Известный уровень техники
В данной заявке заявлен приоритет предварительной заявки США с серийным номером 60/249710, зарегистрированной 17 ноября 2000 г., и предварительной заявки США с серийным номером 60/322859, зарегистрированной 17 сентября 2001 г., обе из которых специально включены здесь в качестве ссылки во всей своей полноте.
1. Область техники, к которой относится изобретение
Настоящее изобретение относится в целом к области медицины. Более конкретно, изобретение относится к способам ингибирования воспаления с применением композиций монотерпенов, которые тормозят NF-κB.
2. Описание области техники, к которой относится изобретение
Растения и животные, особенно морские животные, представляют собой ценные источники для идентификации новых биологически активных молекул. Одним из разнообразных классов молекул, которые были идентифицированы в растениях, является класс сапонинов. Сапонины представляют собой высокомолекулярные соединения, включающие в себя гликозиды с частью сахаров, присоединенной к тритерпену или стероидному агликону. Тритерпеновые сапонины представляют собой объект особенно большого интереса благодаря своим биологическим свойствам.
Исследовались фармакологические и биологические свойства тритерпеновых сапонинов из различных видов растений, включая фунгицидную, противовирусную, антимитогенную, спермицидную или контрацептивную, сердечно-сосудистую и противовоспалительную активности (Hostettmann et al., 1995). Сапонины, как известно, образуют комплексы с холестерином, связывая липиды плазмы, изменяя, таким образом, метаболизм холестерина (Oakenfull et al., 1983). Тритерпеновые гликозиды, добавленные в пищу также, как показано, снижают количество холестерина в крови и тканях экспериментальных животных (Cheeke, 1971). Обнаружено, что сапонины являются составляющими многих народных медицинских средств и некоторых сравнительно недавно разработанных растительных лекарств.
Тритерпенглицирретиновая кислота и ее определенные производные, как известно, имеют противоязвенное, противовоспалительное, антиаллергическое, антигепатитное и противовирусное действие. Например, определенные производные глицирретиновой кислоты могут предотвращать или заживлять язвы желудка (Doll et al., 1962).
К таким соединениям, известным в данной области техники, относятся карбеноксолон (патент США No. 3070623), эфирные производные глицирретиновой кислоты, имеющие заместители в 3'-положении (патент США No. 3070624), аминокислотные соли глицирретиновой кислоты (патентная публикация Японии JP-A-44-32798), амидные производные глицирретиновой кислоты (патент Бельгии No. 753773) и амидные производные 11-дезоксоглицирретиновой кислоты (патент Великобритании No. 134871). Показано, что глицирретиновая кислота ингибирует ферменты, вовлеченные в биосинтез лейкотриенов, включая активность 5-липоксигеназы, и, как предполагается, в результате этого ответственны за описанную противовоспалительную активность (Inoue et al., 1986).
Бетулиновая кислота, пентациклический тритерпен, как описано, является селективным ингибитором роста меланомной опухоли человека в моделях ксенотрансплантатов бестимусным голым мышам, и было показано, что она индуцирует цитотоксичность путем индукции апоптоза (Pisha et al., 1995). Тритерпеновый сапонин из китайского лекарственного растения семейства Cucurbitaceae проявлял противоопухолевую активность (Kong et al., 1993). Моногликозиды тритерпенов, как показано, проявляют сильную и избирательную цитотоксичность в отношении лейкозных клеток MOLT-4 человека (Kasiwada et al., 1992), а определенные тритерпеновые гликозиды семейства Iridaceae тормозили рост опухолей и увеличивали продолжительность жизни мышей с имплантированной асцитной карциномой Эрлиха (Nagamoto et al., 1988). Препарат сапонинов из растения Dolichos falcatus, которое принадлежит семейству Leguminosae, как описано, эффективен против клеток саркомы-37 in vitro и in vivo (Huang et al., 1982). Сапонин сои также из семейства Leguminosae, как показано, эффективен против ряда опухолей (Tomas-Barbaren et al., 1988). Гликозиды олеаноловой кислоты и гипсогенина, проявляющие гемолитическую и моллюскицидную активности, были выделены из земляных фруктовых стручков Swartzia madagascariensis (Leguminosae) (Borel and Hostettmann, 1987).
Генистеин, природно существующий изофлавоноид, выделенный из соевых продуктов, представляет собой ингибитор тирозинкиназы, который, как было показано, ингибирует пролиферацию эстроген-позитивных и эстроген-негативных линий клеток рака молочной железы (Akiyama et al., 1987). Инозитолгексафосфат (фитиновая кислота), который имеется в изобилии в царстве растений и который является природным диетическим ингредиентом зерновых и бобов, как показано, вызывает терминальную дифференцировку клеточной линии карциномы ободочной кишки. Фитиновая кислота проявляет также противоопухолевую активность в отношении экспериментального канцерогенеза ободочной кишки и молочной железы in vivo (Yang et al., 1995). Некоторые тритерпеновые агликоны также, как было продемонстрировано, имеют цитотоксические или цитостатические свойства, т.е. кора ствола растения Crossopteryx febrifuga (Rubiaceae), как показано, является цитостатической в отношении линии клеток Co-115 карциномы ободочной кишки человека в диапазоне нг/мл (Tomas-Barbaren et al., 1988).
В то время как в предшествующих сообщениях идентифицировались тритерпеновые соединения, которые имели любое из множества применений, на данном уровне техники все еще сохраняется большая потребность в идентификации новых биологически активных тритерпеновых соединений. Многие из этих соединений являются токсичными для нормальных клеток млекопитающих. Более того, биологическая активность идентифицированных ранее тритерпенов варьируется в широких пределах, и многие обладают ограниченной или варьирующей степенями эффективности при лечении любого такого состояния человека или млекопитающих. Огромное разнообразие различных тритерпенов, которые были идентифицированы, и огромный диапазон различий и непредсказуемости их биологической активности, наблюдаемой даже среди близкородственных тритерпеновых соединений, подчеркивает сложности, с которыми сталкиваются при получении тритерпенов, которые являются потенциальными терапевтическими агентами. Достижение трудной задачи идентификации новых тритерпенов с выгодными вариантами биологической активности может создать полностью новые пути лечения разнообразного набора болезней человека, для которых терапевтические возможности в настоящее время ограничены.
NF-κB представляет собой повсеместный транскрипционный фактор и регулирует транскрипцию ряда генов, вовлеченных в иммунные и воспалительные пути, таких как различные провоспалительные цитокины, молекулы адгезии и апоптоз, и таким образом представляет собой один из центральных регуляторов ответов организма на различные стрессогенные сигналы. Нарушение регуляции NF-κB вносит вклад в разнообразные патологические состояния, такие как септический шок, острое воспаление, репликация вирусов и некоторые виды рака.
Наиболее преобладающими и активными формами NF-κB являются димерные комплексы p50/relA (p50/p65). В нестимулированных клетках данные факторы удерживаются в цитоплазме в комплексе с ингибиторными белками (IκB), которые маскируют их сигнал ядерной транслокации. В ответ на внеклеточный сигнал, такой как воспалительные цитокины, митогены, бактериальные продукты или окислительный стресс, IκB подвергается фосфорилированию по специфическим сериновым остаткам, которые затем служат сигналом для присоединения к ним убиквитина и деградации по протеосомному пути. Деградация IκB позволяет свободному от ингибитора комплексу NF-κB транслоцироваться в ядро, связываться с ДНК и активировать транскрипцию специфических генов.
Из их роли в воспалении и канцерогенезе, а также в других иммунологических нарушениях, должно следовать, что модуляторы, негативно регулирующие NF-κB, должны иметь огромное терапевтическое применение. Более того, некоторые последующие эффекты, обусловленные ингибированием активности NF-κB, представляют собой снижение уровней экспрессии индуцибельной синтазы оксида азота (iNOS) и циклооксигеназы-2 (COX-2). Как iNOS, так и COX-2 играют определяющую роль в ответе тканей на воспаление, повреждение и канцерогенез. Таким образом, на данном уровне техники существует потребность в регуляторах NF-κB, а также iNOS и COX-2, так как эти соединения должны обеспечить противовоспалительные и хемопротекторные эффекты.
Краткое изложение существа изобретения
Настоящее изобретение преодолевает существующие недостатки на данном уровне техники и предлагает способы ингибирования воспаления с применением монотерпеновых композиций. В некоторых осуществлениях данные монотерпеновые композиции могут дополнительно включать в себя сахара и могут даже дополнительно включать в себя в виде части носитель, который может вносить монотерпеновые композиции в клетку, придавая мембране растворимость или проницаемость, или может придавать желаемые свойства композиции. Монотерпеновые композиции могут дополнительно включать в себя дополнительные химические заменители, такие как, но, конечно, не ограничиваясь этим, тритерпеновые гликозиды и/или другие монотерпены, и/или сахара.
Монотерпеновые композиции согласно изобретению могут быть получены почти из любого источника. Например, растения и морские животные являются богатым источником таких соединений. В некоторых осуществлениях монотерпеновые композиции могут быть выделены из стручков и корней Acacia victoriae (Benth.) (Leguminosae). В еще одних осуществлениях композиции могут быть даже синтезированы химически или ферментативно. Таким образом, могут быть применены химические способы синтеза, известные специалистам в данной области техники. В противоположном варианте могут быть использованы биохимические способы с применением ферментов, которые вовлечены в синтез монотерпеновых композиций. Ферменты, применяемые в данных путях, могут быть выделены из организма, например, растений, морских животных и т.д. или могут быть созданы генно-инженерным способом.
В изобретении предлагаются способы ингибирования воспаления, включающие в себя введение в клетку монотерпеновой композиции, которая ингибирует NF-κB. В некоторых осуществлениях NF-κB индуцируется TNF. В предпочтительных осуществлениях способа монотерпеновая композиция дополнительно включает в себя в виде части носитель. Часть носителя определяется здесь как часть, которая придает мембране растворимость, придает мембране проницаемость или обеспечивает внутриклеточную доступность для монотерпеновой композиции. Специалист в данной области техники должен знать, что может быть использована любая молекула, которая обеспечивает внутриклеточную доступность или клеточную проницаемость, и некоторые неограничивающие примеры части носителя включают в себя липид, липофильный белок, который может пересекать/переходить мембрану, тритерпеновый гликозид, тритерпеновый гликозид, дополнительно присоединенный к другим молекулам, таким как сахара, и/или другие монотерпеновые единицы.
В изобретении также предлагаются способы ингибирования воспаления, включающие в себя введение в клетку монотерпеновой композиции, которая ингибирует NF-κB, где монотерпен дополнительно присоединен к тритерпеновой части и/или к сахару, и/или ко второй монотерпеновой части. Специалист в данной области техники должен понимать, что описанные здесь композиции могут быть дополнительно замещены другими химическими функциональными частями.
Таким образом, монотерпен может дополнительно включать в себя тритерпеновую часть, присоединенную по меньшей мере к одной, предпочтительно - к двум, трем или более дополнительным монотерпеновым частям. Когда присутствует более одной монотерпеновой части, каждая их этих частей может быть присоединена (i) прямо к тритерпеновой части, (ii) к сахару или другой линкерной группе, которая присоединена к тритерпеновой части, или (iii) к монотерпеновой части, которая присоединена к тритерпеновой части напрямую или через сахар или другие линкерные группы. Линкерные группы включают в себя сахара, ацил, амид, алкокси, кетил, алкил, алкилен и другие сходные химические части, которые должны быть очевидны специалисту в данной области техники.
Тритерпеновая часть способа может включать в себя формулу:

или может быть ее изомером, где а) R1 и R2 выбраны из группы, состоящей из водорода, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара, олигосахарида; b) где каждый из R3-R36 отдельно и независимо выбран из группы, состоящей из места ненасыщенной валентности, водорода, гидроксила, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара, С1-С5-алкилового эфира и монотерпеновой группы, и с) по меньшей мере один из R3-R36 представляет собой монотерпеновую группу. Изомер может представлять собой оптический изомер, стереоизомер или цис-изомер, или транс-изомер.
В некоторых осуществлениях способа каждый из R1 и R2 включает в себя олигосахарид. В некоторых конкретных аспектах данного осуществления каждый из R1 и R2 включает в себя моносахарид, дисахарид, трисахарид или тетрасахарид. В других конкретных аспектах способа каждый из R1 и R2 включает в себя олигосахарид, включающий в себя сахара, которые отдельно и независимо выбраны из группы, состоящей из глюкозы, фукозы, рамнозы, арабинозы, ксилозы, хиновозы, мальтозы, глюкуроновой кислоты, рибозы, N-ацетилглюкозамина и галактозы. Еще в одном конкретном аспекте способа по меньшей мере один сахар метилирован.
В других осуществлениях способа R4 присоединен к тритерпеновой части через один из углеродов метилена, присоединенного к тритерпеновой части. В другом аспекте тритерпеновая часть дополнительно включает в себя по меньшей мере одну двойную связь.
В других осуществлениях способа тритерпеновая часть представляет собой сложный эфир акациевой кислоты, сложный эфир олеаноловой кислоты, сложный эфир бетулиновой кислоты, сложный эфир урзоловой кислоты, сложный эфир хиновиновой кислоты, сложный эфир помоловой кислоты, сложный эфир ротундовой кислоты, сложный эфир ротунгеновой кислоты, сложный эфир мадасиатовой кислоты, сложный эфир азиатовой кислоты, сложный эфир эускафиновой кислоты, сложный эфир торментиновой кислоты, сложный эфир мадекассиевой кислоты, сложный эфир лупеоловой кислоты, сложный эфир циликодисковой кислоты, сложный эфир молликовой кислоты, сложный эфир джессиковой кислоты, сложный эфир эхиноцистовой кислоты или сложный эфир энтагеновой кислоты или другую структурно сходную тритерпеноидную часть.
Монотерпеновая часть композиции, применяемая в способе, включает в себя формулу:

или представляет собой ее изомер, где
а) R3 выбран из группы, состоящей из водорода, гидроксила, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара и монотерпеновой группы;
b) формула дополнительно включает в себя R4, где R4 выбран из группы, состоящей из водорода, гидроксила, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара, С1-С5-алкилового сложного эфира и монотерпеновой группы.
Изомер может представлять собой либо цис-изомер, либо транс-изомер.
В других осуществлениях способа R3 представляет собой сахар. Сахар выбран из группы, состоящей из глюкозы, фукозы, рамнозы, арабинозы, ксилозы, хиновозы, мальтозы, глюкуроновой кислоты, рибозы, N-ацетилглюкозамина и галактозы. Композиция способа может дополнительно включать в себя другую монотерпеновую часть, присоединенную к сахару.
В других осуществлениях способа R3 имеет следующую формулу:

где R5 выбран из группы, состоящей из водорода, гидроксила, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара, С1-С5-алкилового сложного эфира и монотерпеновой группы.
В некоторых осуществлениях R5 представляет собой водород или гидроксил. Изомер может быть стереоизомером или оптическим изомером.
В других осуществлениях способа R3 имеет следующую формулу:

В альтернативных осуществлениях R3 имеет следующую формулу:
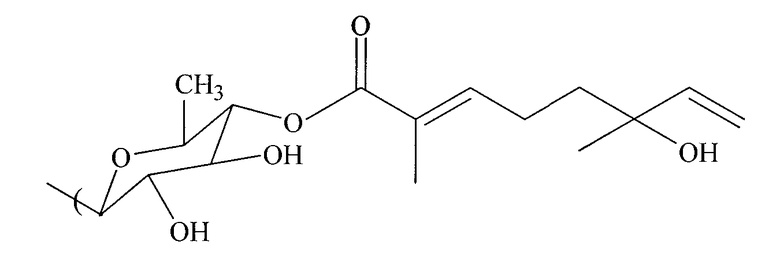
В некоторых конкретных осуществлениях способа монотерпеновая композиция включает в себя формулу:

или ее изомер, где
а) R1 и R2 выбраны из группы, состоящей из водорода, С1-С5-алкила и олигосахарида;
b) R3 выбран из группы, состоящей из водорода, гидроксила, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара и монотерпеновой группы;
с) формула дополнительно включает в себя R4, где R4 выбран из группы, состоящей из водорода, гидроксила, С1-С5-алкила, С1-С5-алкилена, С1-С5-алкилкарбонила, сахара, С1-С5-алкилового эфира и монотерпеновой группы, и где R4 может быть присоединен к тритерпеновой части или монотерпеновой части.
Изомер представляет собой стереоизомер или оптический изомер.
В других конкретных осуществлениях способа монотерпеновая композиция включает в себя формулу:
В других конкретных осуществлениях способа монотерпеновая композиция включает в себя формулу:

В других конкретных осуществлениях способа монотерпеновая композиция включает в себя формулу:
В других аспектах способа воспалительные ответы ингибируются, когда монотерпеновая композиция вводится в клетку в концентрации приблизительно от 0,5 до 2,0 мкг/мл.
Клетка находится у субъекта с воспалительным заболеванием. В предпочтительных аспектах субъект является человеком. В других аспектах субъект может быть животным других видов и может быть мышью или каким-либо другим животным-млекопитающим.
Воспалительное заболевание выбрано из группы, включающей в себя предраковое воспалительное заболевание, атеросклероз, ревматоидный артрит, остеоартрит, рассеянный склероз, болезнь Паркинсона и болезнь Альцгеймера.
Предраковое воспалительное заболевание может представлять собой эзофагит Барретта, воспалительное заболевание кишечника, хронический панкреатит, хронический простатит, семейный полипоз или лучевой кератоз. Часто данные предраковые состояния переходят в рак при отсутствии лечения. Таким образом, важна профилактика или лечение указанных предраковых состояний. Например, больные с определенными типами желудочно-пищеводного рефлюкса склонны к колоночной метаплазии нормальной сквамозной выстилки. Данное состояние, обозначаемое как эзофагит Барретта, имеется приблизительно у 10% больных с желудочно-пищеводным рефлюксом, и оно связано с наличием сужения, глубоких язв и последующим развитием аденокарциномы. Лучевой кератоз представляет собой предраковое состояние кожи, часто вызываемое действием солнечных лучей, и оно может вести к карциноме кожи. Воспалительные заболевания кишечника включают в себя состояния, подобные болезни Крона и язвенному колиту, которые могут приводить к раку ободочной кишки.
В определенных осуществлениях монотерпеновые композиции способа ингибируют фермент циклооксигеназу-2 (COX-2). В других осуществлениях монотерпеновые композиции способа ингибируют iNOS. Оба данных фермента являются нижележащими эффекторами NF-κB и вовлечены в разнообразные воспалительные и хемопревентивные ответы. Оба данных фермента индуцируются в ответ на различные цитокины, такие как интерферон гамма, митогены, микробные продукты, такие как липополисахариды и т.п. Например, монотерпеновые композиции согласно изобретению существенно снижают активацию NF-κB и экспрессию iNOS и COX-2 в ответ на провоспалительные агенты, такие как TNF и микробные продукты, такие как липополисахариды (LPS).
В других аспектах способа введение монотерпеновых композиций осуществляют локальным, местным или системным путями. Способ введения может быть путем инъекции, путем перорального употребления или путем местной аппликации. В других аспектах монотерпеновая композиция представляет собой фармацевтическую композицию в фармакологически приемлемой среде. Фармакологически приемлемая среда представляет собой буфер, растворитель, разбавитель, инертный носитель, масло, мазь или пригодный в пищу материал. Фармацевтическая композиция может дополнительно включать в себя направляющий агент. Направляющий агент может прямо доставлять фармацевтическую композицию к конкретному типу клеток, например к воспалительной клетке.
В данном описании или формуле изобретения термин "включающий в себя" может означать также "включающие в себя". Применяемый здесь термин "другой" может означать по меньшей мере второй или более.
Другие цели, характеристики и преимущества согласно изобретению станут ясными из последующего подробного описания. Должно быть понятно, однако, что подробное описание и конкретные примеры, представляя предпочтительные осуществления изобретения, даются только с целью иллюстрации, так как разнообразные вариации и модификации в пределах концепции и объема изобретения должны быть очевидными для специалистов в данной области техники из подробного описания.
Краткое описание чертежей
Следующие чертежи составляют часть настоящего описания и включены для дополнительной иллюстрации определенных аспектов согласно изобретению. Изобретение может быть лучше понято при ссылке на один или более из данных чертежей в сочетании с подробным описанием представленных здесь конкретных осуществлений.
Фиг. 1: Действие UA-BRF-004-DELEP-F001 на линии опухолевых клеток человека. На фиг. 1 представлено ингибирование роста линий клеток яичника (SK-OV-3, HEY, OVCAR-3), молочной железы (MDA-468), меланомы (A375-M, Hs294t) и эпидермоида человека (A431), обработанных сырым экстрактом стручковых растений.
Фиг. 2: Действие UA-BRF-004-DELEP-F023 (фракция 23) на трансформированные и нетрансформированные клеточные линии. На фиг. 2 представлена цитотоксичность, проявляемая фракцией 23, на линии клеток яичника (SK-OV-3, OCC1, HEY, OVCAR-3), Т-клеточного лейкоза (Jurkat), простаты (LNCaP), свежих опухолевых клеток яичника человека (FTC), фибробластов человека (FS) и эндотелиальных клеток (HUVEC). Была обнаружена только 15-17% цитотоксичность в отношении нетрансформированных клеток по сравнению с 50-95% цитотоксичностью в отношении опухолевых клеток.
Фиг. 3: Действие фракции 35 ("UA-BRF-004-DELEP-F035" или F035) на опухолевые клеточные линии человека. На фиг. 3 представлена цитотоксичность, проявляемая при обработке фракцией 35 линий клеток яичника человека (HEY, OVCAR-3, С-1, SK-OV-3), поджелудочной железы (PANC-1) и почек (769-P, 786-O, A498). IC50 для клеточных линий была заключена в диапазоне 1-6 мкг/мл.
Фиг. 4: Действие фракции 35 на лейкозные клеточные линии. На фиг. 4 показано, что фракция 35 проявляет сильную цитотоксичность в отношении клеток Jurkat (Т-клеточного лейкоза) с IC50 130 нг/мл и IC50 для клеток REH, KG-1 и NALM-6 (B-клеточного лейкоза) в диапазоне 1-3 мкг/мл.
Фиг. 5: Действие фракции 35 на пролиферацию эндотелиальных клеток. На фиг. 5 показано, что фракция 35 является сильным ингибитором пролиферации эндотелиальных клеток в присутствии или в отсутствие bFGF.
Фиг. 6: Действие фракции 35 на миграцию эндотелиальных клеток капилляров. На фиг. 6 показано отсутствие действия на миграцию эндотелиальных клеток капилляров, что предполагает отсутствие токсичности.
Фиг. 7: Представлена тонкослойная хроматография экстрактов сеянцев и наплывов. Дорожка 1, наплыв стебля, выращенный на среде BA-IAA; дорожка 2, наплыв корня, выращенный на среде BA-IAA; дорожка 3, наплыв гипокотиля; дорожка 4, сеянцы, обработанные метилжасмонатом (100 мкМ) на полутвердой среде; дорожка 5, контрольные сеянцы, растущие на полутвердой среде; дорожка 6, стандарт F023; дорожка 7, побег, выращенный на среде BA; дорожка 8, сеянцы, обработанные 50 мкМ метилжасмоната; дорожка 9, сеянцы, обработанные 100 мкМ метилжасмоната; дорожка 10, сеянцы, обработанные 200 мкМ метилжасмоната; дорожка 11, контрольные сеянцы; дорожка 12, стандарт F023.
Фиг. 8: Представлена фотография мыши SENCAR слева и мыши, полученной в результате скрещивания SENCAR и C57B1, справа. Обеих обрабатывали повторяющимися дозами 100 нмоль DMBA в течение 8 недель. К 15 неделям обе имели множественные папилломы, но мышь, полученная в результате скрещивания SENCAR и C57B1, имела меньшее количество папиллом и меньших размеров. Линия C57B1 является устойчивой к канцерогенезу и не дает опухолям развиться.
Фиг. 9A-F: Представлены срезы эпидермиса мышей, обработанных ацетоном, DMBA или DMBA + UA-BRF-004-DELEP-F035. Фиг. 9А: обработка ацетоном в течение 4 недель. Фиг. 9В: обработка ацетоном в течение 8 недель. Фиг. 9С: обработка DMBA в течение 4 недель. Фиг. 9D: обработка DMBA в течение 8 недель. Фиг. 9E: обработка DMBA + UA-BRF-004-DELEP-F035 в течение 4 недель. Фиг. 9F: обработка DMBA + UA-BRF-004-DELEP-F035 в течение 8 недель.
Фиг. 10A,B: Представлено антиоксидантное действие на ДНК UA-BRF-004-DELEP-F035 после 4 недель. Фиг. 10A: представлены антиоксидантные эффекты после обработки низкой концентрацией UA-BRF-004-DELEP-F035 (0,1 мг/0,2 мл). Фиг. 10B: представлены антиоксидантные эффекты после обработки высокой концентрацией UA-BRF-004-DELEP-F035 (0,3 мг/0,2 мл).
Фиг. 11A,B: Представлена толщина эпидермиса после 4-недельной обработки DMBA и UA-BRF-004-DELEP-F035. Фиг. 11A: представлено действие на толщину эпидермиса после обработки низкой концентрацией UA-BRF-004-DELEP-F035 (0,1 мг/0,2 мл). Фиг. 11B: представлено действие на толщину эпидермиса после обработки высокой концентрацией UA-BRF-004-DELEP-F035 (0,3 мг/0,2 мл).
Фиг. 12: Представлено процентное увеличение толщины эпидермиса через 4 недели после обработки DMBA при низкой (0,1 мг/0,2 мл) или высокой (0,3 мг/0,2 мл) концентрации UA-BRF-004-DELEP-F035.
Фиг. 13: Представлено процентное снижение папиллом через 8 недель обработки DMBA при низкой (0,1 мг/0,2 мл) или высокой (0,3 мг/0,2 мл) концентрации UA-BRF-004-DELEP-F035.
Фиг. 14: Представлена авторадиограмма реакции ПЦР, представляющая амплификацию мутации кодона 61 H-ras мыши.
Фиг. 15: Представлена первоначальная стратегия, применяемая для очистки и выделения биологически активных тритерпеновых соединений из Acacia victoriae.
Фиг. 16: Представлена общая улучшенная схема очистки, выделения и характеристики активных составляющих Acacia victoriae.
Фиг. 17A,B. Фиг. 17A: Представлен ВЭЖХ спектр ацетилированных сахаров, выделенных из гидролизованных активных составляющих, обнаруженных во фракции 94 ("UA-BRF-004Pod-DELEP-F094" или F094). Фиг. 17В: Представлен ВЭЖХ спектр ацетилированных сахаров, выделенных из гидролизованных активных составляющих, обнаруженных в F094.
Фиг. 18A-F. Фиг. 18A: Представлены ВЭЖХ спектры UA-BRF-004-DELEP-F035 и F035-B2. Фиг. 18B: Представлены ВЭЖХ спектры UA-BRF-004Pod-DELEP-F094. Фиг. 18C: Представлены ВЭЖХ спектры F140. Фиг. 18D: Представлены ВЭЖХ спектры F142. Фиг. 18E: Представлены ВЭЖХ спектры F144. Фиг. 18F: Представлены ВЭЖХ спектры F145.
Фиг. 19A,B: Анализ клеточного цикла клеток OVCAR-3 до и после обработки (48 час) фракцией 35. На фиг. показано, что наблюдается 8% увеличение числа клеток в фазе G1 и 10% снижение числа клеток в фазе S клеточного цикла после обработки фракцией 35, что отражает приостановку вхождения в фазу G1. Фиг. 19A: анализ клеточного цикла необработанных опухолевых клеток OVCAR-3. Фиг. 19B: анализ клеточного цикла опухолевых клеток OVCAR-3, обработанных фракцией 35.
Фиг. 20: EMSA, демонстрирующий заметное ингибирование NF-κB, активированного TNF, в результате экспозиции клеток с UA-BRF-004-DELEP-F035 и UA-BRF-004Pod-DELEP-F094. Применялась следующая обработка: дорожка 1, необработанные; дорожка 2, TNF (100 пМ); дорожка 3, UA-BRF-004-DELEP-F035 (1 мкг/мл); дорожка 4, TNF + F035 (1 мкг/мл); дорожка 5, F035 (2 мкг/мл); дорожка 6, TNF + F035 (2 мкг/мл); дорожка 7, F094 (1 мкг/мл); дорожка 8, TNF + F094 (1 мкг/мл); дорожка 9, F094 (2 мкг/мл); дорожка 10, TNF + F094 (2 мкг/мл).
Фиг. 21: Определение киназы липидов, демонстрирующее ингибирование PI3-киназы UA-BRF-004-DELEP-F035 и вортманнином.
Фиг. 22: Гель после ДДС-Na ПААГ-электрофореза, проанализированный с помощью иммуноблоттинга-ECL с применением антител, специфичных в отношении фосфата AKT и общего AKT. Последующая обработка клеток 1 и 2 мкг/мл UA-BRF-004-DELEP-F035 вызывала отчетливое ингибирование фосфорилирования AKT (активный AKT), которое было сходным с таковым после 2 часовой обработки клетки 1 мкМ вортманнином.
Фиг. 23: Раскрывает амплификацию PCRTM части гена rol B из четырех независимо трансформированных клонов корня. (Дорожки L-R, 1: маркеры в т.п.н., 2: позитивный контроль (ДНК плазмиды из штамма R1000), 3: негативный контроль (ДНК из нетрансформированного корня). 4-7: четыре независимо трансформированных клона корня. Обратить внимание на амплификацию фрагмента 645 п.н. в позитивном контроле и трансформированных корнях.
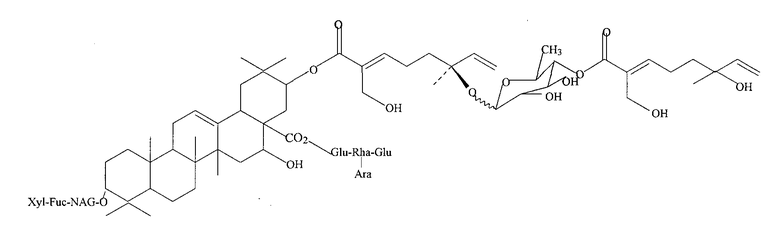
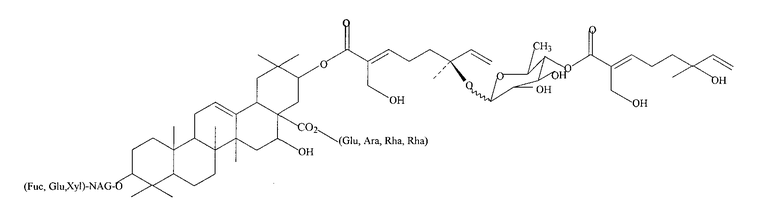
Фиг. 24: Структура эллиптозида A и эллиптозида E (Beutler, 1997).
Фиг. 25: Разделение ВЭЖХ составляющих в F094.
Фиг. 26: Разделение ВЭЖХ составляющих в F035.
Фиг. 27: Первое фракционирование с помощью полупрепаративной ВЭЖХ F094.
Фиг. 28: Второе фракционирование с помощью полупрепаративной ВЭЖХ F094.
Фиг. 29: Препаративное фракционирование F094.
Фиг. 30: Анализ препаративной фракции D.
Фиг. 31: Анализ препаративной фракции G/H.
Фиг. 32: Соединение G1 после второй очистки на колонке PFP.
Фиг. 33: Соединение G1 после окончательной очистки С-18.
Фиг. 34: Соединение D1 после очистки на колонке С-18 Waters.
Фиг. 35: Соединение D1 после окончательной очистки С-18-Aq.
Фиг. 36: Изображены соединения, получаемые после деградации соединения D1.
Фиг. 37: Изображены соединения, получаемые после деградации соединения G1.
Фиг. 38: Изображены соединения, получаемые после деградации соединения B1.
Фиг. 39: Структура тритерпенового гликозида D1.
Фиг. 40: Структура тритерпенового гликозида G1.
Фиг. 41: Структура тритерпенового гликозида B1.
Фиг. 42: Действие тритерпеновых гликозидов (F035) на линии раковых и нормальных клеток. F035 оценивалась на цитотоксичность с помощью процедур, описанных в примерах. Активность F035 определяли с помощью панели линий раковых и нормальных клеток. IC50 находилась в диапазоне 0,2-5,8 мкг/мл для раковых клеток. Не наблюдалось заметной цитотоксичности (IC50 от 15 мкг/мл до > 25 мкг/мл) для нормальных и иммортализированных клеточных линий.
Фиг. 43: Профиль цитотоксичности очищенных тритерпеновых гликозидов D1 и G1 на линиях клеток рака человека. Очищенные экстракты оценивали на их активность в отношении следующих линий клеток рака человека: Jurkat (Т-клеточного лейкоза), варианта С-2 Hey (яичников), 769-P (почек), MDA-MB-231, MDA-MB-453 (молочной железы). Результаты представлены как среднее + ср. кв. ошибка.
Фиг. 44: Действие очищенных соединений D1 и G1 и смеси тритерпеновых гликозидов (F035) на апоптоз: апоптоз измеряли с применением теста связывания аннексина V, в котором клетки окрашивали аннексином-V-ФИТЦ и для содержания ДНК иодидом пропидия (PI) и анализировали с помощью проточной цитометрии. Клетки инкубировали в течение 16 часов с 0,5-1,0 мкг/мл экстрактов. После 16-часовой обработки наблюдались три популяции клеток. Клетки, которые погибли или были на поздней стадии апоптоза (позитивные по аннексину-V-ФИТЦ и PI), клетки, подвергающиеся апоптозу (позитивные по аннексину-V-ФИТЦ и негативные по PI), и клетки, которые были жизнеспособными и не подвергающимися апоптозу (негативные по аннексину-V-ФИТЦ и по PI; нижний левый квадрат).
Фиг. 45 A,B: Ингибирование активности PI3-киназы и фосфорилирования AKT. Способность фосфорилировать фосфатидилинозитол (PI) измеряли для иммунопреципитатов белка p85 из клеточных лизатов. Авторадиограммы на активность киназы in vitro с разделением с помощью тонкослойной хроматографии для иммунопреципитатов р85 с применением клеток Jurkat. Фиг. 45B: ингибирование фосфорилирования AKT по Ser-473 и Thr-308 сырыми и чистыми тритерпеновыми гликозидами. Клетки Jurkat инкубировали с сырым (F035) и очищенным экстрактами D1 и G1 в течение 16 часов при 37°С. Клеточные лизаты разделяли на 9% ДДС-Na ПААГ-электрофорезе и анализировали с помощью иммуноблоттинг-ECL анализа с применением в качестве зондов антител против Ser-473, Thr-308 и против общего AKT.
Фиг. 46A-D. Ингибирование NF-κB, индуцированного TNF, и индукция iNOS тритерпеновыми гликозидами: клетки Jurkat экспонировали с различными концентрациями F035 (1-4 мкг/мл; фиг. 46А) и 2 мкг/мл чистых экстрактов (D1 и G1; фиг. 46В) в течение 16 часов и NF-κB активировали 100 пМ TNF в течение 15 мин при 37°С. ДНК-белковый комплекс разделяли на 7,5% неденатурирующих полиакриламидных гелях и полосы радиоактивности визуализировали и оценивали количественно с помощью PhophoImager. NOS индуцировали в U-937 (фиг. 46С) и клетках Jurkat (фиг. 46D), как описано в методах. Клеточные белки разделяли на ДДС-Na ПААГ-электрофорезе и анализировали с помощью иммуноблоттинг-ECL анализа с применением антител против iNOS.
Фиг. 47: Действие F035 и D1 на расщепление PARP в клетках Jurkat.
Фиг. 48: Действие z-vad fmk на индуцируемое F035 расщепление PARP в клетках Jurkat.
Фиг. 49: Действие F035, F094, D1 и G1 на активность каспазы в клетках Jurkat.
Фиг. 50: Действие F035 на высвобождение цитохромов из митохондрий клеток Jurkat.
Фиг. 51А и фиг. 51В: Действие F094 и монотерпенового/тритерпенового гликозида G1 на активацию NF-κB, индуцированную TNF. Клетки Jurkat (1 х 106/мл) обрабатывали 2 мкг/мл F094 (фиг. 51А) или монотерпеновым/тритерпеновым гликозидом G1 (фиг. 51В) в течение 1-16 часов при 37°С. В конце обработки клетки промывали, ресуспендировали в количестве 2 х 106/мл в полной среде и обрабатывали 1 нМ TNF в течение 15 мин при 37°С. Получали ядерный экстракт и исследовали его на активацию NF-κB, как описано в разделе примеров.
Фиг. 52А и фиг. 52В: Фиг. 52А. Дозозависимость ингибирования монотерпеновым/тритерпеновым гликозидом G1 активации NF-κB, индуцированной TNF. Клетки Jurkat (1 х 106/мл) обрабатывали различными концентрациями монотерпенового/тритерпенового гликозида G1 в течение 16 часов при 37°С. В конце обработки клетки промывали, ресуспендировали в количестве 2 х 106/мл в полной среде и обрабатывали 1 нМ TNF в течение 15 мин при 37°С. Получали ядерный экстракт и исследовали его на активацию NF-κB, как описано в разделе примеры. Фиг. 52В. Суперсдвиг и анализ специфичности NF-κB. Ядерный экстракт клеток, обработанных TNF, инкубировали в течение 15 мин с мутантным олигонуклеотидом NF-κB, немеченым олигонуклеотидом NF-κB, анти-р65 антителом и неиммунной сывороткой кролика. Затем оценивали активацию NF-κB, как описано в разделе примеров.
Фиг. 53А и фиг. 53В: Действие монотерпенового/тритерпенового гликозида G1 на индуцированные TNF IκBα (фиг. 53А) и ядерную транслокацию р65 (фиг. 53В). Клетки Jurkat обрабатывали монотерпеновым/тритерпеновым гликозидом G1 (2 мкг/мл, 16 часов), промывали и инкубировали с 1 нМ TNF в течение различных промежутков времени. Применяли цитоплазматический и ядерный экстракты данных клеток для исследования уровней IκBα и р65, соответственно, с помощью иммуноблоттинга, как описано в разделе примеров.
Фиг. 54А, фиг. 54В и фиг. 54С: фиг. 54А. Действие добавления in vitro монотерпенового/тритерпенового гликозида G1 на связывание NF-κB с ДНК. Экстракты стимулированных TNF клеток Jurkat обрабатывали различными концентрациями монотерпенового/тритерпенового гликозида G1 в течение 30 мин при 37°С и анализировали на связывание NF-κB с помощью ENSA. Фиг. 54В. Действие монотерпенового/тритерпенового гликозида G1 на индуцированную дезоксихолатом (DOC) активацию NF-κB. Цитоплазматические экстракты необработанных клеток обрабатывали DOC в присутствии или в отсутствие монотерпенового/тритерпенового гликозида G1 и затем анализировали на активацию NF-κB. Фиг. 54С. Действие DTT на индуцированное монотерпеновым/тритерпеновым гликозидом G1 ингибирование активации NF-κB.
Фиг. 55А и фиг. 55В: фиг. 55А. Действие монотерпенового/тритерпенового гликозида G1 на активность экспрессии гена люциферазы, зависимого от NF-κB. Клетки Jurkat трансфицировали pGL3-NF-κB с помощью электропорации. NF-κB активировали, применяя LPS (100 нг/мл), PMA (5 нг/мл) или TNF (1 нМ). Активность люциферазы определяли, применяя набор для определения люциферазы (Promega, Madison, WI), придерживаясь инструкций производителя. Фиг. 55В. Действие F094 и монотерпенового/тритерпенового гликозида G1 на индуцированную LPS экспрессию iNOS и COX-2. Клетки RAW 264.7, предварительно обработанные F094 и монотерпеновым/тритерпеновым гликозидом G1, обрабатывали LPS, как описано в методах. Экспрессию iNOS и COX-2 тестировали в цитоплазматических экстрактах данных клеток, применяя анализ с помощью иммуноблоттинга.
Фиг. 56: (Фиг. 56А) Профиль ВЭЖХ авицинов. (Фиг. 56В) Химическая структура авицина D и авицина G.
Фиг. 57: Связывание аннексина-V-ФИТЦ в клетках Jurkat, обработанных F094 или авицином. Клетки Jurkat (1 х 106/мл) обрабатывали 2 мкг/мл F094, авицином D и авицином G в течение различных периодов времени. Клетки окрашивали и анализировали с помощью проточной цитометрии.
Фиг. 58: Действие F094 или авицинов на высвобождение цитохрома c из митохондрий. Клетки Jurkat (1 х 107) обрабатывали F094, авицином D или авицином G (все в концентрации 2 мкг/мл) при 37°С в течение указанных периодов времени. Клетки гомогенизировали и лизаты исследовали на уровни цитохрома с с помощью иммуноблоттинга.
Фиг. 59: (Фиг. 59А). Кинетика высвобождения цитохрома c из фракции митохондрий клеток Jurkat в бесклеточной системе. Митохондрии выделяли, как описано в методах. Обработку авицином G (2 мкг/мл) проводили в течение 0, 1, 2, 5, 10 и 20 мин при 37°С. (Фиг. 59В). Дозовая зависимость высвобождения цитохрома с, индуцированного авицином G. Выделенные митохондрии обрабатывали различными концентрациями авицина G в течение 10 мин при 37°С. (Фиг. 59С). Действие ингибиторов каспазы на высвобождение цитохрома с, индуцированное авицином G. Выделенные митохондрии предварительно обрабатывали DEVD-CH2F (25 мкМ) и zVAD-fmk (25 мкМ) в течение 5 мин при 37°С. Высвобождение цитохрома с анализировали с помощью иммуноблоттинга.
Фиг. 60: (Фиг. 60А). Кинетика активации каспазы-3, индуцированной F094 или авицинами. Клетки Jurkat (1 х 106) обрабатывали 2 мкг/мл F094, авицином D или авицином G в течение различных периодов времени. Определяли активность каспазы-3 в цитозольных экстрактах данных клеток. (Фиг. 60В). Расщепление PARP под действием F094 или авицинов. Клетки Jurkat (3 х 106) обрабатывали 2 мкг/мл агентов в течение указанных периодов времени. Получали клеточные лизаты и тестировали на расщепление PARP. (Фиг. 60С). Действие обработки zVAD-fmk на расщепление PARP, индуцированное F094 или авицинами. Клетки (3 х 106) культивировали ± zVAD-fmk (100 мкМ) в течение 1 часа при 37°С и затем обрабатывали 2 мкг/мл F094 или авицинов в течение 4 часов при 37°С.
Фиг. 61: Действие F094 или авицинов на потенциал митохондриальной мембраны. Клетки Jurkat (1 х 106/мл) обрабатывали 2 мкг/мл F094, авицином D или авицином G в течение различных периодов времени. Клетки окрашивали DiOC6 и анализировали с помощью проточной цитометрии.
Фиг. 62: Действие F094 или авицинов на генерацию активных форм кислорода. Клетки Jurkat (5 х 104/лунка) обрабатывали 1, 2 и 4 мкг/мл каждым из F094, авицина D или авицина G.
Подробное описание изобретения
Настоящее изобретение направлено на преодоление ограничений предшествующего уровня техники путем предложения способов, которые тормозят воспаление с помощью обеспечения клетки, нуждающейся в этом, монотерпеновой композицией, которая ингибирует NF-κB. В некоторых осуществлениях монотерпеновые композиции согласно изобретению ингибируют ферменты, такие как iNOS и COX-2.
Монотерпеновые композиции согласно изобретению могут придавать мембранам растворимость/проницаемость путем дополнительного присоединения молекулы-носителя. Данная молекула-носитель может представлять собой любую часть, которая может обеспечивать внутриклеточную доступность или мембранную проницаемость для монотерпеновых композиций. Молекулы-носители в некоторых осуществлениях могут быть тритерпеновыми частями.
Поскольку монотерпеновые композиции согласно изобретению могут включать в себя тритерпеновые части, они как в этой, так и в других частях описания могут быть обозначены как тритерпеновые соединения или тритерпенгликозидные композиции. Альтернативно, здесь и в других частях описания они также обозначаются как монотерпеновые/тритерпеновые соединения или композиции.
В изобретении применяются способы ингибирования опосредуемого NF-κB воспаления. Поскольку NF-κB представляет собой транскрипционный фактор, который регулирует транскрипцию ряда генов, участвующих в иммунных и воспалительных путях, таких как провоспалительные цитокины, молекулы адгезии и апоптоз, нарушение регуляции NF-κB вносит свой вклад во множество патологических состояний, таких как септический шок, острое воспаление, репликация вирусов и некоторые виды рака. Следовательно, ингибиторы и модуляторы NF-κB важны для предотвращения и лечения воспаления и предраковых состояний. В дополнение к ингибированию NF-κB монотерпеновые композиции согласно изобретению также ингибируют ферменты iNOS и COX-2.
Таким образом, монотерпеновые композиции данного изобретения обеспечивают лечение многочисленных состояний, включая воспаление, таких как предраковые воспалительные заболевания, атеросклероз, ревматоидный артрит, остеоартрит, рассеянный склероз, болезнь Паркинсона и болезнь Альцгеймера. Примеры предраковых воспалительных заболеваний включают в себя такие заболевания, как эзофагит Барретта, воспалительное заболевание кишечника, хронический панкреатит, хронический простатит, семейный полипоз и лучевой кератоз.
I. Очистка и идентификация тритерпеновых соединений
Также описаны идентификация и очистка монотерпеновых/тритерпеновых соединений из Acacia victoriae. Идентифицированные соединения проявляют высокую противоопухолевую активность в концентрациях, которые оказывают слабую цитотоксичность или не обладают цитотоксичностью в отношении нормальных клеток человека.
Тритерпеновые соединения были идентифицированы в результате направленного скрининга 60 растительных экстрактов из выбранных стручковых видов, естественных для засушливых и полузасушливых регионов. В результате первоначального скрининга был выделен один экстракт, обозначенный UA-BRF-004-DELEP-F001 из Acacia victoriae (Benth.) (Leguminosae), проявивший высокую противоопухолевую активность против множества линий опухолевых клеток человека. Данный экстракт далее был очищен с получением отдельных фракций. В двух циклах очистки был идентифицирован экстракт, который содержал очищенные противоопухолевые соединения. Было установлено, что данный экстракт содержит очищенные тритерпенгликозидные сапонины. В последствии был разработан способ эффективного выделения активных соединений.
Дополнительное тестирование более очищенного экстракта дополнительно выявило биологические активности экстракта. Очищенный экстракт проявил повышенную противоопухолевую активность по сравнению с сырым экстрактом в концентрациях, которые не проявляли или проявляли слабую токсичность в отношении нормальных клеток человека. Дополнительно было показано, что экстракт обладает хемопротекторным действием у мышей, экспонированных с канцерогенами.
Растение, из которого был получен экстракт, Acacia victoriae, было выбрано с учетом факторов, включающих в себя природную среду и ограниченную предшествующую изученность видов. Acacia victoriae происходит из Австралии, но была внедрена в качестве садовой культуры широко в мире и широко известна как колючий плетень или элегантный плетень. Дерево растет со скоростью от 60 до 120 см в год, медленно сбрасывает листву при засухе и выдерживает по меньшей мере -15°С. Взрослые растения вырастают до 10-15 футов и имеют голубовато-зеленые дважды перистые листья. На юго-западе Соединенных Штатов растение обычно цветет с апреля по май с созреванием стручков в июне. Acacia victoriae имеет ряд сельскохозяйственных применений, включая защиту от ветра, защитные полосы, пищу, стабилизацию критических областей и в качестве декоративного растения с низким потреблением воды. Семена различных видов Acacia применялись в качестве источника пищевого материала туземцами Австралии в течение многих поколений (Lister et al., 1996). Среди Acacia вид Acacia victoriae является наиболее обычным и широко распространенным повсеместно в Австралии и, следовательно, является наиболее широко применяемым видом. Семена Acacia, обычно называемые семенными сережками, пользуются большой популярностью при применении в качестве основного продукта в кондитерских изделиях и хлебобулочных продуктах, а также в качестве отдушки в десертах, особенно в мороженом. Их также применяют для получения высококачественного напитка, подобного кофе, и среди видов Acacia вид Acacia victoriae (Benth.) обычно рассматривается как обладающий наилучшим запахом (Lister et al., 1996). Однако нет сообщений о применении стручков и корней этого растения.
Важным аспектом в применении растительных экстрактов в качестве фармакологических препаратов является характеристика и определение индивидуальных активных составляющих. То же самое относится к случаю тритерпенсапониновых препаратов, для которых требуются изощренные способы выделения, выяснения структуры и анализа их компонентов и гликозидов. Когда производят биологическое тестирование чистых соединений, необходимо их выделение в достаточном количестве и с достаточной чистотой.
Поскольку тритерпены и другие родственные сапонины обладают относительно высокой молекулярной массой и являются высоко полярными, их выделение может быть проблематичным. Проблема, связанная с выделением чистых сапонинов, заключается в наличии сложных смесей близкородственных соединений, слегка различающихся природой агликоновой или сахарной части (природа, количество, положения и хиральность присоединения моносахаридов). Сложности также встречаются в связи с лабильными заместителями, такими как сложные эфиры. Например, главный сапонин подлинного соевого боба, γ-пироновое производное (BOA) экстрагируется лишь водным этанолом при комнатной температуре. Экстракция при нагревании (80°С) ведет к отщеплению эфирной части и образованию сапонина сои I (Bb) (Kudou et al., 1992). В растениях сапонинам сопутствуют очень полярные вещества, такие как сахариды и красящие вещества, включая фенольные соединения и тому подобное, плохо кристаллизуются и могут быть гигроскопичными, что еще более затрудняет получение кристаллов.
Характеристика чистых сапонинов также является затрудненной из-за отсутствия кристаллического материала. Точки плавления являются неточными и часто определяются при распаде. Следовательно, определения чистоты образца не могут быть выполнены лишь на основе точки плавления, величины оптического вращения или другой физической константы. Лучшим тестом чистоты сапонина является проверка с помощью ТСХ или ВЭЖХ, если возможно, путем совместной хроматографии с аутентичным образцом. Окраска точек на пластинах ТСХ после опрыскивания подходящими реагентами служит дополнительным показателем потенциально индивидуальных компонентов. Например, один из тритерпеновых гликозидов изобретения, D1, имеет время удерживания 15,2 минуты при ВЭЖХ. Это отличает его от другого родственного соединения, эллиптозида E, выделенного John Beutler et al., 1997, из Archidendron ellipticum, который обладает временем удерживания 12,5 минуты при ВЭЖХ. Дополнительная характеристика тритерпенов согласно изобретению показывает, что данное различие во времени удерживания обусловлено по меньшей мере различиями в хиральности и в двойных связях D1 и в сообщавшихся свойствах эллиптозида E.
(i) Химическая очистка
Способы химической очистки хорошо известны специалистам в данной области техники. Данные способы включают в себя на первой стадии грубое фракционирование растительного экстракта на описанные здесь тритерпенгликозидные соединения. При наличии в общем виде разделенных соединений согласно изобретению из растительного материала интересующие тритерпеновые гликозиды могут быть дополнительно очищены с помощью описанных здесь способов, например хроматографических способов, для достижения частичной или полной очистки (или очистки до гомогенности). Аналитические способы, особенно пригодные для получения чистой композиции тритерпенового гликозида, специально раскрыты здесь ниже.
Определенные аспекты согласно изобретению касаются очистки и - в конкретных осуществлениях - существенной очистки монотерпеновых/тритерпеновых гликозидов из растительного материала. В предпочтительном осуществлении изобретения монотерпеновые/тритерпеновые гликозиды очищают из растения семейства Leguminosae или более предпочтительно из рода Acacia, и наиболее предпочтительно из вида Acacia victoriae, и еще более предпочтительно из вида Acacia victoriae (Benth.). Применяемый здесь термин "выделенный монотерпеновый/тритерпеновый гликозид" предназначен для обозначения композиции, отделяемой от других компонентов, где композиция очищается в какой-либо степени относительно имеющегося в природе состояния.
В целом "выделенная" будет относиться к органической молекуле или группе сходных молекул, которые были подвергнуты фракционированию для удаления различных других компонентов, и когда композиция в значительной мере сохраняет выраженную биологическую активность. Когда применяют термин "в значительной мере очищенная", данное обозначение будет относиться к композиции, в которой монотерпеновые или тритерпеновые композиции составляют главный компонент композиции, такой, который составляет приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90%, приблизительно 95% и более молекул в композиции.
Нет необходимости в том, чтобы монотерпеновые/тритерпеновые композиции согласно изобретению всегда выделялись и предлагались в их наиболее чистой форме. Действительно, ожидается, что менее существенно очищенные продукты найдут свое применение в определенных осуществлениях. Например, заявители допускают применение высушенных корней и стручков Acacia victoriae и их экстрактов в качестве пищевых добавок. Пищевые добавки по определению содержат смесь различных биологически активных соединений, которые синергично оказывают благоприятное действие на здоровье. Пищевые добавки согласно изобретению могут находиться в форме таблеток или капсул и могут приниматься перорально или альтернативно могут содержать экстракты растения в мази, которая может наноситься местно. Частичная очистка может быть осуществлена с применением немногочисленных стадий очистки в сочетании или путем использования различных форм одной и той же общей схемы очистки. Например, ожидается, что катионообменная колоночная хроматография, проведенная с применением аппарата для ВЭЖХ, обычно должна приводить к большей "кратности" очистки, чем тот же способ с применением системы хроматографии низкого давления. Способы, проявляющие меньшую степень относительной очистки, могут обладать преимуществами в общем выходе продукта или сохранении биологической активности монотерпеновых/тритерпеновых соединений.
(ii) Экстракция и предварительная очистка
Процедуры экстракции должны быть мягкими, насколько это возможно, поскольку некоторые сапонины могут подвергаться трансформациям, включая ферментативный гидролиз во время водной экстракции, этерификацию кислых сапонинов во время обработки спиртом, гидролиз лабильных сложноэфирных групп и трансацилирование. Следовательно, следует проявлять осторожность при осуществлении отдельных этапов процедуры выделения, например, при тонкослойной хроматографии.
Хотя возможны многочисленные вариации, настоящие общие процедуры для получения сырых смесей сапонинов обычно включают в себя экстракцию метанолом, этанолом, водой или водным спиртом; стадию обезжиривания обычно с помощью пертролейного эфира, выполняемую перед стадией экстракции, или самого экстракта; растворение или суспендирование экстракта в воде; встряхивание или промывку раствора или суспензии н-бутанолом, насыщенным водой, и осаждение (необязательное) сапонинов диэтиловым эфиром или ацетоном. Может также быть включена стадия диализа для удаления небольших водорастворимых молекул, таких как сахара (см., например, Zhou et al., 1981; Massiot et al., 1988).
Наиболее эффективная экстракция сухого растительного материала достигается с помощью метанола или водного метанола. Метанол также применяют для свежего растительного материала. Хотя вода обычно является менее эффективным растворителем сапонинов (если не требуются конкретные водорастворимые гликозиды), она имеет преимущества в виде легкого лиофилизирования и обеспечения более чистого экстракта. В зависимости от пропорции воды, применяемой для экстракции, могут быть получены либо монодесмозидные, либо бидесмозидные сапонины (Domon and Hostettmann, 1984; Kawamura et al., 1988). Свежий вегетативный материал содержит активные ферменты (эстеразы), которые при гомогенизации с растворителем способны превращать бидесмозиды в монодесмозиды. Даже сухой материал может содержать эстеразы, которые активируются в присутствии воды. В случае момордина I (монодесмозидного сапонина олеаноловой кислоты) было обнаружено, что превращение в момордин II (соответствующий бидесмозиду) происходит в воде и в 30% и 60% растворах метанола, но не в 80% и 100% растворах метанола. Напротив, гомогенаты свежих корней в метаноле сохраняли ферментативную активность. Однако ферменты могли быть инактивированы путем исходного замачивания свежих корней в 4% хлористоводородной кислоте и было показано, что бидесмозид является главным компонентом. Таким образом, ясно, что правильный выбор процедуры экстракции является чрезвычайно важным первым этапом.
Обычно применяемые для очистки белков такие способы, как диализ, ионообменная хроматография и хроматография исключения по размеру, пригодны для частичного отделения сапонинов в водном растворе от несапониновых компонентов, но в целом являются неэффективными в разделении отдельных сапонинов из-за склонности сапонинов образовывать смешанные мицеллы. В связи с этим для эффективного разделения обычно требуется применение органических растворителей или систем растворитель/вода, которые солюбилизируют амфифильные сапонины в виде мономеров, в результате чего образование смешанных мицелл не мешает разделению.
Общей проблемой, характерной для фуростаноловых сапонинов, является образование 22-OCH3 производных во время экстракции метанолом. Однако генуиновые 22-гидроксифуростанолы могут быть получены либо путем экстракции другим растворителем (например, пиридином), либо с помощью обработки метоксилированных искусственных продуктов кипящим водным ацетоном (Konishi and Shoji, 1979).
(iii) Тонкослойная хроматография (ТСХ)
Качественный анализ тритерпеновых сапонинов с помощью ТСХ имеет большое значение для всех аспектов исследований сапонинов. Пластины ТСХ (обычно силикагелевые) можно использовать как для чистых сапонинов, так и для сырых экстрактов, они являются недорогими, быстрыми в применении и не требуют специального оборудования. Доступен ряд визуализирующих реагентов для опрыскивания пластин (Таблица 2). Способы получения наиболее общих реагентов являются следующими.
• Ванилин-серная кислота (реагент Година). 1% раствор ванилина в этаноле смешивают в соотношении 1:1 с 3% раствором перхлорной кислоты в воде и опрыскивают им пластину ТСХ. После этого следует 10% раствор серной кислоты в этаноле и нагревание при 110°С.
• Реагент Либерманна-Бурчарда. Концентрированную серную кислоту (1 мл) смешивают с уксусным ангидридом (20 мл) и хлороформом (50 мл). Нагревание при 85-90°С дает требуемое окрашивание на пластине ТСХ.
• Хлорид антимония (III). Пластину ТСХ опрыскивают 10% раствором хлорида антимония в хлороформе и нагревают до 100°С.
• Анисальдегид-серная кислота. Анисальдегид (0,5 мл) смешивают с ледяной уксусной кислотой (10 мл), метанолом (85 мл) и концентрированной серной кислотой (5 мл). Данным раствором опрыскивают пластину ТСХ, которую затем нагревают при 100°С.
Опрыскивание ванилин-серной кислотой в присутствии этанола и перхлорной кислоты, например, дает синее или фиолетовое окрашивание с тритерпеновыми сапонинами. С анисальдегид-серной кислотой получается синее или фиолетово-синее окрашивание при нагревании пластины ТСХ. Опрыскивание пластин ТСХ раствором сульфата церия в серной кислоте дает фиолетово-красные, синие или зеленые флуоресцентные зоны под УФ-светом при 365 нм (Kitagawa et al., 1984b). В некоторых случаях для выявления присутствующих сапонинов достаточно простое опрыскивание пластин водой. Дополнительные реагенты для опрыскивания можно найти, например, у Stahl (1969).
Наиболее часто применяемым растворителем для ТСХ является смесь хлороформ-метанол-вода (65:35:10), но также применяют и другие растворители. Растворитель - смесь н-бутанол-этанол-аммиак (7:2:5) - особенно пригоден для гликозидов, содержащих остатки уроновой кислоты, т.е. для очень полярных смесей. Другие широко применяемые растворители включают в себя смесь н-бутанол-уксусная кислота-вода (4:1:5; верхний слой) или смесь хлороформ-метанол-уксусная кислота-вода (60:32:12:8).
Системы, применяемые для ТСХ гликоалкалоидов, обычно включают в себя смесь этилацетат-пиридин-вода (30:10:30; верхняя фаза). Визуализацию осуществляют с помощью стероидных реагентов (анисальдегид-серная кислота) или с применением алкалоидных реагентов (реагент Драгендорффа, сульфат церия (IV)). Другие системы ТСХ и реагенты визуализации приведены у Jadhav et al. (1981) и Baerheim Svendsen and Verpoorte (1983).
ТСХ позволяет провести многочисленные количественные определения. Например, плотность пятен, полученных с подходящим распыляемым реагентом, может быть прямо измерена с помощью денситометра. Альтернативно, количественные определения возможны путем проведения ТСХ разделений, соскребания соответствующей полосы с пластины (локализация которой определена, например, с помощью паров иода), элюирования сапонина и измерения поглощения УФ после добавления подходящего реагента (например, концентрированной серной кислоты).
В продаже имеются пластины ТСХ с обращенной фазой (ОФ-ТСХ), которые обеспечивают прекрасный способ анализа сапонинов, который является дополняющим к ТСХ на пластинах силикагеля. Почти исключительное применение смесей метанол-вода и ацетонитрил-вода используется для проявления пластин с обращенной фазой (например, пластин Merck RP-8 или RP-18 ВЭТСХ). Альтернативно, в случае DIOL ВЭТСХ могут быть применены пластины со стеклянной подложкой. Их можно применять с растворителями ТСХ-типа с нормальным силикагелем или с растворителями метанол-вода и ацетонитрил-вода, как для ОФ-ТСХ.
Иллюстративные реагенты для определения с помощью ТСХ и для спектрофотометрического и колориметрического определения сапонинов перечислены ниже в таблице 2.
1. Центрифужная тонкослойная хроматография (Ц-ТСХ)
Процедура Ц-ТСХ представляет собой метод плоской хроматографии, относящийся к препаративной тонкослойной хроматографии (ТСХ), но без необходимости соскребать полосы с пластины ТСХ (Hostettmann, 1980). Ц-ТСХ основана на действии центрифужной силы для ускорения прохождения подвижной фазы через круглую пластину ТСХ. Пластина, покрытая подходящим растворителем (толщиной в 1, 2 или 4 мм), вращается при приблизительно 800 об/мин под действием электрического мотора, в то время как внесение образца осуществляется в центре, и элюент нагнетается по поверхности сорбента. Элюирование растворителя дает концентрические полосы по поверхности пластины. Происходит центробежный слив по краям, и он собирается для анализа ТСХ. На 2 мм слое сорбента возможно разделение смеси в 50-500 мг.
Сочетание Ц-ТСХ смесью хлороформ-метанол-вода (100:30:3) и колоночной хроматографии описано для выделения гинсенозидов (Hostettmann, 1980). Сапонины также получали со смесями хлороформ-метанол-вода на пластинах силикагеля. Два гликозида протопримулагенина А из корней Eleutherococcus senticosus (Araliaceae) были очищены с помощью Ц-ТСХ (смесь хлороформ-метанол-вода, 65:35:7) после колоночной хроматографии на силикагеле и гель-фильтрации на сефадексе LH-20 (Segiet-Kujawa and Kaloga, 1991). Для выделения циклоартановых гликозидов из Passiflora quadrangularis (Passifloraceae) применяли систему растворителей этилацетат-этанол-вода (8:2:1 или 16:3:2) при скорости прохождения либо 1 мл/мин (Orsini et al., 1987) или 1,5 мл/мин (Orsini and Verotta, 1985).
Центрифужный жидкостный хроматограф Hitachi, модель CLC-5, описан для применения при разделении сапонинов. Хроматографию осуществляют с помощью данного устройства на пластинах силикагеля с элюентом хлороформ-метанол-вода (7:3:1(нижняя фаза) → 65:35:10 (нижняя фаза)). Применяя данную процедуру, был хроматографирован суммарно 1 г полуочищенной фракции сапонинов на круговой пластине (Kitagawa et al., 1988; Taniyama et al., 1988).
(iv) Хроматография на открытой колонке
Ряд классических систем растворителей, применяемых для колоночной хроматографии сапонинов на силикагеле, описан ранее и может быть найден, например, у Woitke et al., 1970, и Adler and Hiller, 1985. Хроматографию на открытой колонке часто применяют в качестве первой стадии фракционирования для сырой смеси сапонинов, но в определенных случаях она может давать чистые продукты. В целом все-таки разрешение не является высоким, и сложные смеси разделяются только частично. Другими проблемами являются потеря материала из-за необратимой адсорбции и продолжительность времени, требуемого для выполнения разделения.
Хроматография на силикагеле с элюентами хлороформ-метанол-вода является одним из наиболее широко распространенных способов. Когда применяют двухфазную систему, фаза насыщенный водой хлороформ является элюентом. Таким образом, для первоначального разделения метанольного экстракта растительной ткани на силикагеле может быть применен градиент смеси хлороформ-метанол-вода (например, 65:35:5 → 65:40:10). Далее может быть применена хроматография на колонках низкого давления с выходом, например, монодесмозидного моллюскицидного сапонина, в то время как бидесмозидный сапонин может быть получен путем хроматографирования на колонках силикагеля с такой системой растворителей как смесь ацетон-н-пропанол-вода (35:35:5) (Borel et al., 1987).
Сложная смесь тритерпеновых гликозидов была выделена из клубнелуковиц Crocosmia crocosmiiflora (Iridaceae). Три из них, гликозиды 2,9,16-тригидроксипальмитиновой кислоты, полигалациновой кислоты, были получены с помощью стратегии, включая хроматографию на открытой колонке силикагеля 60 (60-230 мкм) сырой смеси сапонинов с применением смеси н-бутанол-этанол-вода (5:1:4, верхний слой) и смеси хлороформ-метанол-вода (60:29:6) в качестве элюентов. Окончательная очистка осуществлялась с помощью ВЭЖХ (Asada et al., 1989).
Широкое использование хроматографии на силикагеле позволяет также разделить даммарановые гликозидные актинстеммозиды A-D из Actinostemma lobatum (Cucurbitaceae). После колонки MCI (Mitsubishi Chemical Industries) с полистироловым гелем требуемые фракции хроматографировали с различными растворителями: смесями хлороформ-метанол-вода (7:3:0,5, 32:8:1), хлороформ-метанол (9:1, 1:1), хлороформ-этанол (17:3), этилацетат-метанол (4:1) и хлороформ-метанол-этилацетат-вода (3:3:4:1,5, нижний слой). С помощью данных средств был получен чистый актиностеммозид С, в то время как актиностеммозиды А и В требовали дополнительной стадии ЖХ низкого давления, а актиностеммозид D требовал окончательного разделения на колонке С-18 с элюированием 70% метанолом (Iwamoto et al., 1987).
Определенные сложные эфиры сапонинов хроматографировали на силикагеле, пропитанном 2% борной кислотой (Srivastava and Kulshreshtha, 1986; 1988).
В качестве дополнения к нормальному силикагелю в хроматографии сапонинов на открытых колонках сейчас применяются грубые сорбенты RP (ОФ). Поскольку гранулометрия не является слишком хорошей и колонки не слишком длинными, вполне подходят колонки c подачей самотёком. ОФ-хроматографию обычно вводят после первоначальной стадии разделения на силикагеле, и она позволяет достичь изменений селективности для веществ, предназначаемых для разделения. Другой возможностью является введение разделения с обращенной фазой после стадии DCCC (Higuchi et al., 1988).
1. Хроматография на открытой колонке с полимерными сорбентами
Применение декстрановых подложек, как найдено для упаковки колонок сефадекса, являлось постоянной практикой в течение ряда лет. Сефадекс LH-20 находит наиболее частое применение в течение ряда лет, но серия "G" полимеров небезынтересна.
В недавней работе по выделению сапонинов была разработана новая генерация полимеров, особенно в Японии. Diaion HP-20 (Mitsubishi Chemical Industries, Tokyo), например, является высокопористым полимером, который широко используется для начальных стадий очистки.
Обычно полимерные подложки промывают водой после нанесения образца, для элюирования моносахаридов, малых заряженных молекул, таких как аминокислоты, и других высоко растворимых в воде веществ. Затем начинается элюирование градиентом метанол-вода (или одним метанолом) для получения фракций сапонинов. Применяют другие хроматографические процедуры для выделения чистых сапонинов.
Описано также элюирование гелей HP-20 смесями ацетон-вода. Например, при выделении бидесмозидных гликозидов кислоты мыльного дерева из клубней Thladiantha dubia (Cucurbitaceae), метанольные экстракты пропускали через колонку Diaion CHP-20P и промывали водой. Сырой экстракт сапонинов элюировали 40% ацетоном. Дополнительное разделение включало в себя хроматографию на силикагеле (смесь этилацетат-метанол-вода 6:2:1) и ВЭЖХ (Nagao et al., 1990).
Для выделения фибринолитических сапонинов из семян Luffa cylindrica (Cucurbitaceae) водный экстракт хроматографировали на колонке Амберлита XAD-2, элюируя метанолом, затем на второй колонке XAD-2, элюируя 40-70% метанолом. Активные составные части получали в чистом виде после хроматографии на колонках силикагеля, элюируя смесью хлороформ-метанол-вода (65:35:10, нижний слой → 65:40:10) (Yoshikawa et al., 1991).
(v) Жидкостная хроматография при среднем давлении (MPLC)
Когда требуются относительно большие количества чистых сапонинов, очень выгодна MPLC. В отличие от имеющегося в продаже оборудования для LPLC граммовые количества образца могут быть нанесены на колонки, в то время как разделение происходит при давлении до 40 бар. Гранулометрия подложки обычно лежит в диапазоне 25-40 мкм, и разделение проходит быстро, требуя значительно меньше времени, чем хроматография на открытой колонке. Прямой перенос условий разделения от аналитической ВЭЖХ к MPLC может быть достигнут на подложках с обратной фазой, ускоряя таким образом выбор растворителя (Hostettmann et al., 1986).
В качестве примера, моллюскицидные сапонины из Cussonia spicata (Araliaceae) были получены в достаточных количествах для биологического тестирования с помощью MPLC на сорбенте С-8 с применением смеси метанол-вода (2:1) (Gunzinger et al., 1986). Действительно, данный способ требовал только двух стадий (одна на силикагельной подложке и вторая на материале RP) для выделения сапонинов из бутанольного экстракта коры ствола.
Выделение сапонинов также может быть достигнуто с помощью сочетания MPLC, например, с применением колонки LiChroprep RP-8 (25-40 мкм, 46 х 2,6 см) с использованием смесей метанол-вода в сочетании с ротационной камерной противоточной хроматографией (RLCC) (Dorsaz and Hostettmann, 1986). В другом способе MPLC применяются сжатые по оси колонки (Jobin-Yvon) (Elias et al., 1991).
Примеры сочетаний подложка-растворитель, которые пригодны для разделения тритерпенов из растительных экстрактов, даны в таблице 1, ниже.
Применение MPLC в разделении тритерпеновых сапонинов
25:25:2 и 70:30:3)
(vi) Высокоэффективная жидкостная хроматография (ВЭЖХ)
Хроматография в виде ВЭЖХ является прекрасным способом получения мультимиллиграммовых количеств сапонинов из смесей близкородственных соединений и в данном случае очень часто применяется в качестве конечной стадии очистки. В то время как при MPLC используются более крупные частицы (25-100 мкм), в сорбентах при полупрепаративной ВЭЖХ гранулометрический диапазон лежит в интервале 5-30 мкм и, следовательно, позволяет достичь более высокой эффективности разделения.
Полупрепаративная ВЭЖХ применялась для разделения тригликозида олеаноловой кислоты из продуктов их частичного гидролиза. Это было необходимо для того, чтобы определить, будет ли галактозная часть присоединяться в положении С-3 или С-4 глюкозного остатка. Выделение изомерных сапонинов осуществлялось на 7 мкм - колонке LiChrosorb RP-8 (250 х 16 мин) с элюированием смесью ацетонитрил-вода (38:62) при скорости прохождения 10 мл/мин. Определение осуществляли при 206 нм и от 50 мг смеси (Decosterd et al., 1987).
Широкомасштабное разделение саикосапонинов а, с и d из корней Bupleurum falcatum (Umbelliferae) было достигнуто на сжатых по оси колонках, размеры 100 х 11 см I.D. Предварительная очистка метанольного экстракта выполнялась с помощью распределения между растворителями и хроматографии на полимере HP-20. Препаративную колонку ВЭЖХ упаковывали силикагелем С-18 (размер частиц 20 мкм; 5 кг) и элюировали при скорости прохождения 210 мл/мин со ступенчатым градиентом ацетонитрила. Нагрузки в 10 г было достаточно для получения 400 мг саикосапонина с, 1200 мг саикосапонина а и 1600 мг саикосапонина d (Sakuma and Motomura, 1987).
Гинзенозиды выделяли из Panax trifolius (Araliaceae) c помощью двухступенчатой процедуры, включающей в себя хроматографию на системе Waters Prep 500 (радиально сжатые колонки) с тремя серийно расположенными силикагельными картриджами (300 х 57 мин). Элюент представлял собой верхнюю фазу смеси н-бутанол-этилацетат-вода (4:1:5) и вводили нагрузку в 4 г. Полупрепаративную ВЭЖХ на колонке углеводов (Waters, 300 x 7,8 мм) с элюированием смесью ацетонитрил-вода (86:14 или 80:20) при скорости прохождения 2 мл/мин применяли для окончательной очистки (Lee and der Marderosian, 1988).
Единственной самой большой трудностью в определении компонентов элюирования при ВЭЖХ является отсутствие подходящего хромофора для УФ-детекции большинства сапонинов, хотя обычно это может быть преодолено с помощью применения способов, включающих в себя определение индекса преломления, определения массы и дериватизации.
Однако в предположении, что изменения градиента являются небольшими, обычно может применяться УФ-детекция в районе 203-210 нм с подходящими чистыми растворителями. Успешное разделение также осуществлялось с применением градиентов ацетонитрил-вода с УФ-детекцией. Ацетонитрил предпочтительнее метанола при низких длинах волн из-за его более низкого УФ-поглощения. Если различие в полярности не является слишком большим в тестируемых сериях сапонинов (например, только небольшие изменения в цепи сахаров), возможно изократическое элюирование.
Пригодный метод для разделения смесей сапонинов включает в себя разделение на октил-связанной колонке с применением градиентного элюирования водным ацетонитрилом. Количество ацетонитрила увеличивается от 30% до 40% в течение 20 мин, давая относительно небольшое базальное смещение при УФ-поглощении. Более полярные бидесмозидные сапонины обычно элюируют намного быстрее, чем монодесмозидные сапонины, и глюкурониды задерживаются меньше других гликозидов. Неполярная октилсилильная подложка может быть использована для отбора липофильной части сапонинов. При применении данного способа гликозиды гедерагенина элюировались перед теми же гликозидами менее полярной олеаноловой кислоты (Domon et al., 1984).
1. Применение дериватизированных тритерпенов
Определение при низких длинах волн, которое ведет к проблемам нестабильности базального поглощения, обусловленного интерференцией следовых количеств сильно поглощающего УФ-материала, может быть улучшено с помощью ВЭЖХ-анализа дериватизированных тритерпенов. Одной возможностью является присоединение функциональных групп к свободным карбоксильным группам, найденным в сапонине, как сообщалось для количественного определения монодесмозидных сапонинов. Обработка гликозидов олеаноловой кислоты 4-бромофенацилбромидом в присутствии бикарбоната калия и кронового простого эфира ведет к образованию бромофенацильных производных. 4-Бромофенацильные производные сильно поглощают при 254 нм, и определение может быть осуществлено при данной длине волны без интерференции растворителя (Slacanin et al., 1988). Дериватизация осуществляется, как показано ниже.

Альтернативный способ определения заключается в получении флуоресцентных кумариновых производных с помощью этерификации части карбоновой кислоты. С помощью данного способа были проанализированы сапонины сои и количественно определены в различных сортах и различных органах соевых бобов с антраценом в качестве внутреннего стандарта (Kitagawa et al., 1984a; Tani et al., 1985).
2. Очистка образцов
Для того чтобы удалить интерферирующий материал, который часто является сильно поглощающим УФ, может быть необходима стадия предварительной очистки. Это может быть достигнуто, например, с помощью применения картриджей Sep-PakR С18 (Guedon et al., 1989) или ExtrelutR (Sollorz, 1985).
В случае ионных соединений, таких как те, которые содержат свободную гидроксильную группу на агликоне или части глюкуроновой кислоты, требуется некий метод подавления образования ионов, если планируется избежать расширения пика. Это может быть достигнуто путем добавления кислоты с низким поглощением УФ к подвижной фазе, такой как фосфорная кислота или трифторуксусная кислота. Другой возможностью является применение ВЭЖХ пар ионов с противоионом, добавляемым к подвижной фазе. Фактор емкости ионных соединений увеличивается в результате образования комплексов ионов с сопрягающим реагентом. Дериватизация карбоксильных групп (как указывалось выше) является альтернативной по отношению к добавкам в подвижную фазу, приводя к существенному увеличению разрешения пиков.
Преимущество количественной ВЭЖХ по сравнению с фотометрическими способами заключается в том, что может быть измерено количество индивидуальных сапонинов в смеси или экстракте. Во многих случаях ВЭЖХ дает лучшие результаты по сравнению с получаемыми колориметрическим, газохроматографическим и ТСХ-флуориметрическим способами.
В тех случаях, когда разделение пиков смесей сапонинов на колонках ВЭЖХ с обращенной фазой является недостаточным, может быть применен ряд других способов, включая использование гидроксилапатитных колонок, колонок с химически модифицированным пористым стеклом, колонок силикагеля и ВЭЖХ боратных комплексов.
3. Гидроксилапатит
Гидроксилапатит (Ca10(PO4)6(OH)2) является более гидрофильным по сравнению с силикагелем и может быть применен с простыми бинарными водными системами растворителей, в результате чего облегчается определение с помощью УФ. Он стабилен в нейтральной и щелочной средах. Недавно были получены тяжелые сферические частицы гидроксилапатита, которые устойчивы к высокому давлению (до 150 кг/см2), что расширяет применение ВЭЖХ. Сапонины, которые различаются лишь по концевой пентозной единице и которые не могут быть разделены с помощью ОФ-ВЭЖХ, могут быть разделены с применением данного способа (Kasei et al., 1987b). Разделение гинсенозидов из Panax ginseng (Araliaceae) было достигнуто изократическим способом (смесь ацетонитрил-вода, 80:20) или лучше с помощью линейного градиента (смесь ацетонитрил-вода 70:30→90:10) (Kasei et al., 1987b). Как это наблюдалось для силикагеля, гликозиды элюируются в порядке увеличения полярности, т.е. обратно наблюдаемому при ОФ-ВЭЖХ.
4. Боратная ионообменная ВЭЖХ
Данный способ нашел применение в анализе моно- и олигосахаридов. Наилучшие результаты данным способом получают с применением анионообменной колонки, например, Asahipak ES-502NTM, 100 х 7,6 мин колонка от Asahi Kasei Kogyo Co. с 0,4 М H3BO3 в 20% (об/об) ацетонитриле (рН 8) при 75°С. Хроматографические характеристики зависят от образования боратных комплексов с цис-диолами сахаридной части. После разделения борат может быть удален в виде летучего метилбората путем повторной совместной перегонки элюата с метанолом.
5. Химически модифицированное пористое стекло
Микропористое стекло (MPG) обладает высокой химической стойкостью и стабильно при рН между 2 и 12. Октадецильное пористое стекло (MPG-ODS) было получено в виде набивки для ВЭЖХ с обращенной фазой и применено для быстрого и эффективного разделения сапонинов. Например, возможно разделить и гинсенозиды, и саикосапонины одновременно из экстрактов сочетанных лекарств, содержащих корень женьшеня и bupleurum с применением для разделения смеси ацетонитрил-вода (25,5:74,5) (Kanazawa et al., 1990a). Сравнение колонок MPG-ODS и оксида кремния-ODS для ВЭЖХ экстракта женьшеня и смесей гинсенозидов показало, что характер задержки сходен, но показатели емкости были ниже для колонки MPG-ODS. Разрешение некоторых пар гинсенозидов было лучше на колонках MPG-ODS (Kanazawa et al., 1993).
6. Силикагель
Применение водосодержащих подвижных фаз является часто неизбежным при разделении сапонинов, и силикагелевая ВЭЖХ не может быть обычно применена сама по себе с такими элюентами. Однако модификация набивки колонки дала возможность разделения водорастворимых гликозидов без ухудшения колонки. Способ включает в себя первую промывку колонки метанолом, затем смесью хлороформ-метанол-этанол-вода (62:16:16:6) и, наконец, системой растворителей, применяемой для разделения (Kaizuka and Takahashi, 1983). С применением, например, 5 мкм силикагелевых колонок с водосодержащим элюентом: смесью гексан-этанол-вода (8:2:0,5), можно было достичь эффективного анализа сапонинов женьшеня и саикосапонинов из Bupleurum falcatum.
(vii) Другие хроматографические способы
Для выделения чистых сапонинов требуются одна или, что более типично, более одной стадии хроматографического разделения для удаления других полярных составляющих спиртовых или водных растительных экстрактов.
Множество способов разделения было описано и может быть применено для разделения тритерпеновых сапонинов, включая моментальную хроматографию, DCCC, жидкостную хроматографию низкого давления (LPLC), жидкостную хроматографию среднего давления (MPLC), ВЭЖХ и традиционную хроматографию с открытой колонкой (см., например, Hostettmann et al., 1986, 1991; Marston and Hostettmann, 1991b). Смысл условий разделения систем растворителей и т.д. должен быть известен специалистам в данной области техники в свете настоящего раскрытия. Наилучшие результаты обычно достигаются с помощью стратегий, в которых используется сочетание способов, таких как конкретно раскрытые здесь ниже.
Поскольку некоторые из сапонинов являются кислыми, могут образовываться соли и по завершении хроматографии может иметься необходимость обработки ионообменной смолой для получения свободного сапонина. Примеры подходящих смол включают в себя Dowex 50Wx8 (H3 форма) (Kitagawa et al., 1988; Yoshikawa et al., 1991), Amberlite IRC 84 (Okabe et al., 1989; Nagao et al., 1990) и Amberlite MB-3 (Mizutani et al., 1984). Однако, если для предотвращения распада необходимы нейтральная среда или тщательный контроль рН, следует исключить этапы, включающие в себя фильтрацию на ионообменных смолах.
В определенных случаях сырые сапониновые фракции были метилированы (принимая, что присутствуют свободные COOH-группы) для достижения удовлетворительного разделения близкородственных продуктов (Okabe et al., 1989; Nagao et al., 1989, 1990).
1. Моментальная хроматография
Моментальная хроматография является способом препаративной жидкостной хроматографии под давлением, который обеспечивает значительное сокращение времени по сравнению с традиционной хроматографией на открытой колонке. Применяют обычные стеклянные колонки, но элюент продавливается через сорбент под действием сжатого воздуха или азота с достижением максимального давления, равного приблизительно 2 бар на вершине колонки. Гранулометрия сорбента в некоторой степени снижена из-за того, что растворитель доставляется под давлением; разрешение, следовательно, выше.
Моментальная хроматография может быть применена в качестве быстрой альтернативы хроматографическим способам предварительного фракционирования на открытой колонке. С применением данного способа можно достичь разделения образца от 10 мг до 10 г в течение такого короткого промежутка времени, как 10 мин. Например, моллюскицидные и фунгицидные хедерагениновый, байогениновый и медикагениновый гликозиды из корней Dolichos kilimandscharicus (Leguminocae) были выделены с помощью данного способа. Метанольный экстракт (3,3 г) фракционировали на силикагеле (63-200 мкм гранулометрии) на колонке 60 х 4 см с системой растворителей хлороформ-метанол-вода (50:10:1) при скорости тока 15 мл/мин. Этого было достаточно для удаления загрязняющего материала и получения двух обогащенных сапонинами фракций. Чистые тритерпеновые гликозиды были получены сочетанием DCCC и LPLC на подложках С-8 (Marston et al., 1988a).
Хотя в большинстве применений использовались силикагелевые сорбенты, наблюдается усиливающаяся тенденция в сторону ОФ-материалов. Моментальная хроматография ОФ обеспечивает отделение сапонинов от других более полярных компонентов, таких как олигосахариды.
2. Жидкостная хроматография низкого давления (LPLC)
LPLC пригодна для выделения чистых сапонинов, благодаря скорости разделения и простоте выполнения. Для LPLC применяют колонки, содержащие сорбенты с размером частиц 40-60 мкм. Возможна высокая скорость потока при давлении до 10 бар, и колонки чаще всего делают из стекла. Имеющиеся в продаже предварительно упакованные колонки (например, ряд "Lobar" от Merck) разного размера являются идеальными для препаративной хроматографии сапонинов в диапазоне образцов 50-500 мг. Высокая и равномерная плотность упаковки гарантирует хорошую эффективность разделения. Более того, сравнительно просто перенести условия аналитической ВЭЖХ на разделение LPLC, принимая во внимание то, что химия сорбентов является сходной (Marston and Hostettmann, 1991b).
Наибольшее количество применений было осуществлено на сорбентах ОФ, элюируемых смесями метанол-вода. Обычно в данном случае наносят лишь предварительно очищенные образцы. Хорошей иллюстрацией LPLC является разделение моллюскицидных и гемолитических гликозидов олеаноловой кислоты и гипсогениновых гликозидов из Swartzia madagascariensis (Leguminocae). Высушенные измельченные плодовые стручки экстрагировали водой и данный экстракт подвергали распределению между н-бутанолом и водой. После хроматографии органической фазы на открытой колонке сапонины разделяли на колонке Lobar LiChroprep C-8 (40-63 pro; 27 x 2,5 см) с применением смеси метанол-вода (75:25) в качестве элюента (Borel and Hostettmann, 1987).
Соединение колонок LPLC в серии позволяет повысить нагрузочную емкость и/или эффективность разделения. Данный подход был применен при разделении даммарановых гликозидов из Actinostemma lobata (Cucurbitaceae), при котором были соединены три колонки Lobar 27 x 2,5 см. Элюент также содержал небольшое количество воды (этилацетат-н-пропанол-вода 20:3:0,3) (Iwamoto et al., 1987).
3. Противоточная хроматография
Способы распределения между жидкостями оказались идеальными для применения в области сапонинов. Очень полярные сапонины особенно хорошо приспособлены для противоточного хроматографического разделения, особенно - поскольку отсутствует потеря вещества за счет необратимой адсорбции на материалах для забивки. Данный аспект был особенно применим для прямого фракционирования сырых экстрактов.
4. Капельная противоточная хроматография (DCCC)
DCCC относится к непрерывному прохождению капель подвижной фазы через несмешивающуюся жидкую стационарную фазу, содержащуюся в большом количестве вертикальных стеклянных трубок. Растворенное вещество подвергается непрерывному распределению между двумя фазами. В зависимости от того, вносится ли подвижная фаза на вершину или основание данных трубок, хроматография является "нисходящего" или "восходящего" типа, соответственно. Было возможным разделение близкородственных сапонинов с помощью DCCC и даже выделение чистых продуктов (Hostettmann et al., 1984). Действительно, данным способом достигались некоторые разделения, которые не были возможны с применением хроматографии жидкость-твердое вещество. С помощью DCCC можно было разделить изомерные сапонины, различающиеся лишь по положениям заместителя ацетатных групп на остатках сахаров (Ishii et al., 1984).
Для разделения сапонинов с помощью DCCC применяли ряд систем растворителей (см., например, Hostettmann et al., 1986) и из них в наибольшем количестве применений использовалась система хлороформ-метанол-вода (7:13:8). Системы хлороформ-метанол-вода могут быть применены либо в восходящем способе для очень полярных сапонинов, либо в нисходящем способе для сапонинов, содержащих один или два сахара и небольшое количество свободных гидроксильных групп.
Был описан крупномасштабный DCCC способ предварительной очистки с применением колонок 18 (30 см х 10 мм, внутр. диам.) с насыщенной н-бутанолом водой в качестве стационарной фазы и насыщенным водой н-бутанолом в качестве подвижной фазы (Komori et al., 1983). В некоторых случаях для получения сапонинов производят два (или более) DCCC разделения.
5. Центрифужная распределительная хроматография (CPC)
Недавно введенный в практику способ CPC является многообещающим благодаря его скорости и многосторонности (Marston et al., 1990). CPC относится к полю центрифугирования, образуемого путем вращения при 800-2000 об/мин или быстрее, а не к полю гравитации для удержания стационарной фазы. Принцип способа включает в себя непрерывный процесс неравновесного распределения растворенного вещества между двумя несмешивающимися фазами, содержащимися во вращающихся кольцах или картриджах.
Аппараты на основе вращающихся колец могут включать в себя либо планетарное, либо непланетарное движение вокруг центральной оси. Один из них, высокоскоростной противоточный хроматограф (HSCCC), состоит из тефлоновой трубки с внутренним диаметром 1,6 или 2,6 мм, обернутой в виде кольца вокруг бобины. Одна, две или три бобины составляют сердцевину аппарата. В случае картриджных аппаратов картриджи расположены по периметру центрифужного ротора с длинными осями, параллельными направлению силы центрифугирования. Количество и объем картриджей может варьировать в зависимости от применения, для которого установлен аппарат. По сравнению с DCCC и RLCC, при которых разделение может занимать 2 дня или более, CPC может дать те же результаты в течение часов. Аппараты на основе вращающихся колец или картриджей обладают емкостями до граммового масштаба. Многослойный спиральный планетный аппарат был применен, например, для предварительной очистки циклоартановых гликозидов из Abrus fruticulosus (Leguminosae) (Fullas et al., 1990). Моллюскицидные тритерпеновые гликозиды из Hedera helix (Araliaceae) были отделены на другом аппарате, хроматографе Sanki LLN (шесть картриджей; общий объем 125 мл). Метанольный экстракт фрукта подвергали распределению между н-бутанолом и водой. Бутанольную фракцию прямо инъецировали на аппарат в количествах по 100 мг с применением нижнего слоя системы растворителей хлороформ-метанол-вода (7:13:8) в качестве подвижной фазы.
Два главных сапонина, асиатикозид и мадекассозид, из Centella asiatica (Umbelliferae) были разделены с помощью многослойного кольцевого сепаратора-экстрактора Ito (P.C. Inc.), снабженного колонкой с внутренним диаметром 66 м х 2,6 мм (емкость 350 мл) с вращением при 800 об/мин. Образец в количестве 400 мг мог быть разделен с помощью системы растворителей хлороформ-метанол-2-бутанол-вода (7:6:3:4; подвижной фазой была нижняя фаза). Определение осуществляли с помощью включенной в систему ТСХ (Diallo et al., 1991). Тот же аппарат применяли при выделении тритерпенового дисахарида из Sesamum alatum (Pedaliaceae). Нижняя фаза растворителя хлороформ-метанол-изопропанол-вода (5:6:1:4) была выбрана в качестве подвижной фазы, и инъецировали наносимый материал в количестве 1,25 г (Potterat et al., 1992).
6. Сочетание способов
Для выделения чистого сапонина из экстракта редко бывает достаточно одной хроматографической стадии. Общим правилом является необходимость серий из нескольких препаративных способов для получения необходимого продукта. Для разделения многих сапонинов была показана пригодность сочетания классических способов (таких как хроматография на открытой колонке) и современных способов с высоким разрешением (таких как ВЭЖХ).
Например, применяют сочетание MPLC на силикагеле и ОФ-материала, LPLC и центрифужной ТСХ для разделения сапонинов (Hamburger and Hostettmann, 1986). Сходным образом для выделения пяти тритерпеновых сапонинов из Swartzia madagascariensis (Leguminosae) потребовались хроматография с открытой колонкой, LPLC и MPLC (Borel and Hostettmann, 1987).
CPC применяли в сочетании с моментальной хроматографией и OPLC для выделения тритерпеновых гликозидов из Abrus fruticulosus (Leguminosae). Многослойный спиральный аппарат (растворитель хлороформ-метанол-вода (7:13:8), нижняя фаза в качестве подвижной фазы) обеспечил исходную очистку, тогда как моментальная хроматография и OPLC были эффективными для получения чистых веществ (Fullas et al., 1990).
Прямое сочетание моментальной хроматографии на немодифицированном силикагеле с либо моментальной хроматографией, либо хроматографией на открытой колонке с веществом RP иногда может быть достаточным для очистки сапонинов (Schopke et al., 1991).
Другая стратегия включает в себя пропускание экстрактов (после предварительного распределения) через высокопористые полимеры и сопровождающее данную стадию дальнейшее фракционирование сырых смесей сапонинов. Данный подход был применен для выделения гликозидов 3β-гидроксиолеан-12-ен-28,29-диоевой кислоты из Nothopanax delavayi (Araliaceae). Метанольный экстракт листьев и стеблей подвергали распределению между гексаном и водой. Водный слой хроматографировали на колонке Diaion HP-20 и элюировали водой, 10% метанолом, 50% метанолом, 80% метанолом, метанолом и хлороформом. Гликозиды получали последующей колоночной хроматографией 80% метанольного элюата на силикагеле с помощью смеси этилацетат-этанол-вода (7:2:1) (Kasai et al., 1987a). Для выделения тритерпеновых и нетритерпеновых сапонинов из Acanthopanax senticosus (Araliaceae) процедуру начинали с фракционирования метанольного экстракта листьев на полимере Diaion HP-20. Фракцию, элюированную метанолом, хроматографировали на силикагеле (смесь хлороформ-метанол-вода, 30:10:1), и все полученные фракции подвергали колоночной хроматографии на LiChroprep RP-8. Конечная очистка была достигнута с помощью ВЭЖХ на TSK-GEL ODS-120T (300 х 21 мин; смесь метанол-вода, 70:30; 6 мл/мин; определение RI) или хроматографией на колонке гидроксилапатита (смесь ацетонитрил-вода 85:15) (Shao et al., 1988).
Процедура разделения гликозидов олеаноловой кислоты включает в себя применение сочетания сефадекса LH-20 (метанол), DCCC (смесь хлороформ-метанол-вода, 7:13:8) и ВЭЖХ (C-18, смесь метанол-вода, 65:35) (De Tommasi et al., 1991).
(viii) Цветные реакции
Для получения окрашенных соединений для количественного или качественного определения тритерпенов могут быть применены реакции тритерпенов с любым из множества агентов. Например, ароматические альдегиды, такие как эйсальдегид и ванилин в сильной неорганической кислоте, например серной, фосфорной и надхлорной кислотах, дают окрашенные продукты с агликонами, имеющие максимумы поглощения между 510 и 620 нм. Полагают, что в данных реакциях происходит дегидрирование с образованием ненасыщенных метиленовых групп, которые дают окрашенные продукты конденсации с альдегидами. С ванилин-серной кислотой тритерпеновые сапонины с С-23 гидроксильной группой имеют пик, расположенный между 460 и 485 нм (Hiai et al., 1976).
Ненасыщенные и гидроксилированные тритерпены и стероиды дают красное, синее или зеленое окрашивание с уксусным ангидридом и серной кислотой (Abisch and Reichstein, 1960). Поскольку терпеноидные сапонины склонны к образованию розового или пурпурного оттенка, а стероидные сапонины - сине-зеленого окрашивания, с помощью данного способа возможна дифференцировка двух классов.
Для детекции тритерпенов может быть применен большой ряд других агентов, включая сульфат церия(IV) или соли железа(III) и неорганические кислоты, такие как серная кислота, которая дает фиолетово-красное окрашивание раствора; 30% раствор хлорида антимония(III) в реагенте уксусный ангидрид-уксусная кислота, который дает цветные реакции с гидрокситритерпенами и гидроксистероидами; хлорид антимония(III) в смеси нитробензол-метанол, который может быть применен для дифференцировки 5,6-дегидропроизводных стероидных гликозидов (диосгениновых и соласодиновых гликозидов) и 5α- или 5β-H-производных (например, томатина), и карбазол, который в присутствии бората и концентрированной серной кислоты обычно указывает на наличие уроновых кислот (Bitter and Muir, 1962).
Иллюстративные реагенты для детекции и для спектрофотометрического и колориметрического определения сапонинов перечислены ниже в таблице 2.
Реагенты визуализации для тритерпеновых сапонинов
Wagner et al., 1984
(ix) Выделение монотерпеновых/тритерпеновых гликозидов из Acacia victoriae
Экстракты бобов получали с помощью экстракции смесью хлороформ:метанол или дихлорметан:хлороформ в университете Аризоны (Tucson, AZ). Были выделены смеси монотерпеновых/тритерпеновых гликозидов из Acacia victoriae (Benth.) (Leguminosae). Первый сбор фракции UA-BRF-004-DELEP-F001 был обработан следующим образом: (1) измельчение до частиц размером 3 мм в мельнице Wiley, (2) упаковка в двухлитровый агрегат для процеживания, (3) экстракция измельченной биомассы смесью дихлорметан:метанол (1:1) в течение 4 час и затем в течение ночи и сушка объединенных фракций в вакууме с получением UA-BRF-004-DELEP-F001 (52 г). F001 (51,5 г) экстрагировали этилацетатом с получением активного нерастворимого (34,7 г) вещества, обозначенного как F004. Мгновенную хроматографию с применением 1,7 кг силикагеля (Merck, размер частиц 23-220 микрон) применяли для фракционирования F004 (34,2), 51 670-мл фракции элюировали смесью дихлорметан:метанол (ступенчатый градиент 95-0% : метанол 5-100%). Колонку промывали девятью литрами метанола и затем шестью литрами смеси метанол:вода (80:20) и затем шестью литрами того же элюента с добавлением 1% муравьиной кислоты. На основе ТСХ фракции 23-34 и 39-40 объединяли, получая 17,2 г F023. Применяли жидкостную хроматографию среднего давления (MPLC, система Bushi 632) дважды с использованием каждый раз 8 г F023 на колонке 4,9 х 46 см, упакованной Lichroprep C-18, с размером частиц 15-25 микрон, с применением ступенчатого градиента смеси ацетонитрил:вода (0, 10, 20, 30, 50% ацетонитрила в воде) с последующей промывкой 100% метанолом. Из 16 г при применении 0-20% ацетонитрила получено семь грамм F027, которая была неактивна. Остальной материал объединяли и подвергали повторной MPLC с той же системой с применением 30-40% ацетонитрила для сведения к минимуму взаимного перекрывания и получения фракций F028-F036. Хотя большинство данных фракций демонстрировало противоопухолевую активность, для дальнейшего тестирования и оценки была выбрана F035 (фракция 35) (наибольший выход 2,19 г).
II. Определение структуры монотерпеновых/тритерпеновых композиций
Для качественного и количественного определения монотерпенов/тритерпенов и их активности могут быть применены различные способы, включая токсическую активность в отношении рыб, гравиметрию, спектрофотометрию, ТСХ, GC, ВЭЖХ, HMQC, HMBC, NOESY, COSY, ЯМР, рентгеноструктурную кристаллографию и т.д. Определения, основанные на классических свойствах тритерпеновых сапонинов (поверхностная активность, токсичность в отношении рыб), были преимущественно заменены фотометрическими способами, такими как денситометрия, колориметрия производных и, позднее, GC, ВЭЖХ и в особенности ЯМР. Спектрофотометрические способы очень чувствительны, но обычно не пригодны для определения тритерпенов в сырых растительных экстрактах, поскольку реакции не являются специфичными, и окрашенные продукты могут образовываться с соединениями, которые сопутствуют тритерпенам, такими как фитостеролы и флавоноиды. Другой проблемой, общей для большинства аналитических работ по сапонинам, является их неполная экстракция из растительного материала. Однако широко применяется ряд способов, которые пригодны для количественного определения тритерпенов.
Существует несколько основных проблем, которые должны быть разрешены при выяснении структуры сапонинов: структура истинного агликона; состав и последовательность составляющих моносахаридов в углеводной части; каким образом моносахаридные единицы соединены между собой; аномерная конфигурация каждой связанной гликозидной связью моносахаридной единицы и локализация углеводной части на агликоне.
Необходимый подход заключается в применении сочетания способов, с тем чтобы прийти к конечному заключению о структуре. Исследования структуры представляют собой обычно поэтапный процесс, в котором сапонин постепенно разрушают до более мелких фрагментов, которые сами по себе анализируются спектроскопически. С помощью логического построения данных для фрагментов создается концепция состава сапонина.
Количества чистых выделенных сапонинов часто малы, так что предпочтительно применение высокочувствительных с высоким разрешением и, если возможно, неразрушающих способов для того, чтобы обеспечить определение структуры сапонина. Инновации в области ЯМР-спектроскопии и масс-спектрометрии (MS) обеспечили такие необходимые возможности для исследований сложных сапонинов. Благодаря сочетаниям этих и других способов может быть осуществлено определение структуры. Например, FAB-MS дает информацию относительно молекулярной массы и во многих случаях последовательности сахаров, а способы I-D и 2-D ЯМР позволяют локализовать связи сахаров и вносят свой вклад в выяснение структуры агликона. Такие исследования определения структуры и химии были подробно обсуждены в обзоре Tanaka and Kasai (1984).
(i) Ядерный магнитный резонанс (ЯМР)
Из всех современных способов выяснения структуры олигосахаридов и гликозидов ЯМР-спектроскопия обеспечивает наиболее полную информацию при наличии или в отсутствие предварительного знания структуры (Agrawal, 1992). Это является единственным подходом, который может в принципе дать полную структуру без обращения к любому другому способу.
1. 13С-ядерный магнитный резонанс
Углерод-13-ЯМР-спектроскопия, в настоящее время широко применяемая для определения структуры сапонинов, является быстрым и неразрушающим способом, но требует весьма больших количеств образца (мг количества). Анализ спектров позволяет делать заключение о местах присоединения гликозидных цепей к агликону; последовательности, природе и количестве моносахаридов; конфигурации и конформации межгликозидных связей; присутствии ацилгликозидов в цепях; природе агликона и структурах, присоединенных эфиров кислот.
Для оценки химических сдвигов полезно сравнить наблюдаемые данные с данными, сообщавшимися для модели и родственных соединений. В качестве руководства в отношении некоторых типичных химических сдвигов в спектре 13С-ЯМР тритерпенового сапонина можно использовать известные сдвиги байогенинового гликозида (Domon and Hostettmann, 1984). Кроме того, были собраны данные о распределениях 13С-ЯМР-сигналов для олеананового (Patra et al., 1981; Agrawal and Jain, 1992), урсанового, люпанового (Wenkert et al., 1978; Sholichin et al., 1980), хопанового (Wenkert et al., 1978; Wilkins et al., 1987) и ланостанового (Parrilli et al., 1979) тритерпенов (Nakanishi et al., 1983). Близкие данные для даммарановых гликозидов были собраны в обзоре (Tanaka and Kasai, 1984), и была описана 13С-ЯМР-спектроскопия сайкогенинов (Tori et al., 1976a) и сайкосапонинов (Tori et al., 1976b). Были также исследованы сапогенины женьшеня и родственные даммарановые тритерпены (Asakawa et al., 1977). Была также описана 13С-ЯМР-спектроскопия акациевой кислоты (Kinjo et al., 1992).
При 13С-ЯМР, когда гидроксильные группы дериватизированы, т.е. гликозилированы, метилированы (или ацетилированы), α- и β-углероды и сахарной, и агликоновой частей подвергаются характерным сдвигам. Например, сигналы α-CH сдвинуты в нижележащую область, а сигналы β-С сдвинуты в вышележащую область, сдвиг, возникающий из общего γ-сдвига в вышележащую область). Таким образом, гликозилирование агликона вызывает сдвиг в нижележащую область α-углерода и сдвиг в вышележащую область (сдвиги гликозидирования) соседних углеродных атомов (Tory et al., 1976b; Kasai et al., 1977). В олеананах гликозидирование 3β-OH группы вызывает сдвиг С-3 в нижележащую область на приблизительно 8,0-11,5 м.д., сдвиг С-2 и С-4 на +0,9 или от -0,9 до -1,9 м.д., сдвиг С-23 в вышележащую область на 0,5-5,1 м.д. и сдвиг С-24 на от -0,2 до 1,6 м.д. Гликозидирование 28-COOH группы вызывает смещение резонанса карбоксильного углерода в вышележащую область (2,5-5,0 м.д.) и смещение С-17 сигнала в нижележащую область (1,0-2,5 м.д.) (Agrawal and Jain, 1992). Таким образом, сравнение данных 13С-ЯМР агликона и сапонина указывает на место присоединения сахара (Seo et al., 1978; Tanaka, 1985).
Сходным образом 13С-ЯМР должен давать указание (в более простых сапонинах на межгликозидные связи путем рассмотрения смещений химических сдвигов при сравнении с модельными соединениями (Konishi et al., 1978). Данные углерод-13-ЯМР для метил-β-D-фукопиранозида были сведены в таблицу Seo et al. (1978), а сигналы 13С-ЯМР для более сложных сахаров перечислены Gorin and Mazurek (1975) и Dorman and Roberts (1970). Апиоза дает характерные сигналы 13С-ЯМР, которые были документированы (Sakuma and Shoji, 1982; Adinolfi et al., 1987; Reznicek et al., 1990).
2. 1Н-Ядерный магнитный резонанс
Хотя 13С-ЯМР-спектральный анализ и распределение сигнала стало особенно применимой процедурой при определении структуры сапонинов, лишь редко сообщалось о полном распределении их 1Н-ЯМР-спектров. Спектры 1Н-ЯМР показали характерную сложность и трудность для анализа. Подавляющее большинство протоновых резонансов углеводной части обнаруживается в очень узком диапазоне спектра шириной 3,0-4,2 м.д. с вытекающими проблемами перекрытия. Это происходит из-за большого количества протонов неаномерных метина и метилена сахара, которые имеют очень близкие химические сдвиги в различных моносахаридных остатках.
Однако метильные пики тритерпенов легко различимы, и большинство позиций протонного резонанса в олеаненовом, урсеновом и сходных скелетах были определены, начиная с 1960-х годов (Kojima and Ogura, 1989) с помощью множества способов. Например, полные 1Н- и 13С-ЯМР спектральные распределения соясапогенола В (33) и конфигурация С-4 гидроксиметильного заместителя были установлены с помощью сочетания 13С-DEPT, 13C-APT, 2-D коррелированной спектроскопии (COSY) (1H-13C-COSY, 1H-1H COSY) и 1H-1H ROESY (2-D ядерное усиление Оверхаузера (NOE) во вращающейся рамке) способов (Baxter et al., 1990). Распределения резонансов четвертичного углерода в данном сапогенине были подтверждены с помощью определяемой по 1Н-гетероядерной множественной связи (HMBC) и одиночной связи (HMQC) спектроскопии (Massiot et al., 1991b). Была также достигнута полная интерпретация спектров 1Н-ЯМР диосгенина и соласодина (Puri et al., 1993).
Ряд полезных данных может быть получен из 1Н-ЯМР-спектров для аномерных конфигураций и связей сахарной цепи. Например, константа взаимодействия протона С-1 α-связанных единиц глюкозы составляет приблизительно 3 Гц, тогда как β-связанные единицы обладают константой взаимодействия 6-7 Гц. Более подробную информацию о константах взаимодействия аномерных протонов сахаров можно найти в других источниках (Lemieux et al., 1958; Capon and Thacker, 1964; Kizu and Tomimori, 1982).
Когда возникают сложности при определении конфигураций гидроксильных групп при С-2, С-3 и С-23, С-24 олеаненовых и урсеновых тритерпенов, анализ 1Н-ЯМР сигнальных пиков протонов на несущих кислород атомах углерода дает ценную информацию (Kojima and Ogura, 1989).
(ii) Способы 1-D и 2-D ЯМР
На практике некоторые спектры 1Н- и 13С-ЯМР могут быть идентифицированы и приписаны на основе аргументов сдвига, но для интерпретации результатов исследований ЯМР строгим образом спектр ЯМР должен быть оценен однозначно, что означает установление того, какие пики ассоциированы с каким углеродом и/или водородом в структуре. Данная информация в большинстве случаев не может быть получена на основе спектральных данных одномерных 1Н- и 13С-ЯМР, но могут быть лучшим образом определены с помощью двумерных исследований. Данные исследования упрощают спектральный анализ путем распределения информации в два частотных домена и выявления взаимодействий между ядрами. Несмотря на тот факт, что механизмы, на которых установлены различные пульсовые последовательности, могут быть сложными, интерпретация двумерных спектров ЯМР обычно является прямолинейной. Было проведено большое количество различных исследований с двумерным ЯМР для определения химических структур. Примеры таких способов, а также других способов ЯМР, конкретно рассматриваемых изобретателями для применения в выяснении химии тритерпеновых сапонинов согласно изобретению, описаны ниже и в таблице 3.
1. HMBC, HMQC
Применение HMQC и HMBC 13С-многоквантовых когерентных спектров ценно не только для установления агликона, но и для выяснения деталей сахарной последовательности. Применение HMBC и HMQC аналогично 13С-1Н гетероядерной коррелированной спектроскопии (HETCOR), но вместо наблюдения 13С определяют более обогащенный 1Н. Например, в случае беллиссапонинов из Bellis perennis (Asteraceae) было возможным приписать все химические сдвиги в спектре 1Н-ЯМР путем рассмотрения данных 13С-ЯМР в связи с 2-D 1Н-определенными HMQC и HMBC спектрами. Перекрестные пики, соответствующие двум и трем взаимодействиям связей, наблюдали для почти всех возможных корреляций в молекуле. Сходным образом 1Н-13С корреляции в длинном диапазоне в спектрах HMQC и HMBC могут быть применены для определения последовательности и положений присоединения сахарных частей (Schopke et al., 1991).
2. 2-D-NOESY
Данный способ был применен, например, при определении последовательности сахаров в гликозидах цикламиретина А (ардисиакриспинов А и В) (Jansakul et al., 1987) и моносахаридной последовательности саксифрагифолина А из Androsace saxifragifolia (Primulaceae) (Waltho et al., 1986). Положение рамнозильных и глюкозильных связей на арабинозной части сапонина С калопанакса было подтверждено с помощью NOESY после анализа последовательности сахара полностью метилированного сапонина. Перекрывающиеся пики наблюдались между Н-1 рамнозильной части и Н-2 арабинозильной части, а также между Н-1 глюкозильной части и Н-3 арабинозильной части (Shao et al., 1989b). Структуры сахарных частей фуростаноловых сапонинов из Balanites aegyptiaca (Balanitaceae) были выяснены с помощью 2-D NOESY на аппарате ЯМР с 400 Мгц (Kamel et al., 1991).
Совместное применение способов 2-D ЯМР привело к полному приписанию 13С и 1Н для олигосахаридного сегмента сарсасапогенинового гликозида 3-О-[{α-L-рамнопиранозил(1→4)}{β-D-глюкопиранозил(1→2)}-β-D-глюкопиранозил]-(25S)-5β-спиростан-3β-ола. Сочетание DEPT, HETCOR, длиннодиапазонного HETCOR, различных гомоядерных способов, NOESY и INEPT применяли для выяснения структуры при решении проблем, вызванных сверхтесным протонным спектром (Pant et al., 1988d).
Идентификация и определение последовательности сахаров в пентасахаридном сапонине 3-О-[β-D-ксилопиранозил(1→3)-α-L-арабинопиранозил(1→4)-β-D-глюкопиранозил(1→3)-α-L-рамнопиранозил(1→2)-α-L-арабинопиранозил]-хедерагенина из Blighia welwitschii (Sapindaceae) было возможно с применением только способов ЯМР с использованием аппарата на 500 Мгц. Сначала сапонин ацетилировали и последующий анализ DQF-COSY, NOESY и ROESY спектров позволил приписать структуру. Для установления последовательности сахаров была особенно полезна информация, полученная из данных NOE (Penders et al., 1989).
С помощью ЯМР, основанного на многоступенчатых исследованиях RCT, был идентифицирован сапонин, содержащий 10 остатков сахара, из Solidago gigantea (Asteraceae). Это включало в себя исследования COSY, гетероядерный COSY, H-H-C когерентный перенос COSY-типа и 2-D NOESY. Таким образом, удалось избежать исследований со значительной деградацией, и определение структуры было возможным с 30 мг продукта (Reznicek et al., 1989a; 1989b). Сходные способы были применены для определения структуры четырех других гликозидов, гигантеасапонинов 1-4 (бидесмозидов байогенина, содержащих девять или десять сахарных единиц), из того же растения (Reznicek et al., 1990а).
Сочетание ЯМР-исследований 2-D COSY, HMBC и ROESY было достаточным для получения последовательности и положений связей гексасахарида в мимонозиде А, сапонине олеаноловой кислоты из Mimosa tenuiflora (Leguminosae) (Jiang et al., 1991).
2-D ЯМР полностью ацетилированного и неизмененного хрисантеллина А обеспечило приписание протонов и определение последовательности сахаров. Было показано, что этерифицированная ксилозная часть находится в β-форме и имеет 1С4-конформацию. Среди использованных способов были HMQC, HMBC и ROESY (или точнее CAMELSPIN (перекрывающаяся релаксация, подходящая для минимолекул, эмулированная запертыми спинами)) на полностью ацетилированном производном и HOHAHA, TOCSY на нативном сапонине. Для определения последовательности сахаров особенно пригодным было исследование ROSEY (Massiot et al., 1991a).
Последовательности сахаров и межгликозидные связи тритерпеновых гликозидов из морских организмов были установлены на основе данных NT1 и исследований NOESY (Miyamoto et al., 1990), но данная методология ограничивается сложностью спектров 1Н-ЯМР в области 3-5 м.д., которая обычно препятствует измерению NOE для большого количества протонов. Однако сочетание COSY, NOESY и прямой и XHCORR ЯМР спектроскопии обеспечило полное приписание сигналов и структурный анализ пентасахаридных тритерпеновых сапонинов из морского огурца Holothurea forskalii (Rodriguez et al., 1991).
При определении структуры сантиагозида, астеросапонина из антарктической морской звезды Neosmilaser georgianus, активно применяли способы COSY, TOCSY, HMQC и ROESY ЯМР спектроскопии, причем для выяснения точной последовательности сахаров, точек их присоединения и стереохимии применяли исследования ROESY (Vazquez et al., 1992).
3. COSY
Существует два основных типа 2-D ЯМР спектроскопии: спектроскопия с J-разрешением, при которой одна частотная ось содержит спаривание спинов (J) и другую информацию о химических сдвигах, и коррелированная спектроскопия, при которой обе частотные оси содержат информацию о химическом сдвиге (δ) (Agrawal, 1992). Одно из главных преимуществ 2-D анализа заключается в том, что он обеспечивает способ преодоления проблемы тесноты спектра. В 1H-COSY с сильным полем это особенно верно для области 2,5-4,0 м.д., что тем самым упрощает приписание сахаридных протонов. При благоприятных условиях могут быть идентифицированы все протоны, имеющиеся в данном сахарном остатке.
На основе спектров COSY может быть сделано несколько общих заключений. Например, положения замещения моносахаридных единиц могут быть определены по наличию или отсутствию соответствующего гидроксильного протона; размеры колец моносахаридов могут быть определены прямо, и природа перекрывающихся пиков выявляет множественность перекрывающихся пиков, что обеспечивает установление констант взаимодействия.
В определенных случаях выяснение структуры сапонина вместе с его сахарной последовательностью было достигнуто с помощью лишь 1Н-ЯМР 1-D- и 2-D-спектроскопии (Massiot et al., 1986; 1988b). Сначала сапонин полностью ацетилируют и, если напряженность поля достаточна (>300 Мгц), протонные резонансы распадаются на две зоны, одна между 4,75 и 5,40 м.д., приписываемая CHOAc, и другая между 3,0 и 4,3 м.д., приписываемая CH2OAc, CHOR и CH2OR. Аномерные протоны расположены между данными двумя зонами в случае простых эфирных связей или при большей частоте, чем 5,5 м.д., для сложноэфирных связей.
Полное ацетилирование также дает производные, которые растворимы в хлороформе, бензоле или ацетоне. В эквивалентных полностью дейтерированных растворителях подвижность молекул такова, что сигналы наблюдаются более четко, и константы взаимодействия могут быть измерены с высокой точностью. Для ацетилированного сапонина корня люцерны исследования COSY и COSY c длинным диапазоном были достаточными для идентификации структуры как Ara-2Glc-2Ara-3хедерагенин28-Glc (Massiot et al., 1986).
Структуры следующих полностью ацетилированных сапонинов из листьев люцерны, Medicago sativa (Leguminosae), и из Tridesmostemon claessenssi (Sapotaceae) были выяснены с помощью способов, сходных с указанными выше. Подтверждение приписаний и последовательностей сахаров было получено в исследованиях HMQC (для 1J взаимодействий) и HMBC (для 2J и 3J взаимодействий) и гомоядерного тройного родственного COSY Хартманна-Хана (HOHAHA) и ROESY (Massiot et al., 1990; 1991b). Сложноэфирные цепи сахаров сапонинов из T. claessenssi содержат β-D-ксилозную часть вне обычной 1С4-конфигурации (все заместители являются аксиальными). При 600 МГц 1Н-ЯМР спектр может быть разрешен достаточно хорошо, чтобы обеспечить приписание всех 1Н химических сдвигов без полного ацетилирования (Schopke et al., 1991).
4. COSY с длинным диапазоном
Данный способ применяли для приписания протонов сахаров в стероидных сапонинах из Allium vineale (Liliaceae) (Chen and Snyder, 1987; 1989). 1H-13С COSY с длинным диапазоном применяли для определения структуры агликона в сапонине циклоастрагенола (Wang et al., 1989b) и для нахождения 4J межсахарного взаимодействия между аномерным протоном внутренней глюкозы и Н-2 внутренней арабинозы сапонина Medicago sativa, описанного выше (Massiot et al., 1986).
5. Фазочувствительная COSY с двойной квантовой фильтрацией (DQF-COSY, DQ-COSY)
Данный способ применяли для приписания протонов сахаров в стероидных сапонинах Allium (Chen and Snyder, 1987; 1989) и для приписания 1Н химических сдвигов в гликозидах 16α-гидроксипрото-липовой кислоты из корней Crossopteryx febrifuga (Rubiaceae) (Gariboldi et al., 1990). Тот же способ применяли для обеспечения полного приписания протонов сахаридов в ацетилированном производном хедерагенина из плодов Sapindus rarak (Sapindaceae) (Hamburger et al., 1992). Межгликозидные связи устанавливали с помощью NOE дифференциальной спектроскопии (Hamburger et al., 1992).
6. HOHAHA
Сети протонного взаимодействия агликонов гипсогенинового гликозида и гликозида мыльного дерева были полностью выяснены с помощью исследований HOHAHA. Имеются сходные с COSY исследования (и близкие к общей коррелированной спектроскопии - TOCSY) за исключением того, что наблюдаемые корреляции между пиками находятся в фазе, что тем самым предотвращает случайное обнуление перекрывающихся пиков. Для раскрытия углеводных цепей соседние константы взаимодействия, полученные из исследований HOHAHA, обеспечивают определение относительной стереохимии каждого асимметрического центра, тем самым создавая возможность идентификации моносахаридов. Исследования, связанные с гетероядерным H - C, могут быть применены для приписания 13С резонансов в сахаридных частях и связей сахаров, определенных из спектров HMBC (Frechet et al., 1991).
7. FLOCK, COLOC и NOE
Гетероядерную коррелированную спектроскопию с длинным диапазоном, включающую в себя двухлинейные вращательные нарушающие взаимодействие импульсы (FLOCK), применяли для наблюдения взаимодействий 1Н-13С на дальнем расстоянии в алатозиде А из Sesamum alatum (Pedaliaceae). Таким образом, наблюдали взаимодействия между протоном при С-18 и атомами углерода С-13, С-17 и С-28. Вместе с гетероядерной 13С-1Н-корреляцией с длинным диапазоном (XHCORR) много информации было собрано относительно структуры нового секо-урсенового агликона (Potterat et al., 1992).
Пример 13С-1Н 2-D-коррелированной спектроскопии (COLOC), оптимизированной для взаимодействий с длинным диапазоном (2JCH и 3JCH), должен быть найден в выявлении структуры сапонинов из Crossopteryx febrifuga (Rubiaceae) (Gariboldi et al., 1990).
NOE нашло широкое применение в определении структуры сапонинов, например в приписании сахаридных протонов и установлении последовательностей сахаров люперозида I (Okabe et al., 1989) и камеллидинов I и II (Nishino et al., 1986). NOE между H-2 арабинозы и аномерным протоном рамнозы помог подтвердить Rha-2Ara-дисахаридную связь в зизифине (Yoshikawa et al., 1991b). Способ имеет широкое применение, поскольку между аномерным протоном и агликоновым протоном часто наблюдаются соединения в положении связи. Негативное NOE наблюдалось между протоном в положении С-3 и аномерным протоном 3-О-гликозидного остатка в циклоастрагеноле и других сапонинах (Wange et al., 1989b).
Выбранные подходы ЯМР для применения в установлении структуры тритерпеновых сапонинов
(iii) Спектроскопические и другие способы для выяснения структуры
Выяснение структуры сапонинов и соответствующих агликонов основано не только на химических методах, но также на спектроскопических и сходных с ними способах, например ИК, УФ, ЯМР, MS, дисперсия оптического вращения (ORD), круговой дихроизм (КД) и рентгеновский анализ. Современные достижения в отношении некоторых из данных способов, наиболее заметные в случае ЯМР-спектроскопии и MS, ускорили решение задачи анализа сапонинов и их соответствующих фрагментов, получаемых при реакции расщепления, так что информация может сравниваться и могут определяться относящиеся к этому структуры. Более того, ЯМР-спектроскопия является способом, не ведущим к деструкции, и как ЯМР, так и MS позволяют исследовать интактный сапонин.
Для решения вопроса о структуре сапонинов необходим интегральный подход с применением различных спектроскопических способов, каждый из которых вносит определенный вклад в суммарные данные.
1. Масс-спектрометрия (MS)
Выбор способа ионизации при MS зависит от полярности, доступности и молекулярной массы соединения, подвергаемого анализу. В принципе это так называемые "мягкие" способы ионизации, такие как FAB и десорбция/химическая ионизация (D/CI), которые применяют для получения информации о молекулярной массе и последовательности сахаров природно существующих гликозидов (Wolfender et al., 1992). Это позволяет анализировать гликозиды без дериватизации. В определенных случаях наблюдается фрагментации агликонов, но для данной цели более полезны масс-спектры (EI-MS), получаемые при бомбардировке электронами.
2. MS при бомбардировке быстрыми атомами (FAB-MS)
При FAB исследованиях ускоренный пучок атомов (или ионов) выстреливается из пушки в направлении мишени, в которую предварительно загружают вязкую жидкость ("матрикс" - обычно глицерин или 1-тиоглицерин), содержащую образец, подвергаемый анализу (Barber et al., 1981; 1982). Когда пучок атомов сталкивается с матриксом, кинетическая энергия передается поверхностным молекулам, большое число которых выбрызгивается из жидкости в высокий вакуум, являющийся источником ионов. Ионизация многих из этих молекул наступает во время их разбрызгивания, давая как положительные, так и отрицательные ионы. Любые из них могут регистрироваться при подходящем выборе параметров прибора, но отрицательные ионы показали себя в целом как более пригодные для работы с сапонинами.
3. Масс-спектрометрия со вторичными ионами (SIMS)
Она представляет собой другой способ десорбции, индуцируемой частицами, в котором keV ионы, сталкиваясь на поверхности тонкой пленки биомолекулы, индуцируют такую же десорбционную ионизацию, что и при PD-MS (Benninghoven and Sichtermann, 1978). Была продемонстрирована применимость данного способа для исследования структуры трех новых бидесмозидов, ацетил-соясапонинов А1, А2 и А3, выделенных из семян американской сои (Glycine max, Leguminosae). Существенный фрагмент ионных пиков давал информацию в отношении типа ацетилирования в моносахаридных единицах, а также о последовательности данных единиц (Kitagawa et al., 1988).
4. Лазерная десорбция (LD)
C помощью LD было показано, что возбуждение под действием лазерных импульсов с короткой продолжительностью (<10 нсек) дает паттерны десорбированных молекулярных ионов, сходные с PD и SIMS. Масс-спектрометрия с лазерной десорбцией/преобразованием Фурье (LD/FTMS), способ, который также подходит для анализа сложных гликозидов, дает спектры, которые и отличаются от и комплементарны FAB-MS.
5. MS с полевой десорбцией (FD-MS)
Данный способ практичен для определения молекулярных масс сапонинов совместно с числом, природой и последовательностью остатков сахаров (Komori et al., 1985). Однако экспериментальная сложность FD-MS и тот факт, что FAB-MS дает более продленные спектры, означает, что в настоящее время популярность подхода FD-MS снижается. FD масс-спектры имеют дополнительный недостаток в том, что они усложняются в результате присутствия катионизированных фрагментов, затрудняющих интерпретацию. Тем не менее FD-MS успешно применяли для выяснения структуры сапонинов (Hostettmann, 1980).
(iv) Жидкостная хроматография-масс-спектрометрия (LC-MS)
В настоящее время разработано несколько типов эффективных средств для прямого и непрямого введения прохождения через колонки ВЭЖХ для масс-спектрометрического анализа. Например, качественный анализ сырых фракций сапонинов выполняли с помощью сопряжения сочетания полу-микро ВЭЖХ с бомбардировкой летающими быстрыми атомами (FRIT-FAB) (Hattori et al., 1988). При данном применении использование колонки NH2 (например, μS-Finepak SIL NH2, Jasco; 25 см х 1,5 мм внутренний диаметр (I.D.)) предпочтительнее, чем колонки октадецил-двуокиси кремния, с разделительным отношением для элюата 1:20 (100 мк/мин → 5 мкл/мин). Элюция линейным градиентом ацетонитрила и воды, содержащей 1% глицерин, обычно дает лучшую остроту пиков, чем получаемая при изократической элюции. Негативные FAB масс-спектры записывали для сапонинов с молекулярной массой до 1235. При данном способе наблюдали псевдомолекулярные [M - 1]- ионы, а также ионные фрагменты, приписываемые отщеплению сахарных частей (Hattori et al., 1988).
Система FRIT-FAB LC-MS описана для разделения смеси изомерных сапонинов розамултина и арджунетина (оба с молекулярной массой 650) из Rosa rugosa (Rosaceae). Розамултин (урсановый гликозид) и арджунетин (олеанановый гликозид), оба имеют единственный остаток глюкозы у С-28, и их анализировали с помощью как негативного, так и позитивного способов FAB с ксеноном в качестве нейтрального газа. ВЭЖХ выполняли на колонке октадецил-двуокиси кремния (250 х 1,5 мм) со смесью ацетонитрил-вода (7:3, содержащей 0,5% глицерин) в качестве растворителя при скорости тока 1 мл/мин. Наблюдали псевдомолекулярные ионы [M + 1]- и [M + 1]+ вместе с сильными пиками, обусловленными родительскими агликонами в негативных масс-спектрах FAB (Young et al., 1988).
Можно также определить сапонины с помощью динамической масс-спектрометрии с вторичными ионами (SIMS) способом, сходным со средствами динамической FAB, в котором элюент проходит прямо в источник. Таким образом, ВЭЖХ в сочетании с УФ (206 нм) и SIMS-определением применяли для анализа смеси одного моно- и двух бидесмозидных тритерпеновых гликозидов (Marston et al., 1991).
Недостатком интерфейсов типа FRIT-FAB и CF-FAB является требуемая низкая скорость тока (около 1-5 мкл/мин). После разделения ВЭЖХ необходима разбивка элюата. Интерфейс термоспрея (TSP) (Blackley and Vestal, 1983), однако, характеризуется своей простотой и своей способностью поддерживать скорости тока в 1-2 мл/мин. Это делает способ более привлекательным для решения проблем, включающих в себя анализ растительных составляющих компонентов. Основой способа TSP является мягкая ионизация молекул, сходная с химической ионизацией MS. Это позволяет анализировать нелетучие и термолабильные моно-, ди- и даже тригликозиды. Получают информацию о молекулярной массе сапонина и природе и последовательности сахарных цепей. TSP LC-MS применяли для анализа моллюскицидных сапонинов в метанольном экстракте плодов Tetrapleura tetraptera (Leguminosae) (Maillard an Hostettmann, 1993). С добавлением после колонки 0,5 М ацетата аммония (0,2 мл/мин) для обеспечения летучим буфером для ионизации ионов при упаривании суммарное содержание ионов при TSP LC-MS (диапазон массы от 450 до 1000 единиц атомной массы) хорошо соответствовало анализу ВЭЖХ-УФ при 206 нм. Следы ионов при m/z 660, 676, 880 и 822 давали сигналы, соответствующие псевдомолекулярным [M + H]+ ионам главных сапонинов. TSP масс-спектр, приобретенный каждым сапонином в экстракте, проявлял главный пик для псевдомолекулярного [M + H]+ иона. Фрагментация сахарных частей наблюдалась для главного моллюскицидного сапонина, ариданина, в котором потеря N-ацетилглюкозильной части давала увеличение [A + H]+ пика для агликона (Maillard and Hostettmann, 1993).
LC-MS при использовании для исследования сапонинов имеет большую потенциальную применимость, так как GC-MS имеет минимальное практическое применение и в одной ВЭЖХ идентичности пиков могут только подтверждаться по их времени удерживания. LC-MS не только подходит для анализа тритерпеновых гликозидов в растительных экстрактах, но является также ценной, через MS-MS, для определения структуры индивидуальных сапонинов в экстрактах.
(v) Инфракрасная спектроскопия (ИК)
Не считая обычного применения ИК, существуют одна или две черты, которые особенно подходят для выяснения структуры сапонинов. ИК пригодна для характеристики стероидных сапогенинов, потому что несколько сильных полос между 1350 и 875 см-1 являются диагностическими для спирокетальной боковой цепи (Jones et al., 1953). Четыре полосы, 980 (полоса А), 920 (полоса В), 900 (полоса С) и 860 см-1 (полоса D), были определены как характерологические для колец E и F. У 25R-сапогенинов полоса В имеет более сильное поглощение, чем полоса С, в то время как в 25R-сериях данное отношение является обратным. В сапогенинах, имеющих кислородные заместители в кольцах E и F или в положении 27, четыре полосы существенно изменены (Takeda, 1972).
Присутствие в сапонинах ионизированных карбоксильных групп может быть установлено с помощью полос в ИК-спектре при 1610 и 1390 см-1 (Numata et al., 1987). Данная информация полезна в течение процедуры выделения, когда важно знать, будут ли карбоксильные группы в молекуле ионизированы.
(vi) Рентгеноструктурная кристаллография
Рентгеноструктурная кристаллография применялась для выяснения молекулярной геометрии трисахаридного тритерпенового азиатикозида из Centella asiatica (Umbelliferae). Осуществляли кристаллизацию из диоксана (Mahato et al., 1987). Анализ рентгеновского преломления также был успешным для подтверждения структуры 3-β-D-гликозида молевой кислоты (Pegel and Rogers, 1985).
Рентгеноструктурная кристаллография особенно пригодна для решения структурных проблем агликонов. Полезная информация для определения структуры агликона алатозида А из Sesamum alatum (Pedaliaceae) была получена с помощью анализа рентгеновского преломления кристаллического триацетата искусственного продукта, полученного после кислотного гидролиза (Potterat et al., 1992). Исследование с помощью рентгеноструктурной кристаллографии медикагеновой кислоты родительского агликона 3-О-глюкозида медикагеновой кислоты из клубней Dolichos kilimandscharicus (Leguminosae) показало, что молекула имеет цис-слитые кольца D и Е. Кольцо С имело слегка искаженную конформацию софы, в то время как кольца A, B, D и Е имели конформацию кресла (Stoeckti-Evans, 1989).
(vii) Реакции расщепления
Тритерпеновые сапонины представляют собой гликозиды, в которых гемиацетальные гидроксильные группы сахаридов в их циклических пиранозных или фуранозных формах образуют ацетали с тритерпеновым или стероидным остатком. Эфирная связь между гемиацетальным гидроксилом и тритерпеном или стероидом известна как гликозидная связь. Моносахаридные составляющие олигосахаридов также связаны эфирными связями (межгликозидные связи).
При полном гидролизе гликозида гликозидная связь разрушается с высвобождением моносахаридных компонентов и неуглеводной части (агликона или генина). Неуглеводная часть, получаемая при гидролизе сапонинов, обозначается как сапогенол или сапогенин. Все известные сапонины являются О-гликозидами с эфирными или сложноэфирными связями.
Многочисленные химические реакции и способы применялись для разрушения сапонинов до более мелких единиц для более удобного анализа (см., например, Kitagawa, 1981). Такие способы должны найти особое применение в определении структуры тритерпеновых сапонинов.
1. Кислотный гидролиз
Кислотный гидролиз может быть выполнен путем кипячения с обратным холодильником сапонина в кислоте в течение фиксированного промежутка времени, например 4 час, в 2-4 М хлористоводородной кислоте. Водный раствор, остающийся после гидролиза, экстрагируют диэтиловым эфиром, хлороформом или этилацетатом для получения агликона. Экстракция сахаров из водного слоя выполняется с помощью пиридина после нейтрализации раствора (щелочной или основной ионообменной смолой) (Tshcesche and Forstmann, 1957; Sandberg and Michel, 1962) и упаривания досуха. Сапонины полностью разрушаются до их составляющих частей с помощью данного способа так, что получают информацию как об индивидуальном агликоне, так и о числе и природе присутствующих моносахаридов. Если просапогенин (полученный после расщепления сложноэфирной связи с помощью гидролиза основанием) гидролизуют кислотой, может быть установлена природа цепей сахаров, которые связаны эфирной связью. Водная реакционная среда может быть заменена спиртовой или диоксановой.
В добавление к хлористоводородной кислоте для гидролиза сапонинов может быть также применена серная кислота. При применении серной кислоты существует меньше шансов для деградации или реорганизации молекулы, но расщепление сложноэфирных связей не настолько эффективно. Удобным способом получения гипсогеновой кислоты из сапонинов Dianthus, например, включал гидролиз 1 М серной кислотой в диоксане (Oshima et al., 1984). Сравнительное исследование условий гидролиза хлористоводородной кислотой и серной кислотой в воде и в смеси вода-этанол показало, что наилучшая открываемость сахаридов достигается при нагревании сапонина в течение 2 час с 5% смесью серная кислота/вода в запаянной ампуле под вакуумом (Kikuchi et al., 1987). Несколько более мягкие условия гидролиза могут быть достигнуты с помощью трихлоруксусной кислоты, например, при кипячении с обратным холодильником в течение 3 час в 1 М трифторуксусной кислоте.
Альтернативой гидролиза сапонинов в растворе является их прямой гидролиз на пластине ТСХ путем обработки парами хлористоводородной кислоты. Как только кислота выпарена, выполняется обычное элюирование растворителем ТСХ, с тем, чтобы идентифицировать присутствующие моносахариды (Kartnig and Wegschaider, 1987). С помощью данного способа были идентифицированы концевые сахара, ксилоза и галактоза, после частичного гидролиза агавесида В. Была создана пластина ТСХ с растворителем в виде смеси хлороформ-метанол-вода (8:5:1) и определение осуществляли с помощью смеси анилин-дифениламин-H3PO4-метанол (1:1:5:48) (Uniyal et al., 1990).
2. Гидролиз основанием
Расщепление О-ацилгликозидных сахарных цепей достигают в условиях гидролиза основанием, обычно с помощью кипячения с обратным холодильником с 0,5 М гидроксидом калия. Альтернативно, могут быть использованы 1-20% этанольный или метанольный растворы гидроксида калия, но здесь существует риск метилирования, особенно карбоксильной группы тритерпеновых кислот. Ионообменники, такие как Dowex 1, создают мягкие условия гидролиза основанием (Bukharov and Karlin, 1970). Другим способом является применение иодида лития в коллидине (Kochetkov et al., 1964).
При строгом контроле условий реакций возможно избирательно расщеплять различные сложноэфирные части. Например, гидролиз сапонина К11 кизута с помощью кипячения с обратным холодильником в 0,5 М гидроксиде калия в течение 30 мин удаляет сахар при С-28 бидесмозида. Однако перемешивание сапонина в течение 20 час в 0,1 М гидроксиде калия при комнатной температуре избирательно удаляет ацетатную группу у С-28 сложноэфирной гликозидной цепи (Kizu et al., 1985b).
3.Частичный гидролиз
В определенных примерах, когда сапонины имеют высокоразветвленные или длинные сахарные цепи, необходима процедура, включающая в себя частичный гидролиз, с тем чтобы получить фрагменты, более поддающиеся выяснению структуры. Это может быть достигнуто с применением кислоты или, конечно, ферментов. Олигосахаридные и/или оставшиеся сапониновые части выделяют и затем характеризуют.
Например, сапонин из Phytolacca dodecandra (Phytolaccaceae) гидролизовали 0,1 М хлористоводородной кислотой в течение 45 мин для получения смеси трех продуктов. Данные соединения разделяли с помощью RP-LPLC и их сахарные последовательности определяли с помощью MS, 13C-ЯМР и GC-MS ацетатов алдитолов. Собирая всю эту информацию вместе, можно приписать химическую формулу соединению, производному олеаноловой кислоты (Dorsaz and Hostettmann, 1986).
Гидролиз в диоксане дает более мягкие условия и возможен частичный гидролиз. В данном примере сапонин кипятили с обратным холодильником в течение 6 час в смеси диоксан-0,1 М хлористоводородная кислота (1:3) (Ikram et al., 1981). Другой способ частичного гидролиза сапонинов представляет собой обработку раствора тритерпенового гликозида в спирте щелочным металлом (натрием или калием) и затем добавление следов воды (Ogihara and Nose, 1986).
4. Гидротермолиз
Гидротермолиз тритерпеновых гликозидов ведет к образованию соответствующих агликонов и таким образом может помочь в определении структуры. Способ включает в себя нагревание гликозида с водой или смесью вода-диоксан при температуре от 100 до 140°С в течение периода от 10 до 140 час в зависимости от образца. Например, гидротермолиз тритерпеновых 3,28-О-бисгликозидов дает соответствующие 3-О-гликозиды (Kim et al., 1992).
5. Ферментативный гидролиз
Очень эффективным и мягким способом отщепления остатков сахаров от сапонинов без образования побочных продуктов является ферментативный гидролиз. Хотя подходящие гидролазы для всех сахаров не имеются в продаже, отщепление остатков β-глюкозы с помощью β-глюкозидазы является особенно простым. Дополнительной выгодой отщепления специфическими ферментами является то, что автоматически подтверждается аномерная конфигурация сахарной части. Определенные препараты ферментов, которые особенно предлагаются для применения при гидролизе тритерпеновых гликозидов, представляют собой β-галактозидазу, гидролазы, целлюлазу, сырую хесперидиназу, пектиназу и нарингиназу.
Систематическое исследование, включающее сырые препараты хесперидиназы, нарингиназы, пектиназы, целлюлазы, амилазы и эмульсина, показали, что хесперидиназа, нарингиназа и пектиназа были наиболее эффективными при гидролизе гинсенозидов (Kohda and Tanaka, 1975).
(viii) Анализ агликонов после гидролиза
После окончания гидролиза агликоны могут быть отделены от гидролизата либо путем простой фильтрации, либо с помощью распределения между водой и органической фазой и могут анализироваться путем сопоставления с известными тритерпенами. Наиболее обычным методом является ТСХ c применением растворителей, таких как смесь диизопропиловый эфир-ацетон (75:30). Реагенты для опрыскивания часто являются теми же, что и применяемые для анализа сапонинов (см. таблицу 2).
Газожидкостная хроматография требует дериватизации тритерпенов. Например, метиловые эфиры олеаноловой и урсолевой кислот разделяли с помощью GC на стеклянной колонке, упакованной 30% 0V-17 или SE-30 (Fokina, 1979). Тритерпены могут быть определены с помощью GC после дериватизации N,O-бис(триметилсилил)ацетамидом и хлортриметилсиланом, как это происходит в случае соясапогенолов А-Е и медикагеновой кислоты люцерны (Jurzysta and Jurzysta, 1978).
Способ GC-MS также представляет ценность для характеристики сапогенинов. Обычно получают триметилсилильные производные и затем анализируют их на спектрометре. Примером является приложение этого к исследованию тритерпенов типа олеанана и урсана. Девять дериватизированных силилом тритерпенов разделяли с помощью GC на упакованной OV-101 колонке и исследовали паттерны их масс-спектров; те, которые содержали 12-ен двойную связь, подвергали характерной реакции ретро-Dies-Alder (Burnouf-Radosevich et al., 1985). Данный способ применяли для определения тритерпенов из лакричника (Bombardelli et al., 1979).
Анализ ВЭЖХ не требует дериватизации и дает отличную воспроизводимость и чувствительность для анализа тритерпенов. Может быть применена как ВЭЖХ с нормальной фазой (анализ сапогенинов лебеды; Burnouf-Radosevich and Delfel, 1984), так и RP-ВЭЖХ (Lin et al., 1981), но недостатком RP-ВЭЖХ является то, что соединения имеют тенденцию к преципитации в водных подвижных фазах.
(ix) Анализ сахаров после гидролиза
Анализ моносахаридов может быть осуществлен с помощью ТСХ на, например, силикагельных пластинах с растворителями, такими как смесь этилацетат-метанол-вода-уксусная кислота (65:25:15:20) и смесь н-бутанол-этилацетат-изопропанол-уксусная кислота-вода (35:100:60:35:30) (Shiraiwa et al., 1991). Определение обычно проводят с пара-анисидин-фталатом, нафторезорцином, тимолсерной кислотой (Kartnig and Wegschaider, 1971) или с хлоридом трифенилтетразолия (Wallenfels, 1950; Kamel et al., 1991). Альтернативно, количественный анализ моносахаридов возможен с помощью GC или ВЭЖХ.
Был описан ряд способов ВЭЖХ для анализа сахаров, включающий в себя анализ на NH2-связанных колонках со смесью ацетонитрил-вода (75:25) (Glombitza and Kurth, 1987); анализ на колонках С-18 (смесь ацетонитрил-вода 4:1) с определением индекса преломления для целей количественного определения, интеграцию пиков ВЭЖХ сравнивали со стандартами (Adinolfi et al., 1987); анализ на колонке HPX-87H c исключением ионов Aminex (BioRad) с 0,005 М серной кислотой в качестве элюента (0,4 мл/мин) (Adinolfi et al., 1990) и анализ п-бромбензоатов сахаров (образованных путем метанолиза сапонина 5% смесью хлористоводородная кислота-метанол и последующего пара-бромбензоилирования метилированных сахаров) с помощью ВЭЖХ и идентификации путем сравнения с подлинными производными (Kawai et al., 1988; Sakamoto et al., 1992).
Для GC применяют предварительно дериватизированные силилом сахара (Wulff, 1965) или выполняют GC-MS анализ алдитол-ацетатных производных. GC-преобразование Фурье ИК (FTIR) анализ подходящим образом дериватизированных моносахаридов является альтернативным способом (Chen and Snyder, 1989).
Наиболее обычно встречающимися сахарами являются D-глюкоза, D-галактоза, L-арабиноза, D-ксилоза, D-фукоза, L-рамноза, D-хиновоза, D-глюкуроновая кислота и D-рибоза.
III. Производные соединений
Как здесь подробно описано, предполагается, что определенные преимущества могут быть достигнуты при манипуляциях с монотерпеновыми/тритерпеновыми гликозидами для придания им новых характеристик, более длинного полупериода жизни in vivo или других благоприятных свойств. Такие способы включают в себя, но не ограничиваются этим, манипуляции или модификацию смесей монотерпеновых/тритерпеновых гликозидов или индивидуальной монотерпеновой/тритерпеновой молекулы самой по себе, модификацию или удаление сахаров и конъюгацию монотерпеновых/тритерпеновых соединений с инертными носителями, такими как композиции, которые придают мембранам растворимость/проницаемость или дают доступ в клетки, включая липиды, белки-носители, липофильные белки, другие тритерпеновые части, сахара и т.д. до различных белковых или небелковых компонентов, включая иммуноглобулины и Fc-части. Должно быть понятно, что более длинный полупериод жизни не является одинаково протяженным с фармацевтическими композициями для применения в "медленном высвобождении". Составы с медленным высвобождением обычно разрабатываются для создания постоянного уровня лекарства в течение продолжительного периода. Увеличение полупериода жизни лекарства, такого как монотерпеновый/тритерпеновый гликозид в соответствии с настоящим изобретением, предназначается для возникновения в результате его высоких уровней в плазме при введении, причем его уровни поддерживаются в течение более длительного времени, но уровни обычно снижаются в зависимости от фармакокинетики соединения.
(i) Конъюгаты монотерпенов/тритерпенов и линкерных молекул
Как описано выше, идентифицированные здесь монотерпеновые/тритерпеновые соединения могут быть присоединены к особым молекулам для того, чтобы улучшить эффективность действия монотерпеновых/тритерпеновых гликозидов у больных, подвергаемых лечению, в отношении любого заболевания, которое лечится соединениями согласно изобретению. Иллюстративное осуществление таких молекул включает в себя направляющие агенты и агенты, которые должны увеличивать полупериод жизни монотерпеновых/тритерпеновых соединений in vivo, или агенты, которые делают монотерпеновую композицию проницаемой для мембраны. Монотерпеновые/тритерпеновые соединения могут быть присоединены к таким вторичным молекулам любым действенным способом, который позволяет каждой области выполнять предназначенную для нее функцию без существенного нарушения биологической активности, например противоопухолевой активности раскрытых здесь соединений.
Монотерпеновые/тритерпеновые композиции согласно изобретению могут быть прямо связаны со вторым соединением или могут быть присоединены через линкерную группу. Под термином "линкерная группа" подразумевается одна или более бифункциональных молекул, которые могут быть использованы для ковалентного присоединения монотерпеновых/тритерпеновых соединений или смеси тритерпенов к агенту и которые не вмешиваются в биологическую активность монотерпеновых/тритерпеновых соединений. Линкерная группа может быть присоединена к любой части монотерпенов/тритерпенов до тех пор, пока точка присоединения не будет вмешиваться в биологическую активность, например противоопухолевую активность соединений.
Примером осуществления связывания монотерпеновых/тритерпеновых соединений согласно изобретению со вторым агентом является получение активного сложного эфира монотерпенов/тритерпенов с последующей реакцией активного сложного эфира с нуклеофильной функциональной группой агента, предназначенного для присоединения. Активные сложные эфиры могут быть получены, например, путем реакции карбоксильной группы тритерпена со спиртом в присутствии агента дегидрирования, такого как дициклогексилкарбодиимид (DCC), 1-(3-диметиламинопропил)-3-этилкарбодиимид (EDC) и метиодид 1-(3-диметиламинопропил)-3-этилкарбодиимида (EDCI). Применение EDC для образования конъюгатов раскрыто в патенте США No. 4526714; заявка РСТ No. WO 91/01750 и Arnon et al., 1980, раскрытия которых специально включены здесь в качестве ссылки во всей своей полноте. Агент, предназначенный для присоединения к монотерпену/тритерпену, например специфичное для опухоли антитело или антитело, специфичное для клетки с воспалением, затем смешивают с активированным эфиром в водном растворе для получения конъюгата.
Когда желательны линкерная группа между монотерпеном/тритерпеном и агент, активный сложный эфир монотерпенового/тритерпенового гликозида, может быть получен как описано выше, и введен во взаимодействие с линкерной группой, например 2-аминоэтанолом, алкилендиамином, аминокислотой, такой как глицин, или карбокси-защищенной аминокислотой, такой как трет-бутиловый сложный эфир глицина. Если линкер содержит защищенную карбоксильную группу, защитную группу удаляют и получают активный сложный эфир линкера (как описано выше). Активный сложный эфир затем вводят в реакцию со второй молекулой для получения конъюгата. Альтернативно, второй агент может быть дериватизирован янтарным ангидридом для получения конъюгата агент-сукцинат, который может быть конденсирован в присутствии EDC или EDCI с монотерпеновым/тритерпеновым линкерным производным, имеющим в линкере свободную амино- или гидроксильную группу (см., например, патентную заявку WO 91/01750, раскрытие которой специально включено здесь в качестве ссылки во всей своей полноте).
Можно также получить монотерпеновый/тритерпеновый гликозидный конъюгат, включающий в себя линкер со свободной аминогруппой, и перекрестно сшить свободную аминогруппу с гетеробифункциональным кросслинкером, таким как сульфосукцинимидил-4-(N-малеимидоциклогексан)-1-карбоксилат, который будет взаимодействовать со свободными сульфгидрильными группами белковых антигенов.
Монотерпеновый/тритерпеновый гликозид может быть также присоединен к линкерной группе с помощью реакции альдегидной группы с аминолинкером с образованием промежуточного иминного конъюгата с последующим восстановлением с помощью борогидрида натрия или цианоборогидрида натрия. Примеры таких линкеров включают в себя аминоспирты, такие как 2-аминоэтанол, и диаминосоединения, такие как этилендиамин, 1,2-пропилендиамин, 1,5-пентандиамин, 1,6-гександиамин и тому подобное. Монотерпеновый/тритерпеновый гликозид может быть затем присоединен к линкеру путем первоначального образования сукцинатного производного с янтарным ангидридом с последующей конденсацией конъюгата монотерпеновый/тритерпеновый гликозид-линкер с DCC, EDC или EDCI.
Кроме того, монотерпеновый/тритерпеновый гликозид или агликон может окисляться периодатом, и получаемый в результате этого диальдегид может конденсироваться с аминоспиртом или диаминосоединением, указанным выше. Свободная гидроксильная или аминогруппа линкера может затем конденсироваться с сукцинатным производным антигена в присутствии DCC, EDC или EDCI. Специалистам в данной области техники известны многие типы линкеров, и они могут быть применены при создании тритерпеновых конъюгатов. Перечень примеров линкеров для применения в изобретении дан ниже в таблице 4.
Гетеробифункциональные кросслинкеры
Сульфгидрилы
Сульфгидрилы
• Расщепляемая поперечная сшивка
Сульфгидрилы
Сульфгидрилы
• Водорастворимый
Сульфгидрилы
• Конъюгация фермент-антитело
• Конъюгация гаптен-белок-носитель
Сульфгидрилы
• Водорастворимый
• Конъюгация фермент-антитело
Сульфгидрилы
• Конъюгация гаптен-белок-носитель
Сульфгидрилы
Сульфгидрилы
Сульфгидрилы
Сульфгидрилы
• Конъюгация фермент-антитело
Сульфгидрилы
• Водорастворимый
сульфо-NHS
Карбоксильные группы
(ii) Рациональный дизайн лекарств
Цель рационального дизайна лекарств заключается в получении структурных аналогов биологически активных соединений. Путем создания таких аналогов возможно смоделировать лекарства, которые являются более эффективными или стабильными, чем природные молекулы, которые имеют различную восприимчивость к изменению или которые могут влиять на функции различных других молекул. При одном подходе можно создать описанную здесь трехмерную структуру монотерпеновых/тритерпеновых соединений или их фрагментов. Это может быть осуществлено с помощью рентгеноструктурной кристаллографии, компьютерного моделирования или сочетания обоих подходов. Альтернативный подход включает в себя случайное замещение функциональных групп в монотерпеновой/тритерпеновой молекуле и определение результирующего влияния на функцию.
Возможно также выделить антитело, специфичное по отношению к монотерпеновому/тритерпеновому соединению, отобранное с помощью функционального теста, и затем выяснить его кристаллическую структуру. В принципе данный подход дает фармакологическую основу, на базе которой может быть основан последующий дизайн лекарства. Можно обойти кристаллографию белков вообще путем создания антиидиотипических антител против функционального фармакологически активного антитела. В качестве зеркального отражения от зеркального отражения связывающий сайт антиидиотипа, как ожидается, должен быть аналогом исходного антигена. Антиидиотип может потом использоваться для идентификации выделенных пептидов из банка пептидов, полученных химическим или биологическим путем. Отобранные пептиды должны затем служить в качестве фармакологической основы. Антиидиотипы могут быть созданы с применением описанных здесь способов продукции антител с применением антитела в качестве антигена.
Таким образом, можно создать дизайн лекарств, которые имеют улучшенную биологическую активность, например противовоспалительную активность или противоопухолевую активность, по отношению к исходному монотерпеновому/тритерпеновому соединению. Посредством процедур химического выделения и представленных здесь описаний могут быть получены количества монотерпеновых/тритерпеновых соединений согласно изобретению, достаточные для выполнения кристаллографических исследований. Кроме того, знание химических характеристик данных соединений позволяет применять компьютерное предсказание структурно-функциональных взаимоотношений.
IV. Лечение рака тритерпеновыми соединениями
При развитии рака клетки млекопитающих проходят через серию генетически детерминированных изменений, которые ведут к нетипичной пролиферации. Это может происходить через стадии, обычно обозначаемые как (1) инициация, когда внешний агент или стимул является триггером генетического изменения в одной или более клеток и (2) промоция, включающая дальнейшие генетические и метаболические изменения, которые могут включать в себя воспаление. В течение "стадии промоции" клетки начинают метаболический переход к стадии клеточного роста, при котором апоптоз блокируется.
Раковые клетки характеризуются потерей контроля апоптоза в добавление к потере контроля за регуляторными стадиями клеточного цикла. Раковые клетки (малигнизированные клетки) избавляются от нормальных механизмов контроля роста в результате серий метаболических изменений в течение стадий инициации и промоции при наступлении малигнизации. Данные изменения являются следствием генетических изменений в клетках. Данные генетические изменения могут включать в себя (i) активирующие мутации и/или увеличенную экспрессию протоонкогенов и/или (ii) инактивирующие мутации и/или пониженную экспрессию одного или более генов-супрессоров опухолей. Главными онкогенными продуктами и продуктами генов-супрессоров опухолей являются компоненты путей передачи сигнала, которые контролируют вхождение в клеточный цикл и выход из него, стимулируют дифференцировку, улавливают повреждение ДНК и инициируют механизмы репарации и/или регулируют программы клеточной смерти. Почти все опухоли имеют мутации множественных онкогенов и генов-супрессоров опухолей. Можно заключить, что клетки применяют множественные параллельные механизмы для регуляции клеточного роста, дифференцировки, контроля над повреждениями ДНК и апоптоза.
Описанные здесь тритерпеновые соединения могут быть введены нуждающемуся в них субъекту для лечения субъекта либо при профилактическом предотвращении рака, либо при терапии после обнаружения опухоли. Для ингибирования инициации и промоции рака, уничтожения раковых/малигнизированных клеток, ингибирования роста клеток, индукции апоптоза, ингибирования метастазов, снижения размеров опухоли и обратного изменения или редукции малигнизированного фенотипа опухолевых клеток другими способами с применением способов и композиций согласно изобретению обычно необходимо ввести в контакт клетку-"мишень" с описанными здесь тритерпеновыми композициями. Это может быть достигнуто путем контактирования опухоли или опухолевой клетки с одной композицией или фармакологическим составом, который включает в себя тритерпеновые соединения, или путем контактирования опухоли или опухолевой клетки с более чем одной отдельной композицией или составом в одно и то же время, где одна композиция включает в себя тритерпен, а другая включает в себя второй агент.
Предпочтительные раковые клетки для лечения соединениями согласно изобретению включают в себя эпителиальные раковые клетки, такие как клетки кожи, ободочной кишки, матки, яичников, поджелудочной железы, легких, мочевого пузыря, молочной железы, почки и простаты. Другие раковые клетки-мишени включают в себя раковые клетки мозга, печени, желудка, пищевода, головы и шеи, яичек, шейки матки, лимфатической системы, гортани, пищевода, околоушной железы, желчных путей, прямой кишки, матки, эндометрия, почки, мочевого пузыря и щитовидной железы, включая сквамозно-клеточные карциномы, аденокарциномы, мелкоклеточные карциномы, глиомы, нейробластомы и тому подобное. Однако данный перечень предназначен только для иллюстрации и не является ограничивающим, так как потенциально любая раковая клетка может подвергаться лечению тритерпеновыми соединениями. Методы тестирования для выяснения относительной эффективности соединений при лечении указанных выше типов опухолевых клеток и других опухолевых клеток конкретно раскрыты здесь и должны быть понятны специалистам в данной области техники в свете настоящего раскрытия.
Описанные здесь соединения предпочтительно вводят в виде композиции пищевой добавки или фармацевтической композиции, включающей в себя фармацевтически или фармакологически приемлемый разбавитель или носитель. Природа носителя зависит от химических свойств применяемого(ых) соединения(ий), включая характер растворимости и/или способ введения. Например, если желательно пероральное введение, может быть выбран твердый носитель, и для в/в введения может быть использован жидкий носитель в виде солевого раствора.
Фразы "фармацевтически или фармакологически приемлемый" относятся к молекулярной сущности и композициям, которые не дают неблагоприятную, аллергическую или другую нежелательную реакцию при введении животному или человеку, как назначено. Применяемый здесь термин "фармацевтически приемлемый носитель" включает в себя любой и все растворители, дисперсионные среды, оболочки, антибактериальные и противогрибковые агенты, изотонические и замедляющие абсорбцию агенты и тому подобное. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно специалистам в данной области техники. За исключением тех случаев, когда общепринятые среды или агент не совместимы с активным ингредиентом, рассматривается их применение в терапевтических композициях. В композиции могут быть также включены добавочные активные ингредиенты.
а. Пищевые добавки
Композиции в виде пищевых добавок являются препаратами природных ингредиентов, которые представляют собой мультикомпонентные системы, состоящие из предпочтительно синергичных природных продуктов и добавок для стимуляции хорошего самочувствия. Соединения пищевых добавок могут происходить из лекарственных растений. Информация о многочисленных растениях и травах, применяемых для получения композиций в виде пищевых добавок, составлена и доступна из публикаций, включая German Commission E Monographs, Botanical Safety Handbook и HerbalGram, ежеквартальную публикацию американского ботанического совета, который ссылается на многочисленные клинические испытания, которые были выполнены с применением пищевых добавок.
Информация по описанию, составляющим частям, современному использованию, дозировке (в разнообразных формах), действию, противопоказаниям, побочным эффектам, взаимодействию с традиционными лекарствами, способу введения, продолжительности применения, регуляторному статусу, рейтинг ботанической безопасности AHPA и комментарии имеются для ряда растений и включают в себя среди прочего, чернику, жостер, акацию Грегги, красный стручковый перец, клюкву, лютик полевой, dong quai, эхинацею, масло энотеры, пиретрум девичий, чеснок, имбирь, гинко, азиатский женьшень, сибирский женьшень, желтокорень канадский, gotu kola, семена винограда, зеленый чай, боярышник, kava, лакричник, молокан ядовитый, пальму сереноа, St. John's ясменник и валериану.
Действие данных соединений в виде пищевых добавок может быть быстрым и/или кратковременным или может помочь достигнуть цели продолжительного здорового состояния. Описанные здесь фитомедицинские средства происходят из Acacia victoriae. Композиции в виде пищевых добавок, включающие в себя высушенные и грунтовые корни Acacia victoriae и стручки или экстракты из данных тканей в фармакологически приемлемой среде, служат в качестве природных подходов, среди прочего, к предотвращению и лечению воспалительных заболеваний и рака. Композиции в виде пищевых добавок могут быть использованы для предотвращения инициации и промоции канцерогенеза и также для индукции апоптоза в малигнизированных раковых клетках. Раскрытые здесь композиции в виде пищевых добавок могут также применяться в качестве противовоспалительных, противогрибковых, противовирусных, антимутагенных, спермицидных или контрацептивных, действующих на сердечно-сосудистую систему и регулирующих метаболизм холестерина агентов. Композиции в виде пищевых добавок могут содержаться в среде, такой как буфер, растворитель, разбавитель, инертный носитель, масло, мазь или потребляемый в пищу материал.
Пищевые добавки могут вводиться перорально и могут быть в форме таблетки или капсулы. Пероральный прием может быть предпочтительным для лечения эзофагита Барретта, воспалительных заболеваний кишечника, рака ободочной кишки и других опухолей внутренних органов.
Альтернативно пищевые добавки могут быть в форме мази, которая содержит экстракты корней или стручков Acacia victoriae в масле, или в виде крема, который может быть нанесен местно на кожу. Данная форма композиции в виде пищевой добавки пригодна для предотвращения инициации рака кожи. Применение данных композиций в виде пищевых добавок обеспечивает способом ингибирования инициации и промоции превращения эпителиальных клеток млекопитающих в премалигнизированное или малигнизированное состояние, когда терапевтически эффективное количество композиции в виде пищевой добавки подводят к данной клетке млекопитающего. Это особенно пригодно для видов рака эпителиальных клеток, таких как рак кожи.
b. Фармацевтические композиции
Описываемыми здесь дополнительно являются композиции, выбеленные из Acacia victoriae, которые частично или полностью очищены и структурно охарактеризованы. Очистка и характеристика данных монотерпеновых/тритерпеновых соединений подробно описана в примерах. D1, G1 и B1 представляют собой три композиции, которые были полностью очищены и их структурная характеристика является почти полной (фиг. 39, 40 и 41). Биологические тесты, выполненные с данными соединениями на клеточных линиях рака, продемонстрировали ингибирование клеточного роста и индукцию апоптоза в малигнизированных клетках (фиг. 43, 44). Более того, частично очищенные композиции данных сапонинов, выделенных из Acacia victoriae, также проявляют хемопротекторные эффекты у мышей, экспонированных с канцерогеном DMBA (фиг. 8, 9, 11, 12 и 13). Таким образом, данные композиции обладают противораковой активностью и действуют с помощью нескольких механизмов, индуцируя апоптоз в раковых клетках. Фармацевтические композиции данных соединений рассматриваются как сильные химиотерапевтические лекарства, которые могут быть использованы сами по себе или в сочетании с другими формами противораковой терапии, такими как химиотерапия, лучевая терапия, хирургия, генная терапия и иммунотерапия. Сочетание видов терапии подробно описано ниже. Специалист в данной области техники должен определить эффективные дозировки и режим сочетанной терапии.
с. Способы введения
(i) Парентеральное введение
В одном осуществлении изобретения предлагаются составы для парентерального введения монотерпеновых/тритерпеновых композиций, например, составленные для введения внутривенным, внутримышечным, подкожным или другим таким способом, включая прямое вливание в опухоль или место заболевания. Получение водной композиции, которая содержит монотерпеновую/тритерпеновую композицию, должно быть известно специалистам в данной области техники в свете настоящего раскрытия. Обычно такие композиции могут быть получены в виде инъецируемых либо как жидкие растворы, либо как суспензии; могут быть получены также твердые формы, подходящие для применения при получении растворов или суспензий при добавлении жидкости перед введением, и препараты могут быть также эмульгированными.
Растворы активных соединений в виде свободных оснований или фармацевтически приемлемых солей могут быть получены в воде, подходящим образом смешанной с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Диспергированные составы также могут быть получены в глицерине, жидких полиэтиленгликолях и их смесях и в маслах. При обычных условиях хранения и использования данные препараты содержат консервант для предотвращения роста микроорганизмов.
Фармацевтические формы, подходящие для использования в виде инъекций, включают в себя стерильные водные растворы или диспергированные составы; составы, включающие в себя кунжутное масло, арахисовое масло или водный пропиленгликоль, и стерильные порошки для спонтанного приготовления стерильных растворов или диспергированных составов для инъекций. Во всех случаях форма должна быть стерильной и должна быть жидкой до такой степени, чтобы легко отбираться шприцем. Она должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы.
Монотерпеновые/тритерпеновые соединения могут быть составлены в композицию в нейтральной или солевой форме. Фармацевтически приемлемые соли включают в себя аддитивные соли кислоты (образуемые со свободными аминогруппами белка), которые образуются с неорганическими кислотами, такими как, например, хлористоводородная или фосфорная кислоты, или с такими органическими кислотами, как уксусная, щавелевая, винная, миндальная и тому подобное. Соли, образуемые со свободными карбоксильными группами, также могут происходить от неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и от таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и тому подобное.
Носитель также может представлять собой растворитель или среду для диспергирования, содержащие, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное), их подходящие смеси и растительные масла. Надлежащее жидкое состояние может поддерживаться, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае диспергированного состава и путем применения поверхностно-активных веществ. Предотвращение действия микроорганизмов может быть осуществлено с помощью различных антибактериальных и противогрибковых агентов, например парабенов, хлорбутанола, фенола, сорбиновой кислоты, тимерозала и тому подобного. Во многих случаях предпочтительным является включение изотонических агентов, например сахаров или хлорида натрия. Продолжительное всасывание вводимых композиций может быть осуществлено с помощью применения в композициях агентов, задерживающих всасывание, например моностеарата алюминия и желатина.
Стерильные растворы для введения получают путем включения активных соединений в требуемом количестве в подходящем растворителе с различными другими ингредиентами, перечисленными выше, как это требуется, с последующей стерилизацией после фильтрации. Обычно диспергированные составы получают путем включения различных стерильных активных ингредиентов в стерильный носитель, который содержит основную среду для диспергирования и другие требуемые ингредиенты из тех, которые перечислены выше. В случае стерильных порошков для получения стерильных растворов для введения предпочтительными методами получения являются способы высушивания в вакууме и лиофилизирования, которые дают порошок активного ингредиента плюс любой дополнительный желаемый ингредиент из его ранее отфильтрованного и стерилизованного раствора.
(ii) Другие способы введения
Другие способы введения также находят применение с предметом изобретения. Например, монотерпеновые/тритерпеновые соединения согласно изобретению могут быть составлены в виде суппозиториев и в некоторых случаях аэрозолей и интраназальных композиций. Для суппозиториев композиция носителей должна включать в себя традиционные связующие агенты и носители, такие как полиалкиленгликоли или триглицериды. Такие суппозитории могут образовываться из смесей, содержащих активный ингредиент в диапазоне приблизительно от 0,5 до 10% (мас./мас.), предпочтительно приблизительно от 1 до 2%.
Пероральные композиции могут быть получены в форме растворов, суспензий, таблеток, пилюль, капсул, составов с постоянным высвобождением или порошков. Данные композиции могут быть введены, например, путем проглатывания или ингаляции. Когда фармацевтическая композиция предназначена для ингаляции, композиция должна предпочтительно включать в себя аэрозоль. Примеры процедур получения водных аэрозолей для применения в данном изобретении можно найти в патенте США No. 5049388, раскрытие которого специально включено здесь в качестве ссылки во всей своей полноте. Получение препаратов сухих аэрозолей описывается, например, в патенте США No. 5607915, раскрытие которого специально включено здесь в качестве ссылки во всей своей полноте.
Также пригодным является введение описанных здесь соединений прямо в трансдермальных составах с усилителями проницаемости, такими как ДМСО. Данные композиции сходно могут включать в себя любые другие подходящие носители, наполнители или разбавители. Другие местные составы могут быть введены для лечения по показаниям определенных заболеваний. Например, могут быть получены интраназальные составы, которые включают в себя наполнители, которые не вызывают ни раздражения слизистой носа, ни значительного нарушения функции ресничек. Разбавители, такие как вода, водный солевой раствор или другие известные вещества, могут быть применены с объектом изобретения. Составы для интраназального введения также могут содержать консерванты, такие как, но не ограничиваясь этим, хлорбутанол и хлорид бензалкония. Поверхностно-активное вещество может присутствовать для увеличения всасывания соединений согласно изобретению слизистой носа.
(iii) Составы и лечение
При составлении растворы должны вводиться способом, совместимым с дозировкой состава и в таком количестве, которое является терапевтически эффективным. Выбранному составу могут сопутствовать разнообразные наполнители, включая, например, фармацевтически чистые маннит, лактозу, крахмал, стеарат магния, натрий сахарин целлюлоза, карбонат магния и тому подобное.
Обычно соединения согласно изобретению должны содержать от менее чем 1% приблизительно до 95% активного ингредиента, предпочтительно приблизительно от 10% до 50%. Больному должно вводиться предпочтительно между приблизительно 10 мг/кг массы тела больного в день и приблизительно 25 мг/кг массы тела больного в день. Частота введения должна быть определена лечащим врачом на основе ответной реакции больного. Другие эффективные дозировки могут быть легко определены специалистом в данной области техники и путем обычных испытаний, устанавливающих кривые дозозависимости.
Независимо от способа введения подходящие фармацевтические композиции в соответствии с изобретением должны обычно включать в себя количество монотерпеновой/тритерпеновой композиции, смешанной с приемлемым фармацевтическим разбавителем или наполнителем, таким как стерильный водный раствор, для получения диапазона конечных концентраций, зависящих от предназначения при применении. Способы получения обычно хорошо известны в данной области техники, примером чего является Remington's Pharmaceutical Sciences, 16th Ed. Mack Publishing Company, 1980, ссылка на который специально включена здесь в качестве ссылки во всей своей полноте. Должно быть понятно, что загрязнение эндотоксинами должно быть сведено к минимуму по степени безопасности, например менее 0,5 нг/мг белка. Более того, для введения человеку препараты должны соответствовать стандартам стерильности, пирогенности, общей безопасности и чистоты, как требуется управлением по биологической стандартизации FDA.
Терапевтически эффективные дозы легко определить с применением животной модели, как представлено здесь в подробных исследованиях. Например, экспериментальные животные, несущие солидные опухоли, часто используются для оптимизации приемлемых терапевтических доз до переноса их на клинические испытания. Такие модели, как известно, являются очень надежными для предсказания эффективной противораковой стратегии. Могут быть также применены сходные животные модели для воспаления и различных воспалительных состояний.
В определенных осуществлениях может быть желательным обеспечение постоянного поступления больному терапевтических композиций. Для внутривенного и внутриартериального путей это достигается с помощью системы капельницы. Для местного применения должно быть использовано повторное нанесение. Для различных подходов могут быть использованы составы с замедленным высвобождением, которые обеспечивают ограниченные, но постоянные количества терапевтического агента в течение продолжительного периода времени. Для внутреннего применения может быть предпочтительной непрерывная перфузия интересующей области. Это может быть достигнуто с помощью катетеризации, в некоторых случаях постоперационной, с последующим непрерывным введением терапевтического агента. Период времени для перфузии должен быть выбран врачом для конкретного больного и ситуации, но диапазон времени должен составлять приблизительно от 1-2 часов до 2-6 часов, до приблизительно 6-10 часов, до приблизительно 10-24 часов, до приблизительно 1-2 дней, до приблизительно 1-2 недель или дольше. Обычно доза терапевтической композиции при непрерывной перфузии должна быть эквивалентной дозе, даваемой путем одной или множественных инъекций, с учетом периода времени, в течение которого вводятся инъекции. Считается, однако, что более высокие дозы могут быть достигнуты при перфузии.
1. Протокол лечения
Два первичных подхода предложены авторами для применения монотерпеновых/тритерпеновых соединений либо индивидуально, либо в сочетанной терапии. Первый представляет собой применение при метастазирующем раке либо у больных, которые не получали до этого химио-, радио- или биологической терапии, либо у больных, ранее не подвергавшихся лечению. Больных необходимо лечить с помощью системного введения, которое представляет собой внутривенное, подкожное, пероральное введение, или с помощью внутриопухолевой инъекции. Вводимая(ые) терапевтическая(ие) доза(ы) должна предпочтительно содержать между 10 и 25 мг монотерпеновых/тритерпеновых композиций на 1 кг массы тела больного в день, включая приблизительно 13, 16, 19 и 22 мг/кг/день. Альтернативно, больного можно лечить одной или более терапевтическими композициями, включающими в себя приблизительно от 1 мг/кг/день монотерпеновых/тритерпеновых композиций до приблизительно 100 мг/кг/день, включая приблизительно 3, 6, 9, 12, 15, 18, 21, 28, 30, 40, 50, 60, 70, 80 и 90 мг/кг/день монотерпеновых/тритерпеновых композиций.
Курс лечения обычно состоит из ежедневного лечения в течение минимум восьми недель или одной инъекции в неделю в течение минимально восьми недель. По выбору врача режим может быть продлен по той же схеме, пока развитие опухоли прогрессирует или если наблюдается отсутствие ответа.
Другим применением соединений согласно изобретению является применение у лечащихся больных, которых привели в свободное от клинических проявлений заболевания состояние с помощью хирургической операции, химиотерапии и/или лучевой терапии. Вспомогательная терапия должна вводиться в том же самом режиме, что и описанный выше, в течение минимум одного года для предотвращения ремиссии заболевания.
2. Предотвращение рака
Другим применением соединений и смесей монотерпеновых/тритерпеновых гликозидных композиций является предотвращение рака у групп высокого риска. Такие больные (например, те, у которых определена генетическая предрасположенность к опухолям, таким как рак молочной железы, рак ободочной кишки, рак кожи и другие) должны лечиться путем перорального приема (опухоли желудочно-кишечного тракта), местного или накожного применения (кожные) или путем системного введения с минимальным периодом в течение одного года и предпочтительнее более длительное время для установления предотвращения рака. При данном применении должны быть включены больные и должны быть хорошо определены пренеопластические повреждения, такие как колоректальные полипы или другие предраковые повреждения кожи, молочной железы, легкого или других органов. Часто это бывают больные с воспалительными заболеваниями, которые представляют собой предраковые воспалительные состояния, например, больные с такими состояниями как эзофагит Барретта, воспалительное заболевание кишечника, хронический панкреатит, хронический простатит, семейный полипоз или лучевой кератоз.
3. Клинический протокол
Клинический протокол описан здесь для облегчения лечения рака с применением тритерпеновых соединений согласно изобретению. В соответствии с данным протоколом должны быть отобраны больные, имеющие гистологическое доказательство наличия рака, например рака яичников, рака поджелудочной железы, рака почки, рака простаты, легкого или мочевого пузыря. Больные могут, но необязательно, ранее подвергаться химио-, лучевой или генной терапии. Оптимально, чтобы больные имели достаточную функцию костного мозга (определяемую как абсолютное число периферических гранулоцитов > 2000/мм3 и число тромбоцитов 100000/мм3), достаточную функцию печени (билирубин ≤ 1,5 мг/дл) и достаточную функцию почек (креатинин < 1, 5 мг/дл).
Протокол предусматривает введение единичной дозы путем внутриопухолевой инъекции фармацевтической композиции, содержащей приблизительно от 10 до 25 мг тритерпеновых соединений на 1 кг массы тела больного. Для опухолей ≥ 4 см вводимый объем должен составлять 4-10 мл (предпочтительно 10 мл), в то время как для опухолей < 4 см должен применяться объем 1-3 мл (предпочтительно 3 мл). Множественные инъекции должны доставляться в виде единичной дозы в объеме 0,1-0,5 мл на расстоянии 1 см или более.
Курс лечения включает в себя приблизительно шесть доз, доставляемых в течение двух недель. По выбору врача схема может быть продолжена, шесть доз каждые две недели или не так часто (ежемесячно, раз в два месяца, ежеквартально и т.п.).
Когда больные подходят для хирургической резекции, опухоль надо обрабатывать, как описано выше, в течение по меньшей мере двух последовательных двухнедельных курсов лечения. В течение одной недели после завершения второго (или более, например, третьего, четвертого, пятого, шестого, седьмого, восьмого и т.д.) курса больной подвергается хирургической резекции. Перед закрытием разреза 10 мл фармацевтической композиции, содержащей описанные здесь тритерпеновые соединения, должно быть доставлено в место хирургической операции (операционное поле) и ей надо позволить оставаться в контакте в течение по меньшей мере 60 минут. Рану закрывают и туда помещают дренаж или катетер. На третий постоперационный день вводят дополнительные 10 мл фармацевтической композиции через дренаж и дают ей оставаться в контакте с операционным полем в течение по меньшей мере двух часов. Затем производят удаление путем отсасывания, и дренаж удаляют в клинически подходящее время.
4. Лечение искусственных и природных полостей тела
Одним из главных источников рецидивов рака являются остаточные, микроскопические очаги заболевания, которые остаются в месте первоначальной опухоли, как локально, так и регионально, после удаления опухоли. Кроме того, существуют аналогичные ситуации, когда в природных полостях тела рассеиваются микроскопические опухолевые клетки. Эффективное лечение таких микроскопических очагов заболевания должно обеспечить существенный прогресс в способах терапии.
Таким образом, в определенных осуществлениях раковая опухоль может быть удалена путем хирургической операции, что создает "полость". Как во время хирургической операции, так и после нее (периодически или постоянно) описанные здесь терапевтические композиции вводятся в полость тела. В сущности, это представляет собой "местное" лечение поверхности полости. Объем композиции должен быть достаточным для того, чтобы была уверенность, что вся поверхность полости находится в контакте с экспрессионным конструктом.
В одном осуществлении введение просто должно представлять собой инъекцию терапевтической композиции в полость, образованную в результате удаления опухоли. В другом осуществлении может быть желательным механическое нанесение с помощью губки, тампона или другого приспособления. Любой из данных подходов может быть использован после удаления опухоли, также как и во время первоначальной хирургической операции. В еще одном осуществлении катетер вводят в полость до закрытия места хирургического вмешательства. Полость может затем постоянно перфузироваться в течение желаемого периода времени.
При другой форме данного лечения "местное" применение терапевтической композиции направлено на природную полость тела, такую как рот, глотка, пищевод, гортань, трахея, плевральная полость, брюшная полость или пустые органные полости, включая полости мочевого пузыря, ободочной кишки или других висцеральных органов. В данной ситуации может быть или не быть существенным наличие первичной опухоли в полости. Лечение направлено на микроскопический очаг заболевания в полости, но среди прочего может также влиять на первичную опухолевую массу, если она не была предварительно удалена, или на пренеопластическое повреждение, которое может присутствовать в данной полости. Опять различные способы могут быть применены при влиянии "местного" применения в данных висцеральных органах или на поверхность полостей. Например, на ротовую полость в глотке можно влиять просто путем орального продувания и полоскания растворами. Однако местное применение в гортани и трахее может требовать эндоскопической визуализации и местной доставки терапевтической композиции. Висцеральные органы, такие как мочевой пузырь или слизистая ободочной кишки, могут требовать постоянно присутствующих катетеров с инфузией или опять прямой визуализации цистоскопом или другим эндоскопическим приспособлением. Такие полости как плевральная и брюшная полость, могут быть доступны благодаря постоянно присутствующим катетерам или хирургическим подходам, которые создают доступ к данным областям.
(iv) Терапевтические наборы
Здесь также описаны терапевтические наборы, включающие в себя монотерпеновые/тритерпеновые композиции. Такие наборы должны обычно включать в себя в подходящих контейнерных устройствах фармацевтически приемлемый состав по меньшей мере одного монотерпенового/тритерпенового соединения. Наборы также могут содержать другие фармацевтически приемлемые составы, такие как те, которые содержат компоненты для направленной доставки монотерпенового/тритерпенового соединения к определенным областям тела больного, когда необходимо лечение, или любое одно или более лекарство, которые могут работать совместно с монотерпеновыми/тритерпеновыми соединениями, например химиотерапевтические агенты.
Наборы могут иметь одно контейнерное устройство, которое содержит монотерпеновые/тритерпеновые соединения с или без любых дополнительных компонентов, или они могут иметь отдельное контейнерное устройство для каждого желаемого агента. Когда компоненты набора предлагаются в одном или более жидком растворе, жидкий раствор представляет собой водный раствор, причем стерильный водный раствор особенно предпочтителен. Однако компоненты набора могут быть предоставлены в виде сухого порошка(ков). Когда реагенты или компоненты предлагаются в виде сухого порошка, порошок может быть воссоздан с помощью добавления подходящего растворителя. Предполагается, что растворитель также может предоставляться в другом контейнерном устройстве. Контейнерное устройство набора должно обычно включать в себя по меньшей мере один флакон, пробирку для тестирования, колбу, сосуд, шприц или другое контейнерное устройство, в которое может быть помещен и предпочтительно подходящим образом аликвотирован монотерпеновый/тритерпеновый гликозид и любой другой желаемый агент. Когда включены дополнительные компоненты, набор должен также обычно дополнительно содержать второй флакон или другой контейнер, в который они помещаются, позволяя вводить отдельные предназначенные дозы. Наборы также могут включать в себя второе/третье контейнерные устройства, в которых содержатся стерильный, фармацевтически приемлемый буфер или другой разбавитель.
Наборы могут также содержать средства для введения монотерпеновых/тритерпеновых композиций животному или больному, например одну или более игл или шприцев, или даже глазную пипетку, или другое такое подобное этому приспособление, из которого состав может быть введен животному или нанесен на пораженную болезнью область тела. Наборы согласно изобретению должны также обычно включать в себя приспособления для хранения сосудов или подобного этому и другой компонент в закрытой упаковке для коммерческой продажи, такой как, например, пластиковые контейнеры для инъекций или форм для разбивания, в которых размещаются и хранятся желаемые флаконы и другие приспособления.
V. Химиотерапевтические сочетания и лечение
В определенных осуществлениях согласно изобретению может быть желательным введение тритерпеновых композиций в сочетании с одним или более другими агентами, обладающими противоопухолевой активностью, включая химиотерапевтические средства, облучение, терапевтические белки или гены. Это может увеличить общую противоопухолевую активность, достигаемую лечением только соединениями согласно изобретению, или они могут быть использованы для предотвращения или борьбы с множественной устойчивостью опухоли к лекарствам.
Для применения соединений согласно изобретению в сочетании с введением второго химиотерапевтического агента можно просто вводить животному тритерпеновую композицию в сочетании со вторым химиотерапевтическим агентом способом, эффективным для получения в результате их сочетанного противоопухолевого действия у животного. Данные агенты могут, следовательно, быть предложены в эффективном количестве и в течение эффективного периода времени для достижения их сочетанного присутствия в сосудах опухоли и их сочетанного действия в опухолевом окружении. Для достижения данной цели тритерпеновая композиция и химиотерапевтические агенты могут вводиться животному одновременно либо в единой композиции, либо в виде двух отдельных композиций с применением разных путей введения.
Альтернативно, лечение тритерпеновой композицией может предшествовать или следовать за химиотерапевтическим агентом, облучением или белковой или генной терапией через интервалы, находящиеся в диапазоне от минут до недель. В тех осуществлениях, когда второй агент и тритерпеновая композиция вводятся животному отдельно, обычно следует быть уверенным, что не истек существенный промежуток времени между временем каждой доставки, так что дополнительный агент и тритерпеновая композиция должны все еще быть способными проявлять благоприятный совместный эффект на опухоль. В таких случаях предполагается, что опухоль должна контактировать с обоими агентами в пределах приблизительно от 5 минут до одной недели друг с другом и более предпочтительно в пределах приблизительно 12-72 часов друг с другом с временем задержки только приблизительно 24-48 часов, что наиболее предпочтительно. В некоторых ситуациях может быть желательным существенно продлить период времени лечения, когда требуется промежуток времени в несколько дней (2, 3, 4, 5, 6 или 7) или даже несколько недель (1, 2, 3, 4, 5, 6, 7 или 8) между соответствующими введениями. Также возможно, чтобы было желательным более чем одно введение либо тритерпенового гликозида, либо второго агента. Для достижения регрессии опухоли оба агента доставляются в объединенном количестве, эффективном для ингибирования ее роста, независимо от времени введения.
Различные агенты подходят для применения в раскрытых здесь способах комбинированной терапии. Химиотерапевтические агенты, рассматриваемые как примеры, включают в себя, например, этопозид (VP-16), адриамицин, 5-фторурацил (5-FU), камптотецин, актиномицин D, митомицин С и цисплатин (CDDP).
Как должно быть понятно специалисту в данной области техники, подходящие дозы химиотерапевтических агентов должны обычно быть близки к тем, которые уже применялись в клинической терапии, при которой химиотерапевтические агенты вводились индивидуально или в сочетании с другими химиотерапевтическими агентами. Только в качестве примера, могут быть использованы такие агенты как цисплатин и другие алкилирующие ДНК агенты. Цисплатин широко использовался для лечения рака с эффективными дозами, используемыми для клинического применения, 20 мг/м2 в течение 5 дней каждые три недели в течение суммарно трех курсов. Цисплатин не всасывается перорально и должен, следовательно, доставляться путем инъекций, внутривенных, подкожных, внутриопухолевых или внутрибрюшинных.
Дополнительные полезные агенты включают в себя соединения, которые вмешиваются в репликацию ДНК, митоз и расхождение хромосом. Такие химиотерапевтические агенты включают в себя адриамицин, также известный как доксорубицин, этопозид, верапамил, подофиллотоксин и тому подобное. Широко используемые в клинической практике для лечения неоплазм данные соединения вводятся путем болюсных инъекций внутривенно в диапазоне доз от 25-75 мг/м2 через 21-дневные интервалы для адриамицина, до 35-50 мг/м2 для этопозида внутривенно или в виде сдвоенной внутривенной дозы перорально.
Также могут быть использованы агенты, которые нарушают синтез и соответствие полинуклеотидных предшественников. Особенно пригодными являются агенты, которые подвергались интенсивному тестированию и легко доступны. Как таковые такие агенты, как 5-фторурацил (5-FU), предпочтительно используются неопластической тканью, что делает данный агент особенно пригодным для направленного воздействия на неопластические клетки. Несмотря на существенную токсичность, 5-FU применяют в широком спектре носителей, включая местные, с внутривенным введением в обычно применяемом интервале доз от 3 до 15 мг/кг/день.
Примеры химиотерапевтических агентов, которые полезны в связи с сочетанной терапией, перечислены в таблице 5. Каждый из перечисленных там агентов является примером и не предназначен для ограничения. В данном отношении специалиста в данной области техники направляют к "Remington's Pharmaceutical Sciences" 15th Edition, chapter 33, на конкретные страницы 624-652. Некоторые вариации дозировки должны обязательно присутствовать в зависимости от состояния субъекта, подвергаемого лечению. Человек, ответственный за введение, должен в любом случае определить подходящую дозу для индивидуального субъекта. Более того, для введения человеку препараты должны удовлетворять требованиям стерильности, пирогенности, общей безопасности и стандартам чистоты, как это требуется управлением по биологической стандартизации FDA.
Химиотерапевтические агенты, пригодные при неопластическом заболевании
Ифосфамид
Флоксуридин (фторид-оксиуридин; FUdR)
Тертипозид
Карбоплатин
Медроксипрогестерона ацетат
Мегэстрола ацетат
Этинилэстрадиол (доступны другие препараты)
Флуоксиместерон(доступны другие препараты)
Другие факторы, которые вызывают повреждение ДНК и интенсивно используются, включают в себя то, что обычно известно как γ-лучи или рентгеновские лучи, и/или направленную доставку радиоизотопов к опухолевым клеткам. Рассматриваются также другие формы факторов, повреждающих ДНК, такие как микроволны и УФ-облучение. Наиболее вероятно, что все данные факторы в широком диапазоне вызывают повреждения ДНК, предшественников ДНК, репликации и репарации ДНК, а также сборки хромосом и ее поддержания. Диапазон доз рентгеновских лучей колеблется в интервале от суточных доз от 50 до 200 рентген в течение продолжительных периодов времени (от 3 до 4 недель) до единичных доз от 2000 до 6000 рентген. Диапазоны доз для радиоизотопов широко варьируются и зависят от периода полужизни изотопа, силы и типа испускаемого излучения и захвата неопластическими клетками.
VI. Направленная противораковая терапия
Описанные здесь монотерпеновые/тритерпеновые соединения могут быть присоединены к одной или более молекулам, которые направляют соединения к специфическим клеткам, например воспалительным клеткам, опухолевым клеткам и т.п. Направленное действие выгодно тем, что может быть использовано для увеличения суммарных уровней лекарства в месте лечения, например в местах воспаления или местах опухолевого роста, в то же время при сведении к минимуму системной экспозиции с лекарством.
В целом в отношении обсуждаемых выше химиотерапевтических агентов допустимо, чтобы применение направляемого тритерпенового соединения могло быть использовано в сочетании со вторым агентом, таким как химиотерапевтический агент. Как тритерпен, так и второй агент будут направляться к одной и той же или к различным мишеням в пределах опухолевого окружения. Это должно давать в результате аддитивные, более высокие, чем аддитивные, или даже значительные синергические эффекты.
Примерами направляющих агентов, применяемых в сочетании с монотерпеновыми/тритерпеновыми соединениями являются те направляющие агенты, которые способны доставлять монотерпеновые/тритерпеновые молекулы к затронутой болезнью области, т.е. способны локализоваться в месте воспаления или в месте опухолевого роста. Сходно желательными должны быть те агенты, которые направляются в сосудистое ложе опухолевой области. Направленная доставка монотерпеновых/тритерпеновых гликозидных соединений конкретно рассматривается, как позволяющая достичь более высоких эффективных концентраций в областях, затронутых болезнью, без или со сведением к минимуму потенциальных побочных эффектов, которые могут наблюдаться при в некоторой степени более широком или системном распределении монотерпеновых/тритерпеновых соединений. Конкретно, направляющий агент может представлять собой антитело, направленное против клетки с воспалением.
(i) Направленная доставка к опухолевым клеткам и антитела
Таким образом, направленная доставка к воспалительным или предраковым, или опухолевым клеткам может быть осуществлена с применением биспецифического антитела, которое обладает областью, способной связываться с относительно специфическим маркером или антигеном опухолевой клетки. Например, специфическое ингибирование или уничтожение опухолевой клетки может быть достигнуто с помощью связывания конъюгата антитело- монотерпеновая/тритерпеновая композиция с опухолевой клеткой-мишенью.
Описаны многие так называемые "опухолевые антигены", любой из которых может быть применен в качестве мишени в связи с аспектами направленной доставки согласно изобретению. Большое количество примеров антигенов, ассоциированных с солидными опухолями, перечислено здесь ниже. Получение и применение антител против таких антигенов хорошо известно специалистам в данной области техники и конкретно раскрыто здесь. Примеры антител включают в себя те, которые локализованы в местах роста гинекологических опухолей (см., например, каталог ATCC): OC 125; OC 133; OMI; Mo v1; Mo v2; 3C2; 4C7; ID3; DU-PAN-2; F 36/22; 4F7/7A10; OV-TL3; B72.3; DF3; 2C8/2F7; MF 116; Movl8; CEA 11-H5; CA 19-9 (1116NS 19-9); H17-E2; 791T/36; NDOG2; H317; 4D5, 3H4, 7C2, 6E9, 2C4, 7F3, 2H11, 3E8, 5B8, 7D3, SB8; HMFG2; 3.14.A3; в местах роста опухоли молочной железы: DF3; NCRC-l1; 3C6F9; MBE6; CLNH5; MAC 40/43; EMA; HMFG1; HFMG2; 3.15.C3; M3, M8, M24; M18; 67-D-11; D547Sp, D75P3, H222; анти-EGF; LR-3; TA1; H59; l0-3D-2; HmABl,2; MBR 1,2,3; 24·l7·l; 24·l7·2 (3E1·2); F36/22.M7/105; C11, G3, H7; B6·2; Bl·l; Cam 17·l; SM3; SM4; C-Mul (566);4D5 3H4, 7C2, 6E9, 2C4, 7F3, 2H11, 3E8, 5B8, 7D3, 5B8; OC 125; MO v2; DU-PAN-2; 4F7/7A10; DF3; B72·3; cccccCEA 11; H17-E2; 3·14·A3; F023C5; в местах роста опухоли прямой кишки: B72·3; (17-1A) 1083-17-1A; CO17-1A; ZCE-025; AB2; HT-29-15; 250-30.6; 44X14; A7; GA73·3; 791T/36; 28A32; 28.19.8; X MMCO-791; DU-PAN-2; ID3; CEA 11-H5; 2C8/2F7; CA-19-9 (1116NS 19-9); PR5C5; PR4D2; PR4D1; в местах роста меланомы: 4·1; 8·2 M17; 96·5; 118·1, I33·2, (113.2); L1, L10, R10(R19); I12;K5; 6·1; R24; 5·1; 225.28S; 465.12S; 9·2·27; F11; 376.96S; 465.12S; 15·75; 15·95; Mel-14; Mel-12; Me3-TB7; 225.28SD; 763.24TS; 705F6; 436910; M148; в местах роста опухолей желудочно-кишечного тракта: ID3; DU-PAN-2; OV-TL3; B72·3; CEA 11-H5; 3·l4·A3; C COLI; CA-19-9 (1116NS 19-9) и CA50; OC125; в местах роста опухолей легких: 4D5 3H4, 7C2, 6E9, 2C4, 7F3, 2H11, 3E8, 5B8, 7D3, SB8; MO v2; B72·3; DU-PAN-2; CEA 11-H5; MUC 8-22; MUC 2-63; MUC 2-39; MUC 7-39; и в местах роста других опухолей: PAb 240; PAb 246; PAb 1801; ERIC·1; M148; FMH25; 6·1; CA1; 3F8; 4F7/7A10; 2C8/2F7; CEA 11-H5.
Другой способ определения и направленной доставки к опухоли заключается в характеристике самой опухолевой клетки, а не в описании биохимических свойств антигена, экспрессируемого клеткой. Известен ряд примеров опухолевых клеточных линий, и они могут быть использованы для получения направляющих агентов. Например, целые клетки или гомогенаты клеток из известных опухолевых линий могут быть использованы для получения противоопухолевых антител для направленной доставки в относящиеся к этому типы опухолей. Сходно, такие опухолевые клеточные линии могут найти применение в осуществлении различных тестов in vitro. В данном отношении специалиста в данной области техники отсылают к каталогу ATCC с целью выяснения примеров линий опухолевых клеток человека, которые имеются в свободном доступе (из каталога ATCC). Примеры клеточных линий включают в себя


Можно справиться в каталоге ATCC за любой последующий год для идентификации других подходящих клеточных линий. Также, если желателен особый клеточный тип, средства для получения таких клеток и/или их постоянно доступный источник должны быть известены специалистам в данной области техники. Анализ научной литературы должен, таким образом, легко выявить подходящий источник клеток для любого клеточного типа опухоли, в отношении которого желательна направленная доставка.
Как объяснялось выше, антитела представляют собой простое средство для узнавания опухолевого антигена-мишени. Известно большое число антител, которые направлены против антигенов солидных опухолей. Определенные пригодные противоопухолевые антитела перечислены выше. Однако, как известно специалистам в данной области техники, некоторые из перечисленных антител не будут иметь подходящих биохимических свойств или могут быть недостаточно специфичными для опухоли, чтобы их использовать в терапевтических целях. Примером является MUC8-22, которое узнает цитоплазматический антиген. Антитела, подобные этим, обычно используются только для исследовательских целей, таких как модельные системы или тесты для скрининга.
Вообще говоря, антитела для применения в данных аспектах согласно изобретению должны предпочтительно узнавать антигены, которые доступны на клеточной поверхности и которые предпочтительно или специфично экспрессируются клетками с воспалением. Такие антитела должны также предпочтительно проявлять такие свойства, как высокое сродство, такое как характеризующееся Kd < 200 нМ и предпочтительно < 100 нМ, и не должны демонстрировать существенного взаимодействия с жизненно важными нормальными тканями, такими как одна или более тканей, выбранных из сердца, почек, мозга, печени, костного мозга, ободочной кишки, молочной железы, простаты, щитовидной железы, желчного пузыря, легкого, надпочечников, мышцы, нервных волокон, поджелудочной железы, кожи или другого жизненно важного органа или ткани организма человека. "Жизненно важные" ткани, которые являются наиболее важными для целей согласно изобретению с точки зрения низкого взаимодействия, включают в себя сердце, почку, ткани центральной и периферической нервной системы и печень. Применяемый здесь термин "существенное взаимодействие" относится к антителу или фрагменту антитела, которые при нанесении на конкретную ткань в условиях, подходящих для иммуногистохимии, не будут проявлять либо никакого окрашивания, либо незначительное окрашивание только с несколькими позитивными клетками, разбросанными среди поля главным образом негативных клеток.
Особенно многообещающими антителами, предназначенными для применения при терапии рака, описанной выше, являются те, которые имеют высокую избирательность в отношении солидной опухоли. Например, антитела, связывающиеся с TAG 72 и HER-2 протоонкогенным белком, который избирательно выявляется на поверхности многих видов рака молочной железы, легкого и прямой кишки (Thor et al., 1986; Colcher et al., 1987; Shepard et al., 1991); Mov18 и OV-TL3 и антитела, которые связываются с основным белком муцина молока и жировой глобулой человеческого молока (Miotti et al., 1985; Burchell et al., 1983); и антитело 9.2.27, которое связывается с антигенами меланомы высокой Mr (Reisfeld et al., 1982). Дополнительными пригодными антителами являются те, которые направлены против фолатсвязывающего белка, который, как известно, гомогенно экспрессируется почти во всех карциномах яичника; те, которые направлены против семейства erb онкогенов, которые гиперэкспрессируются в сквамозно-клеточных карциномах и в большинстве глиом; и другие антитела, известные как объект текущих предклинических и клинических испытаний.
Антитела B3, KSI/4, CC49, 260F9, XMMCO-791, D612 и SM3, как считается, особенно пригодны для применения в клинических осуществлениях после стандартного предклинического испытания, обычно практикуемого в данной области техники. B3 (патент США 5242813; Brinkmann et al., 1991) имеет No. поступления в ATCC HB 10573; KS1/4 может быть получен, как описано в патенте США 4975369, и D612 (патент США No. 5183756) имеет No. поступления в ATCC HB 9796.
Другой способ определения связанной с опухолью мишени заключается в характеристике опухолевой клетки, а не в описании биохимических свойств антигена, экспрессируемого клеткой. Соответственно, авторы предполагают, что любое антитело, которое предпочтительно связывается с опухолевой клеткой или воспалительной клеткой, может быть применено в виде направляющего компонента монотерпен/тритерпен-направляющего конъюгата. Предпочтительное связывание с опухолевой клеткой опять основывается на антителе, которое имеет высокое сродство к опухолевой клетке и не характеризуется существенным взаимодействием с жизненно важными нормальными клетками или тканями, как указано выше.
В изобретении предлагается также несколько средств для выработки антитела для применения в направленной доставке монотерпеновых/тритерпеновых гликозидов к воспалительным клеткам или опухолевым клеткам, как здесь описано. Для выработки антитела, специфичного в отношении клетки с воспалением, надо иммунизировать животное композицией, включающей в себя антиген воспалительной клетки или антиген опухолевой клетки и, как описано более полно ниже, отобрать полученное в результате антитело с подходящей специфичностью. Композиция для иммунизации может содержать очищенный или частично очищенный препарат любого из антигенов, перечисленных выше; такую композицию, как препарат мембран, обогащенный любым из антигенов, перечисленных выше; любую из клеток, перечисленных выше, или смесь, или популяцию клеток, которые включают в себя любой из клеточных типов, перечисленных выше.
Конечно, не взирая на источник антитела, практикой изобретения при лечении человека должна быть предпочтительная уверенность заранее в том, что место воспаления или опухолевого роста экспрессирует антиген, выбранный в конечном счете. Это достигается с помощью средств довольно простого анализа, включающего тестирование антигенов в образце ткани, например, отобранной при хирургической биопсии, или возможно тестирование циркулирующего в результате шеддинга антигена. Это может быть легко осуществлено в тесте иммунологического скрининга, такого как ИФА (иммуноферментный анализ на иммуносорбенте), в котором тестируется связывающее сродство антител из "банка" гибридом в отношении взаимодействия с опухолью. Антитела, проявляющие подходящую селективность и аффинность, затем отбираются для получения биспецифических антител согласно изобретению.
На основе хорошо известного феномена перекрестной реактивности, предполагается, что пригодные антитела могут быть получены в результате протоколов иммунизации, в которых исходно применяемые антигены происходили от животного, такого как мышь или примат, в дополнение к тем, в которых исходные антигены получали из клетки человека. Когда применяют антигены, происходящие от человека, их можно получить из клеточной линии опухоли человека или их можно получить при получении биологического образца от конкретного больного, лечение которого предполагается. Конечно, известны способы выработки антител, которые "сделаны на заказ" к опухоли больного (Stevenson et al., 1990), и они рассматриваются как применяемые в связи с настоящим изобретением.
1. Способы получения антител
Как указывалось, антитела могут найти применение в конкретных осуществлениях согласно изобретению. Например, могут быть получены антитела, которые специфичны в отношении конкретной области больного или конкретного типа ткани. Данные антитела могут затем быть конъюгированы с тритерпеновым соединением согласно изобретению, в результате чего создается специфическая направленная доставка тритерпеновых соединений к ткани, против которой направлено антитело. Примером осуществления такого антитела является такое, которое связывается с опухолевой клеткой. В предпочтительном осуществлении изобретения антитело является моноклональным антителом. Средства для получения и характеристики моноклональных и поликлональных антител хорошо известны в данной области техники и конкретно раскрыты здесь ниже (см., например, Howel and Lane, 1988).
Вкратце, поликлональное антитело получают путем иммунизации животного иммуногеном, включающим в себя желаемый антиген-мишень, и сбора антисывороток от иммунизированного животного. Может быть использован широкий диапазон видов животных для получения антисывороток. Обычно животное, применяемое для продукции антисывороток, представляет собой животное, не являющееся человеком, включая кроликов, мышей, крыс, хомячков, свиней или лошадей. Благодаря относительно большому объему крови у кроликов, выбор кролика является предпочтительным для получения поликлональных антител.
Антитела, как поликлональные, так и моноклональные, специфичные в отношении изоформ антигена, могут быть получены с применением общепринятых способов иммунизации, как это обычно должно быть известно специалистам в данной области техники. Композиция, содержащая антигенные эпитопы конкретных клеточных типов, или, альтернативно, соединения согласно изобретению, может быть использована для иммунизации одного или более экспериментальных животных, таких как кролик или мышь, которые затем будут поддерживаться для получения специфических антител против антигенов. После периода времени, необходимого для выработки антител, могут быть получены поликлональные антисыворотки просто путем взятия крови у животного и получения образцов сыворотки из цельной крови.
Предполагается, что моноклональные антитела согласно изобретению должны найти полезное применение в иммунохимических способах, которые могут быть применены для скрининга на присутствие тритерпеновых соединений согласно изобретению в видах, отличных от Acacia victoriae, или в других способах, в которых могут применяться антитела, специфичные в отношении конкретных антигенов. Как обсуждалось, пример осуществления с применением антител согласно изобретению включает в себя получение антител, направленных к специфическим для опухоли антигенам, присоединение антител к тритерпеновым соединениям согласно изобретению и лечение больных людей антиген-тритерпеновым конъюгатом, в результате чего тритерпеновые соединения согласно изобретению специфически направляются к опухолевым клеткам или другим клеткам, которые вовлечены в состояние, которое может быть излечено с помощью тритерпеновых соединений согласно изобретению. В целом, как поликлональные, так и моноклональные антитела против различных антигенов могут быть применены в различных осуществлениях изобретения. Например, они могут быть применены при очистке тритерпеновых соединений на аффинной колонке с антителом. Средства для получения и характеристики таких антител хорошо известны в данной области техники и раскрыты, например, в Harlow and Lane, 1988, раскрытие которых специально включено здесь в качестве ссылки во всей своей полноте.
Как хорошо известно в данной области техники, данная композиция может варьироваться по своей иммуногенности. Часто является необходимым, следовательно, стимулировать иммунную систему хозяина, что может быть достигнуто с помощью присоединения пептидного или полипептидного иммуногена к носителю. Предпочтительными носителями и их примерами являются гемоцианин моллюска фиссурелии (KLH) и бычий сывороточный альбумин (BSA). Другие альбумины, такие как овальбумин, мышиный сывороточный альбумин или кроличий сывороточный альбумин, также могут быть использованы в качестве носителей. Вещества для конъюгации полипептида с белком-носителем хорошо известны в данной области техники и включают в себя глутаральдегид, м-малеимидбенкоил-N-гидроксисукцинимидный сложный эфир, карбодиимид и бис-биазотизированный бензидин.
Как также хорошо известно в данной области техники, иммуногенность конкретной иммуногенной композиции может быть увеличена путем применения неспецифических стимуляторов иммунного ответа, известных как адъюванты. Предпочтительные адъюванты и их примеры включают в себя полный адъювант Фрейнда (неспецифический стимулятор иммунного ответа, содержащий убитые Micobacterium tuberculosis), неполные адъюванты Фрейнда и адъювант на основе гидроксида алюминия.
Количество композиции иммуногена, применяемое для продукции поликлональных антител, варьируется в зависимости от природы иммуногена, а также от животного, используемого для иммунизации. Для введения иммуногена могут быть использованы разнообразные способы (подкожное, внутримышечное, внутрикожное, внутривенное и внутрибрюшинное введение). Продукция поликлональных антител может прослеживаться с помощью отбора образцов крови у иммунизированного животного в разные периоды времени после иммунизации. Может быть также дана вторая, бустерная, инъекция. Процесс бустерного введения и титрования повторяют до тех пор, пока не будет достигнут подходящий титр. Когда получают желаемый уровень иммуногенности, у иммунизированного животного можно получить кровь, и сыворотку выделить и хранить, и/или животное может быть использовано для выработки mAb.
mAb можно легко получить путем хорошо известных способов, таких как проиллюстрированные в патенте США 4196265, раскрытие которого специально включено здесь в качестве ссылки во всей своей полноте. Обычно данный способ включает в себя иммунизацию подходящего животного выбранной композицией иммуногена, например очищенным или частично очищенным специфичным для опухоли антигеном, полипептидом или пептидом или опухолевой клеткой. Композицию для иммунизации вводят с помощью способа, эффективного для стимуляции клеток, продуцирующих антитела. Предпочтительными животными являются грызуны, такие как мыши и крысы, однако также возможно применение клеток кроликов, овец или лягушек. Применение крыс может давать определенные преимущества (Goding, 1986), но мыши являются предпочтительными, причем наиболее предпочтительны мыши BALB/c, так как они наиболее часто применяются и обычно дают более высокий процент стабильных слияний.
После иммунизации соматические клетки, обладающие потенциалом в отношении выработки антител, конкретно В-лимфоциты (В-клетки) отбирают для применения в протоколе выработки mAb. Данные клетки могут быть получены из биопсийного материала селезенки, миндалин или лимфатических узлов или из образца периферической крови. Предпочтительными являются клетки селезенки и периферической крови, первые, потому что они являются богатым источником клеток, продуцирующих антитела, которые находятся в стадии делящихся плазмобластов, а последние, потому что периферическая кровь является легко доступной. Часто иммунизируется ряд животных и удаляется селезенка животного с самым высоким титром антител, и получают лимфоциты селезенки путем гомогенизации селезенки с помощью шприца. Обычно селезенка иммунизированной мыши содержит приблизительно от 5 х 107 до 2 х 108 лимфоцитов.
Продуцирующие антитела В-лимфоциты от иммунизированного животного затем сливают с клетками иммортализованной клеточной линии миеломы, обычно одного из тех же самых видов, что и животное, которое иммунизировали. Миеломные клеточные линии, подходящие для применения в способах слияния с получением гибридомы, предпочтительно не продуцируют антитела, характеризуются высокой эффективностью слияния и недостаточностью ферментов, которая делает их неспособными расти в определенных селективных средах, которые поддерживают рост только желаемых слитых клеток (гибридом).
Может быть использован любой номер линии миеломных клеток, как это известно специалистам в данной области техники. Например, когда иммунизированным животным является мышь, можно применять P3-X63/Ag8, P3-X63-Ag8.653, NS1/1.Ag4 1, Sp210-Ag14, FO, NSO/U, MPC-11, MPC11-X45-GTG 1.7 и S194/5XX0 Bul; в случае крыс можно применять R210.RCY3, Y3-Ag 1.2.3, IR983F и 4B210; и U-266, GM1500-GRG2, LICR-LON-HMy2 и UC729-6, все являются пригодными в связи со слияниями клеток (см., например, Goding, 1986; Campbell, 1984, и каталог ATCC).
Способы создания гибридом из антителопродуцирующих клеток селезенки или лимфатических узлов и миеломных клеток включают в себя смешивание соматических клеток с миеломными клетками в отношении 2:1, хотя отношение может варьироваться приблизительно от 20:1 до приблизительно 1:1, соответственно, в присутствии агента или агентов (химических или электрических), которые стимулируют слияние клеточных мембран. Описаны способы слияния с применением вируса Сендай (Kohler and Milstein, 1975; 1976), и способы с применением полиэтиленгликоля (ПЭГ), такого как 37% ПЭГ (об./об.) Gefter et al., (1977). Подходит также применение электрически индуцируемых способов слияния (Goding, 1986).
Способы слияния обычно дают жизнеспособные гибриды с низкой частотой около от 1 х 10-6 до 1 х 10-8. Однако это не создает проблемы, так как жизнеспособные слитые гибриды отличаются от родительских, неслитых клеток (конкретно неслитых миеломных клеток, которые должны продолжать нормально, неограниченно делиться) в результате культивирования в селективной среде. Селективная среда представляет собой обычно среду, содержащую агент, который блокирует синтез нуклеотидов de novo в среде тканевой культуры. Предпочтительными агентами и примерами являются аминоптерин, метотрексат и азасерин. Аминоптерин и метотрексат блокируют синтез de novo как пуринов, так и пиримидинов, в то время как азасерин блокирует только синтез пуринов. Когда применяют аминоптерин или метотрексат, среда дополняется гипоксантином и тимидином в качестве источника нуклеотидов (среда HAT). Когда применяют азасерин, среду дополняют гипоксантином.
Предпочтительной селективной средой является HAT. Только клетки, способные использовать имеющиеся нуклеотиды с применением обходных путей, способны выживать в среде HAT. Клетки миеломы имеют дефект ключевых ферментов обходного пути, например гипоксантинфосфорибозилтрансферазы (HPRT), и они не могут выжить. В-клетки могут использовать данный путь, но они имеют ограниченную продолжительность жизни в культуре и обычно умирают в течение приблизительно двух недель. Следовательно, клетки, которые могут выжить в селективных средах, представляют собой только гибриды, образованные из клеток миеломы и В-клеток.
Такое культивирование дает популяцию гибридом, из которых отбираются специфические гибридомы. Обычно отбор гибридом выполняют путем культивирования клеток при разнесении по одному клону в лунку планшетов для микротитрования с последующим тестированием супернатантов индивидуальных клонов (после приблизительно от двух до трех недель) на желаемое взаимодействие. Тест должен быть чувствительным, простым и быстрым, таким как радиоиммунный анализ, иммуноферментный анализ, анализ цитотоксичности, анализ бляшек, дот-иммуносвязывающий анализ и тому подобное.
Выбранные гибридомы должны быть затем серийно разведены и клонированы с получением индивидуальных клеточных линий, продуцирующих антитела, причем клоны можно затем размножать неограниченно долго для получения mAb. Клеточные линии могут быть применены для продукции mAb в двух основных вариантах. Образец гибридомы может быть введен (часто в брюшную полость) гистосовместимому животному, типа такого, которое было применено для получения соматических и миеломных клеток для первоначального слияния. У животного после инъекции развиваются опухоли, секретирующие специфичное моноклональное антитело, продуцируемое слитым клеточным гибридом. Жидкости организма животного, такие как сыворотка или асцитная жидкость, могут затем откачиваться для получения mAb в высокой концентрации. Индивидуальные клеточные линии могут также культивироваться in vitro, когда mAb естественным образом секретируются в культуральную среду, из которой они могут быть легко получены в высоких концентрациях. mAb, полученные любым способом, могут быть дополнительно очищены, если это желательно, с применением фильтрации, центрифугирования и различных хроматографических способов, таких как ВЭЖХ или аффинная хроматография.
(iii) Дополнительные опухолевые клетки-мишени и связывающие лиганды
В дополнение к применению антител для направленной доставки монотерпеновых/тритерпеновых соединений к месту опухолевого роста путем связывания с антигеном опухоли могут применяться другие лиганды. Для антигенов опухолей, которые представляют собой гиперэкспрессируемые рецепторы (например, эстрогенный рецептор, рецептор EGF) или мутантные рецепторы, в качестве направляющих агентов могут быть применены соответствующие лиганды.
Аналогичным образом в отношении лигандов рецепторов эндотелиальных клеток, среди них могут быть компоненты, которые специфично или предпочтительно связываются с опухолевыми клетками. Например, если опухолевый антиген представляет собой гиперэкспрессируемый рецептор, опухолевая клетка может быть покрыта специфическим лигандом in vivo. Следовательно, лиганд может затем быть мишенью либо для антитела против лиганда, либо представлять мишень сам в комплексе с рецептором. Конкретными примерами направляющих агентов данного типа являются антитела против лигандов TIE-1 или TIE-2, антитела против фактора 4 тромбоцитов и связывающий белок адгезии лейкоцитов.
(iv) Токсины
Для определенного применения предполагается, что вторыми терапевтическими агентами, применяемыми в сочетании с описанными здесь монотерпеновыми/тритерпеновыми соединениями, должны быть фармакологические агенты, конъюгированные с антителами или ростовыми факторами, особенно цитотоксические агенты или агенты, негативно действующие на клетку другим способом, обладающие способностью уничтожать клетку или подавлять рост или клеточное деление эндотелиальных клеток. В целом изобретение охватывает применение любого фармакологического агента, включая и в дополнение к описанным здесь монотерпеновым/тритерпеновым соединениям, который может быть конъюгирован с направляющим агентом, предпочтительно антителом, и доставлен в активной форме к опухолевым клеткам-мишеням. Примеры агентов, негативно действующих на клетку, включают в себя химиотерапевтические агенты, радиоизотопы, а также цитотоксины. В случае химиотерапевтических агентов, такие агенты как стероидный гормон; антиметаболит, такой как цитозинарабинозид, фторурацил, метотрексат или аминоптерин; антрациклин; митомицин С; алкалоид барвинка; демеколцин; этопозид; митрамицин; или противоопухолевый алкилирующий агент, такой как хлорамбуцил или мелфалан, должны быть особенно предпочтительными. Другие осуществления могут включать в себя такие агенты, как цитокин, ростовой фактор, бактериальный эндотоксин или липидную А часть бактериального эндотоксина. В любом случае предполагается, что такие агенты, как данные, могут, если это желательно, успешно присоединяться совместно с монотерпеновыми/тритерпеновыми соединениями к направляющим агентам, предпочтительно к антителу, таким способом, который должен обеспечивать их направленную доставку, интернализацию, высвобождение или презентацию компонентам крови в области клеток-мишеней, как это требуется, при применении известных способов конъюгации (см., например, Ghose et al., 1983, и Ghose et al., 1987).
Разнообразные химиотерапевтические и другие фармакологические агенты в настоящее время успешно присоединяются к антителам и проявляют свою фармакологическую функцию (см., например, Vaickus et al., 1991). Примеры исследованных противоопухолевых агентов включают в себя доксорубицин, дауномицин, метотрексат, винбластин и различные другие (Dillman et al., 1988; Pietersz et al., 1988). Более того, описано присоединение других агентов, таких как неокарзиностатин (Kimura et al., 1983), макромицин (Manabe et al., 1984), тренимон (Ghose, 1982) и α-аманитин (Davis & Preston, 1981). Здесь описаны конкретные средства для получения конъюгатов между монотерпеновыми/тритерпеновыми соединениями и подходящие направляющие молекулы конкретно раскрыты здесь выше.
VII. Местные композиции и косметические средства
Определенные аспекты изобретения касаются местных композиций, которые включают в себя описанные здесь соединения, а также способы их применения. С помощью включения соединений согласно изобретению в местные композиции могут быть получены благоприятные физиологические активности соединений в отношении профилактики и/или лечения. Например, соединения согласно изобретению, как здесь показано, снижают различные типы клеточного повреждения, включая, но не ограничиваясь этим, различные типы окислительного повреждения, повреждение, опосредуемое канцерогенами и повреждение, опосредуемое УФ.
В частности, одним способом применения соединений согласно изобретению является применение в качестве активных ингредиентов в местных композициях, включая косметические, предназначенных для лечения старения кожи. Специфические изменения кожи, которые могут быть результатом старения и которые могут быть предотвращены или подвержены лечению в соответствии с изложенным здесь, индуцированы ли они в связи с возрастом или в связи с внешними причинами, включают в себя морщины, кожистость, пожелтение, слабость, шершавость, сухость, появление пятен (гиперпигментацию) и различные типы предракового роста, которые часто являются субклиническими; дегенерацию микрососудистой системы; вялость и развитие морщин, частично обусловленное снижением поперечных сшивок коллагена, аккумуляцией глюкозаминогликанов (основного вещества) и инсоляционным эластозом (агрегацией эластина); уплощение сети пучков; ограниченный регенеративный оборот эпидермиса, связанный с дефектным развитием ороговевшего слоя (нарушенным ороговением), ведущим к высушиванию кожи, к шершавости кожи и к растрескиванию кожи; дефектную регуляцию деления клеток (пролиферации) и созревания клеток (дифференциации) в эпидермисе, что ведет к клеточной атипии и атрофии, а также потере полярности; местную гипер- и гипопигментацию и необычную пигментацию (старческие пятна). Следовательно, авторы конкретно рассматривают лечение любых таких состояний соединениями, предлагаемыми в изобретении.
Как таковые, определенные аспекты изобретения касаются способов восстановления повреждения кожи или его результатов. Например, в изобретении предлагаются способы профилактического и/или терапевтического утолщения кожи для снижения атрофии кожи, для регуляции эластичности кожи и для регуляции видимой и/или ощущаемой неоднородности структуры кожи, включая тонкие линии, морщины, увеличенные поры, шершавость, сухость и другие виды неоднородности структуры кожи, связанные со старением кожи; причем способы включают в себя нанесение на кожу, нуждающуюся в этом, безопасного и эффективного количества местной композиции, включающей в себя соединения согласно изобретению. Предлагаются также способы снижения гиперпигментации кожи млекопитающих, включающие в себя нанесение на кожу, нуждающуюся в таком лечении, безопасного и эффективного количества композиции по уходу за кожей, включающей в себя соединение согласно изобретению. Такая гиперпигментация может быть результатом не связанного с меланином нарушения окрашивания кожи.
Настоящее изобретение также относится к продуктам для лечения и профилактики старения кожи, в частности кожи, которая постарела в результате действия света, и кожи, которая хронологически является старой в результате действия эндогенных механизмов, а также для лечения повреждения, вызванного другими экзогенными факторами. В целом желательно вводить соединения согласно изобретению в косметических или дерматологических местных составах, разработанных для нанесения на кожу.
(i) Защита кожи
Описанные здесь местные композиции должны найти применение в защите кожи от различных агентов, способных повреждать кожу. В частности, соединения согласно изобретению, как здесь показано, предотвращают повреждение от различных форм экзогенных повреждений, включая повреждение, вызываемое облучением, канцерогенами и окислительным стрессом. Таким образом, один аспект изобретения касается предотвращения повреждения кожи или ее восстановления после повреждения, вызываемого экзогенным агентом, включающего в себя контактирование кожи с безопасным и эффективным количеством композиций согласно изобретению.
Так как соединения согласно изобретению, как здесь показано, активируют элементы, чувствительные к антиоксидантам (ARE), соединения должны найти особое применение для предотвращения или сведения к минимуму повреждения, вызываемого активными формами кислорода, радикалами, свободными радикалами, окислительным стрессом, окислительным повреждением и их сочетаниями. Многочисленные болезнетворные процессы относятся к побочной реакции организма на присутствие повышенных уровней активных форм кислорода (АФК), как здесь описывается, и, таким образом, могут лечиться композициями согласно изобретению.
Переносимые по воздуху промышленные загрязнения и загрязнения в результате нефтехимических процессов, такие как озон, оксид азота, радиоактивные частицы и галогенизированные углеводороды, также индуцируют окислительное повреждение кожи, легких, желудочно-кишечного тракта и других органов. Радиационные отравляющие агенты из промышленных источников, включая утечки из ядерных реакторов и воздействие ядерного оружия, представляют собой другие источники облучения и повреждения радикалами. Другие пути экспозиции могут осуществляться в результате жизни или работы в непосредственной близости от источников электромагнитного излучения, таких как электростанции и линии высокого напряжения, рентгеновские аппараты, ускорители частиц, радарные антенны, радиоантенны и тому подобное, а также в результате использования продуктов и приспособлений электроники, которые испускают электромагнитное излучение, таких как телевизор и мониторы компьютеров. Таким образом, желательна защита клеток включая клетки кожи, от таких этиологических агентов и это составляет часть настоящего изобретения.
(ii) Местные и косметические составы
Сочетания активных веществ или активные вещества, предлагаемые в настоящем изобретении, могут присутствовать в местных составах в количествах приблизительно от 0,001 до приблизительно 99% по массе и, например, также в количествах приблизительно от 0,001 до приблизительно 50% по массе, в каждом случае на основе суммарной массы составов. Сочетания активных веществ или активные вещества в соответствии с изобретением могут предпочтительно присутствовать в местных составах в количествах от 0,01 до 10% по массе, в частности в количествах от 0,1 до 1% по массе, в каждом случае на основе суммарной массы составов. Соотношение массы двух компонентов в сочетаниях может варьироваться в широких пределах, например их применяют в соотношении от 1:100 до 100:1, предпочтительно в соотношении от 1:10 до 10:1. Компоненты могут также присутствовать, например, в соотношении масс от 1:2 до 2:1 или 1:1.
В целом для местных растворов желательными ингредиентами могут быть смягчающие или смазывающие носители, такие как маслянистые вещества, которые помогают увлажнять кожу. Применяемый здесь термин "смягчающий" должен быть понят как относящийся к композиции нераздражающего характера в целом. Это значит, что природа носителя и количество в ней активного соединения должны быть выбраны так, чтобы обеспечить дозу ниже порога раздражения для местного нанесения. Уровни смягчающих агентов могут варьироваться в диапазоне от 0,5 до 50% или более, предпочтительно между 5 и 30%, от суммарной массы композиции. Смягчающие агенты могут классифицироваться на такие общие химические категории как сложные эфиры, жирные кислоты и спирты, полиолы и углеводороды.
Сложные эфиры могут представлять собой моно- и диэфиры. Подходящие примеры диэфиров жирных кислот включают в себя дибутиладипат, диэтилсебацинат, дизиопропилдимерат и диоктилсукцинат. Подходящие эфиры жирных кислот с разветвленной цепью включают в себя 2-этилгексилмиристат, изопропилстеарат и изостеарилпальмитат. Подходящие эфиры трехосновных кислот включают в себя триизопропилтрилиноат и трилаурилцитрат. Подходящие эфиры жирных кислот с линейной цепью включают в себя лаурилпальмитат, миристиллактат и стеарилолеат. Предпочтительные сложные эфиры включают в себя кококаприлат/капрат (смесь кококаприлата и кококапрата), пропиленгликольмиристиловый эфир уксусной кислоты, диизопропиладипат и цетилоктаноат.
Среди полиолов, которые могут служить смягчающими агентами, находятся алкилполигидроксильные соединения с линейной и разветвленной цепью. Например, предпочтительными являются пропиленгликоль, сорбит и глицерин. Пригодными могут быть также полимерные полиолы, такие как полипропиленгликоль и полиэтиленгликоль. Бутилен- и пропиленгликоль также особенно предпочтительны в качестве усилителей проницаемости.
Для составов, разработанных для применения в зимнее время и у субъектов с очень сухой кожей, может быть желательной наличие основы мази (без воды). Примерами подходящих основ мазей являются вазелин, вазелин плюс летучие кремнийорганические соединения, ланолин и вода в масляных эмульсиях, таких как Eucerin (Beiersdorf). В теплую погоду и часто для более молодых людей может быть желательным применение основ в виде масла в водной эмульсии (крема). Примерами подходящих основ крема являются крем Nivea (Beiersdorf), кольдкрем (USP), Purpose Cream (Johnson & Johnson), гидрофильная мазь (USP) и Lubriderm (Warner-Lambert).
Местные составы или композиции в соответствии с изобретением, содержащие сочетания и активные вещества в соответствии с изобретением, включают в себя все обычно применяемые формы, например кремы (W/O, O/W или W/O/W), гели, лосьоны или молочко. Местные составы в соответствии с изобретением могут быть составлены в виде жидкости, мази или твердых составов, например в виде водных или спиртовых растворов, водных суспензий, эмульсий, мазей, кремов, масел, порошков или карандашей. В зависимости от желаемого состава активные вещества могут быть включены в фармацевтическую и косметическую основы для местного нанесения, которые включают в себя в качестве дополнительных компонентов, например, компоненты масел, жиры и воск, эмульгаторы, анионные, катионные, амфолитные, цвиттерионные и/или неионные поверхностно-активные вещества, низшие одно- или многоатомные спирты, воду, консерванты, забуферивающие вещества, загустители, ароматизаторы, окрашивающие вещества и агенты, снижающие прозрачность. Активные вещества в соответствии с изобретением могут быть также удачно применены в трансдермальных терапевтических системах.
Дополнительно может быть выгодным добавление к составам антиоксидантов (например, альфа-токоферола, витамина Е и С), флавонов, флавоноидов, имидазолов, альфа-гидроксикарбоновых кислот (например, яблочной кислоты, гликолевой кислоты, глюконовой кислоты, салициловой кислоты и их производных) и/или агентов, комплексирующихся с железом (например, ЭДТА и альфа-гидрокси-жирных кислот), и/или известных фильтров, защищающих от УФ-излучения, в количествах, например, от 0,1 до 10% по массе, для того чтобы быть уверенными в стабильности активных соединений, чувствительных к окислению.
Также может быть выгодным добавление к составам, в частности, от 0,01-10% по массе веществ или сочетания веществ аэробного энергетического метаболизма клетки, например агентов клеточного транспорта энергии (таких как креатин, гуанин, гуанозин, аденин, аденозин, никотин, никотинамид или рибофлавин), коферментов (например, пантотеновой кислоты, пантенола, липоевой кислоты или никотиновой кислоты), вспомогательных факторов (например, L-карнитина, долихола или уридина), субстратов (например, гексоз, пентоз или жирных кислот) и продуктов промежуточного метаболизма (например, лимонной кислоты или пирувата) и/или глутатиона.
Также может быть выгодным добавление к составам усилителей проникновения, в частности олеиновой кислоты, цис-6-гексадеценоевой кислоты или пальмитолеиновой кислоты. Усилители проникновения могут присутствовать в составах в количествах от 0,01% по массе до 1,0% по массе.
Выгодно, чтобы составы в соответствии с изобретением могли дополнительно включать в себя дополнительные вещества, которые поглощают УФ излучение в UVA- и/или в UVB-диапазоне, причем суммарное количество фильтрующих веществ составляет, например, от 0,1% по массе до 30% по массе, предпочтительно от 0,5 до 10% по массе, в частности от 1,0% до 6,00% по массе, на основе суммарной массы составов, для того чтобы косметические составы обеспечивали защиту кожи от всей области ультрафиолетового облучения. Они могут быть также использованы как солнцезащитные для кожи. В составах вещества, поглощающие УФ, могут действовать как антиоксиданты в отношении активных веществ.
Если композиции в соответствии с изобретением включают в себя вещества, действующие как фильтры UVB, они могут быть растворимыми в масле или водорастворимыми. Примерами растворимых в масле фильтров UVB, которые благоприятны в связи с изобретением, являются производные 3-бензилиденкамфоры, предпочтительно 3-(4-метилбензилиден)камфора и 3-бензилиденкамфора. Примерами благоприятных водорастворимых фильтров UVB являются соли 2-фенилбензимидазол-5-сульфокислоты, такие как ее натриевая, калиевая или ее триэтаноламмонийная соль, и сульфокислота сама по себе. Также потенциально находящими применение являются производные PABA, циннамата и салицилата. Например, могут быть применены октилметоксициннамат и 2-гидрокси-4-метоксибензофенон (также известный как оксибензон). Октилметоксициннамат и 2-гидрокси-4-метоксибензофенон имеются в продаже под торговыми марками Parsol MCX и бензофенон-3, соответственно.
Может быть также выгодным сочетание комбинаций активных веществ в соответствии с изобретением с фильтрами UVA, которые до настоящего времени присутствуют в косметических составах. Данные вещества являются предпочтительно производными дибензоилметана, в частности 1-(4'-трет-бутилфенил)-3-(4'-метоксифенил)пропан-1,3-дионом и 1-фенил-3-(4'-изопропилфенил)пропан-1,3-дионом. Данные сочетания и составы, которые включают в себя данные комбинации, также предлагаются изобретением. Могут быть применены количества, используемые для сочетания UVB.
Изобретение также относится к сочетаниям активных веществ в соответствии с изобретением с антиоксидантами, веществами аэробного энергетического метаболизма клетки и/или агентами, поглощающими УФ, с которыми, например, стабильность и действие состава могут быть улучшены. Представленные выше примеры активных веществ, которые можно сочетать с установленными группами активных веществ, служат для описания изобретения без намерения ограничить изобретение данными примерами.
Более того, возможно применение защитных форм составов; веществ в соответствии с изобретением, включенных (инкапсулированных), например, в липосомы, мицеллы, наносферы и т.п., например гидрогенизированные амфифилы, такие как, например, керамиды, жирные кислоты, сфингомиелин и фосфоглицериды или циклодекстраны. Может быть достигнута дополнительная защита с помощью применения защитного газа (например, N2, CO2) в процессе составления и применение упаковки со сжатым газом. Еще одними вспомогательными и дополнительными агентами могут быть связывающие воду вещества, загустители, наполнители, ароматизаторы, красители, эмульгаторы, активные вещества, такие как витамины, консерванты, вода и/или соли.
В процессе получения активных веществ или других веществ, потенциально чувствительных к кислороду, температура не должна обычно превышать приблизительно 40°С. Обычная практика получения композиций, которая известна специалистам в данной области техники, особо описывается.
Соединения согласно изобретению могут быть включены во все косметические основы. Может присутствовать масло или маслянистый материал совместно с эмульгатором для обеспечения либо эмульсии «вода в масле» (W/O), либо эмульсии «масло в воде» (O/W), в зависимости главным образом от среднего баланса гидрофильности-липофильности (HLB) применяемого эмульгатора. В частности, выгодными могут быть эмульсии W/O, O/W и W/O/W. Сочетания в соответствии с изобретением могут обычно выгодно применяться в продуктах по уходу за кожей, таких как, например, кремы O/W, кремы W/O, лосьоны O/W, лосьоны W/O и т.п.
Носители, отличные от воды или в дополнение к воде, которые могут быть применены, включают в себя смягчающие агенты, растворители, увлажнители, загустители и порошки. Особенно пригодным неводным носителем является полидиметилсилоксан и/или полидиметилфенилсилоксан. Применяемые кремнийорганические соединения могут обычно представлять собой такие, вязкость которых колеблется в диапазоне приблизительно от 10 до 10000000 мм2/сантистокс при 25°С. Также желательными могут быть смеси кремнийорганических соединений с низкой и высокой вязкостью. Данные кремнийорганические соединения имеются в продаже от General Electric Company под торговыми марками Vicasil, SE и SF и от Dow Corning Company под сериями 200 и 550. Количества кремнийорганических соединений, которые могут быть применены в композициях согласно изобретению, могут колебаться в диапазоне примерно от 5 до 95%, предпочтительно от 25 до 90%, от массы композиции.
Примерами углеводородов, которые могут служить в качестве смягчающих агентов, являются те, которые имеют углеводородные цепи примерно от 12 до 30 углеродных атомов. Конкретные примеры включают в себя минеральное масло, вазелин, сквален и изопарафины.
Другой категорией функциональных ингредиентов косметических композиций согласно изобретению являются загустители. Загуститель должен обычно присутствовать в количествах примерно от 0,1 до 20% от массы, предпочтительно приблизительно от 0,5% до 10% от массы композиции. Примерами загустителей являются поперечно-сшитые полиакрилатные материалы, имеющиеся в продаже под торговой маркой Carbopol от B. F. Goodrich Company. Могут быть применены камеди, такие как ксантан, каррагинан, желатин, карайа, пектин и камедь бобов робинии. В определенных условиях функция загущения может быть присуща материалу, служащему также в качестве кремнийорганического соединения или смягчителя. Например, силиконовые камеди с избытком в 10 сантистокс и сложные эфиры, такие как стеарат глицерина, имеют двойную функцию.
В косметическую композицию согласно изобретению могут быть включены порошки. Такие порошки включают в себя мел, тальк, каолин, крахмал, смектитовую глину, химически модифицированный силикат магния и алюминия, органически модифицированная монтмориллонитная глина, гидратированный силикат алюминия, парообразный кремнезем, алюминий-крахмал-октенилсукцинат и их смеси.
Другие дополнительные минорные компоненты могут быть также включены в косметические композиции. Данные ингредиенты могут включать в себя окрашивающие агенты, агенты, снижающие прозрачность, и ароматизаторы. Количества данных других дополнительных минорных компонентов может находиться в диапазоне примерно от 0,001 до 20% от массы композиции.
Косметически приемлемый носитель должен обычно составлять от 5 до 99,9%, предпочтительно от 25 до 80% от массы композиции, и может в отсутствие других косметических дополнительных средств образовывать сбалансированную композицию. Предпочтительно носитель состоит по меньшей мере из 80 мас.% воды от массы носителя. Предпочтительно вода составляет по меньшей мере 50 мас.% композиции согласно изобретению, более предпочтительно от 60 до 80 мас.% от массы композиции.
(iii) Применение местных и косметических композиций
Местные и косметические композиции согласно изобретению должны быть предназначены в первую очередь в качестве продукта для местного нанесения на кожу человека, особенно в качестве агента для улучшения, увлажнения и смягчения кожи и предотвращения или снижения появления линий, морщин или стареющей кожи. При применении маленькое количество композиции, например от 1 до 100 мл, наносится на подвергаемые воздействию области кожи из подходящего контейнера или аппликатора и, если это необходимо, затем распределяется по коже и/или втирается в нее с применением руки или пальцев, или подходящего устройства. Альтернативно, композиции могут быть доставлены любым другим местным способом, включая введение через слизистые, такое как пероральное, ректальное и интраназальное введение.
(iv) Форма продукта и упаковка
Местные кожные и косметические композиции согласно изобретению могут быть составлены, например, в виде лосьона, крема или желе. Композиция может быть упакована в подходящий контейнер для соответствия ее вязкости и предназначенного применения потребителем. Например, лосьон или крем могут быть упакованы в сосуд или аппликатор с роликовым шариком или в устройство с аэрозолем и движущим газом, или в контейнер, снабженный насосом, подходящим для использования с помощью пальца. Когда композиция представляет собой крем, она может просто храниться в недеформируемом сосуде или контейнере для выдавливания, таком как тюбик или закрытая банка. Композиция может быть также включена в капсулы, например, такие, которые описаны в патента США No. 5063507, включенном здесь в качестве ссылки. В изобретении, соответственно, также предлагается закрытый контейнер, содержащий косметически приемлемую композицию, как здесь представлено.
VIII. Применение монотерпеновых/тритерпеновых соединений
Описанные здесь монтерпеновые/тритерпеновые соединения рассматриваются как полезные для широкого спектра применения в дополнение к противовоспалительному и противоопухолевому действию, например, в качестве растворителей, противогрибковых и противовирусных агентов, в качестве пестицидов или моллюскицидов, в качестве контрацептивов, антигельминтных средств, в качестве агентов, защищающих от УФ, отхаркивающих агентов, диуретиков, в качестве регуляторов метаболизма холестерина, средств, действующих на сердечно-сосудистую систему, противоязвенных агентов, анальгетиков, седативных средств, иммуномодуляторов, жаропонижающих агентов, регуляторов ангиогенеза, агентов, снижающих ломкость капилляров, агентов, борющихся с признаками старения, и агентов для улучшения когнитивных функций и памяти.
В изобретении предлагаются сильные противовоспалительные агенты, примерами которых являются описанные здесь монотерпеновые/тритерпеновые соединения. Авторы показали, что монотерпеновые/тритерпеновые соединения согласно изобретению являются сильными ингибиторами транскрипционного фактора NF-κB, который играет важную роль в воспалительном ответе. Эти данные особенно важны в связи с наличием увеличивающегося количества доказательств, предполагающих, что воспалительные процессы играют центральную роль в канцерогенезе. Лечение больных предлагаемыми здесь монотерпеновыми/тритерпеновыми соединениями может, следовательно, существенно облегчить широкий диапазон нарушений, связанных с воспалением, включая развитие опухолей и повреждение тканей.
Первоначальные стадии воспалительного ответа характеризуются увеличенной проницаемостью кровеносных сосудов и высвобождением (эксудацией) гистамина, серотонина и основных полипептидов и белков. Это сопровождается гиперемией и образованием отека. В последующем наблюдается клеточная инфильтрация и образование новой соединительной ткани. Авторы считают, что лечение монотерпеновыми/тритерпеновыми соединениями должно ограничить данные ранние стадии воспаления и в результате этого снизить негативные эффекты, связанные с воспалительным состоянием.
Соединения играют роль в регуляции ангиогенеза. Ангиогенез или неоваскуляризация определяется как рост новых кровеносных сосудов. Опухоли и злокачественные новообразования индуцируют ангиогенез для обеспечения поступления кислорода и питательных веществ в опухоль, что жизненно важно для ее разрастания. Развитие новых кровеносных сосудов также обеспечивает выход малигнизированных раковых клеток для распространения в другие части тела. Следовательно, торможение ангиогенеза благоприятно для раковых больных. С другой стороны, ангиогенез требуется в определенный момент, такой как заживление ран. Данные раны могут быть внешними ранами или ранами внутренних органов, которые возникают в результате несчастных случаев, ожогов, повреждений и хирургических операций. Таким образом, агенты, которые стимулируют ангиогенез, имеют большие возможности в отношении использования при заживлении ран.
Рассматривается также применение данных соединений для модуляции метаболизма холестерина. В частности, соединения и пищевые добавки рассматриваются как применяемые в снижении уровня холестерина сыворотки у больных людей. Следовательно, при лечении больных тритерпеновыми соединениями согласно изобретению, либо пероральном, либо внутривенном, считается, что заболеваемость, связанная с высоким холестерином и относящаяся к болезням сердечно-сосудистой системы, может быть снижена.
В отношении сердечно-сосудистых нарушений считается, что описанные здесь соединения могут быть использованы для лечения аритмии и дополнительно могут быть использованы в качестве сосудистых релаксантов, что приводит в результате к их антигипертензивному действию.
Вид растений, в котором идентифицированы соединения согласно изобретению, Acacia victoriae, был выбран частично потому, что он является природным для засушливых областей. Важной функцией метаболизма растений из данных областей является продукция соединений, которые защищают клетки от ультрафиолетового излучения. Авторы изобретения конкретно считают, что монотерпеновые/тритерпеновые соединения способны служить в качестве агентов, защищающих от УФ. Следовательно, предполагается, что соединения согласно изобретению должны найти широкое применение в способах применения, в которых желательна защита против ультрафиолетового облучения. Например, подходящее применение включает в себя использование монотерпеновых/тритерпеновых соединений в качестве ингредиентов в солнцезащитных или других сходных лосьонах для нанесения на кожу человека.
Существенное преимущество такой композиции доказывается хемопротекторными эффектами, продемонстрированными для представленных здесь соединений. Лосьоны и солнцезащитные средства, содержащие монотерпеновые/тритерпеновые соединения, должны, следовательно, быть особенно пригодны для тех, кто предрасположен к различным формам рака кожи. Примеры такого рода включают в себя светлокожих и особенно тех, кто имеет генетическую предрасположенность к раку кожи. Такая предрасположенность включает в себя наследуемые мутации онкогенов или мутации в клеточных механизмах, которые опосредуют репарацию ДНК после индуцированного УФ повреждения. Особенно существенными являются мутации в генах, контролирующих механизмы генетической репарации, например удаление индуцированных УФ димеров тимин-тимин. Сходно монотерпеновые/тритерпеновые соединения могут быть добавлены к любой другой композиции, для которой желательна увеличенная защита от УФ, и данные соединения применимы к любому животному объекту или объекту, не являющемуся животным, для которого ищется защита от УФ.
Другое возможное применение монотерпеновых/тритерпеновых соединений включает в себя защиту от повреждений центральной нервной системы при действии в случае потери памяти или для улучшения когнитивной функции, применение в качестве антиоксиданта (мониторинг уровней антиоксидантных молекул в крови) или для увеличения оксида азота (NO) для лечения гипертензии или атеросклероза и для лечения или предотвращения катаракты. Кроме того, авторы конкретно рассматривают местное применение монотерпеновых/тритерпеновых соединений согласно изобретению для усиления функции полового члена. Изобретение также включает в себя местное введение соединений для увеличения коллагена кожи, в результате чего возникают эффекты борьбы со старением кожи.
IX. Ингибирование воспаления
NF-κB, повсеместный транскрипционный фактор, эволюционно консервативный от мух до млекопитающих, представляет собой один из центральных регуляторов ответов организма на различные стрессогенные сигналы. NF-κB регулирует транскрипцию ряда генов, вовлеченных в иммунные и воспалительные пути. Например, гены различных провоспалительных цитокинов, молекулы адгезии и факторы апоптоза регулируются NF-κB и, следовательно, дисрегуляция NF-κB вносит вклад в разнообразные патологические состояния, такие как септический шок, острое воспаление, репликация вирусов и некоторые виды рака.
NF-κB в своей наиболее преобладающей и активной форме включает в себя димерные комплексы двух белков p50/RelA (также называемых p50/p65). В клетках, которые не являются активированными, эти факторы удерживаются в цитоплазме в комплексе с другими ингибиторными белками, такими как IκB, которые маскируют сигнал ядерной транслокации белкового комплекса. Когда клетка получает внеклеточный сигнал, такой как сигнал, опосредованный воспалительным цитокином, митогеном, микробным продуктом или сигналом окислительного стресса, ингибиторный белок, IκB, подвергается фосфорилированию по специфическим сериновым остаткам. Это служит сигналом для присоединения убиквитина и деградации ингибиторных белков по протеосомному пути. При деградации IκB домен ядерной локализации NF-κB экспонируется, что позволяет теперь свободному от ингибитора комплексу NF-κB транслоцироваться в ядро, связываться с ДНК и активировать транскрипцию специфических генов. Так как NF-κB является важным для контроля/модуляции транскрипции генов в сигнальных путях воспаления, канцерогенеза и ряда иммунологических нарушений, авторы считают, что негативные модуляторы NF-κB должны служить в качестве терапевтического средства при лечении воспалительных нарушений. Таким образом, например, введение монотерпеновой композиции, которая ингибирует NF-κB в соответствии со способами согласно изобретению, ингибирует активацию воспалительных ответов в клетке и, следовательно, предотвращает/ингибирует/снижает воспаление. Во многих случаях продолжительное или хроническое воспаление часто ведет к другим состояниям. В одном примере воспалительное заболевание кишечника (IBD) (описанное ниже более подробно), которое начинается как воспаление толстой кишки, может вести в некоторых случаях к раку ободочной кишки. Таким образом, авторы рассматривают применение монотерпеновых/тритерпеновых композиций, описанных в настоящем изобретении, для исцеления и лечения воспалительных нарушений, предраковых состояний и т.п. с помощью применения указанных здесь способов.
X. Апоптоз
Кругооборот гибели клеток млекопитающих характеризуется двумя основными путями апоптоза. Один представляет собой путь, опосредованный рецепторами, включающий в себя Fas и другие члены семейства рецепторов фактора некроза опухолей (TNF), которые активируют каспазу-8, в то время как другой вовлекает цитохром с, Apaf-1 и каспазу-9. Недавно было показано, что ряд событий в митохондриях существеннен для программируемой клеточной гибели. Одна из ранних решающих стадий в процессе апоптоза заключается в высвобождении цитохрома с из внешней мембраны митохондрий в цитозоль. В цитозоле цитохром с запускает активацию каспаз, которые представляют собой цистеиновые протеазы с аспартатной специфичностью. Описано большое число субстратов для каспаз, включая полимеразу поли(АДФ-рибозы) (PARP), 116 кДа фермент репарации ДНК, который расщепляется в ходе апоптоза. Авторы также описывают здесь эффекты монотерпеновых/тритерпеновых композиций согласно изобретению на митохондриальные события апоптоза.
XI. Предраковые воспалительные заболевания
Некоторые примеры из воспалительных состояний, которые можно лечить монотерпеновыми/тритерпеновыми композициями, описываются ниже.
Эзофагит Барретта. Больные с гастроэзофагитным рефлюксом склонны к эзофагиту, изъязвлению, образованию сужения и колончатой метаплазии нормальной сквамозной выстилки. Эзофагит Барретта наступает приблизительно у 10% больных с гастроэзофагитным рефлюксом и связан с наличием сужения, глубоких язв и с развитием аденокарциномы. Нарушения в виде гастроэзофагитного рефлюкса диагностируются более чем у 500000 человек в США каждый год, и только приблизительно у 35000 проводится антирефлюксная коррекция.
Лучевой кератоз. Предраковый рост кожи обычно вызван инсоляцией. Лучевой кератоз наиболее обычно возникает у белокожих людей, особенно у пожилых и у молодых индивидуумов со светлым цветом лица. Рост возникает в областях кожи, подвергаемых инсоляции. Рост начинается в виде плоских чешуйчатых областей, которые позже развиваются в твердую бородавкообразную поверхность. Он классифицируется как предраковый рост. Приблизительно 20% лучевых кератозов развивается в сквамозно-клеточную карциному. С этим состоянием ассоциируются старческая кожа, инсоляционный эластоз и другие изменения кожи, индуцированные солнцем.
Воспалительное заболевание кишечника (IBD) относится к хронических заболеваниям, которые вызывают воспаление толстой кишки, такое как язвенный колит (UC) и болезнь Крона (CD). UC вызывает воспаление и изъязвление внутренней выстилки ободочной и прямой кишки. Внутренняя выстилка называется мукозным слоем. Болезнь Крона (CD) вызывает воспаление, которое распространяется в более глубокие слои стенки кишечника. UC является относительно обычным в западном мире и по меньшей мере 250000 человек только в США страдают данным заболеванием. Оно наступает чаще всего у людей в возрасте от 15 до 30 лет, хотя у детей и пожилых людей иногда развивается болезнь. Болезнь Крона (CD) представляет собой воспалительный процесс, который может поражать любую часть пищеварительного тракта, но чаще всего наблюдается (приблизительно в половине всех случаев) в последней части тонкого кишечника, иначе называемой концом подвздошной кишки и слепой кишкой. В других случаях может поражаться одно или более из: только ободочной кишки, только тонкого кишечника (двенадцатиперстная кишка, тощая кишка и/или подвздошная кишка), ануса, желудка или пищевода. Многие из данных состояний ведут к раку ободочной кишки.
XII. Активация элементов антиоксидантного ответа
Большинство химических канцерогенов должно подвергнуться метаболической активации перед инициацией канцерогенеза. Активация наступает в результате образования электрофильных реагентов (Ramos-Gomez et al., 2001). Две главные линии защиты от канцерогенеза в клетках млекопитающих представляют собой (1) ферменты фазы 2, способные детоксицировать электрофильные соединения и служащие таким образом в качестве антиоксидантов и (2) глутатион. Уровни ферментов фазы 2 и глутатиона способны индуцироваться под действием различных природных и синтетических агентов. Авторы специально рассматривают вопрос о том, что соединения согласно изобретению могут быть использованы для такой индукции. Это значит, что считается, что хемопротекторная эффективность соединений согласно изобретению может быть обусловлена индукцией ферментов фазы 2, которые нейтрализуют реактивные электрофилы и действуют как непрямые антиоксиданты, и/или потенциально активируют ферменты фазы 1.
Путем введения соединения согласно изобретению можно модулировать уровни ферментов фазы 2 и будут реализоваться физиологические эффекты модуляции, такие как увеличенная или сниженная детоксикация и устойчивость к химическому возмущению. Элементы антиоксидантного ответа вовлечены в экспрессию ферментов фазы 2, опосредующих защиту против разнообразных канцерогенов и других токсинов. Семейство индуцибельных белков фазы 2 в пределах данного класса является разнообразным, причем различные ферменты играют различную роль в защите клеток. В дополнение к индукции "классических" ферментов фазы 2, метаболизирующих лекарства, таких как глутатион-S-трансферазы (GST) и УДФ-глюкуронозилтрансферазы (Benson et al., 1978; Cha et al., 1979), которые конъюгируют ксенобиотики с эндогенными жидкостями, группа индуцируемых белков также включает в себя НАД(Ф)Н:хинон-редуктазу (NQO1) (Benson et al, 1980); эпоксидгидролазу (Benson et al., 1979); гемоксигеназу 1 (Prestera et al., 1995; Primiano et al., 1996); ферритин (Primiano et al., 1996), γ-глутамилцистеинсинтетазу (Mulcahy et al., 1997; Moinova et al., 1998, и Otieno et al., 2000); афлатоксинальдегидредуктазу (Ellis et al., 1996; Kelly et al., 2000); каталазу и супероксиддисмутазу (Otieno et al., 2000); дигидродиолдегидрогеназу (Ciaccio et al., 1994); лейкотриен B4 дегидрогеназу (Primiano et al., 1999); семейство альдокеторедуктаз, включающее в себя альдозоредуктазу и альдегидредуктазу, и откачивающие насосы для глутатион-S-конъюгатов (см. для обзора Hayes et al., 1999). Модуляция одного, всех или некоторых сочетаний данных или других ферментов, опосредующих элементы антиоксидантного ответа, может быть достигнута путем введения раскрытых здесь соединений, и такие применения конкретно составляют часть согласно изобретению.
В дополнение к раку другие многочисленные заболевания связаны с измененной функцией одного или двух ферментов фазы 2. Соответственно, лечение таких заболеваний с применением раскрытых здесь соединений составляет часть изобретения. Например, недостаточность детоксикации токсических альдегидов под действием таких ферментов, как альдокеторедуктазы, связана с заболеваниями, включающими гигантоклеточный височный артериит и болезнь Паркинсона. Гемоксигеназа, как известно, является важной для защиты нейронов и других тканей, включая клетки почек и легких, от окислительного стресса. Далее, функция откачивающих насосов для глутатион-S-конъюгатов, как известно, вовлечена в множественную устойчивость к лекарствам (MDR) раковых клеток, к таким как химиотерапевтические агенты, и в устойчивость к антибиотикам у бактерий. Лечение соединением согласно изобретению может таким образом найти применение в модуляции функции откачивающих насосов для глутатион-S-конъюгатов для того, чтобы уничтожить или минимизировать MDR или устойчивость к антибиотикам. Соединения согласно изобретению могут быть также использованы для модуляции сигнальных путей, опосредуемых откачивающими насосами для глутатион-S-конъюгатов, у бактерий.
Как указывалось, гены, кодирующие многие ферменты фазы 2, содержат в 5'-вышележащей области элементы, чувствительные к антиоксидантам (ARE/EpRE; консенсусная последовательность TGACNNNGC), которые регулируют как базальную, так и индуцибельную экспрессию. Исследовали идентичность транскрипционных факторов, которые взаимодействуют с ARE элементами (см. Hayes et al., 1999 для обзора), но считается, что член семейства транскрипционных факторов с основной лейциновой застежкой играет центральную роль в экспрессии ферментов фазы 2 (Ramos-Gomez et al., 2001). Экспозиция клеток с индукторами разрушает комплекс Keap1-Nrf2 и Nrf2 мигрирует в ядро, где он связывается, в гетеродимерных формах с другими транскрипционными факторами, с ARE энхансерных областей генов фазы 2 и стимулирует их транскрипцию. Недавно было показано, что акцепторы реакции Михаеля индуцируют ферменты фазы 2 и их хемопротекторные эффекты зависят от их взаимодействия с сульфгидрильными группами (Dinkova-Kostova et al., 2001). Таким образом, структура соединений согласно изобретению может отвечать за ингибирование связывания ядерного фактора-κB-p65 с ДНК, а также за индукцию хемопротекторных ферментов. Как таковые, как указано выше, соединения согласно изобретению могут найти применение в качестве важных профилактических агентов во многих клинических ситуациях, характеризуемых хроническим воспалением, окислительным стрессом и высоким риском развития неоплазии.
Одним конкретным применением соединений согласно изобретению является предотвращение клеточного повреждения, вызываемого активными формами кислорода (АФК) и активными формами азота (АФА). Например, при введении соединений согласно изобретению в клетки или в ткани, включающие в себя клетки, для активации элементов, чувствительных к антиоксидантам (ARE), повреждение от АФК и АФА могут быть предотвращены или уменьшены. Сходно такие способы могут включать в себя одновременное снижение воспаления. Способность лечить или сводить к минимуму повреждение, обусловленное АФК и АФА, является существенной, так как многие патологические состояния связаны с активностью АФК/АФА. Некоторые конкретные примеры данных состояний, каждое из которых может лечиться в соответствии с изобретением путем введения описанных здесь соединений нуждающемуся в этом больному, представлены в таблице 6.
Некоторые из клинических состояний, связанных с АФК/АФА
Сокращения: ARDS, респираторный синдром взрослых; COPD, хроническое обструктивное заболевание легких; NSAID, нестероидное противовоспалительное лекарство.
Состояния, связанные с окислительным стрессом, могут быть вызваны в общем одним из следующих факторов (или обоими): (1) снижением уровня антиоксидантов, например, мутациями, влияющими на ферменты антиоксидантной защиты (таким как CuZnSOD, MnSOD и глутатионпероксидаза), либо заболеваниями, которые снижают такую защиту. Многие ксенобиотики метаболизируются путем конъюгации с GSH; высокие дозы могут снижать GSH и вызывать окислительный стресс, даже если ксенобиотик сам не является генератором АФК или АФА. Снижение поступающих с пищей антиоксидантов и других существенных составляющих диеты может также вести к окислительному стрессу; (2) увеличенной продукцией АФК/АФА, например, в результате экспозиции с увеличенным уровнем O2, присутствием токсинов, которые сами являются активными формами (например, NO2) или метаболизируются, генерируя АФК/АФА, или избыточной активацией "природных" систем, продуцирующих АФК/АФА (например, непропорциональной активацией фагоцитарных клеток при хронических воспалительных заболеваниях).
Тканевое повреждение, в частности, является одним из источников окислительного стресса. Примеры повреждений, которые могут вести к окислительному стрессу и которые, следовательно, могут лечиться соединениями согласно изобретению, включают в себя ишемию-реперфузию, жар, травму, переохлаждение, избыточные упражнения, токсины, облучение и инфекцию. Многочисленные модальности указывались для создания окислительного стресса, включая вовлечение фагоцитов и активацию (производство O2, H2O2, NO-, HOCl); высвобождение арахидоновой кислоты, ферментативное образование пероксида (путем активации ферментов - липоксигеназы и циклооксигеназы); разрушение образованных с помощью ферментов и образованных без помощи ферментов пероксидов до пероксильных/алкоксильных радикалов, распространяющих повреждение до других липидов/белков; высвобождение ионов металла из мест хранения (Fe2+, Cu2+), стимулирующих превращение H2O2 в OH-, расщепление липидного пероксида до RO2/RO- и "аутоокислительные" реакции; высвобождение содержащего гем белка (миоглобина, гемоглобина, цитохромов); взаимодействие содержащих гем белков с пероксидами для стимуляции повреждения свободными радикалами и (если пероксид находится в избытке) для высвобождения Fe2+ и гема, оба из которых могут разрушать пероксиды до RO2 и RO; вмешательство в антиоксидантные системы защиты (например, потеря GSH и аскорбата клетками); потерю аскорбата во внеклеточных жидкостях; превращение ксантиндегидрогеназы в оксидазу в определенных тканях, возможное высвобождение ксантиноксидазы из поврежденных клеток, вызывающее системное повреждение, увеличенные уровни гипоксантина, обусловленные нарушенным энергетическим метаболизмом; повреждение митохондрий, увеличенную утечку электронов с образованием O2 -; и повышенный внутриклеточный Ca2+, стимулирующий кальпаины, Ca2+-зависимые нуклеазы и Ca2+/кальмодулин-зависимую синтазу оксида азота, производящую больше NO и создающую увеличенный риск образования ONOO-.
XIII. Тесты и способы скрининга активных соединений
Ряд тестов известен специалистам в данной области техники, и они могут быть использованы для дальнейшей характеристики описанных здесь монотерпеновых/тритерпеновых соединений. Они включают в себя тесты биологической активности, а также тесты химических свойств. Результаты данных тестов позволяют сделать важные выводы как о свойствах соединений, так и о их потенциальном применении при лечении человека или других больных млекопитающих. Считается, что тесты, особенно применимые в данном отношении, включают в себя скрининг биологической активности in vivo и in vitro и иммунологические тесты.
(i) Тесты in vivo
Настоящее изобретение охватывает применение различных животных моделей. В данном случае идентичность, наблюдаемая между человеком и мышью, дает прекрасную возможность исследования функции потенциального терапевтического агента, например монотерпенового/тритерпенового соединения, описанного в настоящем изобретении. Можно применять модели воспаления у мышей, которые будут с высокой точностью предсказывать воспалительные заболевания у человека и других животных. Могут быть также применены модели рака, и данные модели могут применять ортотопическое или системное введение опухолевых клеток для имитации первичного рака и/или метастазов рака. Альтернативно, можно индуцировать воспалительное заболевание или рак у животных путем обработки агентами, известными как ответственные за определенные события, связанные с конкретным типом заболевания.
Обработка животных тестируемыми соединениями должна включать в себя введение животному соединения в подходящей форме. Введение должно осуществляться любым путем, который может использоваться для клинических или неклинических целей, включая, но не ограничиваясь этим, пероральное, интраназальное, защечное, ректальное, вагинальное или местное. Альтернативно, введение может осуществляться с помощью внутритрахеального нанесения, бронхиального нанесения, внутрикожного, подкожного, внутримышечного, внутрибрюшинного или внутривенного введения. Специально рассматриваемым является системное внутривенное введение, региональное введение путем поступления через кровь или лимфу и внутриопухолевое введение.
Определение эффективности соединения in vivo может включать в себя множество различных критериев. Такие критерии включают в себя, но не ограничиваются этим, выживаемость, снижение воспаления, элиминацию воспалительного ответа, ингибирование или предотвращение развития рака, обусловленное предраковым воспалительным состоянием, увеличенный уровень активности, улучшение иммунной эффекторной функции и улучшение потребления пищи.
Один особенно пригодный тип тестирования противоопухолевой активности in vivo включает в себя применение модели кожи мыши. Модель кожи мыши, которая представляет собой одну из наиболее хорошо разработанных экспериментальных моделей многостадийного канцерогенеза, позволяет разделить три различных стадии развития рака: инициацию, промоцию и прогрессию. В настоящее время очевидно, что переход клетки в малигнизированное состояние включает в себя последовательное изменение протоонкогенов и/или генов-супрессоров опухолей, генные продукты которых участвуют в решающих путях передачи сигналов и/или регуляции экспрессии генов. Стадии промоции и прогрессии опухоли кожи характеризуются селективной и поддерживаемой гиперплазией, изменениями дифференцировки и генетической нестабильностью, ведущей к специфической экспансии инициированных клеток и превращению в папилломы и карциномы. Было показано, что индукция поддерживаемой гиперплазии коррелирует с активностью в отношении промоции опухоли кожи под действием различных агентов, таких как форболовые эфиры, некоторые перекиси и крисаробин. На модели кожи мыши показано, что все известные канцерогены и факторы промоции опухолей вызывают поддерживаемую гиперплазию эпидермиса. В целом этому предшествует воспалительный ответ.
Интенсивные исследования выявили хорошую корреляцию между канцерогенностью и мутагенностью. Большинство агентов инициации опухоли либо генерируют, либо метаболически превращаются в электрофильные реагенты, которые ковалентно связываются с клеточной ДНК. Некоторые свободные радикалы и модифицированные основания ДНК, являющиеся свободными радикалами, вовлечены в канцерогенез на стадии инициации опухоли и/или промоции опухоли. Убедительное доказательство показало, что активация гена Ha-ras рано наступает в процессе канцерогенеза кожи мыши и, вероятно, эквивалентна моменту инициации. Например, показано, что присутствие активированного гена Ha-ras в папилломах и карциномах кожи мыши, индуцированных 7,12-диметилбенз[a]антраценом, связано с высокой частотой трансверсий A-T в кодоне 61. Последующие исследования показали, что данный тип мутации зависел от химического инициатора и не зависел от промотора, что предполагает наличие прямого эффекта инициатора на c-Ha-ras. Более того, инфицирование кожи мыши активированным вирусом геном Ha-ras (v-Ha-ras) может служить инициацией даже при двустадийном канцерогенезе. Необходимо подчеркнуть, что все химические канцерогены кожи и инициаторы опухоли кожи, как показано, вызывают мутацию онкогена Ha-ras. Однако промоторы опухоли кожи не вызывают мутации Ha-ras.
(ii) Подтверждение опытов in vivo и клинических исследований
Специалистам в данной области техники должно быть понятно, что терапевтические агенты, включая монотерпеновые/тритерпеновые соединения согласно изобретению или сочетания их с дополнительными агентами, должны обычно тестироваться в условиях in vivo перед применением у субъекта, являющегося человеком. Такое предклиническое тестирование на животных является обычным в данной области техники. Для проведения таких подтверждающих тестов все, что требуется, это приемлемые для данного уровня техники животные модели исследуемого заболевания, такие как модели животных, имеющих воспалительное заболевание или солидную опухоль. В данном контексте может быть использовано любое животное, такое как, например, мышь, крыса, морская свинка, хомячок, кролик, собака, шимпанзе или тому подобное. В плане лечения воспалительных заболеваний и рака широко используется применение небольших животных, таких как мыши, как предсказывающих клиническую эффективность у человека, и такие животные модели являются, следовательно, предпочтительными в контексте настоящего изобретения, так как они легко доступны и относительно недороги по меньшей мере по сравнению с другими экспериментальными животными.
Способ проведения экспериментального теста на животных является ясным для специалиста в данной области техники. Все, что требуется для проведения такого теста, это определение эквивалентных групп для обработки и введение тестируемых соединений одной группе, в то время как различные контрольные исследования проводятся параллельно на эквивалентных животных в оставшейся группе или группах. Ведется наблюдение за животными в течение курса исследования, и в конечном итоге животные забиваются для анализа эффектов лечения.
В плане лечения воспаления считается, что эффективные количества монотерпеновых/тритерпеновых соединений должны быть такими, чтобы обычно по меньшей мере приблизительно в 10% клеток в пределах области воспаления вызывать снижение воспаления. Предпочтительно, чтобы по меньшей мере в приблизительно 20%, приблизительно 30%, приблизительно 40% или приблизительно 50% клеток вызывалось снижение воспаления. Более предпочтительно, чтобы в 100% клеток в пределах данной области вызывалось снижение воспаления.
Предпочтительно применять дозы монотерпеновых/тритерпеновых соединений, способные индуцировать по меньшей мере приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95% и вплоть до 100% снижение воспаления, до тех пор пока применяемые дозы не приведут к значительным побочным эффектам или к другим неблагоприятным реакциям у животного. Все такие определения могут быть легко осуществлены и должным образом оценены специалистом в данной области техники. Например, обслуживающий персонал, ученые и врачи могут применять такие данные, полученные на экспериментальных животных, для оптимизации подходящих доз при лечении человека. У субъектов с далеко зашедшим заболеванием может допускаться определенная степень побочных эффектов. Однако больных на ранних стадиях заболевания можно лечить более умеренными дозами, с тем чтобы получить терапевтический эффект при отсутствии побочных эффектов. Эффекты, наблюдаемые при исследованиях на таких экспериментальных животных, должны предпочтительно быть статистически достоверными по сравнению с контрольными уровнями и должны воспроизводиться от опыта к опыту.
Специалисты в данной области техники должны также понимать, что сочетания и дозы соединений согласно изобретению, вызывающих снижение воспаления, сдвинутые в направлении нижнего предела эффективного диапазона доз, могут тем не менее быть все еще пригодными в связи с настоящим изобретением. Например, осуществления, в которых рассматривается продолжительное нанесение активных агентов, начальная доза, которая вызывает только приблизительно 10% снижение воспаления, будет тем не менее полезной. Еще дополнительно рассматривается, что доза монотерпеновых/тритерпеновых соединений, которая предотвращает или снижает вероятность либо образования рака как продвинутой стадии воспалительного состояния, либо метастазирования, либо канцерогенеза de novo, должна также быть терапевтически выгодной для больного, получающего лечение.
Как обсуждалось выше в связи с системой тестирования in vivo, естественно должно быть понятно, что сочетания агентов, предназначенных для совместного применения, должны быть протестированы и оптимизированы совместно. Соединения согласно изобретению могут быть просто проанализированы в сочетании с одним или более терапевтических лекарств, иммунотоксинов, коагулянтов или тому подобное. Анализ сочетанных эффектов таких агентов должен быть проведен, и результаты определены и оценены в соответствии с правилами, установленными выше.
(iii) Тесты in vitro
В одном осуществлении изобретения скрининг растительных экстрактов проводят in vitro для идентификации тех соединений, которые способны ингибировать воспаление и/или рост опухолевых клеток или вызывать их гибель.
Определение эффективности соединения в отношении снижения воспаления in vitro может быть достигнуто, например, с помощью тестирования экспрессии и индукции различных генов и/или белков, вовлеченных в воспаление, следовательно, в отношении снижения NF-κB, транскрипционного фактора, который вовлечен в контроль числа генов воспалительных путей, или снижения ферментов, таких как iNOS и COX-2. Оба данных фермента индуцируются в ответ на различные цитокины, такие как интерферон гамма, митогены, микробные продукты, такие как липополисахариды и т.п. Таким образом, они являются важными компонентами при воспалительных ответах, при репарации повреждений и при канцерогенезе.
В отношении уничтожения опухолевых клеток или цитотоксичности эти процессы обычно проявляются в виде некроза и апоптоза. Некроз является относительно общим путем, запускаемым внешними сигналами. В течение данного процесса теряется целостность клеточной мембраны и клеточных органелл. С другой стороны, апоптоз, или программируемая клеточная смерть, является высоко организованным процессом морфологических изменений, которые синхронизируются в результате активации и инактивации специфических генов (Thompson et al., 1992; Wyllie, 1985).
Эффективный способ определения цитотоксичности in vitro включает в себя систематическую экспозицию панели опухолевых клеток с различными растительными экстрактами. Такие тесты и линии опухолевых клеток, подходящие для проведения тестов, хорошо известны специалистам в данной области техники. Особенно выгодные линии опухолевых клеток человека для применения в тестах на противоопухолевую активность in vitro включают в себя линии клеток рака яичников человека SKOV-3, HEY, OCC1 и OVCAR-3; Т-лейкозные клетки Юрката; линия клеток рака молочной железы человека MDA-468; клетки рака простаты человека LNCaP, линии клеток меланомы человека A375-M и Hs294t; и клетки рака почки человека 769-P, 786-0, A498. Предпочтительный тип линии нормальных клеток для применения в качестве контроля представляет собой линии фибробластов крайней плоти человека FS или Hs27.
Определение эффективности соединения в плане уничтожения опухолевых клеток in vitro может быть также достигнуто с помощью тестирования экспрессии и индукции различных генов, вовлеченных в остановку клеточного цикла (p21, p27; ингибиторы циклин-зависимых киназ) и апоптоза (bcl-2, bcl-xL и bax). Для выполнения данного тестирования клетки обрабатывают тестируемым соединением, лизируют, белки выделяют и затем разгоняют на ДДС-Na ПААГ гель-электрофорезе и связанные с гелем белки переносят на нитроцеллюлозные мембраны. Мембраны первоначально обрабатывают первыми антителами (например, антителами к p21, p27, bax, bcl-2, bcl-xL и т.п.) и затем проявляют с помощью разведенных вторых антител, конъюгированных с пероксидазой хрена, и мембраны экспонируют с реагентом, определяющим ECL, с последующей визуализацией на ECL-фотографической пленке. Путем анализа относительной пропорции белков может быть сделано заключение о проценте клеток в данной фазе, например фазе G0/G1, фазе S или фазе G2/M.
Цитотоксичность соединения в отношении раковых клеток может быть эффективно обнаружена in vitro с применением MTT или окрашивания кристаллическим фиолетовым. В данном способе клетки рассевают на плашках, экспонируют с различными концентрациями образцов соединений, инкубируют и окрашивают либо MTT (3-(4,5-диметилэтиазол-2-ил)-2,5-дифенилтетразолийбромид; Sigma Chemical Co.) или кристаллическим фиолетовым. Плашки, обработанные MTT, обрабатывают лизисным буфером (20% додецилсульфат натрия в 50% ДМФ) и подвергают дополнительной инкубации перед считыванием ОП при 570 нм. Окрашенные кристаллическим фиолетовым плашки отмывают для удаления красителя буфером Соренсона (0,1 М цитрат натрия (рН 4,2), 50% об./об. этанол) и считывают при ОП 570-600 нм (Mujoo et al., 1996). Относительное поглощение обеспечивает определение результирующей цитотоксичности.
(iv) Тесты на ингибирование NF-κB
Определения in vitro эффективности соединения в отношении ингибирования NF-κB может быть достигнуто, например, с помощью анализа экспрессии и индукции различных генов и/или белков, которые индуцируются/модулируются/регулируются NF-κB, например, с помощью определения генной экспрессии или экспрессии белков для генов, вовлеченных в воспалительные пути. В одном конкретном примере с помощью определения снижения ферментов, таких как iNOS и COX-2, которые индуцируются в результате нижележащих ответов на активацию NF-κB в ответ на стимуляцию клетки цитокинами, такими как интерферон гамма, митогены, микробные продукты, такие как липополисахариды (LPS) и т.п. Данные определения могут включать в себя способы, известные в данной области техники, такие как иммуноблоттинг, тестирование экспрессии генов, определение ферментов и тому подобное.
Например, тесты на ингибирование NF-κB включают в себя анализ с помощью иммуноблоттинга, когда цитоплазматический и ядерный белковые экстракты клеток, обработанных ингибиторным соединением, применяют для изучения деградации ингибиторных белков NF-κB и ядерной транслокации субъединицы NF-κB, соответственно. Для такого теста цитоплазматический или ядерный белковые экстракты разделяют на геле и переносят с помощью электропереноса на подложку/мембрану. Мембраны могут быть протестированы подходящим образом антителом, например антителом кролика против NF-κB или антителом кролика против p65 (Santa Cruz, CA), затем антителом, конъюгированным с проявляющей частью, такой как пероксидаза хрена (HRPO), связывание флуоресцентных меток или радионуклиды. Полосы белков затем могут быть определены и идентифицированы с помощью подходящих средств, таких как хемилюминисценция, флуоресценция, радиоактивный способ и т.п.
В других осуществлениях тест может включать в себя трансфекцию и определение репортерного гена, такого как люцифераза или GFP. Таким образом, клетки могут быть трансфицированы репортерным конструктом, включающим NF-κB, с помощью электропорации или других способов. Клетки затем обрабатывают ингибиторным соединением (таким, как описанные здесь как моно/тритерпеновые композиции). NF-κB затем активируют, применяя стимуляторные агенты, такие как цитокины или микробный LPS, и измеряют активность репортерного гена.
Индукция и измерение ферментов, активированных NF-κB, таких как iNOS и COX-2, может быть осуществлено с помощью рассеивания на плашках клеток, обработанных ингибиторами (такими как F094 или монотерпеновые/тритерпеновые гликозиды), и активации NF-κB в клетках, как описано выше, например, путем экспозиции клеток с LPS/цитокинами. Клетки затем лизируют и белки экстрагируют, клеточный белок наносят на гель и осуществляют электроперенос на мембрану. Уровни iNOS и COX-2 анализировали с помощью иммуноблоттинга с применением подходящих антител, таких как кроличьи антитела против iNOS (Santa Cruz) и козлиные антитела против COX-2, в одном примере. В других осуществлениях активность ферментов может быть также измерена в виде метода определения ингибирования NF-κB.
(v) Иммунологические тесты
Иммунологические тесты могут найти применение в настоящем изобретении, например, при скрининге экстрактов из растительных видов, отличных от Acacia victoriae, на наличие описанных в изобретении монотерпеновых/тритерпеновых соединений. Иммунологические тесты, охватываемые настоящим изобретением, включают в себя, но не ограничиваются, описанными в патенте США No. 4367110 (тест с применением способа сэндвича с двумя моноклональными антителами) и патенте США No. 4452901 (иммуноблоттинг). Другие тесты включают в себя иммунопреципитацию меченых лигандов и иммуногистохимию, оба как in vitro, так и in vivo.
Иммунологические тесты в их наиболее простом и прямом смысле являются тестами связывания. Определенные предпочтительные иммунологические тесты представляют собой различные типы иммуноферментного анализа с применением иммуносорбента (ИФА) и радиоиммунного анализа (РИА), известные в данной области техники. Иммуногистохимическое определение с применением тканевых срезов также особенно пригодно.
В одном примере ИФА антитела против монотерпеновых/тритерпеновых соединений иммобилизуют на выбранной поверхности, имеющей сродство к белку, такой как лунка в полистироловом планшете для микротитрования. Затем тестируемую композицию, проверяемую на содержание монотерпеновых/тритерпеновых соединений, такую как растительный экстракт из растения, относящегося к Acacia victoriae, добавляют в лунки. После связывания и промывания для удаления неспецифически связывающих иммунных комплексов может быть определен связанный антиген. Определение обычно достигается с помощью добавления другого антитела, специфичного в отношении желаемого антигена, которое соединено с определяемой меткой. Данный тип ИФА является простым "ИФА сэндвичного типа". Определение также может быть достигнуто путем добавления второго антитела, специфичного в отношении желаемого антигена, с последующим добавлением третьего антитела, которое обладает связывающим сродством ко второму антителу, причем третье антитело связано с определяемой меткой.
Варианты способов ИФА известны специалистам в данной области техники. В одном таком варианте образцы, проверяемые на содержание желаемого антигена, иммобилизуются на поверхности лунок и затем вводятся в контакт с полученными антителами. После связывания и подходящей промывки определяются связанные иммунные комплексы. Когда первоначальные специфические для антигена антитела присоединяются к определяемой метке, иммунные комплексы могут быть определены прямо. Опять иммунные комплексы могут быть определены с применением второго антитела, которое обладает связывающим сродством к первому антиген-специфическому антителу, причем второе антитело связано с определяемой меткой.
Возможны также конкурентные ИФА, в которых тестируемые образцы конкурируют за связывание с известными количествами меченых антигенов или антител. Количество взаимодействующих видов в неизвестном образце определяется путем смешивания образца с известными видами метки перед или в течение инкубации с покрытыми лунками. Присутствие взаимодействующих видов в образце действует как снижение количества меченых видов, доступных для связывания с лункой и, таким образом, снижает конечный сигнал.
Независимо от применяемого типа ИФА имеют определенные общие черты, такие как покрытие, инкубация или связывание, промывание для удаления неспецифически связанных видов и определение связанных иммунных комплексов. Это описывается ниже.
Антиген или антитела также могут быть привязаны к твердой подложке, такой как в форме планшета, шариков, погруженных стержней, мембраны или матрикса колонки, и образец, подвергаемый анализу, наносят на иммобилизованный антиген или антитело. При покрытии планшета антигеном или антителом обычно лунки планшета инкубируют с раствором антигена или антитела либо в течение ночи, либо в течение конкретного периода. Лунки планшета затем отмывают для удаления не полностью адсорбированного материала. Любые оставшиеся доступные поверхности лунок затем "покрываются" неспецифическим белком, который по антигенам нейтрален в отношении тестируемых антисывороток. Такие белки включают в себя бычий сывороточный альбумин (БСА), казеин, и растворы порошкового молока. Покрытие позволяет блокировать места неспецифической адсорбции на иммобилизующей поверхности и в результате этого снижает фон, обусловленный неспецифическим связыванием антисывороток на поверхности.
В ИФА, видимо, более обычным является использование способов определения со вторым или третьим слоем, чем прямое определение. Таким образом, после связывания антигена или антитела с лункой, покрытия нереакционноспособным материалом для снижения фона и промывания для удаления несвязанного материала, поверхность с иммобилизованным материалом вводят в контакт с клиническим или биологическим образцом, предназначенным для тестирования в условиях, эффективных для индукции образования иммунного комплекса (антиген/антитело). Определение иммунного комплекса требует затем второго меченого лиганда или антитела, или второго связывающего лиганда или антитела в сочетании с третьим меченым антителом или третьим связывающим лигандом.
"В условиях, эффективных для индукции образования иммунного комплекса (антиген/антитело)" означает, что условия предпочтительно включают в себя разведение антигенов и антител растворами, такими как БСА, бычий гамма-глобулин (BGG) и забуференный фосфатом физиологический раствор (ЗФР)/твин. Данные добавляемые агенты также имеют тенденцию способствовать снижению неспецифического фона.
Подходящие условия также означают, что инкубацию проводят при температуре и в течение периода времени, достаточного для достижения эффективного связывания. Стадии инкубации обычно проходят приблизительно от 1 до 2 - 4 часов при температуре предпочтительно порядка от 25 до 27°С или могут проходить в течение ночи приблизительно при 4°С или около того.
В ИФА после всех стадий инкубации поверхность контакта промывают так, чтобы удалить несвязанный материал. Промывание часто включает в себя промывание раствором ЗФР/твин или боратным буфером. После образования специфических иммунных комплексов между тестируемым образцом и первоначально связанным материалом и последующего промывания может быть определено наличие даже минимальных количеств иммунных комплексов.
Для обеспечения средств определения второе или третье антитело должно иметь связанную с ним метку для возможности определения. Предпочтительно, чтобы это был фермент, который должен вызывать развитие окраски при инкубации с подходящим хромогенным субстратом. Таким образом, например, должно быть желательным введение в контакт и инкубация первого или второго иммунного комплекса с антителом, конъюгированным с уреазой, глюкозоксидазой, щелочной фосфатазой или гидрогенпероксидазой, в течение периода времени и в условиях, которые благоприятствуют развитию образования дополнительных иммунных комплексов, например при инкубации в течение 2 часов при комнатной температуре в растворе, содержащем ЗФР, таком как ЗФР-твин.
После инкубации с меченым антителом и последующего промывания для удаления несвязанного материала количественно определяется количество метки, например, с помощью инкубации с хромогенным субстратом, таким как мочевина и бромкрезол фиолетовый или 2,2'-азиноди(3-этилбензтиазолин-6-сульфокислота [ABTS] и H2O2 в случае использования пероксидазы в качестве ферментной метки. Затем достигается количественная оценка с помощью измерения степени развития окраски, например, с применением спектрофотометра для видимых спектров. Альтернативно, метка может быть хемилюминесцентной. Применение таких меток описано в патентах США No. 5310687, 5238808 и 5221605.
Способы анализа in vitro и in situ хорошо известны и включают в себя оценку связывания специфичных в отношении антигена антител с тканями, клетками или клеточными экстрактами. Они являются традиционными способами, имеющимися в распоряжении специалистов в данной области техники. Например, антитела против антигенов опухолевых клеток могут быть применены в связи с свежезамороженными и фиксированными в формалине, заключенными в парафин блоками тканей, полученными для исследования с помощью иммуногистохимии (IHC). Каждый блок ткани может состоять из 50 мг оставшейся "размельченной" опухоли. Способ получения блоков ткани из данных конкретных образцов был успешно применен в предшествующих IHC-исследованиях различных прогностических факторов, например, при раке молочной железы, и хорошо известен специалистам в данной области техники.
Вкратце, замороженные срезы могут быть получены путем регидратации 50 нг замороженной размельченной опухоли при комнатной температуре в ЗФР в маленьких пластиковых капсулах; осаждения частиц центрифугированием; их ресуспендирования в вязкой среде для заключения (OCT); переворачивания капсулы и повторного осаждения центрифугированием; моментального замораживания в изопентане при -70°С; разрезания пластиковой капсулы и удаления замороженного цилиндра ткани; закрепления тканевого цилиндра на зажимном патроне криостатного микротома и нарезки 25-50 серийных срезов, содержащих в среднем приблизительно 500 совершенно интактных опухолевых клеток.
Непрерывный ряд срезов может быть получен сходным способом, включая регидратацию 50 мг образца в пластиковой пробирке для микроцентрифуги; осаждение; ресуспендирование в 10% формалине в течение 4-часовой фиксации; промывку/осаждение; ресуспендирование в теплом 2,5% агаре; осаждение; охлаждение в ледяной воде для затвердевания агара; удаление тканевого/агарового блока из пробирки; инфильтрацию и заключение блока в парафин; нарезку до 50 серийных непрерывных срезов.
В свете настоящего раскрытия можно было бы применять скрининговые тесты для идентификации соединений, обладающих по существу такими же химическими свойствами и биологической активностью, что и описанные здесь. В частности, настоящее раскрытие могло бы обеспечить применение тестов для биологически активных монотерпеновых/тритерпеновых гликозидов из растений, которые являются близко родственными Acacia victoriae, например, членов рода Acacia. Данные тесты могут быть применены во множестве различных вариантов и могут зависеть от типа "активности", в отношении которой производится скрининг. Предпочтительные тесты включают в себя такие, которые предназначены для скрининга противовоспалительной активности и противоопухолевой активности, такие как описанные здесь из экстрактов Acacia victoriae. Применяемый здесь термин "противоопухолевая активность" относится к ингибированию в опухолевых клетках межклеточной сигнализации, роста, метастазирования, клеточного деления, клеточной миграции, образования колоний в мягком агаре, контактного ингибирования, инвазивности, ангиогенеза, опухолевой прогрессии или другого злокачественного фенотипа или к индукции апоптоза. Особенно охватываются функциональные тесты, которые включают в себя критерии применения соединений согласно изобретению в качестве противогрибковых и противовирусных агентов, агентов, токсичных в отношении рыб, или моллюскицидов, контрацептивов, противогельминтных агентов, средств защиты от УФ, отхаркивающих средств, диуретиков, противовоспалительных агентов, регуляторов метаболизма холестерина, средств, влияющих на сердечно-сосудистую систему, противоязвенных агентов, анальгетиков, седативных средств, иммуномодуляторов, жаропонижающих средств, регуляторов ангиогенеза и в качестве агентов для снижения ломкости капилляров. Такие тесты должны быть хорошо известны специалистам в данной области техники в свете настоящего раскрытия. Так же как и прямые тесты активности in vitro и in vivo, данные тесты могут включать в себя измерения ингибирования связывания с субстратом, лигандом, рецептором или другим партнером по связыванию с соединением согласно изобретению.
XIV. Рост и тканевые культуры Acacia victoriae
Важным аспектом в получении описанных здесь монотерпеновых/тритерпеновых соединений должна быть доступность ткани Acacia victoriae. Как описано выше, соединения концентрируются в корнях и стручках Acacia victoriae, доступность данных тканей должна быть особенно важной. Другим источником для выделения соединений согласно изобретению являются молодые сеянцы. Acacia victoriae растет на юго-западе Соединенных Штатов и в Австралии и, следовательно, растительная ткань доступна для населения. Кроме того, авторами в американской коллекции типов культур (ATCC), 10801 University Belvd., Manassas, VA 20110-2209 (7 мая 1998 г) был создан депозит из 2500 семян Acacia victoriae. Данным, положенным на хранение семенам был присвоен No. 209835 поступления в ATCC. Депозит был сделан в соответствии с условиями и положениями Будапештского Договора, касающегося депозитов микроорганизмов, и сделан с условием по меньшей мере тридцати (30) лет и по меньшей мере пяти (05) лет после самого последнего запроса на предоставление образца депозита, полученного депозитарием, или в течение эффективного времени действия патента, каким бы длинным он не был, и должен быть заменен, если он становится нежизнеспособным в течение данного периода.
Таким образом, в свете настоящего раскрытия специалист в данной области техники мог бы посадить данные депонированные семена, вырастить из них растение и выделить из растений ткань для получения тритерпеновых соединений и пищевых добавок согласно изобретению. Также он мог бы выделить ткань из природно существующих популяций Acacia victoriae. Однако получение ткани для выделения монотерпеновых/тритерпеновых соединений могло бы быть более легко достигнуто, если бы были разработаны подходящие способы культивирования для размножения ткани Acacia victoriae. Одной из возможностей для получения ткани могло бы быть культивирование видов в большом масштабе. Более предпочтительные возможности, однако, включают в себя тканевые культуры Acacia victoriae и реализацию аэропонной системы выращивания.
(i) Способы аэропонного выращивания
Ряд преимуществ может быть реализован путем применения аэропонной системы культивирования Acacia victoriae. Во-первых, скорость роста растений приблизительно в два раза выше достигаемой с применением традиционных способов выращивания. Во-вторых, корни могут быть легко собраны, если это необходимо, без повреждения растений. Обрезка корней приводит в дальнейшем к интенсивному латеральному росту волокнистых корней. Поэтому корни можно было бы собирать несколько раз в год. В диких популяциях Acacia victoriae сбор стручков ограничен несколькими неделями в год, а сбор корней затруднен без повреждения или гибели растения.
Аэропонная система выращивания представляет собой закрытую систему, в которой корни растения подвешены в воздухе, затуманенном полным питательным раствором. Корни помещают в водонепроницаемый ящик, затуманиваемый с интервалами питательным раствором. Питательный раствор содержит все необходимые элементы, в которых нуждается растение, для его полного жизненного цикла. Несмотря на тот факт, что разные растения нуждаются в разном уровне и составах для оптимального роста, удовлетворительные результаты дает общий единожды сбалансированный раствор.
(ii) Тканевые культуры Acacia victoriae
Тканевые культуры представляют собой другую возможность для культивирования Acacia victoriae. Для создания тканевых культур семена Acacia victoriae тщательно промывают в водопроводной воде с антимикробным мылом и обрабатывают 20% раствором имеющегося в продаже отбеливателя в течение 15 минут. После повторной промывки в деионизированной воде семена обрабатывают кипящей водой для индукции прорастания и инкубируют в течение ночи. На следующее утро семена вновь один раз дезинфицируют имеющимся в продаже отбеливателем и ополаскивают 2-3 раза в стерильной деионизированной воде. Семена после удаления загрязнений культивируют на среде MS (Murashige et al., 1962) с добавкой витаминов MS и 2% сахарозы (для эксплантантных культур применяли 3% сахарозу) и среду превращают в гель либо с помощью 0,7% агара, либо 0,2% гельрита.
Применяемые для культивирования эксплантаты могут включать в себя потенциально любой тип ткани, включая кончики побегов, узловые сегменты, гипокотили и сегменты корней. Эксплантаты обычно культивируют только на среде MS или MS с добавкой регуляторов роста, таких как IAA, NAA, IBA, 2,4-D и BAP (по отдельности или в сочетании). Культуры обычно поддерживают при 25±2°С при 16-часовом световом фотопериоде при освещенности 1000 люкс, создаваемой холодными белыми флуоресцентными трубками. Полученные проростки содержат во влажной среде в теплице 1 месяц, для того чтобы они окрепли перед переносом их в теплицу, поле или аэропонную систему выращивания.
В настоящем изобретении были разработаны культуры волосистых корней Acacia victoriae. Инфицирование растения штаммом R-1000 Agrobacterium rhizogenes ведет к интеграции и экспрессии T-ДНК в растительном геноме, что вызывает развитие волосистых корней. Культуры волосистых корней быстро растут, проявляют плагиотропный рост корней и являются сильно разветвленными на свободной от гормонов среде, а также проявляют высокую степень генетической стабильности (Aird et al., 1988). Генетическая трансформация и индукция волосистых корней у Acacia victoriae и оптимальные условия для роста подробно описаны в разделе примеров. Культуры волосистых корней обеспечивают быстрый рост ткани в большом масштабе, что может быть использовано для выделения монотерпеновых/тритерпеновых соединений.
Преимущество культивирования ткани заключается в том, что могут быть потенциально получены клональные культуры, которые экспрессируют монотерпеновые/тритерпеновые соединения. Данные культуры могли бы выращиваться в большом масштабе и потенциально могли бы быть расширены до ростовой системы в промышленном масштабе для получения растительной ткани для выделения монотерпеновых/тритерпеновых соединений. Кроме того, растения, регенерированные из тканевых культур, часто проявляют значительную вариабельность. Следовательно, с применением тканевых культур могут быть получены клональные клеточные линии или растения, регенерированные из таких культур, которые являются "элитными" в отношении продукции ими описанных здесь монотерпеновых/тритерпеновых соединений. Полученные растения могли бы себя проявить среди своей генерации и быть отобранными при каждой генерации потомков для получения истинно селекционированных элитных линий.
Однако элитные сорта не обязательно должны происходить из тканевых культур, поскольку значительная генетическая вариабельность существует в диких популяциях Acacia victoriae. Поэтому авторы предусматривают, что генетическая вариация, обнаруженная в диких популяциях Acacia victoriae, включает в себя вариации генов, контролирующих эндогенный уровень продукции монотерпеновой/тритерпеновой композиции. Если так, должна быть возможна идентификация тех представителей популяций Acacia victoriae, которые продуцируют повышенный уровень монотерпенов/тритерпенов по сравнению с другими представителями диких популяций, и отбор данных вариантов для применения в системах выращивания, направленных на получение ткани для выделения монотерпеновых/тритерпеновых соединений. Система выращивания может составлять, например, традиционное выращивание на ферме, аэропонные способы выращивания, культивирование ткани или любой другой подходящий способ для размножения ткани Acacia victoriae. Далее данные растения могут быть отобраны для применения в протоколах размножения для получения вариантов, которые являются более элитными и которые являются также истинно селекционированными.
XV. Определения
В данном описании термин «часть» может относиться к одной, двум, трем или более частям.
Активные составляющие относятся к самым чистым экстрактам, которые сохраняют активность. В настоящем изобретении "активный компонент" или "активное соединение" относится к активным тритерпеновым соединениям, идентифицированными авторами согласно изобретению. Данные соединения очищали и идентифицировали, например, во фракции UA-BRF-004-DELEP-F094.
Стручки определяются как семенные стручки Acacia victoriae.
Термин "цитотоксический" определяется как вызывающий гибель клетки, в то время как термин "цитостатический" определяется как ингибирующий рост и/или пролиферацию клеток.
Апоптоз определяется как нормальный физиологический процесс запрограммированной гибели клеток, который наступает в процессе эмбрионального развития и в процессе поддержания тканевого гомеостаза. Процесс апоптоза может быть подразделен на серии метаболических изменений в клетках, подвергающихся апоптозу. Могут быть определены индивидуальные ферментативные стадии нескольких регуляторных путей или путей передачи сигнала для демонстрации того, что в клетке или в популяции клеток наступает апоптоз, или того, что процесс клеточной гибели нарушается в раковых клетках. Программу апоптоза наблюдают также по морфологическим характеристикам, которые включают в себя изменения в плазматической мембране (такие как потеря асимметрии), конденсацию цитоплазмы и ядра, внутринуклеосомное расщепление ДНК. Это является кульминационной стадией гибели клетки, так как клетка дегенерирует в "апоптотические тела".
Для мультипараметрического исследования апоптоза разработаны способы определения нескольких ферментативных и сигнальных процессов, вовлеченных в апоптоз, в виде стандартных протоколов. Одним примером ранней стадии апоптоза является высвобождение цитохрома с из митохондрий с последующей активацией пути каспазы-3 (PharMingen, San Diego, CA). Индукция каспаз (групп цитозольных протеаз) является одной из наиболее постоянно наблюдаемых черт апоптоза. В частности, каспаза-3 играет центральную роль в данном процессе. Когда каспазы активируются, они расщепляют белки-мишени; одним из наиболее важных из них является PARP (поли-(АДФ-рибоза)-полимераза, которая является белком, локализованном в ядре). Следовательно, тесты, определяющие высвобождение цитохрома с, определяющие активность каспазы-3 и определяющие деградацию PARP, являются эффективными определяющими способами тестирования апоптоза.
Более того, агенты, которые вызывают высвобождение цитохрома с из митохондрий раковых клеток, могут рассматриваться как весьма пригодные для терапии с целью восстановления по меньшей мере в некоторых аспектах клеточного контроля над запрограммированной гибелью клеток.
Другим тестом апоптоза является определение аннексина-V (BioWhitaker, Walkerville, MD). Обычно фосфотидилсерин (PS) локализован на внутренней поверхности плазматической мембраны. Однако в течение ранних стадий апоптоза может иметь место экстернализация PS. Аннексин-V является Ca-связывающим белком, который связывается с PS, и он может быть обнаружен с помощью окрашивания ФИТЦ на аннексин-V и проточной цитометрии (Martin et al., 1995). Способность клеток, обработанных соединениями Acacia victoriae, описанными в данном изобретении, связывать аннексин-V рассматривается как указание на то, что клетки подверглись процессу апоптоза.
В других примерах авторы применяли тест с PI-3-киназой для определения активности в отношении апоптоза клеток, обработанных смесями противораковых соединений, выделенных из Acacia victoriae. Фосфоинозитид-3-киназа (PI3K), фермент, связанный с клеточной мембраной, способна фосфорилировать фосфатидилинозитол в положении 3 инозитольного кольца, определяя в результате этого новый липидный путь проведения сигнала в тех клетках, где активна PI3K. Когда PI3K активна, киназа, называемая AKT, рекрутируется к клеточной мембране. AKT является продуктом онкогена, который каталитически активируется после перехода в мембрану. Полностью активированная AKT играет решающую роль в выживании клеток. Путь PI3K/AKT создает механизм, с помощью которого клетки избегают апоптоза. Таким образом, средства, ингибирующие PI3K в малигнизированных клетках, рассматриваются как весьма пригодные для терапии с целью восстановления по меньшей мере в некоторых аспектах клеточного контроля над апоптозом.
Нетипичная пролиферация определяется как группа генетически определяемых изменений, которые наступают в клетках млекопитающих при патологическом состоянии, известном как рак. Данный процесс в конечном итоге ведет к потере контроля над апоптозом раковых клеток. Это может наступать в течение стадий, обычно обозначаемых как 1) инициация, которая определяется как стадия, при которой внешний агент или стимул запускает генетическое изменение в одной или более клеток, и 2) промоция, которая определяется как стадия, при которой вовлекаются дополнительные генетические и метаболические изменения, которые могут включать в себя воспаление. В течение "стадии промоции" клетки начинают метаболический переход в стадию клеточного роста, в которой блокируется апоптоз.
Малигнизированные клетки определяются как раковые клетки, в которых обойдены нормальные механизмы контроля роста в процессе серий метаболических изменений в течение стадий инициации и промоции наступления злокачественного перерождения. Данные изменения являются следствием генетических изменений в клетках (либо активирующих мутаций и/или увеличенной экспрессии протоонкогенов - и/или инактивирующих мутаций и/или пониженной экспрессии одного или более генов - супрессоров опухолей). Большинство продуктов онкогенов и генов - супрессоров опухолей являются компонентами путей передачи сигналов, которые контролируют вход или выход клетки из клеточного цикла, стимулируют дифференцировку, повреждение смысловой ДНК и инициируют механизмы репарации, и/или регулируют программы клеточной смерти. Клетки применяют множественные параллельные механизмы для регуляции клеточного роста, дифференцировки, контроля повреждений ДНК и апоптоза. Приблизительно все опухолевые и малигнизированные клетки имеют мутации во множестве онкогенов и генов-супрессоров опухолей.
Экстракт или фракция относится к последовательным образцам, собранным из тканей с помощью разных способов. Данные "экстракты" или "фракции" могут быть проанализированы на желаемую противоопухолевую активность и дополнительно "проэкстрагированы" или "отфракционированы" для получения существенно более чистых компонентов, соответствующих активному компоненту.
Монотерпеновые/тритерпеновые соединения или монотерпеновый/тритерпеновый гликозид относятся к идентифицированным здесь новым и/или биологически активным сапониновым соединениям из Acacia victoriae. Монотерпеновое/тритерпеновое соединение или монотерпеновые/тритерпеновые гликозиды не обязательно должны быть выделены из Acacia victoriae, так как специалист в данной области в свете текущего раскрытия может выделить соединения из родственных видов или химически синтезировать аналоги монотерпеновых/тритерпеновых соединений или гликозидов, как здесь раскрыто. Альтернативно, можно биохимически синтезировать соединения. "Монотерпены" согласно изобретению включают в себя описанные здесь сапониновые соединения, которые имеют по меньшеймере монотерпеновую единицу(ы). Монотерпены могут быть дополнительно необязательно присоединены к "несущей части", которая позволяет им транспортироваться в клетку, и носитель может представлять собой тритерпеновые гликозиды, сахар или сахарид, липид, липофильный белок или любую часть, которая придает мембране проницаемость для монотерпена. Дополнительно монотерпен может быть связан с дополнительными химическими функциями, в результате чего модификации не нарушают биологическую активность соединений, как описано в разделе, обозначенном как краткое изложение существа изобретения. Так как монотерпены могут быть присоединены к тритерпенам, соединения обозначаются как "монотерпеновые/тритерпеновые соединения или гликозиды". "Тритерпены" данного изобретения включают в себя описанные здесь сапониновые соединения, которые имеют по меньшеймере тритерпеновую(вые) единицу(ы), и, в случае тритерпеновых гликозидов, сахар или сахарид и по меньшей мере один монотерпен. Данные термины относятся также к соединениям, содержащим дополнительные части или химические функции, включая, но не ограничиваясь этим, монотерпеновые единицы, как будет ясно из последующего описания. Таким образом, тритерпены данного изобретения также включают в себя агликоны, образованные в результате гидролиза сахарных единиц, и они дополнительно включают в себя другие модификации тритерпеноидных соединений, в результате чего модификации не нарушают биологической активности соединений.
XVI. Примеры
Следующие примеры включены для демонстрации предпочтительных осуществлений изобретения. Специалистам в данной области техники должно быть ясно, что раскрытые в следующих примерах способы представляют собой способы, которые, как обнаружили заявители, хорошо работают при осуществлении изобретения на практике, и таким образом могут рассматриваться в качестве предпочтительных способов его реализации. Однако в свете настоящего раскрытия специалистам в данной области техники должно быть понятно, что в конкретных раскрытых осуществлениях могут быть сделаны многочисленные изменения, и при этом могут быть получены подобные или сходные результаты без выхода за пределы концепции, духа и объема изобретения. Конкретнее, должно быть ясно, что определенные агенты, близкие между собой и химически, и физиологически, могут быть заменены описанными здесь агентами для получения тех же самых или сходных результатов. Все такие сходные замены и модификации, очевидные для специалистов в данной области техники, рассматриваются как находящиеся в пределах духа, объема и концепции изобретения, как это определено в прилагаемой формуле изобретения.
Пример 1
Предварительный скрининг и очистка активных противоопухолевых компонентов из Acacia victoriae
Из Проекта пустынных видов стручковых (DELEP) было выбрано шестьдесят видов растений с целью идентификации новых соединений, обладающих благоприятными биологическими свойствами. DELEP (Университет Аризоны, Tucson) является коллекцией пустынных видов стручковых, созданной благодаря сотрудничеству между Университетом Аризоны и Юго-западным питомником Boyce Thompson. Образцы с экспериментальных делянок для каждого из видов растений собирали, сушили на воздухе в течение 3-4 дней, перемалывали до частиц с размером три миллиметра с помощью мельницы Wiley (сито с размером 3 мм) и экстрагировали два или три раза путем процеживания со смесью 1:1 дихлорметана (ДХМ) и метанола (MeOH). Каждая экстракция процеживанием длилась по меньшей мере в течение 5 часов и часто продолжалась ночью. Большую часть экстрагированной биомассы собирали из двух первых процеживаний. После этого биомассу промывали метанолом в объеме, равным половине пустого объема, и выделяли сырой экстракт, содержащийся в аликвотах метанола. Обычно образцы выделяли и готовили для биологического тестирования путем удаления метанола в вакууме, пропускания водной фазы через частицы RP-C18, обнаружения активных компонентов в MeOH и последующего выпаривания MeOH на роторном испарителе для сбора экстракта в виде твердого вещества. После этого сырой экстракт ресуспендировали в H2O, ДМСО или их смесях (менее полярные соединения ресуспендировали в ДМСО, а более полярные соединения ресуспендировали в воде или в смесях воды и ДМСО; агликоны ресуспендировали в ДМСО).
Каждый экстракт затем подвергали скринингу с применением панели опухолевых и неопухолевых клеток человека, включая линии клеток рака яичников, T-лейкозные клетки, эпидермоидные клетки человека, клетки рака молочной железы человека, клетки рака предстательной железы человека, фибробластные клетки крайней плоти человека, эндотелиальные клетки человека и раковые клетки почек человека. Клетки сначала помещали в 96-луночные планшеты на 18-24 часа при 37°С. Затем клетки экспонировали с различными концентрациями растительных экстрактов, инкубировали в течение 72 часов при 37°С и окрашивали либо MTT (3-(3,5-диметилэтиазол-2-ил)-2,5-дифенилтетразолийбромид; Sigma Chemical Co.) в течение 4 часов или кристальным фиолетовым (Sigma Chemical Co.) в течение 20 минут при комнатной температуре. В планшеты с MTT добавляли лизирующий буфер (20% додецилсульфат натрия в 50% ДМФ) и инкубировали еще в течение 6 часов перед проведением считывания ОП при 570 нм. Планшеты с кристальным фиолетовым промывали, краситель экстрагировали в течение 3-4 часов буфером Соренсона (0,1 М цитрат натрия (pH 4,2), 50% об/об этанол) и планшеты считывали при 570-600 нм (Mujoo et al., 1996). Указанием на цитотоксичность подвергаемых скринингу экстрактов служило сравнение показаний ОП для пустой среды обработки и клеток, обработанных растительным экстрактом. Рассчитывали процент цитотоксичности по формуле 100 - % от контроля, где % от контроля = [((ОП клеток, обработанных растительным экстрактом (обработанный образец))/(ОП клеток, экспонированных с пустой средой (необработанный образец))) x 100].
По результатам первоначального скрининга один растительный экстракт проявил сильное ингибирование роста раковых клеток и при этом показал слабую токсичность в отношении нормальных фибробластных клеток человека. Данный экстракт, обозначенный как UA-BRF-004-DELEP-F001, был выделен из стручкового растения Acacia victoriae. Экстракт проявлял IC50 при приблизительно 12 мкг/мл (SKOV-3), 26 мкг/мл (OVCAR-3) и 13 мкг/мл (HEY) при применении линий раковых клеток яичников человека; при более 50 мкг/мл (A375-M) и при приблизительно 38 мкг/мл (HS294T) при применении клеток меланомы человека; при приблизительно 15 мкг/мл для эпидермоидных клеток человека (A431) и при более 50 мкг/мл для клеточной линии MDA-468 рака молочной железы (фиг. 1) (см. описание клеточных линий в примере 13). В отношении нормальных фибробластных клеток крайней плоти человека (FS) и фибробластных клеток мыши (L929), обработанных тем же экстрактом, цитотоксичность не наблюдалась.
По результатам ТСХ данный экстракт, похоже, содержал много составляющих. Поэтому исходно усилия были направлены на очистку данного экстракта для выделения активных компонентов, обеспечивающих избирательную цитотоксичность. Из исходного экстракта в соответствии с общей схемой, показанной на фиг. 15, были выделены хроматографические фракции, обогащенные активными компонентами.
Исходный экстракт, UA-BRF-004-DELEP-F001, получали из 538 г растительного вещества из Acacia victoriae с помощью процеживания (дважды), как описано выше. Экстракт затем сушили в вакууме, что дало приблизительно 52,0 г порошка. 51,5 г высушенного вещества обрабатывали 3 раза 1 л этилацетата (EtOAc). Приблизительно 15,75 г растворимого в EtOAc вещества подвергали колоночной хроматографии на силикагеле (1,5 кг). Было получено пятьдесят четыре подфракции по 670 мл при элюировании с применением смесей с возрастающей полярностью гексана, EtOAc и MeOH. 54 подфракции были собраны в тринадцать отдельных фракций, обозначенных как UA-BRF-004-DELEP-F006 по UA-BRF-004-DELEP-F018. Данные фракции были подвергнуты скринингу на противоопухолевую активность с применением описанной выше процедуры. Ни одна из проверенных фракций не проявила высокой противоопухолевой активности, наблюдавшейся в случае UA-BRF-004-DELEP-F001.
Нерастворимое в EtOAc вещество (приблизительно 34,7 г) также было подвергнуто хроматографии на силикагеле (1,7 кг). Была получена пятьдесят одна подфракция по 670 мл и три дополнительные подфракции общим объемом 21 л при элюировании с применением смесей с возрастающей полярностью ДХМ, MeOH и воды. Данные подфракции были собраны в восемь отдельных фракций, обозначенных как UA-BRF-004-DELEP-F019 по UA-BRF-004-DELEP-F026, согласно таблице 7.
Элюирование фракций UA-BRF-004-DELEP-F019 по UA-BRF-004-DELEP-F026
10% MeOH/ДХМ (7-12)
20% MeOH/ДХМ (13)
35% MeOH/ДХМ (19)
39-40
65% MeOH/ДХМ (39)
100% MeOH (40)
41-51
100% MeOH (41-51)
20% вода/MeOH (6 л)
После этого каждую фракцию подвергали скринингу на противоопухолевую активность с применением панели опухолевых клеток человека, как описано выше для сырого экстракта. Одна из фракций, UA-BRF-004-DELEP-F023, проявила противоопухолевую активность, более высокую, чем активность UA-BRF-004-DELEP-F001. Данные результаты показали, что 6 мкг/мл фракции UA-BRF-004-DELEP-F023 проявляют 50% (OCC1), 63% (SKOV-3), 83% (HEY) и 48% (OVCAR-3) цитотоксичность на клетках рака яичников человека; приблизительно 60% цитотоксичность на раковых клетках предстательной железы человека (LNCaP); приблизительно 92% цитотоксичность на лейкозных клетках (Jurkat) и приблизительно 73% цитотоксичность на свежих раковых клетках яичников человека из асцитов больных (FTC). Биотесты на нетрансформированных клетках показали IC50 10,6 мкг/мл для клеток FS и 23 мкг/мл для клеток HUVEC (фиг. 2).
Биологически активный(е) компонент(ы) в UA-BRF-004-DELEP-F023 дополнительно очищали с помощью множественных разделений способом жидкостной хроматографии при среднем давлении (MPLC) с обращенной фазой с целью выделения и характеристики активного(ых) компонента(ов). Образцы элюировали из дегазированных смесей с возрастающей концентрацией ацетонитрила (ACN) в воде инкрементом 10% на 4 л согласно следующим ступеням: 0, 10%, 20%, 30%, 40% смеси ACN/вода. После этого фракцию объемом 2-4 л элюировали MeOH. После повторных разделений были собраны десять фракций, обозначенные UA-BRF-004-DELEP-F027 по UA-BRF-004-DELEP-F036 согласно таблице 8.
Элюирование фракций UA-BRF-004-DELEP-F027 по UA-BRF-004-DELEP-F036
При ТСХ некоторые из данных фракций выглядели сходным образом. Было обнаружено, что одна из фракций с наибольшим выходом, UA-BRF-004-DELEP-F035 (фракция 35), проявляет противоопухолевую активность.
Скрининг UA-BRF-004-DELEP-F035 на противоопухолевую активность показал IC50 3,0, 1,2, 2,0 и 3,5 мкг/мл, соответственно, против линий клеток рака яичников человека HEY, SKOV-3, OVCAR-3 и C-1 (резистентная к цисплатину OVCAR-3); IC50 2,4 мкг/мл против раковых клеток поджелудочной железы (Panc-1); IC50 1,2 мкг/мл, 3,0 мкг/мл и 3,7 мкг/мл, соответственно, против линий клеток рака почки 769-P, 786-0 и A498; IC50 130 нг/мл против T-лейкозных клеток Jurkat и IC50 от 1 до 3 мкг/мл против линий B-лейкозных клеток KG1, REH и NALM-6 (фиг. 3, 4). Как показано в таблице 9, очистка сырого растительного экстракта резко повышала биологическую активность.
Цитотоксичность сырых экстрактов по сравнению с UA-BRF-004-DELEP-F035
Фракция 35 проявляла IC50 приблизительно 4,7 мкг/мл в отношении нормальных клеток человека FS и IC50 приблизительно 13,3 мкг/мл в отношении нормальных клеток человека Hs27. При оценке действия фракции 35 (F035) на нормальные эритроидные и миелоидные колонии человека (клетки выделяли из костного мозга) наблюдалось 12-18% ингибирование при 3,0 мкг/мл (таблица 10).
Действие фракции 35 на эритроидные и миелоидные колонии
В свете представленных выше результатов, указывающих на противоопухолевую активность фракции 35, было проведено биологическое тестирование, как описано выше, с применением низких концентраций фракции 35, таких низких как 0,095 мкг/мл. В данном исследовании применяли различные концентрации фракции 35 против расширенной панели опухолевых линий. Результаты скрининга показывают, что даже в концентрации 1,56 мкг/мл фракция 35 проявляла значительную противоопухолевую активность в отношении ряда клеточных линий (таблица 11).
Таблица 11
Цитотоксичность различных концентраций UA-BRF-004-DELEP-9F035 против различных линий опухолевых клеток
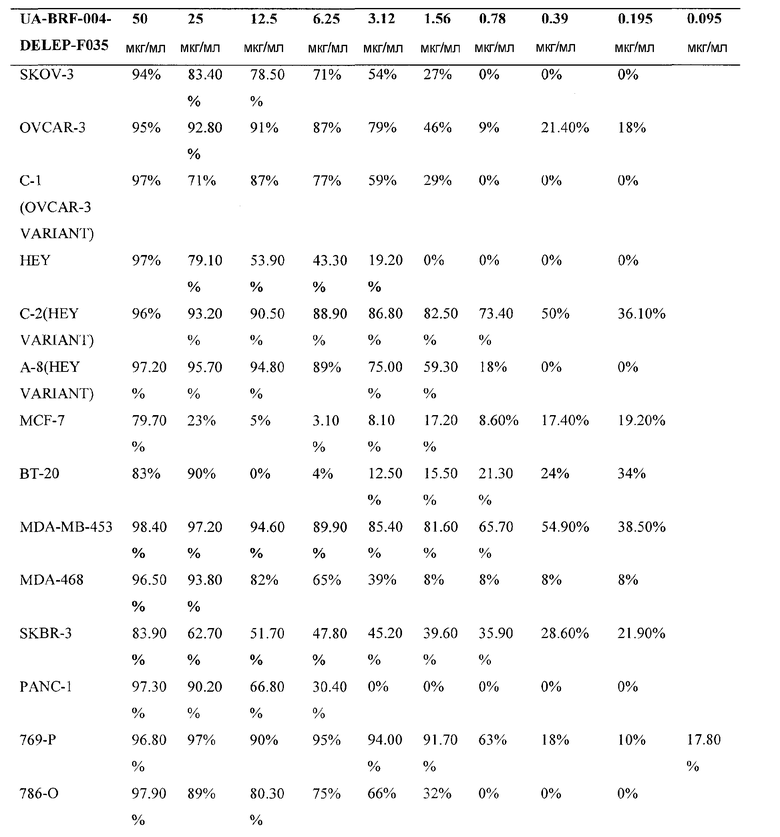
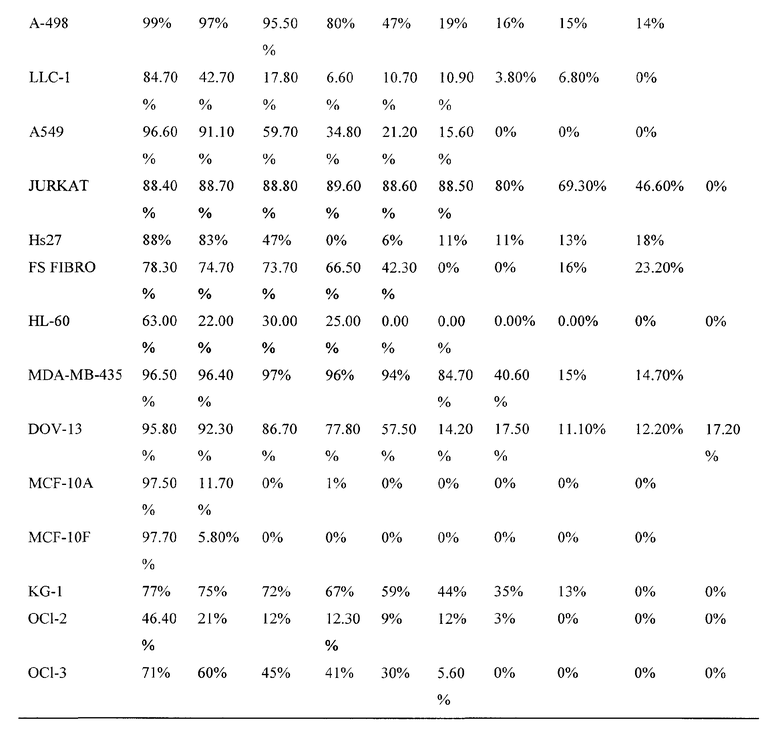
Пример 2
Способы выделения активных составляющих из Acacia victoriae
Была разработана процедура прямого получения фракций, содержащих активные составляющие, содержащиеся в UA-BRF-004-DELEP-F035, выделенной в ходе предварительной очистки, подробно описанной в примере 1. Приблизительно 9665 г свежесобранной ткани кожуры от Acacia victoriae перемалывали в бильной мельнице с ситом 3 мм и затем экстрагировали 80% MeOH в H2O (3 раза) с последующей фильтрацией. Было отброшено 8200 г жмыха. Отдельно были собраны три промывки, и фракциям были присвоены следующие обозначения: F068 в 21,5 л (первая промывка); F069 в 24 л (вторая промывка) и F070 в 34,3 л (третья промывка). F068 дополнительно очищали путем разделения на аликвоты по 1 л, добавляя к каждой аликвоте 400 мл H2O и промывая CHCl3 (2 x 250 мл). Объединенным полярным фазам (28,5 л) было присвоено обозначение F078, а объединенным органическим фазам - F079 (давшим 42 г после удаления органического растворителя с помощью роторного испарителя).
MeOH удаляли из F078 в вакууме и F078 далее фракционировали с помощью RP MPLC с применением колонки 29 x 460 мм, заполненной 530 г регенерированного Bakerbond RP-C18 с частицами 40 мкм. На колонку наносили 500 мл водного раствора и собирали фракции согласно таблице 12.
Элюирование фракций F091 по F094
Каждую фракцию затем сушили путем удаления MeOH, пропуская через частицы C-18, захватывающие MeOH, и выделяли в виде твердого вещества в вакууме. Твердое вещество ресуспендировали в воде и подвергали тестированию на противоопухолевое действие (для некоторых менее полярных фракций к воде добавляли ДМСО; агликоны ресуспендировали в ДМСО). Результаты показали, что биологическая активность элюата 100% MeOH, обозначенного F094, по существу эквивалентна таковой фракции UA-BRF-004-DELEP-F035 (таблица 13). F093 также содержала активные составляющие. Химическое сходство фракций F094 и F095 было подтверждено с помощью ТСХ и ВЭЖХ, хотя F094, по-видимому, содержала дополнительные компоненты.
Таблица 13
Цитотоксичность варьирующих концентраций F094 против различных линий опухолевых клеток

F094 дополнительно фракционировали согласно таблице 14, анализировали с помощью ТСХ и подвергали биологическому тестированию для получения очищенного(ых) активного(ых) компонента(ов). Результаты биотестирования варьирующих количеств полученных фракций (F138-F147) даны в таблице 14.
Элюирование фракций F138 по F147
7-8 (50)
70% MeOH в воде (7-8)
18 (100)
100% MeOH в воде (18)
Таблица 15
Биотестирование фракций F137, F140, F144 и F145
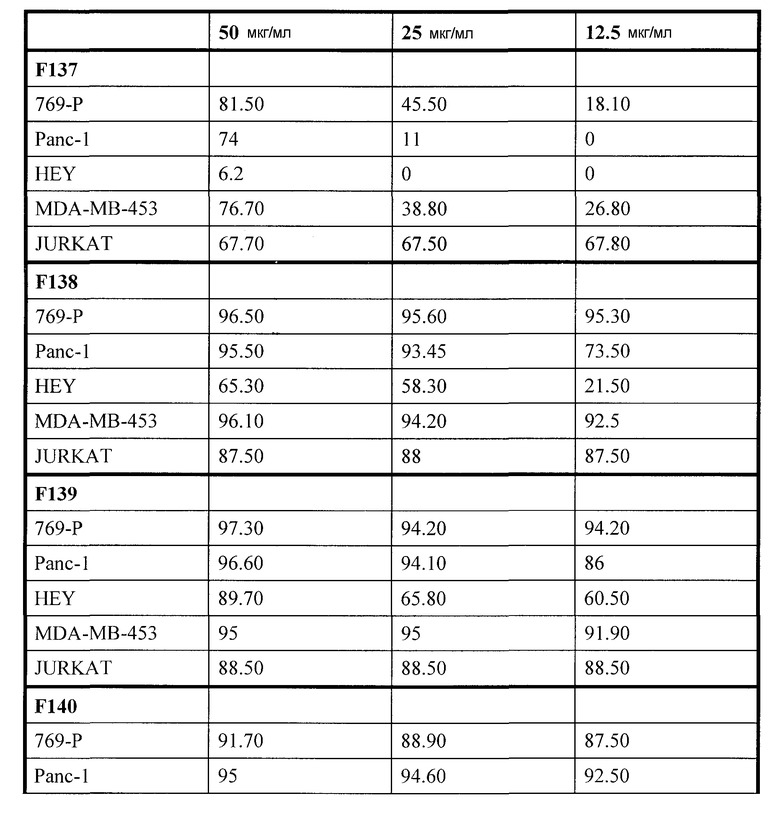


Процент ингибирования роста
Хотя указанные выше процедуры были ориентированы на выделение активных составляющих из стручков Acacia victoriae, активные составляющие могут быть также экстрагированы из корней. В данном случае корни измельчают в течение 1/2 часа и заливают 100% MeOH. Смесь затем фильтруют и разбавляют до 80% MeOH в воде. Если экстракции подлежат большие количества корней, то может быть предпочтительным экстрагировать посредством продавливания, как описано выше. Причина различий в данных процедурах экстракции заключается в том, что корни обычно экстрагируют свежими, тогда как стручки часто сушат перед экстракцией.
Пример 3
Процедура выделения активных составляющих из фракции UA-BRF-004-DELEP-F094 в препаративном масштабе
Для получения смесей активных составляющих из фракции UA-BRF-004Pod-DELEP-F094 (F094) в более крупном масштабе применяли модифицированную процедуру экстракции/разделения. Данную процедуру повторяли много раз с постоянным выходом высокоактивных фракций. Обычно 20-25 г F094 или ее эквивалента растворяли в 150-175 мл 50% MeOH в H2O и затем наносили на колонку ((26 мм x 460 мм) + (70 мм x 460 мм) RP-C18, 40 мкм, 1200 г, уравновешенную 60% MeOH/H2O). Фракции элюировали ступенчато 8 л 60% MeOH/H2O; 7,5 л 70% MeOH/H2O и 2 л MeOH, и фракции обозначали, как показано в таблице 16. Фракция F035-B2 содержит смесь активных составляющих, содержащихся в F094, F133-F136 (выделенных из F093) и F138-147 (выделенных из F094), как показано на фиг. 18A-18F. F094 является приемлемой заменой F035 со снижением эффективности от однократного до двукратного, а F035-B2 обладает меньшей эффективностью по сравнению с F094.
Выделение F035-B2
Была разработана процедура для дополнительной очистки активных компонентов фракции F035-B2 для получения фракций, обладающих характеристиками UA-BRF-004-DELEP-F035 при аналитической ВЭЖХ. Процедура состоит в следующем. Проводят дополнительную препаративную ВЭЖХ с фракцией F035-B2 с применением 10-микронных хроматографических колонок с обращенной фазой для элюирования ступенчатым градиентом смесей ацетонитрила и воды, варьирующим от 26% ацетонитрила в воде до 40% ацетонитрила в воде при шаговом градиенте 1-2% с последующей промывкой 100% ацетонитрилом и 100% метанолом, что обычно дает уникальное разделение F035-B2 на несколько фракций, содержащих от 0 до 20 пиков в минуту (при стандартном шестимикронном аналитическом способе ВЭЖХ RP-18), которые не содержатся в исходной фракции F035 (фиг. 18A). Остальные полученные фракции обеспечивали фракционирование от одного до трех компонентов F035. Как показано тестированием, приведенным выше, фракции F139-F147 сходны с данными фракциями при некотором повышении противоопухолевой активности в отношении определенных линий опухолевых клеток относительно других. Уникальная смесь активных компонентов, имеющихся во фракции F035, может быть получена в большом количестве путем модификации исходной композиции растворителей заднего фронта между 16% и 26% ацетонитрила в воде с последующей очисткой с помощью MPLC для получения многограммовых количеств эквивалента UA-BRF-004-DELEP-F035.
Могут быть применены дополнительные усовершенствования описанной выше процедуры экстракции, а также другие раскрытые здесь процедуры экстракции, с использованием смесей из трех растворителей, ацетонитрила, метанола и воды. Диапазоны процентного содержания могут быть получены и оптимизированы в процессе работы персоналом, знакомым со стандартными способами хроматографии. Сходным образом силикаты неподвижной фазы можно варьировать путем применения комбинированных систем ОФ, включая, но не ограничиваясь этим, C8, CN, диметилдиол и C18. На конечных стадиях даже двуокись кремния с обычной фазой может быть применена для процедур конечной очистки.
Пример 4
Альтернативные процедуры выделения активных составляющих из Acacia victoriae
Фракции F094 (250 г), F035 (50 мг) и измельченные стручки Acacia victoriae (1 кг) (т.е. порошок семенных стручков) получали, как описано выше. Примененная для анализа фракции F094 аналитическая методология и последующее фракционирование F094 описаны в данном примере.
4.1 Аналитическая методология
В попытке разделения фракции F094 было применено несколько способов, включая колонки C8 и C18 при градиентных и изократических условиях. Мониторинг включал как УФ при 220 нм, так и определение рассеяния света при испарении (ELSD). Лучшее разделение пиков наблюдалось с подвижными фазами, содержащими трифторуксусную кислоту (ТФУ). Ниже описан способ, названный здесь Acacia 257. Данный способ обеспечивает хорошее разрешение при малом времени разделения.
ВЭЖХ снабжали детектором диодной матрицы (DAD) или детектором варьируемой длины волны и колонкой 4,6 X 150 мм Inertsil C18 3 мкм (MetaChem). Детектор устанавливали на 220 нм.
Пропускали следующий градиент:
На фиг. 25 показана хроматограмма F094, полученная данным способом. F094 состоит из трех групп или семейств с рядом пиков в каждом семействе. Семейство-1 (от 8 до 20 мин; пики A-D), семейство-2 (от 22 до 35 мин; пики E-H) и семейство-3 (от 36 до 47 мин; пики I-L). Фракция F035 также была проанализирована данным способом, и хроматограмма показана на фиг. 26. В F035 пики второго семейства обогащены по сравнению с F094, в большей степени представлены пики первого семейства.
4.2 Фракционирование
4.2.1 Первое фракционирование
Первое фракционирование было направлено на пики семейства-1. Для данной цели применяли пропорциональную C8 полупрепаративную колонку (7,8 X 300 мм, 7 мкм) (Waters) с программой градиентного элюирования, как показано ниже. Было получено семь подфракционных частей, как показано на фиг. 27. Последняя часть фракции (# 2160-007-31) включала все пики семейства-2 и семейства-3. Данные фракции, а также исходное вещество (F094) были отосланы для биологического тестирования:
4.2.2 Второе фракционирование
Целью данного фракционирования являлось разделение пиков второго семейства соединений. Этого достигали путем применения тех же полупрепаративных колонок C8. Подвижной фазой служил изократический 32% ацетонитрил в воде, содержащей 0,1% ТФУ. Запись хроматографии с указанием границ семи фракций показана на фиг. 28. Отделенная здесь первая фракция включает в себя все пики семейства-1.
4.2.3 Биотесты
Результаты биологического тестирования подфракций первого и второго фракционирования приведены в таблицах 17 и 18, соответственно.
Цитотоксичность подфракций первого фракционирования в отношении клеток Jurkat
Цитотоксичность подфракций второго фракционирования в отношении клеток Jurkat
Из фракции F094 Acacia было получено два тритерпеноидных гликозида, а именно D1 и G1. Кислотный гидролиз D1 дает агликон.
4.4 Фракционирование в препаративном масштабе для получения пиков области D и G/H
F094 (2,3 г) фракционировали на колонке ВЭЖХ Prep PFP (пентафторфенила) (50x250 мм, 10 мкм). Подвижной фазой служила смесь ацетонитрил/вода, содержащая 0,1% трифторуксусную кислоту (ТФУ), пропускаемая градиентным способом от 27% до 32% ацетонитрила на протяжении 38 мин. Как показано на фиг. 29, данным способом были разделены фракции, содержащие пики D и G/H. Разделенные фракции данного препаративного разделения были отосланы для биологического тестирования. Аналитические тесты D и G/H показаны на фиг. 30 и 31. Примененный здесь способ представляет собой способ Acacia 257, который был описан выше в этом же разделе.
Фракцию G/H дополнительно очищали сначала на препаративной колонке PFP для получения G1 с 68% хроматографической чистотой, и данное вещество далее очищали на полупрепаративной колонке C-18 для получения чистого G1.
Приблизительно 100 мг смеси G/H наносили на описанную выше колонку PFP. Пропускали следующий градиент:
Было собрано пять фракций (G1, G2, G3, H1 и H2). Анализ G1 (фиг.32) показал хроматографическую чистоту 68%. Данную фракцию дополнительно очищали на полупрепаративной колонке.
Применяли колонку YMC C18-Aq (10x250 мм, 5 мкм). Конечный продукт G1 имел 100% хроматографическую чистоту (фиг. 33).
Фракцию D (45% D1) с препаративной колонки PFP сначала фракционировали на колонке C-18 Waters (25 x 100 мм). Подвижной фазой служил 61% метанол в воде с 0,1% ТФУ. Анализ ВЭЖХ показал, что чистота D1 составляет 78% (фиг. 34) и что она содержит дополнительный пик (обозначенный D1.5). Образец D1 со 100% хроматографической чистотой получали дальнейшим фракционированием нечистого D1 на колонке YMC C18-Aq с применением смеси 33% ацетонитрил/вода с 0,1% ТФУ в качестве подвижной фазы. Было обнаружено, что D1 более стабилен в разбавленных растворах кислоты, чем в воде при 40°С. Поэтому во время очистки D1 в растворители включали 0,1% ТФУ.
4.4.1. Биотесты
Биотесты проводили на клеточных линиях Jurkat, и эффекты различных подфракций и чистых D1 и G1 показаны в таблицах 19, 20 и 21, соответственно. D1 и G1 тестировали при двух разных величинах pH. Результаты показали слегка повышенную активность при pH 6,5 по сравнению с pH 7,5. Однако клеточный рост ингибировался приблизительно на 40% при более низких значениях pH.
4.4.2 Кислотный гидролиз D1
Сапонин D1 в этаноле гидролизовали 3 н. HCl в течение 3 час при 100°С. Полученный агликон очищали с помощью ВЭЖХ. Масс-спектральный анализ показал молекулярную массу агликона, равную 652.
Цитотоксичность фракций с препаративной колонки PFP
Цитотоксичность фракций, полученных при фракционировании G/H
Цитотоксичность D1 и G1 при pH 6,5 и 7,5
Пример 5
Структуры D1, G1 и B1
5.1 Структура D1
D1 является главным компонентом стручков Acacia victoriae. Тестирование данного соединения показывает, что оно обладает значительной биологической активностью.
5.1.1 Целая молекула D1
D1 был выделен в виде бесцветного аморфного твердого вещества из частично очищенного экстракта F094, полученного с помощью нескольких разделений препаративной ВЭЖХ, как описано в примерах выше. Его молекулярная масса по данным масс-спектроскопии MALDI составляет 2104 атомных единиц массы, что представляет собой натриевый аддукт истинной молекулярной массы 2081. Масс-спектрометрия FAB высокого разрешения подтвердила данную молекулярную массу и дала молекулярную формулу C98H155NO46. Такая молекула слишком велика для определения структуры посредством одной спектроскопии, в связи с чем была предпринята программа деградации, изображенная на схеме 1, показанной на фиг.36. На фиг. 36 D1 представлен структурой, обозначенной '(1)'.
Протонный и углеродный ЯМР D1 показали наличие тритерпена, двух монотерпенов и приблизительно восьми сахаров (см. в таблице 22 отдельные значения 13C-ЯМР под (1)).
Таблица 22
Значения 13C ЯМР (MeOH-d4) D1(1), G1(14), B1(21), агликона (2) и акациевой кислоты (3). (Числа в скобках, т.е. (1), (14), (21), (2) и (3), относятся к структурам D1, G1, B1, агликона и акациевой кислоты, изображенные на фиг.36, 37, 38 соответственно.)
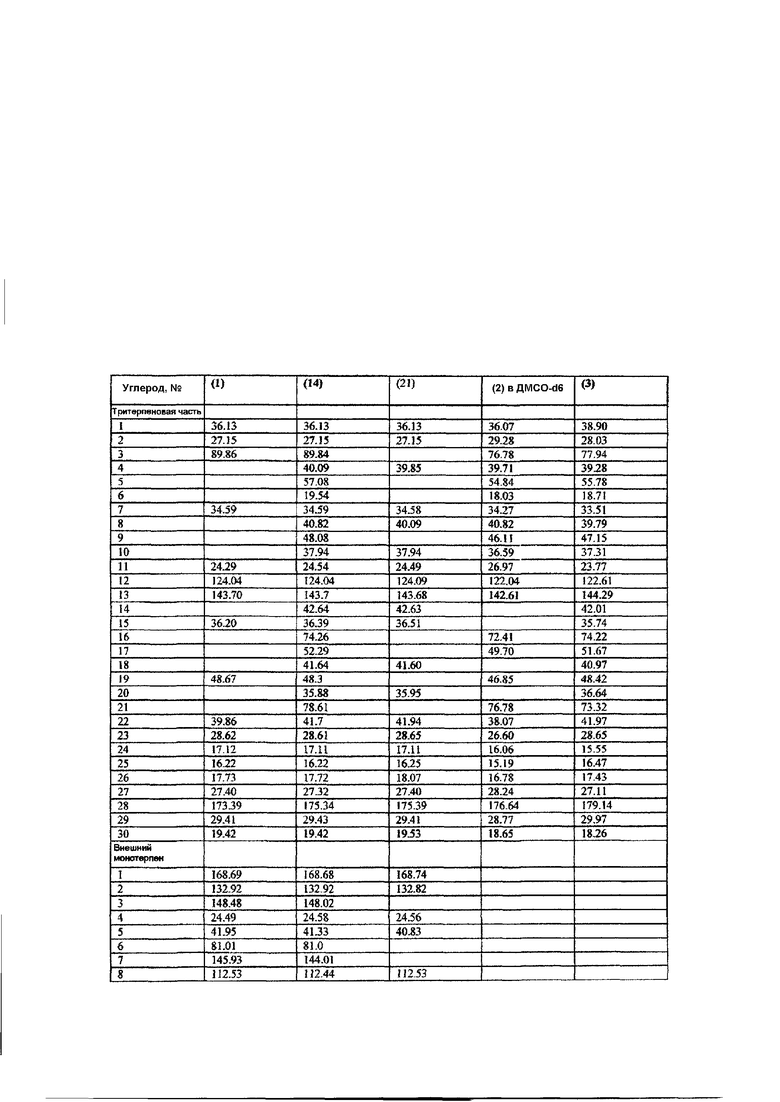

5.1.2 Энергичный кислотный гидролиз D1
Гидролиз D1 в 3 н. HCl при 100°С в течение 2 час дал «агликон D1», обозначенный как (2) на фиг. 36, который, как было показано масс-спектроскопией, имеет молекулярную массу 652. ЯМР агликона D1 показал наличие тритерпена и модифицированного монотерпена, но не сахаров. Данное вещество подвергали дальнейшей деградации путем сапонификации (1,3 н. NaOH при 100°С в течение 30 мин в MeOH), откуда было выделено следующее.
5.1.2.a. Тритерпен. C-13 ЯМР данного вещества был идентичен ранее сообщавшемуся для акациевой кислоты (см. на фиг. 36 структуру, обозначенную как (3), и см. в таблице 22 значения 13C-ЯМР в графе (3)), и его молекулярная масса, составляющая по данным масс-спектроскопии 488, соответствует данной структуре.
5.1.2.b. Циклизированный монотерпен. Молекулярная масса и ЯМР данного соединения указывают на наличие карбоновой кислоты, двух метильных групп, присоединенных к двойной связи, и двух винильных протонов, что ведет к указанной пирановой структуре. Хотя данная структурная единица со структурой (4), показанной на фиг. 36, присутствовала также в «агликоне D1», ее не было в исходном D1. D1 содержит ациклический монотерпен, показанный в виде структуры (5) на фиг. 36, и данная структура подвергается циклизации во время кислотного гидролиза, как показано ниже.
Данные структуры, наряду с исходной молекулярной массой и спектральными характеристиками D1, хорошо укладываются в структуру агликона D1, показанную на фиг. 36 как структура, обозначенная (2). Смотри в таблице 22 отдельные значения 13C-ЯМР в графе (2).
5.1.3. Мягкая сапонификация D1
При обработке D1 0,5 н. NH4OH при комнатной температуре в течение 1 часа происходило полное превращение в два новых соединения.
5.1.3.a. Монотерпен. Данная молекула имела молекулярную массу 200 и спектр ЯМР, которые указывали на то, что она имеет ациклическую монотерпеновую структуру, обеспечивающую ожидаемую деградацию. Данная структура показана на фиг. 36 и обозначена как (5).
5.1.3.b. Тритерпеновый монотерпеновый олигосахарид. Данное соединение является более полярным, чем D1, и его ЯМР согласуется с содержанием в нем акациевой кислоты, одного монотерпена и нескольких моносахаридов. Данная структура показана на фиг. 36 и обозначена как (6).
5.1.4. Анализ сахаров D1
Интенсивный кислотный гидролиз D1 (2 н. HCl при 100°С в течение 2 часов) с последующим получением производных (триметилсилильных эфиров) и анализом GC/MS подтвердил присутствие в исходной молекуле восьми сахарных остатков: арабинозы, рамнозы, фукозы, ксилозы, 6-дезоксиглюкозы (т.е. хиновозы), N-ацетилглюкозамина и двух молекул глюкозы.
5.1.5. Более энергичная сапонификация тритерпенового монотерпенового олигосахарида
В результате экспозиции тритерпенового монотерпенового олигосахарида с 0,3 н. NaOH в течение 1 часа при 60°С образовались три соединения.
5.1.5.a. Олигосахарид. Выделение и анализ данного очень полярного фрагмента позволили предположить, что им является олигосахарид. Анализ сахаров, выполненный путем кислотного гидролиза (2 н. HCl при 100°С в течение 2 часов) и GC/MS анализа триметилсилильных эфиров моносахаридов, подтвердил, что олигосахарид является тетрасахаридом, построенным из двух молекул глюкозы и по одной из молекул арабинозы и рамнозы.
5.1.5.b. Монотерпеновый гликозид. Данное вещество имеет спектр ЯМР, согласующийся со структурой (8), изображенной на фиг. 36. Кислотный гидролиз (2 н. HCl при 100°С в течение 2 часов) данного соединения привел к идентификации сахара как 6-дезоксиглюкозы. Обработка данного монотерпенового гликозида β-глюкозидазой дала монотерпен со структурой, изображенной под обозначением (9) на фиг. 36, которая имеет спектр ЯМР, согласующийся со структурой транс-2-гидроксиметил-6-гидрокси-6-метил-2,7-октидиеновой кислоты. Гидролиз данной связи «бета»-глюкозидазой указывает на то, что связь между двумя данными группами представляет собой бета-связь.
5.1.5.c. Тритерпеновый гликозид. Данное соединение имеет молекулярную массу 951 и спектр ЯМР, которые согласуются со структурой лактона акациевой кислоты, содержащего трисахарид в положении C-3, изображенного как структура (10b) на фиг. 36. Кислотный гидролиз (2 н. HCl при 100°С в течение 2 часов) данного соединения позволил идентифицировать с помощью GC/MS входящие в него сахара как N-ацетилглюкозамин, фукозу и ксилозу в виде триметилсилильных производных. Данная молекула обнаруживалась и в открытой кислотно-спиртовой форме, изображенной на фиг. 36 в виде структуры, обозначенной как (10a), и в закрытой лактонной форме, изображенной на фиг. 36 в виде структуры, обозначенной как (10b).
Анализ сахаров и молекулярная масса фрагментов при сравнении с таковыми целой молекулы D1 подтвердили то, что все части D1 ответственны за фрагменты, изображенные в виде структур, обозначенных как (5), (7), (8) и (10a) на фиг. 36.
5.1.6. Мягкий кислотный гидролиз D1
Мягкий кислотный гидролиз D1 (1 н. HCl в течение 16 час при 25°С) обеспечил образование двух новых молекул.
5.1.6.a. Монотерпеновый сахар. Молекулярная масса, спектры ЯМР и анализ сахаров согласовались с монотерпен-6-дезоксиглюкозой. Структура данной молекулы изображена на фиг. 36 в виде структуры, обозначенной как (11).
5.1.6.b. Тритерпен-монотерпен-гликозид. Вторую молекулу идентифицировали как тритерпен-монотерпен-гликозид, и структура данной молекулы изображена на фиг. 36 в виде структуры, обозначенной как (12).
5.1.7. Соединение подгрупп внутри D1
Исследования ЯМР указывают на то, что карбоновая кислота наружного монотерпена этерифицирована C-4 6-дезоксиглюкозы (хиновозы). Исследования с помощью ЯМР и гидролиза показали, что аномерный углерод хиновозы присоединен к C-6 гидроксильной группе внутреннего монотерпена. Стереохимия аномерного углерода хиновозы указывает на «бета»-связь.
Гидролиз (2 н. HCl в течение 2 час при 100°С) и исследования изомеризации сахаров указывают на то, что сахарами в тетрасахариде являются две молекулы глюкозы и по одной молекуле рамнозы и арабинозы. Единица прямо этерифицирована C-28 карбоновой кислоты тритерпена, как показано на фиг. 39. Масс-спектроскопические исследования с железной ловушкой показывают, что тетрасахаридная структура содержит две молекулы глюкозы и одну молекулу арабинозы, присоединенные к центрально расположенной рамнозе, как показано на фиг. 39. Связь данных сахаров друг с другом пока неизвестна.
Исследования с помощью ЯМР показывают, что N-ацетилглюкозамин (NAG) прямо присоединен к углероду C-3 тритерпена. В перечне сахаров остаются фукоза в середине списка и ксилоза в его конце, выявленные в исследованиях с помощью LC-MS и частичного гидролиза (1 н. HCl в течение 1 час при 60°С в 50% MeOH). Связь данных сахаров друг с другом остается неизвестной.
5.1.8. Эллиптозид E
D1 содержит тритерпен и два монотерпена, обычно обнаруживаемые в описанных сапонинах других видов, включая другие виды Acacia. В то же время структура D1 сходна с сообщавшейся структурой эллиптозида E (фиг. 24) из Archidendron ellipticum (Beutler et al., 1997). В настоящем изобретении было определено удельное вращение D1, равное [α]D = -30,0°, что отлично от сообщавшейся величины -24,3° для эллиптозида E.
Эллиптозид E, описанный Beutler et al. (1997), и D1 имеют разное время удерживания при ВЭЖХ (D1 - 15,2 мин, эллиптозид E - 12,5 мин). Следовательно, данные две молекулы должны различаться каким-то образом, таким как специфичное соединение их субъединиц или присутствие оптических или структурных изомеров.
Заявители наблюдали, что удельное вращение внутреннего монотерпена, изображенного в виде структуры (9) на фиг. 36, составляет +11,2° в MeOH и +16° в хлороформе. Для того же фрагмента в эллиптозиде E сообщалась величина -9,1° в хлороформе. Более того, было определено, что единственный хиральный центр внутреннего монотерпена D1 имеет "S" конфигурацию, что является противоположным обнаруженной в эллиптозиде E. В настоящее время определяется удельное вращение внешнего монотерпена D1. Более того, протонный ЯМР показывает, что монотерпеновые двойные связи в D1 являются «транс», тогда как монотерпеновые двойные связи в эллиптозиде E являются «цис», как показано Beutler et al., 1997. Данные две особенности составляют первые структурные различия, обнаруженные между D1 и эллиптозидом E. Ферментативный каталитический гидролиз конкретных сахаров показал, что расположение сахаров является таким же, как и в эллиптозиде E.
5.2. Структура G1
Биологические тесты данного вещества показывают, что G1 является более активным, чем D1.
5.2.1. Целая молекула G1 (14)
Второй структурой, определенной в настоящем изобретении, был G1. Он был также выделен из F094 с помощью препаративной ВЭЖХ, но с низким выходом соединения. G1 немного менее полярен, чем D1. Молекулярная масса, по результатам масс-спектрометрии MALDI, составляет 2065, что на 16 атомных единиц массы меньше, чем D1. Было обнаружено, что удельное вращение G1 составляет -26,9° (MeOH). Протонный ЯМР показывает, что G1 также является сапонином, очень похожим на D1, и указывает на то, что он отличается от D1 только меньшим на единицу содержанием кислорода в наружном монотерпене, который теперь представляет собой транс-2,6-диметил-6-гидрокси-2,7-октадиеновую кислоту. Смотри структуру, обозначенную (14), на фиг. 37 и отдельные значения 13C-ЯМР в графе (14) таблицы 22. G1 подвергали деградации, как показано на схеме 2, фиг. 37.
5.2.2. Мягкая сапонификация G1
При обработке G1 0,5 н. NH4OH при комнатной температуре даже в течение нескольких минут происходит полное превращение в более полярный продукт мягкой сапонификации и монотерпен.
5.2.2.a. Монотерпен. Молекулярная масса и спектр ЯМР данного вещества указывают на то, что оно содержит метильную группу в положении C-2, где находился гидроксиметил. Это изображено на фиг. 37 в виде структуры, обозначенной (15).
5.2.2.b. Тритерпеновый/монотерпеновый олигосахарид. Спектр ЯМР данного соединения указывает на то, что оно по времени удерживания при ВЭЖХ и по спектру протонного ЯМР было идентично структуре, обозначенной как (16), изображенной на фиг. 37, которая сходна со структурой, обозначенной как (6) на фиг. 36, полученной из D1, которая содержит акациевую кислоту, один монотерпен и восемь моносахаридов, как это наблюдалось в случае D1. Сходство (16) и (6) указывает на сходство стереохимии с наблюдавшейся для внутреннего монотерпена D1.
5.2.3. Анализ сахаров G1
Энергичный кислотный гидролиз G1 (2 н. HCl при 100°С в течение 2 часов) дал те же моносахаридные единицы, которые имеются в D1: арабинозу, рамнозу, фукозу, ксилозу, 6-дезоксиглюкозу, N-ацетилглюкозамин и две молекулы глюкозы.
5.2.4. Кислотный гидролиз G1
Кислотный гидролиз продукта мягкой сапонификации дал возможность выделить три молекулы таким же образом, что и в случае D1. Каждый из них был проанализирован с помощью ЯМР и анализа сахаров (2 н. HCl при 100°С в течение 2 часов). Это изображено на фиг. 37 в виде структуры, обозначенной (16).
5.2.4.a. Олигосахарид, содержащий две молекулы глюкозы и по одной молекуле арабинозы и рамнозы, изображен на фиг. 37 в виде структуры, обозначенной (17).
5.2.4.b. Монотерпеновый гликозид, содержащий ациклический монотерпен (изображенный на фиг. 37 в виде структуры, обозначенной (5)) и 6-дезоксиглюкозу, и полная структура изображена на фиг. 37 в виде структуры, обозначенной (18).
5.2.4.c. Тритерпеновый гликозид, содержащий акациевую кислоту и по одной молекуле каждого из N-ацетилглюкозамина, фукозы и ксилозы. Сахара в данных фрагментах расположены в том же порядке, что и в D1. Данная структура изображена на фиг. 37 в виде структуры, обозначенной (19).
5.2.5. Эллиптозид A
G1 имеет такие же терпеновый состав и сахара, что и эллиптозид A (см. фиг. 24 и Beutler, 1997). Однако эллиптозид A, как было обнаружено, имеет заметно иное время удерживания при ВЭЖХ (G1 - 29,09 мин, а эллиптозид A - 26,04 мин), что указывает на то, что две молекулы должны каким-то образом различаться, таким как специфичное соединение их субъединиц или присутствие оптических или структурных изомеров или того и другого. Сравнение протонных и углеродных ЯМР-спектров G1 и эллиптозида A также показывает различия в химических сдвигах. Это позволяет предполагать, что удельные вращения внутреннего и наружного монотерпенов данных соединений также могут различаться. На фиг. 37 структура (14) представляет структуру G1.
5.3. Структура B1
Данные о биологической активности показывают, что B1 является значительно менее активным, чем D1 или G1.
5.3.1. Целая молекула B1(21)
Выделение B1 было осуществлено с помощью экстракции растения и моментальной хроматографией на C-18 с последующей препаративной и полупрепаративной хроматографией на C-18. Спектр ЯМР B1 указывает на ту же тритерпеновую/монотерпеновую/хиновоза/монотерпеновую структуру, которая наблюдалась в данном семействе сапонинов. ЯМР также указывает на четыре дезоксисахара и одну N-ацетильную группу, что является указанием на то, что данная молекула должна отличаться от D1 по своим сахарным частям. Конкретные значения 13C-ЯМР смотри в графе (21) таблицы 22. Данную молекулу подвергали деградации, как показано на фиг. 38.
5.3.2. Анализ сахаров B1
Данные ЯМР указывают на наличие более одной копии одного из 6-дезоксиметилсахаров (т.е. фукозы, рамнозы, 6-дезоксиглюкозы). Анализ сахаров полной молекулы после гидролиза (2 н. HCl при 100°С в течение 2 часов) показал наличие девяти сахаров: по одной молекуле фукозы, арабинозы, ксилозы, хиновозы и глюкозамина и по две молекуле глюкозы и рамнозы. В исходной молекуле находится глюкозамин, являющийся ремнантом N-ацетилглюкозамина. Структура B1 изображена на фиг. 38 в виде структуры (21).
5.3.3. Мягкая сапонификация B1
При обработке B1 0,5 н. NH4OH при комнатной температуре даже в течение нескольких минут происходит полное превращение в более полярное соединение, являющееся продуктом мягкой сапонификации, и монотерпен.
5.3.3.a. Монотерпен. Молекулярная масса и спектр ЯМР данного вещества указывают на то, что оно имеет ту же структуру, что и монотерпен из D1, изображенный на фиг. 37 в виде структуре, обозначенной (22).
5.3.3.b. Тритерпеновый монотерпеновый олигосахарид. Спектр ЯМР данного соединения указывает на то, что оно содержит акациевую кислоту, один монотерпен и несколько моносахаридов. Это изображено на фиг. 38 в виде структуры, обозначенной (28).
5.3.4. Более жесткая сапонификация тритерпенового монотерпенового олигосахарида
Более жесткая сапонификация (0,3 н. NaOH при 60°С в течение 1 часа) продукта мягкой сапонификации позволила выделить три молекулы сходным с описанным выше для D1 и G1 образом. Для каждой из них были получены данные анализа сахаров и ЯМР.
5.3.4.a. Олигосахарид, содержащий глюкозу, арабинозу и две молекулы рамнозы. Он изображен на фиг. 38 в виде структуры, обозначенной (24).
5.3.4.b. Монотерпеновый гликозид, содержащий 6-дезоксиглюкозу и монотерпен. Он изображен на фиг. 38 в виде структуры, обозначенной (25).
5.3.4.c. Тритерпеновый гликозид, содержащий акациевую кислоту с тетрасахаридом, присоединенным в положении C-3. Тетрасахарид построен из молекул N-ацетилглюкозамина, фукозы, глюкозы и ксилозы, каждая из которых представлена один раз. Он изображен на фиг. 38 в виде структуры, обозначенной (26).
Пример 6
Деэтерификация тритерпеновых соединений
F094 подвергали деэтерификации и деэтерифицированные продукты подвергали биотестированию для выявления активных компонентов. 1,00 г F094 растворяли в 100 мл H2O, затем добавляли 1 г KOH и нагревали с обратным холодильником в течение 1,5 час. Раствору давали остыть до комнатной температуры, и его pH доводили до 7 с помощью 1 н. HCl, после чего производили промывку гексанами (2 x 50 мл). После этого водный раствор подвергали дополнительным ступенчатым экстракциям с получением фракций 159-162. Например, раствор сначала экстрагировали н-бутанолом (2 x 50 мл) с получением 0,127 г растворимого в органических растворителях твердого вещества (F159) после сушки в вакууме. Водный слой подкисляли до pH 5 с помощью 1 н. HCl и экстрагировали EtOAc (2 x 50 мл) с получением 0,397 г растворимого в EtOAc твердого вещества (F160) и затем н-бутанолом (2 x 50 л) с получением 0,338 г твердого вещества (F161) после удаления органических растворителей. Наконец, водный слой нейтрализовали до pH 7 с помощью 1 н. NaOH. Из конечного водного слоя было выделено 1,808 г твердого вещества (F162).
Биотесты показали, что деэтерифицированные продукты обладают низкой активностью или не обладают ей вовсе. F159-162 подвергали биотестам на цитотоксичность против клеточных линий 769-P, Panc-1, HEY, MDA-MB-453 и Jurkat. Активность была обнаружена лишь для F161, которая проявляла цитотоксичность, равную 1,6%, против MDA-MB-453 при 50 мкг/мл, и для F159, которая проявляла цитотоксичность, равную 15,50%, 6,60% и 3,80%, против клеток Jurkat при 50 мкг/мл, 25 мкг/мл и 12,5 мкг/мл, соответственно. Данные результаты указывают на то, что для биологической активности необходима сложноэфирная боковая цепь. Полагают, что сложноэфирная боковая цепь соединений согласно изобретению проявляет противоопухолевую активность и/или действует совместно с тритерпеновым скелетом соединений согласно изобретению для получения проявляемой высокой противоопухолевой активности.
Пример 7
Гидролиз сахаров соединений
Сахара, содержащиеся в F094, также гидролизовали с целью характеристики активных компонентов. 12 г F094 растворяли в 400 мл 2 н. H2SO4 и нагревали с обратным холодильником в течение 75 мин, за это время образовывалось нерастворимое вещество. Раствор охлаждали и фильтровали через пористое стекло. Остаток промывали водой, что дало 4,8 г агликона(ов) (F191), как это было определено с помощью анализа ТСХ. Темно-янтарный фильтрат нейтрализовали KOH или NaOH. Образовавшийся белый осадок собирали. Добавление изопропанола к янтарному фильтрату дало второй белый осадок. Растворитель удаляли в вакууме и растворитель ресуспендировали в MeOH, что привело к образованию белого осадка, который, возможно, соответствовал сульфатным солям, как это было определено по реакции в пламени. Почти прозрачный фильтрат сушили в вакууме и остаток ацетилировали и анализировали с помощью ВЭЖХ на содержание сахаров, как показано на фиг. 17A и 17B. Данная смесь, возможно, содержит по меньшей мере 5 сахаридов. Данные сахара могут быть дополнительно охарактеризованы путем выделения TMS производных для характеристики с помощью GC-MS; бумажной хроматографии; выделения бензильных производных для разделения с помощью ВЭЖХ или DEPT ЯМР или C13 ЯМР, как это полностью описано здесь выше. С помощью стандартной 1-D и 2-D целлюлозной, бумажной и ТСХ с нормальной фазой главные сахара были идентифицированы как глюкоза, ксилоза, рамноза и арабиноза, наряду с неким минорным дополнительным сахаром и значительным потенциальным аминогликозидом, особенно ацетамидозамещенным сахаром.
Ряд различных сахаров, возможно, объясняет усложненные спектры ВЭЖХ, которые показывают наличие множества близкородственных соединений. В частности, некоторые активные составляющие, по-видимому, гликозилированы по двум разным сайтам (спиртовому и карбоксикислотному сайту). Сочетания шести сахаров, присоединенных к двум сайтам, таким образом, должны были бы давать большое число близкородственных соединений, которые было бы трудно разделить.
Альтернативно, условия более мягкого гидролиза, проводимого в 100% этаноле (азеотропе с 5% воды) и от 0,1 до 2 н. H2SO4 (при сохранении неизменной остальной части работы) при медленном нагревании до точки кипения, но не при интенсивном кипячении с обратным холодильником, ведут к образованию сходной смеси агликонов. Некоторые компоненты отсутствуют, что указывает на то, что при более жестких условиях имеет место некая изомеризация.
После этого агликон(ы) F191 (1 г) метилировали путем нагревания с обратным холодильником в течение 5-7 часов с метилиодидом (1 мл) и безводным K2CO3 (1 г) в безводном ацетоне (10 мл). Это дало 0,315 г нерастворимого вещества и 0,54 г метиловых эфиров, обозначенных как F197. 500 мг F197 дополнительно разделяли с помощью MPLC с применением колонки 15 x 460 мм (45 г SiO2, 15-25 мкм), где образец предварительно адсорбировали на 1,5 г SiO2, 15-25 мкм. Соединения элюировали 790 мл 7% изопропилового спирта (IPA) в гексанах (подфракции 1-10), 470 мл 10% IPA в гексанах (подфракции 1-14), 275 мл 20% IPA/гексаны (подфракции 14-15), 200 мл дихлорметана и 100 мл смеси ДХМ/MeOH (1:1) в соответствии с таблицей 23.
Биотесты F191 и F197 показали цитотоксичность в отношении раковых клеток яичника (линия HEY), раковых клеток почек (линия 769-P), раковых клеток поджелудочной железы (линия Panc-1), T-лейкозных клеток Jurkat и раковых клеток молочной железы MDA-MB-453 при соответствующих дозах, как указано в таблице 23.
Фракционирование F197 на фракции с F198 по F208
Таблица 24
Биотестирование фракций F191 и F197


F199 (116 мг) дополнительно фракционировали с помощью той же колонки, которую применяли для фракционирования F191, и элюировали 100 мл 2% IPA/гексаны, 525 мл 4% IPA/гексаны и 250 мл 10% IPA/гексаны согласно таблице 25.
Фракционирование F199 на фракции с F209 по F214
F212 (85 мг) дополнительно фракционировали с помощью ВЭЖХ на колонке 22 x 500 мм Waters Prep LC 4000 (Alltech Econosil C18, 10 мкм, уравновешенной 75% ACN/вода) с элюированием 80% ACN/вода и промывкой ACN со скоростью 40 мл/мин при ограничении детекции 220 нм и сбором подфракций каждые 30 сек (20 мл) согласно таблице 26.
F223 сначала очищали в виде метилэфирного производного с получением C-191. C-191 подвергали классической процедуре ацетилирования. Конкретно, C-191 (47 мг) перемешивали в течение ночи при комнатной температуре в смеси 2:1 уксусного ангидрида и пиридина. Реакцию гасили водой и раствор подвергали распределению с диэтиловым эфиром и 5 н. HCl. После этого органический слой промывали до нейтральной реакции, упаривали на роторном испарителе и остаток подвергали PTLC - 20 см на 20 см препаративная пластина, элюируемая смесью 90:10 гексан : изопропиловый спирт с последующим элюированием PTLC смесью дихлорметан : метанол (98:2) для получения ацетата C-191 (F229, которой позднее был присвоен номер C-194).
Фракционирование F212 на фракции с F220 по F228
F201 (85 мг) также дополнительно фракционировали с помощью MPLC с применением колонки, сходной с использованной для фракционирования F199, с элюированием и сбором в 2% IPA/гексаны (120 мл), 4% IPA/гексаны (330 мл), 7% IPA/гексаны (460 мл), 20% IPA/гексаны (150 мл), ДХМ (50 мл) и смеси ДХМ/MeOH (1:1) (70 мл) согласно таблице 27.
Фракционирование F201 на фракции с F215 по F219
Пример 8
Биологические свойства активных тритерпенов
Ангиогенез или неоваскуляризация представляет собой процесс, посредством которого клетки рекрутируются фактором(ами), продуцируемым(и) опухолью для обеспечения опухоли питающей сосудистой системой. Ингибирование ангиогенеза тормозит экспансию опухоли за счет ограничения кровоснабжения опухоли. Данная функция может быть проверена с применением теста пролиферации клеток эндотелия капилляров быка на клетках, обработанных фракцией 35 (UA-BRF-004-DELEP-F035). Тест проводили следующим образом: получали клетки эндотелия капилляров быка и выращивали их с применением стандартных процедур (Folkman et al., 1979). Клетки промывали ЗФР и диспергировали в 0,05% растворе трипсина. Готовили клеточную суспензию (25000 клеток/мл) с DMEM + 10% BCS + 1% GPS, наносили ее на 24-луночные планшеты для культивирования (0,5 мл/лунка) и суспензию инкубировали в течение 24 час при 37°С. Среду заменяли 0,25 мл DMEM + 5% BCS + 1% GPS и наносили разные концентрации UA-BRF-004-DELEP-F035. Через 20 мин инкубации добавляли среду и bFGF с получением конечного объема 0,5 мл DMEM + 5% BCS + 1% GPS + 1 нг/мл bFGF. Через 72 час клетки диспергировали в трипсине, ресуспендировали в Hematall (Fisher Scientific, Pittsburg, PA) и подсчитывали с помощью режущего счетчика (O'Reilly et al., 1997).
Результаты тестирования показали значительное ингибирование пролиферации эндотелиальных клеток в присутствии или в отсутствие основного фактора роста фибробластов (фиг.5). Данные результаты показывают, что активные компоненты растительного экстракта являются сильными ингибиторами пролиферации эндотелиальных клеток, что часто служит фактором, предсказывающим подавление ангиогенеза iv vivo. Кроме того, фракция не оказывала влияния на подвижность клеток эндотелия капилляров, что позволяет предполагать отсутствие токсичности в отношении нормальных клеток (фиг. 6).
Общей проблемой, встречающейся при работе со стероидными сапонинами (т.е. дигитонином и генин-диосгенином из ямса), является лизис красных кровяных клеток. В тесте простой культуральной пробирки с кровью имелся очень слабо определяемый лизис после обработки 1 мг/мл UA-BRF-004-DELEP-F035. Альтернативно, обработка от 10 до 25 мкг/мл дигитонина вызывали 100% лизис в тесте культуральной пробирки с кровью.
Далее, для дополнительного изучения механизмов, посредством которых активные компоненты ингибируют опухолевые клетки, анализировали индуцированную TNF-альфа активацию транскрипционного фактора NF-κB в клетках Jurkat (3 x 106), которые обрабатывали 1-2 мкг/мл UA-BRF-004-DELEP-F035 и UA-BRF-004Pod-DELEP-F094. Исследование проводили следующим образом: клетки Jurkat предварительно обрабатывали 1-2 мкг/мл F035 или F094 в течение 15 час при 37°С. Клетки собирали, ресуспендировали в 1 мл RPMI и обрабатывали 100 пМ TNF-альфа в течение 30 мин при 37°С. После обработки TNF-альфа получали ядерные экстракты по Schreiber et al. (1989). Вкратце, клетки промывали охлажденным до температуры льда ЗФР и суспендировали в 0,4 мл буфера для лизиса (10 мМ HEPES, pH 7,9, 10 мМ KCl, 0,1 мМ ЭДТА, 1 мМ ЭДТА, 0,1 мМ ЭГТА, 1 мМ дитиотрейтол, 0,5 мМ PMSF, 2 мкг/мл лейпептин, 2 мкг/мл апротинин и 0,5 мг/мл бензамидин). Клеткам давали осесть на льду в течение 15 мин и к клеткам добавляли 25 мкл 10% Nonidet-40. Пробирки перемешивали на вортексе и центрифугировали на микроцентрифуге в течение 30 сек. Ядерный осадок ресуспендировали в 25 мкл охлажденного до температуры льда буфера для ядерной экстракции (20 мМ HEPES, pH 7,9, 0,4 М NaCl, 1 мМ ЭДТА, 1 мМ ЭГТА, 1 мМ дитиотрейтол, 1 мМ PMSF, 2 мкг/мл лейпептин, 2,0 мкг/мл апротинин и 0,5 мг/мл бензамидин) и пробирки инкубировали на льду при периодическом перемешивании. Ядерный экстракт центрифугировали на микроцентрифуге 5-кратно при 4°С и супернатанты хранили при -70°С.
Анализ сдвига электрофоретической подвижности проводили, инкубируя ядерные экстракты (7 мкг белка) с 32P-мечеными олигонуклеотидами NF-κB (SEQ ID NO:1; консенсусный олигонуклеотид NF-κB; Santa Cruz Biotechnology) в присутствии 0,5 мкг поли di-dc в буфере для связывания (25 мм HEPES, pH 7,9, 0,5 мМ ЭДТА, 0,5 мМ дитиотрейтол, 1% Nonidet P-40, 5% глицерин и 50 мМ NaCl в течение 20 мин при 37°С) (Nabel and Baltimore, 1987; Collart et al., 1990; Hassanain et al., 1993). Образовавшийся комплекс ДНК-белок отделяли от свободного олигонуклеотида на 5% неденатурирующем полиакриламидном геле с применением буфера, содержащего 50 мМ трис, 200 мМ глицин и 1 мМ ЭДТА. Гель фиксировали в 10% уксусной кислоте и сушили, полосы визуализировали с помощью авторадиографии с усиливающим экраном при -70°С.
Результаты EMSA показывают, что в необработанных клетках имеется низкий базальный уровень NF-κB, который активируется TNF (фиг. 20, дорожки 1 и 2). Предварительная обработка клеток 1 мкг/мл F035 или F094 с последующей активацией TNF (фиг. 20, дорожки 4 и 8) приводила к ингибированию активации NF-κB. Когда клетки обрабатывали 2 мкг/мл UA-BRF-004-DELEP-F035 или UA-BRF-004Pod-DELEP-F094 (фиг. 20, дорожки 6 и 10), наблюдалось значительное ингибирование NF-κB, активированного TNF. Кроме указания на активные тритерпеновые соединения как потенциально противовоспалительные соединения, результаты особенно важны с учетом возрастающих доказательств того, что воспаление играет центральную роль в канцерогенезе (Sieweke et al., 1990; Prehn, 1997; Schuh et al., 1990).
Пример 9
Исследования пути проведения сигнала F035
Для дальнейшего выяснения молекулярных мишеней активных компонентов растительного экстракта Acacia victoriae было проведено исследование влияния F035 на активность фосфатидилинозитол-3-киназы (PI3-киназы), а также на активность AKT (протеинкиназы B, серин-треониновой киназы), являющейся нижележащим эффектором PI3-киназы. PI3-киназа является ферментом, участвующим в проведении сигнала ростовых факторов за счет ассоциации с рецепторными и нерецепторными тирозинкиназами. Существуют два известных ингибитора PI3-киназы: метаболит грибов вортманнин и LY294002, синтетическое соединение, которое структурно сходно с растительным биофлавоноидом кверцитином.
Анализ проводили следующим образом: клетки Jurkat (1 x 107) оставляли на ночь без питательной среды и экспонировали с различными концентрациями (1-8 мкг/мл в зависимости от линии клеток) F035 в течение различных промежутков времени (2-16 час) при 37°С. Через различные промежутки времени клетки собирали и промывали ЗФР при 2000 об/мин в течение 10 мин. Клетки лизировали в 1% NP-40 буфере для лизиса в течение 30 мин при 4°С и выделяли лизаты центрифугированием в течение 5 мин при 15000 об/мин при 4°С. Для проведения иммунопреципитации PI3-киназы инкубировали 5 мкл антитела кролика против p85 (адапторного белка тирозинкиназного рецептора; Upstate Biotechnology Inc.) с 1 мл клеточного лизата в течение 90 мин при 4°С. Иммунные комплексы выделяли на 100 мкл 20% шариков протеин A-сефарозы еще в течение 90 мин при 4°С. Иммунопреципитаты промывали последовательно в a) ЗФР, 100 мМ Na3VO4, 1% тритон -X100; b) 100 мМ трис, pH 7,6, 0,5 LiCl, 100 мМ Na3VO4; c) 100 мМ трис, pH 7,6, 100 мМ NaCl, 1 мМ ЭДТА, 100 мМ Na3VO4 и d) 20 мМ HEPES pH 7,5, 50мМ NaCl, 5 мМ ЭДТА, 30 мМ NaPPi, 20 мМ Na3VO4, 1 мМ PMSF, 0,03% тритон X-100. Затем иммунопреципитаты ресуспендировали в 30 мкл буфера для киназной реакции (33 мМ трис, pH 7,6, 125 мМ NaCl, 15 мМ MgCl2, 200 мМ аденозин, 20 мМ АТФ, 30 мкКи [g-32P] АТФ). Фосфатидилинозитол (PI; 50 мкл) сушили в атмосфере газообразного азота и ресуспендировали в 20 мМ HEPES, pH 7,5, в концентрации 2 мг/мл и сонифицировали на льду в течение 10 мин. PI3-киназную реакцию инициировали добавлением 10 мкл суспензии PI и 10 мкл гамма-АТФ. Реакции давали пройти в течение 30 мин при комнатной температуре с последующим прекращением реакции добавлением 100 мкл 1 н. HCl. Липиды экстрагировали 600 мкл смеси хлороформ : метанол (1:1) и разделяли на силикагелях (G60) с помощью тонкослойной хроматографии (ТСХ) в системе хлороформ : метанол : NH4OH : H2O (60:47:2:11,3). Радиоактивно меченый фосфатидилинозитолфосфат визуализировали с помощью ауторадиографии и ингибирование количественно определяли с помощью системы шторм (Okada et al., 1994; Vlahos et al., 1994). Результаты (фиг. 21) указывают на то, что через 2 и 6 часов после обработки F035 (4 мкг/мл) происходило ингибирование активности PI3-киназы. Сходным образом, когда клетки экспонировали с 2 мкг/мл F035 в течение 15 час, наблюдалось 95% ингибирование, сходное с действием вортманнина (метаболита грибов и известного ингибитора PI3-киназы) в клетках Jurkat.
Далее, исследовали влияние F035 на AKT, нижележащий эффектор PI3-киназы. AKT, известный также как протеинкиназа B, является клеточным гомологом вирусного онкогена v-AKT, активируется рядом ростовых факторов и действует на пути, включающем активацию PI3-K, которая чувствительна к вортманнину. AKT кодирует серин-треониновую протеинкиназу, которая, как было показано, повышена в 12,1% карцином яичников и 2,8% раков молочной железы. AKT участвует в антиапоптотическом пути посредством фосфорилирования Bad, антиапоптотической молекулы. Больные раком яичников с нарушениями AKT, по-видимому, имеют плохой прогноз (Bellacosa et al., 1995). Было показано, что AKT активно блокирует апоптоз, отчасти за счет активации p70S6 киназы (Kennedy et al., 1997). p70S6 киназа является активируемой митогенами серин-треониновой протеинкиназой, необходимой для клеточного роста и развития G1 клеточного цикла (Chou and Blenis, 1996). Активность p70S6 киназы контролируется множественными событиями фосфорилирования, приуроченного к каталитической и псевдосубстратной области (Cheatham et al., 1995; Weng et al., 1995).
Влияние F035 на фосфорилирование AKT анализировали следующим образом. Клетки Jurkat (5 x 106) лишали сыворотки и экспонировали с F035 в течение 15 час и с вортманнином в течение 2 час при 37°С. Клетки либо индуцировали cd3XL (поперечно-сшитым cd3), либо оставляли без индукции в течение 10 мин при 37°С и лизировали в AKT буфере для лизиса, белки разделяли в 8% ДДС-Na-ПАГЭ гелях и анализировали с помощью иммуноблоттинга-ECL с применением антител, специфичных в отношении фосфорилированного AKT (Ser 437; New England Biolabs) и в отношении AKT в целом. Тестирование влияния F035 на p70S6 киназу можно проводить сходным образом, но с применением набора антител Phosphoplus p70S6 kinase (New England Biolabs) для анализа фосфорилирования p70S6 киназы (Ser 411, thr421/ser424). Результаты анализа AKT (фиг. 22) показали, что поперечно-сшитый cd3 слегка индуцирует фосфо-AKT. Последующая обработка клеток 1 и 2 мкг/мл F035 вызывала заметное ингибирование фосфорилирования AKT (активного AKT), которое сходно с наблюдаемым при 2-часовой обработке клеток 1 мкМ вортманнином. Не было, однако, изменения в экспрессии суммарного AKT. Сходное ингибирование фосфорилирования AKT также было показано с применением раковых клеток яичника OVCAR-3 и C-2 (варианта HEY) и клеток Jurkat, обработанных 2-4 мкг/мл F094. Данные наблюдения показывают, что F035 ингибирует фосфорилирование AKT клеток Jurkat и раковых клеток яичников. Это является важным, учитывая, что было показано, что путь сигнализации PI3-киназа/AKT доставляет антиапоптотический сигнал (Kennedy et al., 1997). Результаты позволяют предполагать, что F035 и F094 опосредуют апоптоз опухолевых клеток через подавление пути сигнализации PI3-K.
Пример 10
Анализ клеточного цикла и тестирование связывания аннексина-V для детекции апоптоза
Для дальнейшей характеристики механизма ингибирования роста и цитотоксичности активных соединений растительного экстракта приблизительно 1 x 106 опухолевых клеток OVCAR-3 помещали в чашки 60 мм3, обрабатывали различными концентрациями UA-BRF-004-DELEP-F035 и инкубировали в течение 18-24 часов при 37°С. Клетки собирали, промывали дважды ЗФР и ресуспендировали при концентрации 1 x 106 клеток/мл. По каплям к слегка перемешиваемым клеткам добавляли параформальдегид (конечная концентрация 1%). После 15-минутной инкубации на льду клетки вновь промывали ЗФР, осадок ресуспендировали в охлажденном до температуры льда 70% этаноле и инкубировали при -20°С в течение ночи. Наконец, этанол отмывали один раз ЗФР и клетки ресуспендировали в растворе 10 мкг/мл иодида пропидия (Sigma Chemical Co.) с 0,1% РНКазы. Клетки вновь инкубировали при комнатной температуре в течение 30 минут, затем переносили в 4°С и анализировали через 18 часов с помощью проточной цитометрии (Pallavicini, 1987). Результаты показывают, что перед обработкой клеток UA-BRF-004-DELEP-F035 48% клеток находились в фазе G0/G1, 36% клеток находились в S-фазе и 7% клеток находились в G2/M-фазе. Однако через 48 час после обработки клеток OVCAR-3 F035 ˜ 58% клеток находились в фазе G1 и только 27% - в S-фазе клеточного цикла, что указывает на 8% увеличение клеток в G1 и ˜ 10% снижение клеток в S-фазе клеточного цикла (фиг. 19A, B). Результаты определенно свидетельствовали об остановке в G1-фазе клеток карциномы яичника человека OVCAR-3.
Проверяли также влияние F035 на профиль клеточного цикла раковых клеток молочной железы человека. Раковые клетки молочной железы MDA-MB-435 и MDA-MB-453 экспонировали с различными концентрациями F035 и анализировали через 72 час с помощью анализа клеточного цикла, как описано выше. Результаты показывают, что F035 индуцирует апоптоз клеток MDA-MB-435 по появлению пика Sub G0 (таблица 28). Кроме того, регуляция клеточного цикла наблюдается также по снижению процента клеток в S- и G2/M-фазах клеточного цикла.
Анализ клеточного цикла раковых клеток молочной железы MDA-MB-435 после обработки F035
Результаты, полученные с применением MDA-MB-453, показывают, что F035 вызывает остановку G1 клеточного цикла с ˜ 10% увеличением количества клеток в G1-фазе и 4-10% снижением количества клеток в S-фазе клеточного цикла спустя семьдесят два часа после обработки F035 (таблица 29). Данные результаты показывают остановку клеточного цикла и апоптоз опухолевых клеток, индуцированные растительным экстрактом.
Анализ клеточного цикла раковых клеток молочной железы MDA-MB-453
Клетки Jurkat (1 x 106) обрабатывали различными концентрациями UA-BRF-004-DELEP-F035 (50-1000 нг/мл) в течение 18 часов при 37°С. Клетки промывали один раз ЗФР, ресуспендировали в буфере для связывания (10 мМ HEPES/NaOH, 140 мМ NaCl, 2 мМ CaCl2), содержащем 5 мкл конъюгата аннексин-V-ФИТЦ (Biowhittaker, Walkersville, MD) и инкубировали в течение 10 минут в темноте. Клетки промывали и ресуспендировали в буфере для связывания, содержащем 10 мкл 20 мкг/мл пропидийиодида (Sigma Chemical Co.), и анализировали с помощью активируемого флуоресценцией аппарата для сортировки клеток (FACS) (Martin et al., 1995).
Результаты показывают, что очищенные активные соединения были способны вызывать апоптоз клеток Jurkat. Данное наблюдение было дополнительно подтверждено способностью обработанных клеток связывать аннексин-V, что служит указанием на то, что клетки подвергались апоптозу (таблица 30). Обычно фосфатидилсерин (PS) локализован на внутренней мембране плазматической мембраны. Однако на ранних стадиях апоптоза имеет место экстернализация PS. Аннексин-V представляет собой кальций-связывающий белок, который связывается с PS и может выявляться с помощью окрашивания аннексин-V-ФИТЦ при проточной цитометрии (Martin et al., 1995).
Апоптоз, измеренный с помощью связывания аннексина-V, в клетках Jurkat, обработанных различными концентрациями UA-BRF-004-DELEP-F035
Пример 11
UA-BRF-004-DELEP-F035 в качестве хемопротекторного агента
Действие UA-BRF-004-DELEP-F035 на модели многостадийного канцерогенеза кожи у мышей SENCAR. Животных обрабатывали нанесением на кожу ацетона, канцерогена DMBA (7,12-диметилбенз[a]антрацена), DMBA + UA-BRF-004-DELEP-F035 и DMBA + фракция 60 (негативный контроль) при низкой (100 мкг UA-BRF-004-DELEP-F035 или фракции 60 на мышь) и высокой (500 мкг UA-BRF-004-DELEP-F035 или фракции 60 на мышь) дозах растительного экстракта, вводимого дважды в неделю в течение 4 недель. UA-BRF-004-DELEP-F035 или контроль наносили на кожу за 5 минут до нанесения DMBA. Животных наблюдали на предмет образования папиллом, впоследствии их забивали и ткани анализировали гистологически (фиг. 9A-9F). Результаты анализа суммированы на фиг. 10A, 10B, 11A, 11B, 12 и 13.
Через 8 недель хода данных экспериментов в группе мышей, обработанных DMBA, имелось 8 папиллом на мышь, а в группе, обработанной DMBA и UA-BRF-004-DELEP-F035, было 0,66 папиллом на мышь, тогда как у мышей, обработанных DMBA и фракцией 60 (негативный контроль) было 6,9 папиллом на мышь. Данные результаты указывают на значительное защитное действие UA-BRF-004-DELEP-F035 против опухолей, тогда как фракция 60 практически не оказывала защитного действия.
Последующие исследования на мышах in vivo показали, что UA-BRF-004-DELEP-F035 является хемопревентивным против индуцируемых канцерогенами опухолей за счет предотвращения мутации онкогена ras. Стадия инициации канцерогенеза в коже мыши осуществляется прямо действующими канцерогенами (т.е. DMBA) и по существу является необратимой стадией. Ингибирование канцерогенеза было определено через 8 недель по снижению индуцированного DMBA образования папиллом. Молекулярный анализ обработанной кожи показал, что UA-BRF-004-DELEP-F035 предотвращает способность DMBA вызывать мутацию онкогена ras (см. примеры 14, 15 и 16 ниже).
Пример 12
Процедура определения активных тритерпеноидов в Acacia victoriae
Была применена процедура, которая обеспечивает эффективную детекцию активных тритерпенов в образце ткани растения. Процедуру осуществляют следующим образом. Приблизительно 5 г листьев и веточек разрезали на мелкие кусочки ножницами или, альтернативно, образцы корня разрезали ножом для получения мелких ломтиков. Растительный материал обрабатывали в небольшом миксере, объединяли с приблизительно 25 мл 80% метанола (об/об) и давали отстояться по меньшей мере в течение 2 часов при встряхивании каждый 1 час. Нерастворимое вещество удаляли центрифугированием при 10000 g. Экстракт затем использовали для тонкослойной хроматографии с пластинами RP (алюминиевые пластины ТСХ, RP-C18 F254S) и 40% ацетонитрилом (об/об). После экспозиции пластин ТСХ со спреем 0,1% ванилина (4-гидрокси-3-метоксибензальдегидом)/H2SO4 и обжига при 70°С в течение от 15 до 30 минут активные тритерпеноидные соединения становились видимыми в виде коричневато-красных пятен (Rf = 0,2-0,3).
Пример 13
Локализация тритерпеновых соединений в растениях Acacia victoriae
В первоначальных исследованиях для экстракции собирали надземные части растений ранним летом. Последующий повторный сбор осенью не обладал активностью. Затем было проведено систематическое исследование для определения относительного отсутствия или присутствия активных тритерпеновых соединений в различных частях растений Acacia victoriae. В результате мониторинга химии растения было определено, что почти все активные компоненты надземной части растения сконцентрированы в стручках, корнях и ростках, и в значительной мере или полностью отсутствуют в ветвях, коре, листьях и семенах. Следовательно, период активного сбора длится лишь приблизительно три недели от начала образования стручков до их раскрытия. Было также определено, что корни растения продуцируют некое активное вещество с изменяющимися соотношениями сахаров и активных компонентов. Последующая характеристика показывает, что аэропоника, которая обеспечивает энергичный рост корней при сохранении нормального развития растения, может быть хорошо применимой для Acacia victoriae.
Пример 14
Линии опухолевых клеток и их рост
Следующие линии клеток рака человека были получены из American Type Tissue Culture Collection (ATCC, Rockville, MD): SK-OV-3 и OVCAR-3 (яичников), Jurkat (T-клеточного лейкоза), U-937 (гистиоцитарной лимфомы), MDA-MB-468, MDA-MB-453, MDA-MB-435, SK-BP-3, MCF-7, MDA-MB-231, BT-20 (молочной железы), LNCaP, PC-3, DU145 (простаты), 769-P, 786-O, A498 (почек) и PANC-1 (поджелудочной железы). Линии клеток HEY и Dov-13 (яичников) были получены из M. D. Anderson Cancer Center. Следующие линии нетрансформированных клеток человека MCF-10A и 10F (эпителия молочной железы) были получены из M. D. Anderson Cancer Center. Hs 27 (фибробласты крайней плоти человека) и L929 (фибробласты мыши) были получены из ATCC. SK-OV-3, MDA-MB-468, Hs 27 L929 выращивали на минимальной эссенциальной среде. OVCAR-3, Jurkat, U-937, LNCaP, DU-145, PC-3, HEY, Dov-13, PANG-1, MCF-10A, MCF-10F и остальные линии клеток рака молочной железы выращивали в RPMI 1640, а среду F-12 применяли для выращивания 769-P, 786-O и A498. Ко всем примененным средам добавляли 10% сыворотку плодов телят, 200 мМ глутамина и 0,05% гентамицина.
Пример 15
Амплификация Ha-ras с мутацией кодона 61 CAA→CTA с применением специфичных для мутации праймеров (MSP)
Данный протокол ведет свое начало от Nelson et al. (1992). Обратный праймер, обозначенный 3MSP61mut, был создан так, чтобы он обеспечивал образование пары 3'-концевого нуклеотидного (A) основания со средним нуклеотидом (подчеркнутым) трансверсии CAA→CTA в кодоне 61 Ha-ras и избирательную амплификацию ДНК при описанных ниже условиях. Тест основан на том факте, что Taq полимераза лишена 3'-эндонуклеазной активности и в результате не способна репарировать несоответствие на 3'-конце отожженного праймера. Условия теста зависят от неспособности обратного праймера эффективно отжигаться с последовательностью дикого типа, в результате чего удлинение не происходит. При применении такого же прямого праймера проводят одну реакцию с обратным праймером с несовпадением (3MSP61mut), а другую реакцию - с обратным праймером дикого типа (3MP61wt). Данный протокол определяет только трансверсии CAA→CTA; однако данные мутации являются преобладающими среди точечных мутаций кодона 61. При данной трансверсии создается сайт Xba I RFLP (T/CTAGA). Мутации можно подтвердить путем рестрикции амплифицированной ДНК с помощью Xba I или путем прямого секвенирования ДНК с применением праймера 5MSP61, меченного по концу 32P. Реакционные смеси, содержащие продукты с несовпадениями, могут быть разделены в 2% низкоплавкой агарозе для последующей очистки и секвенирования. Последовательность примененных праймеров, 5MSP61, 3MSP61mut и 3MSP61wt, представлена ниже и в SEQ ID NO:2, SEQ ID NO:3 и SEQ ID NO:4, соответственно:
5MSP61 (23-член.) 5'-CTA AGC CTG TTG TTT TGC AGG AC-3' (SEQ ID NO:2)
3MSP61mut (20-член.) 5'-CAT GGC ACT ATA CTC TTC TA-3' (SEQ ID NO:3)
3MSP61wt (20-член.) 5'-CAT GGC ACT ATA CTC TTC TT-3' (SEQ ID NO:4)
Последовательность 3MSP61wt содержит 2 или 3 несовпадения с N-ras и K-ras, соответственно, размер фрагмента составляет 110 п.н. Применяли следующую матричную ДНК и реагенты для амплификации:
Условия циклов амплификации с применением термоциклера Perkin Elmer были следующими.
Предварительно нагреть термоциклер до 95°С
Подтверждение корректности теста получали включением следующих контролей: дикого типа (WT), мутанта дикого типа (MUT) и отрицательного контроля (H2O). В качестве образца положительного контроля применяли MUT ДНК из плазмиды pHras61mut. Плазмида pHras61 содержит клонированный экзон 2 Ha-ras ДНК из опухоли мыши Sencar. Клонированная мутация была подтверждена секвенированием ДНК. Мутация представляет собой трансверсию CA→CTA в кодоне 61 (локализованном в экзоне 2) гена Ha-ras мыши в образце ДНК из опухолевой аденокарциномы, содержащей мутацию Ha-ras в кодоне 61 (см. фиг. 14).
Пример 16
Тестирование мутации кодона 12/13 H-ras мыши с помощью горячего PCRTM-RFLP
Данный тест основан на разрушении природно существующего сайта Mn1 I, охватывающего три п.н. кодона 12 и первую п.н. кодона 13 (GGA GGC, нуклеотиды 34-37 в кодирующей последовательности Ha-ras крысы и мыши). Распознаваемым Mn1 I сайтом является N7GGAG. Мутации в любом из четырех положений ведут к неспособности Mn1 I разрезать фрагмент PCRTM, содержащий данную область. Недостатком теста является наличие неполного переваривания под действием Mn1 I. Этот факт затрудняет различение между небольшим процентным содержанием ДНК дикого типа, резистентной к перевариванию, и низким уровнем истинных мутаций. Это иногда наблюдается в тех случаях, когда источник ДНК содержит смесь ДНК дикого типа и мутантной ДНК и когда в тесте применяют 32P для промечивания фрагмента для повышения чувствительности. Набор праймеров PCRTM, применявшийся для тестирования, приведен ниже и в SEQ ID NO:5, и в SEQ ID NO:6. Размер продукта амплификации H-ras 12 составляет 214 п.н.
Праймер #3 (5'): 5'-CCTTGGCTAAGTGTGCTTCTCATTGG-3' (SEQ ID NO:5).
Праймер #6 (3'): 5'-ACAGCCCACCTCTGGCAGGTAGG-3' (SEQ ID NO:6).
Праймер #6 применяют для секвенирования при 55°С при использовании следующих условий реакции.
Условия циклов амплификации с применением набора Perkin Elmer PCRTM были следующими.
Предварительно нагреть термоциклер до 94°С
68°C
72°C
30 сек
1 мин
60°C
72°C
30 сек
1 мин
Пример 17
Результаты тестирования: выявление мутаций c-Ha-ras
Через четыре дня после последнего введения DMBA, растительного экстракта и контроля ДНК, выделенную из свежезамороженных тканей 5 мышей на группу, анализировали на предмет мутаций в кодонах 12 и 13 и кодоне 61 c-Ha-ras с помощью анализа PCRTM. Для данного анализа авторы применяли 4-дневные образцы, поскольку некоторые 2-дневные ДНК подвергались деградации, и, следовательно, были непригодны для анализа Ha-ras. В кодонах 12 и 13 имеется сайт рестрикции Mn1 I, который включает в себя три нуклеотида кодона 12 и первый нуклеотид кодона 13 в последовательности дикого типа. Мутации любого из данных оснований ведут к утрате сайта Mn1 I. Авторы амплифицировали кодон 1 (который содержит кодоны 12 и 13) гена с-Ha-ras из геномной ДНК с помощью термоциклера Perkin Elmer. Реакционную смесь экстрагировали смесью фенол-CHCl3 и ДНК осаждали этанолом. Затем ДНК ресуспендировали в буфере для фермента, продукт PCRTM разрезали Mn1 I и переваренное вещество подвергали электрофорезу в 8% неденатурирующем полиакриламидном геле. Не наблюдалось утраты сайта рестрикции Mn1 I, и было сделано заключение об отсутствии мутаций в кодонах 12 и 13 тестируемого материала. ДНК для анализа Ha-ras получали также из заключенных в парафин срезов 8 мкм из образцов, собранных через два дня после введения последней дозы. 25 срезов из каждого парафинового блока помещали в пробирки микроцентрифуги, депарафинизировали ксилолом и этанолом, центрифугировали и ресуспендировали в 5% хелексе с протеиназой K.
Первая процедура, примененная для кодона 61 Ha-ras, ведет свое начало от Nelson et al. 1992 (пример 15, выше). При применении такого же прямого праймера проводят одну реакцию с обратным праймером с несовпадением (3MSP61mut), а другую реакцию - с обратным праймером дикого типа (3MSP61wt). Данный протокол определяет только трансверсионные мутации CAA→CTA, которые являются преобладающими среди точечных мутаций кодона 61. При данной трансверсии создается сайт Xba I RFLP (T/CTAGA). Реакционные смеси, содержащие продукты с несовпадениями, разделяли в 2% низкоплавкой агарозе для последующей очистки и секвенирования. Отношение количества разрезаемой (ДНК дикого типа) к неразрезаемой (мутантной ДНК) ДНК определяли путем измерения интенсивности окраски бромидом этидия или метки 32P. В качестве образца положительного контроля применялась ДНК из плазмиды pHras61mut. Плазмида pHras61 содержит клонированный экзон 2 Ha-ras ДНК из опухоли мыши Sencar. Клонированная мутация была подтверждена секвенированием ДНК. Мутация представляет собой трансверсию CAA→CTA в кодоне 61 (локализованном в экзоне 2) гена Ha-ras мыши. Условия реакции были описаны в примере 15.
Пример 18
Влияние F035 на инициацию аберрантных крипт у крыс F344, обработанных азоксиметаном
Самцы крыс (Fishcer, 3444) были получены у Charles River (Raleigh, NC) в возрасте 6 недель. Крыс кормили досыта кормом AIN-76A, полученным в Dyets Inc. (Bethlehem, PA). Корм состоял из 20% казеина, 0,3% DL-метионина, 15% кукурузного крахмала, 50% сахарозы, 5% кукурузного масла, 3,5% смеси солей AIN-76, 1% смеси витаминов AIN-76 и 0,2% битартрата холина. Животным также давали водопроводную воду без ограничения. Азоксиметан (AOM), который индуцирует аберрантные крипты у крыс, получали от Sigma Chemical Company (St. Louis, MO). Животных кормили кормом для крыс в течение трех дней карантина, а затем их кормили AIN-76A до возраста 7 недель. Животных случайным образом разделяли на три группы обработки (по 10 животных на группу). Животных группы 1 кормили только кормом AIN-76A, животные группы 2 получали корм AIN-76A + 5 мг/кг F035, а животных группы 3 кормили кормом AIN-76A + 10 мг/кг F035 (таблица 31).
Обработка групп для изучения влияния F035 на инициацию аберрантных крипт у крыс F344 с применением азоксиметана
Через одну неделю кормления животных инъекцировали внутрибрюшинно AOM (15 мг/кг массы тела). Вторую инъекцию AOM производили спустя одну неделю. На протяжении исследования животных еженедельно взвешивали. Через тридцать один день животных забивали асфиксией с помощью CO2. Ободочную кишку вырезали, обмывали ЗФР, разрезали вдоль продольной средней оси, помещали на бумажный фильтр и фиксировали в 70% спирте по меньшей мере в течение 24 час. Ободочную кишку окрашивали метиленовым синим (0,25% в ЗФР) в течение ˜ 1 мин. Очаги аберрантных крипт подсчитывали с помощью анатомического микроскопа при 20x. Аберрантные крипты отличали от окружающих нормальных крипт по их увеличенному размеру, значительно увеличенному расстоянию между люминальной и базальной поверхностями клеток и расширенной перикриптальной зоне. Все образцы были зашифрованы и обсчитывались вслепую двумя операторами. Статистическую значимость определяли путем оценки различий между группами с применением одностороннего ANOVA. Если обнаруживались различия, применяли T-тест Бонферрони для проверки множественных сравнений обеих доз F035 с контрольной группой. Обсчет ободочной кишки производился двумя операторами, каждый из которых не знал, какую экспериментальную группу он оценивает. Между результатами двух операторов было хорошее соответствие.
Было обнаружено, что F035 значительно снижает общее количество очагов аберрантных крипт при добавлении в корм в количестве 10 мг F035 на 1 кг корма, что в грубом приближении соответствует ежедневному приему 1 мг F035 на 1 кг массы тела (таблица 32, таблица 34). Та же доза также значительно снижала количество аберрантных крипт в категориях синглетов и дублетов (таблица 33, таблица 35). Более низкая доза F035, 5 мг на 1 кг корма (в грубом приближении эквивалентная ежедневному приему 500 мкг F035 на 1 кг массы тела) действительно вызывала снижение в общей, синглетной и дублетной категориях очагов аберрантных крипт, но снижение не отличалось значимо от контрольных величин (таблица 33, таблица 35). В ходе исследования не было разницы в прибавке массы между экспериментальной и контрольной группами (таблица 35, таблица 36, таблица 37).
Влияние F035 на количество аберрантных крипт на ободочную кишку у обработанных AOM крыс
+ Отличается значимо от контроля (p<0,05)
C = убедительное исследование
Влияние F035 на количество аберрантных крипт на очаг у обработанных AOM крыс
ст.ош.ср.
ст.ош.ср.
ст.ош.ср.
Сводка сырых данных из анализа влияния F035 на среднее количество аберрантных крипт на ободочную кишку у обработанных AOM крыс
b = среднее из объединенных данных для инъецированных AOM крыс (n=10) через 4 недели
c = значимое отличие от контроля (p<0,05)
Сводка сырых данных из анализа влияния F035 на количество аберрантных крипт на очаг у обработанных AOM крыс
Масса крыс, обработанных AOM, при скармливании F035, 5 мг/кг корма

Масса крыс, обработанных AOM, при скармливании F035, 10 мг/кг корма

Масса крыс, обработанных AOM
Пример 19
In vivo анализ лечения F035 и F060 с применением полных протоколов канцерогенеза и инициации/промоции опухолей
18.1 Полный протокол канцерогенеза
В полном протоколе канцерогенеза мышей обрабатывали 100 нмоль DMBA в 0,2 мл ацетона, наносимыми дважды в неделю в течение 4 недель. Растворы F035 и F060 наносили на обритую область экспериментальных животных за 15 мин перед нанесением DMBA дважды в неделю в течение 4 недель. F035 и F060 наносили в дозах 0,5 мг и 1,0 мг на аппликацию в 0,2 мл ацетона. Через два дня после последней аппликации тестируемых соединений группы из пяти животных забивали для оценки гиперплазии. Оставшиеся животные не подвергались дальнейшим воздействиям. Еще через 4 недели забивали еще по пять мышей из каждой группы для оценки гиперплазии. Оставшихся мышей оставляли суммарно на 16 недель для оценки тяжести опухоли. Результаты исследования представлены в таблице 39 ниже.
18.2 Протокол инициации/промоции опухоли
В протоколе инициации/промоции опухолей опухоли инициировали у мышей однократной аппликацией 10 нмоль DMBA в 0,2 мл ацетона. Одну неделю спустя мышей начинали обрабатывать дважды в неделю 2 мкг TPA в 0,2 мл ацетона в течение всего эксперимента. Промоцию останавливали через 16 недель. F035 и F060 (0,5 мг и 1,0 мг на аппликацию в 0,2 мл ацетона) наносили на обритую область спины экспериментальных животных за 15 мин до аппликации TPA в течение всего эксперимента. Контрольных мышей обрабатывали 0,2 мл ацетона. По пять мышей на группу забивали через 8 недель обработки. Оставшихся мышей оставляли еще на 8 недель для оценки тяжести опухоли. Степень и множественность опухоли регистрировали еженедельно для каждой отдельной мыши. Результаты представлены в таблице 39 ниже.
18.3 Гистологическая оценка
После забоя мышей образцы кожи стандартно фиксировали в формалине и обрабатывали для гистологического анализа. Ткани для гистологической оценки получали с применением традиционных парафиновых срезов и окрашивания гематоксилином и эозином. Приблизительно 1 см2 каждого образца кожи сохраняли в формалине для получения срезов; остальное быстро замораживали в жидком азоте для выделения ДНК. Толщину эпителия определяли по меньшей мере по 20 случайно выбранным участкам в фиксированных формалином образцах кожи. Толщину дермы, от базальной мембраны до жировой ткани, также определяли минимум в 20 случайно выбранных участках на одно животное. Через 2 дня после прекращения введения дозы определяли относительное соотношение клеток воспаления (полиморфоядерных лейкоцитов и лимфоцитов, макрофагов, фибробластов и тучных клеток) дермы путем измерения количества клеток на 1 мкм2 в 10 полях с большим увеличением (x1000) на животное. Данное количество определяется для аналитических экспериментальных целей как общая клеточность дермы.
Кроме того, опухоли, полученные на 12 и 16 неделях, подвергали хромосомному анализу с применением прямого цитогенетического способа (Conti et al., 1986). Папилломы гомогенизировали и инкубировали с ферментативным раствором, содержащим 0,02 мкг/мл колцемида (GIBCO). Диспергированные клетки промывали дважды и ресуспендировали в гипотоническом растворе 0,075 М KCl в течение 10 мин при 37°С. Клетки осаждали, фиксировали в смеси метанол-уксусная кислота (3:1) и окрашивали Гиемса. Хромосомы считали на Cell Analysis Systems (Elmhurst, IL), как описано (Conti et al., 1986).
18.4 Подавление анэуплоидии
При полном протоколе канцерогенеза через 12 недель папилломы были хорошо дифференцированными гиперпластическими повреждениями с небольшой клеточной атипией или без нее. Клетки были диплоидными. Через 16 недель 10% клеток были диспластическими и гипердиплоидными. У мышей, обработанных тритерпеноидными сапонинами, развившиеся папилломы не были гипердиплоидными. В эксперименте с инициацией/промоцией через 12 недель 30% контрольных пренеопластических опухолей были гипердиплоидными. Через 16 недель 50% опухолей были гипердиплоидными. Примечательно, что не было свидетельств анэуплоидии у мышей, обработанных тритерпеноидными сапонинами (см. таблицу 39 ниже).
Результаты являются важными, поскольку анэуплоидия, дисбаланс хромосом, который может вести к нестабильности генома, характерна для большинства солидных раков человека (Duesberg et al., 2000). Хотя генотоксические и негенотоксические канцерогены сами по себе могут вызывать анэуплоидию, промоторы опухолей являются особенно сильными индукторами хромосомных повреждений отчасти за счет выделения активных форм кислорода (АФК), таких как H2O2 (Conti et al., 1986; Duesberg et al., 2000; Cerutti, 1985; Dutton and Bowden, 1985).
Механизмы репарации повреждений ДНК широко сохраняются в ходе эволюции. Кроме того, поврежденные клетки могут быть удалены путем апоптоза (Whal and Vafa, 2000). Продолжаются споры относительно того, предшествуют ли соматические мутации анэуплоидии или наоборот (Duesberg et al., 1999). Однако геномная нестабильность может быть результатом действия активированных онкогенов, часто связанных с АФК (Whal and Vafa, 2000; Denko et al., 1994; Felsher and Bishop, 1999), аномальным хромосомным набором или мутациями в митотическом аппарате (Cahill et al., 1999; Lengauer et al., 1998). Таким образом, описанные здесь результаты, т.е. то, что авицины подавляют и мутации H-ras (которые имеют место до появления анэуплоидного кариотипа), и анэуплоидию, являются важными, поскольку мутации H-ras и анэуплоидия являются ранними событиями в развитии папилломы кожи мыши и, возможно, многих эпителиальных злокачественных образований человека. Результаты исследования, таким образом, указывают на полезность соединений согласно изобретению для предотвращения хромосомных аномалий. Такие применения включают в себя обработку клетки, ткани или больного соединениями согласно изобретению для предотвращения хромосомной нестабильности и связанных эффектов. Такая нестабильность может быть ассоциирована с канцерогенезом и может быть также ассоциирована с различными типами лечения, которые могут применяться для лечения пролиферативных заболеваний. Путем введения соединений согласно изобретению, профилактического или терапевтического, вредные эффекты, связанные с хромосомной нестабильностью, могут быть сведены к минимуму или предотвращены.
Ингибирование F035 развития папилломы в экспериментах с полным канцерогенезом и инициацией/промоцией у мышей SENCAR
Концентрации тестируемых соединений; * 100 нмоль (2x/нед. 4 нед.); † 1,0 мг (2x/нед. 4 нед.); ‡ 1,0 мг (2x/нед. 4 нед.); § 10 нмоль (1x); ¶ 1,0 мг (2x/нед. 8 нед.); II 2 мкг (2x/нед. 8 нед.); ** 1,0 мг (2x/нед. 8 нед.). В скобках показан % папиллом с анэуплоидией.
Пример 20
Противоопухолевая активность агликонов
Исследования подтверждают важность сахара для биологической активности, поскольку удаление сахаров из базовой тритерпеновой молекулы ведет к значительной потере биологической активности. Как показано в таблице 40, UA-BRF-004Pod-DELEP-F164 (полученная гидролизом сахаров из UA-BRF-004Pod-DELEP-F094 с присоединенными эфирами) и UA-BRF-004Pod-DELEP-F245 (смесь метилового эфира продукта гидролиза UA-BRF-004Pod-DELEP-F094) проявляют заметную потерю противоопухолевой активности в отношении набора линий опухолевых клеток. Сходным образом, UA-BRF-004Pod-DELEP-F194 (очищенный ацетат агликона 1) проявляет значительную потерю противоопухолевой активности в отношении набора линий опухолевых клеток по сравнению с данными для тритерпенового гликозида фракции 35. Таким образом, имеется выраженная потеря биологической активности после гидролиза сахарных единиц из раскрытого здесь тритерпенового гликозида.
Биологическое тестирование фракций F164, F245 и C194
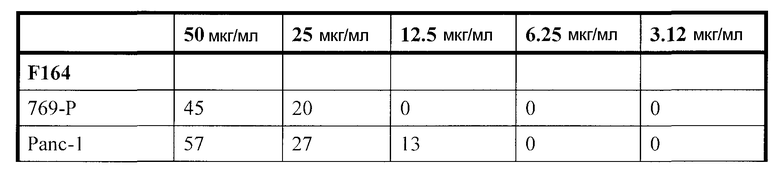

Пример 21
Анализ влияния F035 на метаболизм холестерина
Целью данного исследования является анализ влияния биологически активных тритерпеновых гликозидов согласно изобретению на профилактику сердечно-сосудистого заболевания. Долгосрочная цель данного исследования заключается в демонстрации того, что тритерпеновые соединения, добавленные в пищу млекопитающих, включая человека, обычно снижают холестерин сыворотки. Выбранная для исследования модель гиперлипидемии хомячков представляет собой модель на грызунах, которая в отличие от модели на крысах, близко имитирует изменения и рецептора ЛПНП, и липопротеинов плазмы человека в зависимости от содержания холестерина (Spady et al., 1993).
Тритерпеновый гликозид вводят в двух различных концентрациях в очищенный корм для хомячков без каких-либо изменений уровня кальция, калия, фосфора или других необходимых компонентов пищи. Два разных уровня добавки тритерпенового гликозида применяют для того, чтобы показать дозо-зависимость связи. Животных кормили при свободном доступе к Dyets, очищенному корму для хомячков, составленному в соответствии с рекомендациями NCR (Reevers et al., 1993) с добавкой 1% холестерина или без него (Davis et al., 1989). Очищенный корм для хомячков Dyets, содержащий холестерин, модифицируют тритерпеновым гликозидом, применяя указанные ниже концентрации. Гранулированный корм для исследования и контроля приготовлен Dyets, Inc. (Bethlehem, PA) без изменений содержания кальция, фосфора или любого необходимого питательного микроэлемента. Еженедельно регистрировали потребление пищи и прибавку массы тела животных. Использованные животные являются четырехнедельными аутбредными самцами свободных от вирусов золотистых сирийских хомячков (Charles River Laboratories, Wilmington, MA). Животных рандомизируют по массе, используя генератор случайных чисел программы Statview, помещают по 3 в клетку, в комнате, освещаемой 12 час в день, и содержат при температуре 22±1,0°С.
Через 0, 4 и 8 недель жизни на соответствующей диете с тритерпеновым гликозидом или без него случайным образом отбирают 12 животных на группу и забивают в 9-11 утра. Удаляют печень и почки, взвешивают, обрабатывают и хранят при -70°С для последующих исследований. Кровь получают при забое пункцией сердца до удаления печени и почек и анализируют на профиль липидов. Липидные профили сыворотки крови анализируют в обработанных и контрольных группах. Имеется две контрольные группы (группы 1 и 2) и две обработанные группы (группы 3 и 4). Все группы получают корм NRC для хомячков во время двухнедельного периода карантина. Группу 1 оставляют на корме NRC до окончания исследования. Группы 2-4 питаются кормом NRC плюс 1% холестерин еще в течение двухнедельного периода для индукции гиперхолестеринемии. Затем группу 2 оставляют на данной диете до окончания исследования, а группы 3 и 4 кормят той же диетой с добавкой тритерпенового гликозида (например, F035 или F094). Сводка по обработке групп дана ниже в таблице 41.
Схема модификации диеты
bChol = холестерин
cTG = тритерпеновый гликозид
Через 0 (только контрольная группа), 4 и 8 недель жизни на соответствующей диете с тритерпеновым гликозидом или без него случайным образом отбирали 12 животных на группу и забивали в 9-11 утра. Удаляли печень и почки хомячков, взвешивали и обрабатывали на предмет возможных аномалий. Часть каждого органа, проявляющего аномалии, готовили для гистологического анализа, т.е. замораживали для парафиновых срезов, и срезы окрашивали гематоксилином и эозином. Кровь получали при забое пункцией сердца перед хирургическим удалением печени и почек. Получали сыворотку, которую хранили при -20°С для анализа профиля липидов. Перед забоем хомячки голодали ночь. Данные показаны в таблице 42 ниже.
Собранные в ходе данного исследования образцы крови применяют для определения общего холестерина, триглицеридов, ЛПВП-холестерина и ЛПНП-холестерина плюс ЛПОНП-холестерина (Mackness and Durrington, 1992) в Roche Biomedical Laboratories, Burlington, NC. Статистический анализ данных производят на компьютере Power Macintosh 9600 с помощью программы Macintosh для одностороннего анализа дисперсии, величины p и линейной регрессии (Armitage, 1971). В частности, анализ данных по профилям липидов в каждой группе диета/лекарство проводят с помощью анализа дисперсии с применением разделений среднего Ньюмана-Кейлса (Steel and Torrie, 1980).
Влияние непрерывного скармливания тритерпенового гликозида (TG) хомячкам в течение шести недель
12 хомячков на группу кормили очищенным кормом для хомячков плюс 1% холестерин
Пример 22
Исследование предотвращения индуцированного UVB канцерогенеза фракцией 35
Данное исследование будет сфокусировано на предотвращении активными тритерпеновыми гликозидами согласно изобретению индуцированного UVB канцерогенеза на модели кожи мышей. Долгосрочной целью исследования является демонстрация того, что на модели кожи мышей тритерпеновые гликозиды предотвращают индуцированные UVB повреждения. Экспериментальную модель на мышах применяют потому, что модель близко напоминает ситуацию у человека. В исследовании авторы будут стремиться продемонстрировать, что местное нанесение активных тритерпеновых гликозидов согласно изобретению в ацетоне на кожу спины бесшерстных мышей SKH-1, облученных UVB, должно предотвращать вызываемые UVB повреждения кожи.
В исследовании бесшерстных мышей SKH-1 облучают UVB-облучением в дозе 1,8 кДж/м2 до 15 мин. Мышей предварительно обрабатывают двумя разными дозами F035 (2 мг и 4 мг на дозу), а также отрицательным контролем (F060 или одним ацетоном). Полагают, что для получения статистически значимых результатов необходимо как минимум 10 мышей на группу. Каждое тестируемое соединение наносится местно за 10 мин до облучения 3 раза в неделю на протяжении до 6-10 недель. Исследования проводят в течение короткого промежутка времени для оценки профилактического действия соединений. В указанный отрезок времени не ожидается обнаружения видимых опухолей, даже с одним UVB, а лишь повреждения кожи. Может наблюдаться слабая эритема (слабое покраснение кожи), которая должна исчезнуть на следующий день после облучения.
Применявшийся УФ-прибор содержит восемь солнечных ламп Westinghouse FS40, радиометр/фотометр IL-1400A и присоединенный фототерапевтический радиометр IL-1403 UVB с детектором SEL 240/UVB-1/TD. Средняя часть содержит в себя несколько камер, в каждую из которых помещают одну мышь. В камерах имеются отверстия для надлежащей вентиляции во время облучения мышей. Во время облучения камеры вращаются по кругу, в результате чего все мыши одинаково экспонируются UVB. В данном устройстве имеются дверцы, которые можно закрывать на время включения лампы UVB, так что свет UVB имеется внутри устройства. Доза экспозиции с UVB должна измеряться радиометром UVB. Мышей следует оставлять в камерах не дольше, чем на 10-15 мин.
Целью исследования является установление фотопротекторного действия от повреждения кожи мышей. UVB поглощается непосредственно клеточной ДНК и вызывает повреждения, которые могут быть причиной мутаций гена(ов)-мишени(ей), ведущих в конечном итоге к раку. Раннее обнаружение таких повреждений и профилактика таких повреждений могут указывать на хемопротекторное действие (Berton et al., 1997; Chatterjee et al., 1996; Youn et al., 1997; Shirazi et al., 1996; Baba et al., 1996; Takema et al., 1996).
Режим облучения UVB
Использованные в исследовании обработанные группы указаны в таблице 43. Полагают, что размер групп достаточен для контроля вариации гиперплазии и воспаления кожи в данной группе, вариабельности толщины эпидермиса и воспаления кожи у животных одного возраста и стадии развития. Гиперплазия и воспаление кожи являются главными измеряемыми параметрами в данном исследовании. Оставшуюся кожу оставляют для измерения других биологических маркеров типа модифицированных оснований ДНК (8-OH-dG) и экспрессии онкогенов (онкоген Ha-ras).
На протяжении исследования животные должны иметь свободный доступ к гранулированному корму и питьевой воде. Животные должны подвергаться еженедельному контролю на предмет потребления пищи и прибавки массы тела. Использованные животные являются семинедельными аутбредными самками свободных от вирусов бесшерстных мышей SKH-1 (Charles River Laboratories, Wilmington, MA). Животных рандомизируют по массе, используя генератор случайных чисел программы Statview, помещают по 5 в клетку в комнате, освещаемой 12 час в день, и содержат при температуре 22°С ± 1,0°С.
Статистический анализ данных производят на компьютере Power Macintosh G3 с помощью программы Macintosh для одностороннего анализа дисперсии, величины p и линейной регрессии (Armitage, 1971). В частности, анализ данных по толщине эпидермиса в каждой группе с лекарством проводят с помощью анализа дисперсии (Armitage, 1971).
Пример 23
Влияние биологически активных тритерпенов на экспрессию белков, участвующих в остановке клеточного цикла и апоптозе
Апоптоз определяют как нормальный физиологический процесс запрограммированной гибели клеток, который происходит во время эмбрионального развития и во время поддержания тканевого гомеостаза. Процесс апоптоза может быть разделен на серии метаболических изменений в апоптотических клетках. Для демонстрации того, что апоптоз происходит в клетке или клеточной популяции или для демонстрации того, что процесс гибели клеток нарушен в раковых клетках, могут быть определены отдельные ферментативные этапы нескольких регуляторных путей или путей проведения сигнала. Программа апоптоза обнаруживается также по морфологическим признакам, которые включают в себя изменения в плазматической мембране (такие, как утрата асимметрии), конденсацию цитоплазмы и ядра, межнуклеосомный разрыв ДНК. Кульминацией этого является гибель клетки с вырождением клетки в "апоптотические тела".
Способы анализа нескольких ферментативных и сигнальных процессов, вовлеченных в апоптоз, были разработаны в форме стандартных протоколов для многопараметрического исследования апоптоза. Одним из примеров ранней стадии апоптоза является высвобождение цитохрома c из митохондрий с последующей активацией пути каспазы-3 (PharMingen, SanDiego, CA). Индукция каспаз (ряда цитозольных протеаз) является одной из наиболее постоянно наблюдаемых характерных черт апоптоза. В частности, каспаза-3 играет центральную роль в данном процессе. Активированные каспазы расщепляют белки-мишени, одним из наиболее важных из которых является PARP (белок, локализованный в ядре). Таким образом, тесты, определяющие высвобождение цитохрома c, определяющие активность каспазы-3 и определяющие деградацию PARP, являются эффективными показателями апоптоза.
Более того, можно заключить, что агенты, которые вызывают высвобождение цитохрома c из митохондрий злокачественных клеток, по-видимому, являются терапевтическими для восстановления по меньшей мере некоторых сторон клеточного контроля запрограммированной гибели клеток.
Другой апоптотический тест заключается в определении аннексина-V (BioWhitaker, Walkerville, MD). В норме фосфатидилсерин (PS) локализован на внутренней мембране плазматической мембраны. Однако на ранних стадиях апоптоза происходит экстернализация PS. Аннексин-V представляет собой кальцийсвязывающий белок, который связывается с PS и может выявляться путем окрашивания аннексин-V-ФИТЦ с помощью проточной цитометрии (Martin et al., 1995). Способность клеток, обработанных описанными в данном изобретении соединениями Acacia victoriae, связывать аннексин-V принята как указание на то, что клетки подверглись апоптозу.
В других примерах авторы изобретения применяли тестирование PI-3-киназы для определения апоптотической активности клеток, обработанных противораковыми соединениями, выделенными из Acacia victoriae. Фосфоинозитид-3-киназа (PI3K), ассоциированный с клеточной мембраной фермент, способна фосфорилировать положение 3 кольца инозита фосфатидилинозита, определяя тем самым новый путь липидной сигнализации в тех клетках, где активна PI3K. Когда активна PI3K, киназа, называемая AKT, рекрутируется в клеточную мембрану. AKT является продуктом онкогена, который каталитически активируется после рекрутирования в мембрану. Полностью активированная AKT играет решающую роль в выживании клетки. Путь PI3K/AKT обеспечивает механизм, с помощью которого клетки избегают апоптоза. Таким образом, средства, ингибирующие PI3K в злокачественных клетках, по-видимому, являются терапевтическими для восстановления по меньшей мере некоторых сторон клеточного контроля апоптоза.
Пример 24
Анализ клеточного цикла
Анализ клеточного цикла проводили с помощью проточной цитометрии с применением стандартных способов с некоторыми модификациями. Вкратце, 1 x 106 клеток помещали в чашки 60 мм3 и экспонировали с различными концентрациями F035 в течение 72 час при 37°C. Клетки промывали ЗФР и ресуспендировали в концентрации 1 x 106 клеток/мл. Клетки фиксировали сначала 1% параформальдегидом и затем охлажденным до температуры льда 70% этанолом. После этого клетки окрашивали иодидом пропидия (10 мкг/мл, Sigma Chemical Co., St. Louis, MO), содержащим 0,1% РНКазы (Sigma) в течение 30 минут при комнатной температуре и анализировали на Beckton Dickinson FAC SCAN.
Пример 25
Анализ связывания аннексин-V-фруоресцеинизотиоцианата (ФИТЦ)
Индукцию апоптоза раковых клеток исследовали с помощью анализа связывания аннексин-V-ФИТЦ. Клетки Jurkat (1x106) обрабатывали различными концентрациями смеси тритерпеновых гликозидов (F035) и чистыми экстрактами D1 и G1 (0,5-2,0 мкг/мл) в течение 18 час при 37°C. После промывки в ЗФР клетки ресуспендировали в буфере для связывания (10 мМ HEPES/NaOH, 140 мМ NaCl, 2 мМ CaCl2), содержащем 5 мкл конъюгата аннексин-V-ФИТЦ (BioWhittaker, Walkersville, MD) и инкубировали в течение 10 минут в темноте. Затем клетки окрашивали иодидом пропидия (20 мкг/мл) и анализировали с помощью проточной цитометрии (Martin et al., 1995).
Пример 26
Анализ фосфатидилинозитол-3-киназы (PI3-киназы)
Лишенные сыворотки клетки Jurkat обрабатывали 2 мкг/мл F035 в течение 2-15 час или 0,5 час с вортманнином при 37°С. PI3K-активность определяли, как описано (Whitman et al., 1985; Royal and Park, 1995). PI3-киназу иммунопреципитировали из 1 мг клеточного белка с применением 5 мкл антисыворотки кролика против p85 при 4°С в течение 90 мин. Иммунные комплексы собирали на 20% шариках протеин A-сефарозы в течение 90 мин при 4°С. Затем иммунопреципитат ресуспендировали в 30 мкл буфера для киназной реакции (33 мМ трис, pH 7,6, 125 мМ NaCl, 15 мМ MgCl2, 200 мМ аденозина, 20 мМ АТФ, 30 мкКи [g-32P]аденозинтрифосфата АТФ). PI3-киназную реакцию инициировали добавлением 10 мкл суспензии PI и 10 мкл гамма-АТФ и давали ей пройти в течение 30 мин при комнатной температуре. Реакцию прекращали добавлением 100 мкл 1 н. HCl. Липиды экстрагировали из реакционной смеси смесью хлороформ : метанол (1:1) и разделяли тонкослойной хроматографией (ТСХ) в системе хлороформ : метанол : NH4OH : H2O (60:47:2:11,3) на пластинах силикагеля G60. Радиоактивно меченый фосфатидилинозитол (PI) фосфат визуализировали авторадиографией и ингибирование количественно определяли с помощью системы Storm 860 (Molecular Dynamics).
Пример 27
Анализ общей и фосфорилированной форм AKT
Экспрессию общей и фосфорилированной форм AKT определяли с помощью иммуноблоттинга. Клетки Jurkat, культивированные в среде, содержащей 0,5% FBS, обрабатывали F035 и чистыми экстрактами D1 и G1 (2,0 мкг/мл) в течение 15 час при 37°С. Клетки лизировали в буфере для лизиса AKT (20 мМ трис-HCl, 150 мМ NaCl, 1 мМ ЭДТА, 1 мМ ЭГТА, 1% тритон X-100, 2,5 мМ пирофосфат натрия, 1 мМ 8-глицеринфосфат, 1 мМ Na3VO4, 1 мл лейпептин, 1 мМ PMSF pH 7,5). Клеточный белок (40 мкг) разделяли на 8% ДДС-Na-полиакриламидном геле и переносили электропереносом на нитроцеллюлозную мембрану. Мембраны сначала зондировали фосфоспецифическим AKT (Ser 473) или AKT-антителом и затем антикроличьим антителом козы, конъюгированным с пероксидазой хрена. Белки определяли по хемилюминесценции (ECL, Amersham, Arlington Heights, IL).
Пример 28
Анализ сдвига электрофоретической подвижности (EMSA)
EMSA для изучения влияния сырого (F035) и очищенных экстрактов D1 и G1 на индуцированный TNF (Genetech Inc.) NF-κB проводили, как описано ранее. Клетки Jurkat (1 x 106/мл) обрабатывали различными концентрациями сырого и чистых экстрактов в течение 15 час при 37°С. Затем клетки экспонировали с 100 пМ TNF в течение 15 мин при 37°С. Ядерные экстракты получали, как описано выше. Ядерные экстракты инкубировали с 16 фмоль меченного 32P по концу 45-членного двухцепочечного олигонуклеотида NF-κB из длинного концевого повтора вируса иммунодефицита человека,
5'-TTGTTACAAGGGACTTTCCGCTGGGGACTTTCCAGGGAGGCTGG-3' (SEQ. ID NO.9),
в течение 15 мин при 37°С в присутствии 2 мкг поли (dI-dC). Комплекс белок-ДНК отделяли от свободного олигонуклеотида на 7,5% неденатурирующем полиакриламидном геле. Радиоактивные полосы высушенных гелей визуализировали и количественно определяли с помощью PhosphoImager (Molecular Dynamics, Sunnyvale, CA) с применением программы ImageQuant.
Пример 29
Индукция и анализ индуцибельной синтазы оксида азота (iNOS)
Для изучения iNOS применяли клетки U-937 и Jurkat. Вызывали дифференцировку клеток U-937 в макрофаги путем их культивирования с PMA (100 нМ) в течение 72 час при 37°С. Дифференцированные клетки обрабатывали F035 (2 мкг/мл) в течение 15 час с последующей обработкой LPS (10 мкг/мл) в течение 4 час для индукции iNOS. В клетках Jurkat iNOS индуцировали обработкой 0,5 x 106/мл клеток PHA (10 мкг/мл) и PMA (10 мл) в течение 24 час при 37°C. Лизаты клеток получали повторным замораживанием и размораживанием в буфере RIPA (1% NP-40, 0,5% дезоксихолат Na, 0,1% ДДС-Na в ЗФР). Клеточный белок (200 мкг) разделяли на 7,5% ДДС-Na-полиакриламидном геле, переносили электропереносом на нитроцеллюлозную мембрану, зондировали анти-iNOS-антителом кролика и затем антикроличьим антителом козы, конъюгированным с пероксидазой хрена. Белковые полосы определяли по хемилюминесценции (ECL, Amersham, Arlington Heights, IL).
Пример 30
Смесь и чистые тритерпеновые гликозиды оказывают цитотоксическое действие на опухолевые клетки
Влияние смеси тритерпеновых гликозидов (F035) на жизнеспособность панели раковых и нетрансформированных клеток исследовали, как описано в способах. Как показано на фиг. 42, клетки Jurkat (T-клеточная лимфома) были высокочувствительными к F035 с IC50, равной 0,2 мкг/мл. Сходным образом F035 ингибировала рост ряда линий раковых клеток ингибирующей концентрацией IC50 в диапазоне 1,7-2,8 (для раковых клеток яичников), 2,0-3,3 (почек), 0,93 (поджелудочной железы), 1,2-6,5 (простаты) и 0,72-4,0 мл (некоторых раковых клеток молочной железы). Однако остальные линии клеток рака молочной железы были резистентны к цитотоксическому действию F035. Последние четыре столбика на фиг. 42 показывают, что чтобы вызвать гибель 50% нетрансформированных клеток (фибробластов человека и мыши и иммортализованного эпителия молочной железы) требуется более 25 мкг/мл F035, что позволяет предполагать, что F035 цитотоксична специфично в отношении раковых клеток.
Кроме того, на 5 клеточных линиях тестировали цитотоксичность двух чистых тритерпеновых гликозидов D1 и G1. На фиг. 43 показано, что D1 имеет IC50, сопоставимую с таковой F035 на трех клеточных линиях (769-P, MDA-MB-453 и MDA-MB-231). На клетках C-2 (варианте HEY) и Jurkat D1 был в два раза эффективнее, чем F035. Однако G1 был значительно цитотоксичнее, чем F035 и D1 в отношении большинства тестированных клеток, что могло бы быть из-за того, что G1 менее полярен, чем экстракт D1 (фиг. 43).
Пример 31
Остановка клеточного цикла и индукция апоптоза под действием смеси тритерпеновых гликозидов
Для изучения влияния F035 на клеточный цикл линии раковых клеток MDA-MB-453 и MDA-MB-435 обрабатывали различными концентрациями F035. В таблице 44 показано увеличение количества клеток в G1 (7-10%) и одновременное снижение % клеток в S-фазе (6-10%), что предполагает остановку G1 в клетках MDA-MB-453. Кроме того, через 72 час после обработки F035 16% клеток MDA-MB-435 (другой линии раковых клеток молочной железы), по-видимому, находились в фазе суб-Go клеточного цикла (таблица 44), что позволяет предполагать, что клетки подвергаются апоптозу. Данное наблюдение было дополнительно подтверждено исследованием апоптоза с помощью теста TUNEL.
Анализ клеточного цикла обработанных F035 клеток
Клетки MDA-MB-453 и MDA-MB-435 обрабатывали различными концентрациями F035 в течение 72 час при 37°С. Анализ клеточного цикла проводили после окраски иодидом пропидия, как описано в способах.
Для того чтобы понять механизм, лежащий в основе индуцированной F035 гибели клеток, авторы изобретения провели анализ связывания аннексина V-ФИТЦ с применением клеток Jurkat, обработанных F035, D1 и G1. В таблице 45 показано связывание аннексина V с клетками, обработанными 1 мл F035, D1 и G1 (15-17%), что тем самым указывает на апоптотический путь, ведущий к клеточной гибели.
Анализ 72-часовой цитотоксичности клеток Jurkat (T-клеточного лейкоза)
Пример 32
Смесь тритерпеновых гликозидов ингибирует активность PI3-киназы
Для изучения молекулярной(ых) мишени(ей) F035 авторы исследовали PI3-киназный путь сигнализации. Результаты иммунопреципитации антителом против p85 (адапторного белка) и последующего тестирования липидкиназной активности показали, что F035 ингибирует активность PI3-киназы в клетках Jurkat. На фиг. 45A показано приблизительно 50-70% ингибирование активности PI3-киназы через 2 час после обработки F035. К 6 час наблюдалось 92-95% ингибирование активности PI3-киназы, которое сохранялось до 15 час после обработки. Вортманнин [1 мкМ, 30 мин после обработки], известный ингибитор PI3-киназы, проявлял сходное ингибирование ферментативной активности в клетках Jurkat (фиг. 45A).
Пример 33
Смесь тритерпеновых гликозидов, D1 и G1, ингибирует фосфорилирование AKT
Авторы изобретения определяли действие F035 и чистых экстрактов на AKT, серин-треонинкиназу, расположенный ниже эффектор PI3-киназного пути сигнализации. В отличие от быстрого ингибирования активности PI3-киназы ингибирование фосфорилирования AKT не происходило до 15 час после обработки. Обработка клеток Jurkat F035 (2 мл) в течение 15 час вела к снижению фосфорилирования AKT. Однако данная обработка вела также к снижению уровня общего белка AKT, как это можно видеть на фиг. 45B. Авторы изобретения подтвердили ингибирование активности AKT с чистыми тритерпеновыми гликозидами. Чистые тритерпеновые гликозиды D1 и G1 (2 мкг/мл) также ингибировали фосфорилирование AKT и экспрессию общего белка AKT (фиг. 45B). Обработка клеток Jurkat LY 294002 и вортманнином (известными ингибиторами PI3-киназы) показала ингибирование фосфорилирования AKT.
Пример 34
Смесь тритерпеновых гликозидов, D1 и G1, ингибирует индуцированный TNF NF-κB
Для дальнейшего изучения посредников апоптотического пути авторы оценивали влияние F035, D1 и G1 на транскрипционный фактор NF-κB, который, как было показано, участвует в апоптозе. Представленные на фиг 46A результаты показывают, что F035 зависимо от дозы ингибировала зависимую от TNF активацию NF-κB. Необработанные клетки и клетки, обработанные только F035, не показали активации NF-κB. Авторы также подтвердили данные результаты с чистыми экстрактами D1 и G1. Предварительная обработка клеток 2 мл G1 и D1 приводила к 54 и 87% снижению уровня NF-κB, соответственно (фиг. 46B). Обработка клеток только одним D1 или G1 не вызывала активацию NF-κB (фиг. 46В). В соответствии с недавно показанной регуляцией NF-κB PI3-киназой предварительная обработка клеток вортманнином (1 мкМ) приводила к почти полному ингибированию NF-κB, индуцированного TNF.
Пример 35
Ингибирование F035 iNOS
Поскольку транскрипция iNOS регулируется NF-κB, авторы исследовали влияние F035 на индукцию iNOS. В клетках U-937, которые были дифференцированы в макрофаги, авторы индуцировали iNOS в ответ на LPS (фиг. 46C). Предварительная обработка этих клеток F035 (1 мкг/мл) полностью блокировала индукцию iNOS. Вортманнин оказывал сходное действие на индуцированную LPS iNOS в данных клетках.
Авторы изобретения также проверяли действие F035 на индукцию iNOS в клетках Jurkat. iNOS индуцировали PHA и PMA, как описано в способах. Результаты показывают, что предварительная обработка клеток Jurkat F035 блокировала индукцию iNOS (фиг. 46D).
Пример 36
Анализ деградации PARP с помощью иммуноблоттинга
Индуцированный F035 и D1 апоптоз проверяли по протеолитическому расщеплению поли(АДФ-рибоза)полимеразы (PARP). Клетки Jurkat (2X106/мл) обрабатывали F035 (2 мкг/мл) и D1 (2 мкг/мл) в течение различных промежутков времени. Клеточные лизаты получали в буфере, содержащем 20 мМ HEPES, 250 мМ NaCl, 2 мМ ЭДТА, 0,1% NP-40, 2 мкг/мл лейпептин, 2 мкг/мл апротинин, 0,5 мкг/мл бензамидин, 1 мМ DTT и 1 мМ PMSF. Клеточные белки (60 мкг/мл) разделяли на 7,5% ДДС-Na полиакриламидном геле и переносили электропереносом на нитроцеллюлозную мембрану. Мембрану сначала зондировали моноклональным антителом против PARP (PharMingen), а затем антимышиным антителом, конъюгированным с пероксидазой хрена (HRPO). Белковые полосы определяли по хемилюминесценции (ECL, Amersham). Степень расщепления 116-кДа PARP до пептидных продуктов 85кДа и 41 кДа использовали в качестве меры апоптоза (Tewari et al., 1995).
Пример 37
Тестирование протеазы каспазы-3
Активность каспазы-3 измеряли, как описано ранее (Enari et al., 1995) с некоторыми модификациями. Вкратце, клетки Jurkat (1X106/мл) обрабатывали F035, D1 и G1 в течение различных промежутков времени. Цитозольные экстракты получали повторным замораживанием-оттаиванием в 300 мкл буфера для экстракции (12,5 мМ трис, pH 7,0, 1 мМ DTT, 0,125 мМ ЭДТА, 5% глицерин, 1 мкМ PMSF, 1 мкг/мл лейпептин, 1 мкг/мл пепстатин и 1 мкг/мл апротинин). Клеточные лизаты разводили 1:2 буфером ICE (50 мМ трис, pH 7,0, 0,5 мМ ЭДТА, 4 мМ DTT и 20% глицерин) и инкубировали с 20 мкМ субстрата каспазы 3 (ацетил-Asp-Glu-Val-Asp-аминометилкумарин) при 37°С. Активность каспазы-3 отслеживали по образованию флуоресцентного аминометилкумарина, который измеряли при возбуждении 355 нм, эмиссии 460 нМ с применением Fluoroscan II (Labsystems, Helsinki, Финляндия).
Пример 38
Определение высвобождения цитохрома c из митохондрий
Высвобождение цитохрома c из митохондрий в ответ на обработку F035 определяли с помощью иммуноблоттинга. Клетки Jurkat (10X106/мл) обрабатывали 2 мкг/мл F035 в течение 4 и 6 час при 37°С. Осадки клеток промывали сахарозным буфером (0,25 М сахароза, 30 мМ трис, pH 7,7, 1 мМ ЭДТА). К осадкам клеток добавляли 20 мкл сахарозного буфера, содержащего 1 мкМ PMSF, 1 мкг/мл лейпептина, 1 мкг/мл пепстатина и 1 мкг/мл апротинина. Клетки разрушали в помощью douncing 120 раз в 0,3 мл Kontes douncer с пестиком B (Kontes Glass company). Клеточный белок (60 мкг) разделяли на 15% ДДС-Na полиакриламидном геле и переносили электропереносом на нитроцеллюлозную мембрану. Мембрану сначала зондировали моноклональным антителом против цитохрома c (PharMingen), а затем антимышиным антителом, конъюгированным с пероксидазой хрена (HRPO). Белковые полосы определяли по хемилюминесценции (ECL, Amersham).
Пример 39
F035 и D1 индуцируют расщепление PARP
F035 и D1 индуцируют расщепление PARP в клетках Jurkat зависимым от времени образом. Результаты на фиг. 47 показывают, что к 4 час F035 и D1 начинают индуцировать расщепление PARP, а близкое к завершению расщепление имеет место к 6-8 час. Это указывает на роль каспаз и тем самым апоптоза в механизме, вовлеченном в индуцированную F035 и D1 гибель клеток.
Пример 40
Влияние z-vad fmk на индуцированную F035 гибель клеток
Для дополнительного подтверждения роли каспаз в опосредуемой F035 гибели клеток изобретатели изучали влияние z-vad fmk, ингибитора каспаз, на клетки, обработанные F035. Предварительная обработка клеток Jurkat 100 мкМ z-vad fmk в течение 1 час при 37°С полностью предотвращала индуцированное F035 расщепление PARP (фиг. 48).
Пример 41
F035 индуцирует активацию каспазы 3
Полученные до этого результаты авторов изобретения позволяли уверенно предполагать роль каспаз в индуцированном F035 апоптозе. Авторы далее исследовали активацию каспазы 3 в обработанных F035, F094, D1 и G1 клеток, поскольку эта протеаза расположена непосредственно перед PARP, зависимым от времени образом (фиг. 49). Активация начинается через 4 час после обработки во всех случаях, с максимумами через 6-8 час, после чего снижается.
Пример 42
Высвобождение цитохрома c из митохондрий под действием F035
Высвобождение цитохрома c рассматривается как причина активации каспазы 3 в некоторых путях апоптоза. Для изучения того, являлось ли это справедливым в случае индуцированного F035 апоптоза, изобретатели проверили уровень цитохрома c в цитозольных экстрактах обработанных F035 клеток. Авторы обнаружили высвобождение цитохрома c из митохондрий этих клеток зависимым от времени образом (фиг. 50). Авторы наблюдали высвобождение цитохрома c через 4 час после обработки F035, что совпадает с временем начала активации каспазы 3 и расщепления PARP. Для того чтобы выяснить, предшествует ли высвобождение цитохрома c активации каспазы 3, необходимо исследовать высвобождение в более ранние периоды времени.
Пример 43
Аэропонная система выращивания
В свете данных о том, что тритерпеновые соединения согласно изобретению концентрировались в корнях и стручках растений Acacia victoriae, было желательным создание способа увеличения подходящей ткани, из которой соединения могут быть выделены. Для достижения этой цели была разработана аэропонная система выращивания для культивирования корней Acacia victoriae. Аэропонная система представляет собой закрытую систему, в которой корни растения подвешены в воздухе и окутаны туманом полного питательного раствора. Был сделан ящик 8 x 4 x 3,5 футов из 3/4-дюймовых щитов клееной фанеры, связанных друг с другом болтами и выстланных стекловолоконными щитами для получения водонепроницаемого ящика. Верх ящика закрывали двумя щитами (2 x 8 футов) пенистого полистирола с 12 круглыми отверстиями, просверленными по всей длине, хотя в качестве крышки камеры для последующей работы рассматривается новый дизайн, включающий в себя покрытую поливинилхлоридом клетку для домашней птицы, покрытую непрозрачным совместно спрессованным белым на черном полиэтиленом. Для окутывания туманом корней на период 12 секунд каждые 4,5 мин применяли таймер с повторяющейся программой.
Дизайн системы водоснабжения и дренажа для аэропонной камеры представляет собой закрытую систему, сконструированную из 3/4-дюймового поливинилхлорида с шестью полыми коническими форсунками для вращательного распыления спрея. На дне камеры расположен резервуар с 720 л питательного раствора, который распыляют на корни растений снизу с использованием расположенного снаружи насоса. Применяли насос Little Giant 4-MD 3250 RPM, 1/12 hp.
Насос контролируется таймером с повторением Tork с установленными интервалами из 30 секунд распыления каждые 4,5 минут. Температуру контролировали с помощью электронного термометра Taylor с функциями контроля внутренней и наружной температуры с минимумом и максимумом и с ручной записью. Для нагревания питательного раствора в зимние месяцы применяли два пригодных для работы в воде аквариумных нагревателя Visi-therm 300W, которых было достаточно для поддержания активного роста растений без нагревания окружающего воздуха в ненагреваемом и неохлаждаемом сарае в Tucson, Аризона.
Питательный раствор содержал все необходимые элементы, в которых нуждается растение для осуществления его жизненного цикла. Несмотря на тот факт, что для разных растений требуются различающиеся уровни и составы для оптимального роста, общий, единообразно сбалансированный раствор дает удовлетворительные результаты. Композиция раствора дана ниже в таблице 46.
Аэропонный питательный раствор
Нитрат калия
Ca
146
S
134
Затем семена Acacia victoriae надрезали и высевали в несодержащую почву смесь, состоящую из 50% торфяникового мха и 50% вермикулита. Посевы поливали дважды в день и оплодотворяли однократной дозой осмакота. Когда саженцы достигали длины между 15 и 20 см, что обычно происходило через 3-4 месяца выращивания, корневые комья тщательно промывали для удаления всех следов торфяникового мха и вермикулита. Затем корни пропускали через отверстия в щитах из пенистого полистирола, а верхушки саженцев поддерживали бечевками, спускающимися сверху с конструкций теплицы. Вокруг кроны саженцев оборачивали 7,0 см трубчатый фрагмент пенного материала для предотвращения затуманивания листьев и окружающей среды. Затем ящик заполняли приблизительно на 30 см питательным раствором и включали насос.
Когда саженцы принимали правильное положение и опрыскивались, их содержание ограничивалось подтягиванием растущих саженцев вверх бечевками с помощью пластиковых зажимов и пополнением питательного раствора, когда уровень падал ниже 10 см. Во время роста саженцев в теплице температурный контроль питательного раствора не требовался. Однако, если аэропонный ящик оказывается в условиях окружающей среды, рекомендуется повысить температуру питательного раствора до 70°F, с тем, чтобы растения не засыпали во время зимних месяцев.
Для сбора корней корневую массу одного растения промывают водой непосредственно в аэропонном ящике и корневую массу отрезают ножницами несколькими дюймами выше форсунки. Избыток воды удаляют промоканием бумажными полотенцами с последующим взвешиванием образца. Корневую массу затем разрезают ножницами на фрагменты 3-4 дюйма и подвергают химической экстракции, как описано выше. Альтернативно, для повторяющегося сбора корней отключают насос и корни выстригают из растущей корневой массы. Данные корни затем разрезают на фрагменты 3-4 дюйма и экстрагируют, как описано. Предпринимаются предосторожности, чтобы не повредить несобираемые корни.
При выращивании растений в аэропонной системе был реализован ряд преимуществ. Во-первых, рост растений был приблизительно в два раза выше достигаемого с помощью традиционных способов выращивания. Во-вторых, корни могут быть легко собраны в необходимом количестве без повреждения растений. Данное обрезание корней в дальнейшем ведет к обильному латеральному росту волокнистых корней. Поэтому корни можно было собирать по нескольку раз в год. Кроме того, выращенные аэропонным способом растения цвели на первый год выращивания, тогда как растения, выращенные на открытом воздухе, цвели через 3-5 лет.
Пример 44
Тканевое культивирование и проращивание Acacia victoriae
Семена/субстрат. Семена собирали с растений, растущих в городке Сельскохозяйственного центра университета Аризоны, Tucson, Аризона. Семена тщательно промывали в водопроводной воде и с помощью противомикробного мыла (Vionex, Viro Research International Inc., Durango США, Колорадо), после чего обрабатывали имеющимся в продаже отбеливателем 20% (об/об) в течение 15 мин. После повторной промывки в деионизированной воде их обрабатывали в кипящей воде (приблизительно 200 мл на 100 семян) и оставляли остывать в течение ночи. Затем их обрабатывали 20% (об/об) имеющимся в продаже отбеливателем в течение 20 мин, промывали 2-3 раза стерильной деионизированной водой и культивировали на среде MS (Murashige and Skoog, 1962) и разведенной 1/2 MS. Среда содержала витамины MS, 2% (мас/об) сахарозу и была загущена в гель либо с помощью 0,7% агара, либо 0,2% Gelrite. В одном исследовании семена скарифицировали концентрированной серной кислотой, промывали стерильной водой и культивировали на среде. Все среды автоклавировали при 121°С в течение 15 мин. Культуры поддерживали при 25±2°С при фотопериоде с 16-часовым освещением при 1000 люкс, получаемом от холодных белых флуоресцентных трубок. В каждом исследовании имелось 18 дубликатов.
Размножение: кончики побегов и узловые сегменты, отрезанные от трехнедельных сеянцев, культивировали только в среде MS, а также в среде MS с добавкой 0,1 мг/л ауксинов (IAA, NAA или IBA) и BAB (0,1, 0,3, 0,5, 1,0 и 1,3 мг/л) либо раздельно, либо в сочетаниях. В отношении стимуляции корнеобразования ростков тестировали IAA (0,1 мг/л), IBA (0,1 и 0,6 мг/л) и NAA (0,1 и 0,2 мг/л). Для перенесения на почву ростки вынимали из культуральных пробирок, корни промывали водопроводной водой для удаления питательных веществ, прилипших к корням, и переносили в горшочки, наполненные почвой пустынного типа. Растения накрывали коробками Magenta для поддержания влажности и оставляли в тумане и при слабом освещении в течение 3 недель. Через 3 недели коробки Magenta убирали и растения переносили из тумана в теплицу с более сильным освещением.
Индукция наплыва: Ткань наплыва индуцировали из гипокотильного и корневого сегментов, вырезанных из 3-недельных выращенных in vitro сеянцев. Эксплантаты культивировали на среде MS с добавкой 2,4-D (1 мг/л), NAA (0,5 и 1 мг/л), IAA (0,2 и 1 мг/л), тидиазурона (0,2 мг/л), дикамба (0,2 и 2 мг/л), BAP (0,3 мг/мл) и KN (0,5 и 3 мг/л) раздельно или в сочетаниях.
Проращивание семян. Семена, обработанные горячей водой, прорастали с появлением корешков в течение 3-4 дней, а полные ростки получали через 1 неделю. Семена, культивированные без обработки горячей водой, не прорастали. На среде, отвержденной Gelrite (0,2%), процент прорастания семян был выше, чем на агаре (0,7%). Максимальный процент прорастания, 88,7%, наблюдался в разбавленной в два раза среде MS, отвержденной Gelrite. Результаты прорастания на различных средах суммированы в таблице 47.
Прорастание семян Acacia victoriae
Трансплантируемые сеянцы получали через 3-4 недели. Семена A. victoriae обладают низкой скоростью прорастания in vivo из-за глубокого состояния покоя семян (Kaul and Ganguly, 1965; Grice and Westoby, 1987). Для преодоления состояния покоя оболочка семян должна быть или надрезана острым инструментом, подвергнута кислотной скарификации или они должны быть залиты кипящей водой и оставлены остывать в воде в течение ночи. Авторы обнаружили, что процент прорастания может быть увеличен до 88,7% путем применения обработки кипящей водой с последующим культивированием семян на 1/2 среде MS, превращенной в гель с помощью 0,2% Gelrite. Согласно Larsen (1964) прорастание семян A. victoriae в условиях in vivo в результате обработки кипящей водой может увеличиваться до 36%. Без предварительной обработки процент прорастания составлял 19,4% (Kaul and Ganguly, 1965). Кроме того, появление корешков занимало 12 дней, а полные сеянцы появлялись через 79 дней. В протоколе увеличен процент прорастания (88,7%), и трансплантируемые семена могли быть получены в пределах 3-4 недель.
Культуры верхушек ростков. Для исследования размножения отводками макушки ростков (длиной приблизительно 1,0 см) культивировали либо на одной среде MS, либо на среде MS с добавкой BA и BA в сочетании с IAA. На одной среде MS ростки были слабыми и имели плохой рост корней (1-3 корешка на культуру). На среде, содержащей BA (1,3 мг/л), верхушки ростков образовывали множественные ростки (в среднем 3,94 ростка на культуру). Среди множественных ростков один или два ростка удлинялись и достигали роста 8,6 см через 4 недели. Сочетание BA и IAA также способствовало индукции размножения ростков. Ответы суммированы в таблице 48.
Влияние различных уровней BA и IAA (0,2 мг/л) на индукцию многочисленных ростков Acacia victoriae
BA (мг/л)
IAA (мг/л)
При более высоких концентрациях BA (1,0 и 1,3 мг/л) количество ростков увеличивалось. Было обнаружено, что сочетание BA (1 мг/л) + IAA (0,2 мг/л) дает лучшие результаты по размножению ростков. Наплыв обнаруживался на обрезанных концах при всех сочетаниях BA-IAA. Kaur et al. (1998) сообщали о синергичном действии BA-NAA на индукцию почек ростков Acacia catechu, причем более высокие уровни NAA (1-2 мг/л) не давали преимуществ. Они также утверждали, что IAA была неэффективна для увеличения образования почек ростков, однако из основания эксплантатов образовывался наплыв.
Для исследования корнеобразования полученные in vitro ростки извлекали и переносили на среду, содержащую IAA, NAA или IBA. Ответы суммированы в таблице 49. Было обнаружено, что из тестированных обработок лучшей для формирования корней была 1/2 MS + NAA (0,2 мг/л). Почти 100% ростков давали корни. Ростки достигали высоты 9-11 см в течение 4 недель. Сообщалось (Kaur et al., 1998) об образовании у Acacia catechu промежуточного наплыва в месте соединения корня и ростка, и эти авторы применяли снижение уровня сахарозы с 3 до 1,5% для контролирования наплыва. Сходные результаты сообщались также для Feronia limonia (Purohit and Tak, 1992) и Acacia auriculiformis (Das et al., 1993). В настоящем исследовании слабое образование наплывов также отмечали в 3% сахарозе, и оно сводилось к минимуму при 2% сахарозы. Ростки с корнями переносили в теплицу. Выживаемость после перенесения составляла 100%. Проростки акклиматизировали при отуманивании в течение 3 недель, после чего проростки выращивали в обычной теплице.
Влияние IAA, NA и IBA на формирование корней у ростков Acacia victoriae
Культуры узловых сегментов. Узловые сегменты (котиледонные узлы), вырезанные из проращенных in vitro сеянцев, культивировали на среде MS с добавкой 0,1 мг/л IAA, NAA или IBA. Развивались лишь один или два пазушных ростков на эксплантат. Однако рост данных ростков был медленным. Поэтому узловые эксплантаты не применяли для дальнейших исследований.
Индукция наплыва из гипокотиля и корневых сегментов. Наплыв индуцировали из гипокотильных сегментов, вырезанных из 3-недельных проращенных in vitro сеянцев. Наплыв, образовавшийся на 2,4-D (1 мг/л), Thidiazuron (0,2 мг/л), Dicamba (0,2 мг/л), был зеленоватым, компактным и твердым. Количество образовавшихся наплывов было умеренным при большинстве испробованных концентраций (таблица 50). Изобильное образование наплывов отмечалось в среде MS с добавкой 2,4-D (4 мг/л) + IAA (1 мг/мл) + NAA (1 мг/л).
Сегменты корней, вырезанные из трехнедельных проращенных in vitro сеянцев и культивированные на среде MS с добавкой только 2,4-D (1 мг/л) и 2,4-D в сочетании с KN (0,5 мг/мл), показали развитие светло-желтоватых мягких наплывов с формированием нескольких корней из наплыва. Формирование наплывов отмечалось в 100% культур. На среде с добавкой BA (0,3 мг/л) + IAA (0,2 мг/л) отмечалось образование из корневых сегментов беловатых, мягких, ломких и обильных наплывов. Формирование светло-желтоватых обильных наплывов отмечалось на корневых сегментах, культивированных на среде с добавкой 2,4-D (4 мг/л) в сочетании с 1 мг/л каждого из IAA или NAA. Сходный тип формирования наплывов отмечался при применении Thidiazuron (0,2 мг/л) + Dicamba (2 мг/л) и IAA (0,1 мг/л). Сегменты корней, культивированные на среде с Dicamba (2 мг/л) + IAA (0,1 мг/л), образовывали светло-зеленые компактные твердые наплывы. Попытки регенерировать проростки из наплывов были безуспешными. Для нескольких древесных видов, таких как Albiizzia lebbeck (Lakshmana Rao and De, 1989) и Lonicera japonica (Georges et al., 1993), сообщалось о различиях между типами эксплантатов в отношении индукции наплывов. В исследованиях авторов изобретения также было обнаружено, что имеется различие между наплывами, образованными из гипокотиля и корней, развивавшихся на одинаковой среде. Наплывы, образовавшиеся из гипокотиля на сочетаниях BA-IAA, были светло-зеленоватыми, твердыми и компактными, тогда как образовавшиеся из сегментов корней были беловатыми, мягкими, ломкими, и также проявляли корневую дифференцировку из наплыва при некоторых сочетаниях. Наплыв Dalbergia latifolia на среде регенерации становился компактным, твердым и темно-зеленым, и происходила дифференцировка ростковых почек (Pradhan et al., 1998). В исследованиях изобретателей отмечался сходный тип развития наплыва, но такой наплыв был неспособен к регенерации. В данном исследовании авторы показали, что A. victoriae может быть размножена in vitro из верхушек ростков. Стандартизированный способ пригоден для микроразмножения элитных особей, обнаруженных среди неоднородных популяций сеянцев, и сохранения элитных линий для последующих исследований.
Развитие наплывов из гипокотиля и корневых сегментов Acacia victoriae
Пример 45
Индукция волосистых корней Acacia victoriae для получения противораковых соединений
Инфицирование растительного материала Acacia victoriae Agrobacterium rhizogenes ведет к интеграции и экспрессии T-ДНК генома растения, что вызывает развитие фенотипа волосистых корней (Grant et al., 1991). Культуры волосистых корней растут быстро, проявляют плагиотропный рост корней и сильно ветвятся в свободной от гормонов среде. Волосистые корни также проявляют высокую степень генетической стабильности (Aird et al., 1988). Многие двудольные растения восприимчивы к A. rhizogenes, и растения многих видов регенерировали из культур волосистых корней (Christey, 1997).
Генетическая трансформация и индукция волосистых корней производились изобретателями в качестве способа получения активных тритерпенов из A. victoriae. Для получения культур волосистых корней используется естественная способность почвенной бактерии Agrobacterium rhizogenes трансформировать гены в геном растения-хозяина в корнях, образованных на инфицированной стороне. Волосистые корни характеризуются многочисленными быстрорастущими, сильно ветвящимися добавочными корнями, которые продолжают расти на свободной от гормонов среде.
Авторы изобретения продемонстрировали индукцию волосистых корней у Acacia victoriae с помощью штамма R 1000 Agrobacterium rhizogenes (сконструированного из штамма Agrobacterium tumefaciens, в который вводили плазмиду pRiA4 Agrobacterium rhizogenes, ATCC номер 43056). Продукция в волосистых корнях интересующего соединения была подтверждена с помощью ВЭЖХ. Индукцию волосистых корней производили следующим образом. Сначала собирали семена Acacia victoriae с выращенных на поле растений в Tucson, Аризона. Семена заливали кипящей водой, им давали набухать в течение ночи при остывании воды, и их поверхность стерилизовали в 15% имеющемся в продаже отбеливателе в течение 30 минут. После повторной промывки в стерильной воде семена культивировали на жидкой среде MS (Murashige and Skoog, 1962) с добавкой витаминов MS и 2% сахарозы в 250-мл конических сосудах с 50 мл среды. Культуры поддерживали на вращательном шейкере в помещении для выращивания при 25 ± 2°С в темноте. Через четыре дня культивирования эмбриональные стволы вырезали из прорастающих сеянцев и применяли для агроинфицирования.
Перед агроинфицированием штамм R 1000 Agrobacterium rhizogenes выращивали в течение ночи на среде YENB. Среду YENB готовили добавлением 7,5 г/л дрожжевого экстракта и 8 г/л питательного бульона (Difco Laboratories, Detroit, MI). Эмбриональный ствол эксплантата инфицировали с помощью тонкой иглы из нержавеющей стали, которую погружали в бактериальный раствор. После инфицирования каплю бактериальной суспензии (1:20 в среде MS) наносили на поверхность эксплантатов. Затем эксплантаты переносили на среду MS и среду MS с ацетосирингоном (100 мкМ) (3,5-диметокси-4-гидроксиацетофенон, Aldrich Chem. Co, Milwaukee, WI) для совместного культивирования. Совместное культивирование проводили в течение трех дней в темноте. Через три дня совместного культивирования эксплантаты переносили на среду MS + цефотаксим (500 мг/л, Agri-Bio, North Miami, Fl) для контроля чрезмерно быстрого роста бактерий. Инициация корней отмечалась на стороне инфекции, главным образом из молодых развивающихся листьев от эмбрионального ствола через 3-4 недели. После 4 недель эксплантаты вместе с корнями переносили на среду MS без добавок и инкубацию в темноте продолжали для развития волосистых корней. Развитие волосистых корней отмечалось спустя еще 8 недель. Развившиеся таким образом волосистые корни стандартно размножали в среде MS путем субкультивирования. Трансгенная природа волосистых корней была подтверждена с помощью PCRTM с применением набора праймеров для амплификации части гена rol B. Были применены следующие праймеры:
50 мкл реакционной смеси PCRTM содержали праймеры (конечная концентрация каждого 1 мкМ), Taq полимеразу (1,0 Ед.), 125 мкМ каждого из dNPT, 1 X PCRTM буфер для реакции, 1,5 мМ MgCl2, 300 нг выделенной ДНК. Примененными условиями PCRTM были исходная денатурация при 92°С в течение 5 мин с последующими 35 циклами 92°С 50 секунд, 55°С 1 мин для отжига, 72°С 1 и 1/2 мин для наращивания и 72°С 7 мин окончательного наращивания. Был амплифицирован фрагмент 645 п.н.
Культуры волосистых корней в жидкой среде. Для оптимизации условий роста волосистые корни, растущие на полутвердой среде MS, вырезали и культивировали в жидкой среде MS в сосудах разной емкости (125, 250, 500 и 1000 мл) с 20, 50, 100 и 400 мл среды соответственно. Исходный инокулят волосистых корней составлял 6 г/л. Рост волосистых корней также тестировали в следующих базальных средах: MS, Nitsch and Nitsch (N and N) (1969), Schnek and Hilderbrandt (SH) (1972) и Woody Plant Medium (WPM) (Lloyd and McCown, 1981). Для тестирования влияния различных источников углерода на рост волосистых корней к среде MS добавляли по 2% (мас/об) каждого из сахарозы, глюкозы, фруктозы и маннозы. Влияние гиббереллиновой кислоты (0,1, 0,5 и 1 мг/л) на рост волосистых корней тестировали добавлением стерилизованного на фильтре раствора к среде MS после автоклавирования.
Инициация корней на месте инфицирования отмечалась через 3-4 недели. Четыре независимо трансформированных клона волосистых корней были выведены из эмбриональных стволов, инфицированных штаммом R1000 в присутствии ацетосирингона (100 мкМ). В эмбриональных стволах, совместно культивированных с A. rhizogenes без ацетосирингона, не развивались волосистые корни. При совместном культивировании в течение 3 дней в присутствии ацетосирингона был найден оптимум для индукции волосистых корней. Стимулирующее действие ацетосирингона сообщалось для Salvia militiorrhiza (Hu and Alfermann, 1993). Результаты показали, что ацетосирингон, активатор генов vir Agrobacterium, повышает частоту трансформации. Сходным образом в настоящем исследовании ацетосирингон требовался для индукции волосистых корней.
Трансформированная природа корней была подтверждена с помощью амплификации PCRTM с применением набора праймеров для амплификации части гена rol B. Диагностический фрагмент из 645 п.н. был амплифицирован в четырех тестированных клонах волосистых корней.
Волосистые корни, росшие на жидкой среде, развивались энергично. Было обнаружено, что из тестированных различных базальных сред среда MS является лучшей для роста волосистых корней. В 125-мл сосуде происходил 268-кратный прирост за 4 недели. Со средами WPM и N and N происходило 254- и 196-кратное увеличение, соответственно. B5 и среда SH не способствовали оптимальному росту волосистых корней. На данных двух средах волосистые корни начинали медленно коричневеть. В одном исследовании волосистые корни выращивали в сосудах различной емкости (125, 250, 500 и 1000 мл) с 20, 50, 100 и 400 мл среды MS, соответственно. Кинетика роста суммирована в таблице 53. Исходно рост волосистых корней является энергичным, и достигается 25,77-кратное увеличение за 4 недели в 125-мл сосудах, исходно инокулированных 150 мг. С увеличением объема сосудов рост корней слегка снижался.
Рост волосистых корней может быть чувствителен к составу среды, особенно минеральным ионам и источнику углерода (Wysokinska and Chmiel, 1997). Для Acacia victoriae было тестировано пять различных базальных сред (MS, N and N, SH, WPM и B5) в отношении из влияния на рост волосистых корней. Было обнаружено, что среда MS является лучшей для роста. Sasaki et al. (1998) сравнивали рост волосистых корней Coleus forskohlii на различных питательных средах и обнаружили, что для роста волосистых корней лучшей средой является WPM.
В данном исследовании сахароза способствовала росту волосистых корней по сравнению с другими источниками углерода (фруктозой, глюкозой и маннозой). Максимальный рост (24,52-кратное увеличение) был обнаружен в содержащей сахарозу среде. Содержащая глюкозу среда оказывала слабое ингибирующее действие на рост, а манноза полностью подавляла рост (таблица 54). У Catharanthus roseus продукция катарантина могла быть удвоена применением фруктозы в качестве источника углерода в среде. Однако авторы сообщили, что применение фруктозы приводит приблизительно к 40% снижению роста по сравнению с сахарозой (Jung et al., 1992).
Для продолжительного роста волосистых корней не требуются экзогенные регуляторы роста, поскольку гены, которые повышают чувствительность к ауксину, имеются в плазмиде Ri (Wysokinska and Chmiel, 1997). Однако имеются сообщения, в которых экзогенные гормоны стимулируют рост. Авторы изобретения проверили влияние гиббереллиновой кислоты (0,1, 0,5 и 1,0 мг/л) на рост волосистых корней. Рост волосистых корней был лучшим в среде без GA3 по сравнению с содержащей GA3 средой (15,77-кратное увеличение). Различные уровни GA3 не оказывали существенного влияния на рост (таблица 55). У Artemisia GA3 не повышала накопление общей биомассы, но способствовала более быстрому достижению стационарной фазы по сравнению с культурами, росшими на среде без GA3 (Smith et al., 1997). Rhodes et al., 1994) обнаружили, что ответ волосистых корней Brassica candida на GA3 зависел главным образом от исследуемого клона. Однако они наблюдали, что в целом GA3 оказывала положительное влияние на рост и снижение аккумуляции алкалоидов, сопровождаемые изменениями в характере продукции. Ohkawa et al., (1989) сообщали о том, что GA3 в концентрациях 10 нг/л и 1 мг/л ускоряет рост, увеличивает элонгацию и повышает боковое ветвление волосистых корней Datura innoxia. Zobel (1989) предположил, что GA3 действует как аналог CO2 в отношении роста корней. В случае волосистых корней Acacia victoriae GA3 не повышала рост, что могло бы указывать на различия в ответах разных генотипов.
Применение культур волосистых корней Acacia victoriae должно служить в качестве подходящего источника однородной культуры растительной ткани, из которой могут быть выделены композиции тритерпеновых гликозидов данного изобретения, которые включают в себя выделенные смеси или отдельные очищенные соединения.
Инфицирование штаммом R1000 Agrobacterium rhizogenes эмбриональных стволов Acacia victoriae для получения волосистых корней
MS+ацето.
21
-
-
MS+ацето.
38
9(23)
4(17,3)
a Числа в скобках указывают процент
Влияние различных размеров сосудов на рост волосистых корней Acacia victoriae
Влияние различных базальных сред и размера сосудов на рост волосистых корней Acacia victoriae
Влияние различных источников углерода в среде MS на рост волосистых корней Acacia victoriae
a2% (мас/об). bИсходная свеж. мас. для каждой обработки составляла 300 мг. cДанные представляют среднее для 6 повторов ± ст. ош.
Влияние GA3 на рост волосистых корней Acacia victoriae
В отношении роста волосистых корней тестировали различные среды. Лучший рост был получен на среде MS, содержащей 2% сахарозу. Тестировали влияние сосудов с разной емкостью и гиббереллиновой кислоты на рост волосистых корней. Волосистые корни также выращивали на жидкой среде MS на вращающем шейкере в 125-мл коническом сосуде с 20 мл среды. Через 4 недели было отмечено 84-кратное увеличение роста. Продукция тритерпеновых сапонинов, соответствующих идентифицированным в F035, была подтверждена с помощью анализа ВЭЖХ со стандартным образцом сравнения.
Пример 46
Монотерпеновые композиции ингибируют активацию NF-κB путем ингибирования его ядерной транслокации и способности связываться с ДНК
Анализировали влияние F035 и монотерпенового гликозида G1 на индуцированную фактором некроза опухолей (TNF) активацию NF-κB в клетках Jurkat для выявления роли ядерного транскрипционного фактора каппа B (NF-κB) в индуцированном монотерпеновым гликозидом ингибировании канцерогенеза. NF-κB является одним из главных регуляторов воспаления и контролирует транскрипцию нескольких генов, участвующих в воспалении.
Было обнаружено, что вызываемое свободными радикалами повреждение ДНК снижается в ответ на обработку F035, что позволяет предполагать, что F035 обладает антиоксидантной функцией. Данное наблюдение коррелирует с более ранним исследованием, показавшим, что уровень активных форм кислорода (АФК) снижается в митохондриях клеток, обработанных монотерпеновым гликозидом. Повышение уровней АФК является одним из факторов, приводящих к активации NF-κB. Поэтому один аспект изобретения касается защиты митохондрий от окислительного стресса путем введения соединений согласно изобретению в нуждающиеся в этом клетки.
Способы
Клеточная культура. Клетки Jurkat и RAW 264.7 выращивали на среде RPMI-1640 с добавкой 10% сыворотки плодов телят, 2 мМ глутамина и 0,05% гентамицина.
Обработка клеток монотерпеновыми гликозидами. Клетки Jurkat брали в концентрации 1 x 106/мл в полной среде и обрабатывали 2 мкг/мл F094, монотерпенового/тритерпенового гликозида D1 или монотерпенового/тритерпенового гликозида G1 в течение 8-16 час при 37°С. В конце обработки клетки промывали полной средой и подсчитывали. В различных экспериментах применяли равное количество жизнеспособных клеток.
Получение цитоплазматических и ядерных экстрактов. Клетки Jurkat (2 x 106/мл), обработанные F094 или выделенными монотерпеновыми/тритерпеновыми гликозидами G1 и/или D1, экспонировали с TNF (1 нМ в течение 15 мин) при 37°С. После промывки клеток в холодном ЗФР их суспендировали в 0,4 мл буфера для цитоплазматической экстракции (10 мМ HEPES, pH 7,9, 10 мМ KCl, 0,1 мМ ЭДТА, 0,1 мМ ЭГТА, 1 мМ DTT, 0,5 мМ фенилметилсульфонилфторид (PMSF), 2 мкг/мл лейпептин, 2 мкг/мл апротинин, 0,5 мг/мл бензамидин) в течение 15 мин на льду. Клетки окончательно лизировали путем добавления к клеткам 12,5 мкл 10% нонидета P-40 (NP-40) и цитоплазматические экстракты собирали центрифугированием. Затем к ядерному осадку добавляли 25 мкл холодного буфера для экстракции ядер (20 мМ HEPES, pH 7,9, 0,4 М NaCl, 1 мМ ЭДТА, 1 мМ ЭГТА, 1 мМ DTT, 1 мМ PMSF, 2 мкг/мл лейпептин, 2 мкг/мл апротинин, 0,5 мг/мл бензамидин) и инкубировали на льду в течение 30 мин. Ядерные экстракты собирали путем центрифугирования при 4°С в течение 5 мин.
Анализ сдвига электрофоретической подвижности (EMSA). 4 мкг ядерного экстракта инкубировали с 16 фмоль меченного 32P по концу 45-членного двухцепочечного олигонуклеотида NF-κB из длинного концевого повтора ВИЧ:

(подчеркиванием указаны сайты связывания NF-κB) в течение 15 мин при 37°С. Инкубационная смесь включала 3 мкг поли (dI:dC) в буфере для связывания (25 мМ HEPES, pH 7,9, 0,5 мМ ЭДТА, 0,5 мМ DTT, 1% NP-40, 5% глицерин, 50 мМ NaCl). ДНК-белковый комплекс отделяли от свободного олигонуклеотида на 7,5% полиакриламидном геле. Специфичность связывания NF-kB-ДНК проверяли путем конкуренции двухцепочечного мутантного олигонуклеотида, немеченого олигонуклеотида и с помощью суперсдвига полосы под действием антитела против p65. Радиоактивные полосы высушенных гелей визуализировали с помощью PhosphoImager (molecular Dynamics, Sunnyvale, CA) и количественно измеряли с помощью программы ImageQuant.
Анализ методом иммуноблоттинга. Цитоплазматические и ядерные экстракты клеток Jurkat, обработанных монотерпеновым/тритерпеновым гликозидом G1, применяли для исследования деградации IκB и ядерной транслокации субъединицы p65 NF-κB, соответственно. Для этого 50 мкг цитоплазматического или ядерного белка разделяли на 9% ДДС-Na-полиакриламидном геле и переносили электропереносом на нитроцеллюлозную мембрану. Мембраны зондировали антителом кролика против IκBα или p65 (Santa Cruz, CA) и затем антикроличьим антителом, конъюгированным с пероксидазой хрена (HRPO). Белковые полосы выявляли с помощью хемилюминесценции (ECL, Amersham).
Трансфекция и тестирование люциферазной активности. Клетки Jurkat трансфицировали pGL3-NF-κB с помощью электропорации. Затем клетки обрабатывали монотерпеновым/тритерпеновым гликозидом G1 (1 мкг/мл) в течение 16 час. NF-κB активировали с помощью LPS (100 нг/мл), PMA (5 нг/мл) или TNF (1 нМ). Люциферазную активность измеряли с применением набора для тестирования люциферазы (Promega, Madison, WI) в соответствии с указаниями производителя.
Индукция и измерение iNOS и COX-2. Клетки RAW 264.7 (0,5 x 106/мл) помещали на 100-мм чашки и обрабатывали F094 или монотерпеновыми/тритерпеновыми гликозидами (2 мкг/мл) в течение 16 час. Затем клетки экспонировали с 100 нг/мл LPS в течение 24 часов. Клетки лизировали в буфере, содержащем 50 мМ трис, pH 7,4, 100 мМ NaCl, 0,5% NP-40, 10 мкг/мл лейпептин, 5 мкг/мл апротинин и 100 мкМ PMSF. Затем 70 мкг клеточного белка наносили на 7,5% ДДС-Na-полиакриламидный гель и переносили электропереносом на нитроцеллюлозную мембрану. Уровни iNOS и COX-2 анализировали с помощью теста иммуноблоттинга с применением кроличьих антител против iNOS (Santa Cruz) и козьих антител против COX-2, соответственно.
Результаты
Смесь монотерпеновых гликозидов, таких как тритерпеноидные сапонины и тритерпеновый гликозид G1, ингибирует индуцированный TNF NF-κB зависимым от времени и дозы образом. С помощью анализа сдвига электрофоретической подвижности было продемонстрировано зависимое от времени действие F035 и монотерпенового/тритерпенового гликозида G1 на индуцированный TNF NF-κB в клетках Jurkat, как изображено на фиг. 51A. В обоих случаях максимальное ингибирование наблюдалось через 16 час после обработки. Обработка клеток только F035 или монотерпеновым/тритерпеновым гликозидом G1 в течение различных периодов времени не влияла на базальный уровень NF-κB в данных клетках (фиг. 51A). В предшествующих примерах показано, что различные процессы, ведущие к апоптозу, индуцированному смесью или чистыми монотерпеновыми/тритерпеноидными сапонинами, инициируются уже между 30 мин и 2 час после обработки. Для проверки того, что апоптоз клеток не был ответственен за наблюдаемое снижение активации NF-κB, клетки подсчитывали в конце обработки, и для исследования индуцированной TNF активации NF-κB брали равное количество жизнеспособных клеток. Кроме того, обработка клеток zVAD-fmk проникающим во многие клетки необратимым ингибитором каспаз не влияла на ингибирование уровня активированного NF-κB. Следовательно, данное ингибирование не зависело от активности каспаз. Было обнаружено, что монотерпеновый/тритерпеновый гликозид G1 является более эффективным, чем смесь монотерпеновых/тритерпеноидных сапонинов.
16-часовая обработка монотерпеновым/тритерпеновым гликозидом G1 ингибировала индуцированную TNF активацию NF-κB зависимым от дозы образом. Клетки Jurkat, обработанные 0,5 мкг/мл монотерпеновым/тритерпеновым гликозидом G1 в течение 16 час, показали почти 50% снижение индуцированного TNF NF-κB, а клетки Jurkat, обработанные 2,0 мкг/мл, показали почти полное ингибирование (фиг. 52A). Монотерпеновый/тритерпеновый гликозид G1 сам по себе не оказывал влияния на конститутивный уровень NF-κB в данных клетках (фиг. 52A). За задерживаемую полосу мог конкурировать немеченый олигонуклеотид, и эта полоса подвергалась суперсдвигу под действием антитела против p65; оба этих факта демонстрируют специфичность полосы NF-κB (фиг. 51B).
Монотерпеновый/тритерпеновый гликозид G1 не ингибирует деградацию IκB, но ингибирует ядерную транслокацию субъединицы p65 NF-κB. Активация NF-κB включает в себя две важные стадии: (1) отделение ингибиторной субъединицы IκB и (2) ядерную транслокацию активированного NF-κB. Для выяснения влияния монотерпенового/тритерпенового гликозида G1 на любую из данных стадий или на обе необработанные и обработанные монотерпеновым/тритерпеновым гликозидом G1 клетки Jurkat экспонировали с TNF в течение различных периодов времени. Кинетику деградации IκB исследовали с помощью иммуноблоттинга цитоплазматических экстрактов. Как показано на фиг. 53A, не наблюдалось разницы в характере деградации IκB после обработки монотерпеновым/тритерпеновым гликозидом G1. Вид субъединицы p65 NF-κB в ядерных экстрактах необработанных и обработанных монотерпеновым/тритерпеновым гликозидом G1 клеток (фиг 53B) демонстрирует, что обработка монотерпеновым/тритерпеновым гликозидом G1 значительно замедляет ядерную транслокацию субъединицы p65. Денситометрический анализ полос подтверждает, что количество транслоцированного в ядро p65 снижается после обработки монотерпеновым/тритерпеновым гликозидом G1.
Монотерпеновый/тритерпеновый гликозид G ингибирует связывание NF-κB-ДНК. Поскольку монотерпеновый/тритерпеновый гликозид G1 не полностью блокировал ядерную транслокацию субъединицы p65 NF-κB, авторы изобретения допустили, что монотерпеновый/тритерпеновый гликозид G1 может влиять на активацию NF-κB более чем одним путем. Поэтому авторы изобретения провели анализ действия монотерпенового/тритерпенового гликозида G1 на способность активного NF-κB связываться с ДНК в бесклеточной системе. Ядерные экстракты из стимулированных TNF клеток инкубировали с возрастающими концентрациями монотерпенового/тритерпенового гликозида G1. Монотерпеновый/тритерпеновый гликозид G1 ингибировал связывание NF-κB с ДНК зависимым от дозы образом (фиг. 54A). Известно, что дезоксихолат (DOC) вызывает диссоциацию NF-κB от IκB, тем самым превращая его в активную ядерную форму. После обработки DOC (0,8%) цитоплазматические экстракты клеток, обработанных монотерпеновым/тритерпеновым гликозидом G1, показали сниженное связывание с ДНК по сравнению с экстрактами необработанных клеток (фиг. 54B). Поскольку деградация IκB не затронута, данные результаты показывают, что монотерпеновый/тритерпеновый гликозид G1 модифицирует NF-κB таким образом, что NF-κB не связывается с ДНК.
Клетки Jurkat инкубировали с 100 мкМ DTT в течение 2 час до их обработки монотерпеновым/тритерпеновым гликозидом G (2 мкг/мл в течение 8 час) для определения того, включает ли действие монотерпенового/тритерпенового гликозида G1 необратимое алкилирование свободных сульфгидрильных групп остатков цистеина. После этого клетки экспонировали с TNF, как описано выше. Как показано на фиг. 54C, DTT не модифицировал индуцированную TNF активацию NF-κB; напротив, он снимал действие монотерпенового/тритерпенового гликозида G1. Данные результаты показывают, что монотерпеновый/тритерпеновый гликозид G1 осуществляет свое действие, влияя на важные сульфгидрильные группы, необходимые для индуцированной TNF активации NF-κB.
Монотерпеновый/тритерпеновый гликозид G1 ингибирует зависимую от NF-κB экспрессию генов. Для определения влияния монотерпенового/тритерпенового гликозида G на зависимую от NF-κB экспрессию генов применяли репортерный ген люциферазы, pGL3, соединенный с элементом NF-κB из промотора ИЛ-2. Клетки Jurkat, временно трансфицированные pGL3-NF-κB, обрабатывали LPS, PMA или TNF для индукции люциферазной активности. Предварительная обработка трансфицированных клеток F094 или монотерпеновым/тритерпеновым гликозидом G1 значительно ингибировала люциферазную активность, индуцированную другими агентами (фиг. 55A).
Анализировали также влияние F094 и монотерпенового/тритерпенового гликозида G1 на индукцию iNOS и COX-2, экспрессия которых регулируется NF-κB. В ответ на обработку LPS в клетках RAW264.7 индуцировался значительный уровень iNOS и COX-2. Предварительная обработка клеток F094 или монотерпеновым/тритерпеновым гликозидом G1 резко снижает уровень индуцированных LPS iNOS и COX-2 (фиг. 55B).
Обсуждение
Одной из главных проблем профилактики рака является создание эффективных новых лекарств, которые обладают слабым действием или не оказывают действия на нормальные клетки и ткани. Канцерогенез представляет собой многоступенчатый процесс, и воспаление является его составной частью (Sporn et al., 1986; Ohshima et al., 1994). Механизмы воспаления, относящиеся к канцерогенезу, широко изучаются, и предпринимаются попытки использовать данные механизмы в качестве основы для создания новых химиотерапевтических агентов. О противовоспалительных эффектах тритерпеноидов сообщалось ранее (Singh et al., 1992; D'arcy et al., 1957, и Kim et al.). Поскольку исследования канцерогенеза кожи у мышей (описанные в примере 11) показывают, что F035 является противовоспалительным агентом, были проанализированы эффекты монотерпеновых/тритерпеновых гликозидов на NF-κB, который служит регулятором воспаления.
Здесь показано, что и смесь, и чистые монотерпеновые/тритерпеновые гликозиды являются эффективными ингибиторами индуцированного TNF NF-κB. Обработка клеток Jurkat монотерпеновыми/тритерпеновыми гликозидами приводит к значительно более медленной транслокации субъединицы p65 NF-κB в ядро, тогда как деградация IκB остается без изменений. Монотерпеновые/тритерпеновые гликозиды также нарушают связывание NF-κB с ДНК, как это показано с помощью теста связывания in vitro. В обработанных монотерпеновым/тритерпеновым гликозидом G1 клетках уровень индуцируемого дезоксихолатом (DOC) NF-κB в цитоплазме снижен. Обработка клеток дитиотрейтолом (DTT) полностью снимает индуцированное монотерпеновым/тритерпеновым гликозидом G1 ингибирование активности NF-κB, что указывает на то, что затрагиваются сульфгидрильные группы, важные для активации NF-κB. Исследование экспрессии некоторых генов, регулируемых NF-κB в клетках Jurkat, показывает, что обработка монотерпеновым/тритерпеновым гликозидом G1 резко снижает уровень люциферазной активности, а также индуцированной LPS синтазы оксида азота (iNOS) и циклооксигеназы (COX)-2. Способность эффективно ингибировать активацию NF-κB, а также способность снижать экспрессию генов, участвующих в путях воспаления, которые контролируются NF-κB, объясняют противовоспалительные эффекты монотерпеновых/тритерпеновых гликозидов in vivo.
NF-κB играет важнейшую роль в индуцибельной экспрессии нескольких генов, участвующих в иммунных ответах, воспалении и инфекции, таких как гены интерлейкина-1 (ИЛ-1), интерлейкина-6 (ИЛ-6) и молекул адгезии (Baeuerle P.A. et al., 1997a; 1997b; 1994). Было обнаружено, что все монотерпеновые/тритерпеновые гликозиды F035, F094, монотерпеновый/тритерпеновый гликозид D1, монотерпеновый/тритерпеновый гликозид G1 являются сильными ингибиторами индуцированной TNF активации NF-κB в клетках Jurkat. Ингибирование NF-κB монотерпеновым/тритерпеновым гликозидом G1 было зависимым от дозы и времени. Широкий спектр эффектов NF-κB находится под регуляторным влиянием сложной сети ингибиторов и коактиваторов. Для активации NF-κB воспалительными цитокинами, митогенами, бактериальными продуктами или окислительным стрессом необходима деградация IκBα, которая удерживает NF-κB в цитоплазме в дремлющем комплексированном состоянии. После деградации IκBα NF-κB высвобождается из комплекса для транслокации в ядро, где он связывается с ДНК и активирует различные гены. Авторы данного изобретения показывают, что в то время как монотерпеновые/тритерпеновые гликозиды согласно изобретению не влияют на деградацию IκBα, они ингибируют транслокацию NF-κB в ядро. Исследования связывания с ДНК in vitro показывают, что монотерпеновые/тритерпеновые гликозиды согласно изобретению также подавляют связывание NF-κB с ДНК. Более того, монотерпеновый/тритерпеновый гликозид G1 ингибировал связывание с ДНК индуцированного дезоксихолатом NF-κB in vitro. Данное наблюдение указывает на то, что либо NF-κB не диссоциирует из комплекса, либо его ДНК-связывающие свойства модифицируются. Зависимое от монотерпеновых/тритерпеновых гликозидов ингибирование NF-κB снимается DTT, что указывает на то, что монотерпеновые/тритерпеновые гликозиды модифицируют одну или более сульфгидрильных групп, важных для активации NF-κB.
Среди нескольких регулируемых NF-κB белков, играющих роль в воспалении, находятся iNOS и COX-2, которые активно изучались. Оба эти фермента индуцируются в ответ на различные цитокины (например, интерферон-γ), митогены или продукты микробов (например, липополисахарид). Они формируют необходимые компоненты воспалительного ответа, заживления раны и канцерогенеза (Moncada et al., 1991; Anggard et al., 1994; Seibert et al., 1994). COX-2 играет роль в росте некоторых раковых клеток ободочной кишки из-за ее способности действовать в качестве опухолевого промотора посредством стимуляции ангиогенеза. Было показано, что избыточная экспрессия iNOS или COX-2 участвует в патогенезе ряда хронических заболеваний, таких как рак ободочной кишки (Oshima et al., 1994, и Takahashi et al., 1997), рассеянный склероз (Hooper et al., 1997), болезнь Паркинсона (Hantraye et al., 1996) и болезнь Альцгеймера (Goodwin et al., 1995). Ведутся активные поиски агентов, которые бы избирательно ингибировали индуцируемые формы данных ферментов и не влияли бы на их конститутивные формы. Как показано здесь, обработка клеток RAW 264.7 монотерпеновым/тритерпеновым гликозидом G1 ведет к резкому снижению уровня индуцированных LPS iNOS и COX-2.
Таким образом, авторы демонстрируют здесь, что монотерпеновые/тритерпеновые гликозиды являются противовоспалительными агентами. В ответ на провоспалительные агенты, такие как TNF или LPS, они значительно подавляют активацию NF-κB и экспрессию iNOS и COX-2, которые являются важными участниками процесса воспаления. Данная ингибиторная активность в сочетании с их способностью индуцировать апоптоз опухолевых клеток обеспечивает терапевтические агенты, включая монотерпеновые/тритерпеновые гликозиды согласно изобретению, для таких заболеваний как рак и/или другие хронические заболевания с воспалительными компонентами.
Пример 47
Монотерпеновые/тритерпеновые гликозиды индуцируют апоптоз путем перестройки митохондрий
Здесь описаны очистка двух чистых компонентов из смеси монотерпеновых/тритерпеновых гликозидов, которые содержат основу из акациевой кислоты с двумя ациклическими монотерпеновыми единицами, соединенными хиновозным сахаром, очистка и структура двух монотерпеновых/тритерпеновых сапонинов из F094, которые были названы авицин D и авицин G (тритерпеноидные сапонины Acacia victoriae). Анализ механизмов, лежащих в основе подавления роста опухолевых клеток данными агентами показывает, что первичным объектом проапоптотической функции авицинов являются митохондрии.
Способы
Очистка авицинов: круглые семенные стручки Acacia victoriae экстрагировали 20% MeOH при 60°С. Распределение в системе растворитель/растворитель концентрировало биоактивность в полярной фракции (F094). Анализ ВЭЖХ данной фракции (фиг. 56A) на колонке C-18 с обращенной фазой MetaChem Intersil (3 мкм, 4,6 x 150 мм) с применением программы элюирования градиентом ацетонитрила и подкисленной воды указывал на очень сложную смесь соединений. Первоначальное субфракционирование экстракта на полупрепаративной колонке ВЭЖХ C-18 с обращенной фазой и последующее биологическое тестирование указали области вокруг монотерпеновых/тритерпеновых гликозидов D1 и G1, содержащие наибольшую активность. Выделение данных соединений было достигнуто с помощью двухступенчатой препаративной ВЭЖХ с применением колонки пентафторфенила (50 x 250 мм, 10 мкм, ES Industries) и с использованием системы растворителей водного метанола.
Клеточная культура. Клетки Jurkat выращивали в среде RPMI-1640 с добавкой 10% сыворотки плодов телят, 2 мМ глутамина и 0,05% гентамицина. Для всех обработок клетки брали в концентрации 1 x 106/мл.
Анализ ингибирования роста. Активность в ингибировании роста F094 и авицинов измеряли с помощью теста восстановления MTT [3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромида)] (Deng et al., 1999). Клетки (1 x 104/лунка) культивировали с различными концентрациями F094, авицина D или авицина G в 96-луночных планшетах в течение 72 часов при 37°С. Клетки окрашивали MTT в течение 2 часов и затем инкубировали с буфером для лизиса (20% додецилсульфат натрия в 50% N,N-диметилформамиде) еще в течение 6 часов. В качестве показателя жизнеспособности клеток использовали оптическую плотность при 570 нм.
Анализ связывания аннексина V-ФИТЦ. Индукцию апоптоза изучали с помощью тестирования связывания аннексина V-ФИТЦ. Клетки Jurkat (1 x 106) обрабатывали 2 мкг/мл F094, авицина D или авицина G при 37°С. После промывки клеток холодным ЗФР их ресуспендировали в буфере для связывания (10 мМ HEPES/NaOH, 140 мМ NaCl, 2 мМ CaCl2). Добавляли конъюгат аннексин V-ФИТЦ (Bio Whittaker, Walkersville, MD) (1 мкг/мл) и инкубировали в течение 15 минут при комнатной температуре в темноте. Затем клетки окрашивали иодидом пропидия (5 мкг/мл) и анализировали с помощью проточной цитометрии (Hansen et al., 1989).
Выявление высвобождения цитохрома c из митохондрий. Высвобождение цитохрома c из митохондрий определяли с помощью иммуноблоттинга. Клетки Jurkat (1 x 107) обрабатывали F094, авицином D или авицином G (2 мкг/мл) при 37°С. После промывки осадков клеток в сахарозном буфере (0,25 М сахароза, 30 мМ трис, pH 7,7, 1 мМ ЭДТА) их ресуспендировали в 20 мкл сахарозного буфера, содержащего 1 мкМ PMSF, 1 мкг/мл лейпептина, 1 мкг/мл пепстатина и 1 мкг/мл апротинина. Клетки разрушали с помощью 120-кратного douncing в 0,3 мл douncer Kontes (Kontes Glass Company, Vineland, NJ) с применением пестика B. Клеточный белок (50 мкг) разделяли на 15% ДДС-Na-полиакриламидном геле и переносили с помощью электропереноса на нитроцеллюлозную мембрану. Мембрану зондировали моноклональным антителом против цитохрома c (Pharmingen, San Diego, CA) и затем антимышиным антителом, конъюгированным с пероксидазой хрена (HRPO). Белковые полосы выявляли с помощью хемилюминесценции (ECL, Amersham).
Выделение субмитохондриальной фракции. Клетки Jurkat (1,5-2,0 x 107) суспендировали в сахарозном буфере (250 мМ сахароза в 30 мМ трис-HCl pH 7,4) и переносили в N2 кавитационную камеру (PARR Instruments Company, Moline, IL). Клетки подвергали N2 кавитации (300 фунтов на квадратный дюйм в течение 5 мин), как указано в инструкциях производителя. При данных условиях большая часть клеточных мембран разрушалась без изменения дыхательной активности митохондрий. Затем ДНК и ядерную фракцию удаляли путем центрифугирования (1500g, 30 сек). Супернатант дополнительно центрифугировали (16000g, 10 мин) и осадок использовали как субмитохондриальную фракцию.
Тестирование протеазы каспазы-3. Активность каспазы-3 измеряли, как описано ранее (Martin et al., 1995), с некоторыми модификациями. Вкратце, клетки Jurkat (1 x 106) обрабатывали F094, авицином D или авицином G (2 мкг/мл) при 37°С. Цитозольные экстракты получали повторным замораживанием-размораживанием клеток в 300 мкл буфера для экстракции (12,5 мМ трис, pH 7,0, 1 мМ DTT, 0,125 мМ ЭДТА, 5% глицерин, 1 мкМ PMSF, 1 мкг/мл лейпептин, 1 мкг/мл пепстатин и 1 мкг/мл апротинин). Клеточные лизаты разводили 1:2 буфером ICE (50 мМ трис, pH 7,0, 0,5 мМ ЭДТА, 4 мМ DTT и 20% глицерин) и инкубировали с 20 мкМ субстрата каспазы-3 (ацетил-Asp-Glu-Val-Asp-аминометилкумарин) (Calbiochem, La Jolla, CA) при 37°С. Активность каспазы-3 отслеживали по образованию флуоресцентного аминометилкумарина, который измеряли при возбуждении 355 нм и эмиссии 460 нМ с применением Fluoroscan II (Labsystems, Helsinki, Финляндия).
Анализ деградации PARP методом иммуноблоттинга. Индукцию апоптоза также проверяли по протеолитическому расщеплению PARP (Nicholson and Thornberry, 1997). Клетки Jurkat (3 x 106) обрабатывали 2 мкг/мл F094, авицина D или авицина G при 37°C. Клеточные лизаты получали в буфере, содержащем 20 мМ HEPES, 250 мМ NaCl, 2 мМ ЭДТА, 0,1% NP-40, 2 мкг/мл лейпептина, 2 мкг/мл апротинина, 0,5 мкг/мл бензамидина, 1 мМ DTT и 1 мМ PMSF. Клеточные белки (60 мкг/мл) разделяли на 7,5% ДДС-Na полиакриламидном геле и переносили электропереносом на нитроцеллюлозную мембрану. Мембрану сначала зондировали моноклональным антителом против PARP (PharMingen), а затем антимышиным антителом, конъюгированным с пероксидазой хрена (HRPO). Белковые полосы определяли по хемилюминесценции (ECL). В качестве показателя апоптоза использовали появление 85кДа продукта расщепления.
Измерение мембранного потенциала митохондрий (▵ψm). Митохондриальный ▵ψm измеряли, как описано Zamzami et al., 1995. После обработки F094 авицином D или авицином G (2 мкг/мл) при 37°C клетки Jurkat инкубировали с 80 нМ DiOC6 в течение 15 мин при комнатной температуре. Их затем анализировали с помощью цитофлуориметра (Fascalibar, возбуждение 488 нм и испускание 552 нм).
Тестирование образования активных форм кислорода (АФК). Образование АФК определяли с помощью чувствительного к окислению флуоресцентного красителя 5,6-карбокси-2',7'-дихлорфлуоресцина диацетата (DCFH-DA) (Molecular Probes, Eugene, OR) с помощью описанного ранее способа (Zamzami et al., 1995). Клетки Jurkat, посеянные в 96-луночные планшеты (5 x 104 клеток/лунка), обрабатывали F094, авицином D или авицином G (2 мкг/мл) в буфере Кребса-Рингера, содержащем 20 мМ HEPES, 10 мМ D-глюкозы, 127 мМ NaCl, 5,5 мМ KCl, 1 мМ CaCl2 и 2 мМ MgSO4 (pH 7,4). В лунки добавляли DCFH-DA в концентрации 5 мкг/мл. Необработанные клетки получали только DCFH-DA. Клетки возбуждали при 485 нм и флуоресценцию измеряли каждые 2 минуты при 538 нм на протяжении до 2,5 часов на планшетном ридере Fluoroskan II ELISA, снабженном температурным контролем (Labsystems, Helsinki, Финляндия). Флуоресценцию измеряли в линейном диапазоне.
Результаты
Выявление структуры авицина D и авицина G химическим анализом. Авицин D, основной компонент F094, был выделен в виде бесцветного аморфного твердого вещества. Его молекулярная масса по данным масс-спектрометра MALDI составляла 2104 атомных единиц массы, что является аддуктом натрия молекулярной массы 2081. Масс-спектрометр высокого разрешения FAB дал молекулярную формулу C98H155NO46, подтвердив молекулярную массу 2081. Анализ протонного ядерного магнитного резонанса (ЯМР) авицина D выявил, что он является сапонином с боковой цепью, содержащей две единицы ациклического монотерпена, транс-2-гидроксиметил-6-метил-6-гидрокси-2,7-октадиеновой кислоты, соединенной с помощью хиновозного сахара и присоединенной к углероду 21 акациевой кислоты. Он также содержал трисахарид при углероде 3 и тетрасахарид при углероде 28. С помощью различных экспериментов 2D ЯМР и исследований деградации была получена структура, изображенная на фиг. 56B. Авицин G, другой активный компонент F094, представляет собой сапонин, очень похожий на авицин D. Его масс-спектроскопия MALDI дала молекулярную массу 2065. Протонный ЯМР указал на сходную с авицином D боковую цепь, но с замененным транс-2,6-диметил-6-гидрокси-2,7-октадиеновой кислотой наружным монотерпеном, как показано на фиг. 56B (R=H).
F094 и монотерпеновые/тритерпеновые гликозиды ингибируют рост путем индукции апоптоза. Было обнаружено, что F094 и авицины ингибируют рост клеток Jurkat в культуре. Ингибирующая концентрация 50 (IC50) F094 составляла 0,331-0,407 мкг/мл, тогда как таковая для авицина D и авицина G равнялась 0,320-0,326 мкг/мл и 0,160-0,181 мкг/мл, соответственно. В противоположность этому, при тестировании на нормальных фибробластных клетках человека F094 и авицины имели величины IC50, в 10-35 раз более высокие. Для понимания механизма ингибирования роста клетки Jurkat, обработанные F094 или монотерпеновыми/тритерпеновыми гликозидами, анализировали на предмет связывания аннексин V-ФИТЦ. Клетки одновременно окрашивали иодидом пропидия для контроля их жизнеспособности. Обработка всеми тремя препаратами приводила к зависимому от времени увеличению жизнеспособных аннексин V-позитивных клеток (фиг. 57).
F094 и авицины вызывают высвобождение цитохрома c из митохондрий. Высвобождение цитохрома c из митохондрий в цитозоль, по-видимому, является одним из ранних событий, ведущих к апоптозу. В обработанных F094 клетках показано повышение цитозольного уровня цитохрома c приблизительно через 4 часа после обработки (1,5-кратное) (фиг. 58). Для уровня цитохрома c в цитозоле клеток, обработанных авицином D, показано более быстрое повышение. 1,5-кратное повышение наблюдалось в пределах 30 минут, а 3-кратное повышение - в пределах 4 часов. Интересно, что в цитозоле клеток, обработанных монотерпеновым/тритерпеновым гликозидом G, резкое (3,5-кратное) повышение уровня цитохрома c наблюдалось уже через 30 минут после обработки. К 4 часам наблюдалось 8,4-кратное повышение уровня цитохрома c (фиг. 58).
Для выяснения того, действуют ли монотерпеновые/тритерпеновые сапонины прямо на митохондрии для индукции апоптоза, были проведены эксперименты с применением авицина G в бесклеточной системе. Митохондрии выделяли из клеток Jurkat с помощью способа кавитации N2. Обработка данной митохондриальной фракции 2 мкг/мл авицина G вызывала зависимое от времени высвобождение цитохрома c, начиная с первой минуты обработки с достижением максимума между 5 и 10 мин (фиг. 59A). Исследование зависимости ответа от дозы показало, что наибольшее высвобождение цитохрома с достигается при 0,5-2,0 мкг/мл авицина G и инкубации в течение 10 мин (фиг. 59B). Предварительная обработка митохондриальной фракции DEVD-CH2F необратимым ингибитором каспазы 3 или z-Val-Ala-Asp-CH2F (zVAD-fmk) проникающим в широкий спектр клеток необратимым ингибитором каспаз с широкой специфичностью, не влияла на высвобождение цитохрома c, что вновь указывает на то, что агенты прямо действуют на митохондрии.
F094 и авицины индуцируют активацию каспаз. Высвобождение цитохрома c из митохондрий в цитозоль запускает активацию каскада каспаз, которые играют роль решающих нижележащих эффекторов в различных путях клеточной гибели. Поэтому был проведен анализ состояния каспазы-3 в обработанных клетках Jurkat. Индуцированная F094 и авицином D активация каспазы-3 определялась через 4-6 часов после обработки и позднее (фиг. 60A). Однако с авицином G активность каспазы наблюдалась через 2-4 часа после обработки. К 16 часам активность каспазы снижалась до базального уровня (фиг. 60A).
Одной из нижележащих мишеней каспазы-3 является PARP, который протеолитически расщепляется каспазой-3. И F094, и авицины индуцировали расщепление PARP, начиная с 4 часов после обработки (фиг. 60B). С авицином G расщепление было почти полным к 4 часам, тогда как со смесью и авицином D для полного расщепления требовалось 8-16 часов. Предварительная обработка клеток zVAD-fmk полностью блокировала расщепление PARP (фиг. 60C).
F094 и авицины не влияют на мембранный потенциал митохондрий (▵ψm). Высвобождение цитохрома c в цитозоль обычно предшествует падению или сопровождается падением митохондриального ▵ψm. Обработка клеток Jurkat F094, авицином D или авицином G на протяжении до 8 часов не вызывала существенных изменений ▵ψm. Однако более длительная обработка (16 часов) вызывала значительное падение ▵ψm (фиг. 61).
F094 и авицины снижают образование активных форм кислорода (АФК). Большинство индуцирующих апоптоз агентов высвобождают АФК, ключевой посредник апоптотической сигнализации. Однако обработка клеток Jurkat монотерпеновыми/тритерпеновыми гликозидами на протяжении до 2,5 часов приводила к снижению уровня АФК зависимым от дозы образом (фиг. 62). Данное снижение уровня АФК достигало плато, и при более долгой экспозиции с агентами дальнейшего изменения наблюдать не удавалось (до 24 часов).
Обсуждение
Гомеостаз в эукариотических клетках зависит от тонкого равновесия между сигналами выживания и гибели, исходящими из внеклеточного окружения (Oridate et al., 1997). Любое отклонение в любом из данных путей сигнализации может быть губительным для физиологии клетки. При раке делящиеся клетки не утрачивают способности инициировать апоптоз после стойкого повреждения ДНК (Raff M.C., 1997). Было идентифицировано несколько путей, ведущих к апоптозу, включая индукцию p53 (Raff M.C., 1992), активацию керамидов (Polyack et al., 1997), стимуляцию пути CD95/CD95-лиганд (Basu et al., 1998) и обнаруженные позднее пути сигнализации внутри митохондрий (Hakem et al., 1998; Fujimura et al., 1999; Green et al., 1998; Fulda et al., 1998). Одной из целей химиотерапии рака и его профилактики является выявление новых агентов, которые могут индуцировать апоптоз избирательно в опухолевых клетках посредством влияния на один или более из данных проапоптотических путей.
Здесь продемонстрирована индукция апоптоза в T-клеточной линии человека Jurkat под действием смеси монотерпеновых/тритерпеноидных сапонинов и монотерпеновых/тритерпеновых гликозидов, опосредуемая влиянием на функцию митохондрий. Анализ пути проведения сигнала показал, что цитохром c высвобождается в интервале от 30 минут до 2 часов после обработки. В бесклеточной системе авицин G индуцировал высвобождение цитохрома c в пределах минуты после обработки. Предварительная обработка ингибиторами каспаз DEVD или zVAD-fmk не оказывала влияния на высвобождение цитохрома c, что указывает на то, что авицин G действует прямо на митохондрии без вовлечения каспаз, расположенных выше цитохрома c. После высвобождения цитохрома c происходили активация каспазы-3 и расщепление поли(АДФ-рибоза)полимеразы (PARP) в пределах 2-6 часов после обработки авицинами. Предварительная обработка клеток zVAD-fmk полностью блокировала расщепление PARP. В обработанных клетках высвобождению цитохрома c не предшествовали и его не сопровождали существенные изменения мембранного потенциала. Было также определено небольшое снижение образования активных форм кислорода (АФК). Таким образом, чистые монотерпеновые/тритерпеноидные сапонины, которые являются вторичными метаболитами, служащими в качестве защитных молекул в некоторых растениях и морских организмах, нарушают функцию митохондрий, индуцируя тем самым гибель опухолевых клеток человека.
Как описано в предшествующих примерах, смеси монотерпеновых/тритерпеноидных сапонинов, выделенные из Acacia victoriae (Benth), ингибируют рост множества линий опухолевых клеток человека. Одним из вовлеченных механизмов является ингибирование сигнального пути PI-3-киназы. Однако в клетках Jurkat для данных эффектов требуется продолжительная обработка монотерпеновыми/тритерпеноидными сапонинами (16 часов), период, слишком отдаленный для объяснения раннего наступления апоптоза в клетках Jurkat. Поэтому авторы настоящего изобретения исследовали роль митохондрий, причем не только со смесью монотерпеновых/тритерпеноидных сапонинов, но и с двумя очищенными молекулами. Смесь монотерпеновых/тритерпеноидных сапонинов (F094) разделяли с помощью препаративной ВЭЖХ на несколько компонентов. С помощью мониторинга цитотоксичности на клетках Jurkat для биологической характеристики были выбраны две молекулы, авицин D и авицин G.
В клетках в начале процесса апоптоза происходит перемещение фосфатидилсерина с внутренней стороны плазматической мембраны в ее наружный слой. В данном состоянии экспонирования он может связываться с аннексином V (Friesen et al., 1996), и данное свойство было использовано в качестве маркера апоптоза. Для клеток Jurkat, обработанных F094 или авицинами, было показано зависимое от времени повышение аннексин V-позитивных клеток, начиная с 4 часов после обработки, что указывает на индукцию апоптоза в данных клетках.
Одним из ранних событий, которые инициируют апоптоз, является высвобождение цитохрома c из митохондрий в цитозоль (Schutte et al., 1998; Yang et al., 1997). В цитозоле обработанных клеток Jurkat цитохром c определялся в пределах от 30 минут (для авицина D и авицина G) до 4 часов (для F094). Оказавшись в цитозоле, цитохром c связывается с APAF-1 и прокаспазой-9 в присутствии dATP с образованием апоптосомы (Kluck et al., 1997; Zou et al., 1997). Данный комплекс активирует каспазу-9, которая в свою очередь расщепляет и тем самым активирует каспазу-3. В обработанных клетках высвобождение цитохрома c из митохондрий сопровождалось активацией каспазы-3. Данное событие непосредственно сопровождалось расщеплением PARP, одного из субстратов каспазы-3, являющегося 116 кДа ферментом репарации ДНК. Расщепление PARP приводит к инактивации фермента, что тем самым делает невозможным репарацию ДНК.
Высвобождение цитохрома c может быть инициирующим событием в апоптозе или оно может располагаться ниже по отношению к активации каспаз, как это имеет место в системе CD95 (Fas). Последний вариант зависит от выделения каспазы-8 (Li et al., 1997). После предварительной обработки клеток zVAD-fmk, ингибитора каспаз с широким спектром действия, наблюдалось полное блокирование расщепления PARP при отсутствии влияния на высвобождение цитохрома c. Это показывает, что высвобождение цитохрома c является прямым результатом действия данных агентов на митохондрии и не зависит от активации каспаз. Для подтверждения данных результатов были использованы очищенные митохондрии в бесклеточной системе. В данной системе авицин G индуцировал высвобождение цитохрома c зависимым от времени и дозы образом, и на эту индукцию не влияла предварительная обработка ингибиторами каспаз DEVD-CH2F и zVAD-fmk.
Пока не ясно, каким образом цитохром c, находящийся в пространстве между внешней и внутренней мембраной митохондрий, транслоцируется в цитозоль. Было предложено несколько теорий. Была показана значительная корреляция между апоптозом и феноменом перехода проницаемости митохондрий (PT), который вызывается открытием широкого канала проводимости, что ведет к деполяризации внутренней мембраны (Kuwana et al., 1988). Данная деполяризация в конечном итоге приводит к набуханию митохондрий, разрыву внешней мембраны и высвобождению цитохрома c. Было предположено образование специфических каналов во внешней мембране митохондрий, обеспечивающих высвобождение цитохрома c (Petit et al., 1996). Позднее предположили, что цитохром c может высвобождаться без потери трансмембранного потенциала, но в этом случае происходит гиперполяризация внутренней мембраны (Manon et al., 1997). В обработанных F094 или авицином G клетках высвобождению цитохрома c не предшествовали и его не сопровождали изменения потенциала внутренней мембраны митохондрий. Однако через 16 часов после обработки была показана значительная деполяризация мембраны, что соответствует сообщениям, и это указывает на то, что активированные каспазы прямо индуцируют PT (Vander Heiden et al., 199). Для дальнейшего понимания механизма высвобождения цитохрома c авторы изобретения обдумывают эксперименты с анализом обмена АТФ/АДФ и действия на F0F1-АТФазный протонный насос.
Митохондрии являются богатым источником активных форм кислорода (АФК), которые служат токсичными побочными продуктами аэробного существования и играют важную роль в различных путях проведения сигнала, включая те, которые ведут к апоптозу (Marzo et al., 1998; Korsmeyer S.J., 1995; Buttke and Sandstorm, 1994). Поэтому изучали уровни АФК в клетках Jurkat после обработки монотерпеновыми/тритерпеновыми гликозидами. Наблюдалось снижение образования АФК после обработки монотерпеновыми/тритерпеновыми гликозидами. Некоторые сообщения указывают на то, что образование АФК предшествует высвобождению цитохрома c (Bredesen D.E., 1995), тогда как другие указывают на то, что оно может быть поздним событием, имеющим место даже после инициации активации каспаз. Непрерывные наблюдения уровня АФК до 16 часов после обработки в настоящем изобретении не обнаружили последующего изменения.
Авторы полагают, что снижение уровня АФК после обработки клеток авицинами может быть обусловлено клетками, поддерживающими свой уровень митохондриального АТФ либо за счет удержания некоторого количества цитохрома c в митохондриях, либо за счет клеток, сохранявших гликолиз даже после наступления апоптоза. Таким образом, АТФ может задерживать или ингибировать продукцию АФК. На уровень АФК могли бы прямо или непрямо влиять изменения в регуляции энергообмена, такие как (i) нарушения обмена АТФ/АДФ (Manon et al., 1997), (ii) окисление супероксида до кислорода цитохромом с (Higuchi et al., 1998) или (iii) протеолиз каспазами ингибитора диссоциации D4-ГДФ (Skulachev V.P., 1998). Альтернативным объяснением могло бы быть повышение уровня антиоксидантов, таких как глутатион или супероксиддисмутаза.
Исследования показывают повышение интереса к раскрытию соединений, которые прямо влияют на митохондрии (Na et al., 1996; Ravagnan et al., 1999; Zamzami et al., 1998; Hirsch et al., 1998; Chen et al., 1998). Все данные агенты вызывают апоптоз либо путем нарушения мембранного потенциала, либо за счет высвобождения АФК, что позволяет предполагать, что первичной мишенью служит внутренняя митохондриальная мембрана. Сообщалось, что бетулиновая кислота, пентациклический тритерпен, вызывает апоптоз в нейроэктодермальных опухолях (Bantel et al., 1999), действуя прямо на митохондрии, зависимым от CD95 и p53 образом (Pisha et al., 1995). Было показано, что, в противоположность тритерпеновым гликозидам, бетулиновая кислота вызывает снижение трансмембранного потенциала и повышение уровня АФК одновременно с высвобождением цитохрома c с внешней митохондриальной мембраны (Pisha et al., 1995). Точные химические причины того, почему тритерпеновые гликозиды сохраняют мембранный потенциал и снижают АФК в противоположность действию бетулиновой кислоты, остается неизвестным. Бетулиновая кислота является простым тритерпеновым соединением с очень ограниченной специфичностью в отношении типов клеток. С другой стороны, тритерпеновые гликозиды, проявляющие более широкую клеточную специфичность, содержат гидрофобную акациевую кислоту в качестве стержневого тритерпена и имеют две ациклические монотерпеновые единицы, соединенные хиновозным сахаром. Гидрофобное ядро акациевой кислоты в авицинах, возможно, обеспечивает прохождение ими мембраны и оказание влияния на митохондрии, а свойства остальной части молекулы могут объяснять их биологические эффекты и их отличия от действия бетулиновой кислоты.
Авторы рассматривают возможность изучения укорочения частей авицина D и авицина G, включая боковые цепи и отдельные сахара, для выявления компонента, который является решающим для проапоптотической функции. Тритерпеновые гликозиды структурно очень похожи на эллиптозиды, выделенные ранее из Archidendron ellipticum (Fulda et al., 1997). Однако оценка с помощью ЯМР и исследования хиральности монотерпенов показывают, что имеются небольшие, но существенные различия между сапонинами, о которых сообщается здесь, и теми, которые были описаны ранее как обладающие противоопухолевой активностью.
Авицины являются новым классом монотерпеновых/тритерпеноидных сапонинов, которые индуцируют апоптоз в клетках Jurkat путем нарушения митохондриальной функции независимо от связанных с мембраной рецепторов смерти. Данные соединения избирательно индуцируют апоптоз раковых клеток (например, погибают клетки Jurkat, но не нормальные фибробласты), что делает их полезными химиотерапевтическими агентами, поскольку они не оказывают побочных эффектов в виде индукции гибели других нормальных клеток.
Онкопротеины сенсибилизируют раковые клетки к апоптотическим сигналам на путях, которые резистентны в нормальных клетках (Harrington et al., 1994). Так, эффекты авицинов осуществляются посредством амплификации ранних сигнальных событий, которые ведут к апоптозу. Было показано, что фосфорилирование каспазы-9 под действием AKT ингибирует ее активность (Cardone et al., 1998). Путем ингибирования фосфорилирования AKT авицины могут амплифицировать проапоптотические эффекты, наблюдаемые после высвобождения цитохрома c. Недавно было показано, что APAF и каспаза-9 являются нижележащими эффекторами индуцируемых p53 генов (Soengas et al., 1999). Ингибирование фосфорилирования AKT может также повышать проапоптотическую функцию BAD путем повышения его гетеродимеризации с Bcl-xL (Gross et al., 1999). Альтернативно, авицины могут действовать как детергенты для индукции образования гомодимеров Bax (Hsu et al., 1999). Имея в виду их прямое действие на митохондрии, авторы изобретения полагают, что авицины должны преодолевать резистентность к апоптозу благодаря мутациям в гене p53. Это подтверждается тем фактом, что использованные здесь клетки Jurkat лишены p53. Таким образом, авицины являются эффективными для лечения резистентных раков благодаря их способности замещать функцию утраченных или мутировавших генов супрессоров опухолей.
Пример 48
Снижение эпидермальной гиперплазии, воспаления и мутаций p53
Длины волн UVB спектра, обеспечивающие образование витамина D, ответственны также за повреждение ДНК, в частности за образование пиримидиновых димеров и мутации в ряде генов, а также за продукцию радикалов кислорода, которые дополнительно повреждают ДНК. В данном примере описано ингибирование повреждений кожи мышей, вызванных освещением UVB, монотерпеновыми/тритерпеновыми сапонинами. В примере также описана разработка новой краткосрочной мышиной модели для скрининга модуляторов индуцированного UVB канцерогенеза кожи.
Способы. Бесшерстных мышей SKH-1 облучали UVB в дозе 180 мДж/см2 в течение 15 мин пять раз в неделю в течение 10 недель. Относительную эффективность обработки оценивали по конечной толщине эпидермиса, инвазии лейкоцитов, показателю промечивания (BrdU), мутаций p53 и образованию 8-OH-dG. На кожу спины бесшерстных мышей SKH-1 за 15 мин до облучения UVB наносили только F035 или F035 в сочетании с сильным антиоксидантом витамином E.
Результаты. Смесь монотерпеновых/тритерпеновых сапонинов (F035) снижала повреждение оснований ДНК, а также снижала мутации p53. По данным измерений промечивания 5-бром-2'-дезоксиуридином (BrdU), количество находящихся в цикле клеток, локализованных в базальном и надбазальном слоях кожи, снижалось на 30% у мышей, обработанных только витамином E, 35% у мышей, обработанных только F035, и 55% у мышей, обработанных сочетанием витамина E и F035. Количество позитивных в отношении мутации p53 клеток, расположенных преимущественно в надбазальном слое, снижалось на 60% и 67% у мышей, обработанных во время облучения сочетанием монотерпеновых/тритерпеноидных сапонинов и витамина E. Таким образом, монотерпеновые/тритерпеновые сапонины согласно изобретению снижают эпидермальную гиперплазию, воспаление и мутации p53. Кроме того, в примере предлагается краткосрочная мышиная модель, которая очень полезна для оценки модуляторов индуцированного UVB канцерогенеза кожи.
Таким образом, авторы предвидят способы профилактики индукции мутаций p53 путем введения субъекту композиции, включающей в себя описанные здесь композиции монотерпеновых/тритерпеновых гликозидов. Композиции монотерпеновых/тритерпеновых гликозидов можно вводить многочисленными путями, такими как системный, местный или региональный. В частности, их можно вводить внутривенно, внутримышечно, подкожно, перорально или местно. Таким образом, в изобретении предлагаются способы профилактики индуцированных p53 раков. Мутации p53 участвуют во многих типах рака, и авторы предполагают, что композиции монотерпеновых/тритерпеновых гликозидов, введенные человеку, должны обеспечивать эффективную профилактическую и лечебную терапию против различных видов рака.
Авторы также предусматривают способы профилактики/лечения индуцированного ультрафиолетовым светом раковых и предраковых состояний путем введения индивидууму описанных здесь композиций монотерпеновых/тритерпеновых гликозидов. Например, предраковые состояния, такие как актинический кератоз, и рак, и карцинома кожи могут быть предотвращены с помощью описанных здесь композиций монотерпеновых/тритерпеновых гликозидов. Индуцированные ультрафиолетовым светом повреждения кожи можно лечить местным нанесением кремов, гелей или мазей, включающих в себя композиции монотерпеновых/тритерпеновых гликозидов. Кроме того, авторы предусматривают профилактику индуцированных ультрафиолетовым светом состояний кожи, включая рак, путем местного нанесения средств защиты от солнца, лосьонов, кремов, гелей или мазей, включающих в себя композиции монотерпеновых/тритерпеновых гликозидов. Данные терапевтические композиции могут дополнительно включать в себя дополнительные композиции, такие как витамин E. Способы могут также применяться в дополнение к другим способам лечения, которым подвергается индивидуум для лечения предрака или рака.
Здесь предлагается также новая мышиная модель для скрининга модуляторов индуцированного UVB канцерогенеза кожи. Предлагается бесшерстная мышь SKH-1, которая является прекрасной краткосрочной моделью. Мышь может быть облучена УФ для индукции предраковых или раковых повреждений кожи. Затем кожа спины бесшерстной мыши может быть обработана предполагаемым модуляторным соединением по желанию либо до, либо после обработки. Эффективность обработки может быть затем оценена путем отслеживания таких признаков как окончательная толщина эпидермиса, инвазия лейкоцитов, показатель промечивания (например, BrdU), мутации p53 и образование 8-OH-dG.
Все раскрытые здесь и включенные в формулу изобретения композиции и способы могут выполняться и осуществляться без излишнего экспериментирования в свете настоящего раскрытия. Хотя композиции и способы данного изобретения были описаны в форме предпочтительных осуществлений, специалистам в данной области техники должно быть ясно, что в композиции и способы, а также в стадии или в последовательность стадий описанного здесь способа могут быть внесены вариации, без выхода за пределы концепции, содержания и объема изобретения. Конкретнее, должно быть ясно, что определенные агенты, которые химически и физиологически близки, могут быть заменены описанными здесь агентами, и при этом должны быть достигнуты такие же или сходные результаты. Все такие сходные замены или модификации, очевидные для специалистов в данной области техники, считаются находящимися в пределах смысла, объема и концепции изобретения, как определено в формуле изобретения.
Ссылки
В качестве ссылок здесь включены следующие ссылки в той мере, в которой они обеспечивают методические или другие детали, дополнительные к тем, которые изложены здесь.















| название | год | авторы | номер документа |
|---|---|---|---|
| ТРИТЕРПЕНОВЫЕ КОМПОЗИЦИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 1999 |
|
RU2244547C2 |
| Способ ингибирования нуклеарного фактора каппа В с использованием 5-гидроксиникотината калия в культуре клеток | 2017 |
|
RU2675693C1 |
| СПОСОБ СТИМУЛИРОВАНИЯ ГУМОРАЛЬНОГО И КЛЕТОЧНОГО ИММУННОГО ОТВЕТА НА ЦЕЛЕВОЙ АНТИГЕН | 1996 |
|
RU2127605C1 |
| Способ ингибирования нуклеарного фактора каппа В с использованием 5-гидрокисиникотинат 3-(2,2,2-триметилгидразиний) пропионата калия в культуре клеток | 2017 |
|
RU2674443C1 |
| Способ ингибирования нуклеарного фактора каппа В с использованием 2-этил-6-метил-3-гидроксипиридиния L-2,6-диаминогексаноата в культуре клеток | 2017 |
|
RU2669348C1 |
| ТЕСТ-СИСТЕМА ДЛЯ АНАЛИЗА ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ АНТИТЕЛ ПРОТИВ PD-1 И АНТИТЕЛ ПРОТИВ PD-L1 | 2019 |
|
RU2731896C1 |
| СПОСОБ ПОЛУЧЕНИЯ ФРОНДОЗИДА А И СПОСОБ СТИМУЛИРОВАНИЯ ИММУННОЙ СИСТЕМЫ МЛЕКОПИТАЮЩИХ | 2005 |
|
RU2339644C2 |
| СРЕДСТВО, СТИМУЛИРУЮЩЕЕ АПОПТОЗ КЛЕТОК ЛЕЙКЕМИИ ЧЕЛОВЕКА | 2007 |
|
RU2360692C1 |
| АНТИ-ОХ40 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2753493C2 |
| ПРИМЕНЕНИЯ АНТИ-CD40-АНТИТЕЛ | 2008 |
|
RU2491095C2 |
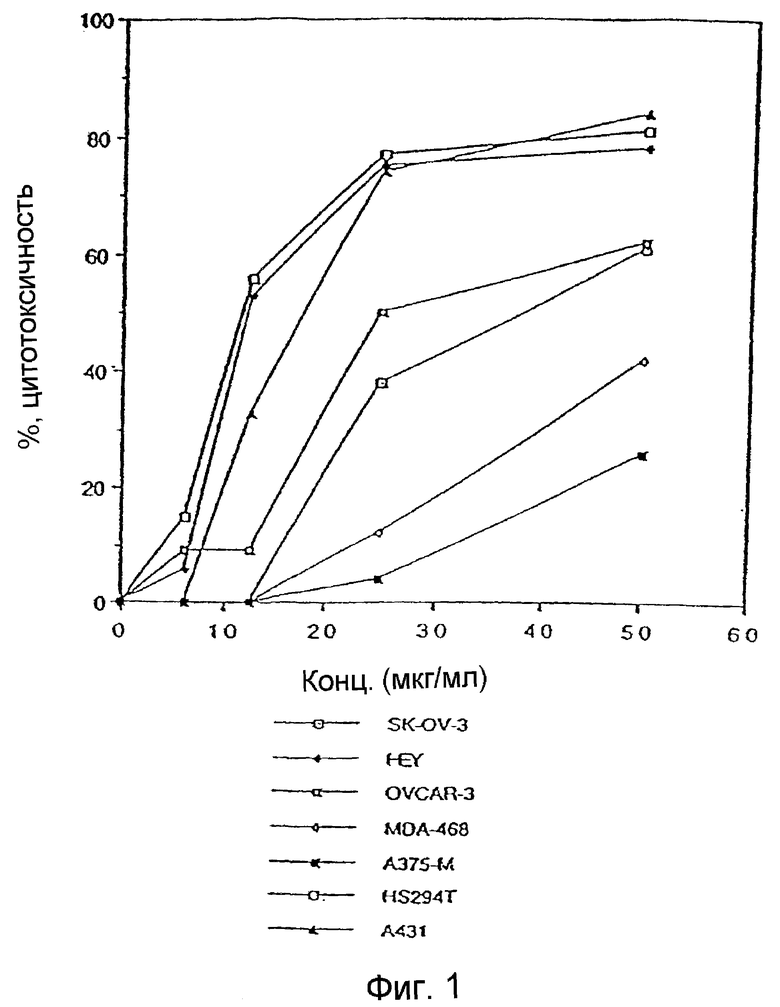




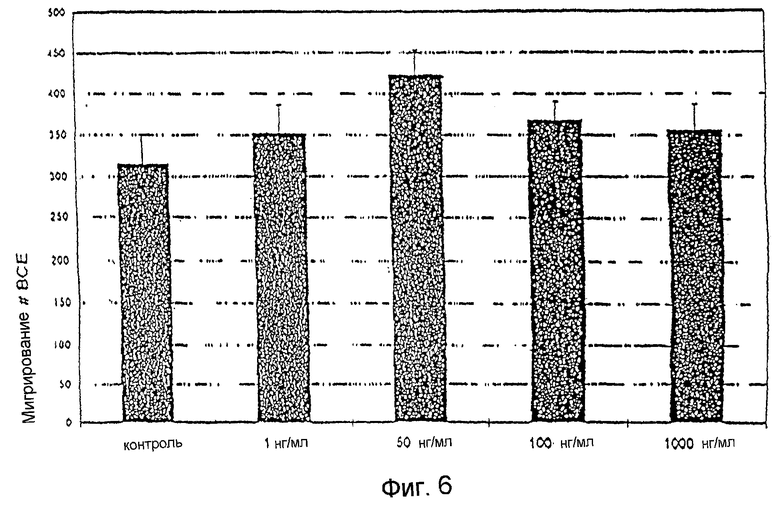
















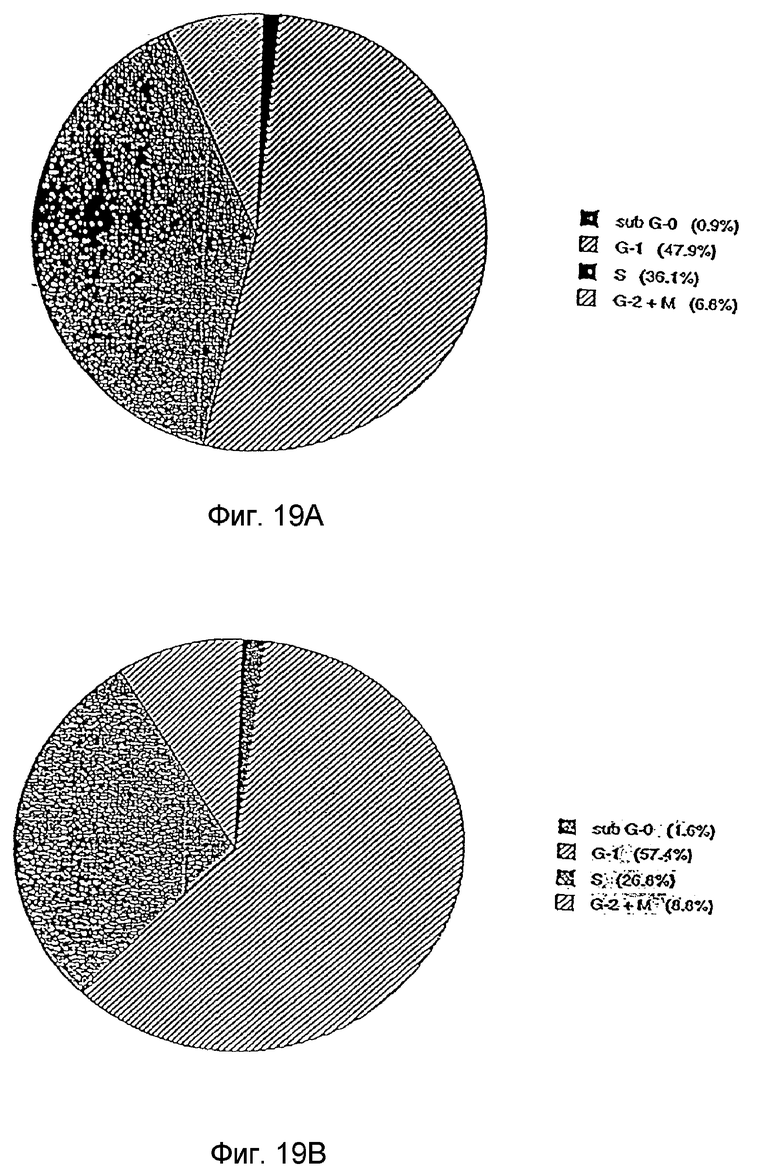
















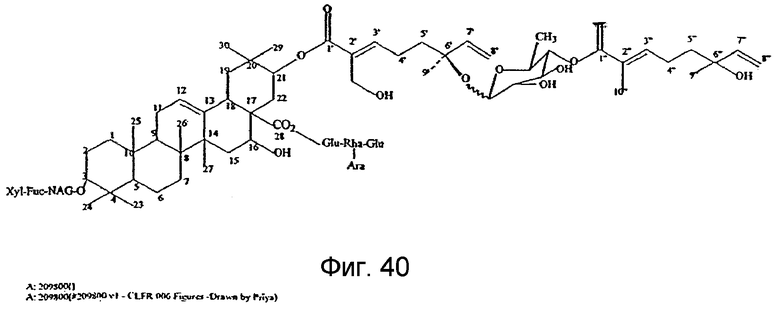








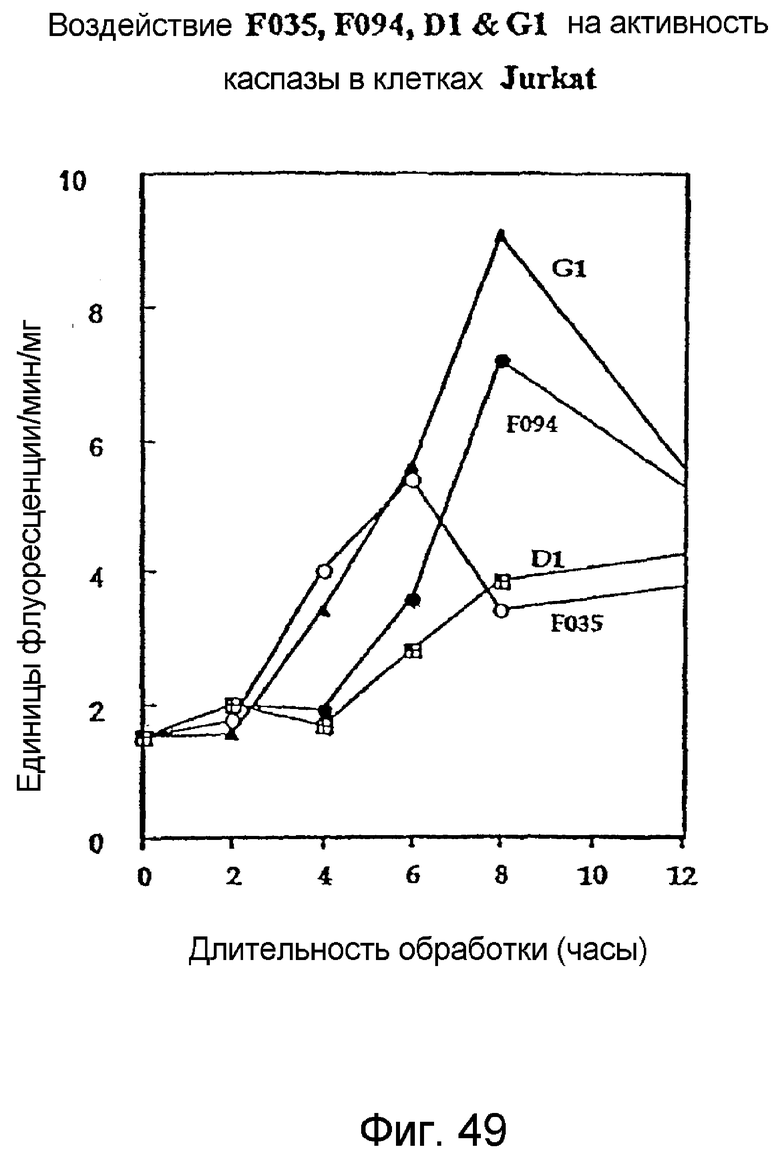

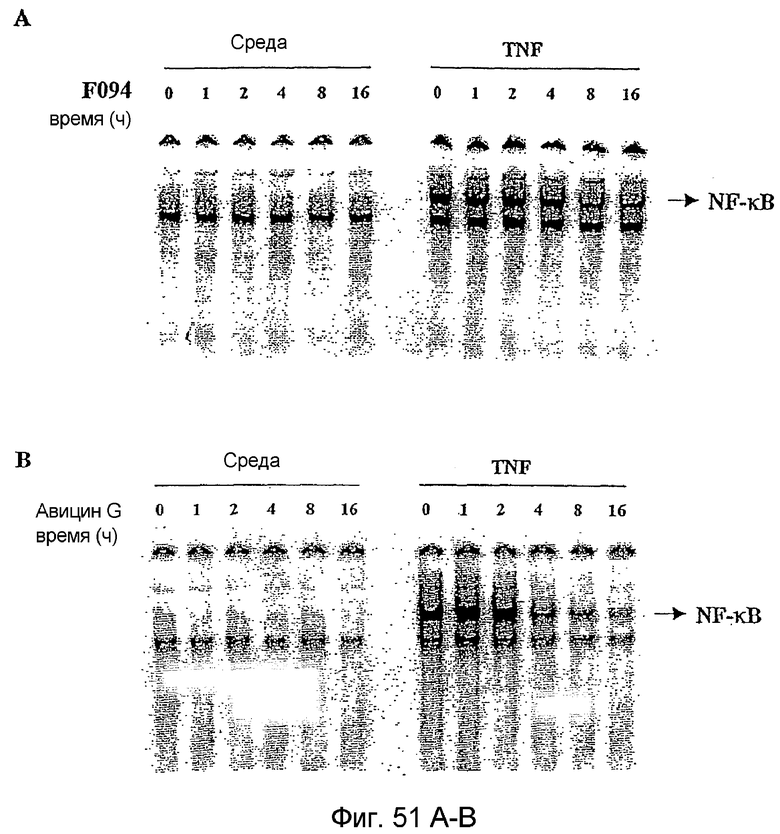
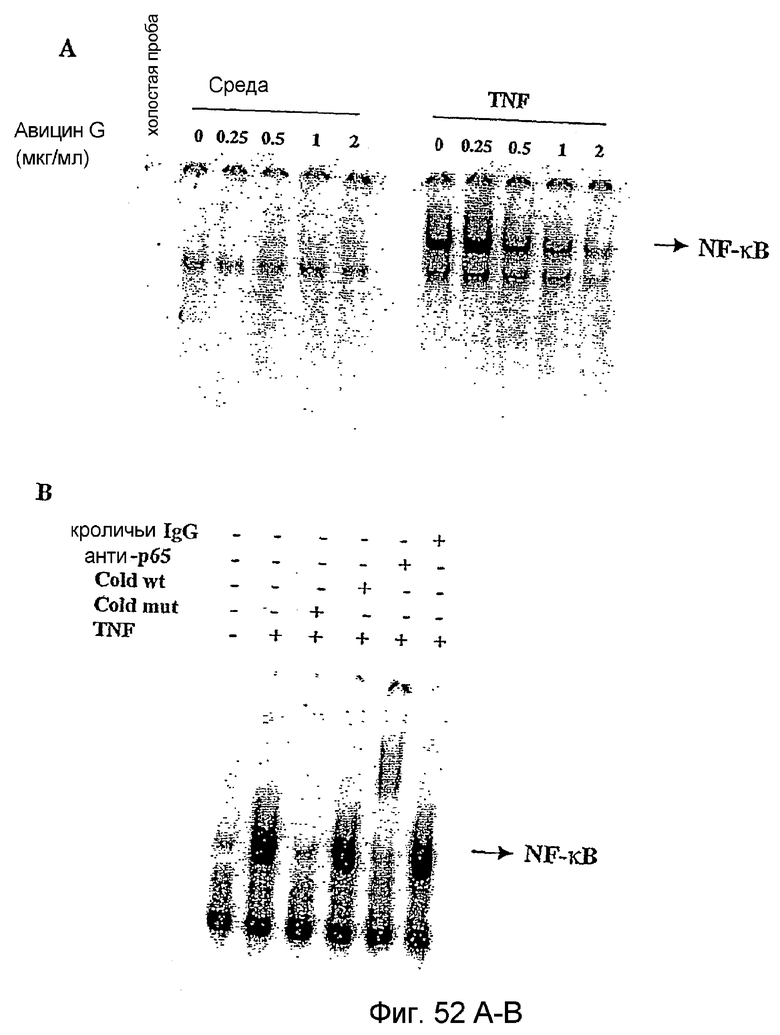




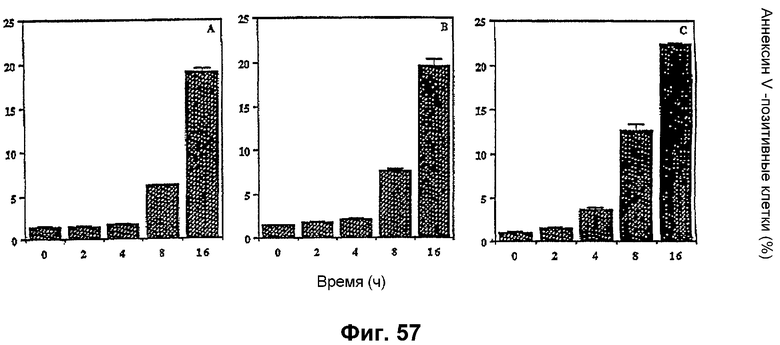



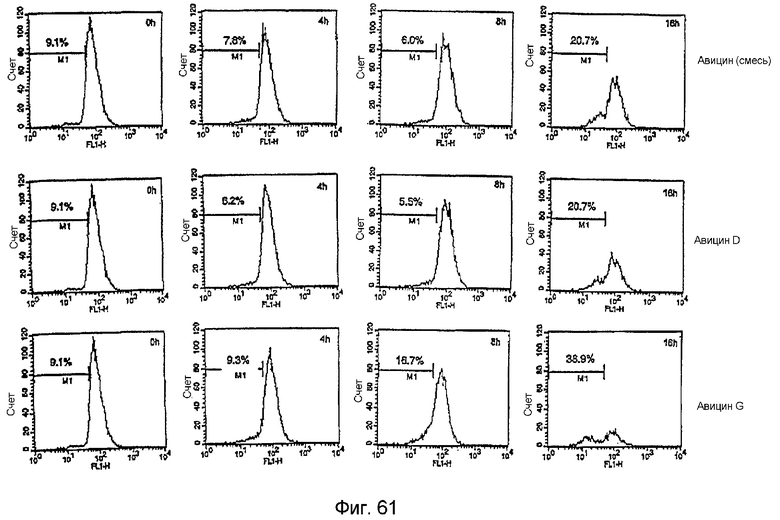







Изобретение относится к области биохимии. Сущность его заключается к разработке способа ингибирования воспаления путем обеспечения клетки монотерпеновыми композициями, которые ингибируют фактор NF-κB. Данные композиции могут также содержать часть в виде носителя, который делает монотерпеновую композицию проницаемой для мембран. Носитель может включать в себя тритерпеноидные части, сахара, липиды или даже дополнительные монотерпеноидные части. Композиция может включать дополнительные химические функциональные агенты. Технический результат - повышение эффективности лечения воспалительных состояний, в частности предраковых. 53 з.п. ф-лы, 55 табл., 62 ил.

или ее изомер, где
a) R1 и R2 выбраны из группы, состоящей из водорода, С1-C5-алкила, С1-C5-алкилена, С1-C5-алкилкарбонила, сахара, олигосахарида;
b) где каждый из R3-R36 отдельно и независимо выбран из группы, состоящей из точки ненасыщенной валентности, водорода, гидроксила, С1-C5-алкила, С1-C5-алкилена, С1-C5-алкилкарбонила, сахара, С1-C5-алкилового эфира и монотерпеновой группы; и
c) по меньшей мере один из R3-R36 представляет собой монотерпеновую группу.

или ее изомер, где
a) R3 выбран из группы, состоящей из водорода, гидроксила, C1-C5-алкила, С1-C5-алкилена, С1-C5-алкилкарбонила, сахара и монотерпеновой группы; и
b) формула дополнительно включает в себя R4, где R4 выбран из группы, состоящей из водорода, гидроксила, С1-C5-алкила, С1-C5-алкилена, С1-C5-алкилкарбонила, сахара, С1-C5-алкилового сложного эфира и монотерпеновой группы.

где R5 выбран из группы, состоящей из водорода, гидроксила, С1-C5-алкила, С1-C5-алкилена, С1-C5-алкилкарбонила, сахара, С1-C5-алкилового сложного эфира и монотерпеновой группы.
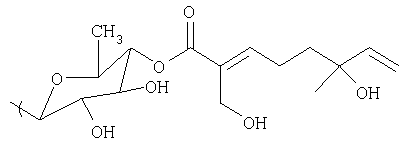


или ее изомер, где
a) R1 и R2 выбраны из группы, состоящей из водорода, С1-C5-алкила и олигосахарида;
b) R3 выбран из группы, состоящей из водорода, гидроксила, С1-C5-алкила, С1-C5-алкилена, С1-C5-алкилкарбонила, сахара и монотерпеновой группы.



| PETIT et al., FEBS Letters, 1996, v.396, p.7-13 | |||
| PEZZUTO et al, Biochem | |||
| Pharmacol., 1997, 53, p.121-133. |

