Описание изобретения Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности, к способу получения L-аминокислоты с использованием бактерии семейства Enterobacteriaceae, в которой ген ltaE инактивирован.
Описание предшествующего уровня техники
Традиционно L-аминокислоты в промышленном масштабе могут быть получены методом ферментации с использованием штаммов микроорганизмов, выделенных из природных источников, или их мутантов, специально модифицированных для того, чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (см., например, патент США 4278765). Указанные методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот и/или уменьшении чувствительности целевого фермента к обратному ингибированию продуцируемой L-аминокислотой (см., например, выложенную патентную заявку Японии №56-18596 (1981), WO 95/16042 или патенты США 5661012 и 6040160).
Известны различные штаммы, использующиеся для производства L-треонина методом ферментации. Это штаммы с увеличенными активностями ферментов, вовлеченных в биосинтез L-треонина (патенты США 5175107; 5661012; 5705371; 5939307; Европейский патент 219027), штаммы, устойчивые к некоторым химических реагентам, таким как L-треонин и его аналоги (заявка РСТ 0114525А1, европейская заявка 301572А2, патент США 5661012), штаммы, устойчивые к соединениям, таким как L-треонин и его аналоги (международная заявка WO 0114525 А1, Европейский патент 301572А2, патент США 5376538), штаммы, в которых устранена чувствительность целевого фермента к ингибированию продуцируемой аминокислотой или ее побочными продуктами по типу обратной связи (патенты США 5175107 и 5661012), штаммы с инактивированными ферментами системы деградации треонина (патенты США 5939307 и 6297031).
Известный штамм-продуцент L-треонина ВКПМ В-3996 (патенты США 5175107 и 5705371) является лучшим на сегодняшний день штаммом-продуцентом L-треонина. Для создания штамма ВКПМ В-3996 несколько мутаций и плазмида, описанная ниже, были введены в родительский штамм Е.coli K-12 (ВКПМ В-7). Мутантный ген thrA (мутация thrA442) кодирует белок аспартокиназа-гомосериндегидрогеназу I, который устойчив к ингибированию треонином по типу обратной связи. Мутантный ген ilvA (мутация ilvA442) кодирует белок треониндеаминазу, обладающую пониженной активностью, которая выражается в пониженном уровне биосинтеза изолейцина и в фенотипе с недостатком по изолейцину типа "leaky". В бактерии с мутацией ilvA442 транскрипция оперона thrABC не репрессируется изолейцином, что дает положительный эффект на продукцию треонина. Инактивация гена tdh приводит к предотвращению деградации треонина. В указанный штамм была введена генетическая детерминанта ассимиляции сахарозы (гены scrKYABR). Для увеличения экспрессии генов, контролирующих биосинтез треонина, в промежуточный штамм TDH6 была введена плазмида pVIC40, содержащая мутантный треониновый оперон thrA442BC. Количество треонина, накопленного в ходе ферментации этого штамма, достигало 85 г/л.
Ранее авторы настоящего изобретения получили исходя из Е.coli K12 мутант, содержащий мутацию thrR (упоминаемый здесь и далее как rhtA23), придающую устойчивость к высокой концентрации треонина и гомосерина на минимальной питательной среде (Astaurova, O.B. et al., Appl. Bioch. And Microbiol., 21, 611-616 (1985)). Фенотипически мутация выражается в повышении продукции L-треонина (патент СССР №974817), гомосерина, и глутамата (Astaurova, O.B. et al., Appl. Bioch. And Microbiol., 27, 556-561,1991, Европейская патентная заявка 1013765А) соответствующим штаммом-продуцентом Е.coli, таким как штамм ВКПМ-3996. Более того, авторы настоящего изобретения показали, что ген rhtA расположен на 18 минуте хромосомы Е.coli, рядом с опероном glnHPQ, кодирующим компоненты транспортной системы глутамина, и что ген rhtA идентичен открытой рамке считывания ORF1 (ген ybiF, номера нуклеотидов с 764 по 1651 в последовательности с инвентарным номером ААА2218541 в GenBank, gi:440181), расположенной между генами рехВ и отрХ. Ген, экспрессирующий белок, кодируемый ORF1, был назван геном rhtA (rht: resistance to homoserine and threonine - устойчивость к гомосерину и треонину). Кроме того, было установлено, что мутация rhtA23 является заменой А на G положении -1 относительно старт кодона ATG (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California August 24-29, 1997, abstract No.457; Европейская патентная заявка 1013765).
Один из подходов, используемых для повышения накопления L-треонина в питательной среде, - это предотвращение деградации указанной аминокислоты. L-алло-треонинальдолаза (ТА) катализирует один из возможных путей деградации L-треонина (Escherichia coli and Salmonella, Second Edition, Editor in Chief: F.C.Neidhardt, ASM Press, Washington D.C., 1996, pp.369-370). L-алло-треонинальдолаза, кодируемая геном ltaA, является низкоспецифичной; она может использовать в качестве субстрата как L-треонин, так и L-алло-треонин. Этот фермент нуждается в пиридоксальфосфате в качестве кофактора; также было показано, что указанный фермент чрезвычайно термостабилен и имеет высокую оптимальную температуру. L-алло-треонинальдолаза может также функционировать в альтернативном пути биосинтеза глицина, в том случае, когда основной путь его биосинтеза неактивен (Liu J.Q. et al, Eur J Biochem.,1998, Jul 1; 255(1):220-6). Но в настоящее время нет сообщений, описывающих использование инактивации гена ltaE для получения L-аминокислот.
Описание изобретения
Целями настоящего изобретения являются повышение продуктивности штаммов-продуцентов L-аминокислот и предоставление способа получения L-аминокислоты с использованием указанных штаммов.
Данная цель была достигнута путем обнаружения того факта, что инактивация гена ItaE может повысить продукцию L-аминокислоты, такой как L-треонин.
Настоящее изобретение предоставляет бактерию семейства Enterobacteriaceae, обладающую способностью к повышенной продукции аминокислоты, такой как L-треонин.
Целью настоящего изобретения является предоставление бактерии-продуцента L-аминокислоты семейства Enterobacteriaceae, при этом указанная бактерия модифицирована таким образом, что активность L-алло-треонинальдолазы уменьшена или полностью отсутствует.
Целью настоящего изобретения является предоставление бактерии-продуцента L-аминокислоты семейства Enterobacteriaceae, при этом указанная активность L-алло-треонинальдолазы уменьшена или отсутствует из-за инактивации гена ltaE.
Также целью настоящего изобретения является предоставление описанной выше бактерии, при этом указанная бактерия принадлежит к роду Escherichia.
Также целью настоящего изобретения является предоставление описанной выше бактерии, при этом указанной L-аминокислота является L-треонином.
Также целью настоящего изобретения является предоставление способа получения L-аминокислоты, который включает в себя:
- выращивание описанной выше бактерии в питательной среде с целью продукции и накопления L-аминокислоты в питательной среде, и
- и выделение указанной L-аминокислоты из культуральной жидкости.
Также целью настоящего изобретения является предоставление описанного выше способа, при этом указанной L-аминокислотой является L-треонин.
Более детально настоящее изобретение будет описано ниже.
1. Бактерия согласно настоящему изобретению.
Бактерия согласно настоящему изобретению - это бактерия - продуцент L-аминокислоты семейства Enterobacteriaceae, модифицированная таким образом, что ген ltaE инактивирован.
Согласно настоящему изобретению «бактерия - продуцент L-аминокислоты» означает бактерию, обладающую способностью к накоплению L-аминокислоты в питательной среде, когда указанная бактерия выращивается в указанной питательной среде. Способность к продукции L-аминокислоты может быть придана или улучшена путем селекции. Используемый здесь термин «бактерия-продуцент L-аминокислоты» также означает бактерию, которая способна к продукции L-аминокислоты и вызывает накопление L-аминокислоты в ферментационной среде в больших количествах по сравнению с природным или родительским штаммом Е.coli, таким, как штамм Е.coli К-12, и предпочтительно означает, что указанный микроорганизм способен накапливать в среде целевую L-аминокислоту в количестве не менее чем 0.5 г/л, более предпочтительно не менее чем 1.0 г/л. Термин "L-аминокислоты" включает в себя L-аланин, L-аргинин, L-аспарагин, L-аспартат, L-цистеин, L-глутаминовую кислоту, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин. Более конкретно L-треонин предпочтителен.
Семейство Enterobacteriaceae включает в себя бактерии, принадлежащие к родам Escherichia, Erwinia, Photorhabdus, Providencia, Salmonella, Serratia, Shigella, Yersinia и т.д. В частности, могут быть использованы бактерии, классифицированные как относящиеся к семейству Enterobacteriaceae, в соответствии с таксономией, используемой в базе данных NCBI (Национальный Центр Биотехнологической Информации - National Center for Biotechnology Information) http://www.ncbi.nlm.nih.gov/htbinpost/Taxonomy/wgetorg?mode=Tree&id=1236&lvl=3&keep=1&srchmode=1&unlock). Бактерия, принадлежащая к роду Escherichia, предпочтительна.
Термин «бактерия, принадлежащая к роду Escherichia» означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, может быть упомянута бактерия Escherichia coli (Е.coli).
Круг бактерий, принадлежащих к роду Escherichia, которые могут быть использованы в настоящем изобретении, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть включены в число бактерий согласно настоящему изобретению.
Термин "активность L-алло-треонинальдолазы" обозначает активность по катализу реакции превращения L-алло-треонина в глицин и ацетальдегид. L-треонин является альтернативным субстратом для L-алло-треонинальдолазы. Активность L-алло-треонинальдолазы может быть измерена с помощью метода, описанного, например, у Liu J.Q. et al (Eur J Biochem.,1998, Jul 1; 255(1):220-6).
Термин "активность L-алло-треонинальдолазы снижена" означает, что указанная удельная активность ниже, чем активность этого фермента в немодифицированном штамме, например, природном. Примеры такой модификации включают уменьшение количества молекул L-алло-треонинальдолазы на бактериальную клетку, снижение специфической активности молекулы L-алло-треонинальдолазы, и так далее. Термин "активность L-алло-треонинальдолазы отсутствует" обозначает, что в модифицированном штамме указанная активность не детектируется. Отсутствие активности L-алло-треонинальдолазы является одним из примеров снижения указанной активности. Кроме того, природным штаммом, используемым для сравнения, может быть, например, штамм Escherichia coli K-12. Согласно настоящему изобретению, в результате снижения или отсутствия внутриклеточной активности L-алло-треонинальдолазы может быть повышено количество накопленного L-треонина в среде из-за предотвращения деградации накопленного L-треонина.
Снижение активности L-алло-треонинальдолазы в бактериальной клетке может быть достигнуто за счет ослабления экспрессии гена, кодирующего L-алло-треонинальдолазу.
В качестве гена, кодирующего L-алло-треонинальдолазу из Escherichia coli, известна нуклеотидная последовательность гена ltaE (номера нуклеотидов с 908517 по 907516 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi:16128838; SEQ ID: 3). Ген ltaE расположен на хромосоме штамма Е.coli K-12 между открытой рамкой считывния ybjT ORF и геном рохВ. Следовательно, ген ltaE может быть получен с помощью ПЦР полимеразная цепная реакция; ссылка на White, T.J. et al., Trends Genet., 5, 185 (1989)) с использованием праймеров, синтезированных на основе нуклеотидной последовательности данного гена. Ген ltaE также имеет другие названия: b0870, EG13690, G6455, ltaA и ybjU. Гены, кодирующие L-алло-треонинальдолазу в других микроорганизмах, могут быть получены сходным образом.
Термины "ген yafA инактивирован" означает, что целевой ген модифицирован таким образом, что такой модифицированный ген кодирует мутантный белок со сниженной активностью, или что такой модифицированный ген кодирует полностью неактивный белок. Также возможно, что естественная экспрессия модифицированного участка ДНК невозможна из-за делеции целевого гена или его части, сдвига рамки считывания данного гена или введения missense/nonsense мутации или модификации прилегающей к гену областей, которые включают последовательности, контролирующие экспрессию гена, такие как промотор(ы), энхансер(ы), аттенуатор(ы), сайт(ы) связывания рибосомы, и т.д.
Инактивация указанного гена может быть произведена традиционными методами, такими как мутагенез с использованием УФ излучения или обработка нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, инактивации гена с помощью гомологичной рекомбинации, или/и инсерционно-делеционного мутагенеза (Yu, D. et al., Proc. Natl. Acad. Sci. USA, 2000, 97:12: 5978-83); (Datsenko K.A. and Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97:12: 6640-45), также называемого "Red-зависимая интеграция".
Методами получения плазмидной ДНК, разрезания и лигирования ДНК, трансформации, выбора олигонуклеотидов в качестве праймеров и подобные им могут являться обычные методы, хорошо известные специалисту в данной области. Эти методы описаны, например, в книге Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Бактерия-продуцент L-аминокислоты.
Бактерии, обладающие способностью к продукции ароматических или неароматических L-аминокислот, могут быть использованы в качестве родительских штаммов, для дальнейшей модификации с инактивацией гена ltaE.
Бактерия согласно настоящему изобретению может быть получена путем инактивации гена ltaE в бактерии, наследственно обладающей способностью к продукции L-аминокислот. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания способности к продукции L-аминокислот бактерии, в которой ген ItaE уже инактивирован.
Бактерия-продуцент L-треонина.
Примеры родительского штамма, используемого для получения бактерии-продуцента L-треонина согласно настоящему изобретению, включают, но не ограничены штаммами - продуцентами треонина, принадлежащими к роду Escherichia, такими как штамм Е.coli TDH-6/pVIC40 (ВКПМ В-3996) (патенты США 5175107 и 5705371), штамм Е.coli NRRL-21593 (патент США 5939307), штамм Е.coli FERM ВР-3756 (патент США 5474918), штаммы Е.coli PERM ВР-3519 и FERM BP-3520 (патент США 5376538), штамм Е.coli MG442 (Гусятинер и др., Генетика, 14, 947-956 (1978)), штаммы Е.coli VL643 и VL2055 (Европейская патентная заявка ЕР1149911А) и подобные им.
Штамм TDH-6 является дефицитным по гену thrC, способен ассимилировать глюкозу и содержит ген ilvA с мутацией типа "leaky". Указанный штамм содержит мутацию в гене rhtA, которая обуславливает устойчивость к высоким концентрациям треонина и гомосерина. Штамм В-3996 содержит плазмиду pVIC40, которая была получена путем введения в вектор, производный от вектора RSF1010, оперона thrA*BC, включающего мутантный ген thrA. Этот мутантный ген thrA кодирует аспартокиназа-гомосериндегидрогеназу I, у которой существенно снижена чувствительность к ингибированию треонином по типу обратной связи. Штамм В-3996 был депонирован 19 ноября 1987 года во Всесоюзном научном центре антибиотиков (РФ, 113105 Москва, Нагатинская ул., 3-А) с инвентарным номером РИА 1867. Указанный штамм также был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 113545 Москва, 1-й Дорожный проезд, 1) с инвентарным номером В-3996.
Предпочтительно, чтобы бактерия согласно настоящему изобретению была далее модифицирована для того, чтобы иметь повышенную экспрессию одного или нескольких следующих генов:
- мутантного гена thrA, кодирующего аспартокиназа-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи;
- гена thrB, кодирующего гомосеринкиназу;
- гена thrC, кодирующего треонинсинтазу;
- гена rhtA, предположительно кодирующего трансмембранный белок.
- гена asd, кодирующего аспартат-Р-семиальдегиддегидрогеназу; и
- гена aspC, кодирующего аспартатаминотрансферазу (аспартаттрансаминазу).
Последовательность гена thrA, кодирующего аспартокиназа-гомосериндегидрогеназу I из Escherichia coli, известна (номера нуклеотидов с 337 по 2799 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrA расположен на хромосоме штамма Е, coli K-12 между генами thrL и thrB. Последовательность гена thrB, кодирующего гомосеринкиназу из Escherichia coli, известна (номера нуклеотидов с 2801 по 3733 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrB расположен на хромосоме штамма Е.coli K-12 между генами thrA и thrC. Последовательность гена thrC, кодирующего треонинсинтазу из Escherichia coli, известна (номера нуклеотидов с 3734 по 5020 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrC расположен на хромосоме штамма Е.coli K-12 между геном thrB и открытой рамкой считывания уааХ. Все три указанных гена функционируют как один треониновый оперон.
Мутантный ген thrA, кодирующий аспартокиназу-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи, так же, как и гены thrB и thrC, могут быть получены в виде единого оперона из хорошо-известной плазмиды pVIC40, которая представлена в штамме-продуценте Е.coli ВКПМ В-3996. Плазмида pVIC40 подробно описана в патенте США 5705371.
Ген rhtA расположен на 18 минуте хромосомы Е.coli рядом с опероном glnHPQ, кодирующим компоненты транспортной системы глутамина, таким образом, ген rhtA идентичен открытой рамке считывания ORF1 (ген ybiF, номера нуклеотидов с 764 по 1651 в последовательности с инвентарным номером ААА2218541 в GenBank, gi:440181), расположенной между генами рехВ и ompX. Участок, экспрессирующий белок, кодируемый ORF1, был назван геном rhtA (rht: resistance to homoserine and threonine - устойчивость к гомосерину и треонину). Кроме того, было установлено, что мутация rhtA23 является заменой А на G положении - 1 относительно старт кодона ATG (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California August 24-29, 1997, abstract No.457; Европейская патентная заявка 1013765).
Нуклеотидная последовательность гена asd из E.coli уже известна (номера нуклеотидов с 3572511 по 3571408 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi: 16131307), и этот ген может быть получен с помощью ПЦР (полимеразная цепная реакция; ссылка на White, T.J. et al., Trends Genet., 5, 185 (1989)) с использованием праймеров, синтезированных на основе нуклеотидной последовательности указанного гена. Гены asd из других микроорганизмов могут быть получены сходным способом.
Также нуклеотидная последовательность гена aspC из Е.coli уже известна (номера нуклеотидов с 983742 по 984932 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi: 16128895), и этот ген может быть получен с помощью ПЦР. Гены aspC из других микроорганизмов могут быть получены сходным способом.
2. Способ согласно настоящему изобретению.
Способом согласно настоящему изобретению является способ получения L-аминокислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-аминокислоты в питательной среде, и выделения L-аминокислоты из культуральной жидкости.
Согласно настоящему изобретению выращивание, выделение и очистка L-аминокислоты из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием бактерии.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от характера ассимиляции используемого микроорганизма могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин и дрожжевой экстракт. При необходимости в среду также могут быть добавлены дополнительные компоненты. Например, если микроорганизму для роста требуется L-аминокислота (ауксотрофность по L-аминокислоте), в питательную среду может быть добавлено необходимое количество указанной L-аминокислоты.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. рН среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и кристаллизации.
Краткое описание чертежей.
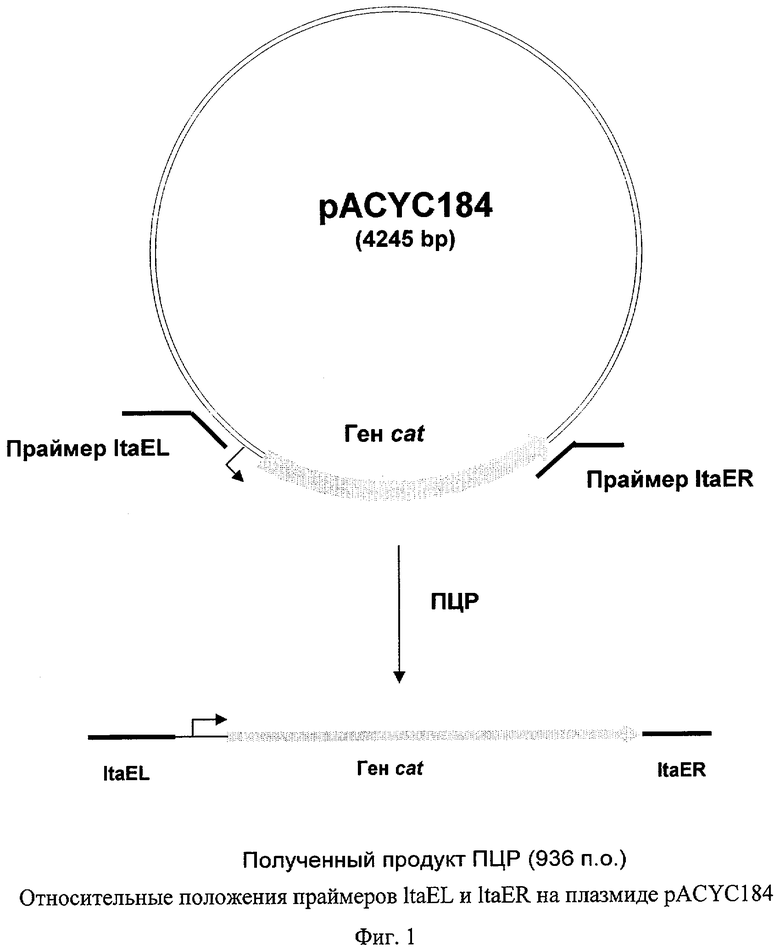
На Фиг.1 изображены относительные положения праймеров ltaEL и ltaER на плазмиде pACYC184, используемой для амплификации гена cat.
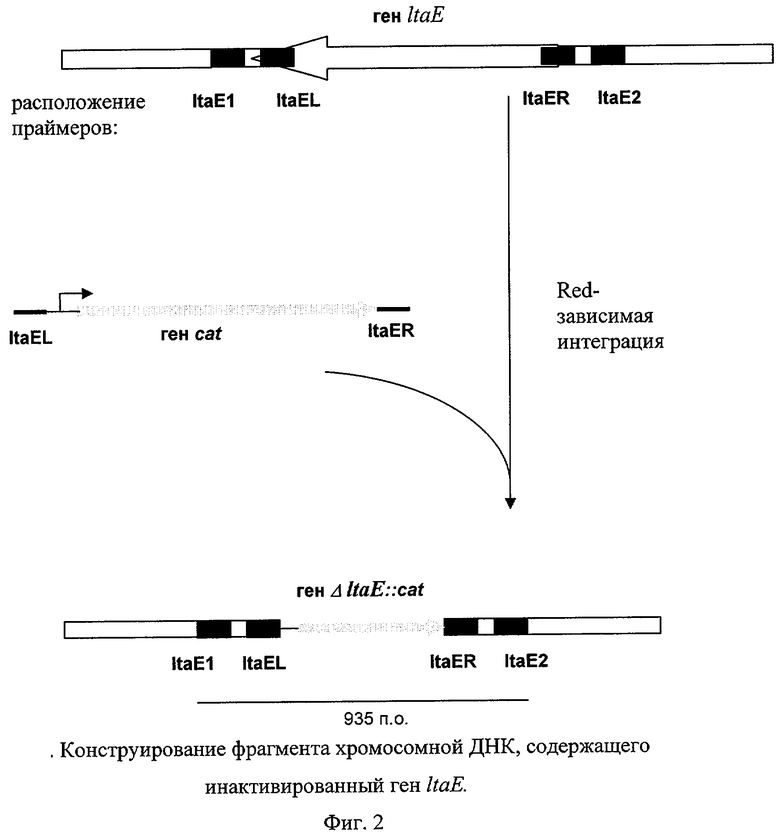
На Фиг.2 изображено конструирование фрагмента хромосомной ДНК, содержащего инактивированный ген ltaE.
Примеры.
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие не ограничивающие настоящее изобретение Примеры.
Пример 1. Конструирование штамма с инактивированньм геном ltaE.
1. Делеция гена ltaE.
Штамм с делетированным геном ltaE был получен с помощью метода, исходно разработанного Datsenko, К.А. и Wanner, B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97(12), 6640-6645), известного как "Red-зависимая интеграция". В соответствии с указанной методикой были синтезированы ПЦР праймеры ltaEL (SEQ ID NO: 1) и ltaER (SEQ ID NO: 2), гомологичные как областям хромосомы, прилегающих к гену ltaE, так и гену, придающему устойчивость к антибиотику на плазмиде, используемой в качестве матрицы. В качестве такой матрицы для ПЦР использовалась плазмида pACYC184 (NBL Gene Sciences Ltd., UK) (инвентарный номер Х06403 в базе данных GenBank/EMBL). Использовался следующий температурный профиль для ПЦР: денатурация при 95°С в течение 3 мин; два первых цикла: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; и последующие 25 циклов: 30 сек при 95°С, 30 сек при 54°С, 40 сек при 72°С; и заключительная полимеризация: 5 мин при 72°С.
Полученный продукт ПЦР - фрагмент длиной 936 п.о. (Фиг.1, SEQ ID NO: 5) был очищен в агарозном геле и мог быть использован для электропорации в штамм Е.coli MG1655(ATCC 700926), содержащий плазмиду pKD46 с термочувствительным репликоном. Плазмида pKD46 (Datsenko, K.A. and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 2000, 97:12:6640-45) содержит фрагмент ДНК фага λ (инвентарный номер последовательности J02459 в базе данных GenBank) длиной 2,154 нуклеотида (31088-33241), а также содержит гены λ Red гомологичной системы рекомбинации (γ, β, ехо гены) под контролем промотора РaraB; индуцируемого арабинозой. Плазмида pKD46 необходима для интеграции продукта ПЦР в хромосому штамма MG1655.
Электрокомпетентные клетки были получены следующим образом: ночную культуру штамма Е.coli MG1655 выращивали при 30°С в LB среде с добавкой ампициллина (100 мг/л), развели в 100 раз при помощи 5 мл среды SOB (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)), содержащей ампициллин и L-арабинозу (1 мМ). Полученную культуру растили с перемешиванием при 30°С до достижения OD600≈0.6, после чего делали клетки электрокомпетентными путем концентрации в 100 раз трехкратного отмывания ледяной деионизированной Н2О. Электропорацию проводили с использованием 70 мкл клеток и ≈100 нг продукта ПЦР. После электропорации клетки инкубировали с 1 мл среды SOC (Sambrook et al, "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2.5 часов, после чего высевали на чашки с L-агаром и выращивали при 37°С для отбора CmR рекомбинантов. Затем для удаления плазмиды pKD46, проводили 2 пассажа на L-агаре с Cm при 42°С и полученные колонии проверяли на чувствительность к ампициллину.
2. Подтверждение делеции гена ltaE с помощью ПЦР.
Мутанты с делетированным геном ItaE и содержащие ген устойчивости к Cm были проверены с помощью ПЦР. Локус-специфичные праймеры ltaE1 (SEQ ID NO: 6) и ltaE2 (SEQ ID NO: 7) использовались для проверки делеции с помощью ПЦР. Использовался следующий температурный профиль для ПЦР проверки: денатурация при 94°С в течение 3 мин; профиль для 30 циклов: 30 сек при 94°С, 30 сек при 54°С, 1 мин при 72°С; заключительный шаг: 7 мин при 72°С. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток родительского штамма ltaE+ MG1655, составляет 1002 п.о. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток мутантного штамма MG1655 ΔltaE::cat, составляет 935 п.о. (Фиг.2).
Пример 2. Продукция L-треонина штаммом Е.coli В-3996-ΔltaE.
Для проверки эффекта делеции гена ltaE на продукцию треонина фрагменты ДНК из хромосомы описанного выше штамма Е.coli MG1655 ΔltaE::cat могут быть перенесены в штамм-продуцент треонина Е.coli VKPM В-3996 Р1 трансдукцией (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY).
Оба штамма - родительский В-3996 и полученный B-3996 ΔltaE::cat, могут выращиваться в течение 18-24 часов при 32°С на чашках с L-агаром, содержащих хлорамфеникол (30 мкг/мл). Для получения посевной культуры каждый из штаммов может быть выращен на роторной качалке (250 об/мин) при 32°С в течение 18 часов в пробирках 20×200 мм, содержащих 2 мл L-бульона с 4% сахарозы. Затем в ферментационную среду может быть внесено 0.21 мл (10%) посевного материала. Ферментация может проводиться в 2 мл минимальной ферментационной среды в пробирках 20×200 мм. Клетки могут выращиваться в течение 48 часов при 32°С на роторной качалке - 250 об/мин.
После выращивания количество накопленного в среде L-треонина может быть определено с помощью бумажной хроматографии, при этом используется следующий состав подвижной фазы: бутанол : уксусная кислота : вода = 4:1:1 (v/v). Раствор (2%) нингидрина в ацетоне может быть использован для визуализации. Пятна, содержащие L-треонина, могут быть вырезаны, L-треонин может быть элюирован 0.5% водным раствором CdCl2, после чего количество L-треонина может быть оценено спектрофотометрическим методом при длине волны 540 нм.
Возможный состав ферментационной среды (г/л):
Глюкозу и сульфат марганца стерилизуют отдельно. СаСО3 стерилизуют сухим жаром при 180°С в течение 2 часов. рН доводят до 7.0. Антибиотик добавляют в среду после стерилизации.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.





| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
| БАКТЕРИЯ РОДА ESCHERICHIA - ПРОДУЦЕНТ L-ТРЕОНИНА И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА L-ТРЕОНИНА С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2515095C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2003 |
|
RU2275424C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТРЕОНИНА (ВАРИАНТЫ) | 2002 |
|
RU2244007C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yfeH | 2006 |
|
RU2337959C2 |
| СПОСОБ ПОЛУЧЕНИЯ НЕАРОМАТИЧЕСКИХ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН csrA | 2005 |
|
RU2311453C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yahN | 2007 |
|
RU2392322C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA - ПРОДУЦЕНТ L-ТРЕОНИНА И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2004 |
|
RU2288264C2 |
Изобретение относится к биотехнологии и представляет собой способ получения L-треонина с использованием бактерии, принадлежащей к роду Escherichia в которой ген ltaE инактивирован. Изобретение позволяет получать L-треонин с высокой степенью эффективности. 2 н. и 1 з.п. ф-лы, 2 ил.
выделение L-треонина из культуральной жидкости.
| US 5939307, 17.08.1999 | |||
| RU 2002123882, 20.04.2004 | |||
| US 6297031, 02.10.2001. |

