ПРЕДПОСЫЛКА ИЗОБРЕТЕНИЯ
ОПИСАНИЕ ФЕДЕРАЛЬНОГО ФИНАНСИРОВАНИЯ
Данное изобретение частично выполнено с использованием средств, полученных от Федерального правительства по гранту № DK-26741. Таким образом, Федеральное правительство имеет определенные права на данное изобретение.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение, в общем, относится к областям нейроэндокринологии и механизмам, вовлеченным в стресс. Более конкретно, данное изобретение относится к новым пептидам, родственным кортикотропин-рилизинг фактору, урокортину II и родственному урокортину белку человека, которые принимают участие в ответе на стресс.
ОПИСАНИЕ СВЯЗАННОЙ ОБЛАСТИ
Кортикотропин-рилизинг фактор (CRF) является пептидом, состоящим из 41 аминокислоты, лучше всего известным ввиду его важной роли в инициации гипофизарно-надпочечниковых ответов на стресс, эффекта, опосредуемого рецепторами CRF типа 1 (1). Кроме того, кортикотропин-рилизинг фактор широко распространен в головном мозге, и многократно было показано, что он принимает участие в мобилизации взаимодополняющих автономных и поведенческих приспособительных реакций в случае множества угрожающих условий (2, 3). Это побудило выдвинуть получившую широкое признание гипотезу о том, что кортикотропин-рилизинг фактор и семейство родственных ему пептидов играют важную роль в регуляции гипоталамо-гипофизарно-надпочечниковой оси (НРА) при основных и стрессовых условиях (4, 5). Также считается, что кортикотропин-рилизинг фактор также участвует в других нейроэндокринных и паракринных ответах во многих тканях. Члены семейства CRF осуществляют интеграцию эндокринных, автономных и поведенческих ответов на стресс-факторы. Указанные пептиды также могут быть вовлечены в осуществление контроля аппетита, возбуждения и познавательных функций. В результате долговременного воздействия стресса могут возникать тяжелые психологические и физиологические последствия, такие как тревожные расстройства, нервная анорексия и меланхолическая депрессия.
Члены семейства кортикотропин-рилизинг фактора опосредуют свои биологические действия в результате специфичного связывания с рецепторами CRF с высокой аффинностью (6, 7). Рецепторы CRF являются рецепторами, связанными с G-белком, которые действуют через аденилатциклазу и являются структурно родственными семейству секретина. Указанное семейство также включает в себя GRF, VIP, PTH и рецептор кальцитонина. Ген рецептора CRF имеет 13 экзонов, и обнаружено несколько вариантов сплайсинга данного рецептора. Рецептор CRF-R1 распределен по всему головному мозгу и обнаружен в местах передачи сенсорных и моторных сигналов (8). CRF-R2α распределен в латеральной перегородке, вентрально-медиальном гипоталамусе, ядре одиночного пути и дорсальном ядре шва, которые представляют собой области, где CRF-R1 экспрессируется очень мало или совсем не экспрессируется (9). CRF-R2β главным образом обнаружен в периферических местах, включая сердце, кровеносные сосуды, желудочно-кишечный тракт, эпидидимис, легкое и кожу (7, 10). Фармакология двух типов рецепторов отличается тем, что кортикотропин-рилизинг фактор обладает низким сродством по отношению к CRF-R2 (Ki=15-100 нМ), но высоким сродством по отношению к CRF-R1 (Ki=1-2 нМ). Другие родственные пептиды, такие как уротензин карпа, саувагин лягушки и урокортин обладают высоким сродством к CRF-R2. Мыши, нокаутированные по CRF-R2, проявляют повышенное похожее на тревожное поведение, вызванное гиперчувствительностью к стресс-факторам (11).
Обнаружено, что в ряде групп клеток, идентифицированных как места действия пептидов, вызывающих стресс-подобные автономные и поведенческие ответы, отсутствует или ослаблена экспрессия необходимого лиганда(дов), рецептора(ров) или и того и другого (12, 13). Это привело к активизации поиска дополнительных родственных CRF сигнальных молекул, которые в настоящее время насчитывают два лиганда, связанные с G-белком рецепторы, получаемые из двух отдельных генов (CRF-R1 и CRF-R2), и связывающий белок, функции которых до конца неизвестны (14, 15).
Недавно обнаружили второй родственный CRF нейропептид млекопитающих, урокортин (Ucn), и показали, что он с высоким сродством связывается с обоими известными типами рецепторов CRF, тогда как CRF с большим предпочтением связывается с CRF-R1. При центральном введении урокортин более эффективен, чем CRF, в подавлении аппетита, но менее эффективен в генерировании острых подобных тревожным эффектов и в генерализованной активации поведения (17). Это использовали для того, чтобы показать, что урокортин может, по меньшей мере, частично опосредовать некоторые связанные со стрессом эффекты, которые сначала приписывали CRF, служа в качестве эндогенного лиганда CRF-R2. Однако данная точка зрения подвергается сомнению в результате таких наблюдений, как наблюдение того, что основные клеточные места экспрессии урокортина в головном мозге не распознаются в виде интегральных компонентов центральной связанной со стрессом схемы и что большинство основных мест экспрессии CRF-R2 слабо иннервированы содержащими урокортин выростами (18). Указанные и другие данные подтверждают возможное существование одного или нескольких дополнительных лигандов рецепторов CRF в головном мозге млекопитающих.
Предшествующий уровень техники не отвечает требованиям из-за отсутствия выявленных дополнительных генов и белков урокортина. Данное изобретение удовлетворяет эту давнюю необходимость и потребность в данной области.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Быстрый прогресс в накоплении данных о последовательностях геномов человека и мыши дает возможность для идентификации новых представителей многих семейств белков. Новая пептидная последовательность, родственный урокортину пептид человека (URP), идентифицировали в опубликованной базе данных генома человека. Последовательность пептида, родственного урокортину, обладает гомологией с урокортином человека (44%), уротензином карпа (39%) и CRF человека (36%). Синтезированный пептид, родственный урокортину, с более высокой аффинностью связывается с CRF-R2 (Ki=0,5 нМ), чем с CRF-R1 (Ki=70 нМ). Пептид человека, родственный урокортину, стимулирует секрецию АКТГ из клеток передней доли гипофиза крыс, хотя значительно менее эффективно по сравнению с урокортином или CRF. Используя средства поиска гомологии последовательностей, также идентифицировали мышиный ген, кодирующий пептид из 38 аминокислот, который представляет собой новый член CRF-семейства нейропептидов. Указанный пептид, названный урокортином II (Ucn II), отличается от других известных представителей семейства тем, что он связывается с высокой избирательностью с CRF-R2. Доказательства наличия урокортина II в головном мозге крыс получены при иммуногистохимических исследованиях и исследованиях гибридизации in situ с использованием антител высокоспецифичных по отношению к урокортину II.
В одном варианте данного изобретения представлена последовательность ДНК, кодирующая урокортин II. Указанную последовательность можно выбрать из группы, состоящей из: изолированной и очищенной ДНК, которая кодирует урокортин II; изолированной и очищенной ДНК, которая в условиях высокой жесткости гибридизуется с антисмысловой последовательностью, комплементарной ДНК урокортина II, (при этом условия высокой жесткости определяют как промывку мембраны при высокой температуре и низкой концентрации соли, функционально эквивалентных 0,1 х SSC при 65°С); и изолированной и очищенной ДНК, кодирующей урокортин II, но которая отличается по последовательности вследствие вырожденности генетического кода. Указанная ДНК предпочтительно кодирует белок-предшественник, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 10.
В другом варианте данного изобретения направлением настоящего изобретения является вектор, способный экспрессировать урокортин II. Такой вектор состоит из ДНК, кодирующей урокортин II, и регуляторных элементов, необходимых для экспрессии урокортина II в клетке. В предпочтительном варианте указанный вектор кодирует белок с аминокислотной последовательностью SEQ ID NO: 11. Направлением данного изобретения также является клетка-хозяин, трансфицированная таким вектором и экспрессирующая урокортин II с указанного вектора. Белок может экспрессироваться в типе клеток, выбранном из бактериальных клеток, клеток млекопитающих, растительных клеток и клеток насекомых. В одном предпочтительном варианте белок экспрессируется в E. coli.
В еще одном варианте данное изобретение относится к выделенному и очищенному белку урокортина II человека, кодируемому описанной выше ДНК. Предпочтительно очищенный родственный урокортину пептид человека имеет аминокислотную последовательность, соответствующую SEQ ID NO: 11.
В другом варианте данное изобретение относится к антителу, направленному против белка урокортина II. Указанное антитело может представлять собой моноклональное антитело.
В еще одном варианте данное изобретение относится к фармацевтической композиции, содержащей белок урокортина II. Такую фармацевтическую композицию можно использовать для снижения температуры тела, подавления аппетита и лечения или профилактики застойной сердечной недостаточности и различных расстройств, связанных со стрессом.
В следующем варианте данное изобретение относится к последовательности ДНК, кодирующей родственный урокортину пептид человека. Указанную последовательность можно выбрать из группы, состоящей из: изолированной и очищенной ДНК, которая кодирует родственный урокортину пептид человека; изолированной и очищенной ДНК, которая в условиях высокой жесткости гибридизуется с антисмысловой последовательностью, комплементарной ДНК родственного урокортину пептида человека (при этом условия высокой жесткости определяют как промывку мембраны при высокой температуре и низкой концентрации соли, функционально эквивалентных 0,1 х SSC при 65°С); и изолированной и очищенной ДНК, кодирующей родственный урокортину пептид человека, но которая отличается по последовательности вследствие вырожденности генетического кода. Указанная ДНК предпочтительно имеет последовательность, показанную в SEQ ID NO: 1 и кодирует белок-предшественник, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 2.
В другом варианте данного изобретения направлением настоящего изобретения является вектор, способный экспрессировать родственный урокортину пептид человека. Такой вектор состоит из ДНК, кодирующей родственный урокортину пептид человека, и регуляторных элементов, необходимых для экспрессии родственного урокортину пептида человека в клетке. В предпочтительном варианте указанный вектор кодирует белок с аминокислотной последовательностью SEQ ID NO: 3. Направлением данного изобретения также является клетка-хозяин, трансфицированная таким вектором и экспрессирующая родственный урокортину пептид человека с указанного вектора. Белок может экспрессироваться в типе клеток, выбранном из бактериальных клеток, клеток млекопитающих, растительных клеток и клеток насекомых. В одном предпочтительном варианте белок экспрессируется в E. coli.
В еще одном варианте данное изобретение относится к выделенному и очищенному белку родственного урокортину пептида человека, кодируемому описанной выше ДНК. Предпочтительно очищенный родственный урокортину пептид человека имеет аминокислотную последовательность, соответствующую SEQ ID NO: 3.
В другом варианте данное изобретение относится к антителу, направленному против белка родственного урокортину пептида человека. Указанное антитело может представлять собой моноклональное антитело.
В еще одном варианте данное изобретение относится к фармацевтической композиции, содержащей белок родственного урокортину пептида человека. Такую фармацевтическую композицию можно использовать для снижения температуры тела, подавления аппетита и лечения или профилактики застойной сердечной недостаточности и различных расстройств, связанных со стрессом.
В другом варианте данного изобретения описаны различные модификации белков урокортина II и родственного урокортину пептида человека, включая модификацию последовательности и отдельных аминокислот белков. Модификации также включают в себя конъюгацию урокортина II и родственного урокортину пептида человека с флуоресцентными метками, комплексными радионуклидами и токсинами.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Чтобы можно было достичь и подробно понять перечисленные выше особенности, преимущества и объекты изобретения, а также другие вопросы, которые станут очевидными, можно представить более конкретные описания изобретения, сущность которого кратко изложена выше, со ссылками на некоторые его варианты, которые иллюстрированы прилагаемыми чертежами. Данные чертежи образуют часть описания. Однако необходимо отметить, что прилагаемые чертежи иллюстрируют предпочтительные варианты изобретения, и поэтому их не следует считать ограничивающими в данных рамках.
На фигуре 1 показана последовательность геномной ДНК человека, на основании которой спрогнозировано существование нового пептида, родственного урокортину и CRF. Геномные последовательности идентифицировали в опубликованной базе данных и использовали для расчета новой последовательности родственного урокортину пептида человека. Предполагаемым сайтом старта является положение 1, и последовательность зрелого пептида показана жирным шрифтом. Рассчитанные сайты отщепления сигнального пептида указаны стрелками.
На фигуре 2 показан предполагаемый предшественник родственного урокортину пептида человека. Подчеркнутая область представляет собой неполную последовательность кДНК, которую выделяли посредством ПЦР из библиотеки кДНК островков поджелудочной железы человека.
На фигуре 3 показано сравнение родственного урокортину пептида человека (URP) (SEQ ID No4) с человеческим Ucn (SEQ ID No:5), уротензином I (SEQ ID No:6), CRF (SEQ ID No:7), саувагином лягушки (SEQ ID No:8) и CRF/Uro морской собаки (SEQ ID No:9). Области самой высокой гомологии находятся внутри белых прямоугольников. Указано количество консервативных аминокислот.
На фигуре 4А показана выведенная аминокислотная последовательность Ucn II. Стартовый метионин, отмеченный жирным шрифтом, локализован выше кодирующей области пептида, которая ограничена рамкой. Полную нуклеотидную последовательность поместили в Genbank (инвентарный No.AF331517).
На фигуре 4В показано выравнивание Ucn II мыши (SEQ ID No:10) с гомологичными пептидами человека (SEQ ID No:11) и рыб (URP)(SEQ ID No:12) и с Ucn крысы (SEQ ID No:13) и CRF крысы/человека (SEQ ID No:14). Остатки, идентичные последовательности Ucn II мыши, ограничены рамкой. Ξ указывает сайт амидирования.
На фигурах 5А и 5В показано опосредованное родственным урокортину пептидом человека замещение 125I-саувагина, связывающегося с CRFR1 и CRFR2β. Аффинность Ucn и пептидов URP по отношению к CRFR1 и CRFR2β, стабильно экспрессированным в клетках СНО, определяли по конкурентному замещению 125I-саувагина. Представлены данные из 3 экспериментов, и значения константы диссоциации/ингибирования (Ki) (95% доверительный интервал) рассчитывали с использованием программы Prism.
На фигуре 6 (А-С) показана экспрессия мРНК урокортина II в головном мозге крыс. Микрофотографии в темном поле, показывающие мечение (белые гранулы), наблюдаемое в отдельных областях при использовании меченного изотопами зонда антисмысловой кРНК, полученной на основе кДНК урокортина II мыши. Позитивные сигналы гибридизации видны в паравентрикулярном ядре гипоталамуса (фигура 6, А), главным образом, в его магноцеллюлярном отделе (pm), при этом более диффузный сигнал виден в парвоцеллюлярной области (mp) и обширно в голубоватом месте (locus coeruleus) (LC; фигура 6, В), моторном ядре лицевого нерва (VII, фигура 6, С) и мозговых оболочках (men) на вентральной поверхности головного мозга. Другие сокращения: CBL, мозжечок; v3, третий желудочек; v4, четвертый желудочек. Увеличения: фигура 6, А и В, Х75; фигура 6, С, Х50.
На фигуре 7 показана авторадиограмма экспрессии родственного урокортину пептида человека в гипоталамусе примата. PVH, паравентрикулярные ядра; SO, супраоптические ядра; CN, хвостатое ядро; och, оптическая хиазма; me, средняя возвышенность; ac, передняя спайка; ic, внутренняя капсула; Sept, перегородка.
На фигурах 8 (А-F) показаны картины клеточной активации в ответ на центральную микроинъекцию урокортина II. Фигура 8, А-С и Е: Микрофотографии в светлом поле иммунопероксидазных препаратов, показывающие индуцированную экспрессию Fos у крыс, забитых через 2 часа после интрацеребровентрикульной (icv) инъекции 1 мкг синтетического урокортина II мыши. Микрофотографии в темном поле, показывающие гистохимические данные о локализации гибридизации мРНК CRF-R2 в областях, соответствующих областям, показанным на фигуре 8, С и Е, представлены на фигуре 8, D и F соответственно. Центральная инъекция урокортина II вызывает индукцию Fos, главным образом, в группе взаимосвязанных структур, участвующих в центральной автономной и нейроэндокринной регуляции, включая парвоцеллюлярный отдел паравентрикулярного ядра (фигура 8, А), центральное ядро миндалевидного тела (фигура 8, В) и ядро одиночного пути (NTS, фигура 8, С). Среди указанных структур только NTS является местом экспрессии CRF-R2 (фигура 8, D). Другие основные места экспрессии CRF-R2, включая вентромедиальное ядро гипоталамуса (фигура 8, F) не проявляют индуцированной урокортином II экспрессии Fos в исследованных пределах доз пептида (1-10 мкг). Все микрофотографии сделаны при увеличении 75Х.
На фигуре 9 показана активация центральных связанных со стрессом групп клеток после центральной инъекции родственного урокортину пептида человека посредством исследования стимуляции ядерной экспрессии FOS в терминальной полоске (stria terminalis) (BST), паравентрикулярном ядре гипоталамуса (PVH), центральном ядре миндалевидного тела (CeA), латеральном парабрахиальном ядре (PBI), голубоватом месте ((locus coeruleus) (LC)) и ядре одиночного пути (NTS). BSTov, ядро ложа stria terminalis (овальное нижнее ядро); ic, внутренняя капсула; CP, хвостатое ядро/скорлупа; ac, передняя спайка; V3, третий желудочек; AHA, передняя область гипоталамуса; pm, задняя магноцеллюлярная часть (паравентрикулярное ядро); fx, свод; CeAm, центральное ядро миндалевидного тела; BLA, базо-латеральное ядро миндалевидного тела; scp, верхняя ножка мозжечка; PBel, парабрахиальное ядро (наружная латеральная часть); V4, четвертый желудочек; ep, эпендима; AP, самое заднее поле; DMX, дорсальное моторное ядро блуждающего нерва; ts, одиночный путь; и cc, центральный канал.
На фигурах 10А и 10В показано влияние центрально введенного урокортина II на потребление пищи и общую моторную активность. На фигуре 10А показано среднее (±SEM; n=3-6 на группу) суммарное потребление пищи в ночное время (g) после icv-введения 1 мкг CRF, урокортина или урокортина II. CRF и урокортин в значительной степени снижали потребление пищи по сравнению с контролями, в которых инъецировали физиологический раствор, начиная с 4 ч после инъекции, тогда как действие урокортина II не проявлялось вплоть до 6 ч после обработки. *р< 0,002 (CRF и Ucn по сравнению с физиологическим раствором), **р< 0,002 (CRF, урокортин и урокортин II по сравнению с физиологическим раствором). На фигуре 10В показаны телеметрические измерения общей моторной активности, которые были в значительной степени повышены у животных, которые получали icv-инъекции CRF; ни урокортин, ни урокортин II не оказывали значимого влияния на моторную активность. *р< 0,001 (CRF по сравнению с физиологическим раствором).
На фигуре 11 показана стимуляция секреции АКТГ из клеток передней доли гипофиза урокортином и родственным урокортину пептидом человека. Клетки передней доли гипофиза крыс вводили в культуру и обрабатывали либо урокортином крыс, либо родственным урокортину пептидом человека. Секретированный АКТГ измеряли, используя набор (Nichols Institute Diagnostics).
На фигуре 12 показано влияние родственного урокортину пептида человека на уровни цАМФ в клетках A7R5, которые экспрессируют нативный CRF-R2β. Дозозависимое действие инкубации с урокортином (светлые кружки) или hURP (закрашенные кружки) в течение 30 минут на продукцию цАМФ. цАМФ измеряли посредством РИА (Biochemical Technologies).
На фигуре 13 показано влияние родственного урокортину пептида человека (hURP) на общую моторную активность у крыс.
На фигуре 14 показано влияние интрацеребровентрикулярной инъекции родственного урокортину пептида человека (URP) на температуру тела крыс.
На фигуре 15 показано влияние интрацеребровентрикулярной инъекции родственного урокортину пептида человека (hURP) на потребление крысами пищи в ночное время.
На фигуре 16 показана модель того, как родственный урокортину пептид человека действует на CRF-R1 и CRF-R2. Родственный урокортину пептид человека с высокой аффинностью связывается с CRF-R2, но не с CRF-R1, тогда как урокортин связывается с обоими рецепторами. CRF с высокой аффинностью связывается с CRF-R1, но не связывается с CRF-R2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно данному изобретению можно использовать традиционные способы молекулярной биологии, микробиологии и технологию рекомбинантной ДНК, разработанные в данной области. Полное объяснение таких способов имеется в литературе. Смотри, например, Maniatis, Fritsch and Sambrook, "Molecular Cloning: A Laboratory Manual (1982); "DNA Cloning: A Practical Approach," Volumes I and II (D.N. Glover ed. 1985); "Oligonucleotide Synthesis" (M.J. Gait ed. 1984); "Nucleic Acid Hybridization" [B.D. Hames and S.J. Higgins Eds. (1985)]; "Transcription and Translation" [B.D. Hames and S.J. Higgins Eds. (1984)]; "Animal Cell Culture" [R.I. Freshney ed. (1986)]; "Immobilized Cells and Enzymes" [IRL Press, (1986)]; B. Perbal, "A Practical Guide To Molecular Cloning" (1984).
Поэтому следующие термины, если они фигурируют в данном описании, будут иметь определения, указанные ниже.
В используемом здесь смысле термин «кДНК» будет относиться к ДНК-копии мРНК-транскрипта гена.
В используемом здесь смысле термин «выведенная аминокислотная последовательность» будет относиться к аминокислотной последовательности, определенной посредством считывания последовательности триплетов оснований нуклеотидов в кДНК.
В используемом здесь смысле термин «скрининг библиотеки» будет относиться к способу использования меченого зонда для того, чтобы при соответствующих условиях проверить, присутствует ли в конкретной библиотеке ДНК последовательность, комплементарная зонду. Кроме того, «скрининг библиотеки» можно выполнить посредством ПЦР.
В используемом здесь смысле термин «ПЦР» относится к полимеразной цепной реакции, которая является предметом патентов США № 4683195 и 4683202 Mullis, а также других модификаций, известных в настоящее время в данной области.
Все последовательности аминокислотных остатков представлены здесь формулами, в которых расположение слева направо соответствуют обычному направлению от аминоконца к карбоксильному концу. Кроме того, следует отметить, что тире в начале или в конце последовательности аминокислотных остатков указывает на пептидную связь с другой последовательностью из одного или нескольких аминокислотных остатков.
Описанные здесь аминокислоты предпочтительно находятся в «L»-изомерной форме. Однако любой остаток L-аминокислоты может быть заменен остатками в «D»-изомерной формой, при условии, что требуемое функциональное свойство полипептида связываться с иммуноглобулином сохраняется. NH2 относится к свободной аминогруппе, присутствующей на аминоконце полипептида. COOH относится к свободной карбоксильной группе, присутствующей на карбоксильном конце полипептида.
Нестандартные аминокислоты можно включить в белки посредством химической модификации существующих аминокислот или в результате искусственного синтеза белка. Нестандартная аминокислота относится к аминокислоте, которая отличается по химической структуре от двадцати стандартных аминокислот, кодируемых генетическим кодом. Посттрансляционная модификация in vivo также может приводить к наличию нестандартной аминокислоты или производного аминокислоты в белке. N-концевые NH2- и С-концевые СООН-группы белка также можно модифицировать в результате природной или искусственной посттрансляционной модификации белка.
Белки можно модифицировать заменами аминокислот. Часто некоторые изменения в результате приводят к значительным изменениям активности белков, тогда как другие оказывают незначительное влияние или не оказывают влияния. Менее всего вероятным является то, что консервативные замены сильно изменяют активность белка. «Консервативная аминокислотная замена» относится к замене аминокислоты химически сходной аминокислотой, т.е. замене неполярной кислоты другой неполярной аминокислотой; замене полярной аминокислоты другой полярной аминокислотой, кислых остатков другими кислыми аминокислотами, и т.д. Примеры предпочтительных консервативных замен указаны в таблице 1:
«Химическое производное» относится к обсуждаемому полипептиду, имеющему один или несколько остатков, химически дериватизированных в результате реакции функциональной боковой группы. Такие дериватизированные полипептиды включают, например, полипептиды, в которых свободные аминогруппы были дериватизированы с целью образования гидрохлорида амина, пара-толуол сульфонильных групп, карбобензоксигрупп, трет-бутилоксикарбонильных групп, хлорацетильных групп или формильных групп. Свободные карбоксильные группы можно дериватизировать с образованием солей, метильных или этильных сложных эфиров или других типов сложных эфиров или гидразидов. Химические производные могут включать такие пептиды, которые содержат одно или несколько производных аминокислот природного происхождения из двадцати стандартных аминокислот. Например, серин можно заместить 4-гидроксипролином; и лизин можно заместить орнитином. Пептиды, охватываемые данным изобретением, также включают в себя пептиды, имеющие одно или несколько присоединений и/или делеций остатков по сравнению с конкретным пептидом, последовательность которого здесь указана, при условии, что модифицированный пептид сохраняет требуемую биологическую активность.
«Репликон» представляет собой любой генетический элемент (например, плазмиду, хромосому, вирус), который функционирует как автономная единица репликации ДНК in vivo; т.е. способна к репликации под своим собственным контролем.
«Вектор» является репликоном, таким как плазмида, фаг или космида, с которым может быть связан другой сегмент ДНК, для того, чтобы осуществлять репликацию связанного сегмента.
«Молекула ДНК» относится к полимерной форме дезоксирибонуклеотидов (адениновых, гуаниновых, тиминовых или цитозиновых), либо в ее однонитевой форме, либо в виде двунитевой спирали. Данный термин относится только к первичной и вторичной структуре молекулы, и не ограничивает ее какими-либо конкретными третичными формами. Таким образом, этот термин включает в себя двунитевую ДНК, обнаруживаемую в числе прочего в виде линейных молекул ДНК (например, фрагментов рестрикции), вирусов, плазмид и хромосом. При обсуждении здесь структуры согласно обычной договоренности приводится только последовательность в направлении от 5' к 3' вдоль нетранслируемой нити ДНК (т.е. нить, имеющая последовательность, гомологичную мРНК).
«Начало репликации» относится к тем последовательностям ДНК, которые принимают участие в синтезе ДНК.
«Кодирующая последовательность» ДНК представляет собой последовательность двунитевой ДНК, которая транскрибируется и транслируется в полипептид in vivo в том случае, когда помещена под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяют по стартовому кодону на 5'-(амино)конце и стоп-кодону трансляции на 3'-(карбоксильном) конце. Кодирующая последовательность может включать, но не ограничена этим, прокариотические последовательности, кДНК с эукариотической мРНК, последовательности геномной ДНК из эукариотической ДНК (например, ДНК млекопитающих) и даже синтетические последовательности ДНК. Сигнал полиаденилирования и последовательность терминации транскрипции, как правило, будут локализованы с 3'-стороны от кодирующей последовательности.
Транскрипционными и трансляционными регуляторными последовательностями являются регуляторные последовательности ДНК, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и тому подобное, которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине.
«Последовательность промотора» представляет собой регуляторную область ДНК, способную связывать РНК-полимеразу в клетке и инициировать транскрипцию кодирующей последовательности, расположенной «справа» (в 3'-направлении). В целях определения согласно данному изобретению промоторная последовательность ограничена на своем 3'-конце сайтом инициации транскрипции и простирается «левее» (в 5'-направлении), включая минимальное количество оснований или элементов, необходимых для инициации транскрипции на уровне, выявляемом выше фона. В промоторной последовательности будет находиться сайт инициации транскрипции, а также домены, связывающие белок (консенсусные последовательности), ответственные за связывание РНК-полимеразы. Эукариотические промоторы часто, но не всегда, содержат «ТАТА»-боксы и «САТ»-боксы. Прокариотические промоторы кроме консенсусных последовательностей -10 и -35 содержат последовательности Шайне-Далгарно.
«Последовательность регуляции экспрессии» представляет собой последовательность ДНК, которая контролирует и регулирует транскрипцию и трансляцию другой последовательности ДНК. Кодирующая последовательность находится «под контролем» транскрипционных и трансляционных регуляторных последовательностей в клетке в том случае, когда РНК-полимераза транскрибирует кодирующую последовательность в мРНК, которая затем транслируется в белок, кодируемый кодирующей последовательностью.
«Сигнальная последовательность» может быть встроена вблизи кодирующей последовательности. Указанная последовательность кодирует сигнальный пептид, N-концевой по отношению к полипептиду, который дает клетке-хозяину сигнал о направлении полипептида к поверхности клетки или секреции полипептида в среду, и указанный сигнальный пептид отрезается клеткой-хозяином перед тем, как белок покинет клетку. Сигнальные последовательности можно обнаружить связанными с различными белками, присущими прокариотам и эукариотам.
Термин «олигонуклеотид» в используемом здесь смысле по отношению к зонду согласно данному изобретению, определяют как молекулу, содержащую два или более рибонуклеотидов, предпочтительно более трех. Его точный размер будет зависеть от многих факторов, которые, в свою очередь, зависят от конечной функции и применения олигонуклеотида.
Термин «праймер» в используемом здесь смысле относится к олигонуклеотиду, либо имеющему природное происхождение как в очищенном продукте рестрикционного расщепления, либо полученному синтетическим путем, который способен действовать как точка инициации синтеза в том случае, когда он помещен в условия, в которых индуцируется синтез продукта удлинения праймера, который комплементарен нити нуклеиновой кислоты, т.е. в присутствии нуклеотидов и индуцирующего агента, такого как ДНК-полимераза, и при подходящей температуре и рН. Праймер может быть либо однонитевым, либо двунитевым, и должен иметь достаточную длину, чтобы праймировать синтез требуемого продукта удлинения в присутствии индуцирующего агента. Точная длина праймера будет зависеть от многих факторов, включая температуру, источник праймера и применение способа. Например, для диагностических применений в зависимости от сложности последовательности-мишени олигонуклеотидный праймер обычно содержит 15-25 или более нуклеотидов, хотя он может содержать меньшее количество нуклеотидов.
Здесь выбраны праймеры в «значительной степени» комплементарные разным нитям конкретной последовательности ДНК-мишени. Это значит, что праймеры должны быть в значительной степени комплементарными для того, чтобы гибридизоваться с соответствующими им нитями. Следовательно, последовательность праймера не должна отражать точную последовательность матрицы. Например, некомплементарный нуклеотидный фрагмент может быть связан с 5'-концом праймера, при этом остальная часть последовательности праймера комплементарна нити. Альтернативно некомплементарные основания или более длинные последовательности могут быть рассеяны в праймере, при условии, что последовательность праймера в достаточной степени комплементарна последовательности или гибридизуется с ней и таким образом образует матрицу для синтеза продукта удлинения.
В используемом здесь смысле термины «эндонуклеазы рестрикции» и «ферменты рестрикции» относятся к ферментам, каждый из которых разрезает двунитевую ДНК в конкретной нуклеотидной последовательности или вблизи нее.
Клетка «трансформирована» экзогенной или гетерологичной ДНК в том случае, когда ДНК была введена внутрь клетки. Трансформирующая ДНК может быть интегрирована (ковалентно связана) или не интегрирована в геном клетки. Например, у прокариот, дрожжей и в клетках млекопитающих трансформирующая ДНК может поддерживаться в эписомном элементе, таком как плазмида. Что касается эукариотических клеток, стабильно трансформированная клетка представляет собой клетку, в которой трансформирующая ДНК оказалась интегрированной в хромосому, так что наследуется дочерними клетками посредством репликации хромосомы. Об указанной стабильности свидетельствует способность эукариотической клетки формировать линии или клоны клеток, состоящие из популяции дочерних клеток, содержащих трансформирующую ДНК. «Клон» является популяцией клеток, полученных от одной клетки или предшественника путем митоза. «Линия клеток» является клоном первичной клетки, которая способна стабильно расти in vitro в течение многих поколений.
Две последовательности ДНК «в значительной степени гомологичны» в том случае, когда, по меньшей мере, примерно 75% (предпочтительно, по меньшей мере, примерно 80% и наиболее предпочтительно, по меньшей мере, примерно 90% или 95%) нуклеотидов совпадают на протяжении определенной длины последовательностей ДНК. Последовательности, которые гомологичны в значительной степени, можно идентифицировать посредством сравнения с использованием стандартной компьютерной программы последовательностей, имеющихся в банках данных последовательностей, или в эксперименте по Саузерн-гибридизации, например, в жестких условиях, которые определяют для данной конкретной системы. Определение соответствующих условий гибридизации известно специалистам в данной области. Смотри, например, Maniatis et al., выше; DNA Cloning, Vols I and II, выше; Nucleic Acid Hybridization, выше.
Гетерологичная область конструкции ДНК является идентифицируемым сегментом ДНК в большей по размеру молекуле ДНК, который в природе не выявляется связанным с большей молекулой. Таким образом, в том случае, когда гетерологичная область кодирует ген млекопитающих, ген, как правило, будет фланкирован ДНК, которая не фланкирует геномную ДНК млекопитающих в геноме исходного организма. В другом примере кодирующая последовательность является конструкцией, в которой сама кодирующая последовательность не выявляется в природе (например, кДНК, когда геномная кодирующая последовательность содержит интроны, или синтетические последовательности, имеющие кодоны, отличные от кодонов нативного гена). Алелльные варианты или мутационные события природного происхождения не вызывают появления гетерологичной области ДНК, определение которой здесь дано.
Метки, обычно используемые для указанных исследований, являются радиоактивными элементами, ферментами, химическими веществами, которые флуоресцируют при воздействии ультрафиолетового излучения, и другими метками. Известно множество флуоресцентных материалов, и их можно использовать в качестве меток. Такие материалы включают, например, флуоресцеин, родамин, аурамин, техасский красный, АМСА синий и люцифер желтый. Конкретным детектирующим материалом является антикроличье антитело, полученное в организме коз и конъюгированное с флуоресцеином с помощью изотиоцианата.
Конкретная система анализа, разработанная и используемая в данной области, известна как рецепторный анализ. В рецепторном анализе материал, который необходимо анализировать, соответствующим образом метят, и затем в определенные тестируемые колонии клеток вносят некоторое количество меток, после чего проводят исследования связывания, чтобы определить степень, с которой меченый материал связывается с клеточными рецепторами. Таким образом можно установить различия между материалами по аффинности.
В используемом здесь смысле подразумевается, что термин «хозяин» включает в себя не только прокариот, но также эукариот, таких как клетки дрожжей, растений и животных. Рекомбинантную молекулу ДНК или ген, который кодирует белок согласно данному изобретению, можно использовать для трансформации хозяина с использованием любого из способов, как правило, известных специалистам в данной области. Прокариотические хозяева могут включать E. coli, S. tymphimurium, Serratia marcescens и Bacillus subtilis. Эукариотические хозяева включают дрожжи, такие как Pichia pastoris, клетки млекопитающих и клетки насекомых.
В общем, экспрессирующие векторы, содержащие промоторные последовательности, которые способствуют эффективной транскрипции встроенного фрагмента ДНК, используют в связи с хозяином. Экспрессирующий вектор обычно содержит начало репликации, промотор(ры), терминатор(ры), а также специфичные гены, которые способны обеспечивать фенотипическую селекцию трансформированных клеток. Трансформированных хозяев можно ферментировать и культивировать согласно способам, известным в данной области для получения оптимального роста клеток.
Для конструирования экспрессирующих векторов, содержащих соответствующие сигналы регуляции транскрипции и трансляции можно использовать способы, хорошо известные специалистам в данной области. Смотри, например, способы, описанные в Sambrook et al., 1989, Molecular cloning: A Laboratory Manual (2nd Ed.), Cold Spring Harbor Press, N.Y. Ген и последовательности регуляции его транскрипции называют «оперативно связанными», если последовательности регуляции транскрипции эффективно контролируют транскрипцию гена. Векторы согласно изобретению включают, но не ограничены указанным, плазмидные векторы и вирусные векторы.
Направлением данного изобретения является последовательность ДНК, кодирующая урокортин II. Указанная последовательность может быть изолированной и очищенной ДНК, которая кодирует урокортин II. Альтернативно она может представлять собой изолированную и очищенную ДНК, которая в условиях высокой жесткости гибридизуется с антисмысловой последовательностью, комплементарной ДНК урокортина II, при условиях высокой жесткости (определяемых как промывка мембраны при высокой температуре и низкой концентрации соли, функционально эквивалентных 0,1 х SSC при 65°С). Наконец, ДНК может представлять собой изолированную и очищенную ДНК, кодирующую урокортин II, но которая отличается по последовательности вследствие вырожденности генетического кода. Указанная ДНК предпочтительно кодирует белок с аминокислотной последовательностью SEQ ID NO: 10 или аминокислотной последовательностью SEQ ID NO: 11.
Направлением данного изобретения также является вектор, способный экспрессировать урокортин II. Такой вектор состоит из ДНК, кодирующей урокортин II, и регуляторных элементов, необходимых для экспрессии урокортина II в клетке. В предпочтительном варианте указанный вектор кодирует белок с аминокислотной последовательностью SEQ ID NO: 10 или аминокислотной последовательностью SEQ ID NO: 11. Направлением данного изобретения также является клетка-хозяин, трансфицированная таким вектором и экспрессирующая урокортин II с такого вектора. Белок можно экспрессировать в типе клеток, выбранном из бактериальных клеток, клеток млекопитающих, растительных клеток и клеток насекомых. В предпочтительном варианте белок экспрессируется в E. coli.
Направлением данного изобретения также является выделенный и очищенный белок урокортина II, кодируемый ДНК, которая описана выше. Предпочтительно очищенный урокортин II имеет аминокислотную последовательность, соответствующую SEQ ID NO: 10 или SEQ ID NO: 11.
Направлением данного изобретения также является антитело, направленное против белка урокортина II. Указанное антитело предпочтительно является моноклональным антителом.
Кроме того, направлением данного изобретения является фармацевтическая композиция, содержащая белок урокортина II и фармацевтически приемлемый носитель. Такую фармацевтическую композицию можно использовать для понижения температуры тела, подавления аппетита, лечения или профилактики застойной сердечной недостаточности, лечения стресса и тревожного состояния и изменения нежелательно низких уровней секреции АКТГ.
Направлением данного изобретения также является последовательность ДНК, кодирующая родственный урокортину пептид человека. Указанная последовательность может представлять собой изолированную и очищенную ДНК, которая кодирует родственный урокортину пептид человека. Альтернативно, она может представлять собой изолированную и очищенную ДНК, которая в условиях высокой жесткости гибридизуется с антисмысловой последовательностью, комплементарной ДНК родственного урокортину пептида человека (при этом условия высокой жесткости определяют как промывку мембраны при высокой температуре и низкой концентрации соли, функционально эквивалентных 0,1 х SSC при 65°С). Наконец, ДНК может представлять собой изолированную и очищенную ДНК, кодирующую родственный урокортину пептид человека, но которая отличается по последовательности вследствие вырожденности генетического кода. Указанная ДНК предпочтительно будет иметь последовательность, показанную в SEQ ID NO: 1 и предпочтительно будет кодировать белок-предшественник, имеющий аминокислотную последовательность SEQ ID NO: 2, который протеолитически процессируется в белок с аминокислотной последовательностью SEQ ID NO: 3.
Направлением данного изобретения также является вектор, способный экспрессировать родственный урокортину пептид человека. Такой вектор состоит из ДНК, кодирующей родственный урокортину пептид человека, и регуляторных элементов, необходимых для экспрессии родственного урокортину пептида человека в клетке. В предпочтительном варианте указанный вектор кодирует белок с аминокислотной последовательностью SEQ ID NO: 3. Направлением данного изобретения также является клетка-хозяин, трансфицированная таким вектором и экспрессирующая родственный урокортину пептид человека с указанного вектора. Белок может экспрессироваться в типе клеток, выбранном из бактериальных клеток, клеток млекопитающих, растительных клеток и клеток насекомых. В предпочтительном варианте белок экспрессируется в E. coli.
Направлением данного изобретения также является выделенный и очищенный белок родственного урокортину пептида человека, кодируемый описанной выше ДНК. Предпочтительно очищенный родственный урокортину пептид человека имеет аминокислотную последовательность, соответствующую SEQ ID NO: 3.
Направлением данного изобретения также является антитело, направленное против белка родственного урокортину пептида человека. Указанное антитело предпочтительно является моноклональным антителом.
Кроме того, направлением данного изобретения является фармацевтическая композиция, содержащая белок родственного урокортину пептида человека и фармацевтически приемлемый носитель. Такую фармацевтическую композицию можно использовать для снижения температуры тела, подавления аппетита, лечения или профилактики застойной сердечной недостаточности, лечения стресса и тревожного состояния и изменения нежелательно низких уровней секреции АКТГ.
Направлением данного изобретения также является урокортин II или родственный урокортину пептид человека, мутированный так, чтобы он содержал остаток тирозина, предназначенный для радиоиодирования белка. Одной из конкретных модификаций является присоединение последовательности, состоящей из Tyr-Gly, к N-концу урокортина II или родственного урокортину пептида человека.
Направлением данного изобретения также являются делеционные мутанты урокортина II или родственного урокортину пептида человека. Особенно пригодной делецией является делеция от одной до пяти аминокислот из N-конца белка.
Направлением данного изобретения также является урокортин II или белок родственного урокортину пептида человека, в котором стандартные изомерные «L-формы» аминокислот заменены изомерными «D-формами» аминокислот. В белке, родственном урокортину человека, особенно пригодна замена остатка изолейцина, соответствующего положению 9 SED ID NO: 3, D-изолейцином, D-фенилаланином и D-лейцином, или D-формой другой аминокислоты. Другая применимая замена представляет собой замену остатка глутаминовой кислоты в положении 17 SEQ ID NO: 3 или 11 D-глутаминовой кислотой.
Направлением данного изобретения также является урокортин II или родственный урокортину пептид человека, в котором различные аминокислоты были заменены нестандартными аминокислотами. Примерами таких нестандартных аминокислот являются Сα-метилированный лейцин, Сα-метилированный аланин, N-im-бензилгистидин, 4-гидроксипролин, 5-гидроксилизин, 3-метилгистидин, гомосерин и орнитин.
Направлением данного изобретения также является урокортин II или белок родственного урокортину пептида человека, имеющий ацилированный N-конец. Указанное ацилирование белка можно использовать для того, чтобы связать молекулу, такую как жирная кислота, с N-концом белка, чтобы защитить Ucn II или URP от ферментативного расщепления или чтобы изменить различные свойства белка, такие как его гидрофильность/гидрофобность. Указанную модификацию можно использовать для изменения длительности существования или биодоступности белка in vivo.
Направлением данного изобретения также является урокортин II или белок родственного урокортину пептида человека, который был модифицирован так, чтобы он содержал флуоресцентную метку, для применения с целью визуализации или в биологическом анализе.
Направлением данного изобретения также является урокортин II или белок родственного урокортину пептида человека, конъюгированный с комплексообразующим агентом для радионуклидов. Ucn II в комплексе с радионуклидом можно использовать для сцинтиграфии или в различных анализах.
Направлением данного изобретения также является урокортин II или родственный урокортину пептид человека, конъюгированный с токсином. Полученный в результате токсичный конъюгат можно использовать для целенаправленного разрешения клеток, несущих рецептор CRF.
Следующие примеры приведены с целью иллюстрации различных вариантов изобретения и никоим образом не подразумевают ограничения данного изобретения.
ПРИМЕР 1
ИДЕНТИФИКАЦИЯ РОДСТВЕННОГО УРОКОРТИНУ ПЕПТИДА ЧЕЛОВЕКА
В ходе попытки идентифицировать новые лиганды CRF-R сконструировали скрытую модель Маркова (НММ) на основе выравнивания Clustal W известных белков семейства CRF, включая CRF крысы/человека, Ucn крысы, Ucn человека, саувагин лягушки и уротензин I рыбы-прилипалы, используя пакет компьютерных программ HMMER (Sean Eddy, Department of Genetics, Washington University, St. Louis, MO; смотри ссылку 19). Указанную НММ использовали для поиска в опубликованной базе данных генома человека, и выявили ВАС (Genbank, инвентарный No: AC005903), полученную из хромосомы 3р21.3-4, которая содержала область из 109 п.н., проявляющую значительную степень гомологии последовательности, но которая не была частью ранее идентифицированного гена. Указанную область удлинили до 621 п.н. при идентификации клона EST человека, который перекрывался с данной последовательностью (Genbank, инвентарный No. BE622276). Однако в последовательности человека отсутствует консенсусный сайт протеолитического расщепления, который обеспечивал бы возможность С-концевого процессинга пептида. Поэтому белок обозначили как последовательность родственного урокортину пептида человека (hURP). На фигуре 1 показана нуклеотидная последовательность (SEQ ID NO: 1) рассчитанной открытой рамки считывания белка URP человека. Указанный ген кодирует пептид с аминокислотной последовательностью SEQ ID NO: 2.
Чтобы подтвердить существование и последовательность гена родственного урокортину пептида человека, использовали олигонуклеотидные праймеры, сходные с праймерами, используемыми для амплификации последовательности родственного урокортину пептида человека из геномного клона, чтобы выделить неполный фрагмент кДНК посредством ПЦР из библиотеки кДНК островков поджелудочной железы человека. Данный фрагмент также субклонировали в векторе pGEM и секвенировали. Последовательность кДНК соответствовала части геномной последовательности. Неполная последовательность кДНК соответствует подчеркнутой последовательности на фигуре 2. Последовательности, показанные на фигурах 1 и 2, кодируют полипептидный предшественник родственного урокортину пептида человека. Первые 19 нуклеотидов родственного урокортину пептида человека кодируют сигнальный пептид, который отщепляется во время посттрансляционной модификации белка, давая зрелый родственный урокортину пептид человека с аминокислотной последовательностью:
I V L S L D V P I G L L Q I L L E Q A R A R A A R E Q A T T N A R I L A R V G H C-NH2 (SEQ ID NO: 3).
На фигуре 3 показаны результаты сравнения гомологии между аминокислотами 72-109 родственного урокортину пептида человека и эквивалентными сегментами урокортина человека, уротензина I человека, кортикотропин-рилизинг фактора (CRF), саувагина лягушки и CRF/Uro морской собаки. Гомология в указанной области находится в пределах от 26 до 42%.
ПРИМЕР 2
ИДЕНТИФИКАЦИЯ УРОКОРТИНА II МЫШИ
Фрагментарные зонды кДНК, полученные на основе последовательности гена человека, специфично перекрестно гибридизовались с тканью крыс (головной мозг), свидетельствуя о том, что существует приемлемая степень гомологии между двумя видами. На основании этой последовательности человека сконструировали праймеры, чтобы идентифицировать гомологичный ген мыши способом быстрой амплификации концов кДНК (RACE). Приготовленную посредством RACE кДНК получали из поли(А+)РНК целого головного мозга мыши с использованием набора для амплификации кДНК RACE SMART (Clontech). Реакции ПЦР проводили в условиях низкой жесткости (низкая Tm), чтобы попытаться обеспечить максимальное гетерологичное праймирование. Первый раунд амплификации выполняли с использованием протокола «touchdown» (94°С, 30 с; ступенчато от 70°С до 55°С, 30 с; 72°С, 3 мин) с последующим вторым раундом амплификации с множественными наборами гнездовых праймеров (94°С, 20 с; 55°С, 20 с; 72°С, 3 мин). Отобранные для испытания продукты ПЦР клонировали в pCRII-TOPO (Invitrogen) для секвенирования обеих нитей. Отобранные для испытания 5'- и 3-продукты реакции идентифицировали на основе их предсказанного размера (рассчитанного на основе последовательности человека), клонировали и секвенировали.
Рассчитанная аминокислотная последовательность Ucn II мыши указана на фиг. 4А. Ген кодирует предшественник из 112 аминокислот, и С-конец содержит кодирующий район предполагаемого зрелого пептида из 38 аминокислот, указанный в области, ограниченной рамкой (фиг. 4А). За С-концевой частью кодирующей последовательности следует глицин и спаренные основные остатки (R-R), предположительно вовлеченные в амидирование и отщепление от предшественника соответственно.
Существуют два других предполагаемых или известных родственных урокортину пептида: пептид человека, пептидную последовательность которого рассчитали на основании опубликованных EST человека, а также недавно клонированный (20) URP рыбы-собаки (из Takifugu rubripes). Выравнивание с родственными урокортину пептидами человека и рыб, Ucn крысы и CRF крысы/человека показано на фиг. 4В. На аминокислотном уровне кодирующая область Ucn II мыши проявляет 77 и 45% гомологии с родственными урокортину пептидами человека и рыб соответственно. Ucn II мыши сравнительно близок известным представителям данного семейства пептидов, проявляя 36 и 44% идентичность аминокислот с CRF крысы и UCN крысы соответственно. Если допустить консервативные замены, сходство возрастает до 62% (с CRF) и 59% (Ucn).
ПРИМЕР 3
СИНТЕЗ ПЕПТИДОВ
Ucn II мыши и Ucn-родственный пептид человека синтезировали вручную, используя твердофазный способ, смолу на основе метилбензгидриламина и Boc-стратегию (21). Для удаления Boc-группы использовали трифторуксусную кислоту, 60% в дихлорметане. Сборку основной цепи осуществляли с помощью диизопропилкарбодиимида. Пептиды отщепляли и удаляли защитные группы во фтористо-водородной кислоте и очищали, используя ОФ-ВЭЖХ и три системы растворителей (фосфат триэтиламмония при рН 2,25 и 6,5 и/или 0,1% ТФУ) (22). Пептиды имели чистоту более 95% согласно независимым критериям ВЭЖХ и CZE. Чтобы подтвердить состав препаратов использовали масс-спектры.
ПРИМЕР 4
АКТИВАЦИЯ РЕЦЕПТОРОВ ПОСРЕДСТВОМ Ucn II
Аффинность Ucn II по отношению к рецепторам CRF-R1 и CRF-R2 оценивали с использованием радиорецепторного анализа. Неочищенные фракции мембран получали из клеток СНО, стабильно экспрессирующих либо клонированный CRF-R1, либо CRF-R2β. Тестируемые пептиды и радиолиганд, 125I-[Tyr0, Glu1, Nle17]-саувагин, разбавляли в буфере для анализа (20 мМ HEPES, 2 мМ EGTA, 0,1% БСА, 10% сахароза, рН 7,6) и смешивали с мембранными препаратами рецепторов в планшетах для микротитрования MAGV (Millipore), предварительно покрытых 0,1% полиэтиленимином. Реакционную смесь инкубировали в течение 90 мин при комнатной температуре, после чего быстро дважды промывали буфером для анализа и фильтровали. Комплекс радиолиганда количественно оценивали посредством счета гамма-излучения. Константы ингибирования связывания определяли с использованием компьютерной программы Prism. Результаты суммированы в таблице 2.
(Связывание)
цАМФ
(Связывание)
цАМФ
Значения определяли из 3-6 независимых экспериментов, используя стабильно трансфицированные клетки СНО или их мембраны для каждого тестируемого пептида. Значения EC50 и Ki определяли, используя компьютерную программу Prism. Значения их log10 усредняли (γ). Средние EC50 или Ki принимали равными 10γ. Рассчитывали стандартное отклонение значений log10 (δ). Пределы принимали равными [(10γ)10δ или 10γ/10δ].
По сравнению с урокортином Ucn II был, по меньшей мере, в 1000 раз менее эффективным при конкурировании с меченым саувагином за связывание с CRF-R1, тогда как он обладал почти одинаковой эффективностью с Ucn при конкурировании за связывание с CRF-R2. Указанную значительную избирательность в отношении рецептора типа 2 также наблюдали в случае активации рецептора, которую измеряли по накоплению внутриклеточного цАМФ. Стабильно трансфицированные клетки СНО (культивируемые в DMEM/10% FBS) высевали в 48-луночные чашки для культуры ткани (Costar) и давали возможность восстановиться в течение 24 часов. Среду заменяли на DMEM/0,1% FBS, по меньшей мере, за два часа до обработки. Клетки предварительно инкубировали в течение 30 мин с 0,1 мМ 3-изобутил-1-метилксантином и затем подвергали воздействию пептидов в течение 20 мин при 370С. Экстрагировали внутриклеточный цАМФ и измеряли в двух повторах из лунок в трех повторах с использованием набора для РИА (Biomedical Technologies). При анализе цАМФ Ucn II проявлял сравнимую с Ucn эффективность в отношении CRF-R2 (таблица 2). Экстремально низкая аффинность Ucn II в отношении CRF-R1 мешала определению его эффективности на данном рецепторе.
ПРИМЕР 5
Связывание выполняли в 96-луночных планшетах Durapore, 0,2 мкм, используя фильтрационную систему для множественного скринингового анализа (Millipore). Каждая лунка содержала общий объем, равный 200 мкл, состоящий из 50 мкл буфера для связывания (10% сахароза, 0,1% БСА, 2 мМ EGTA, 20 мМ буфер HEPES, рН 7,5); 50 мкл немеченого конкурирующего агента (урокортин или родственный урокортину пептид человека) при разных разведениях в буфере для связывания; 50 мкл 125I-саувагина в концентрации 150000 имп/мин/лунку; и 50 мкл клеточных мембран. Планшеты инкубировали в течение 1 часа при комнатной температуре, фильтровали в вакууме, дважды промывали буфером для связывания и давали возможность высохнуть. Отдельные фильтры выбивали и подсчитывали, используя гамма-счетчик.
Опосредованное родственным урокортину пептидом человека замещение 125I-саувагина, связывающегося с CRFR1 и CRFR2β, стабильно экспрессированными в клетках СНО, показано на фигурах 5А и 5В. На основе полученных данных обнаружено, что родственный урокортину пептид человека имеет константу диссоциации (Ki), равную 78 нМ для CRF-R1 и 0,23 нМ для CRF-R2β. С другой стороны, Ucn имел константу диссоциации, равную 0,13 нМ для CRF-R1 и 0,15 нМ для CRF-R2β. Следовательно, родственный урокортину пептид человека гораздо более специфичен в отношении рецептора кортикотропин-рилизинг фактора типа II, чем урокортин.
ПРИМЕР 6
ЭКСПРЕССИЯ мРНК Ucn II
Гистохимический анализ гибридизации выполняли для того, чтобы проанализировать характер экспрессии мРНК Ucn II в головном мозге мыши и крысы. Животных подвергали глубокой анестезии хлоралгидратом (350 мг/кг, в/б) и перфузировали через восходящую часть дуги аорты физиологическим раствором, затем ледяным 4% параформальдегидом в 0,1% боратном буфере, рН 9,5. Головной мозг постфиксировали в течение 16 часов и подвергали криогенной заморозке в течение ночи в 10% сахарозе в 0,1 М фосфатном буфере. Нарезали четыре (мыши) или шесть (крысы) серий замороженных срезов толщиной 30 мкм, используя санный микротом, собирали в холодном криоконсерванте, основанном на этиленгликоле, и хранили при -20°С до гистохимической обработки.
Гибридизацию in situ выполняли, используя 35S-меченые антисмысловые и смысловые (контроль) зонды кРНК (23), сконструированные сначала посредством линеаризации плазмиды TOPO-II, содержащей кДНК мыши. Зонды метили до удельных активностей 1-3 х 109 расп./мин/мкг, наносили на предметные стекла при концентрации примерно 107 имп/мин/мл и гибридизовали в течение ночи при 56°С в условиях высокой жесткости (50% формамид). Конечные промывки проводили в 15 мМ NaCl/1,5 мМ цитрат натрия при 65-68°С. Предметные стекла затем обезвоживали и экспонировали с рентгеновской пленкой (β-Max; Kodak) в течение 16 ч, и затем покрывали жидкой эмульсией Kodak NTB-2 и экспонировали при 4°С в течение 21-28 дней.
Гистохимический анализ гибридизации показал совместимый и ограниченный характер экспрессии мРНК Ucn II в головном мозге мыши и крысы. Зонды на основе смысловых нитей, меченные до сходных удельных активностей, как и антисмысловые зонды, не давали превышающих фон сигналов гибридизации. Регистрируемое распределение мРНК Ucn II преимущественно наблюдали в подкорковой области, при этом основные места экспрессии включали связанные со стрессом группы клеток, такие как паравентрикулярные, супраоптические и дугообразные ядра гипоталамуса, и голубоватое место (locus coeruleus) ростральных отделов моста (фиг. 6). Моторные ядра ствола головного мозга (тройничного, лицевого, подъязычного нервов), а также вентральный рог спинного мозга также идентифицировали как места экспрессии мРНК Ucn II. Среди ненейронных элементов позитивные сигналы гибридизации наблюдали равномерно в оболочках мозга, но не в хориоидном сплетении или эпендиме. Не получено очевидного свидетельства экспрессии мРНК Ucn II глиальными элементами.
ПРИМЕР 7
ЭКСПРЕССИЯ РОДСТВЕННОГО УРОКОРТИНУ ПЕПТИДА В ГОЛОВНОМ МОЗГЕ ПРИМАТОВ
Экспрессию родственного урокортину пептида человека в головном мозге приматов исследовали посредством гибридизации in situ. Гибридизацию in situ выполняли на срезах ткани головного мозга Macaca fascicularis, используя 35S-меченый зонд антисмысловой кРНК, соответствующий примерно 400 парам оснований родственного урокортину пептида человека. Зонд наносили на предметные стекла при концентрации 107 имп/мин/мл, и давали возможность гибридизации продолжаться в течение ночи. После гибридизации предметные стекла обрабатывали 20 мкг/мл рибонуклеазы А в течение 30 минут при 37°С и промывали 15 нМ NaCl/1,5 мМ цитратом натрия/50% формамидом при 70°С. Покровные стекла обезвоживали и экспонировали с рентгеновской пленкой (BetaMax; Kodak) в течение 24 часов. Авторадиограмма образца показана на фиг. 7. Позитивный сигнал для URP наблюдали в паравентрикулярных (PVH) и супраоптических ядрах гипоталамуса приматов.
ПРИМЕР 8
ЭКСПРЕССИЯ Fos, ИНДУЦИРОВАННАЯ Ucn II
Чтобы идентифицировать группы клеток, отвечающие на центральное введение Ucn II, и чтобы оценить соответствующее этим клеткам распространение в местах экспрессии CRF-R2, проводили мониторинг индуцированной экспрессии продукта гена немедленного и раннего ответа Fos в ответ на icv-введение пептида. Перед экспериментом взрослых самцов крыс Sprague-Dawley (250-300 г в начале эксперимента) и мышей С57 BL/6 (25-40 г) содержали в помещении для разведения при цикле свет:темнота 12:12 и при свободном доступе к пище и воде. Для интрацеребровентрикулярных (icv) инъекций крыс анестезировали кетамином/ксилазином/ацепромазином, и стереотаксически имплантировали им направляющую канюлю калибра 26, заканчивающуюся в латеральном желудочке. Для внутривенного (в/в) введения пептидов в яремную вену животным вводили постоянные катетеры. Крысам, получавшим icv-инъекции, также внутрибрюшинно имплантировали датчик, чтобы на расстоянии контролировать уровни общей активности и температуру тела (Mini-Mitter). После хирургической операции животным давали возможность для выздоровления в течение 7 дней перед любым из экспериментов, и в указанный период животных ежедневно брали в руки. Все процедуры были одобрены институтским комитетом по содержанию и использованию животных Salk Institute.
Чтобы проследить характеры индуцированной экспрессии Fos крысам в 10 часов утра либо icv, либо в/в инъецировали синтетический Ucn II (1, 5 или 10 мкг/животное в 2 мкл физиологического раствора для icv-инъекций, или 200 мкл для в/в-введения), или только один наполнитель, и перфузировали спустя два часа. Чтобы проследить влияние введения пептида на потребление пищи животным проводили icv-инъекции синтетического Ucn II мыши, Ucn крысы или CRF крысы/человека за 30 мин до выключения света. Затем измеряли потребление каждый час в течение 6 ч и на 12 ч. Данные анализировали с использованием дисперсионного анализа повторных измерений (ANOVA) с поправками Бонферони для множественных сравнений, применяемыми, когда это оправдано.
Для иммуногистохимического анализа ткань предварительно последовательно обрабатывали 0,3% перекисью водорода и 1% борогидридом натрия. Затем пермеабилизовали в PBS/0,2% тритон Х-100 и инкубировали с первой антисывороткой в течение 48 ч в PBS/2% блокирующей сыворотки. Иммунореактивность Fos локализовали с использованием поликлональной антисыворотки, полученной у кроликов против N-концевого синтетического фрагмента белка Fos человека (Santa Cruz Biotechnology, 1:5К). Локализацию выполняли с использованием традиционного иммунопероксидазного способа с авидином-биотином и усилением никелем, как описано в (24).
Инъекция 1 мкг синтетического Ucn II вызывала активационные ответы, которые наиболее заметны были в группе взаимосвязанных структур, вовлеченных в центральную автономную регуляцию (25, 26). Указанные структуры включали отдельные положения в ядре ложа терминальной полоски (stria terminalis), центральном ядре миндалевидного тела, паравентрикулярном ядре гипоталамуса (PVH), парабрахиальном ядре и ядре одиночного пути (NTS; фиг. 8). Из них только NTS было описано как локус экспрессии CRF-R2 (27). Индукция Fos в других основных местах экспрессии CRF-R2, включая латеральную перегородку, ядра шва среднего мозга и вентромедиальное ядро гипоталамуса (27, 28), не отличалась от индукции, наблюдаемой в контролях с инъекциями физиологического раствора. Более высокие дозы пептида (5 или 10 мкг) вызывали более сильные активационные ответы сходного распределения.
Чтобы проконтролировать потенциальные системные эффекты icv-инъекций, сходные пределы доз Ucn II вводили внутривенно отдельным группам крыс. Только самая высокая доза (10 мкг) вызывала индукцию Fos, которая была заметно выше контрольных уровней. Хотя картина была сходной с таковой, наблюдаемой в ответ на центральные инъекции, ни количество меченых клеток, ни интенсивность их окрашивания не достигали того уровня, который достоверно наблюдали после icv-инъекций 1 мкг Ucn II.
ПРИМЕР 9
ЭКСПРЕССИЯ FOS В ГОЛОВНОМ МОЗГЕ, СТИМУЛИРОВАННАЯ РОДСТВЕННЫМ УРОКОРТИНУ ПЕПТИДОМ
Активацию центральных, связанных со стрессом групп клеток родственным урокортину пептидом человека оценивали посредством выявления продукта гена Fos в клетках после инъекции родственного урокортину пептида человека. Крысам имплантировали направляющую канюлю в латеральный желудочек головного мозга за семь дней до эксперимента. В день тестирования крысам инъецировали 5 мкг синтетического родственного урокортину пептида человека в 5 мкл стерильного физиологического раствора. Крыс забивали через два часа, и готовили покровные текла с различными срезами головного мозга. Покровные стекла окрашивали для локализации иммунореактивности Fos иммунопероксидазным способом с использованием поликлональной сыворотки, полученной у кроликов против остатков 3-16 белка Fos человека.
Как показано на фигуре 9, иммунореактивность Fos выявляли в ядре ложа терминальной полоски (BST), паравентрикулярном ядре гипоталамуса (PVH), центральном ядре миндалевидного тела (CeA), латеральном парабрахиальном ядре (PBI), голубоватом месте (LC) и ядре одиночного пути (NTS). Каждое из указанных мест ранее рассматривали как место активности родственного CRF пептида.
ПРИМЕР 10
ВЛИЯНИЕ Ucn II НА ПОВЕДЕНИЕ
Подобно CRF и Ucn, Ucn II также способен оказывать действие на центральную систему, ингибируя потребление пищи (фиг. 10А). Измерения для отдельных групп крыс, которым инъецировали указанные пептиды (1 мкг, icv) в начале ночной фазы их цикла день-ночь, показывают значимую взаимосвязь между обработкой и временной точкой [F (18,95)=4,22, p<0,0001], при этом оба основных эффекта достигали достоверного значения. Все три пептида в значительной степени снижали потребление пищи на протяжении 12-часового интервала, при этом степень подавления составляла в пределах от 30 (CRF) до 35% (Ucn II) и до 70% (Ucn). Указанные воздействия имели тенденцию к дифференциальному распределению в течение времени, при этом как Ucn-, так и CRF-обработанные животные ели значительно меньше, чем контрольные животные, обработанные физиологическим раствором, в более ранние сроки тестируемого периода (4-5 ч), чем крысы, обработанные Ucn II (6 ч).
У тех же самых указанных субъектов общую моторную активность и температуру тела контролировали телеметрически (фиг. 10В). Анализ данных активности выявил значимую взаимосвязь между действием лекарственного препарата и временной точкой [F (33,110)=1,94, p<0,006], при этом оба основных эффекта достигали достоверного значения. Апостериорные (post-hoc) сравнения показали, что животные, которые получали CRF, были в значительной степени более активными, чем крысы, обработанные носителем, в течение интервала 2-6 ч после инъекции (p< 0,001). Ни обработка Ucn ни обработка Ucn II не вызывали достоверных изменений указанного критерия ни в одной временной точке после инъекции. Также регистрировали внутреннюю температуру тела, причем каждый пептид вызывал сравнительно умеренные (0,5-1°С) и временные (2 ч) гипотермические ответы (данные не показаны).
ПРИМЕР 11
БИОЛОГИЧЕСКИЙ АНАЛИЗ ОПОСРЕДОВАННЫХ hURP ВОЗДЕЙСТВИЙ НА КЛЕТКИ ПЕРЕДНЕЙ ДОЛИ ГИПОФИЗА КРЫС IN VITRO
Для оценки действия на гипофиз измеряли ответ в виде секреции АКТГ на родственный урокортину пептид человека в первичных культурах клеток передней доли гипофиза крыс, как описано в (30). Уровни АКТГ определяли с использованием набора для иммуноанализа АКТГ Nichols Institute Diagnostics. Клетки передней доли гипофиза крыс обрабатывал либо урокортином крыс, либо родственным урокортину пептидом человека, и измеряли уровень секретируемого АКТГ, используя набор (Nichols Institute Diagnostics). Влияние урокортина и родственного урокортину пептида человека на секрецию АКТГ показано на фигуре 11. Обнаружено, что стимуляция секреции АКТГ в клетках передней доли гипофиза менее чувствительна к родственному урокортину пептиду человека, чем к урокортину.
ПРИМЕР 12
БИОАНАЛИЗ ВЛИЯНИЯ hURP НА КЛЕТКИ А7R5 IN VITRO
Определяли влияние hURP на уровни цАМФ в клетках А7R5, которые экспрессируют нативный CRF-R2β. Линию клеток А7R5 поддерживали в DMEM с добавлением 10% фетальной бычьей сыворотки, 2 мМ L-глутамина, 100 мкг/мл стрептомицина. Клетки высевали при плотности 10000 клеток/см2 и выращивали в течение шести дней. Клетки, лишенные сыворотки, предварительно инкубировали с 0,1 мМ 3-изобутил-1-метилксантином в среде для анализа в течение 20 минут и обрабатывали указанными концентрациями пептида в течение 30 минут. Уровни цАМФ измерены посредством РИА (Biochemical Technologies) и показаны на фигуре 12. Родственный урокортину пептид человека оказывал сходное с урокортином влияние на продукцию цАМФ.
ПРИМЕР 13
ВЛИЯНИЕ РОДСТВЕННОГО УРОКОРТИНУ ПЕПТИДА ЧЕЛОВЕКА НА ОБЩУЮ АКТИВНОСТЬ
Чтобы определить играет ли родственный урокортину пептид человека роль в генерации стресс-ответа, исследовали влияние родственного урокортину пептида человека на общую моторную активность. Канюли хирургическим путем вводили в правый латеральный желудочек, тогда как телеметрические устройства имплантировали внутрибрюшинно, чтобы обеспечить непрерывный мониторинг общей моторной активности. Животным давали период выздоровления после хирургической операции, равный семи дням. В это время животных ежедневно брали руками, чтобы животные привыкли к процедуре инъекции. В день инъекции исходную активность регистрировали в течение четырех часов. В 6:00 вечера, во время выключения света, животные получали инъекцию либо 5 мкл физиологического раствора, либо 5 мкл физиологического раствора, содержащего всего 5 мкг родственного урокортину пептида человека. Счет активности суммировали на протяжении периода времени, равного четырем часам. Результаты суммированы на фигуре 13. Не наблюдали значимого отличия в общей моторной активности у животных, которым инъецировали родственный урокортину пептид человека, по сравнению с контрольными животными.
ПРИМЕР 14
ВЛИЯНИЕ РОДСТВЕННОГО УРОКОРТИНУ ПЕПТИДА ЧЕЛОВЕКА НА ТЕМПЕРАТУРУ ТЕЛА
Исследовали влияние родственного урокортину пептида человека на температуру тела крыс. Канюли для инъекции родственного урокортину пептида человека хирургическим путем вводили в правый латеральный желудочек. Телеметрические устройства для непрерывных незаметных анализов температуры тела имплантировали внутрибрюшинно. Животным давали период выздоровления после хирургической операции, равный семи дням. В это время животных ежедневно брали руками, чтобы животные привыкли к процедуре инъекции. В день инъекции исходную температуру регистрировали в течение трех часов. В 6:00 вечера (начало выключения света) животные получали инъекцию либо 5 мкл физиологического раствора, либо 5 мкл родственного урокортину пептида человека в физиологическом растворе в концентрации 1 мкг/мкл. Температуру тела контролировали каждые пять минут в течение двенадцати часов. Как видно на фигуре 14, животные, которым инъецировали родственный урокортину пептид человека, имели более низкие температуры тела сразу и через семь часов после инъекции.
ПРИМЕР 15
ВЛИЯНИЕ РОДСТВЕННОГО УРОКОРТИНУ ПЕПТИДА ЧЕЛОВЕКА НА АППЕТИТ
Также исследовали влияние родственного урокортину пептида человека на аппетит у крыс. Канюли для инъекции родственного урокортину пептида человека хирургическим путем вводили в правый латеральный желудочек и животным давали возможность выздороветь в течение семи дней. В это время животных ежедневно брали руками, чтобы животные привыкли к процедуре инъекции. В день инъекции животные получали инъекцию либо 5 мкл физиологического раствора, либо 5 мкл родственного урокортину пептида человека в физиологическом растворе в концентрации 1 мкг/мкл. Количество съеденной каждым животным пищи регистрировали каждый час в течение шести часов и на четырнадцатый час.
Общее количество потребленной пищи в течение экспериментов показано для каждого периода времени на фигуре 15А. Животные, которым инъецировали родственный урокортину пептид человека, съедали значительно меньше пищи, чем контрольные животные. На фигуре 15В суммировано количество пищи, потребленное в каждый период времени. Животные, обработанные hURP, гораздо меньше съедали пищи во время первого и третьего часа после инъекции, а также во время последних восьми часов эксперимента.
ПРИМЕР 16
ПРИГОДНЫЕ МОДИФИКАЦИИ И ПРОИЗВОДНЫЕ УРОКОРТИНА II И РОДСТВЕННОГО УРОКОРТИНУ ПЕПТИДА ЧЕЛОВЕКА
Заявленные здесь урокортин II и родственный урокортину пептид человека наиболее вероятно представляют собой формы прогормонов указанных белков. Предполагается, что в активацию гормонов будет вовлечен протеолитический процессинг и другой тип модификации белков, такой как модификация, результатом которой являются неамидированные формы белков.
Предыдущие исследования лигандов для других рецепторов CRF показали, что в указанных лигандах можно сделать ряд аминокислотных замен без потери способности к связыванию с рецепторами биологической активности лигандов. Ряд предыдущих исследований с урокортином показали, что бесспорно допустимы одна, две или даже три замены. В некоторых случаях модификации урокортина в результате приводили к белку с более желательными фармакологическими свойствами. Так как урокортин II и родственный урокортину пептид человека являются небольшими белками, такую модификацию легче всего ввести способами пептидного синтеза, хорошо известными специалистам в данной области. Указанные способы включают способ твердофазного синтеза, частичного твердофазного синтеза, конденсацию фрагментов и классическое связывание в растворе. Указанные способы особенно предпочтительны в том случае, если необходимо включить нестандартные аминокислоты в урокортин II или родственный урокортину пептид человека. Альтернативно, если модификации полностью состоят из природных аминокислот, можно использовать технологию рекомбинантной ДНК для мутагенеза и последующей экспрессии модифицированного урокортина II и родственного урокортину пептида человека.
В родственном урокортину пептиде человека отсутствует остаток тирозина. Так как остатки тирозина используют для радиоиодирования белков, одной из возможных модификаций родственного урокортину пептида человека может быть замена тирозином другой аминокислоты в белке. Ранее описано присоединение последовательности, состоящей из Tyr-Gly к N-концу урокортина. Полученный в результате белок сохраняет способность связывать рецептор CRF и биоактивность, тем не менее, может быть пригоден для радиоиодирования белка. Также можно сконструировать другие удлинения N-конца белка согласно данному изобретению для мечения и других целей.
Обнаружено, что делеция первых семи - десяти остатков урокортина приводит к формированию эффективных антагонистов урокортина. Указанные белки способны связываться с рецепторами CRF, но в значительной степени не стимулируют или не активируют рецепторы. Предполагается, что делеция до пяти аминокислот из урокортина II или родственного урокортину пептида человека может также в результате привести к получению эффективных антагонистов. Также можно создать антагонисты из других фрагментов урокортина II и родственного урокортину пептида человека. Указанные антагонисты могут быть эффективными в повышении уровней эндогенных пептидов, в норме клиренс которых осуществляется белком, связывающим CRF. При связывании с CRF-связывающим белком и блокировании связывания CRF, урокортина, урокортина II и родственного урокортину пептида человека с тем же самым белком эффективные концентрации эндогенных CRF, Ucn и Ucn II увеличиваются in vivo. Такие антагонисты можно вводить совместно с другими агонистами или антагонистами CRF, Ucn, Ucn II или URP для усиления их воздействия.
Всесторонний анализ других белков, связывающих рецептор CRF, показал, что замена нормальных аминокислот D-изомерами аминокислот или циклизованными аминокислотами в результате приводит к повышенной аффинности по отношению к CRF-рецепторам. В частности, особенно пригодной заменой является замена остатка изолейцина, соответствующего положению 9 SEQ ID NO: 3 или SEQ ID NO: 11, изомерной «D-формой» аминокислоты, предпочтительно D-изолейцином, D-фенилаланином и D-лейцином. Подобным образом, остаток глутаминовой кислоты, соответствующий положению 17 SEQ ID NO: 3 или SEQ ID NO: 11, можно заменить D-глутаминовой кислотой. Циклизованные аминокислоты можно образовать посредством химических связей между боковыми цепями двух или большего количества остатков. Например, близлежащие остатки глутаминовой кислоты и лизина могут реагировать с образованием амидной связи, давая кольцо лактама. Замену нестандартными аминокислотами, такими как Сα-метилированный лейцин, Сα-метилированный аланин, N-im-бензилгистидин, 4-гидроксипролин, 5-гидроксилизин, 3-метилгистидин, гомосерин и орнитин, также можно использовать для создания агонистов или антагонистов родственного урокортину пептида человека.
Заявленные здесь модификации урокортина II и родственного урокортину пептида человека предназначены для иллюстрации возможной модификации, которую можно осуществить, и никоим образом не означают ограничения изобретения.
ОБСУЖДЕНИЕ
Поиск гомологии по всему геному использовали для того, чтобы идентифицировать новых представителей нейропептидов семейства CRF. Один из новых лигандов, Ucn II, избирательно связывается с CRF-R2, экспрессируется в дискретных областях ЦНС крыс и активирует центральные нейроны, вовлеченные в обработку висцеральной сенсорной информации и в модулирование автономного потока. Кроме того, Ucn II ингибирует потребление пищи без какого-либо влияния на общую моторную активность.
Кроме мышиного пептида, который проявляет структурные характеристики, характеристики связывания, активности и экспрессии, ожидаемые для представителя семейства CRF, идентифицировали URP человека (на основании доступной опубликованной последовательности EST), который на 80% идентичен последовательности мыши на уровне нуклеотидов. Однако явное важное отличие пептида человека состоит в отсутствии какого-либо видимого сайта протеолитического расщепления, который бы обеспечивал С-концевой процессинг гомолога человека. Остается необходимость в определении того, может ли и как создается какой-либо гомологичный пептид человека из данного белка. Тем не менее, тогда как Ucn связывается с высокой аффинностью с обоими рецепторами, CRF-R1 и CRF-R2, и эффективно передает сигналы через эти рецепторы (14-16), Ucn II мыши и URP человека проявляют высокую степень избирательности к CRF-R2 по указанным критериям, и, несомненно, будут иметь значение для раздельных функций, опосредованных двумя типами рецепторов. На фигуре 16 показана модель того, как урокортин II действует на CRF-R1 и CRF-R2. Ucn II с высокой аффинностью связывается с CRF-R2, но не с CRF-R1. Урокортин связывается с обоими рецепторами, тогда как CRF связывается с высокой аффинностью с CRF-R1, но не с CRF-R2.
мРНК Ucn II демонстрирует ограниченное распределение в подкорке в головном мозге грызунов, которое является уникальным, несмотря на видимое частичное перекрывание с распределением CRF (паравентрикулярное ядро; например, ссылка 31) и Ucn (ствол головного мозга и моторные ядра спинного мозга; например, ссылка 18). Особый интерес представляет тот факт, что транскрипт экспрессируется в группе клеток, вовлеченных в связанные со стрессом физиологические и поведенческие функции (смотри ссылку 13). Группа включает голубоватое место (locus coeruleus), которое имеет обширные проекции в коре головного мозга, и считается вовлеченным в создание уровней возбуждения и тревожности (например, 32), паравентрикулярное ядро, в котором размещены множественные популяции важных нейросекреторных нейронов, и которое имеет проекцию в ЦНС, модулируя сенсорный и моторный поток в центральном автономном контуре (например, 33), и дугообразное ядро, которое охарактеризовали как основной компонент широкой системы, способствующей регуляции потребления пищи и энергетического баланса (например, 34). Хотя еще отсутствуют анатомические и функциональные данные для того, чтобы определить место новых пептидов в указанных контекстах, центральная система Ucn II обладает возможностями для участия в связанных со стрессом функциях, долгое время считавшихся компетенцией более широкой центральной сети CRF. Противоположная ситуация существует для Ucn, основное место экспрессии которого в клетках головного мозга, ядро Эдингера-Вестфаля, проявляет очень ограниченные возможности в этом отношении, в значительной степени в силу малого числа документально подтвержденных проекций в передний мозг (16, 18).
С точки зрения параметров связывания и активности неспособность картины активации клеток, вызванной центральным Ucn II, близко имитировать распределение CRF-R2 была неожиданной. Недавнее исследование, в котором сравнили распределение экспрессии Fos, индуцированной icv-введением CRF или UCN, документально подтвердило, что картины активации примерно соответствуют аффинностям связывания указанных пептидов с CRF-R, кодируемыми двумя известными генами (35). То есть CRF в дозах, сходных с применяемыми здесь дозами, активировал места экспрессии CRF-R1 с высоким преимуществом, тогда как UCN вызывал индукцию Fos главным образом в подгруппах групп клеток, которые экспрессируют каждый из рецепторов. Однако оба пептида, кроме того, вовлекали точно такой же набор центральных автономных структур, которые выявлены в данной работе, как преимущественные места индуцированных Ucn II активационных ответов в головном мозге крыс. Это важно по той причине, что элементы центральной автономной системы входят в число наилучшим образом документально подтвержденных мест, в которых могут действовать CRF-подобные пептиды, вызывая связанные со стрессом автономные и поведенческие ответы. Указанные данные, вероятно, свидетельствуют о том, что активация рецептора типа 2, так же как типа 1, способна вовлекать данную систему, хотя основа этого неизвестна. Среди основных точек в центральной автономной сети только парабрахиальное ядро (R1) и NTS (R2) идентифицированы как места экспрессии CRF-R (27, 28, 35), и еще предстоит определить, достаточна ли опосредованная рецептором активация одной из них или обеих указанных точек для того, чтобы вовлечь систему в целом. Важно отметить, что системные инъекции синтетического Ucn II не вызывали сравнительно сильных активационных ответов в группе центральных автономных клеток в тех же пределах доз, которые использовали для исследований в случае icv-инъекций. Это важный элемент контроля, так как активация периферического CRF-R2 может давать заметное и стойкое снижение кровяного давления (16, 17), и провоцирование заметной гипотензии способно активировать те же самые центральные автономные структуры, которые отвечают на центральное введение Ucn II (36, 37).
Начальная характеристика эффектов icv-введенного Ucn II на потребление пищи и активность дополняет недавние попытки разрешить вопрос об отдельной роли индивидуальных CRF-R в связанном со стрессом поведении. Например, в то время как мыши, несущие нуль-мутации одного из двух рецепторов, проявляют нормальный базовый уровень потребления пищи, показано, что животные, дефицитные по CRF-R1, невосприимчивы к анорексическим эффектам UCN во время периода сразу же после инъекции, но не в более поздних временных точках, тогда как для мышей, мутантных по CRF-R2 справедливо обратное (11, 38, 39). Это было признано свидетельством того, что ранняя и поздняя фазы опосредованного Ucn подавления потребления пищи могут представлять собой события, опосредованные соответственно CRF-R1 и CRF-R2. Используя другой принцип (свободное кормление в ночное время, а не индуцированное лишением пищи переедание), получили данные, поддерживающие такой аналитический вывод, так как R2-специфичный лиганд достоверно не подавлял потребление пищи в ранних временных точках, но подавлял позже 6 ч после инъекции.
Показатели измерения моторной активности также подтверждали разделение CRF-R при участии в данном параметре. В соответствии с полученными недавно данными на нокаутированных мышах, свидетельствующих о том, что локомоторная активация является событием, опосредованным CRF-R1 (40), было обнаружено, что избирательный по отношению к R1 агонист, CRF, в значительной степени увеличивал общую моторную активность, тогда как введение UCN II не вызывало увеличения. Интересно, что обработка UCN, который с высокой аффинностью связывается с обоими рецепторами, приводила к незначительному отклонению в сторону повышенной активности, при этом значения были достоверно ниже, чем значения, наблюдаемые в ответ на CRF. Это примерно согласуется с растущей совокупностью доказательств, подтверждая функциональный антагонизм между двумя известными типами рецепторов. Тогда как CRF-R1-дефицитные мыши проявляют пониженные эндокринные ответы и ответы, подобные тревожному состоянию, на стресс (41), линии, мутантные по CRF-R2, демонстрируют увеличения указанных параметров (11, 39, 42), свидетельствуя о том, что основная активация CRF-R2 может играть роль в противостоянии стресс-ответам, управляемым CRF-R1.
Идентификация эндогенного избирательного по отношению к CRF-R2 лиганда позволит более подробно проанализировать роли отдельных родственных CRF сигнальных молекул в связанных со стрессом физиологических и поведенческих функциях. Центральная экспрессия мРНК Ucn II совпадала с группами клеток, которые отвечают на центральное введение пептида, и подтвердила поведенческие ответы, которые согласуются с ранее выдвинутой гипотезой о последствиях активации CRF-R2. Дальнейшее выяснение места указанного пептида в биологии стресса потребует выявления центральных проекций клеток, содержащих Ucn II, и идентификации факторов и условий, которые регулируют экспрессию гена и высвобождение пептида.
В данном описании цитированы следующие ссылки.


Любые патенты или публикации, упоминаемые в данном описании, являются показателем уровней специалистов в данной области, к которым данное изобретение имеет отношение. Указанные патенты и публикации включены здесь в виде ссылок в таком же объеме, в каком они были бы включены, если бы было специально и отдельно указано, что каждая отдельная публикация включена в виде ссылки.
Специалист в данной области легко поймет, что данное изобретение хорошо адаптировано для выполнения целей и получения результатов и преимуществ указанных, а также свойственных изобретению. Представленные примеры наряду с описанными здесь способами, процедурами, обработками, молекулами и конкретными соединениями в настоящее время являются образцами предпочтительных вариантов, являются иллюстративными и не подразумевают ограничений рамок изобретения. Специалисты в данной области могут осуществить изменения в изобретении и другие применения, которые являются составной частью сущности изобретения, определенной рамками формулы изобретения.




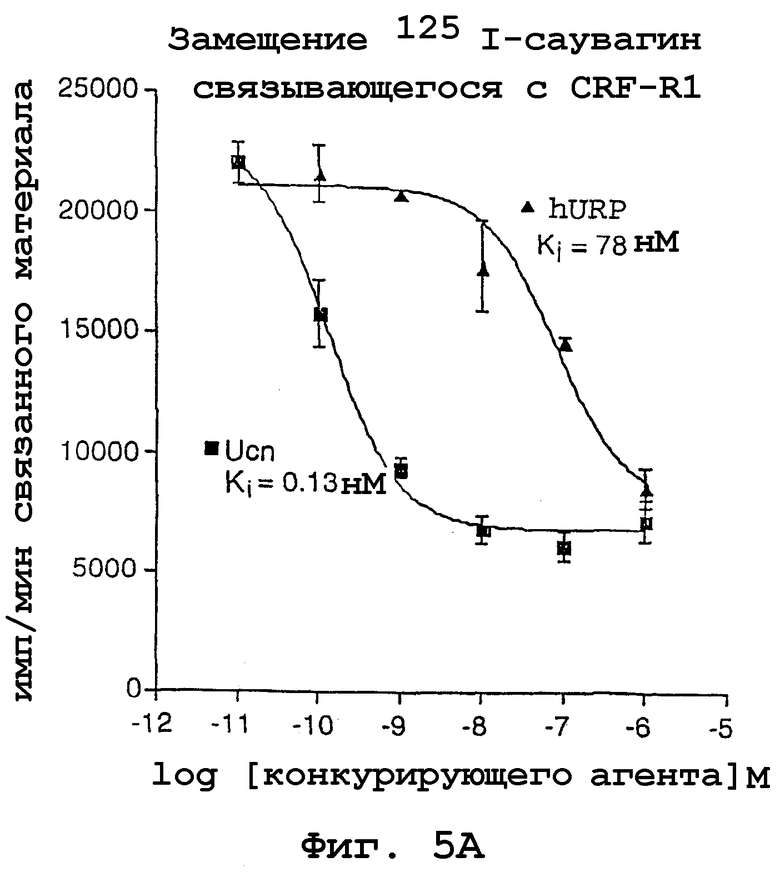
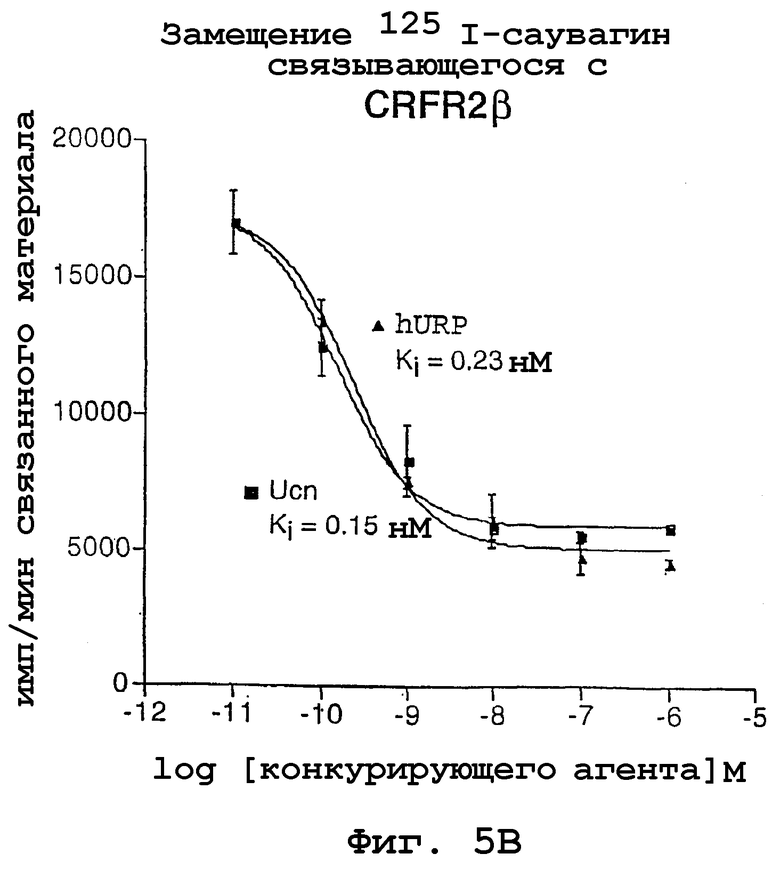
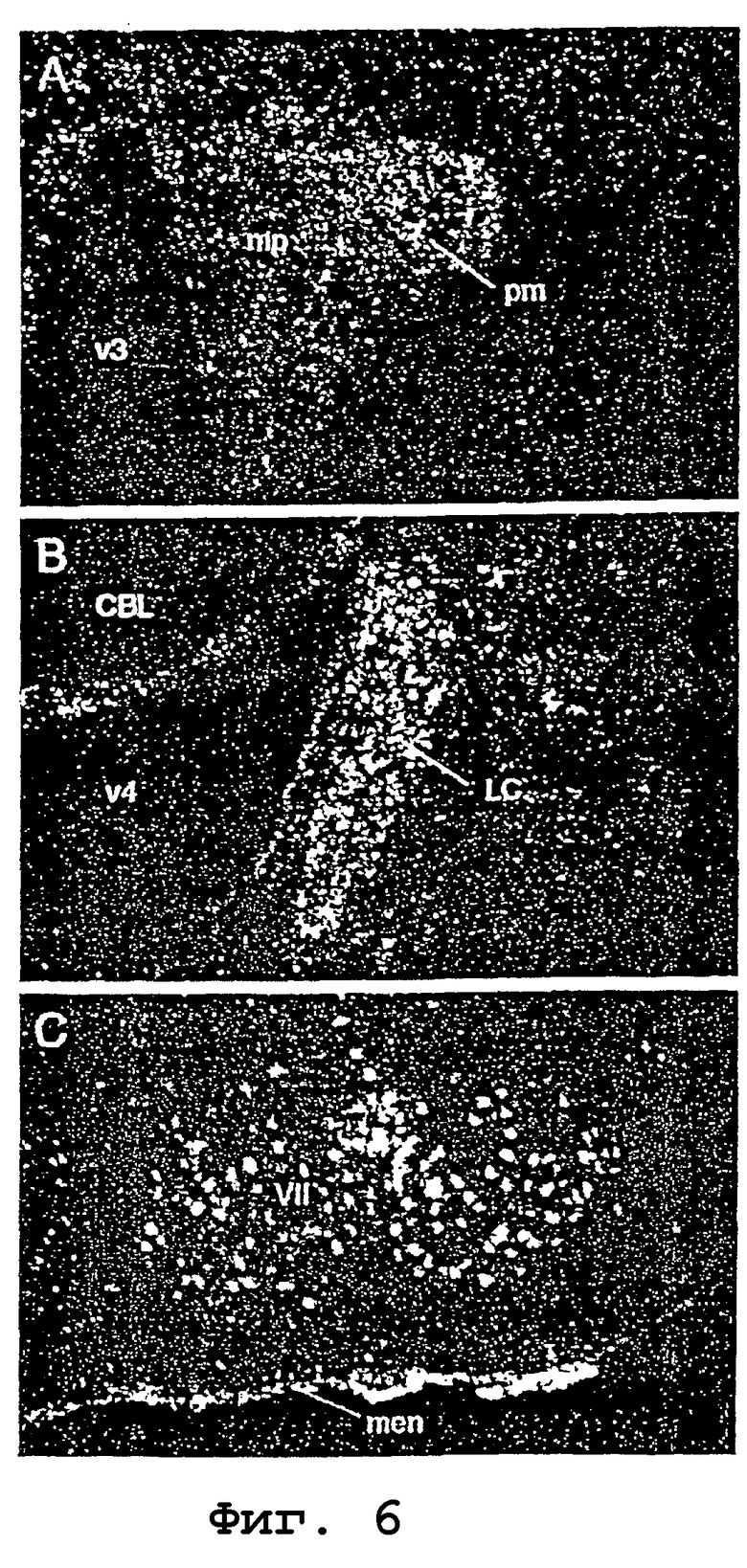
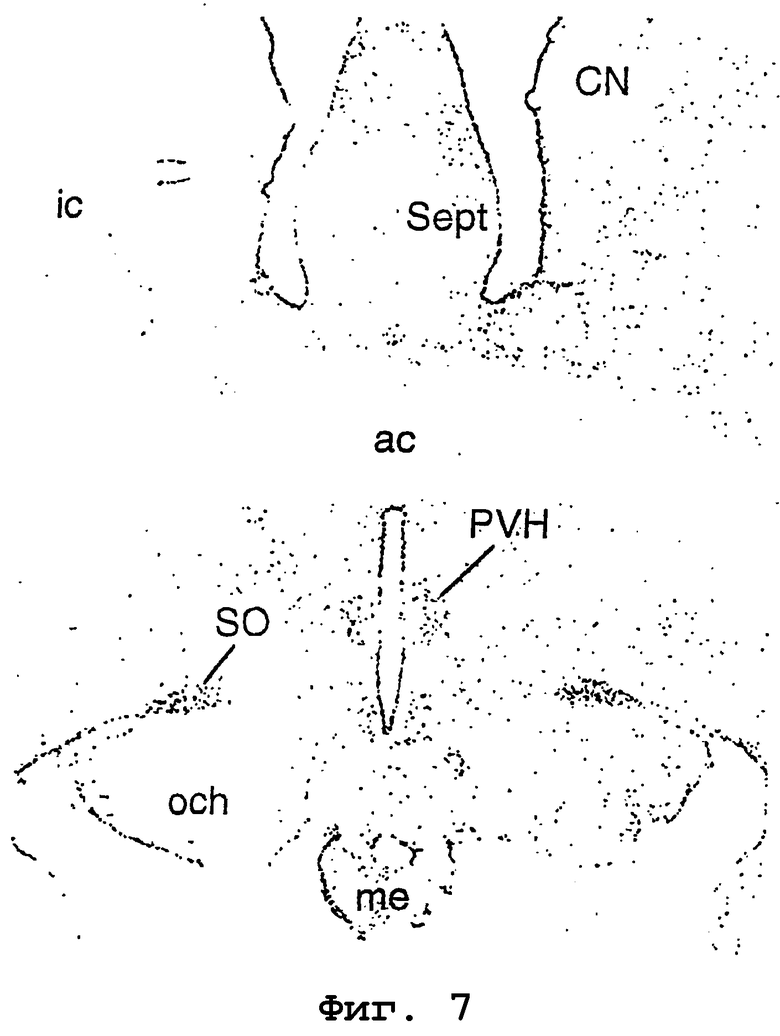
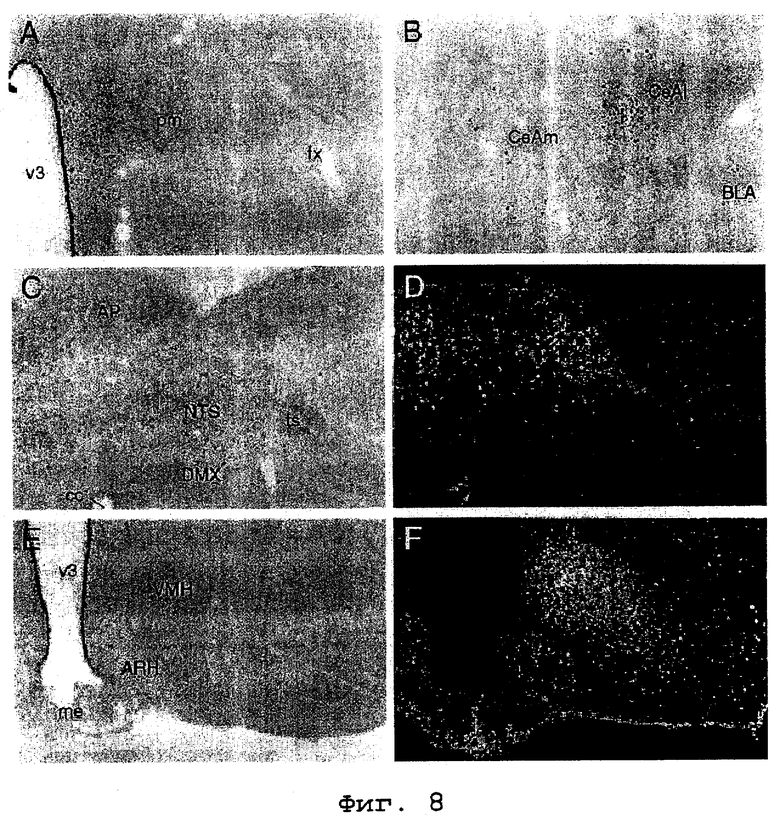
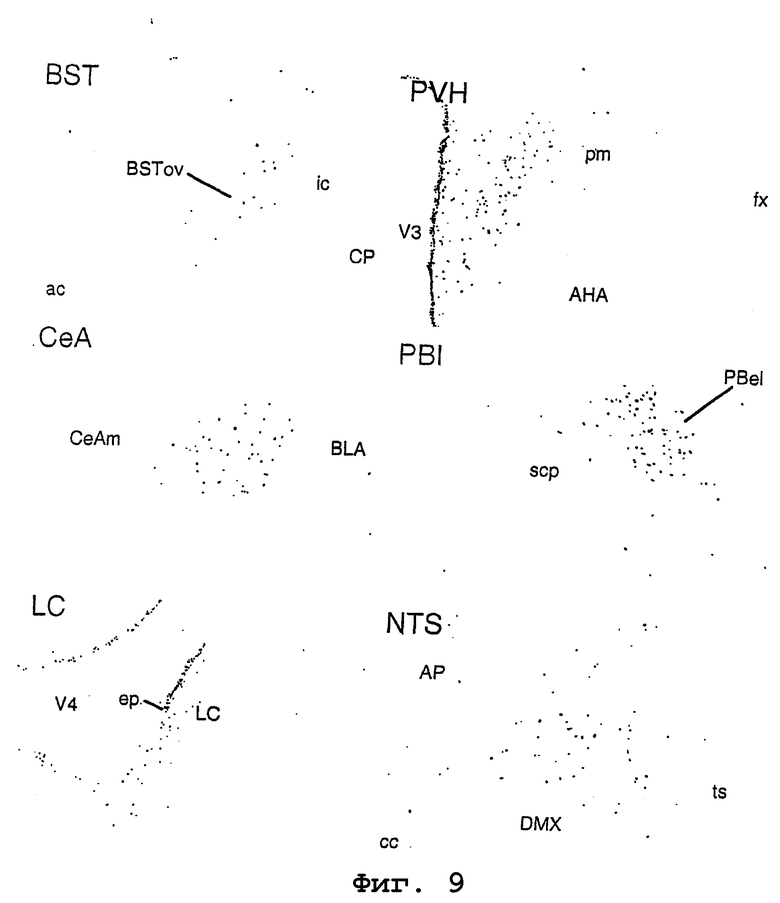
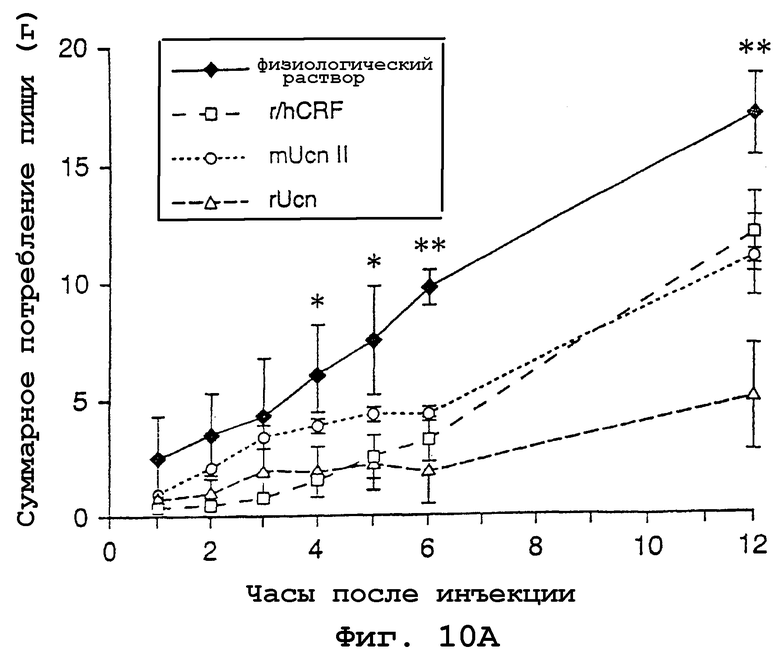
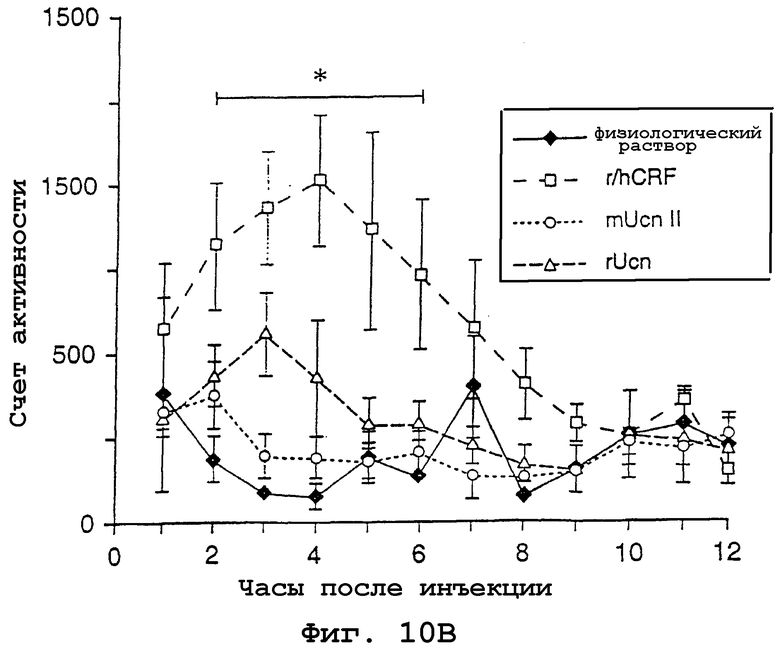
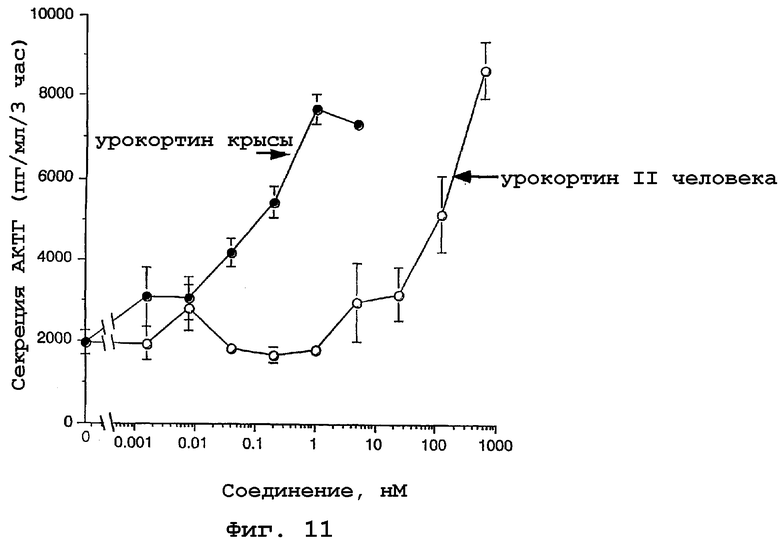
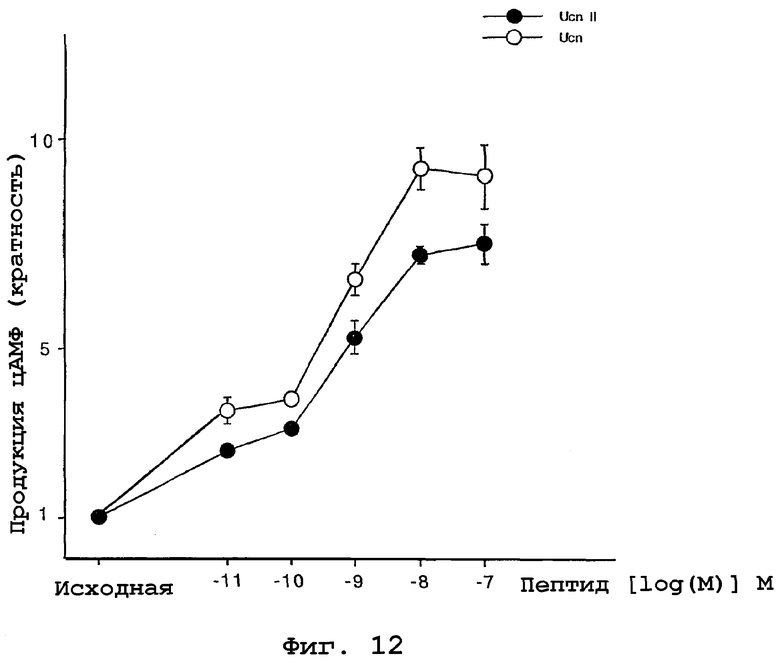
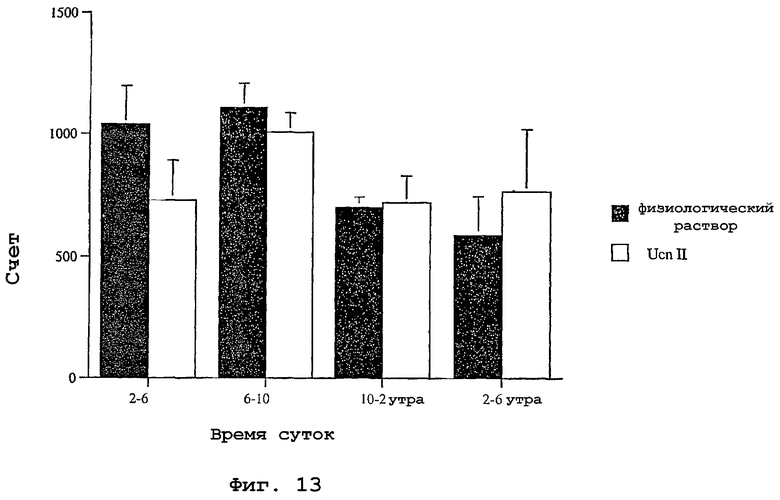
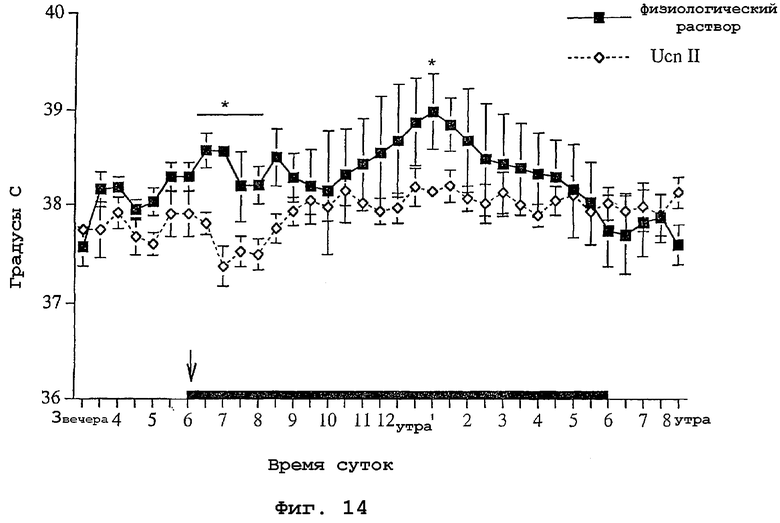
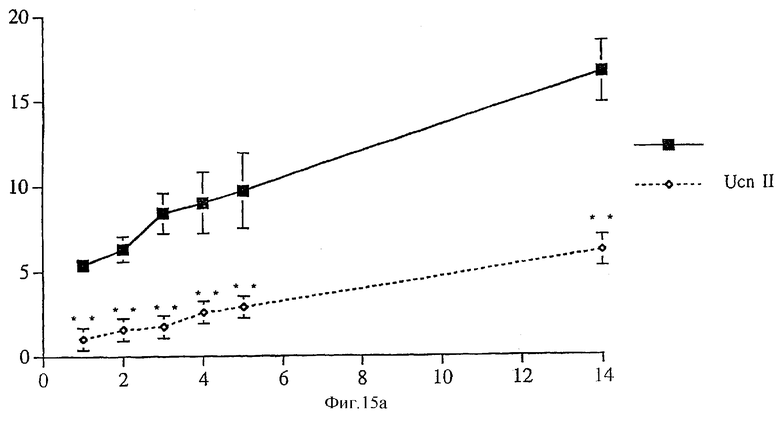
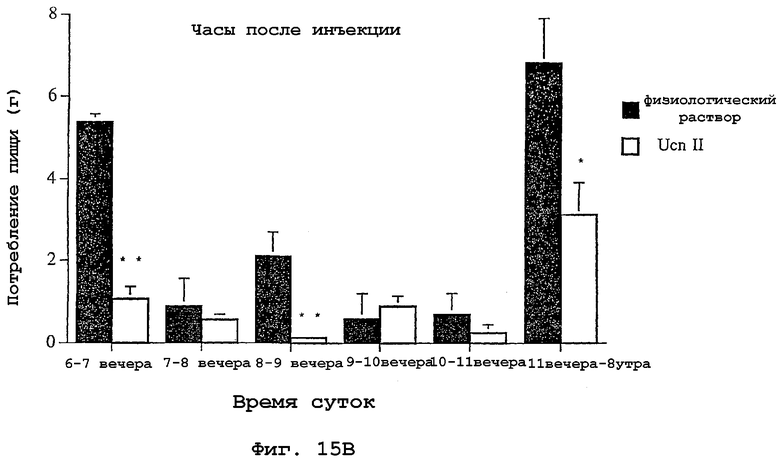
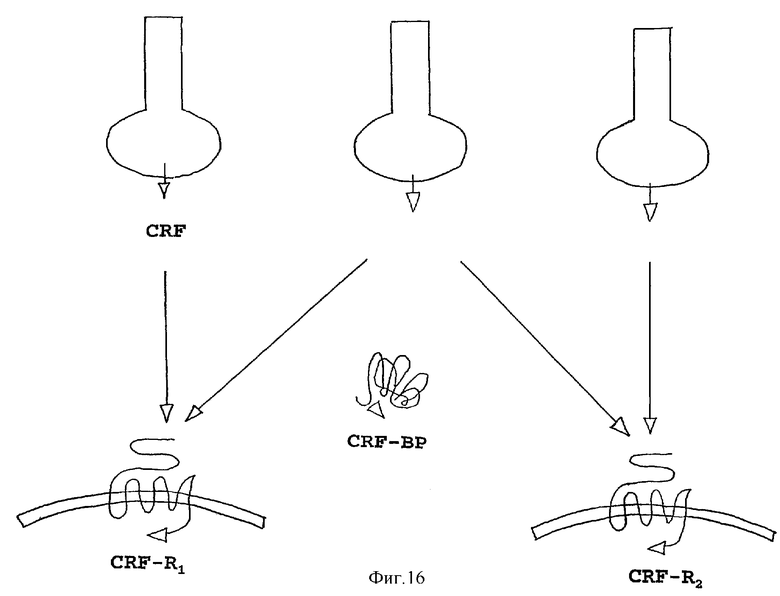
Изобретение относится к генной инженерии и может быть использовано в медицине. Выделена ДНК, кодирующая родственный урокортину пептид человека и пептид, названный урокортином II. Эти пептиды родственны кортикотропин-рилизинг фактору и также вовлечены в механизмы инициации гипофизарно-надпочечниковых ответов на стресс. Фармацевтическая композиция, содержащая такой пептид в сочетании с приемлемым носителем, может быть применена для лечения патофизиологических состояний, таких как повышенная температура тела, нарушение аппетита, застойная сердечная недостаточность, стресс, тревожное состояние и нежелательно низкие уровни АКТГ. 8 н. и 25 з.п. ф-лы, 20 ил., 2 табл.
(a) выделенную и очищенную ДНК, которая кодирует родственный урокортину пептид человека с последовательностью SEQ ID No: 2-4;
(b) выделенную и очищенную ДНК, которая гибридизируется в жестких условиях с антисмысловой последовательностью, комплементарной выделенной ДНК, указанной выше в (а), где жесткие условия определяются промывкой мембраны при высокой температуре и низкой концентрации соли, что функционально эквивалентно 0,1 × SSC при 65°С, где указанная ДНК кодирует родственный урокортину пептид человека с последовательностью SEQ ID No: 2-4; и
(c) выделенную и очищенную ДНК, отличающуюся по последовательности кодонов от указанных выше в (а) и (b) выделенных ДНК в результате вырожденности генетического кода и которая кодирует родственный урокортину пептид человека с последовательностью SEQ ID No: 2-4.
(a) родственный урокортину пептид человека с последовательностью SEQ ID No:2-4 и
(b) родственный урокортину пептид человека, кодируемый ДНК, выбранной из группы, включающей:
(i) выделенную и очищенную ДНК, которая кодирует родственный урокортину пептид человека с последовательностью SEQ ID No:2-4;
(ii) выделенную и очищенную ДНК, которая гибридизируется в жестких условиях с антисмысловой последовательностью, комплементарной выделенной ДНК, указанной выше в (i), где жесткие условия определяются промывкой мембраны при высокой температуре и низкой концентрации соли, что функционально эквивалентно 0,1 × SSC при 65°С, где указанная ДНК кодирует родственный урокортину пептид человека с последовательностью SEQ ID No:2-4; и
(iii) выделенную и очищенную ДНК, отличную по последовательности кодонов от указанных выше в (i) и (ii) выделенных ДНК вследствие вырожденности генетического кода и которая кодирует родственный урокортину пептид человека с последовательностью SEQ ID No:2-4.
(a) добавление последовательности, состоящей из Tyr-Gly, к N-концу указанного белка;
(b) N-концевую делецию, где указанная делеция относится к делециям аминокислот, выбранным из группы, включающей первую аминокислоту, первую и вторую аминокислоты, аминокислоты с первой по третью, аминокислоты с первой по четвертую и аминокислоты с первой по пятую;
(c) замену изолейцинового остатка, соответствующего положению 9 SEQ ID No:3 или SEQ ID No:4, на аминокислоту с D-изомерной формой;
(d) замену глутаминовой кислоты, соответствующей положению 17 SEQ ID No:3 или SEQ ID No:4, на D-глутаминовую кислоту и
(e) ацилирование N-конца указанного белка, по существу, обладающего активностью родственного урокортину пептида человека.
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |

