По данной заявке испрашивается приоритет по предварительной заявке США номер 60/650866, поданной 8 февраля 2005 года, которая включена в настоящее описание в качестве ссылки в полном объеме.
Правительство США обладает правами на настоящее изобретения в соответствии с грантом номер DK 26741 от NIDDK.
I. ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится, главным образом, к способу и композициям, связанным с молекулярной биологией, неврологией и эндокринологией. в определенных аспектах оно относится к композициям, содержащим растворимые сопряженные с G-белком рецепторы (sGPCR) и к способам их применения в качестве модуляторов активности GPCR и/или модуляторов фармакологических эффектов лигандов, которые связывают такие растворимые GPCR.
II. ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Рецепторы, как правило, представляют собой молекулярные структуры, расположенные в клеточной мембране или внутри клетки, которые образуют слабую обратимую связь с веществом, таким как антиген, гормон или нейротрансмиттер. Каждый рецептор предназначен для связывания с конкретным веществом(ами). Конкретное семейство рецепторов представляет собой рецептор с семью трансмембранными участками ("7TM") или сопряженный с G-белком рецептор ("GPCR"). Эти рецепторы связаны со связывающим гуаниновые нуклеотиды G-белком ("G-белок") для передачи сигнала при связывании соответствующего вещества с рецептором. Когда G-белок связан гуаниндифосфатом ("GDP"), то G-белок является неактивным, или "выключенным". Аналогично, если G-белок связан с гуанинтрифосфатом ("GTP"), то G-белок является активным, или "включенным", опосредуя тем самым активацию биологического ответа в клетке.
GPCR обладают общим структурным мотивом. Все эти рецепторы имеют семь последовательностей из от 22 до 24 гидрофобных аминокислот, которые формируют семь альфа-спиралей, каждая из которых пронизывает мембрану (т.е. трансмембранный участок-1 (TM-1), трансмембранный участок-2 (TM-2), и т.д.). Трансмембранные спирали соединены аминокислотными цепями между трансмембранным участком-2 и трансмембранным участком-3, трансмембранным участком-4 и трансмембранным участком-5, и трансмембранным участком-6 и трансмембранным участком-7 на наружной, или на "внеклеточной", стороне клеточной мембраны (их называют "внеклеточными петлями" или "внеклеточными" участками). Трансмембранные спирали также соединены аминокислотными цепями между трансмембранным участком-1 и трансмембранным участком-2, трансмембранным участком-3 и трансмембранным участком-4, и трансмембранным участком-5 и трансмембранным участком-6 на внутренней, или "внутриклеточной", стороне клеточной мембраны (их называют "внутриклеточными петлями" или "внутриклеточными" участками). "Карбокси" ("C") конец рецептора расположен во внутриклеточном пространстве в клетке, и "амино" ("N") конец рецептора расположен во внеклеточном пространстве снаружи клетки.
Как правило, когда лиганд связывается с рецептором и "активирует" рецептор, происходит изменение конформации внутриклеточного участка, которое делает возможным связывание между внутриклеточным участком и внутриклеточным "G-белком". Было описано, что GPCR являются "неразборчивыми" в отношении G-белов, т.е., что GPCR может взаимодействовать более чем с одним G-белком (Kenakin, 1988). Несмотря на то, что существуют другие G-белки, к настоящему времени выявлены G-белки Gq, Gs, Gi и Go. Активированные лигандом GPCR, связанные с G-белком, начинают каскадный процесс передачи сигнала или трансдукцию сигнала. При нормальных условиях трансдукция сигнала в конечном итоге приводит к активации клетки или к ингибированию клетки. Полагают, что с G-белком взаимодействует третья внутриклеточная петля (IC-3), а также C-конец рецептора.
Как правило, активность практически каждой клетки в организме регулируется посредством внеклеточных сигналов. В ряде физиологических процессов у человека, а также в широким диапазоне организмов, используется опосредуемая белком трансмембранная передача сигналов через GPCR. Сигналы от конкретного GPCR приводят к активации G-белка в клетке. Большинство сигналов посредством GPCR передается внутрь клетки. Существует множество различных аспектов процесса передачи сигнала, в который вовлечены разнообразные подтипы рецепторов GPCR и связанные с ними соответствующие G-белки, а также множество связанных с ними внутриклеточных вторичных посредников. Трансдукция сигнала может приводить к общей или частичной активации или инактивации внутриклеточного процесса или процессов, в зависимости от вовлеченных в нее белков. Существенные молекулы для передачи сигнала или нейтротрансмиттеры, которые связываются с GPCR, включают в себя, но не ограничиваются ими кортикотропин-рилизинг фактор, паратиреоидный гормон, морфин, допамин, гистамин, 5-гидрокситриптамин, аденозин, кальцитонин, гастроингибиторный пептид (GIP), глюкагон, рилизинг-фактор гормона роста (GHRH),
паратиреоидный гормон (PTH), PACAP, секретин, вазоактивный интестинальный полипептид (VIP), диуретический гормон, EMR1, латрофилин, специфичный для головного мозга ингибитор ангиогенеза (BAI), кадгерин, EGF, LAG, (CELSR) и другие подобные белки или молекулы.
GPCR формируют суперсемейство белков. В настоящее время существует свыше 2000 описанных в литературе GPCR, которые подразделяются по меньшей мере на три семейства: подобное родопсину семейство (семейство A), рецепторы для кальцитонина (семейство B) и метаботропное семейство для глутамата (семейство C) (Ji et al, 1998). Описанные GPCR включают в себя как охарактеризованные рецепторы, так и одиночные рецепторы, для которых лиганды еще не выявлены. (Wilson et al, 1999; Wilson et al, 1998; Marchese et al, 1999). Несмотря на большое количество GPCR, все GPCR, как правило, обладают сходной молекулярной структурой. Каждый GPCR содержит цепь из аминокислотных остатков различной длины. GPCR расположены в клеточной мембране в виде семи отдельных спиралей, называемых трансмембранными участками. N-конец GPCR расположен снаружи клетки с внеклеточными петлями, в то время как C-конец расположен внутри клетки с внутриклеточными петлями.
Лиганды для GPCR включают в себя низкомолекулярные молекулы, а также пептиды и небольшие белки. Взаимодействие между этими лигандами и их рецепторами варьирует от системы к системе, однако для него может быть необходимым взаимодействие с остатками в нескольких из четырех внеклеточных доменов и на N-конце. В некоторых случаях N-конец один может сохранять способность к взаимодействию и связыванию с лигандами. GPCR и известные лиганды ассоциированы со множеством заболеваний, в том числе с рассеянным склерозом, диабетом, ревматоидным артритом, астмой, аллергией, воспалительным заболеванием кишечника, некоторыми злокачественными опухолями, нарушениями щитовидной железы, заболеванием сердца, пигментным ретинитом, ожирением, неврологическими нарушениями, остеопорозом, инфекцией вирусом иммунодефицита человека ("HIV") и синдромом приобретенного иммунодефицита ("СПИД") (Murphy et al, 2000; Mannstadt et al, 1999; Berger et al, 1999; Jacobson et al, 1997; Meij, 1996).
Таким образом, в данной области существует необходимость в способах продукции модуляторов GPCR и лигандов, которые связывают GPCR для применения в качестве лекарственных средств. Эти лекарственные средства можно использовать для профилактики или лечения ассоциированных с GPCR заболеваний и/или нарушений.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям и способам, связанным со связывающими лиганд доменами sGPCR, а также с эффектами sGPCR на передачу сигнала GPCR и на взаимодействие между лигандами GPCR и их GPCR.
Вариант осуществления этого изобретения включает в себя выделенный связывающий лиганд домен растворимого сопряженного с G-белком рецептора (sGPCR). sGPCR содержит весь внеклеточный домен GPCR или его часть. В одном аспекте этого изобретения sGPCR представляет собой растворимую форму члена семейства B GPCR. В следующем аспекте sGPCR представляет собой член подсемейства B1 GPCR. В следующих аспектах sGPCR представляет собой растворимый рецептор для секретина, рецептор для VPAC1, рецептор для VPAC2, рецептор для PAC1, рецептор для глюкагона, рецептор для рилизинг-фактора гормона роста (GHRH), рецептор для глюкагон-подобного пептида 1 (GLP-1), рецептор для глюкагон-подобного пептида 2 (GLP-2), рецептор для гастроингибиторного пептида (GIP), рецептор для кортикотропин-рилизинг фактора 1 (CRF1), рецептор для кортикотропин-рилизинг фактора 2 (CRF2), рецептор для паратиреоидного гормона 1 (PTH1), рецептор для паратиреоидного гормона 2 (PTH2), подобный рецептору для кальцитонина рецептор или рецептор для кальцитонина. sGPCR может представлять собой растворимый рецептор для PTH1 или рецептор для PTH2. Вариант осуществления этого изобретения также включает в себя sGPCR, который представляет собой растворимую форму рецептора для кортикотропин-рилизинг фактора типа 2α (sCRFR2α). Аминокислотная последовательность sCRFR2α может содержать аминокислотную последовательность, кодируемую экзонами 3, 4 и 5 гена CRFR2α или она не содержит экзон 6 или выше. Рекомбинантный sGPCR по этому изобретению может включать в себя 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 130, 135, 140, 150, 155, 160, 180, 200 или более аминокислот, включая все диапазоны среди этих значений, внеклеточного домена(ов) GPCR, включая весь N-концевой внеклеточный домен или его часть. В определенных аспектах связывающий лиганд домен sGPCR может содержать аминокислотную последовательность по меньшей мере на 70, 75, 80, 85, 90, 95, или 98% сходную с 50, 75, 100, 125, 150 или более аминокислотами SEQ ID NO:4 (sCRFR2α), SEQ ID NO:8 (sCRFR2β), SEQ ID NO:12 (sCRFR2γ) или SEQ ID NO:15 (mCRFR2α). В следующем аспекте sCRFR содержит аминокислотную последовательность SEQ ID NO:4, 8, 12, 15 или их сочетание. В следующем аспекте изобретение относится к выделенному sGPCR, детектируемую аффинный маркер, метку, поддающуюся определению или терапевтическую химическую группу, метку биотин/авидин, радионуклид, детектируемый или терапевтический фермент, флуоресцентный маркер, хемилюминесцентный маркер, домен иммуноглобулина или любое их сочетание. В одном аспекте GPCR содержит домен иммуноглобулина, в частности Fc-домен. sGPCR может быть конъюгирован с полимером, который включает в себя, но не ограничивается этим, полиэтиленгликоль (PEG).
Варианты осуществления этого изобретения включают в себя полинуклеотиды, кодирующие sGPCR по этому изобретению. Полинуклеотид может дополнительно содержать промотор, функционально связанный с полинуклеотидом, кодирующим sGPCR. Кодирующая sGPCR последовательность может быть встроена в экспрессирующую кассету. Экспрессирующая кассета может находится в экспрессирующем векторе. Экспрессирующий вектор может включать в себя, но не ограничивается ими, линейную нуклеиновую кислоту, плазмидный экспрессирующий вектор или вирусный экспрессирующий вектор. В определенных аспектах экспрессирующий вектор находится в векторе для доставки, который может включать в себя, но не ограничиваться ими, липосому, полипептид, поликатион, липид, бактерию или вирус.
Следующие варианты осуществления этого изобретения включают в себя способы модулирования активности сопряженного с G-белком рецептора (GPCR), включающие в себя: a) контактирование ткани-мишени с sGPCR; и b) связывание лиганда GPCR вблизи ткани-мишени, где в ткани модулируется активность GPCR. Лиганд может представлять собой лиганд семейства B GPCR, лиганд подсемейства B1 GPCR. В определенных аспектах лиганд представляет собой кортикотропин-рилизинг фактор (CRF), урокортин 1, урокортин 2, урокортин 3, стресскопин, паратиреоидный гормон, PTH-подобный гормон, TIP39, кальцитонин, амилин, CGRP (CALCA и CALCB),
адреномедуллин, секретин, VIP, PACAP, глюкагон, GHRH, GLP-1, GLP-2, GIP или любое их сочетание.
Способы также могут включать в себя контактирование с тканью-мишенью, включающее в себя стадии: a) получения связывающего лиганд домена sGPCR в виде соответствующего фармацевтического раствора; и b) введения фармацевтического раствора животному, человеку, субъекту и/или пациенту в количестве, необходимом для влияния на связывание лиганда-мишени в ткани-мишени животного. Введение может представлять собой, но не ограничиваться этим, пероральное введение, инъекцию, эндоскопическое введение или перфузию. Инъекция включает в себя, но не ограничивается этим, внутривенную, внутримышечную, подкожную, внутрикожную, внутричерепную или внутрибрюшинную инъекцию. Нарушения, которые можно лечить, ослаблять, модулировать, тяжесть которых можно снижать, включают в себя нарушения, возникающие вследствие гиперактивации GPCR или гиперсекреции лиганда GPCR. В определенных аспектах нарушение представляет собой чувствительность к инсулину или диабет II типа. Нарушение также может включать в себя связанное с тревогой нарушение; нарушение настроения; нарушение, связанное с посттравматическим стрессом; надъядерный паралич; подавление иммунной системы; симптомы отмены лекарственного средства или алкоголя; воспалительные нарушения; боль; астму; псориаз и аллергию; фобии; нарушения сна, вызванные стрессом; фибромиалгию; дистемию; биполярные нарушения; циклотимию; синдром усталости; индуцированную стрессом головную боль; злокачественную опухоль; инфекции вирусом иммунодефицита человека; нейродегенеративные заболевания; желудочно-кишечные заболевания; нарушения питания; геморрагический стресс; индуцированные стрессом психотические эпизоды; эутиреоидный синдром тошноты; синдром неадекватной продукции антидиарейного гормона; ожирение; бесплодие; травмы головы; травму спинного мозга; ишемическое нейрональное повреждение; эксайтоксическое нейрональное повреждение; эпилепсию; сердечно-сосудистые и связанные с сердцем нарушения; дисфункции иммунной системы; мышечные спазмы; недержание мочи; сенильную деменцию по типу болезни Альцгеймера; мультиинфарктную деменцию; боковой амиотрофический склероз; зависимость и привыкание к химическим веществам; психосоциальную карликовость, гиперчувствительность или гипочувствительность к инсулину, гипогликемию, нарушения кожи; или потерю волос. В определенных аспектах нарушение представляет собой связанное с тревогой нарушение; нарушение настроения; биполярное нарушение; нарушение, связанное с посттравматическим стрессом; воспалительное нарушение; зависимость и привыкание к химическим веществам; желудочно-кишечное нарушение; или нарушение кожи. В следующем аспекте связанное с тревогой нарушение представляет собой генерализованную тревогу, или нарушение настроения представляет собой депрессию. В следующем аспекте желудочно-кишечное нарушение представляет собой синдром раздраженного кишечника.
Другие варианты осуществления этого изобретения включают в себя способы выявления лиганда GPCR, включающие в себя: a) контактирование образца, предположительно содержащего лиганд GPCR, с полипептидом sGPCR; и b) оценку наличия или отсутствия связанного с полипептидом sGPCR лиганда. Способы могут дополнительно включать в себя охарактеризовывание связанного лиганда. Охарактеризовывание связанного лиганда включает в себя, но не ограничивается этим, различные способы хроматографии, масс-спектрометрию, секвенирование пептидов и т.п. Полипептид sGPCR может быть функционально связанным с субстратом или поверхностью или может быть не связанным с ними. Способ может дополнительно включать в себя: c) введение радиоактивномеченного лиганда GPCR; и d) оценку связывания или конкуренции за связывание радиоактивномеченного лиганда GPCR с GPCR. Лиганд GPCR может включать в себя, но не ограничиваться ими, кортикотропин-рилизинг фактор (CRF), урокортин 1, урокортин 2, урокортин 3, паратиреоидный гормон, PTH-подобный гормон, TIP39, кальцитонин, амилин, CGRP (CALCA и CALCB), адреномедуллин, секретин, VIP, PACAP, глюкагон, GHRH, GLP-1, GLP-2 или GIP.
Другие варианты осуществления включают в себя способы выявления sGPCR, включающие в себя: a) контактирование образца, предположительно содержащего sGPCR с лигандом, который связывает sGPCR или относящийся к нему связанный с поверхностью GPCR; и b) оценку связывания лиганда GPCR с компонентами образца. Кроме того, способ может включать в себя охарактеризовывание связанного sGPCR, которое может включать в себя хроматографию, масс-спектрометрию, фрагментацию и секвенирование белка и т.п. Лиганд GPCR может быть функционально связан с субстратом или с поверхностью. Кроме того, способы могут включать в себя: c) введение радиоактивномеченного sGPCR; и d) оценку связывания или конкуренции за связывание радиоактивномеченного sGPCR с лигандом GPCR в присутствии или в отсутствии подвергаемого тестированию образца. Иллюстративные лиганды включают в себя кортикотропин-рилизинг фактор (CRF), урокортин 1, урокортин 2, урокортин 3, паратиреоидный гормон, PTH-подобный гормон, TIP39, кальцитонин, амилин, CGRP (CALCA и CALCB), адреномедуллин, секретин, VIP, PACAP, глюкагон, GHRH, GLP-1, GLP-2, GIP или другие известные лиганды GPCR.
Другой вариант осуществления этого изобретения включает в себя антитела, которые специфично связывают sGPCR. В определенных аспектах антитело может связывать N-конец или С-конец sGPCR. Аспекты этого изобретения включают в себя антитело, которое связывает C-концевую последовательность из 5, 10, 15, 20 или более аминокислот, которая может происходить из альтернативной рамки считывания нуклеотидной последовательности, которая кодирует трансмембранный участок GPCR (как правило, она представляет собой результат альтернативного сплайсинга, и ее можно конструировать с получением рекомбинантного полинуклеотида по этому изобретению).
Варианты осуществления этого изобретения включают в себя способы выявления экспрессии sGPCR с использованием анализа или оценки либо белка, либо нуклеиновой кислоты, либо и белка, и нуклеиновой кислоты. Аспекты этого изобретения включают в себя способы выявления мРНК sGPCR, включающие в себя: a) получение образца нуклеиновой кислоты для анализа; и b) оценку наличия нуклеиновой кислоты sGPCR, включающей в себя точки сплайсинга, приводящие к sGPCR. Способ может включат в себя оценку наличия конкретных видов мРНК посредством гибридизации нуклеиновых кислот, амплификации нуклеиновых кислот или других способов анализа нуклеиновых кислот. В конкретном аспекте sGPCR представляет собой растворимый GPCR типа B, растворимый GPCR типа B1, растворимый CRFR, sCRFR1, sCRFR2 или sCRFR2α. Полинуклеотид может включать в себя область соединения экзон/экзон, которая включает в себя N-концевые аминокислоты GPCR и не включает в себя экзон, кодирующий участок трансмембранного домена или включает в себя его часть. В конкретном аспекте точка сплайсинга sCRFR2α представляет собой область соединения экзон 5/экзон 7, где в основе обозначения экзона лежит геномное обозначение экзонов CRFR2. Исходя из транскрипта CRFR2a, экзоны были бы обозначены 3 и 5 соответственно (например, см. фиг.1).
"Растворимый" GPCR (sGPCR) означает GPCR, который содержит весь внеклеточный домен рецептора или его часть, но лишен всех или части из одного или нескольких трансмембранных доменов, которые в норме удерживают полноразмерный рецептор в клеточной мембране, и растворимая форма не интегрирована в клеточную мембрану. Таким образом, например, когда такой растворимый рецептор продуцируют рекомбинантным способом в клетке млекопитающего, он может секретироваться из рекомбинантной клетки-хозяина через плазматическую мембрану, а не оставаться на поверхности клетки. Как правило, растворимый рецептор по этому изобретению является растворимым в водном растворе. Однако в определенных условиях рецептор может находиться в форме внутриклеточного тельца, которое является легко растворимым стандартными способами. Такой sGPCR можно получать из сконструированной нуклеиновой кислоты, процессированного белка (например, подвергнутого протеолизу белка), синтезированного белка, или выделенного варианта по сплайсингу. Полинуклеотид, кодирующий такой sGPCR, можно выделять или получать способами инженерии.
Как используют в настоящем описании, термины "выделенный" и "очищенный" используют взаимозаменяемо и они относятся к нуклеиновым кислотам или полипептидам или их биологически активным участкам, в которых по существу или практически отсутствуют компоненты, которые в норме находятся вместе с нуклеиновой кислотой или полипептидом, или взаимодействуют с ними, в их природных окружающих условиях. Таким образом, выделенная или очищенная нуклеиновая кислота или полипептид по существу не содержит других клеточных компонентов или культуральной среды при продукции рекомбинантными способами, или по существу не содержит химических предшественников или других химических веществ в случае химического синтеза.
"Выделенная" нуклеиновая кислота не содержит последовательностей (предпочтительно кодирующих белок последовательностей), которые в природе фланкируют нуклеиновую кислоту (т.е., последовательностей, расположенных на 5'- и 3'-концах нуклеиновой кислоты) в геномной ДНК организма, являющегося источником нуклеиновой кислоты. Например, в различных вариантах осуществления, выделенные нуклеиновые кислоты могут содержать менее чем приблизительно 5 т.п.н., 4 т.п.н., 3 т.п.н., 2 т.п.н., 1 т.п.н., 0,5 т.п.н. или 0,1 т.п.н. нуклеотидных последовательностей, которые в природе фланкируют нуклеиновые кислоты в геномной ДНК клетки, являющейся источником нуклеиновой кислоты.
Как используют в настоящем описании, термин "выделенный" или "очищенный", когда его применяют в отношении полипептида по этому изобретению, означает, что выделенный белок по существу не содержит клеточных компонентов, и включает в себя препараты белка, обладающие менее чем приблизительно 30%, 20%, 10%, 5% или менее (от сухой массы) белковых примесей. В случае, когда белок по этому изобретению или его биологически активный участок продуцируют рекомбинантными способами, в культуральной среде предпочтительно представлено менее чем приблизительно 30%, 20%, 10% или 5% (от сухой массы) химических предшественников или химических веществ, не являющихся представляющим интерес белком.
Применение формы единственного числа, при использовании совместно с термином "содержащий" в формуле изобретения и/или описании, может означать "один", но также оно соответствует значению "один или несколько", "по меньшей мере один" и "один или более одного".
Применение термина "или" в формуле изобретения используют для обозначения "и/или", если явным образом не указано, что он относится только к альтернативам, или что альтернативы являются взаимоисключающими, хотя описание предполагает определение, которое относится только к альтернативам и "и/или".
Другие цели, свойства и преимущества настоящего изобретения будут очевидны из следующего ниже подробного описания. Однако следует понимать, что подробное описание и конкретные примеры, хотя и демонстрируют конкретные варианты осуществления этого изобретения, приведены только в качестве иллюстрации, поскольку различные изменения и модификации сущности и объема этого изобретения будут очевидными специалистам в данной области из этого подробного описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Для того чтобы сущность приведенных выше свойств, преимуществ и целей этого изобретения, а также прочее, которые будут понятны, достигались и были понятны в деталях, более конкретные описания и определенные варианты осуществления этого изобретения, кратко обобщенные выше, проиллюстрированы на прилагаемых чертежах. Эти чертежи являются частью описания. Однако следует отметить, что прилагаемые чертежи иллюстрируют определенные варианты осуществления этого изобретения и таким образом их не следует рассматривать как ограничивающие его объем.
Фиг.1A-1B. Представлена иллюстративная последовательность нуклеотидов и транслируемых аминокислот растворимого GPCR, рецептора для CRF типа 2α (sCRFR2α) (фиг.1A). Подчеркнутыми аминокислотами указан уникальный C-конец. Заключенными в прямоугольники остатками указаны предполагаемые участки для N-гликозилирования. Представлено схематическое изображение структуры гена CRFR2 мыши (верхние панели), два известных функциональных транскрипта у мыши, α и β (средние панели) и новый вариант сплайсинга sCRFR2α (нижняя панель) (фиг.1B). Указаны расположения участков начала трансляции (ATG). Указаны экзоны, кодирующие N-концевой внеклеточный домен (ECD), семь трансмембранных доменов (7TM) и C-концевой цитоплазматический домен (CD). 5'- и 3'-UTR указаны посредством заштрихованных прямоугольников. Черными прямоугольниками показаны кодирующие участки, и пустыми прямоугольниками показаны экзоны, расположенные ниже стоп-кодона.
Фиг.2A-2C. Представлена экспрессия мРНК CRFR2α и sCRFR2α в головном мозге и гипофизе мыши. На фиг.2A представлено схематическое изображение и расположение олигонуклеотидных праймеров для амплифицированного фрагмента транскриптов CRFR2α мыши (верхняя панель) и sCRFR2α (нижняя панель). Указано расположение олигонуклеотидных праймеров в третьем и седьмом экзонах, которое приводит к амплификации продуктов из 418 и 309, соответствующих CRFR2α и sCRFR2α, соответственно. Фиг.2B представляет собой иллюстративное изображение электрофоретического анализа полуколичественной RT-PCR для мРНК mCRFR2α и sCRFR2α и мРНК рибосомального белка S16 (верхние панели). Также проводили саузерн-блот гибридизацию амплифицированной кДНК mCRFR2α и sCRFR2α и фрагментов кДНК рибосомального белка S16 (нижние панели). Количественное определение для радиоактивных полос проводили посредством PhosphorImager, и нормализованные значения (относительно экспрессии S16) представлены в качестве относительных единиц денситометрии (фиг.2C).
Фиг.3A-3C. Высокоспецифичную антисыворотку, индуцированную у кроликов с использованием синтетического пептидного фрагмента, кодирующего уникальный C-конец белка sCRFR2α (aa 113-143), использовали для проведения радиоиммунного анализа sCRFR2α, использовали для иммуноблоттинга и для иммуноцитохимии. Фиг.3A представляет собой вестерн-иммуноблот sCRFR2α мыши, выделенного из среды клеток COS-M6, временно трансфицированных конструкцией sCRFR2αFLAG, прореагировавшей с сывороткой против sCRFR2α (113-143) (левая панель) или моноклональными M2 против FLAG (правая панель). Дорожки 1, 2 и 3 соответствуют 0,1, 1,0 и 10 мкл экстракта sCRFR2α-FLAG соответственно.
На фиг.3B проиллюстрировано вытеснение [125I]Tyr113 sCRFR2α (а.к. 113-143) из связывания с антителами против sCRFR2α кролика (а.к. 113-143) посредством синтетического sCRFR2α (а.к. 113-143) и посредством очищенного sCRFR2α (а.к. 113-143)-FLAG, экспрессированного COS-M6. На фиг.3C - иммунофлуоресцентное окрашивание клеток COS-M6, временно трансфицированных конструкцией sCRFR2α мыши, визуализированное посредством сыворотки против sCRFR2α (а.к. 113-143), а затем посредством конъюгированного с Cy3 вторичного антитела (фиг.3C(b)). Препараты контрастно окрашивали DAPI для визуализации как трансфицированных, так и нетрансфицированных клеток (фиг.3C(a)). Для клеток, инкубированных с нормальной сывороткой кролика (NRS) в качестве отрицательного контроля, а затем с конъюгированным с Cy3 вторичным антителом, не было показано какого-либо окрашивания (фиг.3C(c)).
Фиг.4A-4G. Представлено наличие подобной sCRFR2α иммунореактивности (ir) в головном мозге мыши с использованием иммуногистохимии и радиоиммунного анализа (RIA). На фиг.4A-4F представлено окрашивание иммунопероксидазой на sCRFR2α в отдельных участках головного мозга мыши. Основные участки клеточной экспрессии включали в себя основные выходные нейроны обонятельной луковицы (фиг.4A); медиальное ядро септума (фиг.4B); и базолатеральное (BLA), но не центральное (CeA) ядро миндалевидного тела (фиг.4C); кору головного мозга, где окрашенные клетки были расположены, главным образом, в слоях 5 и 2/3 (фиг.4D); и красное ядро (фиг.4E). В каждом из этих участков характер мечения клеток был сходным, хотя не обязательно идентичным, с характером мечения для экспрессии мРНК CRFR1. Иммуномеченые волокна и утолщения были ограничены рядом групп клеток, включая паравентрикулярное ядро гипоталамуса (PVH; фиг.4F). На фиг.4G подобную sCRFR2α иммунореактивность экстрагированной кислотой и частично очищенной ткани, выделенной из головного мозга мыши, измеряли посредством радиоиммунного анализа. Экстракты ткани тестировали при 5-7 уровнях дозы и проводили вытеснение связывания sCRFR2α (а.к. 113-143), меченого [125I] Tyr113, с антителом против sCRFR2α кролика (а.к. 113-143) зависимым от дозы образом.
Фиг.5A-5B. Белок sCRFR2α препятствует индукции передачи сигнала cAMP и MAPK, опосредуемой Ucn 1 или CRF. На фиг.5A представлена активация репортера CRE-люциферазы посредством Ucn 1 или CRF, с предварительной инкубацией с sCRFR2α или без нее, в клетках 293T, временно трансфицированных CRFR2α мыши. Люциферазный репортер, содержащий фрагмент промотора CRE гена EVX1, совместно трансфицировали в клетки 293T посредством экспрессирующих CRFR2α векторов. Активность люциферазы измеряли после обработки (4 ч) 0,0001-100 нМ Ucn 1 или CRF в присутствии 0,1 нМ sCRFR2α или в его отсутствии. Данные нормализовали относительно активности котрансфицированной β-галактозидазы. Иллюстрированный способ для шести повторений одного эксперимента представлен на графике. Фиг.5B. Уравновешенные клетки CATHa обрабатывали Ucn 1 (10 нМ) с sCRFR2α (0,4 или 4 нМ) или без него. Через 5 мин после стимуляции рецепторов лизаты клеток собирали и подвергали иммуноблот-анализу SDS-PAGE с использованием антитела фосфо-ERK1/2-p42,44 и антитела ERK2-p44. Активацию ERK вычисляли посредством нормализации уровней фосфорилированной ERK1/2-p42,44 относительно общего уровня ERK2-p44. Репрезентативный способ для трех повторений одного эксперимента представлен на графике. *, P<0,05 для обработки.#, P<0,05 Ucn 1, UD = не определено.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Пригодные терапевтические подходы для лечения заболеваний, связанных с GPCR, и связанными с ними каскадами передачи сигнала, включают в себя ингибирование или модулирование активации или ингибирования GPCR. Одним подходом является получение низкомолекулярных ингибиторов, разработка и внедрение на рынок которых являются дорогостоящими. Недостатком лечения низкомолекулярными ингибиторами или антагонистами GPCR является риск токсичности, особенно при повторном применении. К тому же, многие GPCR не имеют низкомолекулярных антагонистов рецептора. Является целесообразным получение антагониста GPCR, который является менее дорогостоящим и/или менее токсичным, чем низкомолекулярные ингибиторы. Варианты осуществления этого изобретения относятся к композициям и способам, связанным со связывающими лиганд доменами растворимого GPCR (sGPCR), а также с их эффектами на передачу сигнала GPCR и взаимодействие между лигандами GPCR и их GPCR. sGPCR можно использовать для того, чтобы вызывать антагонизм активации или ингибирования GPCR in vitro и/или in vivo.
I. СОПРЯЖЕННЫЕ С G-БЕЛКОМ РЕЦЕПТОРЫ (GPCR)
GPCR составляют суперсемейство белков, которое разделяется на три семейства: подобное родопсину семейство (семейство A), рецепторы для кальцитонина (семейство B) и метаботропное семейство для глутамата (семейство C) (Ji et al, 1998), каждое из которых можно далее разделить на подсемейства. Описанные GPCR включают в себя как охарактеризованные рецепторы, так и одиночные рецепторы, для которых лиганды еще не выявлены (Wilson et al, 1999; Wilson et al, 1998; Marchese et al, 1999). Несмотря на большое количество GPCR, как правило, все GPCR обладают сходной молекулярной структурой. Все GPCR содержат цепь из аминокислотных остатков различной длины. GPCR расположены в клеточной мембране в виде семи отдельных спиралей, называемых трансмембранными участками. N-конец GPCR расположен снаружи клетки, так же как внеклеточные петли, а C-конец расположен внутри клетки совместно с внутриклеточными петлями.
Семейство A GPCR (подобное родопсину) включает в себя, но не ограничивается ими, рецепторные белки для аминов, пептидов, гормональных белков, родопсина, обонятельные рецепторные белки, рецепторы для простаноидов, нуклеотид-подобные рецепторные белки, каннабиноидные рецепторы, рецепторные белки для активирующего тромбоциты фактора, для гонадотропин-рилизинг гормона, для тиротропин-рилизинг гормона и для стимуляторов секреции, для мелатонина, вирусные, лизосфинголипидные и LPA (EDG) рецепторые белки, рецептор для лейкотриена B4 и другие сходные рецепторные белки.
Семейство B GPCR (подобное секретину) включает в себя, но не ограничивается ими, рецепторы для кальцитонина, кортикотропин-рилизинг фактора (CRF), гастроингибиторного пептида (GIP), глюкагона, рилизинг-фактора гормона роста (GHRH), паратиреоидного гормона (PTH), активирующего аденилатциклазу гипофиза полипептида (PACAP), секретина, вазоактивного интестинального полипептида (VIP), диуретического гормона, EMR1, латрофилина, специфичного для головного мозга ингибитора ангиогенеза (BAI), подобные мафусаиловым белки (MTH), кадгерин/EGF/LAG (CELSR) и другие сходные лиганды. Harmar (2001) описывает три подсемейства в семействе B GPCR, подсемейство B1, B2 и B3.
Подсемейство B1 - Подсемейство B1 включает в себя, но не ограничивается ими, классические рецепторы для гормонов, которые у человека кодируются по меньшей мере 15 генами, по меньшей мере с пятью предполагаемыми членами у Drosophila и тремя у C. elegans. Лиганды для рецепторов в этом семействе представляют собой полипептидные гормоны приблизительно из 27-141 аминокислотных остатков; по меньшей мере девять рецепторов млекопитающих отвечают на лиганды, которые являются структурно сходными друг с другом (глюкагон, глюкагон-подобные пептиды (GLP-1, GLP-2), зависимый от глюкозы инсулинотропный полипептид, секретин, вазоактивный интестинальный пептид (VIP), PACAP и рилизинг-фактор гормона роста (GHRH)). Было показано, что члены этого подсемейства способны регулировать внутриклеточные концентрации cAMP посредством присоединения аденилатциклазы через стимуляторный G-белок (Gs). Некоторые члены подсемейства способны передавать сигнал через дополнительные сопряженные с G-белком каскады передачи сигнала, например посредством активации фосфолипазы C.
Подсемейство B2 - Подсемейство B2 состоит из большого количества GPCR семейства B с длинными внеклеточными N-концами, содержащими различные структурные элементы, связанные с основным мотивом 7TM. Прототипными членами для этого подсемейства были содержащий EGF-элемент подобный муцину рецептор для гормонов (EMR1), выделенный из нейроэктодермальной библиотеки кДНК человека (Baud et al., 1995), и лейкоцитарный антиген клеточной поверхности CD97 (Hamann et al, 1995). Подсемейство B2 также включает в себя независимые от кальция рецепторы для α-латротоксина. Выявлено три гена, кодирующих независимые от кальция рецепторы для латротоксина (CL-1, CL-2 и CL-3). Во-вторых, также в это семейство входят специфичные для головного мозга ингибиторы ангиогенеза 1, 2 и 3 (BAI1, BAI2, BAI3), группа белков, которые вовлечены в васкуляризацию глиобластом. В-третьих, также в подсемейство B2 включены белок, кодируемый геном flamingo у Drosophila и его ортологами у людей (кадгерин EGF LAG семичленные рецепторы G-типа Celsr1, Celsr2 и Celsr3) и у C. elegans (F15B9.7). В заключение, подсемейство включает в себя четвертую, отдельную группу рецепторов, которые содержат некоторые мотивы, являющиеся общими для рецепторов подсемейства B2 однако в остальном структурно не сходны с ними (белок эпидидимиса человека 6 (HE6), связанный с EGF-TM7-латрофилином белок (ETL), содержащий повтор рецептор для иммуноглобулина Ig-hepta, сопряженный с G-белком рецептор 56 (GPR56)
и очень крупный сопряженный с G-белком рецептор 1 (VLGR1)). Анализ секвенирования генома человека (1 апреля 2001 года, UCSC Human Genome Project Working Draft (genome.ucsc.edu)) указывает на то, что существует по меньшей мере 18 генов человека, кодирующих члены подсемейства B2, и по меньшей мере четыре существует у Drosophila и три у C. elegans. Недавно была описана структура и функции членов подсемейства B2, Stacey et al. (2000).
Подсемейство B3 - Прототипом третьей группы (подсемейство B3) GPCR семейства B является "methuselah" (mth), ген, выделенный при скрининге единичных мутаций генов, которые увеличивают среднюю длительность жизни D. melanogaster (Lin et al, 1998). Ген кодирует полипептид, который обладает сходством последовательности с другими GPCR семейства B только в участке TM7. В последовательности генома Drosophila закодировано по меньшей мере восемь паралогов "methuselah".
Характерным признаком всех GPCR семейства B является мотив 7TM, отдаленно сходный с аналогичными участками некоторых других семейств GPCR, однако является значительно более консервативным в семействе B. Консервативные остатки цистеина во внеклеточных петлях EC1 и EC2, вероятно, формируют дисульфидные мостики по аналогии с GPCR семейства A, в которых этот признак также является консервативным (Palczewski et al, 2000). Однако в противоположность GPCR семейства A, в основе многих из которых, по-видимому, лежат внутренние гидрофобные последовательности для нацеливания на плазматическую мембрану, большинство GPCR семейства B, по-видимому, обладают N-концевым сигнальным пептидом. Исследования с использованием сайт-направленного мутагенеза и конструирования химер между рецепторами для гормонов из семейства B показали, что N-концевой внеклеточный домен является необходимым для связывания лиганда, но что трансмембранные домены и ассоциированные с ними участки внеклеточных петель рецепторов обеспечивают информацию, необходимую для специфичного взаимодействия с лигандами. Все рецепторы для гормонов семейства B содержат консервативный участок в N-концевом внеклеточном домене вблизи TM1, который может участвовать в связывании лиганда. Было показано, что изменение сплайсинга в этом участке рецептора для PAC1 влияет на специфичность и аффинность в отношении связывания лиганда (Dautzenberg et al, 1999).
Рецепторы подсемейства B2 содержат множество дополнительных структурных мотивов в их крупных N-концевых внеклеточных доменах, которые позволяют сделать предположение об участии этого домена в адгезии клетка-клетка и в передаче сигнала. Они включают в себя домены EGF (в Celsr1, Celsr2, Celsr3, EMR1, EMR2, EMR3, CD97 и Flamingo), повторы ламинина и кадгерина (в Flamingo и его ортологах у человека Celsr1, Celsr2 и Celsr3), подобные ольфактомедину домены (в рецепторах для латротоксина), повторы тромбоспондина типа 1 (в BAI1, BAI2 и BAI3) и, в Ig-hepta, домен иммуноглобулина типа C-2, также представленный в рецепторе для фактора роста фибробластов (FGF) 2 и в молекуле адгезии нейрональных клеток L1. VLGR1 обладает двумя копиями мотива (Calx-beta), представленными в ионообменниках Na+-Ca2+ и в субчастице β4 интегрина.
GPCR cемейства C (метаботропный глутамат/феромон) включают в себя метаботропные глутаматные, подобные чувствительным к кальцию, рецепторы для феромонов, GABA-B, одиночный GPRC5, одиночный GPCR6, белки bride-of-sevenless (BOSS), вкусовые рецепторы (T1R) и другие сходные белки.
В определенных вариантах осуществления sGPCR по этому изобретению представляют собой класс рецепторов B. В одном аспекте sGPCR по этому изобретению представляют собой подсемейство рецепторов B1, а в следующем аспекте sGCPR представляют собой CRFR1 и CRFR2 и рецептор для паратиреоидного гормона. В таблице 1 представлен неограничивающий ряд иллюстративных членов семейства GPCR, регистрационные номера и связанные с ними данные UNIGENE и OMIM включены в настоящее описание в качестве ссылок по дате приоритета и дате подачи этой заявки. Доступ к данным Unigene можно получить с помощью интернет-ссылок, содержащихся на web-странице OMIM. Ряд других GPCR и их регистрационные номера можно найти на web-сайте, определяемом следующим адресом в глобальной сети gpcr.org/7tm/htmls/entries.html.
A. Кортикотропин-рилизинг фактор (CRF) и его рецепторы
В качестве примера GPCR, охватываемых этим изобретением, подробно описаны рецепторы для CRP. Специалист в данной области способен применять эти конкретные данные в отношении других членов семейства GPCR, в частности типа B, и, более конкретно, подсемейства рецепторов B1. В определенных аспектах это изобретение относится, но не ограничивается ими, к sGPCR, происходящих из растворимых рецепторов для кортикотропин-рилизинг фактора (sCRFR), в частности к sCRFR2α. Гипоталамический гипофизиотропный пептидный кортикотропин-рилизинг фактор (CRF), первоначально выделенный из гипоталамуса (Vale et al, 1981), играет важную роль в регуляции гипоталамо-гипофизарно-надпочечниковой (HPA) системы в обычных и стрессовых условиях (River and Vale, 1983; Muglia et al, 1995). Кроме того, CRF оказывает действие посредством интеграции эндокринных, автономных и поведенческих ответов на вызывающие стресс факторы (River and Vale, 1983; Muglia et al, 1995; Koob and Heinrichs, 1999). Семейство пептида CRF млекопитающих включает в себя урокортин 1 (Ucn 1) (Vaughan et al, 1995) и пептиды урокортин 2 (Ucn 2) и урокортин 3 (Ucn 3), также известный как связанный со стресскопином пептид (Reyes et al, 2001; Hsu and Hsueh, 2001) и стресскопин (Hsu and Hsueh, 2001; Lewis et al, 2001) соответственно.
Эффекты связанных с CRF пептидов опосредованы активацией двух высокоаффинных мембранных рецепторов, CRFR1 (Chen et al, 1993; Vita et al, 1993; Chang et al, 1993) и CRFR2 (Perrin et al, 1995; Stenzel et al, 1995; Kishimoto et al, 1995; Lovenberg et al, 1995; Chen et al, 2005), которые принадлежат подсемейству B1 рецепторов с семь-трансмембранным доменом (7TMD), которые передают сигнал посредством связывания с G-белками. Один функциональный вариант гена CRFR1 экспресируется как у человека, так и у грызунов, совместно с некоторыми нефункциональными вариантами, которые получаются посредством дифференциального сплайсинга различных экзонов (Pisarchik and Slominski, 2004; Grammatopoulos et al, 1999). CRFR2 обладает тремя функциональными вариантами по сплайсингу у человека (α, β, и γ) и двумя вариантами у грызунов (α и β), которые возникают вследствие использования альтернативных 5'-экзонов (Perrin et al, 1995; Stenzel et al, 1995; Kishimoto et al, 1995; Lovenberg et al, 1995; Chen et al, 2005; Grammatopoulos et al, 1999; Kostich et al, 1998). мРНК CRFR1 широко экспрессируется в головном мозге и гипофизе млекопитающих, и высокие уровни обнаруживаются в передней доле гипофиза, коре головного мозга, мозжечке, миндалевидном теле, гиппокампе и обонятельной луковице (Van Pett et al, 2000). На периферии, CRFR1 экспрессируется в яичках, яичниках, коже и селезенке. мРНК CRFR2 экспрессируется дискретно в головном мозге с наивысшей плотностью в боковом ядре септума (LS), опорном ядре краевой полосы (BNST), вентромедиальном гипоталамическом ядре (VMH), обонятельной луковице и в ядре шва среднего мозга (Van Pett et al, 2000). CRFR2α представляет собой основной вариант по сплайсингу, экспрессирующийся в головном мозге крысы (Lovenberg et al. 1995) в то время как CRFR2β экспрессируется в периферических тканях, с наиболее высокими уровнями в скелетной мускулатуре, сердце и коже (Perrin et al, 1995).
Распределение CRFR1 и CRFR2 отличается и подразумевает различные физиологические функции, как показано с помощью различных фенотипов мышей с отсутствием CRFR1 или CRFR2. Мыши, дефектные по CRFR1, проявляют поведение со сниженной тревожностью и обладают сниженным ответом на стресс (Smith et al, 1998; Timpl et al, 1998), в то время как мыши с отсутствием CRFR2 обладают поведением с повышенной тревожностью и усиленным ответом на стресс HPA (Zhu et al, 1999; Valerio et al, 2001; Khan et al, 1993). Однако ответы на введение агонистов и антагонистов CRFR2 в определенные участки головного мозга показывают как анксиолитическую, так и анксиогенную роль CRFR2 (Bale and Vale, 2004).
Радиорецепторный и функциональный анализы показали, что CRFR1 и CRFR2 отличаются фармакологически: Ucn 1 обладает равной аффинностью в отношении обоих рецепторов и в отношении CRFR2 является более эффективным, чем CRF, в то время как Ucn 2 и Ucn 3, по-видимому, являются селективными в отношении CRFR2 (Vaughan et al, 1995; Reyes et al, 2001; Lewis et al, 2001). Активация определенных CRFR в различных тканях или типах клеток посредством селективных в отношении рецептора пептидов CRF запускает ряд каскадов передачи сигнала, в том числе присоединение различных G-белков, стимуляцию PKB, PKC, внутриклеточного кальция, и активируемой митогеном протеинкиназы (MAPK) (для обзора см. Bale and Vale, 2004; Perrin and Vale, 1999; Brar et al, 2002).
Как CRFR1, так и CRFR2 существуют в виде множества вариантов по сплайсингу. Авторы изобретения выявили кДНК из головного мозга мыши, кодирующую иллюстративный вариант по сплайсингу sCRFR2α, в котором из нуклеиновой кислоты, кодирующей CRFR2α, удален шестой экзон. Трансляция этой изоформы приводит к ожидаемому растворимому белку из 143 аминокислот. Транслируемый белок включает в себя большую часть внкелеточного домена (ECD1) CRFR2α с последующим уникальным гидрофильным C-концом из 38 аминокислот, который возникает вследствие переключения рамки считывания посредством делеции шестого экзона. В исследованиях были показаны высокие уровни экспрессии sCRFR2α в обонятельной луковице, коре и в участках среднего мозга. Белок, соответствующий sCRFR2α, экспрессированный и очищенный либо из клеточных систем млекопитающих, либо из бактериальных клеточных систем, связывает несколько лигандов из семейства CRF с низкой наномолярной аффинностью. Кроме того, очищенный белок sCRFR2α ингибирует клеточные ответы на CRF и урокортин 1. Таким образом, белок sCRFR2α может представлять собой биологический модулятор лигандов семейства CRF. Модулирование лигандов из семейства CRF не ограничивается головным мозгом, его можно использовать в любой ткани, которая подвержена воздействию одного или нескольких членов лигандов из семейства CRF.
Аспекты этого изобретения, главным образом, относятся к композициям и способам для достижения терапевтического эффекта, включая модулирование активности лиганда GPCR, такого как лиганды семейства CRF, с использованием растворимого связывающего лиганд полипептида GPCR, такого как связывающий CRF
полипептид, в качестве антагониста, либо отдельно, либо совместно с одним или несколькими другими антагонистами гормона (например, низкомолекулярным антагонистом), включая, но не ограничиваясь ими, антагонист лиганда(ов) семейства CRF.
Одним способом антагонизма действию лиганда является воздействие на лиганд рецептора-приманки или растворимого рецептора, чтобы ограничить локальную концентрацию лиганда(ов), который связывает приманку, и модулировать способность лигандов к передаче сигнала через их рецептор на поверхности клеток. Растворимые белки, сходные с мембранными рецепторами, можно получать посредством ферментативного укорочения связанных с мембраной рецепторов, как предложено для рецептора GHRH (Rekaski et al, 2000), рецептора для допамина D3 (Liu et al, 1994) и рецептора для кальцитонина (Seek et al, 2003) или посредством альтернативного сплайсинга в случае рецепторов для глутамата (Malherbe et al, 1999; Zhu et al, 1999; Valerio et al, 2001). Описаны варианты по сплайсингу, содержащие только внеклеточный участок GPCR (Pisarchik and Slominski, 2004; Grammatopoulos et al, 1999; Kostich et al, 1998; Malherbe et al, 1999; Zhu et al, 1999; Valerio et al, 2001; Khan et al, 1993; Graves et al, 1992; You et al, 2000; Schwarz et al, 2000). В большинстве случаев эти белки действуют в качестве связывающих не передающих сигнал молекул, также называемых как рецепторы-приманки. В коже человека выявили два неполных фрагмента кДНК (CRFR1e и CRFR1h), содержащих делецию экзона 3 и 4 и вставку скрытого экзона в CRFR1, и было предсказано, что они существуют в качестве растворимых белков (Pisarchik and Slominski, 2004). Один из этих фрагментов, CRFR1e, проявляет доминантные отрицательные эффекты при котрансфекции с CRFR1 дикого типа.
Kehne and Lombaert (2002) описывают непептидные антагонисты рецептора для CRF для лечения тревоги, депрессии и связанных со стрессом нарушений. CRF вовлечен в психиатрические нарушения, такие как тревожность и депрессия. С момента выявления кортикотропин-рилизинг фактора (CRF) в обширной программе исследований была подтверждена важность этого пептида из 41 аминокислот и сходных с ним членов семейства для осуществления поведенческих, эндокринных и автономных ответов организма на стресс.
Доклинические и клинические данные свидетельствуют об участии CRF в общем и рецепторов для CRF в частности, в тревоге и депрессии. Клинические исследования показали дисфункцию в гипоталамо-гипофизарно-надпочечниковой (HPA) системе и/или повышенные уровни CRF при депрессии и при некоторых связанных с тревогой нарушениях. Доклинические данные, для которых использовали корреляционные способы, генетические модели и способы экзогенного введения CRF у грызунов и не относящихся к человеку приматов, подтверждают связь между сверхактивными каскадами CRF и анксиогенными и подобными депрессивным симптомами. В исследованиях, в которых использовали нокаут рецепторов и селективные непептидные антагонисты CRFR1, были показаны анксиолитические и антидепрессантные эффекты при определенных типах лабораторных состояний. Было описано, что в фазе II открытых клинических испытаний при большом депрессивном расстройстве антагонист CRFR1 является безопасным и эффективным для снижения симптомов тревоги и депрессии.
Различные неограничивающие виды активности антагонистов CRF описаны Owens et al. (1991). Описаны антагонисты CRF, являющиеся эффективными для лечения связанных со стрессом расстройств; нарушений настроения, таких как депрессия, большое депрессивное расстройство, депрессия с одним эпизодом, повторяющаяся депрессия, индуцированная насилием в детском возрасте депрессия, послеродовая депрессия, дистимия, биполярные нарушения и циклотимия; синдрома хронической усталости; нарушений питания, таких как анорексия и нейрогенная булимия; генерализованного связанного с тревогой нарушения; панического нарушения; фобий; обессивно-компульсивного нарушения; нарушения, связанного с посттравматическим стрессом; ощущения боли, такого как фибромиалгия; головной боли; желудочно-кишечных заболеваний; геморрагического стресса; язв; индуцированных стрессом психотических эпизодов; лихорадки; диареи; послеоперационной непроходимости кишечника; гиперчувствительности толстой кишки; синдрома раздраженной кишки; болезни Крона; спастической толстой кишки; воспалительных нарушений, таких как ревматоидный артрит и остеоартрит; боли; астмы; псориаза; аллергии; остеопороза; преждевременных родов; гипертензии, застойной сердечной недостаточности; нарушений сна; нейродегенеративных заболеваний, таких как болезнь Альцгеймера, сенильная деменция по типу болезни Альцгеймера, мультиинфарктная деменция, болезнь Паркинсона и болезнь Гентингтона; травмы головы; ишемического нейронального повреждения; эксайтотоксического нейронального повреждения; эпилепсии; инсульта; травмы спинного мозга; психосоциальной карликовости; эутиреодиного синдрома тошноты; синдрома неадектватной продукции антидиуретического гормона; ожирения; зависимости и привыкания к химическим веществам; симптомов отмены лекарственного средства и алкоголя; бесплодия; злокачественной опухоли; мышечных спазмов; недержания мочи; гипогликемии и дисфункций иммунной системы, включая индуцируемую стрессом дисфункцию иммунной системы, подавление иммунной системы и инфекции вирусом иммунодефицита человека; и индуцированной стрессом инфекции у человека и животных. Эти и другие состояния, подлежащие модулированию посредством CRF, указаны в литературе, которая включает в себя Lovenberg et al. (1995); Chalmers et al. (1996); и патент США 5063245, все из которых включены в настоящее описание в качестве ссылок в полном объеме.
II. ПОЛИПЕПТИДЫ
Полипептиды по этому изобретению включают в себя растворимые формы GPCR или растворимые рецепторы. Растворимые рецепторы по этому изобретению могут содержать субчастицы, которые были изменены с мембраносвязанной на растворимую форму. Таким образом, растворимые пептиды можно получать посредством укорачивания полипептидов с удалением, например, 7-трансмембранных участков и/или цитоплазматического конца. Альтернативно трансмембранные домены можно удалять посредством делеции или посредством замещения гидрофобных в норме аминокислотных остатков, которые содержат трансмембранный домен, гидрофильными остатками. В любом случае, получают по существу гидрофильный или растворимый полипептид со сниженной аффинностью к липидам и повышенной растворимостью в воде. Делеция трансмембранных доменов является предпочтительной по сравнению с замещением гидрофильными аминокислотными остатками, вследствие того, что она предотвращает встраивание потенциально иммуногенных эпитопов. Растворимые рецепторы по этому изобретению могут включать в себя любое количество хорошо известных лидерных последовательностей на N-конце. Такая последовательность позволит пептидам экспрессироваться и направляться по пути секреции в эукариотической системе.
A. Слитые белки
Рецепторы являются эффективными средствами для исследования биологических каскадов и для лечения различных болезненных состояний посредством их простого преобразования в слитые с иммуноглобулином белки. Эти димерные растворимые рецепторные формы представляют собой надежные ингибиторы процессов, опосредуемых либо секретируемыми, либо связанными с поверхностью лигандами. Посредством связывания с этими лигандами они предотвращают взаимодействие лиганда со связанными с клеткой рецепторами. Эти слитые белки рецептор-Ig пригодны не только для экспериментального направления, но и их успешно использовали клинически в качестве TNF-R-Ig для лечения воспалительного заболевания кишечника, ревматоидного артрита и острого клинического синдрома, сопровождающего введение OKT3 (Eason et al, 1996; van Dullemen et al, 1995). Авторы изобретения предположили, что манипулирование множеством процессов, опосредуемых передачей сигнала через GPCR, найдет применение в лечении обусловленных GPCR заболеваний.
Предпочтительно, стабильные белки плазмы, которые, как правило, обладают периодом полураспада в кровотоке млекопитающих более чем часы, можно использовать для конструирования слитых с рецептором белков. Такие белки плазмы включают в себя, но не ограничиваются ими: иммуноглобулины, сывороточный альбумин, липопротеины, аполипопротеины и трансферрин. Также для получения локализованного в конкретной области растворимого слитого белка рецептора к связывающему лиганд домену рецептора можно присоединять последовательности, которые могут направлять растворимые рецепторы в конкретный тип клеток или ткани.
Весь внеклеточный участок GPCR или его функциональный фрагмент, содержащий связывающий лиганд домен GPCR, может быть слитым с константным участком иммуноглобулина, таким как Fc-домен тяжелой цепи IgG1 человека. Растворимые слитые белки рецептор-IgG представляют собой распространенные иммунологические реагенты, и способы для их конструирования хорошо известны в данной области (см., например, патент США 5225538, который включен в настоящее описание в качестве ссылки в полном объеме).
Функциональный связывающий лиганд домен GPCR может быть слитым с Fc-доменом иммуноглобулина (Ig). Источником Fc Ig может быть класс или подкласс иммуноглобулина, включая, но не ограничиваясь этим, IgG1. Fc-домены антител, принадлежащих различным классам или подклассам Ig, могут активировать различные вторичные эффекторные функции. Активация происходит, когда Fc-домен связывается с соответствующим ему Fc-рецептором. Вторичные эффекторные функции включают в себя способность активировать систему комплемента или проходить через плаценту. Свойства различных классов и подклассов иммуноглобулинов описаны в данной области.
Специалист в данной области поймет, что различные аминокислотные остатки, образующие область соединения слитого белка рецептор-Ig могут, изменять структуру, стабильность и конечную биологическую активность слитого белка sGPCR. Для модификации области соединения с выбранным доменом для слияния к C-концу выбранного фрагмента sGPCR можно добавлять одну или несколько аминокислот.
Для встраивания в рекомбинантный экспрессирующий вектор также можно изменять N-конец слитого белка sGPCR посредством замены положения, в котором выбранный фрагмент ДНК sGPCR расщепляется на его 5'-конце. Стабильность и активность каждого слитого белка sGPCR можно тестировать и оптимизировать с использованием общепринятого экспериментирования, включая, но не ограничиваясь этим, оценку связывания лиганда.
С использованием последовательностей связывающего лиганд домена sGPCR во внеклеточном домене, как представлено в настоящем описании, также можно конструировать варианты аминокислотной последовательности для модификации аффинности молекул sGPCR к их лигандам. Растворимые молекулы по этому изобретению могут конкурировать с эндогенными рецепторами за связывание. Предусматривается, что любая растворимая молекула, содержащая связывающий лиганд домен GPCR, который конкурирует с природными рецепторами за связывание лиганда, представляет собой блокирующее рецептор вещество или захватывающее лиганд вещество, которое находится в объеме настоящего изобретения.
B. Конъюгаты белков
В отношении периода полураспада белка, одним способом для повышения периода полураспада белка в кровотоке является обеспечение снижения выведения белка, в частности, посредством почечного выведения и опосредуемого рецептором выведения. Этого можно достигать посредством конъюгации белка с химической группой, которая способна повышать кажущийся размер, таким образом снижая выведение почками и повышая период полураспада in vivo. Более того, присоединение химической группы к белку может эффективно блокировать физический контакт протеолитических ферментов с белком таким образом предотвращая деградацию посредством неспецифичного протеолиза. Полиэтиленгликоль (PEG) представляет собой одну такую химическую группу, которую использовали для получения терапевтических белковых продуктов. Недавно молекула G-CSF, модифицированная одной связанной с N-концом группой PEG массой 20 кДа (Neulastam), была одобрена для продажи в США. Было показано, что эта пегилированная молекула G-CSF обладает повышенным периодом полураспада по сравнению с непегилированным G-CSF и таким образом ее можно вводить менее часто, чем современные продукты G-CSF, однако она не снижает длительность нейтропении по сравнению с непегилированным G-CSF.
Модификация посредством полиэтиленгликоля (PEG) является важной для фармацевтических и биотехнологических способов применения. Пегилирование (ковалентное присоединение PEG) приводит, например, к экранированию антигенных или иммуногенных эпитопов. Более того, оно снижает опосредуемый рецептором захват ретикулоэндотелиальной системой или предотвращает распознавание и деградацию протеолитическими ферментами. Было показано, что пегилирование белков повышает их биодоступность посредством снижения почечной фильтрации.
Подразумевают, что термином "конъюгат" обозначают гетерогенную молекулу, образованную посредством ковалентного присоединения одного или нескольких полипептидов, как правило, одного полипептида, к одному или нескольким неполипептидным группам, таким как молекулы полимера, липофильные соединения, углеводные группы или органические вещества для получения производных. Термин "ковалентное присоединение" означает, что полипептид и неполипептидная группа либо непосредственно ковалентно соединены друг с другом, либо также непрямо ковалентно соединены друг с другом посредством промежуточной группы или групп, таких как мостик, спейсер или связующая группа или группы. Предпочтительно, конъюгат является растворимым в соответствующих концентрациях и условиях, т.е. он растворим в физиологических жидкостях, таких как кровь. Композиции и способы получения конъюгата по этому изобретению описаны в патенте США 6831158, который включен в настоящее описание в качестве ссылки в полном объеме. Способы, описанные в патенте США 6831158, относятся к конъюгации G-CSF, однако их легко можно адаптировать для конъюгации sGPCR по настоящему изобретению.
"Полимерная молекула" представляет собой молекулу, образованную посредством ковалентного соединения двух или более мономеров. Термин "полимер" можно использовать взаимозаменяемо с термином "полимерная молекула". Подразумевается, что термин охватывает углеводные молекулы, включая углеводные молекулы, присоединенные к полипептиду посредством N- или O-гликозилирования in vivo, такую молекулу также называют "олигосахаридной группой". За исключением случаев, когда количество полимерных молекул явно указано, каждое упоминание "полимера", "полимерной молекулы", содержащейся в полипептиде по этому изобретению или иным образом используемой в соответствии с настоящим изобретением, следует относить к одной или нескольким полимерной молекуле(ам).
Подразумевают, что термином "группа для присоединения" обозначают группу аминокислотного остатка полипептида, способную присоединять соответствующую неполипептидную группу. Например, для конъюгации с полимером, в частности с PEG,
часто используемой группой для присоединения является ε-аминогруппа лизина или N-концевая аминогруппа. Другие группы для присоединения полимера включают в себя свободную группу карбоновой кислоты (например, группу C-концевого аминокислотного остатка или остатка аспарагиновой кислоты, или глутаминовой кислоты), активируемые пригодным способом карбонильные группы, окисленные углеводные группы и меркаптогруппы. Пригодные группы для присоединения и соответствующие им непептидные группы проиллюстрированы в таблице 2.
Активированный PEG
PEG с группами трезила
Олигосахаридная группа
присоединение in vitro
Олигосахаридная группа
PEG малеимид
Присоединение
in vitro
PAG со сложным эфиром, простым эфиром, карбаматом, карбонатом
Полимер, например, PEG
Кетон
Гидроксид PEG
WO92/
16655,
WO00/
23114
CRC Press
CRC Press
C. Сайт-специфический мутагенез
В одном варианте осуществления можно получать варианты аминокислотной последовательности полипептида. Они могут представлять собой, например, незначительные варианты последовательности полипептидов, которые возникают вследствие природного разнообразия в популяции или они могут представлять собой гомологи, представленные в других видах. Также они могут представлять собой последовательности, которые в природе не встречаются, однако которые являются достаточно сходными, чтобы сходным образом функционировать и/или вызывать иммунный ответ с перекрестным реагированием с природными формами полипептида. Варианты последовательности можно получать стандартными способами сайт-направленного мутагенеза, такими как способы, описанные ниже.
Варианты аминокислотной последовательности полипептида могут быть вариантами с заменой, вставкой или делецией. В вариантах с делецией отсутствует один или несколько остатков природного белка, которые не являются существенными для выполнения функции или для иммуногенной активности, и их примером являются варианты рецептора, лишенные трансмембранной последовательности. Варианты с заменами, как правило, обладают заменой одной аминокислоты другой аминокислотой в одном или нескольких участках в белке, и они могут быть предназначены для модулирования одного или нескольких свойств полипептида, таких как стабильность в отношении протеолитического расщепления или иммуногенность.
Замены предпочтительно являются консервативными, т.е. одну аминокислоту заменяют аминокислотой со сходной формой и зарядом. Консервативные замены хорошо известны в данной области и включают в себя, например, замены аланина серином; аргинина лизином; аспарагина глутамином или гистидином; аспартата глутаматом; цистеина серином; глутамина аспарагином; глутамата аспартатом; глицина пролином; гистидина аспарагином или глутамином; изолейцина лейцином или валином; лейцина валином или изолейцином; лизина аргинином; метионина лейцином или изолейцином; фенилаланина тирозином, лейцином или метионином; серина треонином; треонина серином; триптофана тирозином; тирозина триптофаном или фенилаланином; и валина изолейцином или лейцином.
Варианты со вставкой включают в себя слитые белки, такие как белки, используемые для обеспечения быстрой очистки полипептида, а также они могут включать в себя гибридные белки, содержащие последовательности из других белков и полипептидов. Например, вариант со вставкой может включать в себя участки аминокислотной последовательности полипептида из одного вида совместно с участком гомологичного полипептида из другого вида. Другие варианты со вставкой могут включать в себя варианты, в которых в кодирующую последовательность полипептида встраивают дополнительные аминокислоты, например можно встраивать участок(и) для расщепления протеазой.
Можно проводить модификацию и изменение структуры полинуклеотида и тем не менее получать функциональную молекулу, которая кодирует белок или полипептид с требуемыми свойствами. Ниже представлено исследование, в основе которого лежит изменение аминокислот в белке с получением эквивалентной, или даже улучшенной, молекулы второго поколения. Аминокислотные изменения можно проводить посредством замены кодонов в последовательности ДНК в соответствии со следующими данными.
Например, определенные аминокислоты можно заменять другими аминокислотами в белковой структуре без существенной потери способности к связыванию при взаимодействии со структурами, например такими, как антигенсвязывающие участки антител или участки для связывания на молекулах субстрата. Поскольку биологическую активность белка определяет способность к взаимодействию и природа белка, можно проводить определенные аминокислотные замены в белковой последовательности, получая тем не менее белок с подобными свойствами. Таким образом, авторы этого изобретения предположили, что различные изменения можно проводить в последовательностях ДНК генов, мРНК или полинуклеотидов без существенной потери их биологических свойств или активности.
При проведении таких изменений можно рассматривать индекс гидрофобности аминокислот. Важность индекса гидрофобности аминокислот для обеспечения биологической функции белка, касающейся взаимодействия, в основном понятна в данной области (Kyte & Doolittle, 1982).
Тот факт, что относительный характер гидрофобности аминокислоты приводит ко вторичной структуре конечного белка, которая в свою очередь определяет взаимодействие белка с другими молекулами, например ферментами, субстратами, рецепторами, ДНК, антителами, антигенами и т.п., является общепринятым. В данной области известно, что определенные аминокислоты можно заменять другими аминокислотами, обладающими сходным индексом или величиной гидрофобности, и тем не менее получать белок со сходной биологической активностью. При проведении таких замен замена аминокислот, индексы гидрофобности которых находятся в пределах ±2 является предпочтительной, замена аминокислот с индексами в пределах ±1 является особенно предпочтительной, и замена аминокислот с индексом в пределах ±0,5 является еще более предпочтительной.
Также в данной области понятно, что замену сходных аминокислот можно проводить эффективно на основе гидрофильности. В патенте США 4554101, включенном в настоящее описание в качестве ссылки, указано, что наибольшая локальная средняя гидрофильность белка, обусловленная гидрофильностью его смежных аминокислот, коррелирует с биологическим свойством белка.
Понятно, что аминокислоту можно заменять другой аминокислотой, обладающей сходным значением гидрофильности, и тем не менее получать биологически эквивалентный и иммунологически эквивалентный белок. При таких изменениях замена аминокислот, величина гидрофильности которых находится в пределах ±2, является предпочтительной, замена аминокислот с величиной гидрофильности в пределах ±1 является особенно предпочтительной, и замена аминокислот с величиной гидрофильности в пределах ±0,5 является еще более предпочтительной.
Сайт-специфический мутагенез представляет собой способ, пригодный для получения определенных пептидов, или биологически функциональных эквивалентов белков или пептидов, посредством специфичного мутагенеза лежащих в их основе ДНК. Кроме того, способ подразумевает быструю возможность получать и тестировать варианты последовательности, включая один или несколько из рассмотренных ранее, посредством внесения одного или нескольких изменений в нуклеотидную последовательность в ДНК. Как правило, способ сайт-специфического мутагенеза хорошо известен в данной области. В способе, как правило, используют вектор на основе бактериофага, который существует как в одноцепочечной и двухцепочечной форме. Типичные векторы, пригодные для сайт-направленного мутагенеза, включают в себя векторы, такие как фаг M13. Эти фаговые векторы являются коммерчески доступными и их применение, главным образом, известно специалистам в данной области. Также для сайт-направленного мутагенеза обычно используют двухцепочечные плазмиды, в случае которых отсутствует стадия переноса представляющего интерес гена из фага в плазмиду.
Получение вариантов последовательностей GPCR, включая, но не ограничиваясь этим, полинуклеотид sCRFR2α с использованием сайт-направленного мутагенеза предусмотрено в качестве способа получения потенциально пригодных образцов, т.е. образцов с измененными свойствами в отношении лиганда, которые включают в себя повышенную аффинность в отношении конкретного лиганда, и не является ограничивающим, поскольку существуют другие способы, посредством которых можно получать варианты последовательностей нуклеиновых кислот. Например, рекомбинантные векторы, кодирующие требуемый ген, можно обрабатывать мутагенными веществами, такими как гидроксиламин, с получением вариантов последовательностей.
D. Экспрессия и очистка полипептидов
Полинуклеотиды по этому изобретению, в частности 100, 150, 200 250, 300, 400, 450, 500, 550 или более смежных нуклеотидов в ДНК, кодирующей GPCR, GPCR семейства B, GPCR семейства B1, или полинуклеотид, который на 70, 75, 80, 85, 90, 95, 98 или 100% является идентичным последовательности, представленной в прилагаемом списке последовательностей, например SEQ ID NO:1, 3, 5, 7, 9, 11, 13 или 14, можно экспрессировать в качестве кодируемых им пептидов или белков. В конкретном аспекте ДНК кодируют весь внеклеточный домен GPCR или его часть и, в частности, N-концевой внеклеточный домен. Конструирование фрагмента(ов) ДНК для экспрессии в прокариотической или эукариотической системе можно проводить посредством способов, в основном известных специалистам в области рекомбинантной экспрессии. Полагают, что для экспрессии заявленных последовательностей нуклеиновых кислот можно использовать практически любую экспрессирующую систему.
В определенных вариантах осуществления настоящее изобретение относится к новым композициям, содержащим по меньшей мере одну белковую молекулу, такую как sGPCR, sCRFR или sCRFR2. Как используют в настоящем описании, "белковая молекула", "белковая композиция", "белковое соединение", "белковая цепь" или "белковое вещество", главным образом, относится, но не ограничивается этим, к белку более чем приблизительно из 200 аминокислот или к полноразмерной эндогенной последовательности, транслируемой с гена; к полипептиду более чем приблизительно из 100 аминокислот; и/или к пептиду от приблизительно 3 до приблизительно 100 аминокислот. Все описанные выше термины, относящиеся к "белковым", могут использоваться в настоящем описании взаимозаменяемо. Более того, эти термины также могут быть использованы в отношении слитых белков.
В определенных вариантах осуществления размер по меньшей мере одной белковой молекулы может составлять, но не ограничиваться этим, приблизительно или по меньшей мере 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 275, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1100, 1200, 1300, 1400, 1500, 1750, 2000, 2250, 2500 или более аминокислотных остатков, и любой диапазон, получаемый из этих значений, в частности 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190 или более последовательностей из смежных аминокислот такой длины для GPCR, GPCR семейства B, GPCR семейства B1 или SEQ ID NO:2, 4, 6, 8, 10, 12 или 15, включая полноразмерную SEQ ID NO:4, 8, 12 или 15. Как кДНК, так и геномные последовательности являются пригодными для экспрессии в эукариотах, поскольку клетка-хозяин, как правило, преобразует геномные транскрипты с получением функциональной мРНК для трансляции в белок.
Как используют в настоящем описании, подразумевается, что термины, "полученные способами инженерии" и "рекомбинантные" клетки, относятся к клетке, в которую вводят фрагмент экзогенной ДНК или полинуклеотида, такой как кДНК или полинуклеотид. Таким образом, полученные способами инженерии клетки отличаются от природных клеток, которые не содержат встроенного рекомбинантным способом сегмента ДНК или гена. Полученные способами инженерии клетки, таким образом, обладают геном или генами, введенными с помощью человека. Рекомбинантные клетки включают в себя клетки, обладающие введенной кДНК или геномной ДНК, и также могут включать в себя гены, расположенные рядом с промотором, не естественным образом ассоциированным с конкретным введенным геном.
Для экспрессии рекомбинантного белка или полипептида как мутантного, так и дикого типа, в соответствии с настоящим изобретением можно получать экспрессирующий вектор, который содержит одну из заявленных выделенных нуклеиновых кислот под контролем одного или нескольких промоторов. Для получения кодирующей последовательности "под контролем" промотора, как правило, помещают 5'-конец точки инициации трансляции рамки считывания приблизительно между 1 и 50 нуклеотидами "ниже" (т.е., с 3'-стороны) выбранного промотора. Расположенный "выше" промотор стимулирует трансляцию встроенной ДНК и обеспечивает экспрессию кодируемого рекомбинантного белка. Это представляет собой значение "рекомбинантной экспрессии" в контексте, используемом в настоящем описании.
Для конструирования экспрессирующих векторов, содержащих соответствующие нуклеиновые кислоты и последовательности для контроля транскрипции/трансляции, в целях достижения экспрессии белка или пептида в разнообразных экспрессирующих системах хозяев, доступно множество стандартных способов. Типы клеток, пригодные для экспрессии, включают в себя, но не ограничиваются ими, бактерии, такие как E. coli, B. subtilis, штамм RR1 E. coli, E. coli LE392, E. coli B, E. coli χ 1776 (ATCC No. 31537) а также E. coli W3110 (F-, лямбда-, прототрофный, ATCC No. 273325); палочковидные бактерии, такие как Bacillus subtilis; и другие энтеробактерии, такие как Salmonella typhimurium, Serratia marcescens и различные виды Pseudomonas, трансформированные рекомбинантными экспрессирующими векторами с фаговой ДНК, плазмидной ДНК или космидной ДНК.
Полинуклеотид или фрагмент полинуклеотида, кодирующий полипептид, можно встраивать в экспрессирующий вектор посредством стандартных способов субклонирования. В одном варианте осуществления используют экспрессирующий вектор E. coli, который продуцирует рекомбинантный полипептид в качестве слитого белка, обеспечивая быструю аффинную очистку белка. Примерами таких экспрессирующих систем для слитого белка являются система глататион-S-трансферазы (Pharmacia, Piscataway, NJ), система связывающего мальтозу белка (New England Biolabs, Beverley, MA), система FLAG (IBI, New Haven, CT) и система 6xHis (Qiagen, Chatsworth, CA). Кроме того, пригодные векторы включают в себя векторы pIN (Inouye et al, 1985); и векторы pGEX для применения для получения растворимых слитых белков глутатион-S-трансферазы (GST). Другие пригодные слитые белки представляет собой белки с β-галактозидазой, убиквитином или сходные с ними.
Для экспрессии в Saccharomyces обычно используют, например, плазмиду YRp7 (Stinchcomb et al, 1979; Kingsman et al, 1979; Tschemper et al, 1980). Эта плазмида содержит ген trp1, который представляет собой селективный маркер для мутантного штамма дрожжей, лишенного способности расти в присутствии триптофана, например ATCC No. 44076 или PEP4-1 (Jones, 1977). Наличие повреждения trp1 в качестве характерной особенности генома дрожжевой клетки-хозяина затем обеспечивает эффективные окружающие условия для выявления трансформации по росту в присутствии триптофана. Пригодные активирующие последовательности в дрожжевых векторах включают в себя промоторы для 3-фосфоглицераткиназы (Hitzeman et al, 1980) или других гликолитических ферментов (Hess et al, 1968; Holland et al, 1978), таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкоза-6-фосфат изомераза, 3-фосфоглицеромутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа. При конструировании пригодных экспрессирующих плазмид в экспрессирующий вектор с 3'-стороны последовательности, подлежащей экспрессии, также лигируют терминирующие последовательности, ассоциированные с этими генами, для обеспечения полиаденилирования мРНК и терминации.
Другие пригодные промоторы, которые обладают дополнительным преимуществом, состоящим в контроле транскрипции условиями роста, включают в себя промоторный участок для алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, ферментов деградации, ассоциированных с метаболизмом азота, и упомянутой выше глицеральдегид-3-фосфатдегидрогеназы, и для ферментов, ответственных за потребление мальтозы и галактозы.
В дополнение к микроорганизмам, также в качестве хозяев можно использовать культуры клеток, полученные из многоклеточных организмов. В принципе, любая такая культура клеток является применимой из культуры клеток как позвоночных, так и беспозвоночных, включая клетки млекопитающих и насекомых (например, патент США No. 4215051).
Примерами пригодных линий клеток-хозяев млекопитающих являются клетки VERO
и HeLa, клеточные линии яичника китайского хомяка (CHO), клеточные линии WI38, BHK, COS-7, 293, HepG2, NIH3T3, RIN и MDCK. Кроме того, можно выбирать клетку-хозяина, которая модулирует экспрессию встроенных последовательностей или модифицирует и преобразует продукт гена конкретным требуемым образом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важными для функционирования кодируемого белка.
Также для эффективной трансляции заявленных кодирующих последовательностей нуклеиновых кислот могут быть необходимы определенные сигналы для инициации. Эти сигналы включают в себя кодон инициации ATG и смежные с ним последовательности. Кроме того, также может быть необходимым предоставление экзогенных сигналов для контроля трансляции, включая кодон инициации ATG. Средний специалист в данной области может легко определить наличие этой необходимости и обеспечить необходимые сигналы. Хорошо известно, что кодон для инициации должен находиться в рамке (или в фазе) с рамкой считывания требуемой кодирующей последовательности для обеспечения трансляции всей вставки. Эффективность экспрессии можно усиливать посредством встраивания соответствующих транскрипционных энхансерных элементов или терминаторов транскрипции (Bittner et al, 1987).
Для длительной продукции рекомбинантных белков с высоким выходом предпочтительной является стабильная экспрессия. Например, можно конструировать клеточные линии, которые стабильно экспрессируют конструкции, кодирующие G-белки. Вместо использования экспрессирующих векторов, которые содержат вирусные ориджины репликации, клетки-хозяева можно трансформировать векторами, контролируемыми соответствующими элементами для контроля экспрессии (например, промоторной, энхансерной последовательностями, терминаторами транскрипции, участками полиаденилирования и т.д.) и селективным маркером. После встраивания чужеродной ДНК полученным способам инженерии клеткам можно позволять расти в течение 1-2 суток в обогащенной среде, а затем переносить на селективную среду. Селективный маркер в рекомбинантной плазмиде придает устойчивость при селекции и позволяет клеткам стабильно встраивать плазмиду в их хромосомы и расти с образованием очагов, которые в свою очередь можно клонировать и расширять в клеточные линии.
Можно использовать ряд систем для селекции, включая, но не ограничиваясь ими, гены тимидинкиназы вируса простого герпеса (Wigler et al, 1977), гипоксантин-гуанин фосфорибозилтрансферазы (Szybalska et al, 1962) и аденин фосфорибозилтрансферазы (Lowy et al, 1980), в клетках tk -, hgprt - или aprt - соответственно. Также можно использовать устойчивость к антиметаболиту в качестве основы селекции посредством dhfr, который придает устойчивость к метотрексату (Wigler et al, 1980; O'Hare et al, 1981); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan et al, 1981); neo, который придает устойчивость к аминогликозиду G-418 (Colbere-Garapin et al, 1981); и hygro, который придает устойчивость к гигромицину.
После определения или получения способами инженерии полинуклеотидной последовательности, кодирующей конкретный полипептид, полинуклеотид можно встраивать в соответствующую экспрессирующую систему. В этом случае, авторы изобретения рассматривают полинуклеотид, кодирующий полипептид связывающего лиганд домена sGPCR. Полинуклеотид можно экспрессировать в любом количестве экспрессирующих систем для рекомбинантных ДНК с получением большого количества полипептидного продукта, который затем можно очищать и/или выделять для применения в качестве лекарственного средства или для вакцинации животных с получением антисыворотки, или в определенных аспектах этого изобретения в качестве антагониста лиганда GPCR и/или активации GPCR. В следующих аспектах sGPCR по этому изобретению можно применять в способах выявления, скрининга или определения лигандов, рецепторов или агониста и/или антагониста GPCR. A полинуклеотид по этому изобретению можно экспрессировать с получением полипептида связывающего лиганд домена GPCR, связывающего лиганд домена GPCR семейства B, связывающего лиганд домена GPCR семейства B1, связывающего лиганд домена sCRFR или связывающего лиганд домена CRFR2, содержащего аминокислотную последовательность, включающую всю или часть аминокислотной последовательности, как указано в списке последовательностей, например, SEQ ID NO:2, 4, 6, 8, 10, 12 или 15.
В качестве альтернативы рекомбинантным полипептидам можно получать синтетические пептиды, соответствующие полипептидам по этому изобретению, включая антигенные пептиды. Такие антигенные пептиды обладают длиной по меньшей мере шесть аминокислотных остатков, и они могут содержать вплоть до приблизительно 35 остатков. Автоматические устройства для синтеза пептидов включают устройства, доступные от Applied Biosystems (Foster City, CA). Применение таких небольших пептидов для вакцинации, как правило, требует конъюгации пептида с иммуногенным белком-носителем, таким как поверхностный антиген гепатита B, гемоцианин лимфы улитки или бычий сывороточный альбумин. Способы проведения такой конъюгации хорошо известны в данной области.
1. Очистка экспрессированных белков
Следующие аспекты настоящего изобретения относятся к очистке и выделению и в конкретных вариантах осуществления к значительной очистке белка или пептида, содержащего весь связывающий лиганд домен sGPCR или его часть. Как используют в настоящем описании, подразумевают, что термин "очищенный или выделенный белок или пептид" относятся к композиции, поддающейся выделению из других компонентов, где белок, полипептид или пептид очищают до любой степени, относительно его доступного природного состояния, т.е. в данном случае, относительно его чистоты в организме или ткани. Очищенный или выделенный белок или пептид, таким образом, также относится к белку или пептиду, не содержащему окружающих его компонентов, с которыми он встречается в природе. Очищенный или выделенный белок или полипептид может обладать чистотой, выше чем или по меньшей мере 70, 75, 80, 85, 90, 95, 98 или 99% чистоты.
Главным образом, "очищенный" относится к композиции белка или пептида, которую подвергали фракционированию для удаления различных других компонентов и где композиция по существу сохраняет свою выраженную активность. В случае, когда используют термин "по существу очищенный", это обозначение относится к композиции, в которой белок или пептид является основным компонентом композиции, составляя, например, приблизительно 50% или более белков в композиции.
Различные способы для количественного определения степени очистки белка или пептида станут известными специалистам в данной области, учитывая настоящее описание. Они включают в себя, например, определение специфичной активности (например, активности связывания с лигандом GPCR, включая, но не ограничиваясь ими, CRF или лиганд из семейства CRF) активной фракции, или оценку количества полипептидов во фракции посредством анализа SDS/PAGE. Предпочтительным способом для оценки чистоты фракции является вычисление специфичной активности фракции для сравнения ее со специфичной активностью исходного экстракта и таким образом для вычисления степени чистоты, оцениваемой в настоящем описании "кратным показателем очистки". Конкретные единицы, используемые для отображения величины активности, которая может включать в себя активность или аффинность связывания, безусловно, зависят от конкретного выбранного способа анализа.
Различные способы, пригодные для применения для очистки белка, хорошо известны специалистам в данной области. Они включают в себя, например, осаждение сульфатом аммония, полиэтиленгликолем, антителами и т.п. или посредством денатурации теплом, с последующим центрифугированием; стадии хроматографии, такие как ионообменная хроматография, гель-фильтрация, хроматография с обращенной фазой, хроматография с гидроксилапатитом и/или аффинная хроматография; изоэлектрическое фокусирование; гель-электрофорез; сочетание этих и других способов. Как известно в данной области, в основном полагают, что порядок проведения различных стадий очистки можно изменять или что определенные стадии можно пропускать и тем не менее иметь в результате пригодный способ получения по существу очищенного белка или пептида.
Не существует общих требований, чтобы белок или пептид всегда был предоставлен в его наиболее очищенном состоянии. Действительно, предполагается, что очищенные в менее значительной степени продукты являются пригодными в определенных вариантах осуществления. Частичную очистку можно проводить с использованием меньшего количества стадий очистки в сочетании или посредством использования различных форм указанной общей схемы очистки. Например, следует учитывать, что катионообменная колоночная хроматография, проводимая с использованием устройства для ВЭЖХ, в большинстве случаев приведет к более высокой очистке, чем тот же способ, проводимый с использованием системы для хроматографии с низким давлением. Способы, характеризующиеся более низкой степенью относительной очистки, могут обладать преимуществами в общем получении белкового продукта или в поддержании активности экспрессированного белка.
E. Получение антител, специфичных в отношении sGPCR
Для некоторых вариантов осуществления является желательной продукция антител, которые с высокой специфичностью связываются с белковым продуктом(ами) выделенной нуклеиновой кислоты, кодирующей sGPCR, включая, но не ограничиваясь этим, sCRFR2α. В определенных аспектах предусмотрено получение антитела, которое распознает или связывается с C-концом GPCR, в частности варианта по сплайсингу, такого как вариант по сплайсингу sCRFR2α, и таким образом его можно использовать для того, чтобы отличать полипептид sGPCR от связанного с мембраной рецептора. Такие антитела можно использовать в любом из множества способов применения, известных специалистам в данной области, включая, но не ограничиваясь ими: способы иммунодетекции, способы иммунопреципитации, способы анализа ELISA,
способы очистки белка, и т.д. Способы получения и охарактеризации антител хорошо известны в данной области (см., например, Harlow and Lane, 1988, включенный в настоящее описание в качестве ссылки).
Способы получения поликлональных антител хорошо известны в данной области. В кратком изложении поликлональное антитело получают посредством иммунизации животного антигенной композицией и забора антисыворотки от этого иммунизированного животного. Для получения антисыворотки можно использовать широкий диапазон видов животных. Как правило, животное, используемое для получения антисыворотки, представляет собой кролика, мышь, крысу, хомяка, морскую свинку, лошадь или козу. Вследствие относительно большого объема крови у кроликов, для получения поликлональных антител предпочтительно выбирать кролика.
Как хорошо известно в данной области, данная композиция может варьировать по своей иммуногенности. Таким образом, часто необходимо стимулировать иммунную систему организма-хозяина, чего можно достигать посредством присоединения к пептидному или полипептидному иммуногену носителя. Иллюстративные и предпочтительные носители представляют собой гемоцианин лимфы улитки (KLH) и бычий сывороточный альбумин (BSA). Также в качестве носителей можно использовать другие альбумины, такие как овальбумин, сывороточный альбумин мыши или сывороточный альбумин кролика. Средства для конъюгации полипептида с белком-носителем хорошо известны в данной области и включают в себя глутаральдегид, сложный эфир м-малеимидобензоил-N-гидроксисукцинимида, карбодиимид и бис-диазотированный бензидин.
Как также хорошо известно в данной области, иммуногенность конкретной композиции иммуногена можно усилить посредством применения неспецифичных стимуляторов иммунного ответа, известных как адъюванты. Иллюстративные и предпочтительные адъюванты включают в себя полный адъювант Фрейнда (неспецифичный стимулятор иммунного ответа, содержащий убитые Mycobacterium tuberculosis), неполные адъюванты Фрейнда и адъювант гидроксид алюминия.
Моноклональные антитела (MAb) можно легко получать посредством применения хорошо известных способов, таких как способы, представленые в патенте США 4196265, включенном в настоящее описание в качестве ссылки. Как правило, этот способ включает в себя иммунизацию пригодного животного выбранной композицией иммуногена, например очищенным или частично очищенным экспрессированным белком, полипептидом или пептидом. Иммунизирующую композицию вводят таким образом, чтобы эффективно стимулировать антителопродуцирующие клетки.
Животным инъецируют антиген, как описано выше. После иммунизации соматические клетки, которые имеют возможность продуцировать антитела, конкретно B-лимфоциты (B-клетки) отбирают для применения в протоколе получения MAb. Часто иммунизируют группу животных и селезенку животного с наивысшим титром антитела удаляют и посредством гомогенизации селезенки с помощью шприца получают лимфоциты селезенки.
Затем проводят слияние продуцирующих антитела B-лимфоцитов из иммунизированного животного с иммортализованными клетками миеломы, как правило, из того же вида, что и животное, которого иммунизировали. Клеточные линии миеломы, пригодные для применения в способах слияния с получением гибридомы, предпочтительно не продуцируют антитело, обладают высокой эффективностью слияния и обладают дефецитом фермента, который приводит к их неспособности к росту в определенной селективной среде, которая поддерживает рост только требуемых слитых клеток (гибридом). Можно использовать любые из множества клеток миеломы, которые известны специалистам в данной области (Goding, 1986). Например, в случае, когда иммунизированное животное представляет собой мышь, можно использовать P3-X63/Ag8, X63-Ag8.653, NS1/1.Ag 4 1, Sp210-Ag14, FO, NSO/U, MPC-11, MPC11-X45-GTG 1.7 и S194/5XX0 Bul; для крыс можно использовать R210.RCY3, Y3-Ag 1.2.3, IR983F и 4B210; и U-266, GM1500-GRG2, LICR-LON-HMy2 и UC729-6 являются пригодными применительно к слиянию с клетками человека.
Одной предпочтительной клеткой миеломы мыши является клеточная линия миеломы NS-1 (также называемая P3-NS-1-Ag4-1), которая широко доступна в NIGMS Human Genetic Mutant Cell Repository при запросе клеточной линии с номером для хранения GM3573. Другая клеточная линия миеломы мыши, которую можно использовать, представляет собой устойчивую к 8-азагуанину клеточную линии миеломы SP2/0, не являющуюся продуцентом.
Большие количества моноклональных антител по настоящему изобретению также можно получать посредством размножения клеток гибридомы in vivo. Клеточные клоны инъецируют в млекопитающего, которое является гистосовместимым с исходными клетками, например, в изогенных мышей для того, чтобы вызвать рост продуцирующих антитело опухолей. Необязательно, перед инъекцией животных подвергают первичному воздействию посредством углеводорода, особенно масел, таких как пристан (тетраметилпентадекан).
В соответствии с настоящим изобретением, можно получать фрагменты моноклонального антитела способами, которые включают в себя расщепление ферментами, такими как пепсин или папаин, и/или расщепление дисульфидных связей посредством химического восстановления. Альтернативно фрагменты моноклонального антитела, охватываемые настоящим изобретением, можно синтезировать с использованием автоматического устройства для синтеза пептидов, или посредством экспрессии полноразмерного полинуклеотида или полинуклеотидных фрагментов, кодирующих полное Mab или его часть.
Конъюгаты антитела можно получать известными в данной области способами, например, посредством проведения реакции антитела с ферментом в присутствии связующего вещества, такого как глутаральдегид или соль йодной кислоты. Конъюгаты с флуоресцеиновыми маркерами получают в присутствии этих связующих веществ или посредством проведения реакции с изотиоцианатом. Конъюгаты с хелатами металлов получают сходным образом. Другие группы, с которыми можно конъюгировать антитела, включают в себя радионуклиды, такие как 3H, 125I, 131I, 32P, 35S, 14C, 51Cr, 36Cl, 57Co, 58Co, 59Fe, 75Se, 152Eu и 99mTc. Меченые радиоактивной меткой антитела по настоящему изобретению можно получать в соответствии с хорошо известными способами. Например, антитела можно йодировать посредством контактирования с иодидом натрия или калия и химическим окислителем, таким как гипохлорит натрия, или с ферментным окислителем, таким как лактопероксидаза. Антитела в соответствии с этим изобретением могут быть мечеными технецием99 посредством процесса лигандного обмена, например посредством восстановления пертехната раствором олова, хелатирования восстановленного технеция в колонке с сефадексом и нанесения антитела в эту колонку, или посредством способов прямого мечения, например посредством инкубации пертехната, восстановителя, такого как SnCl2, буферного раствора, такого как раствор фталата натрия-калия, и антитела.
НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ ПОЛИПЕПТИДЫ SGPCR
Настоящее изобретение относится к нуклеиновым кислотам, которые кодируют весь sGPCR, или его часть, такой как, но не ограничиваясь ими, полипептид GPCR, GPCR семейства B, GPCR семейства B1, CRFR или CRFR2, и может включать в себя различные последовательности нуклеиновых кислот, требуемые для доставки последовательности нуклеиновой кислоты, а также транскрипции и/или трансляции последовательности нуклеиновой кислоты. Молекулы нуклеиновых кислот по этому изобретению могут включать в себя смежные участки нуклеиновой кислоты различной длины, например приблизительно 10, 15, 17, 20, 25, 30, 35, 40, 45, 50, 55, 60, 70, 75, 80, 85, 90, 95, 100, 125, 150, 175, 200, 250, 300, 350, 400, 500, 600, 700, 800, 900, 1000, 1250, 1500, 1750, 2000, 2100, включая все или часть полноразмерных последовательностей нуклеиновых кислот из списка последовательностей, например SEQ ID NO:1, 3, 5, 7, 9, 11, 13 или 14, или предусмотрены полинуклеотиды тех GPCR, которые упомянуты в настоящем описании, их фрагменты, мРНК или кДНК, содержащие последовательности, описанные или упоминаемые в настоящем описании, и мутантные формы каждой из них. Также предусмотрены молекулы, которые являются комплементарными упомянутым выше последовательностям, и которые связываются с этими последовательностями в условиях высокой строгости. Эти зонды являются пригодными в различных вариантах осуществления гибридизации, таких как саузерн-блоттинг и нозерн-блоттинг.
Вблизи описанных нуклеотидных последовательностей можно конструировать различные зонды и праймеры. Длина праймеров может быть любой, но, как правило, она составляет 10-20 оснований. В конкретных аспектах зонд или праймер можно использовать для выявления и скрининга на наличие альтернативно сплайсированных форм GPCR, таких как, но не ограничиваясь ими, ген CRFR2, который включает в себя точку сплайсинга экзон 5/экзон 7 (также она может быть описана как точка соединения экзон 3/экзон 5, если она относится к транскрипции CRFR2α). Эти зонды или праймеры могут либо гибридизоваться с уникальной последовательностью сконструированной нуклеиновой кислоты или областью сплайсинга, или они могут амплифицировать характерный элемент сконструированной нуклеиновой кислоты или область сплайсинга. Приписывая числовые значения последовательности, например первый остаток представляет собой 1, второй остаток представляет собой 2, и т.д., можно предложить следующий алгоритм для определения всех праймеров:
с n по n+y,
где n представляет собой число от 1 до последнего номера последовательности и y представляет собой длину праймера минус один, где n + y не превышает последний номер последовательности. Таким образом, для 10-мера зонды соответствуют основаниям с 1 по 10, со 2 по 11, с 3 по 12... и так далее. Для 15-мера зонды соответствуют основаниям с 1 по 15, со 2 по 16, с 3 по 17... и так далее. Для 20-мера зонды соответствуют основаниям с 1 по 20, со 2 по 21, с 3 по 22... и так далее.
В определенных аспектах последовательности нуклеиновых кислот по этому изобретению можно использовать для кодирования различных полипептидов, описанных в настоящем описании. В одном варианте осуществления настоящего изобретения, последовательности нуклеиновых кислот можно использовать в качестве зондов для гибридизации или праймеров для амплификации. В определенных вариантах осуществления эти зонды и праймеры состоят из олигонуклеотидных фрагментов. Такие фрагменты должны обладать достаточной длиной для обеспечения специфичной гибридизации с образцом РНК или ДНК, экстрагированным из ткани. Последовательности, как правило, обладают 10-20 нуклеотидами, однако они могут быть более длинными. Более длинные последовательности, например, 40, 50, 100, 500 и даже вплоть до полноразмерных последовательностей, являются предпочтительными для определенных вариантов осуществления.
Применение зонда для гибридизации длиной между 17 и 100 нуклеотидами делает возможным образование дуплексной молекулы, которая является как стабильной, так и селективной. Молекулы, обладающие комплементарными последовательностями на протяжении длины более чем 20 оснований, являются в большинстве случаев предпочтительными, в целях повышения стабильности и селективности гибрида и таким образом повышения качества и относительного количества полученных конкретных гибридных молекул. Предпочтительной является конструирование молекул нуклеиновых кислот, обладающих длиной от 20 до 30 нуклеотидов, или, если желательно, более. Такие фрагменты можно легко получать, например, посредством прямого синтеза фрагмента химическими способами или посредством встраивания выбранных последовательностей в рекомбинантные векторы для продукции рекомбинантными способами. Таким образом, нуклеотидные последовательности по этому изобретению можно использовать вследствие их способности селективно формировать дуплексные молекулы с комплементарными участками генов, полинуклеотидов или РНК, или для получения праймеров для амплификации ДНК или РНК из тканей. В зависимости от предусмотренного способа применения, может быть желательным использование различных условий гибридизации для достижения различных степеней селективности зонда в отношении последовательности-мишени.
Для способов применения, требующих высокой селективности, как правило, для получения гибридов может быть желательным использование относительно строгих условий или условий высокой строгости, например, можно выбирать условия с относительно низким содержанием соли и/или с высокой температурой, такие как обеспечивают посредством от приблизительно 0,02 М до приблизительно 0,10 М NaCl при температурах от приблизительно 50°C до приблизительно 70°C. Такие условия высокой строгости допускают небольшое, если допускают, несовпадение в спаривании между зондом и матрицей или цепью-мишенью, и, в частности, являются пригодными для выделения конкретных генов или выявления конкретных транскриптов мРНК. В целом, понятно, что условия можно делать более строгими посредством добавления повышенных количеств формамида.
В определенных вариантах осуществления для определения гибридизации предпочтительным является использование последовательностей нуклеиновых кислот по настоящему изобретению в сочетании с пригодными средствами для определения, такими как флуоресцентная или радиоактивная метка. Широкое множество соответствующих индикаторных средств известно в данной области, включая флуоресцентные, радиоактивные, ферментные или другие лиганды, такие как авидин /биотин, которые поддаются выявлению.
Для способов применения, в которых участки нуклеиновых кислот по настоящему изобретению встраивают в экспрессирующие векторы, такие как плазмиды, космиды или вирусные полинуклеотиды, эти участки можно объединять с другими последовательностями ДНК, такими как промоторы, сигналы для полиаденилирования, участки для рестрикции ферментами, полилинкеры, другие кодирующие участки и т.п., таким образом, их общая длина может значительно варьировать. Предполагается, что можно использовать фрагмент нуклеиновой кислоты практически любой длины, с общей длиной, предпочтительно ограниченной простотой получения и применения в предусмотренном протоколе для рекомбинантных ДНК.
Участки ДНК, кодирующие конкретный полинуклеотид, можно встраивать в рекомбинантные клетки-хозяева и использовать для экспрессии sGPCR, таких как, но не ограничиваясь ими, полипептид sCRFR2α. Альтернативно посредством применения способов генетической инженерии, можно использовать субфрагменты или производные выбранных полинуклеотидов.
На протяжении этой заявки, подразумевают, что термин "экспрессирующая конструкция" включает любой тип генетической конструкции, содержащий нуклеиновую кислоту, обладающую последовательностью, определяющей продукт, такой как, но не ограничиваясь ими, продукт, кодирующий полипептид, в котором часть или вся последовательность нуклеиновой кислоты может быть транскрибирована. Транскрипт может, но не должен, транслироваться в белок. Таким образом, в определенных вариантах осуществления, экспрессия включает в себя как транскрипцию полинуклеотида, так и трансляцию РНК в полипептидный продукт.
В предпочтительных вариантах осуществления нуклеиновая кислота находится под транскрипционным контролем промотора. "Промотор" относится к последовательности ДНК, распознаваемой синтетическим аппаратом клетки, или встроенным синтетическим аппаратом, требуемой для инициации специфичной транскрипции полинуклеотида. Выражение "под транскрипционным контролем" означает, что промотор находится в правильном расположении и ориентации относительно нуклеиновой кислоты, чтобы контролировать инициацию РНК-полимеразы и экспрессию полинуклеотида. Термин промотор используют в настоящем описании в отношении группы элементов транскрипционного контроля, которые сгруппированы в области участка инициации для РНК-полимеразы, в частности РНК полимеразы II. В определенных аспектах, по меньшей мере один элемент каждого промотора функционирует как положение для начала синтеза РНК. Наиболее известным его примером является TATA-бокс, однако в некоторых промоторах отсутствует TATA-бокс, таких как промотор гена терминальной дезоксинуклеотидилтрансферазы млекопитающих и промотор поздних генов SV40, отдельный элемент, лежащий выше точки начала, сам по себе способствует фиксации точки инициации.
Полагают, что конкретный промотор, который используют для контроля экспрессии нуклеиновой кислоты, не является особенно важным, при условии, что он способен экспрессировать нуклеиновую кислоту в клетке-мишени. Таким образом, в случае, когда мишенью является клетка человека, предпочтительным является расположение кодирующего участка нуклеиновой кислоты рядом с промотором и под контролем промотора, который может экспрессироваться в клетке человека. В общем, такой промотор может включать в себя промотор либо человека, либо вируса.
В различных других вариантах осуществления для обеспечения высокого уровня экспрессии трансгенов можно использовать предранний промотор гена цитомегаловируса (CMV) человека, ранний промотор SV40 и длинный концевой повтор вируса саркомы Рауса. Также для достижения экспрессии трансгена предусмотрено применение других промоторов вирусов или клеток млекопитающих или фагов бактерий, которые хорошо известны в данной области, при условии что уровни экспрессии являются достаточными для данной цели. В контексте настоящего изобретения можно использовать несколько элементов/промоторов, как описано ниже, для регуляции экспрессии полинуклеотида, такого как трансген. Подразумевается, что этот перечень не является полным для всех возможных элементов, вовлеченных в запуск экспрессии трансгена, но представляет собой только их иллюстрацию.
Также для запуска экспрессии полинуклеотида можно использовать любое сочетание промотор/энхансер (как в Eucaryotic Promoter Data Base EPDB). Применение цитоплазматической экспрессирующей системы T3, T7 или SP6 представляет собой другой возможный вариант осуществления. Эукариотические клетки могут обеспечивать цитоплазматическую транскрипцию с определенных бактериальных промоторов, если предоставлена соответствующая бактериальная полимераза, либо в качестве части комплекса для доставки, либо в качестве дополнительной генетической конструкции для экспрессии. Применение системы бакуловируса приводит к высокому уровню экспрессии с мощного промотора полиэдрина.
Промоторы включают в себя, но не ограничиваются ими, промоторы тяжелой цепи иммуноглобулина, легкой цепи иммуноглобулина, T-клеточного рецептора, HLA DQ α и DQ β, β-интерферона, интерлейкина-2, рецептора для интерлейкина-2, MHC класса II 5, MHC класса II HLA-DRα, β-актина, креатинкиназы мышц, преальбумина (транстиретин), эластазы I, металлотеонеина, коллагеназы, гена альбумина, α-фетопротеина, α-глобина, β-глобина, c-fos, c-HA-ras, инсулина, молекулы адгезии нейронов (NCAM), α1-антитрипсина, гистона H2B (TH2B), коллагена мыши или I типа, регулируемых глюкозой белков (GRP94 и GRP78), гормона роста крысы, амилоида сыворотки человека A (SAA), тропонина I (TN I), тромбоцитарного фактора роста, мышечной дистрофии Дюшена, SV40, полиомы, ретровирусов, вируса папилломы, вируса гепатита B, вируса иммунодефицита человека, цитомегаловируса, вируса лейкоза гиббонов.
Различные элементы (индуцирующие элементы) включают в себя, но не ограничиваются ими MT II (форболовый эфир (TPA), тяжелые металлы); MMTV (глюкокортикоиды, β-интерферон, poly(rI)X, poly(rc)); аденовирус 5 E2 (Ela); c-jun (форболовый эфир (TPA), H2O2); коллагеназу (форболовый эфир (TPA)); стромелизин (форболовый эфир (TPA), IL-1); SV40 (форболовый эфир (TPA)); ген MX мыши (интерферон, вирус болезни Ньюкасла); ген GRP78 (A23187); α-2-макроглобулин (IL-6); виментин (сыворотка); ген MHC класса I H-2kB (интерферон); HSP70 (Ela, большой T-антиген SV40); пролиферин (форболовый эфир-TPA); фактор некроза опухоли (FMA); и α-ген тиреостимулирующего гормона (тиреоидного гормона).
Как правило, для достижения соответствующего полиаденилирования транскрипта можно добавлять сигнал полиаденилирования. Полагают, что природа сигнала полиаденилирования не является особенно важной для успешного применения на практике этого изобретения и можно использовать любую такую последовательность. Предпочтительные варианты осуществления включают в себя сигнал полиаденилирования SV40 и сигнал полиаденилирования бычьего гормона роста, которые являются удобными и о которых известно, что они хорошо функционируют в различных клетках-мишенях. Также в качестве элемента экспрессирующей кассеты предусмотрено добавление терминатора. Эти элементы могут выполнять функцию повышения уровней транскрипции и сведения к минимуму считывания с кассеты в других последовательностей.
В различных вариантах осуществления этого изобретения, экспрессирующая конструкция может содержать вирус или полученную способами инженерии конструкцию из вирусного генома. Способность определенных вирусов проникать в клетки через рецептор-опосредованный эндоцитоз и встраиваться в геном клетки-хозяина, и экспрессировать вирусные гены стабильно и эффективно, привела к тому, что они стали привлекательными претендентами для переноса чужеродных генов в клетки млекопитающих (Ridgeway, 1988; Nicolas and Rubenstein, 1988; Baichwal and Sugden, 1986; Temin, 1986). Первыми вирусами, используемыми в качестве векторов, были ДНК-вирусы, включающие в себя паповавирусы (вирус обезьяны 40, бычий вирус папилломы и вирус полиомы) (Ridgeway, 1988; Baichwal and Sugden, 1986), и аденовирусы (Ridgeway, 1988; Baichwal and Sugden, 1986), и аденоассоциированные вирусы. Ретровирусы также представляют собой приемлемые носители для переноса генов (Nicolas and Rubenstein, 1988; Temin, 1986), также как вирус коровьей оспы (Ridgeway, 1988) и аденоассоциированный вирус (Ridgeway, 1988). Такие векторы можно использовать для (i) трансформации клеточных линий in vitro в целях экспрессии представляющих интерес G-белков или (ii) трансформации in vitro или in vivo для получения терапевтических полипептидов для генной терапии.
В альтернативном варианте осуществления используемые нуклеиновые кислоты, кодирующие sGPCR, могут по существу кодировать антисмысловые конструкции, которые гибридизуются во внутриклеточных условиях с sGPCR или другой кодирующей нуклеиновой кислотой. Подразумевают, что термин "антисмысловая конструкция" относится к нуклеиновым кислотам, предпочтительно олигонуклеотидам, комплементарным основным последовательностям ДНК-мишени или РНК-мишени.
Как используют в настоящем описании, термин "комплементарный" означает последовательности нуклеиновых кислот, которые являются по существу комплементарными на протяжении всей их длины и обладают очень небольшим количеством ошибок спаривания. Например, последовательности нуклеиновых кислот длиной пятнадцать оснований можно называть комплементарными, если они обладают комплементарным нуклеотидом в тринадцати или четырнадцати положениях только с одной ошибкой спаривания. Понятно, что последовательности нуклеиновых кислот, которые являются "полностью комплементарными" представляют собой последовательности нуклеиновых кислот, которые являются полностью комплементарными по их всей длине и не обладают ошибками спаривания оснований.
A. Выявление и количественное определение нуклеиновых кислот
Один вариант осуществления настоящего изобретения включает в себя способ выявления нуклеиновой кислоты sGPCR, такой как, но не ограничиваясь ими, нуклеиновые кислоты CRFR2α, в биологическом образце посредством амплификации и выявления нуклеиновых кислот, соответствующих sGPCR. Биологический образец может представлять собой любую ткань или жидкость, в которой может находиться полинуклеотид. Нуклеиновую кислоту, используемую в качестве матрицы для амплификации, выделяют из клеток, содержащихся в биологическом образце, в соответствии со стандартными способами (Sambrook et al., 1989). Нуклеиновая кислота может представлять собой фракционированную или целую клеточную РНК.
Пары праймеров, которые селективно гибридизуются с нуклеиновыми кислотами, соответствующими sGPCR, приводят в контакт с выделенной нуклеиновой кислотой в условиях, которые обеспечивают селективную гибридизацию. После гибридизации комплекс нуклеиновая кислота:праймер приводят в контакт с одним или несколькими ферментами, которые способствуют зависимому от матрицы синтезу нуклеиновой кислоты. Многократные раунды амплификации, также называемые "циклами", проводят до тех пор, пока не получат достаточное количество продукта амплификации. Для продуктов амплификации можно проводить детекцию. В определенных способах применения детекцию можно проводить визуально. Альтернативно детекция может включать в себя непрямое выявление продукта посредством хемилюминесценции, радиоактивной сцинтиграфии встроенной радиоактивной метки или флуоресцентной метки или даже посредством системы с использованием электрических или термических импульсных сигналов (Affymax technology; Bellus, 1994).
Для амплификации маркерных последовательностей, представленных в данном образце матрицы, является доступным ряд зависимых от матрицы процессов. Одним из наиболее известных способов амплификации является полимеразная цепная реакция (обозначаемая как ПЦР), которая подробно описана в патентах США 4683195, 4683202 и 4800159, и в Innis et al, 1990, все из которых включены в настоящее описание в качестве ссылок в полном объеме. Технологии полимеразной цепной реакции хорошо известны в данной области.
Другим способом амплификации является лигазная цепная реакция ("LCR"), описанная в EPA No. 320308, которая включена в настоящее описание в качестве ссылки в полном объеме. В патенте США 4883750 описан способ, сходный с LCR, для связывания пар зондов с последовательностью-мишенью. Также в качестве другого способа амплификации по настоящему изобретению можно использовать репликазу Qbeta, описанную в заявке PCT No. PCT 7 US 87/00880. Для амплификации нуклеиновых кислот по настоящему изобретению можно использовать способ изотермической амплификации, в котором для достижения амплификации молекул-мишеней, которые содержат нуклеотид 5'-[альфа-тио]трифосфаты на одной цепи участка рестрикции, используют эндонуклеазы рестрикции и лигазы, Walker et al, (1992), которая включена в настоящее описание в качестве ссылки в полном объеме. Более того, амплификация со смещением цепи (SDA) представляет собой другой способ проведения изотермической амплификации нуклеиновых кислот, которая включает в себя многократные раунды вытеснения цепи и синтеза, т.е. ник-трансляцию. Сходный способ, называемый цепной реакцией с репарацией (RCR), включает в себя отжиг нескольких зондов на протяжении участка, предназначенного для амплификации, с последующей реакцией репарации, в которой представлены только два из четырех оснований. Конкретные последовательности-мишени также можно определять с использованием циклической реакции с зондом (CPR). В соответствии с настоящим изобретением можно использовать другой способ амплификации, описанный в заявке GB No. 2202328 и в заявке PCT No. PCT/US 89/01025, все из которых включены в настоящее описание в качестве ссылок в полном объеме. Другие способы амплификации нуклеиновых кислот включают в себя основанные на транскрипции системы для амплификации (TAS), включая основанную на последовательности нуклеиновой кислоты амплификацию (NASBA) и 3SR (Kwoh et al, 1989); заявка PCT WO 88/10315, включенная в настоящее описание в качестве ссылки в полном объеме.
После амплификации может быть желательным разделение продукта амплификации и матрицы и избыточных праймеров в целях определения того, произошла ли специфичная амплификация. В одном варианте осуществления продукты амплификации разделяют посредством гель-электрофореза в агарозе, агароза-акриламид или полиакриламиде с использованием стандартных способов. См. Sambrook et al., 1989.
Для разделения можно использовать способы хроматографии. Существует множество типов хроматографии, которые можно использовать в соответствии с настоящим изобретением: адсорбция, фракционирование, ионный обмен и молекулярное сито, и множество специализированных способов их применения включают в себя хроматографию на колонке, на бумаге, тонкослойную и газовую хроматографию (Freifelder, 1982).
IV. СПОСОБЫ ЭКСПРЕССИИ ГЕНА SGPCR
В одном варианте осуществления настоящего изобретения предусмотрены способы повышения экспрессии sGPCR в клетке, такого как, но не ограничиваясь этим, экспрессия sCRFR2α. Они особенно пригодны в случае повреждения белка или если экспрессия белка не является достаточной для нормального выполнения функции. Это позволит ослабить симптомы заболевания, которому подвергаются в результате дефицита sGPCR, гиперактивации GPCR или избытка лиганда GPCR.
Основным подходом для повышения sGPCR является контактирование клетки, ткани, животного или субъекта или введение в клетку, ткань, животному или субъекту полипептида sGPCR. Несмотря на то, что предпочтительно, чтобы белок можно было доставлять непосредственно, возможный вариант осуществления включает в себя доставку в клетку или в соседние клетки нуклеиновой кислоты, кодирующей полипептид sGPCR. После этой доставки полипептид sGPCR синтезируется с помощью транскрипционного и трансляционного аппарата клетки-хозяина, а также любого аппарата, который может быть доставлен посредством экспрессирующей конструкции. Предусмотрены совместно воздействующие цис-регуляторные элементы, необходимые для поддержания экспрессии полинуклеотида sGPCR, в форме экспрессирующей конструкции. Также возможно, чтобы экспрессию кодируемой посредством вирусов sGPCR можно было стимулировать или усиливать, или экспрессированный полипептид можно было стабилизировать, таким образом достигая одинакового или сходного эффекта.
В целях достижения экспрессии конструкций, кодирующих полинуклеотиды sGPCR, экспрессирующую конструкцию можно доставлять в клетку посредством вектора для доставки. Одним механизмом доставки является доставка посредством вирусной инфекции, где экспрессирующую конструкцию энкапсидируют в вирусную частицу, которая доставляет либо реплицирующуюся, либо нереплицирующуюся нуклеиновую кислоту.
Способность определенных вирусов проникать в клетки посредством рецептор-опосредуемого эндоцитоза, встраиваться в геном клетки-хозяина и стабильно и эффективно экспрессировать вирусные гены делает их привлекательными претендентами для переноса чужеродных генов в клетки млекопитающих (Ridgeway, 1988; Nicolas and Rubenstein, 1988; Baichwal and Sugden, 1986; Temin, 1986). Первыми вирусами, используемыми в качестве генных векторов, были ДНК-вирусы, включающие в себя паповавирусы (вирус обезьяны 40, бычий вирус папилломы и вирус полиомы) (Ridgeway, 1988; Baichwal and Sugden, 1986) и аденовирусы (Ridgeway, 1988; Baichwal and Sugden, 1986). Они обладают относительно низкой вместимостью для чужеродных последовательностей ДНК и обладают ограниченным диапазоном хозяев. Более того, их онкогенный потенциал и цитопатический эффект в пермиссивных клетках вызывает опасения в отношении безопасности. Они могут вмещать только вплоть до 8 т.п.н. чужеродного генетического материала, однако их можно легко вводить во множество клеточных линий и лабораторных животных (Nicolas and Rubenstein, 1988; Temin, 1986).
Ретровирусы представляют собой группу одноцепочечных РНК-вирусов, характеризующихся способностью превращать РНК в двухцепочечную ДНК в инфицированных клетках; их также можно использовать в качестве векторов. В качестве экспрессирующих конструкций в соответствии с настоящим изобретением можно использовать другие вирусные векторы. Можно использовать векторы из вирусов, таких как вирус коровьей оспы (Ridgeway, 1988; Baichwal and Sugden, 1986; Coupar et al, 1988) аденоассоциированный вирус (AAV) (Ridgeway, 1988; Baichwal and Sugden, 1986; Hermonat and Muzycska, 1984) и вирусы герпеса. Они обладают несколькими привлекательными свойствами для различных клеток млекопитающих (Friedmann, 1989; Ridgeway, 1988; Baichwal and Sugden, 1986; Coupar et al, 1988; Horwich et al, 1990).
Также по этому изобретению предусмотрено несколько невирусных способов переноса экспрессирующих конструкций в культивируемые клетки млекопитающих. Они включают в себя осаждение с фосфатом кальция (Graham and Van Der Eb, 1973; Chen and Okayama, 1987; Rippe et al, 1990) способ с DEAE-декстраном (Gopal, 1985), электропорацию (Tur-Kaspa et al, 1986; Potter et al, 1984), прямую микроинъекцию (Harland and Weintraub, 1985), заполненные ДНК липосомы (Nicolau and Sene, 1982; Fraley et al, 1979) и комплексы липофектамин-ДНК, ультразвуковое воздействие на клетки (Fechheimer et al, 1987), генную бомбардировку с использованием высокоскоростных микроснарядов (Yang et. al, 1990) и опосредуемую рецептором трансфекцию (Wu and Wu, 1987; Wu and Wu, 1988). Некоторые из этих способов можно успешно адаптировать для применения in vivo или ex vivo, как описано ниже.
В другом варианте осуществления этого изобретения экспрессирующая конструкция может просто состоять из депротеинизированной рекомбинантной ДНК или плазмид. Перенос конструкции можно проводить любыми способами, упомянутыми выше, которые физически или химически обеспечивают проницаемость клеточной мембраны. В частности, они является применимыми для переноса in vitro, однако также их можно использовать для применения in vivo. Другой вариант осуществления этого изобретения для переноса экспрессирующей конструкции депротеинизированной ДНК в клетки может вовлекать бомбардировку частицами. В основе этого способа лежит способность ускорять покрытые ДНК микроснаряды до высокой скорости, позволяя им перфорировать клеточные мембраны и проникать в клетки, не вызывая их гибели (Klein et al, 1987). Было разработано несколько устройств для ускорения мелких частиц. В основе одного такого устройства лежит высоковольтный разряд с получением электрического тока, который в свою очередь обеспечивает движущую силу (Yang et al, 1990). Используемые микроснаряды состоят из биологически инертных веществ, таких как вольфрамовые или золотые сферы.
В следующем варианте осуществления этого изобретения экспрессирующие конструкции можно заключать в липосому. Липосомы представляют собой везикулярные структуры, характеризующиеся двухслойной фосфолипидной мембраной и внутренней водной средой. Многослойные липосомы обладают множеством липидных слоев, разделенных водной средой. Они формируются спонтанно, когда фосфолипиды суспендируют в избытке водного раствора. Липидные компоненты претерпевают самостоятельное перераспределение перед образованием закрытых структур и захватывают воду и растворенные вещества между липидными бислоями (Ghosh and Bachhawat, 1991). Также предусмотрены комплексы липофектамин-ДНК. Другие экспрессирующие конструкции, которые можно использовать для доставки нуклеиновой кислоты, кодирующей полинуклеотид sCRFR2α, в клетки, представляют собой опосредуемые рецептором носители для доставки. Они обладают преимуществом селективного захвата макромолекул посредством опосредуемого рецептором эндоцитоза практически во всех эукариотических клетках. Вследствие специфичного в отношении типа клеток распределения различных рецепторов, доставка может быть высокоспецифичной (Wu and Wu, 1993).
V. ФАРМАЦЕВТИЧЕСКИЕ ПРЕПАРАТЫ И СПОСОБЫ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ
В дополнительных вариантах осуществления настоящее изобретение относится к составу из одной или нескольких композиций полинуклеотида, полипептида и/или антитела, описанных в настоящем описании, в фармацевтически приемлемых растворах для введения в клетку, ткань, животному, пациенту или субъекту либо отдельно, либо в сочетании с одним или несколькими другими способами терапии.
Водные фармацевтические композиции по настоящему изобретению обладают эффективным количеством экспрессирующей sGPCR конструкции, экспрессирующей конструкции, которая кодирует терапевтический ген совместно с sGPCR, или белком sGPCR и/или соединением, которое модулирует активность или чувствительность лиганда GPCR, или другую эндокринную функцию. Такие композиции, как правило, растворены или диспергированы в фармацевтически приемлемом носителе или водной среде. "Эффективное количество" в целях терапии определяется как количество, которое приводит к поддающемуся клиническому определению отличию состояния у субъекта. Это количество варьирует в зависимости от вещества, состояния пациента, типа лечения и т.д.
Выражения "фармацевтически или фармакологически приемлемый" относятся к молекулярным системам и композициям, которые не вызывают значительной побочной, аллергической или другой нежелательной реакции при введении животному или человеку. Как используют в настоящем описании "фармацевтически приемлемый носитель" включает в себя любые и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие всасывание вещества и т.п. Применение таких сред и веществ для фармацевтически активных веществ хорошо известно в данной области. За исключением случаев, когда общепринятые среды или вещества являются несовместимыми с активными ингредиентами, предусмотрено их использование в терапевтических композициях.
В дополнение к соединениям, составленным для парентерального введения, таким как соединения для внутривенной или внутримышечной инъекции, другие фармацевтически приемлемые формы включают в себя, например, таблетки или другие твердые вещества для перорального введения; капсулы с замедленным высвобождением; и любую другую используемую в настоящее время форму, включая кремы, лосьоны, вещества для ингаляции и т.п.
Активные соединения по настоящему изобретению часто составляют для парентерального введения, например их составляют для инъекции посредством внутривенного, внутримышечного, подкожного и даже внутрибрюшинного способов. Получение водной композиции, которая содержит sGPCR отдельно или в сочетании с общепринятым лекарственным средством, в качестве активных ингредиентов станет известным специалистам в данной области с учетом настоящего описания. Как правило, такие композиции можно получать в качестве инъецируемых средств, либо в качестве жидких растворов, либо в качестве суспензий; также можно получать твердые формы, пригодные для применения, с получением растворов или суспензий при добавлении жидкости перед инъекцией; а также препараты могут быть эмульгированными.
Фармацевтические формы, пригодные для применения в качестве инъекций, включают в себя стерильные водные растворы или дисперсии; составы, включающие кунжутное масло, арахисовое масло или водный пропиленгликоль; и стерильные порошки для быстрого получения стерильных инъецируемых растворов или дисперсий. Во множестве случаев форма должна быть стерильной и должна быть жидкой, чтобы происходило легкое прохождение в шприц. Она должна быть стабильной в условиях изготовления и хранения и должна быть защищена от загрязняющего воздействия микроорганизмов, таких как бактерии и грибы.
Также носитель может представлять собой растворитель или диспергирующую среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.), их пригодные смеси и растительные масла. Необходимую текучесть можно поддерживать, например, посредством применения покрытия, такого как лецитин, посредством поддержания требуемого размера частиц в дисперсии и посредством применения поверхностно-активных веществ. Воздействие микроорганизмов можно предотвращать посредством различных антибактериальных и противогрибковых веществ, например парабенов, хлорбутанола, фенола, сорбиновой кислоты, тимеросала и т.п. Во многих случаях предпочтительным является добавление изотонических веществ, например сахаров или хлорида натрия. Длительного всасывания инъецируемой композиции можно достигать посредством применения композиций веществ, замедляющих всасывание, например моностеарата алюминия и желатина.
Стерильные инъецируемые растворы получают посредством объединения активного соединения в требуемых количествах в пригодном растворителе с различными другими ингредиентами, перечисленными выше, при необходимости, с последующей стерилизацией посредством фильтрации. Как правило, дисперсии получают посредством добавления различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную диспергирующую среду, и других требуемых ингредиентов среди тех, которые перечислены выше. В случае стерильных порошков, для получения стерильных инъецируемых растворов предпочтительные способы получения представляют собой способы вакуумной сушки и лиофилизации, в результате которых получают порошок активного ингредиента, плюс любые дополнительные требуемые ингредиенты из их предварительно стерилизованного фильтрацией раствора.
При составлении растворы вводят способом, совместимым с дозировкой состава, и в таких количествах, которые являются терапевтически эффективными. Составы легко вводят в виде множества дозированных форм, таких как инъецируемые растворы описанных выше типов, где даже капсулы для высвобождения лекарственного средства и т.п. являются применимыми.
В случае парентерального введения в водном растворе, например, раствор должен быть стабильно буферизованным, если необходимо, и жидкому разбавителю сначала необходимо придать изотоничность посредством пригодного солевого раствора или глюкозы. Эти конкретные водные растворы являются особенно пригодными для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В связи с этим, стерильные водные среды, которые можно использовать, станут известны специалистам в данной области с учетом настоящего описания. Например, одну дозу можно растворять в 1 мл изотонического раствора NaCl и либо добавлять к 1000 мл жидкости для промывания гиподермы или инъецировать в намеченную область инфузии (см. например, "Remington's Pharmaceuticals Sciences" (1980)). В зависимости от состояния субъекта, подвергаемого лечению, необходимо проводить некоторые изменения дозировки. Человек, ответственный за введение, в любом случае, определит соответствующую дозу для конкретного субъекта.
В определенных аспектах способов по этому изобретению, способ введения терапевтической композиции может представлять собой парентеральное введение. Парентеральное введение может представлять собой внутривенную инъекцию, подкожную инъекцию, внутримышечную инъекцию, костномозговую инъекцию, пероральное введение или их сочетание. В определенных аспектах композицию, содержащую sGPCR, вводят в количестве от приблизительно 0,1 до приблизительно 10 микрограмм/кг/масса тела на дозу. В определенных аспектах композицию, содержащую sGPCR, вводят в количестве от приблизительно 1 до приблизительно 5 микрограмм/кг/масса тела на дозу. В определенных аспектах композицию, содержащую sGPCR, вводят в количестве от приблизительно 1,2 до приблизительно 3,6 микрограмм/кг/масса тела на дозу. В определенных аспектах композицию, содержащую sGPCR, вводят в количестве от приблизительно 1,2 до приблизительно 2,4 микрограмм/кг/масса тела на дозу. В предпочтительных аспектах количество sGPCR, вводимых на дозу, может составлять приблизительно 0,1, приблизительно 0,2, приблизительно 0,3, приблизительно 0,4, приблизительно 0,5, приблизительно 0,6, приблизительно 0,7, приблизительно 0,8, приблизительно 0,9, приблизительно 1,0, приблизительно 1,1, приблизительно 1,2, приблизительно 1,3, приблизительно 1,4, приблизительно 1,5, приблизительно 1,6, приблизительно 1,7, приблизительно 1,8, приблизительно 1,9, приблизительно 2,0, приблизительно 2,1, приблизительно 2,2, приблизительно 2,3, приблизительно 2,4, приблизительно 2,5, приблизительно 2,6, приблизительно 2,7, приблизительно 2,8, приблизительно 2,9, приблизительно 3,0, приблизительно 3,1, приблизительно 3,2, приблизительно 3,3, приблизительно 3,4, приблизительно 3,5, приблизительно 3,6, приблизительно 3,7, приблизительно 3,8, приблизительно 3,9, приблизительно 4,0, приблизительно 4,1, приблизительно 4,2, приблизительно 4,3, приблизительно 4,4, приблизительно 4,5, приблизительно 4,6, приблизительно 4,7, приблизительно 4,8, приблизительно 4,9, приблизительно 5,0, приблизительно 5,1, приблизительно 5,2, приблизительно 5,3, приблизительно 5,4, приблизительно 5,5, приблизительно 5,6, приблизительно 5,7, приблизительно 5,8, приблизительно 5,9, приблизительно 6,0, приблизительно 6,1, приблизительно 6,2, приблизительно 6,3, приблизительно 6,4, приблизительно 6,5, приблизительно 6,6, приблизительно 6,7, приблизительно 6,8, приблизительно 6,9, приблизительно 7,0, приблизительно 7,1, приблизительно 7,2, приблизительно 7,3, приблизительно 7,4, приблизительно 7,5, приблизительно 7,6, приблизительно 7,7, приблизительно 7,8, приблизительно 7,9, приблизительно 8,0, приблизительно 8,1, приблизительно 8,2, приблизительно 8,3, приблизительно 8,4, приблизительно 8,5, приблизительно 8,6, приблизительно 8,7, приблизительно 8,8, приблизительно 8,9, приблизительно 9,0, приблизительно 9,1, приблизительно 9,2, приблизительно 9,3, приблизительно 9,4, приблизительно 9,5, приблизительно 9,6, приблизительно 9,7, приблизительно 9,8, приблизительно 9,9, приблизительно 10,0, или более микрограмм/кг/масса тела.
Состав фармацевтически приемлемых растворов эксципиентов и носителей хорошо известен специалистам в данной области, так же как разработка пригодных схем дозирования и введения для применения конкретных композиций, описанных в настоящем описании, в виде множества схем введения, в том числе, например, пероральное, парентеральное, внутривенное, интраназальное и внутримышечное введение и состав для такого введения.
A. АЛИМЕНТАРНАЯ ДОСТАВКА
Термин "алиментарная доставка" относится к введению, прямому или иному введению, в отдел пищеварительного тракта животного. Термин "пищеварительный тракт" относится к трубчатому каналу у животного, который принимает участие в переваривании и всасывании пищи и в удалении остатков пищи, которая проходит от ротовой полости до анального отверстия, и к любым и всем его отделам и сегментам, например к полости рта, пищеводу, желудку, тонкому и толстому кишечнику, а также к их составным частям, таким как, например, желудочно-кишечный тракт. Таким образом, термин "алиментарная доставка" включает в себя несколько способов введения, включая, но не ограничиваясь ими, пероральное, ректальное, эндоскопическое и сублингвальное/буккальное введение. Общим требованием для этих способов введения является всасывание на протяжении некоторой части или всего пищеварительного канала и необходимость в эффективном проникновении через слизистые оболочки нуклеиновой кислоты(кислот), введенной таким образом.
1. Пероральная доставка
В определенных способах применения фармацевтические композиции, описанные в настоящей заявке, можно доставлять посредством перорального введения животному, пациенту или субъекту. По существу, эти композиции можно составлять с инертным разбавителем или с ассимилируемым пищевым носителем, или их можно заключать в желатиновую капсулу с твердой или мягкой оболочкой, или их можно прессовать в таблетки, или их можно вводить непосредственно с едой при питании.
Аналогично, активные компоненты можно вводить с эксципиентами и использовать в форме проглатываемых таблеток, буккальных таблеток, лепешек, капсул, эликсиров, суспензий, сиропов, вафель и т.п. (Mathiowitz et al, 1997; Hwang et al, 1998; патенты США 5641515; 5580579 и 5792451, все из которых прямо включены в настоящее описание в качестве ссылки в полном объеме). Таблетки, лепешки, пилюли, капсулы и т.п. также могут содержать следующие компоненты: связующее вещество, такое как трагакантовая камедь, гуммиарабик, кукурузный крахмал или желатин; эксципиенты, такие как фосфат дикальция; дезинтегрирующее вещество, такое как кукурузный крахмал, картофельный крахмал, альгиновая кислота и т.п.; смазывающее вещество, такое как стеарат магния; и можно добавлять подсластитель, такой как сахароза, лактоза или сахарин, или вкусовую добавку, такую как перечная мята, масло гаультерии или добавку с вишневым вкусом. В случае, когда единичная дозированная форме представляет собой капсулу, она может содержать, в дополнение к веществам указанного выше типа, жидкий носитель. Различные другие вещества могут быть представлены в качестве покрытия или для иной модификации физической формы единицы дозирования. Например, таблетки, пилюли или капсулы можно покрывать шеллаком, сахаром или и тем, и другим. Сироп эликсира может содержать активный компонент, сахарозу в качестве подсластителя, метил и пропилпарабены в качестве консервантов, краситель и вкусовую добавка, такую как добавка со вкусом вишни или апельсина. Безусловно, любое вещество, используемое для получения любой единичной дозированной формы, должно быть фармацевтически чистым и по существу не токсичным в используемых количествах. Кроме того, активные соединения могут входить в состав препарата и составов с замедленным высвобождением.
Как правило, эти составы могут содержать по меньшей мере приблизительно 0,1% активного соединения или более, хотя процентное содержание активного ингредиента(ов), безусловно, может варьировать и может быть пригодным между приблизительно 1 или 2% и приблизительно 60% или 70% или более от общей массы или общего объема всего состава. Безусловно, активное соединение(ия) в каждой терапевтически пригодной композиции можно получать в таком количестве, чтобы достигать пригодной дозировки в любой данной единичной дозе соединения. Факторы, такие как растворимость, биодоступность, биологический период полураспада, способ введения, срок хранения продукта, а также другие фармакологические факторы, будут учитываться специалистом в области изготовления таких фармацевтических составов, и, по существу, могут быть желательными различные дозировки и схемы лечения.
2. Ректальное введение
Лекарственные средства, вводимые посредством перорального способа, часто можно вводить альтернативно посредством введения в нижний отдел кишечника, т.е. через анальное отверстие в прямую кишку или нижний отдел тонкого кишечника. Для этой цели можно использовать ректальные суппозитории, удерживающие клизмы или ректальные катетеры, и они могут быть предпочтительными, когда иным образом трудно достичь комплаентности пациента (например, при педиатрическом и гериатрическом применении, или если пациент страдает рвотой или находится без сознания). Ректальное введение может приводить к более быстрым и высоким уровням в крови, чем пероральный способ, однако также возможно обратное (Harvey, 1990). Вследствие того, что приблизительно 50% лекарственного средства, которое всасывается из прямой кишки, обходит печень, введение этим способом значительно снижает возможность метаболизма при первом прохождении (Benet et al, 1996).
B. Парентеральная доставка
Термин "парентеральная доставка" относится к введению лекарственного средства по этому изобретению животному, пациенту или субъекту способом, отличным от введения через пищеварительный тракт. Способы получения и введения парентеральных фармацевтических композиций известны в данной области (см., например, Avis, 1990).
C. Внутрипросветное введение
Внутрипросветное введение для прямой доставки терапевтического средства в отдельную часть трубчатого органа или ткани (например, такого как артерия, вена, мочеточник или мочеиспускательный канал) может быть желательным для лечения пациентов с заболеваниями или состояниями, поражающими просвет таких органов или тканей. Для осуществления этого способа введения посредством пригодных способов хирургически вводят катетер или канюлю. После изоляции участка трубчатого органа или ткани для введения проводят инфузию композиции, содержащей терапевтическое средство по этому изобретению через канюлю или катетер в изолированный сегмент. После инкубации в течение от приблизительно 1 до приблизительно 120 минут, в процессе которой терапевтическое средство захватывается клетками или контактирует с клетками внутреннего просвета сосуда, канюли или катетер для инфузии удаляют и поток в трубчатом органе или ткани восстанавливают посредством удаления лигатур, которые обеспечивают изоляцию его сегмента (Morishita et al, 1993). Терапевтические композиции по этому изобретению также можно объединять с биологически совместимым матриксом, таким как вещество на основе гидрогеля, и вводить непосредственно в сосудистую ткань in vivo.
D. Внутрижелудочковое введение
Внутрижелудочковое введение для прямой доставки лекарственного средства в головной мозг пациента может быть желательным для лечения пациентов с заболеваниями или состояниями, поражающими головной мозг. Одним способом для осуществления этого способа введения является введение хирургическим способом силиконового катетера в желудочек головного мозга пациента-человека и присоединение к подкожному инфузионному насосу (Medtronic Inc., Minneapolis, Minn.), который хирургическим способом имплантирован в абдоминальную область (Zimm et al, 1984; Shaw, 1993). Насос используют для инъецирования лекарственного средства и обеспечения точного регулирования и изменения схемы дозировки с помощью наружного программирующего устройства. Вместимость насоса составляет 18-20 мл и скорость инфузии может варьировать от 0,1 мл/ч до 1 мл/ч. В зависимости от частоты введения, варьирующей от ежедневного до ежемесячного введения, и дозы лекарственного средства, предназначенного для введения, варьирующей от 0,01 мкг до 100 г на кг массы тела, емкость насоса можно повторно заполнять с интервалами 3-10 недель. Повторное наполнение насоса можно проводить посредством подкожного прокола самогерметизирующейся перегородки насоса.
E. Внутриоболочечное введение лекарственного средства
Внутриоболочечное введение лекарственного средства для введения лекарственного средства в позвоночный столб пациента может быть желательным для лечения пациентов с заболеваниями центральной нервной системы. Для достижения этого способа введения силиконовый катетер можно хирургическим способом имплантировать в L3-4 поясничного пространства спинного мозга пациента и его соединяют с подкожным инфузионным насосом, хирургическим способом имплантированным в верхний абдоминальный отдел (Luer and Hatton, 1993; Ettinger et al, 1978; Yaida et al, 1995). Насос используют для инъецирования лекарственного средства и обеспечения точного регулирования и изменений дозировки в схеме дозирования с помощью наружного программирующего устройства. Вместимость насоса составляет 18-20 мл и скорости инфузии могут варьировать от 0,1 мл/ч до 1 мл/ч. В зависимости от частоты введения лекарственного средства, варьирующей от ежедневного до ежемесячного введения, и дозы лекарственного средства, предназначенного для введения, варьирующей от 0,01 мкг до 100 г на 1 кг массы тела, емкость насоса можно повторно заполнять с интервалами 3-10 недель. Повторное наполнение насоса можно проводить посредством однократного подкожного прокола самогерметизирующейся перегородки насоса.
Для достижения доставки в области, отличные от головного мозга и позвоночного столба посредством этого способа, изменяют форму силиконового катетера для присоединения подкожного инфузионного насоса, например, с печеночной артерией для доставки в печень (Kemeny et al, 1993).
F. Вагинальная доставка
Вагинальная доставка предусматривает локальное введение и позволяет избежать метаболизм при первом прохождении, деградации пищеварительными ферментами и потенциальных системных побочных эффектов. Вагинальные суппозитории (Block, Chapter 87 В: Remington's Pharmaceuticals Sciences, 18th Ed., Gennaro, ed., Mack Publishing Co., Easton, Pa., 1990, стр. 1609-1614) или мази для местного применения используют для осуществления этого способа доставки.
G. Опосредуемая липосомами, нанокапсулами и микрочастицами доставка
В определенных вариантах осуществления авторы настоящего изобретения рассматривают применение липосом, нанокапсул, микрочастиц, микросфер, липидных частиц, везикул и т.п. для введения композиции по настоящему изобретению в пригодные клетки-хозяева. В частности, композиции по настоящему изобретению можно составлять для доставки либо инкапсулированными в липидную частицу, липосому, везикулу, наносферу, либо в качестве наночастицы и.т.п.
Такие составы могут быть предпочтительными для введения фармацевтически приемлемых составов нуклеиновых кислот или конструкций, описанных в настоящем описании. Получение и применение липосом, главным образом, известно специалистам в данной области (см. например, Couvreur et al, 1977; Lasic, 1998; которые описывают применение липосом и нанокапсул в направленной терапии антибиотиками против внутриклеточных бактериальных инфекций и заболеваний). Недавно разработали липосомы с повышенной стабильностью и временем полураспада в кровотоке (Gabizon and Papahadjopoulos, 1988; Allen and Choun, 1987; патент США 5741516, включенный прямо в настоящее описании в качестве ссылки в полном объеме). Кроме того, рассмотрены различные способы получения препаратов липосом и подобных липосомам препаратов в качестве потенциальных носителей лекарственного средства (Takakura, 1998; Chandran et al., 1997; Margalit, 1995; патент США 5567434; 5552157; 5565213; 5738868 и 5795587, все из них прямо включены в настоящее описание в качестве ссылок в полном объеме).
Липосомы образуются из фосфолипидов, которые диспергированы в водной среде и спонтанно формируют многослойные концентрические везикулы из двух слоев (также называемые многослойными везикулами (MLV). MLV, как правило, обладают диаметрами от 25 нм до 4 мкм. Обработка MLV ультразвуком приводит к образованию однослойных везикул малых размеров (SUV), содержащих в центре водный раствор, с диаметрами, находящимися в диапазоне от 200 до 500 Ǻ.
Путь и расположение инъецированных внутривенно липосом зависит от их физических свойств, таких как размер, текучесть и поверхностный заряд. Они могут существовать в тканях в течение часов или суток, в зависимости от их состава, и периоды их полураспада в крови варьируют от минут до нескольких часов. Более крупные липосомы, такие как MLV и LUV, быстро захватываются фагоцитирующими клетками ретикулоэндотелиальной системы, однако физиология системы кровообращения ограничивает выход таких крупных частиц в большинстве областей. Они могут выходить только в участках, где в капиллярном эндотелии существуют крупные отверстия или поры, таких как синусоиды печени или селезенки. Таким образом, эти органы являются основной областью захвата. С другой стороны, для SUV показано более широкое распределение в тканях, однако тем не менее они в высокой степени депонируются в печени и селезенке. Главным образом, этот характер поведения in vivo ограничивает потенциальное нацеливание липосом только теми органами и тканями, которые доступны для их крупного размера. Они включают в себя кровь, печень, селезенку, костный мозг и лимфоидные органы.
Альтернативно это изобретение относится к фармацевтически приемлемым составам композиций по настоящему изобретению для нанокапсул. Нанокапсулы могут, главным образом, захватывать соединения стабильным и воспроизводимым способом (Henry-Michelland et al., 1987; Quintanar-Guerrero et al., 1998; Douglas et al., 1987). Для избежания побочных эффектов вследствие внутриклеточной перегрузки полимерами, такие мельчайшие частицы (размером приблизительно 0,1 мкм) необходимо создавать с использованием полимеров, подлежащих деградации in vivo. Биологически деградируемые наночастицы полиалкилцианоакрилата, которые удовлетворяют этим требованиям, предусмотрены для применения по настоящему изобретению. Такие частицы можно легко получать, как описано (Couvreur et al., 1980; 1988; zur Muhlen et al., 1998; Zambaux et al. 1998; Pinto-Alphandry et al., 1995 и U.S. Pat. No. 5145684, включенные прямо в настоящее описание в качестве ссылок в полном объеме).
ПРИМЕРЫ
Следующие примеры приведены в целях иллюстрации различных вариантов осуществления этого изобретения и не предназначены для ограничения каким-либо образом настоящего изобретения. Специалист в данной области легко поймет, что настоящее изобретение легко адаптируется для осуществления целей и для получения результатов и упомянутых преимуществ, а также целей, результатов и преимуществ, связанных с ними. Данные примеры, наряду с клетками и способами, описанными в настоящем описании, в настоящее время являются репрезентативными для предпочтительных вариантов осуществления, они являются иллюстративными и не предназначены для ограничения объема этого изобретения. Их изменения и другие способы применения, которые охватываются сущностью этого изобретению, как определено объемом формулы изобретения, будут понятны специалистам в данной области.
A. Материалы и способы
Выделение кДНК растворимого CRFR2α мыши. Вариант по сплайсингу растворимого CRFR2a выделяли параллельно с вариантом ортолога CRFR2α мыши. Праймеры для ПЦР были разработаны исходя из гомологии между известными генами CRFR2 млекопитающих. Для скрининга ДНК, полученной из поли(A)+ РНК целого мозга мыши, которую подвергали обратной транскрипции с применением олигонуклеотидов dT или случайных праймеров, использовали следующие олигонуклеотидные праймеры, 5' CCCCGAAGCTGCCCGACTGG 3' (SEQ ID NO: 16) (смысловой) и 5' GGAAGGCTGTAAAGGATGGAGAAG 3' (SEQ ID NO: 17) (антисмысловой). ПЦР проводили при 62°C в течение 35 циклов с удлинением в течение 90 с при 72°C. Амплифицированные фрагменты субклонировали в вектор pCRIITOPO (Invitrogen, Carlsbad, CA), секвенировали и выявляли те, которые кодируют полноразмерный новый вариант CRFR2α по сплайсингу, лишенный шестого экзона, sCRFR2α (Chen et al., 2005).
Полуколичественная RT-PCR и саузерн-анализ. Следующие периферические ткани и ткани CNS мыши рассекали и непосредственно подвергали общему выделению РНК, как описано ранее (Chen et al, 2005): весь головной мозг, обонятельную луковицу, гипоталамус, кору, мозжечок, гиппокамп, средний мозг, мост/продолговатый мозг, спинной мозг и гипофиз. Продукты кДНК использовали в качестве матриц для полуколичественного анализа RT-PCR с использованием специфичных праймеров для CRFR2α, sCRFR2α и рибосомального белка S16. Расположение олигонуклеотидных праймеров в экзонах три и семь обеспечивает амплификацию двух продуктов длиной 418 и 309 п.о., соответствующих CRFR2α и sCRFR2α соответственно. Последовательность олигонуклеотидных праймеров и условия ПЦР можно найти в дополнительном тексте.
Анализ киназы внеклеточного рецептора 1/2 (ERK1/2). Клетки CATH.a уравновешивали DMEM, дополненной 1% (мас./об.) бычьим сывороточным альбумином (BSA) в течение 6 часов, а затем стимулировали посредством 0,1% DMEM/BSA (носитель) или 10 нМ Ucn I при наличии или в отсутствии 0,4 или 4 нМ sCRFR2α, разбавленного 0,1% DMEM/BSA. Клетки сразу собирали и анализировали на фосфорилированную ERK1/2-p42, 44, как описано ранее (Chen et al, 2005).
Временные трансфекции и анализ с люциферазой. Клетки HEK293T трансфицировали люциферазным репортером, содержащим фрагмент гена EVX1, содержащего активный участок CRE. Клетки собирали и активность люциферазного репортера оценивали, как описано ранее (Chen et al, 2005). Через двадцать часов после трансфекции, клетки в течение 4 ч обрабатывали носителем или Ucn 1 (0,0001-100 нМ) в присутствии или отсутствии 0,1 нМ sCRFR2α.
Радиоиммунные анализы (RIA). Антисыворотку индуцировали у кроликов, иммунизированных синтетическим пептидным фрагментом, кодирующим уникальный C-конец (а.к. 113-143) sCRFR2α мыши, конъюгированного с гемоцианином лимфы улитки с использованием протокола, описанного ранее для субчастиц ингибина (Vaughan et al, 1989). Аналог Tyr113 sCRFR2α (113-143) метили радиоактивной меткой посредством Na125I с использованием хлорамина-T и очищали посредством ВЭЖХ (Vaughan et al, 1989) для применения в качестве метки в HA. Способ для RIA sCRFR2α был сходным с подробно описанным ранее способом для субчастиц ингибина (Vaughan et al, 2005). В кратком изложении антитела против sCRFR2α использовали в конечном разведении 1/300000 и синтетический sCRFR2α (113-143) использовали в качестве стандарта. Ткани мыши экстрагировали кислотой и частично очищали с использованием контейнера с октадецилсиликоном, как описано (Vaughan et al, 1989). Лиофилизированные образцы тестировали при от трех до семи уровнях дозы. Свободную метку отделяли от связанной посредством добавления γ-глобулинов овцы против кролика и 10% (мас./об.) полиэтиленгликоля. EC50 и минимальная определяемая доза для sCRFR2α составляли ~5 пг и 100 пг на пробирку соответственно.
Иммуногистохимия. Взрослым самцам мышей C57B6J (Jackson Laboratories) и белых крыс Sprague-Dawley (Harlan Sprague-Dawley) проводили анестезию посредством хлоралгидрата (350 мг/кг, внутрибрюшинно) и перфузию посредством фиксажа Замбони (Bittencourt et al, 1999), с последующей постфикасцией в течение 0-4 ч. Получали расположенные с равными интервалами (1-к-4) серии фронтальных срезов толщиной 30 мкм на всем протяжении головного мозга для усиленной никелем авидин-биотин-иммунопероксидазной локализации sCRFR2α-ir с использованием реагентов Vectastain Elite (Vector Laboratories, Burlingame, CA). Первичную антисыворотку против sCRFR2α адсорбировали относительно носителя, очищали аффинным способом и использовали в разведении 1:2000. Специфичность иммунного окрашивания оценивали с использованием первичной антисыворотки, предварительно инкубированной в течение ночи при 4°C с 0-300 мкМ синтетическим иммуногеном. Мечение также оценивали у мутантных мышей, дефектных по любому из CRFR (Smith et al, 1998; Bale et al 2000). Подробное описание флуоресцентного иммуногистохимического анализа клеток COSM6, трансфицированных sCRFR2α, можно найти в дополнительном тексте.
Экспрессия sCRFR2α в млекопитающих: кДНК, соответствующую аминокислотам 1-143, модифицированную посредством ПЦР для включения эпитопа FLAG после аминокислоты 143, субклонировали в pSec-Tag2 HygroA (Invitrogen, Carlsbad, CA) и использовали для трансфекции клеток COSM6, как описано (Perrin et al, 2001). Через 4 суток среду собирали и содержание sCRFR2α увеличивали посредством очистки с использованием иммунноаффинной хроматографии с FLAG-агарозой (Sigma, St. Louis, MO). Белок выявляли посредством иммуноблот-анализа с использованием либо антитела против FLAG, либо антитела, полученного против уникального C-конца sCRFR2α.
Бактериальная экспрессия sCRFR2α: кДНК, соответствующую аминокислотам 20-143, получали посредством ПЦР с использованием mCRFR2α в качестве матрицы. кДНК субклонировали в pET-32a(+) (Novagen, La Jolla, CA) и белок очищали посредством аффинной хроматографии с S-белком, как описано (Perrin et al, 2001). Белок выявляли посредством иммуноблот-анализа с использованием антитела, полученного против уникального C-конца sCRFR2α.
Радиорецепторные анализы. Растворимый белок, очищенный либо из клеточной среды COSM6, либо E.coli, инкубировали в трех экземплярах в лунках с [125I-DTyrO]-астрессином и с немечеными пептидами в возрастающих концентрациях (Perrin et al, 2003).
B. Результаты
В процессе выделения CRFR2α мыши наблюдали транскрипт кДНК меньшего размера (-100 п.о.) (Van Pett et al, 2000). Этот фрагмент меньших размеров выделили и было обнаружено, что он кодирует вариант CRFR2α, обладающий делецией шестого экзона. Трансляция транскрипта варианта предопределила новый белок из 143 аминокислот, sCRFR2α, содержащий большую часть первого внеклеточного домена CRFR2α с последующим уникальным C-концом из 38 аминокислот (фиг.1A). Скрининг в GenBank показал отсутствие гомологии C-конца с другими белками. Геномное расположение sCRFR2α представлено на фиг.1B.
Если мРНК sCRFR2α является только продуктом ошибки сплайсинга, она должна быть более редко встречающейся, чем правильно сплайсированная РНК. В целях исследования этой проблемы для сравнения относительной распространенности мРНК CRFR2α и sCRFR2α в некоторых участках головного мозга использовали полуколичественную RT-PCR с последующим анализом саузерн-гибридизации. Проводили обратную транскрипцию общей РНК, полученной из тканей, с получением кДНК, которую использовали в качестве матриц для полуколичественного анализа RT-PCR, с последующей саузерн-гибридизацией с использованием специфичных праймеров и зондов для CRFR2α и sCRFR2α (фиг.2). Пара олигонуклеотидных праймеров (расположенная в третьем и седьмом экзонах) обеспечивала одновременную амплификацию как растворимой формы, так и полноразмерного мембраносвязанного рецептора в одной реакции (фиг.2A). sCRFR2α высоко экспрессировался в обонятельной луковице, коре, среднем мозге и гипофизе (фиг.2B и 2C). Более низкие уровни экспрессии обнаружили в гиппокампе, гипоталамусе, мосте, продолговатом мозге и спинном мозге (фиг.2B и 2C). Как представлено на фиг.2, распространенность мРНК sCRFR2α является более низкой, однако сравнимой с распространенностью мРНК CRFR2α. Было обнаружено, что последовательности фрагментов кДНК при RT-PCR кодируют сплайсированный вариант гена CRFR2α мыши (фиг.1A).
С помощью компьютерного анализа последовательности было предварительно определено, что первые 19 аминокислот служат в качестве предполагаемого сигнального пептида. Вследствие того, что последовательность не содержит явных участков для прикрепления к мембране, было предположено, что белок секретируется в качестве растворимой формы. Для исследования этого предположения белок экспрессировали в клетках COSM6. После очистки от среды полосу белка ~30 кДа визуализировали посредством иммуноблот-анализов с использованием либо антисыворотки против FLAG, либо анти-sCRFR2α, антисыворотки, индуцированной против синтетического пептидного фрагмента, кодирующего уникальный C-конец белка sCRFR2α (а.к. 113-143) (фиг.3A). Более крупный размер белка по сравнению с предварительно определенным размером исходя из кДНК, вероятно, является результатом гликозилирования. В целях получения большего количества sCRFR2α, белок, лишенный предполагаемого сигнального пептида, экспрессировали в качестве слитого белка в E. coli (Perrin et al, 2001). После расщепления и очистки белок визуализировали (с использованием анти-sCRFR2α) посредством имммуноблот-анализа в качестве узкой полосы, соответствующей размеру ~20 кДа. Антисыворотка против sCRFR2α позволяет выявить белки sCRFR2α как в радиоиммунном анализе (фиг.3B), так и посредством иммуноцитохимии (фиг.3C).
Иммуногистохимические исследования с использованием антисыворотки против sCRFR2α позволили выявить распределение sCRFR2α-ir в головном мозге грызунов. Клеточное распределение при иммунном мечении для sCRFR2α-ir было широким и более соответствовало расположению области экспрессии мРНК CRFR1, чем расположению CRFR2 (фиг.4A-4F). Описанные результаты представляют собой результаты исследований на мышах; у крыс наблюдался сходный характер мечения. Основные области клеточной экспрессии включают в себя митральные и пучковые клетки обонятельной луковицы, комплекс септо-диагональной полосы, пириформную кору, черную субстанцию, красное ядро, базолатеральное миндалевидное тело, глубокие ядра мозжечка и дорсального столба, все из которых являются областями значительной экспрессии CRFR1. Аналогично CRFR1, клеточные тельца sCRFR2α-ir являются многочисленными на протяжении изокортекса, хотя их распределение по слоям перекрывается только частично. Таким образом, несмотря на то, что экспрессирующие как CRFR1, так и sCRFR2α, клеточные тельца являются многочисленным в слое 2/3, основной кортикальной областью экспрессии CRFR1 является слой 4, в то время как областью экспрессии sCRFR2α является слой 5. Все основные области экспрессии CRFR2, включая боковые ядра перегородки, ядра среднего мозга, вентромедиальные ядра гипоталамуса и медиальные ядра миндалевидного тела, были лишены окрашенных на sCRFR2α клеточных телец, хотя интересно, что последние две области относились к незначительно снабженным мечеными утолщениями, что авторы изобретения отнесли к отражению терминальных областей sCRFR2α-ir. Паравентрикулярное ядро гипоталамуса также содержало предполагаемую терминальную область sCRFR2α-ir с умеренной плотностью.
Мечение во всем головном мозге блокировали посредством предварительной инкубации антисыворотки с низкими микромолярными концентрациями (≥30 мкМ) пептида sCRFR2α (113-143), используемого в качестве иммуногена; конкуренция с соответствующим пептидом, предварительно определенным исходя из последовательности CRFR1, не препятствовала иммунному мечению в таких высоких концентрациях, как 3 мМ. Следующим подтверждением специфичности мечения являются результаты наблюдений, состоящие в том, что все типы иммунной локализации сохранялись у мышей с делецией CRFR1 и/или CRFR2; следует отметить, что ожидалось, что конструкция для нацеливания, используемая для получения каждой из существующих линий с нокаутом рецептора, сохраняет кодирующий участок sCRFR2α (Smith et al, 1998; Timpl et at, 1998; Bale et al, 2000).
В целях определения наличия подобного sCRFR2α ir в головном мозге проводили высокоспецифичный радиоиммунный анализ с использованием анти-sCRFR2α и [125I-Tyr113] sCRFR2α (113-143)] в качестве метки. Ткань из головного мозга мыши подвергали экстракции кислотой, частично очищали на фильтрующих элементах C18 и оценивали во множестве доз в радиоиммунном анализе. Экстракты тканей вытесняли [125I-Tyr113] sCRFR2α (113-143)], связанный с анти-sCRFR2α, зависимым от дозы образом (фиг.4G). Наиболее высокие уровни экспрессии были выявлены в обонятельной луковице, гипоталамусе, коре и среднем мозге, все из которых коррелируют с наличием клеток ir и волокон, определенных в иммуногистохимических исследованиях (фиг.4). Предполагаемая растворимая форма CRFR1 (полученная посредством делеции экзона 5) должна содержать отличающуюся уникальную C-концевую последовательность. Белок, соответствующий этой последовательности, не вытеснял [125I-Tyr113] sCRFR2α (113-143) в радиоиммунном анализе. Эти результаты дополнительно подтверждают существование белка sCRFR2α в ЦНС грызунов.
Взаимодействия sCRFR2α с лигандами семейства CRF оценивали посредством радиорецепторного анализа с использованием конкурентного вытеснения [125I-D TyrO]-астрессина, связанного с sCRFR2α. Растворимые белки, секретируемые клетками COSM6 или продуцируемые бактериями, связывают агонисты, Ucn 1 и CRF, а также антагонист астрессин с наномолярной аффинностью, в то время как аффинность в отношении Ucn 2 и Ucn 3 является значительно более низкой (таблица 2).
sCRFR2α
sCRFR2α
Представлено связывание членов семейства CRF с белками sCRFR2α, очищенными либо из среды клеток COSM6 (mam sCRFR2α), либо E.coli (bact sCRFR2α). Для более подробного описания см. способы.
Для определения возможных функций sCRFR2α авторы изобретения исследовали его эффекты на передачу сигнала посредством лигандов семейства CRF. Белки sCRFR2α, экспрессированные как в клетках млекопитающих, так и в бактериальных клетках, зависимым от дозы образом ингибировали ответ cAMP на Ucn 1 и CRF в клетках HEK293T, трансфицированных CRFR2α мыши, как определяли по люциферазной активности CRE гена EVX1 (фиг.5A). Вследствие того, что урокортины активируют передачу сигнала MAPK (Brar et al, 2002), авторы изобретения определили способность sCRFR2α ингибировать активацию ERK1/2-p42,44 посредством Ucn 1 в клетках CATH.a, которые эндогенно экспрессируют CRFR1 и CRFR2α. sCRFR2α ингибирует индукцию фосфорилированной ERK посредством Ucn 1 в клетках CATH.a (фиг.5B).
Несмотря на то, что настоящее изобретение и его преимущества были подробно описаны, следует понимать, что различные изменения, замены и преобразования можно проводить без отклонения от сущности и объема этого изобретения, которые определены посредством прилагаемой формулы изобретения. Более того, подразумевают, что объем настоящей заявки не ограничивается конкретными вариантами осуществления процесса, устройства, изготовления, композиций для вещества, средств, способов и стадий, описанных в описании. Как будет легко понятно среднему специалисту в данной области из описания настоящего изобретения, в соответствии с настоящим изобретением можно использовать процессы, устройства, изготовление, композиции для вещества, средства, способов или стадий, в настоящее время существующие или те, которые будут разработаны в будущем, которые выполняют по существу такую же функцию или приводят по существу к такому же результату, что и соответствующий вариант осуществления, описанный в настоящем описании. Таким образом, подразумевают, что прилагаемая формула изобретения включает в свой объем такие процессы, устройства, изготовление, композиции для вещества, средств, способов или стадий.
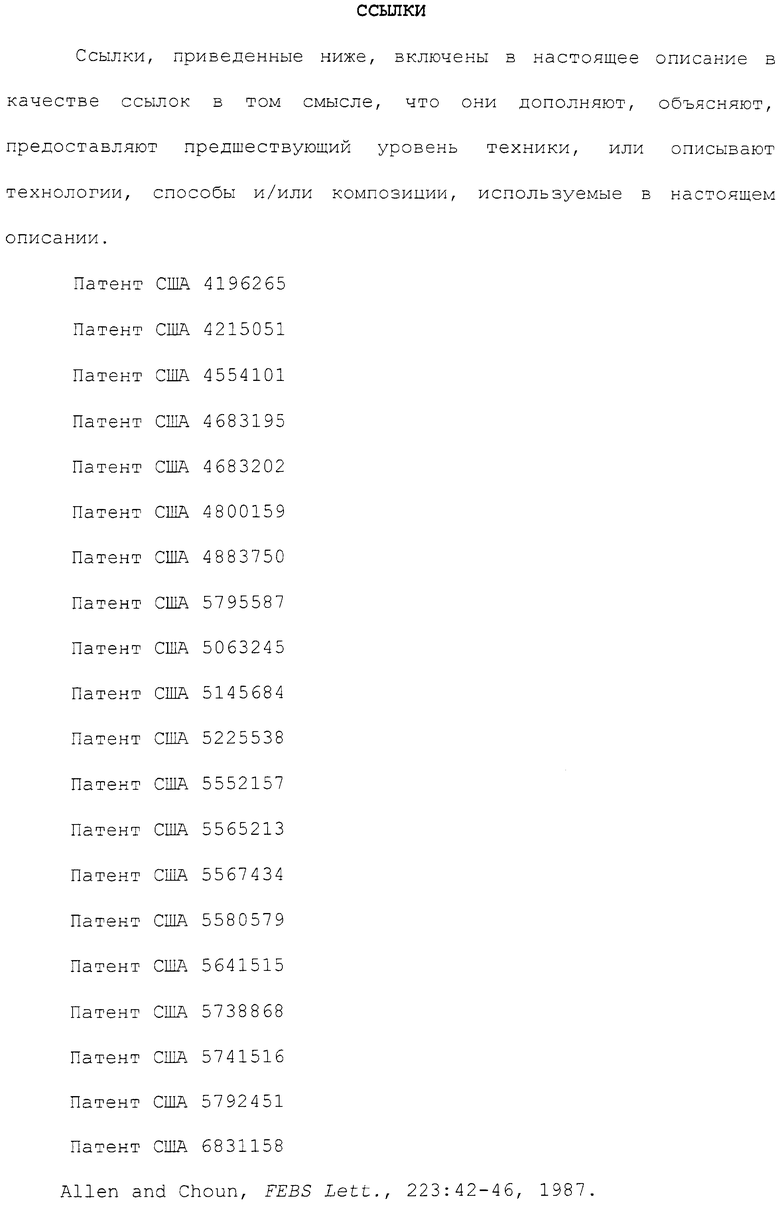








| название | год | авторы | номер документа |
|---|---|---|---|
| ДРОЖЖЕВОЙ ДИСПЛЕЙ БЕЛКОВ В ПЕРИПЛАЗМАТИЧЕСКОМ ПРОСТРАНСТВЕ | 2018 |
|
RU2802770C2 |
| ИДЕНТИФИКАЦИЯ МОЛЕКУЛ, МОДУЛИРУЮЩИХ БЕЛОК-БЕЛКОВОЕ ВЗАИМОДЕЙСТВИЕ | 2008 |
|
RU2476891C2 |
| СОПРЯЖЕННЫЕ С G-БЕЛКОМ РЕЦЕПТОРЫ DROSOPHILA, НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ, ИМЕЮЩИЕ К НИМ ОТНОШЕНИЕ | 2003 |
|
RU2326385C2 |
| СОПРЯЖЕННЫЙ С G-БЕЛКОМ РЕЦЕПТОР, ИМЕЮЩИЙ ИЗМЕНЕННОЕ СРОДСТВО К ЛИГАНДАМ, И ЕГО ПРИМЕНЕНИЕ | 2008 |
|
RU2468035C2 |
| ВЕКТОРЫ, УСЛОВНО ЭКСПРЕССИРУЮЩИЕ ТЕРАПЕВТИЧЕСКИЕ БЕЛКИ, КЛЕТКИ-ХОЗЯЕВА, СОДЕРЖАЩИЕ УКАЗАННЫЕ ВЕКТОРЫ, И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2612788C2 |
| ПОЛУЧЕНИЕ ГЛИКОПРОТЕИНОВ | 2007 |
|
RU2463345C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ ВЕЩЕСТВ | 2003 |
|
RU2325652C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2708311C2 |
| УРОКОРТИН III, ЕГО АНАЛОГ (ВАРИАНТЫ) И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2002 |
|
RU2305109C2 |
| МОДИФИЦИРОВАННЫЕ БЕЛКИ, СКОНСТРУИРОВАННЫЕ ТОКСИНЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2002 |
|
RU2305684C2 |







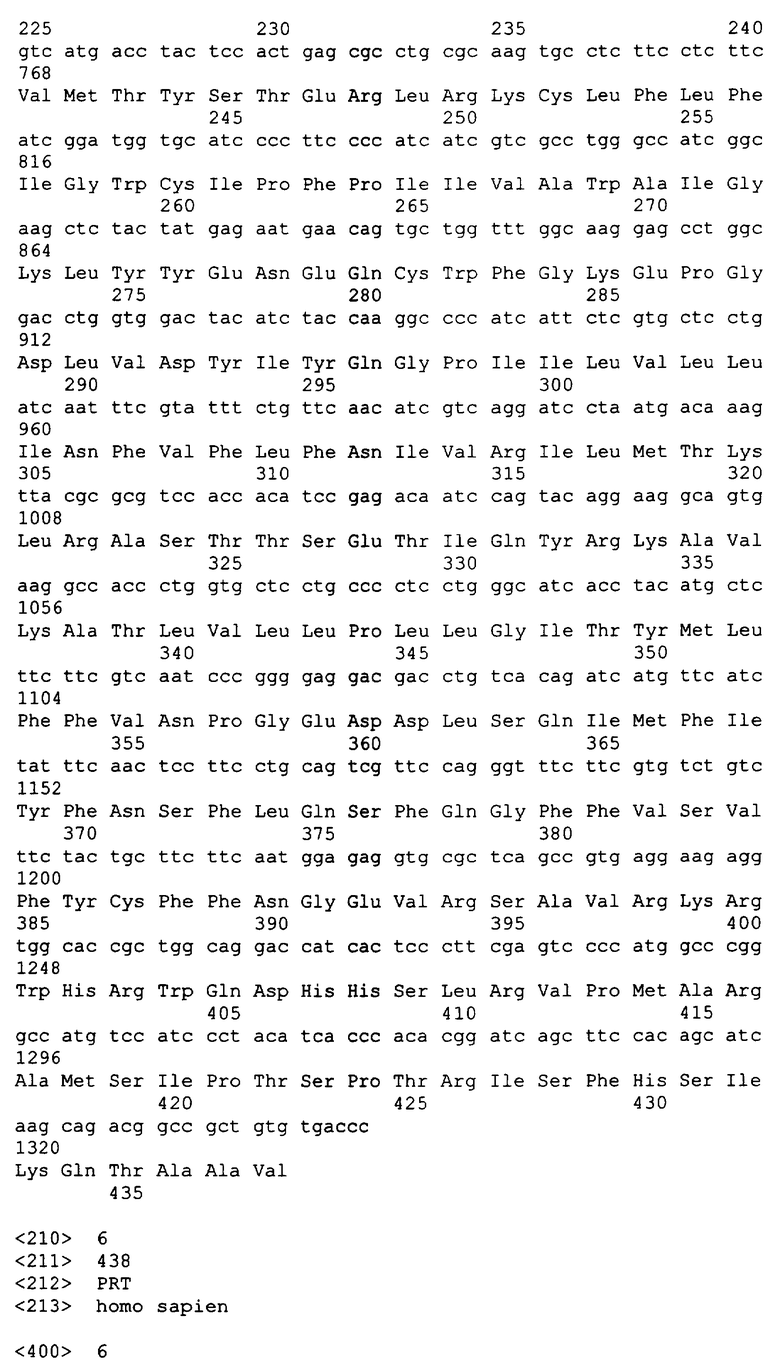



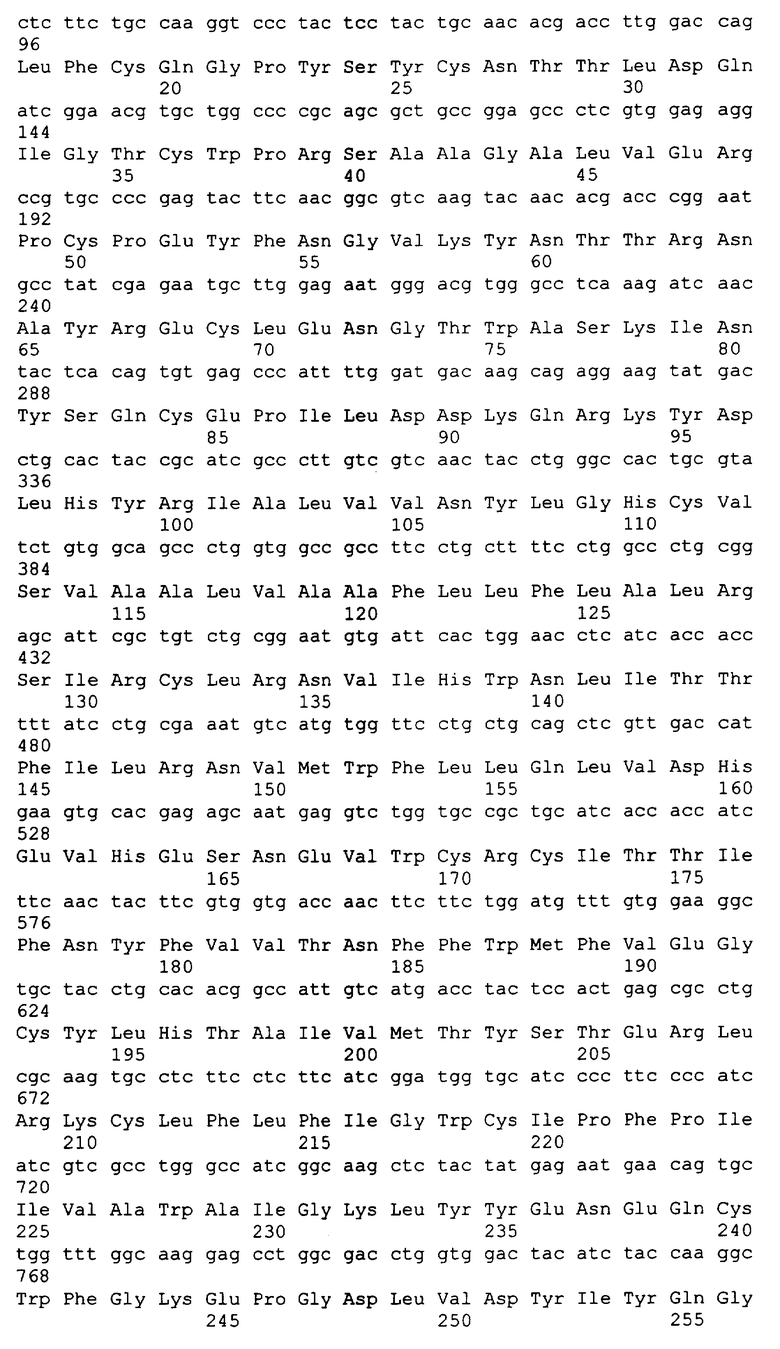























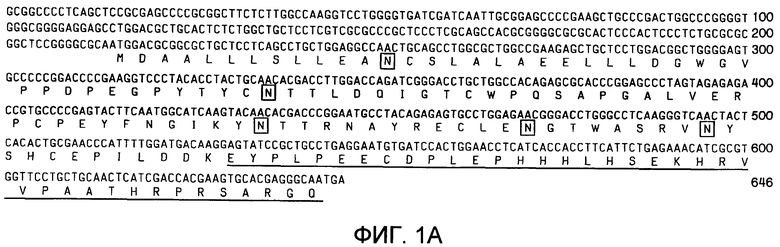
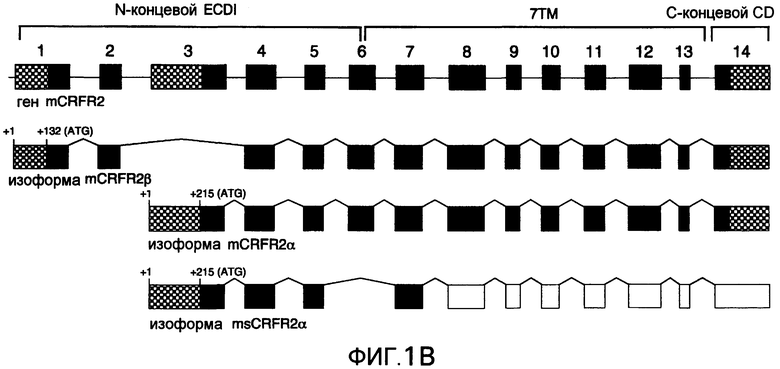

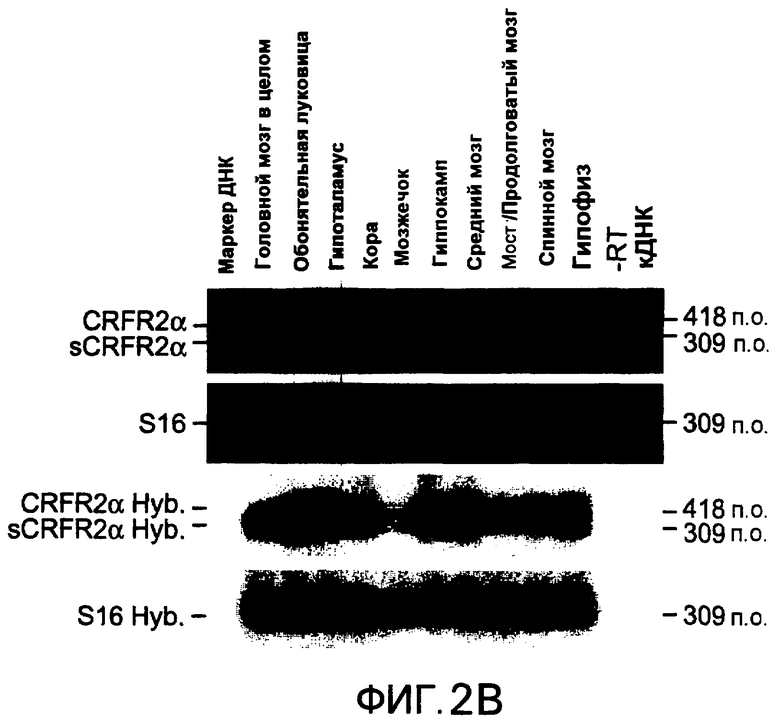
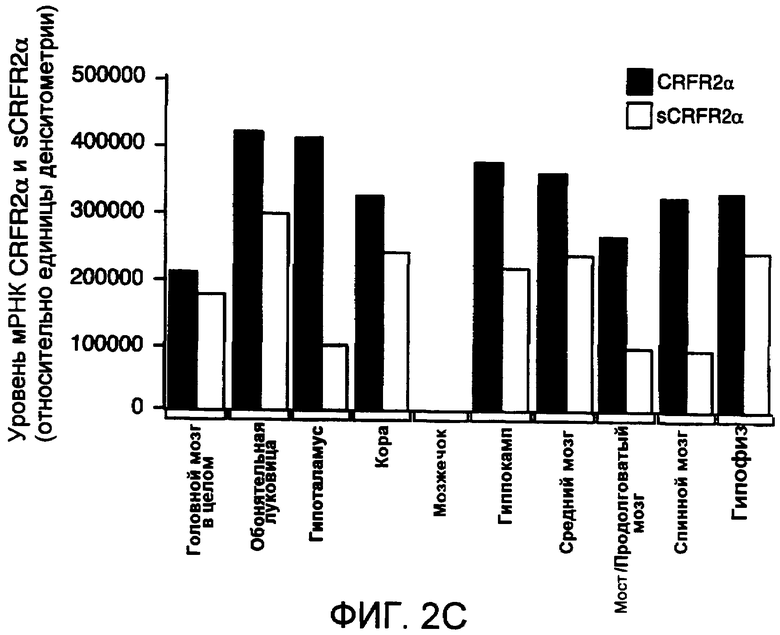
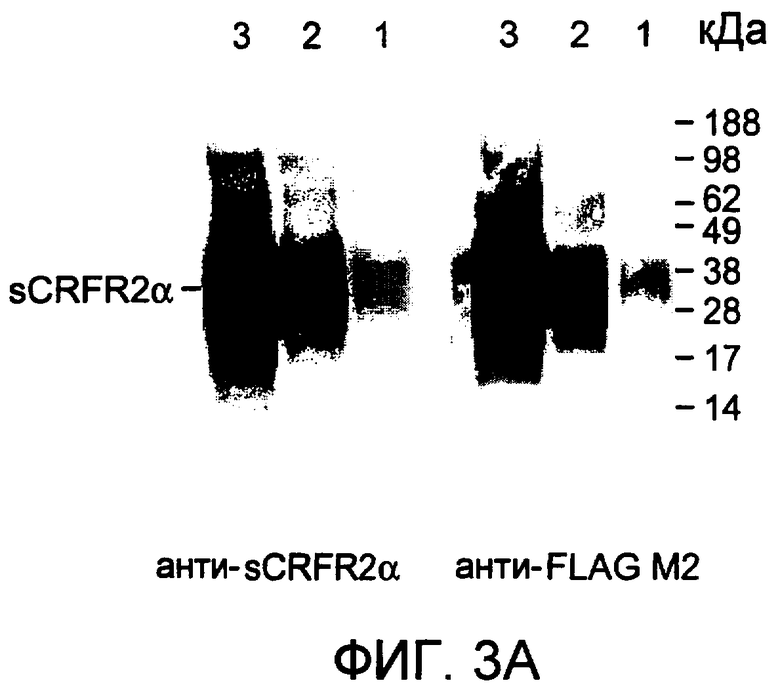
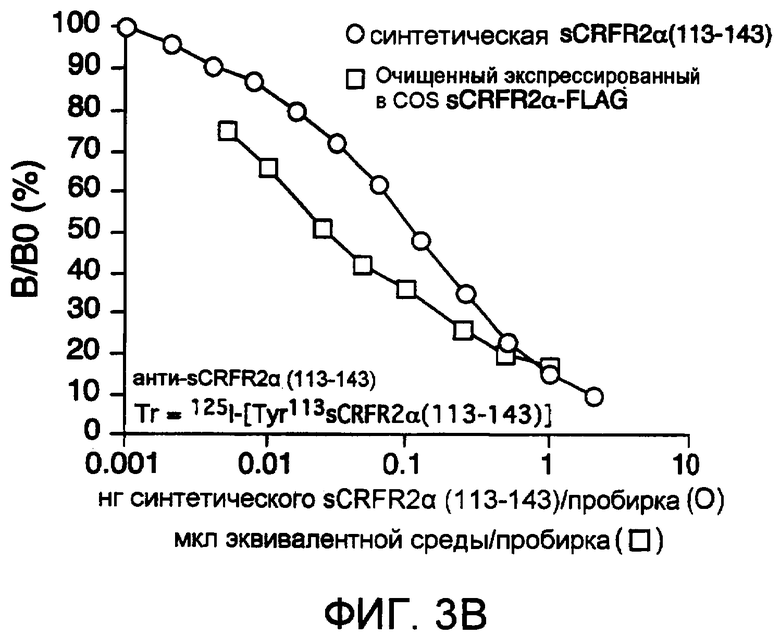
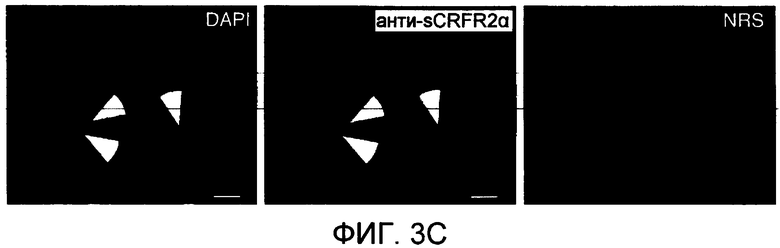






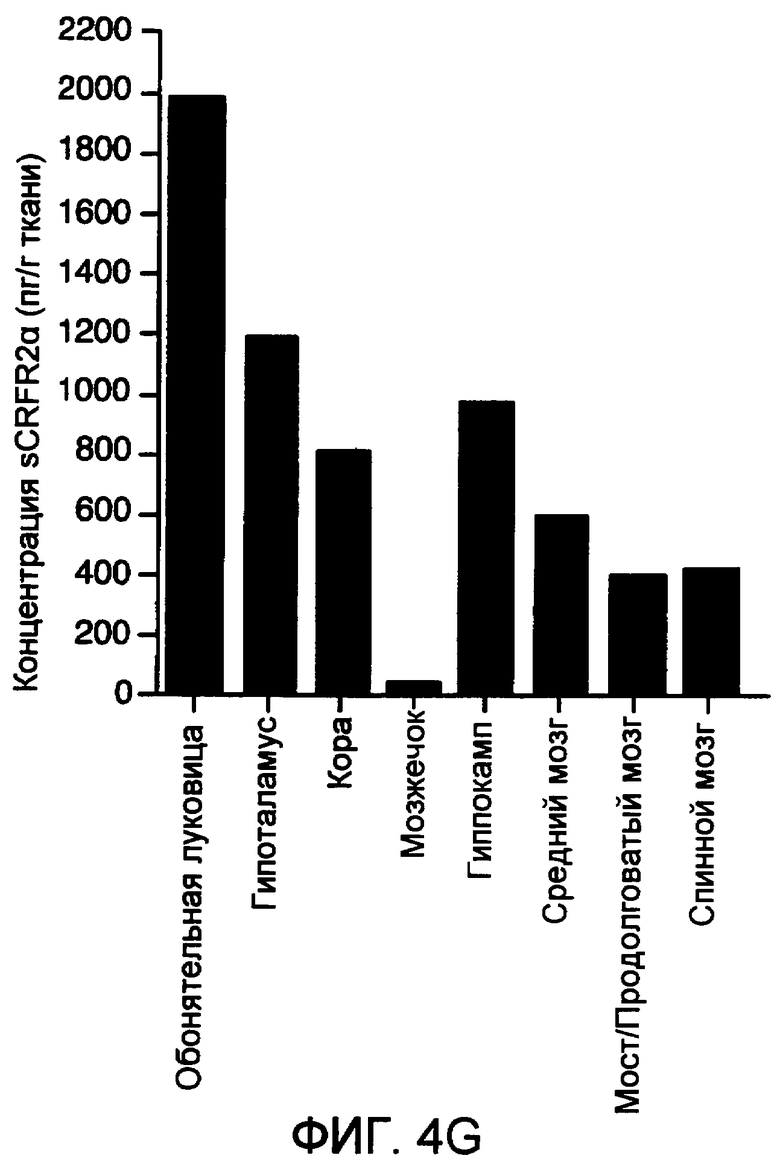
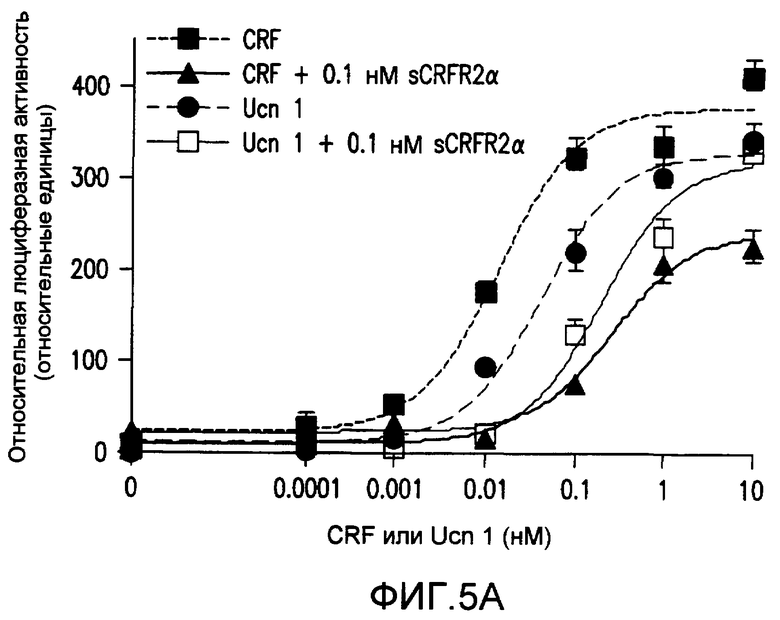
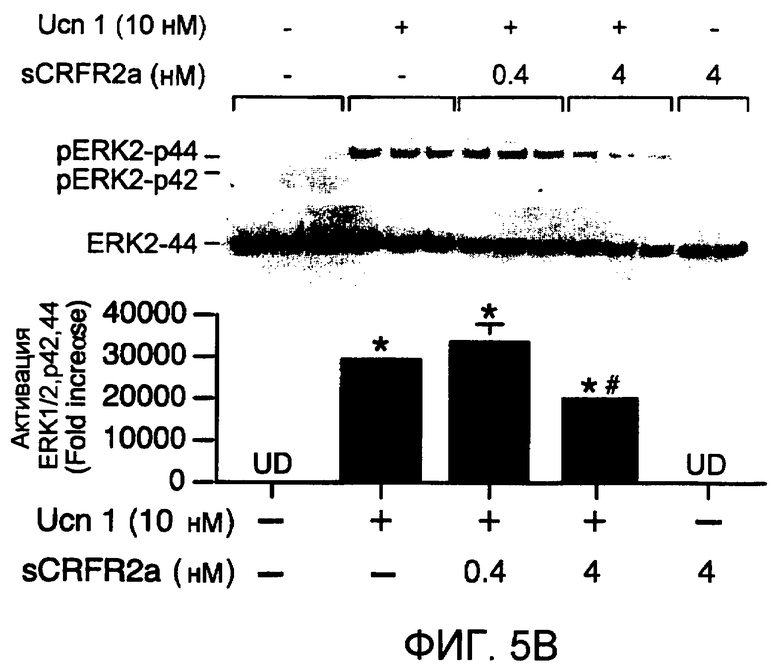
Настоящее изобретение относится к области молекулярной биологии и может быть использовано в неврологии и эндокринологии. Получен лиганд-связывающий домен растворимого рецептора для кортикотропин-рилизинг фактора 2 типа (sCRFR2), который характеризуется способностью ингибировать активность указанного рецептора в присутствии лиганда, выбранного из группы, включающей кортикотропин-рилизинг фактор, урокортин-1, урокортин-2, урокортин-3 и стресскопин. Предложено использовать полученный sCRFR2 или его конъюгат с полиэтиленгликолем в качестве модулятора активности CRFR2 и/или фармакологического эффекта связывающегося с ним лиганда. 4 н. и 2 з.п.ф-лы, 17 ил., 2 табл.
1. Связывающий лиганд домен выделенного растворимого рецептора для кортикотропин-рилизинг фактора 2 типа (sCRFR2), имеющий аминокислотную последовательность SEQ ID NO:4.
2. Конъюгат, содержащий связывающий лиганд домен sCRFR2 по п.1 и аффинный маркер, радионуклид, флуоресцентный маркер, хемилюминесцентный маркер или их сочетание.
3. Конъюгат по п.2, содержащий аффинный маркер.
4. Конъюгат по п.2, содержащий флуоресцентный маркер.
5. Конъюгат, содержащий связывающий лиганд домен sCRFR2 по п.1 и полиэтиленгликоль (PEG).
6. Способ in vitro ингибирования активности рецептора для кортикотропин-рилизинг фактора 2 типа (CRFR2), включающий контактирование клетки, экспрессирующей CRFR2, со связывающим лиганд доменом sCRFR2 по п.1 в присутствии лиганда, выбранного из группы, включающей кортикотропин-рилизинг фактор (CRF), урокортин 1, урокортин 2, урокортин 3 и стресскопин.
| PERRIN М | |||
| ЕТ AL., Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA, 1995, 92(7), 2969-2973 | |||
| PERRIN M | |||
| ET AL., J | |||
| Biol | |||
| Chem., 2003, 278, no.18, 15595-15600 | |||
| KOSTICH W | |||
| ET AL., Mol | |||
| Endocrinology, 1998, 12, 1077-1085 | |||
| RU 2002103717 A, 10.03.2004. |

