ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к детекции нуклеиновокислотной последовательности-мишени с использованием праймера, генерирующего сигнал от мишени (TSG-праймера).
СВЕДЕНИЯ О РОДСТВЕННОМ УРОВНЕ ТЕХНИКИ
Процесс амплификации нуклеиновых кислот-мишеней широко применяется в большинстве технологий детекции нуклеиновокислотных последовательностей-мишеней. Амплификация нуклеиновых кислот - это центральный процесс среди широкого спектра методов молекулярной биологии, поэтому были предложены различные способы амплификации. Например, Miller Н.I. и др. (WO 89/06700) амплифицировали нуклеиновокислотную последовательность, основываясь на гибридизации последовательности промотора/праймера с одноцепочечной ДНК-мишенью ("ssDNA") с последующей транскрипцией многих копий РНК данной последовательности. Другие известные методики амплификации нуклеиновых кислот включают системы амплификации, основанные на транскрипции (Kwoh, D. et al., Proc. Natl. Acad. Sci. U.S.A., 86:1173 (1989); и Gingeras T.R. et al, WO 88/10315).
Наиболее преобладающий способ амплификации нуклеиновых кислот, известный как полимеразная цепная реакция (обозначается далее "ПЦР"), основан на повторяющихся циклах денатурации двухцепочечной ДНК с последующим отжигом олигонуклеотидного праймера с матрицей ДНК и удлинением праймера ДНК-полимеразой (Mullis и др., патенты США №№4683195, 4683202 и 4800159; Saiki et al. (1985) Science, 230, 1350-1354).
Основанные на ПЦР методики широко используются не только для амплификации последовательности ДНК-мишени, но также для научных целей или методов в областях биологических и медицинских исследований. Это такие методики, как ПЦР с обратной. транскрипцией (ОТ-ПЦР), ПЦР с дифференциальным дизайном (ДД-ПЦР), клонирование известных или неизвестных генов с использованием ПЦР, быстрая амплификация концов кДНК (rapid amplification of cDNA ends, RACE), ПЦР с произвольными праймерами (ПП-ПЦР), множественная ПЦР, типирование генома на однонуклеотидные полиморфизмы (SNP) и основанный на ПЦР геномный анализ (McPherson and Moller. PCR. BIOS Scientific Publishers, Springer-Verlag, New York-Berlin-Heidelberg, NY, 2000).
А пока, предложенные к настоящему времени способы детекции нуклеиновых кислот-мишеней, основанные на амплификации нуклеиновых кислот, можно обобщить, как приведено ниже.
1. Способ пост-ПЦР-детекции
Пост-ПЦР-способ, который является типичным видом гетерогенного анализа, включает в себя амплификацию нуклеиновых кислот с последующей детекцией амплифицированных продуктов для анализа нуклеиновокислотной последовательности-мишени. Традиционный способ пост-ПЦР-детекции требует разделения амплифицированных продуктов либо на основе разницы в размерах, чего обычно добиваются путем применения гель-электрофореза, либо посредством иммобилизации продукта. Однако процесс разделения создает серьезные проблемы, такие как контаминация при переносе и низкая производительность.
2. Способы детекции в режиме реального времени
Чтобы преодолеть проблемы, связанные с применением пост-ПЦР-способа, был предложен способ ПЦР в режиме реального времени для детекции амплифицированных продуктов в режиме реального времени и получения их в свободном от загрязняющих веществ состоянии, дающий возможность количественного анализа нуклеиновокислотных последовательностей-мишеней.
2.1. Способы, основанные на применении меченых праймеров
2.1.1. Способ с применением праймеров "sunrise" (Sunrise™ - торговая марка фирмы Oncor)
В этом способе используются праймеры "sunrise", в которых формируются петли-"шпильки" на их 5'-концах, что сближает друг с другом пары флуорофора и гасителя, обеспечивая тем самым низкую флуоресценцию. Когда эти праймеры инкорпорированы в ПЦР-продукт, такие "хвосты" становятся двухцепочечными, и "шпилька" разворачивается, что приводит к увеличению флуоресценции (Nazarenko et al., 2516-2521, Nucleic Acids Research, 1997, v. 25, №12 и патент США №6117635). Однако, способ с применением праймеров "sunrise" очень неудобен из-за сложности конструирования праймеров, содержащих последовательность, комплементарную нуклеиновокислотным последовательностям-мишеням, и последовательность, способную образовывать петли-"шпильки" на своих 5'-концах. Кроме того, присутствие петель-"шпилек" в праймерах ухудшает эффективность их гибридизации с последовательностями-мишенями.
2.1.2. Способ с применением праймеров "scorpion" (Scorpion™ - торговая марка фирмы DxS Ltd)
В этом способе используются праймеры "scorpion", содержащие интегрированную в них сигнальную систему. Данный праймер имеет участок связывания с матрицей и "хвост", содержащий линкер и участок связывания с мишенью. Участок связывания с мишенью гибридизуется с комплементарной последовательностью в продукте удлинения праймера. Затем этот полученный в результате специфичной гибридизации с мишенью продукт, сопрягается с сигнальной системой, при этом гибридизация вызывает детектируемое изменение. Присутствие линкера в таком снабженном "хвостом" праймере предотвращает опосредованное полимеразой копирование цепи хвостового участка матрицы данного праймера (Whitcombe et al., 804-807, Nature Biotechnology, v. 17, AUGUST 1999 и патент США №6326145). Аналогично способу с применением праймеров "sunrise" также имеются трудности при конструировании и синтезе такого снабженного "хвостом" праймера, связанные с введением в праймер линкера для генерирования ампликон-зависимых сигналов, а также участка связывания с мишенью, способного гибридизоваться с продуктом удлинения праймера. Кроме того, присутствие петель-"шпилек" в праймерах снижает эффективность их гибридизации с последовательностями-мишенями.
2.1.3. Способ с применением праймеров с одной меткой (способ Lux (от англ. Light Upon eXtension))
В способе с применением праймеров с одной меткой используются праймеры, содержащие одиночную флуоресцентную метку, для детекции последовательностей-мишеней путем обнаружения изменений в характеристиках флуоресценции праймеров после гибридизации с последовательностями-мишенями (патент США №7537886). Для эффективного генерирования сигнала в этом способе также рекомендуется использовать праймеры, имеющие структуру петель-"шпилек". Более того, характеристики флуоресценции праймеров могут меняться под действием различных факторов, таких как типы меток, последовательности праймеров вокруг флуоресцентной метки, расположение флуоресцентной метки на праймерах и другие окружающие компоненты, что создает значительные трудности для оптимизации процесса конструирования праймеров.
2.1.4. Способ Lion (с использованием 3'→5'-нуклеазной активности)
В этом способе используется меченый праймер с преднамеренным ошибочным спариванием по меньшей мере по одному нуклеотиду на 3'-конце праймера. Меченый праймер инкубируют с образцом в условиях, достаточных для осуществления гибридизации, и затем этот образец обрабатывают полимеразой нуклеиновых кислот, обладающей 3'→5'-нуклеазной корректирующей активностью, в результате чего высвобождается метка или часть системы, содержащей метку (патент США №6248526).
Однако, такой праймер с ошибочным спариванием, содержащий на своем 3'-конце нуклеотид с ошибочным спариванием сконструировать нелегко. Что еще хуже, такой праймер с ошибочным спариванием по всей вероятности будет генерировать ложноположительные сигналы под действием 3'→5'-нуклеазной корректирующей активности, даже когда 3'-конец ошибочно спарен с последовательностями, не являющимися мишенями.
2.2. Способы, основанные на применении меченых зондов
2.2.1. Способы с применением молекулярных "маячков"
Молекулярные "маячки" содержат флуоресцентный и гасящий красители, однако FRET (резонансный перенос энергии флуоресценции) имеет место только тогда, когда гасящий краситель находится в непосредственной близости от флуоресцентного красителя. Конструкция молекулярных "маячков" позволяет принимать шпилечную структуру в свободном состоянии в растворе, в результате чего оба красителя будут находиться в непосредственной близости. Когда молекулярный "маячок" гибридизуется с мишенью, флуоресцентный и гасящий красители разделяются. FRET не осуществляется, и при облучении флуоресцентный краситель испускает свет (Indian J. Med. Res. 124:385-398 (2006) и Tyagi et al., Nature Biotechnology, v. 14, MARCH 1996).
Однако, у способа с применением молекулярных "маячков" имеются некоторые недостатки.
Во-первых, два инвертированных повтора шпилечной структуры должны иметь комплементарные копии в нуклеиновой кислоте-мишени, для чего в свою очередь также необходимо присутствие инвертированных повторов в этой мишени, то есть условие, которое, как правило, невыполнимо. Во-вторых, необходимо, чтобы Тпл петлевого участка шпилечной структуры, содержащего комплементарную нуклеиновокислотную последовательность, и Тпл стволового участка были строго сбалансированы с температурой, при которой проводится анализ, для осуществления специфического разворачивания шпилечного зонда в присутствии мишени и без неспецифического разворачивания. Наконец, в этом способе необходимы дополнительные праймеры, чтобы амплифицировать нуклеиновокислотные последовательности-мишени.
2.2.2. Способы с применением гибридизационных зондов
В этом способе используются четыре олигонуклеотида: два праймера и два зонда. Гибридизационные зонды содержат одиночную метку, один зонд содержит донорный флуорофор, а другой - акцепторный флуорофор. Последовательность этих двух зондов выбрана таковой, что они могут гибридизоваться с последовательностями-мишенями в направлении от головы к хвосту, "подтаскивая" эти красители очень близко друг к другу, что обеспечивает осуществление резонансного переноса энергии флуоресценции (FRET). Акцепторный краситель в одном из зондов переносит энергию, что дает возможность другому красителю рассеивать энергию флуоресценции на другой длине волны. Интенсивность флуоресценции, генерированной в процессе ПЦР, прямопропорциональна количеству ДНК-мишени (385-398, Indian J. Med. Res. 124, обзорная статья October 2006 и 303-308, и Bernad et al., 147-148, Clin. Chem., 2000, 46).
Однако этот способ невозможно адаптировать для множественной детекции, и в этом способе необходимы дополнительные праймеры, чтобы амплифицировать нуклеиновокислотные последовательности-мишени.
2.2.3. Способ с применением TaqMan-зондов (с использованием 5'→3'-нуклеазной активности)
TaqMan-зонды сконструированы для гибридизации с внутренним участком ПЦР-продукта. В процессе ПЦР, когда полимераза делает реплику матрицы, с которой связан TaqMan-зонд, под действием 5'-экзонуклеазной активности полимеразы зонд расщепляется. В результате этого, флуоресцентный и гасящий красители разделяются, и FRET более не происходит (385-398, Indian J. Med. Res., 124, обзорная статья October 2006 и 303-308, патент США №5210015).
Однако ограничения этого способа связаны с использованием в нем трех олигонуклеотидов (зонда с двойной меткой и двух праймеров). Это серьезно усложняет конструирование и синтез зондов и оптимизацию условий реакции.
2.2.4. Способ с применением самогасящегося зонда (с использованием 5'→3'-нуклеазной активности)
В способе с применением самогасящегося зонда используются зонды с двумя метками, содержащие последовательность, гибридизуемую с внутренним участком ПЦР-продукта (патент США №5723591).
Аналогично TaqMan-способу, в способе с применением самогасящегося зонда необходимо применение трех олигонуклеотидов (зонда с двумя метками и двух праймеров) для проведения гомогенного анализа, что серьезно усложняет данный способ в отношении оптимизации конструкции зондов и реакционных условий.
Как описано выше, большинство традиционных способов детекции мишеней, разработанных к настоящему времени, имеют характерные недостатки, которые считаются труднопреодолимыми.
Соответственно, существует давно ощущаемая необходимость в новом подходе к детекции нуклеиновокислотных последовательностей-мишеней с применением способа, более эффективного в техническом, временном и стоимостном выражении.
По всему тексту данной заявки даются ссылки на различные патенты и публикации и упоминания об этом приведены в скобках. Тем самым описание этих патентов и публикаций во всей их полноте включено в данную заявку посредством ссылок с целью более полного описания данного изобретения и состояния предшествующего уровня техники, к которому относится данное изобретение.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения провели интенсивные исследования с целью преодоления недостатков, связанных с традиционными технологиями детекции нуклеиновокислотных последовательностей-мишеней в режиме реального времени. Авторы настоящего изобретения разработали новые TSG (генерирующие сигнал от мишени) праймеры, способные генерировать сигналы в зависимости от гибридизации с нуклеиновокислотными последовательностями-мишенями и удлинения (праймеров), и в свою очередь разработали различные протоколы использования данных праймеров для детекции нуклеиновокислотных последовательностей-мишеней. В результате авторы изобретения убедились, что новые протоколы или способы демонстрируют приемлемые характеристики при детекции нуклеиновокислотных последовательностей-мишеней, в том числе при детекции в режиме реального времени, и с их использованием получаются сигналы, указывающие на присутствие нуклеиновокислотных последовательностей-мишеней как в жидкой фазе, так и в твердой фазе более надежным и быстрым образом.
Соответственно, данным изобретением решается задача разработки способа детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера).
Другой задачей данного изобретения является разработка способа детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера) в реакции амплификации.
Следующей задачей данного изобретения является разработка набора для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера).
Другие задачи и преимущества настоящего изобретения станут очевидны из следующего далее подробного описания в сочетании с прилагаемой формулой изобретения и графическими материалами.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Основные принципы настоящего изобретения приведены на ФИГ.1-4.
На ФИГ.1 схематично показаны стадии анализа с целью детекции нуклеиновокислотной последовательности-мишени с использованием TSG-праймера. На ФИГ.1А показано применение TSG-праймера, имеющего традиционную структуру, для детекции нуклеиновокислотной последовательности-мишени. На ФИГ.1В показано применение TSG-праймера, имеющего структуру олигонуклеотида с двойным праймированием (DPO), для повышения специфичности отжига праймера при детекции нуклеиновокислотной последовательности-мишени.
На ФИГ.2 схематично представлена ПЦР-амплификация в режиме реального времени для детекции нуклеиновой кислоты-мишени в режиме реального времени с использованием TSG-праймера по данному изобретению и матричной полимеразы нуклеиновых кислот, не обладающей 5'→3'-нуклеазной активностью. На ФИГ.2А показано применение TSG-праймера, имеющего традиционную структуру, для ПЦР-амплификации в режиме реального времени. На ФИГ.2В показано применение TSG-праймера, имеющего структуру олигонуклеотида с двойным праймированием (DPO), для повышения специфичности отжига праймера при проведении ПЦР-амплификации в режиме реального времени.
На ФИГ.3 схематично представлена ПЦР-амплификация в режиме реального времени для детекции нуклеиновой кислоты-мишени в режиме реального времени с использованием TSG-праймера по данному изобретению и матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью. На ФИГ.3А показано применение TSG-праймера, имеющего традиционную структуру, для ПЦР-амплификации в режиме реального времени. На ФИГ.3В показано применение TSG-праймера, имеющего структуру олигонуклеотида с двойным праймированием (DPO), для повышения специфичности отжига праймера при проведении ПЦР-амплификации в режиме реального времени.
На ФИГ.4 показаны основные стадии анализа с целью детекции нуклеиновокислотной последовательности-мишени с использованием TSG-праймера, иммобилизованного на твердой подложке. На ФИГ.4А показано применение TSG-праймера, имеющего традиционную структуру, для детекции нуклеиновокислотной последовательности-мишени. На ФИГ.4В показано применение TSG-праймера, имеющего структуру олигонуклеотида с двойным праймированием (DPO), для повышения специфичности отжига праймера при детекции нуклеиновокислотной последовательности-мишени.
На ФИГ.5 показаны результаты детекции только S. pneumoniae посредством гибридизации и удлинения TSG-праймеров во время реакции синтеза нуклеиновой кислоты с использованием матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью, без повторения стадий денатурации, гибридизации и удлинения праймеров.
На ФИГ.6 показаны результаты детекции только S. aureus посредством гибридизации и удлинения TSG-праймеров во время реакции синтеза нуклеиновой кислоты с использованием матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью, без повторения стадий денатурации, гибридизации и удлинения праймеров.
На ФИГ.7 показаны результаты ПЦР-амплификации в режиме реального времени для детекции S. pneumoniae с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.8 показаны результаты ПЦР-амплификации в режиме реального времени для детекции S. aureus с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.9 показаны результаты ПЦР-амплификации в режиме реального времени для детекции N. gonorrhoeae с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.10 показаны результаты ПЦР-амплификации в режиме реального времени для детекции N. meningitidis с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.11 показаны результаты измерения чувствительности ПЦР в режиме реального времени для детекции S. aureus с использованием TSG-праймера и матричной ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.12 показаны результаты генерации и аккумуляции сигнала от TSG-праймеров в зависимости от матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью или не обладающей 5'→3'-экзонуклеазной активностью, во время реакции синтеза нуклеиновой кислоты с повторением стадий денатурации, гибридизации и удлинения праймеров.
На ФИГ.13 показаны результаты ПЦР-амплификации в режиме реального времени для детекции S. pneumoniae с использованием TSG-праймеров и матричной ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.14 показаны результаты ПЦР-амплификации в режиме реального времени для детекции S. aureus с использованием TSG-праймеров и матричной ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.15 показаны результаты ПЦР-амплификации в режиме реального времени для детекции N. gonorrhoeae с использованием TSG-праймеров и матричной ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.16 показаны результаты ПЦР-амплификации в режиме реального времени для детекции N. meningitidis с использованием TSG-праймеров и матричной ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью.
На ФИГ.17 показаны результаты измерения чувствительности ПЦР в режиме реального времени для детекции S. aureus с использованием TSG-праймера и матричной ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новому способу детекции нуклеиновокислотной последовательности-мишени в режиме реального времени с использованием праймера, генерирующего сигнал от мишени (TSG-праймера), представляющего собой систему с двумя метками, и матричной полимеразы нуклеиновых кислот.
Согласно настоящему изобретению, праймер с двумя метками, называемый праймером, генерирующим сигнал от мишени (TSG-праймером), с гашением сигнала гибридизуют с нуклеиновокислотной последовательностью-мишенью для индуцирования отсутствия гашения сигнала (signal unquenching) и удлиняют с целью синтеза последовательности, комплементарной нуклеиновокислотной последовательности-мишени, детектируя в конечном итоге сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени. Другими словами, TSG-праймер претерпевает как конформационное изменение, необходимое для отсутствия гашения сигнала, так и реакцию 3'-удлинения.
Авторы настоящего изобретения впервые обнаружили, что будучи праймером без какой-либо модифицированной структуры, такой как структура петли-"шпильки", TSG-праймер может генерировать сигналы от мишени посредством перехода из состояния с гашением в состояние с отсутствием гашения в зависимости от гибридизации и удлинения праймера. TSG-праймеры и способы в режиме реального времени, в которых они применяются, впервые были предложены авторами настоящего изобретения.
Одно из ключевых преимуществ TSG-праймера состоит в том, что удлинение по 3'-концу TSG-праймера обеспечивает намного меньший разброс в значениях интенсивности сигнала относительно изменения реакционных условий (например, температуры реакции), что приводит авторов изобретения к мысли о том, что более надежные и стабильные результаты, связанные с сигналами, могут быть получены в результате удлинения по 3'-концу TSG-праймера, при этом с незначительным влиянием или без влияния на сигнал изменения в условиях реакции. В частности, удлинение по 3'-концу TSG-праймера дает возможность с использованием TSG-праймера амплифицировать нуклеиновокислотную последовательность-мишень.
TSG-праймер по настоящему изобретению в реакциях амплификации, в том числе в способах детекции, основанных на применении ПЦР в режиме реального времени, вовлечен в амплификацию мишени, а также в генерацию сигнала, что позволяет осуществлять успешный гомогенный анализ последовательностей-мишеней.
Интересно, что новый способ детекции по настоящему изобретению с использованием ПЦР в режиме реального времени, в котором применяются TSG-праймеры, "работает" существенно иным образом по сравнению с традиционными подходами с использованием ПЦР в режиме реального времени, в результате чего преодолеваются недостатки традиционных технологий и повышается эффективность детекции в режиме реального времени.
Наиболее разительным отличием способа, основанного на применении TSG-праймеров, от традиционных способов, основанных на применении зондов, как например, способов с использованием молекулярных "маячков" и TaqMan-зондов, является то, что эти меченые зонды обладают способностью только генерировать сигналы от мишеней, но не амплифицировать последовательности-мишени, a TSG-праймеры обладают способностью амплифицировать последовательности-мишени, равно как и генерировать сигналы от мишеней. В способах, основанных на применении зондов, приходится использовать дополнительные праймеры для амплификации мишени, что явно отличает их от способа по настоящему изобретению. Специалисту в данной области техники будет известно, что при использовании традиционных способов с применением меченых зондов встречаются трудности, обусловленные конструкцией этих зондов и праймеров, выбором последовательности и оптимизацией реакционных условий, поскольку для амплификации необходимы дополнительные праймеры. В противоположность этому, способ с использованием TSG-праймеров не требует дополнительных зондов, поэтому подобные проблемы могут быть сведены к минимуму.
Помимо этого, поскольку TSG-праймеры инкорпорируются в продукты удлинения или амплификации, а меченые зонды не инкорпорируются в какие-либо продукты, в способе с применением TSG-праймеров амплифицированные продукты измеряются напрямую, в то время как сигналы, полученные в способе с применением меченых зондов, нельзя считать непосредственно относящимися к продуктам амплифиции.
Кроме того, гибридизация меченых зондов с последовательностями-мишенями зависит от концентраций меченых зондов и амплифицированных продуктов, что создает трудности при проведении количественного анализа. Даже если для повышения точности количественного анализа может быть использован избыток меченых зондов, это с большой долей вероятности приведет к проблемам, связанным с высоким фоном. В противоположность этому, способ с применением TSG-праймеров позволяет осуществлять непосредственное измерение амплификации мишени с использованием меченых праймеров, что дает возможность проводить количественный анализ последовательностей-мишеней с гораздо большей точностью.
В то же время в различных традиционных, основанных на применении праймеров способах детекции в режиме реального времени, таких как "sunrise"-способ и "scorpion"-способ, перед гибридизацией с последовательностями-мишенями должна образовываться шпилечная структура для гашения их флуоресценции.
Однако, применение шпилечной структуры в способах, основанных на использовании меченых праймеров, снижает эффективность амплификации, а также эффективность гибридизации. Более того, для придания шпилечной структуры меченым праймерам необходимо наличие дополнительной последовательности; ввиду этого, при конструировании и получении праймеров необходимо учитывать наличие как комплементарной последовательности-мишени, так и последовательности, образующей шпильку. В этом контексте, конструирование меченых праймеров со шпилечной структурой будет затруднено. В противоположность этому, TSG-праймер генерирует сигналы от мишеней, не прибегая к помощи шпилечных структур, и позволяет преодолевать недостатки, связанные со шпилечными структурами.
Способ Lux, в котором применяют одиночную метку на праймерах, отличается от способа по настоящему изобретению, в котором используется система двух взаимодействующих меток на праймерах, принципом механизма, лежащего в основе генерации сигнала. На сигнал от одиночной молекулы-метки на праймерах могут оказывать влияние различные факторы, такие как типы меток, последовательности праймеров вокруг метки, расположение этой метки на праймерах и другие окружающие компоненты, что считается недостатками способа Lux.
Как изложено в данном описании выше, способ детекции в режиме реального времени по настоящему изобретению, основанный на применении TSG-праймеров, имеет несколько технических признаков (в частности, принцип генерации сигнала, структура олигонуклеотидов и функция олигонуклеотидов), отличающихся от традиционных способов, основанных на применении праймеров и зондов. Технические признаки изобретения позволяют преодолеть ограничения, присущие традиционным способам в режиме реального времени, и детектировать нуклеиновокислотные последовательности-мишени гораздо более эффективным способом.
Авторы настоящего изобретения неожиданно обнаружили, что праймеры, гибридизованные с нуклеиновокислотными последовательностями-мишенями, подвергаются реакции 5'-расщепления на своем 5'-концевом участке под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью, и реакция 5'-расщепления может быть использована для детекции последовательностей-мишенией посредством генерации сигналов для последовательностей-мишеней (см. PCT/KR2009/007064).
Когда TSG-праймер, содержащий репортерную молекулу или молекулу-гаситель на своем 5'-концевом участке, гибридизуется с последовательностью-мишенью (то есть матрицей), он подвергается реакции 5'-расщепления на своем 5'-концевом участке под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью, и репортерная молекула и молекула-гаситель далее отделяются друг от друга, способствуя отсутствию гашения сигнала от TSG-праймера.
Считается, что мера, в которой TSG-праймеры подвергаются реакции 5'-расщепления, варьирует в зависимости от 5'→3'-нуклеазной активности матричной полимеразы нуклеиновых кислот, реакционных условий и расстояния между репортерной молекулой и молекулой-гасителем.
С учетом того, что реакция 5'-расщепления осуществляется в настоящем изобретении тогда, когда используются матричная полимераза нуклеиновых кислот, обладающая 5'→3'-нуклеазной активностью, и TSG-праймеры, имеющие либо репортерную молекулу, либо молекулу-гаситель на своем 5'-концевом участке, то согласно настоящему изобретению, сигнал, указывающий на присутствие нуклеиновокислотных последовательностей-мишеней, может быть получен двумя различными способами: (1) вследствие генерации сигнала при отсутствии гашения сигнала системой взаимодействующих меток на TSG-праймере, обусловленного конформационным изменением в результате гибридизации с нуклеиновокислотной последовательностью-мишенью; и (2) вследствие генерации сигнала в результате реакции 5'-расщепления на 5'-концевом участке TSG-праймера под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью.
В настоящем изобретении при использовании матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью, предпочтительно, чтобы реакция 5'-расщепления осуществлялась только тогда, когда 5'-концевой участок (более предпочтительно, 5'-конец) TSG-праймера комплементарен нуклеиновокислотной последовательности-мишени.
Согласно настоящему изобретению нуклеиновокислотные последовательности-мишени можно детектировать в режиме реального времени с существенно повышенной эффективность и надежностью. Насколько известно авторам настоящего изобретения, эти научные данные и технологические стратегии впервые предложены авторами настоящего изобретения.
В одном из аспектов настоящего изобретения предложен способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера), включающий стадии:
(a) гибридизации нуклеиновокислотной последовательности-мишени с TSG-праймером; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что реакция 3'-удлинения индуцируется по 3'-концу TSG-праймера; и
(c) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени в ДНК или смеси нуклеиновых кислот.
Авторы настоящего изобретения провели интенсивные исследования с целью преодоления недостатков, связанных с традиционными технологиями детекции нуклеиновокислотных последовательностей-мишеней в режиме реального времени. Авторы настоящего изобретения разработали новые TSG (генерирующие сигнал от мишени) праймеры, способные генерировать сигналы в зависимости от гибридизации с нуклеиновокислотными последовательностями-мишенями и удлинения (праймеров), и в свою очередь разработали различные протоколы использования данных праймеров для детекции нуклеиновокислотных последовательностей-мишеней. В результате авторы изобретения убедились, что новые протоколы или способы демонстрируют приемлемые характеристики при детекции нуклеиновокислотных последовательностей-мишеней, в том числе при детекции в режиме реального времени, и с их использованием получаются сигналы, указывающие на присутствие нуклеиновокислотных последовательностей-мишеней как в жидкой фазе, так и в твердой фазе более надежным и быстрым образом.
Согласно настоящему изобретению TSG-праймер, предназначенный для гибридизации с нуклеиновокислотной последовательностью-мишенью, имеет систему взаимодействующих меток, содержащую репортерную молекулу и молекулу-гаситель. Если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель на TSG-праймере пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; и наоборот, если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель на TSG-праймере пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени. Таким образом, TSG-праймер выполняет две различные функции: (1) генерацию сигнала для нуклеиновокислотной последовательности-мишени; и (2) синтез последовательности, комплементарной нуклеиновокислотной последовательности-мишени.
Ввиду этого, праймер, используемый в настоящем изобретении, называется "праймером, генерирующим сигнал от мишени" (TSG-праймером), а способ по настоящему изобретению называется "анализом для детекции мишеней с применением TSG-праймеров".
Согласно настоящему изобретению сначала осуществляют гибридизацию нуклеиновокислотной последовательности-мишени с TSG-праймером.
Используемый в данном описании термин "нуклеиновая кислота-мишень", "нуклеиновокислотная последовательность-мишень" или "последовательность-мишень" относится к представляющей интерес для детекции нуклеиновокислотной последовательности, на которой отжигают праймер, либо которую гибридизуют с праймером в условиях гибридизации, отжига или амплификации.
Термин "праймер", как он использован в данном описании, относится к олигонуклеотиду, который способен действовать в качестве точки инициации синтеза в случае его помещения в условия, при которых индуцируется синтез продукта удлинения праймера, комплементарного цепи нуклеиновой кислоты (матрице), то есть в присутствии нуклеотидов и агента для полимеризации, такого как ДНК-полимераза, и при подходящих значениях температуры и рН. Предпочтительно праймер является одноцепочечным для максимальной эффективности амплификации. Предпочтительно, чтобы праймер представлял собой олигодезоксирибонуклеотид. Праймер по данному изобретению может содержать природный dNMP (дезоксинуклеозид-монофосфат) (то есть dAMP (дезоксиаденозин-монофосфат), dGMP (дезоксигуанозин-монофосфат), dCMP (дезоксицитидин-монофосфат) и dTMP (дезокситимидин-монофосфат)), модифицированный нуклеотид или неприродный нуклеотид. Праймер также может включать рибонуклеотиды.
Праймер должен иметь достаточную длину, чтобы праймировать синтез продуктов удлинения в присутствии агента для полимеризации. Точная длина праймеров будет зависеть от многих факторов, включая температуру, способ применения и источник праймера. Термин "отжиг" или "праймирование", как он использован в данном описании, относится к присоединению олигодезоксинуклеотида или нуклеиновой кислоты к являющейся матрицей нуклеиновой кислоте, при этом данное присоединению позволяет полимеразе осуществлять полимеризацию нуклеотидов с образованием молекулы нуклеиновой кислоты, которая является комплементарной к являющейся матрицей нуклеиновой кислоте или ее части.
Термин "гибридизация", используемый в данном описании, относится к образованию двухцепочечной нуклеиновой кислоты из комплементарных одноцепочечных нуклеиновых кислот. Гибридизация может осуществляться между двумя цепями нуклеиновой кислоты, полностью спаренными или по существу спаренными с некоторым числом ошибочных спариваний. Комплементарность, необходимая для гибридизация, может зависеть от условий гибридизации, в частности от температуры.
Не предполагается различия между терминами "отжиг" и "гибридизация", и эти термины будут использованы взаимозаменяемо.
Термин "TSG-праймер", используемый в данном описании, означает праймер, обладающий способностями к самогашению и отсутствию самогашения и удлинению. В частности, TSG-праймер, используемый в настоящем изобретении, означает праймер с двумя метками, который может быть использован как для синтеза последовательности, комплементарной последовательности-мишени, так и для генерирования сигналов, указывающих на присутствие нуклеиновокислотных последовательностей-мишеней, так что будучи негибридизованным с нуклеиновокислотной последовательностью-мишенью, он индуцирует гашение системы взаимодействующих меток; будучи же гибридизованным с нуклеиновокислотной последовательностью-мишенью, он индуцирует отсутствие гашения системы взаимодействующих меток, генерируя сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени.
Используемый в данном описании термин "прямой праймер" означает праймер (направление 5'→3'), комплементарный цепи нуклеиновокислотной последовательности-мишени, выравненной в 3'→5'-направлении. Обратный праймер имеет последовательность, комплементарную другой цепи нуклеиновокислотной последовательности.
Используемый в данном описании термин "пространственно близко" в отношении репортерной молекулы и молекулы-гасителя на TSG-праймере означает, что репортерная молекула и молекула-гаситель конформационно приближены друг к другу без помощи каких-либо внутримолекулярных структур праймеров, таких как петля-"шпилька".
TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) систему двух взаимодействующих меток. Термин "комплементарный" используется в данном описании для указания на то, что праймеры являются в достаточной степени комплементарными, чтобы селективно гибридизоваться с нуклеиновокислотной последовательностью-мишенью в желаемых условиях отжига или в жестких условиях, охватывая термины "существенно комплементарный" и "полностью комплементарный", предпочтительно "полностью комплементарный".
Альтернативно, TSG-праймер дополнительно может содержать на своем 5'-конце негибридизующуюся нуклеотидную последовательность, некомплементарную нуклеиновокислотной последовательности-мишени. Предпочтительно, чтобы одна из молекул на TSG-праймере, выбранная из репортерной молекулы и молекулы-гасителя, была расположена на негибридизующейся нуклеотидной последовательности, а другая расположена на гибридизующейся нуклеотидной последовательности. Негибридизующаяся нуклеотидная последовательность TSG-праймера предпочтительно не образует структуру петли-"шпильки" и не участвует в образовании внутримолекулярной структуры, такой как петля-"шпилька". Негибридизующаяся нуклеотидная последовательность TSG-праймера предпочтительно не содержит никакого сайта рестрикции.
Предпочтительно, чтобы TSG-праймер не образовывал структуру петли-шпильки".
Согласно предпочтительному воплощению система с двумя метками расположена на последовательности TSG-праймера, комплементарной мишени.
Согласно предпочтительному воплощению 5'-конец или 5'-концевой участок TSG-праймера имеет последовательность, полностью комплементарную нуклеиновокислотной последовательности-мишени.
TSG-праймер, имеет систему взаимодействующих меток, содержащую репортерную молекулу и молекулу-гаситель.
Система взаимодействующих меток представляет собой генерирующую сигнал систему, в которой энергия передается от донорной молекулы к акцепторной молекуле без участия радиоактивности. В качестве репрезентативной системы взаимодействующих меток система меток при FRET (резонансном переносе энергии флуоресценции) включает флуоресцентную репортерную молекулу (донорную молекулу) и молекулу-гаситель (акцепторную молекулу). При FRET донор энергии является флуоресцентным, а акцептор энергии может быть флуоресцентным или не быть флуоресцентным.
Для другой формы систем взаимодействующих меток донор энергии не является флуоресцентным, например, является хромофором, а акцептор энергии является флуоресцентным. Для еще одной другой формы систем взаимодействующих меток донор энергии является люминесцентным, например биолюминесцентным, хемилюминесцентным, электрохемилюминесцентным, а акцептор является флуоресцентным.
Предпочтительно, чтобы сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, генерировался системами с двумя взаимодействующими метками, наиболее предпочтительно системой FRET-меток.
В том случае, когда в TSG-праймере используют FRET-метку, две метки (флуоресцентная репортерная молекула и молекула-гаситель) отделены друг от друга для индуцирования отсутствия гашения, когда TSG-праймер, гибридизованный с нуклеиновокислотной последовательностью-мишенью, находится в вытянутой конформации. В том случае, когда TSG-праймер, негибридизованный с нуклеиновокислотной последовательностью-мишенью, находится в свернутой конформации, эти две метки располагаются близко друг к другу, индуцируя гашение сигнала.
Используемые в данном описании термины "гашение" и "отсутствие гашения" следует истолковывать как относительные. Например, термин "отсутствие гашения" может быть отнесен к более низкой эффективности или низкому уровню гашения по сравнению с термином "гашение". Другими словами, термин "гашение сигнала от репортерной молекулы" охватывает аннулирование сигнала, а также уменьшение уровней сигнала по сравнению со случаем, при котором гашения не происходит. Кроме того, термин "отсутствие гашения сигнала от репортерной молекулы" охватывает полное восстановление сигнала, а также увеличение уровней сигнала по сравнению со случаем наличия гашения.
Сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, может быть получен на основании изменения уровней гашения сигнала, как описано выше. Например, когда для детекции нуклеиновой кислоты-мишени измеряют относительные интенсивности флуоресценции от флуоресцентной репортерной молекулы, TSG-праймер, негибридизованный с нуклеиновокислотной последовательностью-мишенью, демонстрирует относительно низкую интенсивность флуоресценции (состояние с гашением) ввиду того, что флуоресцентная репортерная молекула и молекула-гаситель территориально (или пространственно) располагаются близко друг к другу. Когда TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то детектируют относительно высокую интенсивность флуоресценции (состояние с отсутствием гашения) ввиду того, что флуоресцентная репортерная молекула и молекула-гаситель территориально отделены друг от друга.
Репортерная молекула и молекула-гаситель могут быть расположены в любом месте TSG-праймера, при условии, что происходит переключение состояний с гашением-отсутствием гашения.
Согласно предпочтительному воплощению репортерная молекула и молекула-гаситель расположены на расстоянии 4-50 нуклеотидов друг от друга.
Согласно предпочтительному воплощению репортерная молекула и молекула-гаситель расположены на расстоянии не более 50 нуклеотидов, более предпочтительно не более 40 нуклеотидов, еще более предпочтительно не более 30 нуклеотидов, существенно более предпочтительно не более 25 нуклеотидов друг от друга.
Согласно предпочтительному воплощению репортерная молекула и молекула-гаситель отделены по меньшей мере 4 нуклеотидами, более предпочтительно по меньшей мере 6 нуклеотидами, еще более предпочтительно по меньшей мере 10 нуклеотидами, существенно более предпочтительно по меньшей мере 15 нуклеотидами.
Согласно предпочтительному воплощению репортерная молекула или молекула-гаситель на TSG-праймере расположена на его 5'-конце или на расстоянии 1-5 нуклеотидов от его 5'-конца. Например, репортерная молекула на TSG-праймере расположена на его 5'-конце или на расстоянии 1-5 нуклеотидов от его 5'-конца, а молекула-гаситель расположена на расстоянии 4-50 нуклеотидов от репортерной молекулы.
Согласно предпочтительному воплощению репортерная молекула на TSG-праймере расположена на его 5'-конце или на расстоянии 1-10 нуклеотидов от его 5'-конца, более предпочтительно на его 5'-конце.
Согласно предпочтительному воплощению молекула-гаситель на TSG-праймере расположена на его 5'-конце или на расстоянии 1-10 нуклеотидов от его 5'-конца, более предпочтительно на его 5'-конце,
Репортерная молекула и молекула-гаситель, полезные в настоящем изобретении, могут включать любые молекулы, известные в данной области техники. Примерами их являются: Су2™ (506), YO-PRO™-1 (509), YOYO™-1 (509), кальцеин (517), FITC (флуоресцеинизотиоцианат) (518), FluorX™ (519), Alexa™ (520), родамин 110 (520), 5-FAM (карбоксифлуоресцеин) (522), Oregon Green™ (Орегон зеленый) 500 (522), Oregon Green™ 488 (524), RiboGreen™ (525), Rhodamine Green™ (родаминовый зеленый) (527), родамин 123 (529), Magnesium Green™ (531), Calcium Green™ (533), TO-PRO™-1 (533), TOT01 (533), JOE (548), BODIPY530/550 (550), Dil (565), BODIPY TMR (568), BODIPY558/568 (568), BODIPY564/570 (570), Су3™ (570), Alexa™ 546 (570), TRITC (тетраметилродамин-изотиоцианат) (572), Magnesium Orange™ (575), фикоэритрин R&B (575), родамин-фаллоидин (575), Calcium Orange™ (576), пиронин Y (580), родамин В (580), TAMRA (тетраметилродамин) (582), Rhodamine Red™ (590), Cy3.5™ (596), ROX (6-карбокси-Х-родамин) (608), Calcium Crimson™ (615), Alexa™ 594 (615), техасский красный (615), нильский красный (628), YO-PRO™-3 (631), YOYO™-3 (631), R-фикоцианин (642), C-фикоцианин (648), TO-PRO™-3 (660), TOT03 (660), DiD DilC(5) (665), Cy5™ (670), тиадикарбоцианин (671) и Су5.5 (694). Числа в скобках представляют собой длину волны в нанометрах, соответствующую максимуму излучения.
Подходящие пары репортер-гаситель изложены в ряде публикаций, приведенных далее: в Pesce et al., editors, Fluorescence Spectroscopy (Marcel Dekker, New York, 1971); White et al., Fluorescence Analysis: A Practical Approach (Marcel Dekker, New York, 1970); Berlman, Handbook of Fluorescence Spectra of Aromatic Molecules, 2nd Edition (Academic Press, New York, 1971); Griffiths, Color and Constitution of Organic Molecules (Academic Press, New York, 1976); Bishop, editor, Indicators (Pergamon Press, Oxford, 1972); Haugland, Handbook of Fluorescent Probes and Research Chemicals (Molecular Probes, Eugene, 1992); Haugland, R. P., Handbook of Fluorescent Probes and Research Chemicals, 6th Edition, Molecular Probes, Eugene, Oreg., 1996; патентах США №№3996345 и 4351760.
Стоит отметить, что в настоящем изобретении может быть использована нефлуоресцентная "черная" молекула-гаситель, способная гасить флуоресценцию в широком диапазоне длин волн или на конкретной длине волны.
Для FRET-меток, адаптированных к TSG-праймеру, термин "репортерная молекула" включает в себя донора FRET, а термин "гаситель" включает в себя другого партнера (акцептора) FRET. Например, в качестве репортерной молекулы используют флуоресцентный краситель, а в качестве гасителя родаминовый краситель.
После гибридизации с нуклеиновокислотной последовательностью-мишенью вещество, полученное на стадии (а), приводят в контакт с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера для удлинения TSG-праймера, гибридизованного с нуклеиновокислотной последовательностью-мишенью.
Фраза "в условиях удлинения праймера" означает условия, достаточные для индуцирования реакции удлинения TSG-праймера по 3'-концу под действием матричной полимеразы нуклеиновых кислот. Такими условиями могут быть условия для удлинения праймера традиционными полимеразами нуклеиновых кислот. Например, эти условия можно найти в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001). В качестве иллюстративныго примера условия включают инкубацию нуклеиновокислотной последовательности-мишени, TSG-праймера, термостабильной ДНК-полимеразы (например, Tag ДНК-полимеразы), dNTP и MgCl2 при относительно высокой температуре (например, 50-75°С) в течение подходящего периода времени.
Удлинение TSG-праймера является важнейшей стадией настоящего изобретения. Когда в результате реакции удлинения TSG-праймера получают продукт удлинения, это приводит к более стабильной гибридизации последовательности TSG-праймера, инкорпорированной в продукт удлинения, с нуклеиновокислотной последовательностью-мишенью. В дополнение к этому, реакция удлинения TSG-праймера дает возможность инкорпорироваться в продукт удлинения меткам на TSG-праймере, тем самым увеличивая интенсивность сигнала соответственно количеству амплифицированного продукта. Такое одновременное событие, полученное в настоящем изобретении, позволяет осуществлять более точный количественный анализ нуклеиновокислотной последовательности-мишени. Согласно настоящему изобретению, в отличие от меченых зондов, когда сам TSG-праймер неспецифически гибридизуется с последовательностями, не являющимися мишенями, невозможна генерация каких-либо ложноположительных сигналов при осуществлении детекции сигнала в условиях высокой жесткости, таких как высокие температуры (например, 50-85°С). Кроме того, результатом удлинения TSG-праймера является амплификация нуклеиновокислотной последовательности-мишени, дающая возможность амплификации сигнала одновременно с амплификацией мишени.
Согласно настоящему изобретению матричная полимераза нуклеиновых кислот включает любую полимеразу нуклеиновых кислот, известную в данной области.
Одно из значительных преимуществ настоящего изобретения заключается в генерации сигналов от мишеней даже при использовании матричной полимеразы нуклеиновых кислот, не обладающей нуклеазными активностями.
Согласно предпочтительному воплощению матричная полимераза нуклеиновых кислот, используемая в настоящем изобретении, не обладает нуклеазной активностью.
Согласно предпочтительному воплощению матричная полимераза нуклеиновых кислот, используемая в настоящем изобретении, не обладает 5'→3'-нуклеазной активностью.
Матричная полимераза нуклеиновых кислот, которая может быть использована в настоящем изобретении, может включать любую из полимераз нуклеиновых кислот, например, фрагмент Кленова ДНК-полимеразы I из Е.coli, термостабильную ДНК-полимеразу и ДНК-полимеразу бактериофага Т7. Предпочтительно, полимераза представляет собой термостабильную ДНК-полимеразу, которая может быть получена из ряда бактериальных видов.
Согласно предпочтительному воплощению, когда матричная полимераза нуклеиновых кислот, используемая в настоящем изобретении, обладает 5'→3'-нуклеазной активностью, TSG-праймер удлиняется по своему 3'-концу под действием полимеразной активности матричной полимеразы нуклеиновых кислот и расщепляется на своем 5'-конце под действием 5'→3'-нуклеазной активности матричной полимеразы нуклеиновых кислот, так что высвобождается либо репортерная молекула, либо молекула-гаситель, генерируя сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени.
Предпочтительно, матричная полимераза нуклеиновых кислот, обладающая 5'→3'-нуклеазной активностью, представляет собой термостабильную ДНК-полимеразу, которая может быть получена из ряда бактериальных видов, включая Thermus aquaticus (Taq), Thermus thermophilus, Thermus filiformis, Thermus flavus, Thermus antranikianii, Thermus caldophilus, Thermus chliarophilus, Thermus igniterrae, Thermus lacteus, Thermus oshimai, Thermus ruber, Thermus rubens, Thermus scotoductus, Thermus silvanus, Thermus species 205 и Thermus species sps 17. Наиболее предпочтительно, чтобы матричная полимераза нуклеиновых кислот, обладающая 5'→3'-нуклеазной активностью, представляла собой Taq ДНК-полимеразу.
Примечательно, что авторы настоящего изобретения обнаружили, что TSG-праймер, гибридизованный с нуклеиновокислотными последовательностями-мишенями, удлиняется по своему 3'-концу и также расщепляется на своем 5'-концевом участке (например, 5'-конце) только в результате контактирования с матричной полимеразой нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью. Реакция расщепления на 5'-концевом участке TSG-праймера также ответственна за генерацию сигнала в настоящем изобретении.
Итак, когда при осуществлении настоящего изобретения предусматривается реакция 5'-расщепления TSG-праймера, тогда согласно настоящему изобретению, сигнал, указывающий на присутствие нуклеиновокислотных последовательностей-мишеней, может быть получен двумя различными способами: (1) вследствие генерации сигнала при отсутствии гашения сигнала системой взаимодействующих меток на TSG-праймере, обусловленного конформационным изменением в результате гибридизации с нуклеиновокислотной последовательностью-мишенью; и (2) вследствие генерации сигнала в результате реакции 5'-расщепления на 5'-концевом участке TSG-праймера под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью.
Согласно предпочтительному воплощению TSG-праймер содержит последовательность, спаренную с нуклеиновокислотной последовательностью-мишенью, на своем 5'-конце или на своем 5'-концевом участке.
Используемый в данном описании в связи с TSG-праймером термин "5'-концевой участок" относится к участку или области, содержащему(ей) любую длинную непрерывную последовательность, начиная от 5'-конца TSG-праймера. Предпочтительно, чтобы 5'-концевой участок TSG-праймера имел 5'-конец и последовательность, содержащую 1-10 нуклеотидов (более предпочтительно 1-7 нуклеотидов, еще более предпочтительно 1-5 нуклеотидов, существенно более предпочтительно 1-3 нуклеотида), помимо 5'-конца.
Альтернативно, в настоящем изобретении можно использовать матричную полимеразу нуклеиновых кислот, обладающую 3'→5'-экзонуклеазной активностью.
Согласно предпочтительному воплощению, если используется матричная полимераза нуклеиновых кислот, обладающая 3'→5'-экзонуклеазной активностью, то TSG-праймер содержит по меньшей мере одну ошибочно спаренную нуклеотидную последовательность на своем 3'-концевом участке или 3'-конце.
Согласно предпочтительному воплощению, если TSG-праймер содержит по меньшей мере одну ошибочно спаренную нуклеотидную последовательность на своем 3'-концевом участке или 3'-конце, то ошибочно спаренная нуклеотидная последовательность не содержит никакой метки.
Количество ошибочно спаренных нуклеотидов может составлять 1-5, предпочтительно 1-4, более предпочтительно 1-3, еще более предпочтительно 1-2 и наиболее предпочтительно 1. Если праймеры несут в себе по меньшей мере 2 ошибочно спаренных нуклеотида, то такие ошибочно спаренные нуклеотиды могут быть расположены непрерывно или с промежутками.
Согласно предпочтительному воплощению, если используется матричная полимераза нуклеиновых кислот, обладающая 3'→5'-экзонуклеазной активностью, то TSG-праймер содержит по меньшей мере один ошибочно спаренный нуклеотид, имеющий остов, устойчивый в отношении 3'→5'-нуклеазной активности матричных полимераз нуклеиновых кислот на своем 3'-концевом участке.
Согласно предпочтительному воплощению, если используется матричная полимераза нуклеиновых кислот, обладающая 3'→5'-экзонуклеазной активностью, то TSG-праймер содержит на своем 3'-конце единственный определяющий спаривание нуклеотид, имеющий остов, устойчивый в отношении 3'→5'-нуклеазной активности матричных полимераз нуклеиновых кислот.
Этот единственный определяющий спаривание нуклеотид на 3'-конце TSG-праймера будет образовывать пару оснований только тогда, когда он гибридизуется со спаривающимся с ним нуклеотидом на противоположной цепи. Однако этот единственный определяющий спаривание нуклеотид TSG-праймера не может образовывать пару оснований, когда нуклеотид, присутствующий на противоположной цепи, не является комплементарным данному единственному определяющему спаривание нуклеотиду.
В настоящем изобретении, ввиду того, что единственный определяющий спаривание нуклеотид на 3'-конце TSG-праймера содержит остов, устойчивый в отношении 3'→5'-нуклеазной активности, реакция расщепления не индуцируется, даже если 3'-конец TSG-праймера не является спаренным, вследствие этого не индуцируется никакой реакции удлинения.
Удлинение TSG-праймера приводит к более стабильной гибридизации с последовательностями-мишенями по сравнению с неудлиненным TSG-праймером. Таким образом, регулируя температуру, можно выполнить детекцию присутствия удлиненного продукта (то есть, последовательности-мишени) путем анализа изменений в сигналах.
Предпочтительно, настоящее изобретение используют для детекции вариабельности нуклеотидов. Более предпочтительно, вариабельность нуклеотидов, детектируемая в данном изобретении, представляет собой замену оснований, еще более предпочтительно, SNP (однонуклеотидный полиморфизм) и точечную мутацию.
Согласно предпочтительному воплощению вариабельность нуклеотидов может присутствовать в сайте, расположенном напротив единственного определяющего спаривание нуклеотида на 3'-конце TSG-праймера.
Остов, устойчивый в отношении 3'→5'-нуклеазной активности, включает любой, известный специалисту в данной области. Например, он включает различные фосфоротиоатные связи, фосфонатные связи, фосфороамидатные связи и углеводы с модификациями по 2'-положению. Согласно предпочтительному воплощению нуклеотиды, имеющие остов, устойчивый к 3'→5'-нуклеазе, содержат фосфоротиоатную связь, алкилфосфотриэфирную связь, арилфосфотриэфирную связь, алкилфосфонатную связь, арилфосфонатную связь, гидрофосфонатную связь, алкилфосфороамидатную связь, арилфосфороамидатную связь, фосфороселенатную связь, модификацию 2'-O-аминопропил, модификацию 2'-O-алкил, модификацию 2'-O-аллил, модификацию 2'-O-бутил, α-аномерный олигодезоксинуклеотид и модификацию 1-(4'-тио-β-D-рибофуранозил).
Согласно предпочтительному воплощению матричная полимераза нуклеиновых кислот, обладающая 3'→5'-экзонуклеазной активностью, представляет собой термостабильную ДНК-полимеразу, которая может быть получена из ряда бактериальных видов, включая Thermococcus litoralis, Thermococcus barossi, Thermococcus gorgonarius, Thermotoga maritima, Thermotoga neapolitana, Thermosipho africanus, Pyrococcus furiosus(Pfu), Pyrococcus woesei, Pyrococcus horikoshii, Pyrococcus abyssi, Pyrodictium occultum, Aquifex pyrophilus и Aquifex aeolieus. Наиболее предпочтительно, чтобы матричная полимераза нуклеиновых кислот, обладающая 3'→5'-нуклеазной активностью, представляла собой Pfu ДНК-полимеразу.
Согласно предпочтительному воплощению настоящее изобретение дополнительно включает повторение по меньшей мере дважды стадий (а)-(b) или (а)-(с) вместе со стадией денатурации между повторяющимися циклами с целью усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени. Число повторений циклов конкретно не ограничивается, в типичном случае составляет по меньшей мере два, предпочтительно по меньшей мере пять, более предпочтительно по меньшей мере десять. В результате денатурации двухцепочечные дуплексы, образованные на стадии (а), превращаются в одноцепочечные нуклеиновые кислоты. Способы денатурации включают, но не ограничиваются этим, нагревание, обработку щелочью, формамидом, мочевиной и глиоксалем, ферментативные методы (например, действие геликазы) и применение связывающих белков. Например, денатурации можно достичь посредством нагревания при температуре, изменяющейся в диапазоне от 80°С до 105°С. Общие способы осуществления такой обработки приведены в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001).
В конце осуществляют детекцию сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени. Детекцию сигналов можно осуществлять для каждого цикла такого повторения (то есть в режиме реального времени), в конце повторения (то есть в режиме конечной точки) или в каждый из предварительно заданных интервалов времени в процессе повторения. Предпочтительно, чтобы детекция сигнала могла быть осуществлена для каждого цикла повторения с целью улучшения точности детекции.
Детекцию или измерение сигнала можно осуществить традиционными для каждой метки методами. Например, детекцию или измерение сигнала флуоресценции можно осуществить традиционными методами, например, с использованием флуориметров.
Преимущества настоящего изобретения становятся очевидными на стадии детекции сигнала. Когда детекцию сигнала осуществляют в условиях высокой жесткости, таких как высокие температуры (например, 50-85°С), ложноположительные сигналы, обусловленные гибридизацией TSG-праймера с нуклеиновокислотными последовательностями, не являющимися мишенями, могут быть полностью исключены.
Согласно предпочтительному воплощению детекцию сигнала осуществляют путем измерения или анализа изменения сигнала от репортерной молекулы системы меток, состоящей из этой репортерной молекулы и молекулы-гасителя.
Согласно предпочтительному воплощению, когда молекула-гаситель представляет собой флуоресцентную молекулу, детекцию сигнала осуществляют путем измерения изменения сигнала от молекулы-гасителя или изменений сигнала как от молекулы-гасителя, так и от репортерной молекулы.
Согласно предпочтительному воплощению флуоресцентной является молекула-гаситель, и подлежащий детекции сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, представляет собой сигнал от флуоресцентной молекулы-гасителя.
Одним из значимых признаков настоящего изобретения является возможность успешного получения сигналов, как в жидкой фазе, так и в твердой фазе. Настоящее изобретение может быть осуществлно в двух фазах, то есть в жидкой фазе и в твердой фазе.
I. Детекция мишени в жидкой фазе
1. Детекция мишени с использованием TSG-праймера и полимеразы нуклеиновых кислот
В соответствии с первым протоколом детекцию нуклеиновокислотной последовательности-мишени осуществляют, используя TSG-праймер и матричную полимеразу нуклеиновых кислот (см. Фиг.1А и 1В).
Первый протокол включает стадии:
(a) гибридизации нуклеиновокислотной последовательности-мишени с TSG-праймером; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что реакция 3'-удлинения индуцируется по 3'-концу TSG-праймера; и
(c) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени в ДНК или смеси нуклеиновых кислот.
Согласно предпочтительному воплощению детекцию на стадии (с) осуществляют в режиме реального времени, в режиме конечной точки или в предварительно заданный интервал времени.
2. Детекция мишени с использованием TSG-праймера и полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью
В соответствии с первым протоколом детекцию нуклеиновокислотной последовательности-мишени осуществляют, используя TSG-праймер и матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-нуклеазной активностью.
Предпочтительно, второй протокол включает стадии:
(a) гибридизации нуклеиновокислотной последовательности-мишени с TSG-праймером; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью, в условиях удлинения и расщепления праймера, так что индуцируются реакция 3'-удлинения по 3'-концу TSG-праймера и реакция 5'-расщепления на 5'-концевом участке TSG-праймера, в результате чего из TSG-праймера высвобождается либо репортерная молекула, либо молекула-гаситель, и генерируется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени; и
(c) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени в ДНК или смеси нуклеиновых кислот.
Согласно предпочтительному воплощению настоящее изобретение дополнительно включает повторение по меньшей мере дважды стадий (а)-(b) или (а)-(с) вместе со стадией денатурации между повторяющимися циклами с целью усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени.
Согласно предпочтительному воплощению детекцию на стадии (с) осуществляют в режиме реального времени, в режиме конечной точки или в предварительно заданный интервал времени.
3. Детекция мишени с использованием TSG-праймера, взаимодополняющего праймера и полимеразы нуклеиновых кислот
В соответствии с третьим протоколом детекцию нуклеиновокислотной последовательности-мишени осуществляют, используя (1) матричную полимеразу нуклеиновых кислот и (2) пару праймеров, состоящую из TSG-праймера и взаимодополняющего праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень, так что сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, амплифицируется одновременно с амплификацией мишени (см. Фиг.2А и 2В).
Взаимодополняющий праймер может быть использован в качестве TSG-праймера или не TSG-праймера,
Предпочтительно, в соответствии с третьим протоколом детекцию нуклеиновокислотной последовательности-мишени в ДНК или смеси нуклеиновых кислот осуществляют посредством реакций амплификации с применением TSG-праймера, включающих стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с парой праймеров, состоящей из двух праймеров в качестве прямого праймера и обратного праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень; при этом по меньшей мере один из этих двух праймеров представляет собой TSG-праймер; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что индуцируется реакция 3'-удлинения по 3'-концу этих двух праймеров;
(c) денатурации продукта со стадии (b);
(d) повторения стадий (а)-(с) по меньшей мере дважды с целью как амплификации нуклеиновокислотной последовательности-мишени, так и усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени; и
(e) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, при этом детекцию осуществляют для каждого цикла повторения стадии (d), в конце повторения стадии (d) или в каждый из предварительно заданных интервалов времени в процессе повторения, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени.
4. Детекция мишени с использованием TSG-праймера, взаимодополняющего праймера и полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью
В соответствии с четвертым протоколом детекцию нуклеиновокислотной последовательности-мишени осуществляют, используя (1) матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-нуклеазной активностью, и (2) пару праймеров, состоящую из TSG-праймера и взаимодополняющего праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень, так что сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, амплифицируется одновременно с амплификацией мишени (см. Фиг.3А и 3В).
Предпочтительно, четвертый протокол включает стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с парой праймеров, состоящей из двух праймеров в качестве прямого праймера и обратного праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень; при этом по меньшей мере один из этих двух праймеров представляет собой TSG-праймер; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения и расщепления праймера, так что индуцируются реакция 3'-удлинения по 3'-концу TSG-праймера и реакция 5'-расщепления в 5'-концевой части TSG-праймера, в результате чего из TSG-праймера высвобождается либо репортерная молекула, либо молекула-гаситель, и генерируется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени; и
(c) денатурации продукта со стадии (b);
(d) повторения стадий (а)-(с) по меньшей мере дважды с целью как амплификации нуклеиновокислотной последовательности-мишени, так и усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени; и
(е) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, при этом детекцию осуществляют для каждого цикла повторения стадии (d), в конце повторения стадии (d) или в каждый из предварительно заданных интервалов времени в процессе повторения, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени.
II. Детекция мишени на твердой фазе
Значительное преимущество настоящего изобретения заключается в эффективной детекции нуклеиновокислотных последовательностей-мишеней даже на твердой фазе, такой как микрочип.
Согласно предпочтительному воплощению настоящее изобретение осуществляют на твердой фазе и TSG-праймер иммобилизуют через его 5'-конец на поверхности твердой подложки.
1. Детекция мишени на чипе с использованием TSG-праймера и полимеразы нуклеиновых кислот
В соответствии с первым протоколом для твердой фазы детекцию нуклеиновокислотной последовательности-мишени осуществляют, используя TSG-праймер и матричную полимеразу нуклеиновых кислот на твердой фазе (см. Фиг.4А и 4В).
Предпочтительно, первый протокол для твердой фазы включает стадии:
(a) гибридизации нуклеиновокислотной последовательности-мишени с TSG-праймером; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом TSG-праймер иммобилизован через свой 5'-конец на поверхности твердой подложки; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что индуцируется реакция 3'-удлинения по 3'-концу TSG-праймера; и
(c) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени на твердой подложке, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени в ДНК или смеси нуклеиновых кислот.
2. Детекция мишени на чипе с использованием TSG-праймера, взаимодополняющего праймера и полимеразы нуклеиновых кислот
В соответствии со вторым протоколом для твердой фазы по настоящему изобретению детекцию нуклеиновокислотной последовательности-мишени осуществляют, используя (1) матричную полимеразу нуклеиновых кислот и (2) пару праймеров, состоящую из TSG-праймера и взаимодополняющего праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень, так что сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, амплифицируется одновременно с амплификацией мишени.
Другими словами, второй протокол на твердой фазе осуществляют с использованием технологии ПЦР на чипе в режиме реального времени.
Предпочтительно, второй протокол на твердой фазе включает стадии:
(a) гибридизации нуклеиновокислотной последовательности-мишени с парой праймеров, состоящей из двух праймеров в качестве прямого праймера и обратного праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень; при этом по меньшей мере один из этих двух праймеров представляет собой TSG-праймер; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом по меньшей мере один из этих двух праймеров иммобилизован через свой 5'-конец на поверхности твердой подложки и иммобилизованный праймер представляет собой TSG-праймер; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что индуцируется реакция 3'-удлинения по 3'-концам этих двух праймеров; и
(c) денатурации продукта со стадии (b);
(d) повторения стадий (а)-(с) по меньшей мере дважды с целью как амплификации нуклеиновокислотной последовательности-мишени, так и усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени; и
(е) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, при этом детекцию осуществляют для каждого цикла повторения стадии (d), в конце повторения стадии (d) или в каждый из предварительно заданных интервалов времени в процессе повторения, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени.
Согласно предпочтительному воплощению TSG-праймер иммобилизован через свой 5'-конец на поверхности твердой подложки, а другой праймер не иммобилизован и не является TSG-праймером.
Согласно предпочтительному воплощению TSG-праймер и взаимодополняющий праймер имеют структуру олигонуклеотида с двойным праймированием (DPO), представленную следующей общей формулой I:
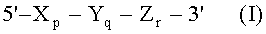
где Xp представляет собой 5'-концевой первый праймирующий участок, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; Yq представляет собой разделительный участок, содержащий по меньшей мере три универсальных основания, Zr представляет собой 3'-концевой второй праймирующий участок, имеющий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени; p, q и r представляют количество нуклеотидов, и X, Y и Z представляют собой дезоксирибонуклеотиды или рибонуклеотиды; Тпл 5'-концевого первого праймирующего участка выше, чем у 3'-концевого второго праймирующего участка, и разделительный участок имеет самую низкую Тпл среди этих трех участков; разделительный участок отделяет 5'-концевой первый праймирующий участок от 3'-концевого второго праймирующего участка с точки зрения событий отжига с нуклеиновой кислотой-мишенью, в силу чего специфичность отжига олигонуклеотида определяется двойственно 5'-концевым первым праймирующим участком и 3'-концевым вторым праймирующим участком, так что общая специфичность отжига данного праймера является повышенной.
Структура DPO в качестве варианта праймера DSO (олигонуклеотида с двойной специфичностью) была впервые предложена авторами настоящего изобретения (см. WO 2006/095981; Chun et al., Dual priming oligonucleotide system for the multiplex detection of respiratory viruses and SNP genotyping of CYP2C19 gene, Nucleic Acid Research, 35: 6e40 (2007)).
DPO является воплощением новой концепции, согласно которой его гибридизация или отжиг двойственно определяется 5'-концевым определяющим специфичность участком с высокой Тпл (или 5'-концевым первым праймирующим участком) и 3'-концевым определяющим специфичность участком с низкой Тпл (или 3'-концевым вторым праймирующим участком), отделенными друг от друга разделительным участком, демонстрируя значительно повышенную специфичность гибридизации (см. WO 2006/095981; Kim et al., Direct detection of lamivudine-resistant hepatitis В virus mutants by multiplex PCR using dual-priming oligonucleotide primers, Journal of Virological Methods, 149:76-84 (2008); Kim, et al., Rapid detection and identification of 12 respiratory viruses using a dual priming oligonucleotide system-based multiplex PCR assay, Journal of Virological Methods, doi: 10.1016/j.jviromet.2008.11.007 (2008); Horii et al., Use of dual priming oligonucleotide system to detect multiplex sexually transmitted pathogens in clinical specimens, Letters in Applied Microbiology, doi:10.111/j.1472-765X2009.02618x (2009)). По сути, DPO содержит в конечном счете два сегмента праймера с различными свойствами в отношении гибридизации: 5'-концевой первый праймирующий участок, который инициирует стабильную гибридизацию, и 3'-концевой второй праймирующий участок, который преимущественно определяет специфичность к мишени.
Амплификация (в частности, множественная амплификация) с использованием праймеров, имеющих структуру DPO, в настоящем изобретении обеспечивает получение ампликонов без ложноположительных и ложноотрицательных результатов.
Согласно предпочтительному воплощению универсальное основание в разделительном участке выбрано из группы, состоящей из дезоксиинозина, инозина, 7-деаза-2'-дезоксиинозина, 2-аза-2'-дезоксиинозина, 2'-ОМе-инозина, 2'-F-инозина, дезокси-3-нитропиррола, 3-нитропиррола, 2'-ОМе-3-нитропиррола, 2'-F-3-нитропиррола, 1-(2'-дезокси-бета-D-рибофуранозил)-3-нитропиррола, дезокси-5-нитроиндола, 5-нитроиндола, 2'-ОМе-5-нитроиндола, 2'-F-5-нитроиндола, дезокси-4-нитробензимидазола, 4-нитробензимидазола, дезокси-4-аминобензимидазола, 4-аминобензимидазола, дезокси-небуларина, 2'-F-небуларина, 2'-F-4-нитробензимидазола, 5-нитроиндола, входящего в состав PNA (пептидо-нуклеиновой кислоты), небуларина, входящего в состав PNA, инозина, входящего в состав PNA, 4-нитробензимидазола, входящего в состав PNA, 3-нитропиррола, входящего в состав PNA, морфолино-5-нитроиндола, морфолино-небуларина, морфолино-инозина, морфолино-4-нитробензимидазола, морфолино-3-нитропиррола, фосфорамидат-5-нитроиндола, фосфорамидат-небуларина, фосфорамидат-инозина, фосфорамидат-4-нитробензимидазола, фосфорамидат-3-нитропиррола, 2'-O-метоксиэтил-инозина, 2'-O-метоксиэтил-небуларина, 2'-O-метоксиэтил-5-нитроиндола, 2'-O-метоксиэтил-4-нитро-бензимидазола, 2'-O-метоксиэтил-3-нитропиррола и их комбинаций. Более предпочтительно, чтобы универсальное основание представляло собой дезоксиинозин, 1-(2'-дезокси-бета-O-рибофуранозил)-3-нитропиррол или 5-нитроиндол, наиболее предпочтительно, дезоксиинозин.
Предпочтительно, чтобы разделительный участок содержал следующие друг за другом нуклеотиды, имеющие по меньшей мере три, более предпочтительно по меньшей мере четыре, наиболее предпочтительно по меньшей мере пять универсальных оснований.
Предпочтительно, чтобы в структуре DPO длина 5'-концевого первого праймирующего участка превышала длину 3'-концевого второго праймирующего участка. Длина 5'-концевого первого праймирующего участка составляет предпочтительно 15-60 нуклеотидов, более предпочтительно 15-40 нуклеотидов, еще более предпочтительно 15-25 нуклеотидов. Предпочтительно, чтобы длина 3'-концевого второго праймирующего участка составляла 3-15 нуклеотидов, более предпочтительно 5-15 нуклеотидов, еще более предпочтительно 6-13 нуклеотидов. Длина разделительного участка составляет предпочтительно 3-10 нуклеотидов, более предпочтительно 4-8 нуклеотидов, наиболее предпочтительно 5-7 нуклеотидов. Согласно предпочтительному воплощению Тпл 5'-концевого первого праймирующего участка находится в диапазоне от 40°С до 80°С, более предпочтительно от 45°С до 65°С. Тпл 3'-концевого второго праймирующего участка предпочтительно находится в диапазоне от 10°С до 40°С. Предпочтительно, чтобы Тпл разделительного участка находилась в диапазоне от 3°С до 15°С.
Согласно предпочтительному воплощению Тпл 3'-концевого первого праймирующего участка находится в диапазоне от 40°С до 80°С, более предпочтительно от 45°С до 65°С. Тпл 5'-концевого второго праймирующего участка предпочтительно находится в диапазоне от 10°С до 40°С. Предпочтительно, чтобы Тпл разделительного участка находилась в диапазоне от 3°С до 15°С.
Согласно предпочтительному воплощению оба эти праймера, используемые для амплификации в настоящем изобретении, имеют структуру DPO.
При использовании традиционных технологий с применением праймеров для детекции нуклеиновой кислоты-мишени нельзя не получить ложных сигналов достаточно высокого уровня ввиду ограничений, присущих используемым праймерам и зондам. Однако праймеры, имеющие структуру DPO с такой обоснованной конструкцией, гибридизуются с нуклеиновокислотной последовательностью-мишенью с существенно повышенной специфичностью, давая возможность детектировать нуклеиновокислотную последовательность-мишень без каких-либо ложных сигналов.
Использованный в данном описании термин "традиционный" по отношению к праймерам означает любой праймер, не имеющий структуры DPO. Они описаны в данном изобретении в качестве традиционных праймеров.
Согласно предпочтительному воплощению одна из молекул, выбранная из репортерной молекулы и молекулы-гасителя, расположена на 5'-концевом первом праймирующем участке, а другая на 5'-концевом первом праймирующем участке, 3'-концевом втором праймирующем участке или разделительном участке.
В настоящем изобретении не требуется, чтобы нуклеиновокислотные последовательности-мишени, подлежащие детекции и/или амплификации, имели какую-либо определенную последовательность или длину.
Если в качестве исходного материала используют мРНК, перед проведением стадии отжига необходима стадия обратной транскрипции, детали которой можно найти в Joseph Sambrook, et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001); и Noonan, K. F. et al., Nucleic Acids Res., 16:10366 (1988)). Для обратной транскрипции можно использовать случайный гексамер или олигонуклеотид dT в качестве праймера, способный гибридизоваться с мРНК.
Олиго-dT-праймер состоит из dTMP, один или более чем один из которых могут быть заменены на другие dNMP, пока этот содержащий dT праймер может еще служить в качестве праймера. Обратную транскрипцию можно осуществить с использованием обратной транскриптазы, обладающей активностью РНКазы Н. Если используется фермент, обладающий активностью РНКазы Н, можно при тщательном выборе условий реакции избежать отдельной стадии переваривания РНКазой Н.
В частности, нуклеиновокислотные последовательности-мишени, которые могут подлежать детекции и/или амплификации, включают любую прокариотическую, эукариотическую (например, из простейших и паразитов, грибов, дрожжей, высших растений, низших и высших животных, в том числе млекопитающих и человека), вирусную (например, из вируса герпеса, ВИЧ, вируса гриппа, вируса Эпштейна-Барр, вируса гепатита, полиовируса и т.д.) или вироидную нуклеиновую кислоту природного происхождения.
Преимущества настоящего изобретения могут быть отмечены при одновременной (множественной) детекции по меньшей мере двух нуклеиновокислотных последовательностей-мишеней.
Согласно предпочтительному воплощению нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) нуклеиновокислотных последовательностей, и TSG-праймеры включают по меньшей мере два типа (более предпочтительно по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров. Согласно предпочтительному воплощению взаимодополняющий праймер, способный амплифицировать нуклеиновокислотную последовательность-мишень вместе с TSG-праймером, включает по меньшей мере два типа (более предпочтительно, по меньшей мере три типа, еще более предпочтительно по меньшей мере пять типов) праймеров.
Например, если настоящее изобретение осуществляют с использованием реакционного сосуда, содержащего пять TSG-праймеров (каждый из которых содержит флуоресцентную репортерную молекулу, характеризующуюся своей длиной волны излучения), пять взаимодополняющих праймеров и образец нуклеиновой кислоты, то генерируется пять разных сигналов флуоресценции, соответствующих пяти разным нуклеиновым кислотам-мишеням, что дает возможность одновременно детектировать пять разных нуклеиновокислотных последовательностей-мишеней в режиме реального времени. В этом случае все используемые молекулы-гасители могут быть выбраны такими, чтобы они отличались по свойствам друг от друга. Альтернативно, все используемые молекулы-гасители или некоторые из них могут быть выбраны такими, чтобы они имели одинаковые свойства.
Кроме того, настоящее изобретение является очень полезным для детекции вариабельности нуклеотидов. Термин "вариабельность нуклеотидов", используемый в данном описании, относится к нуклеотидному полиморфизму в последовательности ДНК в конкретном положении среди непрерывных сегментов ДНК, которые в других случаях являются одинаковыми в последовательности. Такие непрерывные сегменты ДНК включают ген или любую другую часть хромосомы. Например, вариабельность нуклеотидов, детектируемая в настоящем изобретении, включает SNP (однонуклеотидный полиморфизм), делецию, вставку, замену и транслокацию. Примером вариабельности нуклеотидов являются многочисленные вариации в геноме человека (например, вариации в гене MTHFR (метилентетрагидрофолат-редуктазы)), вариации, вовлеченные в возникновение лекарственной устойчивости у патогенных микроорганизмов, и приводящие к онкогенезу вариации.
Согласно предпочтительному воплощению, нуклеиновокислотная последовательность-мишень, используемая в настоящем изобретении, представляет собой предварительно амплифицированную нуклеиновокислотную последовательность. Использование предварительно амплифицированной нуклеиновокислотной последовательности позволяет значительно повысить чувствительность и специфичность способа детекции мишени по настоящему изобретению. Предварительно амплифицируют небольшое количество нуклеотидной последовательности-мишени, получая подходящее количество, и затем осуществляют детекцию способом по настоящему изобретению, повышая чувствительность и специфичность способа детекции мишени по настоящему изобретению. Интересно, что если в способе по настоящему изобретению применяют TSG-праймер, подлежащий гибридизации с последовательностью, расположенной "вниз по течению" по отношению к праймерам, используемым для предварительной амплификации, то он служит в качестве "гнездового" праймера для повышения специфичности способа детекции мишени по настоящему I изобретению.
Согласно предпочтительному воплощению, если в настоящем изобретении для детекции нуклеиновокислотных последовательностей-мишеней с применением реакции амплификации используют предварительно амплифицированную нуклеиновокислотную последовательность, то пара праймеров, используемых в настоящем изобретении, представляет собой пару праймеров для "гнездовой" амплификации.
В другом аспекте данного изобретения предложен набор для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера), содержащий:
(а) TSG-праймер, содержащий (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени; и
(b) матричную полимеразу нуклеиновых кислот, способную индуцировать реакцию 3'-удлинения по 3'-концу TSG-праймера в результате действия на продукт гибридизации TSG-праймера и нуклеиновокислотной последовательности-мишени.
Поскольку набор по данному изобретению составлен для выполнения описанного выше способа детекции по настоящему изобретению, их общее описание опущено, чтобы избежать излишнего дублирования, приводящего к усложнению данного описания.
Наборы по настоящему изобретению возможно могут содержать реагенты, необходимые для проведения амплификации мишени с использованием реакций ПЦР (например, ПЦР-реакций), такие как буферы, кофакторы ДНК-полимеразы и дезоксирибонуклеотид-5-трифосфаты. Возможно, что наборы также могут содержать молекулы различных полинуклеотидов, обратную транскриптазу, различные буферы и реагенты, и антитела, ингибирующие ДНК-полимеразную активность. Наборы также могут содержать реагенты, необходимые для проведения реакций положительного и отрицательного контроля.
Данные наборы также могут включать реагенты, необходимые для выполнения реакций положительного и отрицательного контроля. Оптимальные количества реагентов, которые будут использоваться в заданной реакции, могут быть легко определены специалистом на основе информации, содержащейся в данном описании. Эти наборы обычно устроены так, чтобы содержать описанные выше компоненты в разных упаковках или отделах).
Признаки и преимущества настоящего изобретения будут обобщены ниже.
(a) Настоящее изобретение относится к новому способу ПЦР в режиме реального времени для детекции нуклеиновокислотной последовательности-мишени. TSG-праймер способен генерировать сигналы, указывающие на присутствие нуклеиновокислотной последовательности-мишени, от системы двух взаимодействующих меток и амплифицировать нуклеиновокислотную последовательность-мишень путем его 3'-удлинения во время ПЦР-реакций. Таким образом, стало очевидно, что согласно настоящему изобретению может быть предложен новый способ детекции нуклеиновокислотных последовательностей-мишеней одновременно с амплификацией мишени посредством ПЦР в режиме реального времени.
(b) Реакция удлинения TSG-праймера дает возможность инкорпорировать данный TSG-праймер с двумя метками в продукт удлинения, усиливая ввиду этого интенсивность сигнала соответственно количеству амплифицированного продукта. Такая одновременная стратегия, принятая в настоящем изобретении, позволяет осуществлять более точный количественный анализ нуклеиновокислотных последовательностей-мишеней.
(c) В отличие от меченых зондов, когда сам TSG-праймер неспецифически гибридизуется с последовательностями, не являющимися мишенями, без какого-либо удлинения праймера, невозможна генерация ложноположительных сигналов при осуществлении детекции сигнала в условиях высокой жесткости, таких как высокая температура.
(d) Генерация сигнала в настоящем изобретении может быть выполнена только в результате гибридизации с нуклеиновокислотными последовательностями-мишенями без осуществления реакций расщепления под действием нуклеаз. В этой связи настоящее изобретение не требует, чтобы полимеразы нуклеиновых кислот обязательно обладали 5'→3'-нуклеазной активностью или 3'→5'-нуклеазной активностью, и таким образом дает возможность применения широкого ряда полимераз нуклеиновых кислот для разнообразных приложений.
(e) При использовании полимераз нуклеиновых кислот, обладающих 5'→3'-нуклеазной активностью, согласно настоящему изобретению можно получать сигналы в результате реакции 5'-расщепления TSG-праймера, гибридизованного с нуклеиновокислотными последовательностями-мишенями, увеличивая эффективность детекции мишени.
(f) Традиционные способы ПЦР в режиме реального времени требуют применения меченых зондов или праймеров со сложно модифицированной структурой, такой как шпилечная структура, что затрудняет конструирование, синтез или выбор последовательности зонда и праймера. Однако, поскольку TSG-праймер по настоящему изобретению используют не только для амплификации мишени, но также и для амплификации сигнала без дополнительных меченых зондов или праймеров со сложно модифицированной структурой, его конструирование, синтез и выбор последовательности для ПЦР в режиме реального времени являются очень простыми и легкими. Следовательно, согласно настоящему изобретению предложен новый способ ПЦР в режиме реального времени, который может быть исполнен гораздо более удобным образом, чем традиционные технологии ПЦР в режиме реального времени.
(g) Оптимизация традиционных способов ПЦР в режиме реального времени, основанных на применении зондов, затруднена ввиду необходимости оптимизации условий гибридизации как для зондов, так и для праймеров. Для оптимизации реакционных условий с учетом образования и изменения формы петель-"шпилек" в праймерах предложены традиционные способы ПЦР в режиме реального времени с использованием снабженных "хвостом" праймеров для образования петель-шпилек". В противоположность этому в настоящем изобретении могут быть полностью сняты проблемные вопросы и недостатки, связанные с оптимизацией таких традиционных способов ПЦР в режиме реального времени, поскольку его оптимизация может быть проведена только в отношении праймеров без какой-либо структурной модификации.
(h) Традиционные способы ПЦР в режиме реального времени с малой долей вероятности применимы для множественного анализа ввиду наличия трудностей в конструировании и оптимизации праймеров или зондов. В противоположность этому, поскольку в настоящем изобретении в ПЦР в режиме реального времени используется только меченый праймер без дополнительных зондов или праймера со сложно модифицированной структурой, это дает возможность осуществлять превосходную детекцию мишени в режиме реального времени множественным способом.
(i) По сравнению с традиционными зондами для ПЦР в режиме реального времени TSG-праймер удлиняется в ходе процесса, и удлиненный TSG-праймер сильнее связывается с нуклеиновокислотными последовательностями-мишенями. Для традиционных праймеров для ПЦР в режиме реального времени требуется сложно модифицированная структура, такая как петля-"шпилька", что затрудняет связывание с нуклеиновокислотной последовательностью-мишенью. В противоположность этому, для TSG-праймера таких модификаций не требуется, поскольку TSG-праймер обладает лучшей эффективностью связывания с нуклеиновокислотными последовательностями-мишенями. Этот признак отчасти обуславливает повышенную эффективность детекции мишени в способе по настоящему изобретению.
(j) В способе по настоящему изобретению легко может быть реализована ПЦР-реакция в режиме реального времени путем простого введения метки в праймеры, сконструированные для использования в традиционных ПЦР-реакциях, и применения этих меченых праймеров в качестве TSG-праймеров для данной ПЦР-реакции в режиме реального времени. Кратко, в праймеры, используемые для генерации ампликонов в традиционных ПЦР-реакциях, легко вводят подходящие метки и затем их используют для детекции нуклеиновокислотных последовательностей-мишеней в настоящем изобретении в ПЦР-реакциях в режиме реального времени. В этом отношении способ по настоящему изобретению считается более эффективным во временном и стоимостном выражении для разработки анализа с использованием ПЦР в режиме реального времени.
(k) Как рассмотрено выше, используемые в настоящем изобретении праймеры, имеющие структуру DPO, приводят к улучшению специфичности их связывания, благодаря чему устраняются ложноположительные сигналы, обусловленные нецелевым связыванием праймеров в ПЦР-реакциях в режиме реального времени.
Теперь настоящее изобретение будет описано далее подробно посредством примеров. Специалистам в данной области будет очевидно, что эти примеры предназначены для более конкретной иллюстрации изобретения и объем настоящего изобретения, который изложен в прилагаемой формуле изобретения, не ограничивается данными примерами.
ПРИМЕРЫ
ПРИМЕР 1. Оценка применимости TSG-праймера с ДНК-полимеразой, не обладающей 5'→3'-экзонуклеазной активностью, в детекции нуклеиновокислотных последовательностей-мишеней
TSG-праймер по данному изобретению оценивали в отношении того, можно ли с помощью TSG-праймера генерировать сигнал, достаточный для детекции нуклеиновокислотной последовательности-мишени только в результате его гибридизации с мишенью и удлинения, при котором используется матричная полимераза нуклеиновых кислот без 5'→3'-экзонуклеазной активности.
Для проведения этой оценки авторы изобретения в качестве матриц-мишеней использовали ген Streptococcus pneumoniae (SP) или ген Staphylococcus aureus (SA). Для удобства эксперимента в качестве матриц для гена S. pneumoniae и гена S. aureus использовали синтетические олигонуклеотиды. В качестве ДНК-полимеразы использовали фрагмент Штоффеля, не обладающий собственной 5'→3'-экзонуклеазной активностью. Два TSG-праймера, характеризующихся разными расстояниями между репортерной молекулой и молекулой-гасителем, проверяли соответственно для каждой мишени. В качестве флуоресцентной репортерной молекулы, расположенной на 5'-конце TSG-праймеров, использовали 6-FAM (6-карбоксифлуоресцеин). В качестве молекулы-гасителя использовали гаситель Black Hole 1 (BHQ-1). Реакцию синтеза нуклеиновой кислоты проводили без повторения стадий денатурации, гибридизации и удлинения праймеров. Сигналы измеряли в предварительно определенные интервалы времени.
А. Реакция синтеза нуклеиновой кислоты для детекции гена S. pneumoniae
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень гена S. pneumoniae, последовательности синтетической матрицы и TSG-праймеров, использованных в этом Примере, представляют собой:
(Число 9 или 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
Реакцию синтеза нуклеиновой кислоты с TSG-праймером проводили в конечном объеме 20 мкл, содержащем 2 пмоль матрицы (SEQ ID NO:1), 2 мкл 10х буфера Штоффеля [содержащего 100 мМ Трис-HCl (рН 8,3) и ЮОмМ KCl], 1 единицу ДНК-полимеразы AmpliTaq®, фрагмент Штоффеля (Applied BioSystems, USA), каждый из четырех dNTP (дезоксинуклеозид-трифосфат) (dATP (дезоксиаденозин-трифосфат), dCTP (дезоксицитидин-трифосфат), dGTP (дезоксигуанозин-трифосфат) и dTTP (дезокситимидин-трифосфат)) в концентрации 200 мкМ, 5 мМ MgCl2 и 5 пмоль TSG-праймера (SEQ ID NO:2 или 3); пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и инкубации в течение 40 мин при 50°С. Детекцию генерированного сигнала осуществляли с интервалом 1 мин.
Как представлено на ФИГ.5, каждый из двух TSG-праймеров демонстрировал более высокую интенсивность флуоресценции в присутствии матрицы (NO:1 и 3), чем в отсутствие матрицы (NO:2 и 4). Таким образом, стало ясно, что с помощью TSG-праймеров можно получить сигналы, достаточные для детекции гена S. pneumonia, посредством гибридизации и удлинения TSG-праймеров в процессе реакции синтеза нуклеиновой кислоты.
Примечательно, что TSG-праймер (SEQ ID NO:3), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя, демонстрировал более отчетливые изменения интенсивности сигнала (то есть изменение в величинах RFU (относительные единицы флуоресценции)) в присутствии и в отсутствие матрицы, чем TSG-праймер (SEQ ID NO:2), содержащий репортерную молекулу на расстоянии 9 нуклеотидов от молекулы-гасителя.
В. Реакция синтеза нуклеиновой кислоты для детекции гена S. aureus
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень гена S. aureus, последовательности синтетической матрицы и TSG-праймеров, использованных в этом Примере, представляют собой:
(Число 6 или 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
Реакцию синтеза нуклеиновой кислоты проводили в соответствии с протоколом, использованным для S. pneumoniae, за исключением условий для матрицы (0,2 пмоль S. aureus) и TSG-праймеров (SEQ ID NO:5 или 6).
Как представлено на ФИГ.6, каждый из двух TSG-праймеров демонстрировал более высокую интенсивность флуоресценции в присутствии матрицы (NO:1 и 3), чем в отсутствие матрицы (NO:2 и 4). Таким образом, стало ясно, что с помощью TSG-праймеров можно получить сигналы, достаточные для детекции гена S. aureus, посредством гибридизации и удлинения TSG-праймеров в процессе реакции синтеза нуклеиновой кислоты.
Примечательно, что TSG-праймер (SEQ ID NO:6), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя, демонстрировал более отчетливые изменения интенсивности сигнала (то есть изменение в величинах RFU) в присутствии и в отсутствие матрицы, чем TSG-праймер (SEQ ID NO:5), содержащий репортерную молекулу на расстоянии 6 нуклеотидов от молекулы-гасителя.
ПРИМЕР 2. Проверка применимости TSG-праймера с ДНК-полимеразой, не обладающей 5'→3'-экзонуклеазной активностью, для детекции нуклеиновой кислоты-мишени в условиях ПЦР-рекции в режиме реального времени
Далее авторы изобретения проверяли, можно ли с помощью TSG-праймера генерировать сигнал, достаточный для детекции нуклеиновокислотной последовательности-мишени во время проведения ПЦР-реакции в режиме реального времени с использованием матричной полимеразы нуклеиновых кислот, не обладающей 5'→3'-экзонуклеазной активностью.
Для осуществления этой проверки ПЦР-реакции в режиме реального времени для детекции генов S. pneumoniae, S. aureus, Neisseria gonorrhoeae и Neisseria meningitidis проводили соответственно с использованием пар праймеров, включая TSG-праймеры. Фрагмент Штоффеля, не обладающий собственной 5'→3'-экзонуклеазной активностью, использовали в качестве ДНК-полимеразы. Для каждого гена-мишени проверяли соответственно два TSG-праймера, характеризующихся разными расстояниями между репортерной молекулой и молекулой-гасителем.
А. ПНР в режиме реального времени для детекции гена S. pneumoniae
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень гена S. pneumoniae, последовательности прямого праймера и TSG-праймеров (в качестве обратных праймеров), использованных в этом Примере, представляют собой:
(I представляет собой дезоксиинозин, а число 9 или 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
ПЦР-реакцию в режиме реального времени для детекции S. pneumoniae проводили в конечном объеме 20 мкл, содержащем Юнг геномной ДНК S. pneumoniae, 2 мкл 10× буфера Штоффеля, содержащего 100 мМ Трис-HCl (рН 8,3) и 100 мМ KCl, 1 единицу ДНК-полимеразы AmpliTaq®, фрагмент Штоффеля (Applied BioSystems, USA), каждый из четырех dNTP (dATP, dCTP, dGTP и dTTP) в концентрации 200 мкМ, 5 мМ MgCl2, 5 пмоль прямого праймера (SEQ ID NO:7) и 5 пмоль TSG-праймера (SEQ ID NO:2 или 3) в качестве обратного праймера; пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 30 циклам по 30 с при 94°С, 60 с при 55°С и 10 с при 72°С. Детекцию генерированного сигнала осуществляли на стадии отжига (55°С) каждого цикла.
Как показано на Фиг.7, флуоресцентные сигналы для S. pneumoniae наблюдали в присутствии матрицы S. pneumoniae (NO:1 и 3) в ПЦР-реакции в режиме реального времени с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-нуклеазной активностью, в то время как в отрицательных контролях без матрицы-мишени (NO:2 и 4) не было никакого флуоресцентного сигнала.
TSG-праймер (SEQ ID NO:3), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:2), содержащий репортерную молекулу на расстоянии 9 нуклеотидов от молекулы-гасителя.
В. ПЦР в режиме реального времени для детекции гена S. aureus
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень S. aureus, последовательности прямого праймера и TSG-праймеров (в качестве обратных праймеров), использованных в этом Примере, представляют собой:
(I представляет собой дезоксиинозин, а число 6 или 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
ПЦР-реакцию проводили в соответствии с протоколом, использованным для S. pneumoniae, за исключением применения других матрицы (S. aureus), прямого праймера (SEQ ID NO:8) и TSG-праймеров (SEQ ID NO:5 или 6) в качестве обратных праймеров.
Как показано на Фиг.8, флуоресцентные сигналы для S. aureus наблюдали в присутствии матрицы S. aureus (NO:1 и 3) в ПЦР-реакции в режиме реального времени с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-нуклеазной активностью, в то время как в отрицательных контролях без матрицы-мишени не было никакого флуоресцентного сигнала (NO:2 и 4).
TSG-праймер (SEQ ID NO:6), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:5), содержащий репортерную молекулу на расстоянии 6 нуклеотидов от молекулы-гасителя.
С. ПЦР в режиме реального времени для детекции гена N. gonorrhoeae
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень N. gonorrhoeae (NG), последовательности прямого праймера и TSG-праймеров (в качестве обратных праймеров), использованных в этом Примере, представляют собой:
(I представляет собой дезоксиинозин, а число 4 или 22 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
ПЦР-реакцию проводили в соответствии с протоколом, использованным для S. pneumoniae, за исключением использования других матрицы (N. gonorrhoeae), прямого праймера (SEQ ID NO:9) и TSG-праймеров (SEQ ID NO:10 или 11) в качестве обратных праймеров.
Как показано на Фиг.9, флуоресцентные сигналы для N. gonorrhoeae наблюдали в присутствии матрицы N. gonorrhoeae (NO:1 и 3) в ПЦР-реакции в режиме реального времени с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-нуклеазной активностью, в то время как в отрицательных контролях без матрицы-мишени не было никакого флуоресцентного сигнала (NO:2 и 4).
Кроме того, TSG-праймер (SEQ ID NO:11), содержащий репортерную молекулу на расстоянии 22 нуклеотидов от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:10), содержащий репортерную молекулу на расстоянии 4 нуклеотидов от молекулы-гасителя, как показано на Фиг.9.
D. ПЦР в режиме реального времени для детекции гена N. meningitidis
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень N. meningitides (NM), последовательности обратного праймера и TSG-праймеров (в качестве прямых праймеров), использованных в этом Примере, представляют собой:
(I представляет собой дезоксиинозин, а число 7 или 20 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
ПЦР-реакцию проводили в соответствии с протоколом, использованным для S. pneumoniae, за исключением использования других матрицы (N. meningitidis), обратного праймера (SEQ ID NO:12) и TSG-праймеров (SEQ ID NO:13 или 14) в качестве прямых праймеров.
Как показано на Фиг.10, флуоресцентные сигналы для N. meningitidis наблюдали в присутствии матрицы N. meningitidis (NO:1 и 3) в ПЦР-реакции в режиме реального времени с использованием TSG-праймеров и матричной ДНК-полимеразы, не обладающей 5'→3'-нуклеазной активностью, в то время как в отрицательных контролях без матрицы-мишени не было никакого флуоресцентного сигнала (NO:2 и 4).
Кроме того, TSG-праймер (SEQ ID NO:14), содержащий репортерную молекулу на расстоянии 20 нуклеотидов от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:13), содержащий репортерную молекулу на расстоянии 7 нуклеотидов от молекулы-гасителя, как показано на Фиг.10.
ПРИМЕР 3. Чувствительность ПЦР в режиме реального времени для детекции S. aureus с использованием TSG-праймера и ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью
Чувствительность ПЦР в режиме реального времени тестировали, используя TSG-праймер и ДНК-полимеразу, не обладающую 5'→3'-экзонуклеазной активностью, путем детекции нуклеиновокислотных последовательностей-мишеней гена S. aureus. В этом исследовании в ПЦР-реакции в режиме реального времени использовали TSG-праймер (SEQ ID NO:6) в качестве обратного праймера. В качестве матрицы использовали приготовленную серийным разведением (10-кратное разведение) геномную ДНК S. aureus в количестве от 100 пг до 10 фг.
В этом Примере используют следующие последовательности прямого праймера и TSG-праймера (в качестве обратного праймера):
(I представляет собой дезоксиинозин, а число 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
ПЦР-реакцию в режиме реального времени проводили в конечном объеме 20 мкл, содержащем разведенную геномную ДНК (от 100 пг до 10 фг; 10-кратное разведение) S. aureus, 2 мкл 10× буфера Штоффеля, содержащего 100 мМ Трис-HCl (рН 8,3) и 100 мМ KCl, 1 единицу ДНК-полимеразы AmpliTaq®, фрагмент Штоффеля (Applied BioSystems, USA), каждый из четырех dNTP (dATP, dCTP, dGTP и dTTP) в концентрации 200 мкМ, 5 мМ MgCl2, 5 пмоль прямого праймера (SEQ ID NO:8) и 5 пмоль TSG-праймера (SEQ ID NO:6) в качестве обратного праймера; пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 30 с при 94°С, 60 с при 55°С и 10 с при 72°С. Детекцию генерированного сигнала осуществляли на стадии отжига (55°С) каждого цикла.
Как показано на ФИГ.11, когда ПЦР в режиме реального времени проводили с использованием серийного разведения геномной ДНК S. aureus, как описано на Фиг.11, можно было осуществить детекцию нуклеиновокислотной последовательности-мишени в количестве до 100 фг включительно (NO:1-4).
ПРИМЕР 4. Генерация сигнала в результате реакции 5'-расщепления на 5'-концевом участке TSG-праймера под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-экзонуклеазной активностью
Авторы изобретения обнаружили, что меченые праймеры, гибридизованные с нуклеиновокислотными последовательностями-мишенями, подвергаются реакции 5'-расщепления на своем 5'-концевом участке под действием 5'→3'-нуклеазной активности матричной полимеразы нуклеиновых кислот, и эту реакцию 5'-расщепления сложно адаптировать для детекции нуклеиновокислотных последовательностей-мишеней посредством генерации сигналов для последовательностей-мишеней (см. PCT/KR2009/007064).
По этой причине авторы изобретения проверили, генерируются ли сигналы в результате реакции 5'-расщепления на 5'-концевом участке TSG-праймера под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-экзонуклеазной активностью.
Для осуществления этой проверки авторы изобретения использовали S. pneumoniae в качестве матрицы-мишени и для удобства эксперимента в качестве матрицы для гена S. pneumoniae использовали синтетический олигонуклеотид. Соответственно проверяли два TSG-праймера, характеризующихся разными расстояниями между репортерной молекулой и молекулой-гасителем.
В качестве флуоресцентной репортерной молекулы, расположенной на 5'-конце TSG-праймеров, использовали 6-FAM (6-карбоксифлуоресцеин). В качестве молекулы-гасителя использовали гаситель Black Hole 1 (BHQ-1). Реакцию синтеза нуклеиновой кислоты проводили с повторением стадий денатурации, гибридизации и удлинения праймеров. Сигналы измеряли на стадии гибридизации для каждого повторения. Фрагмент Штоффеля, не обладающий собственной 5'→3'-экзонуклеазной активностью, использовали в качестве ДНК-полимеразы.
В этом Примере используют следующие последовательности синтетической матрицы и TSG-праймера (в качестве обратного праймера):
(Число 9 или 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
А. Реакция синтеза нуклеиновой кислоты с повторением стадий денатурации, гибридизации и удлинения праймеров с использованием ДНК-полимеразы, не обладающей 5'→3'-экзонуклеазной активностью
Реакцию синтеза нуклеиновой кислоты проводили в конечном объеме 20 мкл, содержащем 2 пмоль матрицы (SEQ ID NO:1), 2 мкл 10× буфера Штоффеля [содержащего 100 мМ Трис-HCl (рН 8,3) и 100 мМ KCl], 1 единицу ДНК-полимеразы AmpliTaq®, фрагмент Штоффеля (Applied BioSystems, USA), каждый из четырех dNTP (dATP, dCTP, dGTP и dTTP) в концентрации 200 мкМ, 5 мМ MgCl2 и 5 пмоль TSG-праймера (SEQ ID NO:2 или 3); пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 2 мин при 95°С и подвергали 40 циклам по 30 с при 94°С и 60 с при 50°С. Детекцию генерированного сигнала осуществляли на стадии отжига (50°С) каждого цикла.
На Фиг.12, отображающем результаты после нормирования, изменений сигнала не наблюдали даже в присутствии матрицы (NO:1 и 5), когда в качестве ДНК-полимеразы использовали фрагмент Штоффеля, не обладающий собственной 5'→3'-экзонуклеазной активностью.
Эти результаты указывают на то, что интенсивность сигнала, генерированного только в результате гибридизации и удлинения праймера, не изменялась на протяжении циклов реакции, поскольку матрица не амплифицировалась в процессе реакции синтеза нуклеиновой кислоты.
Ввиду этого следует признать, что только гибридизации и удлинения TSG-праймера в реакциях синтеза нуклеиновой кислоты циклами недостаточно, чтобы аккумулировать детектируемые сигналы даже в присутствии нуклеиновокислотных последовательностей-мишеней (NO:1 и 5).
В. Реакция синтеза нуклеиновой кислоты с повторением стадий денатурации, гибридизации и удлинения праймеров с использованием ДНК-полимеразы. обладающей 5'→3'-экзонуклеазной активностью
Реакцию синтеза нуклеиновой кислоты проводили в конечном объеме 20 мкл, содержащем 2 пмоль матрицы (SEQ ID NO:1), 10 мкл смеси 2х Master Mix для ПЦР DiastarTaq (Solgent, Korea), содержащей [12 мМ MgCl2, буфер для ПЦР DiastarTaq, 2 единицы ДНК-полимеразы DiaStarTaq и смесь dNTP], 5 пмоль TSG-праймера (SEQ ID NO:2 или 3); пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 15 мин при 95°С и подвергали 40 циклам по 30 с при 94°С и 60 с при 50°С. Детекцию генерированного сигнала осуществляли на стадии отжига (50°С) каждого цикла.
Как показано на Фиг.12, изменения сигнала для S. pneumoniae наблюдали (NO:3 и 7), когда в качестве ДНК-полимеразы использовали ДНК-полимеразу Taq, обладающую 5'→3'-экзонуклеазной активностью.
Эти результаты указывают на то, что использование матричной полимеразы, обладающей 5'→3'-экзонуклеазной активностью, дает возможность индуцировать реакцию 5'-расщепления на 5'-концевом участке TSG-праймера. Соответственно, сигналы от меченых фрагментов, высвободившихся из TSG-праймеров, генерируются и аккумулируются в каждом цикле, результатом чего является получение наблюдаемых сигналов, указывающих на присутствие нуклеиновокислотной последовательности-мишени (NO:3 и 7).
Между тем, TSG-праймер (SEQ ID NO:2), содержащий репортерную молекулу на расстоянии 9 нуклеотидов от молекулы-гасителя, демонстрировал более низкие величины Ct, чем TSG-праймер (SEQ ID NO:3), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя. Эти результаты подтверждают, что TSG-праймеры с более коротким расстоянием между репортером и гасителем, демонстрируют более значительные изменения в степени гашения до и после реакции 5'-расщепления по сравнению с праймерами, характеризующимися большим расстоянием между репортером и гасителем.
ПРИМЕР 5. Проверка применимости TSG-праймера с ДНК-полимеразой, обладающей 5'→3'-экзонуклеазной активностью, для детекции нуклеиновой кислоты-мишени в условиях ПЦР-реакции в режиме реального времени
Далее авторы изобретения проверяли, можно ли с помощью TSG-праймера генерировать сигнал, достаточный для детекции нуклеиновокислотной последовательности-мишени во время проведения ПЦР-реакции в режиме реального времени с использованием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-экзонуклеазной активностью.
Для осуществления этой проверки использовали те же матрицы и праймеры, что и в Примере 2, за исключением того, что применяли матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной активностью, и определенные реакционные условия.
А. ПЦР в режиме реального времени для детекции S. pneumoniae
ПЦР в режиме реального времени проводили в конечном объеме 20 мкл, содержащем 10 нг геномной ДНК S. pneumoniae, 10 мкл смеси 2× Master Mix для ПЦР DiastarTaq (Solgent, Korea), содержащей [12 мМ MgCl2, буфер для ПЦР DiastarTaq, 2 единицы ДНК-полимеразы DiaStarTaq и смесь dNTP], 5 пмоль прямого праймера (SEQ ID NO:7) и 5 пмоль TSG-праймера (SEQ ID NO:2 или 3) в качестве обратного праймера; пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 15 мин при 95°С и подвергали 30 циклам по 30 с при 94°С, 60 с при 55°С и 10 с при 72°С. Детекцию генерированного сигнала осуществляли на стадии отжига (55°С) каждого цикла.
Как показано на Фиг.13, флуоресцентные сигналы для S. pneumoniae наблюдали в присутствии матрицы S. pneumoniae (NO:1 и 3), в то время как в отрицательных контролях без матрицы (NO:2 и 4) не было никакого флуоресцентного сигнала.
Кроме того, TSG-праймер (SEQ ID NO:3), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:2), содержащий репортерную молекулу на расстоянии 9 нуклеотидов от молекулы-гасителя.
В. ПЦР в режиме реального времени для детекции S. aureus
ПЦР-реакцию в режиме реального времени проводили в соответствии с протоколом, использованным для детекции S. pneumoniae, за исключением использования других матрицы (S. aureus), прямого праймера (SEQ ID NO:8) и TSG-праймеров (SEQ ID NO:5 или 6) в качестве обратных праймеров.
Как показано на Фиг.14, флуоресцентные сигналы для S. aureus наблюдали в присутствии матрицы S. aureus (NO:1 и 3), в то время как в отрицательных контролях без матрицы (NO:2 и 4) не было никакого флуоресцентного сигнала. Кроме того, TSG-праймер (SEQ ID NO:6), содержащий репортерную молекулу на расстоянии 21 нуклеотида от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:5), содержащий репортерную молекулу на расстоянии 6 нуклеотидов от молекулы-гасителя.
С. ПЦР в режиме реального времени для детекции N. donorrhoeae
ПЦР-реакцию в режиме реального времени проводили в соответствии с протоколом, использованным для детекции S. pneumoniae, за исключением использования других матрицы (N. gonorrhoeae), прямого праймера (SEQ ID NO:9) и TSG-праймеров (SEQ ID NO:10 или 11) в качестве обратных праймеров.
Как показано на Фиг.15, флуоресцентные сигналы для N. gonorrhoeae наблюдали в присутствии матрицы N. gonorrhoeae (NO:1 и 3), в то время как в отрицательных контролях без матрицы (NO:2 и 4) не было никакого флуоресцентного сигнала. Кроме того, TSG-праймер (SEQ ID NO:11), содержащий репортерную молекулу на расстоянии 22 нуклеотида от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:10), содержащий репортерную молекулу на расстоянии 4 нуклеотидов от молекулы-гасителя.
D. ПЦР в режиме реального времени для детекции N. meningitidis
ПЦР-реакцию в режиме реального времени проводили в соответствии с протоколом, использованным для детекции S. pneumoniae, за исключением использования других матрицы (N. meningitidis), обратного праймера (SEQ ID NO:12) и TSG-праймеров (SEQ ID NO:13 или 14) в качестве прямых праймеров.
Как показано на Фиг.16, флуоресцентные сигналы для N. meningitidis наблюдали в присутствии матрицы N. meningitidis (NO:1 и 3), в то время как в отрицательных контролях без матрицы (NO:2 и 4) не было никакого флуоресцентного сигнала. Кроме того, TSG-праймер (SEQ ID NO:14), содержащий репортерную молекулу на расстоянии 20 нуклеотидов от молекулы-гасителя, демонстрировал более низкие величины Ct и более высокие величины RFU, чем TSG-праймер (SEQ ID NO:13), содержащий репортерную молекулу на расстоянии 7 нуклеотидов от молекулы-гасителя.
И в заключение, когда используется матричная ДНК-полимераза, обладающая 5'→3'-нуклеазной активностью, то сигналы, указывающие на присутствие нуклеиновокислотной последовательности-мишени, могут быть представлены двумя различными способами: (1) вследствие генерации сигнала при отсутствии гашения сигнала системой взаимодействующих меток на TSG-праймере, обусловленного конформационным изменением в результате гибридизации с нуклеиновокислотной последовательностью-мишенью; и (2) вследствие генерации сигнала в результате реакции 5'-расщепления на 5'-концевом участке TSG-праймера под действием матричной полимеразы нуклеиновых кислот, обладающей 5'→3'-нуклеазной активностью.
ПРИМЕР 6. Чувствительность ПЦР в режиме реального времени для детекции S. aureus с использованием TSG-праймера и ДНК-полимеразы, обладающей 5'→3'-экзонуклеазной активностью
Чувствительность ПЦР в режиме реального времени тестировали, используя TSG-праймер и ДНК-полимеразу, обладающую 5'→3'-экзонуклеазной активностью, путем детекции нуклеиновокислотных последовательностей-мишеней гена S. aureus. Для осуществления этой проверки использовали те же матрицы и праймеры, что и в Примере 3, за исключением того, что применяли матричную полимеразу нуклеиновых кислот, обладающую 5'→3'-экзонуклеазной активностью, и определенные реакционные условия.
ПЦР в режиме реального времени проводили в конечном объеме 20 мкл, содержащем разведенную геномную ДНК (от 100 пг до 10 фг; 10-кратное разведение) из геномной ДНК S. aureus, 10 мкл смеси 2× Master Mix для множественной ПЦР QuantiTect (Qiagen), содержащей [11 мМ MgCl2, буфер для множественной ПЦР QuantiTect, ДНК-полимеразу HotstartTaq и смесь dNTP], 5 пмоль прямого праймера (SEQ ID NO:8) и 5 пмоль TSG-праймера (SEQ ID NO:6) в качестве обратного праймера; пробирку, содержащую реакционную смесь, помещали в термоциклер для ПЦР в режиме реального времени (CFX96, Bio-Rad); реакционную смесь подвергали денатурации в течение 15 мин при 95°С и подвергали 40 циклам по 30 с при 94°С, 60 с при 55°С и 10 с при 72°С. Детекцию генерированного сигнала осуществляли на стадии отжига (55°С) каждого цикла.
Как показано на ФИГ.17, когда ПЦР в режиме реального времени проводили с использованием серийного разведения геномной ДНК S. aureus, как описано на Фиг.11, можно было осуществить детекцию нуклеиновокислотной последовательности-мишени в количестве до 100 фг включительно (NO:1-4).
ПРИМЕР 7. Оценка применимости TSG-праймера для детекции нуклеиновокислотной последовательности-мишени на чипе
Когда в качестве матрицы используют нуклеиновокислотную последовательность-мишень гена S. pneumoniae, последовательности синтетической матрицы и TSG-праймеров, использованных в этом Примере, представляют собой:
(Число 21 в круглых скобках означает расстояние в нуклеотидах между репортерной молекулой и молекулой-гасителем).
А. Иммобилизация TSG-праймера на пластинке чипа
TSG-праймер (SEQ ID NO:15) растворяли до конечной концентрации 50 мкМ в растворе для точечного нанесения, тип I (Genorama). Растворенный TSG-праймер наносили в виде пятен на стеклянную пластинку (Genorama, Estonia) при комнатной температуре и относительной влажности 70%. Пластинку инкубировали в камере с увлажнением при 37°С в течение 2 часов. Затем пластинку выдерживали в 1%-ном растворе аммиака в течение 10 минут с последующей промывкой дистиллированной водой при комнатной температуре.
В. Реакция синтеза нуклеиновой кислоты на чипе
Реакцию синтеза нуклеиновой кислоты проводили в конечном объеме 20 мкл, содержащем 2 пмоль матрицы (SEQ ID NO:1), 2 мкл 10× буфера Штоффеля [содержащего 100 мМ Трис-HCl (рН 8,3) и 100 мМ KCl], 1 единицу ДНК-полимеразы AmpliTaq®, фрагмент Штоффеля (Applied BioSystems, USA), каждый из четырех dNTP (dATP, dCTP, dGTP и dTTP) в концентрации 200 мкМ, 5 мМ MgCl2; реакционную смесь переносили на пластинку. Пластинку помещали in situ в установку для ПЦР (GeneAmp in situ, Perkin Elmer); пластинку инкубировали в течение 2 мин при 95°С для осуществления денатурации и выдерживали в течение 40 мин при 50°С. После проведения реакции синтеза нуклеиновой кислоты пластинку промывали (при 70°С) и детекцию сигналов с пластинки осуществляли, используя сканер для микрочипов (ScanArray4000, Perkin Elmer), с последующим анализом изображений.
Основываясь на описании предпочтительного воплощения настоящего изобретения, следует понимать, что его варианты и модификации, соответствующие сущности изобретения, могут стать очевидными специалистам в данной области, и объем данного изобретения следует определять прилагаемой формулой изобретения и ее эквивалентами.








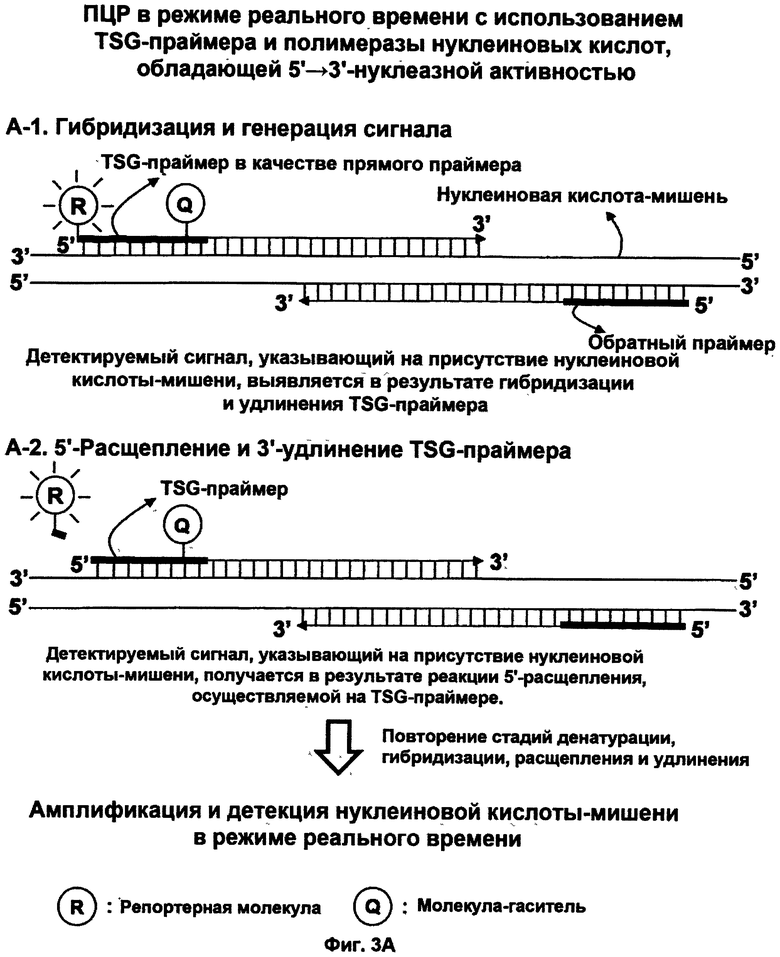

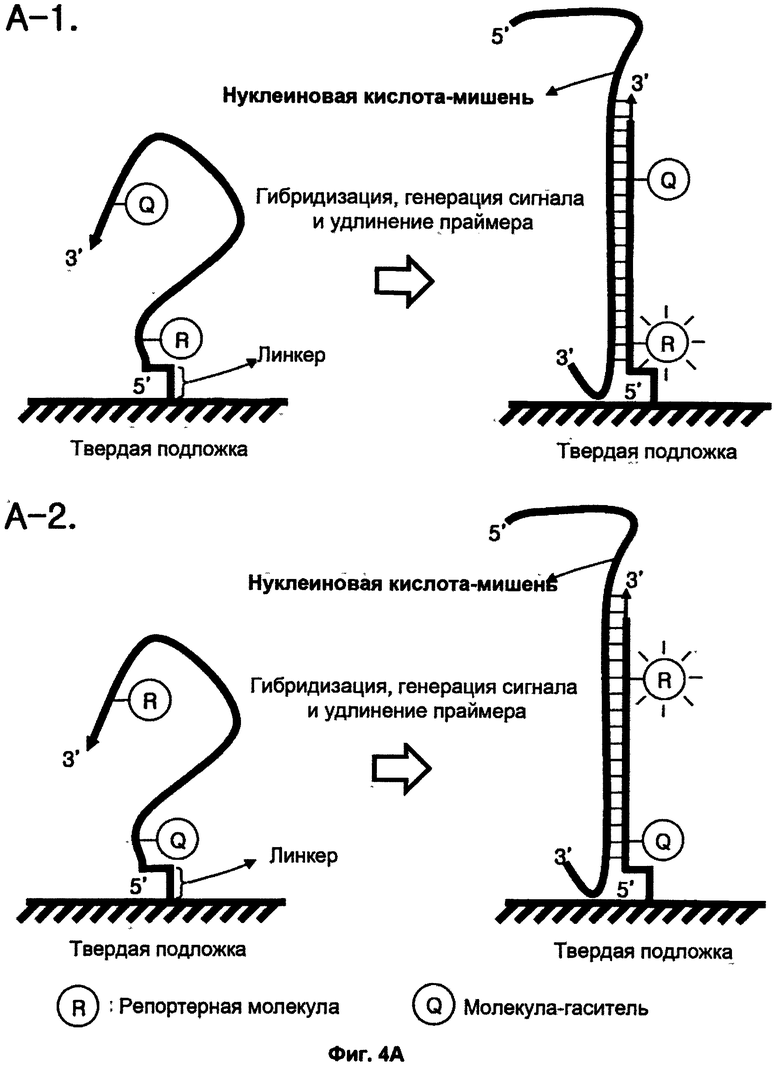

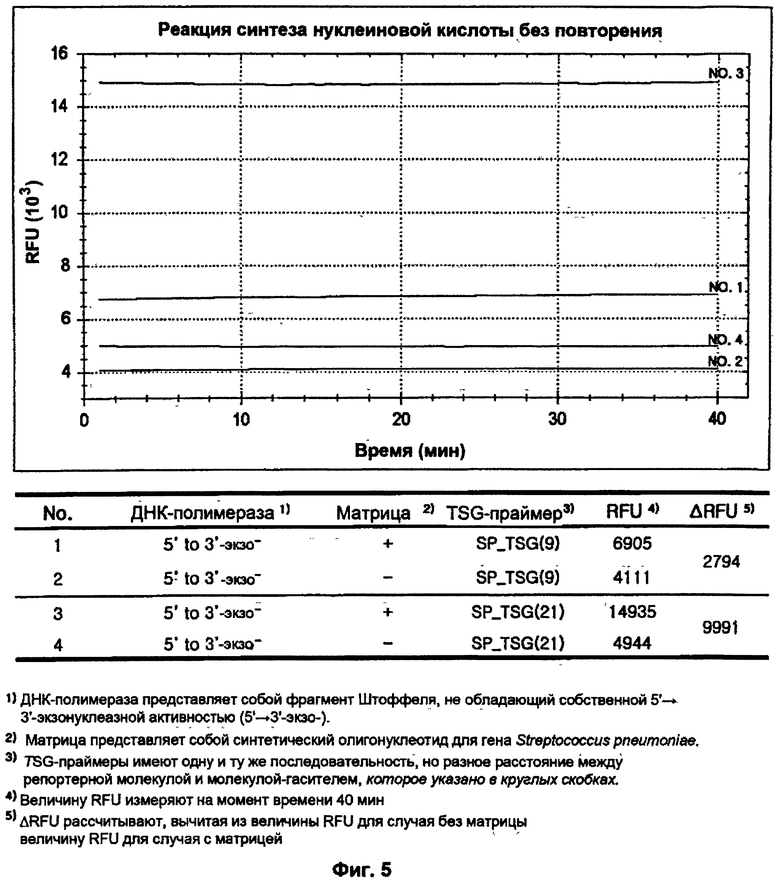
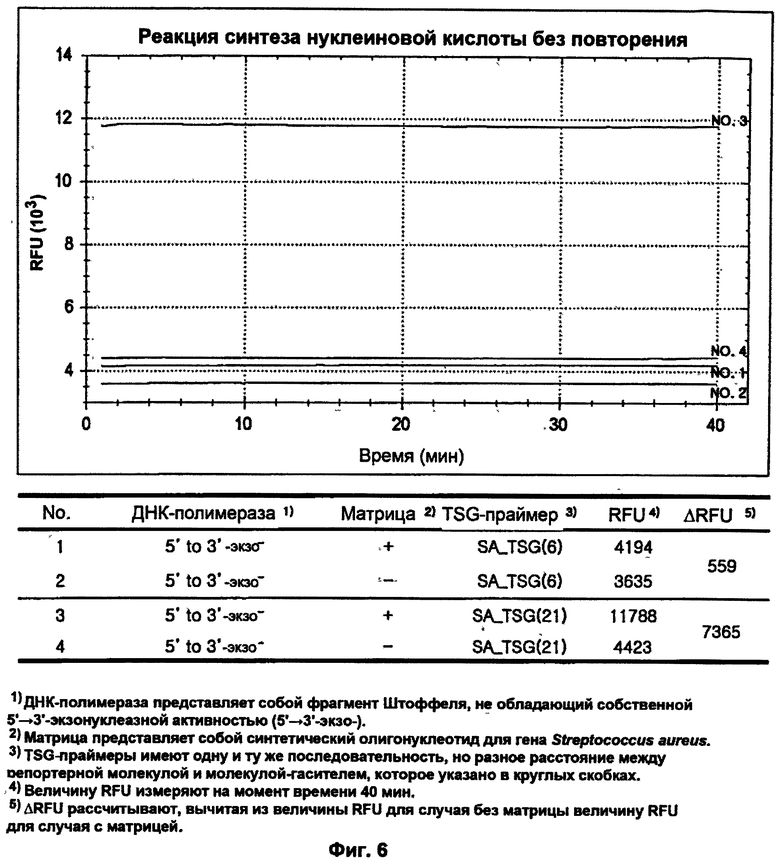
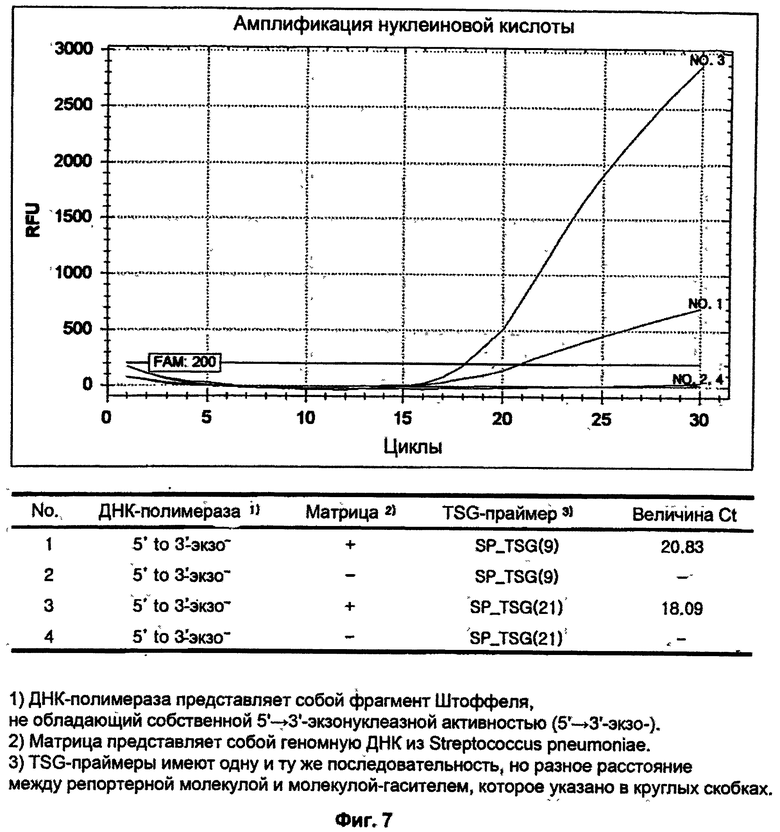
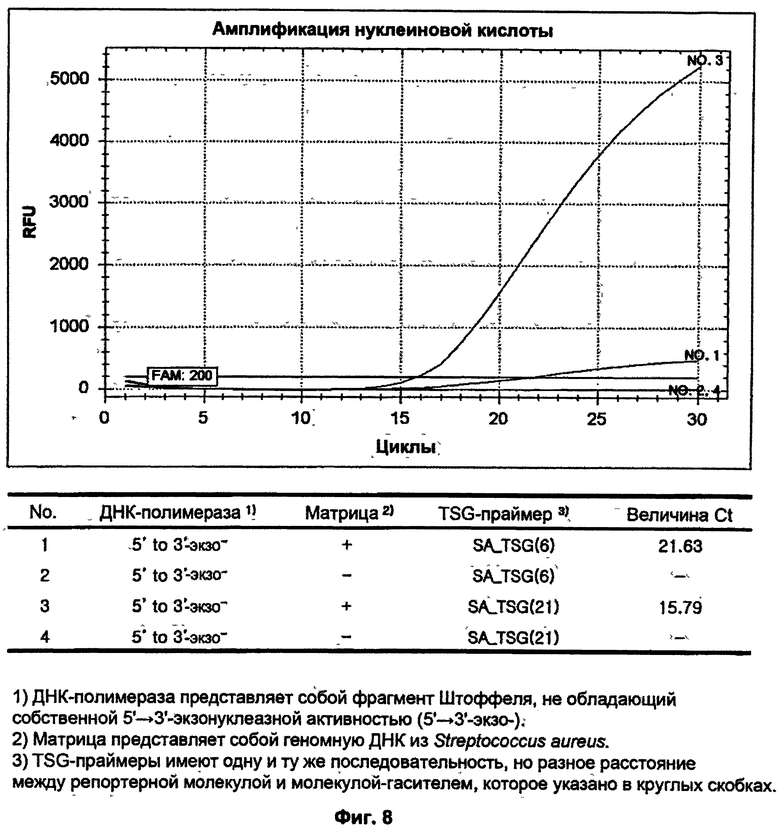

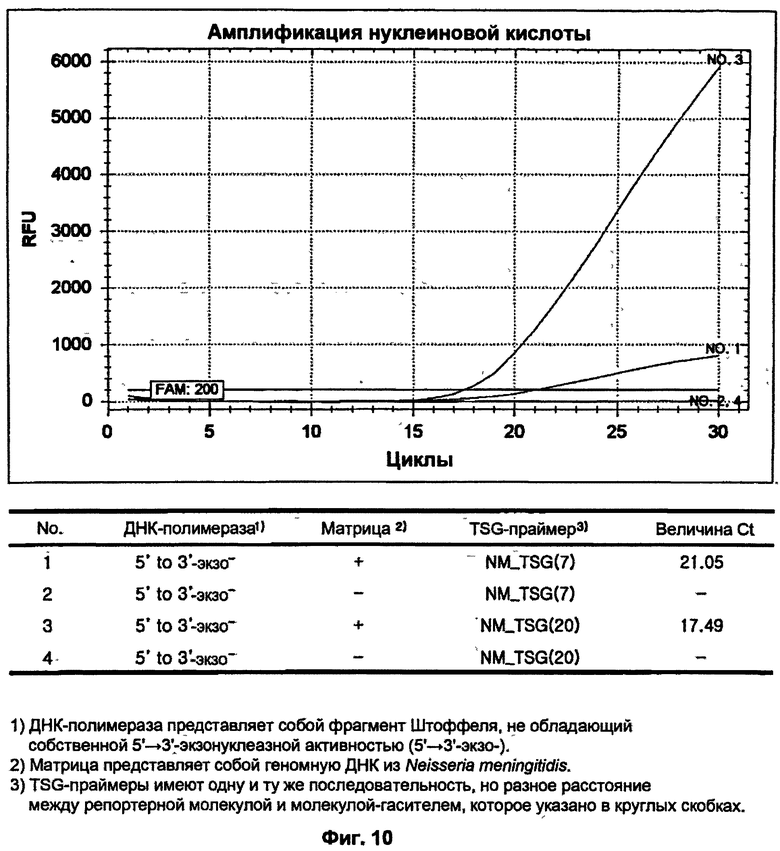
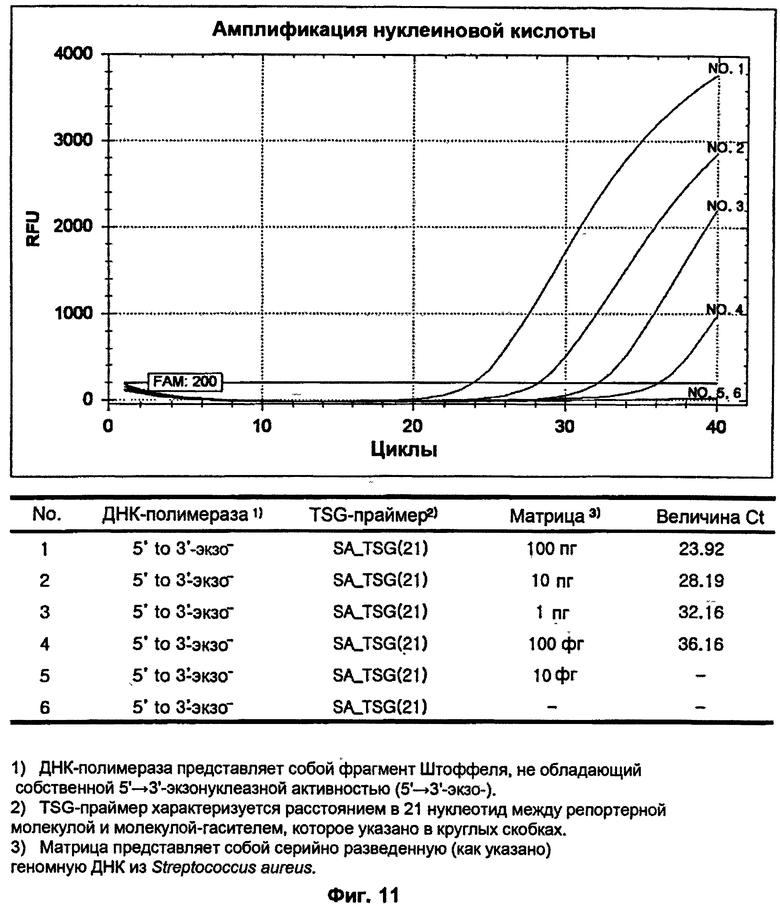
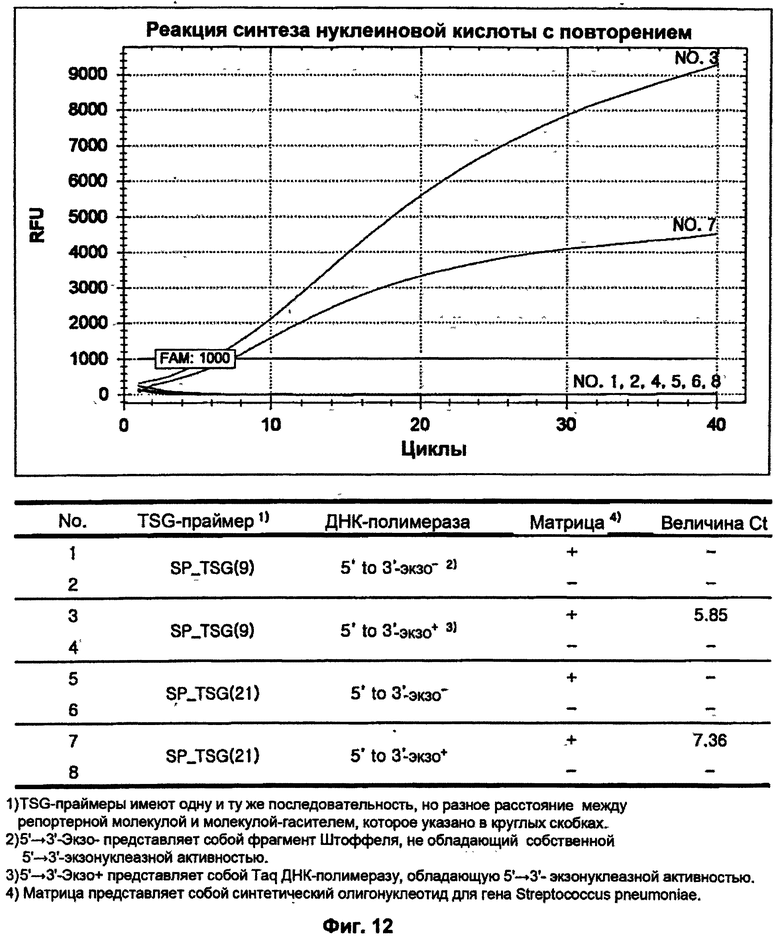
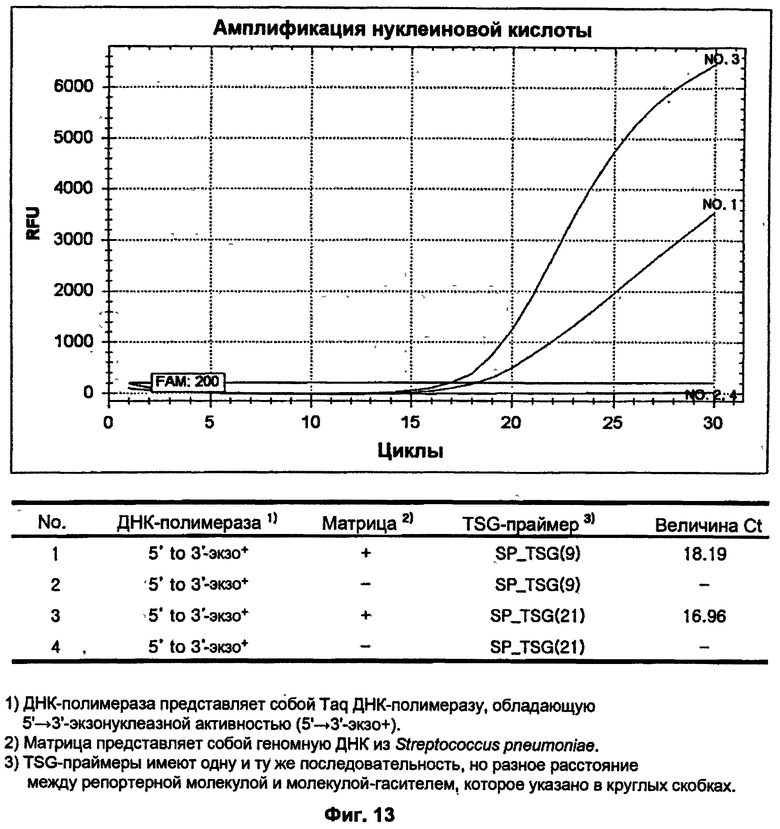
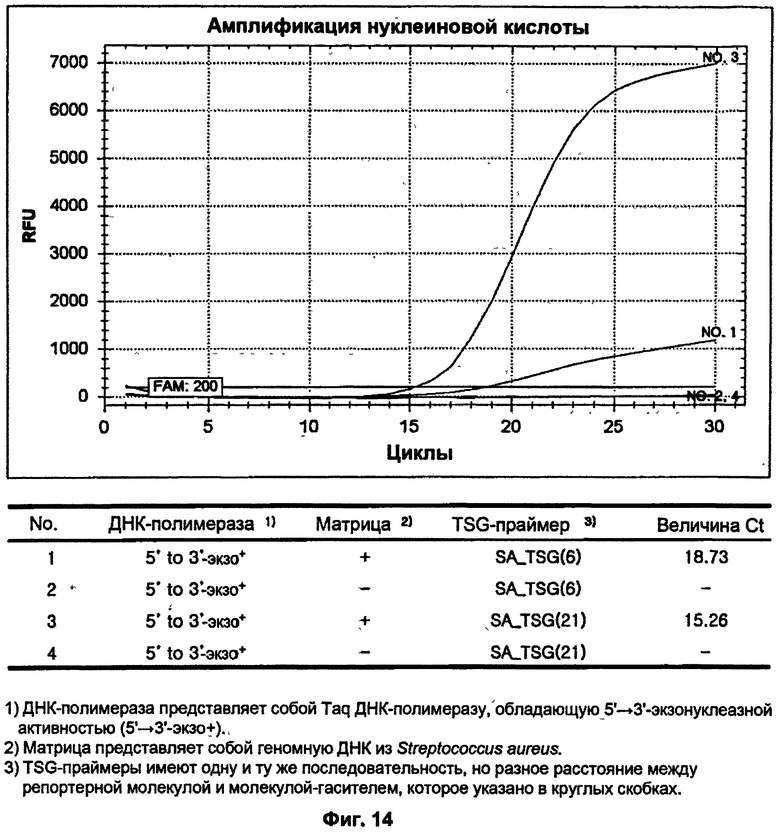

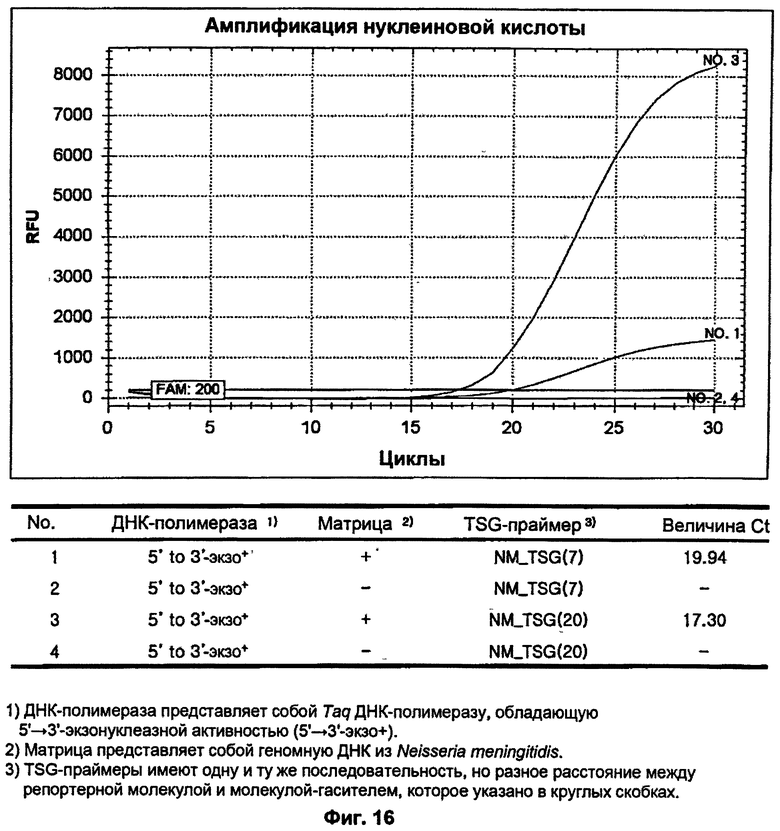
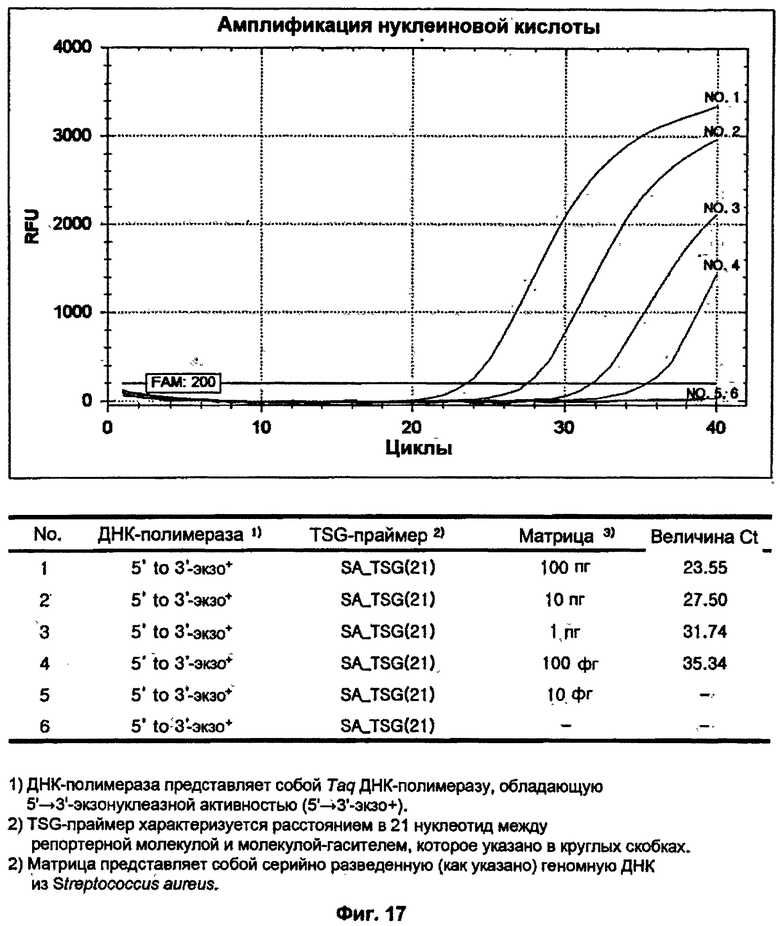
Изобретение относится к биотехнологии, а именно к способу детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера). Способ включает гибридизацию нуклеиновокислотной последовательности-мишени с TSG-праймером, который содержит гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и репортерную молекулу и молекулу-гаситель. TSG-праймер не является структурой петля-шпилька. В случае если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу без образования структур петля-шпилька, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы. В случае если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени. Приведение в контакт продукта с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера так, что реакция 3′-удлинения индуцируется по 3′-концу TSG-праймера. Детекция сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени. Предложенное изобретение позволяет с высокой чувствительностью и специфичностью детектировать нуклеиновокислотную последовательность-мишень. 3 н. и 18 з.п. ф-лы, 21 ил., 7 пр. 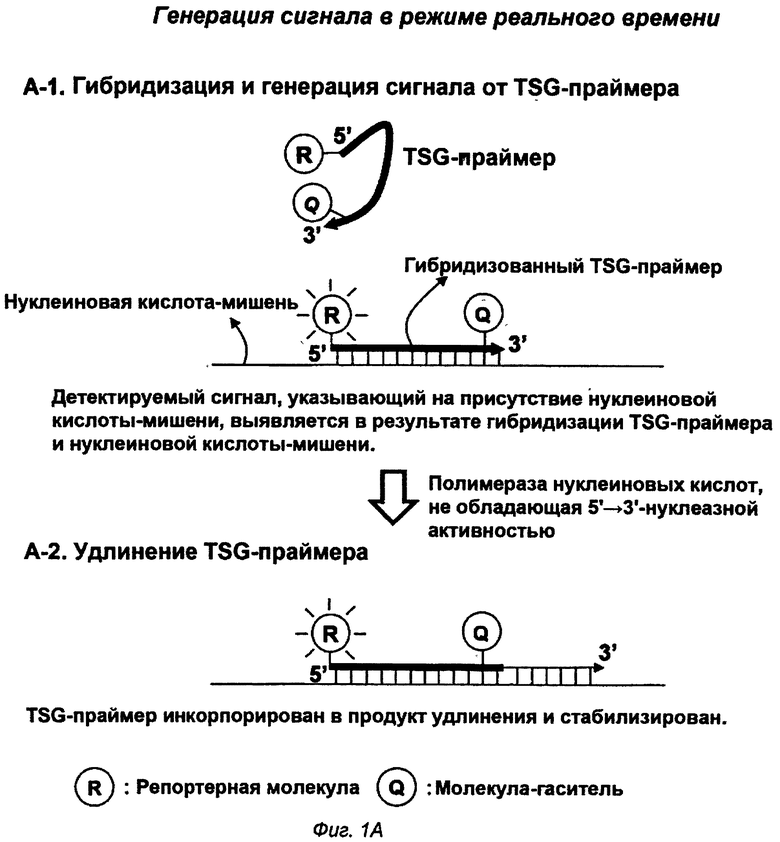
1. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера), включающий стадии:
(a) гибридизации нуклеиновокислотной последовательности-мишени с TSG-праймером; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; где TSG-праймер не является структурой петля-шпилька, при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу без образования структур петля-шпилька, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что реакция 3′-удлинения индуцируется по 3′-концу TSG-праймера; и
(c) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени в ДНК или смеси нуклеиновых кислот.
2. Способ по п. 1, где матричная полимераза нуклеиновых кислот представляет собой матричную полимеразу нуклеиновых кислот, обладающую 5′→3′-нуклеазной активностью, и TSG-праймер удлиняется по своему 3′-концу под действием полимеразной активности матричной полимеразы нуклеиновых кислот и расщепляется на своем 5′-конце под действием 5′→3′-нуклеазной активности матричной полимеразы нуклеиновых кислот, в результате чего из TSG-праймера высвобождается либо репортерная молекула, либо молекула-гаситель, генерируя сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени.
3. Способ по п. 2, дополнительно включающий повторение по меньшей мере дважды стадий (а)-(b) или (а)-(с) вместе со стадией денатурации между повторяющимися циклами с целью усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени.
4. Способ по п. 1 или 2, где репортерная молекула или молекула-гаситель на TSG-праймере расположена на его 5′-конце или на расстоянии 1-5 нуклеотидов от его 5′-конца.
5. Способ по п. 1, где молекула-гаситель представляет собой флуоресцентную молекулу и где сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, детектируемый на стадии (с), представляет собой сигнал от флуоресцентной молекулы-гасителя.
6. Способ по п. 1, где матричная полимераза нуклеиновых кислот представляет собой матричную полимеразу нуклеиновых кислот, обладающую 3′→5′-экзонуклеазной активностью.
7. Способ по п. 6, где TSG-праймер содержит по меньшей мере один ошибочно спаренный нуклеотид, имеющий остов, устойчивый в отношении 3′→5′-экзонуклеазной активности матричной полимеразы нуклеиновых кислот на своем 3′-концевом участке.
8. Способ по п. 1, который осуществляют на твердой фазе и где TSG-праймер иммобилизован через свой 5′-конец на поверхности твердой подложки.
9. Способ по п. 1 или 8, где нуклеиновокислотная последовательность-мишень включает по меньшей мере два типа нуклеиновокислотных последовательностей и TSG-праймеры включают по меньшей мере два типа праймеров.
10. Способ по п. 1 или 8, где нуклеиновокислотная последовательность-мишень имеет вариабельность нуклеотидов.
11. Способ детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера), в реакции амплификации, включающий стадии:
(а) гибридизации нуклеиновокислотной последовательности-мишени с использованием пары праймеров, состоящей из двух праймеров в качестве прямого праймера и обратного праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень; при этом по меньшей мере один из этих двух праймеров представляет собой TSG-праймер; при этом TSG-праймер содержит (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; где TSG-праймер не является структурой петля-шпилька, при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу без образования структур петля-шпилька, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) приведения в контакт продукта со стадии (а) с матричной полимеразой нуклеиновых кислот в условиях удлинения праймера, так что реакция 3′-удлинения индуцируется по 3′-концам этих двух праймеров;
(c) денатурации продукта со стадии (b);
(d) повторение стадий (а)-(с) по меньшей мере дважды с целью как амплификации нуклеиновокислотной последовательности-мишени, так и усиления сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени; и
(e) детекции сигнала, указывающего на присутствие нуклеиновокислотной последовательности-мишени, при этом детекцию осуществляют для каждого цикла повторного проведения стадии (d), в конце повторного проведения стадии (d) или в каждый из предварительно заданных интервалов времени в процессе повторного проведения, тем самым данный сигнал указывает на присутствие нуклеиновокислотной последовательности-мишени.
12. Способ по п. 11, где матричная полимераза нуклеиновых кислот представляет собой матричную полимеразу нуклеиновых кислот, обладающую 5′→3′-нуклеазной активностью, и TSG-праймер удлиняется по своему 3′-концу под действием полимеразной активности матричной полимеразы нуклеиновых кислот и расщепляется на своем 5′-конце под действием 5′→3′-нуклеазной активности матричной полимеразы нуклеиновых кислот, в результате чего из TSG-праймера высвобождается либо репортерная молекула, либо молекула-гаситель, генерируя сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени.
13. Способ по п. 11 или 12, где репортерная молекула или молекула-гаситель на TSG-праймере расположена на его 5′-конце или на расстоянии 1-5 нуклеотидов от его 5′-конца.
14. Способ по п. 11, где молекула-гаситель представляет собой флуоресцентную молекулу и где сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени, детектируемый на стадии (е), представляет собой сигнал от флуоресцентной молекулы-гасителя.
15. Способ по п. 11, где матричная полимераза нуклеиновых кислот представляет собой матричную полимеразу нуклеиновых кислот, обладающую 3′→5′-экзонуклеазной активностью.
16. Способ по п. 11, который осуществляют на твердой фазе, где по меньшей мере один из этих двух праймеров иммобилизован через свой 5′-конец на поверхности твердой подложки и где иммобилизованный праймер представляет собой TSG-праймер.
17. Набор для детекции нуклеиновокислотной последовательности-мишени из ДНК или смеси нуклеиновых кислот с использованием праймера, генерирующего сигнал от мишени (TSG-праймера), содержащий:
(a) TSG-праймер, содержащий (1) гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновокислотной последовательности-мишени, и (2) репортерную молекулу и молекулу-гаситель; где TSG-праймер не является структурой петля-шпилька, при этом если TSG-праймер не гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно располагаются близко друг к другу без образования структур петля-шпилька, что позволяет молекуле-гасителю гасить сигнал от репортерной молекулы; при этом если TSG-праймер гибридизуется с нуклеиновокислотной последовательностью-мишенью, то репортерная молекула и молекула-гаситель пространственно разделяются, что не позволяет молекуле-гасителю гасить сигнал от репортерной молекулы, в результате чего генерируется и выявляется сигнал, указывающий на присутствие нуклеиновокислотной последовательности-мишени;
(b) матричную полимеразу нуклеиновых кислот, способную индуцировать реакцию 3′-удлинения по 3′-концу TSG-праймера в результате действия на продукт гибридизации TSG-праймера и нуклеиновокислотной последовательности-мишени.
18. Набор по п. 17, содержащий пару праймеров, состоящую из двух праймеров в качестве прямого праймера и обратного праймера, способных амплифицировать нуклеиновокислотную последовательность-мишень; при этом по меньшей мере один из этих двух праймеров представляет собой TSG-праймер.
19. Набор по п. 17 или 18, где матричная полимераза нуклеиновых кислот обладает 5′→3′-нуклеазной активностью для индукции как реакции 3′-удлинения по 3′-концу TSG-праймера в результате проявления ее полимеразной активности, так и реакции 5′-расщепления на 5′-конце TSG-праймера в результате проявления ее 5′→3′-нуклеазной активности.
20. Набор по любому из пп. 17 и 18, где репортерная молекула или молекула-гаситель на TSG-праймере расположена на его 5′-конце или на расстоянии 1-5 нуклеотидов от его 5′-конца.
21. Набор по п. 17 или 18, где матричная полимераза нуклеиновых кислот представляет собой матричную полимеразу нуклеиновых кислот, обладающую 3′-5′-экзонуклеазной активностью.
| WO 2002103050 A2, 27.12.2002 | |||
| HART K W ET AL., Novel method for detection, typing, and quantification of human papillomaviruses in clinical samples JOURNAL OF CLINICAL MICROBIOLOGY, 2001, v | |||
| Машина для изготовления проволочных гвоздей | 1922 |
|
SU39A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| УСТРОЙСТВО ДЛЯ ВЫГРУЗКИ СЫПУЧИХ МАТЕРИАЛОВ ИЗ ВАГОНОВ ИЛИ ДРУГИХ ПОМЕЩЕНИЙ | 1925 |
|
SU3204A1 |
| US2009047214 A1, 19.02.2009 | |||
| WO2004092406 A1, 28.10.2004 | |||
| JORDENS J.Z | |||
| ET AL., Amplification with molecular beacon primers and reverse | |||

