Область техники
Настоящее изобретение относится к микробиологической промышленности, конкретно к способу получения L-аминокислоты, такой как L-треонин, L-лизин, L-гистидин, L-фенилаланин, L-аргинин, L-триптофан, L-глутаминовая кислота и L-лейцин путем ферментации с использованием бактерии с повышенной активностью алкогольдегидрогеназы.
Описание предшествующего уровня техники
Традиционно L-аминокислоты получают в промышленном масштабе методами ферментации с использованием штаммов микроорганизмов, выделенных из природных источников, или их мутантов. В большинстве случаев микроорганизмы модифицируют таким образом, чтобы увеличить производственный выход L-аминокислот.
Описано много технологий для увеличения производственного выхода L-аминокислот, включая трансформацию микроорганизмов рекомбинантной ДНК (см., например, патент США US 4,278,765). Другие технологии увеличения производственного выхода включают увеличение активности ферментов, участвующих в биосинтезе аминокислот и/или снижение чувствительности ферментов-мишеней к ингибированию по типу обратной связи образующейся L-аминокислотой (см., например, патенты США 4,346,170; 5,661.012 и 6,040,160).
Для оптимизации главного пути биосинтеза необходимого компонента может быть выполнено дальнейшее улучшение штаммов-продуцентов L-аминокислот. Обычно это может быть достигнуто путем внесения в питательную среду бактерий повышенного количества источника углерода, такого как сахар, например глюкоза. Несмотря на то, что система PTS обеспечивает эффективный транспорт глюкозы в клетку, для высокопродуктивного штамма доступность этого источника углерода может быть недостаточной. Другим способом увеличения продуктивности штаммов-продуцентов L-аминокислоты и уменьшения стоимости целевой L-аминокислоты является использование альтернативных источников углерода, таких как спирты, в частности этанол.
Алкогольдегидрогеназа (этанолоксидоредуктаза, AdhE) бактерии Escherichia coli является многофункциональным ферментом, который катализирует ферментативный синтез этанола путем двух последовательных реакций NADH-зависимого восстановления ацетил-коэнзима А, а также дезактивирует пируватформиатлиазу, которая расщепляет пируват с образованием ацетил-коэнзима А и формиата.
AdhE синтезируется в больших количествах (примерно 3·104 копий на клетку) при анаэробном росте бактерий в среде, содержащей глюкозу, и образует спиральные структуры, называемые спиросомами, длиной примерно 0.22 мкм, которые содержат 40-60 молекул AdhE (Kessler, D., Herth, W., and Knappe, J., J. Biol. Chem., 267, 18073-18079 (1992)). Когда клетки Е. coli переводят из анаэробных условий культивирования в аэробные, транскрипция гена adhE уменьшается и поддерживатся в пределах 10% от уровня, наблюдаемого в анаэробных условиях (Chen, Y. М., and Lin, Е. С.С., J. Bacteriol. 173, 8009-8013 (1991); Leonardo, М. R., Cunningham, P. R., and dark, D. P., J. Bacteriol. 175, 870-878 (1993); Mikulskis, A., Aristarkhov, A., and Lin, Е. C. C., J. Bacteriol. 179, 7129-7134 (1997); Membrillo-Hemandez, J., and Lin, Е. C. C., J. Bacteriol. 181, 7571-7579 (1999)). Трансляция мРНК AdhE также является регулируемой, для чего необходима РНКаза III (Membrillo-Hernandez, J., and Lin, Е. С.С., J. Bacteriol. 181, 7571-7579 (1999); Aristarkhov, A. et al, J. Bacteriol. 178, 4327-4332 (1996)). Было также обнаружено, что AdhE является основной мишенью при стрессе, вызванном перекисью водорода (Tamarit, J., Cabiscol, E., and Ros, J., J. Biol. Chem. 273, 3027-3032 (1998)).
Несмотря на обратимый характер двух NADH-зависимых реакций, катализируемых AdhE, выделенные из природных источников штаммы Е. coli не обладают способностью к росту на этаноле, когда он используется в качестве единственного источника углерода и энергии, поскольку в аэробных условиях понижается транскрипция гена adhE (Chen, Y. М. and Lin, Е. С.С., J. Bacteriol. 73, 8009-8013 (1991); Leonardo, М. R., Cunningham, P. R. & Clark, D. P., J. Bacteriol. 175 870-878 (1993)), тогда как период полураспада белка AdhE уменьшается при аэробном метаболизме за счет катализируемого металлами окисления [metal-catalyzed oxidation (MCO)].
Мутантные штаммы Е. coli, способные расти в аэробных условиях на этаноле, который используется в качестве единственного источника углерода и энергии, были получены и описаны ранее [мутанты с заменой Ala267Thr растут на этаноле со скоростью удвоения 240 мин; мутанты с заменами Ala267Thr и Glu568Lys - со скоростью удвоения 90 мин при 37°С) (Membrillo-Hemandez, J. et al, J. Biol. Chem. 275, 33869-33875 (2000); Holland-Staley, С.A. et al, J. Bacteriol. 182, 6049-6054 (2000))]. Очевидно, что при катализе двух последовательных реакций в направлении, противоположном физиологическому, синтез ацетил-коэнзима А становится лимитирующей стадией для белка AdhE природного типа. В противовес увеличению скорости реакции Vmax при замене А267Т в белке AdhE снижается термостабильность этого фермента и повышается его чувствительность к повреждению окислением, катализируемым металлами [МСО]. Вторая аминокислотная замена, Е568К, в AdhE (A267T/E568K) частично восстанавливала стабильность белка и устойчивость к повреждению МСО без улучшения каталитической активности при окислении субстрата.
Однако в настоящее время отсутствуют сообщения об использовании бактерий семейства Enterobacteriaceae с повышенной активностью как нативной алкогольдегидрогеназы, так и мутантной алкогольдегидрогеназы, устойчивой к аэробной инактивации, для целей увеличения продуцирования ими L-аминокислот путем ферментации в питательной среде, содержащей этанол.
Описание изобретения
Цели настоящего изобретения включают повышение продуктивности штаммов-продуцентов L-аминокислоты и предоставление способа получения неароматических или ароматических L-аминокислот с использованием указанных штаммов.
Данная цель была достигнута путем обнаружения того факта, что экспрессия природного или мутантного гена adhE, кодирующего алкогольдегидрогеназу, под контролем промотора, функционирующего в аэробных условиях культивирования, приводит к увеличению выработки L-аминокислот, таких как L-треонин, L-лизин, L-гистидин, L-фенилаланин, L-аргинин, L-триптофан, L-глутаминовая кислота и/или L-лейцин.
Целью настоящего изобретения является предоставление мутантной алкогольдегидрогеназы, устойчивой к аэробной инактивации, выбранной из группы, состоящей из:
(А) белка, включающего аминокислотную последовательность, представленную в SEQ ID NO: 2, за тем исключением, что указанный белок содержит мутацию в положении 568 в SEQ ID NO: 2, и содержащего по крайней мере одну дополнительную мутацию, способствующую улучшению роста бактерии в жидкой среде, содержащей этанол в качестве единственного источника углерода, отличающейся тем, что указанная дополнительная мутация находится в положении, отличном от положения 267 в SEQ ID NO: 2; и
Б) варианта белка (А), обладающего активностью алкогольдегидрогеназы. Также целью настоящего изобретения является предоставление мутантной алкогольдегидрогеназы, описанной выше, отличающейся тем, что указанная дополнительная мутация выбрана из группы, состоящей из:
A) замены остатка глутаминовой кислоты в положении 560 в SEQ ID NO: 2 на остаток другой аминокислоты;
Б) замены остатка фенилаланина в положении 566 в SEQ ID NO: 2 на остаток другой аминокислоты;
B) замены остатка глутаминовой кислоты, остатка метионина, остатка тирозина, остатка изолейцина и остатка аланина в положениях 22, 236, 461, 554 и 786 соответственно в SEQ ID NO: 2 на остатки других аминокислот; и
Г) комбинации этих замен.
Также целью настоящего изобретения является предоставление мутантной алкогольдегидрогеназы, описанной выше, отличающейся тем, что указанная дополнительная мутация выбрана из группы, состоящей из:
A) замены остатка глутаминовой кислоты в положении 560 в SEQ ID NO: 2 на остаток лизина;
Б) замены остатка фенилаланина в положении 566 в SEQ ID NO: 2 на остаток валина;
B) замены остатка глутаминовой кислоты, остатка метионина, остатка тирозина, остатка изолейцина и остатка аланина в положениях 22, 236, 461, 554 и 786 в SEQ ID NO: 2 на остаток глицина, остаток валина, остаток цистеина, остаток серина и остаток валина соответственно; и
Г) комбинации этих замен.
Также целью настоящего изобретения является предоставление фрагмента ДНК, кодирующего мутантную алкогольдегидрогеназу, описанную выше.
Также целью настоящего изобретения является предоставление бактерии-продуцента L-аминокислоты семейства Enterobacteriaceae, содержащей мутантную алкогольдегидрогеназу, описанную выше.
Также целью настоящего изобретения является предоставление бактерии-продуцента L-аминокислоты семейства Enterobacteriaceae, отличающейся тем, что указанная бактерия модифицирована таким образом, что активность мутантной алкогольдегидрогеназы, описанной выше, в этой бактерии увеличена.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что активность мутантной алкогольдегидрогеназы в этой бактерии увеличена путем усиления экспрессии гена, кодирующего указанную мутантную алкогольдегидрогеназу.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что активность указанной мутантной алкогольдегидрогеназы в этой бактерии увеличена путем модификации последовательности, контролирующей экспрессию гена, кодирующего указанную алкогольдегидрогеназу, или путем увеличения количества копий гена, кодирующего указанную алкогольдегидрогеназу.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия выбрана из группы, состоящей из Escherichia. Enterobacter, Erwinia, Klebsiella, Pantoea, Providencia, Salmonella, Serratia, Shigella и Morganella.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-треонина.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия дополнительно модифицирована таким образом, что у этой бактерии усилена экспрессия гена, выбранного из группы, состоящей из:
- мутантного гена thrA, кодирующего аспартокиназу-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи;
- гена thrB, кодирующего гомосеринкиназу;
- гена thrC, кодирующего треонинсинтазу;
- гена rhiA, кодирующего предполагаемый трансмембранный белок;
- гена asd, кодирующего аспартат-β-полуальдегиддегидрогеназу;
- гена aspC, кодирующего аспартатаминотрансферазу (аспартаттрансаминазу); и
- комбинации этих генов.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия модифицирована таким образом, что экспрессия указанного мутантного гена thrA, указанного гена thrB, указанного гена thrC и указанного гена rhtA в этой бактерии усилена.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-лизина.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-гистидина.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-фенилаланина.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-аргинина.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-триптофана.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-глутаминовой кислоты.
Также целью настоящего изобретения является предоставление бактерии, описанной выше, отличающейся тем, что указанная бактерия является продуцентом L-лейцина.
Также целью настоящего изобретения является предоставление способа получения L-аминокислоты, включающего:
А) выращивание в питательной среде, содержащей этанол, бактерии семейства Enterobacteriaceae - продуцента L-аминокислоты, содержащей алкогольдегидрогеназу; и
Б) выделение L-аминокислоты из культуральной жидкости,
причем ген, кодирующий указанную алкогольдегидрогеназу, экспрессируется под контролем ненативного промотора, функционирующего в аэробных условиях культивирования.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанный ненативный промотор выбран из группы, состоящей из Ptac, Plac, Ptrc, PR и PL.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная алкогольдегидрогеназа устойчива к аэробной инактивации.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная алкогольдегидрогеназа синтезируется бактерией, выбранной из группы, состоящей из Escherichia coli, Erwinia carotovora, Salmonella typhimurium, Shigella flexneri, Yersmia pestis, Pantoea ananatis, Lactobacillus plantarum и Lactococctis lactis.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная алкогольдегидрогеназа содержит аминокислотную последовательность, представленную в SEQ ID NO: 2, за исключением остатка глутаминовой кислоты в положении 568, замещенного остатком другой аминокислоты, иным, чем остаток аспарагиновой кислоты.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная алкогольдегидрогеназа содержит аминокислотную последовательность, представленную в SEQ ID NO: 2 за исключением остатка глутаминовой кислоты в положении 568, замещенного остатком лизина.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная алкогольдегидрогеназа содержит по крайней мере одну дополнительную мутацию, способствующую улучшению роста указанной бактерии в жидкой среде, содержащей этанол в качестве единственного источника углерода.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная дополнительная мутация выбрана из группы, состоящей из:
А) замены остатка глутаминовой кислоты в положении 560 в SEQ ID NO: 2 на остаток другой аминокислоты;
Б) замены остатка фенилаланина в положении 566 в SEQ ID NO: 2 на остаток другой аминокислоты;
В) замены остатка глутаминовой кислоты, остатка метионина, остатка тирозина, остатка изолейцина и остатка аланина в положениях 22, 236, 461, 554 и 786 соответственно в SEQ ID NO: 2 на остаток другой аминокислоты; и
Г) комбинации этих замен.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная дополнительная мутация выбрана из группы, состоящей из:
A) замены остатка глутаминовой кислоты в положении 560 в SEQ ID NO: 2 на остаток лизина;
Б) замены остатка фенилаланина в положении 566 в SEQ ID NO: 2 на остаток валина;
B) замены остатка глутаминовой кислоты, остатка метионина, остатка тирозина, остатка изолейцина и остатка аланина в положениях 22, 236, 461, 554 и 786 в SEQ ID NO: 2 на остатки глицина, валина, цистеина, серина и валина соответственно; и
Г) комбинации этих замен.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная бактерия принадлежит к роду, выбранному из группы, состоящей из Escherichia, Enterobacter, Erwinia, Klebsiella, Pantoea, Providencia, Salmonella, Serratia, Shigella и Morganella.
Также целью настоящего изобретения является предоставление способа, описанного выше, отличающегося тем, что указанная L-аминокислота выбрана из группы, состоящей из L-треонина, L-лизина, L-гистидина, L-фенилаланина, L-аргинина, L-триптофана, L-глутаминовой кислоты и L-лейцина.
Подробное описание наилучшего способа осуществления изобретения
Алкогольдегидрогеназа является Fe2+-зависимым многофункциональным белком с активностью ацетальдегид-СоА-дегидрогеназы на N-концевом участке, железо-зависимой алкогольдегидрогеназы на С-концевом участке и пируват-формиатлиазы-дезактивазы. Синонимы включают В 1241, AdhC, Ana. В аэробных условиях период полураспада активного белка AdhE уменьшается за счет катализируемого металлами окисления при аэробном метаболизме.
Фраза «мутантная алкогольдегидрогеназа, устойчивая к аэробной инактивации» означает, что мутантная алкогольдегидрогеназа сохраняет свою активность в аэробных условиях или что ее активность в аэробных условиях снижается незначительно по сравнению со снижением активности алкогольдегидрогеназы природного типа.
Фраза «активность алкогольдегидрогеназы» означает способность фермента катализировать реакцию NAD-зависимого окисления спиртов с образованием альдегидов или кетонов. Алкогольдегидрогеназа (ЕС 1.1.1.1) обладает активностью по отношению к этанолу, н-пропанолу и н-бутанолу. Активность алкогольдегидрогеназы может быть обнаружена и измерена, например, методом, описанным Membrillo-Hemandez, J. и соавт.(J. Biol. Chem. 275, 33869-33875 (2000)).
Алкогольдегидрогеназа кодируется геном adhE, и в качестве гена алкогольдегидрогеназы в настоящем изобретении может быть использован любой ген adhE бактерий, принадлежащих к родам Escherichia, Erwinia, Klebsiella, Salmonella, Shigella, Yershinia, Pantoea, Lactobacillus и Lactococcus. Конкретные примеры источника гена adhE включают такие бактериальные штаммы, как Escherichia coli, Erwinia carotovora, Salmonella enterica. Salmonella typhimurium, Shigella flexneri, Yersinia pseudotuberculosis, Pantoea ananatis, Lactobacillus plantarum и Lactococcus lactis. Ген adhE дикого типа, кодирующий алкогольдегидрогеназу из Escherichia coli, изучен (нуклеотиды, комплементарные нуклеотидам с 1294669 по 1297344 в последовательности с инвентарным номером NC_000913.2, gi: 49175990 в базе данных GenBank). Ген adhE расположен между открытыми рамками считывания ychG и ychE на хромосоме штамма E.coli K-12. Другие гены adhE, кодирующие алкогольдегидрогеназу, были также расшифрованы: ген adhE Erwinia carotovora (нуклеотиды с 2634501 по 2637176 в последовательности с инвентарным номером NC_004547.2; gi: 50121254 в базе данных GenBank); ген adhE Salmonella enterica (нуклеотиды с 1718612 по 1721290 в последовательности с инвентарным номером NC_004631.1; gi: 29142095 в базе данных GenBank); ген adhE из Salmonella typhimurium (нуклеотиды с 1 по 2637 в последовательности с инвентарным номером U68173.1; gi: 1519723 в базе данных GenBank); ген adhE из Shigella flexneri (нуклеотиды, комплементарные нуклеотидам с 1290816 по 129349 в последовательности с инвентарным номером NC_004741.1, gi: 30062760 в базе данных GenBank); ген adhE из Yersinia pseudotuberculosis (нуклеотиды, комплементарные нуклеотидам с 2478099 по 2480774 в последовательности с инвентарным номером NC_006155.1; gi: 51596429 в базе данных enBank); ген adhE из Pantoea ananatis (SEQ ID NO: 29); ген adhE из Lactobaccillus plantarum (UniProtKB Entry: Q88RY9_LACPL); ген adhE из Lactococcus lactis MG1363 (инвентарный номер AJ001007 в EMBL) и т.п. (см. Фиг.2). Нуклеотидная последовательность гена adhE из Escherichia coli представлена в Перечне последовательностей под номером SEQ ID NO. 1. Аминокислотная последовательность, кодируемая этим геном adhE, представлена в Перечне последовательностей под номером SEQ ID NO.2.
Следовательно, ген adhE может быть получен методом ПЦР (полимеразная цепная реакция; White, T.J. et al,. Trends Genet., 5, 185 (1989)) с использованием праймеров, сконструированных на основе известной нуклеотидной последовательности гена хромосомы Е. coli. Гены других организмов, кодирующие алкогольдегидрогеназу, могут быть получены сходным образом.
Ген adhE из Escherichia coli представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 2; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 2, который обладает активностью алкогольдегидрогеназы.
Ген adhE из Pantoea ananatis представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б);
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 30; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 30, который обладает активностью алкогольдегидрогеназы.
Ген adhE из Shigella flexneri представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 53; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 53, который обладает активностью алкогольдегидрогеназы.
Ген adhE из Yersinia pestis представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 54; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 54, который обладает активностью алкогольдегидрогеназы.
Ген adhE из Erwinia carotovora представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 55; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 55, который обладает активностью алкогольдегидрогеназы.
Ген adhE из Salmonella typhimurium представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 56; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 56, который обладает активностью алкогольдегидрогеназы.
Ген adhE из Lactobacillus plantarum представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 57; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 57, который обладает активностью алкогольдегидрогеназы.
Ген adhE Lactobacillus lactis представлен фрагментом ДНК, который кодирует следующий белок (А) или (Б):
(А) белок, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 58; или
(Б) вариант белка с аминокислотной последовательностью, показанной в SEQ ID NO: 58, который обладает активностью алкогольдегидрогеназы.
Термин «вариант белка» в значении, в котором он используется в настоящем изобретении, означает белок, имеющий изменения в аминокислотной последовательности, а именно делеции, вставки, добавления или замены аминокислот, при условии, что при этом сохраняется необходимый уровень активности алкогольдегидрогеназы. Количество изменений в варианте белка зависит от положения в трехмерной структуре или типа аминокислотного остатка. Количество изменений может быть от 1 до 30, предпочтительно от 1 до 15 и наиболее предпочтительно от 1 до 5 изменений для белка (А). Данные изменения в вариантах белка являются консервативными мутациями, при которых сохраняется функция белка. Другими словами, данные изменения могут иметь место в областях белка, некритичных для его функции. Это становится возможным благодаря тому, что некоторые аминокислоты обладают высокой гомологией друг к другу, и поэтому третичная структура или активность белка не нарушаются при таких заменах. Поэтому вариантом белка (Б) может быть белок, имеющий гомологию не менее 70%, предпочтительно не менее 80%, более предпочтительно не менее 90% и наиболее предпочтительно не менее 95% по отношению к полной аминокислотной последовательности алкогольдегидрогеназы, представленной в SEQ ID NO. 2, при условии сохранения активности алкогольдегидрогеназы.
Гомология между двумя аминокислотными последовательностями может быть установлена с использованием хорошо известных методов, например компьютерной программы BLAST 2.0, которая определяет три параметра: число аминокислот, идентичность и сходство.
Замена, делеция, вставка или добавление одного или нескольких аминокислотных остатков должны представлять собой консервативную(-ые) мутацию(-и), позволяющую(-ие) сохранить активность фермента. Характерными консервативными мутациями являются консервативные замены. Примеры консервативных замен включают замену Аlа на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или А1а, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Туг, замену Не на Leu, Met, Val или Phe, замену Leu на Не, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Аlа, замену Thr на Ser или Аlа, замену Тrp на Phe или Туr, замену Туr на His, Phe или Trp и замену Val на Met, Ile или Leu.
Результаты сравнения первичных последовательностей алкогольдегидрогеназ из Escherichia соli, Shigella flexneri, Pantoea ananatis, Yersinia pestis, Erwinia carotovora, Salmonella typhimurium (грамотрицательные бактерии) и Lactobacillus plantarum, Lactococcus lactis (грамположительные бактерии) свидетельствуют о высоком уровне гомологии этих белков (см. Фиг.2). С этой точки зрения замены или делеции аминокислотных остатков, которые являются идентичными (отмеченные звездочками), во всех вышеупомянутых белках могут быть значимыми для их функции. Допустимой является замена сходных (отмеченных двоеточием) аминокислотных остатков на сходные аминокислотные остатки без снижения активности белка. Но модификации других неконсервативных аминокислотных остатков могут не приводить к изменению активности алкогольдегидрогеназы.
ДНК, кодирующая по существу тот же белок, что и алкогольдегидрогеназа, описанная выше, может быть получена, например, путем модификации нуклеотидной последовательности ДНК, кодирующей алкогольдегидрогеназу (SEQ ID NO: 1), например, посредством сайт-направленного мутагенеза таким образом, что удаляют, заменяют, вставляют или добавляют нуклеотидные последовательности, кодирующие один или несколько аминокислотных остатков в определенном сайте. Фрагмент ДНК, модифицированной, как описано выше, может быть получен с помощью общеизвестных мутагенных воздействий. Такие воздействия включают обработку препарата ДНК, кодирующей белки настоящего изобретения, гидроксиламином или обработку бактерии, содержащей указанную ДНК, УФ-облучением или реагентами, такими как N-метил-N'-нитро-N-нитрозогуанидин или азотистая кислота.
Белок, по существу такой же, что и алкогольдегидрогеназа, может быть получен путем экспрессии ДНК с описанной выше мутацией в соответствующей клетке и исследования активности продукта экспрессии. ДНК, кодирующая по существу тот же белок, что и алкогольдегидрогеназа, может также быть получена путем выделения ДНК, гибридизующейся в жестких условиях с зондом, имеющим нуклеотидную последовательность, которая содержит, например, нуклеотидную последовательность, показанную в SEQ ID NO: 1, и кодирует белок, обладающий активностью алкогольдегидрогеназы. Упомянутые «жесткие условия» включают такие условия, при которых так называемые специфические гибриды образуются, а неспецифические гибриды не образуются. Например, жесткие условия могут включать условия, при которых ДНК с высокой степенью гомологии, например ДНК с идентичностью не менее 50%, предпочтительно не менее 60%, более предпочтительно не менее 70%, еще более предпочтительно не менее 80%, более предпочтительно не менее 90%, наиболее предпочтительно не менее 95%, могут гибридизоваться друг с другом, а ДНК с идентичностью ниже вышеуказанной не могут гибридизоваться друг с другом. С другой стороны, примером жестких условий могут быть условия, при которых ДНК может гибридизоваться при концентрации соли, соответствующей стандартным условиям отмывки при гибридизации по Саузерну, например 1XSSC, 0.1% SDS, предпочтительно 0.1XSSC, 0.1% SDS, при 60°С. Продолжительность отмывки зависит от типа используемой для блоттинга мембраны и, как правило, такова, как рекомендовано производителем. Например, рекомендуемая продолжительность отмывки для нейлоновой мембраны HybondTM N+ (Amersham) при жестких условиях - 15 минут. Предпочтительна двух-трехкратная отмывка.
Часть нуклеотидной последовательности SEQ ID NO: 1 также может быть использована в качестве зонда. Зонды могут быть приготовлены методом ПЦР с использованием праймеров, основанных на нуклеотидной последовательности SEQ ID NO: 1, и фрагмента ДНК, содержащего нуклеотидную последовательность SEQ ID NO: 1, в качестве матрицы. Когда в качестве зонда используется фрагмент ДНК, имеющий длину около 300 п.о., условия отмывки для гибридизации включают, например, 50°С, 2XSSC и 0.1% SDS.
Описанные выше замена, делеция, вставка или добавление нуклеотидов также включают мутации, имеющие место в природе (мутант или вариант), например мутации, которые обусловлены изменчивостью вида или рода бактерии и которые затрагивают алкогольдегидрогеназу.
Алкогольдегидрогеназа дикого типа может подвергаться катализируемому металлами окислению. Хотя такая алкогольдегидрогеназа дикого типа также может использоваться, в настоящем изобретении предпочтительно использование мутантной алкогольдегидрогеназы, устойчивой к аэробной инактивации. Фраза «мутантная алкогольдегидрогеназа, устойчивая к аэробной инактивации» означает, что мутантная алкогольдегидрогеназа сохраняет свою активность в аэробных условиях или ее активность снижается в меньшей степени по сравнению с алкогольдегидрогеназой дикого типа.
В случае экспрессии гена adhE из Е. coli алкогольдегидрогеназа дикого типа включает аминокислотную последовательность, представленную в SEQ ID NO: 2. Пример мутации алкогольдегидрогеназы SEQ ID NO: 2, которая ведет к устойчивости к аэробной инактивации, является замена остатка глутаминовой кислоты в положении 568 остатком лизина. Однако введение мутации в ген adhE, например, в положении 568 в последовательности SEQ ID NO: 2 может приводить к замедлению роста в жидкой среде, содержащей этанол в качестве источника углерода, и в этом случае предпочтительно, чтобы мутантная алкогольдегидрогеназа содержала по крайней мере одну дополнительную мутацию, способную улучшить рост бактерии в жидкой среде, содержащей этанол в качестве единственного источника углерода. Например, рост E.coli улучшается, когда остаток глутаминовой кислоты в положении 568 в алкогольдегидрогеназе с последовательностью SEQ ID NO: 2 замещается другой аминокислотой с введением дополнительной мутации, выбранной из группы, состоящей из:
A) замены остатка глутаминовой кислоты в положении 560 в последовательности SEQ ID NO: 2 остатком другой аминокислоты, например остатком лизина;
Б) замены остатка фенилаланина в положении 566 в последовательности SEQ ID NO: 2 остатком другой аминокислоты, например остатком валина;
B) замены остатка глутаминовой кислоты, остатка метионина, остатка тирозина, остатка изолейцина и остатка аланина в положениях 22, 236, 461, 554 и 786 соответственно в последовательности SEQ ID NO: 2 остатками других аминокислот, например остатком глицина, остатком валина, остатком цистеина, остатком серина и остатком валина соответственно; и
Г) комбинации этих замен.
Ссылка на номера положения в последовательности, например фраза «аминокислотные остатки в положениях 22, 236, 554, 560, 566, 568 и 786» относится к положениям этих остатков в аминокислотной последовательности AdhE дикого типа E.coli. Однако положение аминокислотного остатка может изменяться. Например, если в N-концевой области встраивается аминокислотный остаток, то аминокислотный остаток, изначально находящийся в положении 22, приобретает положение 23. В таком случае аминокислотным остатком в положении 22 согласно настоящему изобретению является аминокислотый остаток, изначально находящийся в положении 22.
Мутантный белок AdhE может включать делецию, замену, вставку или добавление одной или нескольких аминокислот в одном или во множестве положений, отличных от определенных в А) - В), при условии, что активность алкогольдегидрогеназы при этом не теряется или не снижается.
Мутантный белок AdhE и мутантный ген adhE согласно настоящему изобретению могут быть получены из гена adhE природного типа, например, путем сайт-специфического мутагенеза с использованием общеизвестных методов, таких как ПЦР (полимеразная цепная реакция; ссылка на White, T.J. et al.. Trends Genet., 5, 185 (1989)) с использованием праймеров, сконструированных на основе нуклеотидной последовательности гена.
Транскрипция гена adhE в клетках Е. coli дикого типа индуцируется только в анаэробных условиях, главным образом в ответ на повышение уровня восстановленного NADH (Leonardo, M. R., Cunningham, P. R. & Clark, D. P, J. Bacteriol. 175 870-878 (1993)).
В настоящем изобретении бактериальный штамм, используемый для продукции L-аминокислоты, модифицирован таким образом, что экспрессия гена adhE контролируется ненативным промотором, т.е. промотором, который не контролирует экспрессию гена adhE в штамме дикого типа. Такая модификация может быть достигнута заменой нативного промотора гена adhE на хромосоме ненативным промотором, который функционирует в аэробных условиях культивирования таким образом, что ген adhE оказывается функционально связанным с ненативным промотором. В качестве ненативного промотора, функционирующего в аэробных условиях культивирования, может использоваться любой промотор, обеспечивающий экспрессию на определенном уровне вышеописанного гена adhE в аэробных условиях. Что касается уровня белка AdhE в настоящем изобретении, активность алкогольдегидрогеназы в экстракте клеток, измеренная методом, описанным Clark and Cronan (J. Bacteriol. 141 177-183 (1980)), должна быть 1.5 ед или более, предпочтительно 5 ед или более и более предпочтительно 10 ед или более на мг белка. Аэробные условия культивирования могут быть такими, какие обычно используются для культивирования бактерий, в которых кислород поставляется такими методами, как встряхивание, аэрация и перемешивание. Конкретно, может использоваться любой промотор, о котором известно, что он способствует экспрессии гена в аэробных увловиях культивирования. Например, могут использоваться промоторы генов, вовлеченных в гликолиз, пентозофосфатный путь, цикл трикарбоновых кислот, пути биосинтеза аминокислот и т.д. Кроме того, предпочтительно использовать промоторы, о которых известно, что они являются сильными промоторами, функционирующими в аэробных условиях: Рtac промотор, Рlac промотор, Ptrp промотор, Рtrc промотор, PR или PL промоторы фага λ.
Использование ненативного промотора можно сочетать с увеличением числа копий гена. Например, встраивание гена adhE, функционально связанного с ненативным промотором, в вектор, способный функционировать в бактерии семейства Enterobacteriaceae, и введение вектора в бактерию увеличивает число копий гена в клетке. Предпочтительно использование низкокопийных векторов. Примеры низкокопийных векторов включают, но не ограничиваются ими, pSClOl, pMW118, pMW119 и т.п. Термин «низкокопийный вектор» используется для векторов, число копий которого - до 5 на клетку. Увеличение числа копий гена adhE также может быть достигнуто введением множества копий гена в хромосомную ДНК бактерии путем, например, гомологичной рекомбинации, µ-интеграции и т.п. Гомологичная рекомбинация осуществляется с использованием последовательности, которая присутствует на хромосомной ДНК в качестве целевой во множестве копий. Последовательности, имеющие множество копий на хромосомной ДНК, включают, но не ограничиваются ими, повторяющиеся ДНК или инвертированные повторы, присутствующие на концах транспозона. Также, как раскрыто в U.S. Patent No. 5,595,889, можно включить ген adhE в транспозон для переноса и включения множества копий гена в хромосомную ДНК. В этих примерах ген adhE может быть помещен под контроль промотора, функционирующего в аэробных условиях культивирования. Кроме того, действие промотора может быть усилено, например, введением в промотор мутации для увеличения уровня транскрипции гена, локализованного за промотором. Кроме того, известно, что замена нескольких нуклеотидов в области между сайтом связывания рибосомы (ribosome binding site (RBS)) и инициирующим кодоном, особенно в последовательности, локализованной непосредственно перед инициирующим кодоном, значительно влияет на уровень трансляции РНК. Например, было обнаружено 20-кратное увеличение уровня экспрессии в зависимости от природы трех нуклеотидов, предшествующих инициирующему кодону (Gold et al., Annu. Rev. Microbiol., 35, 365-403, 1981; Hui et al., EMBO J., 3, 623-629, 1984). Ранее было показано, что мутация rhtA23 - замена G на А в положении -1 относительно инициирующего кодона ATG (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California August 24-29, 1997, abstract No. 457). Следовательно, можно предположить, что мутация rhtA23 увеличивает экспрессию гена rhtA и, как следствие, увеличивает устойчивость к треонину, гомосерину и некоторым другим веществам, транспортируемым из клетки.
Более того, также можно ввести нуклеотидную замену в область промотора гена adhE на бактериальной хромосоме, которая ведет к усилению функции промотора. Изменение контролирующей экспрессию последовательности может быть осуществлено, например, таким же способом, как замена гена, с использованием чувствительной к температуре плазмиды, как раскрыто в заявке РСТ WO 00/18935 и в выложенной японской патентной заявке No. 1-215280.
Согласно настоящему изобретению «бактерия, обладающая способностью к продукции L-аминокислоты» означает бактерию, способную при культивировании продуцировать и накапливать L-аминокислоту в питательной среде. Способность к продукции аминокислоты может быть придана или усилена искусственно. Используемый здесь термин «бактерия, обладающая способностью к продукции L-аминокислоты» также означает бактерию, способную при культивировании продуцировать и накапливать L-аминокислоту в питательной среде в больших количествах по сравнению с диким типом или родительским штаммом Е. coli, таким как штамм Е. coli К-12, и предпочтительно означает, что бактерия способна накапливать в среде целевую L-аминокислоту в количестве не менее 0.5 г/л, более предпочтительно не менее 1.0 г/л.
Термин «L-аминокислота» включает в себя L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин, L-глутаминовую кислоту, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин. Предпочтительным, в частности, являются L-треонин, L-лизин, L-гистидин, L-фенилаланин, L-аргинин, L-триптофан, L-глутаминовая кислота и L-лейцин.
Семейство Enterobacteriaceae включает бактерии, принадлежащие к роду Escherichia, Enterobacter, Erwinia, Klebsiella, Pantoea, Photorhabdus, Providencia, Salmonella, Serratia, Shigella, Morganella, Yersinia и так далее. Более конкретно, могут быть использованы бактерии, классифицируемые как принадлежащие к семейству Enterobacteriaceae в соответствии с таксономией, используемой в базе данных NCBI (National Center for Biotechnologylnformation) (http://www.ncbi.nlm.nih.gov/htbinpost/Taxonomy/wgetorg?mode=Tree&id=1236&lvl=3&keep=1&srchmode=1&unlock). Бактерия, принадлежащая к роду Escherichia или Pantoea, предпочтительна.
Фраза «бактерия, принадлежащая к роду Escherichia» означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, но не ограничивается только ею, может быть упомянута бактерия Escherichia coli (Е. coli).
Круг бактерий, принадлежащих к роду Escherichia, которые могут быть использованы в настоящем изобретении, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть включены в число бактерий согласно настоящему изобретению.
Фраза «бактерия, принадлежащая к роду Pantoea» означает, что бактерия относится к роду Pantoea в соответствии с классификацией, известной специалисту в области микробиологии. Недавно несколько видов Enterobacter agglomerans были заново классифицированы как Pantoea agglomerans, Pantoea ananatis, Pantoea stewartii или подобные им на основании анализа нуклеотидной последовательности 16S рРНК и т.д. (Int. J. Syst. Bacteriol., 43, 162-173 (1993)).
Бактерия согласно настоящему изобретению включает штамм бактерии семейства Enterobacteriaceae, который обладает способностью продуцировать L-аминокислоту и модифицирован таким образом, что ген, кодирующий алкогольдегидрогеназу, экспрессируется под контролем промотора, функционирующего в аэробных условиях. Кроме того, бактерия согласно настоящему изобретению включает штамм бактерии семейства Enterobacteriaceae, который способен продуцировать L-аминокислоту и не обладает при этом активностью нативной алкогольдегидрогеназы, но трансформирован фрагментом ДНК, кодирующим алкогольдегидрогеназу.
Согласно настоящему изобретению количество накапливаемой L-аминокислоты, например L-треонина, L-лизина, L-гистидина, L-фенилаланина, L-аргинина, L-триптофана, L-глутаминовой кислоты или L-лейцина, может быть значительно увеличено в культуральной среде, содержащей этанол в качестве единственного источника углерода, в результате экспрессии гена, кодирующего алкогольдегидрогеназу, под контролем промотора, функционирующего в аэробных условиях культивирования.
Методы получения плазмидной ДНК включают, но не ограничиваются ими, разрезание и лигирование ДНК, трансформацию, выбор олигонуклеотида в качестве праймера и т.п.или другие методы, хорошо известные специалисту в данной области. Эти методы описаны, например, в книге Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Бактерия согласно настоящему изобретению может быть получена путем введения вышеуказанных фрагментов ДНК в бактерию, уже обладающую способностью к продукции L-аминокислоты. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, уже содержащей указанные фрагменты ДНК, способности к продукции L-аминокислоты.
Бактерии-продуценты L-аминокислот
Согласно настоящему изобретению в качестве бактерии, которая модифицируется таким образом, чтобы содержать мутантную алкогольдегидрогеназу, могут быть использованы бактерии, которые способны продуцировать ароматические или неароматические L-аминокислоты.
Бактерия согласно настоящему изобретению может быть получена путем введения гена, кодирующего мутантную алкогольдегидрогеназу настоящего изобретения, в бактерию, уже обладающую способностью к продукции L-аминокислот. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, которая уже обладает мутантной алкогольдегидрогеназой, способности к продукции L-аминокислот.
Бактерии-продуценты L-треонина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-треонина, согласно настоящему изобретению включают, но не ограничиваются только штаммами бактерий-продуцентов L-треонина, принадлежащих к роду Escherichia, таких как штамм Е. coli TDH-6/pVIC40 (VKPM В-3996) (патенты США 5, 175, 107 и 5,705,371), штамм Е. coli 472T23/pYN7 (ATCC 98081) (патент США 5,631,157), штамм Е. coli NRRL-21593 (патент США 5,939,307), штамм Е. coli FERM ВР-3756 (патент США 5,474,918), штамм Е. coli FERM BP-3519 and FERM BP-3520 (патент США 5,376,538), штамм Е. coli MG442 (Гусятинер М.М. и др., Генетика, 1978, 14: 947-956), штаммы Е. coli VL643 и VL2055 (Европейская патентная заявка ЕР 1149911 А) и подобные им штаммы.
Штамм TDH-6 является дефектным по гену thrC, способен ассимилировать сахарозу и содержит ген ilvA с мутацией типа "leaky". Указанный штамм содержит мутацию в гене rhtA, которая обусловливает устойчивость к высоким концентрациям треонина и гомосерина. Штамм В-3996 содержит плазмиду pVIC40, которая была получена путем введения в вектор, производный от вектора RSF1010, оперона thrA*BC, включающего мутантный ген thrA, кодирующий аспартокиназу-гомосериндегидрогеназу I, у которой существенно снижена чувствительность к ингибированию треонином по типу обратной связи. Штамм В-3996 был депонирован 19 ноября 1987 года во Всесоюзном научном центре антибиотиков (РФ, 117105 Москва, Нагатинская ул., 3-А) с инвентарным номером РИА 1867. Указанный штамм также был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-й Дорожный проезд, 1) с инвентарным номером В-3996.
В качестве родительского штамма для получения бактерии-продуцента L-треонина согласно настоящему изобретению может быть также использован штамм Е. coli VKPM В-5318 (Европейский патент заявка ЕР 0593792 В). Штамм В-5318 является прототрофным в отношении изолейцина, а репрессор температурно-чувствительного фага лямбда С1 и промотор PR замещает регуляторную область треоинового оперона в плазмиде pVIC40. Штамм VKPM В-5318 был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-й Дорожный проезд, 1) 3 мая 1990 года с инвентарным номером В-5318.
Предпочтительно, чтобы бактерия согласно настоящему изобретению была далее модифицирована таким образом, чтобы иметь повышенную экспрессию одного или нескольких следующих генов:
- мутантного гена thrA, кодирующего аспартокиназу-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи;
- гена thrB, кодирующего гомосеринкиназу;
- гена thrC, кодирующего треонинсинтазу;
- гена rhtA, предположительно кодирующего трансмембранный белок;
- гена asd, кодирующего аспартат-β-семиальдегиддегидрогеназу, и
- гена aspC, кодирующего аспартатаминотрансферазу (аспартаттрансаминазу).
Нуклеотидная последовательность гена thrA, кодирующего аспартокиназа-гомосериндегидрогеназу I из Escherichia coli, известна (номера нуклеотидов с 337 по 2799 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrA расположен на хромосоме штамма Е. coli К-12 между генами thrL и thrB. Нуклеотидная последовательность гена thrB, кодирующего гомосеринкиназу из Escherichia coli, известна (номера нуклеотидов с 2801 по 3733 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrB расположен на хромосоме штамма Е. coli К-12 между генами thrA и thrC. Нуклеотидная последовательность гена thrC, кодирующего треонинсинтазу из Escherichia coli, известна (номера нуклеотидов с 3734 по 5020 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrC расположен на хромосоме штамма Е. coli К-12 между геном thrB и открытой рамкой считывания уааХ. Все три указанных гена функционируют как один треониновый оперон. Для того чтобы усилить экспрессию треоининового оперона, область аттенюатора, который регулирует транскрипцию, удалена из оперона (заявки по РСТ WO 2005/049808, WO 2003/097839).
Мутантный ген thrA, кодирующий аспартокиназу-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи, также, как и гены thrB и thrC, может быть получен в виде единого оперона из хорошо известной плазмиды pVIC40, которая представлена в штамме-продуценте Е. coli VKPM В-3996. Плазмида pVIC40 подробно описана в патенте США 5,705,371.
Ген rhtA расположен на 18 минуте хромосомы Е. coli около оперона glnHPQ, который кодирует компоненты транспортной системы глутамина, ген rhtA идентичен ORF1 (rwybiF, номера нуклеотидов с 764 по 1651 в последовательности с инвентарным номером ААА218541 в базе данных GenBank, gi:440181), расположен между генами рехВ и отрХ. Участок ДНК, экспрессирующийся с образованием белка, кодируемого рамкой считывания ORF1, был назван геном rhtA (rht: resistance to homoserine and threonine). Также было показано, что мутация rhtA23 представляет собой замену А на G в положении -1 по отношению к старт-кодону ATG (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California August 24-29, 1997, abstract No. 457; Европейская патентная заявка ЕР 1013765 А). В дальнейшем мутация rhtA23 обозначается как rhtA*.
Нуклеотидная последовательность гена asd из E.coli известна (номера нуклеотидов с 3572511 по 3571408 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi: 16131307) и может быть получена с помощью ПЦР (полимеразная цепная реакция; ссылка на White, T.J. et al.. Trends Genet., 5, 185 (1989)) с использованием праймеров, синтезированных на основе нуклеотидной последовательности указанного гена. Гены asd из других микроорганизмов могут быть получены сходным образом.
Также нуклеотидная последовательность гена aspC из E.coli известна (номера нуклеотидов с 983742 по 984932 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi: 16128895) и может быть получена с помощью ПЦР. Гены aspC из других микроорганизмов могут быть получены сходным образом.
Бактерии-продуценты L-лизина
Примеры бактерий-продуцентов L-лизина, принадлежащих к роду Escherichia, включают мутанты, обладающие устойчивостью к аналогу L-лизина. Аналог L-лизина ингибирует рост бактерий, принадлежащих к роду Escherichia, но это ингибирование полностью или частично снимается, когда в среде также присутствует L-лизин. Примеры аналога L-лизина включают, но не ограничиваются оксализином, лизингидроксаматом, S-(2-аминоэтил)-L-цистеином (АЕС), γ-метиллизином, α-хлорокапролактамом и так далее. Мутанты, обладающие устойчивостью к указанным аналогам лизина, могут быть получены путем обработки бактерий, принадлежащих к роду Escherichia, традиционными мутагенами. Конкретные примеры бактериальных штаммов, используемых для получения L-лизина, включают штамм Escherichia coli AJ11442 (PERM BP-1543, NRRL В-12185; патент США 4,346,170) и штамм Escherichia coli VL611. В этих микроорганизмах аспартокиназа устойчива к ингибированию L-лизином по типу обратной связи.
Штамм WC196 может быть использован в качестве бактерии-продуцента L-лизина Escherichia coli. Данный бактериальный штамм был получен путем селекции фенотипа устойчивости к АЕС у штамма W3110, производного от штамма Escherichia coli K-12. Полученный штамм был назван Escherichia coli AJ13069 и был депонирован в Национальном Институте Биологических Наук и Человеческих Технологий, Агенство Промышленной Науки и Технологии (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology), в настоящее время называющийся Национальный Институт Прогрессивной Промышленной Науки и Технологии, Международный Депозитарий Организмов для Целей Патентования, Централ 6, 1-1, Хигаши 1-Чоме, Тсукуба-ши, Ибараки-кен, 305-8566, Япония (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan), 6 декабря 1994 года и получил инвентарный номер PERM P-14690. Затем было произведено международное депонирование этого штамма согласно условиям Будапештского Договора 29 сентября 1995 года, и штамм получил инвентарный номер PERM BP-5252 (патент США 5,827,698).
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-лизина согласно настоящему изобретению, включают также штаммы, в которых экспрессия одного или более генов, кодирующих ферменты биосинтеза L-лизина, усилена. Примеры таких генов включают, но не ограничиваются только ими, гены, кодирующие дигидродипиколинатсинтазу (ген dapA), аспартокиназу (ген lysC), дигидродипиколинатредуктазу (ген dapB), диаминопимелатдекарбоксилазу (ген tysA), диаминопимелатдегидрогеназу (ген ddh) (патент США 6,040,160), фосфоенолпируваткарбоксилазу (ген ррс), аспартатполуальдегиддегидрогеназу (ген asd) и аспартазу (ген aspA) (Европейская патентная заявка ЕР 1253195 А). Кроме того, родительские штаммы могут иметь повышенный уровень экспрессии гена, включенного в энергетический поток (ген суо) (Европейская патентная заявка ЕР 1170376 А), гена, кодирующего никотинамиднуклеотидтрансгидрогеназу (ген pntAB) (патент США 5,830,716), гена ybjE (заявка РСТ WO 2005/073390), или комбинации этих генов.
Примеры родительских штаммов для получения бактерий-продуцентов L-лизина согласно настоящему изобретению включают штаммы, имеющие сниженную или элиминированную активность фермента, который катализирует реакцию образования компонента, отличного от L-лизина, путем ответвления от пути биосинтеза L-лизина. Примеры ферментов, которые катализирует реакцию образования компонента, отличного от L-лизина, путем ответвления от пути биосинтеза L-лизина, включают гомосериндегидрогеназу, лизиндекарбоксилазу (патент США 5,827,698) и пируват-малаткарбоксилазу/малатдегидрогеназу (заявка РСТ W02005/010175).
Примеры штаммов - продуцентов L-лизина включают Е. coli WC196AcadAAldc/pCABD2 (W02006/078039). Этот штамм был получен введением плазмиды pCABD2, которая описана в U.S. Patent No. 6,040,160, в штамм WC196 с разрушенными генами cadA и IdcC, кодирующими лизиндекарбоксилазу. Плазмида pCABD2 содержит ген dapA E.coli, кодирующий дигидродипиколинатсинтазу с мутацией, обеспечивающей устойчивость к ингибированию L-лизином по типу обратной связи, ген lysC E.coli, кодирующий аспартокиназу III с мутацией, обеспечивающей устойчивость к ингибированию L-лизином по типу обратной связи, ген dapB E.coli, кодирующий дигидродипиколинатредуктазу, и ген ddh Corynebacterium glutamicum, кодирующий диаминопимелатдегидрогеназу.
Бактерии-продуценты L-цистеина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-цистеина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм E.coli JM15, трансформированный различными аллелями гена cysE, кодирующими устойчивые к ингибированию по типу обратной связи серинацетилтрансферазы (патент США 6218168, патентная заявка РФ 2003121601); штамм E.coli W3110, содержащий гены с повышенной экспрессией, кодирующие белок, способный к секреции соединений, токсичных для клетки (патент США 5972663); штаммы E.coli, содержащие цистеиндесульфогидразу со сниженной активностью (патент Японии JP 11155571 А2); штамм E.coli W3110 с повышенной активностью позитивного транскрипционного регулятора цистеинового регулона, кодируемого геном cysB (международная заявка РСТ WO0127307 A1) и подобные им.
Бактерии-продуценты L-лейцина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-лейцина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штаммы E.coli, устойчивые к аналогам лейцина, включающие, например, 3-2-тиенилаланин, 3-гидроксилейцин, 4-азалейцин и 5,5,5-трифлуоролейцин (выложенные патентные заявки Японии 62-34397 и 8-70879), штаммы E.coli, полученные с помощью генно-инженерных методов, описанных в заявке РСТ 96/06926; E.coli штамм Н-9068 (JP 08-70879A) и подобные им.
Бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, вовлеченных в биосинтез L-лейцина. Примеры таких генов включают в себя гены оперона leuABCD и предпочтительно представлены мутантным геном leuA, кодирующим изопропилмалатсинтазу со снятым ингибированием L-лейцином по типу обратной связи (патент США 6403342). Кроме того, бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, кодирующих белки, которые экспортируют L-аминокислоту из бактериальной клетки. Примеры таких генов включают в себя гены b2682 и b2683 (гены ygaZH) (европейская патентная заявка ЕР 1239041 А2).
Бактерии-продуценты L-гистидина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-гистидина, согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штаммы Е. coli 24 (VKPM В-5945, патент РФ 2003677); Е. coli 80 (VKPM В-7270, патент РФ 2119536); Е. coli NRRL В-12116 - В12121 (патент США 4,388,405); Е. coli H-9342 (FERM ВР-6675) и Е. coli Н-9343 (FERM ВР-6676) (патент США 6,344,347); Е. coli H-9341 (FERM BP-6674) (Европейский патент ЕР1085087); Е. coli AI80/pFM201 (патент США 6,258,554) и им подобные.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-гистидина, согласно настоящему изобретению также включают штаммы, в которых экспрессия одного или более генов, кодирующих ферменты биосинтеза L-гистидина, усилена. Примеры таких генов включают гены, кодирующие ферменты биосинтеза L-гистидина, которые включают АТФ-фосфорибозилтрансферазу (ген hisG), фосфорибозил-АМФ-циклогидролазу (ген hisi), фосфорибозил-АТФ-пирофосфогидролазу (ген hisIE), фосфорибозилформимино-5-аминоимидазолкарбоксамидриботидизомеразу (ген hisA), амидотрансферазу (ген hisff), гистидинолфосфатаминотрансферазу (ген hisC), гистидинолфосфатазу (ген hisB), гистидинолдегидрогеназу (ген hisD), и так далее.
Известно, что гены, кодирующие ферменты биосинтеза L-гистидина (hisG, hisBHAFI), угнетаются L-гистидином и поэтому способность продуцировать L-гистидин может быть также эффективно усилена путем введения мутации, придающей устойчивость к ингибированию по типу обратной связи, в фермент фосфорибозилтрансферазу (HisG) (патенты РФ 2003677 и 2119536).
Отдельные примеры штаммов, обладающих способностью продуцировать L-гистидин, включают штаммы Е. coli FERM-P 5038 и 5048, в которые был введен вектор, несущий ДНК, кодирующую ферменты биосинтеза L-гистидина (патентная заявка Японии JP 56-005099 А), штамм Е. coli 80, которому придана устойчивость к сульфагуанидину, DL-1,2,4-триазол-3-аланину и стрептомицину (ВКПМ-7270, патент РФ 2119536), и так далее.
Бактерии-продуценты L-глутаминовой кислоты.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-глутаминовой кислоты, согласно настоящему изобретению включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как Е. coli VL334thrC+(Европейский патент ЕР 1172433). Штамм Е. coli VL334 (ВКПМ В-1641) является ауксотрофным по L-изолейцину и L-треонину и имеет мутации в генах thrC и UvA (патент США 4,278,765). Аллель гена thrC природного штамма переносили методом основной трансдукции с использованием бактериофага Р1, растущего на клетках штамма природного типа Е. coli К 12 (ВКПМ В-7). В результате был получен штамм, ауксотрофный по L-изолейцину VL334thrC+ (ВКПМ В-8961). Этот штамм способен продуцировать L-глутаминовую кислоту.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-глутаминовой кислоты, согласно настоящему изобретению включают в себя, но не ограничиваются штаммами, в которых экспрессия одного или более генов, кодирующих ферменты биосинтеза L-глутаминовой кислоты, усилена. Примеры таких генов включают гены, кодирующие глутаматдегидрогеназу (ген gdh), глутаминсинтетазу (ген glnA), глутаматсинтетазу (gItAB), изоцитратдегидрогеназу (ген icdA), аконитатгидратазу (гены acnА, acnВ), цитратсинтазу (ген gltA), фосфоенолпируваткарбоксилазу (ген ррс), пируватдегидрогеназу (гены aceEF, IpdA), пируваткиназу (гены pykA, pykF), фосфоенолпируватсинтазу (ген ppsA), енолазу (ген епо), фосфоглицеромутазу (гены pgmA, pgmf), фосфоглицераткиназу (ген pgk), глицеральдегид-3-фосфатдегидрогеназу (ген gapA), триозофосфатизомеразу (ген tpiA), фруктозобисфосфатальдолазу (ген fbp), фосфофруктокиназу (гены pfkA, pfkB) и глюкозофосфатизомеразу (renpgi) и т.д.
Примеры штаммов, модифицированных так, что экспрессия гена цитратсинтетазы, гена фосфоенолпируваткарбоксилазы и/или гена глутаматдегидрогеназы, усилена/ы, включают штаммы, описанные в Европейских патентных заявках ЕР 1078989 А, ЕР 955368 А и ЕР 952221 А.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-глутаминовой кислоты, согласно настоящему изобретению также включают в себя штаммы со сниженной или элиминированной активностью фермента, который катализирует синтез компонента, другого чем L-глутаминовая кислота, но ответвляющийся от пути биосинтеза L-глутаминовой кислоты. Примеры таких генов включают гены, кодирующие изоцитратлиазу (ген асеА), α-кетоглутаратдегидрогеназу (ген sucA), фосфотрансацетилазу (ген pta), ацетаткиназу (ген ack), ацетогидроксикислую синтазу (ген ilvG), ацетолактатсинтазу (ген ilvI), форматацетилтрансферазу (ген pfl), лактатдегидрогеназу (ген Idh) и глутаматдекарбоксилазу (ген gadAB). Бактерии, принадлежащие к роду Escherichia, дефицитные по активности фермента α-кетоглутаратдегидрогеназы или имеющие сниженную активность фермента α-кетоглутаратдегидрогеназы, и способы их получения описаны в патентах США 5,378,616 и 5,573,945. Конкретно, они включают нижеследующие штаммы:
E. coli W3110sucA::KmR
Е. coli АJ 12624 (PERM BP-3853)
Е. coli АJ 12628 (PERM BP-3854)
Е. coli АJ 12949 (PERM BP-4881)
Е. coli W3110sucA::K.mR является штаммом, полученным путем разрушения гена α-кетоглутаратдегидрогеназы (в дальнейшем именуемого как ген "sucA)"в штамме Е. coli W3110. Этот штамм полностью дефицитен по α-кетоглутаратдегидрогеназе.
Другие примеры бактерий-продуцентов L-глутаминовой кислоты включают штаммы, которые принадлежат к роду Escherichia и обладают устойчивостью к антиметаболитам аспарагиновой кислоты. Эти штаммы могут быть также лишены активности фермента α-кетоглутаратдегидрогеназы и включают, к примеру, штаммы Е., coli АJ 13199 (PERM ВР-5807) (патент США 5.908,768), FFRM Р-12379, который к тому же имеет низкую способность расщеплять L-глутаминовую кислоту (патент США 5,393,671); АJ 13138 (PERM BP-5565) (патент США 6,110,714) и им подобные.
Примеры бактерий-продуцентов L-глутаминовой кислоты включают мутантные штаммы, принадлежащие к роду Pantoea, которые лишены активности фермента α-кетоглутаратдегидрогеназы или имеют сниженную активность фермента α-кетоглутаратдегидрогеназы и могут быть получены, как описано выше. Такие штаммы включают штамм Pantoea ananatis AJ13356 (патент США 6,331,419). Штамм Pantoea ananatis АJ 13356 был депонирован в Национальном Институте Биологических Наук и Человеческих Технологий, Агенство Промышленной Науки и Технологии (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology), в настоящее время называющийся Национальный Институт Прогрессивной Промышленной Науки и Технологии, Международный Депозитарий Организмов для Целей Патентования, Централ 6, 1-1, Хигаши 1-Чоме, Тсукуба-ши, Ибараки-кен, 305-8566, Япония (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan), 19 февраля, 1998 года и получил инвентарный номер PERM P-16645. Затем было произведено международное депонирование этого штамма согласно условиям Будапештского Договора 11 января 1999 года, и штамм получил инвентарный номер FERM ВР-6615. Штамм Pantoea ananatis АJ 13356 лишен активности фермента α-кетоглутаратдегидрогеназы в результате разрушения субъединицы αKGDH-E1 гена sucA. При получении вышеописанный штамм был идентифицирован как штамм Enterobacter agglomerans и депонирован как штамм Enterobacter agglomerans AJ13356. Однако в последнее время на основе анализа нуклеотидной последовательности 16S pRNA он был переклассифицирован как принадлежащий к виду Pantoea ananatis. Хотя штамм AJ13356 был депонирован в вышеупомянутый депозитарий как штамм Enterobacter agglomerans, в целях данной спецификации они описываются как штаммы Pantoea ananatis.
Бактерии-продуценты L-фенилаланина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-фенилаланина, согласно настоящему изобретению включают в себя, но не ограничиваются только ими, штаммы, принадлежащие к роду Escherichia, такие как штаммы Е. coli АJ 12739 (tyrA::Tn10, tyrR) (ВКПМ В-8197); Е. coli HW1089 (АТСС 55371) несущий ген pheA34 (патент США 5,354,672); Е. coli MWEC101-b (KR8903681); Е. coli NRRL B-12141, NRRL B-12145, NRRL В-12146 и NRRL В-12147 (патент США 4,407,952). Также в качестве родительских штаммов могут быть использованы штаммы Е. coli K-12 [W3110 (tyrA)/pPHAB (FERM BP-3566), Е. coli K-12 [W3110 (tyrA)/pPHAD] (FERM BP-12659), Е. coli K-12 [W3110 (tyrA)/pPHATerm] (FERM BP-12662) и Е. coli K-12 [W3110 (tyrA)/pBR-aroG4, pACMAB] названный AJ12604 (PERM BP-3579) (Европейский патент ЕР 488424 В1). Кроме того, могут использоваться бактерии-продуценты L-фенилаланина, принадлежащие к роду Escherichia с усиленной активностью белка, кодируемого геном yedA или геном yddG (патентные заявки США 2003/0148473 А1 и 2003/0157667 А1).
Бактерии-продуценты L-триптофана
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-триптофана, согласно настоящему изобретению включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штаммы Е. coli JP4735/pMU3028 (DSM10122) и JP6015/pMU91 (DSM10123), лишенными фермента триптофанил-тРНК-синтетаза, кодируемого мутантным геном trpS (патент США 5,756,345); штамм Е. coli SV164 (pGH5) с аллелем serA, кодирующим фосфоглицератдегидрогеназу со снятым ингибированием серином по типу обратной связи и аллелем trpE, кодирующим антранилатсинтазу со снятым ингибированием триптофаном по принципу обратной связи (патент США 6,180,373); штаммы Е. coli AGX17 (pGX44) (NRRL В-12263) и AGX6(pGX50)aroP (NRRL В-12264), лишенные фермента триптофаназы (патент США 4,371,614); штамм Е. coli AGX 17/pGX50,pACKG4-pps, в котором усилена способность продуцировать фосфоенолпируват (заявка РСТ W09708333, патент США 6,319,696) и подобные им могут быть использованы. Бактерии-продуценты L-триптофана, принадлежащие к роду Escherichia с увеличенной активностью белка, кодируемого геном yedA или геном yddG, могут быть также использованы (патентные заявки США 2003/0148473 А1 и 2003/0157667 А1).
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-триптофана, согласно настоящему изобретению, также включают в себя штаммы, в которых увеличена активность одного или более ферментов, выбранных из группы, состоящей из антранилатсинтазы, фосфоглицератдегидрогеназы и триптофансинтазы. Оба фермента антранилатсинтаза и фосфоглицератдегидрогеназа подвержены ингибированию L-триптофаном и L-серином по типу обратной связи таким образом, что мутация десенсибилизации по типу обратной связи может быть введена в эти ферменты. Конкретные примеры штаммов, обладающих такой мутацией, включают штамм Е. coli SV164, несущий антранилатсинтазу со снятым ингибированием, и трансформированный штамм, полученный путем введения в штамм Е. coli SV164 плазмиды pGH5 (WO 94/08031), которая содержит мутантный ген sera, кодирующий фосфоглицератдегидрогеназу со снятым ингибированием по типу обратной связи.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-триптофана, согласно настоящему изобретению также включают в себя штаммы, в которые был введен триптофановый оперон, который содержит ген, кодирующий антранилатсинтазу со снятым ингибированием (патентные заявки Японии JP 57-71397 А, JP 62-244382 А, патент США 4,371,614). Сверх того, способность продуцировать L-триптофан может быть придана бактерии путем усиления экспрессии гена, который кодирует триптофансинтазу в составе триптофановых оперонов (trpBA). Белок триптофансинтаза состоит из α- и β-субъединиц, которые кодируются генами trpA и trpB соответственно. Кроме того, способность продуцировать L-триптофан может быть улучшена путем усиления экспрессии оперона изоцитратлиазмалатсинтазы (заявка РСТ WO2005/103275).
Бактерии-продуценты L-пролина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-пролина, согласно настоящему изобретению включают, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli 702ilvA (VKPM В-8012), который лишен гена ilvA и способен продуцировать L-пролин (Европейский патент ЕР 1172433). Бактерия согласно настоящему изобретению может быть усовершенствована путем усиления экспрессии одного или более генов, участвующих в биосинтезе L-пролина. Примеры таких генов для бактерий-продуцентов L-пролина, которые являются предпочтительными, включают ген proВ, кодирующий фермент глютаматкиназу, для которого регуляция L-пролином по типу обратной связи десенсибилизирована (патент Германии DE 3127361). Дополнительно, бактерия согласно настоящему изобретению может быть усовершенствована путем усиления экспрессии одного или более генов, кодирующих белки, экспортирующие L-аминокислоту из бактериальной клетки. Примером таких генов могут служить гены b2682 и b2683 (гены ygaZH) (Европейская патентная заявка ЕР 1239041 А2).
Примеры бактерий-продуцентов L-пролина, принадлежащих к роду Escherichia, включают следующие штаммы Е. coli: NRRL В-12403 и NRRL В-12404 (патент Великобритании GB 2075056), VKPM В-8012 (патентная заявка РФ 2000124295), плазмидные мутантные штаммы, описанные в патенте Германии DE 3127361, плазмидные мутантные штаммы, описанные Bloom F.R. и соавт. (The 15th Miami winter symposium, 1983, р.34) и им подобные.
Бактерии-продуценты L-аргинина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-аргинина, согласно настоящему изобретению включают, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli 237 (VKPM В-7925) (патентная заявка США 2002/058315 А1) и производные от него штаммы, несущие мутантную N-ацетилглутаматсинтазу (патентная заявка РФ 2001112869), штамм Е. coli 382 (VKPM B-7926) (Европейская патентная заявка ЕР1170358А1), штамм-продуцент аргинина, в который введен ген argA, кодирующий N-ацетилглутаматсинтетазу (Европейская патентная заявка ЕР1170361А1) и им подобные.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-аргинина, согласно настоящему изобретению включают также штаммы, в которых экспрессия одного или более генов, кодирующих ферменты биосинтеза L-аргинина, усилена. Примеры таких генов включают гены, кодирующие N-ацетилглютамилфосфатредуктазу (ген argC), орнитинацетилтрансферазу (ген argJ), N-ацетилглутаматкиназу (ген argB), ацетилорнитинтрансаминазу (ген argD), орнитинкарбамоилтрансферазу (ген argF), синтетазу аргининоянтарной кислоты (ген argG), лиазу аргининоянтарной кислоты (ген argH) и карбамоилфосфатсинтетазу (ген carАВ}.
Бактерии-продуценты L-валина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-валина, согласно настоящему изобретению включают, но не ограничиваются штаммами, которые модифицированы таким образом, что экспрессия оперона ilvGMEDA усилена (патент США 5,998,178). Желательно удалить область оперона ilvGMEDA, необходимую для аттенуации, для того, чтобы экспрессия оперона не снижалась вырабатываемым L-валином. Кроме того, ген ilvA в опероне желательно разрушить для того, чтобы активность треониндезаминазы снизилась.
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-валина, согласно настоящему изобретению включают также мутантные штаммы, имеющие мутацию фермента аминоацил-т-РНК-синтетазы (патент США 5,658,766). К примеру, можно использовать штамм Е. coli VL1970, несущий мутацию в гене ileS, который кодирует фермент изолейцин т-РНК синтетазу. Штамм Е. coli VL1970 депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 113545 Москва, 1-й Дорожный проезд, 1) 24 июня 1988 года с инвентарным номером VKPMB-4411.
Кроме того, мутантные штаммы, требующие для своего роста присутствия липоевой кислоты и/или отсутствия H+-АТФ-азы, могут быть также использованы в качестве родительских штаммов (заявка РСТ WO96/06926).
Бактерии-продуценты L-изолейцина
Примеры родительских штаммов, используемых для получения бактерий-продуцентов L-изолейцина, согласно настоящему изобретению включают, но не ограничиваются штаммами, имеющими мутации устойчивости к 6-диметиламинопурину (патентная заявка Японии JP 5-304969 А), имеющими мутации устойчивости к аналогам изолейцина, таким как тиоизолейцин и гидроксамат изолейцина, и мутантными штаммами, имеющими дополнительно устойчивость к DL-этионину и/или гидроксомату аргинина (патентная заявка Японии JP 5-130882 А). Дополнительно в качестве родительских штаммов могут быть использованы рекомбинантные штаммы, трансформированные генами, кодирующими белки, участвующие в биосинтезе L-изолейцина, такие как треониндезаминаза и ацетогидроксатсинтаза (патентная заявка Японии JP 2-458 А, патент Франции FR 0356739 и патент США 5,998,178).
Способ согласно настоящему изобретению
Способом согласно настоящему изобретению является способ получения L-аминокислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-аминокислоты в культуральной жидкости и выделения L-аминокислоты из культуральной жидкости. Кроме того, способ согласно настоящему изобретению включает способ получения L-треонина, L-лизина, L-гистидина, L-фенилаланина, L-аргинина, L-триптофана и L-глутаминовой кислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-треонина, L-лизина, L-гистидина, L-фенилаланина, L-аргинина, L-триптофана и L-глутаминовой кислоты в культуральной жидкости и выделения L-треонина, L-лизина, L-гистидина, L-фенилаланина, L-аргинина, L-триптофана и L-глутаминовой кислоты из культуральной жидкости.
Согласно настоящему изобретению выращивание, выделение и очистка L-аминокислоты из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием бактерии.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты и спирты, такие как этанол. Согласно настоящему изобретению в качестве единственного источника углерода может быть использован этанол или смесь этанола с карбогидратами, такими как глюкоза и сахароза.
В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и подобные им соединения. При необходимости в питательную среду могут быть добавлены дополнительные питательные вещества. Например, если для роста бактерии необходимо присутствие L-аминокислоты (ауксотрофность по L-аминокислоте), то в питательную среду может быть добавлено значительное количество L-аминокислоты.
Выращивание осуществляется предпочтительно в аэробных условиях, например при перемешивании культуральной жидкости на качалке, взбалтывании с аэрацией, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30°С до 38°С. рН среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды можно доводить аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и/или кристаллизации.
Краткое описание фигур
На Фиг.1 представлена структура участка, предшествующего гену adhE по направлению транскрипции, на хромосоме Е. coli и структура интегрированного фрагмента ДНК, содержащего последовательность гена cat и последовательность рекомбинантного промотора PL-tac.
На Фиг.2 представлен результат выравнивания первичных последовательностей алкогольдегидрогеназы из Escherichia coli (ADHE_ECOLI, SEQ ID NO: 2), Shigella flexneri (Q83RN2_SHIFL, SEQ ID NO: 53), Pantoea ananatis (ADHE PANAN, SEQ ID NO: 30), Yersinia pestis (Q66AM7_YERPS, SEQ ID NO: 54), Erwinia carotovora (Q6D4R4_ERWCT, SEQ ID NO: 55), Salmonella typhimurium (P74880_SALTY, SEQ ID NO: 56), Lactobacillus plantarum (Q88RY9_LACPL, SEQ ID NO: 57) и Lactococcus lactis (086282_9LACT, SEQ ID NO: 58). Выравнивание было произведено с использованием компьютерной программы PIR Multiple Alignment (http://pir.georgetown.edu). Идентичные аминокислоты отмечены звездочкой (*), похожие аминокислоты отмечены двоеточием (:).
На Фиг.3 представлены кривые роста модифицированных штаммов, выросших на минимальной среде М9, содержащей этанол (2% или 3%) в качестве единственного источника углерода.
На Фиг.4 представлены кривые роста модифицированных штаммов, выросших на минимальной среде М9, содержащей смесь глюкозы (0.1% по весу) с этанолом (0.1% по объему).
На Фиг.5 представлены кривые роста штаммов, имеющих мутацию в гене adhE*, при этом сам ген находится под контролем нативного промотора или рекомбинантного промотора PL-tac, выросших на минимальной среде М9, содержащей этанол (2% или 3%) в качестве единственного источника углерода.
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Получение штамма Е. coli MG1655 Δtdh, rhtA*.
Штамм-продуцент L-треонина Е. coli MG1655 Δtdh, rhtA* (pVIC40) был сконструирован путем инактивации нативного гена tdh, кодирующего треониндегидрогеназу, в штамме Е. coli MG1655(ATCC 700926) с использованием гена cat и последующего введения мутации rhtA23(rhtA*), которая придает штамму устойчивость к высоким концентрациям треонина (>40 мг/мл) и гомосерина (>5 мг/мл). Затем полученный штамм был трансформирован плазмидой pVIC40 из штамма Е. coli VKPM В-3996. Плазмида pVIC40 детально описана в U.S. Patent No. 5,705,371.
Для замещения нативного гена tdh фрагмент ДНК, содержащий маркер устойчивости к хлорамфениколу (CmR), кодируемый геном cat, был интегрирован в хромосому штамма Е. coli MG1655 (АТСС 700926) вместо нативного гена методом, описанным Datsenko К.А. и Wanner B.L. (Proc.Natl.Acad.Sci.USA, 2000, 97, 6640-6645), который также называется "Red-зависимая интеграция " и/или "Red-управляемая интеграция". Рекомбинантная плазмида pKD46 (Datsenko, К.А., Wanner, B.L., Proc.Natl.Acad.Sci.USA, 2000, 97, 6640-6645) с термочувствительным репликоном была использована в качестве донора генов фага X, ответственных за Red-зависимую систему рекомбинации. Штамм Escherichia coli BW25113 с инвентарным номером CGSC7630, содержащий рекомбинантную плазмиду pKD46, может быть получен в The Е. coli Genetic Stock Center, Yale University, New Haven, USA.
Фрагмент ДНК, содержащий маркер CmR, кодируемый геном cat, был получен с помощью ПЦР с использованием коммерческой плазмиды pACYC184 (GenBank/EMBL инвентарный номер Х06403, "Fermentas", Литва) в качестве матрицы и праймеров PI (SEQ ID NO: 3) и Р2 (SEQ ID NO: 4). Праймер Р1 содержит последовательность из 35 н., гомологичную 5'-области гена tdh, введенную в праймер для дальнейшей интеграции в хромосому бактерии. Праймер Р2 содержит последовательность из 32 н., гомологичную 3'-концу гена tdh, введенную в праймер для дальнейшей интеграции в хромосому бактерии.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера с 25 мМ MgCl2 ("Fermentas", Литва), по 200 мкМ каждого из dNTP, по 25 пкМ каждого из используемых праймеров и 1 Ед Taq-полимеразы ("Fermentas", Литва). Приблизительно 5 нг ДНК плазмиды было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 25 циклов: денатурация в течение 30 сек при 95°С, отжиг в течение 30 сек при 55°С, элонгация в течение 40 сек при 72°С; заключительный шаг: 5 мин при 72°С. Затем полученный продукт ПЦР-амплифицированный фрагмент ДНК был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом.
Полученный фрагмент ДНК был использован для электропорации и Red-зависимой интеграции в хромосому бактерии штамма Е. coli MG1655/pKD46.
Клетки MG1655/pKD46 выращивали в течение ночи при 30°С в жидкой LB среде, содержащей ампициллин (100 мкг/мл), разводили в 100 раз при помощи среды SOB (Дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; Триптон, 20 г/л; КСl, 2.5 мМ; MgCl2, 10 мМ), содержащей ампициллин(100 мкг/мл) и L-арабинозу (10 мМ) (арабиноза необходима для индуцирования плазмиды, содержащей гены Red-системы) и выращивали с аэрацией при 30°С до достижения значений оптической плотности OD600≈0.4-0.7.
Выросшие клетки из 10 мл бактериальной культуры отмывали 3 раза ледяной деионизированной Н2O и ресуспендировали в 100 мкл воды. Продукт ПЦР в объеме 10 мкл (что соответствует приблизительно 100 нг ДНК, растворенным в деионизованной воде) добавляли к суспензии клеток. Электропорацию проводили в электропораторе фирмы "Bio-Rad" (США) (No. 165-2098, version 2-89) в соответствии с инструкцией производителя.
После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2 часов, после чего высевали на чашки с L-агаром, содержащим хлорамфеникол в концентрации 25 мкг/мл.
Через 24 часа инкубации при 37°С выросшие колонии проверяли на присутствие в них вместо нативного гена tdh маркера CmR методом ПЦР с использованием праймеров РЗ (SEQ ID NO: 5) и Р4 (SEQ ID NO: 6). Для этой цели свежевыделенную колонию ресуспендировали в 20 мкл воды, после чего 1 мкл полученной суспензии тестировали в ПЦР. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 30 циклов: денатурация в течение 30 секунд при 95°С, отжиг в течение 30 сек при 55°С, элонгация в течение 30 сек при 72°С; заключительный шаг: 5 мин при 72°С. Несколько колоний с CmR маркером содержали желаемый фрагмент ДНК размером 1104 п.о., подтверждая присутствие ДНК-маркера СmR вместо фрагмента нативного гена tdh размером 1242 п.о. Одна из полученных колоний была излечена от термочувствительной плазмиды pKD46 путем культивирования при 37°С, и полученный таким образом штамм был назван Е. coli MG1655Δtdh.
Затем в полученный штамм MG1655 Δtdh была введена мутация rhtA23 из штамма VL614rhtA23 (Livshits V.A. et al, 2003, Res. Microbiol., 154:123-135), в результате чего был получен штамм MG1655 Δtdh, rhtA*. Мутация rhtA23 обусловливает устойчивость бактерии к высоким концентрациям треонина (>40 мг/мл) и гомосерина (>5 мг/мл). С этой целью штамм MG1655 Δtdh был инфицирован фагом Pivir. выращенным на донорском штамме VL614rhtA23. Трансдуктанты были отобраны на минимальной среде М9, содержащей 8 мг/мл гомосерина и 0.4% глюкозы в качестве единственного источника углерода.
Пример 2. Конструирование штамма Е. coli MG1655::PL-tacadhE. Штамм Е. coli MG1655::PL-tacadh был получен путем замены области нативного промотора гена adhE в штамме MG1655 на промотор РL-tac.
Для замены области нативного промотора гена adhE фрагмент ДНК, содержащий промотор PL-tac и маркер устойчивости к хлорамфениколу (CmR), кодируемый геном cat, был интегрирован в хромосому штамма Е. coli MG1655 вместо области нативного промотора методом, описанным Datsenko К.А. и Wanner B.L. (Proc.Natl.Acad.Sci.USA, 2000, 97, 6640-6645), который также называется "Red-зависимая интеграция " и/или "Red-управляемая интеграция".
Фрагмент, содержащий промотор РL-tac и ген cat, был получен методом ПЦР с использованием хромосомной ДНК штамма Е. coli MG1655PL-tacXylE (заявка РСТ WO2006/043730) в качестве матрицы. Нуклеотидная последовательность промотора PL-tac показана в Перечне последовательностей (SEQ ID NO: 7). Праймеры Р5 (SEQ ID NO: 8) и Р6 (SEQ ID NO: 9) были использованы для амплификации в ПЦР. Праймер Р5 содержит последовательность из 40 нуклеотидов, комплементарную области, расположенной за 318 п.о. до инициирующего кодона гена adhE, встроенную в праймер для дальнейшей интеграции в хромосому бактерии. Праймер Р6 содержит последовательность из 39 нуклеотидов, идентичную 5'-области гена adhE.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера с 15 мМ MgCl2 ("Fermentas", Литва), по 200 мкМ каждого из dNTP, по 25 пкМ каждого из используемых праймеров и 1 Ед Taq-полимеразы ("Fermentas", Литва). Приблизительно 20 нг геномной ДНК штамма E.coli МG1655РL-tacxуlЕ было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации.
Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 35 циклов: денатурация в течение 30 секунд при 95°С, отжиг в течение 30 сек при 54°С, элонгация в течение 90 сек при 72°С; заключительный шаг: 5 мин при 72°С. Затем полученный продукт ПЦР-амплифицированный фрагмент ДНК был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом. Полученный фрагмент ДНК был использован для электропорации и Red-зависимой интеграции в хромосому бактерии штамма Е. coli MG1655/pKD46.
Клетки MG1655/pKD46 выращивали в течение ночи при 30°С в жидкой LB среде, содержащей ампициллин (100 мкг/мл), разводили в 100 раз при помощи среды SOB (Дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; Триптон, 20 г/л; КСl, 2.5 мМ; MgCl2, 10 мМ), содержащей ампициллин(100 мкг/мл) и L-арабинозу (10 мМ) (арабиноза необходима для индуцирования плазмиды, содержащей гены Red-системы), и выращивали с аэрацией при 30°С до достижения значений оптической плотности OD600≈0.4-0.7. Выросшие клетки из 10 мл бактериальной культуры трижды отмывали ледяной деионизированной Н2O и ресуспендировали в 100 мкл воды. Продукт ПЦР в объеме 10 мкл (что соответствует приблизительно 100 нг ДНК, растворенным в деионизованной воде) добавляли к суспензии клеток. Электропорацию проводили в электропораторе фирмы "Bio-Rad" (США) (No. 165-2098, version 2-89) в соответствии с инструкцией производителя.
После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2 часов, после чего высевали на чашки с L-агаром, содержащим хлорамфеникол в концентрации 25 мкг/мл.
Около 100 полученных клонов было отобрано на чашках со средой М9, содержащей 2% этанола в качестве единственного источника углерода. Несколько клонов, которые выросли на чашках с М9-средой, содержащей 2% этанола, в течение 36 часов, были отобраны и протестированы на присутствие в них вместо области нативного промотора гена adhE маркера CmR методом ПЦР с использованием праймеров Р7 (SEQ ID NO: 10) и Р8 (SEQ ID NO: 11). Для этих целей свежевыделенную колонию ресуспендировали в 20 мкл воды, после чего 1 мкл полученной суспензии тестировали в ПЦР. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 10 мин; профиль для последующих 30 циклов: денатурация в течение 30 секунд при 95°С, отжиг в течение 30 сек при 54°С, элонгация в течение 1.5 мин при 72°С; заключительный шаг: 1 мин при 72°С. Несколько протестированных колоний с СmR маркером содержали желаемый фрагмент ДНК размером ~1800 п.о., подтверждая, таким образом, присутствие ДНК-маркера СmR вместо области нативного промотора гена adhE размером 520 п.о. Одна из полученных колоний была излечена от термочувствительной плазмиды pKD46 путем культивирования при 37°С, и полученный таким образом штамм был назван Е. coli MG1655::PL-tacadhE (см. Фиг.1).
Пример 3. Конструирование штамма Е. coli MG1655Δtdh, rhtA, PL-tacadhE.
Штамм Е. coli MG1655Δtdh, rhtA*, PL-tacadhE был получен путем трансдукции промотора РL-tac из штамма MG1655::PL-tacadhE в штамм MG1655Δtdh, rhtA*.
Штамм MG1655 Δtdh, rhtA был инфицирован фагом Pivir, выросшим на донорском штамме MG1655::PL-tacadhE, и в результате был получен штамм MG1655Δtdh, rhtA*, pL-tacadhE. Ростовые качества этого штамма были проверены на чашках со средой М9, содержащей 2% этанола в качестве единственного источника углерода. Скорость роста была той же, что и у штамма MG1655::PL-tacadhE.
Пример 4. Влияние усиления экспрессии гена adhE на продукцию треонина.
Для оценки влияния усиления экспрессии гена adhE на продукцию L-треонина, оба штамма E.coli, MG1655Δtdh, rhtA*, PL-tacadhE и MG1655Δtdh, rhtА*, были трансформированы плазмидой pVIC40.
Полученные штаммы MG1655Δtdh, rhtA*, PL-tacadhE (pVIC40) и MG1655Δtdh, rhtA* (pVIC40) выращивали при 37°С в течение 18 часов в питательном бульоне и по 0.3 мл каждой из полученных культур были инокулированы в 3 мл среды для ферментации нижеследующего состава в пробирки размером 20х200 мм и выращивались при 34°С в течение 48 часов на роторной качалке. Результаты по крайней мере 10 независимых ферментации в пробирках показаны в Таблицах 1 и 2.
Состав среды для ферментации, г/л:
MgSO4·7H2O и СаСО3 стерилизуются раздельно.
Как можно видеть из Таблиц 1 и 2, штамм MG1655Δtdh, rhtA*, PL-tacadhE был способен накапливать более высокое количество L-треонина по сравнению со штаммом MG1655Δtdh, rhtA*. Кроме того, штамм MG1655Δtdh, rhtA*, PL-tacadhE был способен расти на среде, содержащей этанол в качестве единственного источника углерода и вызывать при этом накопление треонина, тогда как штамм MG1655Δtdh, rhtA* рос очень медленно и имел низкую продуктивность в среде, содержащей этанол в качестве единственного источника углерода.
Пример 5. Конструирование штамма Е. coli MG1655ΔadhE.
Этот штамм был сконструирован путем инактивации нативного гена adhE в штамме Е. coli MG1655 с использованием гена kan.
Для инактивации (или разрушения) нативного гена adhE фрагмент ДНК, содержащий маркер устойчивости к канамицину (KmR), кодируемый геном kan, был интегрирован в хромосому Е. coli MG1655 (АТСС 700926) вместо нативного гена методом, описанным Datsenko K.A. and Wanner B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645), который также называется "Red-зависимая интеграция" и/или "Red-управляемая интеграция".
Фрагмент ДНК, содержащий маркер КmR, кодируемый геном kan, был получен методом ПЦР с использованием коммерческой плазмиды pACYC177 (GenBank/EMBL инвентарный номер Х06402, "Fernentas", Литва) в качестве матрицы и праймеров Р9 (SEQ ID NO: 12) и P10 (SEQ ID NO: 13). Праймер Р9 содержит последовательность из 40 н,, гомологичную области, расположенной за 318 п.о. до инициирующего кодона гена adhE, введенную в праймер для дальнейшей интеграции в хромосому бактерии. Праймер Р10 содержит последовательность из 41 н., гомологичную 3'-области гена adhE, введенную в праймер для дальнейшей интеграции в хромосому бактерии.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера с 25 мМ MgCl2 ("Fermentas", Литва), по 200 мкМ каждого из dNTP, по 25 пкМ каждого из используемых праймеров и 1 ед Taq-полимеразы ("Fermentas", Литва). Приблизительно 5 нг плазмидной ДНК было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации.
Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 25 циклов: денатурация в течение 30 секунд при 95°С, отжиг в течение 30 сек при 55°С, элонгация в течение 40 сек при 72°С; заключительный шаг: 5 мин при 72°С. Затем полученный продукт ПЦР-амплифицированный фрагмент ДНК был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом.
Полученный фрагмент ДНК был использован для электропорации и Red-зависимой интеграции в хромосому бактерии штамма Е. coli MG1655/pKD46.
Клетки MG1655/pKD46 выращивали в течение ночи при 30°С в жидкой LB среде, содержащей ампициллин (100 мкг/мл), разводили в 100 раз при помощи среды SOB (дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; Триптон, 20 г/л; КСl, 2.5 мМ; MgCl2, 10 мМ), содержащей ампициллин(100 мкг/мл) и L-арабинозу (10 мМ) (арабиноза необходима для индуцирования генов плазмиды из Red-системы) и выращивали с аэрацией при 30°С до достижения значений оптической плотности OD600≈0.4-0.7.
Выросшие клетки из 10 мл бактериальной культуры трижды отмывали ледяной деионизированной Н2О и ресуспендировали в 100 мкл воды. Продукт ПЦР в объеме 10 мкл (что соответствует приблизительно 100 нг ДНК, растворенным в деионизованной воде) добавляли к суспензии клеток. Электропорацию проводили в электропораторе фирмы "Bio-Rad" (США) (No. 165-2098, version 2-89) в соответствии с инструкцией производителя.
После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°C в течение 2 часов, после чего высевали на чашки с L-агаром, содержащим канамицин в концентрации 20 мкг/мл.
Колонии, выросшие в течение 24 часов, тестировали на присутствие в них вместо области нативного гена adhE маркера КmR методом ПЦР с использованием праймеров Р11 (SEQ ID NO: 14) и Р12 (SEQ ID NO: 15). Для этой цели свежеприготовленную колонию ресуспендировали в 20 мкл воды и затем 1 мкл полученной суспензии тестировали в ПЦР. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°C в течение 5 мин; профиль для последующих 30 циклов: денатурация в течение 30 секунд при 95°C, отжиг в течение 30 сек при 55°C, элонгация в течение 30 сек при 72°С; заключительный шаг: 5 мин при 72°C. Несколько колоний с KmR маркером содержали желаемый фрагмент ДНК размером 1030 п.о., подтверждая присутствие ДНК-маркера KmR вместо области нативного гена adhE размером 3135 п.о. Одна из полученных колоний была излечена от термочувствительной плазмиды pKD46 путем культивирования ее при 37°C, и полученный таким образом штамм был назван штамм Е. coli MG1655ΔadhE.
Пример 6. Конструирование штамма Е. coli strain MG1655::PL-tacadhE* Штамм Е. coli MG1655::РL-tас adhE* был получен путем введения мутации Glu568Lys (Е568К) в ген adhE. На первой стадии методом ПЦР с использованием геномной ДНК штамма Е. coli MG1655 в качестве матрицы и праймеров Р13 (SEQ ID NO: 16) и Р12 (SEQ ID NO: 15) был получен фрагмент гена adhE размером 1.05 тыс. п.о., несущий мутацию Е568К. Праймер Р13 содержит последовательность, гомологичную областям 1662-1701 п.о. и 1703-1730 п.о. в гене adhE и включает замену g/a (в положении 1702 п.о.), выделенную жирным шрифтом, а праймер Р12 содержит последовательность, гомологичную 3'-концу гена adhE.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера с MgCl2 ("ТаКаКа", Япония), по 250 мкМ каждого из dNTP, по 25 пкМ каждого из используемых праймеров и 2.5 ед Pyrobest ДНК-полимеразы ("TaKaRa", Япония). Приблизительно 20 нг геномной ДНК штамма MG1655 было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации.
Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 35 циклов: денатурация в течение 30 секунд при 95°С, отжиг в течение 30 сек при 54°С, элонгация в течение 1 мин при 72°С; заключительный шаг: 5 мин при 72°С. Затем полученный продукт ПЦР-амплифицированный фрагмент ДНК был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом.
На второй стадии фрагмент ДНК, содержащий область соединения промотора
РL-tac c мутантным геном adhE и маркированный геном cat, который обеспечивает устойчивость к хлорамфениколу, был получен методом ПЦР с использованием геномной ДНК штамма Е. coli MG1655::PL-tacadhE в качестве матрицы (см. Пример 2), праймера P11 (SEQ ID NO: 14) и фрагмента ДНК размером 1.05 тыс.п.о., несущего мутантную последовательность (см.выше) в качестве второго праймера. Праймер Р11 содержит последовательность, гомологичную области, расположенной за 402-425 п.о. до инициирующего кодона гена adhE.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера ("TaKaRa", Япония), 25 мМ MgCl2, по 250 мкМ каждого из dNTP, 10 нг праймера Р11, 1 мкг фрагмента размером 1.05 тыс.п.о. в качестве второго праймера и 2.5 Ед TaKaRaLA ДНК-полимеразы ("TaKaRa", Япония). Приблизительно 20 нг геномной ДНК штамма Е. coli MG1655::PL-tacadhE было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации.
Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 35 циклов: денатурация в течение 30 секунд при 95°С, отжиг в течение 30 сек при 54°С, элонгация в течение 3.5 мин при 72°С; заключительный шаг: 7 мин при 72°С. Затем полученный продукт ПЦР-амплифицированный фрагмент ДНК был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом.
Для того чтобы заменить нативную область гена adhE, фрагмент ДНК, несущий промотор PL-tac с мутантным геном adhE и маркером устойчивости к хлорамфениколу (СmR), кодируемому геном cat (cat-PL-tacadhE*, 4.7 kbp), был интегрирован в хромосому штамма Е. coli MG1655ΔadhE методом, описанным Datsenko К.A. and Wanner B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97, 6640-6645), который также называется "Red-зависимая интеграция " и/или "Red-управляемая интеграция". Клетки штамма MG1655 ΔadhE/pKD46 выращивали в течение ночи при 30°С в жидкой LB-среде, содержащей ампициллин (100 мкг/мл), и затем разводили в 100 раз при помощи 5 мл среды SOB (Дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; Триптон, 20 г/л; КСl, 2.5 мМ; MgCl2, 10 мМ), содержащей ампициллин(100 мкг/мл) и L-арабинозу (10 мМ) (арабиноза необходима для индуцирования плазмиды с генами Red-системы). Полученную культуру выращивали с аэрацией при 30°С до достижения значений оптической плотности OD600≈0.4-0.7. Выросшие клетки из 10 мл бактериальной культуры отмывали 3 раза ледяной деионизированной H2O и ресуспендировали в 100 мкл воды. Продукт ПЦР в объеме 10 мкл (что соответствует приблизительно 300 нг ДНК, растворенным в деионизованной воде) добавляли к суспензии клеток. Электропорацию проводили в электропораторе фирмы "Bio-Rad" (США) (No. 165-2098, version 2-89) в соответствии с инструкцией производителя.
После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°C в течение 2 часов, после чего высевали на чашки с L-агаром, содержащим хлорамфеникол в концентрации 25 мкг/мл.
Выросшие колонии были отобраны на чашках со средой М9, содержащей 2% этанола в качестве единственного источника углерода.
Среди выросших клонов был выбран быстрорастущий клон, в котором было проведено полноразмерное секвенирование гена adhE. В результате был выявлен следующий ряд мутаций: Glu568Lys (gag-aag), Ile554Ser (ate-age), Glu22Gly (gaa-gga), Met236Val (atg-gtg), Ala786Val (gca-gta). Этот клон был назван MG1655::PL-tacadhE*.
Пример 7. Конструирование штамма Е. coli MG1655Δtdh, rhtA, PL-tacadhE*.
Штамм Е. coli MG1655Δtdh, rhtA, PL-tacadhE* был получен путем трансдукции мутации PL-tac adhE* из штамма MG1655::PL-tacadhE*.
Штамм MG1655 Δtdh, rhtA был инфицирован фагом Plvir, выросшим на донорском штамме MG1655::PL-tacadhE*, и в результате был получен штамм MG1655Δtdh, rhtA,
PL-tacadhE*. Ростовые качества этого штамма были проверены на чашках со средой М9, содержащей 2% этанола в качестве единственного источника углерода. Скорость роста вновь полученного штамма была той же, что и штамма MG1655::PL-tacadhE*.
Пример 8. Конструирование штамма Е. coli strain MG1655Δtdh, rhtA, PL-tacadhE-Lys568.
Была осуществлена вторая попытка получения точечной мутации Glu568Lys в гене adhE. Для этой цели был сконструирован штамм Е. coli MG1655Δtdh, rhtA, PL-tacadhE-wtΔ34.
Штамм Е., coli MG1655Δtdh, rhtA, PL-tacadhE-wtΔ34 был получен путем замены гена kan в штамме Е. coli MG1655Δtdh, rhtA, PL-tacadhE-wt (wt означает природный тип) на фрагмент гена adhE размером 34 п.о. (область 1668-1702 п.о., включающая триплет, кодирующий Glu568). Ген kan был интегрирован в хромосому Е. coli MG1655Δtdh, rhtA, PL-tacadhE-wt методом, описанным Datsenko K.A. and Wanner B.L. (Proc.Natl.Acad.Sci.USA, 2000, 97, 6640-6645), который также называется "Red-зависимая интеграция " и/или "Red-управляемая интеграция".
Фрагмент ДНК, содержащий маркер KmR, кодируемый геном kan, был получен методом ПЦР с использованием коммерческой плазмиды pACYC177 (GenBank/EMBL инвентарный номер Х06402, "Fermentas", Литва) в качестве матрицы и праймеров Р14 (SEQ ID NO: 17) и P15 (SEQ ID NO: 18). Праймер Р14 содержит последовательность из 41 н., идентичную области в гене adhE 1627-1668 п.о., а праймер Р15 содержит последовательность из 39 н., комплементарную области в гене adhE 1702-1740 п.о., введенные в праймеры для дальнейшей интеграции в хромосому бактерии.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера с 25 мМ MgCl2 ("Fermentas", Литва), по 200 мкМ каждого из dNTP, по 25 пкМ каждого из используемых праймеров и 1 Ед Taq-полимеразы ("Fermentas", Литва). Приблизительно 5 нг ДНК плазмиды было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 25 циклов: денатурация в течение 30 сек при 95°С, отжиг в течение 30 сек при 55°С, элонгация в течение 50 сек при 72°С; заключительный шаг: 5 мин при 72°С. Затем полученный ПЦР-амплифицированный фрагмент ДНК был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом.
Выросшие колонии проверяли на присутствие в них маркера КmR методом ПЦР с использованием праймеров Р16 (SEQ ID NO: 19) и P17 (SEQ ID NO: 20). Для этой цели свежевыделенную колонию ресуспендировали в 20 мкл воды, после чего 1 мкл полученной суспензии тестировали в ПЦР. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 30 циклов: денатурация в течение 30 сек при 95°С, отжиг в течение 30 сек при 55°С, элонгация в течение 45 сек при 72°С; заключительный шаг: 5 мин при 72°С. Несколько колоний с KmR маркером содержали желаемый фрагмент ДНК размером 1200 п.о., подтверждая, таким образом, присутствие ДНК-маркера KmR вместо фрагмента нативного гена adhE размером 230 п.о. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 путем культивирования при 37°С и полученный таким образом штамм был назван Е. coli MG1655Δtdh, rhtA, PL-tacadhE-wtΔ34.
Затем, для того, чтобы осуществить замену кодируемого геном kan маркера устойчивости к канамицину (KmR) на фрагмент гена adhE, кодирующего мутацию Glu568Lys, в хромосому штамма Е. coli MG1655Δtdh, rhtA, PltacadhE-wt Δ34 методом «Red-зависимой» и/или «Red-управляемой» интеграции (Yu D., Sawitzke J. et al., Recombineering with overlapping single-stranded DNA oligonucleotides: Testing of recombination intermediate, PNAS, 2003, 100(12), 7207-7212) были интегрированы олигонуклеотиды Р18 (SEQ ID NO: 21) и P19 (SEQ ID NO: 22), несущие соответствующую мутацию. Праймер Р18 содержит последовательность из 75 н., идентичную участку нативного гена adhE 1627-1702 п.о., а праймер Р19 содержит последовательность из 75 н., комплементарную участку гена adhE 1668-1740 п.о., при этом оба праймера включают триплет, кодирующий Lys568 вместо Glu568.
Колонии были отобраны на минимальной среде М9, содержащей в качестве источника углерода 2% этанола и 25 мг/мл сукцината.
Полученные колонии были протестированы на отсутствие в них области маркера KmR методом ПЦР с использованием праймеров Р16 (SEQ ID NO: 19) и P17 (SEQ ID NO: 20). Для этих целей свежевыделенную колонию ресуспендировали в 20 мкл воды, после чего 1 мкл полученной суспензии тестировали в ПЦР. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 30 циклов: денатурация в течение 30 сек при 95°С, отжиг в течение 30 сек при 55°С, элонгация в течение 25 сек при 72°С; заключительный шаг: 5 мин при 72°С. Несколько колоний, протестированных на наличие области маркера KmR, содержали желаемый фрагмент ДНК размером 230 п.о. из гена adhE, подтверждая тем самым отсутствие ДНК маркера KmR вместо фрагмента ДНК размером 1200 п.о. Несколько полученных штаммов были излечены от термочувствительной плазмиды pKD46 путем культивирования их при 37°С и полученный таким образом штамм был назван E.coli MG1655Δtdh, rhtA*, PL-tacadhE-Lys568.
Присутствие в полученных штаммах мутации Glu568Lys было подтверждено результатами секвенирования. Кроме того, было обнаружено, что некоторые клоны (№1,13) содержали дополнительную мутацию: клон №1 - Glu568Lys, Phe566Val; клон №13 - Glu568Lys, Glu560Lys.
Для штаммов MG1655Δtdh, rhtA*, PL-tacadhE-Lys568 (кл.18), MG1655Δtdh, rhtA*,
РL-tacadhE-Lys568, Val566 (кл.1), MG1655Δtdh, rhtA*, PL-tacadhE-Lys568, Lys560 (кл.13) и MG1655Δtdh, rhtA*, PL-tacadhE* были определены кривые роста.
Штаммы выращивали в среде М9, содержащей этанол в качестве единственного источника углерода, и в среде М9, содержащей глюкозу и этанол (в молярном соотношении 1:3).
Пример 9. Конструирование штамма Е. coli MG1655Δtdh, rhtA*, adhE*.
Штамм Е. coli MG1655Δtdh, rhtA*, adhE* был получен путем реконструкции нативного промотора гена adhE в штамме MG1655Δtdh, rhtA*, PL-tacadhE*. Фрагмент ДНК, несущий промотор PL-tac и маркер устойчивости к хлорамфениколу (CmR), кодируемый геном cat, был заменен на фрагмент ДНК, несущий нативный промотор гена adhE и маркер устойчивости к канамицину (KmR), кодируемый геном kan. Нативный промотор гена adhE PadhE был получен методом ПЦР с использованием хромосомной ДНК из штамма MG1655 в качестве матрицы и праймеров Р20 (SEQ ID NO: 23) и Р21 (SEQ ID NO: 24). Праймер Р20 содержит на 5'-конце сайт узнавания рестриктазой ЕсоRI, который необходим для дальнейшего лигирования с геном kan, а праймер Р21 содержит последовательность из 30 н., гомологичных 5'-области гена adhE (50-20 п.о.)
Фрагмент ДНК, содержащий последовательность маркера КmR, кодируемого геном kan, был получен методом ПЦР с использованием коммерческой плазмиды pACYC177 (GenBank/EMBL инвентарный номер Х06402, "Fermentas", Литва) в качестве матрицы и праймеров Р22 (SEQ ID NO: 25) и Р23 (SEQ ID NO: 26). Праймер Р22 содержит последовательность из 41 н., гомологичную области, расположенной за 425 п.о. до инициирующего кодона гена adhE и включенную в состав праймера для дальнейшей интеграции в хромосому бактерии, а праймер Р23 содержит на 3'-конце сайт узнавания рестриктазой EcoRI, который необходим для дальнейшего соединения с промотором PadhE.
ПЦР проводили в амплификаторе "Gene Amp PCR System 2700" (Applied Biosystems, США). Реакционная смесь (общий объем - 50 мкл) состояла из 5 мкл 10-кратного ПЦР-буфера с 25 мМ MgCl2 ("Fermentas", Литва), по 200 мкМ каждого из dNTP, по 25 пкМ каждого из используемых праймеров и 1 Ед Таq-полимеразы ("Fermentas", Литва). Приблизительно 20 нг геномной ДНК или 5 нг ДНК плазмиды было добавлено в реакционную смесь в качестве матричной ДНК для ПЦР-амплификации. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 35 циклов в случае с геномной ДНК или для последующих 25 циклов в случае плазмидной ДНК: денатурация в течение 30 сек при 95°С, отжиг в течение 30 сек при 55°С, элонгация при 72°С в течение 20 сек в случае с геномной ДНК или в течение 50 сек в случае с плазмидной ДНК; заключительный шаг: 5 мин при 72°С. Затем полученные ПЦР-амплифицированные фрагменты ДНК были очищены в агарозном геле, выделены с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осаждены спиртом.
Каждый из двух описанных выше фрагментов ДНК был обработан рестриктазой EcoRI и затем фрагменты лигировали. Продукт лигирования был амплифицирован методом ПЦР с использованием праймеров Р21 и Р22. Амплифицироанный фрагмент ДНК kan-PadhE был очищен в агарозном геле, выделен с помощью колонок "GenElute Spin Columns" ("Sigma", США) и осажден спиртом. Полученный фрагмент ДНК был использован для электропорации и Red-зависимой интеграции в хромосому бактерии штамма E.coli MG1655Δtdh:: rhtA*, PL-tacadhE*/pKD46.
Клетки MG1655Δtdh::rhtA*, PL-tacadhE*/pKD46 выращивали в течение ночи при 30°С в жидкой LB среде, содержащей ампициллин (100 мкг/мл), разводили в 100 раз при помощи среды SOB (Дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; Триптон, 20 г/л; КСl, 2.5 мМ; MgCl2, 10 мМ), содержащей ампициллин(100 мкг/мл) и L-арабинозу (10 млМ) (арабиноза необходима для плазмиды с генами Red-системы). Полученную культуру выращивали с аэрацией при 30°С до достижения значений оптической плотности CD600≈0.4-0.7. Выросшие клетки из 10 мл бактериальной культуры отмывали 3 раза ледяной деионизированной Н2O и ресуспендировали в 100 мкл воды. Продукт ПЦР в объеме 10 мкл (что соответствует приблизительно 100 нг ДНК, растворенным в деионизованной воде) добавляли к суспензии клеток. Электропорацию проводили в электропораторе фирмы "Bio-Rad" (США) (No. 165-2098, version 2-89) в соответствии с инструкцией производителя. После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2 часов, после чего высевали на чашки с L-агаром, содержащим канамицин в концентрации 20 мкг/мл. Через 24 часа инкубации при 37°С выросшие колонии проверяли на присутствие в них маркера PgdhE-KmR вместо маркера РL-tac-СmR методом ПЦР с использованием праймеров Р24 (SEQ ID NO: 27) и Р25 (SEQ ID NO: 28). Для этой цели свежевыделенную колонию ресуспендировали в 20 мкл воды, после чего 1 мкл полученной суспензии тестировали в ПЦР. Использовали следующие температурные условия для амплификации с помощью ПЦР: первоначальная денатурация ДНК при 95°С в течение 5 мин; профиль для последующих 30 циклов: денатурация в течение 30 сек при 95°С, отжиг в течение 30 сек при 54°С, элонгация в течение 1 мин при 72°С; заключительный шаг: 5 мин при 72°С.
Несколько колоний с KmR маркером содержали желаемый фрагмент ДНК размером 1200 п.о., подтверждая, таким образом, присутствие промотора PadhE и ДНК-маркера КmR. Некоторые из этих фрагментов были просеквенированы. Структура нативного промотора PadhE была подтверждена. Один из полученных штаммов, содержащий мутантный ген adhE под контролем нативного промотора, был излечен от термочувствительной плазмиды pKD46 путем культивирования при 37°С, и полученный таким образом штамм был назван Е. coli MG1655Δtdh, rhtA*, adhE*.
Была изучена способность всех полученных штаммов MG1655Δtdh, rhtA*, pl-tacadhE; MG1655Δtdh, rhtA*, PL-tacadhE*; MG1655Δtdh, rhtA*, PltacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PltacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*, adhE* и родительского штамма MG1655Δtdh, rhtA* расти на минимальной среде М9, содержащей этанол в качестве единственного источника углерода. Было показано, что родительский штамм MG1655Δtdh, rhtA* и штамм с усиленной экспрессией гена алкогольдегидрогеназы природного типа не способны расти на среде, содержащей этанол (2% или 3%) в качестве единственного источника углерода (фиг.3. A и В). Штамм MG1655Δtdh, rhtA*, PltacadhE-Lys568 (кл.18), содержащий точечную мутацию в алкогольдегидрогеназе, описанную ранее (Membrillo-Hemandez, J. et al, J. Biol. Chem. 275, 33869-33875 (2000)), рос плохо в среде вышеуказанного состава. Однако штаммы, содержащие дополнительные мутации в алкогольдегидрогеназе (помимо мутации Glu568Lys), росли хорошо (Фиг.3, А и В). Все вышеописанные штаммы росли на минимальной среде М9, содержащей смесь глюкозы с этанолом, однако штаммы с усиленной экспрессией мутантной алкогольдегидрогеназы, содержащие дополнительные мутации (помимо мутации Glu568Lys), росли лучше (Фиг.4).
Было показано, что штамм MG1655Δtdh, rhtA*, adhE*, содержащий алкогольдегидрогеназу с пятью мутациями под контролем нативного промотора, не способен расти на минимальной среде М9, содержащей этанол (2% или 3%) в качестве единственного источника углерода. Для хорошего роста необходимым условием является усиленная экспрессия гена, кодирующего алкогольдегидрогеназу (Фиг.5).
Пример 10. Влияние усиления экспрессии мутантного гена adhE на продукцию L-треонина.
Для оценки влияния усиления экспрессии мутантного гена adhE на продукцию L-треонина штаммы Е. coli MG1655Δtdh, rhtA*, PL-tacadhE; MG1655Δtdh, rhtA*, PL-tacadhE*; MG1655Δtdh, rhtA*, PltacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PitacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*, adhE* и родительский штамм MG1655Δtdh, rhtA* были трансформированы плазмидой pVIC40.
Полученные штаммы и родительский штамм MG1655Δtdh, rhtA* (pVIC40) выращивали при 37°С в течение 18 часов в питательном бульоне и 0.3 мл каждой из полученных культур инокулировали в 3 мл среды для ферментации (см. Пример 4) в пробирки размером 20×200 мм и выращивали при 34°С в течение 48 часов на роторной качалке. Результаты 10 независимых ферментаций в пробирках показаны в Таблицах 1 и 2.
Как можно видеть из Таблиц 1 и 2, наличие в штамме мутантной алкогольдегидрогеназы приводит к накоплению этим штаммом более высокого количества L-треонина по сравнению со штаммом MG1655Δtdh, rhtA*, в котором не была усилена экспрессия алкогольдегидрогеназы ни природного типа, ни мутантной, или даже по сравнению со штаммами MG1655Δtdh, rhtA* или PL-tacadhE, в которых экспрессия алкогольдегидрогеназы природного типа была усилена. Более высокое накопление L-треонина в процессе ферментации наблюдалось как в среде, содержащей смесь глюкозы с этанолом, так и в среде, содержащей только этанол в качестве единственного источника углерода.
Пример 11. Влияние усиления экспрессии гена adhE на продукцию L-лизина.
Для проверки влияния усиления экспрессии гена adhE под контролем промотора
PL-tac на продукцию лизина, фрагменты ДНК из хромосомы описанных выше штаммов Е. coli MG1655Δtdh, rhtA*, PL-tacadhE; MG1655Δtdh, rhtA*, PL-tacadhE*; MG1655Δtdh, rhtA*, PitacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PltacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*,adhE* переносили в штамм-продуцент лизина Е. coli WC196 (pCABD2) с помощью Р1 трансдукции (Miller, J.H. Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, 1972, Plainview, NY). pCABD2 - это плазмида, содержащая ген dapA, кодирующий мутантную дигидропиколинатсинтазу, устойчивую к ингибированию L-лизином по типу обратной связи, ген lysC, кодирующий мутантную аспартокиназу III, устойчивую к ингибированию L-лизином по типу обратной связи, ген dapB, кодирующий дигидропиколинатредуктазу, и ген ddh, кодирующий диаминопимелатдегидрогеназу (патент США 6,040,160).
Полученные штаммы и родительский штамм WC196(pCABD2) высевали на чашки с L-средой, содержащей 20 мг/л стрептомицина, и инкубировали при 37°С, выросшие клетки с 1/8 поверхности чашки инокулировали в 20 мл среды для ферментации, содержащей необходимые антибиотики, в колбы объемом 500 мл.
Культивирование производили при 37°С в течение 16 часов с использованием возвратно-поступательной качалки со скоростью перемешивания 115 об/мин. После выращивания количества L-лизина и остаточного этанола в среде измеряли известным способом (Biotech-analyzer AS210, производитель - Sakura Seiki Co.). Затем для каждого из штаммов рассчитывали выход L-лизина в пересчете на потребленный этанол. Состав использованной среды для ферментации, г/л:
рН доводили до 7.0 с помощью КОН, среду автоклавировали при 115°С в течение 10 мин. Глюкозу и MgSO4·7H2O стерилизуют раздельно. СаСО3 стерилизовали сухим жаром при температуре 180°С в течение 2 часов и добавляли в среду до конечной концентрации 30 г/л. Данные двух параллельных экспериментов представлены в Таблице 3.
Как видно из Таблицы 3, экспрессия мутантной алкогольдегидрогеназы и алкогольдегидрогеназы природного типа способствовала усилению роста и накапливанию большего количества L-лизина по сравнению с WC196ΔcadAΔldc (pCABD2), в котором не усилена экспрессия ни алкогольдегидрогеназы природного типа, ни мутантной алкогольдегидрогеназы.
Пример 12. Конструирование штамма E. coli MG1655ΔargR, PL-tacadhE*.
1. Конструирование штамма MG1655ΔargR.
Данный штамм был сконструирован путем инактивации нативного гена argR, кодирующего репрессор пути биосинтеза L-аргинина в Е. coli MG1655 геном kan. Для замены нативного гена argR фрагмент ДНК, содержащий маркер устойчивости к канамицину (КmR), кодируемый геном kan, был интегрирован в хромосому Е. coli MG1655 (АТСС 700926) на место нативного гена argR методом Red-зависимой интеграции. Штамм Е. coli BW25113, содержащий рекомбинантную плазмиду pKD46, может быть получен в E.coli Genetic Stock Center, Yale University, New Haven, USA (инвентарный номер штамма CGSC7630).
Фрагмент ДНК, содержащий маркер KmR, кодируемый геном kan, был получен методом ПЦР с использованием коммерческой плазмиды pACYC177 (GenBank/EMBL accession number X06402, "Fermentas", Lithuania) в качестве матрицы и праймеров Р26 (SEQ ID NO: 31) и Р27 (SEQ ID NO: 32). Праймер Р26 содержит 40 н., гомологичных 5'-области гена argR, введенных в праймер для дальнейшей интеграции в бактериальную хромосому. Праймер Р27 содержит 41 н., гомологичных 3'-области гена argR, введенных в праймер для дальнейшей интеграции в бактериальную хромосому. Использовали следующие температурные условия: начальная денатурация ДНК в течение 5 мин при 95°С, следующие 25 циклов: денатурация при 95°С в течение 30 сек, отжиг при 55°С в течение 30 сек, элонгация при 72°С в течение 40 сек; заключительная элонгация в течение 5 мин при 72°С. Затем амплифицированный фрагмент ДНК был очищен с использованием электрофореза в агарозном геле, выделен с использованием "GenElute Spin Columns" ("Sigma", USA) и осажден этанолом.
Полученный фрагмент ДНК использовали для электропорации и Red-зависимой интеграции в бактериальную хромосому Е. coli MG1655/pKD46.
Клетки MG1655/pKD46 выращивали в течение ночи при 30°С в жидкой среде LB, содержащей ампициллин (100 мкг/мл), затем разводили в отношении 1:100 средой SOB (дрожжевой экстракт, 5 г/л; NaCl, 0.5 г/л; триптон, 20 г/л; КСl, 2.5 мМ; MgCl2, 10 мМ) с добавлением ампициллина (100 мкг/мл) и L-арабинозы (10 мМ) (арабиноза используется для индукции плазмиды с генами Red системы) и выращивали при 30°С до достижения оптической плотности бактериальной культуры OD600≈0.4-0.7. Выросшие клетки из 10 мл бактериальной плазмиды трижды отмывали ледяной деионизированной водой, затем суспендировали в 100 мкл воды. 10 мкл содержащего ДНК раствора, что соответствует 100 нг фрагмента ДНК, растворенного в деионизированной воде, добавляли к суспензии клеток. Электропорацию осуществляли на электропораторе "Bio-Rad" (USA) (No. 165-2098, version 2-89) в соответствии с инструкцией производителя. Затем клетки добавляли 1 мл среды SOC, инкубировали в течение 2 ч при 37°С, высевали на чашки с L-агаром, содержащие 25 мкг/мл хлорамфеникола. Колонии, выросшие за 24 ч, тестировали на присутствие маркера КmR вместо нативного гена argR методом ПЦР с использованием праймеров Р28 (SEQ ID NO: 33) и Р29 (SEQ ID NO: 34). С этой целью свежевыделенные колонии суспендировали в 20 мкл воды и затем 1 мкл полученной суспензии использовали для ПЦР. Использовали следующие температурные условия: начальная денатурация ДНК в течение 5 мин при 95°С; следующие 30 циклов: денатурация при 95°С в течение 30 сек, отжиг при 55°С в течение 30 сек и элонгация при 72°С в течение 30 сек; заключительная элонгация в течение 5 мин при 72°С. Несколько протестированных клонов KmR содержали желаемый фрагмент ДНК длиной 1110 п.о., подтверждая присутствие ДНК маркера KmR вместо фрагмента гена argR длиной 660 п.о.. Один из полученных штаммов был излечен от термочувствительной плазмиды pKD46 культивированием при 37°С, полученный штамм был назван E.coli MG1655ΔargR.
2. Конструирование штамма Е. coli MG1655ΔargR, PL-tacadhE*.
Штамм Е. coli MG1655ΔargR,PL-tacadhE* был получен путем трансдукции мутации
PL-tac adhE* из штамма MG1655::PL-tacadhE*.
Штамм MG1655ΔargR был инфицирован фагом Pivir, выросшим на донорном штамме MG1655::PL-tacadhE*, и был получен штамм MG1655ΔargR,PL-tacadhE*. Этот штамм был проверен на способность расти на чашках со средой М9, содержащей 2% этанола в качестве единственного источника углерода. Скорость роста была такая же, как у MG1655::PL_tacadhE*(364).
Пример 13. Конструирование плазмиды pMW119-ArgA4.
Ген ArgA с единственной мутацией, обеспечивающей фенотип fbr (устойчивость к ингибированию по типу обратной связи) (JP2002253268, ЕР 1170361) под контролем собственного промотора был клонирован в векторе pMW119.
Ген argA был получен методом ПЦР с использованием плазмиды pKKArgA-r4 (JP2002253268, ЕР1170361) в качестве матрицы и праймеров Р30 (SEQ ID NO: 35) и Р31 (SEQ ID NO: 36). Последовательность праймера Р30 гомологична 5'-области гена гена argA, локализованной в 20 п.о. до и в 19 п.о. после инициирующего кодона гена argA е. Праймер Р31 содержит 24 н., гомологичный 3'-области гена argA сайт узнавания рестриктазой HindIII, введенный для дальнейшего клонирования в векторе pMW119/BamHI-HindIII.
Последовательность промотора PargA была получена методом ПЦР с использованием штамма E.coli MG1655 в качестве матрицы и праймеров Р32 (SEQ ID NO: 37) и Р33 (SEQ ID NO: 38). Праймер Р32 содержит 30 н., гомологичных 5'-нетранслируемой области гена argA, локализованной в 245 п.о. перед инициирующим кодоном, кроме этого, эта последовательность включает сайт узнавания рестриктазой ВаmHI. Праймер Р33 содержит 24 н., гомологичных 5'-области гена argA, локализованной в 20 п.о. перед инициирующим кодоном, и самому инициирующему кодону. Использовали следующие температурные условия: начальная денатурация ДНК в течение 5 мин при 95°С, следующие 25 циклов: денатурация при 95°С в течение 30 сек, отжиг при 54°С в течение 30 сек, элонгация при 72°С в течение 1 мин 20 сек (для гена ArgA) или 20 сек (для промотора PargA) и конечная элонгация в течение 5 мин при 72°С. Затем амплифицированные фрагменты ДНК были очищены методом электрофореза в агарозном геле, выделены с использованием "GenElute Spin Columns" ("Sigma", USA) и осаждены этанолом.
Фрагмент PargAArgA был получен методом ПЦР с использованием вышеописанных фрагментов ДНК: промотор PargA и ген ArgA. Во-первых, реакционная смесь (общий объем - 100 мкл) состояла из 10 мкл 10-кратного ПЦР буфера с 25 мМ MgCl2 (Sigma, USA), по 200 мкМ каждого dNTP и 1 Ед Accu-Taq ДНК-полимеразы (Sigma, USA). Фрагмент argA (25 нг) и PargA (5 нг) использовали как ДНК матрицу и как праймеры одновременно. Затем в реакционную смесь добавляли праймеры Р31 и Р32. Использовали следующие температурные условия: первая стадия - начальная денатурация ДНК в течение 5 мин при 95°С, следующие 10 циклов: денатурация при 95°С в течение 30 сек, отжиг при 53°С в течение 30 сек, элонгация при 72°С в течение 1 мин, вторая стадия - 15 циклов: денатурация при 95°С, отжиг при 54°С в течение 30 сек, элонгация при 72°С в течение 1 мин 30 сек. Амплифицированные фрагменты ДНК очищали методом электрофореза в агарозном геле, выделяли с использованием "GenElute Spin Columns" ("Sigma", USA), осаждали этанолом, обрабатывали рестриктазами BamHI и HindIII и легировали с предварительно обработанным этими же рестриктазами вектором pMW119. В результате была получена плазмида pMW119- rgA4.
Пример 14. Влияние усиления экспрессии гена adhE на продукцию L-аргинина.
Для оценки влияния усиления экспрессии мутантного гена adhE на продукцию L-аргинина, каждый из штаммов Е. coli MG1655ΔargR Pi-tacadhE* и MG1655ΔargR был трансформирован плазмидой pMW119-ArgA4. По 10 колоний трансформантов каждого вида культивировали при 37°С в течение 18 часов в питательном бульоне с добавлением 150 мг/л Ар, по 0.1 мл каждой из полученных культур инокулировали в 2 мл среды для ферментации, в пробирки 20×200 мм, и культивировали при 32°С в течение 96 часов на роторной качалке.
После культивирования количество накопленного в среде L-аргинина определяли методом бумажной хроматографии с использованием подвижной фазы следующего состава: бутанол: уксусная кислота: вода=4:1:1 (v/v). 2%-ный раствор нингидрина в ацетоне использовали в качестве проявляющего реагента. Пятно, содержащее L-аргинин, вырезали, L-аргинин элюировали 0.5% водным раствором CdCl2, а количество L-аргинина определяли спектрофотометрически при длине волны 540 nm.
Результаты десяти независимых пробирочных ферментаций представлены в Таблице 4. Как следует из Таблицы 4, MG1655ΔargR PL-tacadhE* продуцировал большее количество L-аргинина по сравнению с MG1655ΔargR PL-tacadhE*, как на среде с добавлением глюкозы, так и без добавления.
Состав среды для ферментации был следующим, г/л:
Этанол и сульфат магния стерилизовали раздельно. СаСО3 стерилизовали сухим жаром при 180°С в течение 2 часов.
Пример 15. Конструирование штамма- продуцента L-лейцина E. coli NS1391.
Штамм NS1391 был получен следующим образом.
Во-первых, был сконструирован штамм с инактивированными генами ацетолактатсинтазы (комбинация делеций ΔilvIH и ΔilvGM). Делеция генов ilvIH (ΔilvIH::cat) в диком штамме Е. coli K12 (VKPM В-7) была осуществлена с использованием Р1 трансдукции (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989). В качестве донорного штамма использовали E.coli MG1655 ΔilvIH::cat. Делеция оперона ilvIH в штамме MG1655 была проведена с использованием метода Red-зависимой интеграции. В соответствии с этой процедурой были сконструированы праймеры для ПЦР Р34 (SEQ ID NO: 39) и Р35 (SEQ ID NO: 40), гомологичные областям, прилегающим к оперону ilvIH, и гену, обеспечивающему устойчивость к хлорамфениколу, на плазмиде-матрице. В качестве матрицы для ПЦР использовали плазмиду pMW-attL-Cm-attR (заявка РСТ WO 05/010175). Использовали следующие температурные условия: стадия денатурации в течение 3 мин при 95°С; условия для первых двух циклов: 1 мин при 95°С, 30 сек при 34°С, 40 сек при 72°С; условия для последних 30 циклов: 30 сек при 95°С, 30 сек при 50°С, 40 сек при 72°С; финальная стадия: 5 мин при 72°С. Полученный продукт ПЦР размером 1713 п.о. очищали методом электрофореза в агарозном геле и использовали для электропорации клеток штамма Е. coli MG1655/pKD46. После электропорации были отобраны устойчивые к хлорамфениколу рекомбинанты, которые были протестированы методом ПЦР с использованием локус-специфических праймеров Р36 (SEQ ID NO: 41) и Р37 (SEQ ID NO: 42). Использовали следующие температурные условия: стадия денатурации в течение 3 мин при 94°С; условия для 30 циклов: 30 сек при 94°С, 30 сек при 53°С, 1 мин 20 сек при 72°С; конечная стадия: 7 мин при 72°С. Продукт ПЦР, полученный в реакции с хромосомной ДНК родительского штамма IlvIH+ MG1655 в качестве матрицы, был длиной 2491 н. Продукт ПЦР, полученный в реакции с хромосомной ДНК мутантного штамма MG1655 ΔilvIH::cat в качестве матрицы, был длиной 1823 н. В результате был получен штамм MG1655 ΔilvIH::cat. После делеции генов ilvIH (ΔilvIH::cat) в штамме E.coli К.12 (VKPM В-7) с использованием Р1 трансдукции были отобраны СmR трансдуктанты. В результате был получен штамм В-7 ΔilvIH::cat. Для удаления маркера устойчивости к хлорамфениколу из клеток штамма В-7 ΔilvIH::cat клетки трансформировали плазмидой pMW118-int-xis (ApR (WO2005/010175). АрR клоны выращивали на чашках с LB-агаром, содержащим 150 мг/л ампициллина, при 30°С. Несколько десятков АрR клонов проверили на чувствительность к хлорамфениколу. Плазмиду pMW118-int-xis удалили из CmS клеток путем культивирования клеток на чашках с LB-агаром при 42°С. В результате был получен штамм В-7 ΔilvIH.
Делению генов ilvGM (ΔilvGM::cat) из клеток Е.coli B-7 ΔilvIH осуществили методом Р1 трансдукции. В качестве донорного штамма использовали Е. coli MG1655 ΔilvGM::cat. Оперон ilvGM был делегирован из штамма MG1655 с использованием Red-зависимой интеграции. В соответстви с данной процедурой, были сконструированы праймеры для ПЦР Р38 (SEQ ID NO: 43) и Р39 (SEQ ID NO: 44), гомологичные областям, прилегающим к оперону ilvGM, и гену, обеспечивающему устойчивость к хлорамфениколу, на плазмиде-матрице. В качестве матрицы в ПЦР использовали плазмиду pMW-attL-Cm-attR (заявка РСТ WO 05/010175). Использовали следующие температурные условия: стадия денатурации в течение 3 мин при 95°С; условия для первых двух циклов: 1 мин при 95°С, 30 сек при 34°С, 40 сек при 72°С; условия для последних 30 циклов: 30 сек при 95°С, 30 сек при 50°С, 40 сек при 72°С; конечная стадия: 5 мин при 72°С.
Полученный продукт ПЦР размером 1713 п.о. очищали методом электрофореза в агарозном геле и использовали для электропорации клеток штамма Е. coli MG1655/pKD46. После электропорации отбирали устойчивые к хлорамфениколу рекомбинанты и проверяли их методом ПЦР с использованием локус-специфических праймеров Р40 (SEQ ID NO: 45) и Р41 (SEQ ID NO: 46). Для ПЦР-верификации использовали следующие температурные условия: стадия денатурации в течение 3 мин при 94°С; условия для 30 циклов: 30 сек при 94°С, 30 сек при 54°С, 1 мин 30 сек при 72°С; финальная стадия: 7 мин при 72°С. Продукт ПЦР, полученный в реакции с использованием в качестве матрицы хромосомной ДНК родительского штамма MG1655, был длиной 2209 н. Продукт ПЦР, полученный в реакции с использованием в качестве матрицы хромосомной ДНК мутантного штамма MG1655 ΔilvGM::cat, был длиной 1941 н.. В результате был получен штамм MG1655 ΔilvGM::cat.
После делеции генов ilvGM (ΔilvGM::cat) в клетках штамма Е.coli B-7 ΔilvIH путем Р1 трансдукции были отобраны СmR трансдуктанты. В результате был получен штамм B-7 ΔilvIH ΔilvBN ΔilvGM::cat. Маркер устойчивости к хлорамфениколу был удален из клеток штамма B-7 ΔilvIH ΔilvBN ΔilvGM::cat описанным выше способом. В результате был получен штамм B-7 ΔilvIH ΔilvGM.
Нативная регуляторная область оперона ilvBN была заменена на PL промотор фага λ с использованием Red-зависимой интеграции. С этой целью в качестве начального штамма для такой модификации использовали штамм В7 ΔilvIH ΔilvGM с единственной AHAS I. В соответствии с процедурой Red-зависимой интеграции были сконструированы праймеры для ПЦР P42(SEQ ID NO: 47) и Р43 (SEQ ID NO:48). Олигонуклеотид Р42 (SEQ ID NO: 47) гомологичен области перед геном ilvB и области, прилегающей к гену, обеспечивающему устойчивость к антибиотику на хромосомной ДНК штамма BW25113
cat-PL-yddG. Олигонуклеотид Р43 (SEQ ID NO: 48) гомологичен области ilvB и области после промотора PL на хромосоме штамма BW25113 cat-PL-yddG. Полученный штамм BW25113 cat-PL-yddG был детально описан ранее (европейская заявка ЕР1449918А1, патент РФ RU2222596). В качестве матрицы для ПЦР использовали хромосомную ДНК штамма BW25113 cat-PL-yddG. Использовали следующие температурные условия: денатурация в течение 3 мин при 95°С; условия для первых двух циклов: 1 мин при 95°С, 30 сек при 34°С, 40 сек при 72°С; условия для последних 30 циклов: 30 сек при 95°С, 30 сек при 50°С, 40 сек при 72°С; конечная стадия: 5 мин при 72°С. В результате получили продукт ПЦР (SEQ ID NO: 49), который очищали путем электрофореза в агарозном геле и использовали для электропорации клеток штамма Е. coli B-7 ΔilvIH ΔilvGM, содержащего плазмиду pKD46, репликация которой чувствительна к температуре. Электрокомпетентные клетки приготовили следующим образом: штамм Е. coli B-7 ΔilvIH ΔilvGM выращивали в течение ночи при 30°С в среде LB, содержащей ампициллин (100 мг/л), затем культуру разводили в 100 раз в 5 мл среде SOB (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) с ампициллином и L-арабинозой (1 мМ). Клетки выращивали с аэрацией при 30°С до достижения значения оптической плотности OD600 of≈0.6, затем концентрировали в 100 раз и трижды отмывали ледяной деионизированной водой. Электропорацию осуществляли с использованием 70 мкл клеток и ≈100 нг продукта ПЦР. После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2.5 ч, затем высевали на чашки с L-агаром и выращивали при 37°С для отбора СmR рекомбинантов. Затем для элиминации плазмиды pKD46 проводили 2 пассажа на L-агаре с CmR при 42°С и полученные колонии тестировали на чувствительность к ампициллину.
Полученный штамм В7 ΔilvIH ΔilvGM cat-PL-ilvBN был чувствительным к валину. Из этого штамма получили новые спонтанные мутанты по AHAS I. Были охарактеризованы штаммы, растущие при содержании в среде валина в концентрации 1 г/л.
Были получены мутации устойчивости к валину, которые обеспечивали также устойчивость к изолейцину. Были получены варианты со специфической активностью, большей, чем у клеток дикого типа. Были определены нуклеотидные последовательности мутантных оперонов для мутанта ilvBN4. Было обнаружено, что IlvBN4 содержал одну точечную мутацию в IlvN: N17K Asn-Lys (кодон аас был заменен на aag). Полученный штамм В7 ΔilvIH ΔilvGM cat-PL-ilvBN4 использовали для последующего конструирования.
Затем фрагмент cat-PL-ilvBN4 DNA переносили из клеток штамма Е.coli B7 ΔilvIH ΔilvGM cat-PL-ilvBN4 в клетки штамма Е.coli MG1655 mini-Mu::scrKYABR (европейская заявка ЕР 1149911) методом Р1 трансдукции. В результате был получен штамм ESP214. Маркер устойчивости к хлорамфениколу был удален из штамма ESP214 описанным выше способом. В результате был получен штамм ESP215.
Затем фрагмент ДНК, показанный в (SEQ ID NO: 50), использовали для электропорации штамма ESP215/pKD46 с целью последующей интеграции в хромосому. Этот фрагмент ДНК содержал области, комплементарные 3' области гена b1701 и 5' области гена b1703 (данные гены прилегают к гену pps), которые необходимы для интеграции в хромосому. Фрагмент ДНК также содержал вырезаемый маркер устойчивости к хлорамфениколу cat и мутантный оперон ilvBN4 под контролем конститутивного промотора PL. Электропорацию осуществляли, как описано выше. Отобранные рекомбинанты CmR содержали делецию гена pps как результат интеграции фрагмента cat-PL-ilvBN4 в хромосому. Таким образом был получен штамм ESP216, маркер устойчивости к хлорамфениколу был удален из штамма ESP216 описанным выше способом. В результате был получен штамм ESP217.
На следующем этапе мутантный ген leuA (Gly479 -> Cys) под контролем конститутивного промотора PL вводили в штамм ESP217. Фрагмент ДНК, представленный в (SEQ ID NO: 51), использовали для электропорации штамма ESP217/pKD46 с целью последующей интеграции в хромосому. Этот фрагмент ДНК содержал область из 35 н., необходимую для интеграции в хромосому и гомологичную области перед геном leuA. Фрагмент ДНК также содержал вырезаемую область, комплементарную последовательности маркера устойчивости к хлорамфениколу cat, и мутантный ген leuA (Gly479 -> Cys) под контролем конститутивного промотора PL. Электропорацию осуществляли, как описано выше. Отобранные рекомбинанты CmR содержали мутантный ген leuA (Gly479 -> Cys) под контролем конститутивного промотора PL, интегрированный в хромосому. Таким образом был получен штамм ESP220. Маркер устойчивости к хлорамфениколу был удален из штамма ESP220 описанным выше способом. В результате был получен штамм ESP221.
Затем фрагмент ДНК, представленный в SEQ ID NO: 52, использовали для электропорации штамма ESP221/pKD46 с целью последующей интеграции в хромосому. Этот фрагмент ДНК содержал область из 35н., гомологичную области перед геном tyrB, которая необходима для интеграции в хромосому. Фрагмент также содержал вырезаемую область, комплементарную последовательности маркера устойчивости к хлорамфениколу cat, и ген tyrB с модифицированной регуляторной (-35) областью. Электропорацию осуществляли, как описано выше. Отобранные CmR рекомбинанты содержали ген tyrB с модифицированной регуляторной (-35) областью. Таким образом был получен штамм NS1390, маркер устойчивости к хлорамфениколу удаляли из штамма NS1390 описанным выше способом. В результате был получен штамм NS1391. Штамм-продуцент лейцина NS1391 использовали в дальнейшей работе.
Пример 16. Влияние усиления экспрессии мутантного гена adhE на продукцию L-лейцина.
Для оценки влияния экспрессии гена adhE под контролем промотора РL-tac на продукцию L-лейцина фрагменты ДНК хромосомы описанного выше штамма MG1655 PL-tacadhE* переносили в штамм-продуцент L-лейцина Е. coli NS1391 методом Р1 трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY) чтобы получить штамм NS1391 PL-tacadhE*.
Оба штамма Е. coli, NS1391 и NS1391 PL-tacadhE*, культивировали в течение 18-24 ч при 37°С на чашках с L-агаром. Для получения посевной культуры штаммы выращивали на роторной качалке (250 об/мин) при 32°С в течение 18 ч в пробирках 20×200-мм, содержащих 2 мл L-бульона с добавлением 4% сахарозы. Затем 0.21 мл посевной культуры (10%) инокулировали в среду для ферментации. Ферментацию проводили в 2 мл минимальной среды для ферментации в пробирках 20×200-мм. Клетки выращивали в течение 48-72 ч при 32°С со встряхиванием при 250 об/мин. Количество L-лейцина измеряли методом бумажной хроматографии (состав жидкой фазы: бутанол-уксусная кислота-вода=4:1:1). Результаты десяти независимых пробирочных ферментаций представлены в Таблице 5. Как следует из Таблицы 5, NS1391 PL-tacadhE* продуцировал большее количество L-лейцина по сравнению с NS1391 в средах, содержащих различные количества этанола.
Использовали среду для ферментации следующего состава, г/л (рН 7.2):
Глюкозу, этанол и СаСО3 стерилизовали раздельно.
Пример 17. Влияние усиления экспрессии гена adhE на продукцию L-фенилаланина.
Для оценки влияния усиления экспрессии гена adhE под контролем промотора PL-tac на продукцию L-фенилаланина, фрагменты ДНК из хромосомы вышеописанных штаммов MG1655Δtdh, rhtA*, PL-tacadhE; MG1655Δtdh, rhtA*, PL-tacadhE*; MG1655Δtdh, rhtA*, PL-tacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PL-tacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*, adhE* могут быть перенесены в штамм-продуцент L-фенилаланина E.coli AJ12739 с помощью Р1 трансдукции (Miller, J.H. Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, 1972, Plainview, NY). Штамм AJ12739 был депонирован во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-ый Дорожный проезд, 1) 6 ноября 2001 года с регистрационными номерами VK.PM В-8197 и затем было произведено международное депонирование этого штамма согласно условиям Будапештского Договора 23 августа 2002 года.
Каждый из полученных штаммов и родительский штамм AJ12739 могут быть выращены при 37°С в течение 18 часов в питательном бульоне и по 0,3 мл каждой из полученных культур могут быть инокулированы в 3 мл среды для ферментации в пробирки размером 20×200 мм и культивированы при 37°С в течение 48 часов со встряхиванием на роторной качалке. После культивирования количество накопленного в среде фенилаланина может быть определено методом тонкослойной хроматографии (ТСХ). Могут быть использованы пластинки для ТСХ размером 10×15-см, покрытые слоями стационарной фазы из силиконового геля Sorbfil толщиной 0.11 мм, не содержащего флуоресцентный краситель (Stock Company Sorbpolymer, Краснодар, Россия). Разделение на пластинках Sorbfil может проводиться в подвижной фазе следующего состава: пропан-2-ол: этилацетат: 25% жидкий аммоний: вода=40:40:7:16 (v/v). В качестве проявляющего реагента может быть использован 2% раствор нингидрина в ацетоне. Может быть использована среда для ферментации следующего состава, г/л:
Этанол и сульфат магния стерилизуются раздельно. СаСО3 стерилизуется сухим жаром при 180°С в течение 2 часов. рН доводится до 7.0.
Пример 18. Влияние усиления экспрессии гена adhE на продукцию L-триптофана.
Для оценки влияния усиления экспрессии гена adhE под контролем промотора РL-tac на продукцию L-триптофана фрагменты ДНК из хромосомы вышеописанных штаммов MG1655Δtdh, rhtA*. PL-tacadhE; MG1655Δtdh, rhtA*, PL-tacadhE*; MG1655Δtdh, rhtA*. PL-tacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PL-tacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*, adhE* могут быть перенесены в штамм-продуцент L-триптофана Е. coli SV164 (pGH5) с помощью Р1 трансдукции (Miller, J.H. Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, 1972, Plainview, NY). Штамм SV164 имеет аллель гена trpE, который кодирует антранилатсинтазу со снятым ингибированием триптофаном по принципу обратной связи. Плазмида pGH5 является носителем мутантного гена serA, кодирующего фосфоглицератдегидрогеназу со снятым ингибированием серином по принципу обратной связи. Штамм SV164 (pGH5) был подробно описан в патенте США 6,180,373 и в Европейском патенте 0662143.
Каждый из полученных штаммов и родительский штамм SV164(pGH5) могут быть выращены при температуре 37°С в течение 18 часов с перемешиванием в 3 мл питательного бульона с добавлением тетрациклина (20 мг/мл, маркер плазмиды pGH5).
Полученные культуры (каждая в объеме 0.3 мл) может быть инокулирована в 3 мл среды для ферментации, содержащей тетрациклин (20 мг/мл) в пробирки размером 20×200-мм и выращиваться при 37°С в течение 48 часов на роторной качалке (250 об/мин). После культивирования количество накопленного в среде триптофана может быть установлено методом ТСХ, как описано в Примере 17.
Компоненты среды для ферментации перечислены в Таблице 6, и должны стерилизоваться отдельными группами (А, В, С, D, Е, F и С), как показано, для предотвращения взаимодействий в процессе стерилизации.
Пример 19. Влияние усиления экспрессии гена adhE на продукцию L-гистидина.
Для проверки влияния усиления экспрессии гена adhE под контролем промотора
PL-tac на продукцию гистидина фрагменты ДНК из хромосомы описанных выше штаммов Е. coli MG1655Δtdh, rhtA*, PL-tacadhE; MG1655Δtdh, rhtA*, PL-tacadhE*; MG1655Δtdh, rhtA*, PL-tacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PL_tacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*, adhE* могут быть перенесены в штамм-продуцент гистидина Е. coli 80 с помощью Р1 трансдукции (Miller, J.H. Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, 1972, Plainview, NY). Штамм Е. Coli 80 описан в патенте РФ 2119536 и депонирован во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-ый Дорожный проезд, 1) 15 октября 1999 года с регистрационным номером VKPM В-7270 и затем было произведено международное депонирование этого штамма согласно условиям Будапештского Договора, 12 июля 2004 года.
Полученные штаммы Е. coli и родительский штамм Е. coli 80 могут быть выращены на L-бульоне при 29°С в течение 6 часов. Затем 0.1 мл полученной культуры может быть инокулировано в 2 мл среды для ферментации в пробирки размером 20×200 мм и выращиваться при 29°С в течение 65 часов с перемешиванием на роторной качалке (350 об/мин). После выращивания количество гистидина, которое накопилось в среде, может быть определено с помощью бумажной хроматографии. Подвижная фаза может быть следующего состава: п-бутанол:уксусная кислота:вода=4:1:1 (v/v). Раствор нингидрина (0.5%) в ацетоне может быть использован в качестве проявляющего реагента.
Состав среды для ферментации (рН 6.0) может быть следующим, г/л:
Этанол, пролин, бетаин и СаСО3 стерилизуются раздельно. Значение рН доводится до 6.0 перед стерилизацией.
Пример 20. Влияние усиления экспрессии гена adhE на продукцию L-глутаминовой кислоты.
Для оценки влияния усиления экспрессии гена adhE под контролем промотора РL-tac на продукцию L-глутамина фрагменты ДНК из хромосомы вышеописанных штаммов MG1655Δtdh, rhtA*, PL-tacadhE; MG1655Δtdh, rhtA*, PL.iacadhE*; MG1655Δtdh,rhtA*, pl-tacadhE-Lys568 (кл.18); MG1655Δtdh, rhtA*, PL-tacadhE-Lys568, Val566 (кл.1); MG1655Δtdh, rhtA*, adhE* могут быть перенесены в штамм-продуцент L-глутаминовой кислоты Е. coli VL334thrC+(Европейский патент ЕР1172433) с помощью Р1 трансдукции (Miller, J.H. Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, 1972, Plainview, NY). Штамм VL334thrC"1" депонирован во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-ый Дорожный проезд, 1) 6 декабря 2004 года с регистрационными номерами VKPM В-8961, и затем было произведено международное депонирование этого штамма, согласно условиям Будапештского Договора, 8 декабря 2004 года.
Каждый из полученных штаммов и родительский штамм VL334thrC+ могут быть выращены в питательном бульоне при температуре 37°С в течение 18 часов с перемешиванием. Полученные культуры (каждая в объеме 0.3 мл) может быть инокулирована в 3 мл среды для ферментации в пробирки размером 20×200-мм и выращена при 37°С в течение 48 часов на роторной качалке (250 об/мин). Среда для ферментации может быть следующего состава (рН 7.2), г/л:
Этанол и СаСО3 стерилизуются раздельно.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения. Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.
Ферментация в пробирках проводилась без восполнения испарившегося этанола.
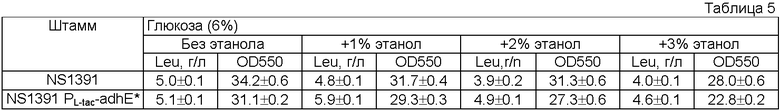
Раствор А имеет рН 7.1, доводится с помощью аммиака.























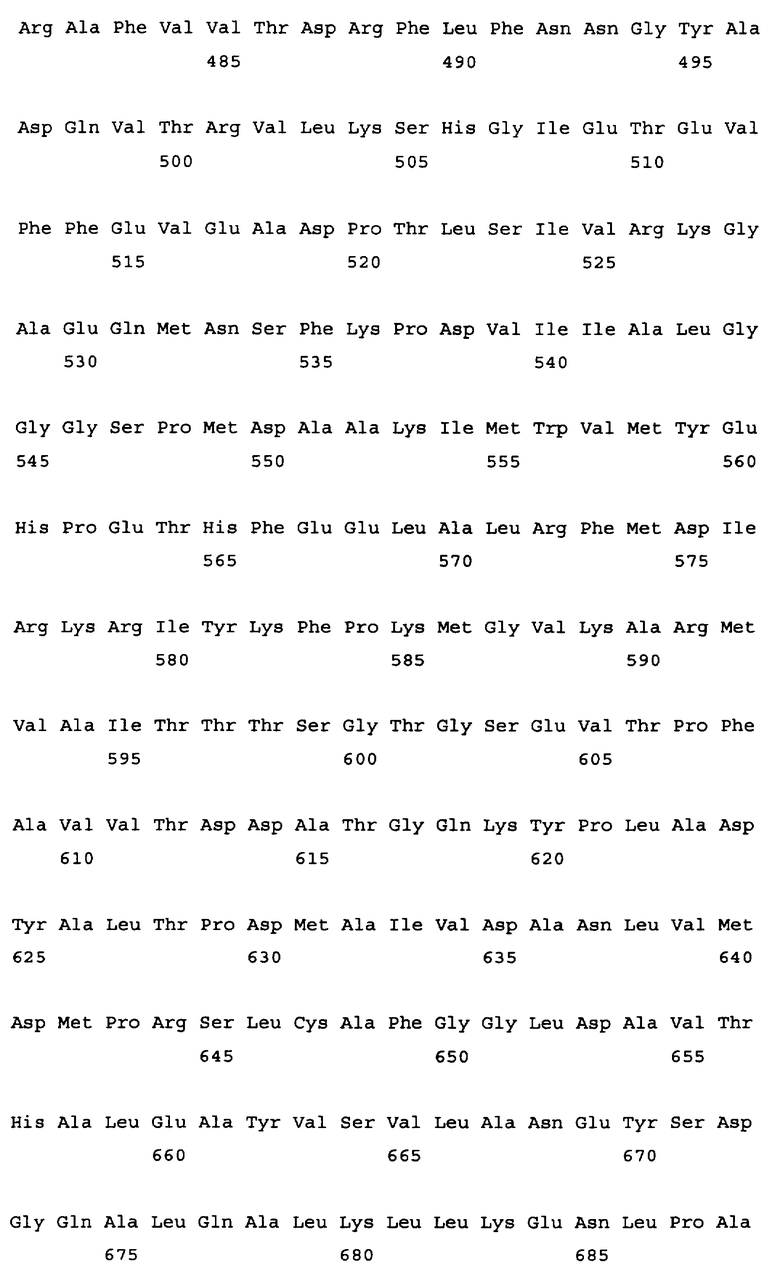













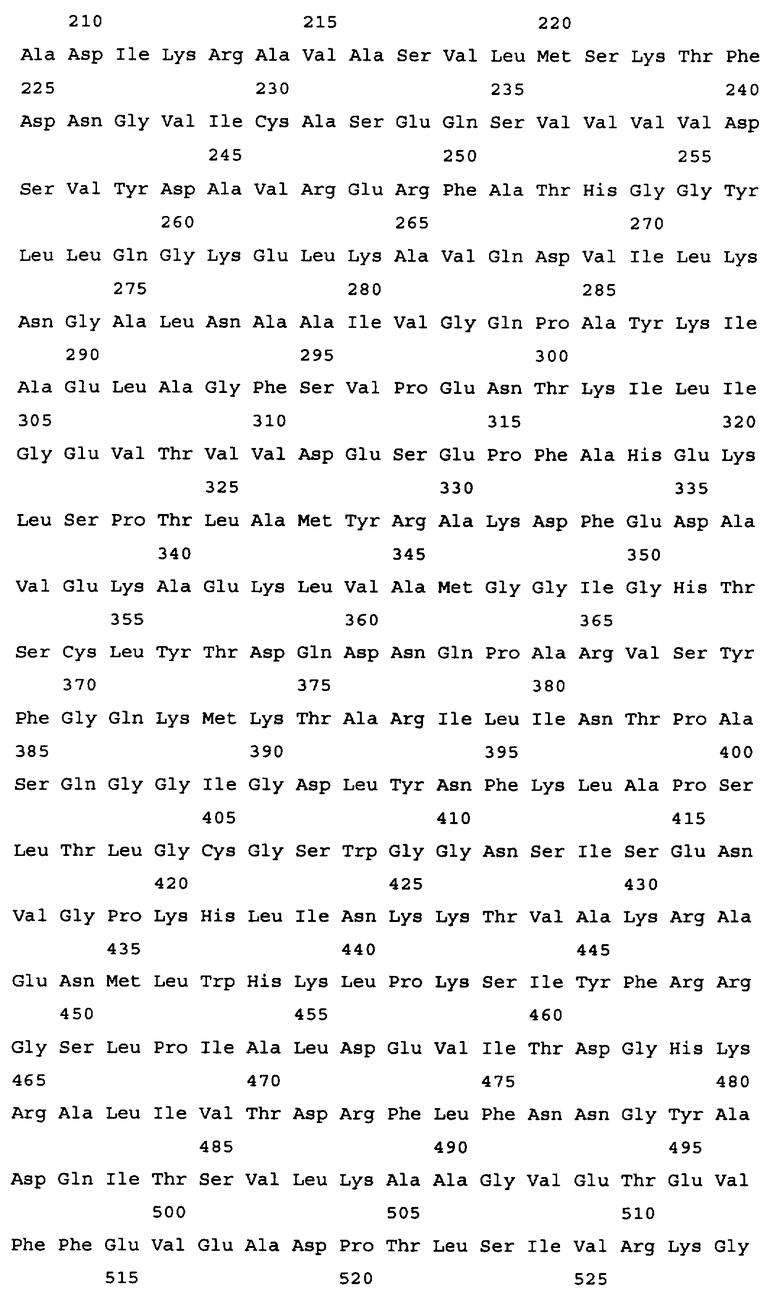


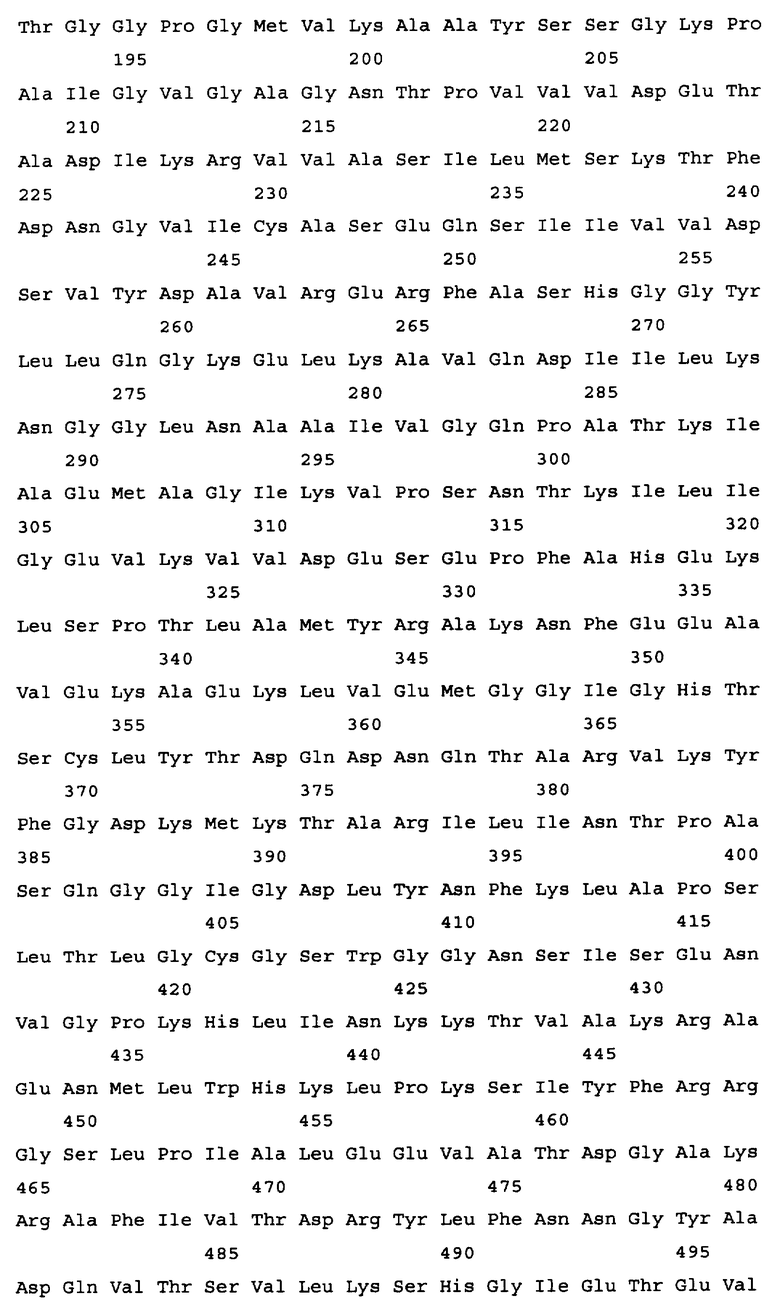


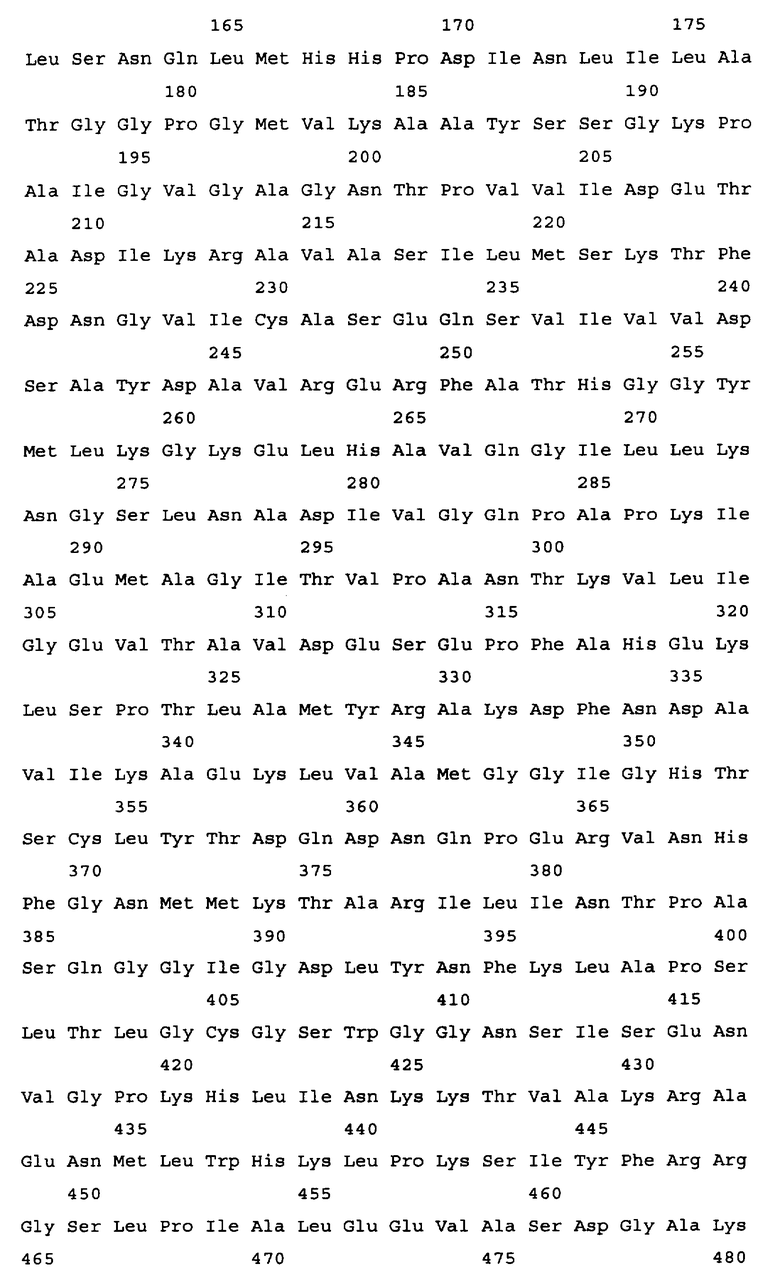



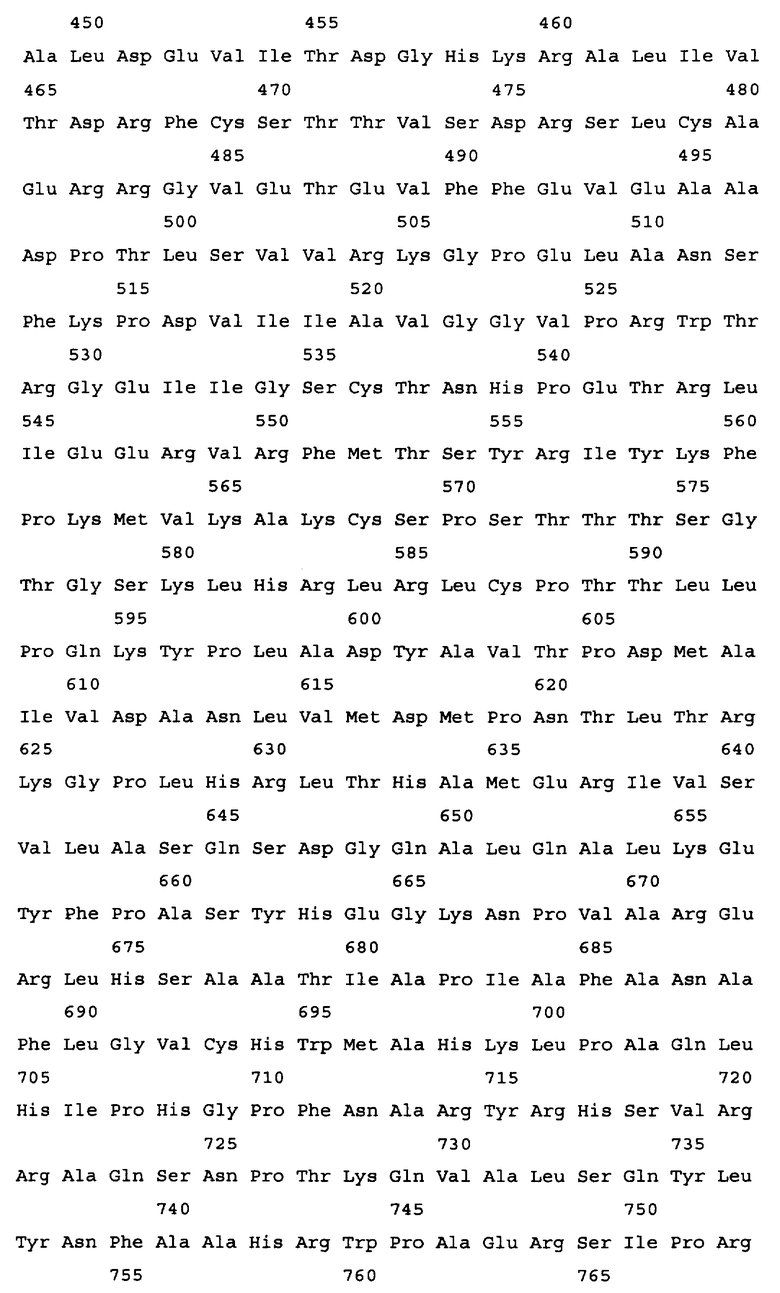







| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae, ОБЛАДАЮЩЕЙ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНОВ КАСКАДА ОБРАЗОВАНИЯ ФЛАГЕЛЛ И КЛЕТОЧНОЙ ПОДВИЖНОСТИ | 2012 |
|
RU2508404C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yahN | 2007 |
|
RU2392322C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2460793C2 |
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА И L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН cpxR | 2005 |
|
RU2320723C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН hipA | 2005 |
|
RU2320718C2 |
Изобретение относится к биотехнологии и представляет собой способ получения L-аминокислоты с использованием бактерии рода Escherichia, причем бактерия модифицирована таким образом, что активность алкогольдегидрогеназы, кодируемой геном adhE, у этой бактерии увеличена. Изобретение позволяет получать L-аминокислоты с высокой степенью эффективности. 4 н. и 15 з.п. ф-лы, 5 ил., 6 табл.
1. Мутантная алкогольдегидрогеназа, устойчивая к аэробной инактивации, выбранная из группы, состоящей из:
А) белка, включающего аминокислотную последовательность, представленную в SEQ ID NO:2, за тем исключением, что указанный белок содержит мутацию в положении 568 в SEQ ID NO:2, и по крайней мере одну дополнительную мутацию, способствующую улучшению роста бактерии в жидкой среде, содержащей этанол в качестве единственного источника углерода, отличающаяся тем, что указанная дополнительная мутация выбрана из группы, состоящей из:
а) замены остатка глутаминовой кислоты в положении 560 в SEQ ID NO:2 на остаток другой аминокислоты;
б) замены остатка фенилаланина в положении 566 в SEQ ID NO:2 на остаток другой аминокислоты;
в) замены остатка глутаминовой кислоты, остатка метионина, остатка тирозина, остатка изолейцина и остатка аланина в положениях 22, 236, 461, 554 и 786 соответственно, в SEQ ID NO:2 на остатки других аминокислот; и
г) комбинации этих замен; и
Б) белка, содержащего мутации, указанные в подпункте (А), а также дополнительные мутации в положениях, отличных от указанных в подпункте (А), и при этом обладающего активностью алкогольдегидрогеназы.
2. Фрагмент ДНК, кодирующий указанную мутантную алкогольдегидрогеназу по п.1.
3. Бактерия, принадлежащая к роду Escherichia, продуцент L-аминокислоты, содержащая мутантную алкогольдегидрогеназу по п.1.
4. Бактерия по п.3, отличающаяся тем, что указанная бактерия модифицирована таким образом, что активность мутантной алкогольдегидрогеназы по п.1 в этой бактерии увеличена.
5. Бактерия по п.4, отличающаяся тем, что активность мутантной алкогольдегидрогеназы увеличена путем усиления экспрессии гена, кодирующего указанную мутантную алкогольдегидрогеназу.
6. Бактерия по п.4, отличающаяся тем, что активность указанной мутантной алкогольдегидрогеназы увеличена путем модификации последовательности, контролирующей экспрессию гена, кодирующего указанную алкогольдегидрогеназу, или путем увеличения количества копий гена, кодирующего указанную алкогольдегидрогеназу.
7. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-треонина.
8. Бактерия по п.7, отличающаяся тем, что указанная бактерия дополнительно модифицирована таким образом, что у этой бактерии усилена экспрессия гена, выбранного из группы, состоящей из:
мутантного гена thrA, кодирующего аспартокиназу-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи;
гена thrB, кодирующего гомосеринкиназу;
гена thrC, кодирующего треонинсинтазу;
гена rhtA, кодирующего предполагаемый трансмембранный белок;
гена asd, кодирующего аспартат-β-семиальдегиддегидрогеназу;
гена aspC, кодирующего аспартатаминотрансферазу (аспартаттрансаминазу); и
- комбинации этих генов.
9. Бактерия по п.8, отличающаяся тем, что указанная бактерия модифицирована таким образом, что экспрессия указанного мутантного гена thrA, указанного гена thrB, указанного гена thrC и указанного гена rhtA у этой бактерии усилена.
10. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-лизина.
11. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-гистидина.
12. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-фенилаланина.
13. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-аргинина.
14. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-триптофана.
15. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-глутаминовой кислоты.
16. Бактерия по п.3, отличающаяся тем, что указанная бактерия является продуцентом L-лейцина.
17. Способ получения L-аминокислоты, включающий:
А) выращивание бактерии - продуцента L-аминокислоты по п.3, содержащая мутантную алкогольдегидрогеназу по п.1, устойчивую к аэробной инактивации, в питательной среде, и
Б) выделение L-аминокислоты из культуральной жидкости, отличающийся тем, что ген, кодирующий указанную алкогольдегидрогеназу, экспрессируется под контролем ненативного промотора, который функционирует в аэробных условиях культивирования.
18. Способ по п.17, отличающийся тем, что указанный ненативный промотор выбран из группы, состоящей из Ptac, Plac, Ptrp, Ptrc, PR и PL.
19. Способ по п.17, отличающийся тем, что указанная L-аминокислота выбрана из группы, состоящей из L-треонина, L-лизина, L-гистидина, L-фенилаланина, L-аргинина, L-триптофана, L-глутаминовой кислоты и L-лейцина.
| KR 20030062847, 28.07.2003 | |||
| CHEN Y.M | |||
| Regulation of the adhE gene, which encodes ethanol dehydrogenase in Escherichia coli, J.Bacteriol 1991 Dec; 173(24): 8009-13 | |||
| KOO O.K | |||
| et | |||
| al | |||
| Cloning and characterization of the bifunctional alcohol/acetaldehyde dehydrogenase gene (adhE) in Leuconostoc mesenteroides isolated from kimchi | |||
| Biotechnol Lett | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |

