Предпосылки изобретения
Перечень федерального финансирования
Настоящее изобретение сделано, отчасти, с использованием фондов Федерального правительства в рамках гранта P01-DK-26741. В этой связи Федеральное правительство имеет определенные права на данное изобретение.
Область изобретения
Настоящее изобретение относится в основном к области нейроэндокринологии и химии нейропептидов. Более конкретно, настоящее изобретение относится к белковым факторам, участвующим в регуляции нейроэндокринной и паракринной реакции на стресс. И наиболее конкретно, настоящее изобретение раскрывает пептид, относящийся к кортикотропин-рилизинг-фактору, обозначенному как урокортин III.
Описание связанной области техники
Кортикотропин-рилизинг-фактор (CRF) и относящиеся к его семейству пептиды характеризуются, в первую очередь, их способностью к регуляции системы гипоталамус-гипофиз-надпочечники оси (ГГН) в спокойных и стрессовых условиях (1, 2). Кортикотропин-рилизинг-фактор (CRF) представляет собой пептид, состоящий из 41 аминокислоты, который был впервые выделен из гипоталамуса овцы (3) и в отношении которого было показано, что он играет важную роль в регуляции системы гипофиз-надпочечники, а также в осуществлении эндокринной, автономной и поведенческой реакций на стресс (4). Семейство CRF-нейропептидов также включает структурно родственные пептиды млекопитающих и не млекопитающих, такие как урокортин (Ucn), состоящий из 40 аминокислот, пептид, который был первоначально идентифицирован в мозге крысы (5), уротензин I (Uro) рыб (6) и соваджин (Svg) земноводных (7).
Была высказана гипотеза о том, что представители CRF-семейства участвуют в нейроэндокринной и паракринной реакциях, протекающих в разных тканях. Как было показано, представители CRF-семейства, в дополнение к их воздействию на гипофиз и центральную нервную систему, способны модулировать функции сердечно-сосудистой системы и желудочно-кишечного тракта, а также процессы воспаления у млекопитающих, приводя к интеграции эндокринной, автономной и поведенческой реакций на стрессовые факторы. Указанные пептиды могут также участвовать в контроле аппетита, возбуждения и познавательных функций. В результате длительного воздействия факторов стресса могут возникать тяжелые психологические и физиологические последствия, такие как расстройства, связанные с беспокойством, нервная анорексия, дисфункция желудочно-кишечного тракта и меланхолическая депрессия.
Представители CRF-семейства оказывают свое биологическое действие посредством специфического связывания с CRF-рецепторами с высокой степенью аффинности (8, 9). CRF-рецепторы представляют собой связанные с G-белком рецепторы, которые действуют через аденилатциклазу и структурно связаны с семейством рецептора секретина. Указанное семейство также включает рецепторы GRF (соматотропин-рилизинг-фактора), VIP (вазоактивного интестинального пептида), PTH (паратгормона) и кальцитонина. CRF-рецепторы происходят из двух обособленных генов, CRF-рецептора типа 1 (CRF-R1) (10-12) и CRF-рецептора типа 2 (CRF-R2) (13-15). CRF-R1 и CRF-R2 характеризуются разной фармакологией и различаются по анатомической локализации (16). Ген CRF-рецептора типа 1 (CRF-R1) содержит 13 экзонов; при этом было обнаружено несколько вариантов сплайсинга указанного рецептора. CRF-R1 локализуется в мозге и обнаружен в сенсорных и двигательных сайтах (17). Рецептор грызунов типа 2α (CRF-R2α) локализуется в латеральной перегородке, в срединной вентральной части гипоталамуса, в ядре одиночного пути и в заднем ядре шва, которые являются зонами очень малой экспрессии CRF-R1 или полного ее отсутствия (18). Рецептор грызунов типа 2β (CRF-R2β) обнаружен преимущественно в периферических участках, включая сердце, кровеносные сосуды, желудочно-кишечный тракт, придаток яичка, легкие и кожу (9, 19).
Фармакология двух типов рецепторов отличается тем, что CRF характеризуется умеренной аффинностью в отношении CRF-R2 [Ki=5-100 нМ], однако обладает высокой аффинностью в отношении CRF-R1 [Ki=1-2 нМ]. Другие родственные пептиды, такие как уротензин карпа, соваджин лягушки и урокортин характеризуются высокой аффинностью в отношении как CRF-R1, так и CRF-R2. Нокаутированные по CRF-R2 мыши, демонстрирует усугубление похожего на тревогу состояния, вызываемого гиперчувствительностью к стрессовым факторам (5, 20).
В последнее время исследования доступных баз данных генома человека привели к идентификации участка, характеризующегося существенной гомологией последовательностей с семейством CRF-нейропептидов. Была амплифицирована и секвенирована полная последовательность человека. Однако последовательность человека не содержит консенсус-сайт протеолитического расщепления, наличие которого позволило бы осуществить С-концевой процессинг пептида, и в этой связи она была обозначена как последовательность урокортин-родственного пептида (URP). С использованием гомологичных праймеров, выведенных на основе последовательности генома человека, из всей поли-(А+)-РНК мозга мыши была выделена к-ДНК, которая кодирует предполагаемый пептид, состоящий из 38 аминокислот, обозначенный как урокортин II, который характеризуется структурным родством с другими известными представителями из семейства млекопитающих, CRF и урокортином (Ucn). Вопрос о том, представляет ли человеческий урокортин-родственный пептид ортолог мышиного Ucn II, остается открытым до получения информации об идентификации дополнительных мышиных генов. Ucn II селективно связывается с CRF-рецептором типа 2 (CRF-R2) и практически не проявляет заметной активности в отношении CRF-R1. Транскрипты, кодирующие Ucn II, экспрессируются в отдельных участках ЦНС грызунов, включая относящиеся к стрессовым группы клеток в гипоталамусе (паравентрикулярные и дугообразные ядра) и стволе мозга (голубоватое место). Указанные открытия позволяют идентифицировать Ucn II в качестве нового представителя семейства CRF-нейропептидов, которые экспрессируются централизованно и селективно связываются с CRF-R2. Первоначальные функциональные исследования соответствуют представлению об участии Ucn II в центральном автономном контроле и в контроле аппетита, но не в генерализованной активации поведенческих функций (21).
На достигнутом уровне техники отсутствуют данные об обнаружении гена человеческого урокортина-III и соответствующего белка и их использовании. Настоящее изобретение отвечает на потребность и желания такого рода, давно назревшие на достигнутом уровне техники.
Краткое описание изобретения
Человеческий урокортин, урокортин-III (Ucn-III), имеющий гомологию с известными урокортинами из иглобрюхих рыб, был идентифицирован с использованием доступной базы данных человеческого генома. С помощью последовательности человеческого гена был выделен ортолог мыши. Настоящее изобретение относится к указанным новым генам и их использованию.
В одном аспекте настоящее изобретение относится к выделенному и очищенному белку урокортину III, который может представлять собой либо мышиный, либо человеческий урокортин III. Мышиный белок предпочтительно имеет аминокислотную последовательность SEQ ID No.5, которая получена из пептида-предшественника с последовательностью SEQ ID No.4. Человеческий белок предпочтительно имеет аминокислотную последовательность SEQ ID No.3, полученную из пептида-предшественника с последовательностью SEQ ID No.2.
Настоящее изобретение также относится к человеческому урокортину III, содержащему одну или более замен в аминокислотной последовательности, происходящих из аминокислотной последовательности мыши. Последовательность мышиного урокортина III (SEQ ID No.5) отличается от последовательности человеческого урокортина III (SEQ ID No.3) четырьмя аминокислотами, а именно Ile14, Asp19, Lys27 и Gln33. Замещение остатка Leu14 в человеческом белке изолейцином рассматривается как особенно полезное.
Настоящее изобретение также относится к фармацевтической композиции, включающей белок урокортина III, и к методу лечения патофизиологического состояния с использованием указанной фармацевтической композиции. Указанная фармацевтическая композиция может быть введена для активации рецептора CRF-R2 для лечения патофизиологического состояния, такого как высокая температура тела, нарушение аппетита, застойная сердечная недостаточность, сосудистые заболевания, стресс и беспокойство.
Настоящее изобретение также относится к модификации белка урокортина III. N-конец урокортина III может быть удлинен за счет дополнительных аминокислот или пептидов, таких как треонин-лизин (предшествующие два остатка в белке-предшественнике), D-тирозин, L-тирозин, D-тирозин-глицин или L-тирозин-глицин. Кроме того, один или более остатков метионина в урокортине III, таких как остатки в позициях 12 и 35 последовательности SEQ ID No.3, могут быть замещены остатками Nle. Альтернативно N-конец может быть удлинен с помощью D-иодтирозина, L-иодтирозина, D-иодтирозин-глицина и L-иодтирозин-глицина, а остатки метионина в положении 12 и 35 могут быть замещены Nle. Остатки иодтирозина могут быть помечены с помощью 125I.
Предполагаются также другие замещения аминокислотными остатками, консервативными для другого урокортина и урокортин-родственных белков, которые отличаются у урокортина III. Такие аналоги урокортина могут включать урокортин III с замещением одной или более аминокислот, выбранных из группы, состоящей из Ile3, Nle3, CαMe-Leu3, Ile5, Nle5, CαMe-Leu5, Leu7, Nle7, Thr8, Ile9, Phe9, Gly10, His10, Leu11, Nle11, Leu12, Nle12, Arg13, Gln13, Nle14, CαMe-Leu14, Nle15, CαMe-Leu15, Leu16, Nle16, Glu17, Asp17, Arg20, Nle24, CαMe-Leu24, Arg32, Ile34, Nle34, CαMe-Leu34, Leu35, Nle35, Asp36, Glu36 и Val38.
Настоящее изобретение также относится к антагонисту рецептора CRF-R2, включающему белок урокортина III или аналог урокортина III, причем первые пять-восемь N-концевых аминокислот указанного белка подвергаются делеции. Указанный антагонист может быть включен в фармацевтическую композицию и может использоваться для лечения застойной сердечной недостаточности, сосудистых заболеваний, дисфункции желудочно-кишечного тракта и головных болей при мигрени или как ингибитор ангиогенеза.
В еще одном варианте осуществления настоящего изобретения урокортин III может быть также модифицирован с получением соединения, содержащего флуоресцентную метку или комплексообразующий агент для радионуклидов. Полученный меченый урокортин III может использоваться для идентификации клеток, экспрессирующих рецепторы урокортина III. Альтернативно урокортин III может быть связан с молекулой токсина.
В еще одном варианте осуществления настоящего изобретения предлагается антитело, направленное против урокортина III. В предпочтительном варианте осуществления изобретения указанное антитело представляет собой моноклональное антитело. Антитело может быть конъюгировано с молекулярной меткой, такой как флуоресцентная метка, фотоаффинная метка или радиоактивные маркеры. Альтернативно указанные антитела могут быть конъюгированы с цитотоксичным соединением с образованием иммунотоксина.
Краткое описание чертежей
С тем чтобы указанные выше особенности, достоинства и цели настоящего изобретения, а также другие его характеристики, которые будут описаны ниже, были достигнуты и поняты во всей полноте, более конкретное описание изобретения, которое было в общем виде приведено выше, может быть получено посредством ссылок на конкретные варианты его осуществления, которые проиллюстрированы на прилагаемых чертежах. Указанные чертежи образуют часть описания. Следует, однако, отметить, что прилагаемые чертежи иллюстрируют предпочтительные варианты осуществления изобретения, и их не следует рассматривать как ограничивающие его область.
На фиг.1 приведены нуклеотидные и пептидные последовательности человеческого урокортина III.
На фиг.2А приведена предсказанная аминокислотная последовательность, кодирующая человеческий Ucn III, а на фиг.2В показана аминокислотная последовательность мышиного Ucn III. Аминокислоты пронумерованы, начиная с инициирующего метионина. Предположительный регион, кодирующий зрелый пептид, указан внутри прямоугольника. Полные нуклеотидные последовательности были депонированы в банк генов (Genbank, номер доступа № AF 361943 для человеческого Ucn III и AF361944 для мышиного Ucn III).
На фиг.2С показано выравнивание предположительных участков зрелого пептида человеческого и мышиного Ucn III с гомологичными урокортинами иглобрюхих рыб, человеческим и мышиным Ucn II, человеческим и овечьим CRF, уротензином (Uro) иглобрюхих рыб, соваджином лягушки, человеческим и мышиным Ucn. Остатки, идентичные последовательности человеческого Ucn III, заключены в прямоугольник. Выравнивание было осуществлено с использованием метода Clustal Method of Megalign в DNASTAR. ▪, обозначает место амидирования (предположительное для человеческого Ucn II).
На фиг.2D показано филогенетическое древо, на котором сгруппированы человеческий и мышиный Ucn III с урокортинами иглобрюхих рыб и человеческим и мышиным Ucn II. Более отдаленные родственные группы включают овечий и человеческий CRF, человеческий и мышиный Ucn, Uro иглобрюхих рыб и соваджин лягушки. Шкала внизу древа соответствует измерению расстояний в последовательностях. Филогенетическое древо получено с использованием DNASTAR.
Фиг.3А и 3В иллюстрируют влияние Ucn-родственных пептидов на накопление ц-АМФ в клеточной линии, экспрессирующей CRF-R2β (фиг.3А), и в первичных клетках передней части гипофиза крыс (фиг.3В). На фиг.3А показаны результаты, полученные на клетках A7r5 гладкой мышцы аорты крысы. ЕС50 (ЭК50): mUcn II (Ucn II мыши) - 0,18 нМ; mUcn III (Ucn III мыши) - 3,7 нМ; hUcn III(Ucn III человека) - 80,9 нМ. На фиг.3В показаны результаты, полученные на первичных клетках передней части гипофиза крысы, которые были переведены в культуру и подвергнуты стимулированию различными пептидами в течение 45 минут. ЕС50: rUcn (Ucn крысы) - 2,3 нМ; hUcn II - 1 мкМ*; mUcn II - 0,75 мкМ* (*: установлено с использованием значения выхода на плато для rUcn).
На фиг.4 показана экспрессия м-РНК мышиного Ucn III в мозге и периферических тканях. Показана репрезентативная картина теста на защиту м-РНК Ucn III от действия РНказы. Суммарная РНК, выделенная из каждой перечисленной ткани, была гибридизована с антисмысловым относительно мышиного Ucn III зондом и антисмысловым относительно мышиного GAPDH зондом. Защищенные фрагменты разделяли на 6% полиакриламидном геле с мочевиной. Сокращения: BnST - ложе ядра терминальной полоски.
На фиг.5A-5F показана гистохимическая локализация при гибридизации м-РНК Ucn III в мозге крыс.Положительный гибридизационный сигнал был наиболее заметен в трех участках вентральной части переднего мозга. Они включают медиальное предоптическое ядро (фиг.5А, 5В), растральную перифорникальную область, которая включает зоны, находящиеся сбоку от паравентрикулярного ядра (фиг.5С), заднюю часть ложа ядра терминальной полоски (фиг.5D) и радиальное миндалевидное ядро (фиг.5Е). В стволе мозга положительные гибридизационные сигналы обнаруживаются в основном в верхнем параоливном ядре (фиг.5F). Сокращения: 3V - третий желудочек; ac - передняя спайка; BSTp - задняя часть ложа ядра терминальной полоски; fx - свод; MeA - медиальное ядро миндалины; MePO - медиальное предоптическое ядро; OVLT - сосудистый орган (терминальной пластинки); opt - зрительный тракт; PVH - паравентрикулярное ядро гипоталамуса; SPO - верхнее параоливное ядро; Tz - ядро трапециевидного тела. Деление шкалы = 50 мкм.
Подробное описание изобретения
В соответствии с настоящим изобретением могут использоваться традиционные методики молекулярной биологии, микробиологии и технологии рекомбинантной ДНК, которыми владеют специалисты в данной области техники. Указанные методики наиболее полно описаны в литературе. См., например, Maniatis, Fritsch & Sambrook, "Molecular Cloning: A Laboratory Manual" (1982); "DNA Cloning: A Practical Approach," Volumes I and II (D.N.Clover ed. 1985); "Oligonucleotide Synthesis" (M.J.Gait ed. 1984); "Nucleic Acid Hybridization" [B.D.Hames & S.J.Higgins Eds. (1985)]; "Transcription and Translation" [B.D.Hames & S.J.Higgins Eds. (1984)]; "Animal Cell Culture" [R.I.Freshney, ed. (1986)]; "Immobilized Cells and Enzymes" [IRL Press, (1986)]; B.Perbal, "A Practical Guide to Molecular Cloning", (1984). Другие используемые методики могут относиться к пептидному синтезу (Stewart J.M.; Young J.D. Solid Phase Peptide Synthesis. In Solid Phase Peptide Synthesis; Eds.; Pierce Chemical Co.: Rockford, IL, 1984; V. pp. 176), к аналитической химии (Miller C.; Rivier J. Peptide Chemistry: Development of high-performance liquid chromatography and capillary zone electrophoresis. Biopolymers, 1996, 40, 265-317), к методам стратегии, основанной на соотношении структуры и активности (включая, в числе других, тестирование in vivo и in vitro и структурный анализ с использованием ЯМР, КД, кристаллографии в рентгеновских лучах) (Gulyas J; Rivier C.; Perrin M.; Koerber S.C.; Sutton S.; Corrigan A.; Lahrichi S.L.; Craig A.G.; Vale W.W.; Rivier J. Potent, structurally constrained agonists and competitive antagonists of corticotropin releasing factor (CRF). Proc. Natl. Acad. Sci. USA 1995, 92, 10575-10579).
В связи с этим следующие термины, используемые в настоящем описании, имеют указанные ниже значения.
В контексте настоящего описания термин «кДНК» относится к ДНК-копии м-РНК-транскрипта гена.
В контексте настоящего описания термин «выведенная аминокислотная последовательность» обозначает аминокислотную последовательность, определяемую посредством считывания триплетной последовательности нуклеотидных оснований в кДНК.
В контексте настоящего описания термин «скрининг библиотеки» относится к процессу использования меченого зонда для проверки того, существует ли в соответствующих условиях, в конкретной библиотеке ДНК последовательность, комплементарная к имеющемуся зонду. Кроме того, «скрининг библиотеки» может быть проведен посредством ПЦР.
В контексте настоящего описания термин "ПЦР" относится к полимеразной цепной реакции, которая является предметом патентов США №№4683195 и 4683202 (Mullis), а также к другим усовершенствованиям, известным в данной области техники.
Следует отметить, что все последовательности аминокислотных остатков изображаются в настоящем описании формулами, в которых ориентация слева направо соответствует традиционному направлению от аминоконца к карбоксиконцу. Кроме того, следует отметить, что пунктирная линия в начале или в конце последовательности аминокислотных остатков обозначает пептидную связь со следующей последовательностью из одного или более аминокислотных остатков.
Представленные в настоящем описании аминокислоты предпочтительно имеют "L"-изомерную форму. Однако, любой L-аминокислотный остаток может быть заменен остатками в "D"-изомерной форме при условии, что такой модифицированный полипептид сохраняет желательные функциональные свойства в отношении связывания иммуноглобулина. NH2 относится к свободной аминогруппе, присутствующей на аминоконце полипептида. СООН относится к свободной карбоксигруппе, присутствующей на карбоксиконце полипептида. Сокращения названий аминокислотных остатков приводятся в соответствии со стандартной номенклатурой полипептидов (J. Biol. Chem., 243:3552-59 (1969)), которые хорошо известны в данной области техники.
Нестандартные аминокислоты могут быть включены в белки посредством химической модификации имеющихся аминокислот или посредством синтеза de novo соответствующего белка/пептида. Термин "нестандартная аминокислота" относится к аминокислоте, которая отличается по химической структуре от двадцати стандартных аминокислот, кодируемых генетическим кодом. Посттрансляционная модификация in vivo также может привести к присутствию нестандартной аминокислоты или производного аминокислоты в белке. N-концевая группа NH2 и С-концевая СООН-группа белка также могут быть модифицированы, например, посредством естественной или искусственной посттрансляционной модификации белка.
Белки/пептиды могут быть модифицированы посредством замещений аминокислот. Зачастую некоторые изменения приводят к значительным изменениям активности (агонисты вместо антагонитов) и потенциала/аффинности белков/пептидов, тогда как другие оказывают небольшой эффект или вовсе не оказывают никакого эффекта. Консервативные замещения с наименьшей вероятностью приводят к резкому изменению активности белка. Термин «консервативное замещение аминокислоты» относится к замещению аминокислоты химически подобной ей аминокислотой, то есть к замещению неполярных аминокислот другими неполярными аминокислотами, к замещению полярных аминокислот другими полярными аминокислотами, кислых аминокислотных остатков другими кислыми аминокислотами и т.п. Примеры предпочтительных консервативных замещений приведены в таблице 1.
Консервативные замещения аминокислот
Термин "химическое производное" относится к рассматриваемому полипептиду, который содержит один или более остатков, химически модифицированных посредством реакции на основе боковой функциональной группы. Такие модифицированные полипептиды включают, например, те их них, в которых свободные аминогруппы были модифицированы с образованием конкретных солей или были модифицированы посредством алкилирования и/или ацилирования, в том числе паратолуолсульфонильными группами, карбобензокси группами, трет-бутилоксикарбонильными группами, хлорацетильными группами, формильными и ацетильными группами. Свободные карбоксильные группы могут быть модифицированы с образованием органических или неорганических солей, метиловых и этиловых сложных эфиров или других типов сложных эфиров или гидразидов и, предпочтительно амидов (первичных или вторичных). Химические производные могут включать те пептиды, которые содержат одно или более встречающихся в природе аминокислотных производных двадцати стандартных аминокислот. Так, например, серин может быть замещен 4-гидроксипролином, а лизин может быть замещен орнитином. Пептиды, рассматриваемые в рамках настоящего изобретения, также включают те пептиды, которые имеют один или более дополнительных остатков и/или делеций по сравнению с конкретным пептидом, последовательность которого приводится в настоящем описании, при условии, что такой модифицированный пептид сохраняет необходимую биологическую активность.
Термин "репликон" представляет собой любой генетический элемент (например, плазмиду, хромосому, вирус), который функционирует как автономная единица при репликации ДНК in vivo, то есть как элемент, способный к репликации под собственным контролем.
Термин "вектор" представляет собой репликон, такой как плазмида, фаг или космида, к которому может быть присоединен другой сегмент ДНК, с тем чтобы осуществлять репликацию присоединенного сегмента.
Термин "молекула ДНК" относится к полимерной форме дезоксирибонуклеотидов (аденина, гуанина, тимина или цитозина) в ее либо одноцепочечной форме, либо в форме двухцепочечной спирали. Указанный термин относится только к первичной и вторичной структуре молекулы и не ограничивается какими-либо особыми третичными формами. Таким образом, данный термин включает двухцепочечную ДНК, встречающуюся, в том числе, в виде линейных молекул ДНК (например, рестрикционные фрагменты), в виде вирусов, плазмид и хромосом. В настоящем описании структура рассматривается в соответствии с принятым соглашением, согласно которому последовательность указывается только в направлении от 5' к 3'-концу вдоль нетранскрибируемой цепи ДНК (то есть цепи, имеющей последовательность, гомологичную мРНК).
Термин "начало репликации" относится к тем последовательностям ДНК, которые участвуют в синтезе ДНК.
Термин "кодирующая последовательность" ДНК представляет собой последовательность двухцепочечной ДНК, которая транскрибируется и транслируется in vivo в полипептид в условиях контроля соответствующими регуляторными последовательностями. Границы кодирующей последовательности определяются стартовым кодоном на 5' (амино) конце и кодоном терминации трансляции на 3' (карбокси) конце. Кодирующая последовательность может включать, не ограничиваясь приведенными последовательностями, прокариотические последовательности, кДНК из эукариотической мРНК, последовательности геномной ДНК из эукариотической (например, млекопитающих) ДНК и даже последовательности синтетической ДНК. Сигнальная последовательность для полиаденилирования и последовательность терминации транскрипции обычно локализованы на 3' конце кодирующей последовательности.
Контрольные последовательности транскрипции и трансляции представляют собой регуляторные последовательности ДНК, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и другие, которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине.
"Промоторная последовательность" представляет собой регуляторный участок ДНК, способный к связыванию РНК-полимеразы в клетке и инициирующий транскрипцию находящейся «правее» (в 3'-направлении) кодирующей последовательности. В контексте настоящего изобретения, промоторная последовательность ограничивается на ее 3'-конце сайтом инициации транскрипции и простирается «левее» (в сторону 5'-конца) до включения минимального числа оснований или элементов, необходимых для инициации транскрипции на уровнях, превышающих фон. Внутри промоторной последовательности находится сайт инициации транскрипции, а также домены связывания белка (консенсусные последовательности), ответственные за связывание РНК-полимеразы. Эукариотические промоторы зачастую, хотя и не всегда, содержат блоки "ТАТА" и блоки "САТ". Прокариотические промоторы содержат последовательности Шайн-Дальгарно в дополнение к -10 и -35 консенсусным последовательностям.
"Последовательность, контролирующая экспрессию", представляет собой последовательность ДНК, которая контролирует и регулирует транскрипцию и трансляцию другой последовательности ДНК. Кодирующая последовательность находится "под контролем" последовательностей, контролирующих транскрипцию и трансляцию в клетке, когда РНК-полимераза осуществляет транскрипцию кодирующей последовательности в м-РНК, которая затем в свою очередь транслируется в белок, кодируемый указанной кодирующей последовательностью.
"Сигнальная последовательность" может быть включена рядом с кодирующей последовательностью. Указанная последовательность кодирует сигнальный пептид, являющийся N-концевым в полипептиде, который взаимодействует с клеткой-хозяином, определяя перемещение полипептида к поверхности клетки или секрецию полипептида в среду, при этом указанный сигнальный пептид отрезается от хозяйской клетки прежде, чем указанный белок покидает клетку. Сигнальные последовательности могут быть обнаружены в ассоциации со множеством белков, естественных для прокариотов и эукариотов.
Термин "олигонуклеотид", применяемый в настоящем описании по отношению к зонду настоящего изобретения, определяется как молекула, содержащая два или более рибонуклеотида, предпочтительно более чем три. Его точный размер зависит от многих факторов, которые в свою очередь зависят от конечной функции и от использования указанного олигонуклеотида.
Термин "праймер" в контексте настоящего описания относится к олигонуклеотиду либо естественного происхождения, в виде очищенного продукта расщепления рестриктазами, либо полученного синтетическим путем, который способен действовать в качестве точки инициации синтеза при помещении в условия, при которых индуцируется синтез продукта удлинения праймера, являющегося комплементарным к цепи нуклеиновой кислоты, то есть в присутствии нуклеотидов и индуцирующего агента, такого как ДНК-полимераза, и при подходящей температуре и подходящем значении рН. Праймер может быть либо одноцепочечным, либо двухцепочечным и должен быть достаточно длинным для начала процесса синтеза желательного продукта удлинения в присутствии индуцирующего агента. Точная длина праймера зависит от многих факторов, включая температуру, источник праймера и использование метода. Так, например, для применения с диагностической целью, в зависимости от сложности целевой последовательности, олигонуклеотидный праймер в типичном случае содержит 15-25 или более нуклеотидов, хотя он может содержать и меньшее число нуклеотидов.
Праймеры согласно настоящему описанию выбирают так, чтобы они были "по существу" комплементарны к различным цепям конкретной целевой последовательности ДНК. Это означает, что праймеры должны быть достаточно комплементарными, чтобы гибридизоваться с соответствующими им цепями. В этой связи праймерная последовательность не обязательно отражает точную последовательность матрицы. Так, например, некомплементарный нуклеотидный фрагмент может быть присоединен к 5'-концу праймера, при этом остаток праймерной последовательности является комплементарным к указанной цепи. Альтернативно некомплементарные основания или более длинные последовательности могут быть введены в праймер при условии, что праймерная последовательность обладает достаточной комплементарностью к указанной последовательности или гибридизуется с ней и при этом образует матрицу для синтеза продукта удлинения.
В контексте настоящего описания термин "рестрикционные эндонуклеазы" и "ферменты рестрикции" относится к ферментам, каждый из которых разрезает двухцепочечную ДНК в области специфической нуклеотидной последовательности или рядом с ней.
Клетка становится "трансформированной" экзогенной или гетерологичной ДНК в том случае, когда такая ДНК вводится внутрь клетки. Трансформирующая ДНК может интегрироваться (ковалентное связывание) в геном клетки или не интегрироваться в него. В прокариотах, дрожжах и клетках млекопитающих, например, трансформирующая ДНК может сохраняться на эписомальном элементе, таком как плазмида. В отношении эукариотических клеток следует отметить, что устойчиво трансформированная клетка представляет собой такую клетку, в которой трансформирующая ДНК интегрирована в хромосому, так что она наследуется дочерними клетками при репликации хромосомы. Указанная стабильность демонстрируется способностью эукариотической клетки образовывать клеточные линии или клоны, включающие популяцию дочерних клеток, содержащих трансформирующую ДНК. "Клон" представляет собой популяцию клеток, полученную из единственной клетки или предшественника посредством митоза. "Клеточная линия" представляет собой клон первичной клетки, который способен к стабильному росту in vitro в течение многих генераций.
Две последовательности ДНК рассматриваются как "по существу гомологичные", когда по меньшей мере около 75% (предпочтительно по меньшей мере около 80% и еще более предпочтительно по меньшей мере около 90 или 95%) нуклеотидов совпадают на протяжении определенной длины последовательностей ДНК. Последовательности, которые являются по существу гомологичными, могут быть идентифицированы путем сравнения последовательностей с использованием стандартного программного обеспечения, доступного в банках данных последовательностей, или в эксперименте по гибридизации по методу Саузерна, например, в жестких условиях, определяемых для конкретной системы. Установление соответствующих условий гибридизации доступно специалистам в данной области. См., например, Maniatis et al., supra; DNA Cloning, Vols. I & II, supra; Nucleic Acid Hybridization, supra.
"Гетерологичный" участок ДНК конструкции представляет собой идентифицируемый сегмент ДНК внутри более крупной молекулы ДНК, который в природе не встречается в сочетании с более крупной молекулой. Таким образом, когда гетерологичный участок кодирует ген млекопитающего, указанный ген обычно ограничивается ДНК, которая не прилегает к геномной ДНК млекопитающего в геноме данного организма. В другом примере кодирующая последовательность представляет собой конструкцию, в которой сама кодирующая последовательность не встречается в природе (например, кДНК, в которой геномная кодирующая последовательность содержит интроны или синтетические последовательности, имеющие иные кодоны, чем исходный ген). Аллельные вариации или природные мутации не приводят к появлению гетерологичных участков ДНК согласно настоящему определению.
Метки, чаще всего используемые для указанных исследований, представляют собой радиоактивные элементы, ферменты, химические вещества, способные к флуоресценции под действием ультрафиолетового освещения, и другие. В технике известно множество флуоресцентных материалов, и они могут использоваться в качестве меток. Указанные материалы включают, например, флуоресцеин, родамин, аурамин, техасский красный, АМСА голубой и люцифер желтый. Конкретный материал, применяемый для обнаружения, представляет собой антикроличьи антитела, полученные иммунизацией коз и конъюгированные с флуоресцеином через изотиоцианат.
Конкретная система тестирования, которая была разработана и использовалась в технике, известна как рецепторный анализ. В указанном рецепторном анализе исследуемый материал подвергают соответствующему мечению и затем определенные тестируемые клеточные колонии инокулируют определенным количеством меченого и немеченого материала, после чего проводят исследования на наличие связывания с целью определения уровня связывания меченого материала с клеточными рецепторами. Таким способом могут быть установлены различия в аффинности между материалами.
В контексте настоящего описания термин "хозяин" включает не только клетки прокариотов, но также и клетки эукариотов, таких как дрожжевые, растительные и животные клетки. Рекомбинантная молекула ДНК или ген, который кодирует белок согласно настоящему изобретению, могут использоваться для трансформации клетки-хозяина с использованием любой известной в технике методики. Прокариотические клетки-хозяева включают E.coli, S.tymphimurium, Serratia marcescens и Bacillus subtilis. Эукариотические клетки-хозяева включают дрожжи, такие как Pichia pastoris, клетки млекопитающих и клетки насекомых.
В целом в сочетании с клеткой-хозяином используют экспрессирующие векторы, содержащие промоторные последовательности, которые облегчают эффективную транскрипцию введенного фрагмента ДНК. Экспрессирующий вектор в типичном случае содержит начало репликации, промотор(ы), терминатор(ы), а также специфические гены, которые делают возможным осуществление фенотипического отбора в трансформированных клетках. Трансформированные клетки-хозяева могут затем ферментироваться и культивироваться с использованием способов, известных в технике для достижения оптимального роста клеток.
Могут использоваться методы, известные специалистам в данной области техники, для конструирования экспрессирующих векторов, содержащих соответствующие сигналы контроля транскрипции и трансляции. См., например, технические приемы, описанные в Sambrook et al., 1989, Molecular Cloning: A Laboratory Manual (2nd Ed.), Cold Spring Harbor Press, N.Y. Ген и последовательности контроля его транскрипции определяются как "оперативно связанные", если последовательности контроля транскрипции эффективно контролируют транскрипцию гена. Векторы согласно настоящему изобретению включают, не ограничиваясь этим, плазмидные векторы и вирусные векторы.
В одном варианте осуществления настоящего изобретения предлагается выделенный и очищенный белок урокортин III. Указанный белок может представлять собой человеческий или мышиный белок урокортин III. Человеческий белок кодируется ДНК, частично включенной в состав человеческого EST (GenBank, номер доступа AW293249), со значительной гомологией последовательности с урокортинами иглобрюхих рыб.
Другой вариант осуществления настоящего изобретения относится к человеческому белку урокортину III, кодируемому пептидом-предшественником с последовательностью SEQ ID No.2. После осуществления посттрансляционной модификации очищенный человеческий урокортин III предпочтительно имеет аминокислотную последовательность, соответствующую SEQ ID No:3.
Настоящее изобретение также относится к модификации белков урокортина III. N-концы белков урокортина III могут быть модифицированы различными ацилирующими агентами, такими как карбоксилсодержащие группировки, сульфонилсодержащие группировки и изоцианаты. Альтернативно N-конец урокортина III может быть химически связан с молекулой токсина. N-конец урокортина III может также быть удлинен с помощью дополнительных аминокислот или пептидов, таких как D-тирозин, L-тирозин, D-тирозин-глицин или L-тирозин-глицин. Кроме того, один или более остатков метионина в урокортине III, таких как остатки, расположенные в положении 12 и 35 последовательности SEQ ID No.3, могут быть замещены остатками Nle. Альтернативно N-конец может быть удлинен с использованием D-иодтирозина, L-иодтирозина, D-иодтирозин-глицина и L-иодтирозин-глицина, а метиониновые остатки в положении 12 и 35 могут быть замещены Nle. Остатки иодтирозина могут быть помечены 125I.
В другом варианте осуществления настоящего изобретения предлагается фармацевтическая композиция, включающая белок урокортин III, а также способ лечения патофизиологического состояния с использованием указанной фармацевтической композиции. Фармацевтическая композиция может быть введена, например, с целью активации рецептора CRF-R2 у индивидуума и может таким образом способствовать лечению различных патофизиологических состояний, таких как высокая температура тела, нарушение аппетита, застойная сердечная недостаточность, сосудистые заболевания, стресс и беспокойство.
Другой вариант осуществления настоящего изобретения относится к модификации белка урокортин III. N-конец урокортина III может быть удлинен с использованием дополнительных аминокислот или пептидов, таких как D-тирозин, L-тирозин, D-тирозин-глицин или L-тирозин-глицин. Кроме того, один или более остатков метионина в урокортине III, таких как остатки, расположенные в положении 12 и 35 последовательности SEQ ID No.3, могут быть замещены остатками Ile, Val, Leu или предпочтительно Nle. Альтернативно N-конец может быть удлинен с использованием D-иодтирозина, L-иодтирозина, D-иодтирозин-глицина и L-иодтирозин-глицина, а метиониновые остатки в положении 12 и 35 могут быть замещены Nle. Остатки иодтирозина могут быть помечены 125I.
Предполагаются дополнительные замещения аминокислотными остатками, сохраняющимися в другом урокортине и родственных урокортину белках, которые отличаются у урокортина III. Такие аналоги урокортина могут включать урокортин III с одним или более аминокислотными замещениями, выбираемыми из группы, состоящей из Ile3, Nle3, CαMe-Leu3, Ile5, Nle5, CαMe-Leu5, Leu7, Nle7, Thr8, Ile9, Phe9, Gly10, His10, Leu11, Nle11, Leu12, Nle12, Arg13, Gln13, Nle14, CαMe-Leu14, Nle15, CαMe-Leu15, CαMe-Leu16, Leu16, Nle16, Glu17, Asp17, Nle18, Leu18, Arg20, Nle24, CαMe-Leu24, Arg32, Ile34, Nle34, CαMe-Leu34, Leu35, Nle35, Asp36, Glu36 и Val38.
В другом варианте осуществления настоящего изобретения предлагается антагонист рецептора CRF-R2. Указанный антагонист включает белок урокортина III или аналог урокортина III, причем первые пять-восемь N-концевых аминокислот в белке делетированы. Указанный антагонист может быть включен в состав фармацевтической композиции и использоваться для лечения застойной сердечной недостаточности, сосудистых заболеваний, дисфункции желудочно-кишечного тракта и головных болей мигрени или может использоваться как ингибитор ангиогенеза.
В другом варианте осуществления настоящего изобретения урокортин III модифицируется таким образом, что он содержит флуоресцентную метку или комплексующий агент для радионуклидов. Полученный в результате этого меченый урокортин может быть использован для идентификации клеток, экспрессирующих рецепторы урокортина III. Альтернативно урокортин III может быть связан с молекулой токсина.
В еще другом варианте осуществления настоящего изобретения предлагается антитело, направленное против урокортина III. В предпочтительном варианте осуществления изобретения указанное антитело представляют собой моноклональное антитело. Антитело может быть конъюгировано с молекулярной меткой, такой как флуоресцентная метка, фотоаффинная метка или радиоактивные маркеры.
Приведенные ниже примеры даны с целью иллюстрации различных вариантов осуществления настоящего изобретения и не должны рассматриваться как ограничивающие каким-либо образом настоящее изобретение.
Пример 1
Идентификация человеческого урокортина III
В базе данных экспрессируемых последовательностей tag (EST) человека GenBank был проведен поиск с использованием последовательности иглобрюхих рыб (Takifugu rubripes) (GenBank, номер доступа AJ251323), относящийся к урокортину (урокортин-родственный пептид (URP)), в качестве зонда. Указанный поиск позволил идентифицировать человеческий EST (GenBank, номер доступа AW293249) со значительной гомологией последовательности относительно урокортинов иглобрюхих рыб. Сопоставления гомологии новой последовательности с человеческим урокортином I и урокортином II показали, что указанная последовательность кодирует новый человеческий урокортин, который был обозначен как урокортин III. Частичный человеческий EST содержал последовательность-предшественник и первые 29 аминокислот участка зрелого пептида. Были сконструированы на основе частичной последовательности человеческого EST гнездовые праймеры для урокортина III, состоящие из последовательностей: 5'-AAG AGT CCC CAC CGC ACC AAG TTC ACC-3' (SEQ ID No.16) и 5'-TCC CTC GAC GTC CCC ACC AAC ATC ATG-3' (SEQ ID No.17). Указанные праймеры были использованы совместно с «гнездовыми» закрепленными праймерами для скрининга набора Human Genome Walker Kit (Clontech) методом ПЦР. «Гнездовой» ПЦР проводили в течение 35 циклов, состоящих из денатурации при 95°С в течение пяти минут с последующим удлинением последовательности при 66°С в течение 12 минут. Амплифицированные фрагменты подвергали субклонированию в вектор pCRIITOPO (Invitrogen), затем секвенировали и определили, что они кодируют зрелый пептид полной длины.
Для расширения информации о последовательности на С-конце были разработаны праймеры на основании использования частичной последовательности EST человека для скрининга набора Human Genome Walker Kit (Clontech) методом ПЦР. Библиотечный набор геномных ДНК человека подвергли скринингу с использованием ген-специфических и закрепленных праймеров с использованием стратегии гнездовой ПЦР. Идентифицировали человеческий ген полной длины, кодирующий белок предположительного пептида. Указанный белок, обозначенный как урокортин III (Ucn III), родственен другим известным представителям семейства CRF.
Пример 2
Идентификация мышиного урокортина III посредством гибридизационного скрининга
На основании последовательности человеческого Ucn III полной длины был получен зонд длиной 144 п.н., перекрывающий участок зрелого пептида, с целью поиска мышиного ортолога. Указанный зонд был использован для скрининга геномной библиотеки мыши λFIXII (Stratagene) посредством гибридизации в условиях низкой жесткости. Гибридизацию проводили в течение ночи при 42°С в смеси 20% формамида/5XSSC/5X Denhardts/0,5% СДС/5% сульфата декстрана. Промывку осуществляли в 1X SSC при 55°С. Очищенные бляшки подвергали субклонированию в pBluescript и секвенировали. Полноразмерный клон мышиного генома идентифицировали и секвенировали. кДНК мышиного Ucn III выделяли из полной кДНК мозга посредством ПЦР с использованием праймеров, созданных на основе последовательности мышиного генома. ПЦР проводили при 55°С в течение 35 циклов с 2-минутным выдерживанием при 72°С.
Пример 3
Анализ генов человеческого и мышиного Ucn III
Гены человеческого и мышиного Ucn III оба содержат два потенциальных сайта инициации трансляции. Нуклеотидная последовательность человеческого гена кодирует белок, который согласно подсчетам должен включать 161 или 159 аминокислот, в зависимости от использованного N-концевого метионина, где предпочтительным является первый метионин в соответствии с программой предсказания Netstart 1.0 (22) (фиг.2А). Мышиный ген кодирует белок, который по подсчетам должен включать 164 или 162 аминокислот предшественника, также с предпочтительным началом трансляции от первого метионина (фиг.2В). Считается, что процессинг молекулы предшественника с получения предлагаемого зрелого пептида из 38 аминокислот происходит посредством расщепления в положении Lys 119 с С-концевой стороны в случае человеческого гена и в положении следующего Lys 122 в случае мышиного Ucn III.Таким образом, Ucn III подчиняется правилам процессинга в одноосновных остатках (23), сходных у Ucn и мышиного Ucn II. С-концевая последовательность содержит пару основных остатков (R-K) в случае последовательности человека и (K-K) в случае мышиной последовательности, которым непосредственно предшествует глицин, предположительно участвующий в С-концевом амидировании. Предполагаемые участки зрелого пептида в человеческом и мышином Ucn III показаны в виде участков, заключенных в прямоугольники (фиг.2А и 2В). Поскольку участки зрелого пептида человеческого и мышиного Ucn III отличаются только по 4 аминокислотным остаткам, они, скорее всего, являются ортологами. Указанные последовательности депонированы в банк данных GenBank [номер доступа AF361943 (человеческий Ucn III) и AF361944 (мышиный Ucn III)].
Пример 4
Сравнение Ucn III с другими представителями семейства CRF
На фиг.2С приведено сравнение участка зрелого пептида из 38 аминокислот последовательности человеческого Ucn III с такими же участками других представителей семейства. Человеческий и мышиный Ucn III демонстрируют 40% идентичность с мышиным Ucn II. Человеческий и мышиный Ucn III демонстрируют 37% идентичность с человеческим URP. Они демонстрируют более отдаленное сходство с Ucn и CRF. Человеческий и мышиный Ucn III демонстрируют 21% и 18% идентичность с человеческим и мышиным Ucn, соответственно. Человеческий и мышиный Ucn III демонстрируют 32% и 26% идентичность с человеческим/крысиным CRF. Урокортины II и III млекопитающих, по всей видимости, представляют собой отдельную, но эволюционно родственную ветвь CRF-семейства, при этом они более тесно связаны с двумя урокортинами иглобрюхих рыб, чем с Ucn или CRF млекопитающих (фиг.2D). В настоящее время в базе данных GenBank имеются три последовательности из иглобрюхих рыб, близкие к CRF-семейству. На аминокислотном уровне урокортин-родственный пептид (URP) иглобрюхих рыб (GenBank, номер доступа AJ251323) наиболее близок (76% идентичности) как человеческому, так и мышиному Ucn III и более удален от человеческого URP (42%) и мышиного Ucn II (37%). Вторая последовательность урокортина иглобрюхих рыб (GenBank, AL175143) также более близка к Ucn III (53%), чем к любому другому представителю семейства CRF млекопитающих. Третий пептид иглобрюхих рыб (GenBank, AL218869) наиболее сходен с рыбьими уротензинами типа I и имеет наивысшую идентичность по аминокислотам с уротензином I камбалы (63%).
Пример 5
Синтез урокортина III
Человеческий и мышиный Ucn III синтезировали вручную с использованием твердофазного метода на основе 4-метилбензгидриламиновой смолы и Вос-подхода (24). Для удаления Вос-групп использовали трифторуксусную кислоту (ТФУ, 60% в дихлорметане). Основную сборку цепи проводили с использованием диизопропилкарбодиимида. Пептид расщепляли и подвергали депротекции во фтористоводородной кислоте и очищали с использованием ВЭЖХ-ОФ и трех систем растворителей (фосфат триэтиламмония при рН 2,25 и рН 6,5 и/или 0,1% ТФУ) (25). Получали пептиды с чистотой выше 95% по независимым критериям ВЭЖХ и капиллярного зонного электрофореза. Данные масс-спектрального анализа подтверждали состав полученных препаратов.
В дополнение к описанному выше синтезированному урокортину III (расчетная масса 4136,34; найдено: 4136,2), с использованием того же метода конструировали следующие синтетические аналоги: [Tyr-Gly]-урокортин III (расчетная масса 4356,34; найдено: 4356,3); [Nle12,35]-урокортин III (расчетная масса 4100,34; найдено: 4100,2); [Tyr0,Nle12,35]-урокортин III (расчетная масса 4263,41; найдено: 4263,2) и [Tyr-Gly0,Nle12,35]-урокортин III (расчетная масса 4320,431; найдено: 4320,4).
Пример 6
Тест по связыванию Ucn III
Синтезированный урокортин III анализировали с точки зрения его способности связываться с рецепторами CRF-R1, CRF-R2α и CRF-R2β с помощью конкурентного замещения 125I[Tyr0,Glu1,Nle17]соваджина из клеток яичников китайского хомячка, стабильно экспрессирующих каждый из указанных типов рецепторов. Стабильно трансфицированные клеточные линии, экспрессирующие CRF-R2α, были любезно предоставлены Demitri Grigoriadis, Neurocrine Biosciences Inc. Неочищенные мембранные фракции были получены из клеток яичников китайского хомячка, стабильно экспрессирующих либо клонированный человеческий CRF-R1, либо мышиный CRF-R2β, как было описано ранее (5). Исследуемые пептиды и радиолиганд, [125Ityr0,Glu1,Nle17]-соваджин, разведенные в буфере для тестирования (20 мМ Hepes/2 мМ EGTA/0,1% БСА/10% сахароза; рН 7,6), объединяли с мембранными фракциями (10 мкг) на микротитрационных планшетах MAGV (Millipore), предварительно покрытых 0,1% полиэтиленимином. После выдерживания в течение 90 минут при комнатной температуре реакционную смесь фильтровали и дважды промывали буфером для анализа. Определение количества полученного комплекса с радиолигандом осуществляли с помощью гамма-счетчика. Константы ингибирования связывания - Ki - определяли с помощью программы GRAPHPAD. Аффинности Ucn II и Ucn III в отношении CRF-связывающего белка (CRF-BP) вычисляли на основании данных по замещению [D125ITyr0]-hCRF с использованием методики, описанной в цитируемой литературе (26).
Данные, полученные на основе по меньшей мере трех экспериментов, объединяли и значения константы ингибирования диссоциации (Ki) вычисляли с использованием значений ЕС50, полученных с помощью программы Graphpad Prism. Подсчитывали среднее из трех значений их log10 и использовали его в качестве экспоненты числа 10 с целью определения "среднего" значения ЕС50. Определяли ошибку путем подсчета стандартного отклонения значений log10, с помощью которого число 10 экспонировали с определением фактора ошибки. Вычисленные значения ЕС50 затем делили или умножали на коэффициент ошибки с целью определения нижней и верхней границ ошибки соответственно.
В таблице 2 приведены данные по сравнению аффинностей к связыванию и потенциалов по стимулированию накопления цАМФ в клетках, стабильно экспрессирующих человеческий CRF-R1, крысиный CRF-R2α и мышиный CRF-R2β. Ucn представляет собой значительно более эффективный фактор по связыванию с CRF-R1, чем Ucn II или Ucn III. И Ucn II, и Ucn III селективно связывают оба варианта сплайсинга CRF-R2 по сравнению с CRF-R1. Человеческий Ucn III проявляет значительно более низкую аффинность к любому рецептору типа 2, чем Ucn или Ucn II. Мышиный Ucn III демонстрирует значительно более высокую аффинность к рецепторам мышиного CRF-R2, чем человеческий Ucn III. И Ucn II, и Ucn III оба проявляют несколько большую аффинность в отношении к CRF-R2β по сравнению с CRF-R2α. Наблюдается также превышение потенциала мышиного Ucn III над человеческим Ucn III в отношении связывания с человеческим CRF-R2α (данные не показаны).
Связывающие свойства и функциональные активности лигандов CRF-семейства
Пример 7
Активация аденилатциклазы в клетках, трансфицированных рецепторами
Клетки яичника китайского хомячка, стабильно трансфицированные человеческим CRF-R1 или мышиным CRF-R2, помещали в 48-луночные планшеты для тканевых культур (Costar) и оставляли для восстановления в течение 24 часов. Среду заменяли по меньшей мере за два часа до обработки на среду DMEM/0,1% фетальной бычьей сыворотки. Клетки предынкубировали с 0,1 мМ 3-изобутил-1-метилксантином в течение 30 минут и затем обрабатывали пептидами в течение 20 минут при 37°С. Внутриклеточное накопление цАМФ в клетках СНО, стабильно трансфицированных CRF-R1 или CRF-R2, использовали в качестве показателя активации рецепторов. И Ucn II, и Ucn III оба обладают очень низкой способностью активировать CRF-R1 (>100 нМ), что резко отличает их от Ucn, значение ЕС50 для которого составляет ˜0,15 нМ. И действительно, Ucn III не показывает активации CRF-R1 даже при очень высоких дозах (1 мкМ). Способности Ucn II и III активировать CRF-R2α и CRF-R2β приблизительно равны и почти эквивалентны таковой у Ucn. Таким образом, в тесте на накопление цАМФ и Ucn II, и Ucn III демонстрируют селективность в отношении CRF-R2, превышающую селективность в отношении CRF-R1, но не проявляют предпочтения в отношении CRF-R2α и CRF-R2β. Кроме того, относительные способности мышиного и человеческого Ucn II и Ucn III к функциональной активации CRF-R2 перекрываются, несмотря на низкое сродство человеческого Ucn III в отношении связывания с CRF-R2.
Пример 8
Активация аденилатциклазы в клетках, экспрессирующих эндогенные рецепторы
Способности Ucn II и Ucn III активировать аденилатциклазу были оценены в клетках, экспрессирующих эндогенный CRF-R1 (культура первичных клеток передней доли гипофиза, (5)) или CRF-R2β (клетки A7r5, (27)). Клетки гладкой мускулатуры аорты крыс, A7r5, помещали в 48-луночные культуральные планшеты и выдерживали для восстановления в течение 48 часов. Клетки выдерживали в среде DMEM/0,2% фетальной бычьей сыворотки в течение ночи перед экспериментом. Полученные клетки преинкубировали с 0,1 мМ 3-изобутил-1-метилксантином в течение 30 минут и затем обрабатывают пептидами в течение 20 минут при 37°С. Выводили в культуру клетки передней доли гипофиза крысы (28) и обрабатывали их исследуемыми пептидами в течение 45 минут при 37°С. Внутриклеточную цАМФ экстрагировали из всех клеток и измеряли в тройной повторности с использованием набора для радиоиммуноанализа (Biomedical Technologies). Активности определяли с использованием программы PRISM GRAPHPAD. Результаты представлены на фиг.3.
В соответствии с результатами, полученными на трансфицированных рецепторах, Ucn II и Ucn III способны активировать эндогенный CRF-R2β при суб- или низких наномолярных концентрациях лигандов (фиг.3А). Как ожидалось, Ucn II демонстрирует низкую способность увеличивать уровень циклического АМФ в культивируемых клетках гипофиза, экспрессирующих CRF-R1, тогда как Ucn III неактивен в данном тесте даже в концентрациях >1 мкМ (фиг.3В).
Пример 9
Связывание с CRF-связывающим белком
В отличие от CRF и урокортина, которые характеризуются субнаномолярными аффинностями в отношении CRF-BP, ни Ucn II, ни Ucn III не проявляют заметной аффинности в отношении данного белка (данные не приведены). Урокортин III не проявляет высокой аффинности ни в отношении CRF-R1, ни в отношении CRF-R2. Однако аффинность в отношении CRF-R2 выше, чем аффинность в отношении CRF-R1. В связи с его высокой способностью стимулировать образование циклического АМФ в клетках, экспрессирующих CRF-R2α или β, аффинность его, очевидно, достаточно высока, чтобы урокортин III мог действовать как нативный агонист CRF-R2. Также вполне вероятно, что урокортин III является более мощным агонистом еще не идентифицированного рецептора.
Пример 10
Экспрессия мРНК Ucn III
Тест на защиту от РНКазы проводили для определения тканевого распределения мРНК мышиного Ucn III. Анализ на защиту от РНКазы проводили по ранее описанной методике (29). Суммарную РНК экстрагировали с использованием TRI REAGENT (Molecular Research Center, Inc., Cincinnati, OH). Уровни мРНК мышиного Ucn III и глицеральдегид-3-фосфатдегидрогеназы (GAPDH) измеряли одновременно посредством реакции защиты от РНКазы с использованием мышиной GAPDH в качестве внутреннего контроля нанесения.
Антисмысловой по Ucn III рибонуклеотидный зонд из 528 нуклеотидов, специфичный к мРНК Ucn III мыши, синтезировали с использованием РНК-полимеразы Т3. Антисмысловой рибонуклеотидный зонд из 200 нуклеотидов, специфичный к мРНК GAPDH мыши, синтезировали с использованием РНК-полимеразы Т3. Все рибонуклеотидные зонды синтезировали в присутствии [α-32P]UTP (3000 Ки/ммоль) и либо 20 мкМ UTP в случае Ucn III или 200 мкМ UTP в случае GAPDH, как было описано ранее (29). Размеры фрагментов, защищенных с помощью рибонуклеотидных зондов к Ucn III и GAPDH, составляют 415 и 135 нуклеотидов соответственно.
Образцы РНК (50 мкг периферической ткани или 20-25 мкг мозговой ткани) гибридизовали в 24 мкл деионизованного формамида плюс 6 мкл буфера для гибридизации, содержащего 107 имп/мин в случае Ucn III и 3х104 имп/мин в случае GAPDH-антисмысловых рибонуклеотидных зондов. После нагревания до 85°С в течение 5 минут образцы гибридизовали при 42°С в течение 12 часов и последовательно расщепляли РНКазой (175 мкг/мл РНКазы А и 500 Ед/мл РНКазы Т1) при 24°С в течение 60 минут. Образцы разделяли на 5% полиакриламидных гелях с мочевиной. Анализы полученных картин разделения проводили с использованием системы PhosphorImager (Molecular Dynamics, Inc., Sunnyvale, CA) и пакета программного обеспечения ImageQuant 4.0.
Суммарную РНК из тканей, охватывающих несколько участков ЦНС, и периферических тканей гибридизовали с кРНК из 528 п.н. Указанный зонд охватывает участок зрелого пептида и приводит к получению защищенного фрагмента из 415 п.н. В ЦНС сайты экспрессии мРНК включают гипоталамус, ствол мозга и латеральную перегородку (LS)/ложе ядра терминальной полоски (BnST) (фиг.4). В гипофизе, мозжечке и коре головного мозга не отмечается заметной экспрессии мРНК. В периферических тканях мРНК Ucn III экспрессируется в области тонкого кишечника и кожи и при этом не обнаруживается заметной экспрессии в сердце, аорте, печени или легком (фиг.4).
Пример 11
Гибридизация in situ
Для выявления более детального распределения экспрессии Ucn III в мозге проводили гибридизацию in situ на сериях срезов головного мозга крыс и мышей с использованием антисмысловых и смысловых зондов кРНК, полученных на основе матрицы кДНК мышиного Ucn III размером 415 п.н. Мозговые и периферические ткани получали из взрослых мышей (C57BL/6) и крыс линии Sprague-Dawley путем их быстрого извлечения и замораживания в сухом льду. Замороженные блоки нарезали срезами толщиной 20 мкм на криостате, оттаивали при помещении на предметные стекла и сохраняли при -80°С до использования. Гибридизацию in situ проводили с мечеными 32Р антисмысловыми и смысловыми (контроль) зондами кРНК, транскрибированными из линеаризованной плазмиды (pCRIITOPO), содержащей кДНК мышиного Ucn III (30). Зонды метили до достижения удельной активности 1-3×109 (расп/мин)/мкг, наносили на предметные стекла до концентрации примерно 2,8×107 (имп/мин)/мл и гибридизовали в течение ночи при 56°С. Стекла промывали в SSC повышающейся жесткости и затем обрабатывали РНКазой при 37°С и, в завершение, 0,1Х SSC при 65°С. После дегидратации стекла выкладывали на рентгеновскую пленку (β-Max; Kodak) на 4 дня при 4°С, а затем покрывали жидкой эмульсией Kodak NTB-2 и экспонировали при 4°С в течение 10 дней.
Позитивный сигнал мРНК Ucn III наблюдали только в срезах после гибридизации с антисмысловым зондом. Распределение экспрессии мРНК Ucn III, как было показано, ограничивается в основном несколькими отдельными участками вентральной части переднего мозга (фиг.5). Один из них находится в медиальном предоптическом ядре, где наблюдается непрерывная полоса положительно меченых клеток, включающих инвертированную Y-образную срединную группировку. Ни одна из циркумвентрикулярных групп клеток, с которыми тесно связано медиальное предоптическое ядро (сосудистый орган терминальной пластинки и субфорникальный орган) не содержит положительных гибридизационных сигналов и только дополнительные сайты экспрессии мРНК Ucn III на данном уровне отмечались над рассеянными клетками в медиальном и латеральном предзрительных полях. Второе основное место экспрессии мРНК Ucn III отмечалось в виде продольно организованного кластера меченых клеток, связанных с (и по существу окруженных) стволами свода по всему ростральному гипоталамусу. Указанный кластер включает клетки, расположенные в задней части ложа ядра терминальной полоски передней и латеральной области гипоталамуса и в пораженной заболеванием области сразу же сбоку от (но иногда внутри) паравентрикулярного ядра гипоталамуса. Хвостовое расширение указанной группировки наблюдается в аналогичном положении, в основном в дорсальном и латеральном относительно ростральных участков дорсомедиального гипоталамического ядра. В стороне от указанной "ростральной перифорникальной" группы отмечаются рассеянные положительно гибридизующиеся нейроны, явно отмечаемые в вентральной части переднего перивентрикулярного ядра и обратноперекрестного поля. Третий основной сайт экспрессии мРНК Ucn III находился над подмножеством клеток в переднедорсальной части медиального миндалевидного ядра. В стволе головного мозга единственный достоверный сайт экспрессии Ucn III локализован дискретно в клеточной группе, относящейся к слуховой системе, в верхнем параоливаном ядре (фиг.5).
Пример 12
Ацилирование урокортина III
N-конец белков урокортина III согласно настоящему изобретению может быть удлинен с помощью ацилирующих агентов, получаемых на основе большого множества карбоксил- и сульфонилсодержащих группировок, включая дополнительные аминокислоты или пептиды, выбираемые из группы, состоящей из D-тирозина, L-тирозина, D-тирозин-глицина и L-тирозин-глицина. N-конец белка урокортина III может быть также удлинен с использованием токсинов или изоцианатов. Добавление ацилирующих агентов может повысить устойчивость к деградации (химической и ферментативной), может модулировать растворимость белка до достижения медленного высвобождения терапевтических форм указанных белков и позволяет осуществлять выборочное мечение (радиоактивное, флуоресцентное, хелаторами, токсинами, фотоаффинное, иммуноспецифическое и др.).
Пример 13
Аналоги урокортина III
Предшествующие исследования с лигандами для других CRF-рецепторов показали, что в указанных лигандах могут быть проведены различные замещения без потери лигандами их способности связываться с соответствующими рецепторами или их биологической активности. Многочисленные предшествующие исследования урокортинов показали, что достаточно легко может переноситься проведение одного, двух и даже трех замещений. В некоторых случаях модификации урокортина приводили к получению белка с более желательными фармакологическими свойствами. Поскольку урокортин представляет собой небольшой белок, такие модификации могут быть наиболее легко введены с помощью методов пептидного синтеза, известных специалисту в данной области техники. Такие способы включают твердофазные технологии, способы синтеза в неполностью твердой фазе, конденсацию фрагментов и классическое добавление в растворе. Указанные способы особенно предпочтительны, если в урокортин III нужно включить нестандартные аминокислоты. Альтернативно, если модификации состоят полностью из природных аминокислот, то для мутагенеза и последующей экспрессии модифицированного урокортина III может использоваться методика рекомбинантной ДНК.
Зрелый урокортин III теряет остаток тирозина. Поскольку тирозиновые остатки полезны для радиоактивного иодирования белков, одна возможная модификация урокортина III могла бы состоять в замещении тирозином другой аминокислоты в белке. В предшествующих примерах было описано добавление последовательности, состоящей из Tyr-Gly, к N-концевому хвосту урокортина. Полученный в результате этого белок сохраняет связывание с CRF-рецептором и биологическую активность и может быть полезен в радиоактивном иодировании указанного белка.
Было показано, что делетирование первых пяти-восьми остатков урокортина приводит к образованию эффективных антагонистов урокортина. Указанные белки способны к связыванию с CRF-рецепторами, но не стимулируют и не активируют в значительной мере указанные рецепторы. Ожидается, что делеция от четырех до восьми аминокислот из урокортина III также привела бы к получению эффективных антагонистов. Существует также вероятность обнаружения антагонистов CRF-связывающих белков среди других фрагментов урокортина III. Указанные антагонисты могут быть эффективны в плане повышения уровней эндогенных пептидов, которые обычно удаляются CRF-связывающим белком. Посредством ассоциации с CRF-связывающим белком и блокирования связывания CRF урокортина, урокортина II и урокортина III с тем же белком, увеличиваются эффективные in vivo концентрации эндогенного CRF, урокортина, урокортина II и урокортина III. Такие антагонисты могут быть введены совместно с другими агонистами или антагонистами CRF, урокортина, урокортина II или урокортина III с целью повышения их эффективности.
На фиг.2С приведены результаты сравнения на предмет гомологии, существующей между урокортином III и эквивалентными сегментами урокортин-родственного пептида из иглобрюхих рыб, мышиного урокортина II, человеческого урокортин-родственного пептида, урокортина крысы, кортикотропин-рилизинг-фактора крысы, кортикотропин-рилизинг-фактора овцы и соваджина лягушки. Уровни гомологии составляют от 20% до 77%. Анализ, проиллюстрированный на фиг.2С, позволил выявить наличие консервативных аминокислот в других урокортинах и урокортин-родственных белках. В то время как множество аминокислотных остатков, которые являются консервативными в других урокортинах и урокортин-родственных белках, сохраняются в урокортине III, другие аминокислотные остатки отличаются от тех, которые являются консервативными в других указанных белках. В этой связи замещение указанных отличающихся аминокислотных остатков теми, которые совпадают у других урокортин-родственных белков, представляет способ, посредством которого можно создавать аналоги урокортина для использования их в качестве агонистов и антагонистов. Так, например, лизиновый остаток находится в позиции 20 урокортина II, а остаток аргинина находится в аналогичной позиции в крысином урокортине, мышином урокортине II, человеческом урокортин-родственном пептиде и в кортикотропин-рилизинг-факторе крысы. В этой связи замещение Lys20 в урокортине остатком аргинина должно привести к получению аналога урокортина III, который все еще связан с CRF-рецепторами, но имеет модифицированную связывающую активность. Другие предполагаемые замещения в урокортине III включают Ile3, Ile5, Leu7, Thr8, Ile9, Phe9, Gly10, His10, Leu11, Leu12, Arg13, Gln13, Leu16, Glu17, Asp17, Arg20, Arg32, Ile34, Leu35, Asp36, Glu36 и Val38. Поскольку многие из указанных замещений приводят к получению белка урокортина III, более близкого к консенсусной последовательности всех урокортин-родственных белков, ожидается, что многие из них будут более сильно связываться с CRF-рецепторами и таким образом будут образовывать мощные агонисты CRF-рецептора. Альтернативно посредством N-концевой делеции первых четырех-восьми аминокислот, как указано выше, такие агонисты могут быть превращены в антагонисты CRF-рецептора.
Другие возможные замены включают замещение остатков метионина в положениях 12 и 35 и/или остатков лейцина в положениях 3, 5, 14, 15, 24 и 34 остатками Nle или CαMe-Leu15. Поскольку сравнение последовательности урокортина III с другими позициями урокортина свидетельствует о замещении лейцина в положениях 7, 11, 12, 16 и 35 в урокортине III, то в указанных положениях вместо Leu могут быть включены Nle или CαMe-Leu15.
Синтезируют аналог урокортина III, содержащий одно или более изменений, описанных выше. Анализ в соответствии с общей процедурой, указанной в примере 5, показывает улучшение связывания с рецепторами CRF-R1, CRF-R2α и/или CRF-R2β в процессе конкурентного замещения 125I[Tyr0,Glu1,Nle17]соваджина из клеток яичника китайского хомячка, стабильно экспрессирующих каждый тип рецептора, и изменение внутриклеточных уровней цАМФ. Дальнейшее исследование показывает, что аналог урокортина III оказывает благоприятные эффекты при лечении высокой температуры тела, расстройства аппетита, застойной сердечной недостаточности, сосудистых заболеваний и других сердечно-сосудистых состояний, дисфункции желудочно-кишечного тракта, стресса и беспокойства, а также головной боли при мигрени, и может действовать как мощный ингибитор ангиогенеза.
Пример 14
Фармацевтическое введение урокортина III и его аналогов
Урокортин III, его аналоги или их нетоксичные аддитивные соли, объединенные с фармацевтически или ветеринарно приемлемыми носителями с образованием фармацевтической композиции, могут вводиться млекопитающим, включая людей и других животных, множеством способов: внутривенно, подкожно, внутримышечно, чрескожно, например, интраназально, внутрилегочно, интрацереброспинально, подъязычно или перорально. Пептиды должны быть по меньшей мере примерно 90% чистоты и предпочтительно должны характеризоваться чистотой по меньшей мере порядка 97%; однако пептиды меньшей степени чистоты являются эффективными и также могут использоваться для введения животным, отличным от людей. Указанная степень чистоты означает, что рассматриваемые пептиды составляют указанный весовой процент от всех имеющихся пептидоподобных компонентов и пептидных фрагментов.
Введение урокортина III или агонистов урокортина III людям может проводиться врачом с целью лечения высокой температуры тела, расстройства аппетита, застойной сердечной недостаточности, заболеваний сосудов, дисфункции желудочно-кишечного тракта, стресса и беспокойства. Антагонисты урокортина III могут вводиться с целью лечения застойной сердечной недостаточности, сосудистых заболеваний, дисфункции желудочно-кишечного тракта и мигрени или для ингибирования ангиогенеза. Требуемая дозировка варьирует в зависимости от конкретного состояния, подлежащего лечению, от тяжести состояния и от длительности предполагаемого лечения. Указанные пептиды могут также использоваться для оценки функции системы гипоталамус-гипофиз-надпочечники у животных, у которых имеется подозрение на патологию эндокринной или центральной нервной системы, посредством приемлемого введения с последующим мониторингом функций организма.
Такие пептиды зачастую вводятся в виде фармацевтически или ветеринарно приемлемых нетоксичных солей, таких как аддитивно-кислотные соли или комплексы с металлами, например с цинком, железом, кальцием, барием, магнием, алюминием или др. (которые рассматриваются как аддитивные соли в контексте настоящего описания). Иллюстративными примерами таких аддитивных-кислотных солей являются гидрохлорид, гидробромид, сульфат, фосфат, таннат, оксалат, фумарат, глюконат, альгинат, малеат, ацетат, цитрат, бензоат, сукцинат, малат, аскорбат, тартрат и им подобные. Если активный ингредиент должен вводиться в виде таблетки, то указанная таблетка может содержать связующее вещество, такое как трагакант, кукурузный крахмал или желатин, дезинтегрирующее средство, такое как альгиновая кислота, и замасливатель, такой как стеарат магния. Если желательно введение в жидкой форме, то могут использоваться подсластитель и/или корригент, а внутривенное введение может производиться в изотоническом растворе, растворах фосфатного буфера или др.
Рассматриваемые пептиды должны вводиться под контролем врача, при этом указанные фармацевтические композиции обычно содержат пептид в сочетании с традиционным, фармацевтически или ветеринарно приемлемым носителем. Обычно дозировка составляет от примерно 1 до примерно 200 микрограмм пептида на килограмм веса тела животного-хозяина.
Пример 15
Ucn III и функция поджелудочной железы
Изучение экспрессии мРНК Ucn III у грызунов показало, что Ucn III экспрессируется и в центральной нервной системе, и на периферии. На периферии основными органами экспрессии Ucn III являются поджелудочная железа и желудочно-кишечный тракт. Дополнительные гистологические исследования показали, что внутри поджелудочной железы Ucn III локализуется совместно с инсулином и, следовательно, содержится в β-клетках островков Лангерганса.
Для определения функции указанного нового продукта β-клеток Ucn III вводили крысам внутривенно, и при этом было показано, что он быстро стимулировал секрецию глюкагона и затем повышал уровень сахара в крови. Другими авторами было показано, что глюкагон может действовать непосредственно на β-клетки, стимулируя секрецию инсулина паракринным способом. Таким образом, Ucn III может служить средством передачи сигнала от β-клетки к α-клетке, которая продуцирует глюкагон, который, в свою очередь, будет воздействовать на β-клетку. Следовательно, Ucn III (или любой агонист CRF-R2) может использоваться для стимуляции и поддержания функции островков.
Поскольку инсулин и глюкагон оказывают резко противоположное воздействие на уровень сахара в крови, возможно, что Ucn III может оказывать пролонгированные благоприятные эффекты на функцию островков. С другой стороны, ввиду возможной роли Ucn III, локализованного внутри островка, существует вероятность того, что антагонист CRF-R2 может быть также полезен для снижения образования глюкагона, который является основным гипергликемическим фактором, известным в плане осложнения контроля глюкозы при диабете.
Пример 16. Сравнение различных аналогов урокортина III
На таблице 3 показаны результаты замещения лиганда различными аналогами Unc III. Эти исследования были осуществлены в соответствии с методикой, описанной в статье River et al., 1999.
Вкратце, были получены указанные аналоги и проведены исследования их способности замещать радиоактивно меченный неспецифический агонист урокортина (савагин лягушки), либо рецептора кортикотропин-релизинг фактора типа 1 (CRF-R1), либо рецептора кортикотропин-релизинг фактора типа 2 (CRF-R2).
В таблице две колонки справа обозначают наномолярную концентрацию аналога, необходимую для замены агониста двух указанных рецепторов, с учетом стандартного отклонения, указанного в скобках. Номера слева представляют собой интервал обозначения несущественной релевантности. В описании указана номенклатура обозначений аналогов. Дополнительно ТК hU3 обозначает Thr-Lys-UIII человека, YG mU3 обозначает Tyr-Gly-UIII мыши, a GTK mU3 обозначает Gly-Thr-Lys-UIII мыши.
Из приведенной таблицы видно, что каждый из аналогов обладает способностью специфично связываться с CRF-R2b, по сравнению с CRF-R1.
Обсуждение
Ucn III был идентифицирован в процессе скрининга на гомологию последовательностей человеческой базы данных GenBank. Анализы пептидной или нуклеотидной последовательностей в полных белках или на предположительно зрелых пептидных участках Ucn III и Ucn II позволяют предположить, что они представляют собой отдельную ветвь CRF-семейства, характеризуясь более тесным родством друг с другом, чем с другими представителями (фиг.2D). С учетом 76% идентичности в предположительно зрелом пептидном домене можно предполагать, что Ucn III и URP иглобрюхих рыб являются ортологами. В отсутствие идентификации любых иных новых урокортин-родственных пептидов, вполне можно предположить, что ген человеческого урокортин-родственного пептида, который наиболее близок к мышиному Ucn II, является ортологом человеческого Ucn II. Указанный факт позволяет предположить, что ген человеческого URP должен быть переименован в человеческий Ucn II. Тот факт, что человеческий Ucn II не содержит консенсусных остатков протеолитического процессинга на предположительном С-концевом участке (хотя имеется потенциальный донор амида - глицин) поднимает вопрос о возможности того, что прогормон человеческого Ucn II может не образовывать пептид, аналогичный по размеру другим представителям семейства. Химическая природа и функции урокортина II у человека, несмотря на ортологическое родство между мышиным и человеческим Ucn II, остается не выясненной.
И мышиный, и человеческий Ucn III характеризуются высокой селективностью в отношении CRF-рецепторов типа 2 и аналогично Ucn II демонстрируют низкую аффинность в отношении рецепторов типа 1, а также минимальную способность индуцировать образование циклического АМФ в клетках, экспрессирующих либо эндогенный (кортикотропы передней доли гипофиза; фиг.3В), либо трансфицированный (таблица 2) CRF-R1. Человеческий Ucn III характеризуется меньшей аффинностью в отношении рецепторов типа 2, чем либо мышиный Ucn III, либо человеческий или мышиный Ucn II. Существует отличие только по четырем остаткам между человеческим и мышиным Ucn III, предоставляющее возможность исследования взаимоотношения структура/функция с точки зрения потребностей в высокой аффинности в отношении связывания CRF-R2. Человеческий Ucn III также менее активен в отношении человеческого CRF-R2α (данные не приведены), что указывает на то, что различия в аффинности между мышиным и человеческим Ucn III не связаны с типом источника рецептора. Однако несмотря на относительно низкую связывающую способность в отношении CRF-R2, человеческий Ucn III функционально достаточно активен (ЕС50<1 нМ) для стимуляции образования циклического АМФ клетками, экспрессирующими указанный рецептор. В этой связи следует отметить, что человеческий и мышиный Ucn III демонстрируют достаточную активность для того, чтобы действовать в качестве нативных лигандов для CRF-R2. Ни один из селективных лигандов для CRF-R2, Ucn III или Ucn II не связывается с CRF-BP с высокой аффинностью. И наоборот, два лиганда с высокой аффинностью в отношении CRF-R1 обладают также высокой аффинностью в отношении CRF-BP.
На основании результатов анализа по защите от расщепления РНКазой можно видеть, что урокортин III может быть доступен для рецепторов, происходящих из CRF-R2 в мозге и на периферии. Что касается периферии, то мРНК Ucn III была обнаружена в тонком кишечнике и в коже, хотя еще предстоит определить типы клеток, в которых экспрессируется мРНК Ucn III. Было показано, что в ЖК тракте CRF-R2 участвует в модуляции перистальтики кишечника (31).
В мозге мРНК Ucn III была найдена в дискретных подкорковых участках, где ее распределение отличается от такового для CRF (32), Ucn (33) и Ucn II (21). Несмотря на то, что для определения условий, в которых может действовать Ucn III, необходимо дождаться результатов подробных иммуногистохимических исследований и функционального анализа, некоторые предположения могут быть сделаны на основании его основных сайтов экспрессии, локализованных в медиальном предоптическом ядре, называемом в рамках настоящего описания ростральным перифорникальным участком, и в медиальном ядре миндалин. За счет притока из циркумвентрикулярных структур терминальной пластинки и ствола головного мозга медиальное предоптическое ядро можно рассматривать как ключевой сайт для интеграции нервных и гуморальных сигналов от внутренних органов в процессе поддержания гомеостаза жидкости и сердечно-сосудистой системы (34, 35). Среди основных мишеней его воздействия имеются множественные нейросекреторные и предавтономные популяции паравентрикулярно и/или супраоптического ядер гипоталамуса (36), в отношении которых, по меньшей мере для некоторых из них, известно, что они экспрессируют CRF-R2 (37).
Оказалось определенно труднее присвоить какие-либо вероятные функции ростральной перифорникальной группе клеток, экспрессирующих Ucn III, поскольку указанный кластер охватывает несколько цитоархитектурно определенных клеточных групп, и он отличается от перифорникального ядра гипоталамуса, идентифицированного некоторыми авторами (38). Можно указать, однако, что перифорникальный участок был идентифицирован как чувствительный сайт воздействия ряда нейроактивных агентов в отношении стимуляции пищеварительной функции (39-41), а также воздействия аминокислотных возбудителей при осуществлении сердечно-сосудистых реакций (42). Перифорникальный участок был идентифицирован как достигающий таких основных сайтов экспрессии CRF-R2, как ядро боковой перегородки и вентромедиальное ядро гипоталамуса (43-45), а источники локальных ингибиторных (GABA-эргических) проекций, достигающие стресс-родственных нейроэндокринных и автономных эффекторов в паравентрикулярном ядре, включают ростральный перифорникальный компонент (46), локализация которого очень близка к локализации экспрессирующих Ucn III нейронов.
Что касается третьего основного сайта экспрессии Ucn III, идентифицированного в настоящем изобретении, то анатомические и функциональные исследования указывают на то, что медиальное ядро миндалины простирается интенсивно в сторону гипоталамуса (включая вентромедиальное ядро) и другие лимбические структуры переднего мозга (47), а также участвует в модуляции поведенческих функций (48, 49) и нейроэндокринной функции (50, 51), в частности имеющие отношение к репродукции и стрессу. В целом, приведенные в настоящем описании центральные сайты экспрессии Ucn III, соответствуют возможной роли указанного пептида в модуляции относящихся к стрессу автономных, нейроэндокринных и поведенческих функций, возможно включающих некоторые из тех, что ранее рассматривались как придаток других членов пептидного семейства.
Распределение каждого из трех мышиных урокортинов демонстрирует определенный потенциал с точки зрения взаимодействия с CRF-рецепторами типа 2, при этом каждый из них демонстрирует высокую аффинность к указанным рецепторам. В противоположность этому сам CRF высоко селективен в отношении CRF-R1 и обладает меньшей аффинностью в отношении CRF-R2. В рамках предположения о том, что номенклатура для трех урокортин-родственных пептидов будет принята, резонно рассматривать CRF-R2 как рецептор урокортина и по функции, и по названию. Термин "CRF-система" в настоящее время включает лиганды, обладающие селективностью для каждого типа рецептора, а также бивалентный лиганд-урокортин.
Литература
1 Rivier C. and W.Vale (1983) Nature 305, 325-327.
2. Rivier J., C.Rivier, and W.Vale (1984) in European Peptide Symposium. Djuronaset Sweden, pp.104.
3. Vale et al., (1981) Science 213, 1394-1397.
4. Koob G.F. & Heinrichs, S.C. (1999) Brain Res. 848, 141-152.
5. Vaughan et al., (1995) Nature 378, 287-292.
6. Lederis et al., (1982) Proc. West.Pharmacol. Soc.25, 223-227.
7. Montecucchi et al., (1980) Int. J.Pept. Protein Res.16, 191-199.
8. Chen R. et al. (1993) Proc. Natl. Acad. Sci. USA90, 8967-8971.
9. Perrin et al. (1995) Proc. Natl. Acad. Sci. USA 92, 2969-2973.
10. Chen et al., (1993) in 23rd Annual Meeting of The Society for Neuroscience, Washington, DC, pp.238.
11. Vita et al., (1993) FEBS 335, 1-5.
12. Chang et al., (1993) Neuron 11, 1187-1195.
13. Perrin et al., (1993) Endocrinology 133, 3058-3061.
14. Kishimoto et al., (1995) Proc. Natl. Acad. Sci. USA 92, 1108-1112.
15. Lovenberg et al., (1995) Proc. Natl. Acad. Sci. USA 92, 836-840.
16. Van Pett et al., (2000) J. Comp. Neurol. 428, 191-212.
17. Potter E. et al. (1994) in 76th Annual Meeting of The Endocrine Society Anaheim, CA p.217.
18. Lovenberg T.W. et al., (1995) Endocrinology 136, 3351-3355.
19. Rohde, E. et al. (1996) Bigchem Pharmacol 52(6), 829-33.
20. Bale T.L. et al. (1999) Nat. Genet. 24(4), 410-414.21.
21. Reyes et al., (2001) Proc. Natl. Acad. Sci. USA 98, 2843-2848.
22. Pedersen A.G. & Nielsen, H. (1997) Proc. Int. Conf. Intell. Syst. Mol. Biol. 5, 226-233.
23. Devi L. (1991) Fed. Eur. Biochem. Soc. 280, 189-194.
24. Miranda et al., E. (1994) J. Med. Chem. 37, 1450-1459.
25. Miller C. & Rivier J. (1996) Biopolymers 40, 265-317.
26. Sutton et al., (1995) Endocrinology 136, 1097-1102.
27. Kageyama K., Suda T. & Vale W.W. (2001).
28. Vale et al., (1983) in Methods in Enzymology:
Neuroendocrine Peptides, ed. Conn P.M. (Academic Press, New York), Vol.103, pp.565-577.
29. Bilezikjian et al., (1996) Endocrinology 137, 4277-4284.
30. Simmons D.M., Arriza J.L. & Swanson, L.W. (1989) J. Histotechnol, in press.
31. Nozu T., Martínez, V. Rivier, J. Taché, Y. (1999) Am. J.Physiol.: Gastrointest. Liver Physiol. 39, G867-G874.
32. Swanson et al., (1983) Neuroendocrinol 36, 165-186.
33. Bittencourt et al., (1999) J. Comp. Neurol. 415, 285-312.
34. Johnson A.K. & Loewy A.D. (1990) in Central Regulation of Autonomic Functions, eds. Loewy A.D. & Spyer K.M. (Oxford University Press, New York), pp.246-267.
35. Saper C.B. & Levisohn D. (1983) Brain Res. 288, 21-31.
36. Sawchenko et al., (1983) J. Comp. Neurol. 218, 121-144.
37. Chalmers et al., (1995) J. Neurosci. 15, 6340-6350.
38. Paxinos, G. & Watson, C. (1986) Academic Press, San Diego, California.
39. Gillard et al., (1998) J. Neurosci. 18, 2646-2652.
40. Stanley et al., (1993) Brain Res. 604, 304-317.
41. Leibowitz, S.F. & Rossakis, C. (1979) Brain Res. 172, 101-113.
42. Allien et al., (1993) J. Comp. Neurol. 330, 421-438.
43. Kita H. & Oomura, Y. (1982) Brain Res. Bull. 8, 53-62.
44. Onteniente B., Menetrey D., Arai R. & Calas A. (1989) Cell Tissue Res. 256, 585-592.
45. Szeidemann et al., (1995) J. Comp. Neurol. 358, 573-583.
46. Roland et al., (1993) J. Comp. Neurol. 332, 123-143.
47. Canteras et al., (1995) J. Comp. Neurol. 360, 213-245.
48. Newman S.W. (1999) Ann. N. Y. Acad. Sci. 877, 242-257.
49. Rajendren G. & Moss, R. L. (1993) Brain Res. 617, 81-86.
50. Feldman et al., (1990) Neuroscience 37, 775-779.
51. Dayas et al., (1999) Eur. J. Neurosci. 11, 2312-2322.
Любые указанные в настоящем описании патенты или публикации соответствуют уровню специалистов в данной области техники, на которых рассчитано настоящее изобретение. Указанные патенты и публикации включены в качестве ссылок в той же степени, как если бы каждая индивидуальная публикация, выполненная в качестве ссылки, была указана индивидуально.
Специалист в данной области может без труда оценить, что настоящее изобретение хорошо адаптировано для осуществления всех заявленных в нем целей и получения результатов и указанных преимуществ, а также для осуществления других свойственных ему целей. Приведенные в настоящем описании примеры вместе с методами, процедурами, обработками, молекулами и конкретными соединениями, приведенными в настоящем описании, являются репрезентативными для предпочтительных вариантов его осуществления. Они являются иллюстрацией и не рассматриваются как ограничивающие область настоящего изобретения. Специалистами в данной области техники могут быть внесены изменения и предприняты другие варианты использования, которые охватываются рамками и принципами настоящего изобретения, определяемыми формулой изобретения.











| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛКИ УРОКОРТИНА И ИХ ПРИМЕНЕНИЯ | 2001 |
|
RU2278871C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, СВЯЗАННЫЕ С РАСТВОРИМЫМИ СОПРЯЖЕННЫМИ С G-БЕЛКОМ РЕЦЕПТОРАМИ (SGPCR) | 2006 |
|
RU2423378C2 |
| СПОСОБ ПОЛУЧЕНИЯ НЕПРИРОДНОЙ ТРАНСГЕННОЙ МЫШИ С ДЕФИЦИТОМ ГЕНА РЕЦЕПТОРА 2 КОРТИКОЛИБЕРИНА И ЕЕ ИСПОЛЬЗОВАНИЕ | 2000 |
|
RU2251838C2 |
| АГОНИСТЫ РЕЦЕПТОРА КОРТИКОТРОПИН-РИЛИЗИНГ ФАКТОРА 2 | 2003 |
|
RU2286995C2 |
| АГОНИСТЫ РЕЦЕПТОРА КОРТИКОТРОПИН-РИЛИЗИНГ ФАКТОРА 2 (CRFR) ПРИМЕНЕНИЕ | 2003 |
|
RU2294333C2 |
| АГОНИСТЫ РЕЦЕПТОРА КОРТИКОТРОПИН-РИЛИЗИНГ ФАКТОРА 2 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2305110C2 |
| СИСТЕМНАЯ ДОСТАВКА И РЕГУЛИРУЕМАЯ ЭКСПРЕССИЯ ПАРАКРИННЫХ ГЕНОВ ДЛЯ ЛЕЧЕНИЯ СЕРДЕЧНО-СОСУДИСТЫХ И ИНЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2642605C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТВОРИМОГО FcR В ВИДЕ Fc-ГИБРИДА С ИНЕРТНОЙ Fc-ОБЛАСТЬЮ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2630659C2 |
| АНТИ-CXCR3 АНТИТЕЛА | 2013 |
|
RU2663141C2 |
| СПОСОБ И НАБОР ДЛЯ ВЫЯВЛЕНИЯ ГЕНА, КОДИРУЮЩЕГО МЕМБРАНОСВЯЗАННЫЙ БЕЛОК, ВЕКТОР (ВАРИАНТЫ) | 1999 |
|
RU2246538C2 |
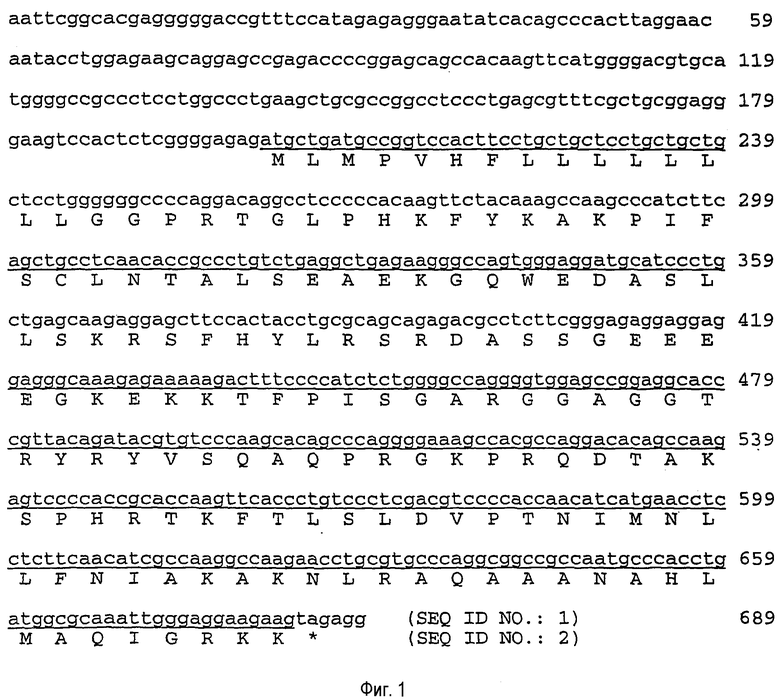


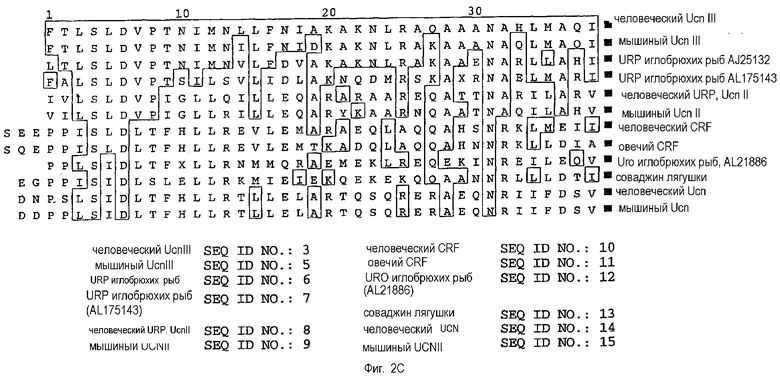
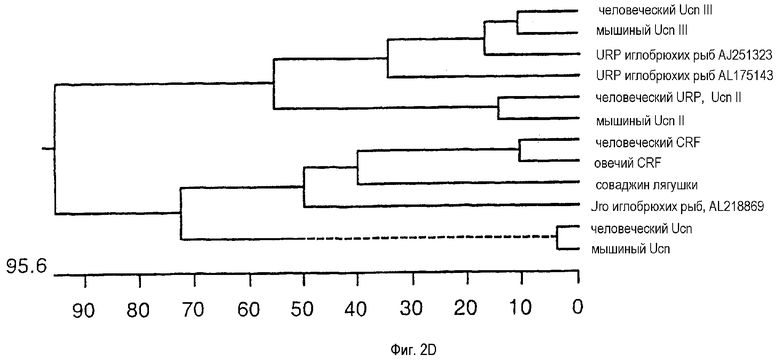
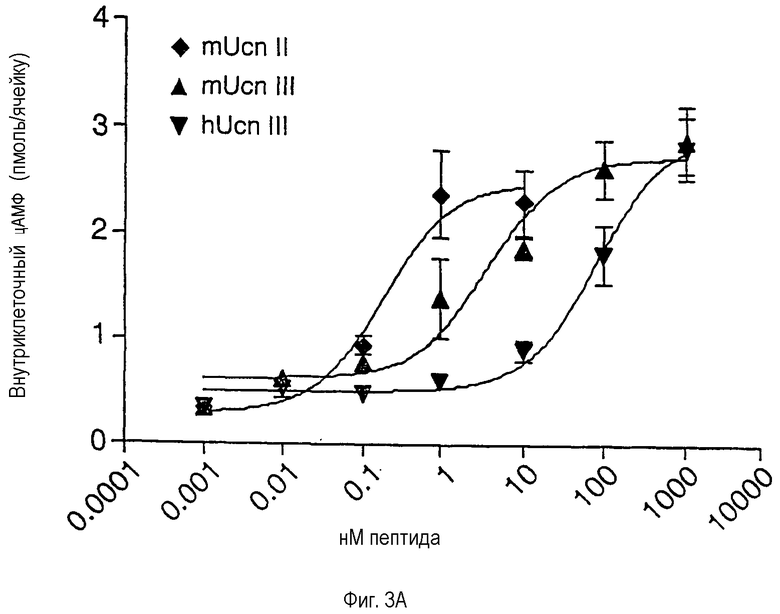
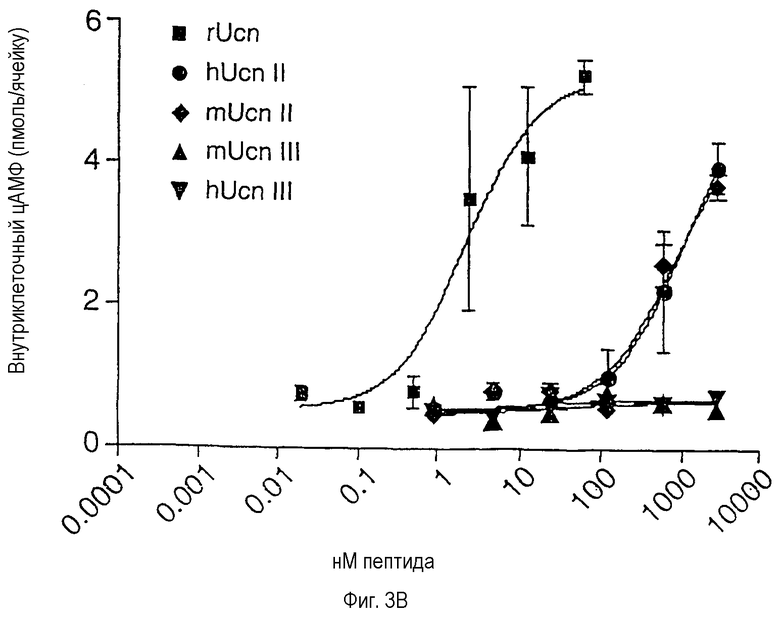
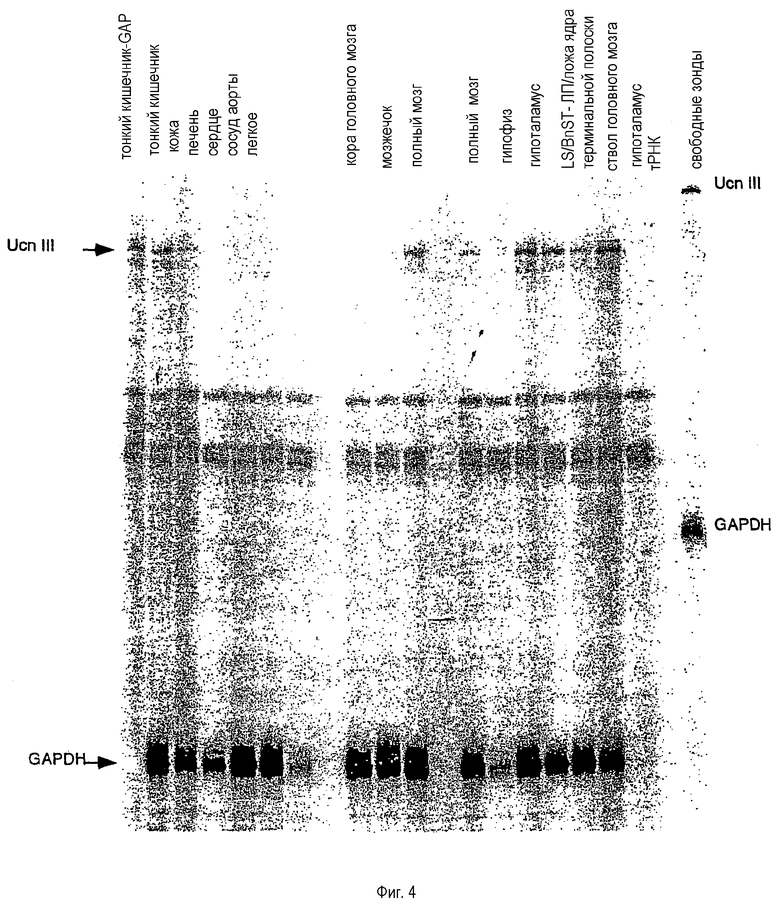
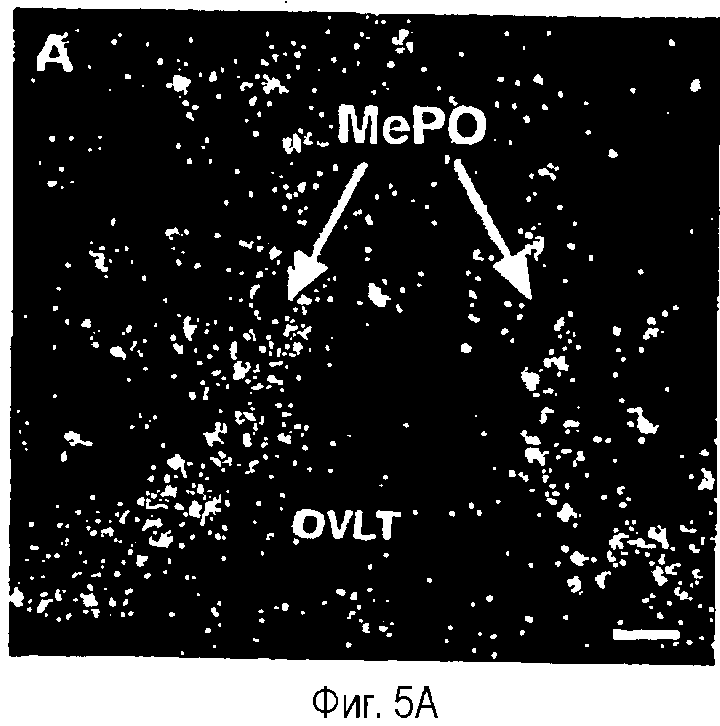
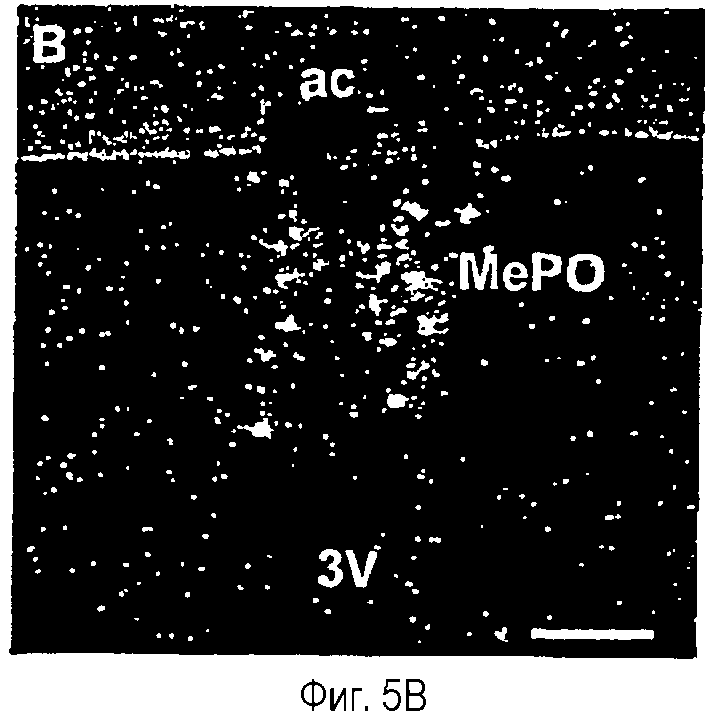
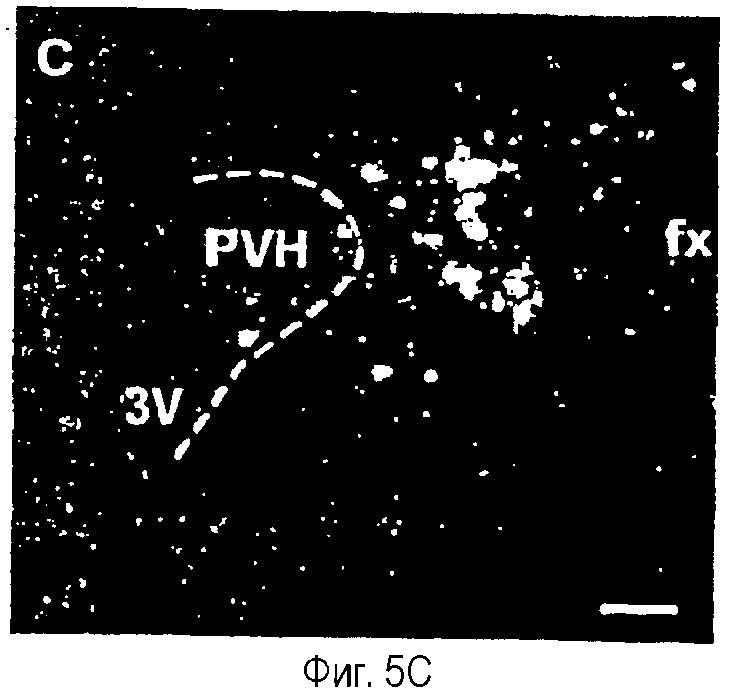
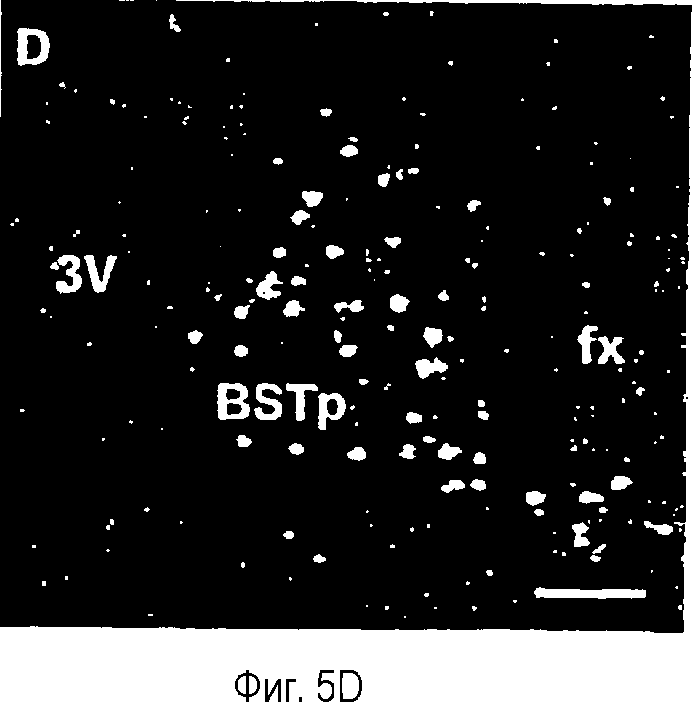
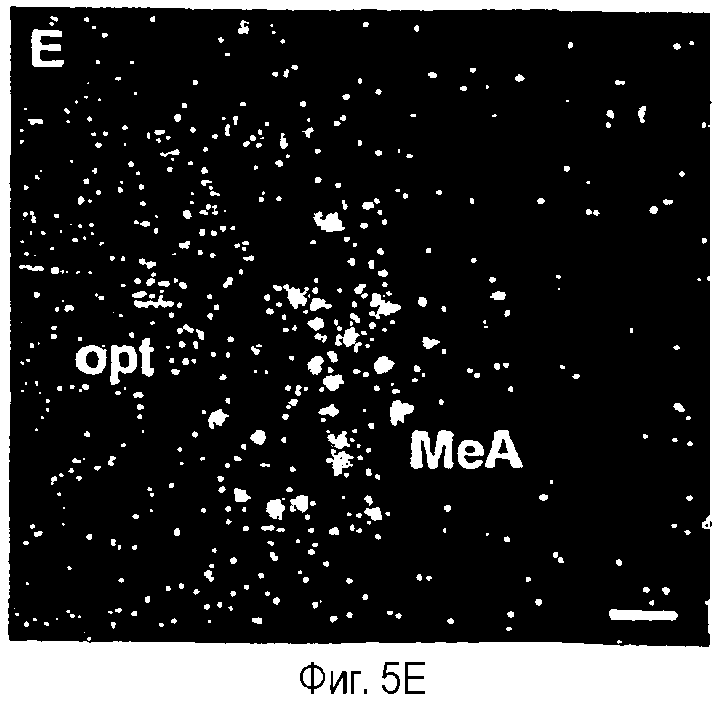
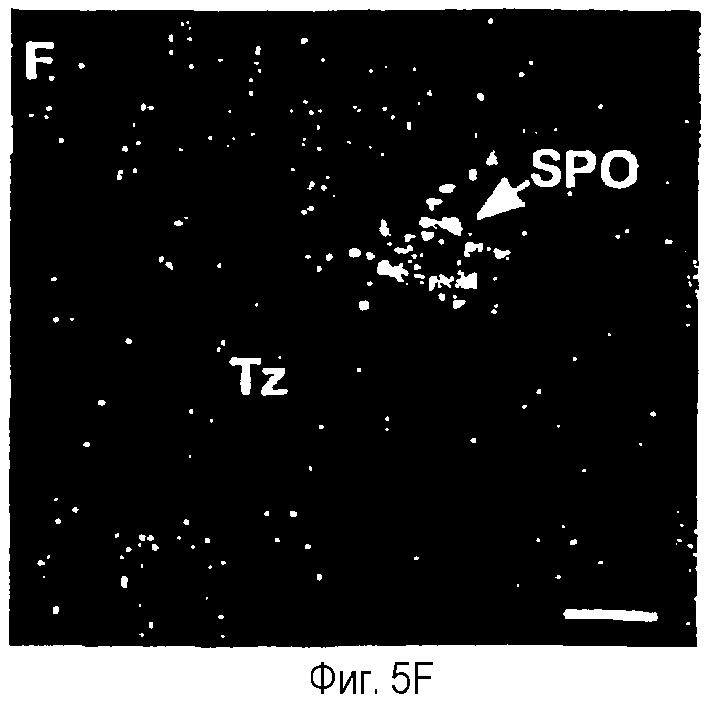
Изобретение относится к области молекулярной биологии и биохимии и может быть использовано в медицине. Из геномной ДНК человека и мыши получены и определены полные последовательности, кодирующие новый белок семейства кортикотропин-рилизинг-факторов, получивший название урокортина III (Исп III). Основной особенностью Исп III является повышенное сродство к рецептору CRF-2 α или β в сравнении с рецептором CRF-1, которое определяет возможность его функционирования в качестве агониста рецептора CRF-2. Получены синтетические аналоги Исп III, сохраняющие способность нативного белка к избирательному взаимодействию с CRF-2. Предлагается использовать новый белок и его аналоги в составе фармацевтических композиций, которые предназначаются для лечения патофизиологических состояний, связанных с необходимостью активации CRF-2, в частности застойной сердечной недостаточности, дисфункции желудочно-кишечного тракта, сахарном диабете, мигрени и т.п. 5 н. и 9 з.п. ф-лы, 10 ил., 3 табл.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| RU 97117102 А, 10.08.1999. | |||

