В настоящей заявке, в соответствии со ст. 35 § 119 Кодекса законов США, испрашивается приоритет на основании предварительной заявки на патент США №61/588936, поданной 20 января 2012 года, которая во всей своей полноте включена в настоящее описание посредством ссылки.
Настоящее изобретение относится к антителам и к способам применения антител для лечения расстройств, ассоциированных с передачей CXCR3-сигнала, таких как сахарный диабет типа 1 (диабет типа I; T1D).
Диабет характеризуется хронической гипергликемией, вызываемой отсутствием инсулиновой активности, а также различными характерными метаболическими расстройствами. Диабет может быть подразделен на две широкие категории: диабет типа I и типа II. T1D характеризуется потерей панкреатических β-клеток островков Лангерганса, а диабет типа II характеризуется снижением уровня секреции инсулина и чувствительности к инсулину (инсулинорезистентностью). В Соединенных Штатах диабетом страдает приблизительно 2-4 процента населения, причем на диабет типа I (также известный как инсулинзависимый диабет или ИЗСД) приходится приблизительно 7-10 процентов от всех случаев заболевания диабетом.
Диабет типа I характеризуется низким продуцированием инсулина, то есть на уровне, недостаточном для поддержания гомеостаза глюкозы. Такое расстройство, вероятно, инициируется разрушением панкератических β-клеток, опосредуемых аутоиммунным заболеванием. В развитии аутоиммунного заболевания, ассоциированного с диабетом типа I, участвуют аутореактивные B- и T-лимфоциты. Действительно, до 98% пациентов, страдающих диабетом типа I, имеют антитела против одного или более собственных β-клеточных антигенов, включая инсулин, (глутаминовая кислота)декарбоксилазу (GAD), антиген-2 инсулиномы и антиген-2b инсулиномы (IA-2 и IA-2β), и гетерогенные цитоплазматические антигены островковых клеток (ICA). Уровень одного или более аутоантител обычно коррелирует с уровнем деструкции β-клеток, хотя это не всегда может быть определено. Irvine, et al., Diabetes, 26:138-47 (1997); Riley, et al., N. Engl. J. Med., 323:1167-72 (1990). В соответствии с этим, аутоантитела могут служить индикаторами развития аутоиммунного диабета и, вместе с метаболическими расстройствами, позволяют предсказать риск развития диабета T1D у родственников пациентов.
Развитие диабета типа I может опосредоваться аутореактивными T-клетками, о чем свидетельствуют тканевые биоптаты, полученные на время диагностирования T1D и указывающие на инфильтрацию островков активированными Т-клетками. Bottazzo et al., N. Engl J. Med., 313:353-60 (1985); Hanninen et al., J. Clin. Invest, 90:1901-10 (1992); Itoh et al., J. Clin, invest, 92:2313-22 (1993); Imagawa, et al., Diabetes, 50:1289-73 (2001).
Хемокиновый (мотив C-X-C) рецептор 3 (CXCR3), также известный как связанный с G-белком рецептор 9 (GPR9), CD183, рецептор IP-10 и рецептор Mig представляют собой хемокиновые рецепторы, экспрессированные на аутореактивных T-клетках, которые участвуют в различных физиологических процессах и в развитии ассоциированных с ними расстройств, таких как T1D. CXCR3 в основном отсутствует в «необученных» T-клетках, но индуцируется после активации этих клеток антигеном и рекрутинга активированных клеток на участки воспаления тканей в ответ на действие первичных лигандов: CXCL9, CXCL10 и CXCL11. Было обнаружено, что β-клетки преимущественно экспрессируют CXCL10, а также CXCL9 на низком уровне у мышей с моделью T1D (Christen et al The Journal of Immunology, 2003, 171:6838-6845; Morimoto et al. J. Immunol 2004; 173;7017-7024; Sarkar et al. Diabetes. 2012 Feb; 61 (2):436-46); и в островковых клетках у пациентов с T1D, страдающих инсулитом (Uno et al 2010; Roep et al. Clinical and Experimental Immunology, 2003, 159: 338-343; Sarkar et al. Diabetes. 2012 Feb; 61 (2):436-46). Кроме того, было обнаружено, что T-клетки, инфильтрирующиеся в поджелудочную железу, экспрессируют CXCR3 у мышей с моделью T1D и в образцах поджелудочной железы, взятых у пациентов с диабетом типа 1 (Christen et al., The Journal of Immunology, 2003, 171: 6838-6845; Van Halteren et al., Diabetologia 48:75-82 (2005); Uno et al 2010; Roep et al., Clinical and Experimental immunology, 2003, 159: 338-343; Sarkar et al., Diabetes. 2012 Feb; 61 (2):436-46). Кроме того, у мышей, дефицитных по CXCR3 («нокаут»-мышей), наблюдалось значительное замедление начала развития T1D и снижение заболеваемости T1D (Frigerio et al., Nature Medicine 8:1414-1420 (2002)), а сверхэкспрессия CXCL10 в островках трансгенных мышей приводила к стимуляции инфильтрации Т-клеток и к ускорению начала развития T1D (Rhode et al., J. Immunol. 175(6): 3516-24 (2005)). Было показано, что нейтрализация CXCL10 путем лечения антителом оказывала протективное действие (Christen et al., The Journal of immunology, 2003, 171: 6838-6845).
Существуют три изоформы CXCR3, обозначенные A, B и Alt., которые были идентифицированы у человека (Lasagni et al., J. Exp. Med. 2003 197:1537; Ehlert et al, J. Immunol. 2004; 173; 6234-6240). CXCR3-A связывается с CXC-мотивом хемокинов CXCL9 (MIG), CXCL10 (IP-10) и CXCL11 (I-TAC); CXCR3-B также связывается с этими мишенями, но он также связывается с CXCL4; а CXCR3-Alt, очевидно, взаимодействует с CXCL11. Хотя альтернативный сплайсинг приводит к генерированию нескольких изоформ белка CXCR3, однако, доминирующей формой in vivo является CXCR3-A, тогда как GXCR3-B и CXCR3-Alt экспрессируются на гораздо более низких уровнях белка. Lasagni et al., J. Exp. Med. 2003 197:1537; Ehlert et al., J. Immunol. 2004; 173; 6234-6240.
Попытки нарушить CXCR3-путь с использованием низкомолекулярных ингибиторов CXCR3 оказались абсолютно неэффективными. Christen et al., Clin Exp. Immunol. 165: 318-328 (2011). В соответствии с этим были исследованы антитела и другие методы, направленные на разрушение CXCL10, главным образом, до начала развития диабета. Morimoto et al., J. Immun. 173: 7017-7024 (2004); Oikawa et al., Rev. Diabet Stud. 7: 209-224 (2010).
Поскольку T1D и другие расстройства, в развитии которых участвует CXCR3, являются широко распространенными, то необходимо разработать дополнительные способы, нацеленные на подавление передачи CXCR3-сигнала, например, для лечения или подавления прогрессирования у пациента такого расстройства, как T1D.
В настоящей заявке описаны антитела и способы применения антител, обладающих способностью связываться с CXCR3. В некоторых вариантах осуществления изобретения, антитела могут быть использованы для предупреждения, лечения или подавления прогрессирования T1D на ранней стадии у индивидуума посредством ингибирования CXCR3-пути. При проведении исследований с применением антител и соответствующих способов было неожиданно обнаружено, что нейтрализующие антитела, направленные на CXCR3, по меньшей мере отчасти, могут предупреждать начало развития T1D у NOD-мышей при введении им этих антител до начала развития заболевания, либо они могут индуцировать обратное развитие заболевания при введении этих антител NOD-мышам на начальной стадии развития T1D. Кроме того, нейтрализация активности CXCR3 не связана со значительным нарушением нормальной функции иммунной системы пациента, а поэтому она не будет вызывать значительные нежелательные побочные эффекты при терапии антителами.
В соответствии с этим, в одном из своих аспектов настоящее изобретение относится к антителам и к их антигенсвязывающим фрагментам, способным нейтрализовать активность CXCR3. В некоторых вариантах осуществления изобретения, CXCR3-нейтрализующие антитела могут быть охарактеризованы как антитела, обладающие способностью связываться с пептидами, выбранными из остатков 1-58, 1-16 или 1-37 SEQ ID NO:1. В некоторых вариантах осуществления изобретения указанные антитела содержат все клоны антител или их части (Cl), обозначенные Cl 12, Cl 135, Cl 82, Cl 53 и/или Cl 4. В некоторых своих вариантах настоящее изобретение относится к вариантам антител Cl 12, Cl 135, Cl 82, Cl 53 и/или Cl 4, включая CDR-привитые варианты, гуманизированные варианты, варианты с обратной мутацией и полностью человеческие варианты описанных антител. В конкретных вариантах осуществления изобретения антитело содержит одну или более определяющих комплементарность областей (CDR), например, одну или более CDR1, CDR2 и CDR3 тяжелой цепи, и/или одну или более CDR1, CDR2 и CDR3 легкой цепи, происходящих от клонов Cl 12, C1 135, Cl 82, Cl 53 и/или Cl 4 или любых вариантов клонов 4, 12, 53, 82 и 135, описанных в настоящей заявке. В некоторых вариантах осуществления изобретения, антитела, происходящие от Cl 12, Cl 135, Cl 82, Cl 53 и/или Cl 4, или их химерные или гуманизированные варианты обладают некоторыми более предпочтительными свойствами, чем клоны анти-CXCR3 антител 5H7, 7H5, V44D7, 1C6 и/или 49801. Так, например, описанные здесь антитела могут обладать более высокой аффинностью связывания по сравнению с клонами анти-hCXCR3 антител 5H7, 7H5, V44D7, 1C6 и 49801. Так, например, указанное антитело может обладать аффинностью, которая в 1, 2, 3, 4, 5 или более раз (или в любое промежуточное число раз) превышает аффинность анти-CXCR3 антител, таких как 1C6, например, как было измерено методом поверхностного плазмонного резонанса (например, с помощью анализа BiACORE™). Описанные здесь гуманизированные антитела также обладают прогнозируемой способностью к снижению иммуногенности по сравнению с клонами мышиных анти-hCXCR3 антител 5H7, 7H5, V44D7, 1C6 и 49801. Кроме того, тяжелые цепи описанных здесь клонов 4.7-4.11 были оптимизированы для удаления сайта дезамидирования в положениях 58 и 59 (в соответствии с нумерацией IMGT) и для повышения стабильности по сравнению с последовательностью CDR2 вариабельного домена (VH) тяжелой цепи исходного мышиного анти-hCXCR3 антитела.
В другом своем аспекте настоящее изобретение относится к способам профилактики начала развития T1D, а также к способам лечения или замедления прогрессирования впервые выявленного T1D у индивидуума путем введения эффективного количества CXCR3-нейтрализующего антитела. В конкретных вариантах осуществления изобретения указанным индивидуумом является млекопитающее, например, человек.
В некоторых вариантах осуществления изобретения, индивидуума, у которого был впервые выявлен T1D, подвергают лечению описанными здесь способами через 6 месяцев после установления клинического диагноза. В других вариантах осуществления изобретения, индивидуума подвергают лечению в течение более 6 месяцев после установления клинического диагноза, где у указанного индивидуума сохраняются общие остаточные уровни С-пептидов в сыворотке натощак, составляющие по меньшей мере приблизительно 0,2 нмоль/л.
В некоторых вариантах осуществления изобретения индивидуумы могут быть охарактеризованы как индивидуумы, у которых наблюдаются повышенные уровни глюкозы в крови натощак в отсутствие экзогенного инсулина, превышающие 120 мг/дл, или аномально низкий общий уровень С-пептидов в сыворотке натощак, составляющий приблизительно 0,033-1,0 нмоль/л в минуту во время стимуляции C-пептидом. В конкретных вариантах осуществления изобретения, CXCR3-нейтрализующее антитело вводят в дозе приблизительно 0,03-3,7 мг/кг/дозу. В некоторых вариантах осуществления изобретения индивидууму вводят по меньшей мере одну дозу антитела. В некоторых вариантах осуществления изобретения индивидууму повторно вводят дозы антитела (например, по меньшей мере ежегодно, один раз в три месяца, один раз в два месяца, один раз в месяц, один раз в две недели, один раз в неделю или ежедневно). В других вариантах осуществления изобретения описанные выше способы могут также включать стадию введения иммунодепрессанта и/или агента, стимулирующего β-клетки, и одновременное или последовательное введение (перед этой стадией или после нее) CXCR3-нейтрализующего антитела.
В различных вариантах осуществления изобретения описанные здесь анти-CXCR3 антитела вводят для лечения состояния, характеризующегося аномальной экспрессией CXCR3. В некоторых вариантах осуществления изобретения, анти-CXCR3 антитела вводят для лечения любого состояния, которое может поддаваться лечению посредством ингибирования и/или нейтрализации активности CXCR3. В некоторых вариантах осуществления изобретения описанные здесь анти-CXCR3 антитела вводят для лечения T1D.
Другие варианты и преимущества настоящего изобретения будут представлены ниже и будут отчасти более очевидны из нижеследующего описания либо из описания практического осуществления настоящего изобретения. Такие варианты и преимущества настоящего изобретения могут быть реализованы и достигнуты с использованием элементов и их комбинаций, указанных, в частности, в прилагаемой формуле изобретения.
Совершенно очевидно, что представленное выше общее описание и представленное ниже подробное описание имеют лишь иллюстративный характер и приводятся для лучшего понимания настоящего изобретения, но при этом они не должные рассматриваться как ограничение заявленного объема изобретения.
Прилагаемое описание графического материала, которое вводится в описание настоящей заявки и составляет ее часть, представлено в целях иллюстрации одного (или нескольких) из вариантов осуществления изобретения, и вместе с таким описанием позволяет лучше понять принципы настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
На фиг. 1 проиллюстрирована экспрессия инсулина (левая панель), CXCL10 (центральная панель) и CD3 (правая панель) в срезах поджелудочной железы, взятых у 6-недельных самок NOD-мышей (первый ряд), у 10-недельных самок NOD-мышей (второй ряд), и у самок NOD-мышей с только что зарегистрированным диабетом (третий ряд).
На фиг. 2 проиллюстрирован проточный цитометрический анализ на экспрессию CXCR3 на T-клетках поджелудочной железы самок NOD-мышей с только что зарегистрированным диабетом. CD4+- и CD8+-T-клетки идентифицировали и окрашивали на экспрессию CXCR3, как показано сплошной линией на двух нижних графиках. Окрашивание контрольного изотипа показано на заштрихованной кривой на тех же самых двух графиках.
На фиг. 3 указаны проценты самок NOD-мышей с отсутствием диабета в зависимости от времени, которым, по достижении ими 10-недельного возраста, вводили PBS, анти-CXCR3 антитело и контрольный IgG до начала развития диабета. Результаты двух независимых исследований представлены на фиг. 3A и 3B.
На фиг. 4 показаны срезы поджелудочной железы 26-недельных самок NOD-мышей с отсутствием диабета, которым, по достижении ими 10-недельного возраста, вводили анти-CXCR3 антитело в профилактических целях, где указанные срезы были окрашены на инсулин (левая панель) или CD3/Foxp3 (центральная и правая панели). На правой панели показана увеличенная фотография среза, представленного на центральной панели.
На фиг. 5 указаны величины уровней глюкозы в крови, ежедневно регистрируемые утром у самок NOD-мышей, обработанных PBS, анти-CXCR3 антителом, контрольным IgG, мышиным антителом против глобулина тимоцитов (мышиного тимоглобулина, mATG), через 3-4 дня после предполагаемого развития диабета у мышей. Каждая линия представляет отдельную мышь. Стрелками указаны дни обработки.
На фиг. 6A представлены гистограммы, на которых указан процент T-клеток самок NOD-мышей, обработанных PBS, анти-CXCR3 антителом, контрольным IgG и антителом против mATG, где указанными Т-клетками являются CD4+-клетки (левая панель) и CD8+-клетки (правая панель). Поджелудочную железу брали у мышей во время обработки после 5-й инъекции тестируемого препарата и у mATG-обработанных мышей соответствующего возраста. На фиг. 6B представлен график зависимости уровня экспрессии CD44 (вертикальная ось) и экспрессии CD62L (горизонтальная ось) на CD4+-клетках, выделенных из поджелудочной железы мышей, обработанных PBS, контрольным антителом или анти-CXCR3 антителом. G1 и G2 относятся к гейтированным Т-клеткам с высоким уровнем CD44/низким уровнем CD62L и с низким уровнем CD44/низким уровнем CD62L соответственно. На фиг. 6С проиллюстрирован уровень экспрессии CXCR3 на CD4+-клетках в G1 и G2, по сравнению с уровнем экспрессии CXCR3 на клетках, окрашенных антителом контрольного изотипа и гейтированных на лимфоцитах.
На фиг.7 показаны срезы поджелудочной железы самок NOD-мышей, обработанных контрольным IgG (левые панели), анти-CXCR3 антителом (центральные панели) и mATG (правые панели), где указанные срезы были окрашены на инсулин (верхний ряд) или CD3/Foxp3 (нижний ряд).
На фиг. 8A-D представлен график зависимости уровней глюкозы в крови, регистрируемых после введения глюкозы самкам NOD-мышей соответствующего возраста с отсутствием диабета (фиг. 8A), NOD-мышам с диабетом, обработанных PBS (фиг. 8В), NOD-мышам с ремиссией заболевания, обработанных анти-CXCR3 антителом (фиг. 8С), и NOD-мышам с диабетом, обработанных контрольным антителом IgG (фиг. 8D). Введение глюкозы мышам проводили через 100 дней после появления первых признаков диабета и распределения мышей по группам исследования. Каждая линия представляет данные для каждого отдельного животного.
На фиг. 9A-B указаны проценты мышей с отсутствием диабета в зависимости от времени для NOD.Scid-мышей, которым вводили пулы донорных CD4+- и CD8+-Т-клеток, выделенных у самок NOD-мышей, обработанных PBS, анти-CXCR3 антителом, контрольным IgG или анти-mATG антителом. Т-клетки были выделены у самок NOD-мышей с диабетом приблизительно через 80-90 дней после введения PBS или контрольного IgG, или у самок NOD-мышей с ремиссией заболевания приблизительно через 80-90 дней после введения анти-CXCR3 антитела или анти-mATG антитела. Результаты этих двух независимых исследований представлены на фиг. 9A и 9B.
На фиг. 10A указан процент донорных CD4+- и CD8+-Т-клеток, выделенных у самок NOD-мышей, обработанных PBS, анти-CXCR3 антителом, контрольным IgG или анти-mATG антителом (левая панель), как описано на фиг. 9. Процент эффекторных клеток и центральных клеток памяти в пулах донорных CD4+- и CD8+-Т-клеток для каждой группы обработки представлен на правых панелях фиг. 10А. На фиг. 10B указан процент регуляторных Т-клеток в пулах донорных Т-клеток, идентифицированных по экспрессии CD4 и CD25 или по экспрессии CD4, CD25 и Foxp3. На фиг. 10C указан процент CD8+-клеток (левая панель) и CD4+-клеток (правая панель) в пулах донорных Т-клеток, которые также экспрессируют CXCR3.
На фиг. 11A-B указан процент мышей с отсутствием диабета в зависимости времени после адоптивного переноса Т-клеток, взятых у донорных OVA-специфических TCR-трансгенных мышей, мышам-реципиентам RIP-OVA, которых либо не обрабатывали, либо обрабатывали анти-CXCR3 антителом или контрольным IgG. Результаты этих двух исследований представлены на фиг. 11А и 11В.
На фиг. 12A проиллюстрирована экспрессия CXCR3 на донорных Т-клетках, проанализированная с помощью проточной цитометрии до адоптивного переноса этих клеток мышам-реципиентам RIP-OVA (заштрихованная кривая). Окрашивание антителом контрольного изотипа проиллюстрировано на затененной кривой. На фиг. 12B указан процент донорных клеток в крови, селезенке и лимфоузлах поджелудочной железы мышей-реципиентов, обработанных анти-CXCR3 антителом или контрольным антителом IgG на дни 2, 4, 7, 9 и 15 после адоптивного переноса. На фиг. 12C указан процент пролиферирующих донорных клеток в крови, селезенке и лимфоузлах поджелудочной железы мышей-реципиентов, обработанных анти-CXCR3 антителом или контрольным антителом IgG после адоптивного переноса. На фиг. 12D указан процент донорных CXCR3+-клеток в крови, селезенке и лимфоузлах поджелудочной железы мышей-реципиентов, обработанных анти-CXCR3 антителом или контрольным антителом IgG после адоптивного переноса.
На фиг. 13 показаны срезы поджелудочной железы необработанных мышей-реципиентов RIP-OVA, и эти срезы были окрашены на инсулин (вверху слева) или CD3 (вверху справа), или срезы мышей, которые были обработаны анти-CXCR3 антителом, и эти срезы были окрашены на инсулин (внизу слева) или CD3 (внизу справа). Поджелудочную железу выделяли через 60 дней после адоптивного переноса донорных Т-клеток.
На фиг. 14A-C представлен уровень ингибирования CXCR3-опосредуемого хемотаксиса CXCL11, опосредуемого клонами Cl 4, 12, 53, 82 и 135. Данные выражены как средние относительные единицы флуоресценции (RFU) мигрирующих клеток в анализе на хемотаксис. На фиг. 14D указана концентрация антитела, необходимая для ингибирования мобилизации кальция на 50% клонами антител Cl 4, 12, 53 и 135.
На фиг. 15A-C представлен уровень ингибирования CXCR3-опосредуемого хемотаксиса CXCL9 (фиг. 15A), CXCL10 (фиг. 15B) и CXCL11 (фиг. 15C), опосредуемого клонами Cl 4, 12, 53, 82 и 135. Данные выражены как средние относительные единицы флуоресценции (RFU) мигрирующих клеток в анализе на хемотаксис.
На фиг. 16 графически представлены гистограммы, на которых проиллюстрировано связывание антитела с клетками, экспрессирующими различные другие хемокиновые рецепторы. Концентрация связанного антитела увеличивается по горизонтальной оси на каждой из гистограмм.
На фиг. 17A проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 4.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначенных VH1-3 и 7-11 и VK1-3). На фиг. 17А указаны последовательности тяжелой цепи, представленные как SEQ ID NO:18, 20, 22, 24, 29-33 и 659, и последовательности легкой цепи, представленные как SEQ ID NO:19, 25, 21, 23 и 660, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. На фиг. 17B проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 12.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначенных VH1-3 и VK1-3). На фиг. 17В указаны последовательности тяжелой цепи, представленные как SEQ ID NO:2, 4, 6, 8 и 661, и последовательности легкой цепи, представленные как SEQ ID NO:3, 5, 7, 9 и 662, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. На фиг. 17C проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 53.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначенных VH1-6 и VK1-9). На фиг. 17С указаны последовательности тяжелой цепи, представленные как SEQ ID NO:38, 40, 42, 44, 46-48 и 663, и последовательности легкой цепи, представленные как SEQ ID NO:39, 41, 43, 45,49-54 и 664, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. На фиг. 17D проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 82.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначаемых VH1-3 и VK1-3). На фиг. 17D указаны последовательности тяжелой цепи, представленные как SEQ ID NO:55, 57, 59, 61 и 665, и последовательности легкой цепи, представленные как SEQ ID NO:56, 58, 60, 62 и 666, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. На фиг. 17E проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 135.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначенных VH1-3 и VK1-3). На фиг. 17E указаны последовательности тяжелой цепи, представленные как SEQ ID NO:10, 12, 14, 16 и 667, и последовательности легкой цепи, представленные как SEQ ID NO:11, 13, 15, 17 и 668, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. На фиг. 17F проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 4.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначенных VH4-8 и VK4-7). На фиг. 17F указаны последовательности тяжелой цепи, представленные как SEQ ID NO:19, 34-37 и 669, и последовательности легкой цепи, представленные как SEQ ID NO:18, 27, 28 и 26, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. На фиг. 17G проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клона 53.0 (называемого «родительским» клоном) и некоторых гуманизированных вариантов (обозначенных VH7-10 и VK10-13). На фиг. 17G указаны последовательности тяжелой цепи, представленные как SEQ ID NO:38, 63-66 и 663, и последовательности легкой цепи, представленные как SEQ ID NO:39, 67-70 и 664, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. Нижняя последовательность в каждом выравнивании на фиг. 17A-G представляет собой последовательность, наиболее сходную с последовательностями человеческой зародышевой линии. В черных рамках указаны домены CDR, в которых затененные остатки в этой последовательности отличаются от соответствующих остатков зародышевой линии (фиг. 17A-E) или от соответствующих родительских остатков (фиг. 17F-G), где остатки пронумерованы в соответствии с IMGT и где используется делимитация границ CDR. На фиг. 17H проиллюстрировано выравнивание вариабельных доменов тяжелой (VH) и легкой (VK) цепей клонов 4.0, 12.0, 82.0 и 135, а также клонов антител 5H7 и 7H5. На фиг. 17H указаны последовательности тяжелой цепи, представленные как SEQ ID NO:18, 2, 38, 55, 10 и 670-671, и последовательности легкой цепи, представленные как SEQ ID NO:19, 3, 39, 56, 11 и 672-673, где все указанные последовательности, соответственно, приводятся для иллюстрации выравнивания. В черных рамках указаны домены CDR, в которых затененные остатки в этой последовательности отличаются от предыдущей выравниваемой последовательности, где остатки пронумерованы в соответствии с IMGT.
На фиг. 18 показаны границы остатков минимальных эпитопов для клонов антител 4, 12, 53, 82 и 135. Остатки, играющие важную роль в активности связывания, обозначены X. На фиг. 18 указана SEQ ID NO:81.
На фиг. 19 представлены гистограммы, иллюстрирующие связывание антитела с клетками 300.19, трансфицированным человеческим CXCR3 для химерных клонов 4, 12, 53, 82 и 135, а также для гуманизированных вариантов Hu1, Hu2, Hu3. Антитело вводили в концентрации 5 мкг/мл (черная линия), 0,5 мкг/мл (темно-серая линия) или 0,1 мкг/мл (черная пунктирная линия), либо вводили 5 мкг/мл только «второго» антитела (гистограмма, закрашенная серым), и по этим данным строили график зависимости числа клеток (по горизонтальной оси) от процента максимальной флуоресценции.
На фиг. 20A-C указан процент ингибирования миграции (вертикальная ось) клеток, трансфицированных человеческим CXCR3, в направлении CXCL9 (фиг. 20A), CXCL10 (фиг. 20B) и CXCL11 (фиг. 20C) в отсутствие или в присутствии 10 мкг/мл химерных (Chim) или гуманизированных (Hu1, Hu2 или Hu3) вариантов клонов антител 4, 12, 53, 82 и 135, или коммерчески доступного клона 1C6.
На фиг. 21 представлен график, на котором проиллюстрирована способность химерных (Chim) и гуманизированных (Hu1, Hu2 или Hu3) вариантов клонов антител 4, 12, 53, 82 и 135 и коммерчески доступного клона 1C6 ингибировать мобилизацию кальция в GXCR3-Gqi4qi4-трансфицированных клетках CHO. На графике представлена зависимость концентрации антитела (горизонтальная ось) от процента максимального ингибирования (вертикальная ось).
На фиг. 22A-D проиллюстрировано влияние обработки анти-CXCR3 антителом на процентное соотношение CD3+/CD4+-T-клеток (фиг. 22A), CD3+/CD8+ -T-клеток (фиг. 22D) у NOD-scid IL2rγnull (NSG)-мышей. HulgG1 означает человеческий IgG1 (герцептин), а термин «клоны 4, 12, 53, 82 и 135» означает клоны химерного антитела.
На фиг. 23A представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 12.0. На фиг. 23B представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 12.1. На фиг. 23C представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 12.2. На фиг. 23D представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 12.3.
На фиг. 24A представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 135.0. На фиг. 24B представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 135.1. На фиг. 24C представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 135.2. На фиг. 24D представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 135.3.
На фиг. 25A представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 4.0. На фиг. 25B представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 4.1. На фиг. 25C представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 4.2. На фиг. 25D представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 4.3. На фиг. 25E представлены аминокислотные последовательности тяжелой цепи клона антитела 4.4. На фиг. 25F представлены аминокислотные последовательности тяжелой цепи клона антитела 4.5. На фиг. 25G представлены аминокислотные последовательности тяжелой цепи клона антитела 4.6. На фиг. 25H представлены аминокислотные последовательности тяжелой цепи клона антитела 4.7. На фиг. 25I представлены аминокислотные последовательности тяжелой цепи клона антитела 4.8. На фиг. 25J представлены аминокислотные последовательности тяжелой цепи клона антитела 4.9. На фиг. 25K представлены аминокислотные последовательности тяжелой цепи клона антитела 4.10. На фиг. 25L представлены аминокислотные последовательности тяжелой цепи клона антитела 4.11. На фиг. 25M представлены аминокислотные последовательности легкой цепи клона антитела 4.4. На фиг. 25N представлены аминокислотные последовательности легкой цепи клона антитела 4.5. На фиг. 25О представлены аминокислотные последовательности легкой цепей клона антитела 4.6. На фиг. 25P представлены аминокислотные последовательности легкой цепи клона антитела 4.7.
На фиг. 26A представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 53.0. На фиг. 26B представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 53.1. На фиг. 26C представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 53.2. На фиг. 26D представлены аминокислотные последовательности тяжелой и легкой цепей клонов антитела 53.3. На фиг. 26E представлены аминокислотные последовательности тяжелой цепи клона антитела 53.4. На фиг. 26F представлены аминокислотные последовательности тяжелой цепи клонов антитела 53.5. На фиг. 26G представлены аминокислотные последовательности тяжелой цепи клона антитела 53.6. На фиг. 26H представлены аминокислотные последовательности легкой цепи клона антитела 53.4. На фиг. 26I представлены аминокислотные последовательности легкой цепи клона антитела 53.5. На фиг. 26J представлены аминокислотные последовательности легкой цепи клона антитела 53.6. На фиг. 26K представлены аминокислотные последовательности легкой цепи клона антитела 53.7. На фиг. 26L представлены аминокислотные последовательности легкой цепи клона антитела 53.8. На фиг. 26М представлены аминокислотные последовательности легкой цепи клона антитела 53.9. На фиг. 26N представлены аминокислотные последовательности тяжелой цепи клона антитела 53.7. На фиг. 26О представлены аминокислотные последовательности тяжелой цепи клона антитела 53.8. На фиг. 26P представлены аминокислотные последовательности тяжелой цепи клона антитела 53.9. На фиг. 26Q представлены аминокислотные последовательности тяжелой цепи клона антитела 53.10. На фиг. 26R представлены аминокислотные последовательности легкой цепи клона антитела 53.10. На фиг. 26S представлены аминокислотные последовательности легкой цепи клона антитела 53.11. На фиг. 26T представлены аминокислотные последовательности легкой цепи клона антитела 53.12. На фиг. 26U представлены аминокислотные последовательности легкой цепи клона антитела 53.13.
На фиг. 27A представлены аминокислотные последовательности тяжелой и легкой цепей клона антитела 82.0. На фиг. 27B представлены аминокислотные последовательности тяжелой и легкой цепей клона антитела 82.1. На фиг. 27C представлены аминокислотные последовательности тяжелой и легкой цепей клона антитела 82.2. На фиг. 27D представлены аминокислотные последовательности тяжелой и легкой цепей клона антитела 82.3.
На фиг. 28A-P представлены последовательности нуклеиновой кислоты тяжелой цепи клонов антител 12.0-12.3 и легкой цепи клонов антител 12.0-12.3, тяжелой цепи клонов антител 135.0-135.3 и легкой цепи клонов антител 135.0-135.3, тяжелой цепи клонов антител 4.0-4.11 и легкой цепи клонов антител 4.0-4.7, тяжелой цепи клонов антител 53.0-53.6 и легкой цепи клонов антител 53.0-53.9, и тяжелой цепи клонов антитела 82.0-82.3 и легкой цепи клонов антител 82.0-82.3.
РЕПРЕЗЕНТАТИВНЫЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже подробно описаны некоторые репрезентативные варианты осуществления изобретения, некоторые примеры которых проиллюстрированы в прилагаемом графическом материале.
CXCR3
CXCR3 (MIM: 300574, человеческий GeneID: 2833, хемокиновый (мотив C-X-C) рецептор 3; также известный как CD182, CD183, CKR-L2, CKAR3, GPR9, IP10-R, Mig-R, MigR, рецептор 9, связанный с G-белком; рецептор IP-10, рецептор Mig, хемокиновый (C-X-C) рецептор 3, рецептор индуцируемого интерфероном белка 10) представляет собой хемокиновый рецептор, который в основном отсутствует в «необученных» T-клетках, но индуцируется после активации антигеном и осуществляет рекрутинг этих клеток в сайты воспаления ткани в ответ на первичные лиганды: CXCL9 (человеческий GeneID: 4283), CXCL10 (человеческий GeneID: 3627) и CXCL11 (человеческий GeneID: 6373). β-клетки островков Лангерганса экспрессируют CXCL9 и CXCL10 (Frigerio et al., Nature Medicine 8:1414-1420 (2002), а T-клетки, которые инфильтрируют поджелудочную железу, экспрессируют CXCR3 (Christen et al, The Journal of Immunology, 2003, 171: 6838-6845; Van Halteren et al., Diabetologia 48:75-82 (2005); Uno et al 2010; Roep et al., Clinical and Experimental Immunology, 2003, 159: 338-343; Tanaka et al., Diabetes 58: 2285-2291 (2009); Sarkar et al., Diabetes. 2012 Feb; 61(2):436-46).
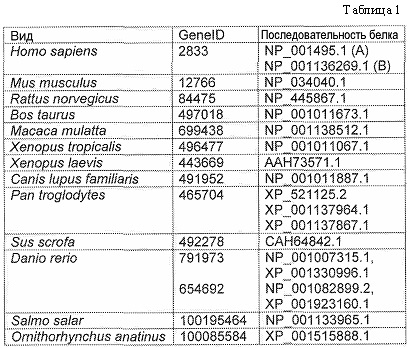
CXCR3 экспрессируется в различных организмах, включая, например, человека, мышей, крыс, коров, шимпанзе, макак, собак, лягушек, утконосов, свиней и рыбу-зебру. В таблице 1 приводится список последовательностей, имеющихся в банке GeneID Национального центра биотехнологической информации США (U.S. National Center for Biotechnology Information (NCBI)), и эталонная последовательность белка CXCR3 различных организмов. SEQ ID NO:1 представляет собой полноразмерную человеческую последовательность CXCR3 (вариант сплайсинга A). Пептидная последовательность варианта сплайсинга B представлена как эталонная последовательность NP_001136269.1. Предсказанные внеклеточные домены варианта сплайсинга А человеческого CXCR3 описаны в публикации Colvin et al., Mol. Cell. Bio., 26: 5838-49 (2006) и включают остатки 1-58, 1-16, 111-126, 190-223, 278-301 SEQ ID NO:1, указанные ниже.
SEQ ID NO:1 NP_001495 изоформы А человеческого CXCR3:
1 mvievsdhqv lndaevaall enfsssydyg enesdsccts ppcpqdfsin fdraflpaly
61 sllfllgllg ngavaavlls rrtalsstdt fllhlavadt llvltlpiwa vdaavqwvfg
121 sglckvagal fninfyagal llacisfdry lnivhatqly rrgpparvtl tclavwglcl
181 lfalpdfifl sahhderlna thcqynfpqv grtalrvlgl vagfllpllv maycyahila
241 vllvsrgqrr lramrlvvvv vvafalcwtp yhlwlvdil mdlgalarnc gresrvdvak
301 svtsglgymh cclnpllyaf vgvkfrermw mlllrlgcpn qrglqrqpss srrdsswset
361 seasysgl
CXCR3 и CXCL10 экспрессируются у человека, страдающего T1D. Uno et al., Endocrine J. 57: 991-996 (2010); Roep et al., Clin, and Exp. Immun. 159: 338-343 (2009); Tanaka et al., Diabetes 58: 2285-2291 (2009). У этих пациентов, CXCL10 экспрессируется в имеющихся инсулин-продуцирующих бета-клетках островков. CXCR3 экспрессируется в инвазивных Т-клетках, окружающих островки. Аналогичные паттерны экспрессии были репродуцированы у мышей с диабетом, не страдающих ожирением (NOD), и у мышей с моделями диабета. Morimoto et al., J. Immun., 173: 7017-7024 (2004); Li et al., World J. Gastroenterol, 11 (30): 4750-4752 (2005); Sarkar et al. Diabetes, 2012 Feb; 61 {2):436-46).
CXCR3 также экспрессируется в T-клетках, присутствующих в воспаленных тканях некоторых типов, а CXCL9, CXCL10 и CXCL11 часто продуцируются клетками, присутствующими на участках воспаления. В соответствии с этим, в некоторых вариантах осуществления изобретения описана терапия диабета T1D, проводимая посредством разрушения CXCR3.
Антитела
Используемый здесь термин «антитело» означает любой полипептид, содержащий антигенсвязывающий сайт, независимо от его источника, вида и происхождения, от метода его продуцирования и/или свойств, и охватывает иммуноглобулины или их антигенсвязывающие части или фрагменты. Термин «антитело» включает, но не ограничивается ими, антитела, происходящие от человека, орангутангов, мышей, крыс, коз, овец и кур. Этот термин включает, но не ограничивается ими, поликлональные антитела, моноклональные антитела, моноспецифические антитела, полиспецифические антитела, неспецифические антитела, гуманизированные антитела, полностью человеческие антитела, верблюжьи антитела, одноцепочечные антитела, химерные антитела, синтетические антитела, рекомбинантные антитела, гибридные антитела, мутированные антитела, антитела с обратной мутацией и CDR-привитые антитела. Термин «антитело», употребляемый в настоящем изобретении, также включает, если это не оговорено особо, фрагменты антител, такие как Fab, F(ab')2, Fv, scFv, Fd, dAb, VHH (также называемые наноантителами), и другие фрагменты антител, сохраняющие антигенсвязывающую функцию исходных антител, включая биспецифические или мультиспецифические антитела. Термин «антитело» также означает антигенсвязывающие молекулы, которые не происходят от иммуноглобулинов. Так, например, не-иммуноглобулиновыми каркасами, известными специалистам, являются небольшие модульные иммунофармацевтические средства (смотрите, например, публикации заявок на патент США №№20080181892 и 20080227958, опубликованные 31 июля 2008 года и 18 сентября 2008 года, соответственно), тетранектины, домены фибронектина (например, AdNectins, смотрите публикацию заявки на патент США №2007/0082365, опубликованную 12 апреля 2007 года), белок A, липокалины (смотрите, например, патент США № 7118915), анкириновые повторы и тиоредоксин.
Термин «антигенсвязывающий домен» означает часть молекулы антитела, которая включает область, специфически связывающуюся с частью антигена или с целым антигеном, или комплементарную этой части или целому антигену. В случае крупного антигена, антитело может связываться только с конкретной частью антигена. В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его антигенсвязывающий фрагмент содержат по меньшей мере один антигенсвязывающий домен. В некоторых вариантах осуществления изобретения, антитело или его фрагмент являются мультиспецифическими и содержат два или более (например, 2, 3, 4, 5 или более) антигенсвязывающих доменов, а поэтому такое антитело или его фрагмент обладают способностью связываться с двумя или более молекулами CXCR3 в одном и том же эпитопе или в различных эпитопах, или с CXCR3 и по меньшей мере с одним другим антигеном с высокой аффинностью. Антигенсвязывающая часть антитела может содержать один или более фрагментов антитела, которые сохраняют свою способность специфически связываться с антигеном. Эти фрагменты могут включать вариабельную область тяжелой и/или легкой цепи родительского антитела или варианта родительского антитела.
Термины «эпитоп» или «антигенная детерминанта» означают часть молекулы антигена, ответственную за специфическое взаимодействие с антигенсвязывающим доменом антитела. Антигенсвязывающий домен может содержать один или более вариабельных доменов антитела. Антигенсвязывающий домен может содержать по меньшей мере одну вариабельную область легкой цепи антитела (VL) и по меньшей мере одну вариабельную область тяжелой цепи антитела (VH). Антигенсвязывающий домен может также содержать только области VH или только области VL. Так, например, антитела, происходящие от верблюдов и лам (Camelidae, camelids), представляют собой уникальный тип антитела, которое образовано только тяжелыми цепями и не содержит легких цепей. Антигенсвязывающим сайтом таких антител является один единственный домен, обозначаемый VHH. Такие антитела называются «камелизированными антителами» или «наноантителами». Смотрите, например, патенты США №№5800988 и 6005079 и публикации международных заявок №№ WO 94/04678 и WO 94/25591, которые вводятся в настоящее описание посредством ссылки.
Описанные здесь анти-CXCR3 антитела могут быть продуцированы любым подходящим методом, известным специалистам. Так, например, такими антителами могут быть поликлональные антитела или моноклональные антитела. Методы получения поликлональных антител известны специалистам (Harlow et al., Antibodies; a Laboratory Manual, Cold Spring Harbor Laboratory Press, 2nd ed. (1988)). Для продуцирования анти-CXCR3 антител могут быть использованы иммуногены, содержащие полипептиды CXCR3, их фрагменты (например, один или более внеклеточных доменов или 58 N-концевых аминокислот, или 37 N-концевых аминокислот, или 20 N-концевых аминокислот, или 16 N-концевых аминокислот и т.п.), гибридные белки или их варианты.
Иммуноген может продуцироваться клеткой, которая продуцирует или сверхпродуцирует CXCR3, и такой клеткой может быть природная клетка, природная мутантная клетка или генетически сконструированная клетка. Иммуноген, в зависимости от природы полипептидов (например, процента гидрофобности, процента гидрофильности, стабильности, суммарного заряда, изоэлектрической точки и т.п.), может быть модифицирован или конъюгирован в целях изменения его иммуногенности. Так, например, CXCR3 или его часть могут быть конъюгированы с носителем. Такое конъюгирование может включать химическое конъюгирование посредством дериватизации активными химическими функциональными группами или конъюгирование методами получения гибридных белков или другими известными методами. Примерами носителей и/или других белков, влияющих на иммуногенность, являются, но не ограничиваются ими, KLH, овальбумин, сывороточный альбумин, бычий тироглобулин, соевый ингибитор трипсина и неспецифические T-хелперные пептиды.
Для усиления иммунологического ответа могут быть также использованы различные адъюванты в комбинации с иммуногеном CXCR3. Примерами адъювантов являются, но не ограничиваются ими, адъювант Фрейнда (полный и неполный), минеральные масла, гели, квасцы (гидроксид алюминия), поверхностно-активные вещества, такие как лизолецитин, полиолы плюроники, полианионы, пептиды, масляные эмульсии, гемоцианины лимфы улитки (KLH), динитрофенол и человеческие адъюванты, такие как БКГ (бацилла Кальметта-Герена) и Corynebacterium parvum. Дополнительными примерами адъювантов, которые могут быть использованы, являются адъювант MPL-TDM (монофосфориллипид A, синтетический дикориномиколят трегалозы). Протоколы иммунизации хорошо известны специалистам, и такая иммунизация может быть осуществлена любым методом, позволяющим вырабатывать иммунный ответ у выбранного животного-хозяина. Таким образом, могут быть применены различные способы введения в различные периоды времени в соответствии с выбранной схемой введения.
Так, например, описанный здесь иммуноген может быть введен различным животным-хозяевам, включая, но не ограничиваясь ими, кролики, мыши, верблюды, крысы и т.п., для индуцирования продуцирования сыворотки, содержащей поликлональные антитела, специфичные к CXCR3. Введение иммуногена может включать одну или более инъекций иммунизирующего агента и, необязательно, адъюванта. В некоторых вариантах осуществления изобретения, иммуноген (с адъювантом или без адъюванта) инъецируют млекопитающему путем введения множества подкожных или внутрибрюшинных инъекций, либо внутримышечных или внутривенных инъекций. В некоторых вариантах осуществления изобретения, после получения подходящего поликлонального препарата, а в частности, антитела, этот препарат может быть выделен известными методами разделения, такими как аффинная хроматография, пэннинг, абсорбция и т.п., в результате чего может быть получено антитело определенного вида. В некоторых вариантах осуществления изобретения, антитело определенного вида подвергают дополнительному исследованию, например, секвенированию в целях идентификации аминокислотных последовательностей одной или более CDR.
В некоторых вариантах осуществления изобретения, анти-CXCR3 антителами являются моноклональные антитела. Моноклональным антителом является любое антитело, происходящее от одного эукариотического клона, фага или прокариотического клона, экспрессирующего такое антитело. Моноклональные антитела могут быть получены, например, традиционными гибридомными методами (смотрите публикации Kohler and Milstein, Nature 256: 495-499 (1975) и патент США № 4376110, которые вводятся в настоящее описание посредством ссылки), методами рекомбинантных ДНК (смотрите патент США № 4816567, который вводится в настоящее описание посредством ссылки) или методами фагового представления, осуществляемыми с использованием библиотек антител (Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1991)). Различные другие методы продуцирования антител можно найти в публикации Antibodies: A Laboratory Manual, eds. Harlow et al., Cold Spring Harbor Laboratory, 1988. Другими примерами методов продуцирования моноклональных антител являются, но не ограничиваются ими, гибридомная технология на основе человеческих В-клеточных гибридом (Kosbor et al., Immunology Today 4:72 (1983) и Cole et al., Proc. Natl. Acad. Sci. USA 80:2028 (1983)) и EBV-гибридомная технология (Cole et al., Monoclonal Antibodies and Cancer Therapy, pp. 77-96, Alan R, Liss (1985)). Такими антителами могут быть иммуноглобулины любого класса, включая IgG, IgM, IgE, IgA и IgD, и любого подкласса, или их варианты. Гибридомы, продуцирующие mAb согласно изобретению, могут быть культивированы in vitro или in vivo.
В некоторых вариантах осуществления изобретения, иммуноген, содержащий полипептиды CXCR3, их фрагменты (например, один или более внеклеточных доменов или 58 N-концевых аминокислот, или 37 N-концевых аминокислот, или 20 N-концевых аминокислот, или 16 N-концевых аминокислот и т.п.), гибридные белки или их варианты могут быть использованы для иммунизации животного-хозяина (например, кроликов, мышей, верблюдов, крыс и т.п.) в целях получения гибридом, продуцирующих моноклональные антитела. Лимфоциты, которые продуцируют или обладают способностью продуцировать антитела, специфически связывающиеся с CXCR3, могут быть выделены у иммунизованного хозяина и подвергнуты слиянию с миеломными клетками с использованием подходящего агента для слияния, такого как полиэтиленгликоль, с получением гибридомных клеток (Coding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp. 59-103 (1986)).
Может быть получено множество гибридом, продуцирующих моноклональные антитела, после чего могут быть отобраны гибридомы, обладающие желательными свойствами, или, предположительно, терапевтическими свойствами, например, способностью предотвращать связывание лиганда CXCR3 с его рецептором. Выбранные антитела могут быть дополнительно модифицированы для сообщения или усиления желательных свойств, таких как повышенная стабильность in vivo. Так, например, после идентификации гибридомных клеток, продуцирующих антитела с нужной специфичностью, аффинностью и/или активностью, клоны могут быть подвергнуты субклонированию методами разведения и культивированы стандартными методами (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp. 59-103 (1986)). Подходящими культуральными средами являются, например, модифицированная по способу Дульбекко среда Игла (D-MEM) или среда RPMI-1640. Кроме того, гибридомные клетки могут быть культивированы in vivo в виде опухолей у животных. Субклоны могут быть проанализированы на специфичность, аффинность и/или активность, и субклоны, обладающие многими желательными свойствами, могут быть отобраны для последующей характеризации.
Существует ряд альтернативных методов продуцирования моноклональных антител, и любой из этих методов может быть применен для продуцирования описанных здесь анти-CXCR3 антител. Так, например, моноклональные антитела могут быть получены методами рекомбинантных ДНК, такими как методы, описанные в патенте США № 4816567, который во всей своей полноте вводится в настоящее описание посредством ссылки.
ДНК, кодирующая моноклональные антитела, может быть выделена и секвенирована с использованием стандартных процедур (например, с использованием олигонуклеотидных зондов, способных связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител или цепи человеческих, гуманизированных или других антител) (Innis et al., PCR Protocols, A Guide to Methods and Applications, Academic (1990), и Sanger et al., Proc. Natl. Acad. Sci. USA 74:5463 (1977)). Источником такой ДНК могут служить гибридомные клетки. Для синтеза моноклональных антител в рекомбинантных клетках-хозяевах, ДНК, после ее выделения, может быть введена в экспрессионные векторы, которые могут быть перенесены в клетки-хозяева, такие как клетки E. coli, клетки NSO, клетки COS, клетки яичника китайского хомячка (CHO) или миеломные клетки, которые, по своей природе, не продуцируют белок иммуноглобулин. ДНК может быть также модифицирована, например, путем замены гомологичных мышиных последовательностей, последовательностью, кодирующей константные домены тяжелой и легкой цепи человеческого антитела (патент США №4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851 (1984)) или путем ковалентного связывания со всей иммуноглобулин-кодирующей последовательностью или ее частью, происходящей от последовательности, кодирующей полипептид, не являющийся иммуноглобулином. В некоторых вариантах осуществления изобретения, полипептид, не являющийся иммуноглобулином, может быть заменен на константные домены антитела, либо на вариабельные домены одного CXCR3-связывающего сайта антитела с получением химерного двухвалентного антитела.
В некоторых вариантах осуществления изобретения, описанные здесь антитела могут быть модифицированы с получением CDR-привитых антител и/или антител, гуманизированных каким-либо другим способом. CDR-прививка представляет собой одну из форм гуманизации, но могут быть также применены и другие методы гуманизации, известные специалистам. Методы CDR-прививки известны специалистам, и такие CDR-привитые последовательности могут быть пронумерованы в соответствии со схемой нумерации CDR, включая схему нумерации по системе IMGT (Международной информационной системе ImMunoGeneTics®, Montpellier, France), нумерации по Кэбату, нумерации по Чотию и нумерации по Чотию с модификациями. Смотрите, например, imgt.org (где приводится краткое описание применения системы сплошной нумерации IMGT, в которой определены и объединены данные о каркасных областях и гипервариабельных областях, структурные данные, полученные в рентгеноструктурных анализах, и данные характеризации гипервариабельных петель, в целях создания уникальной системы нумерации для всех V-областей IG и TR всех видов); Abhinandan and Martin, Mol. Immunol., 45:3832-9 (2008); смотрите, также Abhinandan and Martin, J. Mol. Biol., 369(3):852-62 (2007) (описание методов оценки «степени гуманизации» химерного антитела); Retter et al., Nucleic Acids Res. 33 (Database issue):D671-4 (2005) (описание базы данных VBASE2 генов вариабельных доменов); и Johnson and Wu, Int. Immunol., 10(12): 1801-5 (1998) (описание распределения длин CDRH3).
Так, например, в соответствии с системой нумерации IMGT, консервативные аминокислоты всегда имеют одно и то же положение. Гидрофобные аминокислоты каркасных областей также пронумерованы в консервативных положениях, что позволяет сравнивать каркасные аминокислоты (и кодоны), находящиеся в одних и те же положениях в различных последовательностях, без выравнивания последовательностей. В другом примере, в системе нумерации по Кэбату, CDR-H1 начинается приблизительно с аминокислоты 31 {то есть приблизительно с 9-го по счету остатка после первого цистеинового остатка), включает приблизительно 5-7 аминокислот и заканчивается в положении следующего тирозинового остатка. CDR-H2 начинается с пятнадцатого остатка от конца CDR-HI, включает приблизительно 16-19 аминокислот и заканчивается в положении следующего аргининового или лизинового остатка. CDR-H3 начинается приблизительно с тридцать третьего аминокислотного остатка от конца CDR-H2; включает 3-25 остатков и заканчивается у последовательности W-G-X-G, где X означает любую аминокислоту. CDR-L1 начинается приблизительно с остатка 24 (то есть после цистеинового остатка); включает приблизительно 10-17 остатков и заканчивается в положении следующего тирозинового остатка. CDR-L2 начинается приблизительно с шестнадцатого остатка от конца CDR-L1 и включает приблизительно 7 остатков. CDR-L3 начинается приблизительно с тридцать третьего остатка от конца CDR-L2; включает приблизительно 7-11 остатков и заканчивается у последовательности F-G-X-G, где X означает любую аминокислоту. Антитела, содержащие по меньшей мере одну из этих CDR, могут быть использованы в способах согласно изобретению.
CDR-привитые антитела могут содержать последовательности вариабельной области тяжелой и легкой цепей, происходящие от человеческого антитела, где одна или более CDR-областей VH и/или VL заменены CDR-последовательностями донорных антител, например, описанных ниже мышиных антител, которые связываются с CXCR3. Каркасная последовательность, происходящая от любого человеческого антитела, может служить в качестве матрицы для CDR-прививки. Однако прямая замена CDR-цепи такой каркасной последовательностью может приводить к некоторой потере аффинности связывания с антигеном. Чем выше гомология человеческого антитела с исходным антителом, например, мышиным антителом, тем меньше вероятность того, что объединение донорных CDR с человеческой каркасной областью будет вносить нарушения в CDR, которые могут снижать аффинность. Поэтому, в некоторых вариантах осуществления изобретения, CDR-привитые анти-CXCR3 антитела согласно изобретению включают человеческую вариабельную каркасную область, которая имеет последовательность, по меньшей мере на 65% идентичную каркасной последовательности вариабельной области донорного мышиного CXCR3-нейтрализующего антитела. Методы продуцирования таких антител известны специалистам (смотрите EP 239400; публикацию заявки PCT № WO 91/09967; и патенты США №№5225539; 5530101 и 5585089) и включают «маскировку» или реконструирование (EP 592106; EP 519596; Padlan (1991) Mol. Immunol. 28(4/5): 489-498; Studnicka et al. (1994) Prot. Engineer. 7(6): 805-814; и Roguska et al. (1994) Proc. Acad. Sci. USA 91: 969-973), перестановку цепей (патент США № 5585352) и применение антиидиотипических антител.
В некоторых вариантах осуществления изобретения, описанные здесь антитела могут быть гуманизированными. Термин «гуманизированные антитела» означает молекулы антител, которые связываются с нужным антигеном, имеют одну или более CDR, происходящих от не-человеческих антител, и имеют каркасные области и/или константные домены человеческой молекулы иммуноглобулина. Известные человеческие последовательности Ig описаны, например, на сайтах: www.ncbi.nlm.nih.gov/entrez- /query.fcgi; www.atcc.org/phage/hdb.html; www.sciquest.com/; www.abcam.com/; www.antibodyresource.com/onlinecomp.html; и в публикации Kabat et al., Sequences of Proteins of Immunological Interest, U.S. Dept. Health (1983). «Импортные» человеческие последовательности могут быть использованы для снижения иммуногенности, либо для ослабления, усиления или модификации связывания, аффинности, скорости ассоциации, скорости диссоциации, авидности, специфичности, времени полужизни или любых других подходящих свойств, известных специалистам. Антитела могут быть гуманизированы различными известными методами, такими как, но не ограничивающимися ими, методы, описанные в публикациях Jones et al. (1986) Nature 321:522; Verhoeyen et al. (1988) Science 239:1534; Sims et al. (1993) J. Immunol. 151: 2296; Chothia and Lesk (1987) J. Mol. Biol. 196: 901; Carter et al. (1992) Proc. Natl. Acad. Sci. USA 89: 4285; Presta et al. (1993) J. Immunol. 151:2623; в патентах США №№ 5589205; 565332; 6180370; 6632927; 7241877; 7244615; 7244832; 72620505; и в публикации заявки на патент США № 2004/0236078 (поданной 30 апреля, 2004), которые во всей своей полноте вводятся в настоящее описание посредством ссылки.
В некоторых вариантах осуществления изобретения, каркасные остатки в гуманизированных или CDR-привитых антителах могут быть заменены соответствующим остатком CDR донорного антитела, например, каркасными остатками антимышиного CXCR3-нейтрализующего антитела для изменения, например, для повышения уровня связывания с антигеном. Смотрите Queen et al., Proc. Natl. Acad. Sci. USA 86:10029-33 (December 1989). Такие замены в каркасных областях идентифицируют хорошо известными методами, например, путем моделирования взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, играющих важную роль в связывании с антигеном, и путем сравнения последовательностей для идентификации редко встречающихся каркасных остатков в конкретных положениях. Смотрите, например, патент США № 5585089; и публикацию Riechmann et al. (1988) Nature 332:323, которые во всей своей полноте вводятся в настоящее описание посредством ссылки. Трехмерные модели иммуноглобулина являются общедоступными и известны специалистам. Существуют компьютерные программы, которые иллюстрируют и представляют возможные трехмерные конформационные структуры выбранных последовательностей-кандидатов иммуноглобулина. Оценка таких представлений позволяет проанализировать вероятную роль этих остатков в функционировании последовательности-кандидата иммуноглобулина, то есть проанализировать остатки, влияющие на способность иммуноглобулина-кандидата связываться с антигеном. Таким образом, каркасные остатки могут быть отобраны и объединены с консенсусными и «импортными» последовательностями для сообщения антителу нужных свойств, таких как повышенная аффинность по отношению к CXCR3.
Антитела могут быть гуманизированы или CDR-привиты, и каркасные остатки, происходящие от CDR-доноров, которые могут быть использованы для повышения уровня связывания с антигеном, могут быть идентифицированы различными известными методами, такими как, но не ограничивающимися ими, методы, описанные в публикациях Jones et al. (1986) Nature 321: 522; Verhoeyen et al. (1988) Science 239: 1534; Sims et al. (1993) J. Immunol. 151: 2298; Chothia and Lesk (1987) J. Mol. Biol. 198: 901; Carter et al. (1992) Proc. Natl. Acad. Sci. USA 89: 4285; Presta et al. (1993) J. Immunol. 151: 2823; и в патентах США №№ 5585332; 5723323; 5976882; 5824514; 5817483; 5814476; 5783192; 5723323; 5788886; 5714352; 8204023; 6180370; 5693762; 5530101; 5585089; 5225539 и 4816587. В некоторых вариантах осуществления изобретения, 4D-гуманизацию осуществляют для получения вариантов антител согласно изобретению (например, для получения 4D-гуманизированных вариантов клона 4, содержащих любую из тяжелых цепей 4.4-4.6 и любую из легких цепей 4.4-4.7). Смотрите заявку WO 2009/032661 (которая во всей своей полноте вводится в настоящее описание посредством ссылки), например, параграфы [0037]-[0044], где описаны методы 4D-гуманизации. Вкратце, 4D-гуманизация может включать: a) построение 3-D-модели вариабельного домена, который должен быть гуманизирован; b) идентификацию гибких остатков в вариабельном домене посредством имитации молекулярной динамики 3-D модели указанного домена; c) идентификацию наиболее близкородственной человеческой зародышевой линии путем сравнения молекулярно-динамического пути 3-D-модели с молекулярно-динамическими путями 49 человеческих зародышевых линий; и d) введение мутации гибких остатков, которые не являются частью CDR, в их аналог человеческой зародышевой линии (идентифицированной в стадии c).
В некоторых вариантах осуществления изобретения, CDR-привитые и/или как-либо иначе гуманизированные антитела могут содержать CDR-привитые и/или гуманизированные варианты клонов 4, 12, 53, 82 и 135. Так, например, соответствующие области тяжелой и легкой цепей, происходящие от любого из клонов 4, 12, 53, 82 и 135 (например, клон 4 тяжелой цепи и клон 4 легкой цепи), могут быть присоединены к человеческим константным доменам с получением химерных антител. Химерные антитела могут быть дополнительно гуманизированы путем замены одного или более каркасных остатков или аминокислот CDR на соответствующий человеческий остаток. Аналогичным образом, в некоторых вариантах осуществления изобретения, шесть областей CDR тяжелой и легкой цепей любого из клонов 4, 12, 53, 82 и 135 (например, CDR1, CDR2 и CDR3 тяжелой цепи клона 4, и CDR1, CDR2 и CDR3 легкой цепи клона 4) или любого из вариантов клонов 4, 12, 53, 82 и 135 могут быть субклонированы в человеческие каркасные и/или константные домены с получением гуманизированных антител. Гуманизация может быть проведена с использованием человеческих вариабельных доменов, за исключением аминокислот CDR и/или любых остатков в положении Верньера. Гуманизированные антитела могут также включать обратные мутации в остатках, расположенных в четырех аминокислотах CDR и/или в положениях, идентифицированных как остатки, которые в исходной последовательности антитела и в человеческих последовательностях «очень сильно отличаются» друг от друга, например, в случае применения IMGT-моделирования. Для повышения стабильности или терапевтической эффективности антитела, в каркасную область или в CDR-области могут быть введены дополнительные мутации, например, путем внесения мутаций для удаления сайта дезамидирования в положениях 58 и 59 (IMGT-нумерация) CDR2 VH клона 4.
Так, например, описанные здесь антитела, химерные антитела и гуманизированные антитела могут содержать шесть CDR и/или вариабельные домены тяжелой и легкой цепей, происходящие от любого из клонов 4, 12, 53, 82 и 135 и их химерных или гуманизированных вариантов. Так, например, антитело или его фрагмент, способные связываться с CXCR3, могут содержать три CDR, происходящих от любой из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3 тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3. Аналогичнным образом, антитело или его фрагмент могут содержать три CDR, происходящих от любой из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3. В некоторых вариантах осуществления изобретения, CDR тяжелой и легкой цепей происходят от одного и того же клона, но могут происходить и от различных вариантов этого клона (например, три CDR тяжелой цепи 4.1, присоединенные к трем CDR легкой цепи 4.2). Тяжелая и легкая цепи 4.0, 12.0, 82.0 и 135.0 представляют собой вариабельный домен в клонах мышиного антитела и в химерных антителах (где указанные антитела содержат мышиные вариабельные домены и человеческие каркасные области). Остальные тяжелая и легкая цепи представляют собой гуманизированные цепи, указанные в таблице 11.
В некоторых вариантах осуществления изобретения, антитело или его фрагмент, способные связываться с CXCR3, могут содержать любую из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3, тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3. Аналогичным образом, антитело или его фрагмент могут содержать любую из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3.
В некоторых вариантах осуществления изобретения, тяжелую и легкую цепи выбирают так, чтобы три CDR тяжелой цепи конкретного клона (например, CDR тяжелой цепи клона 4) были присоединены к трем CDR любой из легких цепей этого клона (например, CDR легкой цепи клона 4). В некоторых вариантах осуществления изобретения, тяжелую и легкую цепи выбирают так, чтобы тяжелая цепь конкретного клона (например, тяжелая цепь клона 4) была присоединена к любой из легких цепей этого клона (например, легкой цепи клона 4).
В некоторых вариантах осуществления изобретения, три CDR любого из вариабельных доменов тяжелой цепи 4.0-4.11 могут быть присоединены к трем CDR любого из вариабельных доменов легкой цепи 4.0-4.7; три CDR любого из вариабельных доменов тяжелой цепи 12.0-12.3 могут быть присоединены к трем CDR любого из вариабельных доменов легкой цепи 12.0-12.3; три CDR любого из вариабельных доменов тяжелой цепи 53.0-53.0 могут быть присоединены к трем CDR любого из вариабельных доменов легкой цепи 53.0-53.13; три CDR любого из вариабельных доменов тяжелой цепи 82.0-82.3 могут быть присоединены к трем CDR любого из вариабельных доменов легкой цепи 82.0-82.3; или три CDR любого из вариабельных доменов тяжелой цепи 135.0-135.3 могут быть присоединены к трем CDR любого из вариабельных доменов легкой цепи 135.0-135.3.
В некоторых вариантах осуществления изобретения, любой из вариабельных доменов тяжелой цепи 4.0-4.11 может быть присоединен к любому из вариабельных доменов легкой цепи 4.0-4.7; любой из вариабельных доменов тяжелой цепи 12.0-12.3 может быть присоединен к любому из вариабельных доменов легкой цепи 12.0-12.3; любой из вариабельных доменов тяжелой цепи 53.0-53.10 может быть присоединен к любому из вариабельных доменов легкой цепи 53.0-53.13; любой из вариабельных доменов тяжелой цепи 82.0-82.3 может быть присоединен к любому из вариабельных доменов легкой цепи 82.0-82.3 или любой из вариабельных доменов тяжелой цепи 135.0-135.3 может быть присоединен к любому из вариабельных доменов легкой цепи 135.0-135.3.
Выравнивание некоторых вариабельных доменов тяжелой и легкой цепей проиллюстрировано на фиг. 17. В некоторых вариантах осуществления изобретения, описанное здесь антитело может содержать спаренные вариабельные домены тяжелой и легкой цепей, как показано в таблице 2 (Ch = химерное антитело, Hu = гуманизированное антитело, VH = тяжелая цепь, VK = легкая цепь). Так, например, первое антитело таблицы 2 означает вариант клона 4, содержащий тяжелую цепь 4.0 и легкую цепь 4.0. Второе антитело означает вариант клона 4, содержащий тяжелую цепь 4.1 и легкую цепь 4.1. Каждому антителу, содержащему указанные последовательности тяжелой цепи и легкой цепи, было также присвоено обозначение, указанное во втором столбце таблицы 2. Так, например, первому антителу в данной таблице (содержащему тяжелую цепь 4.0 и легкую цепь 4.0) присвоено обозначение 4Ch, а второму антителу в этой таблице (содержащему тяжелую цепь 4.1 и легкую цепь 4.1) присвоено обозначение 4Hu1.
Термины «специфическое взаимодействие» или «специфически связывается» или т.п. означают, что две молекулы образуют комплекс, который является относительно стабильным в физиологических условиях. Специфическое связывание характеризуется высокой аффинностью и низкой или средней антигенсвязывающей способностью. Неспецифическое связывание обычно характеризуется низкой аффинностью и средней или высокой антигенсвязывающей способностью. Обычно, связывание считается специфическим, если константа аффинности Ka превышает 106 M-1 или предпочтительно превышает 108 M-1. В некоторых вариантах осуществления изобретения, антитела, их варианты и фрагменты связываются с антигеном(ами) с константами ассоциации, составляющими по меньшей мере 106, 107, 108, 109 M-1 или выше. В некоторых вариантах осуществления изобретения, антитела, их варианты и фрагменты связываются с CXCR3 с кинетикой связывания, указанной в таблицах 7A-B, 8-10 и/или 12. При необходимости, уровень неспецифического связывания может быть снижен, например, путем изменения условий связывания, так, чтобы это, по существу, не влияло на специфическое связывание. Такие условия известны специалистам и могут быть выбраны рутинными методами. Эти условия обычно определяют как концентрацию антител, ионную силу раствора, температуру, время, необходимое для связывания, и концентрацию блокирующих молекул, таких как сывороточный альбумин и молочный казеин.
В некоторых вариантах осуществления изобретения, описанные здесь анти-CXCR3 антитела могут нейтрализовать CXCR3. «CXCR3-нейтрализующее антитело» связывается с CXCR3 и блокирует активность рецептора, такую как типичные физиологические и генетические ответы, индуцируемые связыванием лигандов CXCR3 с CXCR3. Нейтрализующая активность может быть полной (100% нейтрализация) или частичной, например, нейтрализация, составляющая приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 95 процентов (или любое промежуточное значение), или более, и такая активность зависит от различных факторов, известных специалистам, таких как концентрация антитела, аффинность и эпитоп, а также от конкретного анализа, проводимого для оценки нейтрализующей активности. Нейтрализующая активность CXCR3-нейтрализующего антитела может быть определена с помощью анализов, проводимых путем измерения ингибирования, например, связывания с лигандом, связывания с GTP, мобилизации кальция, хемотаксиса клеток и/или интернализации рецептора. Специалистам известно множество анализов для определения активности нейтрализующих антител, а в частности, CXCR3-нейтрализующего антитела, и такие анализы могут быть легко адаптированы для подтверждения нейтрализующей активности конкретного антитела.
Так, например, в некоторых вариантах осуществления изобретения, нейтрализующая активность антитела, используемого в способах согласно изобретению, может быть оценена в анализе на хемотаксис, в основном, как описано во вкладыше, вложенном в упаковку с антителом, продуцируемым клоном 49801, которое поставляется R&D Systems® (номер по каталогу AB160). Нейтрализующую дозу-50 (ND50) определяют как концентрацию антитела, необходимую для достижения полумаксимального ингибирования CXCR3-опосредуемого rhl-TAC-ответа на клеточной поверхности отвечающей клеточной линии, в конкретной концентрации rhl-TAC. Для оценки способности антитела блокировать rhl-TAC-индуцированный хемотаксис hCXCR3-трансфицированных клеток BaF/3, rhl-TAC в концентрации 7 нг/мл добавляют в нижнее отделение 96-луночной камеры для хемотаксиса (NeuroProbe, Cabin John, MD). Затем, к камере для хемотаксиса подсоединяют поликарбонатный фильтр без PVP (размер пор 5 микрон). В верхние лунки камеры добавляют серийные разведения антитела (например, от 0,001 до 10000 мкг/мл) и 0,25×106 клеток/лунку. После инкубирования в течение 3 часов при 37°C в инкубаторе с относительной влажностью в атмосфере 5% CO2, камеру демонтируют, и клетки, которые мигрируют в нижнюю камеру, переносят на рабочую пластину и количественно оценивают, например, по уровню флуоресценции резазурина.
В работе Colvin et al., Mol. Cell. Bio., 26: 5838-49 (2006) описаны дополнительные анализы, которые, в некоторых вариантах осуществления изобретения, могут быть проведены для определения нейтрализующей активности CXCR3-нейтрализующих антител, используемых в настоящем изобретении. Вкратце, могут быть использованы клетки 300-19, то есть клеточная линия мышиного пре-B-клеточного лейкоза, которая функционально экспрессирует CXCR4. После трансфекции, эта линия может функционально экспрессировать и другие хемокиновые рецепторы, например, человеческий CXCR3 (смотрите, например, параграфы 201-209 публикации заявки на патент США № 2010/0061983, которые вводятся в настоящее описание посредством ссылки). Клетки 300-19, экспрессирующие человеческий CXCR3, могут быть культивированы в полной среде RPMI, содержащей 10% фетальную бычью сыворотку (FBS). Для оценки связывания лигандов CXCR3 с CXCR3 в присутствии CXCR3-нейтрализующих антител-кандидатов, 400000 клеток CXCR3/300-19 помещают в 96-луночные планшеты для культивирования тканей в общем объеме 150 мкл буфера для связывания (0,5% BSA, 5 мМ MgCl2, 1 мМ CaCl2, 50 мМ HEPES, pH 7,4). В клетки может быть добавлено всего 0,04 нМ 125I-меченного CXCL10 (New England Nuclear, Boston, MA) или CXCL11 (Amersham Biosciences, Piscataway, NJ) и 5×106 нМ - 500 нМ немеченного CXCL10 или CXCL11 (Peprotech, Rocky Hill, NJ), и полученную смесь инкубируют в течение 90 минут при комнатной температуре со встряхиванием. Затем клетки переносят в 96-луночные фильтры-планшеты (Millipore, Billerica, MA), которые были предварительно пропитаны 0,3%-ным полиэтиленимином, и три раза промывают 200 мкл буфера для связывания, в который было добавлено 0,5 M NaCl. После этого, планшеты сушат и измеряют радиоактивность после добавления сцинтиляционной жидкости в сцинтиляционный счетчик WaNac Microbeta (Perkin-Elmer Life Sciences, Boston, MA). Связывание CXCL9 может быть оценено методом, аналогичным методу оценки связывания CXCL10 и 11.
В некоторых вариантах осуществления изобретения, описанные здесь антитела могут блокировать или уменьшать поток кальция в CXCRS-экспрессирующие клетки. В некоторых вариантах осуществления изобретения, поток кальция может быть детектирован в клетках, таких как клетки CXCR3/300-19. Приблизительно 5×106 клеток суспендируют в 2 мл среды RPMI с 1% BSA. После этого добавляют 15 микрограммов Fura-2 (Molecular Probes, Eugene, OR), и клетки инкубируют при 37°C в течение 20 минут. Эти клетки два раза промывают в PBS и ресуспендируют в 2 мл буфера для потока кальция (145 мМ NaCl, 4 мМ KCl, 1 мМ NaHPO4, 1,8 мМ CaCl2, 25 мМ HEPES, 0,8 мМ NaCl2 и 22 мМ глюкозы). Интенсивность флуоресценции измеряли при 37°C на флуориметре DeitaRAM (Photon Technology International, Lawrenceville, NJ). До и после добавления хемокинов (например, CXCL9, 10 или 11), концентрации внутриклеточного кальция регистрируют как интенсивность возбуждения флуоресценции, излучаемой на 510 нм в ответ на последовательное возбуждение на длинах волн 340 нм и 380 нм, и представляют как отношение флуоресценции на 340 нм к флуоресценции на 380 нм.
В некоторых вариантах осуществления изобретения, нейтрализация CXCR3 может быть оценена путем измерения снижения степени интернализации рецептора. В некоторых вариантах осуществления изобретения, анализы на интернализацию рецептора могут быть осуществлены путем инкубирования приблизительно 2,5×106 клеток, таких как клетки CXCR3/300-19, в среде RPMI с 1% BSA с различными концентрациями CXCL10, CXCL11 или CXCL9, в течение 30 минут при 37°C. Затем эти клетки могут быть промыты охлажденным на льду буфером в клеточном сортере с активацией флуоресценции, после чего они могут быть проанализированы на поверхностную экспрессию CXCR3 с использованием ФЭ-конъюгированного анти-CXCR3 антитела.
Дополнительные анализы для оценки нейтрализующей активности описаны, например, в примерах 2-4 патента США № 7405275, которые вводятся в настоящее описание посредством ссылки.
Как было оценено в любом из вышеупомянутых анализов, в некоторых вариантах осуществления изобретения, CXCR3-нейтрализующее антитело может иметь концентрацию ND50, составляющую приблизительно 0,01, 0,02, 0,05, 0,1, 0,2, 0,5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 40, 50 или 100 мкг/мл. В конкретных вариантах осуществления изобретения, ND50 может составлять 0,5-12 мкг/мл, а более конкретно, 1-6 мкг/мл.
Описанными здесь выделенными анти-CXCR3 антителами могут быть антитела, которые специфически связываются с эпитопами CXCR3. Так, например, антитела, используемые в настоящем изобретении, могут связываться с пептидом, содержащим полноразмерную последовательность или часть последовательности (например, фрагмент, состоящий по меньшей мере из 5, 6, 8, 10, 12, 14, 15, 16, 18 или 20 остатков), выбранной из остатков 1-58, 1-16 или 1-37 SEQ ID NO:1. В некоторых вариантах осуществления изобретения описанными здесь антителами являются антитела, которые связываются с одним или более эпитопами, идентифицированными как показано на фиг.18. В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело может включать антитело, которое связывается с эпитопом CXCR3, содержащим последовательность SDHQVLNDAE (SEQ ID NO:71). В некоторых вариантах осуществления изобретения, эпитоп содержит последовательность SDHQVLND (SEQ ID NO:72), DHQVLND (SEQ ID NO:73) и/или VLNDAE (SEQ ID NO:74). В некоторых вариантах осуществления изобретения, эпитоп содержит последовательность VLND (SEQ ID NO:75). В некоторых вариантах осуществления изобретения эпитоп содержит последовательность XDXXVXNDXX (SEQ ID NO:76), где X означает любую аминокислоту. В некоторых вариантах осуществления изобретения эпитоп содержит последовательности XDXXVXND (SEQ ID NO:77), DXXVXND (SEQ ID NO:78) и/или VXNDXX (SEQ ID NO:79), где X означает любую аминокислоту. В некоторых вариантах осуществления изобретения эпитоп содержит последовательность VXND (SEQ ID NO:80), где X означает любую аминокислоту.
Анти-CXCR3 антитела могут быть специфичными для всех последовательностей CXCR3, происходящих от различных видов, или селективными для последовательностей CXCR3, происходящих от конкретных видов, или для последовательности CXCR3 конкретного изотипа. В конкретных вариантах осуществления изобретения, анти-CXCR3 антитело является специфичным для индивидуума конкретного вида, которому вводят такое антитело. В соответствии с этим, в некоторых вариантах осуществления изобретения, анти-CXCR3 антитело может быть специфичным для человеческой последовательности CXCR3 (например, способным связываться с пептидом, содержащим последовательность, гомологичную любой из подпоследовательностей SEQ ID NO:1, перечисленных выше). Гомологичная последовательность может быть легко идентифицирована специалистом в данной области с применением таких средств, как выравнивание белковых последовательностей (например, программы BLASTp, ClustalW и т.п.). В конкретных вариантах осуществления изобретения антитело, используемое в настоящем изобретении, связывается с пептидом, содержащим последовательность, которая по меньшей мере на 90%, 95% или 99% (или на любую промежуточную величину) аналогична или идентична последовательности SEQ ID NO:1 по всей длине данной последовательности или на протяжении соответствующего «окна», составляющего по меньшей мере 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65 или 70 остатков (или любое промежуточное количество остатков). В некоторых вариантах осуществления изобретения, указанное антитело способно связываться с эпитопом, последовательность которого по меньшей мере на 80%, 85%, 90%, 95% или 99% (или на любую промежуточную величину) аналогична или идентична последовательности одного из эпитопов, описанных выше (смотрите также фиг. 18).
Описанными здесь конкретными антителами являются, например, клоны антител (Cl) 12, Cl 135, Cl 82, Cl 53 и Cl 4, а также их химерные и гуманизированные варианты.
В некоторых вариантах осуществления изобретения описанные здесь антитела обладают некоторыми улучшенными свойствами по сравнению с известными антителами, включая антитела 5H7 и 7H5 (описанные, например, в патенте США № 7405275; CDR этих антител указаны в таблицах 1 и 2 и в списке известных последовательностей, которые вводятся в настоящее описание посредством ссылки); антитело V44D7 (описанное в публикации Международной заявки WO 2008/094942), антитело 1C6 (описанное в патенте США № 7407855; где картирование эпитопов описано в примерах 8 и 9, которые вводятся в настоящее описание посредством ссылки) и антитело 49801, поставляемое R&D Systems под каталожным номером MAB160.
В некоторых вариантах осуществления изобретения описанные здесь клоны антител (клоны 4, 12, 53, 82 и 135 и их химерные и гуманизированные аналоги) обладают некоторыми неожиданно обнаруженными предпочтительными свойствами по сравнению с известными антителами 5H7, 7H5, V44D7, 1C6 и 49801. Так, например, описанные здесь клоны обладают повышенной аффинностью связывания по сравнению с клонами анти-hCXCR3 антител 5H7, 7H5, V44D7, 1C6 и 49801. Описанные здесь гуманизированные клоны могут также обладать пониженной иммуногенностью по сравнению с клонами мышиных анти-hCXCR3 антител 5H7, 7H5, V44D7, 1C6 и 49801. Кроме того, описанные здесь антитела, такие как антитела, содержащие клоны тяжелой цепи 4.7-4.11, были оптимизированы путем модификации в положениях 58 и 59 (в соответствии с нумерацией по IMGT) в целях удаления сайта дезамидирования для повышения стабильности. В некоторых вариантах осуществления изобретения описанные здесь антитела сохраняют CXCR3-нейтрализующую активность.
В некоторых вариантах осуществления изобретения описанные здесь антитела или их фрагменты могут содержать последовательности CDR VH и/или VL, которые приблизительно на 80-100% (например, приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%) идентичны последовательностям CDR VH и/или VL любого из антител Cl 12, Cl 135, Cl 82, Cl 53 и Cl 4 или химерных или гуманизированных вариантов этих клонов (например, на 80-100% идентичны шести CDR в Cl 12 или шести CDR в Cl 12.1, и т.п.). В некоторых вариантах осуществления изобретения, антитела или их фрагменты могут содержать последовательности CDR VH и VL, которые имеют 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных замен (включая добавления, делеции и замены, такие как консервативные замены) по сравнению с последовательностями CDR VH и/или VL любого из антител Cl 12, Cl 135, Cl 82, Cl 53 и Cl 4.
Используемые здесь термины «процент (%) идентичности последовательностей» или «гомология» определены как процент аминокислотных остатков или нуклеотидов в последовательности-кандидате, которые идентичны аминокислотным остаткам или нуклеотидам в сравниваемых последовательностях после выравнивания этих последовательностей и введения пробелов, если это необходимо для достижения максимального процента идентичности последовательностей, за исключением консервативных замен нуклеиновых кислот. Оптимальное выравнивание этих последовательностей для сравнения может быть осуществлено вручную, а также с помощью алгоритмов для определения локальной гомологии, известных специалистам, или с помощью компьютерных программ, в которых используются такие алгоритмы (например, BLAST P).
В некоторых вариантах осуществления изобретения описанное здесь выделенное анти-CXCR3 антитело или его антигенсвязывающий фрагмент включают аминокислотную последовательность VH, которая по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% (или на любую промежуточную величину) идентична аминокислотной последовательности любой из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3, тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3. В некоторых вариантах осуществления изобретения антитело или его фрагмент включает аминокислотную последовательность VH, имеющую 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мутаций (включая добавления, делеции и замены, такие как консервативные замены) в аминокислотной последовательности SEQ любой из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3, тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3. Используемый здесь термин «консервативная замена» означает замену одной аминокислоты другой аминокислотой так, чтобы это не приводило к значительному изменению химических, физических и/или функциональных свойства антитела или его фрагмента (например, антитело или его фрагмент должны сохранять те же самые свойства, такие как заряд, структура, полярность, гидрофобность/гидрофильность, и/или те же самые функции, такие как способность распознавать CXCR3, связываться с CXCR3 и/или нейтрализовать его активность).
В некоторых вариантах осуществления изобретения описанное здесь выделенное анти-CXCR3 антитело или его антигенсвязывающий фрагмент включают аминокислотную последовательность VL, которая по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% (или на любую промежуточную величину) идентична аминокислотной последовательности любой из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3. В различных вариантах осуществления изобретения антитело или его фрагмент включает аминокислотную последовательность VL, имеющую 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мутаций (включая добавления, делеции и замены, такие как консервативные замены) в аминокислотной последовательности SEQ любой из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3.
В некоторых вариантах осуществления изобретения описанное здесь выделенное анти-CXCR3 антитело или его антигенсвязывающий фрагмент включают вариабельный домен тяжелой цепи, содержащий последовательность, которая по меньшей мере на 80% идентична аминокислотной последовательности любой из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3, тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3, и вариабельный домен легкой цепи, содержащий последовательность, которая по меньшей мере на 80% идентична аминокислотной последовательности любой из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3. В некоторых вариантах осуществления изобретения тяжелую и легкую цепи отбирают так, чтобы тяжелая цепь конкретного клона (например, тяжелая цепь клона 4) была присоединена к любой из легких цепей этого клона (например, легкой цепи клона 4). В некоторых вариантах осуществления изобретения, тяжелая и легкая цепи были присоединены, как показано в таблице 2. В других вариантах осуществления изобретения, антитело или его фрагмент, содержащие описанные здесь последовательности VH и/или VL, сохраняют способность нейтрализовать активность CXCR3.
В некоторых вариантах осуществления изобретения описанное здесь антитело представляет собой гуманизированный вариант Cl 12, 135, 82, 53 и/или 4. В других вариантах осуществления изобретения указанное антитело является полностью человеческим. В некоторых вариантах осуществления изобретения указанное антитело представляет собой гуманизированное или полностью человеческое производное антитела, выбранного из клонов 12, 135, 82, 53 и 4. В некоторых вариантах осуществления изобретения указанное антитело имеет константу аффинности по меньшей мере 108 M-1 (например, по меньшей мере 108 M-1, по меньшей мере 109 M-1, по меньшей мере 1010 M-1 или по меньшей мере 1011 M-1, или любую промежуточную величину). В некоторых вариантах осуществления изобретения указанное антитело способно связываться со всеми изоформами CXCR3. В некоторых вариантах осуществления изобретения указанное антитело способно связываться с А- и В-изоформами CXCR3. В некоторых вариантах осуществления изобретения указанное антитело не связывается с B-изоформой CXCR3.
В некоторых вариантах осуществления изобретения выделенное анти-CXCR3 антитело или его антигенсвязывающий фрагмент включают VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и/или VL CDR3, содержащие аминокислотные последовательности, которые приблизительно на 90-100% (например, приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичны последовательностям CDR VH и VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения выделенное анти-CXCR3 антитело или его антигенсвязывающий фрагмент включают CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL и/или CDR3 VL, содержащие аминокислотные последовательности, идентичные последовательностям CDR VH и VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов, или имеющие 1, 2, 3, 4 или 5 мутаций аминокислотных остатков (включая добавления, делеции и замены, такие как консервативные замены).
В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его фрагмент включают тяжелую цепь, имеющую три CDR (CDR1, CDR2 и CDR3 тяжелой цепи), и легкую цепь, имеющую три CDR (CDR1, CDR2 и CDR3 легкой цепи). В некоторых вариантах осуществления изобретения, CDR1 VH имеет 1, 2 или 3 аминокислотных мутации в последовательности CDR1 VH любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, CDR2 VH имеет 1, 2 или 3 аминокислотных мутации в последовательности CDR2 VH любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, CDR3 VH имеет 1, 2 или 3 аминокислотных мутации в последовательности CDR3 VH любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, CDR1 VL имеет 1, 2 или 3 аминокислотные мутации в последовательности CDR1 VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, CDR2 VL имеет 1, 2 или 3 аминокислотных мутации в последовательности CDR2 VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, CDR3 VL имеет 1, 2 или 3 аминокислотных мутации в последовательности CDR3 VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, CDR1, CDR2 и CDR3 тяжелой и легкой цепи представляют собой CDR любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов, или включают 1-3 аминокислотных мутации в последовательности CDR, выбранной из любого клона антитела или их химерных/гуманизированных вариантов. В некоторых вариантах осуществления изобретения, указанные мутации находятся в выделенных положениях, показанных на фиг. 17A-H, на которых проиллюстрировано выравнивание последовательностей. В некоторых вариантах осуществления изобретения, мутация присутствует в одном или более положениях 58 и 59 в CDR2 VH любого из клонов 4.0-4.11.
В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его фрагмент включают тяжелую цепь и легкую цепь. В некоторых вариантах осуществления изобретения, тяжелая цепь по меньшей мере приблизительно на 90% (например, по меньшей мере приблизительно на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% или на любую промежуточную величину) идентична любой из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3, тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3, или имеет 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных мутаций. В некоторых вариантах осуществления изобретения легкая цепь по меньшей мере приблизительно на 90% (например, по меньшей мере приблизительно на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% или на любую промежуточную величину) идентична любой из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3, или имеет 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных мутаций. В некоторых вариантах осуществления изобретения тяжелая цепь по меньшей мере приблизительно на 90% (например, по меньшей мере приблизительно на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% или на любую промежуточную величину) идентична любой из тяжелых цепей 4.0-4.11, тяжелых цепей 12.0-12.3, тяжелых цепей 53.0-53.10, тяжелых цепей 82.0-82.3 и тяжелых цепей 135.0-135.3, или имеет 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных мутаций, и/или легкая цепь по меньшей мере приблизительно на 90% (например, по меньшей мере приблизительно на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% или на любую промежуточную величину) идентична любой из легких цепей 4.0-4.7, легких цепей 12.0-12.3, легких цепей 53.0-53.13, легких цепей 82.0-82.3 и легких цепей 135.0-135.3, или имеет 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных мутаций. В некоторых вариантах осуществления изобретения указанные мутации находятся в положениях, показанных на фиг. 17A-H, на которых проиллюстрировано выравнивание последовательностей.
В некоторых вариантах осуществления изобретения выделенное анти-CXCR3 антитело или его антигенсвязывающий фрагмент, содержащие вышеописанные последовательности CDR VH и/или VL, сохраняют свою CXCR3-нейтрализующую активность.
В различных вариантах осуществления изобретения вариабельные домены тяжелой и легкой цепи анти-CXCR3 антитела или его фрагмента могут содержать по меньшей мере одну каркасную область (например, по меньшей мере одну из FR1, FR2, FR3 и FR4). Каркасные области тяжелой цепи обозначены FR VH, а каркасные области легкой цепи обозначены FR VL. В некоторых вариантах осуществления изобретения каркасные области могут иметь замены, инсерции или другие модификации. В некоторых вариантах осуществления изобретения такие модификации позволяют улучшить или оптимизировать аффинность связывания антитела. Неограничивающими примерами остатков каркасной области, которые могут быть модифицированы, являются остатки, которые непосредственно и нековалентно связываются с CXCR3, взаимодействую с конформацией CDR или влияют на такую конформацию, и/или участвуют во VL-VH-взаимодействии.
В некоторых вариантах осуществления изобретения тяжелая цепь (VH) анти-CXCR3 антитела или его фрагмента может включать FR1, FR2, FR3 и/или FR4, имеющие аминокислотные последовательности, которые приблизительно на 80%-100% (например, приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%, или на любую промежуточную величину) идентичны соответствующим каркасным областям VH любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его фрагмент могут включать по меньшей мере одну FR (FR1, FR2, FR3 и/или FR4) VH, имеющую аминокислотную последовательность, идентичную соответствующим областям FR VH любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов или имеющую 1, 2, 3, 4 или 5 мутаций аминокислотных остатков (включая добавления, делеции и замены, такие как консервативные замены).
В некоторых вариантах осуществления изобретения легкая цепь (VL) анти-CXCR3 антитела или его фрагмента может включать FR1, FR2, FR3 и/или FR4, имеющие аминокислотные последовательности, которые приблизительно на 80%-100% (например, приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%, или на любую промежуточную величину) идентичны соответствующим каркасным областям VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его фрагмент могут включать по меньшей мере одну VL FR (FR1, FR2, FR3 и/или FR4) VL, имеющую аминокислотную последовательность, идентичную соответствующии областям FR VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов, или имеющую 1, 2, 3, 4 или 5 мутаций аминокислотных остатков (включая добавления, делеции и замены, такие как консервативные замены).
В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его фрагмент включают области FR (FR1, FR2, FR3 и/или FR4) VH, имеющие аминокислотную последовательность, идентичную соответствующим областям FR VH любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов, или имеющие 1, 2, 3, 4 или 5 аминокислотных мутаций, и включают области FR (FR1, FR2, FR3 и/или FR4) VL, имеющие аминокислотную последовательность, идентичную соответствующим областям VL FR любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов или имеющие 1, 2, 3, 4 или 5 аминокислотных мутаций. В некоторых вариантах осуществления изобретения, анти-CXCR3 антитело или его фрагмент включают области FR (FR1, FR2, FR3 и/или FR4) VH, имеющие аминокислотные последовательности, которые приблизительно на 80%-100% идентичны соответствующим областям VH FR любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов, и области FR (FR1, FR2, FR3 и/или FR4) VL, имеющие аминокислотную последовательность, которая приблизительно на 80%-100% идентична соответствующим областям FR VL любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов.
Описанные здесь области CDR и FR могут быть объединены в различные комбинации, где каждая из областей CDR и FR может быть независимо выбрана и объединена с любой другой областью CDR или FR данного антитела. В некоторых вариантах осуществления изобретения, последовательности CDR и FR VH и/или VL могут присутствовать в любой комбинации в антителе или в его фрагменте, которые сохраняют свою способность нейтрализовать активность CXCR3.
Описанные здесь антитела и их фрагменты могут содержать одну или более аминокислотных последовательностей, которые, по существу, не вносят изменений в описанные здесь аминокислотные последовательности. Аминокислотные последовательности, которые, по существу, являются аналогичными, включают последовательности, содержащие консервативные аминокислотные замены, а также аминокислотные делеции и/или инсерции, не влияющие на способность антитела или его фрагмента нейтрализовать активность CXCR3.
Описанные здесь антитела и их фрагменты могут быть также конъюгированы с одной или более дополнительными молекулами. Так, например, конъюгат может включать антитело, присоединенное непосредственно или посредством линкера к одному или более терапевтическим агентам, солюбилизирующим агентам, стабилизаторам, иммунодепрессантам, рецепторам и их фрагментам, антигенсвязывающим пептидам и/или другим молекулам, нацеленным на лиганд. В некоторых вариантах осуществления изобретения терапевтическим агентом является агент, который может быть использован для лечения T1D и/или других расстройств, ассоциированных с CXCR3. В некоторых вариантах осуществления изобретения антитело или его фрагмент конъюгированы с агентом, стимулирующим β-клетки, или с инсулином.
Нуклеотидные последовательности
В некоторых вариантах осуществления изобретения помимо вышеописанных аминокислотных последовательностей описаны также нуклеотидные последовательности, соответствующие этим аминокислотным последовательностям. В некоторых вариантах осуществления изобретения нуклеотидная последовательность кодирует антитело или его фрагмент, способные нейтрализовать активность CXCR3. В некоторых вариантах осуществления изобретения нуклеотидные последовательности могут быть использованы в целях конструирования экспрессионных векторов для экспрессии анти-CXCR3 антител в клетках (например, экспрессии в клетках млекопитающих).
В некоторых своих вариантах настоящее изобретение также относится к полинуклеотидам, по существу, идентичным полинуклеотидам, кодирующим описанные здесь аминокислотные последовательности. По существу, идентичные последовательности могут представлять собой полиморфные последовательности, то есть альтернативные последовательности или аллели в данной популяции. По существу, идентичные последовательности могут также содержать мутагенные последовательности, включая последовательности, содержащие молчащие мутации. Термин «мутация» может включать одну или более замен нуклеотидных остатков, делеций одного или более нуклеотидных остатков или инсерций одного или более дополнительных нуклеотидных остатков. По существу, идентичные последовательности могут также включать различные нуклеотидные последовательности, кодирующие одну и ту же аминокислоту в любом указанном аминокислотном положении описанной здесь аминокислотной последовательности, что обусловлено вырожденностью кода нуклеиновой кислоты. В настоящем изобретении также рассматриваются, по существу, идентичные последовательности, которые представляют собой последовательности, кодирующие цепь или цепи антитела, сохраняющего свою способность нейтрализовать CXCR3.
В некоторых своих вариантах настоящее изобретение также относится к полинуклеотидам, которые гибридизуются в условиях гибридизации высокой или низкой жесткости с полинуклеотидами, кодирующими CXCR3-нейтрализующее антитело или его фрагмент. Используемый здесь термин «условия жесткости» означает условия (например, температуру и концентрацию соли) эксперимента по гибридизации, проводимого для оценки степени гомологии между двумя нуклеиновыми кислотами, причем, чем выше жесткость, тем выше процент гомологии между двумя нуклеиновыми кислотами. Используемый здесь термин «гибридизующийся» или его грамматические формы относятся к связыванию первой молекулы нуклеиновой кислоты со второй молекулой нуклеиновой кислоты в условиях низкой, умеренной или высокой жесткости или в условиях проведения синтеза нуклеиновых кислот. Понятие «гибридизация» может включать случаи, в которых первая молекула нуклеиновой кислоты связывается со второй молекулой нуклеиновой кислоты, и случаи, в которых первая и вторая молекула нуклеиновой кислоты являются комплементарными.
Условия жесткости гибридизации включают, но не ограничиваются ими, гибридизацию связанной с фильтром ДНК в 6× хлориде натрия/цитрате натрия (SSC) приблизительно при 45°C с последующей одной или более промывками в 0,2×SSC/0,1% ДСН приблизительно при 50-65°C. Другие условия жесткости гибридизации включают гибридизацию связанной с фильтром ДНК в 6×SSC приблизительно при 45°C с последующей одной или более промывками в 0,1×SSC/0,2% ДСН приблизительно при 65°C. Специалистам известны и другие условия жесткости гибридизации, описанные в настоящей заявке.
В некоторых вариантах осуществления изобретения описанная здесь нуклеиновая кислота может кодировать аминокислотную последовательность цепи или цепей антитела или его фрагмента, обладающих способностью нейтрализовать активность CXCR3, либо указанная нуклеиновая кислота может гибридизоваться в жестких условиях с нуклеиновой кислотой, кодирующей аминокислотную последовательность цепи или цепей антитела или его фрагмента.
В некоторых вариантах осуществления изобретения описанная здесь полинуклеотидная последовательность содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность домена VH CXCR3-нейтрализующего антитела или его фрагмента и которая по меньшей мере приблизительно на 80-100% (например, приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% или на любую промежуточную величину) идентична нуклеотидной последовательности, кодирующей тяжелую цепь любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения полинуклеотидная последовательность может включать нуклеотидную последовательность, имеющую 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мутаций (включая добавления, делеции и замены, такие как консервативные замены) по сравнению с нуклеотидной последовательностью, кодирующей тяжелую цепь любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов.
В некоторых вариантах осуществления изобретения описанная здесь полинуклеотидная последовательность содержит нуклеотидную последовательность, которая кодирует аминокислотную последовательность домена VL CXCR3-нейтрализующего антитела или его фрагмента, и которая по меньшей мере приблизительно на 80-100% (например, приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% или на любую промежуточную величину) идентична нуклеотидной последовательности, кодирующей легкую цепь любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов. В некоторых вариантах осуществления изобретения полинуклеотидная последовательность может включать нуклеотидную последовательность, имеющую 0, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мутаций (включая добавления, делеции и замены, такие как консервативные замены) по сравнению с нуклеотидной последовательностью, кодирующей легкую цепь любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов.
В конкретных вариантах осуществления изобретения описанная здесь полинуклеотидная последовательность содержит нуклеотидную последовательность, которая по меньшей мере приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% (или на любую промежуточную величину) идентична нуклеотидной последовательности, кодирующей аминокислотную последовательность VH, и по меньшей мере приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% (или на любую промежуточную величину) идентична нуклеотидной последовательности, кодирующей аминокислотную последовательность VL, где указанные нуклеотидные последовательности кодируют аминокислотные последовательности тяжелой и легкой цепей любого из клонов 12, 135, 82, 53 и 4 и их химерных и гуманизированных вариантов.
Описанные полинуклеотиды могут быть получены любым известным методом. Так, например, если нуклеотидная последовательность антитела является известной, то полинуклеотид, кодирующий антитело, может быть сконструирован с использованием химически синтезированных олигонуклеотидов. Такой способ может включать, например, синтез перекрывающихся олигонуклеотидов, содержащих части последовательности, кодирующей антитело; отжиг и лигирование этих олигонуклеотидов, а затем амплификацию лигированных олигонуклеотидов с помощью ПЦР. Описанные полинуклеотиды могут быть также сконструированы из любого другого подходящего источника нуклеиновых кислот, такого как библиотека кДНК антител или библиотека кДНК, выделенных из любых тканей или клеток, экспрессирующих указанное антитело (например, из выбранных гибридомных клеток, экспрессирующих антитело).
В некоторых вариантах осуществления изобретения любые описанные полинуклеотиды могут быть включены в экспрессионный вектор. Векторы, подходящие для экспрессии в различных клетках человека и животных, известны специалистам. В некоторых своих вариантах настоящее изобретение относится к клеткам-хозяевам, содержащим указанные векторы. Подходящими клетками-хозяевами являются, например, CHO, COS, SF9 и/или другие человеческие или нечеловеческие клеточные линии. В некоторых вариантах осуществления изобретения, клетки-хозяева, при их культивировании в культуральной среде, транзиентно или стабильно экспрессируют нуклеиновую кислоту в векторе, и эти клетки используют в способе продуцирования описанных здесь антител или их фрагментов.
Фармацевтические композиции
Фармацевтическая композиция может содержать любые описанные здесь антитела или их фрагменты. Также описаны фармацевтические композиции, содержащие нуклеиновые кислоты, кодирующие антитела или их фрагменты и используемые, например, в генотерапии и/или для транзиентной или стабильной экспрессии в клетках-хозяевах (например, в CHO, COS, SF9 и/или в других человеческих или нечеловеческих клеточных линиях) с получением белков или их фрагментов.
Описанные здесь фармацевтические композиции могут содержать фармацевтически приемлемый носитель и/или по меньшей мере одну добавку, такую как растворитель, наполнитель, агент для придания объема, дезинтегратор, буфер или стабилизатор. Стандартные способы приготовления фармацевтических композиций известны специалистам (смотрите, например, 2005 Physicians' Desk Reference®, Thomson Healthcare: Montvale, J., 2004; Remington: The Science and Practice of Pharmacy, 20th ed., Gennado et al., Eds. Lippincoft Williams & Wilkins: Philadelphia, PA, 2000). Подходящими фармацевтическими добавками являются, например, маннит, крахмал, глюкоза, лактоза, сахароза, желатин, солод, рис, мука, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое сепарированное молоко, глицерин, пропиленгликоль, вода, этанол и т.п. В некоторых вариантах осуществления изобретения фармацевтические композиции могут также содержать pH-забуферивающие реагенты и смачивающие или эмульгирующие агенты (например, забуференный фосфатом физиологический раствор, стерильный физиологический раствор для инъекций и т.п.). В других вариантах осуществления изобретения указанные композиции могут содержать консерванты или другие стабилизаторы.
В некоторых вариантах осуществления изобретения фармацевтические композиции, включающие любые описанные здесь антитела или их фрагменты или нуклеиновые кислоты, кодирующие такие антитела или их фрагменты, могут также включать один или более из нижеследующих компонентов, таких как маннит, полисорбат 80, двухосновный гептагидрат фосфата натрия и одноосновный моногидрат фосфата натрия. В другом варианте осуществления изобретения, фармацевтические композиции могут содержать 10 мМ гистидина, pH 6,5; до 2% глицина, до 2% маннита и до 0,01% полисорбата 80.
Состав фармацевтических композиций может в значительной степени варьироваться в зависимости от предполагаемого способа их введения и других параметров (смотрите, например, Rowe et al., Handbook of Pharmaceutical Excipients, 4th ed., APhA Publications, 2003). В некоторых вариантах осуществления изобретения фармацевтическая композиция может представлять собой лиофилизованный брикет или порошок. Лиофилизованная композиция может быть разведена для введения путем внутривенной инъекции, например, стерильной водой для инъекций, USP. В других вариантах осуществления изобретения композиция может представлять собой стерильный апирогенный раствор.
Описанные здесь фармацевтические композиции могут содержать описанное здесь антитело или его фрагмент, или нуклеиновые кислоты, кодирующие антитела или их фрагменты, используемые в качестве одного активного соединения, либо фармацевтическая композиция может содержать комбинацию с другим соединением, другой композицией или другим биологическим материалом. Так, например, фармацевтическая композиция может также содержать одну или более небольших молекул или других агентов, используемых для лечения заболевания или расстройства, ассоциированного с CXCR3, такого как T1D. В некоторых вариантах осуществления изобретения, фармацевтическая композиция может содержать агент, стимулирующий β-клетки, инсулин и/или инсулин-продуцирующую клетку. В некоторых вариантах осуществления изобретения фармацевтическая композиция может также содержать один или более иммунодепрессантов, ингибиторов mTOR или ингибиторов аутофагии. Примерами иммунодепрессантов являются рапамицин и велкад. Рапамицин также является ингибитором mTOR.
Введение и дозы
В некоторых своих вариантах настоящее изобретение относится к способу лечения пациента, страдающего заболеванием или расстройством, ассоциированным с CXCR3, где указанный способ включает введение пациенту одного или более описанных здесь анти-CXCR3 антител и/или их фрагментов. В некоторых вариантах осуществления изобретения антитело или его фрагмент способны нейтрализовать CXCR3. В некоторых вариантах осуществления изобретения заболеванием или расстройством является воспалительное расстройство. В некоторых вариантах осуществления изобретения указанным расстройством является T1D. В некоторых вариантах осуществления изобретения введение композиции (например, фармацевтической композиции), содержащей антитело или его фрагмент, осуществляют для предупреждения, лечения, снижения тяжести и/или какого-либо другого ослабления симптомов заболевания или расстройства, ассоциированного с CXCR3. В некоторых вариантах осуществления изобретения антитело или его фрагмент вводят в дозе и с частотой, достаточными для предупреждения, лечения, снижения тяжести и/или какого-либо другого ослабления симптомов заболевания или расстройства, ассоциированного с CXCR3.
В некоторых своих вариантах настоящее изобретение относится к композиции, используемой в целях приготовления лекарственного препарата для лечения заболевания или расстройства, где указанный лекарственный препарат содержит любые из описанных здесь антител или их фрагментов. Так, например, антитело или его фрагмент могут содержать три CDR любого из вариабельных доменов тяжелой цепи 4.0-4.11, присоединенные к трем CDR любого из вариабельных доменов легкой цепи 4.0-4.7; три CDR любого из вариабельных доменов тяжелой цепи 12.0-12.3, присоединенные к трем CDR любого из вариабельных доменов легкой цепи 12.0-12.3; три CDR любого из вариабельных доменов тяжелой цепи 53.0-53.10, присоединенные к трем CDR любого из вариабельных доменов легкой цепи 53.0-53.13; три CDR любого из вариабельных доменов тяжелой цепи 82.0-82.3, присоединенные к трем CDR любого из вариабельных доменов легкой цепи 82.0-82.3; или три CDR любого из вариабельных доменов тяжелой цепи 135.0-135.3, присоединенные к трем CDR любого из вариабельных доменов легкой цепи клона 135.0-135.3.
В некоторых вариантах осуществления изобретения антитело или его фрагмент в указанном лекарственном препарате может содержать любой из вариабельных доменов тяжелой цепи 4.0-4.11, присоединенных к любому из вариабельных доменов легкой цепи 4.0-4,7; любой из вариабельных доменов тяжелой цепи 12.0-12.3, присоединенных к любому из вариабельных доменов легкой цепи 12.0-12.3; любой из вариабельных доменов тяжелой цепи 53.0-53.10, присоединенных к любому из вариабельных доменов легкой цепи 53.0-53.13; любой из вариабельных доменов тяжелой цепи 82.0-82.3, присоединенных к любому из вариабельных доменов легкой цепи 82.0-82.3; или любой из вариабельных доменов тяжелой цепи 135.0-135.3, присоединенных к любому из вариабельных доменов легкой цепи 135.0-135.3.
Дозы анти-CXCR3 антител, используемых в описанных здесь способах, могут варьироваться в зависимости от различных известных параметров, таких как физиологические особенности пациента (телосложение или площадь поверхности тела, масса тела, возраст и метаболизм) и патологическое состояние, а также от фармакологических параметров, таких как механизм доставки лекарственного средства, состав лекарственного препарата и любая сопутствующая или последующая терапия. Термин «эффективное количество» анти-CXCR3 антитела означает количество, обеспечивающее один или более желаемых in vivo эффектов, таких как сохранение или снижение уровней HA1bc (гемоглобина A1c) (приблизительно менее, чем на 7%, например, менее, чем на 7,5, 7,4, 7,3, 7,2, 7,1, 7,0, 6,9, 6,8, 6,7, 6,6 или 6,5%, или на любую промежуточную величину), увеличение уровней продуцирования эндогенного инсулина и/или уровней инсулина в кровотоке; сохранение или увеличение уровней С-пептида натощак; повышение толерантности к глюкозе; снижение уровней глюкозы в крови натощак в отсутствии экзогенного инсулина; снижение степени утилизации экзогенного инсулина; снижение степени β-клеточного воспаления и/или увеличение популяции и/или усиление роста β-клеток. Могут быть также проведены прямые клеточные анализы на ингибирование CXCR3, например, анализы на снижение числа CXCR3+-клеток в крови (включая, но не ограничиваясь ими, Т-клетки), на ингибирование связывания с лигандом CXCR3, связывания с GTP, притока и/или мобилизации кальция, хемотаксиса клеток и/или интернализации рецептора.
В некоторых вариантах осуществления изобретения эффективное количество анти-CXCR3 антитела обеспечивает улучшение любого из вышеописанных параметров in vivo, по сравнению с контролем, приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, или 100% (или на промежуточную величину в процентах) или в 1, 2, 4, 5, 10, 15, 20, 30, 40, 50 или 100 раз (или в любое промежуточное число раз) или более. В некоторых вариантах осуществления изобретения такое улучшение может быть охарактеризовано как снижение уровня глюкозы в крови натощак в отсутствии экзогенного инсулина до величины ниже 100, 110, 120, 130, 140, 150, 160, 180, 200, 220, 240, 250, 300 или 350 мг/дл (или до любой промежуточной величины). В некоторых вариантах осуществления изобретения, такое улучшение может быть охарактеризовано как увеличение базальных уровней С-пептида в сыворотке до величины более, чем 0,2, 0,3, 0,4, 0,5, 0,6, 0,8 или 1,0 нмоль/л (или до любой промежуточной величины). В некоторых вариантах осуществления изобретения, такое улучшение может быть охарактеризовано как увеличение общих уровней С-пептида в сыворотке натощак во время введения С-пептида (в постпероральном тесте на толерантность к глюкозе) до величины приблизительно более, чем 0,03, 0,033, 0,04, 0,05, 0,06, 0,07, 0,08, 0,09, 0,1, 0,2, 0,4, 0,5, 0,6, 0,8 или 1,0 нмоль/л в минуту (или до любой промежуточной величины). В некоторых вариантах осуществления изобретения эффективная доза анти-CXCR3 антитела может быть также охарактеризована как доза, снижающая концентрацию CD4+- и/или CD8+-клеток в поджелудочной железе по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% (или на любой промежуточную процентную величину) или по меньшей мере в 1, 2, 3, 4 или 5 раз (или в любое промежуточное число раз), по сравнению с контрольными индивидуумами.
В различных вариантах осуществления изобретения эффективная доза антитела также выбрана как безопасная доза для введения человеку. В некоторых вариантах осуществления изобретения такая безопасная доза анти-CXCR3 антитела может быть охарактеризована как доза, которая не дает какого-либо заметного истощения Т-клеток или нарушения Т-клеточной активности (но не активности CXCR3) у индивидуума (например, как было определено путем оценки концентрации или активности Т-клеток в крови индивидуума). В конкретных вариантах осуществления изобретения выражение «отсутствие какого-либо заметного истощения Т-клеток или нарушения Т-клеточной активности» означает снижение концентрации и/или активности (за исключением активности CXCR3) CD4+- и/или CD8+-клеток у индивидуума, подвергаемого лечению CXCR3-нейтрализующаим антителом, на 40%, 30%, 25%, 20%, 15%, 10%, 5% (или на любую промежуточную процентную величину) по сравнению с контрольными индивидуумами, которым вводили плацебо, и/или по сравнению с индивидуумами, которые ранее уже проходили курс лечения. В некоторых вариантах осуществления изобретения такая безопасная доза также охарактеризована как доза, обеспечивающая снижение концентрации клеток одного или более типов, выбранных из регуляторных Т-клеток, В-клеток, миелоидных клеток, дендритных клеток и/или гранулоцитов, на 40%, 30%, 25%, 20%, 15%, 10%, 5% (или на любую промежуточную величину) по сравнению с контрольными индивидуумами.
Репрезентативные неограничивающие дозы для индивидуума, такого как человек, составляют приблизительно 0,03, 0,06, 0,12, 0,24, 0,5, 1,0, 1,5, 2,0, 2,5, 3,0, 3,5 или 3,7 мг/кг/дозу (или любое промежуточное значение) антитела с ND50 приблизительно 1-6 мкг/мл в анализе на хемотаксис in vitro. В некоторых вариантах осуществления изобретения, антитело может быть введено в количестве, составляющем приблизительно в пределах 0,03-3,7 мг/кг/дозу, 0,15-0,7 мг/кг/дозу или 0,25-0,5 мг/кг/дозу.
Анти-CXCR3 антитело может быть введено один раз или несколько раз в течение определенного периода времени, например, ежедневно, раз в неделю, раз в две недели, раз в месяц, раз в два месяца, раз в три месяца или ежегодно. В соответствии с этим, в неограничивающем примере, в котором обсуждаются интервалы доз, указанные выше, пациенту может быть введена общая доза приблизительно 0,16-18 мг/кг анти-CXCR3 антитела в течение всего курса лечения.
Анти-CXCR3 антитела могут быть введены любым подходящим способом, известным специалистам, включая, например, внутривенное, внутрибрюшинное, интраназальное, внутриглазное, пероральное, парентеральное, подкожное или трансдермальное введение. В конкретных вариантах осуществления изобретения антитело может быть введено непосредственно в поджелудочную железу индивидуума или в участок, находящийся поблизости от поджелудочной железы, или в определенные области поджелудочной железы, такие как островковые клетки поджелудочной железы.
Дозы, которые являются эффективными для одного животного, могут быть преобразованы так, чтобы они были эффективными для другого животного, включая человека, с использованием коэффициентов преобразования, известных специалистам. Смотрите, например, Reagan-Shaw et al., FASEB J. 22:659-61 (2008); Schein et al., Clin. Pharmacol. Ther. 11: 3-40 (1970); и Freireich et al., Cancer Chemother. Reports 50(4):219-244 (1966). Так, например, эквивалентная для человека доза (HED) в мг/кг может быть рассчитана исходя из дозы для животного по следующему уравнению: HED (мг/кг) = доза для животного (мг/кг)×(Kmживотного/Kmчеловека), где Km = масса/площадь поверхности тела (кг/м2).
Репрезентативные коэффициенты преобразования, полученные исходя из вышеуказанного уравнения, представлены в нижеследующей таблице. Описанные выше репрезентативные дозы для человека могут быть скорректированы для животного другого вида или для другого человека с использованием указанных коэффициентов или другими известными способами.
Индивидуумы
Индивидуумами, подвергаемыми лечению способами согласно изобретению, могут быть человек или животные, такие как крупный рогатый скот, домашние животные и дикие животные. В некоторых вариантах осуществления изобретения, животными являются птицы, коровы, собаки, киты, лошади, кошки, овцы, рыбы (Pisces), свиньи, приматы, грызуны или копытные. Индивидуумами могут быть индивидуумы на любой стадии развития, включая взрослых индивидуумов, молодых индивидуумов, плод или эмбрион. В конкретных вариантах осуществления изобретения пациентом является млекопитающее, а в более конкретных вариантах осуществления изобретения, человек.
В различных вариантах осуществления изобретения индивидуум может быть подвернут лечению либо в профилактических целях, либо после начала развития любого состояния, ассоциированного с нарушением активности CXCR3, или любого состояния, при котором нарушение передачи CXCR3-сигнала может давать терапевтический эффект. В некоторых вариантах осуществления изобретения индивидуум может быть подвернут лечению либо в профилактических целях, либо после начала развития T1D. В некоторых вариантах осуществления изобретения индивидуум может быть подвернут лечению в профилактических целях до начала развития T1D описанными здесь способами, либо индивидуум, у которого был впервые выявлен T1D, может быть подвернут лечению описанными здесь способами.
Выражение «индивидуум, у которого был впервые выявлен T1D», относится к любому индивидууму, у которого наблюдается уже детектируемое снижение инсулин-продуцирующей способности β-клеток поджелудочной железы, независимо от возраста индивидуума, у которого был клинически диагностирован диабет (например, включая взрослых индивидуумов, детей, плод или эмбрион). В некоторых вариантах осуществления изобретения, индивидууму, у которого был впервые выявлен T1D, назначают лечение предпочтительно в течение приблизительно шести месяцев (например, приблизительно в течение 1 дня, 1 недели, 2 недель, 3 недель, 4 недель, 5 недель, 1 месяца, 2 месяцев, 3 месяцев, 4 месяцев, 5 месяцев, 6 месяцев или в течение любого промежуточного периода времени) после самого раннего установления диагноза T1D. В других вариантах осуществления изобретения индивидууму может быть назначено лечение в течение более шести месяцев после самого раннего установления диагноза T1D, где у указанного индивидуума сохраняются минимальные, но еще измеримые базальные уровни С-пептида в сыворотке, составляющие приблизительно 0,2 ммоль/л или более (например, по меньшей мере приблизительно 0,2, 0,3, 0,4, 0,5, 0,6, 0,8 или 1,0 нмоль/л). В некоторых вариантах осуществления изобретения такое лечение включает введение одной или более доз, содержащих одно или более описанных здесь антител. В некоторых вариантах осуществления изобретения указанным антителом является CXCR3-нейтрализующее антитело.
В некоторых вариантах осуществления изобретения у индивидуума, у которого был впервые выявлен T1D, сохраняются общие уровни С-пептида в сыворотке натощак, составляющие по меньшей мере приблизительно 0,03, 0,033, 0,04, 0,05, 0,06, 0,07, 0,08, 0,09, 0,1, 0,2, 0,4, 0,5, 0,6, 0,8 или 1,0 нмоль/л в минуту, например, приблизительно 0,03-1,0 или 0,033-1,0 нмоль/л в минуту во время стимуляции C-пептидом. В конкретных вариантах осуществления изобретения у указанного индивидуума общие уровни С-пептида в сыворотке натощак составляют 0,033-1,0 нмоль/л в минуту во время стимуляции C-пептидом. В некоторых вариантах осуществления изобретения стимуляцию С-пептидом осуществляют после проведения анализа с пероральным введением глюкозы, и такой анализ может включать измерение общих уровней С-пептида в сыворотке в течение 60-150 минут после введения 10,0-13,9 ммоль/л глюкозы. Смотрите Keymeulen et al., Diabetologia 53: 614-623 (2010). В более конкретных вариантах осуществления изобретения у индивидуума, после проведения анализа на постпероральную толерантность к глюкозе, измеримые общие уровни С-пептида в сыворотке увеличиваются до 0,8, 0,7, 0,6, 0,54, 0,5, 0,4, 0,3, 0,2 или 0,1 нмоль/л в минуту или менее. В более конкретных вариантах осуществления изобретения, у индивидуума, после проведения анализа на постпероральную толерантность к глюкозе, измеримые общие уровни С-пептида в сыворотке увеличиваются до 0,54 нмоль/л в минуту или менее. C-пептид соответствует остаткам 57-87 пептида-предшественника инсулина (человеческая эталонная последовательность NP_000198), а остатки 90-110 и 25-54 соответствуют A- и B-цепям инсулина, соответственно.
В некоторых вариантах осуществления изобретения у индивидуума, у которого был впервые выявлен T1D, наблюдаются повышенные уровни глюкозы в крови натощак в отсутствии экзогенного инсулина, превышающие приблизительно 100, 110, 120, 130, 140, 150, 160, 180, 200, 220, 240, 250, 300, 350 мг/дл (или любую промежуточную величину) или более. В некоторых вариантах осуществления изобретения у указанного индивидуума могут наблюдаться повышенные уровни глюкозы в крови натощак, как описано выше, а также пониженные общие уровни С-пептида в сыворотке натощак, как описано выше.
В некоторых вариантах осуществления изобретения, индивидуума подвергают лечению описанными здесь способами сразу после первого диагностирования у него впервые выявленного T1D. В более конкретных вариантах осуществления изобретения, индивидуума подвергают предварительному лечению способами согласно изобретению в течение 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дней, или в течение 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 недель, или в течение 1 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 месяцев (или в течение любого промежуточного периода времени) после впервые установленного клинического диагноза T1D. В более конкретных вариантах осуществления изобретения, индивидуума подвергают предварительному лечению способами согласно изобретению в течение 6 месяцев после впервые установленного клинического диагноза T1D. В других вариантах осуществления изобретения, индивидуума, страдающего T1D, подвергают лечению описанными здесь способами в любой период времени, независимо от того, когда был установлен клинический диагноз T1D, где у указанного индивидуума сохраняется остаточные уровни С-пептида в сыворотке, составляющие по меньшей мере приблизительно 0,2 нмоль/л.
Дополнительные способы
В некоторых вариантах осуществления изобретения описанные здесь способы могут включать дополнительные курсы терапии, которые могут быть проведены одновременно или последовательно с введением (до или после введения) описанного здесь анти-CXCR3 антитела. Так, например, в некоторых вариантах осуществления изобретения, описанные способы включают, помимо введения указанному индивидууму анти-CXCR3 антитела, дополнительную стадию введения иммунодепрессанта. Иммунодепрессантами могут быть, но не ограничиваются ими, один или более препаратов, таких как азатиоприн (имуран), β-интерферон 1a, β-интерферон 1b, базиликсимаб, кортикостероиды, циклоспорин (Sandimmune), циклофосфамид, хлорамбуцил, даклизумаб, дезоксиспергуалин, этанерцепт, ацетат глатирамера, инфликсимаб, лефлуномид, меркаптопурин (6-MP), метотрексат, митоксантрон, муромонаб-CD3, микофенолят (MFM или CellCept), натализумаб, анакинра, канакинумаб, ритуксимаб, белимумаб, абатацепт, альдеслейкин, преднизон, рапамицин, сиролимус, такролимус и устекинумаб.
В некоторых вариантах осуществления изобретения описанные здесь способы могут включать, помимо введения указанному индивидууму анти-CXCR3 антитела, стадию введения агента, стимулирующего β-клетки. Стадия введения агента, стимулирующего β-клетки, может быть проведена одновременно или последовательно с введением (до или после введения) анти-CXCR3 антитела. Репрезентативными агентами, стимулирующими β-клетки, являются, но не ограничиваются ими, один или более агентов, таких как трансплантированные β-клетки (аутологичные, аллогенные или сингенные), трансплантированные инсулин-продуцирующие клетки (аллогенные или сингенные), ингибиторы DDP4 (последовательность эталонного человеческого белка NP_001926.2), пептиды TM4SF20 (последовательность эталонного человеческого белка NP_079071), пептиды TMEM27 (последовательность эталонного человеческого белка NP_065716), аналоги экзендина 1 или GLP-1 (последовательность эталонного человеческого белка NP_002045), лиганды рецептора gp130 и EGF, и агенты, описанные в параграфах 8-11 публикации заявки на патент США №20100130476. В способах согласно изобретению, агент, стимулирующий β-клетки, может быть введен вместе с иммунодепрессантом одновременно или последовательно (до или после введения иммунодепрессанта). В некоторых вариантах осуществления изобретения агент, стимулирующий β-клетки, инсулинпродуцирующие клетки и/или иммунодепрессант могут быть введены путем имплантации устройства, способного доставлять агент, стимулирующий β-клетки, инсулинпродуцирующие клетки и/или иммунодепрессант в нужную ткань или в нужный орган.
В настоящем изобретении также описаны способы детектирования и/или количественной оценки CXCR3 и/или клеток, экспрессирующих CXCR3 (например, CXCR3+-T-клеток). В некоторых вариантах осуществления изобретения указанные способы включают применение одного или более описанных здесь анти-CXCR3 антител для детектирования и/или количественной оценки CXCR3 и/или клеток, экспрессирующих CXCR3. Так, например, одно или более антител могут быть добавлены в образец, взятый у пациента (например, в пробу крови), и детектированы с использованием «включенный» не имеют какой-либо ограничений. При этом подразумевается, что любой описанный здесь интервал значений включает граничные значения и все величины между этими граничными значениями.
Приведенные здесь заголовки разделов даны лишь в организационных целях и не должны рассматриваться как ограничение предмета изобретения. Все цитируемые здесь документы или части документов, включая, но не ограничиваясь ими, патенты, патентные заявки, статьи, книги и монографии точно, во всей своей полноте и в любых целях вводятся в настоящее описание посредством ссылки. Если содержание публикаций и патентов или патентных заявок, включенных в настоящее описание посредством ссылки, противоречит описанию настоящей заявки, то следует отдать предпочтение описанию, представленному в настоящей заявке.
Вся информация об эталонных генных последовательностях, представленная в настоящей заявке, такая как данные GeneID или регистрационные номера, включая, например, геномные локусы, геномные последовательности, описания функций, аллельные варианты и эталонные мРНК (включая, например, границы экзонов) и последовательности белков (такие как структуры консервативных доменов), во всей своей полноте вводится в настоящее описание посредством ссылки.
ПРИМЕРЫ
Нижеследующие примеры служат в качестве иллюстрации, а поэтому никоим образом не должны рассматриваться как ограничение настоящего изобретения.
Пример 1: Материалы и методы
Продуцирование иммуногена. Клетки CHO трансформировали ДНК, кодирующей полноразмерный человеческий CXCR3, и CXCR3 экспрессировали на поверхности клеток («клеток r-CXCR3-CHO»). Была получена последовательность CXCR3, которую использовали для трансформации клеток, и открытую рамку считывания CXCR3 встраивали в экспрессионный вектор pcDNA3.1neo_DEST, а затем переносили в клетки 300-19 (Immunogen). N-концевой пептидный фрагмент внеклеточного домена CXCR3 (EC-домена) с аминокислотной последовательностью: MVLEVSDHQVLNDAEVAALLENFSSSYDYGENESDSC (SEQ ID NO:81) конъюгировали с KLH посредством C-концевого цистеина, и использовали в качестве иммуногена. CXCR3-экспрессирующие клетки выдерживали при 37°C в атмосфере 5% CO2 в среде RPMI (Invitrogen, Carlsbad, Calif.), в которую была добавлена 10% диализованная фетальная бычья сыворотка (FBS) (Invitrogen). Клетки для инъекции получали путем замены вышеописанной культуральной среды забуференным фосфатом физиологическим раствором (без Ca/Mg) (CMF-PBS), в который было добавлено 5 мМ ЭДТА, и клетки собирали в этом буфере. Собранные клетки осаждали путем центрифугирования при 500×g в течение приблизительно 5 минут, а затем один раз промывали путем ресуспендирования осадка в CMF-PBS, и центрифугировали как указано выше, после чего клетки подсчитывали и, для инъекции, доводили до соответствующего объема (например, 5×106 клеток в 0,2 мл) путем ресуспендирования клеточного осадка в CMF-PBS.
Получение антител. 37 N-концевых аминокислот человеческого CXCR3 использовали для продуцирования мышиных моноклональных гибридом, содержащих антитело против человеческого CXCR3, и пять клонов антител (4, 12, 53, 82 и 135) отбирали для последующей характеризации. 37 N-концевых аминокислот человеческого CXCR3 были на 65% гомологичны аминокислотам выровненной области мышиного CXCR3 и содержали остатки, играющие важную роль в связывании с CXCL9, CXCL10 и CXCL11. Приблизительно 6-8-недельных мышей BALB/c (Charles River Laboratories, Wilmington, MA) иммунизовали клетками, экспрессирующими CXCR3 или внеклеточный пептид CXCR3. Группу мышей подкожно (s.c.) примировали на день 0 эмульсией KLH-конъюгированного пептида, смешанного с адъювантом, 1:1 (TiterMax Gold, Sigma Aldrich, #12685-1ML), а затем 3-5 раз с интервалами в 2-3 недели вводили s.c. эмульсию пептида с адъювантом 1:1, или внутрибрюшинно (i.p.) вводили клетки в PBS без адъюванта, после чего два дня подряд вводили i.p. бустер-дозу пептида и/или клеток в PBS без адъюванта, и мышей умерщвляли. Вторую группу мышей примировали i.p. 3-5 раз с интервалами в 2-3 недели и два дня подряд вводили i.p. бустер-дозу клеток в PBS, а затем мышей умерщвляли. Каждая инъекция, вводимая мышам обеих групп, содержала приблизительно 2×106 клеток в объеме приблизительно 100 мкл.
На следующий день после последней инъекции, мышей умерщвляли, удаляли селезенку, и эту селезенку помещали приблизительно в 10 мл бессывороточной DMEM (Gibco) в чашку Петри. Мышиные миеломные клетки Sp2\О (ATCC CRL-1581) подвергали слиянию с клетками селезенки, взятыми у иммунизованных мышей, с использованием 50% (масс/масс) ПЭГ методом, описанным Kohler and Milstein (Nature, 256:495-7 (1975)). По окончании этой процедуры клетки ресуспендировали в 50 мл среды для регенерации гибридомы ClonaCell-Hy (StemCell Technologies), переносили в колбу T75-см2 и инкубировали в течение 16-24 часов при 37°C. После такого инкубирования клетки собирали и добавляли в 100 мл селективной среды с метилцеллюлозой ClonaCell-HY (StemCell Technologies). Затем эту смесь разделяли на десять 100-мм2 аликвот по чашкам с тканевой культурой и инкубировали в течение 10-14 дней. Клональные гибридомы переносили из среды с метилцеллюлозой в жидкую среду и культивировали в 96-луночном планшете в целях проведения анализов для идентификации CXCR3-специфических моноклональных антител.
Если это не оговорено особо, то в этих примерах в качестве варианта служило антитело Hu4.1, то есть антитело, содержащее вариант тяжелой цепи и вариант легкой цепи под одним и тем же номером (например, Hu4.1 должно содержать тяжелую цепь 4.1 и легкую цепь 4.1). Все ссылки на антитела относятся к антителам, которые были пронумерованы и содержали спаренные цепи VH/VK, как указано в таблице 2.
Животные. Самок мышей NOD/LtJ закупали в лаборатории Jackson Laboratory (Bar Harbor, ME), и этих мышей содержали в непатогенных условиях. Мышей скринировали на гликозурию с использованием прибора для измерения уровня глюкозы в крови ACCU-CHEK® Compact Plus Blood Glucose Meter (Roche, Indianapolis, IN) с помощью пункций, вводимых два раза в неделю в хвостовую вену мышей, достигших 10-недельного возраста. Считается, что у мышей имеется диабет, если уровень глюкозы в крови три дня подряд превышает 250 мг/дл. Мышей наблюдали в течение минимум 100 дней после начала обработки. На все эксперименты, проводимые на животных, было получено разрешение регионального Комитета по контролю и наблюдению за экспериментами на животных (IACUC).
Инъекции антител. Для исследований по предупреждению диабета, NOD-мышам с преддиабетом, достигшим 10-недельного возраста, внутрибрюшинно (i.p.) инъецировали 100 мкг антитела один раз в неделю в течение 6 недель. Для исследований по обратному развитию заболевания, животных произвольно распределяли по группам обработки, и в течение 1 недели после предположительного развития диабета у мышей, два раза в день измеряли уровень глюкозы в крови с интервалом по меньшей мере шесть часов между каждым измерением, и во время всего исследования мышам, у которых уровень глюкозы в крови превышал 250 мг/дл, вводили инсулин путем i.p. инъекции. Считалось, что у мышей в группах обработки, у которых сохранялась инсулин-независимость 30 дней подряд, наблюдается обратное развитие заболевания. У пяти мышей в каждой группе, в интервале между пятой и шестой обработками извлекали лимфоидные органы, брали кровь, а также извлекали костный мозг и поджелудочную железу для проведения клеточного анализа. По окончании исследования поджелудочную железу подвергали гистологическому и иммуногистохимическому анализу. Антитело против мышиного CXCR3, клон CXCR3-173 (Uppaluri et al. 2008), и контрольное антитело IgG хомячка закупали у BioLegend (San Diego, CA).
FACS-анализ. Были приготовлены моноклеточные суспензии селезенки, лимфоузлов паховой области, лимфоузлов поджелудочной железы и костного мозга. Поджелудочную железу разрезали на небольшие кусочки и инкубировали в 2 мг/мл коллагеназы D (Roche Diagnostics, Indianapolis, IN) в течение 30 минут при 37°C, фильтровали через клеточный фильтр 70 микрон (BD Biosciences, San Jose, CA), и лимфоциты выделяли из ткани поджелудочной железы путем центрифугирования в градиенте плотности. Клетки окрашивали АФК-меченным антителом против мышиного CD25, ФИТЦ-меченным антителом против мышиного CD4 и ФЭ-меченным антителом против мышиного Foxp3 (eBiosciences, San Diego, CA) для регуляторных T-клеток (T reg); PerCP-меченным антителом против мышиного CD8α, ФЭ-меченным антителом против CD44, АФК-меченным антителом против мышиного CD62L и PeCy7-меченным антителом против мышиного CXCR3 для активированных Т-клеток/Т-клеток памяти; и ФИТЦ-меченным антителом против мышиного CD94, PerCP-меченным антителом против мышиного CD4, АФК-меченным антителом против мышиного B220, PeCy7-меченным антителом против мышиного CD11c, антителом против мышиного CD11b, меченного темно-синим красителем, и ФЭ-меченным антителом против мышиного NKp46 для B-клеток, миелоидных клеток, дендритных клеток, NK-клеток и NKT-клеток. Эти клетки инкубировали в течение 30 минут при 4°C, а затем блокировали антителом против мышиного CD16/32 в течение 20 минут на льду. Для регуляторных T-клеток осуществляли поверхностное окрашивание, а затем клетки промывали, фиксировали и делали проницаемыми в буфере Cytofix/Perm (eBiosciences), после чего окрашивали антителом против мышиного Foxp3 в течение 30 минут на льду. После окрашивания, клетки два раза промывали, фиксировали в параформальдегиде и загружали в цитометр LSRII, и полученные данные анализировали с использованием компьютерной программы Flow Jo (Treestar, Ashland. OR). Если это не оговорено особо, все антитела закупали у BD Biosciences.
Анализ на хемотаксис. Анализ на хемотаксис осуществляли в 24-луночных планшетах (Costar), содержащих проницаемые носители Transwell с 5-мкм мембраной. CXCR3-трансфицированные клетки 300.19 помещали во вставки Transwell при плотности 1×106 клеток в 2,5% термоинактивированной фетальной бычьей сыворотке в среде RPM1640 (общий объем 0,2 мл). Отдельно взятую среду или среду, в которую были добавлены рекомбинантный хемокин в количестве 300 нг/мл CXCL9 (MIG), 100 нг/мл CXCL10 (IP-10) или 100 нг/мл CXCL11 (I-TAC), помещали в нижнее отделение (0,6 мл), и вставки Transwell, содержащие клетки, загружали в нижнее отделение. Планшеты инкубировали в течение 4-5 часов в инкубаторе с повышенной влажностью в атмосфере 5% CO2 при 37°C. После инкубирования, вставки Transwell вынимали, а затем всю среду в нижнем отделении объединяли, и клетки осаждали путем центрифугирования в течение 5 минут при 1200 об/мин. Затем среду отсасывали и клетки окрашивали кальцеином AM (конечный объем 10 мкг/мл) в течение 30 минут при 37°C. После этого клетки осаждали и промывали, добавляли среду (0,1 мл), и суспензию переносили в 96-луночные планшеты с черными стенками и с прозрачным дном. Планшеты центрифугировали при 1200 об/мин для осаждения клеток и измеряли флуоресценцию на длине волны 490/520 нм на устройстве Flexstation. Все эксперименты в соответствующих условиях проводили с тремя повторностями. Полученные данные выражали в средних относительных единицах флуоресценции (RFU) мигрировавших клеток. Смотрите фиг. 14A-C и фиг. 15A-C.
Для осуществления хемотаксиса с блокадой CXCR3, CXCR3-трансфицированные клетки 300.19 предварительно обрабатывали различными количествами блокирующего антитела или контрольного IgG в течение 20-30 минут при 37°C, а затем эти клетки использовали для анализа не хемотаксис. Антитело не промывали, и в таком виде использовали в анализе с инкубированием.
Анализ на мобилизацию кальция, проводимый с использованием FLIPR. Клетки почек человеческого эмбриона 293 (HEK), экспрессирующие HCXCR3, собирали при конфлюэнтности 80% путем обработки PBS+2 мМ ЭДТА. Эти клетки суспендировали в бессывороточной среде HEK-SFM при плотности 1×106 клеток/мл. 15 мкл (15000 клеток) полученной суспензии распределяли по всем лункам 384-луночного планшета. Клетки нагружали красителем в течение 30 минут при комнатной температуре путем добавления 15 мкл разведенного кальций-чувствительного красителя 4 на флуориметре FLIPR. Клоны антитела против человеческого CXCR3 и клоны антитела контрольного изотипа серийно разводили (3-кратно) в HBSS+20 мМ HEPES+1% BSA с получением 10 тестируемых концентраций на клон. Каждую тестируемую концентрацию анализировали с двумя повторностями (n=2) на одном и том же планшете. К клеткам, содержащимся в каждой лунке, добавляли тестируемую 15-мкл концентрацию, и планшет инкубировали при комнатной температуре в течение 1 часа. На устройство FLIPR, содержащееся в каждой лунке, добавляли фиксированную концентрацию CXCL11 (R & D Systems), представляющую EC80 для мобилизации внутриклеточного кальция, и проводили мониторинг изменения флуоресценции в течение определенного периода времени. Максимальный ответ для каждой лунки нормализовали по базовой линии, и данные регистрировали после усреднения по 4-параметрическому уравнению с использованием компьютерной программы GraphPad Prism, а затем определяли IC50 для каждого клона. Смотрите фиг. 14D и фиг. 21.
Анализ Biacore на аффинность. Приготовление поверхности для анализа Biacore. Аффинности связывания клонов 4, 12, 53, 82 и 135 гибридом, содержащих мышиное антитело против человеческого CXCR3, с человеческим пептидом CXCR3 вычисляли с помощью анализа на кинетику/аффинность Biacore T100. На сенсорный чип Biacore CMS Series S (GE #BR-1006-68) иммобилизовали антитело для захвата, а именно, кроличье антитело против мышиного Fc (RAM-Fc) (GE# BR-1008-38) в соответствии со стандартной программой связывания с амином. Поверхность чипа, покрытого карбоксиметилдекстраном, активировали смесью, 1:1, 0,1 M N-гидроксисукцинимида (NHS) и 0,4 M гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) для связывания поверхности с реакционноспособными аминогруппами на антителе для захвата. После иммобилизации антитела реакционноспособную поверхность сенсорного чипа гасили 1 M гидрохлорида этаноламина/NaOH, pH 8,5. После иммобилизации получали 8000 RU антитела RAM-Fc для захвата, находящегося на одной проточной кювете. Другая контрольная проточная кювета была использована в качестве поверхности для вычитания базового значения во время анализа данных.
Условия проведения анализа Biacore. Температуру в камере для образца на приборе Biacore T100 и температуру при проведении анализа устанавливали на 4°C и 25°C, соответственно. Мышиные анти-hCXCR3 антитела разводили до 500 нМ в рабочем HBS-EP+-буфере (10 мМ HEPES, 150 мМ NaCl, 0,05% поверхностно-активное вещество P20, 3 мМ ЭДТА, pH 7,4), и иммобилизовали путем проведения тридцати двух впрысков со скоростью 10 мкл/мин. В этих условиях достигался стабильный захват приблизительно 1200 RU каждого тестируемого клона мышиного анти-hCXCR3 антитела. Пептиды hCXCR3 разводили до концентраций 200, 100, 50, 25, 12,5 и 0 нМ в HBS-EP+. В каждом цикле анализа пептид впрыскивали в течение пяти минут со скоростью потока 50 мкл/мин для измерения степени ассоциации, а затем промывали в HBS-EP+ в течение десяти минут со скоростью потока 50 мкл/мин для измерения степени диссоциации. Поверхность захвата (RAM-Fc) регенерировали между циклами анализа с использованием 10 мМ глицина-HCl, pH 1,7, со скоростью потока 50 мкл/мин в течение трех минут. Анализ проводили с помощью компьютерной программы Biacore T100 Kinetics Evaluation v2.0 (GE Healthcare). Затем строили сенсорограммы для 1:1-модели связывания с эталонной проточной кюветой и с вычитанием концентрации 0 нМ (двойного вычитания фона).
Анализ Biacore для целого рецептора. Полноразмерный белок рецептора человеческого CXCR3 с C-концевой 6× His (SEQ ID NO:82) и HPC4-меткой экспрессировали в клетках Sf9 насекомых с использованием бакуловирусного вектора. Затем белок рецептора очищали путем аффинной очистки на Ni-NTA и HPC4. Конечный продукт подвергали буферному обмену на 10 мМ HEPES, 300 мМ NaCl, 0,5% н-додецил-β-D-мальтопиранозид и 5% глицерин. Белок рецептора иммобилизовали на NTA-чипах посредством образования хелатного комплекса с Ni, а затем стабилизировали посредством связывания с амином с использованием смеси 1:1 0,1 M N-гидроксисукцинимида (NHS) и 0,4 М гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC), разведенной 1:10. Стабилизированную поверхность чипа с рецептором тестировали на активность связывания с лигандом путем впрыска 20 нМ лигандов hCXCL10 и hCXCL11. Для кинетического анализа, лиганды человеческого CXCR3 (hCXCL9, hCXCL10 и hCXCL11) разводили до концентраций 20, 10, 5, 2,5, 1,25, 0,6125, 0,3125 и 0 нМ в HBS-EP+. Анти-CXCR3 антитела разводили до концентраций 80, 40, 20, 10, 5, 2,5, 1,25 и 0 нМ в HBS-EP+. Аналит впрыскивали в течение пяти минут со скоростью потока 50 мкл/мин для измерения степени ассоциации, а затем промывали в HBS-EP+ в течение десяти минут со скоростью потока 50 мкл/мин для измерения степени диссоциации. Поверхность чипа с рецептором регенерировали между циклами анализа с использованием 10 мМ глицина-HCl, pH 1,7, со скоростью потока 50 мкл/мин в течение 1 минуты. Анализ проводили с помощью компьютерной программы Biacore T100 Kinetics Evaluation v2.0 (GE Healthcare). Затем строили сенсорограммы для 1:1-модели связывания с эталонной проточной кюветой и с вычитанием концентрации 0 нМ (двойного вычитания фона).
Тест на толерантность к глюкозе. Вечером перед введением глюкозы оценивали уровень глюкозы в крови после кормления, но животным, страдающим диабетом, инсулин не вводили. Мышей содержали в режиме голодания в течение 12 часов, после чего вводили i.p. инъекцию D-глюкозы (20%; Sigma) при 2 мг/г массы тела. Уровень глюкозы в крови измеряли до инъекции и через 15, 30, 60 и 120 минут после инъекции.
Пример 2: Характеризация мышей NOD
Репрезентативные срезы поджелудочной железы, взятой у 6-10-недельных самок NOD-мышей с преддиабетом и у самок NOD-мышей с впервые выявленным диабетом, заливали в парафин и окрашивали на инсулин, CXCL10 и T-клеточный маркер CD3. Фиг. 1. Экспрессию CXCL10 детектировали в поджелудочной железе NOD-мышей в островках, окруженных инфильтрирующими клетками (показано стрелками в центральном столбце на фиг. 1). У более старых мышей с преддиабетом и у мышей с впервые выявленным диабетом наблюдалось заметное повышение уровня инфильтрации островковых Т-клеток (фиг. 1, правый столбец) и снижение уровня продуцирования инсулина в указанных островках (фиг. 1, левый столбец).
Для того, чтобы определить, присутствуют ли CXCR3+-T-клетки в поджелудочной железе NOD-мышей, был проведен проточный цитометрический анализ тканей поджелудочной железы, собранных у самок NOD-мышей с впервые выявленным диабетом. Фиг.2.
Экспрессию CXCR3 оценивали на CD4+/TCR+- и CD8+/TCR+-T-клетках. Окрашивание антителом контрольного изотипа проиллюстрировано на затененной кривой. Проточную цитометрию проводили на моноклеточных суспензиях пула тканей поджелудочной железы, собранных у нескольких мышей. Полученные данные показали, что в поджелудочной железе NOD-мышей присутствуют цитотоксические и хелперные CXCR3+-T-клетки.
Пример 3: Профилактическая обработка NOD-мышей анти-CXCR3 антителом
Самок NOD-мышей с преддиабетом, достигших 10-недельного возраста, обрабатывали один раз в неделю в течение шести недель 100 мкг антитела хомячка против мышиного CXCR3 (клона CXCR3-173, закупленного у BioLegend, San Diego, CA) или контрольным IgG хомячка или PBS. Мониторинг уровней глюкозы в крови проводили два раза в неделю, и животные, у которых уровень глюкозы в крови превышал 250 мг/дл три дня подряд, рассматривались как животные с диабетом, и этих животных подвергали эвтаназии. На фиг. 3 представлены проценты мышей, у которых развивался диабет в течение определенного периода времени в каждой группе обработки. Каждая линия представляет объединенные результаты для десяти мышей на группу. Представлены результаты двух независимых исследований (фиг. 3A и 3B). На этих графиках показано, что профилактическая обработка анти-CXCR3 антителом способствует предупреждению развития диабета у самок NOD-мышей с преддиабетом.
Для дополнительной оценки эффекта профилактического введения анти-CXCR3 антитела, репрезентативные срезы поджелудочной железы, взятой у самок NOD-мышей, обработанных анти-CXCR3 антителом, окрашивали на инсулин (фиг. 4, левая панель), CD3 и Foxp3 (фиг. 4, центральная и правая панели). На правой панели фиг. 4 представлено увеличенное изображение среза, проиллюстрированного на центральной панели. Ткань поджелудочной железы брали у мышей по окончании исследования (у мышей, достигших возраста 26 недель). На фиг. 4 продемонстрировано, что инсулинположительные островки присутствуют у NOD-мышей, обработанных анти-CXCR3 антителом, при этом большинство Т-клеток окружено островками, но не содержится в этих островках.
Пример 4: Обратное развитие впервые выявленного диабета у NOD-мышей
Самки мышей, у которых уровень глюкозы в крови превышал 250 мг/дл три дня подряд, рассматривались как животные с диабетом, и этих мышей произвольно распределяли на группы обработки. Обработку начинали в первую неделю после распределения на группы. Мышей обрабатывали PBS, антителом против мышиного CXCR3 (100 мкг антитела, клона CXCR3-173, вводимого внутрибрюшинно) или контрольным IgG (100 мкг IgG, вводимого i.p.) один раз в неделю в течение шести недель, или мышиным антителом против глобулина тимоцитов (mATG; 500 мкг, вводимого i.p.) на дни 0 и 4. После распределения на группы, уровень глюкозы в крови измеряли утром и днем (с перерывами по меньшей мере в шесть часов), и вводили i.p. инсулин только тем мышам, у которых уровень глюкозы в крови превышал 250 мг/дл. Для отдельных мышей представлены ежедневные утренние уровни глюкозы в крови (фиг. 5). Данные, полученные в четырех независимых исследованиях по обратному развитию заболевания, объединяли для 8-10 мышей на группу в каждом исследовании. Полученные данные продемонстрировали обратное развитие впервые выявленного диабета у NOD-мышей после обработки этих мышей анти-CXCR3 антителом.
Для оценки изменений в Т-клеточных субпопуляциях поджелудочной железы мышей, обработанных анти-CXCR3 антителом, моноклеточные суспензии поджелудочной железы, взятой у четырех мышей на группу обработки (мышей, обработанных PBS, антителом против мышиного CXCR3, контрольным IgG или mATG), объединяли, окрашивали на Т-клетки и анализировали с помощью проточной цитометрии. Ткань поджелудочной железы брали через несколько дней после пятой обработки мышей дозами PBS, антитела против мышиного CXCR3 или контрольного IgG, и у mATG-обработанных мышей того же возраста. Процент CD4+- и CD8+-T-клеток в суспензиях представлен на фиг. 6A. На фиг. 6B проиллюстрирована экспрессия CD44 и CD82L на CD4+-T-клетках в поджелудочной железе, взятой у мышей, обработанных PBS (слева), контрольным IgG (в середине) и антителом против мышиного CXCR3 (справа). Процент клеток в гейте 1 (G1; CD44hiCD62Llo) указан для каждой группы обработки (67,3% для обработки PBS, 67,1% для обработки контрольным IgG и 30,4% для обработки анти-CXCR3 антителом). На фиг. 6C представлен график экспрессии CXCR3 на CD4+-T-клетках в гейте 1 (G1) или в гейте 2 (G2), как указано на фиг. 6B, по сравнению с экспрессией в клетках, окрашенных антителом контрольного изотипа и гейтированных на лимфоцитах.
Для того, чтобы определить, присутствуют ли инсулинположительные островки у NOD-мышей, у которых наблюдалось обратное развитие заболевания после их обработки анти-CXCR3 антителом, приготавливали залитые в парафин срезы поджелудочной железы самок NOD-мышей, обработанных контрольным IgG (левые панели), антителом против мышиного CXCR3 (средние панели) или мышиным ATG (правые панели), и эти срезы окрашивали на инсулин (верхний ряд) или на CD3 и Foxp3 (нижний ряд). Смотрите фиг. 7. По окончании исследования, у мышей брали ткань поджелудочной железы (приблизительно через 100 дней после распределения на группы). Окрашенные срезы продемонстрировали, что инсулинположительные островки присутствуют у NOD-мышей, у которых наблюдалось обратное развитие заболевания после их обработки анти-CXCR3 антителом, и что Т-клетки окружали островки, а некоторые Т-клетки содержались в этих островках.
Для оценки ответа на введение глюкозы проводили тест на толерантность к глюкозе. Смотрите фиг. 8. Самок NOD-мышей соответствующего возраста, у которых отсутствовал диабет (фиг. 8A), NOD-мышей с диабетом, обработанных PBS (фиг. 8B), NOD-мышей с диабетом, у которых наблюдалось обратное развитие заболевания после их обработки анти-CXCR3 антителом (фиг. 8C), и NOD-мышей с диабетом, обработанных IgG (фиг. 8D), выдерживали в течение ночи в условиях голодания и вводили глюкозу посредством i.p.-инъекции. Уровень глюкозы в крови измеряли до введения глюкозы (время 0) и после введения глюкозы в указанные периоды времени. Были представлены репрезентативные данные, полученные для 4-5 мышей на группу обработки. Полученные данные показали, что обработка анти-CXCR3 антителом приводила к повышению толерантности к глюкозе натощак через 100 дней после включения в группы обработки.
Пример 5: Адоптивный перенос Т-клеток
Для оценки способности T-клеток NOD-мышей, которые были обработаны анти-CXCR3 антителом (клоном CXCR3-173) и у которых наблюдалась ремиссия заболевания, индуцировать диабет у животных-реципиентов, выделенные CD4+- и CD8+-T-клетки подвергали адоптивному переносу мышам-реципиентам NOD.scid (у мышей с диабетом, не страдающих ожирением, и у мышей с тяжелым комбинированным иммунодефицитом) путем внутривенной инъекции. На фиг.9 представлены проценты мышей, не страдающих диабетом, в течение определенного периода времени после адоптивного переноса выделенных CD4+- и CD8+-T-клеток у мышей с диабетом, обработанных PBS или контрольным IgG, или у мышей, у которых наблюдалась ремиссия заболевания после обработки мышиным ATG или антителом против мышиного CXCR3. CD4+- и CD8+-T-клетки выделяли из селезенки, лимфоузлов поджелудочной железы и лимфоузлов паховой области, взятых через 80-90 дней после распределения самок NOD-мышей по различным группам обработки. CD4+- и CD8+-T-клетки объединяли и 8 миллионов клеток подвергали адоптивному переносу реципиентам NOD.Scid, после чего проводили мониторинг развития диабета посредством измерения уровней глюкозы в крови два раза в неделю. На фиг. 9, каждая линия представляет объединенные данные для пяти мышей на группу. Два репрезентативных исследований проиллюстрированы на фиг. 9A и 9B. Выделенные Т-клетки, взятые у мышей, обработанных анти-CXCR3 антителом, обнаруживали замедленную передачу заболевания.
Были дополнительно охарактеризованы выделенные донорные T-клетки. На фиг. 10A указан общий процент CD4+- и CD8+-T-клеток (левая панель) среди донорных Т-клеток, выделенных у мышей, обработанных PBS, антителом против мышиного CXCR3, контрольным IgG или мышиным ATG, как было описано в предыдущем параграфе. На правой панели фиг. 10A указан процент эффекторных клеток и центральных Т-клеток памяти в субпопуляции Т-клеток, представляющих собой CD4+-клетки (верхняя панель) и CD8+-T-клетки (нижняя панель) для каждой суспензии донорных клеток, как было определено по экспрессии CD44 и CD62L. Выделенные пулы CD4+- и CD8+-T-клеток окрашивали на экспрессию CD44 и CD62L до их переноса, загружали в проточный цитометр и анализировали. Также оценивали процент регуляторных Т-клеток в пулах выделенных донорных Т-клеток. На фиг. 10B указан процент регуляторных Т-клеток для каждой группы обработки, определенный по экспрессии CD4 и CD25 или по экспрессии CD4, CD25 и внутриклеточного Foxp3. На фиг. 10C указан процент CD8+-T-клеток (левая панель) и CD4+- Т-клеток (правая панель) в донорных клетках, которые также экспрессировали CXCR3. Эти данные указывали на снижение процента CXCR3+-T-клеток среди донорных клеток у мышей, у которых наблюдалось обратное развитие заболевания после их обработки анти-CXCR3 антителом.
Эффективность CXCR3-обработки после адоптивного переноса донорных CD8+-T-клеток OT-1 оценивали с использованием модели RIP-OVA с диабетом типа 1. Мышами RIP-OVA являются трансгенные мыши, у которых трансген, кодирующий белок овальбумин (OVA), индуцируемый промотором крысиного инсулина (RIP), был введен в мышиный геном, в результате чего в этих мышей экспрессировалась мембранная форма овальбумина в островковых β-клетках. У мышей исходной линии C57BL/6 и у мышей RIP-OVA не наблюдалось спонтанного развития диабета. Мыши RIP-OVA, также обозначаемые C57BL/6-Tg(Ins2-TFRC/OVA)296Wehi/WehiJ, были закуплены в лаборатории Jackson Laboratory. Развитие диабета у этих мышей наблюдалось после адоптивного переноса овальбумин-специфических CD8+-T-клеток, выделенных у трансгенных по TCR (Т-клеточному рецептору) мышей OT-1 (Kurts et al., J. Exp. Med. 184: 923-930), которые были закуплены в лаборатории Jackson Laboratory. Мыши OT-1 содержали трансгенные вставки мышиных генов TCRa-V2 и TCRb-V5 (Hogquist et al. Cell 76:17-27). Трансгенный TCR распознает остатки овальбумина в окружении белков MHCI H2Kb. У мышей ОТ-1, более, чем 95% CD8+-T-клеток экспрессируют TCR, распознают пептид овальбумина и активируются этим пептидом.
На фиг. 11 указан процент мышей, не страдающих диабетом, в течение определенного периода времени после адоптивного переноса CD8+-T-клеток OT-1 мышам-реципиентам RIP-OVA, которые либо не были обработаны, либо были обработаны антителом против мышиного CXCR3, либо были обработаны контрольным IgG. Обработку (100 мкг i.p.) начинали в день 1 (исследования 1 и 2) или на 7-ой день (исследование 2) после адоптивного переноса, и такую обработку проводили два раза в неделю в течение 3 недель. Каждая линия представляет собой объединенные данные, полученные для пяти мышей на группу. Результаты этих двух исследований представлены на фиг. 11A и 11B. Полученные данные показали, что обработка анти-CXCR3 антителом приводила к предотвращению развития диабета у мышей с моделью RIP-OVA.
На фиг. 12 также представлены данные, характеризующие эффективность различных обработок после адоптивного переноса Т-клеток OT-1 мышам-реципиентам RIP-OVA. На фиг. 12A проиллюстрирована экспрессия CXCR3 на донорных Т-клетках, проанализированных с помощью проточной цитометрии до их адоптивного переноса реципиентам RIP-OVA. Окрашивание антителом контрольного изотипа проиллюстрировано на затененной кривой. На фиг. 12B указан процент донорных клеток в крови, селезенке и в лимфоузлах поджелудочной железы (pLN) в указанные периоды времени после адоптивного переноса Т-клеток OT-1 мышам-реципиентам RIP-OVA, обработанным антителом против мышиного CXCR3 или контрольным IgG. Обработку антителом (100 мкг i.p.) начинали в день 1 после переноса Т-клеток и проводили два раза в неделю в течение двух недель. Каждая точка представляет собой данные для одной отдельной мыши. На фиг. 12C указано число донорных клеток в крови, селезенке и в лимфоузлах поджелудочной железы (pLN), которые пролифирировались в ответ на стимуляцию аутоантигеном (OVA) у мышей-реципиентов RIP-OVA, обработанных антителом против мышиного CXCR3 или контрольным IgG в определенные периоды времени после адоптивного переноса. Обработку антителом (100 мкг i.p.) начинали в день 1 после переноса Т-клеток OT-1 и проводили два раза в неделю в течение двух недель. Каждая точка представляет собой данные для одной отдельной мыши. На фиг. 12D указан процент CXCR3-экспрессирующих донорных клеток в крови, селезенке и в лимфоузлах поджелудочной железы (pLN) в определенные периоды времени после адоптивного переноса Т-клеток ОТ-1 мышам-реципиентам RIP-OVA, обработанным антителом против мышиного CXCR3 или контрольным IgG. Обработку антителом (100 мкг i.p.) начинали в день 1 после переноса Т-клеток OT-1 и проводили два раза в неделю в течение двух недель. Каждая точка представляет собой данные для одной отдельной мыши. Обработка анти-CXCR3 антителом приводила к снижению процента CXCR3+-T-клеток у мышей RIP-OVA.
На фиг. 13 показаны репрезентативные залитые в парафин срезы поджелудочной железы, выделенной у необработанных мышей-реципиентов RIP-OVA, где указанные срезы окрашивали на инсулин (фиг. 13A) или на CD3 (фиг. 13B), или у мышей, обработанных антителом против мышиного CXCR3, где указанные срезы окрашивали на инсулин (фиг. 13С) или на CD3 (фиг. 13D). Обработку анти-CXCR3 антителом (100 мкг i.p.) начинали в день 1 после переноса Т-клеток ОТ-1 и проводили два раза в неделю в течение 3 недель. Ткань поджелудочной железы брали у мышей по окончании исследования (приблизительно через 60 дней после переноса Т-клеток). Срезы продемонстрировали отсутствие инфильтрации Т-клеток у мышей RIP-OVA, обработанных антителом против мышиного CXCR3.
Пример 6: Оценка клонов антитела против человеческого CXCR3
Клоны антитела против человеческого CXCR3 Cl 4, 12, 53, 82, и 135 оценивали на их влияние на хемотаксис CXCR3 и мобилизацию кальция с применением методов, описанных выше в разделе «Материалы и методы» (пример 1). Для анализа на хемотаксис, клетки 300.19, трансфицированные человеческим CXCR3, предварительно инкубировали только в одной среде или в среде, содержащей различные концентрации антитела, как показано на фиг. 14A-C, а затем эти клетки использовали в анализе на хемотаксис. На фиг. 14A-C показано, что CXCR3-опосредуемый хемотаксис в CXCL11 ингибировался клонами Cl 4, 12, 53, 82 и 135. Клон 2 на фиг.14 идентичен клону 4.
Для анализа на мобилизацию кальция, клетки HEK, трансфицированные человеческим CXCR3, предварительно инкубировали с различными концентрациями антитела, а затем переносили в FLIPR методами, описанными выше в разделе «Материалы и методы» (пример 1). Концентрация антитела, необходимая для ингибирования мобилизации кальция на 50% для каждого антитела, указана на фиг. 14D. На фиг. 14D показано, что CXCR3-опосредуемая мобилизация кальция в CXCL11 ингибировалась клонами Cl 4, 12, 53 и 135.
Для дополнительной оценки влияния клонов 4, 12, 53, 82 и 135 на хемотаксис, hCXCRS-трансфицированные клетки 300.19 предварительно инкубировали только в одной среде или в среде, содержащей 50 мкг/мл антитела, а затем их использовали в анализе на хемотаксис и анализировали на миграцию в CXCL9 (фиг. 15A), CXCL10 (фиг. 15B) и CXCL11 (фиг. 15C). Полученные данные продемонстрировали, что клоны 4, 12, 53, 82 и 135 ингибируют миграцию в CXCL10 и CXCL11 и частично ингибируют миграцию в CXCL9.
Для оценки специфичности клонов 4, 12, 53, 82 и 135, антитела анализировали на связывание с другими хемокиновыми рецепторами. Клетки 300.19 трансфицировали человеческими CXCR1, CXCR5, CXCR2, CXCR4 или CCR5, и связывание с антителом анализировали путем инкубирования с клонами антитела против человеческого CXCR3, с последующим окрашиванием «вторым» антителом и проведением проточной цитометрии. Введение только «второго» антитела служило в качестве негативного контроля. Клетки 300.19, трансфицированные человеческим CXCR3, служили в качестве позитивного контроля на окрашивание этими клонами. На фиг.16 представлены гистограммы связывания антитела с клетками, экспрессирующими различные хемокиновые рецепторы, и эти гистограммы показали, что клоны 4, 12, 53, 82 и 135 не связываются с другими хемокиновыми рецепторами и являются специфичными к CXCR3.
В анализе на связывание с хемокиновым рецептором проводили стандартные процедуры, применяемые в проточной цитометрии. Вкратце, клеточные линии собирали путем обработки версеном, и каждую клеточную линию разделяли на семь образцов. Каждый образец инкубировали на льду с одним «первым» антителом (5 мкг/мл), а затем окрашивали ФИТЦ-конъюгированным «вторым» антителом для детектирования связанного «первого» антитела. Клетки, используемые в качестве негативного контроля, окрашивали только «вторым» антителом (инкубирование с «первым» антителом не проводили). «Первое» антитело представляет собой клон антитела против человеческого CXCR3 или клон контрольного антитела против человеческого CXCR3 1C6. После окрашивания, клетки загружали на проточный цитометр, и данные анализировали с использованием компьютерной программы FlowJo. Каждая линия на фиг.16 представляет отдельный образец клеток, окрашенных одним «первым» антителом и «вторым» антителом, или только «вторым» антителом.
Аффинность (Ka) и скорость диссоциации (Kd) для клонов 4, 12, 53, 82 и 135 анализировали с помощью анализа Biacore методами, описанными выше (пример 1). Результаты систематизированы ниже в таблице 4.
Пример 7: Картирование эпитопа
Усеченные биотинилированные человеческие пептиды CXCR3 (N-концевые фрагменты из 16 аминокислот) N-концевого внеклеточного домена CXCR3 использовали в целях определения эпитопов для клонов 4, 12, 53, 82 и 135. Была получена серия фрагментов с аланиновыми заменами (смотрите ниже таблицу 5, где аланиновая замена показана жирным шрифтом), и эти фрагменты были биотинилированы. Картирование эпитопов оценивали в анализах Octet® (ForteBio, Menlo Park, CA) и Biacore™ (GE Healthcare).
Для проведения анализа Octet, пептиды ресуспендировали в 80% ДМСО и разводили до 10 мкг/мл в PBS. Клоны антител 4, 12, 53, 82, 135 и коммерчески доступный клон 1C6 (BD Biosciences) разводили до 120 нМ в PBS. Кинетический анализ осуществляли в 96-луночном планшете при 300 мкл/лунку на системе Octet QK (ForteBio). Каждый аналитический планшет содержал биотинилированный у N-конца полноразмерный пептид HCXCR3 ECD дикого типа (Abgent), используемый в качестве позитивного контроля, а также контрольный PBS-буфер для вычитания фона. Стрептавидиновые биосенсоры Octet (ForteBio) предварительно смачивали в PBS в течение по меньшей мере пяти минут, а затем использовали для проведения анализа. Биосенсоры сначала погружали в PBS на пять минут без встряхивания для получения базовых значений. Во всех остальных стадиях, скорость вращения составляла 1000 об/мин. Биосенсоры погружали в пептидные растворы на пять минут для загрузки пептидов. Затем проводили другую основную стадию в PBS в течение пяти минут. Затем биосенсоры погружали в растворы антител на десять минут для оценки ассоциации. И наконец, биосенсоры переносили в PBS на пятнадцать минут для оценки диссоциации. Сенсорограммы анализировали с помощью анализа данных Octet Data Analysis v7.0. Активность связывания выражали как процент максимального уровня ответа каждого антитела по отношению к уровню ответа полноразмерного пептида hCXCRS дикого типа.
Относительные уровни ответов регистрировали на тепловой карте эпитопа. Максимальные ответы на сенсорограмме по сравнению с ответами пептида hCXCRS ECD дикого типа составляли в пределах 4-8 нм. Каждый скринированный клон имел уникальный эпитоп. Ни один из тестированных мутантов полностью не блокировал связывание с клоном 1C6. Валиновый остаток в положении 10 и аспартат в положении 13 играют определенную роль в связывании всех скринируемых антител. Антитела 12 и 1C6 имели наибольшее число N-концевых эпитопов, причем, мутации валина в положении 5 оказывали влияние на активность обоих антител. Антитело 82 имело наибольшее число С-концевых эпитопов и обнаруживало снижение активности связывания, начиная с глутамина в положении 9. Исходя из этих данных были установлены аминокислотные границы эпитопов на последовательности CXCR3 для каждого антитела; Cl 4: аминокислоты 7-13; Cl 12: аминокислоты 5-13; Cl 53: аминокислоты 7-13; Cl 82: аминокислоты 9-15; Cl 135: аминокислоты 7-13; и клон 1C6: аминокислоты 5-13.
Для проведения анализа Biacore, пептиды разводили до 10 нг/мл в рабочем HBS-EP+-буфере (10 мМ HEPES, 150 мМ NaCl, 3 мМ ЭДТА, 0,005% полисорбат 20). С использованием Biacore T100™, пептиды впрыскивали поверх чипов CM5-SA (GE #BR-1005-31) со скоростью 5 мкл/мин до тех пор, пока на был достигнут стабильный захват 20 единиц отклика (RU) на проточную кювету. Проточная кювета 1 служила в качестве контроля для вычитания фона на каждом чипе. Пептид hCXCR3 дикого типа из 37 аминокислот наносили на одну проточную кювету для каждого чипа и использовали в качестве позитивного контроля. Мышиные анти-hCXCR3 антитела 4, 12, 53, 82 и 135 разводили до концентраций 50, 25, 12,5, 6,25 и 3,125 нМ в HBS-EP+. В каждом цикле анализа, антитела впрыскивали в течение трех минут со скоростью потока 50 мкл/мин для измерения степени ассоциации, а затем промывали буфером в течение трех минут со скоростью потока 50 мкл/мин для измерения степени диссоциации. Поверхность чипа с пептидом регенерировали между циклами анализа с использованием 10 мМ глицина-HCl, pH 2,0, со скоростью потока 50 мкл/мин в течение 60 секунд. Затем строили сенсорограммы для 1:1-модели связывания и анализировали с двойным вычитанием с помощью компьютерной программы BiaEvaluation v2.0.1, после чего пептиды иммобилизовали на стрептавидиновых биосенсорах. Типичные уровни ответов составляли 0-500 RU, а также были определены граничные значения для ответов на «сильное», «умеренное» и «слабое» связывание. Относительные уровни ответов регистрировали, и по этим данным строили эпитопную карту. Затем определяли скорости диссоциации, а также регистрировали пептиды, которые давали высокие значения Kd (более чем 0,001 с-1).
Все антитела связывались с hCXCR3 дикого типа, и это связывание происходило в первых 16 аминокислотах последовательности hCXCR3. Данные по связыванию представлены в таблице 6 для клонов 4, 12, 53, 82 и 135, а соответствующая карта, на которой указаны границы минимального эпитопа, необходимого для сообщения активности связывания с каждым клоном антитела, приводится на фиг. 18, где остатки, играющие важную роль в таком связывании, обозначены X. Все эпитопы для антител картированы в человеческой последовательности CXCR3 в остатках 6-15, то есть в области, участвующей в связывании с CXCL10 и CXCL11. Аминокислоты в эпитопе были определены путем детектирования снижения ответной реакции связывания после замены аланина в данном положении пептидного фрагмента CXCR3. Клоны 53 и 135 имели наибольшее число аналогичных эпитопов. Клоны 4 и 12 имели наибольшее число N-концевых эпитопов (сила связывания изменялась после замены валином в положении 5). Клон 82 имел наибольшее число С-концевых эпитопов, причем снижение активности связывания наблюдалось сразу после замены глутамином в положении 9. Аспартат в положении 7 в CXCR3 необходим для каждого клона, за исключением клона 82. Валин в положении 10 и аспартат в положении 13 в CXCR3 играют определенную роль в связывании всех этих клонов. Какого-либо различия в эпитопах для мышиных, химерных и гуманизированных вариантов этих пяти клонов не наблюдалось.
(SEQ ID NO:99-114, соответственно, для иллюстрации)
Примечания:
+++ Сильное ответное связывание (>300 RU)
++ Умеренное ответное связывание (150-300 RU)
+ Слабое ответное связывание (10-150 RU)
- Связывание отсутствует (<10 RU)
* Высокая скорость диссоциации (Kd>0,001), несмотря на более сильный связывающий ответ (>10 RU)
Пример 8: Гуманизация клонов антител против человеческих CXCR3
Для каждого из пяти клонов анти-CXCR3 антител (клонов 4, 12, 53, 82 и 135) было получено четыре варианта (химерный вариант, гуманизированные варианты 1 (Hu1), Hu2 и Hu3). Смотрите фиг. 20. Все химерные варианты были продуцированы посредством «экспрессии пулов» в исследованиях in vivo на животных-моделях. Клетки CHO-DXB11 (Urlaub and Chasin, Proc. Natl. Acad, Sci., 77:4218-20, (1980)), которые были адаптированы для суспензионной культуры, подвергали электропорации с использованием экспрессионных векторов, содержащих тяжелые и легкие цепи клонов химерных анти-CXCR3 антител 4, 12, 53, 82 и 135. Эти экспрессионные векторы также содержали ген дигидрофолат-редуктазы для отбора стабильных трансфектантов CHO. После отбора на 100 нМ метотрексата, стабильно трансфецированные пулы CHO использовали для биопродуцирования культур в шейкерных колбах со средой OptiCHO. Рекомбинантное химерное антитело очищали от кондиционированной среды с помощью стандартной хроматографии на белке А. Клоны Hu1 подвергали CDR-прививке на человеческих каркасных областях, и такие клоны были полностью гуманизированными, за исключением аминокислот мышиных CDR и любых остатков в положении Верньера. Клоны Hu2 представляли собой клоны с «удлиненными CDR», где указанные клоны были получены посредством обратных замен в последовательностях Hu1 в остатках, расположенных в пределах четырех аминокислот мышиных CDR. Клоны Hu3, по сравнению с последовательностями Hu2, включали дополнительные обратные мутации в положениях, идентифицированных как «очень отличающиеся» положения мышиных и человеческих антител, полученных путем моделирования на основе IMGT. В целом, были транзиентно экспрессированы двадцать вариантов в клетках CHO-NRC, и эти варианты были очищены для проведения анализов in vitro.
Клоны 4 и 53 подвергали дополнительной гуманизации. Тяжелые цепи 4.7-4.11 включали дополнительные гуманизированные обратные мутации, находящиеся за пределами CDR, а также модификации, вводимые в целях удаления сайта дезамидирования в положениях 58 и 59 (в соответствии с нумерацией IMGT) в CDR2 VH для повышения стабильности клона. В частности, сайт дезамидирования в VH 4.2 был заменен различными альтернативными остатками. Клоны VH 53.4-53.6 и VL 53.4-53.9 включали дополнительные гуманизированные обратные мутации, находящиеся за пределами CDR, которые были, в частности, введены для придания этим цепям большего сходства с цепями VH 4.1 и VL 4.1.
Пример 9: Кинетика связывания с целым рецептором
Кинетику связывания двадцати вариантов анти-CXCR3 антител оценивали с помощью анализов Octet® и Biacore™, проводимый с использованием интактных пептидов CXCR3. Анализ Octet проводили, как описано выше. В анализе Biacore проводили три стадии колоночной очистки, где очищенные человеческие пептиды CXCR3 дикого типа имели C-концевые His- и HPC4-метки, и эти пептиды иммобилизовали на NTA-чипах посредством реакции образования хелатного комплекса с никелем и связывания с амином. Средние величины RU (приблизительно 1200 RU) на чипах были использованы для улучшения качества данных и для минимизации эффекта двухвалентного связывания. 0-80 нМ образцов антител впрыскивали поверх рецептора. Стандартный оценочный анализ Biacore™ проводили с использованием сенсорограмм с подбором локальных данных Rmax для облегчения построения кривой. Были оценены кривые связывания для четырех вариантов (химерного варианта (Ch) и гуманизированных вариантов (Hu) 1-3) пяти клонов (4, 12, 53, 82 и 135). Для сравнения также оценивали кинетику связывания с человеческими лигандами CXCR3, а именно с CXCL9, CXCL10 и CXCL11, и с мышиным CXCL11 (mCXCL11). Результаты представлены ниже в таблицах 7A (систематизированные по KD) и 7B (систематизированные по Kd). В этих таблицах выделены четыре варианта с наилучшими KD и Kd. В случае гуманизированных вариантов анти-hCXCR3 mAb, систематизированных по скорости диссоциации (Kd), большинство этих вариантов, обозначенных по меньшей мере цифрой 1, имели более низкую скорость диссоциации, чем наиболее активный человеческий лиганд CXCR3, а именно, hCXCL11.
Как показано в таблице 7A, четырьмя вариантами с наилучшими KD являются химерный клон 4 (высокая скорость ассоциации, низкая скорость диссоциации), клон 4Hu3 (высокая скорость ассоциации, низкая скорость диссоциации) клон 82Hu3 (высокая скорость ассоциации, средняя скорость диссоциации), и химерный клон 53 (средняя скорость ассоциации, низкая скорость диссоциации).
Связывание антитела дополнительно оценивали в CXCR3-экспрессирующих клетках. Клетки 300.19, трансфицированные человеческим CXCR3, подвергали контактированию с очищенными гуманизированными вариантами анти-hCXCR3 антитела Hu1, Hu2, Hu3, и с химерным антителом. Как показано на фиг.19, каждый из клонов 4, 12, 53, 82 и 135 вводили в концентрации 5 мкг/мл (черная линия), 0,5 мкг/мл (темно-серая линия) или 0,1 мкг/мл (черная пунктирная линия), или вводили 5 мкг/мл только «второго» антитела (гистограмма, заштрихованная серым). Клетки окрашивали клонами немеченного антитела в течение 30 минут на льду, а затем два раза промывали PBS-1% FBS, и связанное антитело детектировали с использованием ФИТЦ-конъюгированного специфического «второго» антитела против человеческого IgG1 путем инкубирования на льду в течение 30 минут в темноте. Образцы два раза промывали, фиксировали в 2% растворе параформальдегида-PBS и хранили в темноте при 4°C, а затем загружали на проточный цитометр. Гистограммы CXCR3, позитивно гейтированного на жизнеспособных клетках, представлены на фиг.19.
Пример 10: Сравнение с антителом 1C6
Клон анти-hCXCR3 mAb 1C6 (Becton, Dickinson catalog #557183, тот же самый клон, который был описан в патенте США № 6184358), сравнивали с мышиным клоном анти-hCXCR3 mAb и с их гуманизированными вариантами Hu2 и Hu3 методом анализа Biacore на целом рецепторе. Клон 4 обладал приблизительно в 2 раза большей аффинностью (KD). Гуманизированные варианты обладали еще большей аффинностью (приблизительно в 4 раза превышающей аффинность вариантов Hu2 и Hu3). В таблице 8 приводятся величины кинетики и аффинности связывания для клона 1C6 и клона 4 и их гуманизированных вариантов Hu2 и Hu3.
Пример 11: Функциональные анализы
Были оценены функциональные эффекты двадцати вариантов антител. Эти антитела оценивали на их влияние на хемотаксис клеток в ответ на действие CXCL9, CXCL10 и CXCL11 и на ингибирование мобилизации кальция в анализе FLIPR® на кальций.
Хемотаксис клеток 300.19, трансфицированных человеческим CXCR3, к лигандам CXCR3, а именно, CXCL9, CXCL10 и CXCL11, оценивали в отсутствии или в присутствии 10 мкг/мл гуманизированных вариантов антител против человеческого CXCR3, клонов 4, 12, 53, 82 и 135. Трансфицированные клетки предварительно обрабатывали 10 мкг/мл гуманизированных вариантов антител против человеческого CXCR3 или коммерчески доступным антителом 1С6 в течение 20 минут при 37°C. Клетки с антителом переносили в 5-микронные лунки Transwells, и в лунки приемника, содержащие 100 или 300 нг/мл рекомбинантого мышиного CXCL10 и CXCL11 или CXCL9, соответственно, помещали вставки. Планшеты для хемотаксиса инкубировали при 37°C в атмосфере 5% CO2 в течение 4 часов. Вставки Transwells вынимали, и клетки, которые мигрировали в лунки приемника, переносили в 96-луночные планшеты с U-образным дном, осаждали и ресуспендировали в кальцеиновом красителе AM. Клетки инкубировали в течение 30 минут при 37°C в атмосфере 5% CO2, один раз промывали, переносили в черный/прозрачный планшет и данные сразу считывали на приборе FiexStation на длине волны 490/520 нм. Данные представлены как процент ингибирования хемотаксиса. На фиг. 20 представлены репрезентативные данные, иллюстрирующие способность различных вариантов каждого из пяти клонов ингибировать хемотаксис к CXCL9, CXCL10 и CXCL11.
Для оценки ингибирования мобилизации внутриклеточного кальция, поток кальция в клетках СНО, трансфицированных человеческим CXCR3-Gqi4qi4, измеряли в ответ на CXCL10 в отсутствии или в присутствии различных концентраций вариантов анти-CXCR3 антитела или антитела 1С6, используемого в качестве позитивного контроля. Клетки высевали (12000/лунку) в черные 384-луночные планшеты и клетки оставляли на ночь для связывания. На следующий день, клетки нагружали кальций-чувствительным красителем Fluo-4NW за 40 минут до добавления антитела и инкубировали при 37°C. Затем добавляли антитело, и клетки оставляли на 20 минут для инкубирования, после чего добавляли лиганд CXCR3, рекомбинантный человеческий CXCL10, в предварительно определенной концентрации EC80 для CXCL10. Добавление красителя осуществляли вручную с помощью электронной многоканальной пипетки, а добавление антитела и агониста осуществляли автоматизированным методом на FLIPR Tetra®, и после добавления CXCL10, данные сразу считывали с планшета на длине волны 470-495 нм. Образцы получали в дубликате, и строили график зависимости среднего процента ингибирования (± стандартное отклонение) при каждой концентрации антитела. Клон 4 Hu1 не ингибировал мобилизацию кальция и был использован в качестве негативного контроля в этих экспериментах. На фиг. 21 проиллюстрирована способность различных вариантов каждого из пяти клонов ингибировать мобилизацию кальция, и сравнение этой способности с соответствующей способностью коммерчески доступного клона 1C6. Репрезентативные величины IC50 (M) антител против CXCL10 в анализе на мобилизацию кальция представлены ниже в таблице 9.
В таблице 9, (*) указывает на то, что антитело 1C6 не является химерным антителом, а представляет собой полностью мышиное антитело IgG1 против человеческого CXCR3.
Как показано в таблице 10, данные по кинетике связывания (смотрите пример 9) хорошо коррелируют с результатами функционального анализа. Исходя из данных по кинетике связывания и результатов функционального анализа, для дополнительной оценки и 4D-гуманизации были отобраны клоны 4 и 53.
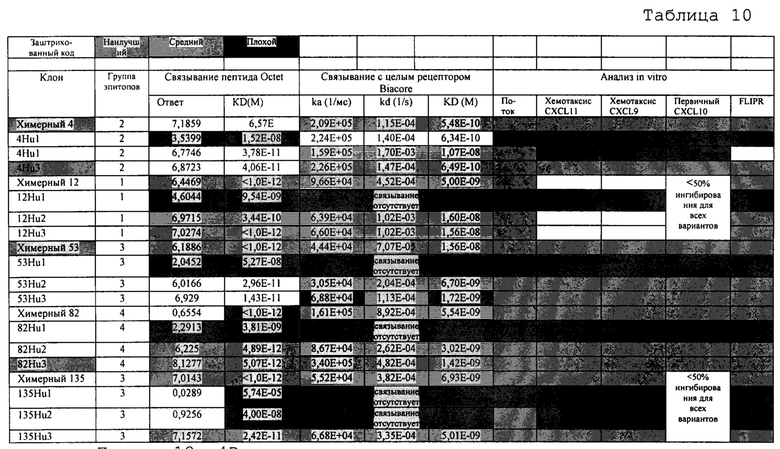
Пример 12: 4D-гуманизация
4D-гуманизацию клонов анти-hCXCR3 антитела осуществляли, как описано в WO 2009/032661 (например, в параграфах [0037]-[0044]). Вкратце, 4D-гуманизация включает: a) построение 3-D-модели вариабельного домена, который должен быть гуманизирован; b) идентификацию гибких остатков в вариабельном домене посредством имитации молекулярной динамики 3-D-модели указанного домена; c) идентификацию наиболее близкородственной человеческой зародышевой линии путем сравнения молекулярно-динамического пути 3-D-модели с молекулярно-динамическими путями 49 человеческих зародышевых линий (а не путем сравнения последовательностей антител, как это обычно осуществляют в случае традиционной гуманизации); и d) введение мутации гибких остатков, которые не являются частью CDR, в их аналог человеческой зародышевой линии (идентифицированной в стадии c). Этим методом были сконструированы тяжелые цепи 4.4-4.6 и легкие цепи 4.4-4.7. В частности, была сконструирована исходная 4D-гуманизированная конструкция (VH 4.4 и VL 4.4) а затем в эти тяжелые и легкие цепи были внесены дополнительные модификации (VH 4,5-4.6 и VL 4,5-4.7) для введения стабилизирующих и антиагрегирующих мутаций и для удаления других нежелательных мотивов. Аналогичные методы были также применены для создания 4D-гуманизированных конструкций VH 53.7-53.10 и VL 53.10-53.13.
В таблице 11 проиллюстрирована стратегия гуманизации (и дополнительных модификаций, если это необходимо), применяемая для получения вариантов тяжелой и легкой цепей клонов 4, 12, 53, 82 и 135, включая гуманизированные и 4D-гуманизированные цепи (VH = тяжелая цепь; VK = легкая цепь).
Несколько 4D-вариантов клона 4 (4Hu6, 4Hu7, 4Hu8, 4Hu9, 4Hu10) анализировали методом Biacore на целом рецепторе для оценки кинетики связывания и аффинности по отношению к CXCR3, и полученные данные сравнивали с данными для клона 4 химерного антитела. Как показано в таблице 2, клон 4Hu6 содержал тяжелую цепь 4.4 и легкую цепь 4.4. Клон 4Hu7 содержал тяжелую цепь 4.4 и легкую цепь 4.7. Клон 4Hu8 содержал тяжелую цепь 4.5 и легкую цепь 4.5. Клон 4Hu9 содержал тяжелую цепь 4.5 и легкую цепь 4.8. И клон 4Hu10 содержал тяжелую цепь 4.6 и легкую цепь 4.4.
Как показано в таблице 12, при сравнении вариантов 4D-модели клона 4 с химерным вариантом (4Ch) было обнаружено, что четыре из этих пяти вариантов обладали повышенной аффинностью (KD). Однако 4D-вариант 4Hu10 обнаруживал снижение аффинности приблизительно на 1 порядок величины.
Пример 13: Мышиная модель NSG-PBL
Мышам NOD-scid IL2rγnull (NSG) инъецировали 27 свежих первичных человеческих мононуклеарных клеток периферической крови (МКПК), выделенных в градиенте фиколла, на день 0. Животных (n=8/группу) обрабатывали 5 мг/кг химерных антител против человеческих CXCR3 или контрольным человеческим IgG1 (герцептином) два раза в неделю в течение всего исследования, начиная со дня 3 после переноса клеток. Кровь, взятую на 14-й день после начала проведения исследования, подвергали проточной цитометрии и окрашивали стандартными методами с использованием антител против человеческого CD45 (hCD45), человеческого CD3 (hCD3), человеческого CD4 (hCD4), человеческого CD8 (hCD8) и человеческого CXCR3. В лимфоцитарном гейте определяли гейт hCD45+-клеток, а затем hCD3+-клеток. Затем оценивали экспрессию hCD4 и hCD8 на hCD45+hCD3+-T-клетках и определяли процент человеческих CD4+CD45+CD3+-Т-клеток и человеческих CD8+CD45+CD3+-Т-клеток. После этого определяли процент CXCR3-экспрессирующих человеческих CD4+-T-клеток и CD8+-T-клеток. Каждая точка на фиг.22 представляет одну мышь, где репрезентативные данные были получены в трех экспериментах. Полученные данные показали, что обработка мышей NSG-PBL с моделью ксеногенной GvHD (реакции «трансплантат против хозяина») анти-CXCR3 антителом приводила к модуляции CXCR3-экспрессирующих Т-клеток, но выявила функциональные различия между этими пятью клонами.
В таблице 12 указана средняя выживаемость мышей NSG-PBL с ксеногенной GvHD после обработки этих мышей химерными антителами-кандидатами против человеческого CXCR3.
*p=0,043; **p=0,016; ***p=0,010 в случае обработки анти-CXCR3 антителом по сравнению с huIgG1-обработкой, как было определено с помощью логрангового критерия.
Вышеописанные примеры приводятся лишь в иллюстративных целях и не ограничивают объема настоящего изобретения. Другие варианты описанных здесь устройств и способов будут очевидны для специалиста исходя из описания и практического применения описанных здесь устройств и способов.
| название | год | авторы | номер документа |
|---|---|---|---|
| Гуманизированные антитела к антигену Томсена-Фриденрайха | 2015 |
|
RU2699717C2 |
| АНТИТЕЛА ПРОТИВ CD40 И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2796413C2 |
| НОВЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С CD40 И FAP | 2018 |
|
RU2766234C2 |
| НОВЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ ДИМЕРИЗАЦИЮ С-МЕТ, И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2552161C2 |
| Анти-LAG-3 антитела | 2016 |
|
RU2760582C2 |
| АНТИТЕЛА ПРОТИВ ИНТЕРФЕРОНА-АЛЬФА | 2002 |
|
RU2314317C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ ДЛЯ ПЕПТИДА-6, ПОЛУЧЕННОГО ИЗ HSP65, СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2010 |
|
RU2596925C2 |
| АНТИТЕЛА ПРОТИВ IL2 | 2006 |
|
RU2425054C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| АНТИТЕЛА ПРОТИВ CTLA4 И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2779312C2 |

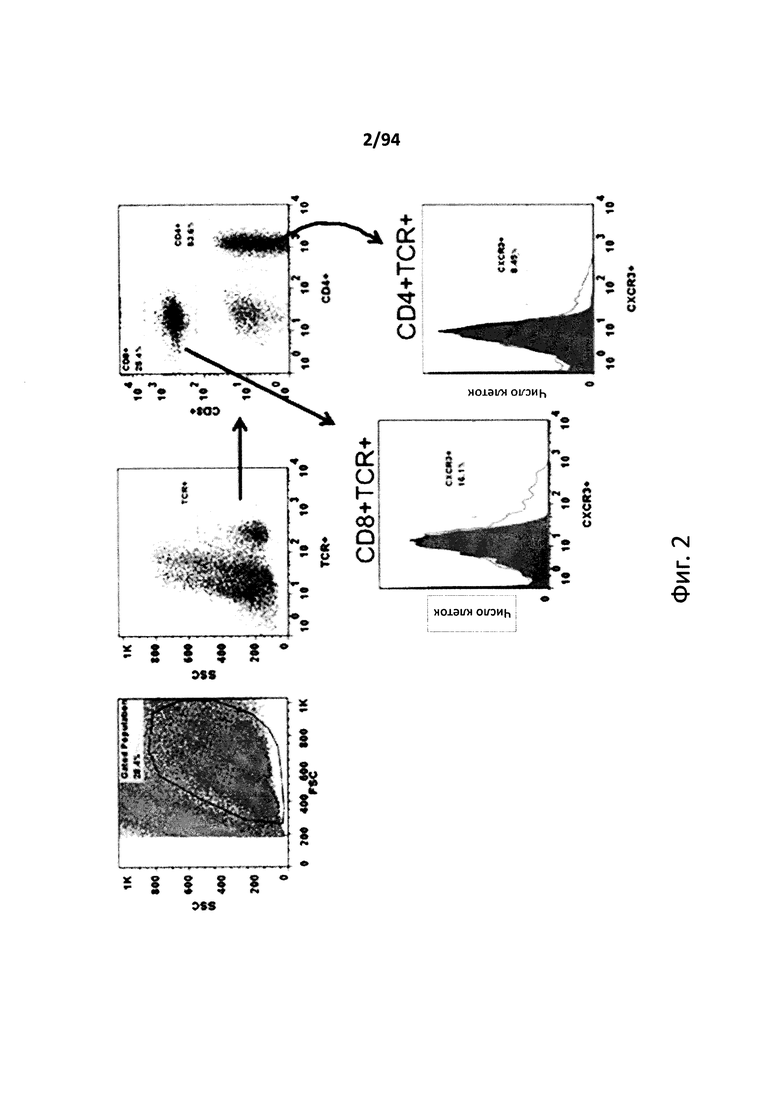
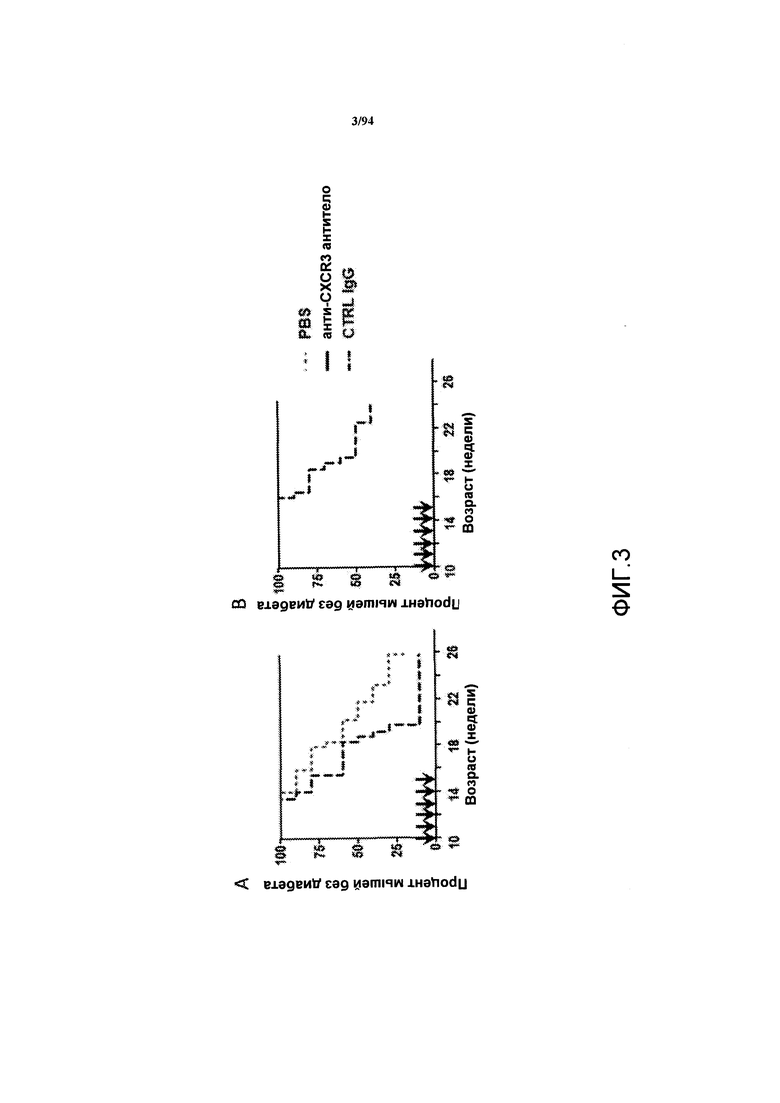
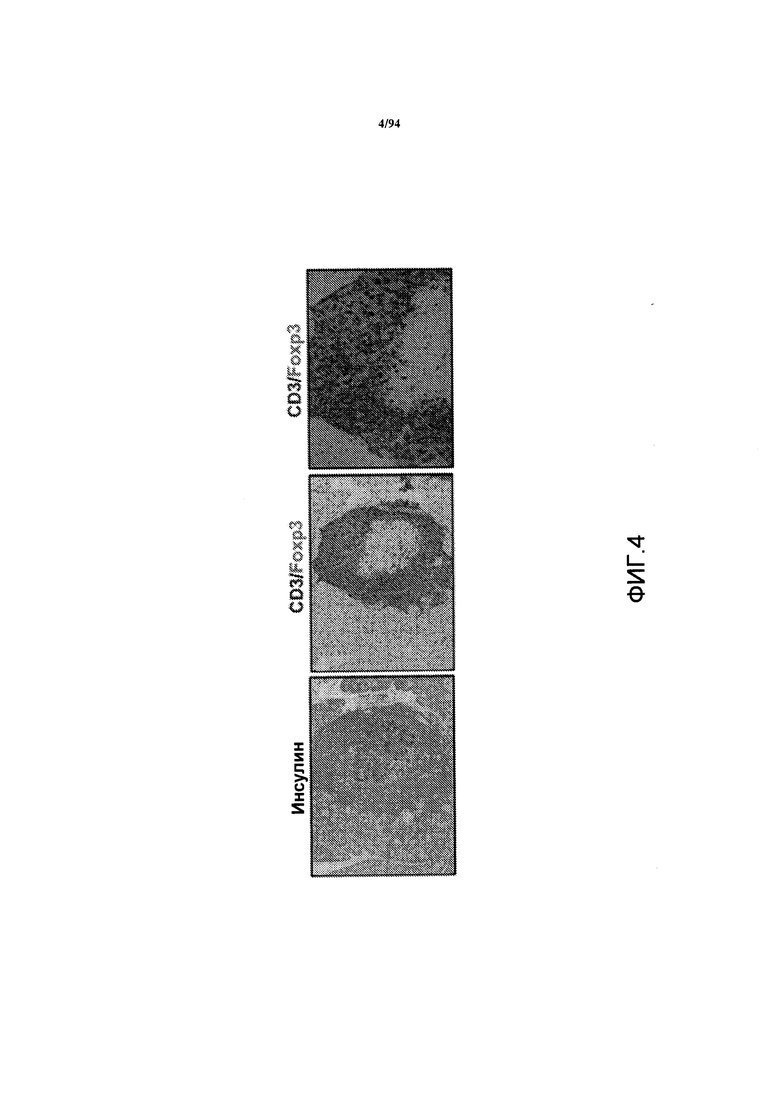
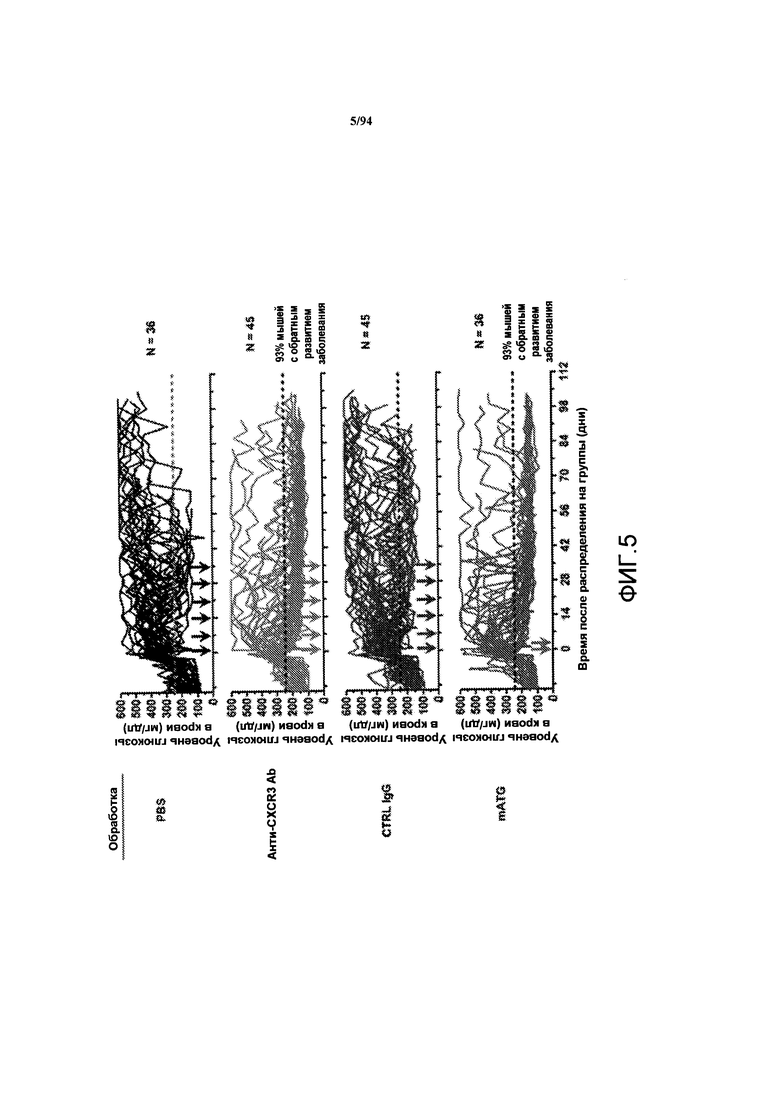
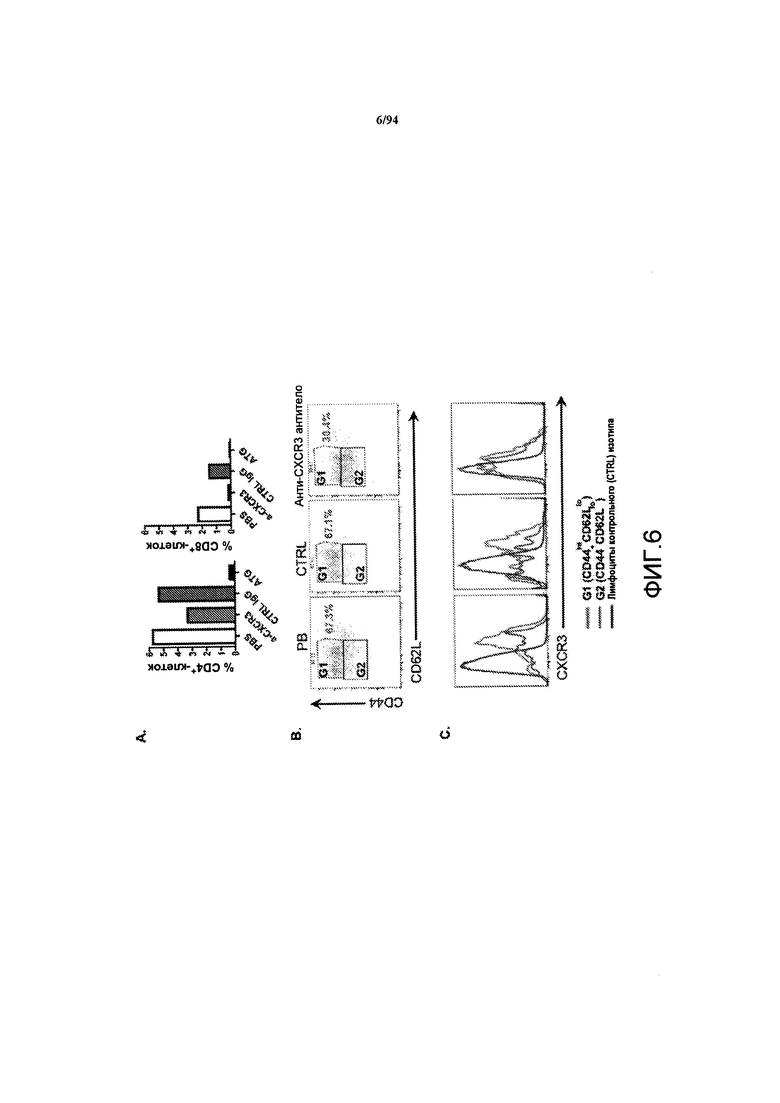
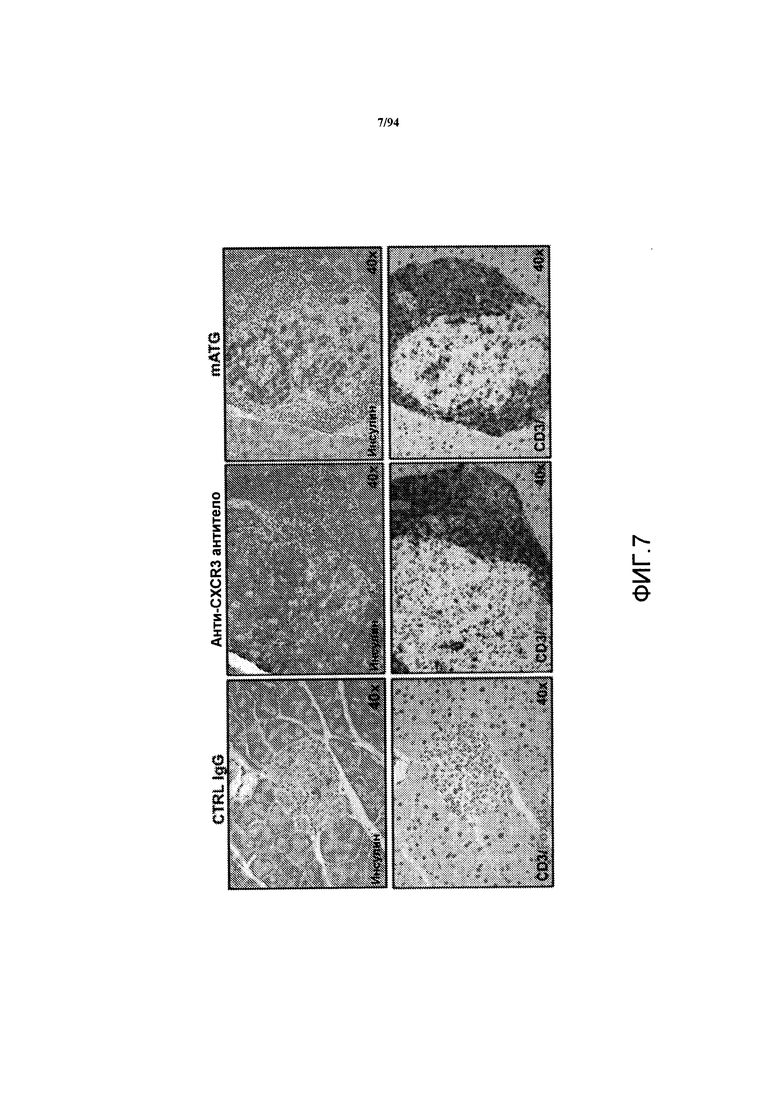
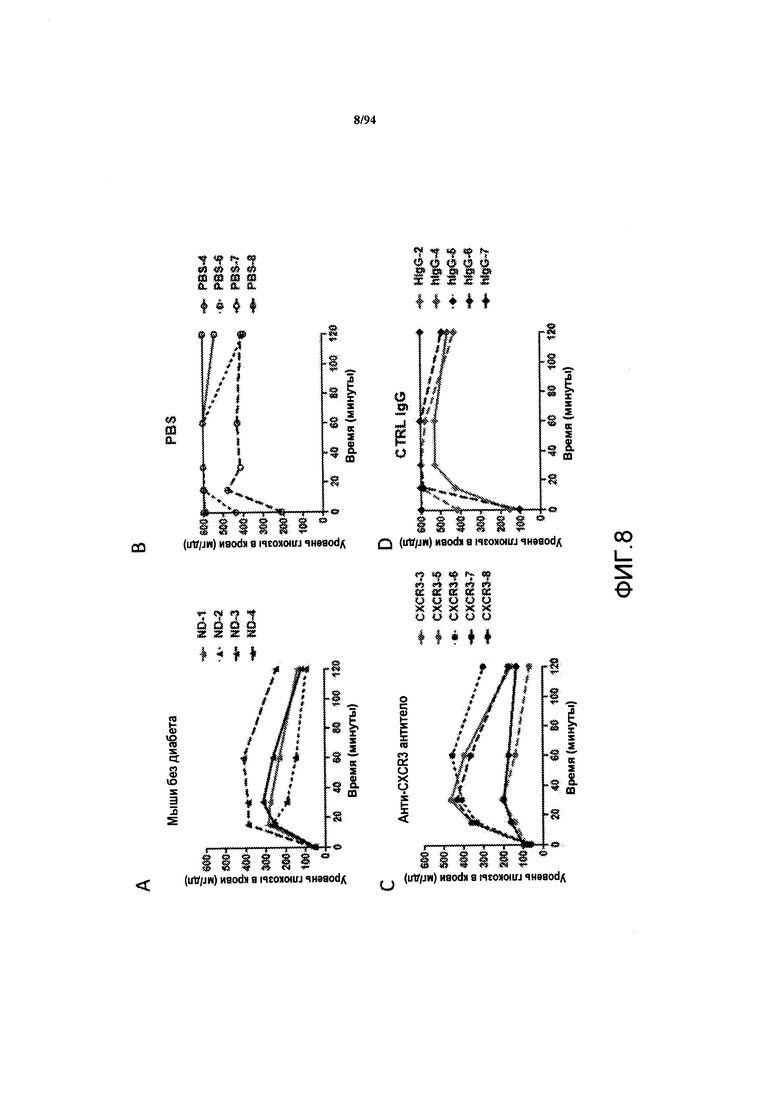
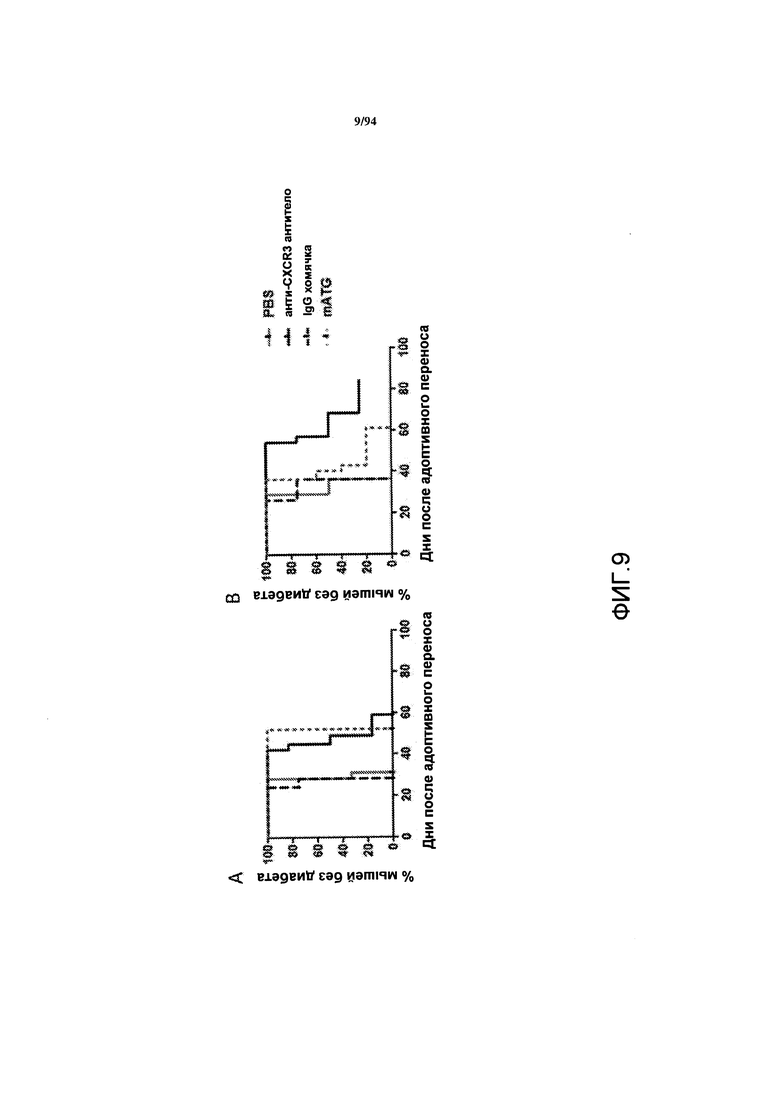
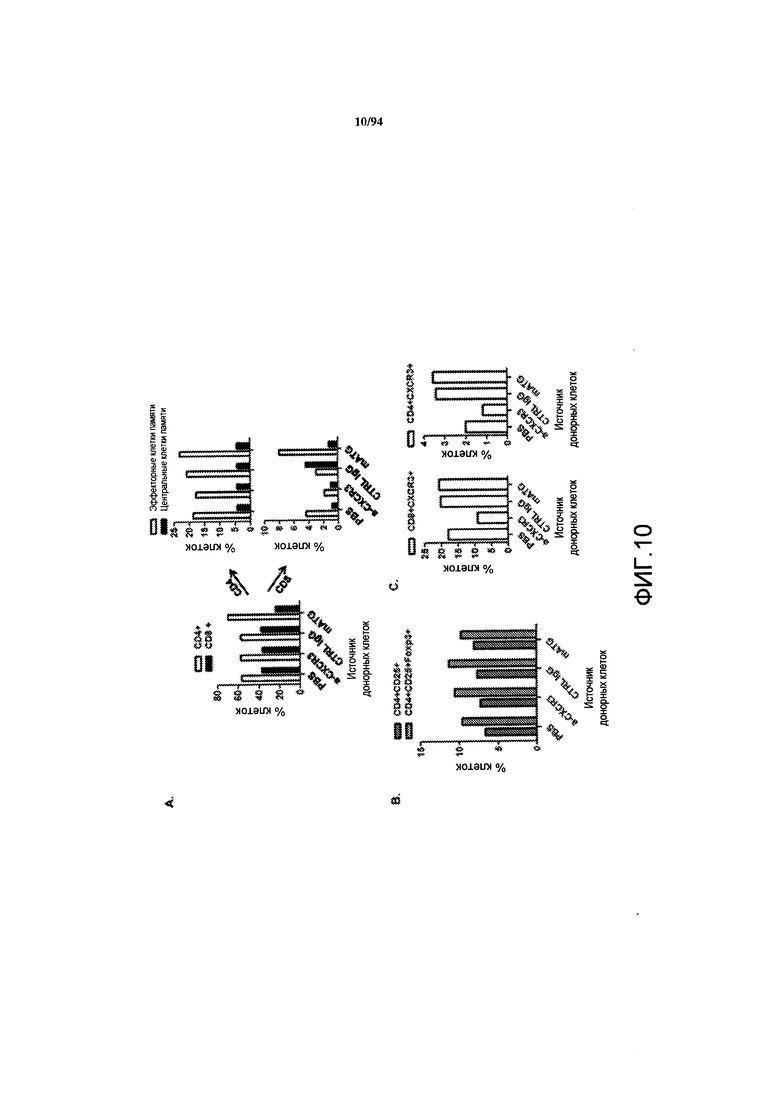
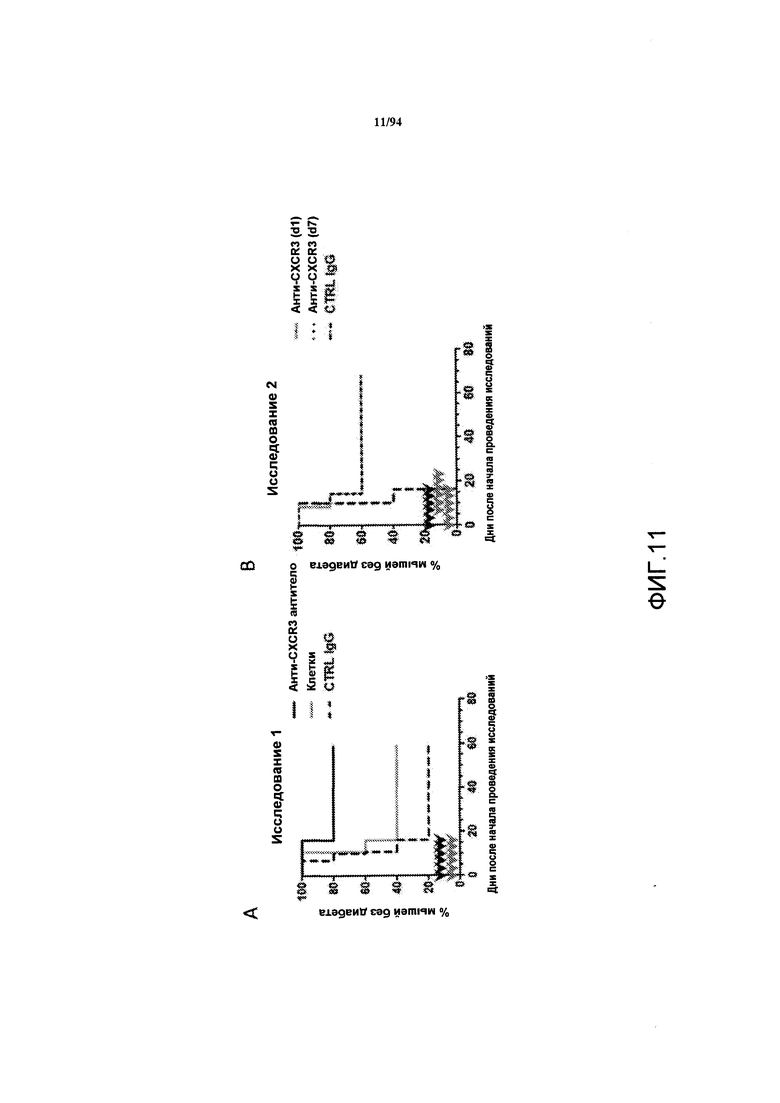
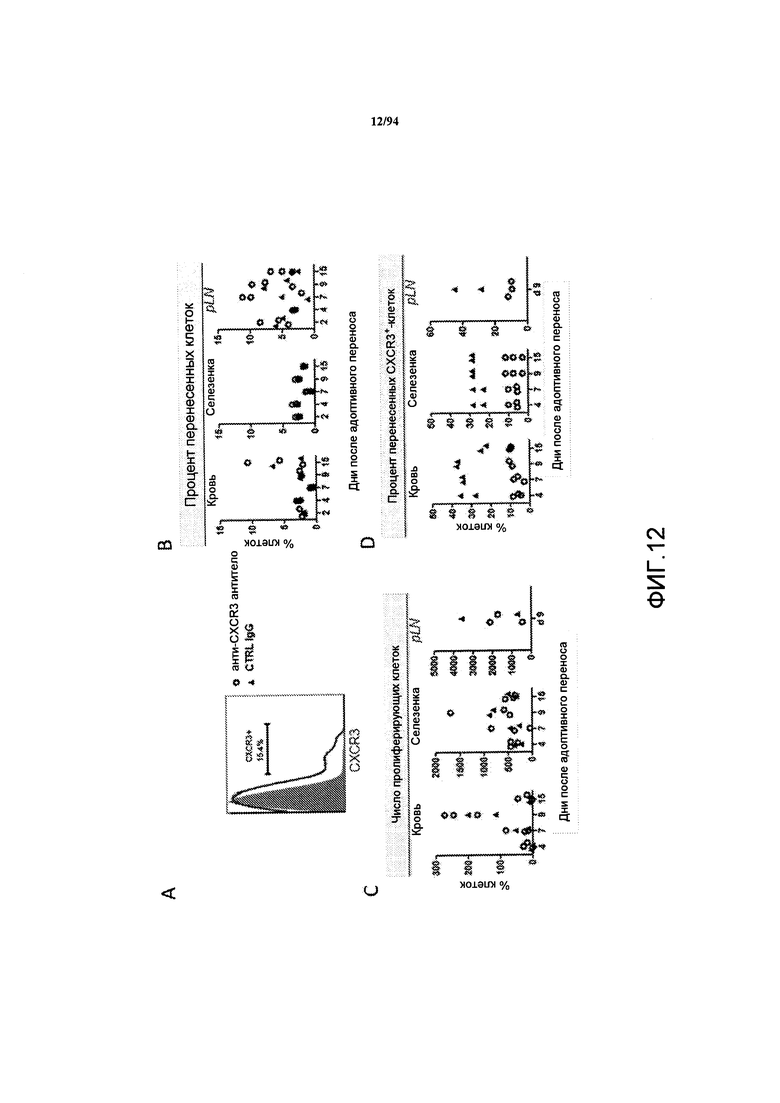
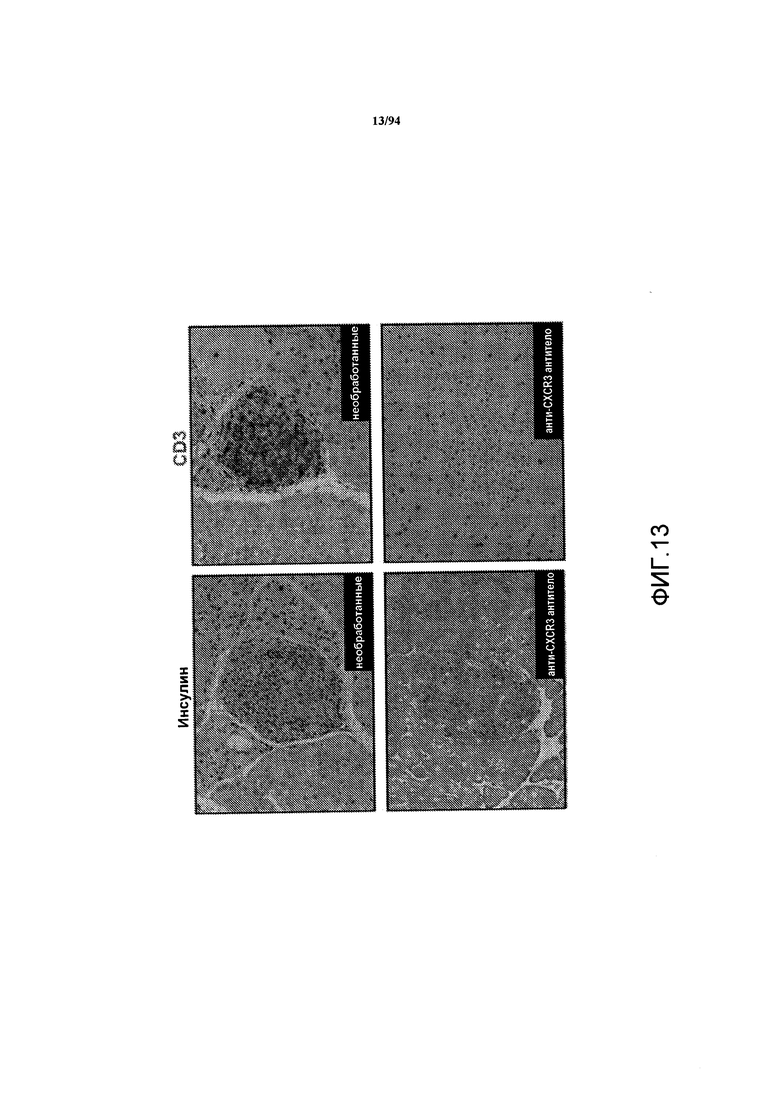
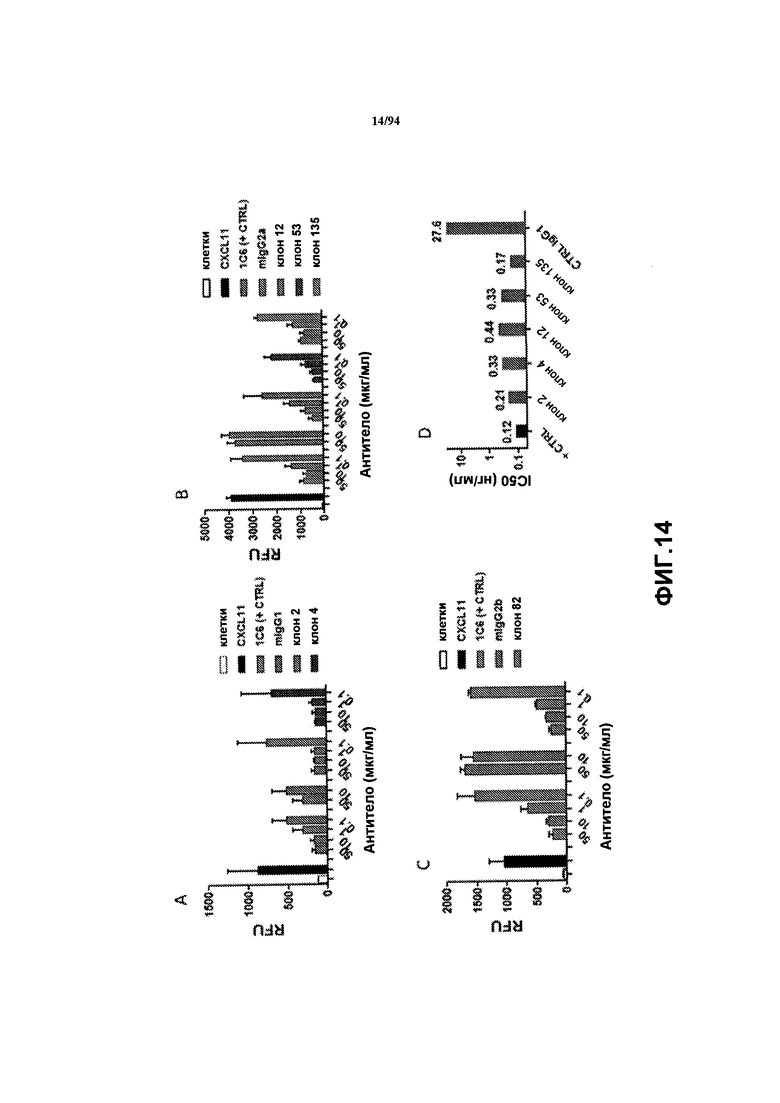
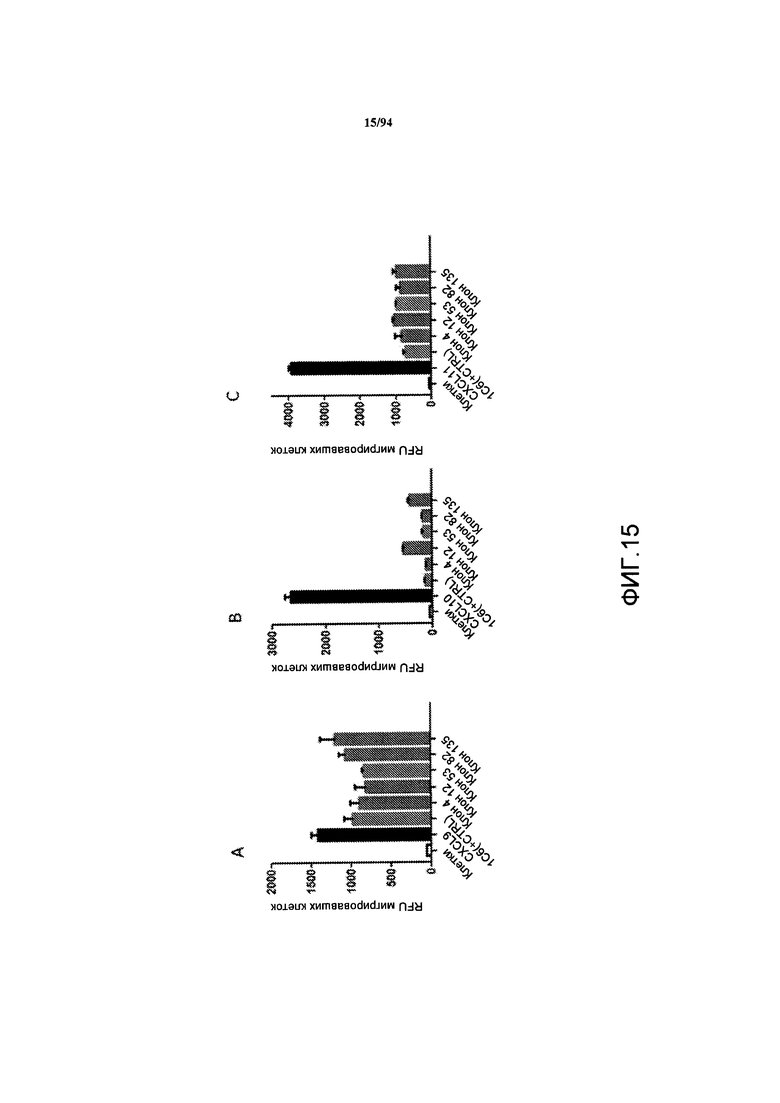
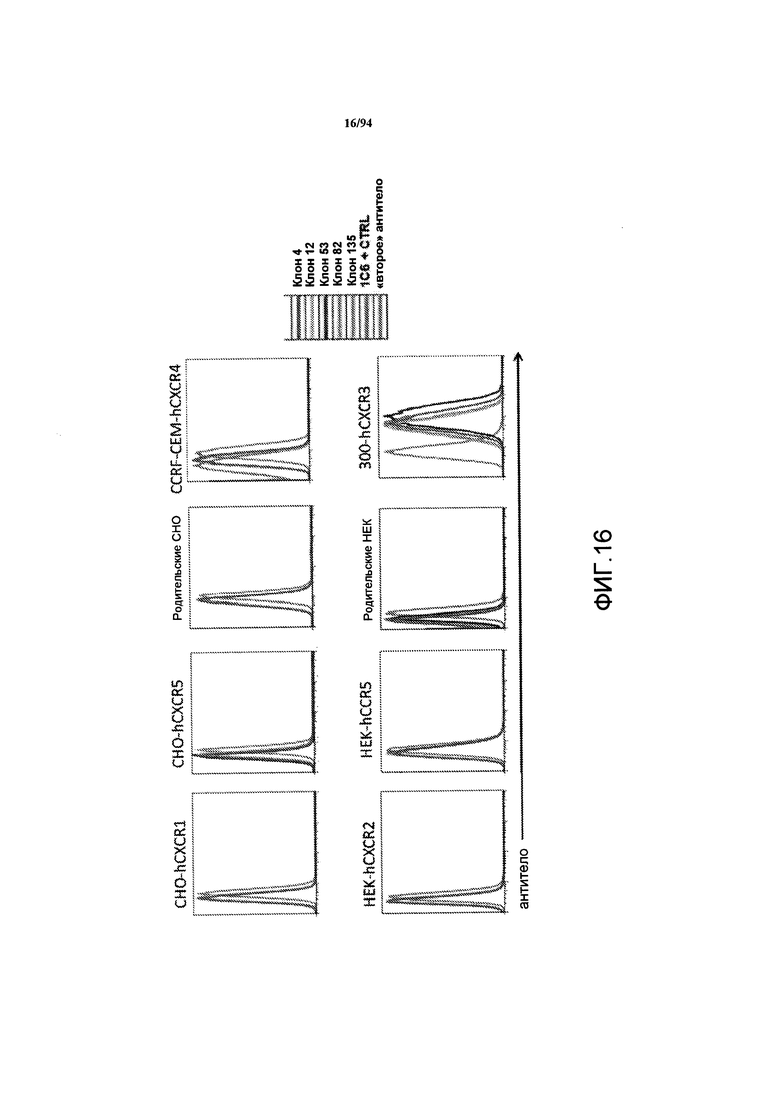
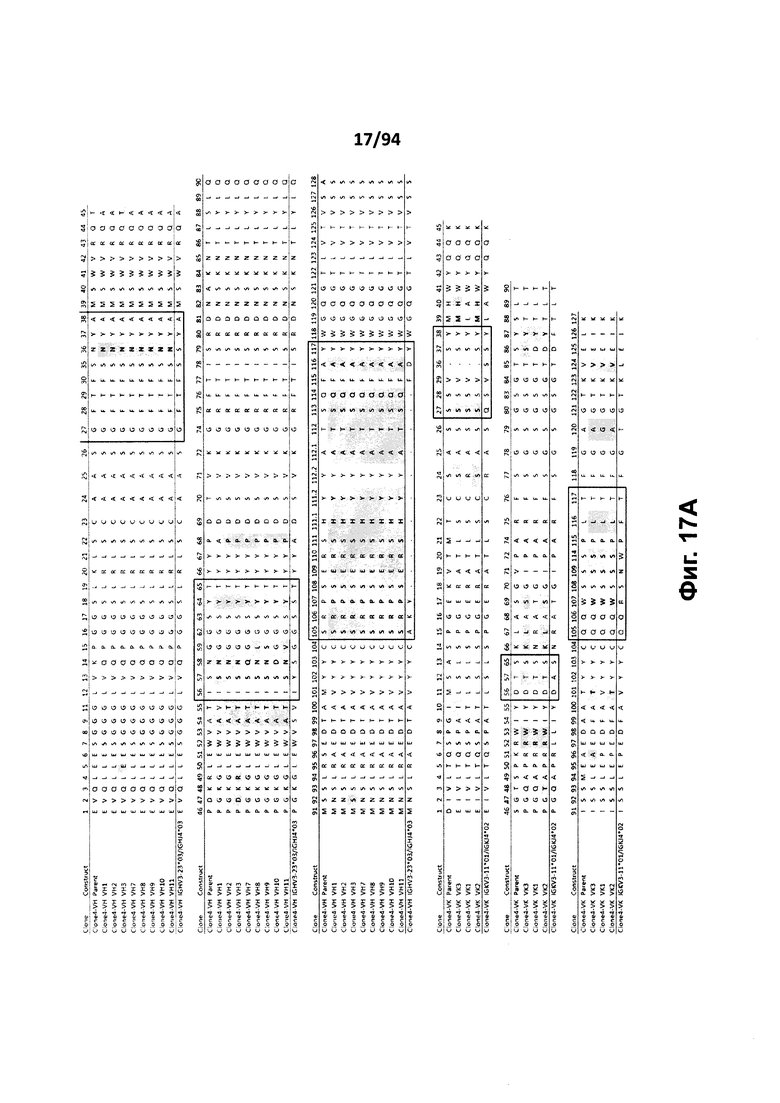
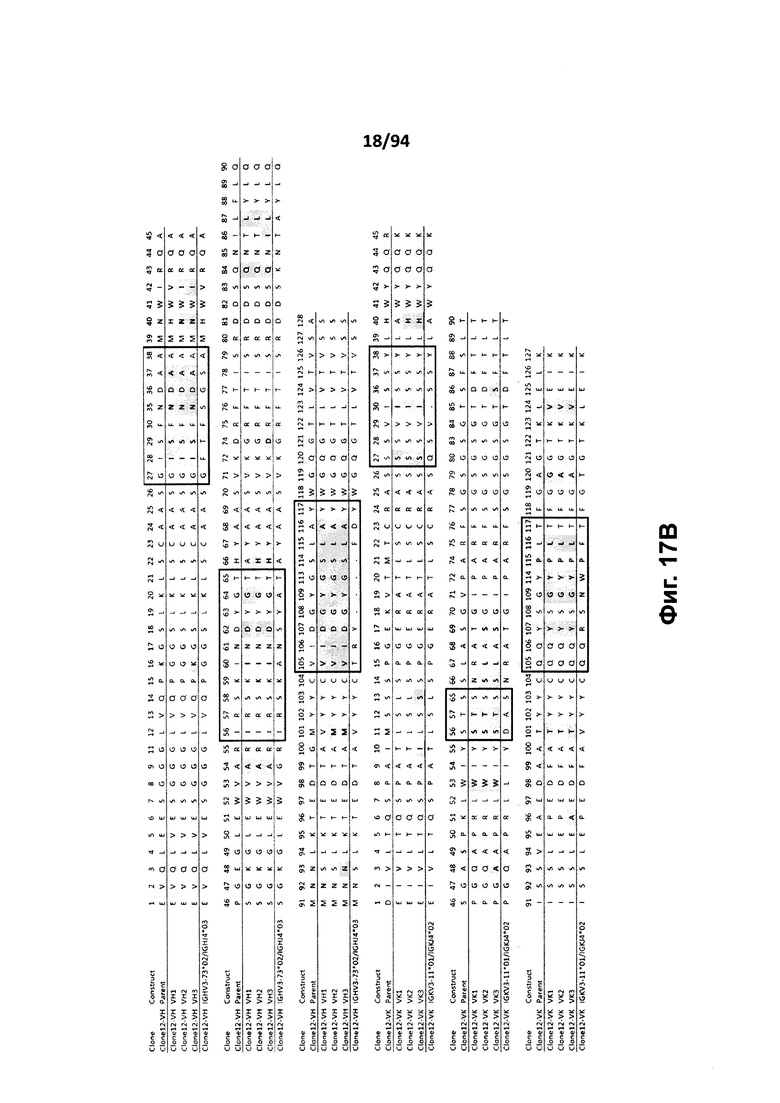
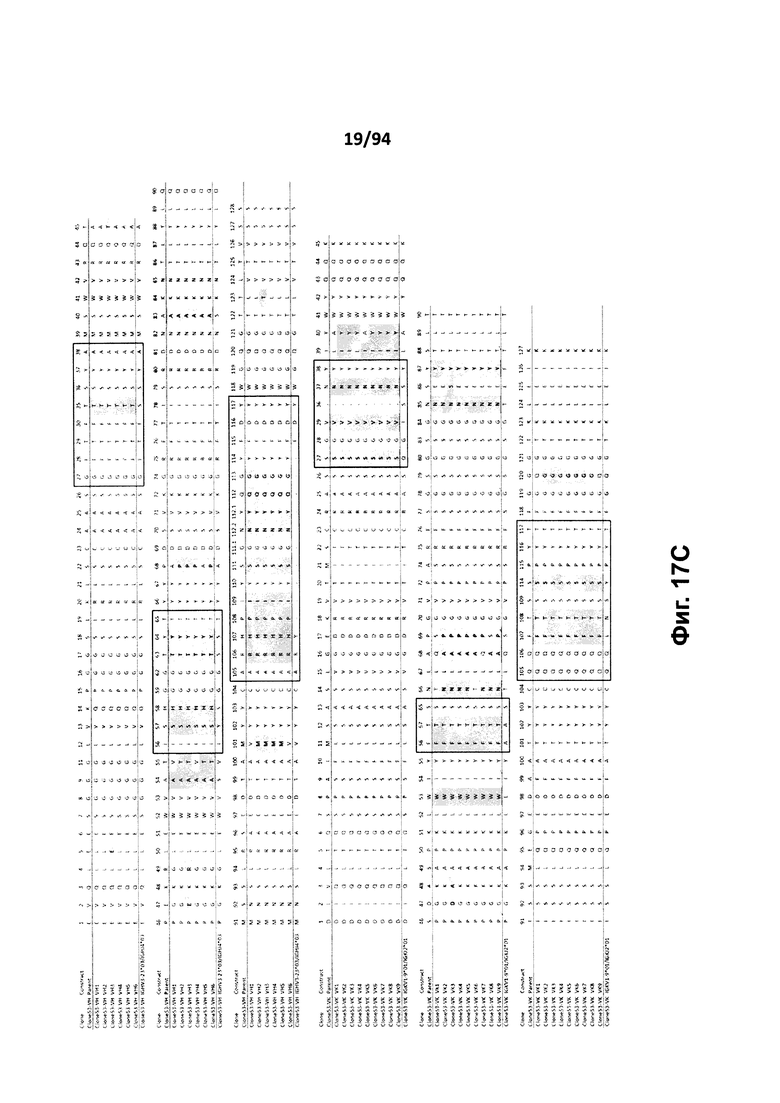
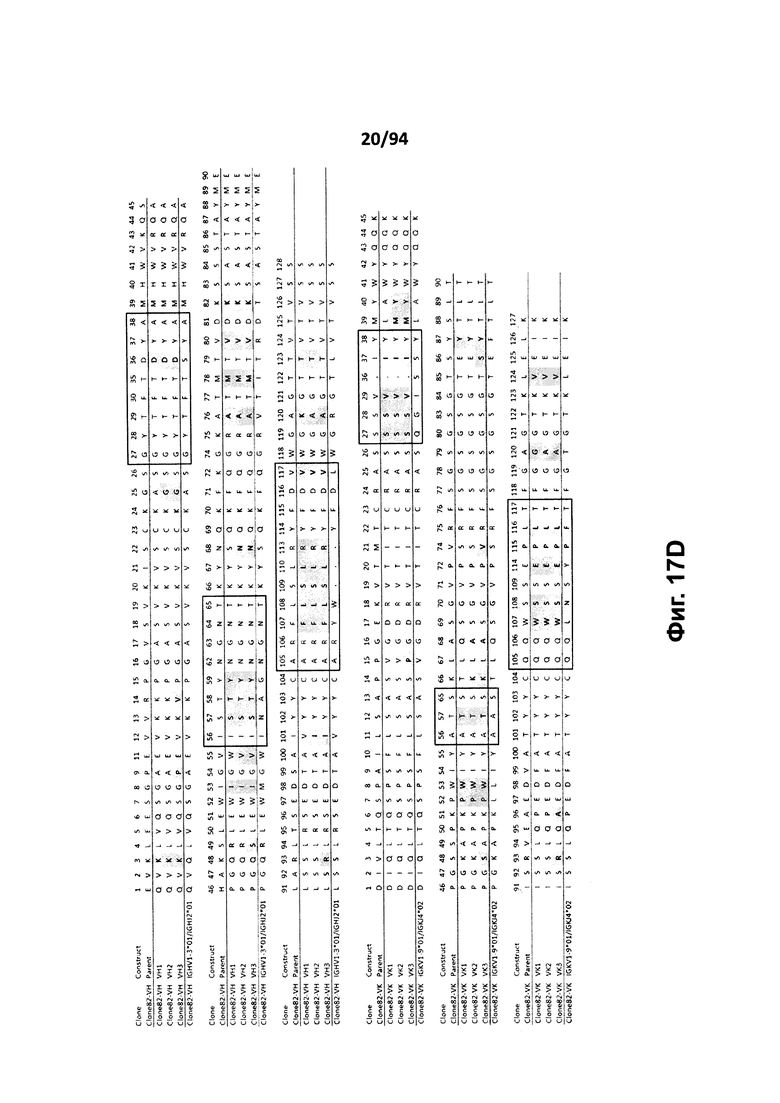
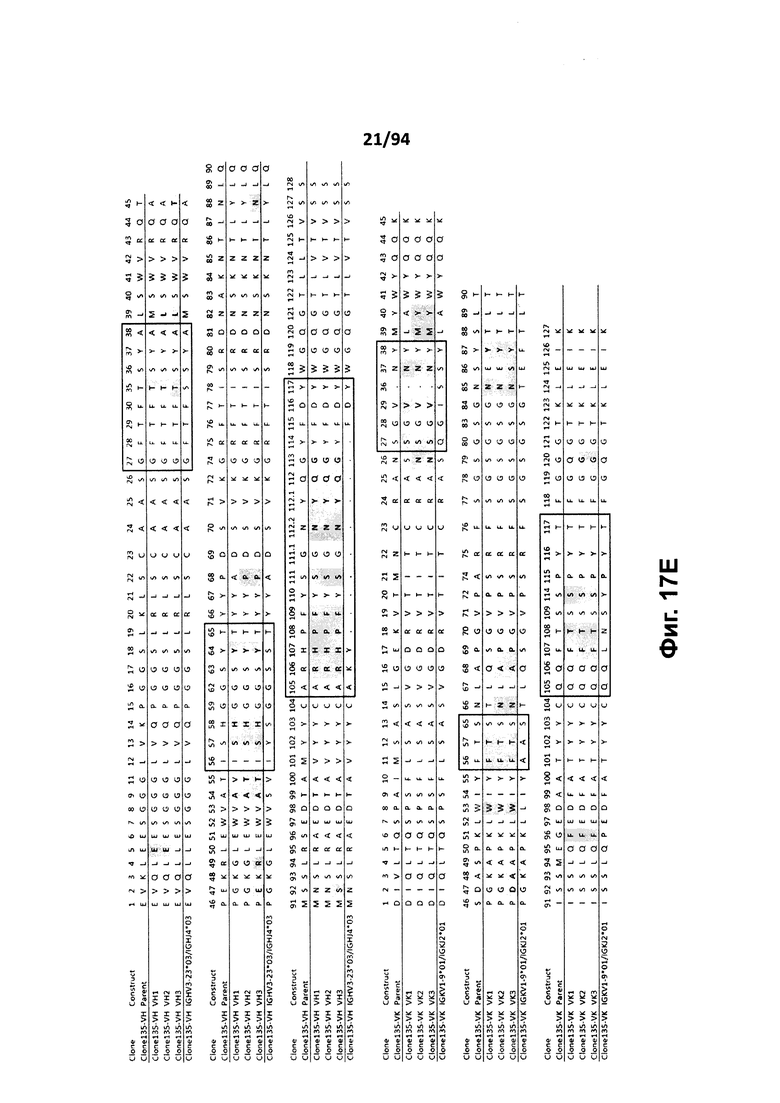
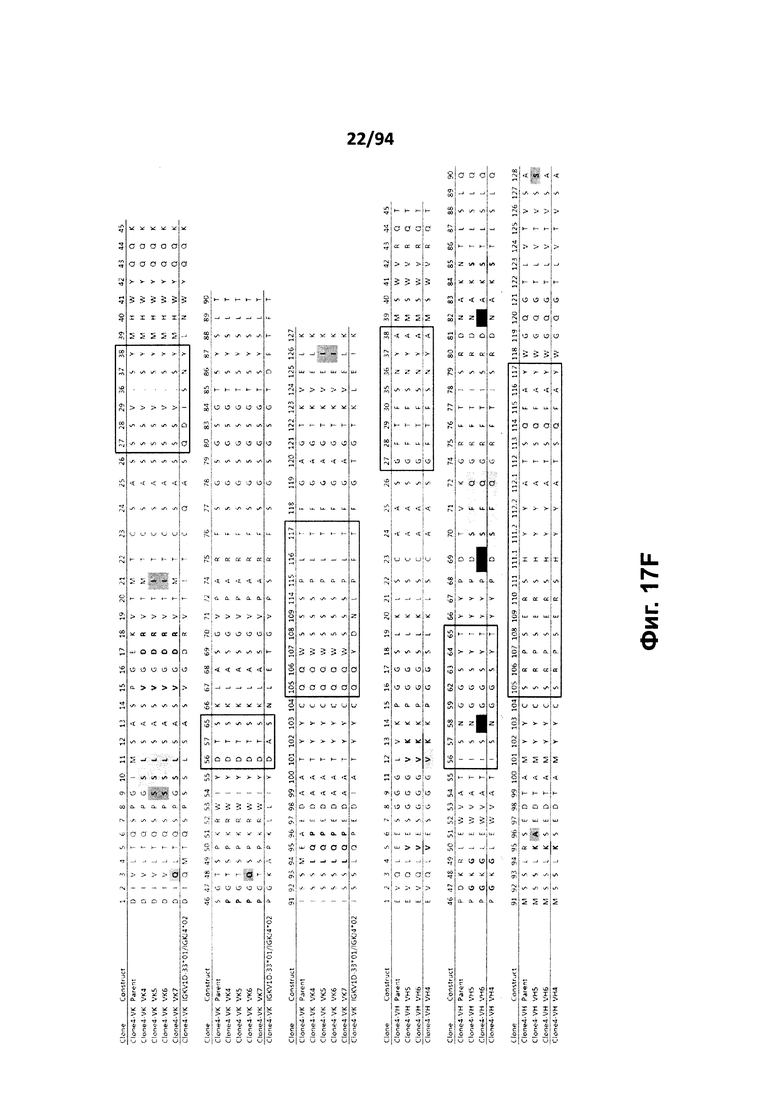
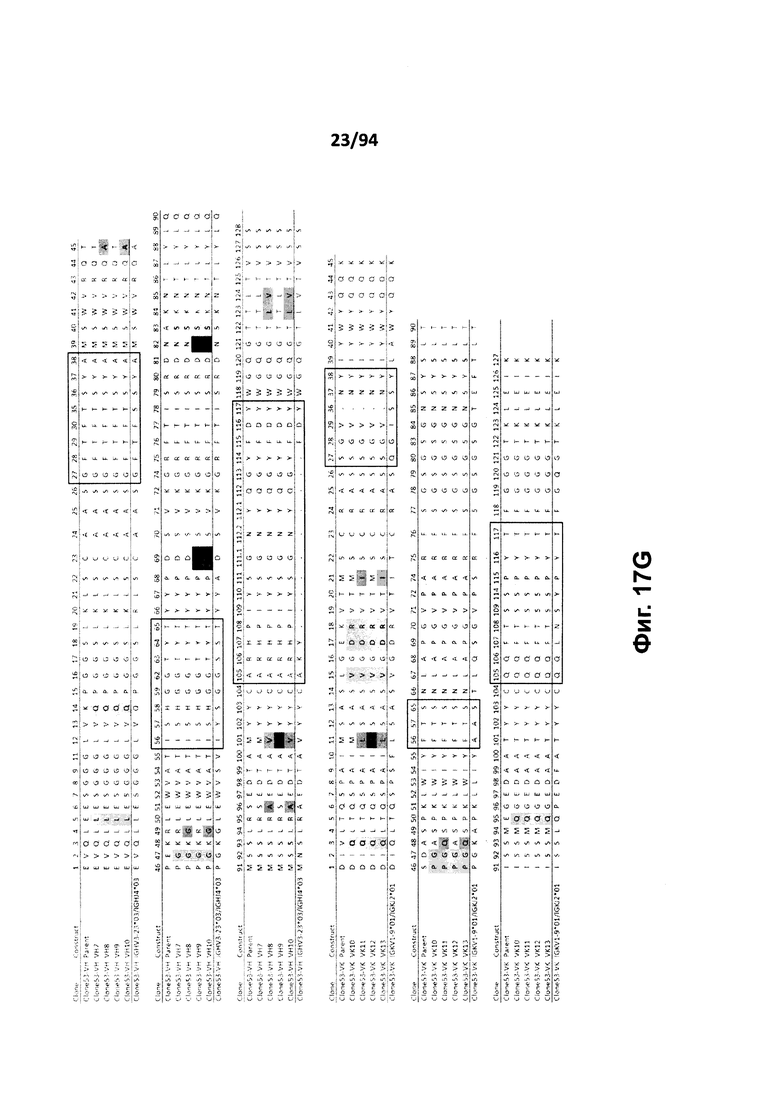
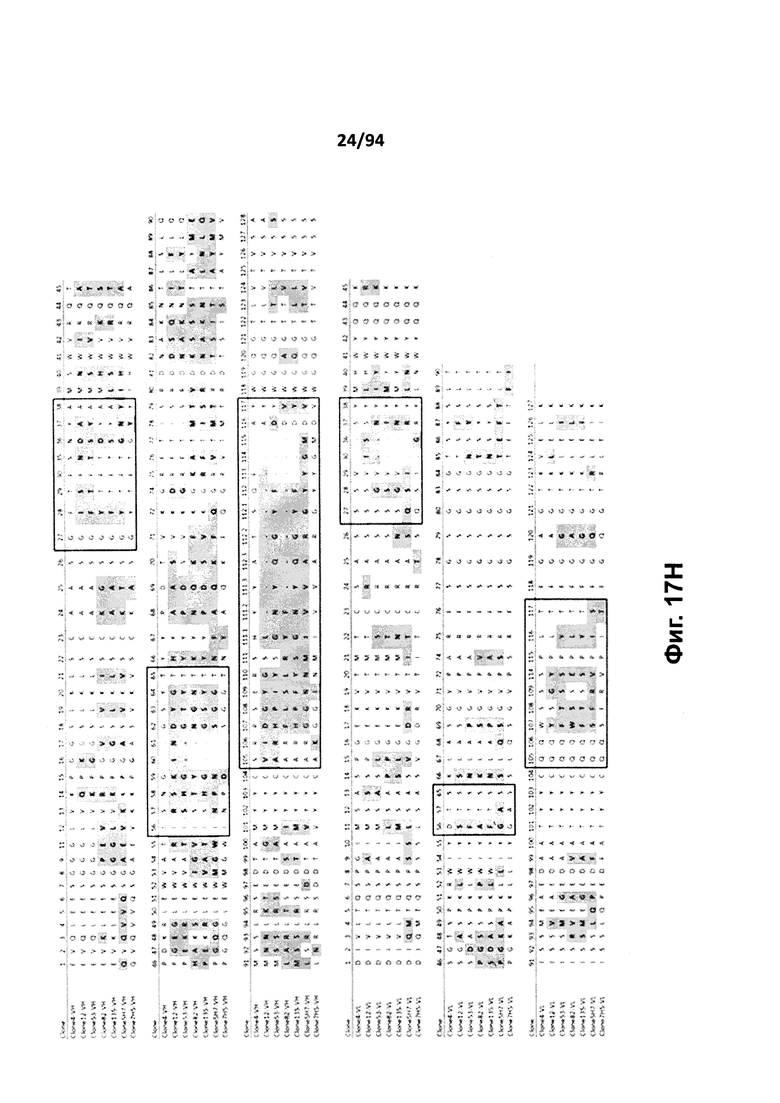
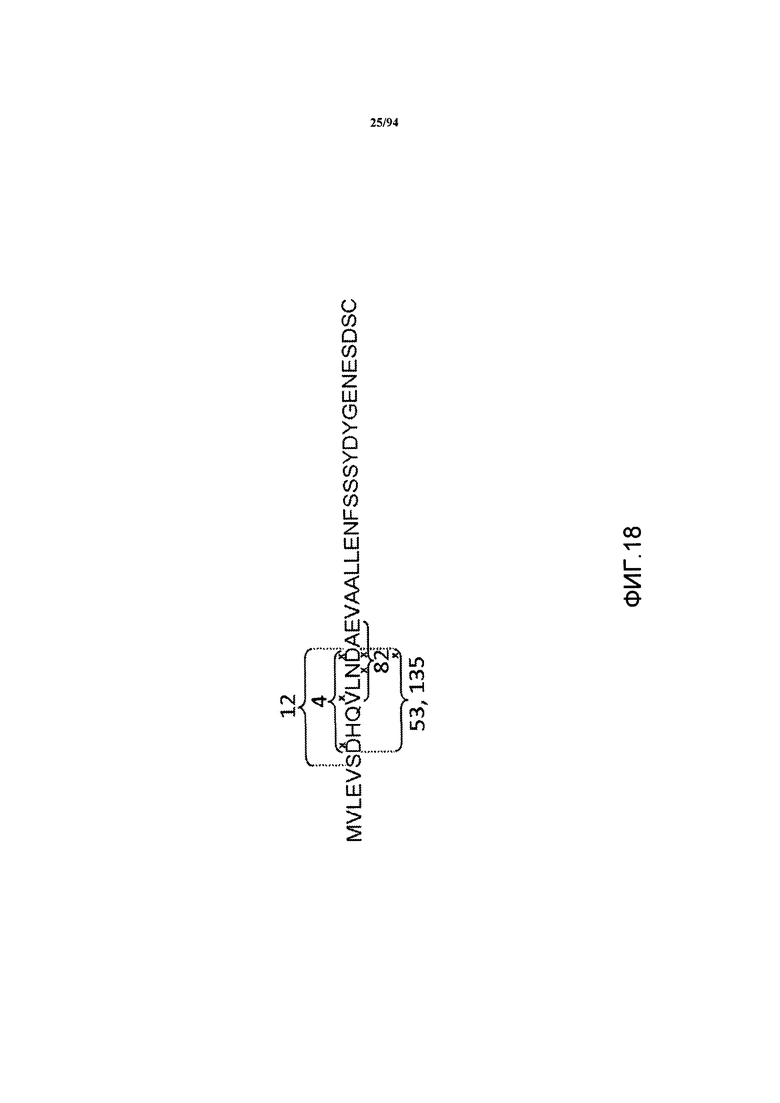
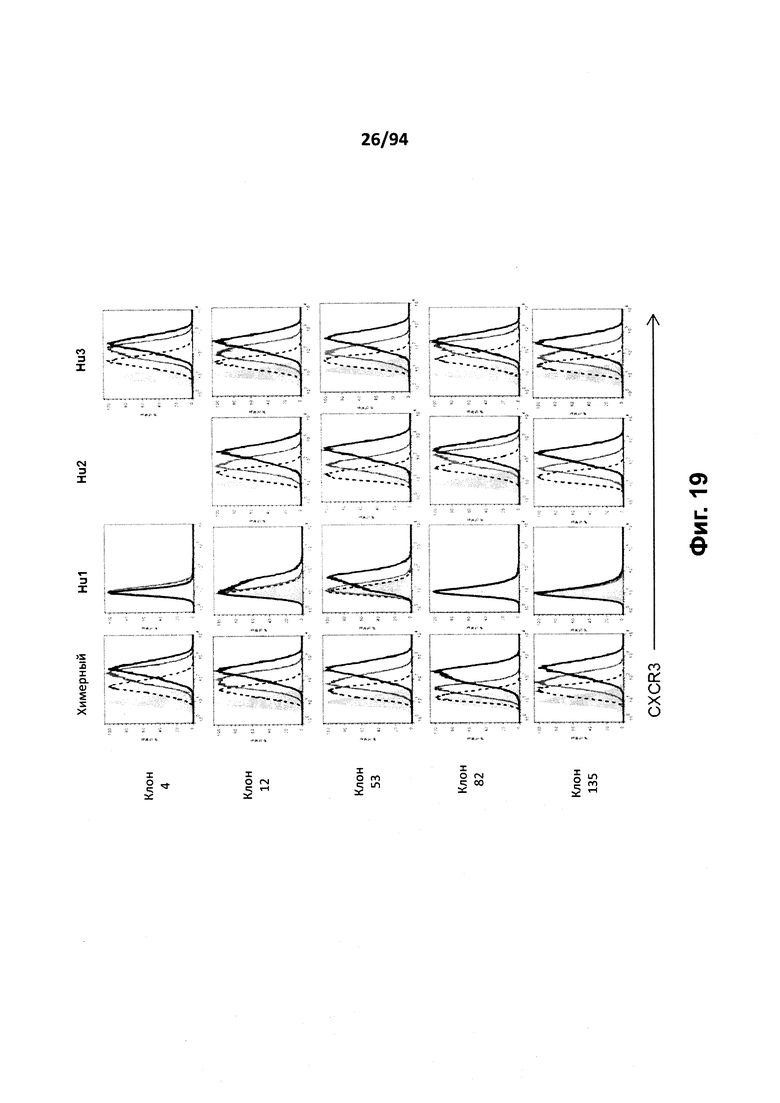
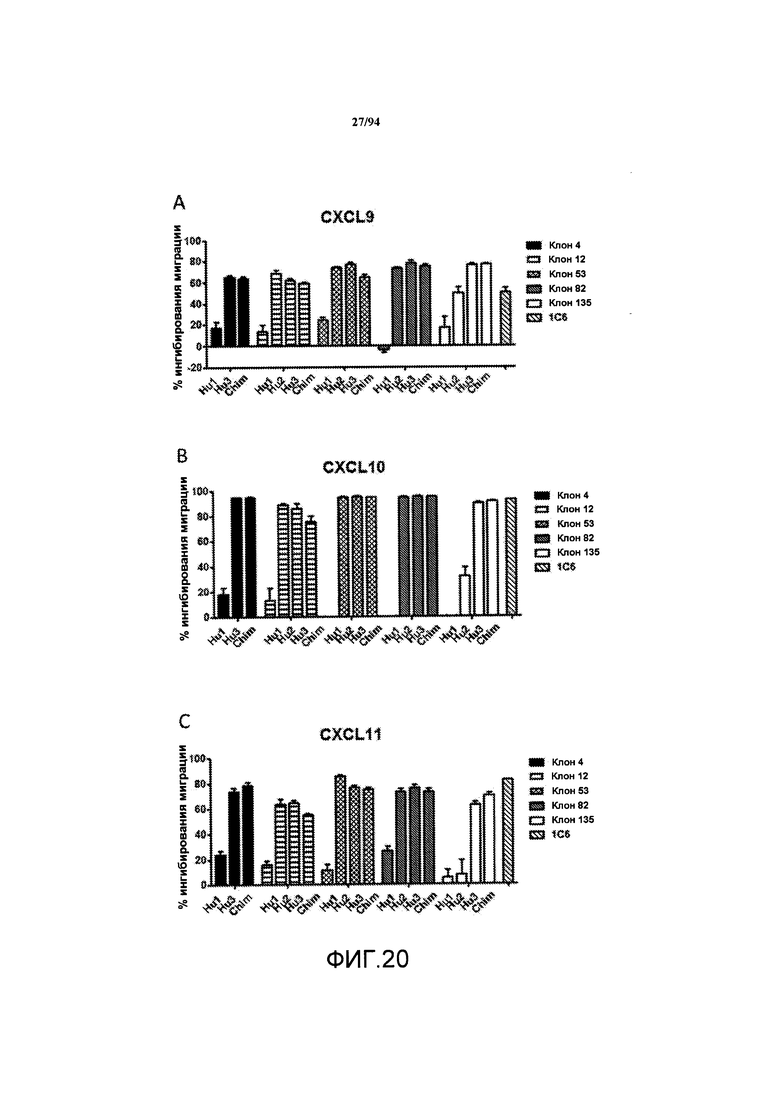
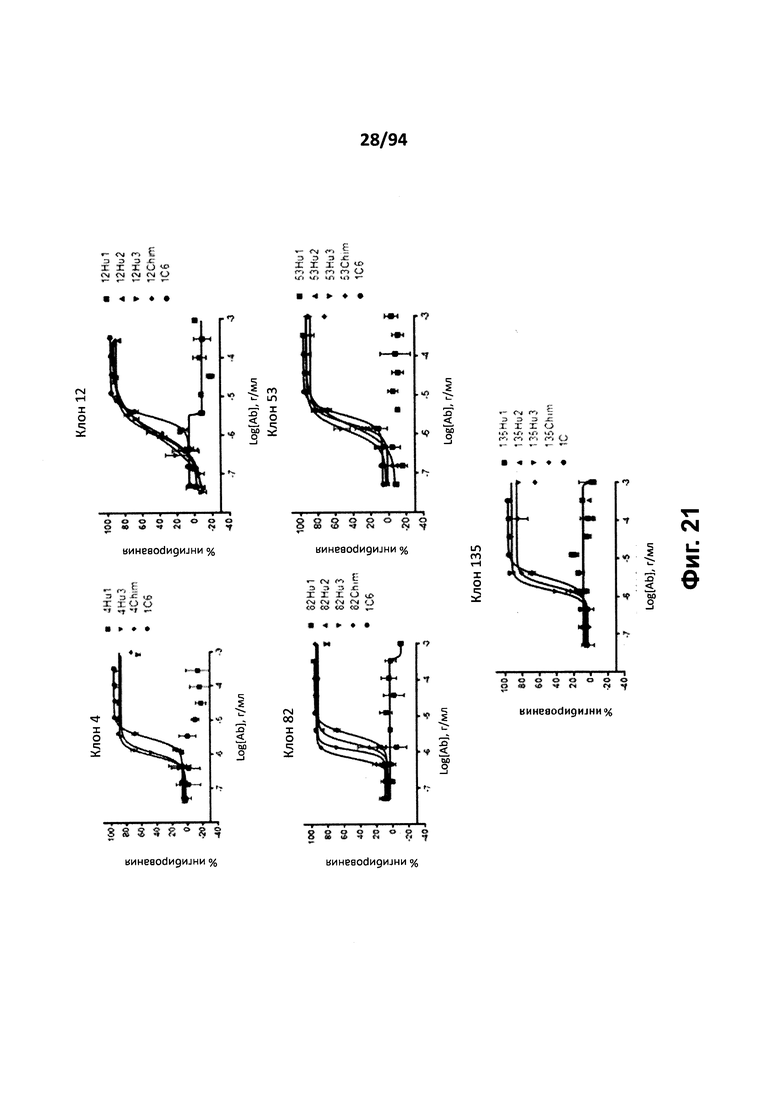
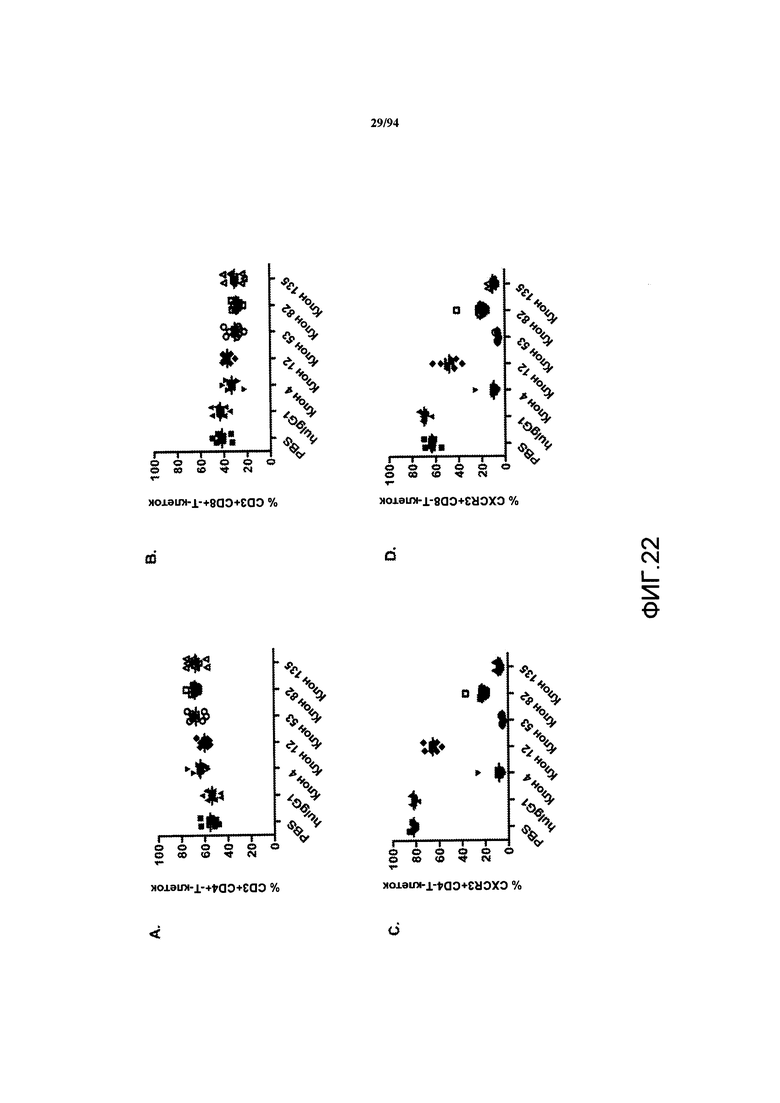

































































Настоящее изобретение относится к иммунологии. Предложены антитело и его антигенсвязывающий фрагмент, способные связываться с CXCR3, а также содержащие их конъюгат и фармацевтическая композиция. Кроме того, описано применение антитела или его фрагмента для предупреждения, лечения или подавления прогрессирования впервые выявленного диабета типа 1 (T1D) и in vitro-способ обнаружения присутствия или определения концентрации CXCR3 в тест-образце. Данное изобретение может найти дальнейшее применение в терапии заболеваний, связанных с CXCR3. 5 н. и 12 з.п. ф-лы, 28 ил., 13 пр., 13 табл.
1. Антитело или его антигенсвязывающий фрагмент, способные связываться с CXCR3, причем указанное антитело или указанный фрагмент включают шесть гипервариабельных областей (CDR): CDR1 вариабельного домена тяжелой цепи (VH), CDR2 VH, CDR3 VH, CDR1 вариабельного домена легкой цепи (VL), CDR2 VL и CDR3 VL, где CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL и VL CDR3 представлены на Фиг. 26A.
2. Антитело или его фрагмент по п. 1, где указанное антитело или его фрагмент являются химерными, CDR-привитыми, мутированными, мутированными для удаления одного или более сайтов дезамидирования, гуманизированными, гуманизированными с обратной мутацией или рекомбинантными.
3. Антитело или его фрагмент по п. 1 или 2, где указанное антитело или его фрагмент способны связываться с полипептидом, содержащим пептид, выбранный из группы, состоящей из:
a) пептида, содержащего остатки 1-58 последовательности MVLEVSDHQVLNDAEVAALLENFSSSYDYGENESDSCCTSPPCPQDFSLNFDRAFLPALYSLLFLLGLLGNGAVAAVLLSRRTALSSTDTFLLHLAVADTLLVLTLPLWAVDAAVQWVFGSGLCKVAGALFNINFYAGALLLACISFDRYLNIVHATQLYRRGPPARVTLTCLAVWGLCLLFALPDFIFLSAHHDERLNATHCQYNFPQVGRTALRVLQLVAGFLLPLLVMAYCYAHILAVLLVSRGQRRLRAMRLVVVVVVAFALCWTPYHLVVLVDILMDLGALARNCGRESRVDVAKSVTSGLGYMHCCLNPLLYAFVGVKFRERMWMLLLRLGCPNQRGLQRQPSSSRRDSSWSETSEASYSGL;
b) пептида, содержащего остатки 1-16 последовательности MVLEVSDHQVLNDAEVAALLENFSSSYDYGENESDSCCTSPPCPQDFSLNFDRAFLPALYSLLFLLGLLGNGAVAAVLLSRRTALSSTDTFLLHLAVADTLLVLTLPLWAVDAAVQWVFGSGLCKVAGALFNINFYAGALLLACISFDRYLNIVHATQLYRRGPPARVTLTCLAVWGLCLLFALPDFIFLSAHHDERLNATHCQYNFPQVGRTALRVLQLVAGFLLPLLVMAYCYAHILAVLLVSRGQRRLRAMRLVVVVVVAFALCWTPYHLVVLVDILMDLGALARNCGRESRVDVAKSVTSGLGYMHCCLNPLLYAFVGVKFRERMWMLLLRLGCPNQRGLQRQPSSSRRDSSWSETSEASYSGL; и
c) пептида, содержащего остатки 1-37 последовательности MVLEVSDHQVLNDAEVAALLENFSSSYDYGENESDSCCTSPPCPQDFSLNFDRAFLPALYSLLFLLGLLGNGAVAAVLLSRRTALSSTDTFLLHLAVADTLLVLTLPLWAVDAAVQWVFGSGLCKVAGALFNINFYAGALLLACISFDRYLNIVHATQLYRRGPPARVTLTCLAVWGLCLLFALPDFIFLSAHHDERLNATHCQYNFPQVGRTALRVLQLVAGFLLPLLVMAYCYAHILAVLLVSRGQRRLRAMRLVVVVVVAFALCWTPYHLVVLVDILMDLGALARNCGRESRVDVAKSVTSGLGYMHCCLNPLLYAFVGVKFRERMWMLLLRLGCPNQRGLQRQPSSSRRDSSWSETSEASYSGL.
4. Антитело или его фрагмент по п. 1 или 2, где указанное антитело или его фрагмент способны связываться с полипептидом, содержащим пептид, выбранный из группы, состоящей из:
a) пептида, содержащего аминокислотную последовательность SDHQVLNDAE;
b) пептида, содержащего аминокислотную последовательность SDHQVLND;
c) пептида, содержащего аминокислотную последовательность DHQVLND;
d) пептида, содержащего аминокислотную последовательность VLNDAE;
e) пептида, содержащего аминокислотную последовательность VLND ;
f) пептида, содержащего аминокислотную последовательность XDXXVXNDXX;
g) пептида, содержащего аминокислотную последовательность XDXXVXND;
h) пептида, содержащего аминокислотную последовательность DXXVXND;
i) пептида, содержащего аминокислотную последовательность VXNDXX; и
j) пептида, содержащего аминокислотную последовательность VXND,
где каждый X независимо означает любую аминокислоту.
5. Антитело или его фрагмент по п. 1 или 2, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где указанная вариабельная область тяжелой цепи содержит последовательность, которая по меньшей мере приблизительно на 85%, 90%, 95% или 99% идентична последовательности, выбранной из группы, состоящей из последовательностей, представленных на Фиг. 26А-26G и 26N-26Q; и где вариабельная область легкой цепи содержит последовательность, которая по меньшей мере приблизительно на 85%, 90%, 95% или 99% идентична последовательности, выбранной из группы, состоящей из последовательностей, представленных на Фиг. 26А-26D, 26H-26M и 26R-26U.
6. Антитело или его фрагмент по п. 1 или 2, где вариабельная область тяжелой цепи содержит последовательность, выбранную из группы, состоящей из последовательностей, представленных на Фиг. 26A-26G и 26N-26Q; и где вариабельная область легкой цепи содержит последовательность, выбранную из группы, состоящей из последовательностей, представленных на Фиг. 26А-26D, 26H-26M и 26R-26U.
7. Антитело или его фрагмент по п. 1 или 2, где указанное антитело или его фрагмент содержат комбинацию вариабельных доменов тяжелой цепи и легкой цепи, выбранных из группы, состоящей из последовательностей VH и VL, представленных на Фиг. 26A, последовательностей VH и VL, представленных на Фиг. 26B, последовательностей VH и VL, представленных на Фиг. 26C, последовательностей VH и VL, представленных на Фиг. 26D, последовательности VH, представленной на Фиг. 26B, и последовательности VL, представленной на Фиг.26C, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26B, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26H, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26I, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26J, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26K, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26L, последовательности VH, представленной на Фиг. 26C, и последовательности VL, представленной на Фиг. 26M, последовательности VH, представленной на Фиг. 26E, и последовательности VL, представленной на Фиг. 26C, последовательности VH, представленной на Фиг. 26F, и последовательности VL, представленной на Фиг. 26C, последовательности VH, представленной на Фиг. 26G, и последовательности VL, представленной на Фиг. 26C, последовательности VH, представленной на Фиг. 26B, и последовательности VL, представленной на Фиг. 26H, последовательности VH, представленной на Фиг. 26B, и последовательности VL, представленной на Фиг. 26J, последовательности VH, представленной на Фиг. 26G, и последовательности VL, представленной на Фиг. 26H, последовательности VH, представленной на Фиг. 26G, и последовательности VL, представленной на Фиг. 26J, последовательности VH, представленной на Фиг. 26N, и последовательности VL, представленной на Фиг. 26R, и последовательности VH, представленной на Фиг. 26N, и последовательности VL, представленной на Фиг. 26S.
8. Антитело или его фрагмент по п. 1 или 2, где указанное антитело или его фрагмент способны связываться преимущественно с А-изоформой CXCR3.
9. Применение антитела или его фрагмента по любому из пп. 1-8 для предупреждения, лечения или подавления прогрессирования впервые выявленного диабета типа 1 (T1D).
10. Применение по п. 9, где указанное антитело или его фрагмент вводят в дозе, составляющей приблизительно 0,03-3,7 мг/кг/дозу.
11. Применение по п. 9 или 10, где антитело или его фрагмент во всех случаях вводят в общей дозе, составляющей приблизительно 0,16-18 мг/кг.
12. Применение по п. 9 или 10, где указанное антитело или его фрагмент вводят человеку, страдающему впервые выявленным диабетом и имеющему базальный уровень С-пептида в сыворотке, превышающий или равный приблизительно 0,2 нмоль/л, и/или имеющего общий уровень С-пептида в сыворотке натощак во время С-пептидной стимуляции приблизительно 0,033-1,0 нмоль/л в минуту.
13. Конъюгат для предупреждения, лечения или подавления прогрессирования впервые выявленного диабета типа 1 (T1D), включающий антитело или его фрагмент по любому из пп. 1-8, и по меньшей мере один дополнительный агент, где указанным дополнительным агентом является терапевтический агент, солюбилизатор, стабилизатор, иммунодепрессант, рецептор или антигенсвязывающий пептид.
14. Фармацевтическая композиция для предупреждения, лечения или подавления прогрессирования впервые выявленного диабета типа 1 (T1D), содержащая в эффективном количестве антитело или его фрагмент по любому из пп.1-8 и фармацевтически приемлемый носитель.
15. Фармацевтическая композиция по п. 14, где композиция дополнительно содержит по меньшей мере один дополнительный терапевтический агент.
16. Фармацевтическая композиция по п. 14, где указанная композиция также содержит по меньшей мере один дополнительный терапевтический агент, выбранный из группы, состоящей из агента, стимулирующего β-клетки, инсулина и инсулинпродуцирующих клеток.
17. In vitro-способ обнаружения присутствия или определения концентрации CXCR3 в тест-образце, где указанный способ включает контактирование указанного тест-образца с антителом или его фрагментом по любому из пп.1-8 и с детектируемой меткой, где присутствие CXCR3 или его концентрация прямо или обратно коррелируют с сигналом, генерируемым детектируемой меткой.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| CLARK-LEWIS I | |||
| et al., "Structure-Function Relationship between the Human Chemokine Receptor CXCR3 and Its Ligands", J Biol Chem., 2003, 278(1):289-95 | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| US 6686175 B1, 03.02.2004 | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |

