ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Группа изобретений относится к дискриминирующему мишень зонду (TD-зонду) и его применениям или приложениям.
СВЕДЕНИЯ О РОДСТВЕННОМ УРОВНЕ ТЕХНИКИ
Гибридизация ДНК представляет собой фундаментальный метод молекулярной биологии. Многие технологии, использующие гибридизацию ДНК, несомненно, будут очень полезными инструментами для детекции конкретной последовательности-мишени и совершенно очевидно будут необходимы в области клинической диагностики, генетического исследования и лабораторной судебно-медицинской экспертизы.
За последнее время предпринято много усилий с целью улучшения специфичности гибридизации олигонуклеотидов, поскольку на гибридизацию ДНК влияют многие условия типа концентрации солей, температуры, органических растворителей, состава оснований, длины комплементарных нитей и количества ошибочных спариваний нуклеотидных оснований между гибридизующимися нуклеиновыми кислотами (Maniatis et al., Molecular Cloning, A Laboratory Manual, Cold Springs Harbor Laboratory, 1982 и Sambrook и соавт, 1989). За прошедшее десятилетие предложено много способов: способ химической модификации оснований ДНК для высокочувствительной гибридизации (Azhikina et al., (1993) Proc. Natl. Acad. Sci., USA, 90: 11460-11462) и способ, в котором стадию промывки после гибридизации проводят при низких температурах в течение длительного периода времени для повышения способности различать (то есть дискриминировать) ошибочное спаривание (Drmanac et al., (1990) DNA and Cell Biology, 9: 527-534). He так давно был введен другой способ повышения разрешающей способности в отношении однонуклеотидных полиморфизмов (SNP) при гибридизации ДНК путем внесения искусственных ошибочных спариваний (Guo et al., (1997) Nature Biotechnology, 15: 331-5). Кроме того, во многих патентах США, включая патенты США №№6077668, 6329144, 6140054, 6350580, 6309824, 6342355 и 6268128, описан зонд для гибридизации и его применения.
Для детекции последовательностей-мишеней с использованием зондов предложено много способов. Среди этих типов способов имеется целый ряд предложенных способов с использованием гибридизационных зондов и нуклеолитических ферментов. Способ с применением зонда TaqMan™ представляет собой один из типичных примеров использования этих принципов. Зонды TaqMan™ представляют собой олигонуклеотиды, которые содержат флуоресцентный краситель, обычно на 5'-конце, и гасящий краситель, обычно расположенный на 3'-конце. После облучения возбужденный флуоресцентный краситель переносит энергию на расположенную поблизости молекулу гасящего красителя, а не флуоресцирует, то есть в результате получается нефлуоресцентный субстрат. Зонды TaqMan™ разработаны для гибридизации с внутренней областью ПЦР-продукта. Во время протекания ПЦР, когда полимераза участвует в репликации матрицы, с которой связаны зонды TaqMan™, в результате проявления 5'→3'-экзонуклеазной активности полимеразы зонды расщепляются. В результате этого происходит разделение флуоресцентного и гасящего красителей, а резонансного переноса энергии флуоресценции (FRET) больше не происходит. Флуоресценция усиливается с каждым циклом, пропорционально степени расщепления зонда. (Parashar et al., Indian J. Med. Res., 124: 385-398 (2006)). To есть, характерной чертой способа с использованием зонда TaqMan™ является использование гибридизации и реакций расщепления, обусловленных 5'→3'-нуклеазной активностью полимеразы. Однако эта технология сама по себе несет внутреннее ограничение. Наиболее острой проблемой, связанной со способом с использованием зонда TaqMan™, является неспецифическая гибридизация зондов, поскольку она неизбежно сопровождает гибридизацию между зондами и последовательностями-мишенями. Кроме того, в этом способе с большой вероятностью получаются ложноположительные сигналы (результаты), в особенности при многократной детекции множества последовательностей-мишеней.
Другой подход к детекции последовательностей-мишеней заключается в применении способов с лигированием зондов (D.Y.Wu, et al., Genomics, 4: 560 (1989); U. Landegren, et al., Science 241: 1077 (1988) и Е.Winn-Deen, et al., Clin. Chem. 37: 1522 (1991)). Реакция лигирования рассматривается как перспективный инструмент для детекции точечных мутаций. В анализе с лигированием олигонуклеотидных зондов (OLA) два зонда, перекрывающие представляющую интерес область-мишень, гибридизуются с данной областью-мишенью. Когда зонды гибридизуются с соседними основаниями мишени, противостоящие концы элементов зондов могут быть соединены путем лигирования, например, посредством обработки лигазой. Лигированный зонд указывает на наличие последовательности-мишени. Имеются сообщения, что ДНК-лигазы катализируют образованием связи между ДНК-субстратами, имеющими ошибочно спаренные нуклеотиды в сайте лигирования (Luo J, et al., Nucleic Acid Res., 24: 3071 (1996)). Даже при использовании основанных на лигировании подходов к детекции мишени остается необходимость предупреждения неспецифического связывания зондов с последовательностями-мишенями. Кроме того, необходимо, чтобы реакция лигирования осуществлялась со значительно большей специфичностью, например с дискриминацией единичного ошибочно спаренного нуклеотида, находящегося в сайте лигирования.
Существуют возрастающие потребности в полезном способе одновременной детекции наличия, уровня или картин экспрессии каждого из большого количества генов или совокупности генов. Одним из наиболее перспективных способов для этих целей являются технологии, основанные на использовании микрочипов. (Schena et al., 1995. Quantitative Monitoring of Gene Expression Patterns with a Complementary DNA Microarray, Science, 270: 467-470; DeRisi et al., 1996, Use of a cDNA Microarray to Analyse Gene Expression Patterns in Human Cancer, Nature Genetics 14: 457-460). Предложенные к настоящему времени технологии, основанные на использовании микрочипов, относятся к детекции генов или вариабельности нуклеотидов и анализу картин их экспрессии.
Как правило, в технологиях, основанных на использовании микрочипов, применяют одноцепочечные олигонуклеотиды (нуклеиновокислотные зонды), комплементарные специфической нуклеиновокислотной последовательности в нуклеиновой кислоте-мишени. Однако, поскольку детекция нуклеотидных последовательностей-мишеней с использованием традиционных ДНК-микрочипов зависит главным образом от гибридизации, у них имеются серьезные недостатки, связанные с высокой частотой ложноположительных сигналов. Нельзя исключить наличие случаев перекрестной гибридизации, особенно когда используется большое число зондов. Такая перекрестная гибридизация может существенно повлиять на качество данных и вызвать получение ложноположительных/ложноотрицательных результатов. Кроме того, микрочип нуждается в многочисленных стадиях обработки с применением жидкости, а температуры инкубации и промывки должны тщательно контролироваться для дискриминации единичного нуклеотидного ошибочного спаривания. Подтверждено, что мультиплексирование этого подхода очень затруднительно ввиду разных оптимальных условий гибридизации среди многих последовательностей зондов. (William E. Bunney, et al., 2003, Microarray Technology: A Review of New Strategies to Discover Candidate Vulnerability Genes in Psychiatric Disorders, Am. J. Psychiatry 160: 4, 657-666).
Несмотря на непрерывное внедрение усовершенствованных подходов к каждому способу, все эти способы и методы, вовлекающие гибридизацию олигонуклеотидов, не могут быть полностью избавлены от ограничений и проблем, связанных с неспецифичностью гибридизации олигонуклеотидов.
По всей этой заявке сделаны ссылки на различные патенты и публикации, и упоминания о них приведены в круглых скобках. Тем самым описание этих патентов и публикаций включено в эту заявку посредством ссылок во всей своей полноте с целью более полного описания данного изобретения и состояния уровня техники, которому соответствует данное изобретение.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
При таких обстоятельствах авторы настоящего изобретения понимали, что для преодоления недостатков традиционных технологий должны быть созданы новые зонды, способные специфически гибридизоваться с последовательностями-мишенями, отличающиеся по механизму гибридизации от традиционных зондов. В частности, авторы настоящего изобретения учитывали, что новые зонды должны обладать особыми характеристиками в отношении дискриминации мишени в нуклеолитических реакциях с участием нуклеазы, а также в реакциях лигирования.
Авторы настоящего изобретения провели интенсивные исследования с целью разработки новых технологий детекции мишеней более удобным способом для детекции или идентификации нуклеиновокислотных последовательностей-мишеней, технологий, не дающих ложноположительных и ложноотрицательных результатов. В результате авторы настоящего изобретения разработали новый дискриминирующий мишень зонд, характеризующийся разными картинами гибридизации для нуклеиновокислотных последовательностей-мишеней и нуклеиновокислотных последовательностей, не являющихся мишенями, и таким образом по сути способный отличать нуклеиновокислотные последовательности-мишени от нуклеиновокислотных последовательностей, не являющихся мишенями. В дополнение к этому, на основе новых дискриминирующих мишень зондов авторы настоящего изобретения предложили новые протоколы детекции нуклеиновокислотных последовательностей-мишеней, достоверно применимые для реакций как в жидкой фазе, так и твердой фазе.
Соответственно, данным изобретением решается задача создания дискриминирующего мишень зонда (TD-зонда), позволяющего отличать (то есть дискриминировать) нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Другой задачей данного изобретения является разработка способа детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) и 5'→3'-экзонуклеолитической реакции в жидкой фазе или твердой фазе.
Еще одной другой задачей данного изобретения является разработка способа детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) и полимеразной цепной реакции (ПЦР).
Следующей задачей данного изобретения является разработка способа детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) и реакции лигирования.
Еще одной следующей задачей данного изобретения является создание наборов для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот.
Другие задачи и преимущества настоящего изобретения станут очевидны из следующего далее подробного описания в сочетании с прилагаемой формулой изобретения и графическими материалами.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг.1 схематично представлена дискриминация нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, с использованием TD-зонда с двумя метками и фермента, обладающего 5'→3'-экзонуклеазной активностью. TD-зонд содержит репортерную молекулу на своем 5'-концевом втором участке гибридизации и молекулу-гаситель на своем 3'-концевом первом участке гибридизации.
На Фиг.2 схематично представлена дискриминация нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, с использованием TD-зонда с двумя метками и фермента, обладающего 5'→3'-экзонуклеазной активностью. TD-зонд содержит как репортерную молекулу, так и молекулу-гаситель на своем 5'-концевом втором участке гибридизации.
На Фиг.3 схематично представлено отсутствие генерации сигнала в ПЦР в режиме реального времени с использованием матричной ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью, когда TD-зонд с двумя метками, содержащий как репортерную молекулу, так и молекулу-гаситель на своем 5'-концевом втором участке гибридизации, гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью.
На Фиг.4 схематично представлена дискриминация нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, с использованием иммобилизованного TD-зонда, имеющего одну метку, и фермента, обладающего 5'→3'-экзонуклеазной активностью, в твердой фазе. На Фиг.4А представлено изменение интенсивности сигнала флуоресценции при специфичной к мишени гибридизации иммобилизованного TD-зонда. На Фиг.4В представлено отсутствие какого-либо изменения интенсивности сигнала флуоресценции при неспецифичной к мишени гибридизации иммобилизованного TD-зонда.
На Фиг.5 схематично представлена дискриминация нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, с использованием иммобилизованного TD-зонда, имеющего две метки, и фермента, обладающего 5'→3'-экзонуклеазной активностью, на твердой фазе. На Фиг.5А представлена генерация сигнала при специфичной к мишени гибридизации иммобилизованного TD-зонда. На Фиг.5В представлено отсутствие какого-либо сигнала при неспецифичной к мишени гибридизации иммобилизованного TD-зонда.
На Фиг.6 схематично представлена дискриминация нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, с использованием неиммобилизованного олигонуклеотида в качестве первого зонда с одной меткой, иммобилизованного TD-зонда в качестве второго зонда с одной меткой и лигазы в твердой фазе. На Фиг.6А представлено лигирование между первым зондом и вторым зондом при специфичной к мишени гибридизации. На Фиг.6В представлено отсутствие какого-либо лигирования зондов при неспецифичной к мишени гибридизации.
На Фиг.7 схематично представлено лигирование при специфичной к мишени гибридизации с использованием неиммобилизованного олигонуклеотида в качестве первого зонда с одной меткой, иммобилизованного TD-зонда в качестве немеченого второго зонда и лигазы в твердой фазе.
На Фиг.8 показаны результаты расщепляющего действия фермента, обладающего 5'→3'-экзонуклеазной активностью, на зонды с 5'-концевым ошибочным спариванием. Символы: 1)матрица представляет собой синтетический олигонуклеотид для гена Staphylococcus aureus; 2)зонд содержит репортерную молекулу на своем 5'-концевом и молекулу-гаситель на своем 3'-концевом участке; 3)SA_P0 имеет спаренную последовательность на своем 5'-концевом участке; 4)SA_P1 имеет единственный ошибочно спаренный нуклеотид на своем 5'-конце; 5)SA_P3 имеет три ошибочно спаренных нуклеотида на своем 5'-концевом участке; 6)SA_P6 имеет шесть ошибочно спаренных нуклеотидов на своем 5'-концевом участке; 7)SA_P9 имеет девять ошибочно спаренных нуклеотидов на своем 5'-концевом участке.
На Фиг.9 показаны результаты дискриминации нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, в зависимости от гибридизации 5'-концевого второго участка гибридизации TD-зонда, имеющего две метки. На Фиг.9А и 9 В показана детекция гена Staphylococcus aureus и гена Neisseria gonorrhoeae, соответственно. Символы на Фиг.9А: 1)матрица представляет собой синтетический олигонуклеотид для гена Staphylococcus aureus; 2)TD-зонд содержит репортерную молекулу на своем 5'-конце и молекулу-гаситель на своем 3'-концевом первом участке гибридизации; 3)SA_TD_M имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации; 4)SA_TD_m имеет ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации. Символы на Фиг.9В: 1)матрица представляет собой синтетический олигонуклеотид для гена Neisseria gonorrhoeae; 2)TD-зонд содержит репортерную молекулу на своем 5'-конце и молекулу-гаситель на своем 3'-концевом первом участке гибридизации; 3)NG_TD_M имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации; 4)NG_TD_m имеет ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации.
На Фиг.10 показаны результаты сравнения TD-зонда и традиционного зонда для детекции гена Staphylococcus aureus. Символы: 1)матрица представляет собой синтетический олигонуклеотид для гена Staphylococcus aureus; 2)зонд содержит репортерную молекулу на своем 5'-концевом и молекулу-гаситель на своем 3'-концевом участке; 3)SA_TD_M представляет собой TD-зонд и имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации; 4)SA_TD_m1 представляет собой TD-зонд и имеет три ошибочно спаренных нуклеотида на своем 5'-концевом втором участке гибридизации; 5)SA_Con_M представляет собой традиционный зонд и имеет спаренную последовательность на своем 5'-концевом участке; 6)SA_Con_m1 представляет собой традиционный зонд и имеет три ошибочно спаренных нуклеотида на своем 5'-концевом участке.
На Фиг.11 показаны результаты ПЦР в режиме реального времени для детекции нуклеиновокислотной последовательности-мишени с использованием TD-зонда, имеющего как репортерную молекулу, так и молекулу-гаситель на своем 5'-концевом втором участке гибридизации. На Фиг.11А и 11В показана детекция гена Staphylococcus aureus и гена Neisseria gonorrhoeae, соответственно. Символы на Фиг.11А: 1)матрица представляет собой геномную ДНК Staphylococcus aureus; 2)TD-зонд содержит и репортерную молекулу, и молекулу-гаситель на своем 5'-концевом втором участке гибридизации; 3)SA_TD2_M имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации; 4)SA_TD2_m имеет ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации. Символы на Фиг.11 В: 1)матрица представляет собой геномную ДНК Neisseria gonorrhoeae; 2)TD-зонд содержит и репортерную молекулу, и молекулу-гаситель на своем 5'-концевом втором участке гибридизации; 3)NG_TD2_M имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации; 4)NG_TD2_m имеет ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации.
На Фиг.12 показаны результаты ПЦР в режиме реального времени для дискриминации единичного нуклеотидного ошибочного спаривания с использованием TD-зонда, имеющего и репортерную молекулу, и молекулу-гаситель на своем 5'-концевом втором участке гибридизации. Символы: 1)матрица представляет собой геномную ДНК Staphylococcus aureus; 2)TD-зонд содержит и репортерную молекулу, и молекулу-гаситель на своем 5'-концевом втором участке гибридизации; 3)SA_TD_S_M имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации; 4)SA_TD_S_m имеет единственный ошибочно спаренный нуклеотид на своем 5'-концевом втором участке гибридизации.
На Фиг.13 показаны результаты дискриминации нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, в зависимости от гибридизации 5'-концевого второго участка гибридизации TD-зонда, имеющего две метки, иммобилизованного на поверхности твердой подложки. Для проверки воспроизводимости каждое пятно анализировали в двух повторах. Интенсивность флуоресценции указывает среднюю величину для дублированных пятен. Символы: SA_TD1_Chip_M относится к TD-зонду, имеющему спаренную последовательность на своем 5'-концевом втором участке гибридизации; SA_TD1_Chipjn относится к TD-зонду, имеющему ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации.
На Фиг.14 показаны результаты сравнения TD-зонда и традиционного зонда в отношении детекции гена Staphylococcus aureus в твердой фазе. Символы: SA_TD1_Chip_M относится к TD-зонду, имеющему спаренную последовательность на своем 5'-концевом втором участке гибридизации; SA_TD1_Chip_m1 относится к TD-зонду, имеющему три ошибочно спаренных нуклеотида на своем 5'-концевом втором участке гибридизации; SA_Con_Chip_M относится к традиционному зонду, имеющему спаренную последовательность на своем 5'-концевом участке; SA_Con_Chip_m1 относится к традиционному зонду, имеющему три ошибочно спаренных нуклеотида на своем 5'-концевом участке.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится (is drawn) к дискриминирующему мишень зонду (TD-зонду) и его применениям или приложениям.
TD-зонды
В одном из аспектов настоящего изобретения предложен дискриминирующий мишень зонд (TD-зонд), имеющий структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей далее общей формулой I, позволяющий отличать (то есть дискриминировать) нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью:
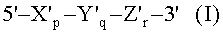
где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл (температура плавления) 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Авторы настоящего изобретения провели интенсивные исследования с целью разработки новых технологий детекции мишеней более удобным способом для детекции или идентификации нуклеиновокислотных последовательностей-мишеней, технологий, не дающих ложноположительных и ложноотрицательных результатов. В результате авторы настоящего изобретения разработали новый дискриминирующий мишень зонд, характеризующийся разными картинами гибридизации для нуклеиновокислотных последовательностей-мишеней и нуклеиновокислотных последовательностей, не являющихся мишенями, и таким образом по сути способный отличать нуклеиновокислотные последовательности-мишени от нуклеиновокислотных последовательностей, не являющихся мишенями. В дополнение к этому, на основе новых дискриминирующих мишень зондов авторы настоящего изобретения предложили новые протоколы детекции нуклеиновокислотных последовательностей-мишеней, достоверно применимые для реакций как в жидкой фазе, так и твердой фазе.
В связи с этим зонд, используемый в настоящем изобретении, называется "дискриминирующим мишень зондом" (TD-зондом) и представленные технологии с использованием TD-зонда называются "методом детекции мишеней с использованием TD-зонда".
TD-зонд по настоящему изобретению имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), содержащую три разные участка с различными свойствами в пределах одной олигонуклеотидной молекулы: 5'-концевой второй участок гибридизации, 3'-концевой первый участок гибридизации и разделительный участок. Такая структура способствует функционированию TD-зонда в качестве зонда, демонстрирующего более высокую специфичность, что интерпретирует настоящее изобретение как являющееся новым и неочевидным по сравнению с предшествующим уровнем техники.
Структура mDSO представляет собой по-новому модифицированную версию структуры DSO (олигонуклеотида с двойной специфичностью), которая впервые была предложена авторами настоящего изобретения (см. WO 2006/095981). Структура DSO также называется DPO (олигонуклеотидом с двойным праймированием), поскольку она служит в качестве праймеров (Chun et al., Dual priming oligonucleotide system for the multiplex detection of respiratory viruses and SNP genotyping of CYP2C19 gene, Nucleic Acid Research, 35: 6e40 (2007)).
DSO является воплощением новой концепции, согласно которой ее гибридизация или отжиг двойственно определяется 5'-концевым определяющим специфичность участком с высокой Тпл (или 5'-концевым первым участком гибридизации, 5'-концевым первым праймирующим участком) и 3'-концевым определяющим специфичность участком с низкой Тпл (или 3'-концевым вторым участком гибридизации, 3'-концевым вторым праймирующим участком), отделенными друг от друга разделительным участком, демонстрируя значительно повышенную специфичность гибридизации (см. WO 2006/095981; Kim et al., Direct detection of lamivudine-resistant hepatitis В virus mutants by multiplex PCR using dual-priming oligonucleotide primers, Journal of Virological Methods, 149: 76-84 (2008); Kirn, et al., Rapid detection and identification of 12 respiratory viruses using a dual priming oligonucleotide system-based multiplex PCR assay, Journal of Virological Methods, doi:10.1016/j.jviromet.2008.11.007 (2008); Horii et al., Use of dual priming oligonucleotide system to detect multiplex sexually transmitted pathogens in clinical specimens, Letters in Applied Microbiology, doi:10.111/j.1472-765X2009.02618x (2009)). По сути, DSO содержит в конечном счете два сегмента с различными свойствами в отношении гибридизации: 5'-концевой первый участок гибридизации, который инициирует стабильную гибридизацию, и 3'-концевой второй участок гибридизации, который преимущественно определяет специфичность к мишени.
Структура mDSO представляет собой структуру, обратную DSO, а именно: 5'-концевой второй участок гибридизации, который преимущественно определяет специфичность к мишени, и 3'-концевой первый участок гибридизации, который инициирует стабильную гибридизацию.
Когда TD-зонд, имеющий структуру mDSO, гибридизуется с последовательностями, не являющимися мишенями, скорее то в гибридизацию скорее не вовлечен его 5'-концевой участок, а не 3'-концевой участок, что резко отличает его от структуры DSO, ранее предложенной автором настоящего изобретения.
Чтобы полностью преодолеть проблемы, связанные с ложноположительными сигналами, в частности, относящимися к зондам, авторы настоящего изобретения приложили интенсивные усилия, чтобы предложить более надежные и точные подходы, при которых генерация сигнала, указывающая на наличие последовательностей-мишеней, была связана не только с гибридизацией зонда, но также и с дополнительными ферментативными реакциями, такими как 5'→3'-экзонуклеазная реакция и лигирование двух зондов. С учетом того, что новые подходы зависят в большей степени от гибридизации 5'-концевого участка зондов, авторы настоящего изобретения разработали зонды, способные проявлять максимальные параметры специфичности для 5'-конца, и модифицировали известную DSO с тем, чтобы предложить TD-зонд.
TD-зонд с характерными особенностями гибридизации 5'-конца позволяет осуществлять детекцию последовательностей-мишеней без получения каких-либо ложноположительных сигналов, чего не было достигнуто с использованием традиционных зондов и DSO-зондов.
Специфичность гибридизации (или специфичность к мишени) TD-зонда, обусловленная структурой mDSO, способствует отсутствию ложной детекции мишени в настоящем изобретении.
Интересно, что TD-зонд, имеющий структуру mDSO, отчетливо демонстрирует различное поведение при гибридизации с каждой из нуклеиновокислотных последовательностей-мишеней и нуклеиновокислотных последовательностей, не являющихся мишенями. Как схематично представлено на Фиг.1-3, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации TD-зонда образуют двойную цепь с этой нуклеиновокислотной последовательностью-мишенью. Когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью (то есть, при неспецифической гибридизации или неспецифическом связывании), его 3'-концевой первый участок гибридизации главным образом связывается с нуклеиновокислотной последовательностью, не являющейся мишенью, но как 5'-концевой второй участок гибридизации, так и разделительный участок оба не гибридизуются с нуклеиновокислотной последовательностью, не являющейся мишенью, так что оба участка образуют одиночную цепь.
Когда происходит отжиг 3'-концевого первого участка гибридизации с последовательностью, не являющейся мишенью, маловероятно, что 5'-концевой второй участок гибридизации с более короткой последовательностью (более низкой величиной Тпл) гибридизуется с последовательностью, не являющейся мишенью, в условиях специфичной к мишени гибридизации TD-зонда. Причина в том, что 3'-концевой первый участок гибридизации и 5'-концевой второй участок гибридизации отделены друг от друга разделительным участком с точки зрения событий гибридизации. Другими словами, 5'-концевой второй участок гибридизации вовлекается в события гибридизации относительно независимым образом от 3'-концевого первого участка гибридизации, и гибридизация 5'-концевого второго участка гибридизации в меньшей степени зависит от гибридизации 3'-концевого первого участка гибридизации. В связи с этим вероятность гибридизации 5'-концевого второго участка гибридизации с последовательностью, не являющейся мишенью, становится намного ниже.
Когда и 3'-концевой первый участок гибридизации, и 5'-концевой второй участок гибридизации TD-зонда имеют последовательность, комплементарную матрице, TD-зонд может специфично гибридизоваться с нуклеиновокислотной последовательностью-мишенью матрицы в условиях специфичной к мишени гибридизации. Однако, когда только 5'-концевой второй участок гибридизации TD-зонда имеет последовательность, комплементарную матрице, тогда TD-зонд не может гибридизоваться с матрицей в условиях специфичной к мишени гибридизации.
Описанные выше характеристики TD-зонда позволяют выявлять последовательности-мишени с очень высокой специфичностью к мишени ввиду следующих двух моментов наблюдения за мишенью. Во-первых, TD-зонд, имеющий в каждом случае разные картины гибридизации для нуклеиновокислотных последовательностей, являющихся мишенями и не являющихся мишенями, как описано выше, способен с гораздо более высокой специфичностью отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью. Во-вторых, наличие последующих ферментативных реакций (5'→3'-экзонуклеолитической реакции или реакции лигирования) определяется в зависимости от картин гибридизации TD-зонда, повышая специфичность к мишени в процедурах детекции мишени.
TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью и образует двойную цепь. Как обсуждалось выше, TD-зонд, имеющий структуру mDSO с такой необычной конструкцией, позволяет идеально отличать нуклеиновокислотные последовательности-мишени от нуклеиновокислотных последовательностей, не являющихся мишенями.
Согласно предпочтительному воплощению универсальное основание в разделительном участке выбрано из группы, состоящей из дезоксиинозина, инозина, 7-деаза-2'-дезоксиинозина, 2-аза-2'-дезоксиинозина, 2'-ОМе-инозина, 2'-F-инозина, дезокси-3-нитропиррола, 3-нитропиррола, 2'-ОМе-3-нитропиррола, 2'-F-3-нитропиррола, 1-(2'-дезокси-бета-D-рибофуранозил)-3-нитропиррола, дезокси-5-нитроиндола, 5-нитроиндола, 2'-ОМе-5-нитроиндола, 2'-F-5-нитроиндола, дезокси-4-нитробензимидазола, 4-нитробензимидазола, дезокси-4-аминобензимидазола, 4-аминобензимидазола, дезокси-небуларина, 2'-F-небуларина, 2'-F-4-нитробензимидазола, 5-нитроиндола, входящего в состав PNA (пептидо-нуклеиновой кислоты), небуларина, входящего в состав PNA, инозина, входящего в состав PNA, 4-нитробензимидазола, входящего в состав PNA, 3-нитропиррола, входящего в состав PNA, морфолино-5-нитроиндола, морфолино-небуларина, морфолино-инозина, морфолино-4-нитробензимидазола, морфолино-3-нитропиррола, фосфорамидат-5-нитроиндола, фосфорамидат-небуларина, фосфорамидат-инозина, фосфорамидат-4-нитробензимидазола, фосфорамидат-3-нитропиррола, 2'-O-метоксиэтил-инозина, 2'-O-метоксиэтил-небуларина, 2'-O-метоксиэтил-5-нитроиндола, 2'-O-метоксиэтил-4-нитро-бензимидазола, 2'-O-метоксиэтил-3-нитропиррола и их комбинаций. Более предпочтительно, чтобы универсальное основание представляло собой дезоксиинозин, 1-(2'-дезокси-бета-D-рибофуранозил)-3-нитропиррол или 5-нитроиндол, наиболее предпочтительно, дезоксиинозин.
Предпочтительно, чтобы разделительный участок содержал нуклеотиды, имеющие по меньшей мере три, более предпочтительно по меньшей мере четыре, наиболее предпочтительно по меньшей мере пять универсальных оснований. Более предпочтительно, чтобы разделительный участок содержал следующие друг за другом нуклеотиды, имеющие по меньшей мере три, более предпочтительно по меньшей мере четыре, наиболее предпочтительно по меньшей мере пять универсальных оснований. Альтернативно, разделительный участок содержит 3-10, 3-8, 4-7 или 4-5 следующих друг за другом универсальных оснований.
Предпочтительно, чтобы длина 3'-концевого первого участка гибридизации превышала длину 5'-концевого второго участка гибридизации. Длина 3'-концевого первого участка гибридизации составляет предпочтительно 15-60 нуклеотидов, более предпочтительно 15-40 нуклеотидов, еще более предпочтительно 15-30 нуклеотидов.
Предпочтительно, чтобы длина 5'-концевого второго участка гибридизации составляла по меньшей мере 3, более предпочтительно 5 и еще более предпочтительно 6 нуклеотидов. Предпочтительно, чтобы длина 5'-концевого второго участка гибридизации не превышала 15, более предпочтительно не превышала 13 и еще более предпочтительно не превышала 12 нуклеотидов.
Предпочтительно, чтобы длина 5'-концевого второго участка гибридизации составляла 3-15 нуклеотидов, более предпочтительно 3-13 нуклеотидов, еще более предпочтительно 4-12 нуклеотидов и наиболее предпочтительно 5-11 нуклеотидов. Длина разделительного участка составляет предпочтительно 3-10 нуклеотидов, более предпочтительно 3-8 нуклеотидов, еще более предпочтительно 4-7 нуклеотидов, наиболее предпочтительно 4-5 нуклеотидов. Длина обоих участков, и 5'-концевого второго участка гибридизации, и разделительного участка, предпочтительно составляет по меньшей мере шесть, более предпочтительно по меньшей мере девять, еще более предпочтительно по меньшей мере двенадцать и наиболее предпочтительно по меньшей мере пятнадцать нуклеотидов.
Согласно предпочтительному воплощению Тпл 3'-концевого первого участка гибридизации находится в диапазоне от 40°С до 80°С, более предпочтительно от 45°С до 70°С. Тпл 5'-концевого второго участка гибридизации предпочтительно находится в диапазоне от 6°С до 40°С и более предпочтительно от 10°С до 40°С. Тпл разделительного участка предпочтительно находится в диапазоне от 2°С до 15°С и более предпочтительно от 3°С до 15°С.
Согласно предпочтительному воплощению TD-зонд имеет метку или систему взаимодействующих меток, содержащую некоторое количество меток для генерирования детектируемого сигнала, указывающего на наличие нуклеиновокислотных последовательностей-мишеней.
Метка, генерирующая детектируемый сигнал, полезная в настоящем изобретении, включает любую метку, известную специалисту в данной области техники. Некоторые метки представляют собой метку, состоящую из одной молекулы или одного атома; однако большинство меток (например, система интерактивных меток) представляют собой метки, состоящие по меньшей мере из двух или более молекул либо двух или более атомов.
Согласно предпочтительному воплощению метка на TD-зонде представляет собой химическую метку, ферментативную метку, радиоактивную метку, флуоресцентную метку, люминесцентную метку, хемилюминесцентную метку или металлическую метку (например, золото).
Химическая метка включает биотин. Специфичность связывания биотина со стрептавидином (или авидином) позволяет осуществлять опосредованное генерирование сигнала, указывающее на наличие нуклеиновокислотных последовательностей-мишеней.
Ферментативная метка включает щелочную фосфатазу, β-галактозидазу, β-глюкозидазу, люциферазу, цитохром Р450 и пероксидазу хрена. Используя субстраты для ферментативных меток, можно получить сигнал, указывающий на наличие нуклеиновокислотных последовательностей-мишеней. Когда используют щелочную фосфатазу, то в качестве субстрата для развития цветной реакции могут быть применены бромхлориндолилфосфат (ВСIР), нитротетразолий синий (NBT) или ECF (усиленная хемифлуоресценция; от англ. enhanced chemifluorescence); в случае использования пероксидазы хрена в качестве субстрата могут быть применены хлорнафтол, аминоэтилкарбазол, диаминобензидин, D-люциферин, люцигенин (нитрат бис-N-метилакридиния), простой бензиловый эфир резоруфина, люминал, реагент амплекс красный (Amplex Red) (10-ацетил-3,7-дигидроксифеноксазин), HYR (п-фенилендиамин-HCl и пирокатехол), ТМВ (3,3',5,5'-тетраметилбензидин), ABTS (2,2'-азино-ди[3-этилбензтиазолина сульфонат]), о-фенилендиамин (OPD) или нафтол/пиронин; и в случае использования глюкозооксидазы, в качестве субстрата могут быть применены тетра-NBT (нитротетразолий синий) или 1-метокси-PMS (феназина метосульфат).
Радиоактивная метка включает С14, I125, P32 и S35.
Согласно предпочтительному воплощению настоящего изобретения метка, соединенная с TD-зондом, представляет собой одиночную метку, способную давать сигнал в режиме реального времени. Например, одиночная метка представляет собой флуоресцентный хелат тербия (Nurmi et al., Nucleic Acids Research, 2000, Vol.28, №8). Nurmi и соавт.сообщают, что данная метка в связанной с зондом форме излучает низкий уровень флуоресценции, но когда в результате проявления 5'→3'-нуклеолитической активности метка высвобождается из дуплекса зонд-матрица, сигнал флуоресценции возрастает. Поэтому флуоресцентный хелат тербия позволяет осуществлять детекцию мишени в режиме реального времени, даже несмотря на то, что с TD-зондом по настоящему изобретению соединена одиночная метка.
Система взаимодействующих меток представляет собой генерирующую сигнал систему, в которой энергия передается от донорной молекулы к акцепторной молекуле без участия радиоактивности.
В качестве репрезентативной системы взаимодействующих меток система меток при FRET (резонансном переносе энергии флуоресценции) включает флуоресцентную репортерную молекулу (донорную молекулу) и молекулу-гаситель (акцепторную молекулу). При FRET донор энергии является флуоресцентным, а акцептор энергии может быть флуоресцентным или не быть флуоресцентным.
Для другой формы систем взаимодействующих меток донор энергии не является флуоресцентным, например, является хромофором, а акцептор энергии является флуоресцентным. Для еще одной другой формы систем взаимодействующих меток донор энергии является люминесцентным, например биолюминесцентным, хемилюминесцентным, электрохемилюминесцентным, а акцептор является флуоресцентным.
Более предпочтительно, чтобы метка на TD-зонде представляла собой систему взаимодействующих меток, еще более предпочтительно систему FRET-меток, наиболее предпочтительно пару, состоящую из репортерной молекулы и молекулы-гасителя.
Предпочтительно, в том случае, когда используют FRET-метку, чтобы две метки (репортерная молекула и молекула-гаситель, расположенные на TD-зонде) были разделены сайтом внутри TD-зонда, чувствительным к нуклеазному расщеплению, что позволяет тем самым, при наличии 5'→3'-экзонуклеазной активности, отделить репортерную молекулу от молекулы-гасителя путем осуществления расщепления по данному чувствительному сайту с получением вследствие этого сигнала, указывающего на наличие нуклеиновокислотной последовательности-мишени.
Метку можно присоединить к TD-зонду в соответствии с традиционными способами. Например, метку можно присоединить к TD-зонду через спейсер, содержащий по меньшей мере три атома углерода (например, 3-углеродный спейсер, 6-углеродный спейсер или 12-углеродный спейсер).
Согласно предпочтительному воплощению репортерная молекула и молекула-гаситель обе расположены на 5'-концевом втором участке гибридизации, либо репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и разделительного участка. Например, репортерная молекула расположена на 5'-концевом втором участке гибридизации, а молекула-гаситель на разделительном участке. Альтернативно, молекула-гаситель расположена на 5'-концевом втором участке гибридизации, а репортерная молекула на разделительном участке.
Более предпочтительно, чтобы одна из молекул, выбранная из репортерной молекулы и молекулы-гасителя, была расположена на 5'-конце TD-зонда, а другая была расположена в сайте 5'-концевого второго участка гибридизации.
Согласно предпочтительному воплощению TD-зонд имеет одну из молекул, выбранную из репортерной молекулы и молекулы-гасителя, на своем 5'-концевом втором участке гибридизации, а другую на своем 3'-концевом первом участке гибридизации.
Более предпочтительно, чтобы одна из молекул, выбранная из репортерной молекулы и молекулы-гасителя, была расположена на 5'-конце TD-зонда, а другая была расположена в сайте 3'-концевого первого участка гибридизации.
TD-зонд по настоящему изобретению имеет широкий спектр приложений для детекции последовательности-мишени, которые приведены ниже.
I. Способ детекции мишени с использованием 5'→3'-экзонуклеолитической реакции в жидкой фазе или на твердой фазе
1. Способ детекции мишени в жидкой фазе
TD-зонд по настоящему изобретению демонстрирует превосходные показатели при детекции последовательности-мишени.
В другом аспекте настоящего изобретения предложен способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с TD-зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
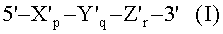
где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит две метки - флуоресцентную репортерную молекулу и молекулу-гаситель, способную гасить флуоресценцию репортерной молекулы; по меньшей мере одна молекула из репортерной молекулы и молекулы-гасителя расположена на 5'-концевом втором участке гибридизации; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, а 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
(b) приведения в контакт продукта со стадии (а) с ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, в результате чего сигнал флуоресценции будет отсутствовать; и
(c) детекции сигнала флуоресценции, так что сигнал флуоресценции, генерируемый в результате переваривания 5'-концевого второго участка гибридизации, является указанием на наличие нуклеиновокислотной последовательности-мишени.
Согласно настоящему изобретению TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью.
Согласно настоящему изобретению детекцию нуклеиновокислотной последовательности-мишени без ложноположительных сигналов можно осуществить с использованием только TD-зонда и фермента, обладающего 5'→3'-экзонуклеазной активностью, что впервые предложено авторами настоящего изобретения.
Как представлено на Фиг.1, TD-зонд демонстрирует отчетливо разное поведение при гибридизации по отношению к каждой из нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью. Когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации TD-зонда образуют двойную цепь с этой нуклеиновокислотной последовательностью-мишенью. В противоположность этому, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью (то есть, при неспецифической гибридизации или неспецифическом связывании), тогда его 3'-концевой первый участок гибридизации связывается главным образом с нуклеиновокислотной последовательностью, не являющейся мишенью, но как 5'-концевой второй участок гибридизации, так и разделительный участок оба не гибридизуются с нуклеиновокислотной последовательностью, не являющейся мишенью, так что оба участка образуют одиночную цепь.
Вследствие этого, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, (например, матричной полимеразой нуклеиновых кислот, обладающей 5'→3'-экзонуклеазной активностью), и флуоресцентная репортерная молекула и молекула-гаситель отделяются друг от друга, генерируя сигнал флуоресценции для нуклеиновокислотной последовательности-мишени. Обычно переваривание TD-зонда происходит первоначально на его 5'-конце и в дальнейшем в 5'→3'-направлении.
В противоположность этому, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, которая не переваривается в результате проявления 5'→3'-экзонуклеазной активности фермента. В результате TD-зонд не генерирует никаких сигналов при неспецифической гибридизации.
В результате такого особого поведения TD-зонда при гибридизации можно осуществить детекцию нуклеиновокислотной последовательности-мишени без ложных сигналов, используя только TD-зонд и фермент, обладающий 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению используемый фермент, обладающий 5'→3'-экзонуклеазной активностью, действует только на 5'-конце двухцепочечных нуклеиновых кислот и катализирует экзонуклеолитическую реакцию в 5'→3'-направлении, не переваривая одноцепочечные нуклеиновые кислоты.
Согласно предпочтительному воплощению фермент, обладающий 5'→3'-экзонуклеазной активностью, является термостабильным ферментом. Согласно предпочтительному воплощению фермент, обладающий 5'→3'-экзонуклеазной активностью, представляет собой матричную полимеразу нуклеиновых кислот, более предпочтительно термостабильную матричную полимеразу нуклеиновых кислот.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и 3'-концевого первого участка гибридизации. Например, флуоресцентная репортерная молекула может быть расположена на 5'-концевом втором участке гибридизации, а молекула-гаситель на 3'-концевом первом участке гибридизации. Альтернативно, молекула-гаситель может быть расположена на 5'-концевом втором участке гибридизации, а флуоресцентная репортерная молекула на 3'-концевом первом участке гибридизации.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула и молекула-гаситель обе расположены на 5'-концевом втором участке гибридизации, либо репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и разделительного участка. Наиболее предпочтительно, чтобы флуоресцентная репортерная молекула и молекула-гаситель обе располагались на 5'-концевом втором участке гибридизации TD-зонда.
Известно, что некоторые ферменты (включая матричные полимеразы нуклеиновых кислот), обладающие 5'→3'-экзонуклеазной активностью, также проявляют эндонуклеазную активность, которая обычно бывает очень низкой. Степень эндонуклеазной активности может зависеть от (1) типа ферментов, (2) реакционных условий, таких как температура, продолжительность реакции и состав реакционной смеси, (3) длины зондов, последовательности зондов и длины последовательности с ошибочным спариванием на 5'-конце зондов или (4) последовательностей мишени. Согласно предпочтительному воплощению, когда в способе по настоящему изобретению используются ферменты, обладающие как 5'→3'-экзонуклеазной активностью, так и эндонуклеазной активностью, его осуществляют в условиях, достаточных для защиты от эндонуклеазной активности. Предпочтительно, чтобы реализация настоящего изобретения осуществлялась с использованием ферментов, обладающих 5'→3'-экзонуклеазной активностью и незначительной эндонуклеазной активностью или не обладающих никакой эндонуклеазной активностью.
Ввиду этого эндонуклеазная активность не является существенным фактором при детекции мишени с использованием TD-зондов вместе с ферментом, обладающим 5'→3'-экзонуклеазной активностью и эндонуклеазной активностью. Тем не менее, для более точной детекции мишени в 3'-концевой первый участок гибридизации TD-зонда может быть введена молекула-блокатор для блокирования катализируемого эндонуклеазной активностью переваривания 3'-концевого первого участка гибридизации TD-зонда, гибридизующегося с нуклеиновокислотной последовательностью, не являющейся мишенью. В частности, когда TD-зонд используют в жидкой фазе, флуоресцентная репортерная молекула и молекула-гаситель обе могут быть расположены на 5'-концевом втором участке гибридизации TD-зонда для более точной детекции мишени.
В настоящем изобретении фермент, обладающий 5'→3'-экзонуклеазной активностью, как правило включает ферменты, обладающие 5'→3'-экзонуклеазной активностью, и обычно включает ферменты, обладающие наряду с 5'→3'-экзонуклеазной активностью дополнительно эндонуклеазной активностью. В настоящем изобретении матричная полимераза нуклеиновых кислот, обладающая 5'->3'-экзонуклеазной активностью, как правило включает полимеразы нуклеиновых кислот, обладающие 5'→3'-экзонуклеазной активностью, и обычно включает полимеразы нуклеиновых кислот, обладающие наряду с 5'→3'-экзонуклеазной активностью дополнительно эндонуклеазной активностью.
Согласно предпочтительному воплощению TD-зонд содержит по меньшей мере одну метку в любом сайте последовательности, содержащей 1-10 нуклеотидов от его 5'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-5 нуклеотидов от его 5'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-3 нуклеотида от его 5'-конца. Наиболее предпочтительно, чтобы TD-зонд содержал по меньшей мере одну метку на своем 5'-конце.
Согласно предпочтительному воплощению стадию (а) выполняют с использованием TD-зонда вместе с прямым праймером, подлежащим гибридизации с сайтом, расположенным "вниз по течению" относительно сайта гибридизации TD-зонда, и ферментом, обладающим 5'→3'-экзонуклеазной активностью, является матричная полимераза нуклеиновых кислот, обладающая 5'→3'-экзонуклеазной активностью, так что прямой праймер удлиняется под действием матричной полимеразы нуклеиновых кислот на стадии (b).
Вслед за гибридизацией прямой праймер, гибридизованный с нуклеиновокислотной последовательностью-мишенью, удлиняется в результате проявления полимеразной активности матричной полимеразы нуклеиновых кислот, и осуществляется переваривание TD-зонда в результате проявления 5'→3'-экзонуклеазной активности для отделения флуоресцентной репортерной молекулы от молекулы гасителя с генерированием сигнала флуоресценции.
В соответствии с предпочтительным воплощением стадию (а) выполняют с использованием TD-зонда вместе с обратным праймером, и ферментом, обладающим 5'→3'-экзонуклеазной активностью, является матричная полимераза нуклеиновых кислот, обладающая 5'→3'-экзонуклеазной активностью, так что на стадии (b) получают нуклеиновокислотную последовательность-мишень, гибридизуемую с TD-зондом благодаря реакции удлинения обратного праймера под действием матричной полимеразы нуклеиновых кислот.
С помощью обратного праймера получают дополнительные нуклеиновокислотные последовательности-мишени для гибридизации с TD-зондом, результатом чего является получение более очевидных и сильных сигналов флуоресценции, указывающих на наличие нуклеиновокислотных последовательностей-мишеней.
Репортерная молекула и молекула-гаситель, полезные в настоящем изобретении, могут представлять собой флуоресцентные соединения. Полезными в данном изобретении являются репортерные молекулы и молекулы-гасители, известные в данной области техники. Примерами их являются: Сy2™ (506), YO-PRO™-1 (509), YOYO™-1 (509), кальцеин (517), FITC (флуоресцеинизотиоцианат) (518), FluorX™ (519), Alexa™ (520), родамин 110 (520), Oregon Green™ (Орегон зеленый) 500 (522), Oregon Green™ 488 (524), RiboGreen™ (525), Rhodamine Green™ (родаминовый зеленый) (527), родамин 123 (529), Magnesium Green™ (531), Calcium Green™ (533), TO-PRO™-1 (533), TOT01 (533), JOE (548), BODIPY530/550 (550), Dil (565), BODIPY TMR (568), BODIPY558/568 (568), BODIPY564/570 (570), Су3™ (570), Alexa™ 546 (570), TRITC (тетраметилродамин-изотиоцианат) (572), Magnesium Orange™ (575), фикоэритрин R&B (575), родамин-фаллоидин (575), Calcium Orange™ (576), пиронин Y (580), родамин В (580), TAMRA (тетраметилродамин) (582), Rhodamine Red™ (590), Cy3.5™ (596), ROX (6-карбокси-Х-родамин) (608), Calcium Crimson™ (615), Alexa™ 594 (615), техасский красный (615), нильский красный (628), YO-PRO™-3 (631), YOYO™-3 (631), R-фикоцианин (642), С-фикоцианин (648), TO-PRO™-3 (660), TOTО3 (660), DiD DilC(5) (665), Cy5™ (670), тиадикарбоцианин (671), Сy5.5 (694), HEX (гексахлорфлуоресцеин) (556), ТЕТ (тетрахлорфлуоресцеин) (536), Biosearch Blue (447), CAL Fluor Gold 540 (544), CAL Fluor Orange 560 (559), CAL Fluor Red 590 (591), CAL Fluor Red 610 (610), CAL Fluor Red 635 (637), FAM (520), флуоресцеин (520), флуоресцеин-С3 (520), Pulsar 650 (566), Quasar 570 (667), Quasar 670 (705) и Quasar 705 (610). Числа в скобках представляют собой длину волны, соответствующую максимуму излучения, в нанометрах.
Подходящие пары репортер-гаситель изложены в ряде публикаций, приведенных далее: в Pesce et al., editors, Fluorescence Spectroscopy (Marcel Dekker, New York, 1971); White et al., Fluorescence Analysis: A Practical Approach (Marcel Dekker, New York, 1970); Berlman, Handbook of Fluorescence Spectra of Aromatic Molecules, 2nd Edition (Academic Press, New York, 1971); Griffiths, Color and Constitution of Organic Molecules (Academic Press, New York, 1976); Bishop, editor, Indicators (Pergamon Press, Oxford, 1972); Haugland, Handbook of Fluorescent Probes and Research Chemicals (Molecular Probes, Eugene, 1992); Pringsheim, Fluorescence and Phosphorescence (Interscience Publishers, New York, 1949); Haugland, R. P., Handbook of Fluorescent Probes and Research Chemicals, 6th Edition, Molecular Probes, Eugene, Oreg., 1996; патентах США №№3996345 и 4351760.
Стоит отметить, что в настоящем изобретении может быть использована нефлуоресцентная "черная" молекула-гаситель, способная гасить флуоресценцию в широком диапазоне длин волн или на конкретной длине волны. Примерами таких гасителей являются BHQ и DABCYL (4-((4-(диметиламино)фенил)азо)бензойная кислота).
Для FRET-меток, адаптированных к TD-зонду, термин "репортерная молекула" включает в себя донора FRET, а термин "гаситель" включает в себя другого партнера (акцептора) FRET. Например, в качестве репортерной молекулы используют флуоресцентный краситель, а в качестве гасителя родаминовый краситель.
Используемый в данном описании термин "нуклеиновая кислота-мишень", "нуклеиновокислотная последовательность-мишень" или "последовательность-мишень" относится к представляющей интерес для детекции нуклеиновокислотной последовательности, на которой отжигают праймер или зонд либо которую гибридизуют с праймером или зондом в условиях гибридизации, отжига или амплификации.
Используемый в данном описании термин "зонд" относится к молекуле одноцепочечной нуклеиновой кислоты, содержащей участок или участки, по существу комплементарную(ые) нуклеиновокислотной последовательности-мишени. Зонды по данному изобретению могут содержать природный dNMP (дезоксинуклеозид-монофосфат) (то есть dAMP (дезоксиаденозин-монофосфат), dGM (дезоксигуанозин-монофосфат), dCMP (дезоксицитидин-монофосфат) и dTMP (дезокситимидин-монофосфат)), модифицированный нуклеотид или неприродный нуклеотид. Зонды также могут включать рибонуклеотиды.
Предпочтительно, 3'-конец меченого зонда блокируют, чтобы препятствовать удлинению зонда. Блокирование может быть достигнуто посредством использования некомплементарных оснований или путем добавления химической группировки, такой как биотин или фосфатная группа, к 3'-гидроксилу последнего нуклеотида. Блокирование также может быть достигнуто посредством удаления 3'-ОН или путем использования нуклеотида, лишенного 3'-ОН, такого как дидезоксинуклеотид.
Термин "праймер", как он использован в данном описании, относится к олигонуклеотиду, который способен действовать в качестве точки инициации синтеза в случае его помещения в условия, при которых индуцируется синтез продукта удлинения праймера, комплементарного цепи нуклеиновой кислоты (матрице), то есть в присутствии нуклеотидов и агента для полимеризации, такого как ДНК-полимераза, и при подходящих значениях температуры и рН. Предпочтительно праймер является одноцепочечным для максимальной эффективности амплификации. Предпочтительно, чтобы праймер представлял собой олигодезоксирибонуклеотид. Праймер по данному изобретению может содержать природный dNMP (то есть dAMP, dGM, dCMP и dTMP), модифицированный нуклеотид или неприродный нуклеотид. Праймер может также включать рибонуклеотиды.
Праймер должен иметь достаточную длину, чтобы праймировать синтез продуктов удлинения в присутствии агента для полимеризации. Точная длина праймеров будет зависеть от многих факторов, включая температуру, способ применения и источник праймера. Термин "отжиг" или "праймирование", как он использован в данном описании, относится к присоединению олигодезоксинуклеотида или нуклеиновой кислоты к являющейся матрицей нуклеиновой кислоте, при этом данное присоединению позволяет полимеразе осуществлять полимеризацию нуклеотидов с образованием молекулы нуклеиновой кислоты, которая является комплементарной к являющейся матрицей нуклеиновой кислоте или ее части.
Термин "гибридизация", используемый в данном описании, относится к образованию двухцепочечной нуклеиновой кислоты из комплементарных одноцепочечных нуклеиновых кислот. Не предполагается различия между терминами "отжиг" и "гибридизация", и эти термины будут использованы взаимозаменяемо.
Отжиг или гибридизация TD-зонда может быть осуществлен(а) с использованием широкого спектра способов гибридизации, известных специалистам в данной области. Подходящие условия гибридизации в настоящем изобретении могут быть определены в установленном порядке путем оптимизации методик. Такие условия, как температура, концентрация компонентов, продолжительность гибридизации и промывки, компоненты буферов, их рН и ионная сила, могут варьироваться в зависимости от различных факторов, включая длину и GC-состав олигонуклеотидов, таких как зонды и нуклеиновокислотные последовательности-мишени. Подробные условия гибридизации можно найти в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001); и M.L.M.Anderson, Nucleic Acid Hybridization, Springer-Verlag New York Inc. N.Y. (1999).
Согласно предпочтительному воплощению температура гибридизации TD-зонда находится в диапазоне от примерно 40°С до 80°С, более предпочтительно от 45°С до 75°С, еще более предпочтительно от 50°С до 72°С.
Используемый в данном описании термин "прямой праймер" относится к праймеру, предназначенному для гибридизации с сайтом, расположенным "вниз по течению" относительно сайта гибридизации TD-зонда, и предназначенному для образования комплементарной последовательности с нуклеиновокислотной последовательностью-мишенью с помощью матричной полимеразы нуклеиновых кислот.
TD-зонд, прямой праймер и обратный праймер каждый содержит гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени. Термин "комплементарный" используется в данном описании для указания на то, что праймеры или зонды являются в достаточной степени комплементарными, чтобы селективно гибридизоваться с нуклеиновокислотной последовательностью-мишенью в желаемых условиях отжига или в жестких условиях, охватывая термины "существенно комплементарный" и "полностью комплементарный", предпочтительно "полностью комплементарный".
Согласно предпочтительному воплощению 5'-концевой второй участок гибридизации TD-зонда является комплементарным нуклеиновокислотной последовательности-мишени. Другими словами, 5'-концевой второй участок гибридизации может содержать полностью совпадающую последовательность или неполностью совпадающую последовательность с нуклеиновокислотной последовательностью-мишенью. Если необходимо, 5'-концевой второй участок гибридизации может быть сконструирован содержащим некоторое количество ошибочно спаренных нуклеотидов.
В соответствии со специфическим воплощением данного изобретения 5'-концевой второй участок гибридизации TD-зонда может содержать от одного до трех дополнительных ошибочно спаренных нуклеотидов на своем 5'-конце. Сообщалось, что среди ферментов (например, полимераз нуклеиновых кислот), обладающих 5'→3'-экзонуклеазной активностью, есть ферменты, способные переваривать от одного до трех нуклеотидов с 5'-конца олигонуклеотидов, гибридизованных с последовательностями-мишенями (см. Murante et al., Journal of Biological Chemistry, Vol.269, 1191-1196 (1994) и Пример 1). Когда используют такие ферменты, TD-зонд может быть сконструирован содержащим от одного до трех искусственных ошибочно спаренных нуклеотидов на своем 5'-конце.
Нуклеиновокислотная последовательность-мишень, предназначенная для детекции в настоящем изобретении, включает любую молекулу нуклеиновой кислоты, например ДНК (гДНК (геномную ДНК) и кДНК (кодирующую ДНК)) и РНК. Нуклеиновокислотная последовательность-мишень включает любую природную прокариотическую, эукариотическую (например, из простейших и паразитов, грибов, дрожжей, высших растений, низших и высших животных, включая млекопитающих и людей) или вирусную (например, из вируса герпеса, ВИЧ, вируса гриппа, вируса Эпштейна-Барр, вируса гепатита, полиовируса и т.д.) или вироидную нуклеиновую кислоту.
Нуклеиновокислотные последовательности-мишени в образце могут представлять собой или ДНК, или РНК. Данная молекула может находиться или в двухцепочечной, или одноцепочечной форме. Когда нуклеиновая кислота в качестве исходного вещества является двухцепочечной, предпочтительно, чтобы две цепи были переведены в одноцепочечную или частично одноцепочечную форму. Известные методы разделения цепей включают нагревание, обработку щелочью, формамидом, мочевиной и глиоксалем, ферментативные методы (например, действие геликазы) и применение связывающих белков, но этим не ограничиваются. Например, разделения цепей можно достичь посредством нагревания при температуре, изменяющейся в диапазоне от 80°С до 105°С. Общие методы осуществления такой обработки приведены в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001).
Когда в качестве исходного вещества используют мРНК, перед проведением стадии отжига необходима стадия обратной транскрипции, детали проведения которой приводятся в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001); и Noonan, K. F. et al., Nucleic Acids Res., 16: 10366 (1988)). Для проведения обратной транскрипции используют случайный гексамер или олигонуклеотидный олиго(dT)-содержащий праймер, гибридизуемый с поли-А-хвостом мРНК. Олигонуклеотидный onnro(dT)-содержащий праймер состоит из dTMP, один или более чем один из которых могут быть заменены на другие dNMP до тех пор, пока данный олиго(dT)-содержащий праймер может служить в качестве праймера. Обратная транскрипция может быть осуществлена с использованием обратной транскриптазы, обладающей активностью РНКазы Н. При использовании фермента, обладающего активностью РНКазы Н, можно исключить отдельную стадию переваривания РНКазой Н, тщательно выбирая реакционные условия.
Зонды или праймеры, используемые в настоящем изобретении, гибридизуют или отжигают с сайтами нуклеиновокислотных последовательностей-мишеней (в качестве матриц), в результате чего образуется двухцепочечная структура. Условия гибридизации или отжига нуклеиновых кислот, подходящие для образования таких двухцепочечных структур, описываются в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001) и Haymes, В. D., et al., Nucleic Acid Hybridization, A Practical Approach, IRL Press, Washington, D.C. (1985).
Согласно предпочтительному воплощению прямой праймер и/или обратный праймер имеет структуру с двойной специфичностью (DSO), представленную следующей общей формулой II:

где Хр представляет собой 5'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; Yq представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Zr представляет собой 3'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; р, q и r представляют количество нуклеотидов, и X, Y и Z представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого первого участка гибридизации выше, чем у 3'-концевого второго участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков; разделительный участок отделяет 5'-концевой первый участок гибридизации от 3'-концевого второго участка гибридизации с точки зрения событий гибридизации с нуклеиновой кислотой-мишенью, в силу чего специфичность гибридизации олигонуклеотида определяется двойственно 5'-концевым первым участком гибридизации и 3'-концевым вторым участком гибридизации, так что общая специфичность гибридизации олигонуклеотида является повышенной.
Описания структуры DSO могут быть сделаны со ссылкой на структуру mDSO.
Предпочтительно, чтобы в структуре DSO длина 5'-концевого первого участка гибридизации была больше, чем у 3'-концевого второго участка гибридизации. Длина 5'-концевого первого участка гибридизации составляет предпочтительно 15-60 нуклеотидов, более предпочтительно 15-40 нуклеотидов, еще более предпочтительно 15-25 нуклеотидов. Предпочтительно, чтобы длина 3'-концевого второго участка гибридизации составляла 3-15 нуклеотидов, более предпочтительно 5-15 нуклеотидов, еще более предпочтительно 6-13 нуклеотидов. Длина разделительного участка составляет предпочтительно 3-10 нуклеотидов, более предпочтительно 4-8 нуклеотидов, наиболее предпочтительно 5-7 нуклеотидов. Согласно предпочтительному воплощению Тпл 5'-концевого первого участка гибридизации находится в диапазоне от 40°С до 80°С, более предпочтительно от 45°С до 65°С. Тпл 3'-концевого второго участка гибридизации предпочтительно находится в диапазоне от 10°С до 40°С. Предпочтительно, когда Тпл разделительного участка находится в диапазоне от 3°С до 15°С.
Предпочтительно, фермент, обладающий 5'→3'-экзонуклеазной активностью, и матричная полимераза нуклеиновых кислот, обладающая 5'→3'-экзонуклеазной активностью, используемые в настоящем изобретении, могут представлять собой любую матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной активностью (например, ДНК-полимеразу I Е. соli, термостабильную ДНК-полимеразу и ДНК-полимеразу бактериофага Т7), наиболее предпочтительно термостабильную ДНК-полимеразу, полученную из ряда бактериальных видов, включая Thermus aquaticus (Taq), Thermus thermophilus, Thermus filiformis, Thermus flavus, Thermus antranikianii, Thermus caldophilus, Thermus chliarophilus, Thermus igniterrae, Thermus lacteus, Thermus oshimai, Thermus ruber, Thermus rubens, Thermus scotoductus, Thermus silvanus, Thermus species Z05 и Thermus species sps 17. Наиболее предпочтительно, чтобы матричная полимераза нуклеиновых кислот, обладающая 5'→3'-экзонуклеазной активностью, представляла собой ДНК-полимеразу Taq.
Под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, (предпочтительно, матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-экзонуклеазной активностью) TD-зонд расщепляется, и генерируется сигнал, указывающий на наличие нуклеиновокислотной последовательности-мишени. Детекцию или измерение сигнала можно осуществить традиционными для каждой метки методами. Например, детекцию или измерение сигнала флуоресценции можно осуществить традиционными методами, например, с использованием флуориметров.
Термин "генерация сигнала" или "генерирование сигнала", используемый в данном описании, охватывает изменение интенсивности сигнала флуоресценции, включая не только увеличение интенсивности сигнала флуоресценции, но также уменьшение интенсивности сигнала флуоресценции. Согласно предпочтительному воплощению подлежащий детекции сигнал, указывающий на наличие нуклеиновокислотной последовательности-мишени, представляет собой сигнал от флуоресцентной репортерной молекулы. В альтернативном случае флуоресцентной является молекула-гаситель, и подлежащий детекции сигнал, указывающий на наличие нуклеиновокислотной последовательности-мишени, представляет собой сигнал от флуоресцентной молекулы-гасителя.
Когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5'-концевой второй участок гибридизации не переваривается под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, результатом чего является отсутствие сигнала флуоресценции.
Термин "отсутствие сигнала флуоресценции" относится к случаю отсутствия сигнала флуоресценции, а также к случаю пренебрежимо малого сигнала флуоресценции. Например, термин охватывает интенсивность флуоресценции, обычно измеряемую или наблюдаемую в случае отрицательного контроля или фона.
Согласно предпочтительному воплощению настоящее изобретение дополнительно включает повторение стадий (а)-(b) или (а)-(с), и при повторении стадий (a)-(b) или (а)-(с) настоящее изобретение дополнительно включает стадию денатурации между повторяющимися циклами.
Методы денатурации включают нагревание, обработку щелочью, формамидом, мочевиной и глиоксалем, ферментативные методы (например, действие геликазы) и применение связывающих белков, но этим не ограничиваются. Например, денатурации можно достичь посредством нагревания при температуре, изменяющейся в диапазоне от 80°С до 105°С. Общие способы осуществления такой обработки приведены в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001).
Повторение дает возможность увеличить интенсивность сигнала флуоресценции от флуоресцентной репортерной молекулы. В частности, повторение, согласно способу по настоящему изобретению с использованием обратных праймеров, дает возможность увеличить количество нуклеиновокислотной последовательности-мишени, вносящей вклад в увеличение интенсивности сигнала флуоресценции от флуоресцентной репортерной молекулы.
Согласно предпочтительному воплощению используемая нуклеиновокислотная последовательность-мишень представляет собой предварительно амплифицированную с применением амплифицирующего праймера нуклеиновокислотную последовательность.
Такая предварительно амплифицированная нуклеиновокислотная последовательность-мишень может включать нуклеиновокислотную последовательность-мишень, предварительно амплифицированную в других реакционных условиях (или другом реакционном сосуде), нежели реакционные условия (или реакционный сосуд) для стадий (а)-(с).
Когда настоящее изобретение дополнительно включает повторение стадий (а)-(b) или (а)-(с), предпочтительно, чтобы детекция сигналов осуществлялась для каждого цикла такого повторения (то есть в режиме реального времени), в конце повторения (то есть в режиме конечной точки) или в каждый из предварительно заданных интервалов времени в процессе повторения. Предпочтительно, чтобы детекция сигнала могла быть осуществлена для каждого цикла повторения для улучшения точности детекции.
Согласно предпочтительному воплощению амплифицирующий праймер (например, включая прямой праймер и обратный праймер) для получения предварительно амплифицированных нуклеиновокислотных последовательностей-мишеней имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную общей формулой II, описанной выше.
Согласно предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью. Согласно предпочтительному воплощению блокирующий сайт расположен в сайте TD-зонда, расщепляемом ферментом, обладающим 5'→3'-экзонуклеазной активностью, и предпочтительно в 3'-концевом участке гибридизации TD-зонда.
Когда используют фермент, обладающий 5'→3'-экзонуклеазной и эндонуклеазной активностями, (например, матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной и эндонуклеазной активностями), для более точного определения метки в 3'-концевой первый участок гибридизации TD-зонда может быть введена молекула-блокатор для блокирования катализируемого эндонуклеазной активностью переваривания 3'-концевого первого участка гибридизации TD-зонда, гибридизующегося с нуклеиновокислотной последовательностью, не являющейся мишенью.
Согласно предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, и этот блокирующий сайт расположен в сайте, подлежащем расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью; при этом, когда TD-зонд, содержащий блокирующий сайт, гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд, содержащий блокирующий сайт, гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, он не переваривается ферментом, обладающим экзонуклеазной активностью, и никакого сигнала флуоресценции не генерируется.
Согласно предпочтительному воплощению блокирующий сайт TD-зонда расположен на 3'-концевом первом участке гибридизации TD-зонда. Более предпочтительно, чтобы блокирующий сайт TD-зонда был расположен на 3'-концевом первом участке гибридизации, прилегающем к 3'-концу разделительного участка.
Согласно предпочтительному воплощению блокирующий сайт содержит 1-15 молекул-блокаторов, более предпочтительно 2-10 молекул-блокаторов, еще более предпочтительно 3-5 молекул-блокаторов.
Нуклеотиды, служащие в качестве молекул-блокаторов, то есть имеющие остов, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, включают любые нуклеотиды, известные специалисту в данной области. Например, они содержат различные фосфоротиоатные связи, фосфонатные связи, фосфороамидатные связи и углеводы с модификациями по 2'-положению. Согласно предпочтительному воплощению нуклеотиды, имеющие остов, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, содержат фосфоротиоатные связи, алкилфосфотриэфирную связь, арилфосфотриэфирную связь, алкилфосфонатную связь, арилфосфонатную связь, гидрофосфонатную связь, алкилфосфороамидатную связь, арилфосфороамидатную связь, фосфороселенатную связь, модификацию 2'-O-аминопропил, модификацию 2'-O-алкил, модификацию 2'-O-аллил, модификацию 2'-O-бутил, α-аномерный олигодезоксинуклеотид и модификацию 1-(4'-тио-β-D-рибофуранозил). Блокирующий нуклеотид может быть представлен в составе TD-зонда один или большее количество раз непрерывно или дискретно.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, и TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов, и прямой праймер включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов, и обратный праймер включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров.
Кроме того, настоящее изобретение является очень полезным для детекции вариабельности нуклеотидов. Термин "вариабельность нуклеотидов", используемый в данном описании, относится к нуклеотидному полиморфизму в последовательности ДНК в конкретном положении среди непрерывных сегментов ДНК, которые в других случаях являются одинаковыми в последовательности. Такие непрерывные сегменты ДНК включают ген или любую другую часть хромосомы. Например, вариабельность нуклеотидов, детектируемая в настоящем изобретении, включает SNP (однонуклеотидный полиморфизм), делецию, вставку, замену и транслокацию. Примером вариабельности нуклеотидов являются многочисленные вариации в геноме человека (например, вариации в гене MTHFR (метилентетрагидрофолат-редуктазы)), вариации, вовлеченные в возникновение лекарственной устойчивости у патогенных микроорганизмов и приводящие к онкогенезу вариации.
Согласно предпочтительному воплощению вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации TD-зонда.
2. Способ детекции мишени на твердой фазе
Настоящее изобретение прекрасно адаптировано для применения на твердой фазе (например, микрочипе), а также в жидкой фазе.
В другом аспекте данного изобретения предложен способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот на твердой фазе с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с TD-зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд иммобилизован через свой 3'-конец на поверхности твердой подложки; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
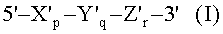
где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд имеет метку, генерирующую детектируемый сигнал, и эта метка расположена на 5'-концевом втором участке гибридизации TD-зонда; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, а 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью;
(b) приведения в контакт продукта со стадии (а) с ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, с высвобождением метки из TD-зонда, в результате чего изменяется сигнал от TD-зонда, иммобилизованного на твердой подложке; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активность, в результате чего не происходит никакого изменения сигнала, в силу чего изменение сигнала на твердой подложке детектируют для определения наличия нуклеиновокислотной последовательности-мишени; и
(с) детекции изменения сигнала на твердой подложке, так что изменение сигнала в результате переваривания на 5'-концевом втором участке гибридизации указывает на наличие нуклеиновокислотной последовательности-мишени.
Поскольку в способе по настоящему изобретению на твердой фазе используется TD-зонд и применяются стадии способа по настоящему изобретению, описанного для осуществления в жидкой фазе, общее для них описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
Для проведения реакции на твердой фазе TD-зонд может быть иммобилизован непосредственно или опосредованно (предпочтительно опосредованно) через свой 3'-конец на поверхности твердой подложки. Кроме того, иммобилизацию зондов на поверхности твердой подложки можно проводить ковалентным или нековалентным образом. В тех случаях, когда иммобилизованные зонды представляют собой зонды, иммобилизованные на поверхности твердой подложки, используют подходящие линкеры. Линкеры, полезные в данном изобретении, могут включать любые линкеры, используемые для иммобилизации зондов на микрочипе. Например, алкильные или арильные соединения с аминной функциональной группой либо алкильные или арильные соединения с тиоловой функциональной группой служат в качестве линкеров для иммобилизации зонда. Помимо этого, в качестве линкеров для сведения к минимуму стерических препятствий в отношении ферментативной реакции (например, реакций ферментативного расщепления) или для повышения эффективности гибридизации, могут быть использованы производные оснований поли-(Т) или поли-(А). Следует понимать, что производные оснований поли-(Т) или поли-(А) не рассматриваются как последовательности, входящие в состав TD-зонда. Например, производные оснований поли-(Т) или поли-(А), соединенные с концом 3'-концевого первого участка гибридизации TD-зонда, не рассматриваются как часть 3'-концевого первого участка гибридизации.
Согласно предпочтительному воплощению твердая подложка, используемая в настоящем изобретении, представляет собой микрочип. Микрочип, обеспечивающий соблюдение условий реакции в данном изобретении, может включать любой из микрочипов, которые известны специалисту в данной области. Все способы по настоящему изобретению, то есть отжиг с нуклеиновой кислотой-мишенью, удлинение/переваривание и детекция флуоресценции, осуществляют на микрочипе. Иммобилизованные на микрочипе зонды служат в качестве гибридизуемых элементов чипа. Твердая подложка для изготовления микрочипа включает металлы (например, золото, сплав золота и меди, аллюминий), оксид металла, стекло, керамику, кварц, кремний, полупроводник, пластинку из Si/SiO2, германий, арсенид галия, углерод, углеродную нанотрубку, полимеры (например, полистирол, полиэтилен, полипропилен и полиакриламид), сефарозу, агарозу и коллоиды, но этим не ограничивается. Большинство используемых в данном изобретении иммобилизованных зондов может быть иммобилизовано на доступном участке или двух или более доступных участках на твердой подложке, которая может содержать 2-1000000 доступных участков. Чтобы получить микрочип или микрочипы для заданного применения, иммобилизованные зонды могут быть изготовлены с использованием традиционных технологий изготовления, таких как фотолитография, технология струйной печати, механическое точечное нанесение пятен и их производные.
Согласно предпочтительному воплощению фермент, обладающий 5'→3'-экзонуклеазной активностью, представляет собой термостабильный фермент. Согласно предпочтительному воплощению фермент, обладающий 5'→3'-экзонуклеазной активностью, представляет собой матричную полимеразу нуклеиновых кислот, более предпочтительно термостабильную матричную полимеразу нуклеиновых кислот.
Согласно предпочтительному воплощению стадию (а) выполняют с использованием TD-зонда вместе с прямым праймером, подлежащим гибридизации с сайтом, расположенным "вниз по течению" относительно сайта гибридизации TD-зонда, и ферментом, обладающим 5'→3'-экзонуклеазной активностью, является матричная полимераза нуклеиновых кислот, обладающая 5'→3'-экзонуклеазной активностью, так что прямой праймер удлиняется под действием матричной полимеразы нуклеиновых кислот на стадии (b).
Согласно предпочтительному воплощению стадию (а) выполняют с использованием TD-зонда вместе с обратным праймером, и ферментом, обладающим 5'→3'-экзонуклеазной активностью, является матричная полимераза нуклеиновых кислот, обладающая 5'→3'-экзонуклеазной активностью, так что на стадии (b) получают нуклеиновокислотную последовательность-мишень, гибридизуемую с TD-зондом благодаря реакции удлинения обратного праймера под действием матричной полимеразы нуклеиновых кислот.
Согласно предпочтительному воплощению метка представляет собой химическую метку, ферментативную метку, радиоактивную метку, флуоресцентную метку, интерактивную метку, люминесцентную метку, хемилюминесцентную метку или металлическую метку.
Как показано на Фиг.4 или 5, способ по настоящему изобретению на твердой фазе может быть осуществлен с использованием единственной метки (например, одной флуоресцентной метки) или интерактивной метки (например, репортерной молекулы и молекулы-гасителя).
Например, когда для детекции нуклеиновокислотной последовательности-мишени используют TD-зонд, имеющий единственную флуоресцентную метку, то данная флуоресцентная метка, находящаяся на 5'-концевом втором участке гибридизации, высвобождается из TD-зонда, иммобилизованного на твердой подложке, что приводит к уменьшению интенсивности сигнала флуоресценции на твердой подложке. Уменьшение или исчезновение сигнала флуоресценции может указывать на наличие нуклеиновокислотной последовательности-мишени.
Согласно предпочтительному воплощению, когда такая единственная метка представляет собой флуоресцентную репортерную молекулу, изменение сигнала представляет собой уменьшение или исчезновение сигналов флуоресценции на твердой подложке.
Согласно предпочтительному воплощению, когда используют TD-зонд, имеющий единственную флуоресцентную метку, перед детекцией на стадии (с) возможно проводят дополнительно стадию промывки. Альтернативно, когда используют TD-зонд, имеющий единственную флуоресцентную метку, перед детекцией на стадии (с) стадию промывки не проводят.
Согласно предпочтительному воплощению такая единственная флуоресцентная молекула расположена в сайте на 5'-концевом втором участке гибридизации, подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Для ясности: должно быть очевидно, что фраза "сайт на 5'-концевом втором участке гибридизации, подлежащий перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью", означает, что перевариванию под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, может подвергаться весь участок, часть участка или положение на 5'-концевом втором участке гибридизации, а метка может быть расположена в любом одном сайте, подлежащем перевариванию на 5'-концевом втором участке гибридизации. Поэтому фраза "сайт на 5'-концевом втором участке гибридизации, подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью", может быть записана как "сайт, подлежащий перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью, на 5'-концевом втором участке гибридизации".
Более предпочтительно, чтобы эта единственная флуоресцентная молекула была расположена в любом сайте последовательности, содержащей 1-10 нуклеотидов от 5'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-5 нуклеотидов от 5'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-3 нуклеотида от 5'-конца TD-зонда. Наиболее предпочтительно, чтобы эта единственная флуоресцентная молекула была расположена на 5'-конце TD-зонда.
Согласно предпочтительному воплощению метка представляет собой систему взаимодействующих меток, содержащую пару, состоящую из флуоресцентной репортерной молекулы и молекулы-гасителя.
Согласно предпочтительному воплощению одна молекула, выбранная из репортерной молекулы и молекулы-гасителя, расположена в сайте на 5'-концевом втором участке гибридизации TD-зонда, а другая в сайте, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению одна молекула, выбранная из репортерной молекулы и молекулы-гасителя, расположена в сайте, подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью, на 5'-концевом втором участке гибридизации TD-зонда, а другая в сайте, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению сайт, не подлежащий перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью, может находиться на 5'-концевом втором участке гибридизации, разделительном участке или 3'-концевом первом участке гибридизации TD-зонда.
Согласно предпочтительному воплощению, когда способ по настоящему изобретению осуществляют на твердой фазе, TD-зонд иммобилизован через свой 3'-конец на поверхности твердой подложки; при этом молекула-гаситель расположена в сайте на 5'-концевом втором участке гибридизации TD-зонда, подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью, а флуоресцентная репортерная молекула расположена в сайте, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции от репортерной молекулы; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, в результате чего сигнал флуоресценции будет отсутствовать, тем самым осуществляется детекция сигнала флуоресценции на твердой подложке для определения наличия нуклеиновокислотной последовательности-мишени.
Когда иммобилизованный TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, он переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, по направлению от 5'-конца к 3'-концу. На данном этапе связывание между фрагментом иммобилизованного TD-зонда и нуклеиновой кислотой-мишенью становится слабым, что приводит к отсоединению нуклеиновой кислоты-мишени от фрагмента TD-зонда, иммобилизованного на твердой подложке. В этом смысле можно считать, что иммобилизованный TD-зонд состоит из двух частей, перевариваемой части и неперевариваемой части под действием фермента, обладающего 5'→3'-экзонуклеазной активностью. Ввиду этого, метка, расположеннная на неперевариваемой части зонда, остается на поверхности твердой подложки.
Принимая во внимание характер переваривания иммобилизованного TD-зонда, можно понять, что TD-зонд также может содержать перевариваемую часть и неперевариваемую часть внутри 5'-концевого второго участка гибридизации. На образование перевариваемой части и неперевариваемой части внутри TD-зонда может влиять разделительный участок.
Согласно предпочтительному воплощению, когда в сайт TD-зонда между молекулой-гасителем и репортерной молекулой вводят молекулу-блокатор, такую как модифицированные нуклеотиды или остовы, устойчивые к проявлению 5'→3'-экзонуклеазной активности, тогда фермент, обладающий 5'→3'-экзонуклеазной активностью, не способен далее переваривать TD-зонд ввиду наличия молекулы-блокатора, так что фрагмент иммобилизованного TD-зонда, содержащий одну метку, остается на твердой подложке как неперевариваемая часть.
Согласно предпочтительному воплощению молекула-гаситель расположена на 5'-концевом втором участке гибридизации, подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула расположена на 5'-концевом втором участке гибридизации, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула расположена на 3'-концевом первом участке гибридизации, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула, расположенная в сайте, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью, остается на поверхности твердой подложки после выполнения стадии (b), что позволяет удобно детектировать сигнал флуоресценции от репортерной молекулы в режиме реального времени без стадий промывки.
Предпочтительно, чтобы молекула-гаситель была расположена на 5'-конце TD-зонда или на расстоянии 1-3 нуклеотидов от 5'-конца, а флуоресцентная репортерная молекула была расположена вплотную к 3'-концу TD-зонда или в середине 3'-концевого первого участка гибридизации TD-зонда.
Согласно предпочтительному воплощению молекула-гаситель расположена в любом сайте последовательности, содержащей 1-10 нуклеотидов от 5'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-5 нуклеотидов от 5'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-3 нуклеотида от 5'-конца TD-зонда. Наиболее предпочтительно, чтобы молекула-гаситель была расположена на 5'-конце TD-зонда.
Согласно предпочтительному воплощению репортерная молекула расположена в любом сайте последовательности, содержащей 1-30 нуклеотидов от 3'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-20 нуклеотидов от 3'-конца, еще более предпочтительно в любом сайте последовательности, содержащей 1-15 нуклеотидов от 3'-конца TD-зонда.
Согласно предпочтительному воплощению прямой праймер и/или обратный праймер имеет структуру с двойной специфичностью (DSO), представленную приведенной выше общей формулой II.
Согласно предпочтительному воплощению настоящее изобретение дополнительно включает повторение стадий (а)-(b) или (а)-(с), и при повторении стадий (а)-(b) или (а)-(с) настоящее изобретение дополнительно включает стадию денатурации между повторяющимися циклами.
Когда настоящее изобретение дополнительно включает повторение стадий (а)-(b) или (а)-(с), предпочтительно, чтобы детекция сигналов осуществлялась для каждого цикла такого повторения (то есть в режиме реального времени), в конце повторения (то есть в режиме конечной точки) или в каждый из предварительно заданных интервалов времени в процессе повторения. Предпочтительно, чтобы детекция сигнала могла быть осуществлена для каждого цикла повторения для улучшения точности детекции и также для количественного определения нуклеиновой кислоты-мишени.
Согласно предпочтительному воплощению используемая нуклеиновокислотная последовательность-мишень представляет собой предварительно амплифицированную с применением амплифицирующего праймера нуклеиновокислотную последовательность.
Согласно предпочтительному воплощению амплифицирующий праймер (например, включая прямой праймер и обратный праймер) для получения предварительно амплифицированных нуклеиновокислотных последовательностей-мишеней, имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную общей формулой II, описанной выше.
Согласно предпочтительному воплощению стадии (а) и (b) проводят одновременно с амплификацией нуклеиновокислотной последовательности-мишени для детекции нуклеиновокислотной последовательности-мишени в режиме реального времени.
Согласно предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью. Согласно предпочтительному воплощению блокирующий сайт расположен в сайте TD-зонда, расщепляемом ферментом, обладающим 5'→3'-экзонуклеазной активностью, и предпочтительно в 3'-концевом участке гибридизации TD-зонда.
Согласно другому предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый в отношении 5'→3'-экзонуклеазной активности фермента (например, матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-экзонуклеазной активностью), и блокирующий сайт расположен в сайте, расщепляемом в результате проявления эндонуклеазной активности фермента, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью
Согласно предпочтительному воплощению блокирующий сайт TD-зонда расположен на 3'-концевом первом участке гибридизации TD-зонда. Более предпочтительно, чтобы блокирующий сайт TD-зонда был расположен на 3'-концевом первом участке гибридизации, примыкающем к 3'-концу разделительного участка.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, и TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов, и прямой праймер включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов, и обратный праймер включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров.
Кроме того, настоящее изобретение является очень полезным для детекции вариабельности нуклеотидов.
Согласно предпочтительному воплощению вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации TD-зонда.
II. Предпочтительное воплощение: ПЦР-анализ в режиме реального времени с использованием TD-зонда
Настоящее изобретение предпочтительно осуществляют одновременно с амплификацией нуклеиновокислотной последовательности-мишени, используя в качестве прямого праймера и обратного праймера пару праймеров, состоящую из двух праймеров, способных амплифицировать нуклеиновокислотную последовательность-мишень. Амплификацию предпочтительно осуществляют в соответствии с ПЦР (полимеразной цепной реакцией), описанной в патентах США №№4683195, 4683202 и 4800159.
В еще одном другом аспекте данного изобретения предложен способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) и полимеразной цепной реакции (ПЦР), включающий стадии:
(а) приготовления смеси для ПЦР, содержащей (1) нуклеиновокислотную последовательность-мишень, (2) TD-зонд, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, (3) пару праймеров, состоящую из двух праймеров в качестве прямого праймера и обратного праймера, каждый из которых имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (4) матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной активностью; где TD-зонд гибридизуется с сайтом, расположенным между двумя этими праймерами; при этом TD-зонд содержит структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:

где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит две метки - флуоресцентную репортерную молекулу и молекулу-гаситель, способную гасить флуоресценцию репортерной молекулы; флуоресцентная репортерная молекула и молекула-гаситель обе расположены на 5'-концевом втором участке гибридизации, или репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и разделительного участка; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков X'p, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, и 5'-концевой второй участок гибридизации переваривается в результате проявления 5'→3'-экзонуклеазной активности матричной полимеразы нуклеиновых кислот; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5'-концевой второй участок гибридизации не переваривается в результате проявления 5'→3'-экзонуклеазной активности матричной полимеразы нуклеиновых кислот, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
(b) амплификации нуклеиновокислотной последовательности-мишени с использованием смеси для ПЦР посредством проведения по меньшей мере двух циклов отжига праймеров, удлинения праймеров и денатурации, при этом данные два праймера удлиняются под действием полимеразной активности матричной полимеразы нуклеиновых кислот для амплификации нуклеиновокислотной последовательности-мишени; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, 5'-концевой второй участок гибридизации переваривается в результате проявления 5'→3'-экзонуклеазной активности матричной полимеразы нуклеиновых кислот с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5'-концевой второй участок гибридизации не переваривается в результате проявления 5'→3'-экзонуклеазной активности матричной полимеразы нуклеиновых кислот, так что флуоресцентная репортерная молекула не отделяется от молекулы-гасителя на TD-зонде, в результате чего сигнал флуоресценции будет отсутствовать; и
(с) детекции сигнала флуоресценции, так что генерируемый сигнал флуоресценции указывает на наличие нуклеиновокислотной последовательности-мишени.
Поскольку в применяемом в настоящем изобретении ПЦР-анализе в режиме реального времени используется TD-зонд и применяются стадии способа по настоящему изобретению, описанного выше, общее для них описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
В ПЦР-анализе в режиме реального времени с использованием 5'→3'-нуклеолитических реакций матричные полимеразы нуклеиновых кислот, обладающие 5'→3'-экзонуклеазной активностью, применяются для амплификации мишени, а также для генерации сигнала (например, в способе с использованием TaqMan-зонда). Как описано выше, матричная полимераза нуклеиновых кислот может обладать двумя видами нуклеолитической активности, включая 5'→3'-экзонуклеазную активность и эндонуклеазную активность. Эндонуклеазная активность может обуславливать генерацию ложноположительных сигналов в процессах, сопровождающих амплификацию мишени.
Чтобы полностью преодолеть проблемы и затруднения, связанные с эндонуклеазной активностью, в настоящем изобретении принята на вооружение уникальная стратегия, согласно которой все сдвоенные метки расположены на 5'-концевом втором участке гибридизации TD-зонда.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула и молекула-гаситель обе расположены на 5'-концевом втором участке гибридизации TD-зонда, или флуоресцентная репортерная молекула и молекула-гаситель - каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и разделительного участка TD-зонда, при проведении ПЦР в режиме реального времени.
Даже если эндонуклеазная активность матричной полимеразы нуклеиновых кислот может проявляться в сайте разветвления, образующемся на 3'-концевом первом участке гибридизации, когда TD-зонд гибридизуется с нуклеиновокислотными последовательностями, не являющимися мишенями, в процессе ПЦР в режиме реального времени, флуоресцентная репортерная молекула и молекула-гаситель, расположенные на 5'-концевом втором участке гибридизации, не отделяются друг от друга, так что сигнал флуоресценции от флуоресцентной репортерной молекулы не генерируется в результате проявления эндонуклеазной активности.
В этом смысле, используемый в настоящем изобретении ПЦР-анализ в режиме реального времени полностью обеспечивает устранение любых возможностей генерации ложных сигналов.
Согласно предпочтительному воплощению детекцию сигналов осуществляют для каждого цикла повторения (то есть в режиме реального времени), в конце повторения (то есть в режиме конечной точки) или в каждый из предварительно заданных интервалов времени в процессе повторения. Предпочтительно, чтобы детекция сигнала могла быть осуществлена для каждого цикла повторения для улучшения точности детекции.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов, прямой праймер включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров, и обратный праймер включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень характеризуется вариабельностью нуклеотидов.
Согласно предпочтительному воплощению вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации TD-зонда.
Согласно предпочтительному воплощению прямой праймер и/или обратный праймер имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную описанной выше общей формулой II.
III. Способ детекции мишени с использованием реакции лигирования в жидкой фазе или на твердой фазе
В следущем аспекте данного изобретения предложен способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с первым зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную первому сайту нуклеиновокислотной последовательности-мишени, и вторым зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную второму сайту нуклеиновокислотной последовательности-мишени, который расположен "вверх по течению" относительно первого сайта; где по меньшей мере один зонд, выбранный из первого зонда или второго зонда, содержит метку для генерации детектируемого сигнала; где второй зонд представляет собой TD-зонд; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
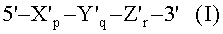
где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной; при этом, когда второй зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации второго зонда оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, что позволяет осуществлять лигирование первого зонда и второго зонда; при этом, когда второй зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок второго зонда оба образуют одиночную цепь, так что первый зонд и второй зонд не подвергаются лигированию, на основании чего второй зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью;
(b) лигирования первого зонда и второго зонда, гибридизованных с нуклеиновокислотной последовательностью-мишенью, с получением лигированного зонда;
(c) денатурации продукта со стадии (b);
(d) детекции сигнала от метки на лигированном зонде, так что сигнал указывает на наличие нуклеиновокислотной последовательности-мишени.
Поскольку в настоящем способе с применением реакции лигирования используется TD-зонд, общее описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
Способ по настоящему изобретению может быть осуществлен в жидкой фазе или на твердой фазе. Способ по настоящему изобретению предпочтительно осуществляют на твердой фазе.
В способе по настоящему изобретению с использованием реакции лигирования первоначально гибридизуют первый зонд и второй зонд с нуклеиновокислотной последовательностью-мишенью. Вторым зондом является описанный выше TD-зонд. Первый зонд имеет гибридизующуюся нуклеотидную последовательность, комплементарную первому сайту нуклеиновокислотной последовательности-мишени, а второй зонд имеет гибридизующуюся нуклеотидную последовательность, комплементарную второму сайту нуклеиновокислотной последовательности-мишени, который расположен "вверх по течению" относительно первого сайта. Первый зонд и второй зонд должны гибридизоваться с нуклеиновокислотной последовательностью-мишенью в порядке, описанном выше. Кроме тех случаев, когда положения гибридизации первого зонда и второго зонда удовлетворяют приведенным выше, специфичной к мишени детекции согласно настоящему изобретению не осуществляется.
Согласно предпочтительному воплощению первый зонд и второй зонд расположены в непосредственной близости по отношению друг к другу, когда они гибридизуются с нуклеиновокислотной последовательностью-мишенью.
Близкое расположение необходимо для осуществления реакции лигирования между данными двумя зондами. Используемый в данном описании термин "близкий" в отношении положений гибридизации первого зонда и второго зонда означает, что 3'-конец одного зонда и 5'-конец другого зонда находятся достаточно близко относительно друг друга, что позволяет соединить данные концы обоих зондов друг с другом.
Согласно предпочтительному воплощению 3'-конец первого зонда содержит гидроксильную группу, а 5'-конец второго зонда содержит фосфатную группу.
Подразумевается, что используемый в данном описании термин "в непосредственной близости" в отношении положений гибридизации первого зонда и второго зонда относится к достаточной близости между двумя зондами, позволяющей привести в соприкосновение 5'-конец второго с 3'-концом первого зонда таким образом, что они могут быть лигированы подходящим агентом, таким как лигаза. Когда 5'-конец второго зонда отстоит на 0 нуклеотидов от 3'-конца первого зонда, между обоими зондами образуется одноцепочечный разрыв, который будет лигирован лигазой.
Либо первый зонд, либо второй зонд имеет метку для генерирования детектируемого сигнала. Альтернативно, метку содержат как первый зонд, так и второй зонд.
Согласно предпочтительному воплощению данная метка представляет собой химическую метку, ферментативную метку, радиоактивную метку, флуоресцентную метку, интерактивную метку, люминесцентную метку, хемилюминесцентную метку или металлическую метку.
Более предпочтительно, чтобы метка представляла собой систему взаимодействующих меток, содержащую пару, состоящую из репортерной молекулы и молекулы-гасителя. Например, первый зонд содержит в качестве метки или репортерную молекулу, или молекулу-гаситель, а второй зонд содержит в качестве метки или молекулу-гаситель, или репортерную молекулу.
Согласно предпочтительному воплощению первый зонд имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную описанной выше общей формулой II.
Более предпочтительно, чтобы первый зонд имел структуру DSO, а второй зонд представлял собой TD-зонд и чтобы 3'-конец первого зонда располагался в непосредственной близости к 5'-концу второго зонда.
Точность при детекции последовательностей-мишеней с использованием зондов, как правило, зависит от специфичности зондов к последовательностям-мишеням. Если используют первый зонд, имеющий структуру DSO, и второй зонд, имеющий структуру mDSO (TD-зонд), то 3'-концевой второй участок гибридизации первого зонда и 5'-концевой второй участок гибридизации второго зонда будут располагаться в непосредственной близости по отношению друг к другу, когда эти два зонда гибридизуются с нуклеиновокислотной последовательностью-мишенью. Тогда происходит лигирование 3'-концевого второго участка гибридизации первого зонда и 5'-концевого второго участка гибридизации второго зонда. Когда первый зонд и второй зонд гибридизуются с последовательностью, не являющейся мишенью, в неспецифическую гибридизацию будут вовлечены только их первые участки гибридизации, а каждый из их вторых участков гибридизации образует одиночную цепь, результатом чего является отсутствие лигирования первого зонда и второго зонда (Фиг.6).
Как описано выше, следует понимать, что для пары зондов, состоящей из первого зонда, имеющего структуру DSO, и второго зонда, имеющего структуру mDSO, ложноположительные результаты при детекции мишени отсутствуют полностью.
После гибридизации первый зонд и второй зонд, гибридизованные с нуклеиновокислотной последовательностью-мишенью, лигируют.
Поскольку ферментативное лигирование является предпочтительным способом ковалентного присоединения первого зонда ко второму зонду, термин "лигирование" будет использован повсеместно в данной заявке. Однако термин "лигирование" является общим термином, и следует понимать, что он включают любой способ ковалентного присоединения обоих зондов друг к другу. Одной альтернативой ферментативному лигированию является фотолигирование, которое описано в ЕР 0324616.
Лигирование по настоящему изобретению может быть осуществлено в соответствии с двумя альтернативными способами: во-первых, лигирование может быть осуществлено способом заполнения бреши (патент США №6004826), когда 3'-конец одного зонда, удлиненный под действием ДНК-полимеразы, лигируют с 5'-концом другого зонда; во-вторых, лигирование может быть осуществлено способом залечивания разрыва без реакций удлинения.
Согласно предпочтительному воплощению лигирование по настоящему изобретению проводят способом залечивания разрыва без каких-либо дополнительных реакций удлинения для присоединения 3'-конца одного зонда к 5'-концу другого зонда.
Реакции лигирования могут быть осуществлены с использованием широкого разнообразия лигирующих агентов, включая агенты ферментативного лигирования и агенты неферментативного лигирования, такие как агенты химического и фотолигирования. Агенты химического лигирования включают, без ограничения, активирующие, конденсирующие и восстанавливающие агенты, такие как карбодиимид, бромистый циан (BrCN), N-цианоимидазол, имидазол, 1-метилимидазол/карбодиимид/цистамин, дитиотреит (DTT) и ультрафиолетовый свет. Аутолигирование, то есть спонтанное лигирование в отсутствие лигирующего агента, также находится в пределах объема разъяснений, изложенных в данном описании. Подробные протоколы способов химического лигирования и описания соответствующих реакционноспособных групп можно найти в Xu et al., Nucl. Acids Res., 27: 875-81 (1999); Gryaznov and Letsinger, Nucl. Acids Res. 21: 1403-08 (1993); Gryaznov et al., Nucleic Acid Res. 22: 2366-69 (1994); Kanaya and Yanagawa, Biochemistry 25: 7423-30 (1986); Luebke and Dervan, Nucl. Acids Res. 20: 3005-09 (1992); Sievers and von Kiedrowski, Nature 369: 221-24 (1994); Liu and Taylor, Nucl. Acids Res. 26: 3300-04 (1999); Wang and Kool, Nucl. Acids Res. 22: 2326-33 (1994)).
Фотолигирование с использованием света соответствующей длины волны в качестве агента лигирования также находится в пределах объема разъяснений, изложенных в данном описании. В некоторых воплощениях фотолигирование относится к зондам, содержащим аналоги нуклеотидов, включая 4-тиотимидин (s4T), 5-винилурацил и его производные или их комбинацию, но этим не ограничиваясь. В некоторых воплощениях лигирующий агент включает: (а) свет в диапазоне UV-A (от примерно 320 нм до примерно 400 нм), диапазоне UV-B (примерно от 290 нм до примерно 320 нм) или их комбинации, (b) свет с длиной волны от примерно 300 нм до примерно 375 нм, (с) свет с длиной волны от примерно 360 нм до примерно 370 нм; (d) свет с длиной волны от примерно 364 нм до примерно 368 нм или (е) свет с длиной волны примерно 366 нм. Описание фотолигирования можно найти, среди прочего, в Fujimoto et al., Nucl. Acid Symp.Ser. 42: 39-40 (1999); Fujimoto et al., Nucl. Acid Res. Suppl. 1: 185-86 (2001); Fujimoto et al., Nucl. Acid Suppl., 2: 155-56 (2002); Liu and Taylor, Nucl. Acid Res. 26: 3300-04 (1998).
Согласно предпочтительному воплощению реакцию лигирования проводят с использованием лигазы, такой как лигаза бактериофага Т4, лигаза E. соli и термостабильная лигаза. Более предпочтительно, реакцию лигирования проводят с использованием термостабильной лигазы, включая лигазу Afu, лигазу Taq, лигазу 777, лигазу Mth, лигазу Tth, лигазу Tth HB8, лигазу вида Thermus AK16D, лигазу Аре, лигазу LigTk, лигазу Аае, лигазу Rm и лигазу Pfu (Housby et al., Nucl. Acids Res. 28: e10 (2000); Tong et al., Nucl. Acids Res. 28: 1447-54(2000); Nakatani et al., Eur., J. Biochem. 269: 650-56 (2002); Zirvi et al., Nucl. Acids Res. 27: e40 (1999); Sriskanda et al., Nucl. Acids Res. 11: 2221-28 (2000)).
Межнуклеотидная связь, образованная в результате лигирования, включает фосфодиэфирную связь и другие виды связей. Например, в результате лигирования с использованием лигаз, как правило, получаются фосфодиэфирные связи. При неферментативных способах лигирования могут образовываться другие виды межнуклеотидных связей. Другие виды межнуклеотидных связей включают, без ограничения, образование ковалентной связи между соответствующими реакционноспособными группами, такими как α-галогеноацильная группа и фосфоротиоатная группа, с образованием тиофосфорилацетиламиногруппы, такими как фосфоротиоатная и тозилатная группа или группа йодид, с образованием 5'-фосфоротиоэфирной и пирофосфатной связей.
После проведения реакции лигирования ее продукт затем подвергают денатурации для отделения от нуклеиновокислотной последовательности-мишени.
В способе по настоящему изобретению, осуществляемом на твердой фазе, первый зонд или второй зонд, используемый в качестве иммобилизованного зонда, иммобилизован на поверхности твердой подложки. Другой зонд, используемый в качестве мобильного зонда, не является иммобилизованным.
Более предпочтительно, чтобы в способе, осуществляемом на твердой фазе, первый зонд был иммобилизован через свой 5'-конец на поверхности твердой подложки, а второй зонд был неиммобилизованным. Альтернативно, в способе, осуществляемом на твердой фазе, второй зонд иммобилизован через свой 3'-конец на поверхности твердой подложки, а первый зонд не иммобилизован (Фиг.6).
Когда на твердой фазе используют молекулу с одной меткой, она предпочтительно расположена на мобильном зонде (Фиг.7).
Когда данные два зонда гибридизуются с нуклеиновокислотной последовательностью, не являющейся мишенью, они не лигируются друг с другом, и мобильный зонд отделяется от иммобилизованного зонда в процессе денатурации, так что никакого сигнала не генерируется.
По этой причине стадия денатурации является одной из контрольных точек для специфической детекции нуклеиновокислотной последовательности-мишени в настоящем изобретении.
В конце осуществляют детекцию сигнала от метки после лигирования первого зонда и второго зонда, идентифицируя наличие нуклеиновокислотной последовательности-мишени.
Согласно предпочтительному воплощению предписанный твердофазный способ дополнительно включает перед стадией (d) промывку продукта со стадии (с) для удаления мобильного зонда, не лигированного с иммобилизованным зондом.
Согласно предпочтительному воплощению способ дополнительно включает повторение стадий (а)-(с) или (a)-(d).
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень, используемая на стадии (а), представляет собой предварительно амплифицированную с применением амплифицирующего праймера нуклеиновокислотную последовательность. Предпочтительно, амплифицирующий праймер имеет структуру олигонуклеотида с двойной специфичностью гибридизации (DSO), представленную общей формулой II.
Такая предварительно амплифицированная нуклеиновокислотная последовательность-мишень может включать нуклеиновокислотную последовательность-мишень, предварительно амплифицированную в других реакционных условиях (или другом реакционном сосуде), нежели реакционные условия (или реакционный сосуд) для стадий (а)-(с). Альтернативно, предварительно амплифицированная нуклеиновокислотная последовательность-мишень может быть получена в тех же реакционных условиях (или том же реакционном сосуде), что и реакционные условия (или реакционный сосуд) для стадий (а)-(с).
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, первый зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов, и второй зонд включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) зондов.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень характеризуется вариабельностью нуклеотидов, более предпочтительно SNP (однонуклеотидным полиморфизмом).
Согласно предпочтительному воплощению вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации второго зонда.
IV. Способ детекции мишени по изменению сигнала флуоресценции в зависимости от гибридизации в жидкой фазе или на твердой фазе
В еще одном следующем аспекте данного изобретения предложен способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с TD-зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:

где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит в качестве метки флуоресцентную репортерную молекулу на 5'-концевом втором участке гибридизации; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, индуцируя изменение флуоресценции от флуоресцентной репортерной молекулы; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, не индуцируя изменения флуоресценции от флуоресцентной репортерной молекулы, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью; и
(b) детекции изменения флуоресценции, так что изменение флуоресценции указывает на наличие нуклеиновокислотной последовательности-мишени.
Поскольку в настоящем изобретении используется разный характер гибридизации 5'-концевого второго участка гибридизации TD-зонда, общее описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
Было установлено, что один и тот же меченый флуоресцентной меткой олигонуклеотид генерирует разное флуоресцентное излучение в одноцепочечном и двухцепочечном состояниях (см. патенты США №№7348141 и 7537886).
Авторы изобретения обнаружили, что, когда TD-зонд содержит в качестве метки единственную флуоресцентную репортерную молекулу на своем 5'-концевом втором участке гибридизации, то генерируется флуоресценция разной интенсивности в зависимости от гибридизации с нуклеиновокислотными последовательностями-мишенями и нуклеиновокислотными последовательностями, не являющимися мишенями.
Изменение флуоресценции от флуоресцентной репортерной молекулы, детекция которого будет осуществлена по окончании, включает ослабление флуоресценции, а также усиление флуоресценции. Параметры флуоресценции, по которым можно осуществлять детекцию такого изменения, включают интенсивность флуоресценции, поляризацию флуоресценции, время жизни флуоресценции и квантовый выход флуоресценции, но этим не ограничиваются. Наиболее предпочтительно, подлежащим детекции параметром флуоресценции является интенсивность флуоресценции от флуоресцентной репортерной молекулы.
Изменение флуоресценции от флуоресцентной репортерной молекулы в процессе гибридизации с последовательностями-мишенями зависит от нескольких факторов, таких как типы и положения меток, которые можно найти в патентах США №№7348141 и 7537886.
Флуоресцентная репортерная молекула, используемая в настоящем изобретении, может быть описана со ссылкой на указанные выше описания. Согласно предпочтительному воплощению флуоресцентная репортерная молекула представляет собой молекулу на основе флуоресцеина (например, JOE, TET или FAM), молекулу на основе родамина (например, TAMRA или ROX) или BODIPY530/550.
Флуоресцентная репортерная молекула расположена на 5'-концевом втором участке гибридизации, который обладает наибольшим потенциалом в TD-зонде в отношении дискриминации последовательностей-мишеней и последовательностей, не являющихся мишенями. Как продемонстрировано по всей данной заявке, поведение при гибридизации 5'-концевого второго участка гибридизации является самым определяющим фактором при дискриминации последовательностей-мишеней и последовательностей, не являющихся мишенями.
Флуоресцентная репортерная молекула расположена на 5'-конце, 3'-конце или внутреннем нуклеотиде 5'-концевого второго участка гибридизации. Более предпочтительно, чтобы флуоресцентная репортерная молекула была расположена на внутреннем нуклеотиде.
Согласно предпочтительному воплощению флуоресцентная репортерная молекула соединена с остатком урацила.
Согласно предпочтительному воплощению изменение флуоресценции наблюдают при заданной температуре или в заданном диапазоне температур.
Согласно предпочтительному воплощению стадию (а) выполняют с использованием TD-зонда вместе с обратным праймером и матричной полимеразой нуклеиновых кислот, так что образуется дополнительное количество нуклеиновокислотной последовательности-мишени, гибридизуемой с TD-зондом, что усиливает изменение флуоресценции, указывающее на наличие нуклеиновокислотной последовательности-мишени.
Согласно предпочтительному воплощению стадию (а) выполняют с использованием TD-зонда вместе с парой праймеров, состоящей из двух праймеров в качестве прямого праймера и обратного праймера, и матричной полимеразой нуклеиновых кислот, так что нуклеиновокислотная последовательность-мишень, гибридизуемая с TD-зондом, амплифицируется в результате ПЦР, что усиливает изменение флуоресценции, указывающее на наличие нуклеиновокислотной последовательности-мишени.
Альтернативно, TD-зонд дополнительно метят молекулой-гасителем, способной гасить флуоресценцию репортерной молекулы, и данный гаситель расположен на TD-зонде для гашения флуоресценции репортерной молекулы, когда TD-зонд или 5'-концевой второй участок гибридизации TD-зонда не вовлечен в гибридизацию с нуклеиновокислотной последовательностью-мишенью.
Согласно предпочтительному воплощению данный гаситель расположен на TD-зонде для конформационного гашения флуоресценции репортерной молекулы, когда TD-зонд или 5'-концевой второй участок гибридизации TD-зонда не вовлечен в гибридизацию с нуклеиновокислотной последовательностью-мишенью.
Согласно предпочтительному воплощению молекула-гаситель является флуоресцентной, и подлежащий детекции сигнал, указывающий на наличие нуклеиновокислотной последовательности-мишени, представляет собой сигнал от флуоресцентной молекулы-гасителя.
Когда реализацию настоящего изобретения осуществляют вместе с обратным праймером или парой праймеров, матричная полимераза нуклеиновых кислот предпочтительно представляет собой термостабильную полимеразу, не обладающую 5'→3'-экзонуклеазной активностью, включая фрагмент Штоффеля Taq-полимеразы (F С Lawyer et al„ Genome Res. 2: 275-287 (1993)) и мутантные формы ДНК-полимеразы из Thermus aquaticus, Thermus flavus или Thermus thermophilus (патент США №5885813). Их примерами являются: ДНК-полимераза (экзо-) KOD (TOYOBO), ДНК-полимераза (экзо-) Vent (NEB), ДНК-полимераза (экзо-) Deep vent (NEB), ДНК-полимераза экзо(-) Platinum™ Tfi (Invitrogen), фрагмент Штоффеля ДНК-полимеразы Amplitaq (ABI), ДНК-полимераза экзо-Pfu (Agilent).
Способ по настоящему изобретению также можно осуществлять, используя термостабильные полимеразы с 5'→3'-экзонуклеазными активностями.
Согласно предпочтительному воплощению способ по настоящему изобретению осуществляют в жидкой фазе или на твердой фазе. Когда способ по настоящему изобретению осуществляют на твердой фазе, TD-зонд иммобилизован через свой 3'-конец на поверхности твердой подложки.
V. Конструирование и приготовление зонда, позволяющего осуществлять дискриминацию последовательностей-мишеней
В другом аспекте данного изобретения предложен способ, позволяющий с помощью молекулы зонда отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью, включающий стадии:
(a) выбора нуклеиновокислотной последовательности-мишени;
(b) конструирования последовательности молекулы зонда, содержащей (1) гибридизующуюся последовательность, комплементарную нуклеиновой кислоты-мишени, и (2) разделительный участок, содержащий по меньшей мере три универсальных основания, так что разделительный участок располагается посередине гибридизующейся последовательности с образованием трех участков в молекуле зонда; и
(c) определения положения разделительного участка в молекуле зонда, позволяющего участку, расположенному в 5'-направлении от разделительного участка, иметь более низкую Тпл, чем участку, расположенному в 3'-направлении от разделительного участка, и позволяющего разделительному участку иметь наиболее низкую Тпл среди этих трех участков, тем самым обеспечивая получение молекулы зонда, имеющей три разных участка с отличающимися друг от друга величинами Тпл, среди которых (1) 5'-концевой второй участок гибридизации молекулы зонда имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени, (2) 3'-концевой первый участок гибридизации молекулы зонда имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; и (3) разделительный участок молекулы зонда, расположенный между 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, содержит по меньшей мере три универсальных основания; и Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков,
при этом, когда молекула зонда гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации второго зонда оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью; при этом, когда молекула зонда гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, на основании чего молекула зонда позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Способ по настоящему изобретению относится к предложению нового подхода для значительного усиления эффективности дискриминации зондов в отношении последовательностей-мишеней. Способ по настоящему изобретению также может быть представлен как способ улучшения дискриминирующей способности зонда в отношении последовательностей-мишеней.
Способ по настоящему изобретению осуществляют для получения TD-зонда, рассмотренного выше. Поэтому, чтобы избежать ненужного дублирования, общие для них описания не повторяются, тем не менее, в данное описание способа они включены, как если бы они были приведены повторно.
Согласно способу по настоящему изобретению предложена новая стратегия повышения дискриминирующей способности посредством придания новых характеристик олигонуклеотидным последовательностям как таковым, обеспечивающая получение новых зондов, демонстрирующих разное поведение при гибридизации с последовательностями, являющимися мишенями и не являющимися мишенями.
Критическим в способе по настоящему изобретению является конструирование последовательности молекулы зонда, имеющей (1) гибридизующуюся последовательность, комплементарную нуклеиновой кислоте-мишени, и (2) разделительный участок, содержащий по меньшей мере три универсальных основания, так что разделительный участок располагается посередине в гибридизующейся последовательности с образованием трех участков в молекуле зонда.
На этой стадии приводится структурная схема олигонуклеотида, имеющая вид: 5'-концевой участок/разделительный участок/3'-концевой участок в олигонуклеотиде. И 5'-концевой, и 3'-концевой участки оба несут гибридизующуюся последовательность, комплементарную нуклеиновой кислоте-мишени, и между ними располагается разделительный участок.
Самая критическая стадия в настоящем изобретении заключается в определении положения разделительного участка в зонде, позволяющего участку, расположенному в 5'-направлении от разделительного участка, иметь более низкую Тпл, чем участку, расположенному в 3'-направлении от разделительного участка, и позволяющего разделительному участку иметь самую низкую Тпл среди этих трех участков, тем самым обеспечивая получение олигонуклеотида, имеющего три разных участка с отличающимися друг от друга величинами Тпл.
Новыми структурными характеристиками, введенными в олигонуклеотиды согласно способу по настоящему изобретению, являются: (1) три отдельных участка (5'-концевой второй участок гибридизации, разделительный участок и 3'-концевой первый участок гибридизации) в олигонуклеотидных последовательностях; (2) отличающиеся друг от друга величины Тпл для данных трех участков; (3) разделительный участок, содержащий по меньшей мере три универсальных основания, между 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации; (4) два участка, вовлеченные в молекулярное взаимодействие с мишенями при гибридизации, которые отделены друг от друга с точки зрения событий гибридизации разделительным участком; (5) величины Тпл, убывающие в порядке от 3'-концевого первого участка гибридизации к 5'-концевому второму участку гибридизации и разделительному участку. Такие структурные особенности гибридизационных зондов обеспечивают обеспечивают протекание гибридизации зондов с последовательностями, являющимися мишенями и не являющимися мишенями, совершенно разным образом, что позволяет резко повысить специфичность гибридизации зондов с их последовательностями-мишенями.
VI. Наборы для детекции мишени
1. Наборы для детекции мишени в жидкой фазе
В еще одном другом аспекте данного изобретения предложен набор для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот, содержащий дискриминирующий мишень зонд (TD-зонд), имеющий структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную общей формулой I, описанной выше, позволяющий отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Поскольку набор по данному изобретению составлен для осуществления описанных выше способов детекции по настоящему изобретению, общее для них описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
Согласно предпочтительному воплощению TD-зонд имеет метку или систему взаимодействующих меток, содержащую некоторое количество меток для генерирования детектируемого сигнала.
Более предпочтительно, чтобы система взаимодействующих меток представляла собой пару, состоящую из репортерной молекулы и молекулы-гасителя, расположенную на TD-зонде.
Согласно предпочтительному воплощению репортерная молекула и молекула-гаситель обе расположены на 5'-концевом втором участке гибридизации, либо репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и разделительного участка.
Согласно предпочтительному воплощению TD-зонд имеет одну из молекул, выбранную из репортерной молекулы и молекулы-гасителя, на своем 5'-концевом втором участке гибридизации, а другую на своем 3'-концевом первом участке гибридизации.
Согласно предпочтительному воплощению набор дополнительно содержит фермент, обладающий 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению набор дополнительно содержит матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной активностью, и по меньшей мере один праймер, выбранный из прямого праймера, подлежащего гибридизации с сайтом, расположенным "вниз по течению" относительно сайта гибридизации TD-зонда, и обратного праймера.
Согласно предпочтительному воплощению используемая нуклеиновокислотная последовательность-мишень представляет собой предварительно амплифицированную с применением амплифицирующего праймера нуклеиновокислотную последовательность, и набор дополнительно содержит амплифицирующий праймер.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, и TD-зонд включает по меньшей мере два типа зондов.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа зондов и прямой праймер включает по меньшей мере два типа праймеров или обратный праймер включает по меньшей мере два типа праймеров.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень характеризуется вариабельностью нуклеотидов.
Согласно предпочтительному воплощению вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации TD-зонда.
Согласно предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, и блокирующий сайт расположен в 3'-концевом первом участке гибридизации TD-зонда.
2. Наборы для детекции мишени на твердой фазе
В другом аспекте данного изобретения предложен набор для детекции нуклеиновокислотной последовательности-мишени на твердой фазе из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), содержащий:
(а) TD-зонд, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд иммобилизован через свой 3'-конец на поверхности твердой подложки; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:

где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд имеет метку, генерирующую детектируемый сигнал, и эта метка расположена на 5'-концевом втором участке гибридизации TD-зонда; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, а 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью; и
(b) твердую подложку.
Поскольку набор по данному изобретению составлен для осуществления описанного выше способа детекции по настоящему изобретению, общее для них описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
Согласно предпочтительному воплощению метка представляет собой химическую метку, ферментативную метку, радиоактивную метку, флуоресцентную метку, интерактивную метку, люминесцентную метку, хемилюминесцентную метку или металлическую метку Более предпочтительно, чтобы метка представляла собой систему взаимодействующих меток, содержащую пару, состоящую из флуоресцентной репортерной молекулы и молекулы-гасителя, и TD-зонд имел одну из молекул, выбранную из репортерной молекулы и молекулы-гасителя, в сайте на 5'-концевом втором участке гибридизации, а другую в сайте, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению молекула-гаситель расположена в сайте на 5'-концевом втором участке гибридизации TD-зонда, подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью, а флуоресцентная репортерная молекула расположена в сайте, не подлежащем перевариванию ферментом, обладающим 5'→3'-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5'-концевой второй участок гибридизации переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5'-концевой второй участок гибридизации не переваривается ферментом, обладающим 5'→3'-экзонуклеазной активностью, в результате чего сигнал флуоресценции будет отсутствовать; тем самым осуществляется детекция сигнала флуоресценции на твердой подложке для определения наличия нуклеиновокислотной последовательности-мишени.
Согласно предпочтительному воплощению набор дополнительно содержит фермент, обладающий 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, и TD-зонд включает по меньшей мере два типа зондов.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень характеризуется вариабельностью нуклеотидов, и вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации TD-зонда.
Согласно предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, и блокирующий сайт расположен в 3'-концевом первом участке гибридизации TD-зонда.
3. Наборы для детекции мишени с использованием ПЦР
В еще одном другом аспекте данного изобретения предложен набор для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) и полимеразной цепной реакции (ПЦР), содержащий:
(a) TD-зонд, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; и
(b) пару праймеров, состоящую из двух праймеров в качестве прямого праймера и обратного праймера, каждый из которых имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени;
где TD-зонд гибридизуется с сайтом, расположенным между двумя этими праймерами; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:

где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит две метки - флуоресцентную репортерную молекулу и молекулу-гаситель, способную гасить флуоресценцию репортерной молекулы; флуоресцентная репортерная молекула и молекула-гаситель обе расположены на 5'-концевом втором участке гибридизации, или репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5'-концевого второго участка гибридизации и разделительного участка; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков X'p, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, а 5'-концевой второй участок гибридизации переваривается в результате проявления 5'→3'-экзонуклеазной активности матричной полимеразы нуклеиновых кислот; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5'-концевой второй участок гибридизации не переваривается в результате проявления 5'→3'-экзонуклеазной активности матричной полимеразы нуклеиновых кислот, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Согласно предпочтительному воплощению набор дополнительно содержит матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной активностью.
Согласно предпочтительному воплощению используемая нуклеиновокислотная последовательность-мишень представляет собой предварительно амплифицированную с применением амплифицирующего праймера нуклеиновокислотную последовательность, и набор дополнительно содержит амплифицирующий праймер.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа зондов, прямой праймер включает по меньшей мере два типа праймеров и обратный праймер включает по меньшей мере два типа праймеров.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень характеризуется вариабельностью нуклеотидов.
Согласно предпочтительному воплощению прямой праймер, обратный праймер или амплифицирующий праймер имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную общей формулой II.
Согласно предпочтительному воплощению в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5'→3'-экзонуклеазной активностью, и блокирующий сайт расположен в 3'-концевом первом участке гибридизации.
Согласно предпочтительному воплощению блокирующий сайт TD-зонда расположен на 3'-концевом первом участке гибридизации TD-зонда. Более предпочтительно, чтобы блокирующий сайт TD-зонда был расположен вблизи 3'-конца разделительного участка.
Согласно предпочтительному воплощению блокирующий сайт содержит 1-10 молекул-блокаторов.
4. Наборы для детекции мишени с использованием реакции лигирования
В следущем аспекте данного изобретения предложен набор для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), с применением реакции лигирования, содержащий:
(a) первый зонд, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную первому сайту нуклеиновокислотной последовательности-мишени; и
(b) второй зонд, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную второму сайту нуклеиновокислотной последовательности-мишени, который расположен "вверх по течению" по отношению к первому сайту;
где по меньшей мере один зонд, выбранный из первого зонда или второго зонда, содержит метку для генерации детектируемого сигнала; где второй зонд представляет собой TD-зонд; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:

где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда второй зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации второго зонда оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, что позволяет осуществлять лигирование первого зонда и второго зонда; при этом, когда второй зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок второго зонда оба образуют одиночную цепь, так что первый зонд и второй зонд не подвергаются лигированию, на основании чего второй зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Согласно предпочтительному воплощению набор дополнительно содержит лигазу, предназначенную для лигирования первого зонда и второго зонда, гибридизованных с нуклеиновокислотной последовательностью-мишенью.
Согласно предпочтительному воплощению данная метка представляет собой химическую метку, ферментативную метку, радиоактивную метку, флуоресцентную метку, интерактивную метку, люминесцентную метку, хемилюминесцентную метку или металлическую метку.
Согласно предпочтительному воплощению метка представляет собой систему взаимодействующих меток, содержащую пару, состоящую из репортерной молекулы и молекулы-гасителя.
Согласно предпочтительному воплощению первый зонд имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную общей формулой II.
Согласно предпочтительному воплощению используемая нуклеиновокислотная последовательность-мишень представляет собой предварительно амплифицированную с применением амплифицирующего праймера нуклеиновокислотную последовательность, и набор дополнительно содержит амплифицирующий праймер.
Согласно предпочтительному воплощению набор применяют для твердой фазы; где первый зонд иммобилизован через свой 5'-конец на поверхности твердой подложки, а второй зонд не иммобилизован.
Согласно предпочтительному воплощению набор применяют для твердой фазы; где второй зонд иммобилизован через свой 3'-конец на поверхности твердой подложки, а первый зонд не иммобилизован.
Согласно предпочтительному воплощению первый зонд и второй зонд расположены в непосредственной близости по отношению друг к другу, когда они гибридизуются с нуклеиновокислотной последовательностью-мишенью.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, и первый зонд и второй зонд каждый включает по меньшей мере два типа зондов.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень характеризуется вариабельностью нуклеотидов.
Согласно предпочтительному воплощению вариабельность нуклеотидов в нуклеиновокислотной последовательности-мишени присутствует в сайте, расположенном напротив 5'-концевого второго участка гибридизации второго зонда.
5. Наборы для детекции мишеней, основанной на гибридизации
В еще одном другом аспекте данного изобретения предложен набор для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), содержащий:
дискриминирующий мишень зонд (TD-зонд), имеющий структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I, позволяющий отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью:

где Х'р представляет собой 5'-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y'q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z'r представляет собой 3'-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит в качестве метки флуоресцентную репортерную молекулу, расположенную на 5'-концевом втором участке гибридизации; р, q и r представляют количество нуклеотидов; и X', Y' и Z' представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого второго участка гибридизации ниже, чем у 3'-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х'р, Y'q и Z'r; разделительный участок отделяет 5'-концевой второй участок гибридизации от 3'-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5'-концевым вторым участком гибридизации и 3'-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5'-концевой второй участок гибридизации, и 3'-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, индуцируя изменение флуоресценции от флуоресцентной репортерной молекулы; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, не индуцируя никакого изменения флуоресценции от флуоресцентной репортерной молекулы, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
Согласно предпочтительному воплощению набор дополнительно содержит обратный праймер и матричную полимеразу нуклеиновых кислот для образования дополнительного количества нуклеиновокислотной последовательности-мишени, гибридизуемой с TD-зондом, что усиливает изменение флуоресценции, указывающее на наличие нуклеиновокислотной последовательности-мишени.
Предпочтительно, набор дополнительно содержит пару праймеров, состоящую из двух праймеров в качестве прямого праймера и обратного праймера, и матричную полимеразу нуклеиновых кислот для амплификации нуклеиновокислотной последовательности-мишени, гибридизуемой с TD-зондом в ПЦР, что усиливает изменение флуоресценции, указывающее на наличие нуклеиновокислотной последовательности-мишени.
Альтернативно, TD-зонд дополнительно содержит в качестве метки молекулу-гаситель, способную гасить флуоресценцию репортерной молекулы, и этот гаситель располагается на TD-зонде, чтобы индуцировать самогашение, когда TD-зонд не вовлечен в гибридизацию с нуклеиновокислотной последовательностью-мишенью.
В тех случаях, когда набор по настоящему изобретению содержит обратный праймер или пару праймеров, матричной полимеразой нуклеиновых кислот предпочтительно является термостабильная полимераза, не обладающая 5'→3'-экзонуклеазной активностью, включая фрагмент Штоффеля Taq-полимеразы и мутантные формы ДНК-полимеразы из Thermus aquaticus, Thermus flaws или Thermus thermophilus (патент США №5885813).
Набор по настоящему изобретению применяют в жидкой фазе или на твердой фазе.
Все наборы по настоящему изобретению, описанные выше, возможно могут содержать реагенты, необходимые для проведения амплификации мишени с использованием реакций ПЦР (например, ПЦР-реакций), такие как буферы, кофакторы ДНК-полимеразы и дезоксирибонуклеотид-5-трифосфаты. Возможно, что наборы также могут содержать молекулы различных полинуклеотидов, обратную транскриптазу, различные буферы и реагенты, и антитела, ингибирующие ДНК-полимеразную активность. Наборы также могут содержать реагенты, необходимые для проведения реакций положительного и отрицательного контроля. Оптимальные количества реагентов, которые будут использоваться в заданной реакции, могут быть легко определены специалистом на основе информации, содержащейся в данном описании. Обычно наборы предназначены для того, чтобы вмещать описанные выше составляющие в отдельных упаковках или отсеках.
Признаки и преимущества данного изобретения будут обобщены ниже.
(a) TD-зонд, имеющий структуру mDSO, гибридизуется с нуклеиновокислотными последовательностями-мишенями на всем протяжении своей общей последовательности, включая 5'-концевой второй участок гибридизации и 3'-концевой первый участок гибридизации. В условиях специфической гибридизации TD-зонда с мишенью, когда TD-зонд гибридизуется с нуклеиновокислотными последовательностями, не являющимися мишенями, его 3'-концевой первый участок гибридизации неспецифически связывается с нуклеиновокислотными последовательностями, не являющимися мишенями, но тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба не гибридизуются с нуклеиновокислотной последовательностью, не являющейся мишенью, образуя одиночную цепь ввиду низких значений их Тпл.
По этой причине 5'-концевой второй участок гибридизации TD-зонда демонстрирует в каждом случае отчетливо разные картины гибридизации для нуклеиновокислотных последовательностей, являющихся мишенями и не являющихся мишенями, с гораздо более высокой специфичностью отличая нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
(b) Приложения, относящиеся к детекции мишени с использованием TD-зонда, демонстрируют очень высокую специфичность к мишени, обусловленную следующими моментами наблюдения за мишенью. Во-первых, TD-зонд, имеющий в каждом случае разные картины гибридизации для нуклеиновокислотных последовательностей, являющихся мишенями и не являющихся мишенями, как описано выше, способен с гораздо более высокой специфичностью отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью. Во-вторых, наличие последующих ферментативных реакций (экзонуклеолитической реакции или реакции лигирования) определяется в зависимости от картин гибридизации TD-зонда, повышая специфичность к мишени в процедурах детекции мишени.
(c) Дискриминирующая способность TD-зонда в отношении мишени, обусловленная разными картинами гибридизации, с успехом применена в способах детекции мишени с использованием 5'→3'-экзонуклеазной активности, что полностью предотвращает генерацию ложноположительных сигналов (результатов). В качестве иллюстрации, когда традиционный зонд, содержащий метку на своем 5'-концевом участке, гибридизуется своим 5'-участком с последовательностями, не являющимися мишенями, то этот 5'-концевой участок переваривается в результате проявления 5'→3'-экзонуклеазной активности, генерируя ложноположительные сигналы. Даже когда TD-зонд гибридизуется с последовательностями, не являющимися мишенями, маловероятно, что его 5'-концевой второй участок гибридизации будет гибридизоваться с последовательностями, не являющимися мишенями, генерируя какие-либо ложноположительные сигналы.
(d) В способе с применением ПЦР в режиме реального времени для детекции нуклеиновокислотных последовательностей-мишеней показано, что TD-зонд с обеими молекулами, репортерной молекулой и молекулой-гасителем, расположенными на 5'-концевом втором участке гибридизации, или с каждой молекулой, расположенной на 5'-концевом втором участке гибридизации и разделительном участке, превосходно предотвращает получение ложных сигналов. Традиционные технологии, такие как способ с использованием зонда TaqMan™, допускают получение ложноположительных сигналов, обусловленных неспецифическим связыванием меченых зондов, в частности, при многократной детекции мишени. Однако настоящее изобретение успешно преодолевает такие проблемы путем использования TD-зонда, в котором обе молекулы, репортерная молекула и молекула-гаситель, расположены на 5'-концевом втором участке гибридизации. Кроме того, TD-зонд, в типичном случае содержащий две метки, как описано выше, позволяет преодолеть недостатки, связанные с 5'→3'-эндонуклеазной активностью полимераз, вызывающие проблемы в зависимости от типов полимераз и реакционных условий.
(e) Уникальная картина гибридизации TD-зонда также позволяет отлично предотвращать получение ложноположительных сигналов в способах детекции мишени с использованием лигирующей активности. Как правило, в способах детекции мишени с использованием лигирования двух зондов (прямого первого зонда и обратного второго зонда) для осуществления лигирования необходимо наличие двухцепочечных форм (дуплекса) близкорасположенных концевых участков этих двух зондов. В анализе с применением лигирования по настоящему изобретению с использованием TD-зонда в качестве второго зонда 5'-концевой второй участок гибридизации TD-зонда образует одиночную цепь, когда TD-зонд гибридизуется с последовательностями, не являющимися мишенями, и ввиду этого предотвращает реакции лигирования, не генерируя никаких ложноположительных сигналов.
(f) TD-зонд демонстрирует отличную специфичность в отношении дискриминации однонуклеотидной вариабельности благодаря использованию разных картин гибридизации 5'-концевого второго участка гибридизации. Даже если между 5'-концевым вторым участком гибридизации TD-зонда и нуклеиновокислотной последовательностью имеется единственный ошибочно спаренный нуклеотид, TD-зонд способен распознавать данную последовательность как последовательность, не являющуюся мишенью, и его 5'-концевой второй участок гибридизации будет образовывать одиночную цепь, не давая в результате никаких ложноположительных сигналов. В частности, TD-зонд обладает приемлемой разрешающей способностью при детекции SNP.
(д) TD-зонд позволяет реализовывать системы микрочипов для точного и высокопроизводительного твердофазного анализа. Традиционные системы микрочипов с использованием традиционных зондов допускают получение ложноположительных сигналов, обусловленных неспецифической гибридизацией традиционных зондов. В противоположность этому, твердофазный анализ по настоящему изобретению с использованием TD-зонда вместе с 5'→3'-экзонуклеазной активностью (или лигирующей активностью) позволяет осуществлять детекцию нуклеиновокислотных последовательностей-мишеней в режиме реального времени, а также детекцию нуклеиновокислотных последовательностей-мишеней более точным и быстрым способом.
Теперь настоящее изобретение будет описано далее подробно посредством примеров. Специалистам в данной области будет очевидно, что эти примеры предназначены для более конкретной иллюстрации изобретения и объем настоящего изобретения, который изложен в прилагаемой формуле изобретения, не ограничивается данными примерами.
ПРИМЕРЫ
ПРИМЕР 1. Оценка расщепляющей активности фермента, обладающего 5'→3'-экзонуклеазной активностью, в отношении зонда с 5'-концевым ошибочным спариванием
Авторы изобретения проверяли, может ли фермент, обладающий 5'→3'-экзонуклеазной активностью, расщеплять зонд, содержащий ошибочно спаренные нуклеотиды на своем 5'-концевом участке.
Для проведения этой оценки в качестве матриц использовали синтетический олигонуклеотид для гена Staphylococcus aureus (SA). Использовали пять разных типов зондов с двумя метками, и они содержат: спаренную последовательность, единственный ошибочно спаренный нуклеотид, три ошибочно спаренных нуклеотида, шесть ошибочно спаренных нуклеотидов и девять ошибочно спаренных нуклеотидов в своих 5'-концевых участках, соответственно. Зонд с двумя метками содержит 6-FAM (6-карбоксифлуоресцеин) в качестве флуоресцентной репортерной молекулы на своем 5'-конце и гаситель Black Hole 1 (BHQ-1) в качестве молекулы-гасителя на своем 3'-концевом участке. Зонд с двумя метками модифицирован С3-спейсером по своему 3'-концу, так что удлинение такого зонда, содержащего две метки, не происходит.
ДНК-полимеразу, обладающую 5'→3'-экзонуклеазной активностью, использовали для 5'→3'-экзонуклеолитических реакций с данными зондами, содержащими две метки, и данной матрицей (ген S. aureus). Сигналы измеряли на стадии гибридизации каждого цикла.
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и зондов с двумя метками для гена S. aureus:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды.)
Экзонуклеолитическую реакцию проводили в конечном объеме 20 мкл, содержащем 0,2 пмоль синтетического олигонуклеотида для S. aureus (SEQ ID NO:1), 5 пмоль зонда с двумя метками (SEQ ID NO:2, 3, 4, 5 или 6) и 10 мкл смеси 2× Master Mix, содержащей 6 мМ MgCl2, 200 мкМ dNTP и 2 единицы ДНК-полимеразы Taq DiaStar™ (Solgent, Korea); пробирку, содержащую реакционную смесь, помещали в термоциклер для проведения ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 20 с при 95°С и 60 с при 60°С. Детекцию генерированного сигнала осуществляли на стадии гибридизации (60°С) каждого цикла.
Как показано на Фиг.8, когда использовали зонды с двумя метками, содержащие спаренную последовательность и имеющие единственный ошибочно спаренный нуклеотид на своих 5'-концевых участках (NO:1 и 3), происходила генерациия флуоресцентных сигналов для SA. С другой стороны, никаких флуоресцентных сигналов для SA не наблюдали в случае использования зондов с двумя метками, имеющих по меньшей мере три ошибочно спаренных нуклеотида в своих 5'-концевых участках (NO:5, 7 и 9). Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2, 4, 6, 8 и 10).
Эти результаты указывают на то, что фермент, обладающий 5'→3'-экзонуклеазной активностью, не расщепляет зонд, имеющий по меньшей мере три ошибочно спаренных нуклеотида на своем 5'-концевом участке.
ПРИМЕР 2. Оценка TD-зонда с двумя метками на предмет дискриминации нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью, с использованием фермента, обладающего 5'→3'-экзонуклеазной активностью
Проводили оценку TD-зонда по данному изобретению на предмет его способности отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью, с использованием фермента, обладающего 5'→3'-экзонуклеазной активностью.
Сначала авторы изобретения продемонстрировали, может ли специфичный к мишени сигнал генерироваться TD-зондом имеющим разные картины гибридизации на своем 5'-концевом втором участке гибридизации.
Для проведения этой оценки в качестве матриц использовали синтетические олигонуклеотиды для генов Staphylococcus aureus (SA) и Neisseria gonorrhoeae (NG). Два разных типа TD-зондов для каждого гена имеют спаренную последовательность и ошибочно спаренную последовательность на своих 5'-концевых участках гибридизации, соответственно. TD-зонд с двумя метками содержит 6-FAM (6-карбоксифлуоресцеин) в качестве флуоресцентной репортерной молекулы на своем 5'-конце и гаситель Black Hole 1 (BHQ-1) в качестве молекулы-гасителя в своем 3'-концевом участке. TD-зонд с двумя метками модифицирован С3-спейсером по своему 3'-концу, так что удлинение такого TD-зонда, содержащего две метки, не происходит.
ДНК-полимеразу, обладающую 5'→3'-экзонуклеазной активностью, использовали для 5'→3'-экзонуклеолитических реакций с TD-зондом, имеющим две метки, и матрицей-мишенью (ген S. aureus или N. gonorrhoeae). Сигналы измеряли на стадии гибридизации каждого цикла.
А. Генерирование специфичного к мишени сигнала для гена S. aureus с использованием TD-зонда
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и TD-зонда с двумя метками для гена S. aureus:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды.)
Экзонуклеолитическую реакцию проводили в конечном объеме 20 мкл, содержащем 0,2 пмоль синтетического олигонуклеотида для S. aureus (SEQ ID NO:1), 5 пмоль TD-зонда с двумя метками (SEQ ID NO:7 или 8) и 10 мкл смеси 2× Master Mix, содержащей 6 мМ MgCl2, 200 мкМ dNTP и 2 единицы ДНК-полимеразы Taq DiaStar™ (Solgent, Korea); пробирку, содержащую реакционную смесь, помещали в термоциклер для проведения ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 20 с при 95°С и 60 с при 60°С. Детекцию генерированного сигнала осуществляли на стадии гибридизации (60°С) каждого цикла.
Как показано на Фиг.9А, флуоресцентный сигнал для нуклеиновокислотной последовательности-мишени для SA генерировался, когда использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:1). С другой стороны, не наблюдали никакого флуоресцентного сигнала для нуклеиновокислотной последовательности-мишени для SA в случае использования TD-зонда с двумя метками, содержащего ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:3). Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2 и 4).
В. Генерирование специфичного к мишени сигнала для гена N. Qpnorrhoeae с использованием TD-зонда
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и TD-зондов с двумя метками для гена N. gonorrhoeae:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды.)
Экзонуклеолитическую реакцию проводили, следуя тому же протоколу, который использовали для S. aureus, за исключением матрицы (2 пмоль N. gonorrhoeae) и TD-зондов с двумя метками (SEQ ID NO:10 или 11).
Как показано на Фиг.9В, флуоресцентный сигнал для нуклеиновокислотной последовательности-мишени для NG генерировался, когда использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:1). С другой стороны, не наблюдали никакого флуоресцентного сигнала для нуклеиновокислотной последовательности-мишени для NG в случае использования TD-зонда с двумя метками, содержащего ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:3). Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2 и 4).
Эти результаты показали, что генерирование специфичного к мишени сигнала TD-зондом, зависящее от гибридизации на его 5'-концевом втором участке гибридизации, указывает на то, что TD-зонд может отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
ПРИМЕР 3. Влияние 5'-концевых участков при сравнении TD-зонда и традиционного зонда
Авторы изобретения проверяли, оказывает ли 5'-концевой участок традиционного зонда такое же влияние, как и 5'-концевой второй участок гибридизации TD-зонда.
Для осуществления этой проверки авторы изобретения использовали TD-зонды двух разных типов: один TD-зонд имеет спаренную последовательность на своем 5'-концевом втором участке гибридизации, а другой имеет три ошибочно спаренных нуклеотида на своем 5'-концевом втором участке гибридизации. Традиционные зонды имеют те же последовательности, что и TD-зонды, за исключением дезоксиинозина.
В качестве матрицы использовали синтетический олигонуклеотид для Staphylococcus aureus (SA). Традиционный зонд и TD-зонд каждый содержит 6-FAM (6-карбоксифлуоресцеин) в качестве флуоресцентной репортерной молекулы на своем 5'-конце и гаситель Black Hole 1 (BHQ-1) в качестве молекулы-гасителя на своем 3'-концевом участке. Все зонды модифицированы С3-спейсером по своим 3'-концам.
ДНК-полимеразу, обладающую 5'→3'-экзонуклеазной активностью, использовали для 5'→3'-экзонуклеолитических реакций с TD-зондом, содержащим две метки, и матрицей-мишенью (S. aureus).
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и традиционных и TD-зондов с двумя метками для гена S. aureus:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды.)
Экзонуклеолитическую реакцию проводили в конечном объеме 20 мкл, содержащем 0,2 пмоль синтетического олигонуклеотида для S. aureus (SEQ ID NO:1), 5 пмоль зонда с двумя метками (SEQ ID NO:7, 12, 13 или 14) и 10 мкл смеси 2× Master Mix, содержащей 6 мМ MgCl2, 200 мкМ dNTP и 2 единицы ДНК-полимеразы Taq DiaStar™ (Solgent, Korea); пробирку, содержащую реакционную смесь, помещали в термоциклер для проведения ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 20 с при 95°С и 60 с при 60°С.Детекцию генерированного сигнала осуществляли на стадии гибридизации (60°С) каждого цикла.
Как показано на Фиг.10, флуоресцентный сигнал для нуклеиновокислотной последовательности-мишени генерировался, когда использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации, (NO:1). Интересно, что в случае TD-зонда, содержащего три ошибочно спаренных нуклеотида на своем 5'-концевом втором участке гибридизации, (NO:3), никакого сигнала не наблюдали. В отличие от этого, когда использовали традиционные зонды, содержащие три ошибочно спаренных нуклеотида, а также содержащие спаренную последовательность в своих 5'-концевых участках, (NO:5 и 7), наблюдали генерирование сигналов. Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2, 4, 6 и 8).
Эти результаты показали, что в противоположность TD-зонду, традиционный зонд генерировал ложноположительный сигнал при неспецифической гибридизации. Таким образом, стало ясно, что с помощью TD-зонда можно осуществлять детекцию нуклеиновокислотной последовательности-мишени без получения ложноположительных сигналов.
ПРИМЕР 4. ПЦР в режиме реального времени с использованием TD-зонда для детекции нуклеиновокислотной последовательности-мишени
Авторы изобретения применили TD-зонд в ПЦР в режиме реального времени для детекции нуклеиновокислотной последовательности-мишени с использованием ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью.
Для этого из каждой клеточной линии экстрагировали и использовали геномные ДНК Staphylococcus aureus и Neisseria gonorrhoeae. TD-зонд содержит либо спаренную, либо ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации. На 5'-концевом втором участке гибридизации TD-зонда с двумя метками располагались и репортерная молекула, и молекула-гаситель.
А. ПЦР в режиме реального времени для детекции гена S. aureus
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень гена S. aureus, в этом Примере применяют приведенные ниже последовательности праймеров и содержащих две метки TD-зондов:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды.)
ПЦР в режиме реального времени проводили в конечном объеме 20 мкл, содержащем 1 нг геномной ДНК S. aureus, 5 пмоль зонда с двумя метками (SEQ ID NO:17 или 18), 10 пмоль прямого праймера (SEQ ID NO:15), 10 пмоль обратного праймера (SEQ ID NO:16) и 10 мкл смеси 2× Master Mix, содержащей 6 мМ MgCl2, 200 мкМ dNTP и 2 единицы ДНК-полимеразы Taq DiaStar™ (Solgent, Korea); пробирку, содержащую реакционную смесь, помещали в термоциклер для проведения ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 20 с при 95°С и 60 с при 60°С. Детекцию генерированного сигнала осуществляли на стадии гибридизации (60°С) каждого цикла.
Как показано на Фиг.11А, флуоресцентный сигнал от мишени для гена S. aureus генерировался, когда использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:1). С другой стороны, не наблюдали никакого флуоресцентного сигнала для нуклеиновокислотной последовательности-мишени для гена S. aureus в случае использования TD-зонда с двумя метками, содержащего ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:3), что указывает на то, что репортерная молекула и молекула-гаситель, расположенные на 5'-концевом втором участке гибридизации, не разделялись. Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2 и 4).
В. ПЦР в режиме реального времени для детекции гена N. gonorrhoeae
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень гена N. gonorrhoeae, в этом Примере применяют приведенные ниже последовательности праймеров и содержащих две метки TD-зондов:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды.)
ПЦР в режиме реального времени проводили, следуя тому же протоколу который использовали для S. aureus, за исключением матрицы (1 нг N. gonorrhoeae), TD-зондов с двумя метками (SEQ ID NO:21 и 22) и праймеров (SEQ ID NO:19 и 20).
Как показано на Фиг.11 В, флуоресцентный сигнал от мишени для гена N. gonorrhoeae генерировался, когда использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:1). С другой стороны, не наблюдали никакого флуоресцентного сигнала для нуклеиновокислотной последовательности-мишени для гена N. gonorrhoeae в случае использования TD-зонда с двумя метками, содержащего ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:3), что указывает на то, что репортерная молекула и молекула-гаситель, расположенные на 5'-концевом втором участке гибридизации, не разделялись. Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2 и 4).
Эти результаты показали, что TD-зонд, содержащий систему взаимодействующих меток на своем 5'-концевом втором участке гибридизации, может быть использован при проведении ПЦР в режиме реального времени для дискриминации нуклеиновокислотной последовательности-мишени и нуклеиновокислотной последовательности, не являющейся мишенью.
ПРИМЕР 5. Дискриминация однонуклеотидной вариабельности с использованием TD-зонда с двумя метками в ПЦР в режиме реального времени
Авторы изобретения проверяли, можно ли с помощью TD-зонда различить однонуклеотидную вариабельность в нуклеиновокислотнй последовательности.
Для проведения этой проверки TD-зонд содержит или спаренную последовательность, или единичный ошибочно спаренный нуклеотид на своем 5'-концевом втором участке гибридизации. Как репортерная молекула, так и молекула-гаситель располагались на 5'-концевом втором участке гибридизации TD-зонда с двумя метками.
В качестве матрицы используют геномную ДНК S. aureus. Ниже приведены использованные в этом Примере последовательности праймеров и TD-зонда с двумя метками:

ПЦР в режиме реального времени проводили в конечном объеме 20 мкл, содержащем 500 пг геномной ДНК S. aureus, 5 пмоль TD-зонда с двумя метками (SEQ ID NO:23 или 24), 10 пмоль прямого праймера (SEQ ID NO:15), 10 пмоль обратного праймера (SEQ ID NO:16) и 10 мкл смеси 2× Master Mix, содержащей 6 мМ MgCl2, 200 мкМ dNTP и 2 единицы ДНК-полимеразы Taq DiaStar™ (Solgent, Корея); пробирку, содержащую реакционную смесь, помещали в термоциклер для проведения ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 20 с при 95°С и 60 с при 63°С. Детекцию генерированного сигнала осуществляли на стадии гибридизации (63°С) каждого цикла.
Как показано на Фиг.12, флуоресцентный сигнал для S. aureus генерировался, когда использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации (NO:1). С другой стороны, не наблюдали никакого флуоресцентного сигнала в случае использования TD-зонда с двумя метками, содержащего единичный ошибочно спаренный нуклеотид на своем 5'-концевом втором участке гибридизации (NO:3). Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (NO:2 и 4).
Эти результаты продемонстрировали разную картину гибридизации TD-зонда, зависящую даже от ошибочного спаривания единичного нуклеотида на его 5'-концевом втором участке гибридизации. Таким образом, стало ясно, что TD-зонд обладает высокой специфичностью в отношении дискриминации однонуклеотидной вариабельности, в том числе SNP, без ложноположительных сигналов в ПЦР в режиме реального времени.
ПРИМЕР 6. Оценка TD-зонда, иммобилизованного на твердой фазе, с использованием фермента, обладающего 5'→3'-экзонуклеазной активностью
Далее авторы изобретения провели оценку того, можно ли с помощью TD-зонда отличить нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью, с использованием фермента, обладающего 5'→3'-экзонуклеазной активностью.
Для проведения этой оценки в качестве матрицы использовали синтетический олигонуклеотид для гена Staphylococcus aureus (SA). TD-зонд содержит или спаренную или ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации. TD-зонд содержит Quasar570 в качестве флуоресцентной репортерной молекулы на своем 3'-концевом первом участке гибридизации, гаситель Black Hole 2 (BHQ-2) в качестве молекулы-гасителя на своем 5'-конце и поли(Т)7 в качестве линкерной ножки. Содержащий две метки TD-зонд иммобилизовали на поверхности твердой подложки, используя аминогруппу (аминоС7) на его 3'-конце. Для проведения 5'→3'-экзонуклеолитических реакций использовали ДНК-полимеразу Bst, обладающую 5'→3'-экзонуклеазной активностью.
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и содержащих две метки TD-зондов:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды).
Для изготовления TD-зондов двух разных типов (SEQ ID NO:25 и 26) использовали NHS-пластинки (N-гидроксисукцинимидил) NSB9 (NSBPOSTECH (NanoSurface Biosciences at Pohang University of Science and Technology), Корея. TD-зонды, растворенные в NSB-буфере для точечного нанесения в конечной концентрации 20 мкм, печатали на NSB9 NHS-пластинках с использованием OmniGrid Accent Microarrayer (DIGILAB, США). Каждый TD-зонд наносили в виде пятен по одной линии в формате 2×1 (пятна в двух повторах), и полученный микрочип инкубировали в камере с поддержанием ~85%-ной влажности в течение ночи. Затем пластинки промывали в буферном растворе, содержащем 2xSSPE (0,3 М хлорид натрия; 0,02 М гидрофосфат натрия и 2,0 мМ EDTA (этилендиаминтетрауксусная кислота)), рН 7,4 и 7,0 мМ SDS (додецилсульфат натрия), при 37°С в течение 10 мин для удаления неспецифически связавшегося TD-зонда и промывали дистиллированной водой. Затем ДНК-функционализированные пластинки сушили с применением центрифуги для пластинок и хранили в темноте при 4°С до применения.
Экзонуклеолитическую реакцию проводили на поверхности ДНК-функционализированной пластинки в конечном объеме 30 мкл, содержащем 10 пмоль синтетического олигонуклеотида для SA (SEQ ID NO:1) и 3 мкл 10× реакционного буфера, по 0,6 мкл каждого из dNTP (10 мМ), 2 единицы ДНК-полимеразы Bst (NEB, США). Всю эту смесь помещали в камеру, собранную на поверхности стеклянной пластинки NSB, к которой были пришиты TD-зонды. Пластинку располагали на блоке в термоциклере (Genepro B4I, Китай). Экзонуклеолитическую реакцию проводили в течение 30 мин при 50°С и останавливали, промывая дистиллированной водой при 95°С в течение 1 мин. Получение изображений осуществляли с применением конфокального лазерного сканирующего устройства Axon GenePix4100A (Molecular Device, США) со сканированием при разрешении с размером пикселя 5 мкм. Интенсивность флуоресценции анализировали с применением программного обеспечения для количественного анализа микрочипов GenePix pro5.1 (Molecular Device, США). Интенсивность флуоресценции выражали в виде средних значений по пятну после вычитания локального фона. Для проверки воспроизводимости каждое пятно анализировали в двух повторах. Интенсивность флуоресценции указывает среднюю величину для дублированных пятен.
Как показано на Фиг.13, флуоресцентный сигнал для S. aureus генерировался, когда вместе с матрицей (SA_TD1_Chip_M, RFU 65472,0±4,2) использовали TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации. С другой стороны, никакого флуоресцентного сигнала для S. aureus не наблюдали в случае с использованием TD-зонда с двумя метками, содержащего ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации (SA_TD1_Chip_m, RFU 3227,0±17,0). He отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (SA_TD1_Chip_M, RFU 2798,0±4,2 или SA_TD1_Chip_m, RFU 3077,0±9,9).
Эти результаты показали, что иммобилизованный TD-зонд может быть применен в анализах с использованием микрочипов, чтобы отличить нуклеиновокислотные последовательности-мишени от нуклеиновокислотных последовательностей, не являющихся мишенями.
ПРИМЕР 7. Влияние 5'-концевого второго участка гибридизации иммобилизованных TD-зондов
Далее авторы изобретения проверяли, можно ли с помощью иммобилизованных TD-зондов устранить получение ложноположительных сигналов на твердой фазе, используя влияние 5'-концевого второго участка гибридизации.
Для проведения этой оценки в качестве матрицы использовали синтетический олигонуклеотид для гена Staphylococcus aureus (SA). TD-зонд содержит или спаренную последовательность, или три ошибочно спаренных нуклеотида на своем 5'-концевом втором участке гибридизации. Традиционные зонды имеют те же самые последовательности, что и TD-зонды, за исключением дезоксиинозина. TD-зонды и традиционные зонды содержат Quasar570 в качестве флуоресцентной репортерной молекулы на своем 3'-концевом первом участке гибридизации, гаситель Black Hole 2 (BHQ-2) в качестве молекулы-гасителя на своем 5'-конце и поли(Т)7 в качестве линкерной ножки. Содержащие две метки TD-зонды иммобилизовали на поверхности твердой подложки, используя аминогруппу (аминоС7) на их 3'-концах. Для проведения 5'→3'-экзонуклеолитических реакций использовали ДНК-полимеразу Bst, обладающую 5'→3'-экзонуклеазной активностью.
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и содержащих две метки TD-зондов и традиционных зондов:


(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды).
Для изготовления зондов (SEQ ID NO:25, 27, 28 и 29) использовали NSB9 NHS-пластинки (NSBPOSTECH, Корея). Каждый зонд, растворенный в NSB-буфере для точечного нанесения в конечной концентрации 20 мкм, печатали на NSB9 NHS-пластинке с использованием OmniGrid Accent Microarrayer (DIGILAB, США). Каждый зонд наносили в виде пятен по одной линии в формате 2×1 (пятна в двух повторах), и полученный микрочип инкубировали в камере с поддержанием ~85%-ной влажности в течение ночи. Затем пластинки промывали в буферном растворе, содержащем 2xSSPE (0,3 М хлорид натрия, 0,02 М гидрофосфат натрия и 2,0 мМ EDTA), рН 7,4 и 7,0 мМ SDS, при 37°С в течение 10 мин для удаления неспецифически связавшихся зондов и промывали дистиллированной водой. Затем ДНК-функционализированные пластинки сушили с применением центрифуги для пластинок и хранили в темноте при 4°С до применения.
Экзонуклеолитическую реакцию проводили на поверхности ДНК-функционализированной пластинки в конечном объеме 30 мкл, содержащем 10 пмоль синтетического олигонуклеотида для SA (SEQ ID NO:1) и 3 мкл 10х реакционного буфера, по 0,6 мкл каждого из dNTP (10 мМ), 2 единицы ДНК-полимеразы Bst (NEB, США). Всю эту смесь помещали в камеру, собранную на поверхности стеклянной пластинки NSB, к которой были пришиты зонды. Пластинку располагали на блоке в термоциклере (Genepro B4I, Китай). Экзонуклеолитическую реакцию проводили в течение 30 мин при 50°С и останавливали, промывая дистиллированной водой при 95°С в течение 1 мин. Получение изображений осуществляли с применением конфокального лазерного сканирующего устройства Axon GenePix4100A (Molecular Device, США) со сканированием при разрешении с размером пикселя 5 мкм. Интенсивность флуоресценции анализировали с применением программного обеспечения для количественного анализа микрочипов GenePix pro5.1 (Molecular Device, США). Интенсивность флуоресценции выражали в виде средних значений по пятнам после вычитания локального фона. Для проверки воспроизводимости каждое пятно анализировали в двух повторах. Интенсивность флуоресценции указывает среднюю величину для дублированных пятен.
Как показано на Фиг.14, флуоресцентный сигнал для нуклеиновокислотной последовательности-мишени SA генерировался, когда вместе с матрицей (SA_TD1_Chip_M, RFU: 65467,0±5,7) использовали иммобилизованный TD-зонд с двумя метками, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации. В случае с использованием TD-зонда, содержащего три ошибочно спаренных нуклеотида на своем 5'-концевом втором участке гибридизации, никакого сигнала не наблюдали (SA_TD1_Chip_m1, RFU: 6679,5±222,7). С другой стороны, наблюдали генерирование сигналов, когда использовали традиционные зонды, имеющие три ошибочно спаренных нуклеотида (SA_Con_Chip_m1, RFU: 65464,0±5,7), а также имеющие спаренную последовательность (SA_Con_Chip_M, RFU: 65464,5±6,4). Не отмечено никаких сигналов в отсутствие матрицы в качестве отрицательного контроля (SA_TD1_Chip_M, RFU: 2716,5±12,0) (SA_TD1_Chip_m1, RFU: 2810,5±14,8) (SA_Con_Chip_m1, RFU: 3216,5±41,7) (SA_Con_Chip_M, RFU: 2749,5±19,1).
Эти результаты показали, что в противоположность иммобилизованному TD-зонду, иммобилизованный традиционный зонд генерирует ложноположительный сигнал при неспецифической гибридизации. Следовательно, стало ясно, что с помощью иммобилизованных TD-зондов можно детектировать нуклеиновокислотные последовательности-мишени без получения ложноположительных сигналов.
ПРИМЕР 8. Детекция нуклеиновокислотных последовательностей-мишеней с использованием TD-зондов с одной меткой, иммобилизованных на поверхности твердой подложки
В дальнейшем авторы изобретения применяли TD-зонды с одной меткой для детекции нуклеиновокислотных последовательностей-мишеней на твердой фазе.
Для этого применения в качестве матрицы использовали синтетический олигонуклеотид для гена Staphylococcus aureus (SA). TD-зонд содержит или спаренную, или ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации. TD-зонд содержит 6-FAM или 6-TAMRA (6-карбокситетраметилродамин) в качестве флуоресцентной репортерной молекулы на своем 5'-конце и поли(Т)7 в качестве линкерной ножки. Содержащий одну метку TD-зонд иммобилизовали на поверхности твердой подложки, используя аминогруппу (аминоС7) на его 3'-конце. Для проведения 5'→3'-экзонуклеолитических реакций использовали ДНК-полимеразы Bst или Taq, обладающие 5'→3'-экзонуклеазной активностью.
А. Генерирование сигнала в результате проведения экзонуклеолитической реакции
Ниже приведены использованные в этом Примере последовательности синтетической матрицы и содержащих одну метку TD-зондов:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды).
Для изготовления зондов (SEQ ID NO:30 и 31) использовали NSB9 NHS-пластинки (NSBPOSTECH, Корея). Каждый зонд, растворенный в NSB-буфере для точечного нанесения в конечной концентрации 20 мкм, печатали на NSB9 NHS-пластинке с использованием OmniGrid Accent Microarrayer (DIGILAB, США). Каждый зонд наносили в виде пятен по одной линии в формате 2×1 (пятна в двух повторах), и полученный микрочип инкубировали в камере с поддержанием ~85%-ной влажности в течение ночи. Затем пластинки промывали в буферном растворе, содержащем 2×SSPE (0,3 М хлорид натрия, 0,02 М гидрофосфат натрия и 2,0 мМ EDTA), рН 7,4 и 7,0 мМ SDS, при 37°С в течение 10 мин для удаления неспецифически связавшихся зондов и промывали дистиллированной водой. Затем ДНК-функционализированные пластинки сушили с применением центрифуги для пластинок и хранили в темноте при 4°С до применения.
Экзонуклеолитическую реакцию проводили на поверхности ДНК-функционализированной пластинки в конечном объеме 30 мкл, содержащем 10 пмоль синтетического олигонуклеотида для SA (SEQ ID NO:1) и 3 мкл 10х реакционного буфера, каждый dNTP (50 мкМ), 2 единицы ДНК-полимеразы Bst (NEB, США). Всю эту смесь помещали в камеру, собранную на поверхности стеклянной пластинки NSB, к которой были пришиты зонды. Пластинку располагали на блоке в термоциклере (Genepro B4I, Китай). Экзонуклеолитическую реакцию проводили в течение 30 мин при 50°С и останавливали, промывая дистиллированной водой при 95°С в течение 1 мин. Получение изображений осуществляли с применением конфокального лазерного сканирующего устройства Ахоn GenePix4100A (Molecular Device, США) со сканированием при разрешении с размером пикселя 5 мкм. Интенсивность флуоресценции анализировали с применением программного обеспечения для количественного анализа микрочипов GenePix pro5.1 (Molecular Device, США). Интенсивность флуоресценции выражали в виде средних значений по пятнам после вычитания локального фона. Для проверки воспроизводимости каждое пятно анализировали в двух повторах. Интенсивность флуоресценции указывает среднюю величину для дублированных пятен.
В. Генерирование сигнала в результате проведения циклической экзонуклеолитической реакции
Последовательности синтетической матрицы и TD-зондов с одной меткой, использованные в этой реакции, представляют собой:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды).
Для изготовления зондов (SEQ ID NO:32 и 33) использовали NSB9 NHS-пластинки (NSBPOSTECH, Корея). Каждый зонд, растворенный в NSB-буфере для точечного нанесения в конечной концентрации 20 мкм, печатали на NSB9 NHS-пластинке с использованием OmniGrid Accent Microarrayer (DIGILAB, США). Каждый зонд наносили в виде пятен по одной линии в формате 2×1 (пятна в двух повторах), и полученный микрочип инкубировали в камере с поддержанием ~85%-ной влажности в течение ночи. Затем пластинки промывали в буферном растворе, содержащем 2×SSPE (0,3 М хлорид натрия, 0,02 М гидрофосфат натрия и 2,0 мМ EDTA), рН 7,4 и 7,0 мМ SDS, при 37°С в течение 10 мин для удаления неспецифически связавшихся зондов и промывали дистиллированной водой. Затем ДНК-функционализированные пластинки сушили с применением центрифуги для пластинок и хранили в темноте при 4°С до применения.
Циклическую экзонуклеолитическую реакцию проводили на поверхности ДНК-функционализированной пластинки в конечном объеме 30 мкл, содержащем 10 пмоль синтетического олигонуклеотида для SA (SEQ ID NO:1) и 3 мкл 10× реакционного буфера, каждый dNTP (50 мкМ), 2 единицы ДНК-полимеразы Bst (Solgent, Корея). Всю эту смесь помещали в камеру, собранную на поверхности стеклянной пластинки NSB, к которой были пришиты зонды. Пластинку располагали на блоке в термоциклере (Genepro B41, Китай). Термоциклирование осуществляли, как приведено ниже: денатурация в течение 2 мин при 95°С и цикл (5, 10, 20, 30, 40 или 50 циклов) при 95°С в течение 20 с и 55°С в течение 20 с. Получение изображений осуществляли с применением конфокального лазерного сканирующего устройства Axon GenePix4100A (Molecular Device, США) со сканированием при разрешении с размером пикселя 5 мкм. Интенсивность флуоресценции анализировали с применением программного обеспечения для количественного анализа микрочипов GenePix pro5.1 (Molecular Device, США). Интенсивность флуоресценции выражали в виде средних значений по пятнам после вычитания локального фона. Для проверки воспроизводимости каждое пятно анализировали в двух повторах. Интенсивность флуоресценции указывает среднюю величину для дублированных пятен.
В случае экзонуклеолитической реакции, когда с матрицей использовали иммобилизованный TD-зонд с одной меткой, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации, сигнал флуоресценции на твердой подложке в конечном итоге исчезал. В случае циклической экзонуклеолитической реакции интенсивность флуоресценции на твердой подложке уменьшалась в зависимости от количества циклов.
ПРИМЕР 9. Детекция однонуклеотидной вариабельности с использованием TD-зонда и лигазы на твердой фазе
Далее авторы изобретения проверяли, можно ли с помощью TD-зонда различить однонуклеотидную вариабельность в нуклеиновокислотной последовательности с применением лигазной реакции на твердой фазе.
Для проведения этого исследования первый зонд, имеющий структуру DSO, содержит Quasar570 в качестве репортерной молекулы на своем 5'-конце, и его используют в качестве мобильного зонда. TD-зонд в качестве второго зонда содержит или спаренную последовательность, или один ошибочно спаренный нуклеотид на своем 5'-концевом втором участке гибридизации. TD-зонд содержит поли(Т)7 в качестве линкерной ножки. TD-зонд иммобилизовали на поверхности твердой подложки, используя аминогруппу (аминоС7) на его 3'-конце. В качестве матрицы использовали синтетический олигонуклеотид для гена Staphylococcus aureus (SA). Для лигирования использовали термостабильную ДНК-лигазу амплигазу (ampligase).
Ниже приведены использованные в этом Примере последовательности синтетической матрицы, первого и второго (TD) зондов:

(Подчеркнутая и выделенная жирным шрифтом буква указывает на ошибочно спаренный нуклеотид).
Для изготовления TD-зондов (SEQ ID NO:35 и 36) использовали NSB9 NHS-пластинки (NSBPOSTECH, Корея). Каждый зонд, растворенный в NSB-буфере для точечного нанесения в конечной концентрации 20 мкм, печатали на NSB9 NHS-пластинке с использованием OmniGrid Accent Microarrayer (DIGILAB, США). Каждый зонд наносили в виде пятен по одной линии в формате 2х1 (пятна в двух повторах), и полученный микрочип инкубировали в камере с поддержанием ~85%-ной влажности в течение ночи. Затем пластинки промывали в буферном растворе, содержащем 2×SSPE (0,3 М хлорид натрия, 0,02 М гидрофосфат натрия и 2,0 мМ EDTA), рН 7,4 и 7,0 мМ SDS, при 37°С в течение 10 мин для удаления неспецифически связавшихся зондов и промывали дистиллированной водой. Затем ДНК-функционализированные пластинки сушили с применением центрифуги для пластинок и хранили в темноте при 4°С до применения.
Лигазную реакцию проводили на поверхности ДНК-функционализированной пластинки в конечном объеме 30 мкл, содержащем 10 пмоль синтетического олигонуклеотида для SA (SEQ ID NO:34), 5 пмоль первого зонда (SEQ ID NO:37) и 3 мкл 10× реакционного буфера для амплигазы, содержащего 20 мМ Трис-НСl (рН 8,3), 25 мМ KCl, 10 мМ MgCl2, 0,5 мМ NAD (никотинамидадениндинуклеотид, окисленная форма) и 0,01% Triton® X-100, 0,2 мкл термостабильной ДНК-лигазы амплигазы (5 ед./мкл) (Epicentre Biotechnologies, США). Всю эту смесь помещали в камеру, собранную на поверхности стеклянной пластинки NSB, к которой были пришиты зонды. Реакцию осуществляли, как приведено ниже: гибридизацию нуклеиновой кислоты-мишени, первого зонда и иммобилизованного TD-зонда проводили при 45°С в течение 5 мин, а затем проводили лигазную реакцию в течение 30 мин при 65°С. Реакцию останавливали и осуществляли денатурацию, промывая дистиллированной водой при 95°С в течение 2 мин. Получение изображений осуществляли с применением конфокального лазерного сканирующего устройства Axon GenePix4100A (Molecular Device, США) со сканированием при разрешении с размером пикселя 5 мкм. Интенсивность флуоресценции анализировали с применением программного обеспечения для количественного анализа микрочипов GenePix pro5.1 (Molecular Device, США). Интенсивность флуоресценции выражали в виде средних значений по пятнам после вычитания локального фона. Для проверки воспроизводимости каждое пятно анализировали в двух повторах. Интенсивность флуоресценции указывает среднюю величину для дублированных пятен.
Флуоресцентный сигнал для нуклеиновокислотной последовательности-мишени SA генерировался, когда вместе с матрицей использовали TD-зонд, содержащий спаренную последовательность на своем 5'-концевом втором участке гибридизации в качестве второго зонда. В случае с использованием TD-зонда, содержащего единственный ошибочно спаренный нуклеотид на своем 5'-концевом втором участке гибридизации в качестве второго зонда, не наблюдали никакого сигнала. Эти результаты демонстрируют, что реакция лигирования согласно настоящему изобретению позволяет осуществлять детекцию однонуклеотидной вариабельности.
ПРИМЕР 10. Детекция нуклеиновокислотной последовательности-мишени по изменению флуоресценции в результате гибридизации 5'-концевого второго участка гибридизации TD-зонда
Далее авторы изобретения проверяли, можно ли применять TD-зонд, содержащий флуоресцентную молекулу на своем 5'-концевом втором участке гибридизации, для детекции мишени на основе изменения сигнала флуоресценции в зависимости от гибридизации меченого участка.
Для этого в качестве матрицы использовали геномную ДНК Staphylococcus aureus. TD-зонд имеет либо спаренную, либо ошибочно спаренную последовательность на своем 5'-концевом втором участке гибридизации. Флуоресцентную молекулу присоединяли к внутреннему нуклеотиду 5'-концевого второго участка гибридизации TD-зонда. Для амплификации мишени использовали матричную полимеразу нуклеиновых кислот, не обладающую никакой 5'→3'-экзонуклеазной активностью.
Ниже приведены использованные в этом Примере последовательности праймеров и TD-зондов с одной меткой:

(Подчеркнутые и выделенные жирным шрифтом буквы указывают ошибочно спаренные нуклеотиды).
ПЦР в режиме реального времени проводили в конечном объеме 20 мкл, содержащем 1 нг геномной ДНК S. aureus, 5 пмоль TD-зонда с одной меткой (SEQ ID NO:38 или 39), 10 пмоль прямого праймера (SEQ ID NO:15), 10 пмоль обратного праймера (SEQ ID NO:16), 2 мкл 10× буфера Штоффеля (содержащего 100 мМ Трис-HCl (рН 8,3) и 100 мМ KCl), каждый из четырех dNTP (dATP, dCTP, dGTP и dTTP) (200 мкМ), 5 мМ MgCl2 и 1 единицу ДНК-полимеразы AmpliTaq®, фрагмент Штоффеля (Applied BioSystems, US); пробирку, содержащую реакционную смесь, помещали в термоциклер для проведения ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 20 с при 95°С, 30 с при 55°С и 10 с при 72°С. Детекцию генерированного сигнала осуществляли на стадии гибридизации (60°С) каждого цикла.
Экспериментальные результаты авторов изобретения указывают на то, что TD-зонд позволяет производить дискриминацию при детекции последовательностей-мишеней посредством регистрации изменения флуоресценции от молекулы с одной меткой в зависимости от гибридизации 5'-концевого второго участка гибридизации.
Основываясь на описании предпочтительного воплощения настоящего изобретения, следует понимать, что его варианты и модификации, соответствующие сущности изобретения, могут стать очевидными специалистам в данной области, и объем данного изобретения следует определять прилагаемой формулой изобретения и ее эквивалентами.














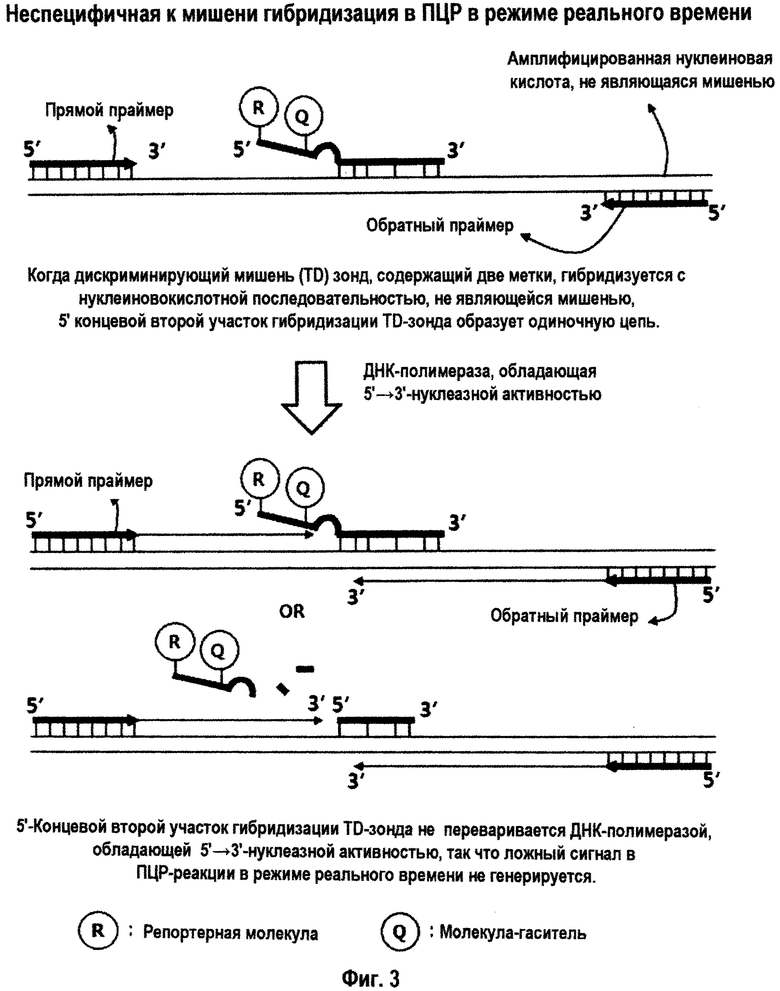





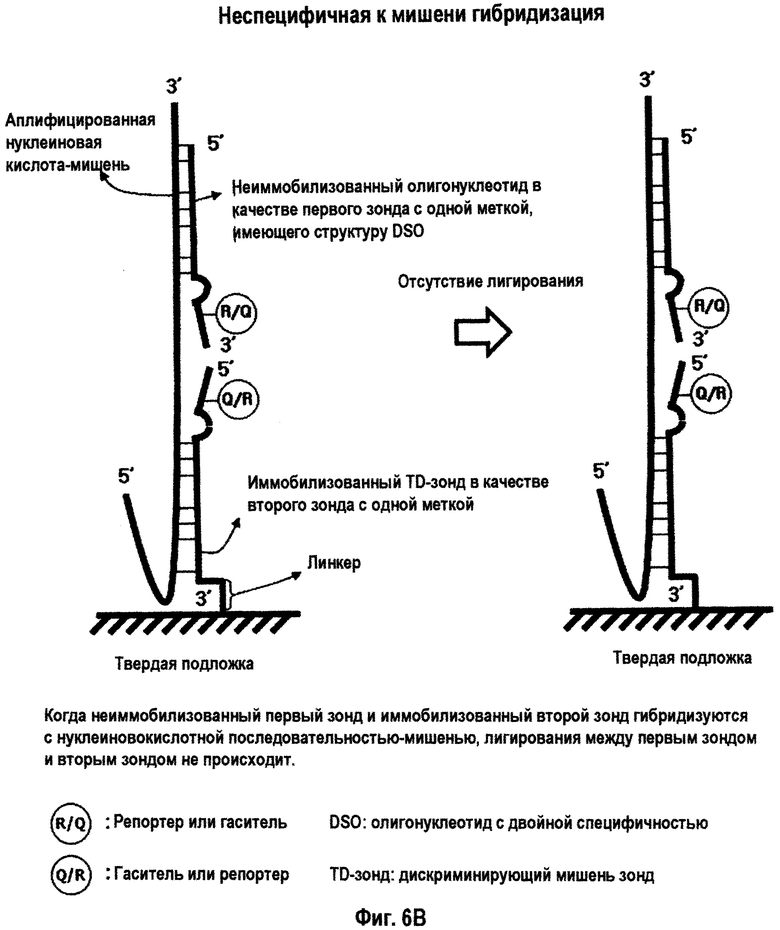
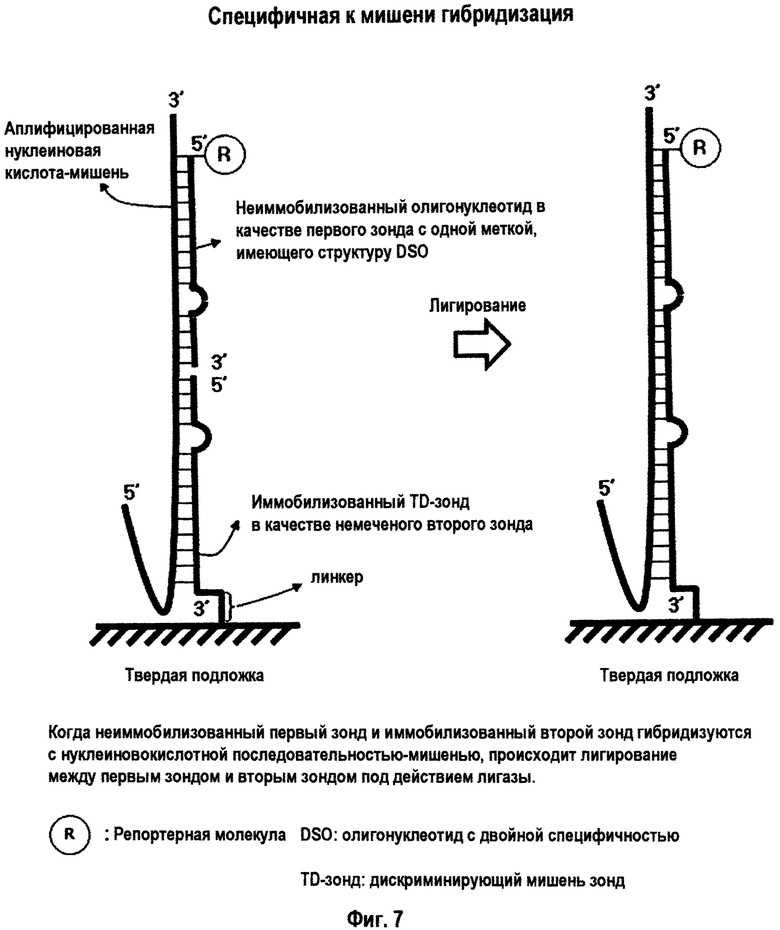

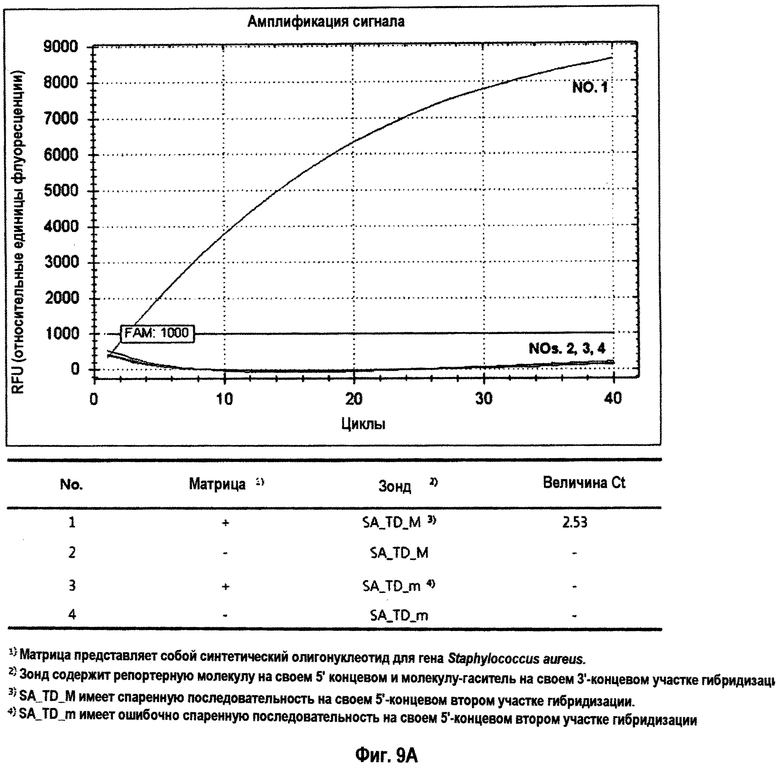





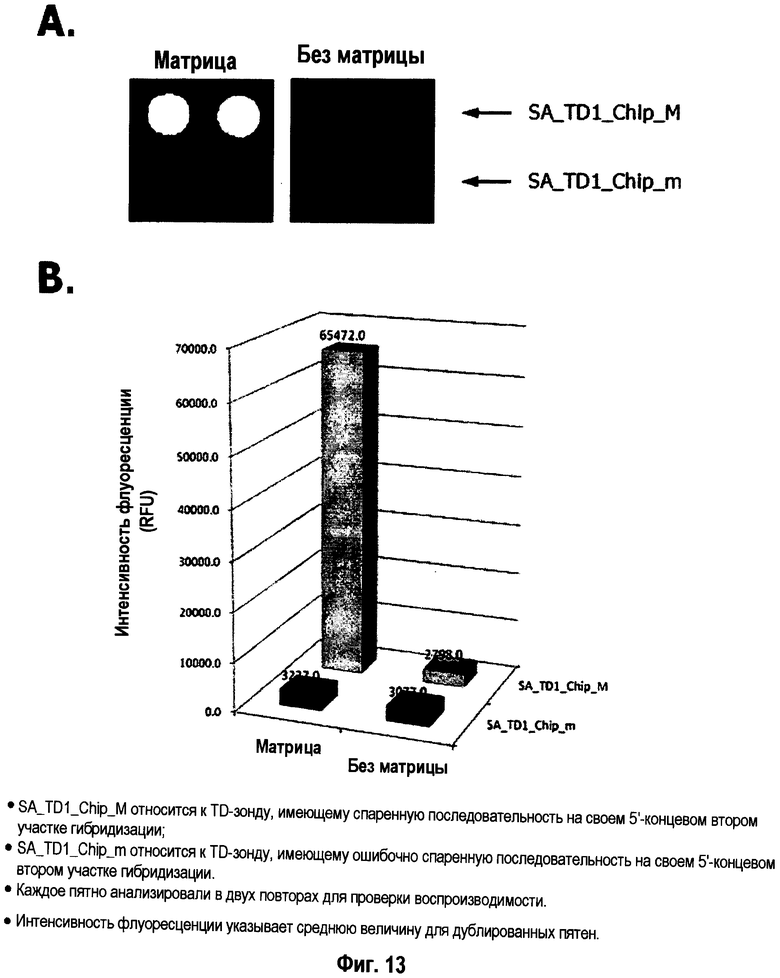

Группа изобретений касается дискриминирующего мишень зонда (TD-зонду), способа его конструирования и способов детекции нуклеиновокислотной последовательности-мишени с его использованием. TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью как через 5'-концевой второй участок гибридизации, так и 3'-концевой первый участок гибридизации. Когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5'-концевой второй участок гибридизации, и разделительный участок оба не гибридизуются с нуклеиновокислотной последовательностью, не являющейся мишенью, так что оба участка образуют одиночную цепь вследствие низкой величины своей Тпл. Представленные изобретения позволяют повысить специфичность гибридизации и могут быть использованы в области клинической диагностики и генетических исследований. 7 н. и 32 з.п. ф-лы, 14 ил., 9 пр.
1. Дискриминирующий мишень зонд (TD-зонд), имеющий структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I, позволяющий отличать (то есть дискриминировать) нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью:
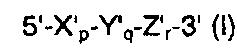 ,
,
где Х′р представляет собой 5′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y′q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z′r представляет собой 3′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; р, q и r представляют количество нуклеотидов; и X′, Y′ и Z′ представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл (температура плавления) 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х′р, Y′q и Z′r; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой второй участок гибридизации от 3′-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
2. TD-зонд по п. 1, где зонд имеет метку или систему взаимодействующих меток, содержащую некоторое количество меток для генерирования детектируемого сигнала.
3. TD-зонд по п. 1, где Тпл 3′-концевого первого участка гибридизации находится в диапазоне от 40°C до 80°C.
4. TD-зонд по п.1, где Тпл 5′-концевого второго участка гибридизации находится в диапазоне от 6°C до 40°C.
5. TD-зонд по п. 1, где Тпл разделительного участка находится в диапазоне от 2°C до 15°C.
6. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации по меньшей мере одной нуклеиновокислотной последовательности-мишени с по меньшей мере одним TD-зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
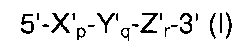 ,
,
где Х′р представляет собой 5′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y′q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z′r представляет собой 3′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит две метки - флуоресцентную репортерную молекулу и молекулу-гаситель, способную гасить флуоресценцию репортерной молекулы; по меньшей мере одна молекула из флуоресцентной репортерной молекулы и молекулы-гасителя расположена на 5′-концевом втором участке гибридизации; р, q и r представляют количество нуклеотидов; и X′, Y′ и Z′ представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х′р, Y′q и Z′r; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой второй участок гибридизации от 3′-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, а 5′-концевой второй участок гибридизации переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5′-концевой второй участок гибридизации не переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
(b) приведения в контакт продукта со стадии (а) с ферментом, обладающим 5′→3′-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, 5′-концевой второй участок гибридизации переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, с целью разъединения флуоресцентной репортерной молекулы и молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5′-концевой второй участок гибридизации не переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, в результате чего сигнал флуоресценции будет отсутствовать; и
(c) детекции сигнала флуоресценции, так что сигнал флуоресценции, генерируемый в результате переваривания 5′-концевого второго участка гибридизации, является указанием на наличие нуклеиновокислотной последовательности-мишени.
7. Способ по п. 6, где фермент, обладающий 5′→3′-экзонуклеазной активностью, представляет собой матричную полимеразу нуклеиновых кислот, обладающую 5′→3′-экзонуклеазной активностью.
8. Способ по п. 6, где стадию (а) выполняют с использованием по меньшей мере одного TD-зонда вместе с по меньшей мере одним прямым праймером, подлежащим гибридизации с сайтом, расположенным ″вниз по течению″ относительно сайта гибридизации TD-зонда, и ферментом, обладающим 5′→3′-экзонуклеазной активностью, является матричная полимераза нуклеиновых кислот, обладающая 5′→3′-экзонуклеазной активностью, так что прямой праймер удлиняется под действием матричной полимеразы нуклеиновых кислот на стадии (b); при этом 3′-конец TD-зонда блокирован для предотвращения удлинения TD-зонда.
9. Способ по п. 6, где стадию (а) выполняют с использованием TD-зонда вместе с обратным праймером, и ферментом, обладающим 5′→3′-экзонуклеазной активностью, является матричная полимераза нуклеиновых кислот, обладающая 5′→3′-экзонуклеазной активностью, так что на стадии (b) получают нуклеиновокислотную последовательность-мишень, гибридизуемую с TD-зондом благодаря реакции удлинения обратного праймера под действием матричной полимеразы нуклеиновых кислот; при этом 3′-конец TD-зонда блокирован для предотвращения удлинения TD-зонда.
10. Способ по п. 6, где флуоресцентная репортерная молекула и молекула-гаситель обе расположены на 5′-концевом втором участке гибридизации, либо флуоресцентная репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5′-концевого второго участка гибридизации и разделительного участка.
11. Способ по п. 6, где флуоресцентная репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5′-концевого второго участка гибридизации и 3′-концевого первого участка гибридизации.
12. Способ по п. 6, дополнительно включающий повторение стадий (а)-(b) или (а)-(с) вместе со стадией денатурации между повторяющимися циклами.
13. Способ по п. 6, где нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, и TD-зонд включает по меньшей мере два типа зондов.
14. Способ по п. 8, где нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа зондов и прямой праймер включает по меньшей мере два типа праймеров или обратный праймер включает по меньшей мере два типа праймеров; при этом 3′-конец TD-зонда блокирован для предотвращения удлинения TD-зонда.
15. Способ по п. 6, где нуклеотидная вариация в нуклеиновокислотной последовательности-мишени присутствует в сайте, соответствующем 5′-концевому второму участку гибридизации TD-зонда.
16. Способ по п. 6, где в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5′→3′-экзонуклеазной активностью, и блокирующий сайт расположен в 3′-концевом участке гибридизации TD-зонда.
17. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот на твердой фазе с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации по меньшей мере одной нуклеиновокислотной последовательности-мишени с по меньшей мере одним TD-зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд иммобилизован через свой 3′-конец на поверхности твердой подложки; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
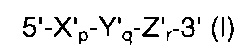
где Х′р представляет собой 5′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y′q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z′r представляет собой 3′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд имеет метку или две взаимодействующих метки, генерирующих детектируемый сигнал, и по меньшей мере одна метка расположена на 5′-концевом втором участке гибридизации TD-зонда; р, q и r представляют количество нуклеотидов; и X′, Y′ и Z′ представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х′р, Y′q и Z′r; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой второй участок гибридизации от 3′-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, а 5′-концевой второй участок гибридизации переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5′-концевой второй участок гибридизации не переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью;
(b) приведения в контакт продукта со стадии (а) с ферментом, обладающим 5′→3′-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5′-концевой второй участок гибридизации переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, с высвобождением метки из TD-зонда, в результате чего изменяется сигнал от TD-зонда, иммобилизованного на твердой подложке; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5′-концевой второй участок гибридизации не переваривается ферментом, обладающим 5′→3′-экзонуклеазной активность, в результате чего не происходит никакого изменения сигнала на TD-зонде, иммобилизованном на твердой подложке, в силу чего изменение сигнала на твердой подложке детектируют для определения наличия нуклеиновокислотной последовательности-мишени; и
(c) детекции изменения сигнала на твердой подложке, так что изменение сигнала в результате переваривания на 5′-концевом втором участке гибридизации указывает на наличие нуклеиновокислотной последовательности-мишени.
18. Способ по п. 17, где метка представляет собой флуоресцентную репортерную молекулу и изменение сигнала представляет собой уменьшение или исчезновение сигналов флуоресценции на твердой подложке.
19. Способ по п. 17, где метка представляет собой систему взаимодействующих меток, содержащую пару, состоящую из флуоресцентной репортерной молекулы и молекулы-гасителя, и TD-зонд имеет одну из молекул, выбранную из репортерной молекулы и молекулы-гасителя, расположенную в сайте на 5′-концевом втором участке гибридизации, подлежащем перевариванию ферментом, обладающим 5′→3′-экзонуклеазной активностью, а другую - в сайте, не подлежащем перевариванию ферментом, обладающим 5′→3′-экзонуклеазной активностью.
20. Способ по п. 19, где молекула-гаситель расположена в сайте на 5′-концевом втором участке гибридизации TD-зонда, подлежащем перевариванию ферментом, обладающим 5′→3′-экзонуклеазной активностью, а флуоресцентная репортерная молекула расположена в сайте, не подлежащем перевариванию ферментом, обладающим 5′→3′-экзонуклеазной активностью; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, его 5′-концевой второй участок гибридизации переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5′-концевой второй участок гибридизации не переваривается ферментом, обладающим 5′→3′-экзонуклеазной активностью, в результате чего сигнал флуоресценции будет отсутствовать; тем самым осуществляется детекция сигнала флуоресценции на твердой подложке для определения наличия нуклеиновокислотной последовательности-мишени.
21. Способ по п. 17, дополнительно включающий повторение стадий (а)-(b) или (а)-(с) вместе со стадией денатурации между повторяющимися циклами.
22. Способ по п. 17, где нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, и TD-зонд включает по меньшей мере два типа зондов.
23. Способ по п. 17, где эта нуклеиновокислотная последовательность-мишень содержит нуклеотидную вариацию и эта нуклеотидная вариация в нуклеиновокислотной последовательности-мишени присутствует в сайте, соответствующем 5′-концевому второму участку гибридизации TD-зонда.
24. Способ по п. 17, где в состав TD-зонда входит блокирующий сайт, содержащий в качестве молекулы-блокатора по меньшей мере один нуклеотид, устойчивый к расщеплению под действием фермента, обладающего 5′→3′-экзонуклеазной активностью, и блокирующий сайт расположен в 3′-концевом участке гибридизации TD-зонда.
25. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) и полимеразной цепной реакции (ПЦР), включающий стадии:
(а) приготовления смеси для ПЦР, содержащей (1) по меньшей мере одну нуклеиновокислотную последовательность-мишень, (2) по меньшей мере один TD-зонд, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, (3) пару праймеров, состоящую из двух праймеров в качестве прямого праймера и обратного праймера, каждый из которых имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (4) матричную полимеразу нуклеиновых кислот, обладающую 5′→3′-экзонуклеазной активностью; где TD-зонд гибридизуется с сайтом, расположенным между двумя этими праймерами; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
 ,
,
где Х′р представляет собой 5′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y′q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z′r представляет собой 3′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит две метки - флуоресцентную репортерную молекулу и молекулу-гаситель, способную гасить флуоресценцию репортерной молекулы; флуоресцентная репортерная молекула и молекула-гаситель обе расположены на 5′-концевом втором участке гибридизации, или репортерная молекула и молекула-гаситель каждая расположена каждый раз на своем участке, выбранном из 5′-концевого второго участка гибридизации и разделительного участка; р, q и r представляют количество нуклеотидов; и X′, Y′ и Z′ представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х′р, Y′q и Z′r; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой второй участок гибридизации от 3′-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, и 5′-концевой второй участок гибридизации переваривается в результате проявления 5′→3′-экзонуклеазной активности матричной полимеразы нуклеиновых кислот; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, вследствие чего 5′-концевой второй участок гибридизации не переваривается в результате проявления 5′→3′-экзонуклеазной активности матричной полимеразы нуклеиновых кислот, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью;
при этом 3′-конец TD-зонда блокирован для предотвращения удлинения TD-зонда;
(b) амплификации нуклеиновокислотной последовательности-мишени с использованием смеси для ПЦР посредством проведения по меньшей мере двух циклов отжига праймеров, удлинения праймеров и денатурации, при этом данные два праймера удлиняются под действием полимеразной активности матричной полимеразы нуклеиновых кислот для амплификации нуклеиновокислотной последовательности-мишени; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, 5′-концевой второй участок гибридизации переваривается в результате проявления 5′→3′-экзонуклеазной активности матричной полимеразы нуклеиновых кислот с отделением флуоресцентной репортерной молекулы от молекулы-гасителя на TD-зонде, в результате чего генерируется сигнал флуоресценции; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, 5′-концевой второй участок гибридизации не переваривается в результате проявления 5′→3′-экзонуклеазной активности матричной полимеразы нуклеиновых кислот, так что флуоресцентная репортерная молекула не отделяется от молекулы-гасителя на TD-зонде, в результате чего сигнал флуоресценции будет отсутствовать; и
(с) детекции сигнала флуоресценции, так что генерируемый сигнал флуоресценции указывает на наличие нуклеиновокислотной последовательности-мишени.
26. Способ по п. 25, где нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, TD-зонд включает по меньшей мере два типа зондов, прямой праймер включает по меньшей мере два типа праймеров и обратный праймер включает по меньшей мере два типа праймеров.
27. Способ по п. 25, где эта нуклеиновокислотная последовательность-мишень содержит нуклеотидную вариацию и эта нуклеотидная вариация в нуклеиновокислотной последовательности-мишени присутствует в сайте, соответствующем 5′-концевому второму участку гибридизации TD-зонда.
28. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда) с применением реакции лигирования, включающий стадии:
(а) гибридизации по меньшей мере одной нуклеиновокислотной последовательности-мишени с по меньшей мере одним первым зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную первому сайту нуклеиновокислотной последовательности-мишени, и по меньшей мере одним вторым зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную второму сайту нуклеиновокислотной последовательности-мишени, который является вышележащим относительно первого сайта таким образом, что 3′-конец первого зонда и 5′-конец второго зонда находятся в такой близости друг от друга, которая необходима для протекания реакции лигирования; где по меньшей мере один зонд, выбранный из первого зонда или второго зонда, содержит метку для генерации детектируемого сигнала; где второй зонд представляет собой TD-зонд; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
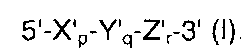 ,
,
где Х′р представляет собой 5′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y′q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z′r представляет собой 3′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; р, q и r представляют количество нуклеотидов; и X′, Y′ и Z′ представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х′р, Y′p и Z′r; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой второй участок гибридизации от 3′-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда второй зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации второго зонда оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, что позволяет осуществлять лигирование первого зонда и второго зонда; при этом, когда второй зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок второго зонда оба образуют одиночную цепь, так что первый зонд и второй зонд не подвергаются лигированию, на основании чего второй зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью;
(b) лигирования первого зонда и второго зонда, гибридизованных с нуклеиновокислотной последовательностью-мишенью, с получением лигированного зонда;
(c) денатурации продукта со стадии (b);
(d) детекции сигнала от метки на лигированном зонде, так что сигнал указывает на наличие нуклеиновокислотной последовательности-мишени.
29. Способ по п. 28, где каждый первый зонд и второй зонд несут метку, и указанные метки образуют систему взаимодействующих меток, содержащую пару, состоящую из репортерной молекулы и молекулы-гасителя.
30. Способ по п. 28, где первый зонд имеет структуру олигонуклеотида с двойной специфичностью (DSO), представленную следующей общей формулой II:
 ,
,
где Хр представляет собой 5′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; Yq представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Zr представляет собой 3′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; р, q и r представляют количество нуклеотидов, и X, Y и Z представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5′-концевого первого участка гибридизации выше, чем у 3′-концевого второго участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой первый участок гибридизации от 3′-концевого второго участка гибридизации с точки зрения событий гибридизации с нуклеиновой кислотой-мишенью, в силу чего специфичность гибридизации олигонуклеотида определяется двойственно 5′-концевым первым участком гибридизации и 3′-концевым вторым участком гибридизации, так что общая специфичность гибридизации олигонуклеотида является повышенной.
31. Способ по п. 28, дополнительно включающий повторение стадий (а)-(с) или (a)-(d).
32. Способ по п. 28, который осуществляют на твердой фазе; где первый зонд иммобилизован через свой 5′-конец на поверхности твердой подложки, а второй зонд не иммобилизован.
33. Способ по п. 28, который выполняют на твердой фазе; где второй зонд иммобилизован через свой 3′-конец на поверхности твердой подложки, а первый зонд не иммобилизован.
34. Способ по п. 28, где нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей, и первый зонд и второй зонд каждый включает по меньшей мере два типа зондов.
35. Способ по п. 57, где эта нуклеиновокислотная последовательность-мишень содержит нуклеотидную вариацию и эта нуклеотидная вариация в нуклеиновокислотной последовательности-мишени присутствует в сайте, соответствующем 5′-концевому второму участку гибридизации второго зонда.
36. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием дискриминирующего мишень зонда (TD-зонда), включающий стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с TD-зондом, имеющим гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; где TD-зонд имеет структуру модифицированного олигонуклеотида с двойной специфичностью (mDSO), представленную следующей общей формулой I:
 ,
,
где Х′р представляет собой 5′-концевой второй участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; Y′q представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания; Z′r представляет собой 3′-концевой первый участок гибридизации, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени; TD-зонд содержит в качестве метки флуоресцентную репортерную молекулу на 5′-концевом втором участке гибридизации; р, q и r представляют количество нуклеотидов; и X′, Y′ и Z′ представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков Х′р, Y′q и Z′r; длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов; разделительный участок отделяет 5′-концевой второй участок гибридизации от 3′-концевого первого участка гибридизации с точки зрения событий гибридизации с нуклеиновокислотной последовательностью-мишенью, в силу чего специфичность гибридизации TD-зонда определяется двойственно 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, так что общая специфичность гибридизации TD-зонда является повышенной;
при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью, индуцируя изменение флуоресценции от флуоресцентной репортерной молекулы; при этом, когда TD-зонд гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, не индуцируя изменения флуоресценции от флуоресцентной репортерной молекулы, на основании чего TD-зонд позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью; и
(b) детекции изменения флуоресценции, так что изменение флуоресценции указывает на наличие нуклеиновокислотной последовательности-мишени.
37. Способ по п. 36, где стадию (а) выполняют с использованием TD-зонда вместе с обратным праймером и матричной полимеразой нуклеиновых кислот, так что образуется дополнительное количество нуклеиновокислотной последовательности-мишени, гибридизуемой с TD-зондом, что усиливает изменение флуоресценции, указывающее на наличие нуклеиновокислотной последовательности-мишени.
38. Способ по п. 36, где стадию (а) выполняют с использованием TD-зонда вместе с парой праймеров, состоящей из двух праймеров в качестве прямого праймера и обратного праймера, и матричной полимеразой нуклеиновых кислот, так что нуклеиновокислотная последовательность-мишень, гибридизуемая с TD-зондом, амплифицируется в результате ПЦР, что усиливает изменение флуоресценции, указывающее на наличие нуклеиновокислотной последовательности-мишени.
39. Способ конструирования зонда, позволяющего детектировать нуклеиновокислотную последовательность-мишень, включающий стадии:
(a) выбора нуклеиновокислотной последовательности-мишени;
(b) конструирования последовательности молекулы зонда, содержащей (1) гибридизующуюся последовательность, комплементарную нуклеиновой кислоты-мишени, и (2) разделительный участок, содержащий по меньшей мере три универсальных основания, так что разделительный участок располагается посередине в гибридизующейся последовательности с образованием трех участков в молекуле зонда; и
(с) определения положения разделительного участка в молекуле зонда, позволяющего участку, расположенному в 5′-направлении от разделительного участка, иметь более низкую Тпл, чем участку, расположенному в 3′-направлении от разделительного участка, и позволяющего разделительному участку иметь наиболее низкую Тпл среди этих трех участков, тем самым обеспечивая получение молекулы зонда, имеющей три разных участка с отличающимися друг от друга величинами Тпл, среди которых (1) 5′-концевой второй участок гибридизации молекулы зонда имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени, (2) 3′-концевой первый участок гибридизации молекулы зонда имеет гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; и (3) разделительный участок молекулы зонда, расположенный между 5′-концевым вторым участком гибридизации и 3′-концевым первым участком гибридизации, содержит по меньшей мере три универсальных основания; и Тпл 5′-концевого второго участка гибридизации ниже, чем у 3′-концевого первого участка гибридизации, и разделительный участок имеет самую низкую Тпл среди этих трех участков, а длина 5′-концевого второго участка гибридизации составляет 3-15 нуклеотидов, а длина 3′-концевого первого участка гибридизации составляет 15-60 нуклеотидов;
при этом, когда молекула зонда гибридизуется с нуклеиновокислотной последовательностью-мишенью, тогда и 5′-концевой второй участок гибридизации, и 3′-концевой первый участок гибридизации второго зонда оба гибридизуются с данной нуклеиновокислотной последовательностью-мишенью; при этом, когда молекула зонда гибридизуется с нуклеиновокислотной последовательностью, не являющейся мишенью, тогда и 5′-концевой второй участок гибридизации, и разделительный участок оба образуют одиночную цепь, на основании чего молекула зонда позволяет отличать нуклеиновокислотную последовательность-мишень от нуклеиновокислотной последовательности, не являющейся мишенью.
| US2003165888 A1, 04.09.2003 | |||
| Переключающее устройство | 1979 |
|
SU777749A1 |
| СПОСОБ ЧАСТИЧНОГО СЕКВЕНИРОВАНИЯ ДНК ДЛЯ ОПРЕДЕЛЕНИЯ МУТАЦИЙ В КОРОТКИХ ФРАГМЕНТАХ ОДНОЦЕПОЧЕЧНОЙ ДНК С ИСПОЛЬЗОВАНИЕМ МИКРОЧИПА | 2001 |
|
RU2206615C1 |

