Область техники.
Настоящее изобретение относится к биотехнологии, конкретно к гену, полученному из бактерии Escherichia coli, кодирующему 6-фосфоглюконолактоназу, и к способу получения L-аминокислот методом ферментации. Указанный ген способствует увеличению продукции L-аминокислот, а именно ароматических L-аминокислот, таких как L-триптофан, L-фенилаланин и L-тирозин.
Предшествующий уровень техники.
Пентозофосфатный путь (РРР) является важной частью центрального метаболизма у большинства организмов. В окислительной ветви пентозофосфатного пути происходит синтез НАДФ-Н, а фосфорилированные карбогидраты - промежуточные соединения неокислительной ветви пентозофосфатного пути, являются предшественниками пути биосинтеза нуклеотидов (рибозо-5-фосфат), ароматических аминокислот и витаминов (эритрозо-5-фосфат). Эритрозо-4-фосфат (Е4р) является основным предшественником общего пути биосинтеза ароматических L-кислот. Таким образом, оптимизация специфических путей биосинтеза фосфоенолпирувата (PEP) и Е4р может положительно повлиять на продукцию ароматических L-аминокислот.
Окислительная ветвь пентозофосфатного цикла включает в себя три реакции. Первая и третья реакции катализируются хорошо изученными ферментами глюкозо-6-фосфатдегидрогеназой (ЕС 1.1.1.49) и глюконат-6-фосфатдегидрогеназой (ЕС 1.1.1.44), которые кодируются генами zwf и gnd соответственно. Вторая реакция - это реакция гидролиза 6-фосфоглюконолактона с образованием 6-фосфоглюконата (Escherichia coli and Salmonella, Second Edition, Editor in Chief: F.C.Neidhardt, ASM Press, Washington D.C., 1996). Фермент, который катализирует эту реакцию, был обнаружен во многих организмах, включая, например, человека (Collard, F., et al, FEBS Lett., 459:2, 223-6 (1999)), Trypanosoma brucei (Duffieux, F., et al, J. Biol. Chem., 275:36, 27559-65 (2000)), Plasmodium berghei (Clarke, J.L., et al, Eur. J. Biochem., 268:7, 2013-9 (2001)), Pseudomonas aeroginosa (Hager P.W. et al, J. Bacteriol., 182:14, 3934-41 (2000)), Pseudomonas putida (Petruschka, L., et al, FEMS Microbiol. Lett., 215:1, 89-95 (2002)), хотя также известно, что указанная реакция может протекать спонтанно.
δ-6-Фосфоглюконолактон, один из продуктов реакции, катализируемой глюкозо-6-фосфатдегидрогеназой, способен образовывать изомер γ-6-фосфоглюконолактон путем внутримолекулярной перегруппировки. Только δ-6-фосфоглюконолактон может спонтанно гидролизоваться с образованием 6-фосфоглюконата, и абсолютно такую же реакцию катализируют известные ферменты 6-фосфоглюконолактоназы (ЕС 3.1.1.31) (Miclet E. et al., J Biol Chem., 276:37, 34840-46 (2001)). Ген pgl из E.coli, гипотетически presumably кодирующий 6-фосфоглюконолактоназу, был картирован на хромосоме E. coli между att-λ и геном chlD (в современных базах данных - ген modC). Мутанты E.coli (pgl-) имеют фенотип "maltose-blue" - колонии синего цвета на среде с мальтозой (Kupor, S.R. and Fraenkel, D.G., J. Bacteriol., 100:3, 1296-1301 (1969)), являющийся отличительной чертой штаммов, накапливающих мальтодекстрин (Adhya S. and Schwartz M., J Bacteriol, 108:2, 621-626 (1971)).
Но в настоящее время, неизвестны ни нуклеотидная последовательность, ни точное местоположение гена pgl на хромосоме E.coli. Из E.coli не были выделены ферменты, обладающие активностью 6-фосфоглюконолактоназы, также не существует публикаций, связывающих увеличение активности 6-фосфоглюконолактоназы в клетке бактерии - продуцента L-аминокислот с повышением продукции L-аминокислот указанной бактерией.
Описание изобретения.
Целью настоящего изобретения является предоставление
6-фосфоглюконолоктоназы из E.coli, для повышения продукции ароматических L-аминокислот штаммами-продуцентами, а также способ получения ароматических L-аминокислот указанными штаммами.
Данная цель была достигнута путем идентификации того факта, что открытая рамка считывания (ORF) ybhE в штамме E. coli К-12 кодирует 6-фосфоглюконолактоназу и усиление экспрессии ybhE ORF (pgl гена) способно повысить продукцию L-триптофана соответствующими штаммами-продуцентами L-триптофана. Таким образом было совершено настоящее изобретение.
Целью настоящего изобретения является предоставление 6-фосфоглюконолактоназы, выбранной из группы, состоящей из следующих белков:
(A) белок, включающий последовательность аминокислот, приведенную в списке последовательностей под номером 2 (SEQ ID NO:2); и
(B) белок, включающий последовательность аминокислот, содержащую делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), который обладает как минимум 70% гомологией по отношению к аминокислотной последовательности, приведенной под номером 2 (SEQ ID NO:2), и который обладает активностью 6-фосфоглюконолактоназы.
Далее по тексту белки, описанные в пунктах (А) или (В), называются "белки согласно настоящему изобретению".
Также целью настоящего изобретения является предоставление ДНК, кодирующей описанную выше 6-фосфоглюконолактоназу.
Также целью настоящего изобретения является предоставление бактерии-продуцента L-аминокислоты, при этом указанная бактерия модифицирована таким образом, что активность 6-фосфоглюконолактоназы повышена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, при этом указанная бактерия принадлежит к семейству Enterobacteriaceae, и к роду, выбранному из группы родов Escherichia, Erwinia, Providencia и Serratia.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой активность 6-фосфоглюконолактоназы повышена путем модификации нуклеотидной последовательности контролирующей экспрессию гена 6-фосфоглюконолактоназы в хромосоме указанной бактерии, таким образом, что экспрессия указанного гена усиливается.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой природный промотор описанного выше гена заменен на более сильный промотор.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой ген 6-фосфоглюконолактоназы получен из бактерии, принадлежащей к роду Escherichia.
Также целью настоящего изобретения является предоставление описанной выше бактерии-продуцента L-аминокислот, в которой ген, кодирующий 6-фосфоглюконолактоназу, выбран из группы, состоящей из:
(a) ДНК, включающей последовательность нуклеотидов с 1 по 996, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1); или
(b) ДНК, которая гибридизуется в жестких условиях с последовательностью нуклеотидов с 1 по 996, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1), или с зондом, приготовленным на основании указанной последовательности нуклеотидов, и кодирует белок, обладающий активностью 6-фосфоглюконолактоназы.
Также целью настоящего изобретения является предоставление описанной выше бактерии, где жесткими условиями являются условия, при которых отмывка осуществляется при 60°С в течение 15 минут, а концентрация соли соответствует 1×SSC и 0.1% SDS.
Также целью настоящего изобретения является предоставление описанной выше бактерии, где бактерия модифицирована таким образом, что экспрессия открытой рамки считывания ybhE усилена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, где L-аминокислота - это ароматическая L-аминокислота, выбранная из группы аминокислот, состоящей из L-триптофана, L-фенилаланина и L-тирозина.
Также целью настоящего изобретения является предоставление способа продукции ароматических L-аминокислот, включающего в себя выращивание описанной выше бактерии и выделение из культуральной жидкости накопленной в ней L-аминокислоты.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором L-аминокислоты - это ароматические аминокислоты, выбранные из группы, состоящей из L-триптофана, L-фенилаланина и L-тирозина.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором в указанной бактерии экспрессия генов путей биосинтеза ароматических аминокислот усилена.
Описанный способ получения L-аминокислот включает в себя способ получения L-триптофана с использованием бактерии-продуцента L-триптофана, в которой активность белка согласно настоящему изобретению повышена. Также описанный способ получения L-аминокислот включает в себя способ получения L-фенилаланина с использованием бактерии-продуцента L-фенилалнина, в которой активность белка согласно настоящему изобретению повышена. Также описанный способ получения L-аминокислот включает в себя способ получения L-тирозина с использованием бактерии-продуцента L-тирозина, в которой активность белка согласно настоящему изобретению повышена.
Краткое описание чертежей
На Фиг.1 показана структура природного фрагмента бактериальной ДНК вокруг открытой рамки считывания (ORF) ybhE.
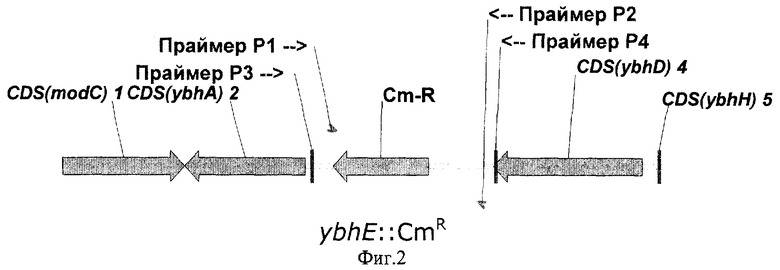
На Фиг.2 показана структура фрагмента бактериальной ДНК с делегированной ybhE ORF.
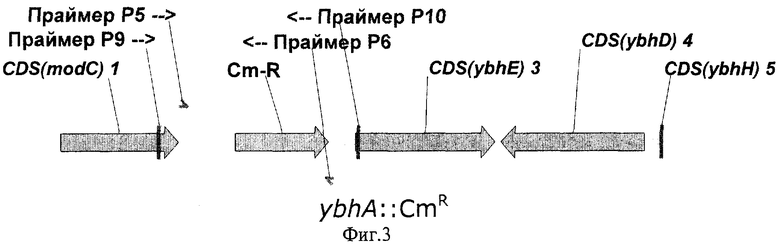
На Фиг.3 показана структура фрагмента бактериальной ДНК с делегированной ybhA ORF.
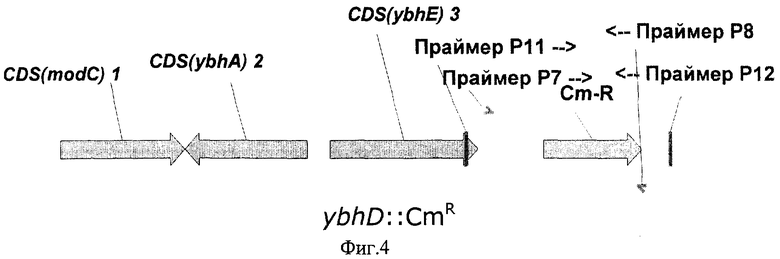
На Фиг.4 показана структура фрагмента бактериальной ДНК с делегированной ybhD ORF.
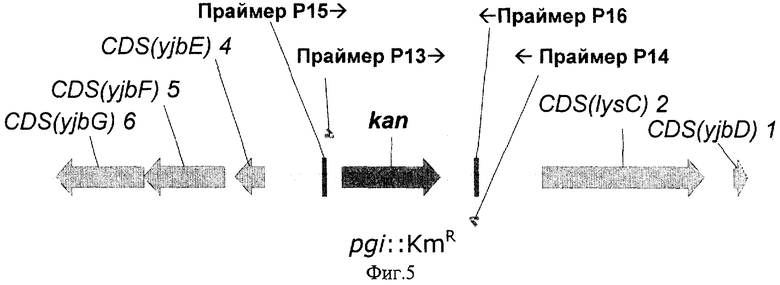
На Фиг.5 показана структура фрагмента бактериальной ДНК с делегированным геном pgi.
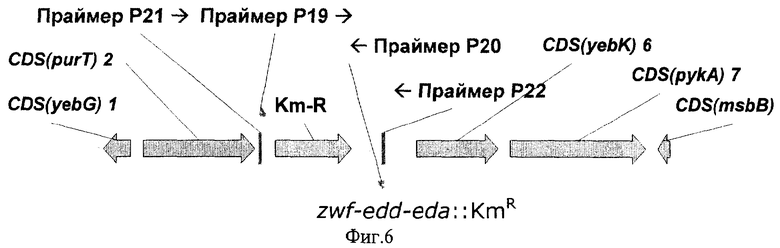
На Фиг.6 показана структура фрагмента бактериальной ДНК с делетированным опероном zwf-edd-eda.
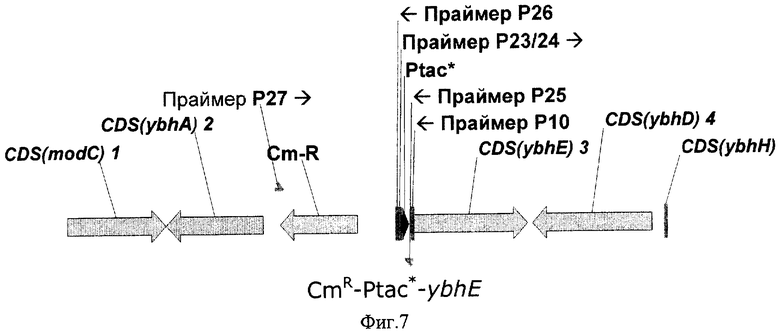
На Фиг.7 показана структура фрагмента бактериальной ДНК с искусственным промоторным регионом (Рtac*) перед геном pgl (ybhE ORF).
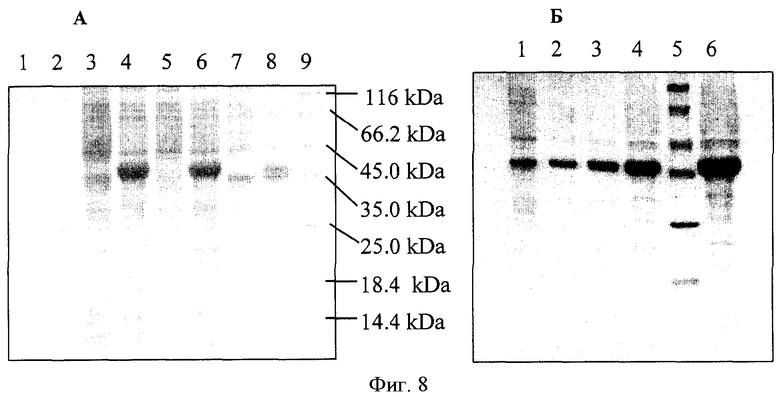
На Фиг.8 показаны разделение в геле белка (His)6-YbhE и его очистка в геле. А. Грубый экстракт клеток штамма BL21(DE3)[pET-HTybhE]. Дорожки 1, 2, 9 - белковый маркер молекулярного веса; дорожки 4, 3 - общий белок из штаммов после индукции с помощью IPTG и без нее; дорожки 6, 5 - растворимая фракция из штаммов после индукции с помощью IPTG и без нее; дорожки 8, 7, - нерастворимая фракция из штаммов после индукции с помощью IPTG и без нее. В. Дорожка 1 - общий белок из штамма BL21(DE3)[pET-HTybhE], дорожки 2, 3, 4, 6 - повышающиеся концентрации очищенного белка (His)6-YbhE; дорожка 5 - белковый маркер молекулярного веса.
Наилучший способ осуществления изобретения.
Согласно настоящему изобретению 6-фосфоглюконолактоназа - это фермент, полученный из бактерии, принадлежащий к роду Escherichia и обладающий активностью 6-фосфоглюконолактоназы. Термин "активность 6-фосфоглюконолактоназы" обозначает активность, каталитизирующую реакцию гидролиза 6-фосфоглюконолактона до 6-фосфоглюконата. Активность 6-фосфоглюконолактоназы может быть измерена с помощью метода, описанного, например, у Kupor, S.R. и Fraenkel, D.G. (J. Bacteriol, 100:3, 1296-1301 (1969)).
Примером 6-фосфоглюконолактоназы, полученной из бактерии, принадлежащей к роду Escherichia является белок, выбранный из группы, состоящей из следующих белков:
(A) белок, включающий последовательность аминокислот, приведенную в списке последовательностей под номером 2 (SEQ ID NO:2); и
(B) белок, включающий последовательность аминокислот, содержащую делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательность аминокислот, приведенную в списке последовательностей под номером 2 (SEQ ID NO:2), которая обладает как минимум 70% гомологией по отношению к аминокислотной последовательности, приведенной под номером 2 (SEQ ID NO:2), и который обладает активностью 6-фосфоглюконолактоназы.
В качестве гена, кодирующего 6-фосфоглюконолактоназу из Escherichia coli (ЕС 3.1.1.31), заявляется ген pgl, включающий в себя открытую рамку считывания ybhE ORF (номера нуклеотидов с 797809 по 798804 в последовательности с инвентарным номером NC_000913.1 в GenBank; gi:16128735). Открытая рамка считывания ybhE расположена в хромосоме штамма Е.coli К 12 между открытыми рамками считывания (ORF) ybhA и ybhD.
Таким образом, ген pgl может быть получен с помощью ПЦР (полимеразной цепной реакции; по White, T.J. et al., Trends Genet., 5, 185 (1989)) с использованием праймеров, синтезированных на основе нуклеотидной последовательности указанного гена.
Примером гена pgl из Escherichia coli является ДНК, выбранная из группы, состоящей из:
(a) ДНК, которая включает последовательность нуклеотидов с 1 по 996, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1); или
(b) ДНК, которая гибридизуется в жестких условиях с последовательностью нуклеотидов с 1 по 996, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1), или с зондом, приготовленным на основании указанной последовательности нуклеотидов, и кодирует белок, обладающий активностью 6-фосфоглюконолактоназы.
ДНК, кодирующая белки согласно настоящему изобретению, включает в себя ДНК, кодирующую белок, содержащий делеции, замены, вставки или добавления одной или нескольких аминокислот в одной или нескольких положениях в белке (А) при условии, что указанный белок не теряет своей активности. Несмотря на то, что количество "нескольких" аминокислот может быть различным в зависимости от положения и типа аминокислотного остатка в трехмерной структуре указанного белка, это количество может варьировать от 2 до 30, предпочтительно от 2 до 20 и более предпочтительно от 2 до 10 аминокислотных остатков для белка (А). Белок согласно настоящему изобретению, содержащий описанные выше делеции, замены, вставки или добавления одной или нескольких аминокислот, должен иметь как минимум 70%-ную гомологию к белку, представленному в списке последовательностей под номером 2 (SEQ ID NO:2). Процент гомологии измеряется с помощью сравнения изучаемой последовательности с приведенной в списке последовательностей под номером 2 (SEQ ID NO:2) по всей длине последовательности и определения количества подобных аминокислотных остатков. Белок согласно настоящему изобретению должен иметь как минимум 70%-ную гомологию к белку, представленному в списке последовательностей под номером 2 (SEQ ID NO:2), более предпочтительна гомология как минимум в 80%, еще более предпочтительна гомология как минимум в 90% и наиболее предпочтительна гомология как минимум в 95% к белку, представленному в списке последовательностей под номером 2 (SEQ ID NO:2).
Процент гомологии белка или ДНК также может быть оценен с помощью известных расчетных методов, таких как BLAST, FASTA или CrustalW. BLAST (Basic Local Alignment Search Tool) является эвристическим поисковым алгоритмом, использованным в программных продуктах blastp, blastn, blastx, megablast, tbiastn, и tbiastx; указанные программы приписывают величины значимости в результате поиска с использованием статистических методов, разработанных Karlin, Samuel и Stephen F. Altschul ("Methods for assessing the statistical significance of molecular sequence features by using general scoring schemes". Proc. Natl. Acad. Sci. USA, 1990, 87:2264-68; "Applications and statistics for multiple high-scoring segments in molecular sequences". Proc. Natl. Acad. Sci. USA, 1993, 90:5873-7). Метод поиска FASTA описан W.R.Pearson ("Rapid and Sensitive Sequence Comparison with FASTP and FASTA", Methods in Enzymology, 1990 183:63-98). Метод ClustalW описан Thompson J.D., Higgins D.G. и Gibson T.J. ("CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice", Nucleic Acids Res. 1994, 22:4673-4680).
Для того чтобы сохранялась активность белка, замены, делеции, вставки или добавки одного или нескольких аминокислотных остатков в указанном белке должны являться консервативной мутацией(ями). Пример консервативной мутации - это консервативная замена. Примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ile на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp, и замену Val на Met, Ile или Leu.
ДНК, кодирующая практически такие же белки, как белок, описанный в пункте (А), может быть получена, например, путем модификации нуклеотидной последовательности ДНК, кодирующей белок, описанный в пункте (А), например, с помощью метода сайт-направленного мутагенеза, таким образом, что один или несколько аминокислотных остатков могут быть делегированы, заменены, вставлены или добавлены. ДНК, модифицированная, как описано выше, может быть получена традиционными известными методами обработки с целью получения мутаций. Такие методы включают обработку ДНК, кодирующей белок согласно настоящему изобретению, гидроксиламином или обработку бактерии, содержащей указанную ДНК, с помощью УФ-излучения или реагентом, таким как N-метил-N'-нитро-N-нитрозогуанидин или азотистая кислота.
ДНК, кодирующая белки согласно настоящему изобретению, включает также ДНК, которая может быть получена из различных штаммов и вариантов бактерий, принадлежащих к роду Escherichia ввиду природного разнообразия. ДНК, кодирующая такие белки, может быть получена путем выделения ДНК, которая гибридизуется геном pgl или его частью в жестких условиях, и кодирует белок, обладающий активностью 6-фосфоглюконолактоназы. Термин "жесткие условия", упомянутый здесь, означает условия, при которых образуются так называемые специфические гибриды, а неспецифические - не образуются. Трудно четко описать такие условия в каких-либо численных выражениях. Например, к жестким условиям относятся условия, при которых гибридизуются ДНК, обладающие высокой степенью гомологии, к примеру ДНК, обладающие гомологией не менее 70%, предпочтительно не менее 80%, более предпочтительно не менее 90%, более предпочтительно не менее 95%, друг относительно друга. С другой стороны, примером жестких условий являются условия, при которых ДНК гибридизуются друг с другом, при концентрации солей, соответствующей стандартным условиям отмывки при гибридизации по Саузерну, например, 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS при 60°С. Продолжительность отмывки зависит от вида фильтра, используемого для блоттинга и, как правило, рекомендована изготовителем. Например, рекомендованная продолжительность отмывки нейлонового фильтра HybondTM N+(Amersham) в жестких условиях составляет 15 минут. Предпочтительно, чтобы отмывка производилась два или три раза.
В качестве зонда для ДНК, кодирующей описанные выше белки, и которая гибридизуется с геном pgl, может быть использована часть нуклеотидной последовательности под номером 1. Зонд подобного рода может быть получен в результате ПЦР с использованием в качестве праймеров олигонуклеотидов, полученных на основе нуклеотидной последовательности под номером 1, и фрагмента ДНК, содержащего нуклеотидную последовательность под номером 1, в качестве матрицы. В случае когда в качестве зонда используется фрагмент ДНК длиной около 300 пар оснований, условия отмывки при гибридизации соответствуют, например, 50°С, 2×SSC и 0.1% SDS.
Трансформация бактерии с помощью ДНК, кодирующей белок, означает введение указанной ДНК в клетку бактерии, например, традиционными методами для увеличения экспрессии указанного гена, кодирующего белок согласно настоящему изобретению, и увеличения активности указанного белка в клетке бактерии.
Бактерия согласно настоящему изобретению - это бактерия, принадлежащая к роду Escherichia, - продуцент L-аминокислот, обладающая повышенной активностью белка, что приводит к увеличению продукции целевой L-аминокислоты. Предпочтительно, бактерией согласно настоящему изобретению является бактерия, принадлежащая к роду Escherichia, - продуцент ароматической L-аминокислоты, обладающая повышенной активностью белка согласно настоящему изобретению. Более предпочтительно, бактерией согласно настоящему изобретению является бактерия, принадлежащая к роду Escherichia, - продуцент ароматических L-аминокислот, такая как бактерия-продуцент L-триптофана, модифицированная таким образом, что активность 6-фосфоглюконолактоназы повышена. Еще более предпочтительно, если бактерия согласно настоящему изобретению содержит ДНК с геном pgl (ybhE ORF), при этом последовательность контроля экспрессии указанного гена в хромосоме этой бактерии модифицирована таким образом, что способность этой бактерии к продукции L-триптофана усилена.
Термин "бактерия - продуцент L-аминокислоты" означает бактерию, обладающую способностью к накоплению этой L-аминокислоты в питательной среде, в условиях, когда бактерия согласно настоящему изобретению выращивается в указанной питательной среде. Способность к продукции L-аминокислоты может быть придана или улучшена путем селекции. Используемый здесь термин "бактерия-продуцент L-аминокислоты" также означает бактерию, которая способна к продукции L-аминокислоты и вызывает накопление L-аминокислоты в питательной среде в количествах больших, чем природный или родительский штамм, и, предпочтительно, означает, что микроорганизм способен производить и накапливать в ферментационной среде целевую L-аминокислоту в концентрациях не менее чем 0.5 г/л, более предпочтительно - не менее 1.0 г/л.
L-аминокислоты включают в себя: L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин (цистин), L-глутаминовую кислоту, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин, и, предпочтительно, включают в себя ароматические L-аминокислоты, такие как L-триптофан, L-фенилаланин и L-тирозин.
Семейство Enterobacteriaceae включает в себя бактерий, принадлежащих к роду Escherichia, Erwinia, Providencia и Serratia. Род Escherichia предпочтителен. Термин "бактерия, принадлежащая к роду Escherichia" означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, может быть упомянута бактерия Escherichia coli (E.coli).
Термин "модифицирована таким образом, что активность 6-фосфоглюконолактоназы увеличена" означает то, что удельная активность становится выше, чем у немодифицированного штамма, например природного штамма. Например, в случае когда количество молекул 6-фосфоглюконолактоназы в клетке увеличено, когда специфическая активность у молекулы 6-фосфоглюконолактоназы увеличена и так далее. Кроме того, в качестве природного штамма, который может служить объектом для сравнения, может быть упомянут штамм Escherichia coli К-12. В результате увеличения внутриклеточной активности 6-фосфоглюконолактоназы наблюдается эффект увеличения количества накопленной L-аминокислоты.
Увеличение активности 6-фосфоглюконолактоназы в клетке бактерии может быть достигнуто путем увеличения экспрессируемого количества гена, кодирующего 6-фосфоглюконолактоназу. Примером гена, кодирующего 6-фосфоглюконолактоназу, может являться ген, выделенный из бактерий, принадлежащих к роду Escherichia.
Бактерия согласно настоящему изобретению включает в себя бактерию, в которой активность белка согласно настоящему изобретению повышена путем замены последовательности, контролирующей экспрессию ДНК, кодирующей белок, описанный в пункте (А) или (В) в хромосоме бактерии, на более сильную. Усиление экспрессии гена может быть достигнуто помещением ДНК согласно настоящему изобретению под контроль более сильного промотора взамен природного. Термин "природный промотор" означает фрагмент ДНК в природном организме, расположенный перед открытой рамкой считывания (ORF) гена и способствующий транскрипции этого гена. Сила промотора определяется частотой акта начала синтеза РНК. Методы оценки силы промотора и примеры сильных промоторов описаны у Deuschle, U., Kammerer, W., Gentz, R., Bujard, H. (Promoters in Escherichia coli: a hierarchy of in vivo strength indicates alternate structures. EMBO J., 1986, 5, 2987-2994).
Усиление трансляции может быть достигнуто с помощью введения в ДНК согласно настоящему изобретению более эффективного сайта связывания рибосомы (Ribosome Binding Site - RBS) взамен природной последовательности RBS. Последовательность RBS - это область, расположенная перед старт-кодоном мРНК, которая взаимодействует с 16S РНК рибосомы (Shine J. and Dalgarno L., Proc. Natl. Acad. Sci. USA, 1974, 71, 4, 1342-6). Термин "природная последовательность RBS" означает последовательность RBS в природном организме. В качестве примера эффективной последовательности RBS можно привести последовательность RBS гена 10 из фага Т7 (Olins P.O. et al, Gene, 1988, 73, 227-235).
Методами получения хромосомной ДНК, гибридизации, ПЦР, получения плазмидной ДНК, разрезания и лигирования ДНК, трансформации, выбора олигонуклеотидов в качестве праймеров и подобные им могут быть обычные методы, хорошо известные специалисту в данной области. Эти методы описаны, например, в книге Sambrook, J., and Russell D., "Molecular Cloning A Laboratory Manual, Third Edition", Cold Spring Harbor Laboratory Press (2001) и подобным ей.
Бактерия согласно настоящему изобретению может быть получена путем введения вышеуказанных ДНК в бактерию, уже обладающую способностью к продукции L-аминокислот. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, уже содержащей указанные ДНК, способности к продукции L-аминокислот.
В качестве родительского штамма, в котором активность белка согласно настоящему изобретению будет повышена, могут быть использованы штаммы, принадлежащие к роду Escherichia, - продуценты L-триптофана, такие как штаммы Е.coli JP4735/pMU3028 (DSM10122) и JP6015/pMU91 (DSM10123), дефицитные по триптофанил-тРНК синтетазе, кодируемой мутантным геном trpS (патент США 5756345); штамм Е.coli SV164 (pGH5) с аллелем гена serA, кодирующим фермент со снятым ингибированием серином по принципу обратной связи (патент США 6180373); штаммы Е.coli AGX17 (pGX44) (NRRL В-12263) и AGX6 (pGX50) aroP (NRRL В-12264) дефицитные по ферменту триптофаназе (патент США 4371614); штамм Е.coli AGX17/pGX50, pACKG4-pps, в котором усилена способность к продукции фосфоенолпирувата (WO 9708333, патент США 6319696) и подобные им.
В качестве родительского штамма, в котором активность белка согласно настоящему изобретению будет повышена, могут быть использованы штаммы, принадлежащие к роду Escherichia, - продуценты L-фенилаланина, такие как штамм AJ12739 (tyrA::Tn10, tyrR) (ВКПМ В-8197); штамм HW1089 (АТСС инвентарный номер 55371), содержащий ген pheA34 (патент США 5354672); мутантный штамм MWEC101-b (патент Южной Кореи 8903681); штаммы NRRL В-12141, NRRL В-12145, NRRL В-12146 и NRRL B-12147 (патент США 4407952) и подобные им. Также в качестве родительского штамма для последующего повышения в нем активности белка согласно настоящему изобретению, могут быть использованы бактерии-продуценты L-фенилаланина, принадлежащие к роду Escherichia, штаммы Е.coli К-12 [W3110 (tyrA)/pPHAB (FERM BP-3566), Е.coli К-12 [W3110 (tyrA)/pPHAD] (FERM BP-12659), E. coli K-12 [W3110 (tyrA)/pPHATerm] (FERM BP-12662) и E. coli K-12 [W3110 (tyrA)/pBR-aroG4, pACMAB], называемый также AJ 12604 (FERM BP-3579) (Европейский патент 488424 B1).
Авторами настоящего изобретения ранее было показано, что ген yddG, кодирующий мембранный белок, не участвующий в путях биосинтеза ни одной из L-аминокислот, в случаях когда аллель природного гена амплифицирован на многокопийном векторе, сообщает микроорганизмам устойчивость к L-фенилаланину и нескольким аналогам аминокислот. Кроме того, ген yddG может повысить продукцию L-фенилаланина или L-триптофана, в случаях когда дополнительные копии этого гена введены в клетки соответствующего штамма-продуцента (патентная заявка РФ 2002121670, WO 03044192). Следовательно, желательно, чтобы бактерия-продуцент L-триптофана была в дальнейшем модифицирована с целью усиления экспрессии открытой рамки считывания yddG.
Гены, важные для биосинтеза L-триптофана, включают в себя гены оперона trpEDCBA, гены, общие для биосинтеза всех ароматических аминокислот - такие как aroF, aroG, aroH, aroB, aroD, aroE, aroK, aroL, aroA и aroC, гены биосинтеза L-серина, такие как serA, serB и serC, и подобные им.
В качестве родительского штамма, в котором активность белка согласно настоящему изобретению будет повышена, могут быть использованы штаммы, принадлежащие к роду Escherichia, - продуценты L-тирозина, такие как штаммы Е.coli, в которых повышена способность к продукции фосфоенолпирувата или повышена активность ферментов общего пути биосинтеза ароматических аминокислот (Европейская патентная заявка ЕР 0877090).
Способ получения L-аминокислоты согласно настоящему изобретению включает стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-аминокислоты в питательной среде, выделения L-аминокислоты из культуральной жидкости. Также способ согласно настоящему изобретению включает в себя способ получения L-триптофана и включает стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-триптофана в питательной среде, выделения L-триптофана из культуральной жидкости. Также способ согласно настоящему изобретению включает в себя способ получения L-фенилалнина, и включает стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-фенилалнина в питательной среде, выделения L-фенилалнина из культуральной жидкости. Также способ согласно настоящему изобретению включает в себя способ получения L-тирозина и включает стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-тирозина в питательной среде, выделения L-тирозина из культуральной жидкости.
Согласно настоящему изобретению выращивание, выделение и очистка L-аминокислот, предпочтительно ароматических аминокислот, таких как L-триптофан, L-фенилалнин или L-триптофан, из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием микроорганизма.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от характера ассимиляции, используемого микроорганизмом, могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. Некоторые питательные добавки могут быть, при необходимости, добавлены в питательную среду. Например, если микроорганизму для роста необходим тирозин (ауксотрофия по тирозину), соответствующее количество тирозина может быть добавлено в питательную среду для выращивания.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией, при температуре в пределах от 20 до 42°С, предпочтительно в пределах от 37 до 40°С. рН среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно, выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем целевая L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и кристаллизации.
Примеры
Более детально настоящее изобретение будет разъяснено ниже со ссылкой на следующие Примеры, не ограничивающие область применения настоящего изобретения.
Пример 1. Идентификация гена pgl из Е.coli и сравнение нуклеотидных последовательностей.
Kupor и Fraenkel картировали мутацию pgl между генами chlD (известный в настоящее время как modC) и bioA в хромосоме Е.coli (Kupor, S.R. and Fraenkel, D.G., J. Bacteriol., 100:3, 1296-1301 (1969)). Это соответствует 17.18 и 17.40 минутам генетической карты Е.coli. В этой области расположены восемь открытых рамок считывания, кодирующих белки с неизвестными функциями. Далее, Е.coli Stock Center Database определил мутацию pgl между минутами 17.20 и 17.22. Эти координаты достаточно точно соответствуют координатам открытой рамки считывания ybhE, расположенной между открытыми рамками считываниями ybhA и ybhD (Фиг.1).
Поиск с помощью программы BLAST показал, что белок YbhE имеет большое количество гомологов с неизвестными функциями в различных организмах, таких как Shigella flexmeri (98.8% схожести), Salmonella typhi (92,8% схожести), Yersinia pestis (68,4% схожести), несколько гомологов с известными функциями, например, домен гема Цитохрома D1 из Bacillus anthracis (28% идентичности), 3-карбоксимуконатциклаза из Pseudomonas fluorescens, предсказанная автоматическим компьютерным анализом (28% идентичности), муконатциклоизомераза из Trichosporon beigelii (26% идентичности), и один гомолог из Bacillus cereus, описанный в базах данных как 6-фосфоглюконатлактоназа под инвентарным номером NP_833107, но без ссылок на печатные экспериментальные работы.
Также с помощью программы NCBI Conserved Domain Search были обнаружены три перекрывающихся консервативных белковых домена. Два из них принадлежат к консервативному белковому семейству с неизвестной функцией, и один домен принадлежит к семейству 3-карбоксимуконатциклаз.
При поиске с помощью программы BLAST в пределах протеома Е.coli не были обнаружены гомологи описанных 6-фосфоглюконатлактоназ, например, из Pseudomonas putida.
Для того чтобы установить, является ли ORF, обозначаемая как ybhE в хромосоме Е.coli, геном pgl, кодирующим 6-фосфоглюконатлактоназу, было проведено последовательное разрушение открытых рамок считывания ybhA, ybhE и ybhD, и полученные мутанты были проверены на предмет фенотипа "maltose blue" (смотри ниже).
Пример 2. Деления открытой рамки считывания (ORF) ybhE. Замена ybhE ORF на фрагмент ДНК, содержащий ген устойчивости к хлорамфениколу (CmR).
Для делеции ybhE ORF в хромосому штамма Е.coli BW25113 [pKD46] взамен природной ybhE ORF был интегрирован фрагмент ДНК, содержащий маркер устойчивости к хлорамфениколу (CmR), кодируемый геном cat, способом, описанным у Datsenko K.A. и Wanner B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645). Этот способ также называют "интеграция с использованием Red-системы" и/или "интеграция за счет Red-системы". Нуклеотидная последовательность исходной природной области ybhE ORF и аминокислотная последовательность, кодируемая указанной ORF, представлены в списке последовательностей под номерами 1 и 2 соответственно (SEQ ID NO: 1 и 2). Штамм Escherichia coli BW25113, содержащий рекомбинантную плазмиду pKD46, может быть получен из Е.coli Genetic Stock Center, Yale University, New Haven, USA, инвентарный номер штамма - CGSC7630.
Фрагмент ДНК, содержащий маркер CmR, может быть получен с помощью ПЦР, при этом в качестве матрицы использовалась коммерчески доступная плазмида pACYC184 (инвентарный номер Х06403 в GenBank/EMBL, "Ферментас", Литва), а также праймеры Р1 (SEQ ID NO: 3) и Р2 (SEQ ID NO: 4). Праймер Р1 содержал 36 нуклеотидов, гомологичных 5'-концу рамки ybhE ORF и праймер Р2 содержал 36 нуклеотидов, гомологичных 3'-концу рамки ybhE ORF. Эти последовательности гена ybhE были включены в праймеры Р1 и Р2 для дальнейшей интеграции полученного фрагмента в хромосому.
ПЦР проводили с использованием амплификатора "TennoHybaid PCR Express". Реакционная смесь общим объемом 50 мкл содержала 5 мкл 10х буфера для ПЦР ("Ферментас", Литва) с добавлением MgCl2 до конечной концентрации в реакционной смеси 15 мМ, 200 мкМ каждого трифосфата dNTP, 25 пМ каждого из используемых праймеров и 1 ед Taq-полимеразы ("Ферментас", Литва). Примерно 5 нг плазмидной ДНК добавляли в реакционную смесь в качестве матрицы для ПЦР амплификации. Температурный профиль ПЦР был следующий: начальная стадия денатурации в течение 5 мин при 95°С с последующими 25 циклами денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд, наращивания цепи при 72°С в течение 30 секунд; финальная стадия полимеризации при 72°С в течение 7 минут.
Затем амплифицированный фрагмент ДНК очищали электрофорезом в агарозном геле, выделяли из геля с помощью "GenElute Spin Columns" ("Sigma", USA) и высаживали этанолом. Нуклеотидная последовательность синтезированного фрагмента ДНК представлена в SEQ ID NO:5.
Полученный фрагмент ДНК, очищенный описанным выше способом, использовали для электропорации и интеграции с использованием Red-системы в бактериальную хромосому штамма Е.coli BW25113 [pKD46]. Рекомбинантная плазмида pKD46 (Datsenko, К.А., Wanner, B.L., Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645) с термочувствительным репликоном использовалась как донор генов фага λ, ответственных за функционирование Red-системы рекомбинации.
Клетки BW25113 [pKD46] выращивали в течение ночи при 30°С в жидкой среде LB, содержащей ампициллин (100 мкг/мл), затем разбавляли в отношении 1:100 средой SOB (дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; Триптон, 20 г/л, KCl, 2,5 мМ; MgCl2, 10 мМ), содержащей ампициллин (100 мкг/мл) и арабинозу (10 мМ) (арабиноза использовалась для индукции плазмиды, кодирующей гены системы гомологичной рекомбинации фага λ), и выращивали при 30°С до достижения оптической плотности бактериальной культуры OD600=0.4-0.7. Клетки из 10 мл подращенной бактериальной культуры отмывали 3 раза деионизированной водой, охлажденной на льду, затем ресуспендировали в 100 мкл воды. 10 мкл (100 нг) фрагмента ДНК, растворенного в деионизованной воде, добавляли к суспензии бактериальных клеток. Электропорацию проводили с помощью прибора для электротрансформации бактерий "Bio-Rad" (США) (№165-2098, версия 2-89) в соответствии с рекомендациями производителя.
После электропорации к клеточной суспензии добавляли 1 мл среды SOC Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)), подращивали в течение 2 часов при 37°С, после чего высевали на чашки с L-агаром, содержащим хлорамфеникол (25 мкг/мл). Колонии, выросшие на этих чашках в течение 24 часов тестировались с помощью ПЦР на наличие маркера CmR, заместившего природную рамку ybhE, при этом использовались праймеры Р3 (SEQ ID NO:6) и Р4 (SEQ ID NO:7). Для этой цели свежевыращенные колонии ресуспендировались в 20 мкл воды и 1 мкл полученной суспензии использовали в качестве матрицы для ПЦР. Температурный профиль ПЦР был следующий: начальная стадия денатурации ДНК - в течение 10 мин при 95°С, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд, наращивания цепи при 72°С в течение 1 минуты; финальная стадия полимеризации при 72°С в течение 7 минут.
Лишь несколько протестированных CmR колоний содержали искомый фрагмент ДНК длиной 1279 п.о., подтверждающий замену природной рамки ybhE ORF на CmR маркер ДНК. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем выращивания штамма при 37°С. Штамм, полученный в результате всех описанных выше процедур, был назван Е.coli BW25113-ΔybhE.
Структура бактериального фрагмента ДНК с разрушенной рамкой ybhE ORF показана на Фиг.2.
Пример 3. Делеция открытых рамок считывания ybhA ORF и ybhD ORF. Замена рамок ybhA и ybhD ORF на фрагмены ДНК, содержащие ген устойчивости к хлорамфениколу (CmR).
Для делеции рамок ybhA ORF и ybhD ORF, в хромосому штамма Е.coli BW25113 [pKD46] взамен природных рамок ybhA ORF и ybhD ORF независимо были интегрированы фрагменты ДНК, содержащие маркер устойчивости к хлорамфениколу (CmR), кодируемый геном cat, способом, описанным в Примере 2.
Для получения с помощью ПЦР фрагментов ДНК, необходимых для электропорации и разрушения рамок ybhA ORF и ybhD ORF, были синтезированы две пары праймеров Р5 (SEQ ID NO:8) и Р6 (SEQ ID NO:9), и Р7 (SEQ ID NO:10) и Р8 (SEQ ID NO:11) соответственно. Праймер Р5 содержал 36 нуклеотидов, гомологичных 3'-концу рамки ybhA ORF. Праймер Р6 содержал 36 нуклеотидов, гомологичных 5'-концу рамки ybhA ORF. Праймер Р7 содержал 36 нуклеотидов, гомологичных 3'-концу рамки ybhD ORF. И праймер Р8 содержал 36 нуклеотидов, гомологичных 5'-концу рамки ybhD ORF. Эти последовательности были включены в праймеры Р5, Р6, Р7 и Р8 для дальнейшей интеграции полученных фрагментов в хромосому.
Нуклеотидные последовательности синтезированных фрагментов ДНК представлены в SEQ ID NO:12 и SEQ ID NO:13 соответственно. Нуклеотидные последовательности замененных природных фрагментов с рамками ybhA ORF и ybhD ORF представлены в базе данных GenBank с инвентарным номером NC_000913.1 (номера нуклеотидов с 796836 по 797654 и с 798845 по 799777, gi:16128734 и gi:33347481 соответственно). Структуры фрагментов бактериальной ДНК с разрушенными рамками ybhA ORF и ybhD ORF показаны на Фиг.3 и Фиг.4 соответственно.
После электропорации полученные колонии были проверены с помощью ПЦР на наличие маркера CmR: для разрушенной рамки ybhA ORF использовались праймеры Р9 (SEQ ID NO:14) и P10 (SEQ ID NO:15), для разрушенной рамки ybhD ORF использовались праймеры P11 (SEQ ID NO:16) и Р12 (SEQ ID NO:17).
В первом случае несколько протестированных CmR колоний содержали искомый фрагмент ДНК в 1424 п.о., подтверждающий замену природной рамки ybhA ORF на ген CmR Во втором случае несколько протестированных CmR колоний содержали искомый фрагмент ДНК в 1386 п.о., подтверждающий замену природной рамки ybhD ORF на ген CmR. В обоих случаях один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем выращивания штамма при 37°С, и полученные в итоге штаммы, были названы Е.coli BW25113-ΔybhA и BW25113-ΔybhD соответственно.
Пример 4. Проверка мутантов ybhE-, vbhA- и ybhD- на предмет фенотипа "maltose blue".
Каждый из трех полученных мутантных штаммов был проверен на фенотип "maltose blue" (синие колонии на среде с мальтозой) способом, описанным у Kupor, S.R. и Fraenkel, D.G. (J.Bacteriol., 100:3, 1296-1301 (1969)). Штаммы были перепечатаны методом реплик на чашках с минимальной средой М9, содержащей 0.8% мальтозы. Спустя 6 часов, инкубированные чашки заливались 5 мл раствора, содержащего 0.01 М I2 и 0.03 М KI, после чего оценивали цвет каждой колонии-реплики как "синий" или "не синий".
Один из полученных штаммов BW25113-ΔybhE был оценен как "синий", в то время как другие штаммы BW25113-ΔybhA, BW25113-ΔybhD и BW25113 (в качестве контроля) были засчитаны как "не синие".
Пример 5. Конструирование штаммов с двойной мутацией - делениями pgi и ybhE или ybhD. Сравнительная характеристика роста таких штаммов на средах с различными источниками углерода.
У мутантного штамма, дефицитного по фосфоглюкозоизомеразе (pgi-), замедлен рост на глюкозе, поскольку работает только окислительная ветвь пентозофосфатного цикла. Двойной мутант, дефицитный также по фосфоглюконолактоназе (pgl), катализирующей второй этап окислительной ветви пентозофосфатного цикла должен расти на глюкозе еще медленнее, поскольку превращение 6-фосфоглюконолактона в глюконат-6-фосфат в этом случае возможно только за счет спонтанного гидролиза. Значит, если рамка ybhE ORF действительно является геном pgl, то двойной мутант pgi, ybhE будет расти медленнее, чем природный штамм и pgi мутант. Для проверки этого предположения был создан двойной мутант pgi, ybhE.
В ген pgi была введена мутация путем замены соответствующего природного участка бактериальной хромосомы в штамме Е.coli BW25113 [pKD46] на фрагмент ДНК, содержащий ген устойчивости к канамицину (KmR), способом, описанным в Примере 2. Нуклеотидная последовательность замененного природного фрагмента тени pgi представлена в базе данных GenBank, инвентарный номер NC_000913.1 (номера нуклеотидов с 4231337 по 4232986; gi:16131851).
Фрагменты ДНК, содержащие ген KmR, были синтезированы с помощью ПЦР, в качестве матрицы использовалась коммерчески доступная плазмида pUC4KAN (инвентарный номер Х06404 в GenBak/EMBL, "Ферментас", Литва), также праймеры Р13 (SEQ ID NO:18) и Р14 (SEQ ID NO:19). Праймер Р13 содержал 36 нуклеотидов, гомологичных 3'-концу гена pgi, и праймер Р14 содержал 36 нуклеотидов, гомологичных 5'-концу гена pgi. Эти последовательности гена pgi были включены в праймеры Р13 и Р14 для дальнейшей интеграции полученного фрагмента в бактериальную хромосому.
ПЦР проводили так же, как описано в Примере 2.
Затем амплифицированный фрагмент ДНК очищали электрофорезом в агарозном геле, выделяли из геля с помощью "GenElute Spin Columns" ("Sigma", США) и высаживали этанолом. Нуклеотидная последовательность синтезированного фрагмента ДНК представлена в SEQ ID NO:20.
Полученный фрагмент ДНК, очищенный описанным выше способом, использовали для электропорации и интеграции с использованием Red-системы в бактериальную хромосому штамма Е.coli BW25113 [pKD46], как описано выше в Примере 2, за исключением того, что клетки после электропорации высевались на чашки с L-агаром, содержащим канамицин в концентрации 50 мкг/мл.
Колонии, выросшие на этих чашках в течение 24 часов, тестировались с помощью ПЦР на наличие маркера KmR, заместившего природный ген pgi, при этом использовались праймеры Р15 (SEQ ID NO:21) и P16 (SEQ ID NO:22). Для этой цели свежевыращенные колонии ресуспендировали в 20 мкл воды и 1 мкл полученной суспензии использовали в качестве матрицы для ПЦР. Условия ПЦР описаны в Примере 2. Несколько протестированных KmR колоний содержали искомый фрагмент ДНК длиной 1286 п.о., подтверждающий замену природного гена pgi на ДНК с KmR маркером. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем выращивания штамма при 37°С. Штамм, полученный в результате всех описанных выше процедур, был назван Е.coli BW25113-Δpgi.
Структура полученного фрагмента бактериальной ДНК с делегированным геном pgi показана на Фиг.5.
Делеция гена pgi была перенесена трансдукцией по Fraenkel (J. Bacteriol. 93 (1967), 1582-1587) в штамм Е.coli MG1655 с последующей селекцией на чашках с канамицином. Полученный штамм был назван MG-Δpgi. Затем, мутации в рамках ybhE ORF и ybhD ORF были также перенесены трансдукцией из штаммов BW25113-ΔybhE и BW25113-ΔybhD, описанных в Примерах 2 и 3, в полученный штамм с последующей селекцией на чашках с хлорамфениколом. Полученные штаммы были названы MG-Δpgi-ΔybhE и MG-Δpgi-ΔybhD соответственно.
Оба полученных штамма, совместно с MG1655 и MG1655-Δpgi, были перепечатаны методом реплик на чашки с минимальной средой М9, содержащей глюкозу или глюконат в качестве источника углерода. После 24 часов инкубирования чашек было проведено визуальное сравнение роста указанных штаммов. Штамм MG-Δpgi-ΔybhE рос хуже по сравнению с другими штаммами на чашке с глюкозой, а на чашке с глюконатом практически не рос.
Пример 6. Конструирование плазмиды, содержащей ген pgi из Pseudomonas putida, и комплементация этой мутации ybhE.
Ген pgl описан для нескольких организмов. Среди них известен также ген 6-фосфоглюконолактоназы из Pseudomonas putida, организма, являющегося близким родственником для Е.coli. Описаны эксперименты по клонированию нескольких генов из Р.putida в Е.coli, приводящие к комплементации соответствующих мутаций в Е.coli (Ramos-Gonzalez, M.I. and Molin, S., J.Bacteriol., v 180, 13, p.3421, 1998).
Ген pgl из Р.putida был клонирован с использованием праймеров 17 (SEQ ID No.23) и 18 (SEQ ID No.24). Праймер P17 содержит последовательность, комплементарную нуклеотидной последовательности гена pgl из Р.putida с 1 по 19 п.о. Этот праймер также содержит сайт связывания рибосомы (RBS) гена lacZ из E.coli, расположенный перед описанной выше комплементарной последовательностью, а также сайт узнавания рестриктазы SacI, введенный в 5'-конец праймера. Праймер Р18 содержит последовательность, комплементарную последовательности гена pgl из Р. putida с 709 по 729 п.о., и сайт узнавания рестриктазы EcoRI, введенный в 5'-конец праймера.
Хромосомная ДНК штамма Р. putida KT2440 (АТСС 47054) (Bagdasarian, M.&Timmis, К.N., в Current Topics of Microbiology and Immunology, eds. Goebel, W.&Hofschneider, P.H. (Springer, Berlin), p.47-67 (1981)) была выделена стандартным способом. ПЦР производился на амплификаторе "Perkin Elmer GeneAmp PCR System 2400" при следующих условиях: 40 сек. при 95°С, 40 сек. при 53°С, 40 сек. при 72°С, 25 циклов с Taq полимеразой (Fermentas). Полученный ПЦР-фрагмент, содержащий ген pgl из Р.putida с RBS гена lacZ, был обработан рестриктазами SacI и EcoRI и клонирован в многокопийный вектор pUC19, предварительно обработанный теми же рестриктазами. Таким образом была получена плазмида pUC19-pgl.
Штамм BW25113-ΔybhE трансформировали полученной плазмидой pUC19-pgl. Трансформированную культуру высеяли на чашки с минимальной средой, содержащие мальтозу и ампициллин 100 мкг/мл, после чего обработали описанным выше раствором и проверили на "maltose blue" фенотип. Трансформанты не имели "maltose blue" фенотипа, в отличие от контрольного штамма BW25113-ΔybhE.
Таким образом, клонированная копия гена pgl из Pseudomonas putida комплементирует мутацию ybhE в Е.coli, еще раз подтверждая нашу гипотезу, что рамка ybhE ORF является кодирующей областью гена pgl.
Пример 7. Измерение активности 6-фосфоглюконолактоназы в штамме-мутанте по ybhE.
Ночные культуры штаммов BW25113 и BW25113-ΔybhE развели в 50 раз минимальной средой М9, содержащей глюкозу. Клетки выращивались до достижения культурой оптической плотности OD540=1. Экстракты клеток готовились из 3 мл культуры. Клетки отмывались физиологическим раствором, ресуспендировались в 400 мкл фосфатно-кальциевого буфера (рН 7.0) и обрабатывались ультразвуком. Фракция супернатанта, полученная после центрифугирования, использовалась для измерений без дополнительного разведения.
Для измерения активности 6-фосфоглюконолактоназы использовали методику, описанную у Collard, F. et al. (FEBS Letters, 459, 223-226 (1999)). Лактон синтезировали непосредственно в реакционной смеси, инкубируя 50 мкМ глюкозо-6-фосфата (Sigma, США) в присутствии 0.2 мМ NADP, 25 мМ HEPES (рН 7.1), 2 мМ MgCl2 и 1.75 ед дрожжевой глюкозо-6-фосфатдегидрогеназы (Sigma, США) при 30°С (общий объем реакционной смеси - 1 мл). Когда оптическая плотность реакционной смеси при А340 достигла плато, в реакционную смесь добавили смесь 0.5 ед/мл 6-фосфоглюконатдегидрогеназы (Sigma, США) с ранее полученными фракциями супернатанта, после чего в течение 10 минут измеряли оптическую плотность при А340. Количество белка измерялось способом, описанным у Bradford, M.M. (Anal. Biochem. 72, 248-254 (1976)). Полученные данные представлены в Таблице 1. Активность измерена в относительных единицах в пересчете на мг общего белка.
Как видно из Таблицы, активность 6-фосфоглюконолактоназы в штамме, мутантном по ybhE как минимум на порядок ниже, чем в природном штамме, и сравнима с уровнем спонтанного гидролиза.
Пример 8. Деления оперона zwf-edd-eda. Замена генов zwf-edd-eda на фрагмены ДНК, содержащие ген устойчивости к канамицину (KmR).
Для того чтобы получить штамм с усиленной экспрессией гена ybhE, было решено осуществить интеграцию с использованием Red-системы (смотри Пример 9) конститутивного промотора, производного от Рtac, между ybhE RBS и природным промотором этого гена.
Но попытка модифицировать таким образом хромосому природного штамма MG1655 оказалась неудачной. Мы не можем объяснить токсический эффект усиления экспрессии гена pgl (ybhE), но мы предполагаем, что это связано с повышением активности фосфоглюконолактоназы, что ведет к дисбалансу пентозофосфатного пути (Pentose-Phosphate Pathway (PPP)) и связанного с этим возможного накопления некоторых токсичных соединений (или недостаток некоторых веществ, необходимых клетке для выживания). Таким образом, мы решили полностью заблокировать пентозофосфатный цикл благодаря делеции гена zwf, кодирующего первый фермент этого пути.
Делецию оперона zwf-edd-eda производили способом, описанным в Примере 5 для гена pgi. Нуклеотидная последовательность природного оперона zwf-edd-eda приведена в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank (номера нуклеотидов с 1932863 по 1934338, gi:16129805; с 1930817 по 1932628, gi:16129804 и с 1930139 по 1930780, gi:16129803 для генов zwf, edd и eda соответственно). Фрагменты ДНК, содержащие ген KmR, были синтезированы с использованием ПЦР - праймеры Р19 (SEQ ID NO:25) и Р20 (SEQ ID NO:26). Праймер P19 содержит 36 нуклеотидов, комплементарных 3'-концу гена eda. Праймер Р20 содержит 36 нуклеотидов, комплементарных 5'-концу гена zwf. Нуклеотидная последовательность синтезированного фрагмента ДНК представлена в SEQ ID NO:27.
Колонии, выращенные в течение 24 часов, были протестированы на наличие маркера KmR вместо оперона zwf-edd-eda с помощью ПЦР - праймеры Р21 (SEQ ID NO:28) и Р22 (SEQ ID NO:29). Несколько протестированных KmR колоний содержали искомый фрагмент ДНК длиной 1287 п.о., подтверждающий замену природного оперона zwf-edd-eda на ДНК с маркером KmR. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем выращивания штамма при 37°С и полученный штамм был назван Е.coli BW25113-Δzwf-edd-eda. Структура фрагмента бактериальной ДНК, содержащей делецию оперона zwf-edd-eda показана на Фиг.6.
Пример 9. Замена природной области ДНК перед геном ybhE, расположенной на хромосоме Е.coli, новым регуляторным элементом, содержащим искусственный Ptac*-промотор.
Для дальнейшей интеграции перед геном pgl (ybhE) искусственных промоторов разной силы Ptac*, была проведена повторная трансформация штамма Е.coli BW25113-Δzwf-edd-eda плазмидой pKD46. Полученный штамм, устойчивый к канамицину и ампициллину был назван Е.coli BW25113-Δzwf-edd-eda [pKD46}. Поскольку плазмида pKD46 является термочувствительной, дальнейший отбор трансформантов производился при температуре 30°С.
Широко известен тот факт, что у мутантов с модифицированной "-35" областью промотора, распознаваемой комплексом Е.coli РНК полимеразы и фактора σ70, эффективность инициации транскрипции существенно изменена. Следовательно, среди промоторов, полученных в результате рандомизации промоторной области, могут быть получены промоторы с разной силой. Таким образом, этот общий подход может быть использован для тонкой настройки уровня экспрессии целевого гена. Авторами настоящего изобретения ранее была получена библиотека модифицированных промоторов Ptac различной силы (с этого места и далее такие модифицированные промоторы Ptac будут помечаться звездочкой). Эти промоторы отличаются друг от друга последовательностью 4 центральных нуклеотидов "-35" области. В настоящей работе были использованы два промотора Рtsc* с различной силой. Основываясь на значениях активности β-галактозидазы, экспрессируемой под контролем соответствующего промотора, они были названы Рtac-3900 (обычный Рtac) и Рtac-10000 (с центральными нуклеотидами TTGC вместо природных TGAC).
Далее, каждый из указанных искусственных промоторов Рtac* был интегрирован перед кодирующей областью гена pgl в хромосоме штамма Е.coli BW25113-Δzwf-edd-eda [pKD4-6] описанным выше способом (смотри Пример 2). Кроме того, перед промоторной областью был интегрирован также фрагмент ДНК с геном устойчивости к хлорамфениколу (CmR) (смотри Фиг.7).
Конструирование вышеупомянутых искусственных фрагментов ДНК, интегрированных в соответствующие области бактериальных хромосом, проводилось в несколько этапов. На первом этапе с помощью ПЦР был синтезирован фрагмент ДНК, содержащий сайт узнавания рестриктазы BglII перед соответствующим промотром Рtac*.
Хромосомные ДНК из штаммов Е.coli MG1655, содержащие интегрированные в хромосому искусственные промоторы Рtac-3900 и Рtac-10000, использовались для ПЦР в качестве матрицы. ПЦР проводился с использованием праймеров Р23 (SEQ ID NO:30) и Р24 (SEQ ID NO:31) для промотора Рtac-3000 и Рtac-10000 соответственно и праймера Р25 (SEQ ID NO:32) для обоих промоторов. Оба праймера Р23 и Р24 содержат сайт узнавания рестриктазы BglII, введенный в 5'-конец. Праймер Р25 содержит 11 нуклеотидов (включая RBS) перед геном pgl и первые 25 нуклеотидов кодирующей области гена pgl. Вышеупомянутые последовательности были введены в праймер Р25 для дальнейшей интеграции в бактериальную хромосому.
ПЦР проводили с использованием амплификатора "TermoHybaid PCR Express". Реакционная смесь общим объемом 50 мкл содержала 5 мкл 10х буфера для ПЦР ("Ферментас", Литва) с добавлением MgCl2 до конечной концентрации в реакционной смеси 15 мМ, 200 мкМ каждого трифосфата dNTP, 25 пМ каждого из используемых праймеров и 1 ед Taq-полимеразы ("Ферментас", Литва). Примерно 0,5 мкг плазмидной ДНК добавляли в реакционную смесь в качестве матрицы для ПЦР амплификации. Температурный профиль ПЦР был следующий: начальная стадия денатурации в течение 5 мин при 95°С с последующими 25 циклами денатурации при 95°С в течение 30 секунд, отжига при 53°С в течение 30 секунд, наращивания цепи при 72°С в течение 30 секунд; финальная стадия полимеризации при 72°С в течение 7 минут.
Далее был проведен второй этап конструирования целевого фрагмента ДНК. Ген CmR был амплифицирован с помощью ПЦР, в качестве матрицы использовалась коммерчески доступная плазмида pACYC184 (инвентарный номер Х06403 в базе данных GenBank/EMBL, "Ферментас", Литва) и праймеры Р26 (SEQ ID NO:33) и Р27 (SEQ ID NO:34). Праймер Р26 содержит сайт узнавания рестриктазы BglII, необходимый для дальнейшего присоединения данного фрагмента ДНК к ранее полученному фрагменту, содержащему промотор Рtac*. Праймер Р27 содержит 46 нуклеотидов, комплементарных нуклеотидам с 58 по 12, расположенным перед старт-кодоном гена pgl (ybhE) на хромосоме Е.coli, эта комплементарная область необходима для дальнейшей интеграции полученного фрагмента в бактериальную хромосому.
Амплифицированные фрагменты ДНК концентрировали электрофорезом в агарозном геле, выделяли из геля с помощью "GenElute Spin Columns" ("Sigma", США) и высаживали этанолом. Затем оба полученных фрагмента ДНК обработали эндонуклеазой BglII, после чего лигировали с использованием Т4 ДНК лигазы (Maniatis Т., Fritsch E.F., Sambrook, J.: Molecular Cloning: A Laboratory Manual. 2nd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1989).
Лигированный продукт был амплифицирован с помощью ПЦР, при этом использовались праймеры Р25 и Р27. Реакционная смесь общим объемом 50 мкл содержала 5 мкл 10х буфера для ПЦР ("Sigma", США), 200 мкМ каждого трифосфата dNTP, 25 пМ каждого из используемых праймеров и 1 ед AccuTaq LA-полимеразы ("Sigma", США). Примерно 50 нг плазмидной ДНК добавляли в реакционную смесь в качестве матрицы для ПЦР амплификации. Температурный профиль ПЦР был следующий: начальная стадия денатурации в течение 5 мин при 95°С с последующими 25 циклами денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд, наращивания цепи при 72°С в течение 4 минут; финальная стадия полимеризации при 72°С в течение 7 минут.
Нуклеотидная последовательность полученного фрагмента ДНК представлена в последовательностях SEQ ID NO:35 и 36 для промоторов Рtac-3900 и Рtac-10000 соответственно.
Полученные фрагменты ДНК, очищенные описанным выше способом, были использованы для электропорации и Red-зависимой интеграции в бактериальную хромомсому штамма Е.coli BW25113Δzwf- -eda [pKD46], как описано в Примере 2.
Колонии, выращенные в течение 24 часов в среде с хлорамфениколом, были протестированы на наличие маркера CmR перед геном pgl с помощью ПЦР - праймеры Р27 (SEQ ID NO:34) и P10 (SEQ ID NO:15). Эти же колонии были также протестированы с помощью ПЦР на наличие промоторной области Рtac* перед геном pgl - праймеры Р23 (SEQ ID NO:30) и Р24 (SEQ ID NO:31) для Ptac-3900 и Рtac-10000 соответственно и Р10 (SEQ ID NO:15). Для этой цели свежевыращенные колонии ресуспендировались в 20 мкл воды и 1 мкл полученной суспензии использовали в качестве матрицы для ПЦР. Температурный профиль ПЦР был следующий: начальная стадия денатурации ДНК - в течение 10 мин при 95°С, затем 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 54°С в течение 30 секунд, наращивания цепи при 72°С в течение 1 минуты; финальная стадия полимеризации при 72°С в течение 7 минут. Несколько протестированных CmR колоний содержали искомые фрагменты ДНК длиной 1192 п.о. и 124 п.о., подтверждающие наличие как всего сконструированного фрагмента ДНК перед геном pgl, и гибридного регуляторного элемента, содержащего промотор Рtac* в тестируемых хромосомах штаммов Е.coli. В обоих случаях, один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем выращивания штамма при 37°С и полученные штаммы были названы Е.coli BW25113-Ptac-3900-ybhE и BW25113-Ptac-10000-ybhE соответственно. Структура сконструированного фрагмента бактериальной ДНК перед геном pgl показана на Фиг.7.
Пример 10. Измерение активности 6-фосфоглюконолактоназы в штаммах с усиленной экспрессией гена pgl.
Активность 6-фосфоглюконолактоназы в штаммах BW25113-Ptac-3900-ybhE и BW25113-Рtac-10000-ybhE была измерена способом, описанным в Примере 7. Полученные данные приведены в Таблице 2. При измерениях активности уровень спонтанного гидролиза вычитался.
Таким образом, усиленная экспрессия гена pgl приводит к повышению активности 6-фосфоглюконолактоназы.
Пример 11. Эффект усиления экспрессии гена pgl на продукцию триптофана.
В качестве родительского штамма для оценки эффекта влияния усиления экспрессии гена pgl на продукцию триптофана был использован штамм-продуцент L-триптофана Е.coli SV164 [pMW-PlacUV5-serA5-fruR, pYDDG2]. Штамм SV164 детально описан в патенте США 6180373. Штамм SV164 [pMW-PlacUV5-serA5-fruR, pYDDG2] является производным от штамма SV164, он дополнительно содержит плазмиды pMW-PlacUV5-serA5-fruR и pYDDG2. Плазмида pMW-PtacUV5-serA5-fruR содержит мутантный ген serA5, кодирующий белок, не ингибируемый серином по типу обратной связи. Амплификация гена serA5 необходима для увеличения пула серина, предшественника L-триптофана (патент США 6180373). Плазмида pYDDG2 сконструирована на базе вектора pAYCTER3 (WO 03/044192) и содержит ген yddG, кодирующий трансмембранный белок (возможный экспортер) положительно влияющий на продукцию L-риптофана. Вектор pAYCTER3 является производным вектора pAYC32, являющегося низкокопийным и очень стабильным вектором, сконструированным на основе плазмиды RSF1010 и содержащим маркер устойчивости к стрептомицину (Christoserdov A.Y., Tsygankov Y.D, Broad-host range vectors derived from a RSF 1010 Tnl plasmid, Plasmid, 1986, v.16, p.161-167). Вектор pAYCTER3 был получен введением в плазмиду pAYC32 полилинкера из плазмиды pUC19 и сильного терминатора rrnB вместо промотора.
Для оценки эффекта влияния на продукцию триптофана усиленной экспрессии гена pgl, находящегося под контролем промоторов Ptac*, фрагменты ДНК из хромосомы вышеупомянутых штаммов Е.coli BW25113-Ptac-3900-ybhE и BW25113-Рtac-10000-ybhE с помощью Р1трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY) были перенесены в штамм-продуцент триптофана Е. Coli SV164 [pMW-PlacUV5-serA5-fruR]. Затем, в полученные трансдуктанты и в штамм SV164 [pMW-PlacUV5-serA5-fruR] ввели плазмиду pYDDG2.
Штаммы SV164 [pMW-Plac-serA5-fruR, pYDDG2],
S164-Ptac-3900-ybhE [pMW-PlacUV5-serA5-fruR, pYDDG2] и
SV164-Ptac-10000-ybhE [pMW-PlacUV5-serA5-fruR, pYDDG2] выращивались в течение ночи на роторной качалке при 37°С в 3 мл питательного бульона, содержащего ампициллин (100 мкг/мл) и стрептомицин (50 мкг/мл). По 0.3 мл полученных культур были инокулированы в пробирки 20×200 мм с 3 мл ферментационной среды, содержащей указанные антибиотики, с выращивались при 37°С в течение 40 часов на роторной качалке при 250 об/мин. Состав среды для ферментации представлен в Таблице 3.
Значение рН 7.1 во фракции А доводилось с помощью NH4OH. Каждую фракцию стерилизовали отдельно.
После выращивания количество накопленного в среде L-триптофана определялось с помощью тонкослойной хроматографии (TLC). Использовались пластинки TLC размером 10×15 см с нанесенными слоями силикатного геля Sorbfil без флуоресцентного индикатора толщиной 0.11 мм (Акционерное Общество Сорбполимер, Краснодар, Россия). Пластинки Sorbfil экспонировались в жидкой фазе: пропан-2-ол: этилацетат: 25%-ный водный раствор аммиака: вода=16:16:3:9 (объемные доли). Раствор (2%) нингидрина в ацетоне использовался в качестве агента для визуализации. Полученные данные представлены в Таблице 4.
Как видно из Таблицы 4, усиление экспрессии гена pgl положительно сказывается на способности штамма SV164 [pMW-PtacUV5-serA5-fruR, pYDDG2] к продукции триптофана.
Пример 12. Выделение белка YbhE с концевой His6 группой и определение его активности.
Все результаты, описанные в Примерах 1-7, могут служить только косвенным доказательством того, что ybhE ORF является геном pgl, кодирующим функционально активный белок 6-фосфоглюконолактоназу в Е.coli. С другой стороны, существует вероятность того, что ybhE ORF кодирует, например, позитивный регулятор экспрессии другого неизвестного гена, который, в свою очередь, и является 6-фосфоглюконолактоназой. Таким образом, окончательный вывод относительно природы ybhE ORF может быть сделан лишь на основе прямого измерения биологической активности белкового продукта.
Для этой цели ybhE ORF была экспрессирована с использованием системы экспрессии фага Т7, штамма Е.coli BL21 (DE3) в качестве штамма-реципиента, содержащего в хромосоме ген РНК полимеразы фага Т7, и плазмидного вектора рЕТ-22b (+) с поздним промотором фага Т7 и эффективным сайтом связывания рибосомы (RBS) гена 10 фага Т7. Для удобства последующей очистки белка 6 гистидиновых кодонов были введены на 5'-конец, ybhE ORF, сразу после инициаторного кодона ATG.
Для клонирования ybhE ORF с добавкой 6 гистидиновых кодонов в экспрессирующей системе фага Т7 был использован метод ПЦР с праймерами Р28 (SEQ ID NO:37) и Р29 (SEQ ID NO:38) и хромосомной ДНК штамма Е.coli MG1655 в качестве матрицы. Праймер Р28 содержит сайт рестрикции NdeI, ATG-кодон, соединенный с 6 дополнительными гистидиновыми кодонами перед вторым кодоном ybhE ORF. Праймер Р29 содержит на 5'-конце сайт рестрикции BamHI, необходимый для дальнейшего клонирования. Амплифицированный фрагмент ДНК был выделен, обработан рестриктазами NdeI и BamHI и лигирован в плазмиды рЕТ-22b (+), предварительно обработанную этими же рестриктазами. Конструкция полученной плазмиды pET-HT-ybhE была подтверждена путем определения нуклеотидной последовательности.
Затем клетки BL21 (DE3), содержащие в хромосоме ген РНК полимеразы фага Т7 под контролем лактозного промотора, были трансформированы плазмидой pET-HT-ybhE. Ночная культура, полученная от одной колонии, была разбавлена в 50 раз средой LB и выращивалась до достижения оптической плотности OD600˜1.0 с последующим добавлением IPTG (1 мМ) для индукции посредством РНК-полимеразы фага Т7 ybhE ORF в полученной рекомбинантной плазмиде. После инкубирования в течение 2 часов отбирались клетки в объеме 20 мл. Клеточный экстракт был получен путем обработки ультразвуком в буфере, содержащем 20 мМ Tris-HCl, pH 8.0 и 2 мМ PMSF. Затем образцы центрифугировали в течение 20 минут при 16,000 g и 4°С и выделяли из супернатанта белок с гистидиновой добавкой с использованием колонок HiTrap Chelating Columns (Amersham Bioscience) в соответствии с рекомендациями производителем.
По истечении 2 часов после индукции системы экспрессии фага Т7 в культуре в логарифмической фазе роста наблюдалось накопление белка с электрофоретической подвижностью, соответствующей подвижности белка YbhE с добавкой 6 гистидиновых остатков (молекулярный вес ≈37 кДа). Количество указанного белка составляло порядка 15% от общего количества полипептидов клетки. Указанный белок обнаруживался главным образом в растворимой фазе (см. Фиг.8А).
Полученный белок His6-YbhE был очищен на колонках Ni-NTA. Определение уровня синтеза рекомбинантного белка и контроль процесса очистки проводили с использованием SDS-PAGE электрофореза в соответствии с методом, описанным Laemmli U.K. (Nature, 227, 680-685 (1970)). Как видно из Фиг.8Б чистота полученного белка составляла более 90%, при этом белок проявлял активность 6-PGL в стандартном тесте с использованием латоназы (Collard, F. et al, FEBS Letters, 459, 223-226 (1999)); Пример 7). Интересно отметить, что измеренная удельная активность 6-PGL у очищенного белка His6-YbhE (780 U/мг) была близкой к ранее описанной активности аналогично модифицированного белка His6-6-PGL человека (Collard, F. et al, FEBS Letters, 459, 223-226 (1999)).
Таким образом был сделан вывод о том, что ybhE ORF с неизвестной функцией из Е.coli в действительности является геном pgl, кодирующим 6-PGL.
Пример 13. Эффект усиления экспрессии гена pgl на продукцию фенилаланина.
Штамм Е.coli AJ12739 - продуцент фенилаланина был использован в качестве исходного штамма для оценки эффекта усиления экспрессии гена pgl на продукцию фенилаланина. Штамм AJ12739 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 113545 Москва, 1-й Дорожный проезд, 1) 6 ноября 2001 года под инвентарным номером ВКПМ В-8197.
Фрагменты хромосомной ДНК из штаммов BW25113-Ptac-3900-ybhE и BW25113-Ptac-10000-ybhE были перенесены в штамм Е.coli AJ12739 - продуцент фенилаланина Р1 трансдукцией с получением штаммов AJ12739 Ptac-3900-ybhE и AJ12739 Рtac-10000 соответственно. Каждый из этих штаммов выращивался при 37°С в течение 18 часов в питательном бульоне, содержащем 25 мг/мл хлорамфеникола, 0.3 мл полученной культуры было перенесено в 3 мл питательной среды для ферментации, содержащей 25 мг/мл хлорамфеникола, в пробирке 20×200 мм и инкубировалось при 34°С в течение 24 часов на роторной качалке. После выращивания, количество фенилаланина, накопленное в среде, определялось методом ТСХ. Использовались пластинки для ТСХ размером 10×15 см, покрытые слоем в 0.11 мм силикагелем Sorbfil без флюоресцентного индикатора (Компания "Сорбполимер", Краснодар, РФ). Пластинки Sorbfil экспонировались с подвижной фазой следующего состава: пропан-2-ол:этилацетат:водный аммиак (25%):вода=40:40:7:16 (v/v). В качестве реагента для визуализации использовался 2%-ный раствор нингидрина в ацетоне.
Состав среды для ферментации (г/л):
Глюкозу и сульфат магния стерилизовали раздельно. СаСО3 стерилизовали при 180°С в течение 2 часов. рН поддерживался в районе 7.0. Антибиотик добавляли в питательную среду после стерилизации.
Результаты представлены в Таблице 5.
Как видно из Таблицы 5, усиление экспрессии гена pgl повышало способность штамма АJ12739 продукции фенилаланина.
Хотя указанное изобретение описано в деталях со ссылкой на конкретные Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения. Каждый из упомянутых выше документов, цитируемых в описании, является частью настоящего описания.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ОБЛАДАЮЩИХ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНА pckA | 2003 |
|
RU2268300C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ С ОПТИМИЗИРОВАННЫМ УРОВНЕМ ГЕННОЙ ЭКСПРЕССИИ | 2003 |
|
RU2268305C2 |
| СПОСОБ ПРОДУКЦИИ АРОМАТИЧЕСКИХ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Methylophilus | 2005 |
|
RU2333950C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2007 |
|
RU2364628C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA, - ПРОДУЦЕНТ L-ЦИСТЕИНА И СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА | 2003 |
|
RU2275425C2 |
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
Изобретение относится к биотехнологии, L-аминокислоты, такие как L-триптофан, L-фенилаланин, L-тирозин и L-гистидин получают культивированием бактерии рода Escherichia, модифицированной таким образом, что активность 6-фосфоглюконолактоназы в ней повышена. Заявленное изобретение позволяет получать указанные аминокислоты с высокой степенью эффективности. 4 н. и 11 з.п. ф-лы, 5 табл., 8 ил.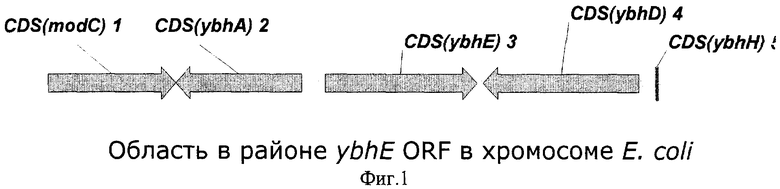
(A) белок, включающий последовательность аминокислот, приведенную в списке последовательностей под номером 2 (SEQ ID NO:2); и
(B) белок, включающий последовательность аминокислот, содержащую делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), который обладает как минимум 70%-ной гомологией по отношению к аминокислотной последовательности, приведенной под номером 2 (SEQ ID NO:2), и который обладает активностью 6-фосфоглюконолактоназы.
(a) ДНК, включающая последовательность нуклеотидов с 1 по 996, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1);
(b) ДНК, которая гибридизуется в жестких условиях с последовательностью нуклеотидов с 1 по 996, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1), или с зондом, приготовленным на основании указанной последовательности нуклеотидов, и кодирует белок, обладающий активностью 6-фосфоглюконолактоназы.
| MICLET Е | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| J Biol Chem | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |

