта длина является длиной наименьших уникальных последовательностей в геноме человека. В вариантах наиболее предпочтительными являются зонды из 6-30 оснований. Однако множество более коротких зондов может быть использовано для обнаружения нуклеотидной последовательности, имеющей в ней множество неуникальных последовательностей-мишеней, которые комбинируются для уникальной идентификации данной нуклеотидной последовательности. Длина зонда может быть выбрана таким образом, что она совпадает с длиной мишени.
Данное изобретение не требует применения радиоактивных зондов, которые являются опасными, применение которых является трудоемким и занимающим много времени, и которые требуют постоянной регенерации. Зонды данного изобретения предпочтительно являются безопасными для применения и стабильными в течение нескольких лет. Таким образом, зонды могут быть получены или заказаны в больших количества и могут храниться.
Предпочтительно зонд и мишень являются немечеными, но в альтернативных вариантах используют интеркалирующий агент, ковалентно связанный с зондом. В таких вариантах интеркалирующий агент предпочтительно связан с зондом на любой стороне.
В других вариантах интеркалирующий агент не является ковалентно связанным с зондом, хотя он может вставляться между зондом и мишенью во время анализа, в некотором смысле связывая этот зонд нековалентным образом.
Предпочтительные интеркалирующие агенты для применения в данном изобретении включают в себя, например, YOYO-1, TOTO-1, бромид этидия, гомодимер-1 этидия, гомодимер-2 этидия и акридин. Обычно интеркалирующий агент является частью молекулы, которая способна интеркалироваться между цепями дуплексного и/или триплексного комплекса нуклеиновых кислот. В предпочтительных вариантах интеркалирующий агент (или его компонент) является по существу нефлуоресцентным в отсутствие нуклеиновых кислот и флуоресцирует при интеркаляции и возбуждении излучением подходящей длины волны, проявляя 100-кратное - 10000-кратное усиление флуоресценции при интеркаляции в дуплексном или триплексном комплексе нуклеиновых кислот.
В альтернативных вариантах интеркалирующий агент может проявлять смещение в длине волны флуоресценции при интеркаляции и возбуждении излучением подходящей длины волны. Точная длина волны флуоресценции может зависеть от структуры нуклеиновых кислоты, которая является интеркалированной, например ДНК vs. РНК, дуплекс vs. триплекса и т.д.
Длину волны возбуждения выбирают (рутинным экспериментированием и/или общепринятым путем) таким образом, чтобы она соответствовала максимуму возбуждения для используемого флуорофора, и она равна предпочтительно 200-1000 нм. Интеркалирующие агенты предпочтительно выбраны таким образом, что они имеют длину волны испускания (эмиссии) 200-1000 нм. В предпочтительном варианте лазер на ионах аргона используют для облучения флуорофора светом, имеющим длину волны в диапазоне 400-540 нм, и флуоресцентную эмиссию детектируют в диапазоне 500-750 нм.
Анализ данного изобретения может выполняться в широком диапазоне температур, таком как, например, от 5 до 85°С. Некоторые анализы прежнего уровня техники требуют повышенных температур, что добавляет расходы и замедляет анализ. С другой стороны, данное изобретение может проводиться при комнатной температуре или более низкой температуре (например, при температурах ниже 25°С).
Анализ по данному изобретению является чрезвычайно чувствительным, устраняя тем самым необходимость проведения ПЦР-амплификации мишени. Например, по меньшей мере, в вариантах с интенсивностью флуоресценции можно анализировать тест-пробу, имеющую объем около 20 микролитров, которая содержит около 10 фемтомоль мишени и около 10 фемтомоль зонда. Варианты данного изобретения являются достаточно чувствительными для анализа мишеней при концентрации 5×10-9 М, предпочтительно при концентрации не более чем 5×10-10 М. Варианты данного изобретения являются достаточно чувствительными для применения зондов при концентрации 5×10-9 М, предпочтительно при концентрации не более чем 5×10-10 М.
Измерения электропроводности могут различать пробы, имеющие мало, как зонда приблизительно 1 пмоль, так и мишени 1 пмоль в 40 мкл. Уменьшение объема пробы позволит использовать даже меньшие количества зонда и мишени.
Нечего говорить о том, что вышеуказанные величины позволяют предполагать, что данный способ может детектировать более высокие концентрации.
Большой диапазон концентраций интеркалатора устойчив при каждой концентрации тестируемых зонда и мишени. Например, при гибридизаци 5×10-10 М зонда и 5×1-10 М мишени оптимальная концентрация интеркалятора YOYO-1 находится в диапазоне от 26 до 2,5 нМ. При концентрации 5×10-8 М как зонда, так и мишени предпочтительный диапазон концентраций YOYO-1 составляет 1000-100 нМ.
Этот анализ является достаточно чувствительным для различения комплекса зонд-мишень с ошибочным спариванием одной пары оснований от комплекса зонд-мишень с ошибочным спариванием двух оснований, предпочтительно комплекса зонд-мишень с ошибочным спариванием двух оснований от комплекса зонд-мишень с ошибочным спариванием трех оснований. Конечно этот анализ является достаточно чувствительным для отличения комплекса зонд-мишень с точным спариванием от любого из вышеуказанных комплексов с ошибочным спариванием.
Гибридизационной средой может быть любая общепринятая среда, о которой известно, что она является пригодной для сохранения нуклеотидов. См., например, Sambrook et al., "Molecular Cloning. A Lab Manual, vol. 2 (1989). Например, жидкая среда может содержать нуклеотиды, воду, буферы и стандартные концентрации солей.
Гибридизация между комплементарными основаниями происходит при большом разнообразии условий, имеющих вариации в температуре, концентрации соли, электростатической силе и составе буфера. Примеры этих условий и способов их применения известны в данной области.
Предпочтительно, чтобы гибридизационные комплексы образовывались при температуре около 15°С - до около 25°С в течение от приблизительно 1 минуты - до приблизительно 5 минут. Более продолжительные времена реакции не требуются, но инкубирование в течение нескольких часов не будет оказывать вредного влияния на гибридизационные комплексы.
Можно (хотя и необязательно, в частности, для вариантов, содержащих интеркалирующий агент) облегчить гибридизацию в растворе посредством использования определенных реагентов. Предпочтительные примеры этих реагентов включают в себя одноцепочечные связывающие белки, такие как белок Rec A, белок гена 32 Т4, одноцепочечный связывающий белок Е. coli, основные или минорные связывающие белки бороздки нуклеиновых кислот, двухвалентные ионы, поливалентные ионы, виологен и интеркалирующие вещества, такие как бромид этидия, актиномицин D, псорален и ангелицин. Такие облегчающие реагенты могут оказаться применимыми при отклоняющихся от нормы уровнях рН или крайне высоких температурах.
Анализ по данному изобретению может быть использован, например, для идентификации доступных районов в уложенных нуклеотидных последовательностях, для определения числа ошибочно спаренных пар оснований в гибридизацонном комплексе и для картирования геномов.
В вариантах, где интенсивность флуоресценции детектируют с использованием интеркалирующего агента, интенсивность увеличивается с увеличением аффинности связывания между зондом и мишенью. В вариантах, где интенсивность флуоресценции детектируют с использованием неинтеркалирующего флуорофора, интенсивность уменьшается по мере увеличения аффинности связывания между зондом и мишенью. Независимо от того, интеркалируется флуорофор или нет, данный способ не требует измерения поляризации флуоресценции в противоположность способам флуоресцентной анизотропии.
Данное изобретение будет иллюстрировано более подробно со ссылкой на следующие примеры, но должно быть понятно, что данное изобретение не должно считаться ограниченным ими.
ПРИМЕРЫ
Пример 1
Смысловые и антисмысловые 50-мерные оцДНК-последовательности-мишени, полученные из экзона 10 гена муковисцидоза (Nature 380, 207 (1996)), синтезировали на ДНК-синтезаторе (Expedite 8909, PerSeptive Biosystems) и очищали при помощи ВЖХ. Эквимолярные количества комплементарных олигонуклеотидов денатурировали при 95°С в течение 10 мин и давали отжигаться постепенно по мере снижения температуры до 21°С на протяжении 1,5 часов. Двухцепочечные ДНК (дцДНК)-олигонуклеотиды растворяли в ddH2О при концентрации 1 пмоль/мкл.
Последовательность для смысловой цепи ДНК-мишени дикого типа (SEQ ID NО:1): 5'-TGG CAC CAT TAA AGA AAA TAT CAT CTT TGG TGT TTC СТА TGA TGA ATA TA-3'.
Последовательность для антисмысловой цепи ДНК-мишени дикого типа (SEQ ID NО:1): 5'-TAT ATT CAT CAT AGG AAA CAC CAA AGA TGA TAT TTT CTT TAA TGG TGC CA-3'.
Температура плавления (Tm) дцДНК (SEQ ID NO:1) равна 65,2°C.
SEQ ID NO: 2 была 50-мерной мутантной дцДНК-последовательностью-мишенью, идентичной ДНК-мишени дикого типа (SEQ ID NО:1), за исключением мутации одной пары оснований (подчеркнутой) в аминокислотном положении 507, в котором CAT последовательности дикого типа был заменен на CGT.
Последовательность для смысловой цепи SEQ ID NO:2; 5'-TGG CAC CAT TAA AGA AAA TAT CGT CTT TGG TGT TTC СТА TGA TGA ATA TA-3'.
Последовательность для антисмысловой цепи SEQ ID NO: 2: 5'-TAT ATT CAT CAT AGG AAA CAC CAA AGA CGA TAT TTT CTT TAA TGG TGC CA-3'.
Температура плавления (Тm) дцДНК (SEQ ID NO: 2) равна 66,0°С.
SEQ ID NО:3 была 50-мерной мутантной дцДНК-последовательностью-мишенью, идентичной ДНК-мишени дикого типа (SEQ ID NO:1), за исключением мутации последовательных двух пар оснований (подчеркнутых) в аминокислотных положениях 506 и 507, в которых CAT последовательности дикого типа был заменен на ACT.
Последовательность для смысловой цепи SEQ ID NО:3: 5'-TGG CAC CAT TAA AGA AAA TAT ACT CTT TGG TGT TTC СТА TGA TGA ATA TA-3'.
Последовательность для антисмысловой цепи SEQ ID NО:3: 5'-TAT ATT CAT CAT AGG AAA CAC CAA AGA ACT TAT TTT CTT TAA TGG TGC CA-3'.
Температура плавления (Tm) вцДНК (SEQ ID NО:3) равна 65,2°C.
ПНК-зонды, использованные в этих примерах, синтезировали, очищали при помощи ВЖХ и подтверждали масс-спектрометрией с использованием Commonwealth Biotechnologis, Inc. (Richmond, VA, USA). ПНК-зонды сначала растворяли в 0,1% ТФУ (трифторуксусной кислоте) до концентрации 10 мг/мл и затем разбавляли до 1 мг/мл добавлением ddH2O. Конечные исходные растворы ПНК готовили в ddH2O в концентрации 1 пмоль/мкл.
Зонд № 1 был 15-мерным антипараллельным ПНК-зондом, сконструированным, чтобы быть полностью комплементарным 15-нуклеотидному сегменту смысловой цепи 50-мерной ДНК-мишени дикого типа (SEQ ID NO:1), перекрывающему аминокислотные положения 505-510 (Nature 380, 207 (1996)). Этот зонд имел следующую структуру (SEQ ID N0:8):
5'-Н-САС САА AGA TGA TAT-Lys-CONH2-3'
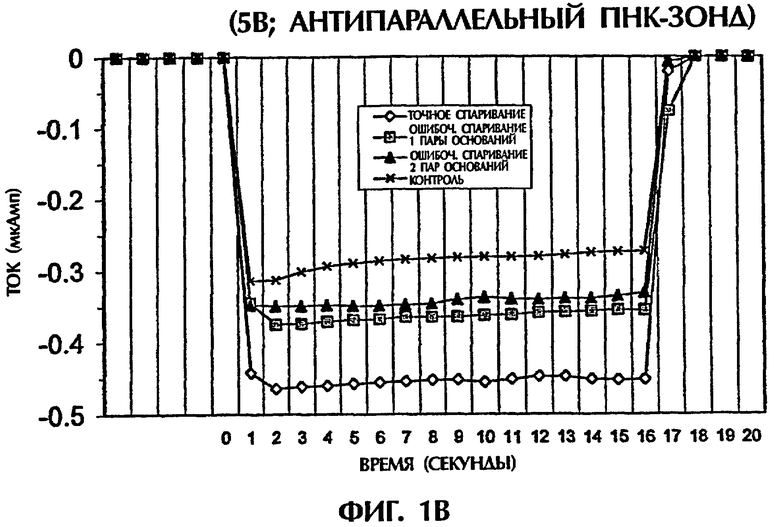
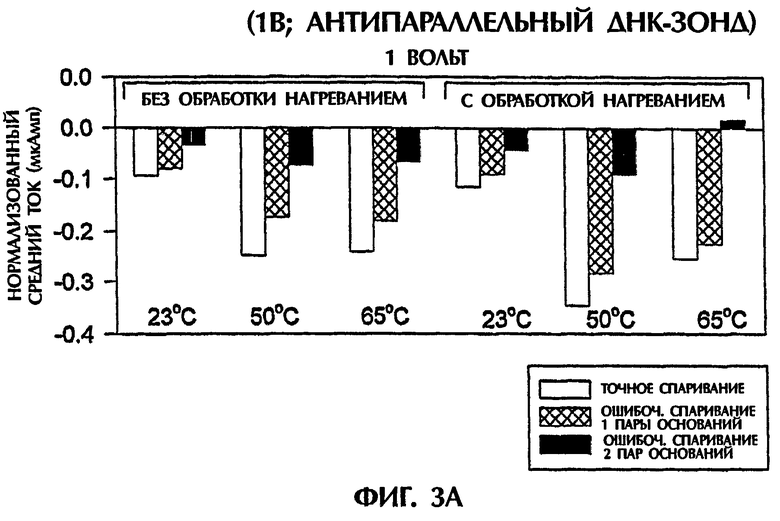
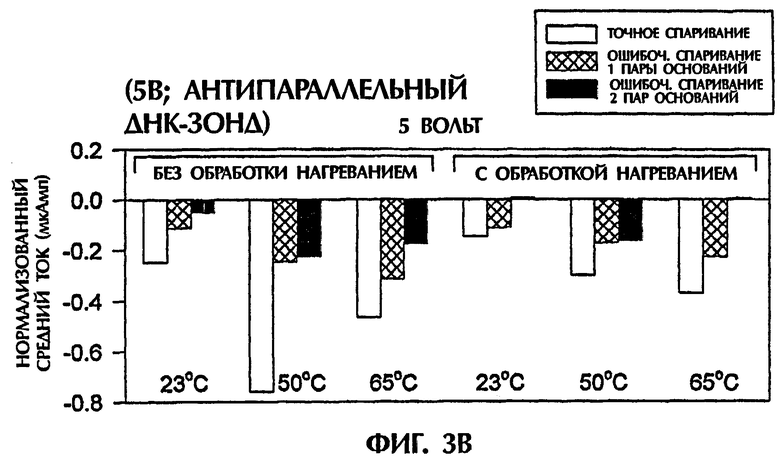
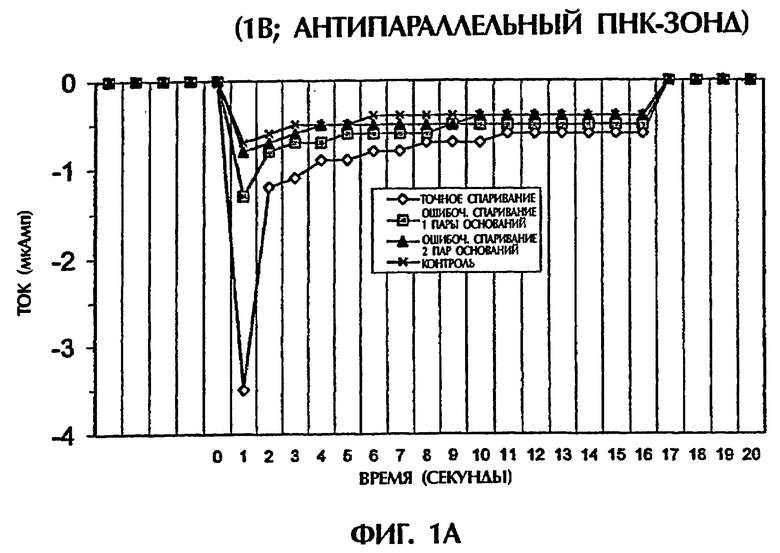
Гибридизационная реакционная смесь (80 мкл) содержала следующее: 2 пмоль дцДНК-мишени, 2 пмоль ПНК-зонда, 0,5Х ТВЕ и 250 нМ интеркалятор ДНК YOYO-1 (Molecular Probes, Eugene, OR, USA). Пробы помещали в кювету 3 мм из кварцевого стекла и подвергали действию 1 или 5 вольт постоянного тока (DC) в течение 15 секунд. Амперометрический анализ состоял из мониторинга тока при подаче напряжения к раствору. Температурный датчик помещали в каждый раствор для измерения температуры во время амперометрической оценки. При 1 В наблюдали постоянный пик во время первых 2 секунд подачи электрического тока. Ток резко уменьшался на протяжении следующих 13 секунд. Эксперименты с подачей 5 вольт давали появление токов, которые оставались относительно стабильными на протяжении всего периода подачи электрического тока (15 секунд).
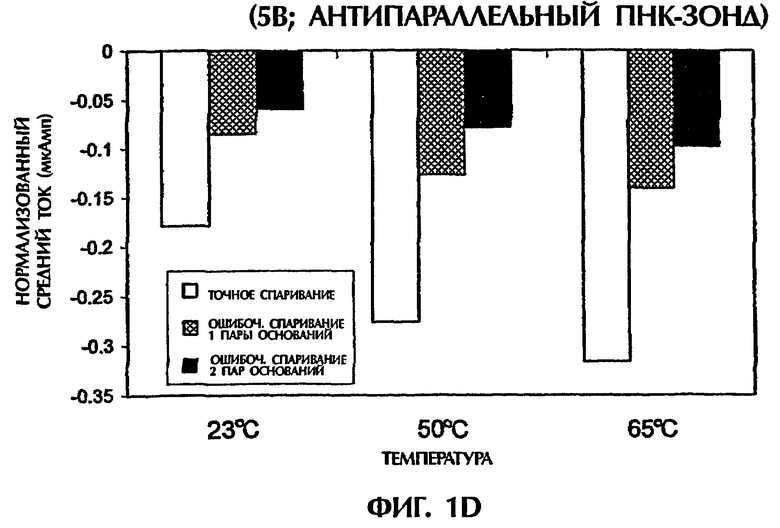
Проводили ряд экспериментов, где величины электропроводности наблюдали, когда не присутствовали ни ДНК, ни ПНК (контроль), или, когда SEQ ID NO:1 дикого типа, мутантная SEQ ID NO: 2 или мутантная SEQ ID NО:3 взаимодействовали с антипараллельным ПНК-зондом № 1. Фигуры 1А и 1В дают графики данных, полученных для электропроводности в отдельных экспериментах. Фигура 1А изображает результаты подачи электрического тока 1 В, а фигура 1В - подачу электрического тока 5 В. Гибридные триплексы двухцепочечная ДНК:ПНК, состоящие из точно комплементарных последовательностей (SEQ ID NO:1 + зонд № 1), делали возможной максимальную интеркаляцию YOYO-1, давая наивысшие величины электропроводности (изображенные на этих фигурах в виде отрицательных величин электропроводности) на протяжении всех 15 секунд подачи 1 В. Нормализованная максимальная электропроводность для триплексной гибридизации антипараллельного ПНК-зонда с дцДНК с 1 ошибочно спаренной парой оснований (SEQ ID NO: 2 + зонд № 1) и с дцДНК с 2 ошибочно спаренными парами оснований была соответственно на 79% и 96% более низкой, чем эта электропроводность, наблюдаемая с точно спаренным триплексным гибридом дцДНК:ПНК (SEQ ID NO:1 + зонд № 1) во время первой секунды подачи напряжения (фиг.1А). Сходные процентные уменьшения в электропроводности между точно комплементарными триплексами и триплексами, содержащими ошибочные спаривания пар оснований, получали, когда величины электропроводности на протяжении всех 15 секунд подачи напряжения усреднялись. На фиг.1А гибриды дцДНК:ПНК с 1 ошибочно спаренной парой оснований и 2 ошибочно спаренными парами оснований приводили к величинам средней электропроводности, которые были на 65% и на 91% более низкими соответственно, чем эти величины для точно спаренного гибрида дцЦНК:ПНК. Все эксперименты, изображенные на фигуре 1А, проводили при комнатной температуре (23°С). По мере увеличения степени ошибочного спаривания между зондом и двухцепочечной мишенью уровень интеркаляции YOYO-1 снижался, и уровень электропроводности уменьшался. Эти взаимосвязи наблюдали также при повторении описанных выше экспериментов и подаче более высокого напряжения (5 В). Во время подачи 5 В величины нормализованной средней электропроводности для триплекса дцДНК: ПНК с 1 ошибочно спаренной парой оснований (SEQ ID NO: 2 + зонд № 1) и триплекса дцДНК: ПНК с 2 ошибочно спаренными парами оснований (SEQ ID NО:3 + зонд № 1) были соответственно на 52% и 67% более низкими, чем эта величина, наблюдаемая для точно спаренного триплекса дцДНК: ПНК (SEQ ID NО:3 + зонд № 1) (фиг.1В).
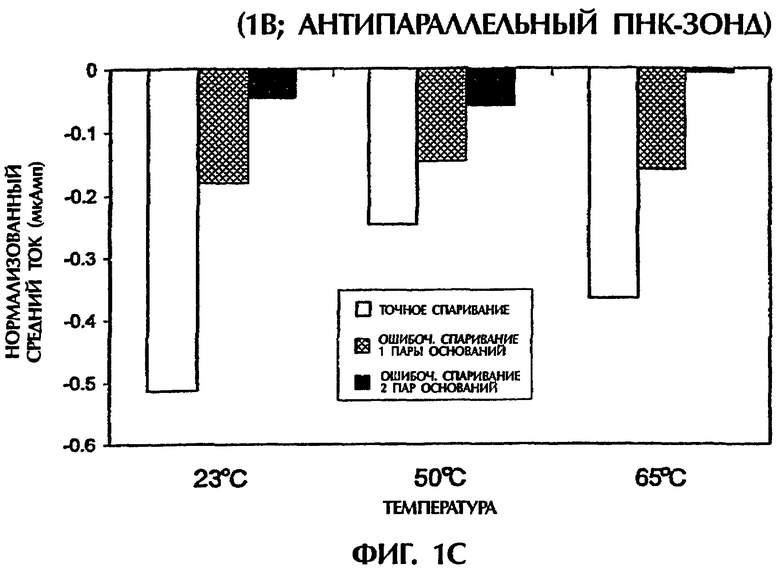
При повторении экспериментов с температурой, увеличенной до 50°С и 65°С, наблюдали сходные амперометрические величины. При 50°С подача 1 В в течение 15 секунд к точно спаренному триплексу дцДНК: ПНК (SEQ ID NO:1 + зонд № 1) давала средний ток -0,25 мкАмп в сравнении с величинами -0,15 мкАмп (40%-ное уменьшение) и -0,06 мкАмп (76%-ное уменьшение для триплекса дцДНК:ПНК с 1 ошибочно спаренной парой оснований (SEQ ID NO:2 + зонд № 1) и триплекса дцДНК:ПНК с 2 ошибочно спаренными парами оснований (SEQ ID NО:3 + зонд № 1) соответственно (фиг.1С). При 65°С сходные наблюдения отмечали, когда напряжение 1 В подавали в течение 15 секунд. Точно спаренные гибриды нуклеиновых кислот давали средний ток -0,37 мкАмп в сравнении с -0,16 мкАмп (57%-ное уменьшение) и -0,01 мкАмп (97% уменьшение) для гибридов с 1 ошибочно спаренной парой оснований и 2 ошибочно спаренными основаниями соответственно (фиг.1С). Применение 5 В при высоких температурах давало аналогичные результаты. Хотя эксперименты, выполненные при 50°С, генерировали средние токи -0,27 мАмп, -0,13 мАмп (52%-ное уменьшение) и -0,08 мАмп (70%-ное уменьшение) для точно спаренных гибридов, гибридов с 1 парой ошибочно спаренных оснований и гибридов с 2 парами ошибочно спаренных оснований соответственно, эксперименты, выполненные при 65°С, приводили к средним величинам тока -0,31 мАмп, -0,14 мАмп (55%-ное уменьшение) и -0,10 мАмп (68%-ное уменьшение) для тех же самых соответствующих групп (фиг.1D). Для всех предыдущих экспериментов дцДНК не денатурировали перед триплексной гибридизацией с антипараллельным ПНК-зондом № 1.
Подобные эксперименты выполняли при меняющихся температурах, после того как гибридизационные смеси нагревали до 65°С и сразу же давали им охлаждаться. После охлаждения до комнатной температуры (23°С) подача 1 В в течение 15 секунд к точно спаренной пробе (SEQ ID NO:1 + зонд № 1) давала средний ток -0,18 мкАмп. Посредством сравнения величины -0,06 мкАмп (67%-ное уменьшение) и 0,05 мкАмп (72% уменьшение) для триплексного гибрида с 1 ошибочно спаренной парой оснований (SEQ ID NO:2 + зонд № 1) и триплексным гибридом с 2 ошибочно спаренными основаниями (SEQ ID NО:3 + зонд № 1) соответственно наблюдали (данные не показаны). При охлаждении этих проб с 65°С до 50°С сходные наблюдения отмечали, когда 1 В подавали после этого в течение 15 секунд. Точно спаренная проба (SEQ ID NO:1 + зонд № 1) давала средний ток -0,23 мкАмп в сравнении с -0,11 мкАмп (52%-ное уменьшение) и -0,01 мкАмп (96%-ное уменьшение) наблюдали для проб с 1 ошибочно спаренной парой оснований и 2 ошибочно спаренными парами оснований соответственно (данные не показаны). Когда 5 В подавали после охлаждения до 23°С или 50°С, средний ток, генерированный в точно спаренном триплексном гибриде (SEQ ID NO:1 + зонд № 1), триплексном гибриде с 1 ошибочно спаренной парой оснований (SEQ ID NO:2 + зонд № 1) и триплексном гибриде с 2 ошибочно спаренными основаниями (SEQ ID NО:3 + зонд № 1), был: -0,15 мАмп, -0,09 мАмп (40%-ное уменьшение) и -0,07 мАмп (53%-ное уменьшение) соответственно при 23°С и -0,23 мАмп, -0,09 мАмп (61%-ное уменьшение) и 0,09 мАмп (61%-ное уменьшение) соответственно при 50°С (данные не показаны).
Предобработка гибридизационных смесей при 65°С (Тm 50-мерных дцДНК-последовательностей) с последующим охлаждением не влияла значимо на различие в электропроводности, наблюдаемое между точно комплементарными триплексами дцДНК: ПНК и теми, которые содержали 1 ошибочно спаренную пару оснований или 2 ошибочно спаренные пары оснований, при измерении непосредственно при 23°С или 50°С (без предварительного нагревания при 65°С) при применении антипараллельного ПНК-зонда. Ясно, что антипараллельный ПНК-зонд в присутствии интеркалятора ДНК YOYO-1 был способен образовывать триплексные структуры с дцДНК мишенями. Подача низких уровней электрического тока (таких как 1 В или 5 В) позволяла отличать точно спаренные последовательности триплекса дцДНК:ПНК от последовательностей, содержащих 1 ошибочно спаренную пару оснований или 2 ошибочно спаренные пары оснований, без предварительной денатурации последовательностей.
Пример 2
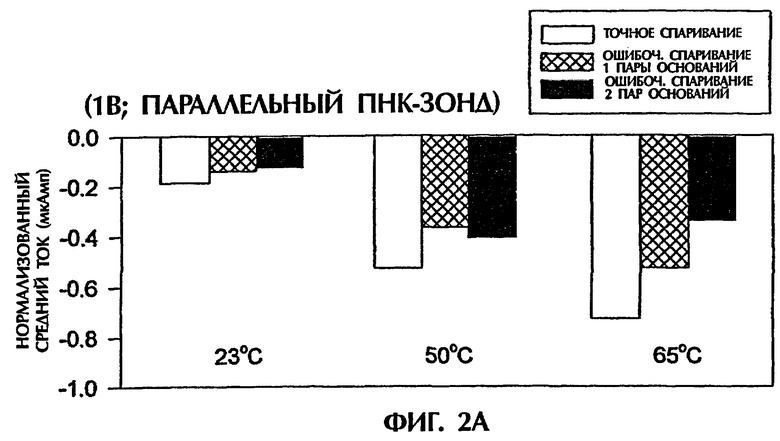
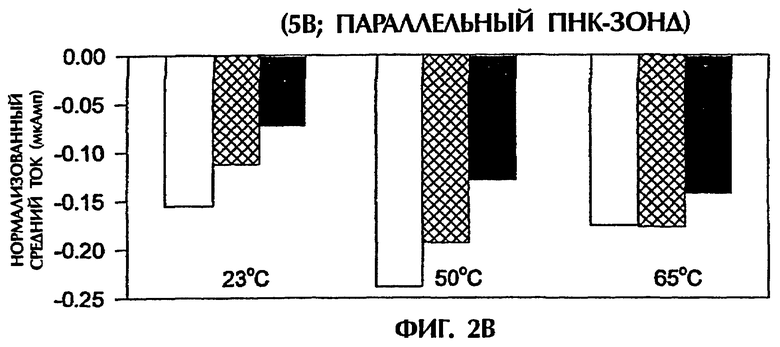
Фигура 2 демонстрирует, что амперометрический анализ по данному изобретению может также различать точно спаренные триплексные гибриды дцДНК: ПНК и триплексные гибриды, содержащие 1 ошибочно спаренную пару оснований или 2 ошибочно спаренные пары оснований при применении ПНК-зонда в параллельной ориентации относительно последовательности ДНК-мишени. Зонд № 2 был 15-мерным ПНК-зондом, идентичным в его последовательности зонду № 1, но был синтезирован для спаривания параллельной ориентации ДНК-мишени вместо общепринятой антипараллельной ориентации. Зонд № 2 имел следующую структуру (SEQ ID NO:9):
5'-Н-ТАТ AGT AGA AAC CAC-Lis-CONH2-3'
Эксперименты с условиями анализа, идентичными условиям, описанным в примере 1, проводили с единственным различием, что использовали зонд № 2 вместо зонда № 1. При приложении 1 В средний ток для триплекса дцДНК: ПНК (SEQ ID NO:2 + зонд № 2) с 1 ошибочно спаренной парой оснований или с последовательными 2 ошибочно спаренными парами оснований был соответственно на 25% и 32% более низким при 23°С, соответственно на 30% и 23% более низким при 50°С и соответственно на 28% и 53% более низким при 65°С, чем средний ток, наблюдаемый с точно спаренным триплексом дцДНК: ПНК (SEQ ID NO:1 + зонд № 2) при совпадающих температурах (фиг.2А).
Сходные результаты получали при подаче 5 В (вместо 1 В) в течение 15 секунд. Точно спаренные гибриды дцДНК:ПНК при 23°С, 50°С и 65°С генерировали средние токи -0,15 мАмп, -0,24 мАмп и -0,17 мАмп соответственно (фиг.2В). Неполностью комплементарные триплексы с 1 ошибочно спаренной парой оснований или с 2 ошибочно спаренными парами оснований давали средние токи, которые были на 27% меньше (-0,11 мАмп) и 53% меньше (-0,07 мАмп) соответственно при 23°С, на 21% меньше (-0,19 мАмп) и 46% меньше (-0,13 мАмп) соответственно при 50°С и неизменные (-0,17 мАмп) и на 18% меньше (-0,14 мАмп) соответственно при 65°С, чем средние токи, достигаемые точно спаренными гибридными пробами (фиг.2В).
Результаты, показанные на фигурах 2А и 2В, свидетельствуют, что при применении параллельного ПНК-зонда № 2 различия в электропроводности, получаемые между точно спаренными триплексами дцДНК:ПНК и триплексами, содержащими 1 ошибочно спаренную пару оснований или 2 ошибочно спаренных пар оснований, были менее драматичными, чем различия, достигаемые с антипараллельным ПНК-зондом № 1 (фиг.1).
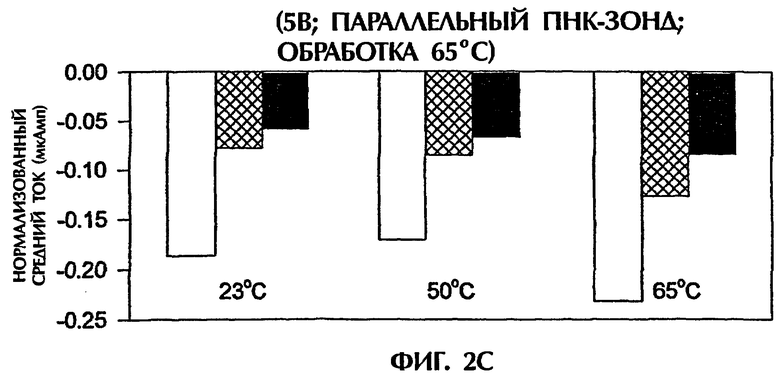
Однако эксперименты с использованием параллельного зонда № 2 и подачи 5 В, после того как эти пробы нагревали до 65°С и сразу же давали им охлаждаться, обнаружили амперометрические измерения, которые продемонстрировали усиленные различия передачи сигналов между точно спаренными триплексами дцДНК:ПНК и триплексами с 1 ошибочно спаренной парой оснований или с 2 ошибочно спаренными парами оснований (фиг.2С). Точно спаренные гибриды (SEQ ID NO:1 + зонд № 2), гибриды с 1 ошибочно спаренной парой оснований (SEQ ID NO:2 + зонд № 2) и гибриды с 2 ошибочно спаренными основаниями (SEQ ID NО:3 + зонд № 2) давали средние величины электропроводности -0,19 мАмп, -0,08 мАмп и -0,06 мАмп соответственно при 23°С, -0,17 мАмп, -0,09 мАмп и 0,07 мАмп соответственно при 50°С и -0,23 мАмп, -0,13 мАмп и -0,08 мАмп соответственно при 65°С. Это соответствовало уменьшениям электропроводности на 58% и 69% при 23°С, 47% и 59% при 50°С и 43% и 65% при 65°С для проб с 1 ошибочно спаренной парой оснований и 2 ошибочно спаренными основаниями соответственно в сравнении с величинами, достигаемыми точно комплементарными пробами (фиг.2С).
Таким образом, как антипараллельные, так и параллельные ПНК-зонды в амперометрическом анализе способны различать точно комплементарные дцДНК-мишени и неполностью комплементарные дцДНК мишени, содержащие мутации 1 пары оснований или 2 пар оснований.
Пример 3
Зонд № 3 был 15-мерным оцДНК-зондом, идентичным по последовательности и ориентации 15-мерному антипараллельному ПНК-зонду № 1 (SEQ ID NO: 8). Зонд № 3 имел следующую структуру:
5'-САС САА AGA TGA ТАТ-3'
Специфичность амперометрического анализа исследовали дополнительно посредством взаимодействия оцДНК-зонда № 3 с 50-мерными дцДНК-последовательностями-мишенями дикого типа и мутантными дцДНК-последовательностями-мишенями в отсутствие предварительной денатурации. Условия анализа были идентичны условиям, описанным в примере 1.
Усиленные интеркалятором ДНК YOYO-1, триплексы дцДНК:оцДНК образовывались между 30 и 65°С. При обработке 1 В точно спаренный ДНК-триплекс, состоящий из SEQ ID NO:1 + зонд № 3, давал наивысшие величины электропроводности (фиг.3А). В противоположность этому комбинации неполностью комплементарного зонда и мишени, генерирующие 1 ошибочно спаренную пару оснований (SEQ ID NO:2 + зонд № 3) и последовательные 2 ошибочно спаренные пары оснований (SEQ ID NО:3 + зонд № 3), приводили к средним величинам электропроводности, которые были на 14% и 64% ниже при 23°С, на 30% и 70% ниже при 50°С и на 25% и 72% ниже при 65°С соответственно, чем величины, наблюдаемые с точно комплементарными последовательностями при совпадающих температурах (фиг.3А). Подача более высокого напряжения (5 В) к этим пробам приводила к более высоким амперометрическим различиям, наблюдаемым между точно спаренными и ошибочно спаренными пробами, чем различия, полученные при 1 В, в частности, при более низких температурах. После обработки 5 В в течение 15 секунд средние токи для триплекса с 1 ошибочно спаренной парой оснований и триплекса с 2 ошибочно спаренными основаниями были на 54% и 78% более низкими соответственно при 23°С, на 68% и 70% более низкими соответственно при 50°С и на 33% и 61% более низкими соответственно при 65°С, чем средний ток, наблюдаемый с точно спаренным ДНК-триплексом при совпадающих температурах (фиг.3В).
В сходных экспериментах с подачей электрического тока гибридизационные смеси нагревали до 65°С и либо поддерживали при этой температуре, либо сразу же давали им охладиться до 50°С или 23°С перед подачей 1 В или 5 В. Обработка 1 В в течение 15 секунд точно спаренных ДНК-триплексных последовательностей (SEQ ID NО:1 + зонд № 3) давала наивысшие величины электропроводности при 23°С, 50°С и 65°С (фиг.3А). ДНК-триплексы, содержащие 1 ошибочно спаренную пару оснований (SEQ ID NO:2 + зонд № 3) или 2 ошибочно спаренные пары оснований (SEQ ID NО:3 + зонд № 3), были менее электропроводными на 21% и 63% соответственно при 23°С, на 18% и 74% соответственно при 50°С и на 12% и 106% соответственно при 65°С (фиг.3А). Подобным образом при подаче 5 В в течение 15 секунд предварительно нагретым пробам средние величины электропроводности для ДНК-триплексов с 1 ошибочно спаренной парой оснований и с 2 ошибочно спаренными парами оснований были уменьшены на 24% и 104% соответственно при 23°С, на 42% и 44% соответственно при 50°С и на 38% и 102% соответственно при 65°С по сравнению со средними величинами электропроводности, генерируемыми точно спаренными ДНК-триплексами (фиг.3В).
Наблюдение, что антипараллельный ПНК-зонд (фиг.1) и оцДНК-зонд (фиг.3) вели себя одинаковым образом в амперометрическом анализе, предполагало, что скелет сегмента нуклеиновой кислоты, используемого в качестве зонда, не был особенно важным. Присутствие YOYO-1 позволило дцДНК-мишеням и оцДНК-зонду образовать конформацию тройной спирали, способную генерировать различные электрические заряды в зависимости от уровня комплементарности последовательностей между мишенью и зондом в растворе. По мере увеличения степени ошибочного спаривания между зондом и мишенью уровень электропроводности уменьшался, что доказывало надежность амперометрического анализа при применении природного ДНК-зонда в отсутствие предварительной денатурации.
Пример 4
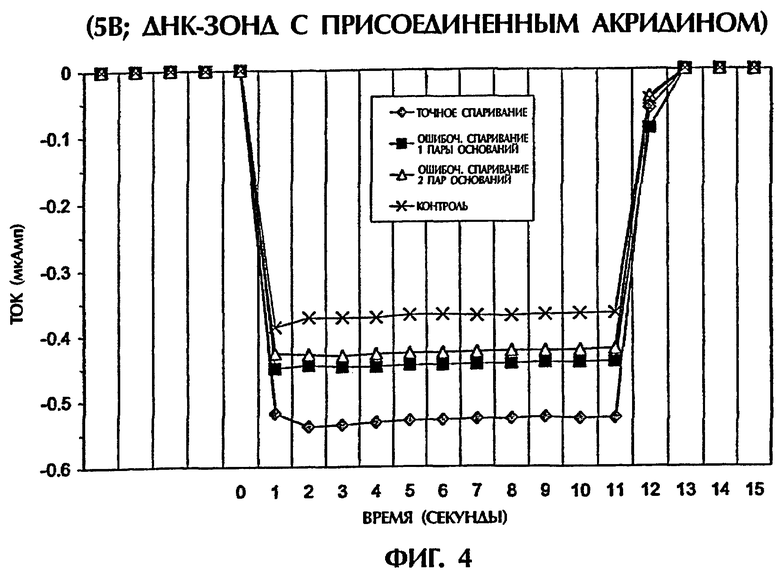
В амперометрических анализах, раскрытых в примерах 1-3, интеркалятор ДНК YOYO-1 добавляли к раствору, содержащему гибридизационные смеси. Интеркаляция при помощи YOYO-1 облегчала образование триплексов дцДНК:ПНК и триплексов дцДНК:оцДНК. Возможность использования интеркаляторной части, ковалентно связанной с оцДНК-зондом, в амперометрическом анализе оценивали в примере 4.
Акридин является альтернативным интеркалятором дцДНК, который также обладает способностью интеркалироваться в триплексные структуры нуклеиновых кислот, стабилизируя тем самым образование тройной спирали. См., например, Kukreti et al., "Extension of the range of DNA sequences available for triple helix formation: stabilization of mismatched triplexes by acridin-containing oligonucleotides." 25 Nucleic Acids Research 4264-4270 (1997). оцДНК-зонд, содержащий молекулу акридина (Glen Research, Sterling, VA, USA), ковалентно присоединенный на 3'-конце, синтезировали на ДНК-синтезаторе (Expedite 8909, PerSeptive Biosystems) и очищали при помощи ВЖХ.
Зонд № 4 был 15-мерным оцДНК-зондом, идентичным по последовательности и ориентации 15-мерному зонду № 3 (и, следовательно, также идентичным по последовательности и ориентации 15-мерному антипараллельному ПНК-зонду № 1 (SEQ ID NO:8)), но с добавлением акридиновой части в 3/-положении. Этот зонд имел следующую структуру:
5'-САС САА AGA TGA ТАТ-акридин-3'
Гибридизационная реакционная смесь (80 мкл) содержала следующее: 2 пмоль дцДНК-мишени, 2 пмоль оцДНК-зонда № 4 и 0,5Х ТВЕ. Пробы помещали в кварцевую кювету 3 мм и подвергали электрическому току 5 В в течение 11 секунд при 23°С. Ток и температуру подвергали мониторингу, как описано в примере 1.
Как показано на фиг.4, оцДНК-зонд № 4 был способен гибридизоваться с 50-мерной точно спаренной дцДНК-мишенью (SEQ ID NO:1) в результате стабильной интеркаляции ковалентно связанной акридиновой части молекулы, генерируя средний ток -0,53 мАмп. В сравнении с этим менее стабильные ДНК-триплексы, содержащие 1 ошибочно спаренную пару оснований (SEQ ID NO:2 + зонд № 4) или 2 ошибочно спаренные пары оснований (SEQ ID NО:3 + зонд № 4) давали средние токи, которые были на 52% и на 66% более низкими соответственно чем ток, достигаемый точно спаренным ДНК-триплексом при нормализации против контроля (зонда № 4 без ДНК-мишени) (фиг.4).
Таким образом, акридин, присоединенный к оцДНК-зонду, был равным образом эффективен, что и неприсоединенный YOYO-1, в образовании тройных ДНК-спиралей, которые генерировали различные токи в зависимости от уровня комплементарности последовательностей между мишенью и зондом в амперометрическом анализе.
Пример 5
Смысловые и антисмысловые 50-мерные оцДНК-последовательности-мишени, полученные из экзона 10 гена муковисцидоза, синтезировали, очищали и отжигали, как описано в примере 1. дцДНК-олигонуклеотиды растворяли в ddH2О в концентрации 1 пмоль/мкл.
SEQ ID NO: 4 была 15-мерной дцДНК последовательностью-мишенью, произведенной из SEQ ID NO:1, сконструированной таким образом, что она была полностью комплементарной зонду № 1.
Последовательность для смысловой цепи ДНК-мишени дикого типа (SEQ ID NО:4): 5'-ATA ТСА ТСТ TTG GTG-3'.
Последовательность для антисмысловой цепи ДНК-мишени дикого типа (SEQ ID NО:4): 5’-САС САА AGA TGA TAT -3'.
Температура плавления (Тm) дцДНК (SEQ ID NО:4) равна 40,0°С.
SEQ ID NO:5 была 50-мерной мутантной дцДНК-последовательностью-мишеныо, идентичной ДНК-мишени дикого типа (SEQ ID NО:4), за исключением мутации одной пары оснований (подчеркнутой), в которой ТТТ последовательности дикого типа был заменен на TAT.
Последовательность для смысловой цепи ДНК-мишени дикого типа (SEQ ID NO:5): 5'-АТА ТСА ТСТ ATG GTG-3'.
Последовательность для антисмысловой цепи ДНК-мишени дикого типа (SEQ ID NO:5): 5'-САС CAT AGA TGA TAT -3'.
Температура плавления (Tm) дцДНК (SEQ ID NO:5) равна 40,0°C.
SEQ ID NO:6 была 50-мерной мутантной дцДНК-последовательностью-мишенью, идентичной ДНК-мишени дикого типа (SEQ ID NO:4), за исключением мутации последовательных двух пар оснований (подчеркнутых), в которой АТС последовательности дикого типа был заменен на GGC.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:6):
5'-АТА TCG GCT TGG GTG-3'.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:6):
5'-САС САА AGC CGA TAT -3'.
Температура плавления (Тm) дцДНК (SEQ ID NO:6) равна 44,0°С.
SEQ ID NO:7 была 50-мерной мутантной дцДНК-последовательностью-мишенью, идентичной ДНК-мишени дикого типа (SEQ ID NO:4), за исключением разделенных мутаций трех пар оснований (подчеркнутых), где три мутации 1 пары оснований были разделены, каждая, 3 парами оснований. Последовательности АТС, ТСТ и TGG были изменены на АСС, ТАТ и TAG соответственно.
Последовательность для смысловой цепи мутантной ДНК-мишени (SEQ ID NO:7): 5'-АТА ССА ТАТ ТТА GTG-3'.
Последовательность для антисмысловой цепи мутантной ДНК-мишени (SEQ ID NO:7): 5'-САС ТАА АТА TGG ТАТ -3'.
Температура плавления (Тm) дцДНК (SEQ ID NO:6) равна 38,0°С.
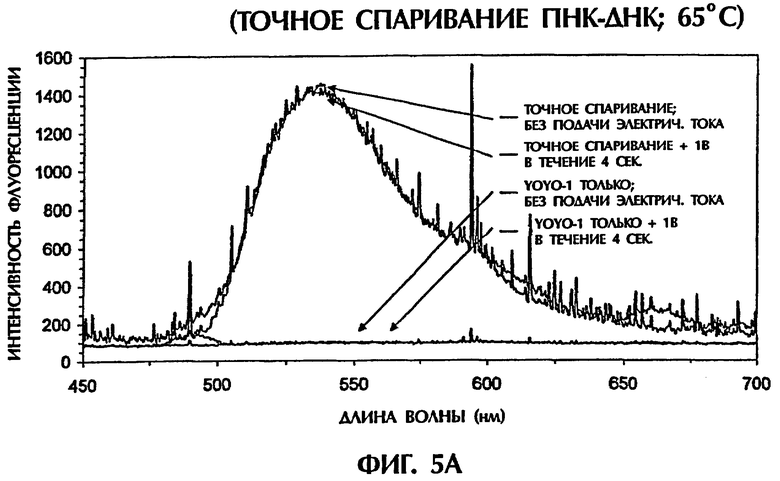
Гибридизационная реакционная смесь (80 мкл) содержала следующее: 2 пмоль дцДНК-мишени, 2 пмоль параллельного ПНК- зонда № 2, 0,5Х ТВЕ и 250 нМ интеркалятор ДНК YOYO-1. Реакционные смеси инкубировали при 95°С в течение 5-10 минут для прохождения денатурации и затем поддерживали при 65°С до анализа. Пробы помещали в кварцевую кювету, облучали лазерным лучом на ионах аргона, имеющим длину волны 488 нм, и подвергали мониторингу на флуоресцентную эмиссию при 65°С. Сопутствующие измерения температуры выполняли с использованием регулируемого программным обеспечением датчика температуры, помещаемого непосредственно в каждую пробу. Максимальная интенсивность флуоресцении имела место при длине волны 536 нм, что указывало на интеркаляцию YOYO-1 в гибриды ПНК:ДНК. В качестве второго анализа после первоначального лазерного облучения каждой пробы те же самые пробы подвергали действию постоянного электрического тока DC 1 В в течение 4 секунд. Во время последней секунды обработки электрическим током пробы облучали второй раз лазером на ионах аргона и подвергали мониторингу на флуоресцентную эмиссию при 65°С. Интенсивности флуоресценции строили в виде графика как функцию длины волны для каждой анализируемой пробы.
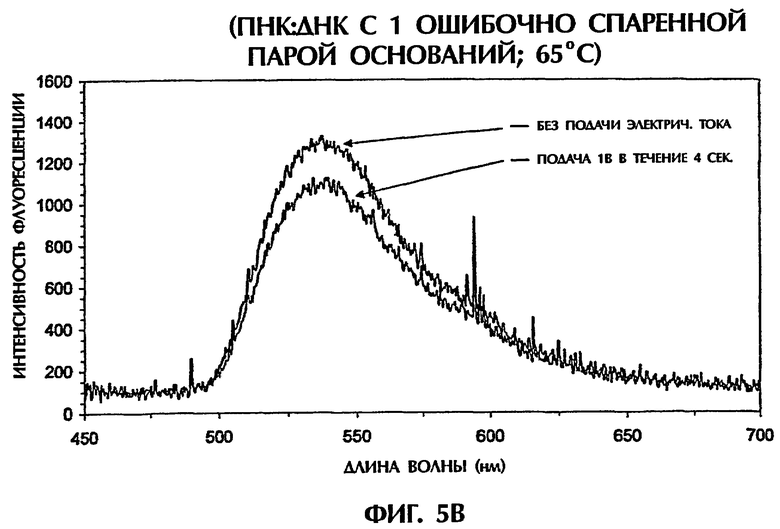
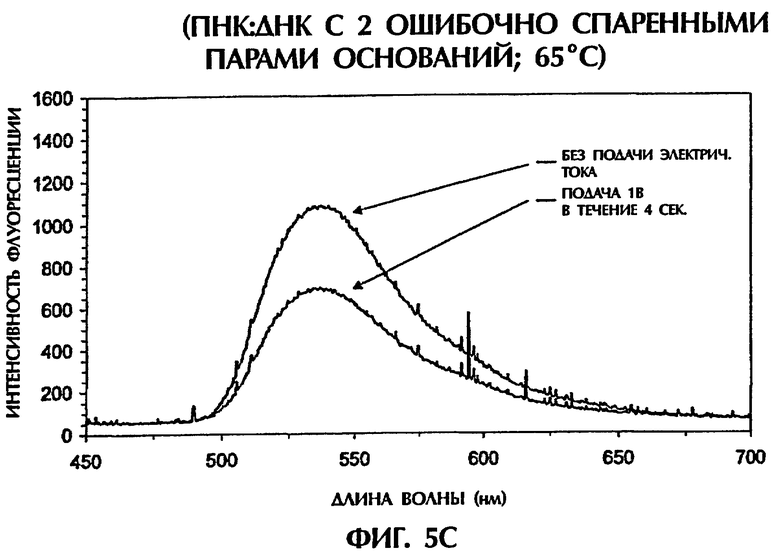
Гибриды оцДНК:ПНК, состоящие из точно комплементарных последовательностей (SEQ ID NО:4 + зонд № 2), позволяли максимальную интеркаляцию YOYO-1, давая наивысшие интенсивности флуоресценции (фиг.5А). Интенсивности флуоресценции для гибрида оцДНК:ПНК (SEQ ID NO:6 + зонд № 2) с 1 ошибочно спаренной парой оснований, гибрида оцДНК: ПНК (SEQ ID NO:6 + зонд № 2) с последовательными 2 ошибочно спаренными парами оснований и гибрида оцДНК: ПНК (SEQ ID NO:7 + зонд № 2) с разделенными 3 ошибочно спаренными парами оснований, все, были более низкими, чем интенсивность флуоресценции, наблюдаемая с точно спаренным гибридом оцДНК:ПНК при 65°С (фиг.5 и неприведенные данные). По мере увеличения степени ошибочных спариваний между зондом и мишенью уровень интеркаляции YOYO-1 уменьшался и, следовательно, уровень интенсивности флуоресценции снижался. Только уровни фона флуоресценции наблюдали, когда ни ДНК, ни ПНК не присутствовали (только YOYO-1) (фиг.5А).
При подвергании точно спаренных гибридов оцДНК:ПНК действию 1 В электрического тока в течение 4 секунд при 65°С интенсивность флуоресценции оставалась относительно постоянной, уменьшаясь только на 2% (фиг.5А). В противоположность этому подача 1 В к неполностью комплементарным дуплексам, содержащим 1 ошибочно спаренную пару оснований (фиг.5В), 2 ошибочно спаренные пары оснований (фиг.5С) и 3 ошибочно спаренные пары оснований (данные не показаны), давала интенсивности флуоресценции, которые были на 18%, 39% и 71% более низкими соответственно чем интенсивность флуоресценции, достигаемая с теми же самыми пробами, облученными в отсутствие электрического тока. Обработка низкими уровнями электрического тока (например, 1 В) дополнительно уменьшала стабильность гибридов дцДНК:ПНК, содержащих ошибочные спаривания пар оснований. По мере уменьшения комплементарности последовательности между зондом и мишенью уровень флуоресценции уменьшался драматически в присутствии электрического тока, обеспечивая высоконадежный и точный второй анализ для различения между точно спаренными последовательностями и последовательностями, содержащими мутации 1 пары оснований, 2 пар оснований или 3 пар оснований.
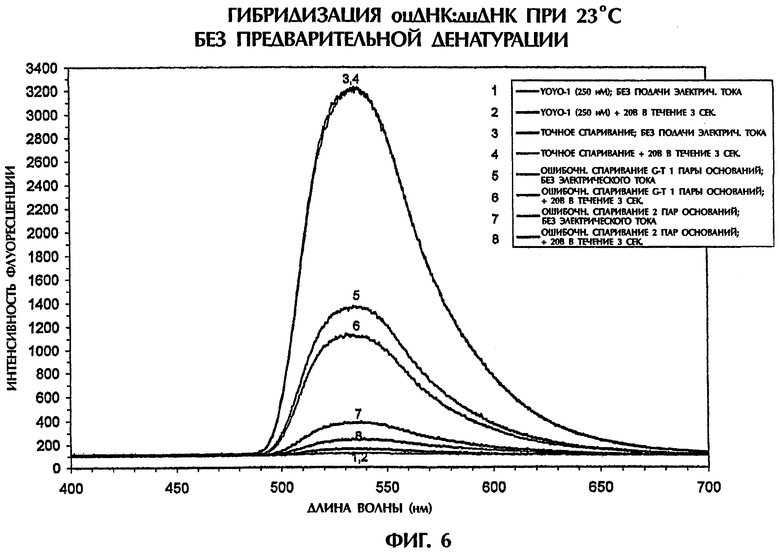
Пример 6
Гибридизационный анализ в примере 5 выполняли после денатурации дцДНК-последовательностей-мишеней и образование гибрида дцДНК:ПНК измеряли при температуре выше точки плавления (Тm) дцДНК-мишеней. Пример 6 демонстрирует надежность анализа интенсивности флуоресценции в отсутствие и в присутствии подаваемого электрического тока для различения между точными спариваниями и ошибочно спаренными парами оснований без необходимости предварительной денатурации.
Гибридизационная реакционная смесь (80 мкл) содержала следующее: 4 пмоль дцДНК-мишени, 4 пмоль антипараллельного ПНК-зонда № 1, 0,5Х ТВЕ и 250 нМ интеркалятор ДНК YOYO-1. Пробы помещали в кварцевую кювету, облучали лазерным лучом на ионах аргона, имеющим длину волны 488 нм, в течение 80 мсек и подвергали мониторингу на флуоресцентную эмиссию при 23°С. Сопутствующие измерения температуры выполняли с использованием регулируемого программным обеспечением датчика температуры, помещаемого непосредственно в каждую пробу. Максимальная интенсивность флуоресцении имела место при длине волны 536 нм, что указывало на интеркаляцию YOYO-1 в гибриды ПНК:ДНК. В качестве второго анализа после первоначального лазерного облучения каждой пробы те же самые пробы подвергали действию постоянного электрического тока DC 20 В в течение 4 секунд. Сразу же после 3 секунд подачи электрического тока пробы облучали второй раз лазером на ионах аргона и подвергали мониторингу на флуоресцентную эмиссию при 23°С. Интенсивности флуоресценции строили в виде графика как функцию длины волны для каждой анализируемой пробы.
Усиленные интеркалатором YOYO-1 триплексы дцДНК:ПНК образовывались при 23°С. Наивысшая интенсивность флуоресценции достигалась, когда 50-мерная дцДНК-последовательность-мишень дикого типа (SEQ ID NO:1) гибридизовалась с 15-мерным антипараллельным ПНК-зондом № 1 (фиг.6). В сравнении с этим интенсивности флуоресценции для триплекса дцДНК:ПНК (SEQ ID NO:2 + зонд № 1) с 1 ошибочно спаренной парой оснований и триплекса дцДНК: ПНК (SEQ ID NО:3 + зонд № 1) с 2 ошибочно спаренными парами оснований были на 60% и на 91% более низкими соответственно чем интенсивность флуоресценции, наблюдаемая с точно спаренным триплексом дцДНК:ПНК при 23°С (фиг.6). В случае отсутствия ДНК или ПНК в реакционной смеси, содержащей YOYO-1, наблюдали только фоновые уровни флуоресценции.
Различие в интенсивностях флуоресценции, полученных при помощи точно комплементарных триплексов и триплексов, содержащих 1 или 2 ошибочно спаренных пар оснований, было значимо более высоким, чем различие, достигаемое между точно спаренными дуплексами и не полностью комплементарными дуплексами (сравните фиг.5 и 6). Ясно, что анализ образования триплексов с использованием интенсивности флуоресценции обладал повышенной дискриминирующей способностью для детектирования ошибочных спариваний пар оснований.
Кроме того, даже дополнительное различение между последовательностями дикого типа и мутированными последовательностями было возможно со вторичной подачей электрического тока. Обработка 20 В в течение 3 секунд точно спаренных триплексов дцДНК:ПНК давала спектр интенсивности флуоресценции, фактически идентичный спектру, достигаемому той же самой пробой, подвергнутой действию электрического тока (фиг.6). Однако подача 20 В в течение 3 секунд к неполностью комплементарным триплексам, содержащим 1 ошибочно спаренную пару оснований и 2 ошибочно спаренные пары оснований, давала интенсивности флуоресценции, которые были на 23% и 71% более низкими соответственно чем интенсивность флуоресценции, полученная с теми же самыми пробами, облучаемыми в отсутствие электрического тока (фиг.6). Обработка электрическим током 20 В не влияла на стабильность точно комплементарных триплексов, но ослабляла стабильность триплексов дцДНК: ПНК, содержащих ошибочно спаренные пары оснований, на уровне, зависимом от степени комплементарности последовательностей между зондом и мишенью. Таким образом, подача электрического тока в анализе интенсивности флуоресценции обеспечивала даже более высоконадежный анализ для различения между последовательностями дикого типа и последовательностями, содержащими 1 или 2 ошибочно спаренных пар оснований, без предварительной денатурации последовательностей.
Хотя данное изобретение описано подробно и со ссылкой на его характерные примеры, специалисту в данной области должно быть понятно, что различные изменения и модификации могут быть произведены без отхода от идеи и объема данного изобретения.





Изобретение относится к способам секвенирования и анализа нуклеиновых кислот. Предложен способ анализа последовательность-специфической гибридизации. Способ предусматривает взаимодействие мишени, содержащей последовательность нуклеиновой кислоты, и зонда, содержащего последовательность нуклеиновой кислоты или аналог нуклеиновой кислоты. Далее к указанной тест-пробе прикладывают первый стимул и детектируют сигнал от стимулированной тест-пробы. Полученный сигнал коррелирует с аффинностью связывания между мишенью и зондом. Затем проводят калибрование первого сигнала относительно первого контрольного сигнала и на его основании определяют степень соответствия между мишенью и зондом. Далее прикладывают второй стимул к тест-пробе и детектируют его сигнал. Второй сигнал также коррелирует с аффинностью связывания мишени и зонда. Затем калибруют второй сигнал относительно второго контрольного сигнала и на его основании определяют степень соответствия мишени и зонда. Далее проводят количественное определение афинности связывания. Предложенный способ позволяет определить степень спаривания между мишенью и зондом. Сигналами, которые коррелируют с аффинностью связывания, могут быть электропроводность и/или интенсивность флуоресценции. Предложенный способ может использоваться для анализа специфической гибридизации между одноцепочечным зондом и неденатурированной двухцепочечной мишенью с образованием триплексов и для дуплексной гибридизации комплексов. 4 с. и 48 з.п. ф-лы, 6 ил.
обеспечение мишени, содержащей по меньшей мере одну последовательность нуклеиновой кислоты, причем указанную мишень при необходимости амплифицируют при помощи ПЦР;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы, причем указанную мишень при необходимости предварительно денатурируют;
приложение первого стимула к указанной тест-пробе для обеспечения первой стимулированной тест-пробы;
детектирование первого сигнала от указанной первой стимулированной тест-пробы, где указанный первый сигнал коррелирует с аффинностью связывания между указанным зондом и указанной мишенью;
калибрование указанного первого сигнала относительно первого контрольного сигнала;
определение на основании указанного калибрования первого сигнала первого определения степени соответствия между указанным зондом и указанной мишенью;
приложение второго стимула к указанной первой стимулированной тест-пробе для обеспечения второй стимулированной тест-пробы;
детектирование второго сигнала от указанной второй стимулированной тест-пробы, где указанный второй сигнал коррелирует с аффинностью связывания между указанным зондом и указанной мишенью;
калибрование указанного второго сигнала относительно второго контрольного сигнала;
определение на основании указанного калибрования второго сигнала второго определения указанной степени соответствия между указанным зондом и указанной мишенью;
количественное определение указанной аффинности связывания, где флуорофор обеспечивают в указанной тест-пробе и где а) указанный первый стимул представляет собой электрическое напряжение, указанный первый сигнал является электрической характеристикой, указанный второй стимул представляет собой возбуждающее излучение и указанный второй сигнал представляет собой интенсивность флуоресценции или б) указанный первый стимул представляет собой возбуждающее излучение, указанный первый сигнал представляет собой интенсивность флуоресценции, указанный второй стимул представляет собой электрическое напряжение и указанный второй сигнал является электрической характеристикой.
обеспечение мишени, содержащей по меньшей мере одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
обеспечение флуорофора;
добавление указанного зонда, указанного флуорофора и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
измерение доэлектризационной интенсивности флуоресценции указанной тест-пробы путем приложения возбуждающего излучения;
приложение напряжения к указанной тест-пробе;
измерение послеэлектризационной интенсивности флуоресценции указанной тест-пробы во время или после указанного приложения напряжения путем приложения указанного возбуждающего излучения и
сравнение указанной доэлектризационной интенсивности флуоресценции с указанной послеэлектризационной интенсивностью флуоресценции для осуществления указанного анализа, где а) указанный зонд и указанную мишень определяют как являющихся полностью комплементарными, когда указанная послеэлектризационная интенсивность флуоресценции эквивалентна указанной доэлектризационной интенсивности флуоресценции, б) указанный зонд и указанную мишень определяют как имеющие мисматч по крайней мере по одному основанию, когда указанная послеэлектризационная интенсивность флуоресценции не эквивалентна указанной доэлектризационной интенсивности флуоресценции, и в) величина разности между указанной доэлектризационной интенсивностью флуоресценции и указанной послеэлектризационной интенсивностью флуоресценции прямо пропорциональна числу мисматчей между указанным зондом и указанной мишенью.
обеспечение мишени, содержащей по меньшей мере одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
измерение первого сигнала первого состояния указанной тест-пробы, где указанный первый сигнал является флуоресцентной эмиссией, для обеспечения первичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанный первый сигнал коррелирует с гибридизацией между указанным зондом и указанной мишенью, причем способ предусматривает подачу напряжения к указанной тест-пробе до или во время указанного измерения указанного первого сигнала;
измерение второго сигнала второго состояния указанной тест-пробы, где указанный второй сигнал является электропроводностью, для обеспечения вторичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанный второй сигнал коррелирует с гибридизацией между указанным зондом и указанной мишенью, при условии, что, когда указанное первое состояние и указанное второе состояние являются одинаковыми, к указанной тест-пробе после измерения указанного первого сигнала и перед измерением указанного второго сигнала прилагают стимул, где указанный стимул значимо влияет на несовершенно комплементарную гибридизацию между указанным зондом и указанной мишенью и не влияет значимо на совершенно комплементарную гибридизацию между указанным зондом и указанной мишенью, и
сравнение указанного первичного определения и указанного вторичного определения для оценки, требует ли какое-либо несовпадение между ними повторного тестирования, причем когда указанное первичное определение отличается от указанного вторичного определения, способ дополнительно предусматривает повторное тестирование указанной тест-пробы.
обеспечение мишени, содержащей по меньшей мере одну последовательность нуклеиновой кислоты;
обеспечение зонда, содержащего последовательность нуклеиновой кислоты или аналога нуклеиновой кислоты;
добавление указанного зонда и указанной мишени к гибридизационной среде для обеспечения тест-пробы;
измерение доэлектризационной интенсивности флуоресценции указанной тест-пробы для обеспечения первичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанная доэлектризационная интенсивность флуоресценции коррелирует с гибридизацией между указанным зондом и указанной мишенью;
приложение напряжения к указанной тест-пробе;
измерение послеэлектризационной интенсивности флуоресценции указанной тест-пробы во время или после указанного приложения напряжения для обеспечения вторичного определения, касающегося гибридизации между указанным зондом и указанной мишенью, где указанная послеэлектризационная интенсивность флуоресценции коррелирует с гибридизацией между указанным зондом и указанной мишенью, и
сравнение указанного первичного определения и указанного вторичного определения для оценки, требует ли какое-либо несовпадение между ними повторного тестирования.
| WO 9838334, 03.09.1998 | |||
| US 5849486, 15.12.1998. |

