Изобретение относится к иммунологии и вирусологии, и может быть использовано для разработают вакцин, в том числе против СПИДа, безопасных в отношении активации вируса иммунодефицита человека (ВИЧ) и предназначенных для использования у ВИЧ-инфицированных людей.
Изобретение направлено на получение высокоэффективных иммуностимуляторов, усиливающих продукцию специфических антител и клеточно-пролиферативные реакции в ответ на введение антигенов без активации ВИЧ, находящегося в латентной форме, или в состоянии низкого уровня репликации.
Разработка и производство современных вакцин, включая вакцины против СПИДа, предполагает использование наряду с специфической антигенной субстанцией адьювантов, усиливающих иммунный ответ на целевой антиген. В современных вакцинах для людей в качестве адьювантов как правило используются соли алюминия (Al) [1, 2]. Однако соли Al способны индуцировать выработку IgE-антител и приводить к возникновению аллергии. Низкая иммуногенность поверхностных белков ВИЧ в присутствии Al явилась дополнительным негативным фактором, стимулировавшим поиск иных адьювантов, пригодных для создания анти-СПИД вакцин [3].
Среди кандидатов в адьюванты для включения в вакцины против СПИДа и вакцины, предназначенные для профилактики иных заболеваний у ВИЧ-инфицированных людей, рассматриваются мурамилпептиды и сапонины. Для многих мурамилпептидов продемонстрирован эффект активирования в различных типах клеток нуклеарного фактора NF-kB, что свидетельствует о потенциальной способности адьювантов этого типа активировать геном ВИЧ [4] и делает вакцины и иные препараты на их основе опасными для ВИЧ-инфицированных людей.
Прототипом настоящего изобретения является применение вещества сапонин Quil А, получаемого из Quillaja saponaria. Этот препарат обычно в композиции с фосфатидилхолином, холестерином и детергентом входит в состав так называемых иммуностимулирующих комплексов (ISCOMs) [5], куда включается и целевой антиген. ISCOMs стимулируют выработку антител к различным фрагментам поверхностных белков ВИЧ-1 активнее, чем соли алюминия, но уступают адьювантной смеси мурамилдипептида (МДП), сквалена, детергента Tween-80 и плюроник блок-полимера, а также полному адьюванту Фрейндa [3]. Недостатками использования Quil А являются: 1) необходимость использования препарата только в составе ISCOM, что делает применение адьюванта дороже, 2) отсутствие сведений о влиянии Quil А на репродукцию ВИЧ, 3) более слабый иммуностимулирующий эффект, чем у наиболее эффективных производных МДП и композиций на их основе. Кроме того, для получения сапонина Quil А используют Quillaja saponaria, растения, практически не произрастающего в Крыму, на Украине, в России и других европейских государствах.
В основу изобретения поставлена задача расширения набора средств, пригодных для применения у ВИЧ-инфицированных людей, конкретно - создание способа стимулирования клеточного и гуморального ответа на целевой антиген, в котором был бы исключен риск активации репродукции ВИЧ, если иммунизации подвергается ВИЧ-инфицированное лицо.
Поставленная задача решается тем, что в способе стимулирования гуморального и клеточного иммунного ответа на целевой антиген, включающем введение в организм адьювантов и целевого антигена, согласно изобретению в качестве адьюванта используют сапонины, выделяемые из различных органов растений рода Hedera L. с общей формулой
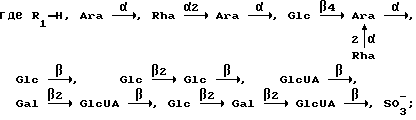
R2 - H, OH;
R3 - H, OH;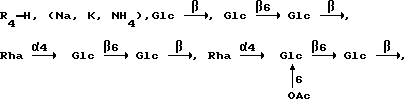
например: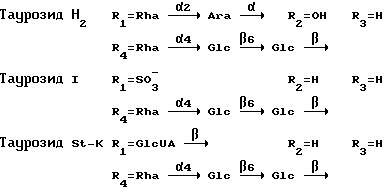
Условные обозначения:
Ara - L-арабинопиранозил, Glc - D-глюкопиранозил, Gal - D-галактопиранозил, GlcUA - D-глюкуронопиранозил, Rha - L-рамнопиранозил, Аc - ацетил
Способ получения тритерпеновых глюкозидов (сапонинов) подробно описан в [6] и заключается в высушивании растительного материала, измельчении, обезжиривании последовательно гексаном и хлороформом, и экстракции гликозидов бутанолом, насыщенным водой. Упаривание бутанольного экстракта дает сырую сумму веществ, содержащую сапонины. Для выделения суммы сапонинов, очищенной от сопутствующих веществ, остаток после упаривания растворяют в метаноле и сапонины осаждают ацетоном с последующим отделением осадка фильтрованием и промывкой сухим ацетоном. Для выделения индивидуальных сапонинов из их смеси проводят хроматографическое разделение на силикагеле при градиентном элюировании смесями хлороформ - этанол, насыщенными водой. Фракции, содержащие индивидуальные гликозиды, упаривают досуха и при необходимости дополнительно хроматографией на силикагеле при элюировании смесью хлороформ-этанол, насыщенной 10% водным аммиаком.
Целевой антиген может вводиться в любой дозировке и любым путем, приемлемым для данного продукта. Отдельные сапонины, или их комбинации могут вводиться в организм иммунизируемого как вместе с целевым антигеном, так и отдельно от него, до введения, во время введения, или после введения антигена. Сапонины могут вводиться в растворителях на водной основе, или дистиллированной воде как пероральным, так и парентеральным путем. Дозировка сапонинов может вариировать в зависимости от способа введения в концентрациях 0,1-5000,0 мкг/мл.
Сведениями, подтверждающими возможность осуществления изобретения, являются нижеперечисленные примеры.
Пример 1. Мышей линии Bald/C внутримышечно иммунизируют смесью антигена - ВИЧ-1 rgp160 (рекомбинантный экспрессирующийся с помощью бакуловируса в культуре клеток лепидоптер гликопротеин из ВИЧ-1 штамма IIIB) в концентрации 1 мкг/мышь, и одного из таурозидов в концентрации 100 мкг/мышь в забуференном изотоническом растворе хлорида натрия (ЗФР) в объеме 0,1 мл/мышь. Мышей реиммунизируют на 14,40 и 62 день этой же иммунизирующей смесью. Взятие крови и селезенок проводят на 10 день после последней реиммунизации. Контрольная группа мышей иммунизируется по той же схеме, но вместо таурозидов в качестве адьюванта используют фосфат алюминия в концентрации 100 мкг/мышь в 0,1 мл раствора антигена на ЗФР/мышь.
Наличие антител против rgp160 и rgp120 анализируют с помощью твердофазного иммуноферментного анализа (ИФА). Лунки микротитровочных 96-луночных пластин (МП) покрывают 100 мкл растворов rgp160 (rgp120)-1 мкг/мл в 0,1 М карбонатном буфере pH 9,6 и оставляют на ночь при комнатной температуре. Затем лунки промывают 4 раза ЗФР, содержащим 0,05% твин-20 (ЗФР-Т) в течение 10 минут. Мышиная сыворотка титруется на ЗФР-Т, содержащем 2% козьей сыворотки (ЗФР-Т-С) и добавляется в лунки по 100 мкл. Пластины инкубируют 1 час при 37o С и затем 5-кратно отмывают с помощью ЗФР-Т. Вносят в лунки меченый пероксидазой хрена конъюгат коза-анти IgG мыши (Bio-Rad), разведенный 1/4000 на ЗФР-по 50 мкл/лунку. Пластины инкубируют 1 час при 37 o С и затем 5-кратно отмывают ЗФР-Т. Вносят субстратный раствор (СР): 1,2 мг/мл ортофенилендиамина в 0,05 М цитратном буфере, pH 5,6, 0,01% перекиси водорода, - по 100 мкл/лунку. Через 30 минут реакцию останавливают 2,5 М серной кислотой (5 мкл/лунку). Оптическую плотность оценивают при длине волны 490 нм с помощью прибора Minireader (Dynatech). Контрольные лунки вместо иммунных сывороток обрабатывают сыворотками интактных мышей.
Результаты изучения выработки антигликопротеиновых антител приведены в табл. 1.
Пролиферацию клеток селезенки в ответ на введение антигена ВИЧ оценивают по следующей методике. После обескровливания стерильно извлекают селезенку и готовят суспензию отдельных клеток. Клетки трижды отмывают средой RPMI-1640 и культивируют в 96-луночных культуральных планшетах (КП) -1 500 000 клеток/мл в RPMI-1640 с добавлением 10% эмбриональной сыворотки коров (ЭСК), 100 мкг/м пеницилина и 100 МЕ/мл стрептомицина. Пролиферацию индуцируют добавлением 0,01 мкг/лунку rgp160. КП инкубируют от 6 дней при 37o С в 6% двуокиси углерода. Затем добавляют меченый тритием тимидин, и клетки оставляют на 4 часа, после чего их собирают и измеряют радиоактивность образцов с помощью сцинтилляционного счетчика (LKB Wallac 81 000). Включение радиоактивной метки определяют как среднее от трехкратного измерения культур, индекс стимуляции рассчитывают как частное от деления количества распадов/мин в экспериментальных образцах на количество распадов/мин в контроле (контрольная среда), результаты представлены в табл. 1.
Таким образом, таурозиды H2 и St-K стимулируют ВИЧ-1-специфический пролиферативный ответ клеток селезенки, хотя и уступают фосфату алюминия в способности активировать выработку антител к гликопротеинам ВИЧ-1 (кроме антителопродукции к rgp120 при использовании таурозида St-K). Таурозид 1 обладает более выраженной, чем у фосфата алюминия способностью стимулировать выработку антител к rgp160.
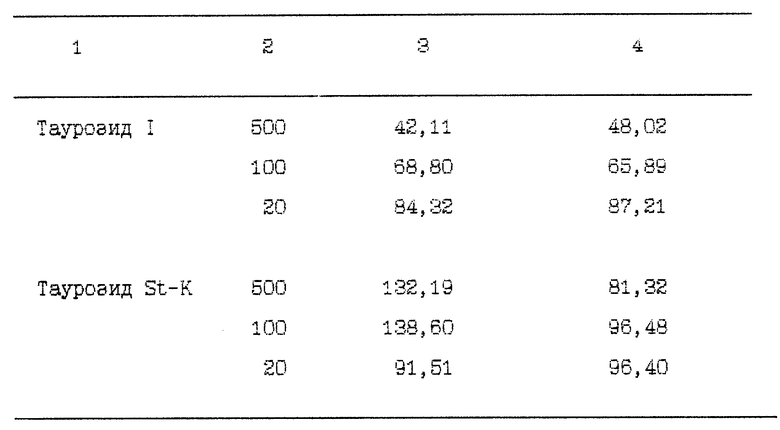
Пример 2. Культуральная жидкость (КЖ), содержания вирус ВИЧ-1 LAI (20 TCID50), вносится по 50 мкл в лунки 96-луночных КП одновременно с разведениями таурозидов (по 25 мкл/лунку) и клетками линии Jurkat-tat (200 мкл/лунку суспензии, содержащей 250 000 клеток/мл). Клетки вносятся в лунки и в последующем культивируются в среде RPMI-1640 с добавлением 10% ЭСК, 2 ммоль/л глютамина, 100 мкг/мл пенициллина и 100 МЕ/мл стрептомицина. Культивирование происходит при 37oС в атмосфере 5% двуокиси углерода. Таурозиды вносят в виде водных растворов в дозах, необходимых для достижения конечных концентраций 500, 100 и 20 мкг/мл. Клетки культивируют в присутствии вируса и таурозидов, или только вируса (контроли), учитывая результаты на 3 и 7 дни. Оценивают количество жизнеспособных клеток в тесте с трипановым синим, а также концентрацию сердцевинного антигена ВИЧ-1 р24 в КЖ. ВИЧ-1 р24 определяют в ИФА [7]. Для этого лунки МП покрывают кроличьими анти-ВИЧ-1 иммуноглобулинами (ИГ) - 100 мкл/лунку раствора с 20 мкг/мл ИГ в 0,1 М карбонатном буфере pH 9,6, в течение ночи при комнатной температуре. МП промывают 4 раза ЭФР-Т, после чего вносят либо стандартный антиген ВИЧ-1 р24, калиброванный по стандарту набора DuPont ELISA, либо исследуемые КЖ. МП инкубируют в течение ночи при комнатной температуре и 5-кратно отмывают ЗФР-Т. Затем вносят конъюгированные с пероксидазой хрена мышиные моноклональные антитела против ВИЧ-1 р24 (Nat. Baot. Lab., Швеция) и инкубируют МП 2 часа при 37o С. Далее следует 5-кратная отмывка ЭФР-Т и добавление СР с последующим перечнем операций, как описано в примере 1.
С целью сопоставления влияний, оказываемых изучаемыми препаратами на репликацию ВИЧ, используют расчет коэффициента изменения скорости репликации ВИЧ (КИСР):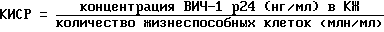
Значение КИСР в контролях (ВИЧ вносится в клетки без тауразидов) оценивают как 100% и все экспериментальные значения коэффициентов соотносят с этим значением, выражая в % (таблица 2).
Таким образом, таурозиды H2 и 1 во всех использованных концентрациях и во все сроки наблюдения демонстрируют тенденцию к ослаблению репликации ВИЧ-1 (кроме таурозида H2 в концентрации 20 мкг/мл-7 день, когда значение КИСР находилось на уровне контрольных значений). Таурозид St-K в концентрациях 100 и 500 мкг/мл на 3 день усиливал репликацию ВИЧ, однако к 7 дню наблюдения этот эффект исчезал.
Основными преимуществами предлагаемого способа являются, во-первых, доступность сырья и средств для получения используемых веществ, во-вторых, его высокие вакциноусиливающие свойства, и, главное - способность стимулировать гуморальные и клеточные иммунные ответы на целевые антигены без активации экспрессии ВИЧ из латентного состояния или состояния низкоуровневой репликации.
Источники информации
1. Veien N. K. , Hattel Т., Justesen O., Norholm A. Aluminium allergy //Contact dermatitis-1986. - Vol. 15. - P. 295-297.
2. Nagel J., Sveo D., Waters T., Fireman P. IgE synthesis in man: Development of specific IgE antibodies after immunisation with tetanus-diphtheria (Td) toxoids //J. Immunol. - 1977. - Vol. 118. - P. 334-341.
3. Levi M. , Ruden U. , Birx D., Loomis L., Redfield R., Lovgren K., Akerblom L. , Sandstrom E., Wahren B. Effects of adjuvants and multiple antigen peptides on humeral and cellular immune responses to gp160 of HIV-1 // Journal of AIDS. - 1993. - Vol. 6. - P. 855-864.
4. Schreck R. , Bevec D., Dukor P., Baeuerle P.A. Chedid L., Bahr G.M. Selection of a muramyl peptide based on its lack of activation of nuclear factor-kB as a potential adjuvant for AIDS vaccines//Clin.exp. Immunol. - 1992. - Vol. 90. - P. 188-193.
5. Bomford R., Stapleton M., Winsor S., Beesley J.E., Jessup E.A., Price K.R., Fenwick G.R. Adjuvanticity and ISCOM formation by structurally diverse saponins // Vaccine. - 1992. - Vol. 10. - P. 572-577.
6. Гришковец В.И., Кондратенко А.E., Толкачева Н.В., Шашков А.С., Чирва В.Я. Тритерпеновые глинозиды Hedera helix. I.//Химия природ. соедин. - 1994. - N 6. - С. 742-746.
7. Sundqvist V. -A. , Albert J., Ohlsson E., Hinkula J., Fenyo E.M., Wahren B. Human immunodeficiency virus type 1 p24 production and antigenic variation in tissue culture of isolated with various growth characteristics //J.Med.Virol. - 1989. - Vol. 29. - P. 170-175.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм гибридных культивируемых клеток животных MUS мUSсULUS L - продуцент моноклональных антител к М - белку вируса гриппа типа А | 1989 |
|
SU1652340A1 |
| ПРОТИВОГРИБКОВОЕ СРЕДСТВО "ТАУРАЗИД Е" И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2003 |
|
RU2241484C1 |
| Штамм гибридных культивируемых клеток животных MUS мUSсULUS-продуцент моноклональных антител к гемагглютинину вируса гриппа А/Ленинград/385/80 (H3N2) | 1987 |
|
SU1479510A1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА 1 ТИПА | 2006 |
|
RU2317107C2 |
| ВАКЦИНА ПРОТИВ ВИЧ/СПИД | 2010 |
|
RU2475264C2 |
| МОНОКЛОНАЛЬНОЕ АНТИИДИОТИПИЧЕСКОЕ АНТИТЕЛО Ab2, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ИСПОЛЬЗОВАНИЯ ПРОТИВ БОЛЕЗНЕЙ, СОПРОВОЖДАЮЩИХСЯ ЭКСПРЕССИЕЙ Lewis Y6 АНТИГЕНА, И ДЛЯ ОЧИСТКИ ВАРИАНТА BR55-2 АНТИТЕЛА, ЛЕЧЕБНО-ПРОФИЛАКТИЧЕСКИЙ СОСТАВ | 1993 |
|
RU2208642C2 |
| СПОСОБ ПОЛУЧЕНИЯ НОСИТЕЛЯ АНТИГЕНОВ НА ОСНОВЕ ЛИПИДОВ ИЗ МОРСКИХ МАКРОФИТОВ И ТРИТЕРПЕНОВОГО ГЛИКОЗИДА КУКУМАРИОЗИДА | 2005 |
|
RU2319506C2 |
| НОСИТЕЛЬ АНТИГЕНОВ | 2006 |
|
RU2322259C1 |
| Рекомбинантный химерный полипептид-иммуноген nTBI, обладающий способностью индуцировать антитела, нейтрализующие вирус иммунодефицита человека 1 типа, и предназначенный для использования в качестве компонента вакцины против ВИЧ-1 | 2016 |
|
RU2642258C1 |
| НОСИТЕЛЬ И АДЪЮВАНТ ДЛЯ АНТИГЕНОВ | 2006 |
|
RU2311926C2 |


Изобретение относится к иммунологии и вирусологии и может быть использовано для разработки вакцин. Способ включает введение в организм целевого антигена и адъюванта. При этом в качестве адъюванта вводят тритерпеновые гликозиды, выделенные из различных органов растений рода Hedera L. Способ позволяет стимулировать гуморальные и клеточные иммунные ответы на целевые антигены без активации экспрессии ВИЧ из латентного состояния или состояния низкоуровневой репликации, способ использует доступное сырье и средства для получения используемых веществ. 2 табл.
Способ стимулирования гуморального и клеточного иммунного ответа на целевой антиген, включающий введение в организм целевого антигена и адъюванта, отличающийся тем, что в качестве адъюванта вводят тритерпеновые гликозиды, выделенные из различных органов растений рода Hedera L. формулы

где Ara - L-арабинопиранозил, Glc - D-глюкопиранозил, Gal - D-галактопиранозил, GlcUA - D-глюкуронопиранозил, Rha - L-рамнопиранозил, Ac-ацетил.
| Bomford R | |||
| Et.al., Adjvanticity and ISCOM formmation by structurally diverse saponins, Vaccine, 1992, v.10, p.572-577 | |||
| Раплав для диффузионного насыщения | 1974 |
|
SU518533A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Машина для посадки высадков сахарной свеклы | 1958 |
|
SU126313A1 |
| СЕЛЕКТОР СКОРОСТЕЙ НЕЙТРАЛЬНЫХ ЧАСТИЦ | 0 |
|
SU255420A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Лекапосидзе Г.Е | |||
| Исследование тритерпеновых гликозидов | |||
| - Тбилиси: Мицниероба, 1982, с.35. | |||
