Область изобретения
Настоящее изобретение относится к соединению или рецептору, связывающему с Helicobacter pylori, и его применению, например, в фармацевтических и пищевых композициях для лечения состояний, обусловленных присутствием Helicobacter pylori. Данное изобретение также направлено на применение рецептора для диагностики Helicobacter pylori.
Предпосылки изобретения
Helicobacter pylori участвуют в развитии нескольких заболеваний желудочно-кишечного тракта, включая хронический гастрит, заболевание желудка, связанное с приемом нестероидных противовоспалительных средств (НСПВС), двенадцатиперстной кишки и язвы желудка, лимфому из лимфоидной ткани слизистой желудка и аденокарциному желудка (Axon, 1993; Blaser, 1992; DeCross and Marshall, 1993; Dooley, 1993; Dunn et al., 1997; Lin et al., 1993; Nomura and Stemmermann, 1993; Parsonnet et al., 1994; Sung et al., 2000; Wotherspoon et al., 1993). Заболевания, совсем или частично не относящиеся к желудочно-кишечным, включают синдром внезапной младенческой смерти (Kerr et al., 2000 и патент США № 6083756), аутоиммунные заболевания, такие как аутоиммунный гастрит, и пернициозную анемию (Appelmelk et al., 1998; Chmiela et al., 1998; Clayes et al., 1998; Jassel et al., 1999; Steininger et al., 1998) и некоторые заболевания кожи (Rebora et al., 1995), заболевания поджелудочной железы (Correa et al., 1990), заболевания печени, включая аденокарциному (Nilsson et al., 2000; Avenaud et al., 2000), и заболевания сердца, такие как атеросклероз (Farsak et al., 2000). Имеется обзор по многочисленным заболеваниям, вызванным или связанным с Helicobacter pylori (Pakodi et al., 2000). Наибольший интерес в отношении бактериальной колонизации и инфицирования представляет механизм(ы) адгезии бактерий на поверхность эпителиальных клеток слизистой желудка.
Сообщалось, что гликоконъюгаты как на основе липидов, так и на основе белков, служат в качестве рецепторов для связывания данных микроорганизмов, например, в виде сиалилированных гликоконъюгатов (Evans et al., 1988), сульфатида и GM3 (Saitoh et al., 1991), Leb-детерминант (Boren et al., 1993), полигликозилкерамидов (Miller-Podraza et al., 1996; 1997a), лактозилкерамида (Engström et al., 1998) и ганглиотетраозилкерамида (Lingwood et al., 1992; Engström et al., 1998). Другие потенциальные рецепторы Helicobacter pylori включают полисахарид гепарансульфат (Ascensio et al., 1993), а также фосфолипид фосфатидилэтаноламин (Lingwood et al., 1992).
В патентах США, Zopf et al.: № 5883079 (март 1999 г.), № 5753630 (май 1998 г.) и № 5514660 (май 1996 г.) описываются Neu5Acα3Gal-содержащие соединения в качестве ингибиторов адгезии Helicobacter pylori. Молекула сиалил-лактозы ингибирует связывание Helicobacter pylori с линиями человеческих клеток желудочно-кишечного тракта (Simon et al., 1999) и также эффективна на модели инфекции на макаках-резусах (Mysore et al., 2000). Соединение проходит клинические испытания.
В патенте США, Krivan et al. № 5446681 (ноябрь 1995 г.) описываются конъюгаты рецептора бактерий и антибиотика, включающие асиалоганглиозид, связанный с антибиотиком пенициллином. В особенности заявляется лечение инфекций, вызванных Helicobacter pylori, с помощью конъюгата амоксиллин-асиало-GM1. Описанные в изобретении олигосахаридные последовательности/гликолипиды не относятся к ганглиопроизводным гликолипидов.
В патентах США, Krivan et al.: № 5386027 (январь 1995 г.) и № 5217715 (июнь 1993 г.) описано применение олигосахаридных последовательностей или гликолипидов для ингибирования нескольких патогенных бактерий, однако, обсуждаемая здесь специфичность связывания там не упомянута, и Helicobacter pylori не входит в перечень рассмотренных или заявленных бактерий.
Сахаридная последовательность GlcNAcβ3Gal была описана в качестве рецептора Streptococcus (Andersson et al., 1986). У некоторых бактерий специфичности связывания могут перекрываться, и невозможно предсказать характер связывания даже у очень близких бактериальных адгезинов. В случае Helicobacter pylori сахаридные связывающие молекулы неизвестны, за исключением связывающего белка Lewis b.
Краткое описание изобретения
Настоящее изобретение относится к применению соединения или рецептора, связывающегося с Helicobacter pylori, включающего олигосахаридную последовательность.
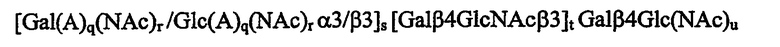
в которой каждый из g, r, s, t и u независимо равен 0 или 1,
так, что когда t=0 и u=0, то тогда олигосахаридная последовательность связана с поливалентным носителем или находится в виде свободного олигосахарида в высокой концентрации, и аналоги или производные указанной олигосахаридной последовательности, обладающие активностью связывания с Helicobacter pylori, для производства композиции, обладающей активностью связывать или ингибировать Helicobacter pylori.
Среди объектов изобретения находится применение связывающихся с Helicobacter pylori олигосахаридных последовательностей, описанных в изобретении, в качестве лекарственного препарата и их применение для производства фармацевтической композиции, в частности, для лечения любого состояния, обусловленного присутствием Helicobacter pylori.
Настоящее изобретение также относится к способам лечения состояний, обусловленных присутствием Helicobacter pylori. Изобретение также направлено на применение рецептора(ов), описанного в изобретении, в качестве связывающего или ингибирующего Helicobacter pylori соединения для диагностики Helicobacter pylori.
Другим объектом изобретения является обеспечение соединений, фармацевтических композиций и пищевых добавок или композиций, содержащих связывающую Helicobacter pylori олигосахаридную последовательность(и).
Другими объектами изобретения являются применение вышеуказанных связывающих Helicobacter pylori соединений для идентификации бактериального адгезина, типирование Helicobacter pylori и анализы связывания Helicobacter pylori.
Еще одним объектом изобретения является применение вышеуказанных связывающих Helicobacter pylori соединений для производства вакцины против Helicobacter pylori.Краткое описание фиг.1А и 1В. EI*
Электроионизация.
/MC перметилированных олигосахаридов, полученных в результате расщепления гексагликозилкерамида под действием эндогликокерамидазы. Хроматограмма, полученная при анализе газовой хроматографией олигосахаридов (вверху) и EI/MC-спектры соответственно пиков А и В (внизу).
Фиг.2А и 2В. Бомбардировка быстрыми атомамиFAB*
Бомбардировка быстрыми атомами.*-масс-спектры отрицательных ионов гекса-(2А) и пентагликозилкерамида (2В).
Фиг.3А и 3В. Протонные ЯМР-спектры, показывающие аномерную область гликолипида из шести сахаров (3А) и гликолипида из пяти сахаров (3В). Спектры были получены в течение ночи для хорошего соотношения сигнал-шум в целях детектирования минорного компонента типа 1.
Фиг.4А, 4В и 4С. Ферментативное расщепление гликосфинголипидов из тимуса кролика. Пластинки для тонкослойной хроматографии в силикагеле развивали в системе C/M/H2O (60:35:8, объемные соотношения). 4А и 4В, пластинки проявляли 4-метоксибензальдегидом. 4С, авторадиограмма после нанесения 35S-меченных Helicobacter pylori. 1, гептагликозилкерамид (структура 1, таблица I); 2, десиалилированный гептагликозилкерамид (полученный после обработки кислотой); 3, десиалилированный гептагликозилкерамид, обработанный β4-галактозидазой; 4, гептагликозилкерамид, обработанный сиалидазой и β4-галактозидазой; 5, стандарты гликосфинголипидов из эритроцитов человека (лактозилкерамид, тригексозилкерамид и глобозид); 6, десиалилированный гептагликозилкерамид, обработанный β4-галактозидазой и β-гексозаминидазой; 7, гептагликозилкерамид, обработанный сиалидазой, β4-галактозидазой и β-гексозаминидазой.
Фиг.5А и 5В. ТСХ продуктов, полученных после частичного кислотного гидролиза гептагликозилкерамида из тимуса кролика (структура 1, таблица I). Развивающий растворитель был такой же, как для фиг.4А, 4В и 4С. 5А, проявленная 4-метоксибензальдегидом пластинка; 5В, авторадиограмма после нанесения 35S-меченных Helicobacter pylori. 1, гептагликозилкерамид; 2, десиалилированный гептагликозилкерамид (обработка кислотой); 3, пентагликозилкерамид; 4, гидролизат; 5, стандарты гликосфинголипидов (как для фиг.4А, 4В и 4С).
Фиг.6А и 6В. Серийные разведения гликосфинголипидов. Связывающую активность на пластинках для ТСХ определяли методом нанесения бактерий. Развивающий растворитель для ТСХ был такой же, как для фиг.4А, 4В и 4С. Различные гликолипиды наносили на пластинки в эквимолярных количествах. Количественное определение гликолипидов проводили по содержанию гексозы. 6А, гекса- и пентагликозилкерамиды (структуры 2 и 3, таблица I); 6В, пента- и тетрагликозилкерамиды (структуры 4 и 5, таблица I). Количества гликолипидов (выраженные в пмолях) были следующими: 1,1280 (каждого); 2,640; 3,320; 4,160; 5,80; 6,40; 7,20 пмоль (каждого).
Фиг.7А и 7В. Тонкослойная хроматограмма разделенных гликосфинголипидов при проявлении 4-метоксибензальдегидом (7А) и авторадиограмма после связывания с меченными радиоактивным изотопом Helicobacter pylori штамма 032 (7В). Гликосфинголипиды разделяли на пластинках с силикагелем 60 для ТСХ на алюминиевой подложке HPTLC (Merck) с использованием смеси хлороформ/метанол/вода 60:35:8 (объемные соотношения) в качестве системы растворителей. Анализ связывания проводили, как описано в разделе «Материалы и методы». Авторадиографию проводили в течение 72 ч. Полосы содержали:
полоса 1) Galβ4GlcNAcβ3Galβ4Glcβ1Cer
(неолактотетраозилкерамид); 4 мкг;
полоса 2) Galα3Galβ4GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид В5), 4 мкг;
полоса 3) Galα3Galβ4GlcNH2β3Galβ4Glcβ1Cer, 4 мкг;
полоса 4) Galα3(Fucα2)Galβ4GlcNAcβ3Galβ4Glcβ1Cer
(гликосфинголипид типа 2 В6), 4 мкг;
полоса 5) GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer, 4 мкг;
полоса 6) Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ1Cer, 4 мкг;
полоса 7) GalNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer
(х2-гликосфинголипид), 4 мкг;
полоса 8) NeuAcα3GalNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer (NeuAc-x2), 4 мкг;
полоса 9) Fucα2Galβ4GlcNAcβ3Galβ4Glcβ2Cer (гликосфинголипид типа 2 H5), 4 мкг;
полоса 10) NeuAcα3Galβ4GlcNAcβ3Galβ4Glcβ1Cer
(сиалилнеолактотетраозилкерамид), 4 мкг.
Источники гликосфинголипидов являются теми же, что представлены в таблице 2.
Фиг.8А, 8В, 8С и 8D.Рассчитанные конформации с минимальной энергией трех гликосфинголипидов, которые связывают Helicobacter pylori:
GalNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer (8А),
GalNAcα3Galβ4GlcNAcβ3Galβ4GlcβCer (8В) и
Galα3Galβ4GlcNAcβ3Galβ4GlcβCer (8С). Также представлена несвязывающая структура Galα3Galβ4GlcNH2β3Galβ4GlcβCer (8D). Также показан вид сверху олигосахаридного фрагмента каждой рассчитанной структуры с минимальной энергией. Несмотря на различия в аномерности, отсутствие или наличие ацетамидной группы, аксиальное или экваториальное положение 4-ОН в концевом сахаре и тот факт, что плоскость кольца соединений с концевой α3-связью несколько приподнята по сравнению с соответствующей плоскостью в соединениях с β3-связью, имеется значительная топографическая аналогия между данными структурами, и также GlcNAcβ3-концевой структурой, полученной из тимуса кролика (фиг.9А), что таким образом объясняет их сходную аффинность для бактериального адгезина. В противоположность ацетамидная группа внутреннего GlcNAcβ3 важна для связывания (сравни 8С и 8D).
Фиг.9А, 9В, 9С и 9D.Рассчитанные конформации с минимальной энергией активных в отношении связывания гликосфинголипидов GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer (9А) и
Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer (9В) и несвязывающих гликосфинголипидов NeuAcα3GalNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer (9С) и Galα3(Fucα2)Galβ4GlcNAcβ3Galβ4GlcβCer (9D). У двух последних удлиненных последовательностей (9С и 9D) отсутствует связывание Helicobacter pylori, в то время как у первой (9В) имеется, но пониженная аффинность. Наряду с установлением того, что де-N-ацилирование ацетамидной группы внутреннего GlcNAc в В5 (фиг.8А, 8В, 8С и 8D) полностью ликвидирует связывание, участок, составляющий связывающий эпитоп, должен состоять из конечного трисахарида В5, показанного на фиг.8С, поскольку ацетамидная группа концевого N-ацетилгалактозамина не является важной.
Фиг.10. Конформер с минимальной энергией соединения NeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer из семи сахаров представлен в двух проекциях, находящихся под углом 90° друг к другу. Концевой атом углерода гликолильной группы сиаловой кислоты, а также углеродные атомы метила ацетамидных групп двух внутренних остатков GlcNAc отмечены черным только для облегчения ориентации зрителя. Для связи GlcβCer обоснованно была выбрана удлиненная конформация для представления, но минимальная связывающая последовательность GlcNAcβ3Galβ4GlcNAcβ3 наиболее подходит для связи с адгезином в конформациях GlcβCer, чем другая показанная здесь.
Фиг.11А, 11В и 11С.Связывание моноклональных антител ТН2 (11В) и лектина из E.cristagalli (11С) с цельными некислотными фракциями гликосфинголипидов из эпителиальных клеток слизистой желудка человека, гранулоцитов человека и эритроцитов человека, разделенных на хроматограммах, полученных тонкослойной хроматографией. На фиг.11А показаны те же фракции, окрашенные 4-метоксибензальдегидом. Авторадиографию в случае (11В) и (11С) проводили в течение 12 ч. Для полос 1-6 наносили 80 мкг цельных некислых фракций из эпителиальных клеток слизистой желудка человека от индивидуумов с пятью различными группами крови А, в то время как для полосы 6 наносили 40 мкг цельной некислотной фракции из гранулоцитов человека и для полосы 7-40 мкг цельной некислотной фракции из эритроцитов человека. Тесты с нанесением проводили, как описано в разделе «Материалы и методы».
Подробное описание изобретения
Настоящее изобретение относится к группе специфических олигосахаридных последовательностей, связывающихся с Helicobacter pylori. Провели скрининг многочисленных естественных гликосфинголипидов тонкослойной хроматографией с нанесением бактерий (таблица 2). Структуры использованных гликосфинголипидов характеризовали протонной ЯМР и масс-спектрометрией. Для сравнительного анализа трехмерной структуры соединений, связывающихся с Helicobacter pylori, использовали молекулярное моделирование.
Новая специфичность связывания была показана при сравнении четырех пентасахаридных гликолипидов. Было установлено, что замена концевого сахарида невосстанавливающего конца в GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer на GalNAcβ3 (краткое название х2GSL), GalNAcα3 или Galα3 (В5) приводило к связыванию Helicobacter pylori, несмотря на имеющиеся различия в аномерности, отсутствие или наличие ацетамидной группы и аксиальное/экваториальное положение 4-ОН. Специфичность также включает структуры с более слабым связыванием Helicobacter pylori: более короткую форму Galβ4GlcNAcβ3Galβ4GlcβCer и β4-удлиненные формы гликолипида с концевым N-ацетилглюкозамином: Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer и
NeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer.
В противоположность ранее известной зависимой от сиаловой кислоты специфичности (Evans et al., 1988; Miller-Podraza et al., 1996; 1997a), N-гликолилнейраминовую кислоту у последнего указанного гликосфинголипида можно удалить, не влияя при этом на связывание Helicobacter pylori.
Связывание с GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer носило в значительной степени воспроизводимый характер, хотя в целом на связывание сахаридов с Helicobacter pylori оказывали влияние фазовые изменения бактерий, и высокую аффинность связывания можно было наблюдать в тесте с нанесением бактерий в низких пикомолярных количествах гликолипида.
Установили длину связывающего эпитопа и было показано, что GlcNAcβ3Galβ4GlcβCer, Galβ4GlcNβ3Galβ4GlcβCer и
Galα3Galβ4GlcNβ3Galβ4GlcβCer (укороченная и N-дезацетилированная формы активных соединений) не связывались с Helicobacter pylori. Этими данными было установлено, что внутренний остаток GlcNAc участвует в связывании, но один он не приводит к достаточно прочному связыванию. Было установлено, что связывающим эпитопом является концевой трисахарид в пентасахаридных эпитопах, которые обсуждались выше. Когда присутствуют только два остатка, как в Galβ4GlcNAcβ3Galβ4GlcβCer, то связывание является более слабым, и в гликолипиде из шести сахаридов,
Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer, концевой Galβ4 ингибирует связывание, что объясняет более слабую активность. Гликолипид из семи сахаридов, имеющий Galα3 в менее активной структуре гликолипида из шести сахаридов,
Galα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer, обладал более высокой активностью, что также указывает на то, что для высокой связывающей активности необходимы концевые трисахаридные эпитопы.
Специфичность связывания исследовали при анализе изомеров и модифицированных форм активных соединений. Удлиненные формы Galβ4GlcNAcβ3Galβ4GlcβCer, имеющие следующие модификации концевого Gal: Fucα2 (краткое название H5-2), Fucα2 и Gal/GalNAcα3 (B6-2, A6-2), Neu5Acα3 или Neu5Acα6 (сиалилпараглобозиды) или Galα4 (P1) были неактивны в анализе связывания с Helicobacter pylori. Связывание также нарушалось при наличии β6-связанного внутреннего ответвления из галактозы, представленного структурой
Galβ4GlcNAcβ3(Galβ4GlcNAcβ6)Galβ4GlcβCer. Было показано, что ответвление изменяет представление Galβ4GlcNAcβ3-эпитопа, и дисахаридный связывающий сайт, возможно, имеет стерические затруднения (Teneberg et al., 1994). (Однако результаты показывают, что внутренний остаток галактозы, с которым связаны дисахаридные или трисахаридные связывающие эпитопы, связан β3-связью, может также вносить свой вклад в связывание.) Кроме того, оказалось, что Neu5Acα3GalNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer (удлиненная форма активного x2-гликосфинголипида) или GalNAcβ3Galα3Galβ4GlcNAсβ3Galβ4Glcβ3Cer (удлиненный GSL B5) не связываются с Helicobacter pylori.
Для сравнения активных связывающих структур и неактивных соединений использовали молекулярное моделирование. На примере рассчитанных конформеров с минимальной энергией четырех пентасахаридных гликосфинголипидов (Galβ4GlcNAcβ3Galβ4GlcβCer с удлинением на GlcNAcβ3, GalNAcβ3, GalNAcα3 или Galα3) было показано, что конформации соединений могут в значительной степени быть сходными друг с другом. Конформации неактивных гликолипидов были другими. Несмотря на то, что концевые сахариды также могут различаться по их аномерной связи (два с альфа- и два с бета-связями), при молекулярном моделировании было установлено, что структуры с минимальной энергией топографически являются очень близкими. Различия концевых структур заключаются в том, что в Galα3 отсутствует ацетамидная группа, имеющаяся в трех других, Gal и GalNAc имеют 4-ОН в аксиальном положении и GlcNAc - в экваториальном положении, плоскости кольца альфа аномерного конца несколько приподняты по сравнению с соответствующей плоскостью бета-аномерных производных. Возможно удлинение конца в положении 4 в GlcNAc, указывая на то, что 4-ОН не имеет такого важного значения для связывания, хотя удлинение Galβ4 оказывает стерическое влияние. В итоге оказалось, что ни положение 4-OH, ни отсутствие/наличие ацетамидной группы, ни аномерная структура концевого моносахаридного остатка не являются решающими для связывания, поскольку все четыре пентасахаридных гликолипида имеют аналогичную аффинность для адгезина Helicobacter pylori.
В свете этих закономерностей связывания четыре других концевых моносахарида в связывающем соединении можно также обеспечить трисахаридными связывающими эпитопами: Galβ3Galβ4GlcNAc, GlcNAcα3Galβ4GlcNAc, Glcβ3Galβ4GlcNAc и Glcα3Galβ4GlcNAc. Они аналогичны исследованным последовательностям и имеют единственные различия в аномерной, 4-эпимерной или C2 NAc/OH структурах. Первая имеется в гликолипиде из эритроцитов человека, в то время как о наличии последних трех в тканях человека до сих пор неизвестно, и в большей степени они могут представлять аналоги естественного рецептора.
Было показано, что связывающий эпитоп включает концевой трисахаридный элемент активных гликолипидов из пяти сахаридов и, по меньшей мере, в более крупных повторяющихся N-ацетилактозаминах эпитоп также может находиться в середине сахаридной цепи. Заявители понимают, что связывающие эпитопы могут быть представлены различными путями в естественных и полученных биосинтетическим путем гликоконъюгатах и олигосахаридах таких, как О-связанные или N-связанные гликаны гликопротеинов, или в олигосахаридах производных поли-N-ацетилактозамина. В результате химического или ферментативного синтеза, особенно в области углеводов, получали почти бесконечное количество производных и аналогов. Размер связывающего эпитопа позволяет провести некоторые модификации, например, в положениях С1, С2 и С4 концевого моносахарида, удалением невосстанавливающего концевого моносахарида или удлинением от положения С4 концевого GlcNAc в GlcNAcβ3Galβ4GlcNAc, например, положение С4 в GlcNAcβ3 может быть связано с олигосахаридной цепью гликозидной связью. Когда олигосахаридной последовательностью является
GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc, то положение С4 в концевом GlcNAcβ3 можно связать с Galβ1- или олигосахаридной цепью гликозидной связью. Для получения производных или олигомерных, или полимерных конъюгатов, обладающих активностью связываться с Helicobacter pylori, особенно можно использовать С2- и С4-положения моносахаридного остатка невосстанавливающего концевого в трисахаридном эпитопе и восстанавливающие концы эпитопа. Также для получения производных и аналогов можно использовать С6-положения моносахаридных остатков, особенно предпочтительным являются С6-положение невосстанавливающего концевого остатка в трисахаридной последовательности и восстанавливающий концевой остаток ди- и трисахаридных связывающих соединений.
В данном изобретении термины «аналог» и «производное» определяются следующим образом. По настоящему изобретению, возможно сконструировать структурные аналоги или производные связывающих Helicobacter pylori олигосахаридных последовательностей. Таким образом, изобретение также относится к структурным аналогам соединений по изобретению. Структурные аналоги по изобретению включают структурные элементы, имеющие значение для связывания Helicobacter pylori с олигосахаридными последовательностями. Для конструирования эффективных структурных аналогов важно знать, какой структурный элемент имеет значение для связывания Helicobacter pylori и сахаридов. Предпочтительно, чтобы важные структурные элементы были не модифицированы, или модифицировались таким образом, чтобы они были очень подобны важному структурному элементу. Данные элементы предпочтительно включают 4- и 6-гидроксильные группы остатка Galβ4 в трисахаридном и дисахаридном эпитопах. Также важным структурным элементом является расположение связей между кольцевыми структурами. Для высокой аффинности связывания предпочтительной является ацетамидная группа или группа, подобная ацетамидной, в положении, соответствующем ацетамидной группе восстанавливающего конца-GalNAc ди- или трисахаридных эпитопов. Группа, подобная ацетамидной, может быть другим амидом, например, алкиламидо, ариламидо, вторичным амином, предпочтительно N-этилом или N-метилом, О-ацетилом, или О-алкилом, например, О-этилом или О-метилом. Для высокой аффинности связывания предпочтительными являются амиды по карбоксильной группе концевой уроновой кислоты и ее аналогов. Полагается, что активность немодифицированной уроновой кислоты возрастает при уменьшении значения рН.
Структурные производные по изобретению являются олигосахаридными последовательностями по изобретению, химически модифицированные таким образом, что связывание с Helicobacter pylori сохраняется или повышается. Предпочтительно по изобретению дериватизировать одну или несколько гидроксильных или ацетамидных групп олигосахаридных последовательностей. В изобретении описывается несколько положений молекул, по которым можно провести изменения для получения аналогов или производных. Гидроксильная или ацетамидная группы, для которых допускается, по меньшей мере, несколько модификаций, обозначены R-группами в формуле 1.
Крупные или кислотные заместители и другие структуры, такие как моносахаридные остатки, являются неустойчивыми по меньшей мере, когда находятся в положении С2-, С3- или С6-гидроксильных групп Galβ4GlcNAc и в С3-гидроксильной группе моносахара невосстанавливающего концевого моносахарида трисахаридных эпитопов. Способы получения олигосахаридных аналогов для связывания лектина являются хорошо известными. Например, были получены многочисленные аналоги сиалил-Lewis × олигосахарида, представляющие различный скелет активных функциональных групп, смотри страницу 12090 у Sears и Wong, 1996. Подобные аналоги олигосахаридов на основе гепарина получены корпорацией Sanofi, и ингибиторы, подобные сиаловой кислоте, такие как занамивир и тамифлу (Relenza) для сиалидазы были получены многочисленными группами. Предпочтительно олигосахаридный аналог получают на основе молекулы, включающей по меньшей мере 5- или 6-членную кольцевую структуру, более предпочтительно аналог включает по меньшей мере две кольцевые структуры, состоящие из 6 или 5 атомов. Предпочтительный тип аналогов олигосахарида включает амид концевой уроновой кислоты или аналог, связанный со структурой, подобной Galβ4GlcNAc-сахариду. Альтернативно амид концевой уроновой кислоты является 1-3-связанным с Gal, который связан со структурой, подобной GlcNAc. В подобных структурах моносахаридные кольца могут быть замещены кольцами такими, как циклогексан или циклопентан, ароматическими кольцами, включая бензольное кольцо, гетероциклические кольцевые структуры могут включать помимо атома кислорода, например, атомы азота и серы. Для сохранения активных конформаций колец кольцевые структуры можно связать между собой устойчивыми линкерами. Типичная подобная структура также может включать пептидные аналоги-структуры для олигосахаридной последовательности или ее части.
Влияние активных групп на связывающую активность является кумулятивным, и отсутствие одной группы может быть компенсировано добавлением активного остатка с другой стороны молекулы. Для получения структур-аналогов для связывающих Helicobacter pylori олигосахаридных последовательностей по изобретению можно использовать молекулярное моделирование, предпочтительно осуществляемое с помощью компьютера. Данные молекулярного моделирования нескольких олигосахаридных последовательностей представлены в примерах, и такие же или подобные методы, помимо ЯМР и рентгеновской кристаллографии, можно применять для получения структур для других олигосахаридных последовательностей по изобретению. Для поиска аналогов олигосахаридные структуры можно «соединить» со связывающей молекулой(ами) углеводорода Helicobacter pylori, наиболее вероятно с лектинами бактерий, и можно найти возможные дополнительные связывающие взаимодействия.
Также следует отметить, что одновалентные, олиговалентные или поливалентные олигосахариды можно активировать для получения более высокой активности в отношении лектинов получением производного олигосахарида комбинаторной химией. Когда библиотека создана замещением одного или нескольких остатков в олигосахаридной последовательности, ее можно рассматривать в качестве библиотеки производных, альтернативно тому, когда библиотека создана из аналогов олигосахаридных последовательностей, описанных в изобретении. Библиотеку комбинаторной химии можно создать на основе олигосахарида или его предшественника, или гликоконъюгатов по изобретению. Например, можно получить олигосахариды с различным восстанавливающим концом с помощью так называемой карбогибридной технологии.
В предпочтительном воплощении библиотека комбинаторной химии конъюгирована со связывающими Helicobacter pylori соединениями, описанными в изобретении. В более предпочтительном воплощении библиотека включает, по меньшей мере, 6 различных молекул. Предпочтительно модификации комбинаторной химии получают с помощью различных амидов по карбоциклической группе в R8 формулы 1. Группа для модификации в R8 может также представлять альдегид или амин, или другой тип реакционноспособной группы. Такая библиотека является предпочтительной для применения при анализе связывания микроорганизмов с олигосахаридными последовательностями по изобретению. Аминокислоты или группы органических амидов являются промышленно доступными, эти соединения можно использовать для синтеза комбинаторной библиотеки амидов уроновой кислоты. С помощью комбинаторной библиотеки можно идентифицировать связывающее соединение с высокой аффинностью, например, при использовании теста ингибирования, в котором соединения из библиотеки применяются для ингибирования связывания бактерий с гликолипидами или гликоконъюгатами, описанными в изобретении. Предпочтительные структурные аналоги и производные по изобретению могут ингибировать связывание Helicobacter pylori со связывающими Helicobacter pylori олигосахаридными последовательностями по изобретению.
Стерическое затруднение за счет липидной части или близость поверхности силикагеля, возможно, ограничивает определение эпитопа GlcNAcβ3Galβ4Gal в имеющемся тесте TCX. В недавно проведенном исследовании невозможно было поставить тест определения активности данной последовательности с токсином А из Clostridium difficile, который специфически распознает такие же четыре трисахаридных эпитопа, описанных для Helicobacter pylori (Teneberg et al., 1996). Однако другими исследователями было показано связывание Galα3Galβ4Glc с токсином А при использовании модифицированного крупным полимерным спейсером конъюгата сахарида (Castagliuolo et al., 1996). Также принимая во внимание вклад, который вносит концевой трисахарид в связывание, можно поместить Glc в восстанавливающий конец эпитопа; возможно, в неактивной N-дезацетилированной форме положительный заряд свободной аминогруппы в большей степени препятствует связыванию, чем присутствие гидроксильной группы. Трисахаридные эпитопы с Glc на восстанавливающем конце рассматриваются как эффективные аналоги связывающего Helicobacter pylori соединения, когда находятся в олиговалентной или более предпочтительно в поливалентной форме. Одним воплощением настоящего изобретения являются сахариды с Glc на восстанавливающем конце, которые используются в виде свободных восстанавливающих сахаридов в высокой концентрации, предпочтительно в пределах 1-100 г/л, более предпочтительно 1-20 г/л. Понятно, что данные сахариды могут обладать минимальной активностью в пределах концентраций 0,1-1 г/л.
В последующем связывающая Helicobacter pylori последовательность описана в виде олигосахаридной последовательности. Олигосахаридная последовательность, определенная здесь, может быть частью естественного или синтетического гликоконъюгата или свободным олигосахаридом, или частью свободного олигосахарида. Подобные олигосахаридные последовательности можно связать с различными моносахаридами или олигосахаридами, или полисахаридами в полисахаридных цепях, например, если сахаридная последовательность экспрессируется в виде части бактериального полисахарида. Кроме того, известны многочисленные естественные модификации моносахаридов, примерами являются О-ацетильное или сульфатированное производное олигосахаридных последовательностей. Связывающее Helicobacter pylori соединение, определенное здесь, может включать описанную олигосахаридную последовательность в виде части естественного или синтетического гликоконъюгата или соответствующего свободного олигосахарида, или части свободного олигосахарида. Связывающее Helicobacter pylori соединение также может включать смесь связывающих Helicobacter pylori олигосахаридных последовательностей.
Несколько производных рецепторных олигосахаридных последовательностей обладают связыванием ниже порога чувствительности используемого в данном случае теста, проявляя тем самым специфичность распознавания. Данные по связыванию показывают, что если в указанных олигосахаридных последовательностях имеется GalNAcβ3, связанный с Galα3Galβ4GlcNAc (замещенная последовательность: GalNAcβ3Galα3Galβ4GlcNAc) или Neu5Acα3, связанный с GalNAcβ3Galβ4GlcNAc (замещенная последовательность: Neu5Acα3GalNAcβ3Galβ4GlcNAc), то соединения неактивны. Когда указанная олигосахаридная последовательность представляет Galβ4GlcNAc, то это не α4-галактозилированная (последовательность не является Galα4Galβ4GlcNAc), α3- или α6-сиалилированная (последовательность не является Neu5Acα3/6Galβ4GlcNAc), α2- или α3-фукозилированная [указанная последовательность не является Fucα2Galβ4GlcNAc или Galβ4(Fucα3)GlcNAc или Fucα2Galβ4(Fucα3)GlcNAc, α3-фукозилирование относится к фукозилированию остатков GlcNAc лактозамина, образующего Lewis х, Galβ4(Fucα3)GlcNAc]. Сахариды, имеющие структуры, в которых Galβ3 связан с GlcNAcβ3 (такие, как Galβ3GlcNAcβ3Galβ4GlcNAc/Glc), имеют другие конформации по сравнению со связывающими Helicobacter pylori соединениями, описанными здесь, и отдельно изучалась специфичность их связывания. Связывающие Helicobacter pylori соединения могут быть частью сахаридной цепи или гликоконъюгата, или смеси гликосоединений, включающих другие известные связывающие Helicobacter pylori эпитопы, с другими сахаридными последовательностями и конформациями такими, как Lewis b (Fucα2Galβ3(Fucα4)GlcNAc) или Nue5Acα3Galβ4Glc/GlcNAc. Для терапии может быть полезным использование нескольких связывающих соединений вместе.
Связывающие Helicobacter pylori олигосахаридные последовательности можно синтезировать ферментативно под действием гликозилтрансфераз, или получить трансгликозилированием, катализируемым гликозидазой или трансгликозидазой (Ernst et al., 2000). Можно заложить специфичность данных ферментов и применение кофакторов. Специфические модифицированные ферменты можно использовать для проведения более эффективного синтеза, например, гликосинтазу модифицируют только для проведения трансгликозилирования. Органический синтез сахаридов и конъюгатов или конъюгатов, описанный здесь, аналогичен известным способам (Ernst et al., 2000). Сахариды можно выделить из естественных источников и превратить химически или ферментативно в связывающие Helicobacter pylori соединения. Естественные олигосахариды можно выделить из молока, продуцированного различными жвачными. Для получения сахаридов можно использовать трансгенных животных таких, как коровы или микроорганизмы, экспрессирующих ферменты гликозилирования.
Моносахаридные остатки уроновой кислоты, описанные в изобретении, можно получить способами, известными в данной области. Например, гидроксильную группу при 6-углероде N-ацетилглюкозамина или N-ацетилгалактозаминов можно окислить химическим путем в карбоновую кислоту. Окисление можно провести с надлежаще защищенным олигосахаридом или моносахаридом.
В предпочтительном воплощении незащищенный полимер или олигомер, включающий гексозы, N-ацетилгексозамины или гексозамины, в которых связь между моносахаридами, не находится между 6 атомами углерода, представляет:
1) окисленный в соответствующий полимер остатков уроновой кислоты или полимер, включающий мономеры 6-альдегидомоносахаридов;
2) необязательно дериватизированный из карбоксильной группы или 6-альдегидной группы предпочтительно в амид или амин и
3) гидролизованный до моносахаридов уроновой кислоты или моносахаридов производных уроновой кислоты.
Способы окисления моносахаридных остатков в уроновые кислоты и гидролиза амина или полимеров уроновой кислоты химическим или ферментативным путем хорошо известны в данной области. Особо предпочтительно использовать способ для олигомеров или полимеров целлюлозы, крахмала или других гликанов с 1-2- или 1-3-, или 1-4-связями, хитина (GlcNAc-полимера) или хитозана (GlcN-полимера), которые являются промышленно доступными в больших количествах, или полисахариды на основе N-ацетилгалактозамина/галактозамина (например, происходящие из бактерий) окисляют в соответствующий 1-4-связанный сахарид. Данный способ можно также применить к полимерам галактана. Производные уроновой кислоты можно также получить из естественных полимеров, включающих уроновые кислоты, таких, как пектины или глюкуроновая кислота, содержащие полисахариды бактерий, включая N-ацетилгепарин, экзополисахариды бактериального типа, производные гиалуроновой кислоты и хондроитина. Данный способ включает:
1) дериватизацию карбоксильных групп полисахарида, предпочтительно по амидной связи и
2) гидролиз полисахарида в моносахариды на основе уроновой кислоты или моносахариды на основе производного уроновой кислоты.
Известны также химические и ферментативные способы окисления первичного спирта по 6 углероду полисахарида в альдегид или карбоновую кислоту. Затем альдегид можно превратить, например, в амин восстановительным аминированием. Предпочтительно окисляют концевые Gal или GalNAc под действием ферментоподобной галактозоксидазы, окисляющей первичные спирты, и затем дополнительно дериватизируют, например, аминами.
Остатки уроновой кислоты можно конъюгировать с дисахаридами или олигосахаридами обычными способами органической химии. Альтернативно GlcA можно связать с глюкуронилтрансферазой, катализирующей перенос GlcA с UDP-GlcA на концевой Lac(NAc).
Производные моносахаридов, подобные N-ацетилгексозаминам, можно получить из полимера или олигомера, содержащего гексозамины или другие моносахариды со свободными первичными аминогруппами способом, включающим:
1) дериватизацию аминогрупп во вторичный или третичный амин или амид;
2) гидролиз полимера в соответствующие моносахариды.
Хитозан и его олигосахариды представляют пример амина, включающего полимер или олигомер.
В основном способ получения содержащего карбоновую кислоту, содержащего 6-альдегидную группу, содержащего амин и/или амид моносахарида/моносахаридов, включает следующие стадии:
1) необязательное введение карбоксильной группы или 6-альдегидной группы в углеводный полимер, в котором имеется первичная гидроксильная группа для модификации;
2) превращение карбоксильных групп или 6-альдегидных групп, или первичных аминогрупп полимера во вторичные или третичные амины, или амиды, когда проводится стадия 1, то стадия 2 является необязательной;
3) гидролиз полимера в соответствующие моносахариды.
Гидролиз до моносахаридов также может быть частичным, и в результате его получают дисахарид или олигосахарид для получения соединений-аналогов. Предпочтительно, чтобы в результате гидролиза получалось 30% моносахаридов. Способы проведения химических стадий известны в данной области. Например, окисление полисахаридов в соответствующие моносахариды можно провести, как описано Muzzarelli et al., 1999 и 2002. Данные способы являются предпочтительными для применения незащищенных моносахаридов, поскольку при этом исключается проведение защиты реакционноспособных восстанавливающих концов моносахаридов.
В предпочтительном воплощении олигосахаридные последовательности, включающие GlcAβ3Lac или GlcAβ3LacNAc, эффективно синтезируют трансгликозилированием с использованием специфической глюкуронидазы такой, как глюкуронидаза из печени крупного рогатого скота. Понятно, что фермент может сайт-специфически переносить с β1-3-связи на Galβ4GlcNAc и Galβ4Glc с неожиданно высоким для реакции трансгликозилирования выходом. Как правило, в реакциях трансгликозилирования не получают подобную избирательность и выход, близкий к 30% или выше.
Одним воплощением настоящего изобретения является применение связывающего Helicobacter pylori соединения или рецептора, включающего олигосахаридную последовательность.
в которой g, r, s, t и u каждый независимо равен 0 или 1,
так, что когда t=0 и u=0, то тогда олигосахаридная последовательность связана с поливалентным носителем или находится в виде свободного олигосахарида в высокой концентрации, и аналоги или производные указанной олигосахаридной последовательности, обладающие активностью связывания с Helicobacter pylori, для производства композиции, обладающей активностью связывать или ингибировать Helicobacter pylori.
А в вышеуказанной олигосахаридной последовательности обозначает уроновую кислоту моносахаридного остатка или производное по 6 углероду моносахаридного остатка, наиболее предпочтительным производным по 6 углероду является амид уроновой кислоты.
Последующие олигосахаридные последовательности находятся среди предпочтительных связывающих Helicobacter pylori соединений для применения по изобретению:


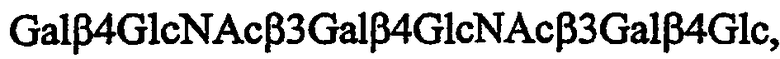
и их поливалентные конъюгаты с восстанавливающим концом,
а также

Другое воплощение изобретения описано формулой 1.
Формула 1:
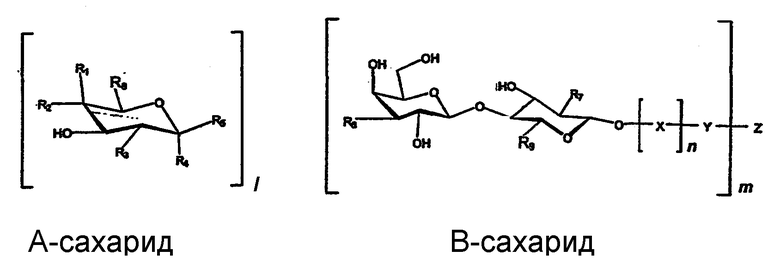
Среди предпочтительных связывающих Helicobacter pylori соединений или смесей соединений по изобретению и для применений по изобретению находятся олигосахаридные структуры формулы 1, в которой целые числа l, m и n имеют значения m≥1, l и n независимо равны 0 или 1, и в которой R1 представляет Н, и R2 представляет ОН, или R1 представляет ОН, и R2 представляет Н, или R1 представляет Н, и R2 представляет моносахаридильную или олигосахаридильную группу, предпочтительно галактозильную группу, связанную бета-гликозидной связью, R3 независимо представляет -ОН или ацетамидо(-NHCOCH3), или аналогичную ацетамидной группу. R7 представляет ацетамидную (-NHCOCH3) или аналогичной ацетамидную группу. Когда l=1, R4 представляет -Н, R5 представляет атом кислорода, связанный со связью R6, и образует бета-аномерную гликозидную связь с сахаридом В, или R5 представляет -Н, и R4 представляет атом кислорода, связанный со связью R6, и образует альфа-аномерную гликозидную связь с сахаридом В, когда l=0, R6 представляет ОН, связанный с В. Х представляет остаток моносахарида или олигосахарида, предпочтительно Х представляет лактозил-, галактозил-, поли-N-ацетил-лактозаминил, или часть олигосахаридной последовательности на основе О-гликана или N-гликана; Y представляет спейсер или концевой конъюгат такой, как липидная группа керамида или связь с Z. Z представляет олиговалентный или поливалентный носитель. Связывающее соединение может также быть аналогом или производным указанного соединения формулы 1, обладающими активностью связывания по отношению к Helicobacter pylori, например, кислородную связь (-О-) между положением С1 сахарида В и Х-остатка сахарида, или спейсер Y можно заменить углеродным (-С-), азотным (-N-) или серным (-S-) мостиком.
В формуле 1 R8 предпочтительно представляет амид карбоновой кислоты такой, как метиламид или этиламид, гидроксиметил (-СН2-ОН) или карбоксильную группу, или ее эфир такой, как метиловый или этиловый эфир. Амид карбоновой кислоты может включать альтернативную связь с поливалентным носителем Z, содержащим амин такой, как полисахарид хитозан или галактозамин, или Z, содержащим первичный амин, включающий спейсер, предпочтительно гидрофильный спейсер. Структура R8 может быть также подобной описанным выше структурой, известной в данной области. Например, можно использовать вторичные или третичные амины, или амидированный вторичный амин.
В формуле 1 R9 предпочтительно представляет гидроксиметил, но его можно использовать для видов дериватизации, описанных для R8.
R3 представляет гидроксил, ацетамидную или подобную ацетамидной группу такую, как С1-6алкиламиды, ариламидо, вторичный амин, предпочтительно N-этил или N-метил, О-ацетил, или О-алкил, например, О-этил или О-метил. R7 имеет те же значения, что R3, но предпочтительно представляет ацетамидную или подобную ацетамидной группу.
R2 может предпочтительно включать шестичленную кольцевую структуру, подобную Galβ4-концу.
Связывающие бактерии соединения предпочтительно представлены в виде кластеров таких, как гликолипиды на клеточных мембранах, мицеллы, липосомы, или на твердых фазах таких, как пластинки для ТСХ в тестах. Представление в виде кластеров с правильным расположением приводит к высокой аффинности связывания.
По изобретению также возможно использовать связывающие Helicobacter pylori эпитопы или их естественный, или полученный синтетическим путем аналог или производное, обладающие такой же или лучшей активностью связывания по отношению к Helicobacter pylori. Также возможно использовать производное, включающее связывающее бактерии соединение такое, как рецепторный активный ганглиозид, описанный в изобретении, или его аналог, или производное, обладающие такой же или лучшей активностью связывания по отношению к Helicobacter pylori. Связывающее бактерии соединение может быть связано гликозидной связью с концевым эпитопом олигосахаридной цепи. Альтернативно связывающий бактерии эпитоп может быть ответвлением олигосахаридной цепи, предпочтительно полилактозамина.
Связывающее Helicobacter pylori соединение может быть конъюгировано с антибиотиком, предпочтительно антибиотиком типа пенициллина. Связывающее Helicobacter pylori соединение целенаправленно доставит антибиотик к Helicobacter pylori. Подобный конъюгат полезен для лечения, поскольку требуется меньшее количество антибиотика для лечения или терапии инфекции, вызываемой Helicobacter pylori, что приводит к меньшему проявлению побочных эффектов антибиотика. Часть конъюгата, приходящаяся на антибиотик, направлена на уничтожение или ослабление бактерий, но конъюгат также может обладать антиадгезивным действием, описанным ниже.
Связывающие бактерии соединения, предпочтительно в олиговалентной форме или в форме кластеров, можно использовать для лечения заболевания или состояния, вызванного присутствием Helicobacter pylori. Это обеспечивается использованием связывающих Helicobacter pylori соединений против адгезии, т.е. для подавления связывания Helicobacter pylori с рецепторными эпитомами клеток- или тканей-мишеней. При введении связывающего Helicobacter pylori соединения или фармацевтической композиции, они будут конкурировать с рецепторными гликоконъюгатами на клетках-мишенях за связывание с бактериями. Тогда некоторые или все бактерии будут связываться со связывающим Helicobacter pylori соединением вместо рецептора на клетках-мишенях или тканях. Затем бактерии, связанные со связывающими Helicobacter pylori соединениями, элиминируют из организма пациента (например, потоком жидкости в желудочно-кишечном тракте), приводя к меньшему влиянию бактерий на здоровье пациента. Предпочтительно используемое соединение является растворимой композицией, включающей связывающие Helicobacter pylori соединения. Соединение можно «посадить» на носитель, который предпочтительно не является белком. При использовании молекулы-носителя несколько молекул связывающего Helicobacter pylori соединения можно соединить с одним носителем и тем самым повысить эффективность ингибирования.
Клетки-мишени в основном представляют эпителиальные клетки ткани-мишени, особенно желудочно-кишечного тракта, другими потенциальными тканями-мишенями являются, например, печень и поджелудочная железа. Гликозилирование ткани-мишени может изменяться под действием инфекции, вызванной патогеном (Karlsson et al., 2000). Клетки-мишени также могут быть злокачественными, трансформированными или раковыми/опухолевыми клетками в ткани-мишени. Трансформированные клетки и ткани имеют измененные типы гликозилирования и могут нести рецепторы для бактерий. Связывание лектинов или сахаридов (углевод-углеводное взаимодействие) с сахаридами на гликопротеиновых или гликолипидных рецепторах может активировать клетки, и в случае раковых/злокачественных клеток это может привести к росту или метастазированию опухолей. Сообщалось, что несколько описанных здесь олигосахаридных эпитопов таких, как GlcNAcβ3Galβ4GlcNAc (Hu J. et al., 1994), Galα3Galβ4GlcNAc (Castronovo et al., 1989) и нейтральные и сиалилированные полилактозамины из злокачественных клеток (Stroud et al., 1996) связаны с опухолями или являются опухолевыми антигенами. Также описаны олигосахаридные цепи, включающие соединения, описанные здесь, происходящие из лимфоцитов (Vivier et al., 1993). Helicobacter pylori связаны с лимфомой желудка. Описанные здесь соединения можно использовать для предупреждения связывания Helicobacter pylori с предопухолевыми или опухолевыми клетками и активации роста опухоли и метастазирования. Подавление связывания может оказывать лечебное действие на опухоли желудка, особенно лимфому. Описана связывающая Helicobacter pylori олигосахаридная последовательность структуры GlcNAcβ3Galβ4GlcNAcβ6GalNAc из муцина желудка человека. Данный эпитоп муцина и аналогичные О-гликановые гликоформы наиболее вероятно являются естественными рецепторами Helicobacter pylori в желудке человека с высокой аффинностью. Также на это указывает высокая аффинность связывания аналогичной последовательности
GlcNAcβ3Galβ4GlcNAcβ6GlcNAc в виде неогликолипида для Helicobacter pylori, и тот факт, что последовательность GlcNAcβ3Galβ4GlcNAcβ6Gal также имеет некоторую активность связывания по отношению к Helicobacter pylori в том же тесте. Следовательно, предпочтительные олигосахаридные последовательности включают О-гликаны и аналоги последовательностей на основе О-гликана, такие как GlcNAcβ3Galβ4GlcNAcβ6GlcNAc/GalNAc/Gal, GlcNAcβ3Galβ4GlcNAcβ6GlcNAc/GalNAc/GalαSer/Thr, GlcNAcβ3Galβ4GlcNAcβ6(Gal/GlcNAcβ3)GlcNAc/GalNAc/GalαSer/Thr и гликопептиды и аналоги гликопептидов, включающие последовательности на основе О-гликана. Даже последовательности без невосстанавливающего конца GlcNAc могут обладать некоторой активностью. Основываясь на этом, все другие связывающие Helicobacter pylori олигосахаридные последовательности (OS) и особенно трисахаридные эпитопы также являются особенно предпочтительными, когда связаны с восстанавливающим концом с образованием структур OSβ6Gal(NAc)0-1 или OSβ6Glc(NAc)0-1, или OSβ6Gal(NAc)0-1αSer/Thr, или OSβ6Glc(NAc)0-1αSer/Thr. Ser- или Thr-соединения или их аналоги, или восстанавливающие олигосахариды также являются предпочтительными, когда связаны с поливалентным носителем. Восстанавливающие олигосахариды можно связать при восстановлении с поливалентным носителем.
Клетки-мишени также включают клетки крови, особенно лейкоциты. Известно, что штаммы Helicobacter pylori, вызывающие пептическую язву, как основной используемый здесь штамм, вызывают ответную воспалительную реакцию гранулоцитов, даже, когда бактерии не опсонизированы (Rautelin et al., 1994 a, b). Наиболее вероятно первоначальное событие при фагоцитозе бактерий заключается в специфических лектиноподобных взаимодействиях, приводящих к агглютинации гранулоцитов (Ofek и Sharon, 1988). После фагоцитоза имеют место реакции дыхательного взрыва, которые могут иметь значение для патогенеза вызываемых Helicobacter pylori заболеваний (Babior, 1978). Было выделено из гранулоцитов и охарактеризовано несколько сиалилированных и некислых гликосфинголипидов, имеющих повторяющиеся единицы N-ацетиллактозамина (Fukuda et al., 1985; Stroud et al., 1996), и они могут функционировать таким образом в качестве потенциальных рецепторов для Helicobacter pylori на поверхности лейкоцитов. Кроме того, из того же источника также был выделен х2-гликосфинголипид (Teneberg, S., неопубликованные данные). Настоящее изобретение подтверждает наличие рецепторных сахаридов на эритроцитах и гранулоцитах человека, которые могут распознаваться специфическим для N-ацетиллактозамина лектином и моноклональными антителами (X2, GalNAcβ3Galβ4GlcNAc-). Связывающие Helicobacter pylori соединения могут быть применимыми для подавления связывания лейкоцитов с Helicobacter pylori и предупреждения дыхательного взрыва и/или воспаления, возникающих после активации лейкоцитов.
Известно, что Helicobacter pylori могут связываться с несколькими видами олигосахаридных последовательностей. В некоторой степени связывание определенными штаммами может представлять симбиотические взаимодействия, которые не приводят к развитию опухолей или тяжелых состояний. Настоящие данные по связыванию с сахаридными эпитопами опухолевого типа указывают на то, что связывающее Helicobacter pylori соединение может предупреждать более патологические взаимодействия, при этом давая возможность менее патогенным Helicobacter pylori/штаммам связываться с другими рецепторными структурами. Следовательно, для профилактики заболеваний, вызываемых Helicobacter pylori, может быть необязательным полное удаление бактерий. Менее патогенные бактерии могут даже оказывать пробитическое действие для профилактики состояний, вызванных более патогенными штаммами Helicobacter pylori.
Также понятно, что Helicobacter pylori имеют крупные олигосахариды на основе полилактозамина на их поверхности, которые по меньшей мере у некоторых штаммов содержат нефукозилированные эпитопы, которые могут связываться с бактериями, как описано по изобретению. Описанное здесь соединение также может предупреждать связывание между бактериями Helicobacter pylori и таким образом подавлять, например, колонизацию бактерий.
По изобретению возможно включать связывающее Helicobacter pylori соединение, необязательно с носителем, в фармацевтическую композицию, которая является подходящей для лечения состояния, вызванного присутствием Helicobacter pylori у пациента, или использовать связывающее Helicobacter pylori соединение в способе лечения подобных состояний. Примерами состояний, которые поддаются лечению по изобретению, являются хронический поверхностный гастрит, язва желудка, язва двенадцатиперсной кишки, не-ходжкинская лимфома желудка человека, аденокарционома желудка и некоторые заболевания поджелудочной железы, кожи, печени или сердца, синдром внезапной младенческой смерти, аутоиммунные заболевания, включая аутоиммунный гастрит, и пернициозную анемию, и заболевание желудка, связанное с приемом нестероидных противовоспалительных препаратов (NSAID), все, по меньшей мере, частично вызванные инфицированием Helicobacter pylori.
Фармацевтическая композиция, включающая связывающее Helicobacter pylori соединение, также может содержать другие соединения, такие как инертный растворитель или фармацевтически приемлемые носители, консерванты и т.д., которые хорошо известны специалистам в данной области. Связывающее Helicobacter pylori соединение можно вводить вместе с другими лекарственными препаратами такими, как антибиотики, используемые для лечения Helicobacter pylori.
Связывающее Helicobacter pylori соединение или фармацевтическую композицию, включающую это соединение, можно вводить любым подходящим путем, хотя, предпочтительным является пероральное введение.
Термин «лечение» в том смысле, в котором он здесь используется, относится к лечению с целью излечения или ослабления заболевания или состояния, и к лечению с целью профилактики развития заболевания или состояния. Лечение может быть кратковременным или продолжительным.
Термин «пациент» в том смысле, в котором он здесь используется, относится к человеку или млекопитающему, отличному от человека, при необходимости лечения по изобретению.
Также возможно использовать связывающее Helicobacter pylori соединение для идентификации одного или более адгезинов при скрининге белков или углеводов (углевод-углеводные взаимодействия), которые связываются со связывающим Helicobacter pylori соединением. Белком, связывающимся с углеводом, может быть лектин или фермент, связывающийся с углеводом. Скрининг можно проводить, например, аффинной хроматографией или аффинным сшиванием (Ilver et al., 1998).
Кроме того, возможно использовать соединения, специфические связывающие или инактивирующие Helicobacter pylori, находящиеся в человеческих тканях, и таким образом предупреждать связывание Helicobacter pylori. Примеры таких соединений включают растительные лектины, такие как Erythrina cristagalli и Erythrina corallodendron (Teneberg et al., 1994). При использовании у человека, связывающее соединение должно быть подходящим для подобного применения таким, как гуманизированное антитело или рекомбинантная гликозидаза человека, которая не является иммуногенной и способна отщеплять концевой моносахаридный остаток/остатки от связывающих Helicobacter pylori соединений. Однако в желудочно-кишечном тракте многие естественные лектины и гликозидазы, происходящие, например, из пищи, являются устойчивыми.
Кроме того, возможно использовать связывающее Helicobacter pylori соединение в виде части пищевой композиции, включая продукты питания и корма. Предпочтительно использовать связывающее Helicobacter pylori соединение в виде части так называемого функционального или функционализированного продукта питания. Указанный функциональный продукт питания оказывает положительное влияние на здоровье человека и животных, подавляя или предупреждая связывание Helicobacter pylori с мишеневыми клетками или тканями. Связывающее Helicobacter pylori соединение может быть частью определенного продукта питания или функциональной пищевой композиции. Функциональный продукт питания может включать другие приемлемые пищевые ингредиенты, разрешенные органами такими, как Управление по надзору за пищевыми продуктами и медикаментами в США. Связывающее Helicobacter pylori соединение также можно использовать в виде пищевой добавки, предпочтительно в виде добавки для продукта питания или напитка для получения функционального продукта питания или функционального напитка. Продукт питания или пищевую добавку также можно получать, например, от домашнего животного такого, как корова, или другого животного, продуцирующего связывающее Helicobacter pylori соединение в больших количествах, естественным путем, в молоко. Это можно осуществить, имея животное с сверхэкспрессией подходящих гликозилтрансфераз в молоко. Можно выбрать определенную линию или вид домашнего животного и обеспечить его разведение для продукции в больших количествах связывающего Helicobacter pylori соединения. Связывающее Helicobacter pylori соединение для пищевой композиции или пищевой добавки также можно получить с использованием микроорганизмов таких, как бактерии или дрожжи.
Особенно применимым является получение связывающего Helicobacter pylori соединения в виде составной части продукта питания для детей первого года жизни, предпочтительно в виде питательной смеси для младенцев. Многих детей первого года жизни кормят специальными питательными смесями вместо натурального женского молока. В питательных смесях могут отсутствовать особые олигосахариды, производные лактозы, имеющиеся в женском молоке, особенно удлиненные формы, такие как лакто-N-неотетроза, Galβ4GlcNAcβ3Galβ4Glc, и ее производные. Известно, что лакто-N-неотетроза и пара-лакто-N-неогексоза (Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc), а также Galβ3Galβ4Glc, имеются в женском молоке и, следовательно, их можно рассматривать в качестве безопасных добавок или ингредиентов в продуктах питания для детей первого года жизни. Helicobacter pylori являются особенно инфекционными для младенцев и маленьких детей, и с учетом заболеваний, которые они могут вызвать впоследствии, разумно предупреждать заражение. Также известно, что Helicobacter pylori вызывают синдром внезапной младенческой смерти, а лечение сильными антибиотиками для уничтожения бактерий в случае маленьких детей и младенцев может быть особенно неподходящим.
Предпочтительные концентрации олигосахаридов женского молока в функциональном продукте питания для потребления (например, в восстановленной питательной смеси для младенцев) являются аналогичными таковым, которые имеются в натуральном женском молоке. Следует отметить, что натуральное женское молоко содержит многочисленные свободные олигосахариды и гликоконъюгаты (которые могут быть поливалентными), включая олигосахаридную последовательность(и), описанную в изобретении, следовательно, возможно использовать даже более высокие, чем естественные концентрации отдельных молекул для получения более высокого ингибирующего действия в отношении Helicobacter pylori без проявления отрицательных побочных эффектов. Натуральное женское молоко содержит лакто-N-неотетрозу по меньшей мере в пределах концентраций 10-210 мг/л с отклонениями в отдельных случаях (Nakhla et al., 1999). Следовательно, лакто-N-неотетрозу предпочтительно использовать в функциональном продукте питания в пределах концентраций 0,01-10 г/л, более предпочтительно 0,01-5 г/л, наиболее предпочтительно 0,1-1 г/л. Когда свободные олигосахариды, описанные здесь, являются трисахаридами или дисахаридами с последовательностью Galβ4Glc на восстанавливающем конце, предпочтительно использовать их для потребления в концентрациях 1-100 г/л, более предпочтительно в пределах концентраций 1-20 г/л. Альтернативно общая концентрация сахаридов, используемая в функциональном продукте питания, является такой же или близкой к общей концентрации сахаридов в натуральном женском молоке, которые связываются с Helicobacter pylori, подобно описанным соединениям, или которые содержат связывающий эпитоп/олигосахаридную последовательность, указанную в изобретении. По меньшей мере, в одном источнике сообщалось, что женское молоко содержит Galβ3Galβ4Glc в качестве основного нейтрального олигосахарида в высокой концентрации (Charlwood et al., 1999).
Кроме того, возможно использовать связывающее Helicobacter pylori соединение при диагностике состояния, вызванного Helicobacter pylori. Использования для диагностики также включают применение связывающего Helicobacter pylori соединения для типирования Helicobacter pylori. Когда соединение используется для диагностики или типирования, его можно включить, например, в зонд или палочку для теста, необязательно составляющую часть набора для теста. Когда этот зонд или палочку для теста приводят в контакт с пробой, содержащей Helicobacter pylori, бактерии будут связываться с зондом или палочкой для теста, и таким образом их можно выделить из пробы и подвергнуть дальнейшему анализу.
Полученные данные также показывают, что остаток концевого моносахарида невосстанавливающего конца в предпочтительных трисахаридных последовательностях по изобретению может включать карбоциклическую группу в 6-ом углероде (остатком концевого моносахарида является уроновая кислота, HexA или HexANAc, где Hex представляет Gal или Glc) или производное по 6-ому углероду остатка HexA (NAc) или производное по 6-ому углероду соответствующего остатка Hex(NAc). Подобные концевые остатки предпочтительно включают связанную β3-связью глюкуроновую кислоту и более предпочтительно 6-амиды, такие как ее метиламид. Следовательно, аналоги и производные последовательности можно получить изменением или дериватизацией концевого 6-положения трисахаридных эпитопов.
Предпочтительные связывающие Helicobacter pylori соединения
Было установлено, что олигосахаридные последовательности по изобретению являются неожиданно эффективными связывающими соединениями, когда находятся на тонкослойной поверхности. Данный способ позволяет обеспечить поливалентное представление гликолипидных последовательностей. Удивительно высокая активность поливалентного представления олигосахаридных последовательностей делает поливалентность предпочтительным способом для представления олигосахаридных последовательностей по изобретению.
Гликолипиды в естественных условиях находятся в поливалентной форме на клеточных мембранах. Данный тип представления можно имитировать твердофазным анализом, описанным ниже, или приготовлением липосом из гликолипидов или неогликолипидов.
Настоящие новые неогликолипиды, полученные восстановительным аминированием гидрофобного гексадециланилина, были способы обеспечить эффективное представление олигосахаридов. Большинство ранее известных конъюгатов неогликолипидов, использованных для связывания бактерий, включали отрицательно заряженные группы, такие как фосфорный эфир фосфадитилэтаноламина неогликолипидов. Проблемы подобных соединений заключаются в наличии отрицательного заряда у соединения и естественном биологическом связывании, вовлекающем фосфолипидную структуру. Известно, что отрицательно заряженные молекулы принимают участие в многочисленных неспецифических связываниях с белками и другими биологическими веществами. Кроме того, многие из этих структур являются лабильными и могут разрушаться под действием ферментов или химическим путем. Настоящее изобретение относится к некислым конъюгатам олигосахаридных последовательностей, т.е. олигосахаридных последовательностей, связанных с некислыми химическими структурами. Предпочтительно некислые конъюгаты являются нейтральными, означая, что олигосахаридные последовательности связаны с нейтральными, незаряженными химическими структурами. Предпочтительные конъюгаты по изобретению являются поливалентными соединениями.
На предшествующем уровне в данной области биологически активные олигосахаридные последовательности часто связывали со структурами носителей восстановлением части структуры рецепторного активного олигосахарида. Использовали гидрофобные спейсеры, содержащие алкильные цепи (-СН2-)n и/или бензольные кольца. Однако хорошо известно, что гидрофобные структуры, как правило, участвуют в неспецифических взаимодействиях с белками и другими биологически активными молекулами.
Данные по неогликолипидам примеров, представленных ниже, показывают, что в экспериментальных условиях, использованных в тесте, гексадециланилин в неогликолипидах не приводил к неспецифическому связыванию с тестированными бактериями. В неогликолипидах гексадециланилин конъюгатов, возможно, образует липидный слой, как структуру, и он становится недоступным для связывания. В изобретении показано, что восстановление моносахаридного остатка, относящегося к связывающему эпитопу, может нарушить связывание. В дальнейшем стало понятно, что восстановленный моносахарид можно использовать в качестве гидрофильного спейсера для связывания эпитопа рецептора и поливалентной презентирующей структуры. По изобретению предпочтительно связывать биологически активный олигосахарид посредством гидрофильного спейсера с молекулой поливалентного или мультивалентного носителя с образованием поливалентной или олиговалентной/мультивалентной структуры. Все поливалентные (включающие более 10 олигосахаридных остатков) и олиговалентные/мультивалентные структуры (включающие 2-10 олигосахаридных остатков) относятся здесь к поливалентным структурам, хотя, в зависимости от применения, олиговалентные/мультивалентные конструкции могут быть более предпочтительными по сравнению с более крупными поливалентными структурами. Предпочтительно гидрофильный спейсер включает, по меньшей мере, одну гидроксильную группу. Более предпочтительно, чтобы спейсер включал, по меньшей мере, две гидроксильные группы, и наиболее предпочтительно, чтобы спейсер включал, по меньшей мере, три гидроксильные группы.
По изобретению гидрофильный спейсер предпочтительно представляет собой гибкую цепь, включающую одну или несколько
-СНОН-групп и/или амидную боковую цепь такую, как ацетамидо,
-NHCOCH3, или алкиламидо. Гидроксильные группы и/или ацетамидная группа также защищает спейсер от ферментативного гидролиза в условиях in vivo. Термин «гибкий» означает, что спейсер включает гибкие связи и не образует кольцевую структуру, не обладающую гибкостью. Восстановленные моносахаридные остатки, такие как образуются при восстановительном аминировании по настоящему изобретению, представляют примеры гибких гидрофильных спейсеров. Гибкий гидрофильный спейсер является оптимальным для того, чтобы избежать неспецифического связывания неогликолипида или поливалентных конъюгатов. Это является важной оптимальной активностью, например, при постановке биотестов и для проявления биологической активности фармацевтических композиций или функциональных продуктов питания.
Общая формула конъюгата с гибким гидрофильным линкером имеет следующую формулу 2:
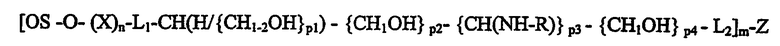
в которой L1 и L2 представляют линкеры, независимо включающие связывающие атомы кислорода, азота, серы или углерода, или два связывающих атома группы, образующей связи, такие как -О-, -S-, -CH2-, -N, -N(COCH3)-, амидные группы -CO-NH- или -NH-CO-, или -N-N-(производное гидразина) или аминокси-связи -O-N- и -N-O-. L1 представляет связь из 1 углерода моносахарида восстанавливающего конца X или, когда n=0, то L1 замещено -O- и связями непосредственно от восстанавливающего конца C1 в OS.
p1, p2, p3 и p4 независимо представляют целые числа 0-7, при условии, что по меньшей мере один из p1, p2, p3 и p4 равен по меньшей мере 1. CH1-2OH в ответвлении {CH1-2OH}р1 означает, что концевая группа цепи представляет CH2OH, и когда p1 больше 1, то имеются вторичные спиртовые группы -CHOH-, связывающие концевую группу с остальной частью спейсера. R предпочтительно представляет ацетильную группу (-COCH3) или R представляет альтернативную связь с Z, и тогда L2 представляет концевую группу цепи из одного или двух атомов, в другом воплощении R является аналогичной образующей группой, включающей С1-4ацил (предпочтительно гидрофильный такой, как гидроксиалкил), содержащий амидоструктуру или Н, или С1-4алкил, образующий амин. И m>1, и Z представляет поливалентный носитель. OS и X имеют значения, определенные для формулы 1.
Предпочтительные поливалентные структуры, содержащие гибкий гидрофильный спейсер, формулы 2 включают связывающую Helicobacter pylori олигосахаридную последовательность (OS)β1-3, связанную через β1-3-связь с Galβ4Glc(красным)-Z, и OSβ6GlcNac(красным)-Z и OSβ6GalNAc(красным)-Z, где «красный» означает структуру с аминной связью, образовавшуюся в результате восстановительного аминирования из моносахаридов восстанавливающего конца и аминогруппы поливалентного носителя Z.
В настоящем изобретении олигосахаридная группа предпочтительно связана в поливалентной или олиговалентной формой с носителем, которая не является белком или пептидом для избежания проявления антигенности и возможных аллергических реакций, предпочтительно скелет представляет натуральный неантигенный полисахарид.
При сравнении активности связывания гликолипидов и неогликолипидов было установлено, что последовательности с Galα3Galβ- обладали меньшей активностью при поливалентном представлении на тонкослойной пластинке. Последовательности с концевой последовательностью Galβ4GlcNAc- были также слабее. Следовательно, оптимальное поливалентное некислотное соединение по изобретению включает концевую олигосахаридную последовательность
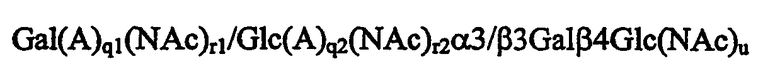
в которой q1, q2, r1, r2 и u каждый независимо равен 0 или 1, при условии, что когда оба q1 и r1 равны 0, то тогда концевой моносахаридный остаток невосстанавливающего конца не является Galα. Более предпочтительно u=0, и наиболее предпочтительно олигосахаридная последовательность, представленная в поливалентной форме, представляет
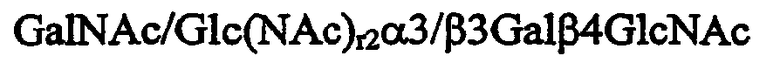
в которой r2 независимо равен 0 или 1, и ее аналог или производное.
Последующие последовательности являются особенно предпочтительными. Таковые представляют структуры, которые не описаны для тканей человека и животных:
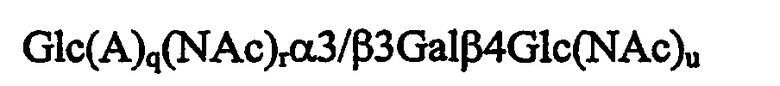
при условии, что когда олигосахаридная последовательность включает β3-связь, то q и r равны 1 или 0; или
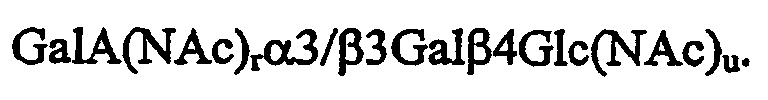
Новизна вышеуказанных олигосахаридных последовательностей делает их особенно предпочтительными. Гликозидазы, расщепляющие подобные последовательности, неизвестны. Следовательно, последовательности являются особенно стабильными и предпочтительными в биологических условиях. За счет естественного типа последовательностей, описанных по изобретению, они могут расщепляться гликозидазами, что снижает их применимость, особенно при использовании у человека и животных. Известно, что гликозидазы, расщепляющие последовательности, проявляют активность в желудочно-кишечном тракте человека. Описано несколько гликозидаз таких, как N-ацетилгексозаминидазы или галактозидазы, в качестве расщепляющих ферментов, и также они находятся в продуктах питания.
Понятно, что новые соединения по изобретению также пригодны для ингибирования токсина А Clostridium difficile S. Teneberg et al., 1996. Характер связывания токсина А с уже известными соединениями в значительной степени аналогичен специфичности Helicobacter pylori, описанной здесь. Таким образом, связывающие Helicobacter pylori соединения можно использовать для лечения, например, вызванной Clostridium difficile диареи.
Номенклатура гликолипидов и углеводов используется здесь согласно рекомендациям Комиссии по биохимической номенклатуре IUPAC-IUB (Carbohydrate Res. 1998, 312, 167; Carbohydrate Res. 1997, 297, 1; Eur. J. Biochem. 1998, 257, 29).
Полагается, что Gal, Glc, GlcNAc и Neu5Ac имеют D-конфигурацию, Fuc - L-конфигурацию, и все моносахаридные звенья находятся в форме пиранозы. Глюкозамин обозначен, как GlcN или GlcNH2, и галактозамин - как GalN или GalNH2. Гликозидные связи показаны частично в более краткой и частично в более длинной номенклатуре, связи Neu5Ac-остатков α3 и α6 представляют тоже, что соответственно α2-3 и α2-6, и для других моносахаридных остатков α1-3, β1-3, β1-4 и β1-6 могут быть обозначены короче соответственно в виде α3, β3, β4 и β6. Лактозамин относится к N-ацетиллактозамину, Galβ4GlcNAc, и сиаловая кислота относится к N-ацетилнейраминовой кислоте (Neu5Ac) или N-гликолилнейраминовой кислоте (Neu5Gc), или любой другой естественной сиаловой кислоте. Термин «гликан» в широком смысле означает здесь олигосахаридные или полисахаридные цепи, имеющиеся в гликоконъюгатах человека или животных, особенно в гликолипидах или гликопротеинах. В краткой номенклатуре жирных кислот и оснований число перед двоеточием означает число атомов углерода в цепи, а число после двоеточия - общее число двойных связей в углеводной цепи. Сокращение GSL относится к гликосфинголипиду. Сокращения или краткие названия, или обозначения гликосфинголипидов представлены в тексте и таблицах 1 и 2. Helicobacter pylori относится также к бактериям, подобным Helicobacter pylori.
В настоящем изобретении гекс(NAc)-уроновая кислота и ее производные, и остатки обозначены следующим образом: GlcA представляет глюкуроновую кислоту и производные по 6 углероду глюкозы или глюкуроновой кислоты, GalA представляет галактуроновую кислоту и производные по 6 углероду галактозы или галактуроновой кислоты, GlcANAc представляет N-ацетилглюкуроновую кислоту и производные по 6 углероду N-ацетилглюкозамина или представляет уроновой кислоты N-ацетилглюкозамин, и GalANAc представляет уроновой кислоты N-ацетилгалактозамин или производные по 6 углероду N-ацетилгалактозамина или уроновой кислоты N-ацетилгалактозамина.
Выражение «концевая олигосахаридная последовательность» указывает, что олигосахарид не замещен по концевому остатку невосстанавливающего конца другим моносахаридным остатком.
Термин «α3/β3» означает, что смежные остатки в олигосахаридной последовательности могут быть соединены друг с другом α3- или β3-связью.
Далее настоящее изобретение иллюстрируется последующими примерами, которые никоим образом не ограничивают объем изобретения.
Примеры
Материалы и методы
Материалы. Пластинки для ТСХ с силикагелем 60 (на алюминиевой подложке) получали от Merck (Darmstadt, Германия). Все испытуемые гликосфинголипиды были получены в лаборатории заявителей. β-галактозидазу (Escherichia coli) получали от Boehringer Mannheim (Германия), среду Ham's F12 - от Gibco (Великобритания), 35S-метионин - от Amersham (Великобритания) и FCS (фетальную телячью сыворотку) - от Sera-Lab (Англия). β4-галактозидаза (Streptococcus pneumoniae), β-N-ацетилгексоаминидаза (Streptococcus pneumoniae) и сиалидаза (Arthrobacter ureafaciens) были от Oxford GlycoSystems (Abington, Великобритания). Клинические изоляты Helicobacter pylori (штаммы 002 и 032), полученные от пациентов с соответственно гастритом и язвой двенадцатиперстной кишки, были безвозмездно предоставлены Dr. D. Danielsson, Örebro Medical Center, Швеция. Штамм типа 17875 был из Культуральной коллекции Университета Гетеборг (CCUG).
Гликосфинголипиды. Чистые гликосфинголипиды для опытов, представленных на фиг.7А и 7В, получали из цельной кислотной или некислотной фракции из источников, перечисленных в таблице 2, как описано (Karlsson, 1987). В основном отдельные гликосфинголипиды получали ацетилированием (Handa, 1963) цельных фракций гликосфинголипидов и разделяли многократной колоночной хроматографией в силикагеле и затем структуру подтверждали масс-спектрометрией (Samuelsson et at., 1990), ЯМР (Falk et al., 1979 a, b, c; Koerner Jr. et al., 1983) и методами разложения (Yang и Hakomori, 1971; Stellner et al., 1973). Гликолипиды, выделенные из тимуса кролика, описаны ниже.
Выделение гликолипидов. Кислые гликосфинголипиды выделяли из 1000 г ацетонового порошка тимуса кролика (Pel-Freeze Biological Inc., North Arkansas, Ark. США). Ацетоновый порошок экстрагировали в аппарате Сокслета смесью хлороформ/метанол 2/1 (об./об., если не указано иначе) в течение 24 ч с последующей экстракцией смесью хлороформ/метанол/вода 8/1/1 в течение 36 ч. Экстрагированные липиды, 240 г, подвергали разделению по Фольчу (Folch et al., 1957) и собранную гидрофильную фазу - для ионообменной гель-хроматографии на целлюлозе DE23 (DEAE, Whatman, Maidstone, Великобритания). В результате этих стадий выделения получали 2,5 г кислых гликосфинголипидов. Ганглиозиды разделяли соответственно числу сиаловых кислот ионообменной гель-хроматографией на стеклянной колонке (внутренний диаметр 50 мм). Колонку подсоединяли к насосу для ВЭЖХ, получая непрерывный градиент (заранее запрограммированный градиент № 4, System Gold Chromatographic Software, Beckman Instruments Inc., CA, США), начиная с метанола и заканчивая 0,5 M CH3COONH4 в метаноле. Скорость потока составляла 4 мл/мин и собирали 200 фракций по 8 мл каждая. 300-400 мг смеси ганглиозидов наносили на 500 г сефарозы DEAE (CL6, Pharmacia, Uppsala, Швеция, высота примерно 130 мм). Моносиалилированные ганглиозиды затем разделяли ВЭЖХ на колонке с силикагелем, 300 мм × 22 мм внутренний диаметр, с размером пор 120 Å, с размером частиц 10 мкм (SH-044-10, Yamamura Ltd., Kyoto, Япония). Примерно 150 мг моносиалилированных ганглиозидов наносили и использовали прямой градиент элюирования (хлороформ/метанол/вода от 60/35/8 до 10/103, 4 мл/мин, 240 фракций).
Частичный кислотный гидролиз. Десиалилирование ганглиозидов проводили 1,5% CH3COOH в воде при 100°C, после чего вещество нейтрализовали NaOH и высушивали в атмосфере азота. Для частичного расщепления углеводного скелета гликолипиды гидролизовали 0,5 M HC1 в течение 7 мин на кипящей водяной бане. Затем вещество нейтрализовали и распределяли в системе C/M/H2O (8:4:3, об./об.). Собирали нижнюю фазу, выпаривали в атмосфере азота и выделенные гликолипиды использовали для анализа.
Получение пентагликозилкерамида из гексагликозилкерамида ферментативным гидролизом. Гексагликозилкерамид (структура 2, таблица 1), полученный из гептагликозилкерамида (4 мг, из тимуса кроликов)(структура 1, таблица 1) кислотным десиалилированием (смотри выше), повторно растворяли в смеси С/М (2:1) и наносили на небольшую колонку с силикагелем (0,4×5 см). Колонку элюировали смесью C/M/H2O (60:35:8, об./об.). Собирали фракции объемом примерно 0,2 мл и анализировали на присутствие углеводов. Полученный гексагликозилкерамид (2,0 мг) растворяли в 1,5 мл 0,1 М фосфатного калиевого буфера, рН 7,2, содержащего тауродезоксихолат натрия (1,5 мг/мл), MgCl2 (0,001 M) и β-галактозидазу (E.coli, 500 Е при анализе с использованием 2-нитрофенил-β-D-галактозида в качестве субстрата) и пробу инкубировали в течение ночи при 37°C. Затем вещество распределяли в системе C/M/H2O (10:5:3) и гликолипид, находящийся в нижней фазе, очищали хроматографией в силикагеле (колонки 0,4×5 см), как описано выше для гексагликозилкерамида. Для удаления всех загрязняющих детергентов хроматографию повторяли дважды. Конечное извлечение пентагликозилкерамида составляло 0,7 мг.
Расщепление гликолипидов эндогликокерамидазой (Ito и Yamagata, 1989). Реакционная смесь состояла из 200 мкг гликолипидов, 80 мкг тауродезоксихолата натрия и 0,8 мЕ фермента в 160 мкл 50 мМ ацетатного буфера, рН 6,0. Пробу инкубировали в течение ночи при 37°C, после чего добавляли воду (140 мкл) и смесь C/M (2:1, объемные соотношения, 1500 мкл), пробу встряхивали и центрифугировали. Верхнюю фазу высушивали в атмосфере азота, повторно растворяли в небольшом объеме воды и обессоливали на колонке с Сефадексом G-25 (0,4×10 см), которую уравновешивали H2O и элюировали водой. Собирали фракции объемом примерно 0,1 мл и анализировали на присутствие сахаров.
Перметилирование сахаридов. Перметилирование проводили по методу Larson et al., 1987. К пробам добавляли гидроокись натрия перед внесением йодистого метила, как предложено Needs и Selvendran, 1993. В некоторых опытах сахариды восстанавливали NaBH4 перед метилированием. В этом случае количество йодистого метила увеличивали до конечного соотношения ДМСО (диметилсульфоксид)/йодистый метил, равного 1:1 (Hansson and Karlsson, 1990).
Газовая хроматография/масс-спектрометрия. Газовую хроматографию проводили на газовом хроматографе Hewlett-Packard 5890A Series II, снабженном инжектором и пламенно-ионизационным детектором. Перметилированные олигосахариды анализировали на сплавленной капиллярной колонке с силикагелем (Fluka, 11 м × 0,25 мм внутренний диаметр), покрытым поперечно-сшитым PS264 (толщина слоя 0,03 мкм). Пробу растворяли в этилацетате и вводили на колонку при 80°C. Температуру программировали в пределах от 80°C до 390°C со скоростью возрастания 10°C/мин и выдерживали в течение 2 мин при верхней температуре. Газовую хроматографию-масс-спектрометрию перметилированных олигосахаридов проводили на газовом хроматографе Hewlett-Packard 5890A Series II, соединенным с масс-спектрометром JEOL SX-102 (Hansson и Karlsson, 1990). FAB-MC проводили на масс-спектрометре JEOL SX-102. Получали FAB-спектр отрицательных ионов с использованием бомбардировки атомами Xe (10 кВ) и триэтаноламина в качестве матрицы.
ЯМР-спектроскопия. Протонные ЯМР-спектры получали при 11,75 Т на спектрометре Jeol Alpha 500 (Jeol, Tokyo, Япония). Пробы дейтерировали перед анализом и затем записывали спектры при 30°С с цифровым разрешением 0,35 Гц/pt. Химические сдвиги представлены относительно TMS (тетраметилсилана) с использованием внутреннего сигнала растворителя.
Ферментативный анализ. Ферментативные тесты Oxford GlycoSystems проводили согласно рекомендациям изготовителя за исключением того, что в каждую инкубационную смесь вносили тритон Х-100 до конечной концентрации 0,3%. Когда для расщепление проводили смесью сиалидазы и β4-галактозидазы использовали буфер для инкубации из набора для β4-галактозидазы. Если в смеси для расщепления находилась β-гексозаминидаза, использовали буфер из того же ферментативного набора. Концентрации ферментов в инкубационных смесях составляли: 80 мЕ/мл для Hexβ4HexNAc-галактозидазы (S.pneumoniae), 120 мЕ/мл для β-N-ацетилгексозаминидазы (S.pneumoniae) и 1 Е/мл для сиалидазы (Arthrobacter ureafaciens). Концентрация субстрата была равной примерно 20 мкМ. Ферментативное расщепление проводили в течение ночи при 37°С. После расщепления пробы высушивали и обессоливали с использованием небольших колонок с сефадексом G-25 (Wells и Dittmer, 1963), 0,3 г, уравновешенных смесью С/М/Н2О (60:30:4,5, объемные соотношения). Каждую пробу наносили на колонку в 2 мл того же растворителя и элюировали 2,5 мл смеси С/М/Н2О (60:30:4,5) и 2,5 мл смеси С/М (2:1). Собирали нанесенные и элюированные растворы, и выпаривали в атмосфере азота.
Другие аналитические методы.Гексозу определяли по методу Dubois et al., 1956.
Де-N-ацилирование. Превращение ацетамидной группы остатков GlcNAc/GalNAc в амин проводили обработкой различных гликосфинголипидов безводным гидразином, как описано ранее (Engström et al., 1998).
Культивирование бактерий.Штаммы Helicobacter pylori хранили при -80°С в триптическом соевом бульоне, содержащем 15% глицерина (по объему). Первоначально бактерии культивировали на агаре GAB-CAMP (Soltesz et al., 1988) во влажных (98%) микроаэрофильных условиях (О2: 5-7%, СО2: 8-10% и N2: 83-87%) при 37°С в течение 48-72 ч. Для введения радиоактивной метки колонии наносили на агар GAB-CAMP за исключением данных, представленных на фиг.1А и 1В, где вместо него использовали агар Brucella (Difco, Detroit, MI) и 50 мккюри 35S-метионина (Amersham, Великобритания), растворенного в 0,5 мл забуференного фосфатом физиологического раствора (PBS), рН 7,3, наносили на чашки. После инкубации в течение 12-24 ч при 37°С в микроаэрофильных условиях, клетки соскребали, три раза отмывали PBS и ресуспендировали до титра 1×108 КОЕ/мл в PBS. Альтернативно колонии высевали (1×105 КОЕ/мл) в среду Ham's F12 (Gibco BPL, Великобритания) с добавлением 10% инактивированной нагреванием фетальной телячьей сыворотки (Sera-Lab). Для введения радиоактивной метки добавляли 50 мккюри 35S-метионина на 10 мл среды и инкубировали при встряхивании в микроаэрофильных условиях в течение 24 ч. Бактериальные клетки собирали центрифугированием и чистоту культур и низкое содержание кокковых форм подтверждали фазово-контрастной микроскопией. После двух промываний PBS клетки ресуспендировали до титра 1×108 КОЕ/мл в PBS. Проведение обеих методик введения метки давало суспензии с удельной активностью, равной примерно 1 импульс в минуту на 100 микроорганизмов Helicobacter pylori.
ТСХ с нанесением бактерий.Тонкослойную хроматографию проводили на пластинках для ТСХ с силикагелем 60 на стеклянной и алюминиевой подложке HPTLC (Merck, Darmstadt, Германия) с использованием смеси хлороформ/метанол/вода 60:35:8 (объемные соотношения) в качестве системы растворителей. Проявление проводили окрашиванием анисовым альдегидом (Waldi, 1962). Тест с нанесением бактерий проводили, как описано ранее (Hansson et al., 1985). Гликосфинголипиды (1-4 мкг/полосу или как указано в надписи к фигурам) хроматографировали на пластинках с силикагелем на алюминиевой подложке и затем обрабатывали 0,3-0,5% полиизобутилметакрилатом в смеси диэтиловый эфир/н-гексан 1:3 (объемные соотношения) в течение 1 мин, высушивали и затем замачивали в PBS, содержащем 2% бычьего сывороточного альбумина и 0,1% твина-20 на 2 ч. Суспензию меченных радиоактивным изотопом бактерий (разбавленную PBS до титра 1×108 КОЕ/мл и 1-5×106 импульсов в минуту/мл) наносили на хроматограммы и инкубировали в течение 2 ч с последующим многократным промыванием PBS. После высушивания хроматограммы покрывали рентгеновской пленкой XAR-5 (Eastman Kodak Co., Rochester, NY, США) на 12-72 ч.
ТСХ с нанесением белков. Введение метки 125I в моноклональные антитела ТН2 и лектин из Erythrina crisragalli (Vector Laboratories, Inc., Burlingame, CA) проводили методом Iodogen (Aggarwal et al., 1985), получив в среднем 2×103 импульсов в минуту/мкг. Методику нанесения проводили так же, как описано выше для бактерий, за исключением того, что не использовали твин, и 125I-меченный белок, разбавленный примерно до 2×138 импульсов в минуту/мкл PBS, содержащего 2% бычьего сывороточного альбумина, использовали вместо бактериальной суспензии.
Молекулярное моделирование. Рассчитывали конформеры с минимальной энергией гликосфинголипидов, перечисленных в таблице 1, с помощью программы для молекулярного моделирования Biograf (Molecular Simulations Inc.) с использованием силового поля Dreining-II (Mayo et al., 1990) на Silicon Graphics 4D/35TG. Получали частичные атомные заряды с использованием метода уравновешивания зарядов (Rappé и Goddard III, 1991) и диэлектрическую проницаемость, зависимую от расстояния (ε=3,5r), использовали для взаимодействий Coulomb. Кроме того, использовали специальные водородные связи, при которых максимальное взаимодействие (Dhb) было установлено на -4 ккалмоль-1 . Дигедральные углы связи GlcβCer определяли следующим образом: Φ=Н-1-С-1-О-1-С-1, ψ=С-1-О-1-С-1-С-2 и θ=О-1-С-1-С-2-С-3, начиная с конца глюкозы (смотри Nyholm и Pascher, 1993).
Олигосахарид GlcNAcβ3Galβ4GlcNAc синтезировали из Galβ4GlcNAc (Sigma, St. Louris, США) и
GlcNAcβ3Galβ4GlcNAcβ6GlcNAc синтезировали из Galβ4GlcNAcβ6GlcNAc при инкубации акцепторного сахарида с β3-N-ацетилглюкозаминилтрансферазой сыворотки крови чеовека и UDP-GlcNAc в присутствии 8 мМ MnCl2 и 0,2 мг/мл АТФ при 37°С в течение 5 суток в 50 мМ Трис-HCl, рН 7,5. Galβ4GlcNAcβ6GlcNAc получали из GlcNAcβ6GlcNAc (Sigma, St. Louis, США) при инкубации дисахарида с β4-галактозилтрансферазой (из коровьего молока, Calbiochem, CA, США) и UDP-Gal в присуствии 20 мМ MnCl2 в течение нескольких часов в 50 мМ MOPS-NaOH, рН 7,4. Гексасахарид Galβ3GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc (1 мг, Dextra labs., Великобритания) обрабатывали 400 мЕ β3/6-галактозидазы (Calbiochem., CA, США) в течение ночи, следуя рекомендациям изготовителя. Олигосахариды очищали хроматографией и их чистоту оценивали масс-спектрометрией MALDI-TOF и ЯМР. Galα3Galβ4GlcNAcβ3Galβ4Glc был от Dextra laboratories, Reading, Великобритания. Гликолипид GlcAβ3Galβ4GlcNAcβ3Galβ4GlcβCer (Wako Pure Chemicals, Osaka, Япония) восстанавливали в Glcβ3Galβ4GlcNAcβ3Galβ4GlcβCer, как описано Lanne et al., 1995. Производное гликолипида
Glc(А-метиламид)β3Galβ4GlcNAcβ3Galβ4GlcβCer получали
амидированием карбоксильной группы глюкуроновой кислоты GlcAβ3Galβ4GlcNAcβ3Galβ4GlcβCer, как описано Lanne et al., 1995.
Результаты
Гептагликозилкерамид NeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer выделяли из тимуса кролика ВЭЖХ, как описано выше. Структуру характеризовывали ЯМР и масс-спектрометрией (данные не представлены). Ганлиозид на основе гептасахарида связывался с большинством изолятов Helicobacter pylori (примерно 60), тестированных в лаборатории заявителей.
Для того чтобы детектировать возможные минорные изомерные компоненты в веществе гептагликозилкерамида, ганглиозид десиалилировали, обрабатывали эндогликокерамидазой, после чего выделенные олигосахариды перметилировали и анализировали газовой хроматографией и EI/MC (фиг.1А и 1В). Установили присутствие двух сахаридов в области, соответствующей шести сахарам, что указывает на предполагаемую углеводную последовательность Hex-HexNAc-Hex-HexNAc-Hex-Hex, что подтверждали фрагментными ионами при m/z 219, 464, 668, 913 и 1118. Когда углеводы превращали в альдитолы (восстановлением NaBH4) перед метилированием, то обнаружили наличие отдельных фрагментных ионов при m/z 235, 684 и 1133 в дополнении к ранее перечисленным ионам (данные не представлены). Для преобладающего сахарида, составляющего 90% от целого вещества (пик B, фиг.1A и 1B), был характерен высокий фрагментный ион при m/z 182, что подтверждает наличие β4GlcNAc (неолактосоединение, углеводная цепь типа 2). Минорный сахарид (пик А, фиг.1А и 1В) давал спектр, характерный для цепи типа-1 (лактосоединение) с очень низким фрагментным ионом при m/z 182 и высоким фрагментным ионом при m/z 228. Препарат также содержал следы других, дающих положительную реакцию на сахара соединений, которые могли представлять 4- и 5-сахарсодержащие сахариды тех же видов. Фукозусодержащие сахариды не были обнаружены в смеси. Чистоту асиалоганглиозидов также определяли FAB/MC и ЯМР-спектроскопией. FAB/MC отрицательных ионов гексагликозилкерамида (фиг.2А) подтверждала предсказанную углеводную последовательность и показывала, что керамиды в основном состояли из сфингозина и C16:0 жирных кислот (m/z 536,5). Полученный ЯМР-спектр гексагликозилкерамида (фиг.3А) показывал четыре основных дублета в аномерной области с β3-связями (J˜8 Гц). Они имели соотношение интенсивности 2:2:1:1. Сигнал при 4,655 ppm (GlcNAcβ3), 4,256 ppm (внутренняя Galβ4), 4,203 ppm (концевая Galβ4) и 4,166 ppm (Glcβ) согласовывались с ранее опубликованными данными для nLcOse6-Cer (Clausen et al., 1986). Имел место также небольшой дублет при 4,804 ppm, который вместе с небольшим сигналом метила при 1,81 ppm (виден в виде плеча на большом резонансе метила типа 2) указывали на присутствие небольшой фракции цепи типа 1. За счет перекрывания в области 4,15-4,25 ppm невозможно было определить положение и распределение связи данного типа 1. Общее количество связи типа 1 приблизительно составляло 10%. Поскольку количество цепи типа 1 в пентагликозилкерамиде, полученном из гексагликозилкерамида расщеплением β-галактозидазой, также составляло примерно 5% (фиг.3В), кажется вероятным, что связь типа 1 равномерно распределялась между внутренней и внешней частями сахаридной цепи, т.е. по 5% гликолипидов могло быть типа 1 и типа 2.
Для исследования того, связана ли активность связывания гликолипидов с преобладающей неолакто (тип 2) структурой, асиалогликолипид обрабатывали β4-галактозидазой и β-гексозаминидазой, и продукты анализировали ТХС с нанесением бактерий (фиг.4А, 4В и 4С). Как и предполагалось, первый фермент превращал гексагликозилкерамид в пентагликозилкерамид (4А, полоса 3) и смесь двух ферментов расщепляла вещество до лактозилкерамида (4В, полоса 6). При визуальной оценке пластинок для ТСХ обе реакции протекали полностью или почти полностью. Такие же результаты получали при обработке вещества сиалидазой и кислотой. Расщепление гексагликозилкерамида β4-галактозидазой сопровождалось потерей связывающей Helicobacter pylori активности в области данного гликолипида на пластинках для ТСХ с одновременным появлением высокой активности в области пентагликозилкерамидов (4С, полоса 3). Дальнейшее ферментативное расщепление пентагликозилкерамида приводило к исчезновению связывающей активности в данной области. Не наблюдали появления связывающей активности в области, соответствующей четырем сахарам. Чувствительность химического окрашивания пластинок ТСХ была слишком низкой для того, чтобы детектировать соединения, находящиеся в следовых количествах.
В отдельном опыте исходный ганглиозид подвергали частичному кислотному расщеплению и высвобожденные гликолипиды тестировали на активность связываться с Helicobacter pylori. На фиг.5А и 5В показаны результаты ТСХ гидролизата (5А) и соответствующая авторадиограмма (5В) после нанесения на гидролизат 35S-меченных Helicobacter pylori. Гликолипиды, расположенные в областях гекса-, пента-, тетра- и дигликозилкерамидов, проявляли связывающую активность, в то время, как тригликозилкерамид был неактивным.
Связывание гекса-, пента-, тетрагликозилкерамидов было аналогичным при тестировании, по меньшей мере, с тремя штаммами Helicobacter pylori (17875, 002 и 032).
Высокую активность связывания пентагликозилкерамида, полученную после удаления концевой галактозы из гексагликозилкерамида и очистки хроматографией в силикагеле, исследовали более детально. Спектр FAB/MC отрицательных ионов данного гликолипида подтвердил наличие углеводной последовательности HexNAc-Hex-HexNAc-Hex-Hex- и показывал тот же состав керамидов, что и для гексагликозилкерамида (фиг.2В). В протонном ЯМР-спектре, полученном для пентагликозилкерамида (фиг.3В), было пять основных β-дублетов в аномерной области: при 4,653 ppm (внутренний GlcNAcβ3), 4,615 ppm (концевой GlcNAcβ3), 4,261 ppm (двойная интесивность, внутренняя Galβ4), 4,166 (Glcβ), согласующихся со структурой GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer, и также они полностью соответствовали соединению из шести сахаров с удаленной концевой Galβ. Также имел место небольшой β-дублет при 4,787, соответствующий 3-замещенному GlcNAcβ (цепь типа 1). Предполагаемый сигнал метила был также виден в виде плеча на значительно большем сигнале метила при 1,82 ppm, но перекрывание не давало возможности оценить эти сигналы количественно. По интегралу аномерного протона можно было высчитать, что 6% гликолипида включало цепь типа 1. Таким образом, относительное соотношение углеводных цепей типа 2 и типа 1 было аналогичным таковому для гликолипида из шести сахаров. Два пятна, видимые на пластинках для ТСХ, соответствующие гекса- и пентагликозильным фракциям, в большей степени отражали гетерогенность керамидов, чем нежели различия в составе сахарной цепи, о чем судили по их чувствительности к воздействию β4-галактозидазы. Верхнее пятно в пентагликозильной области появлялось как после неизбирательного гидролиза асиалоганглиозида, так и при избирательном отщеплении 4-связанной галактозы от асиалопродукта. Кроме того, когда гексагликозилкерамид с высоким содержанием верхних хроматографических субфракций расщепляли с помощью β4-галактозидазы и β-гексоаминидазы, то полученный лактозилкерамид давал две различные хроматографические полосы. Хроматографически гомогенный гексагликозилкерамид давал только одну полосу лактозилкерамида. Как верхние, так и нижние субфракции в области, соответствующей пентасахаридам, были высокоактивными, как было показано в тестах с нанесением бактерий.
Гликосфинголипиды неолактосоединений с 6,5 и 4 сахарами (структуры 2, 4 и 5, таблица 1) оценивали полуколичественными тестами при использовании ТСХ с нанесением бактерий. Гликолипиды наносили на пластинки с силикагелем в серийных разведениях и их связывание с Helicobacter pylori оценивали визуально после нанесения меченых бактерий и авторадиографией (фиг.6А и 6В). Наиболее активным видом соединений был пентагликозилкерамид, который давал положительную реакцию на пластинках для ТСХ в количествах до 0,039 нмоль/пятно (среднее значение из 7 опытов, стандартное отклонение δn-1=0,016 нмоль). Гекса- и тетрагликозилкерамиды связывали Helicobacter pylori, находясь в количестве соответственно примерно 0,2 и 0,3 нмоль гликолипида/пятно.
Связывание Helicobacter pylori с высшими гликолипидами изученных видов было в высокой степени воспроизводимым. Частота связывания Helicobacter pylori, штамма 0,32, составляла для пентагликозил- и гексагликозилкерамидов ˜90% (общее число пластинок равнялось примерно 100).
Анализ связывания, выявляющий изорецепторы и специфичность связывания (фиг.7А и 7В).
В дополнение к гликосфинголипиду из семи сахаров из тимуса кроликов, имеющего неолактоядро
NeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer, и полученным из него тетра-гексагликозилкерамидам, специфичность связывания могла включать другие гликолипиды из неолактосоединений.
Связывание Helicobacter pylori (штамм 032) с очищенными гликосфинголипидами, разделенными на пластинках для ТСХ с нанесением бактерий, представлено на фиг.7А и 7В. Эти данные вместе с таковыми, полученными для дополнительного ряда очищенных гликосфинголипидов, обобщены в таблице 2. Связывание Helicobacter pylori с неолактотетраозилкерамидом (полоса 1) и гликосфинголипидами из пяти и шести сахаров (полосы 5 и 6), полученными из NeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer, было аналогичным вышепредставленным результатам. Однако неожиданно было также установлено связывание для GalNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer (x2 гликосфинголипида, полоса 7) и дефукозилированного гликосфинголипида А6-2 GalNAcα3Galβ4GlcNAcβ3Galβ4GlcβCer (номер 12, таблица 2). В совокупности с установлением того, что Galα3Galβ4GlcNAcβ3Galβ4GlcβCer (гликосфинголипид В5, полоса 2) также является активным в отношении связывания, можно предположить в большей степени о наличии перекрестного связывания, чем о наличии многих адгезинов, специфических для каждого из гликосфинголипидов (смотри ниже). Кроме того, единственным удлинением только что упомянутых различных гликосфинголипидов, содержащих пять сахаров, которое не оказывало влияния на связывание с бактериальным адгезином, было добавление Galβ4 к полученному из тимуса соединению с GlcNAcβ3-концом (полоса 6). Было установлено, что другие удлиненные структуры, такие как NeuAc-х2 (полоса 8) и GalNAcβ3-B5 (номер 25, таблица 2) были неактивными в отношении связывания. Следует также отметить, что ацетамидная группа внутреннего GlcNAcβ3 в В5 важна для связывания, поскольку де-N-ацилирование этой группы обработкой безводным гидразином приводит к полной потере связывания (полоса 3), как и в случае аналогичной обработки неолактотетраозилкерамида (номер 6, таблица 2).
Перекрестное связывание гликосфинголипидов из пяти сахаров.Для того чтобы понять характер связывания различных молекул гликосфинголипидов на основе неолактосоединений, использованных в данном исследовании, изучали конформационные предпочтения активных, а также неактивных структур с помощью молекулярного моделирования. На фиг.8А, 8В, 8С и 8D показано, что х2-гликосфинголипиды вместе с тремя другими последовательностями: дефукозилированным А6-2, В5 и де-N-ацилированным В5, за исключением химически модифицированной структуры В5, обладают аналогичной активностью связывания. Также в данное сравнительное исследование следовало включить гликосфинголипид из пяти сахаров, выделенный из тимуса кролика (смотри фиг.9А), поскольку данная структура отличается только по четвертому положению концевого остатка по сравнению с х2-структурой, и было установлено, что он был также в равной степени активен. Все четыре активные структуры имеют неолактоядро, которое заканчивается соответственно GalNAcβ3, GalNAcα3, Galα3 и GlcNAcβ3. Конформеры этих структур с минимальной энергией получали, как описано ранее (Teneberg et al., 1996). Другие структуры с минимальной энергией, представленные в таблице 2, основаны на ранее полученных данных, имеющихся в литературе (Bock et al., 1985; Meyer, 1990; Nyholm et al., 1989). В отношении гликосфинголипидов, с сиаловой кислотой на конце, то была получена синклинальная конформация для гликозидных дигедральных углов α3-связанных остатков, как показано, например, на фиг.9С, но также оценивали влияние других конформаций (Siebert et al., 1992), в частности, антиклинальной. Также для той же цели получали конформеры с низкой энергией для нескольких α6-связанных вариантов (Breg et al., 1989).
Как указано выше, на основании того факта, что в состав всех четырех активных в отношении связывания гликосфинголипидов из пяти сахаров (номера 10-13, таблица 2), входило неолактоядро, можно предположить, что объяснением этих наблюдений может быть наличие перекрестного связывания с одним и тем же сайтом адгезина. Однако на первый взгляд кажется удивительным, что гликосфинголипид В5, который отличается по концевому положению от соединения из пяти сахаров, полученного из тимуса кролика, первый имеет Galα3, а последний - GlcNAcβ3, является в равной степени активным, и это следует иметь в виду при рассмотрении специфичности связывания неолактосоединений. Несмотря на то, что эти два концевых сахарида различаются также по их аномерной связи, видно (фиг.8С и 9А), что структуры с минимальной энергией топографически очень сходны, различия заключаются в том, что в Galα3 отсутствует ацетамидная группа и имеется 4-ОН в аксиальном положении, и плоскость ее кольца несколько приподнята по сравнению с соответствующей плоскостью в соединении из пяти сахаров. Однако оказалось, что ни положение 4-ОН, ни отсутствие/наличие ацетамидной группы не является решающим для связывания, поскольку также х2 и дефукозилированные гликосфинголипиды А6-2 (фиг.8А, В), которые заканчиваются соответственно GlcNAcβ3 и GalNAcα3, обладают сходными аффинностями в отношении адгезина Helicobacter pylori. В свете этих фактов также следует ожидать, что Galβ3Galβ4GlcNAcβ3Galβ4GlcβCer, который был выделен из эритроцитов человека (Stellner и Hakomori, 1974) будет связываться с бактериальным адгезином. Также в свете закономерностей связывания три другие концевые моносахариды в связывающих Helicobacter pylori эпитопах представляют возможные трисахаридные связывающие эпитопы, а именно GlcNAcα3Galβ4GlcNAc, Glcβ3Galβ4GlcNAc и Glcα3Galβ4GlcNAc. До сих пор подобные соединения неизвестны для тканей человека, и они в большей степени могут представлять аналоги естественного рецептора. К сожалению, заявители не располагали ни Galβ3Galβ4GlcNAc-гликолипидом, ни тремя аналогами для тестирования.
Неолактосоединение из семи сахаров,
NeuGlcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer также подвергали молекулярному моделированию. На фиг.10 представлены две различные проекции структуры с минимальной энергией со связью GlcβCer в удлиненной конформации. Сиаловая кислота находилась в синклинальной конформации, но антиконформеры, вероятно, также представляют неразветвленные структуры (Siebert et al., 1992). Оказалось, что сиаловая кислота оказывает незначительное влияние на связывающую активность в отношении Helicobacter pylori по сравнению с соединением из шести сахаров, 9В. На основании сравнительного анализа первой проекции с фиг.9А и 9В можно предположить, что в структуре из семи сахаров также имеется тот же связывающий эпитоп.
Делинезация связывающего эпитопа неолактосоединений. На основании данных по относительной активности связывания структур, полученных в результате химического и ферментативного расщепления соединений из семи сахаров, выделенных из тимуса кроликов (номера 1, 5, 10 и 21, таблица 2), можно предположить, что последовательность из трех сахаров GlcNAcβ3Galβ4GlcNAcβ3 может быть минимальной связывающей последовательностью. Так, предполагается наличие ингибирующего действия концевого Galβ4 для соединения из шести сахаров, в то время, как отсутствие концевого GlcNAcβ3 в неолактотетраозилкерамиде снижает его связывающую активность, поскольку имеется только два из трех сахаров в эпитопе. Значение внутреннего GlcNAcβ3 четко показано при потере связывания бактерий у неолактотетраозилкерамида и В5 после превращения ацетамидной группы в амин в результате де-N-ацилирования (номера 6 и 14, таблица 2). Отсутствие связывания может иметь место в результате потери необходимого взаимодействия между адгезином и ацетамидной группой, и/или измененных конформационных предпочтений данных гликосфинголипидов. Однако трудно представить ситуацию, при которой измененная ориентация внутренней Galβ4 будет создавать стерическое затруднение для доступа к связывающему эпитопу. Таким образом, установив, что минимальная связывающая последовательность должна включать последовательность GlcNAcβ3Galβ4GlcNAcβ3, легко объяснить отсутствие связывания у структур Р1, Н5-2 и двух сиалилпараглобозидов (номера 15, 18-20, таблица 2), поскольку данные удлинения непосредственно мешают предполагаемому связывающему эпитопу. Также гликосфинголипид из пахты, полученной из коровьего молока (Teneberg et al., 1994), который имеет β6-связанную боковую цепь Galβ4GlcNAcβ, соединенную с внутренней Galβ4 неолактотетраозилкерамида (номер 26, таблица 2), не обладает связывающей активностью за счет блокирования доступа к связывающему эпитопу.
Показано, что удлинение различных связывающих последовательностей из пяти сахаридов, представленных в таблице 2, заключающееся только в добавлении Galβ4 к выделенной из тимуса структуре, не оказывает влияния на связывание, что согласуется с наблюдением о том, что положение 4-ОН может быть либо экваториальным, либо аксиальным, но с потерей аффинности связывания за счет стерического затруднения. Добавление NeuAcα3 к х2 или GalNAcβ3 к В5 приводит к полной потере связывания (номера 24 и 25, таблица 2). Кроме того, наблюдаемое отрицательное влияние единицы Fucα2, как это имеет место в Н5-2, подтверждается отсутствием связывания Helicobacter pylori с А6-2 и В6-2 (номера 22 и 23, таблица 2). В случае удлиненной структуры (номер 28, таблица 2), заканчивающейся тем же трисахаридом, что и в В5, то она должна быть, как в случае В5, концевым трисахаридом, ответственным за наблюдаемое связывание, хотя также имеется второй внутренний связывающий эпитоп. Однако связывание с внутренним эпитопом, вероятно, можно исключить, поскольку предполагается получить предпоследнюю Galβ4, или оно не зависит от типа штамма и условий культивирования (Miller-Podraza et al., 1996, 1997 a,b).
Суммируя вышеизложенное, можно заключить, что связывающий эпитоп неолактопроизводных гликосфинголипидов должен включать последовательность из трех сахаров GlcNAcβ3Galβ4GlcNAcβ3 для того, чтобы получить максимальную активность. На основании сравнительных данных по характеру связывания потенциальных изорецепторов, использованных в данном исследовании, можно придти к выводу из структур, представленных на фиг.8А-D и 9А-D, что почти все в данном трисахариде важно, чтобы имело место связывание, за исключением ацетамидной группы конечного GlcNAcβ3 и 4-ОН в том же остатке, которые не являются решающими.
Биологическое наличие рецепторов. Из четырех гликосфинголипидов, в состав которых входят пять сахаров, которые в условиях in vitro могут функционировать взаимозаменяемо в качестве рецепторов для Helicobacter pylori, в естественных условиях только х2 находится в тканях человека, но еще не было установлено его наличие в слизистой желудка, за исключением опухолей желудка, где он был обнаружен в опухолевой ткани (Kannagi et al., 1982b). В исследовании Thorn et al., 1992, однако, было показано, что х2-гликосфинголипиды и удлиненные структуры, имеющие концевую последовательность GlcNAcβ3Galβ4GlcNAcβ, присутствуют в нескольких тканях человека, однако, к сожалению, эпителиальной ткани желудка среди тестированных не было. Следовательно, провели тонкослойную хроматографию с нанесением специфических моноклональных антител ТН2 к GalNAcβ3Galβ4GlcNAcβ цельной некислотной фракции гликосфинголипидов из эпителиальных клеток слизистой желудка человека от индивидуумов с несколькими группами крови А (полосы 1-6) (фиг.11В). Однако не наблюдали детектируемого связывания с гликосфинголипидами, полученными из эпителия желудка, с использованием данного теста. Результаты соответствующего нанесения Galβ4GlcNAcβ-связывающего лектина из E.cristagalli представлены на фиг.11А, 11В и 11С. Из препаратов различных гликосфинголипидов эпителия желудка на первых трех полосах видно слабое связывание с полосами в области фракции из четырех сахаров, которое, вероятно, относится к неолактотетраозилкерамиду, но не было установлено детектируемого связывания Helicobacter pylori с этими полосами за счет низких количеств данного гликосфинголипида (Teneberg et al., 2001).
Кроме того, последовательность Galα3Galβ4GlcNAcβ, если имеется в гликосфинголипиде В5 или в удлиненной структуре, описанной выше (номер 28, таблица 2), возможно, не обнаруживается в нормальных тканях человека за счет отсутствия экспрессии трансферазы, ответственной за добавление Galα3 (Larsen et al., 1990). Следовательно, остается сделать вывод, что, если рецептор(ы)-мишени(ей), имеющий связывающий эпитоп, определенный выше, находится на поверхности эпителиальных клеток желудка, то он может быть основан на повторяющихся элементах N-ацетиллактозамина в гликопротеинах, но не на липидных структурах.
Однако известно, что штаммы Helicobacter pylori, вызывающие пептическую язву, как основной используемый здесь штамм, вызывают ответную воспалительную реакцию гранулоцитов, даже, когда бактерии не опзонизированы (Rautelin et al., 1994a, b). Наиболее вероятно, что первоначальное событие при фагоцитозе бактерий заключается в специфических лектиноподобных взаимодействиях, приводящих к агглютинации гранулоцитов (Ofek и Sharon, 1988). После фагоцитоза имеют место реакции окислительного «взрыва», которые могут иметь значение для патогенеза вызываемых Helicobacter pylori заболеваний (Babior, 1978). Было выделено из гранулоцитов и охарактеризовано несколько кислотных и некислых гликосфинголипидов, имеющих неолактоядро и повторяющиеся единицы лактозамина, включая номер 21 в таблице 2, и сиалилированное соединение из семи сахаров (номер 27, таблица 2), в которых ацетамидная группа сиаловой кислоты находится в ацетилированной форме (Fukuda et al., 1985; Stroud et al., 1996), и они могут функционировать в качестве потенциальных рецепторов Helicobacter pylori на поверхности лейкоцитов. Кроме того, из того же источника был выделен х2-гликосфинголипид (Teneberg S., неопубликованные данные).
Возвращаясь к фиг.11В, видно, что моноклональные антитела ТН2 действительно связываются с полосами в области фракций из пяти сахаров, полученных как из гранулоцитов, так и из эритроцитов (соответственно полосы 7 и 8), которые могут относиться к х2-гликосфинголипидам (Teneberg S., неопубликованные данные; Thorn et al., 1992; Teneberg et al., 1996). Аналогично было установлено, что неолактотетраозилкерамид находится в гранулоцитах и эритроцитах, когда использовали лектин E.cristagalli в тестах с нанесением (фиг.11С, полосы 7 и 8). В двух этих случаях Helicobacter pylori связывались с неолактотетраозилкерамидом (Bergstöm J., неопубликованные данные). Для гранулоцитов также обнаруживается довольно слабая полоса в области фракций из шести сахаров, соответствующая, вероятно, неолактотетраозилкерамиду с наращением на одну единицу N-ацетиллактозамина (сравни номер 21, таблица 2), что согласуется с данными Fukuda et al., 1985. Однако остается выяснить, насколько эти гликосфинголипиды являются основными мишенями в процессе агглютинации.
Анализ неогликолипидов и новых гликолипидов
Олигосахариды GlcNAcβ3Galβ4GlcNAc, GlcNAcβ3Galβ4GlcNAcβ6GlcNAc, Galα3Galβ4GlcNAcβ3Galβ4Glc и GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc и мальтогептозу (Sigma, Saint Louis, США) подвергали восстановительному аминированию 4-гексадециланилином (сокращение HDA от Aldrich, Stockholm, Швеция) с использованием цианоборогидрида (Halina Miller-Podraza, данные будут опубликованы позднее). Продукты характеризовали масс-спектрометрией, и было подтверждено, что это GlcNAcβ3Galβ4GlcNAc(красный)-HDA,
GlcNAcβ3Galβ4GlcNAcβ6GlcNAc(красный)-HDA, Galα3Galβ4GlcNAcβ3Galβ4Glc(красный)-HDA, GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc(красный)-HDA и мальтогептаоза(красный)-HDA [где «(красный)» означает структуру с аминной связью, полученную в результате восстановительного аминирования глюкозы восстанавливающего конца сахаридов и аминогруппы гексадециланилина (HDA)]. Соединения Galα3Galβ4GlcNAcβ3Galβ4Glc(красный)-HDA и GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc(красный)-HDA обладают выраженной связывающей активностью и GlcNAcβ3Galβ4GlcNAcβ6GlcNAc(красный)-HAD обладает высокой связывающей активностью в отношении Helicobacter pylori в тесте ТСХ с нанесением бактерий, описанном выше, в то время, как для GlcNAcβ3Galβ4GlcNAc(красный)-HDA и мальтогептаозы(красный)-HDA была характерна слабая активность, или они были неактивны. В примере показано, что тетрасахарид GlcNAcβ3Galβ4GlcNAcβ3Gal представляет структуру, связывающуюся с Helicobacter pylori. Glc-остаток восстанавливающего конца, возможно, не требуется для связывания, поскольку в результате восстановления разрушается пиранозная кольцевая структура Glc-остатка. В противоположность интактная кольцевая структура восстанавливающего конца GlcNAc необходима для эффективного связывания трисахарида GlcNAcβ3Galβ4GlcNAc.
Биосинтетический предшественник-аналог гликолипида NHK-1
GlcAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer и новые гликолипиды Glcβ3Galβ4GlcNAcβ3Galβ4GlcβCer и
Glc(А-метиламид)β3Galβ4GlcNAcβ3Galβ4GlcβCer тестировали ТСХ с нанесением бактерий и наблюдали, что они являются активными в отношении связывания Helicobacter pylori. Glc(А-метиламид) означает производное глюкуроновой кислоты, в котором карбоксильная группа амидирована метиламином. Структура Glcβ3Galβ4GlcNAcβ3Galβ4GlcβCer обладала высоким связыванием с Helicobacter pylori и Glc(А-метиламид)β3Galβ4GlcNAcβ3Galβ4GlcβCer обладал очень высоким связыванием с Helicobacter pylori.
Получение GlcAcβ3Galβ4Glc(NAc) трансгликозилированием
Акцепторный сахарид Galβ4Glc или Galβ4GlcNAc (примерно 10-20 мМ) инкубировали с 10-кратным молярным избытком паранитрофенил-бета-глюкуроновой кислоты и β-глюкуронидазы из печени крупного рогатого скота (20000Е, Sigma) в буфере с рН примерно 5 в течение 2 суток при 37°С при перемешивании. Продукт очищали ВЭЖХ.

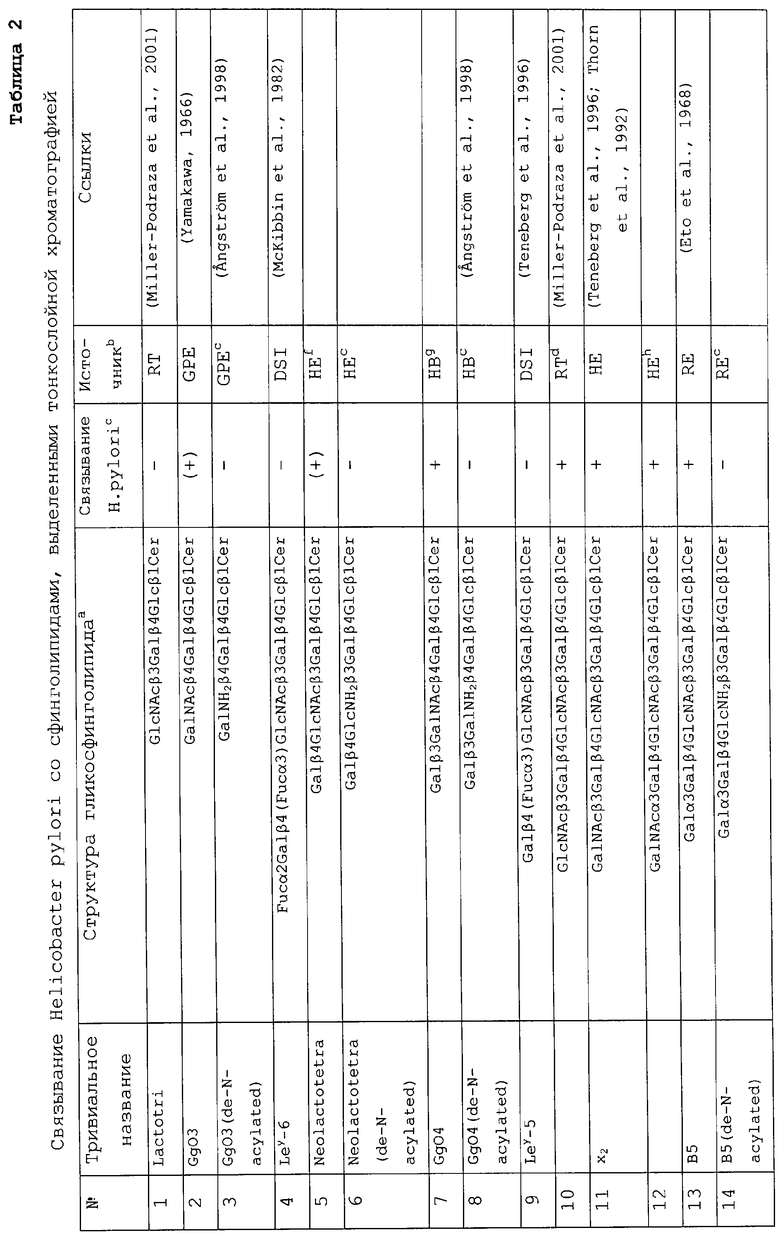
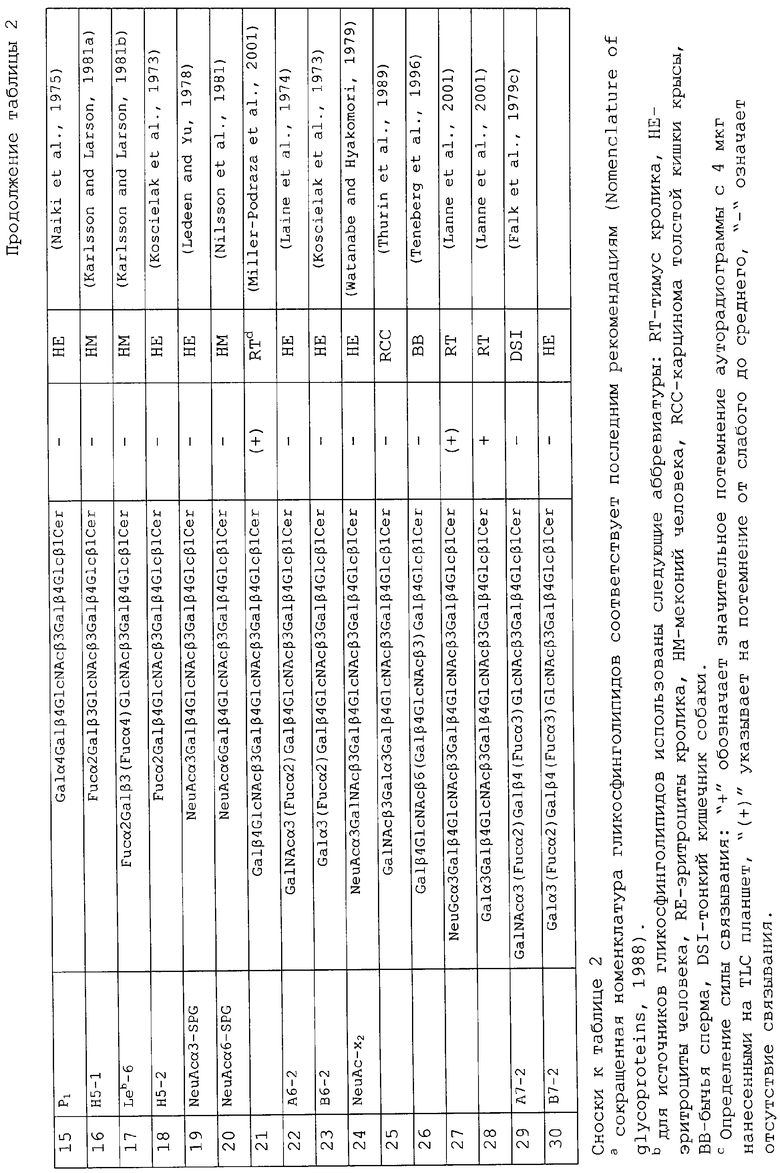
Литература





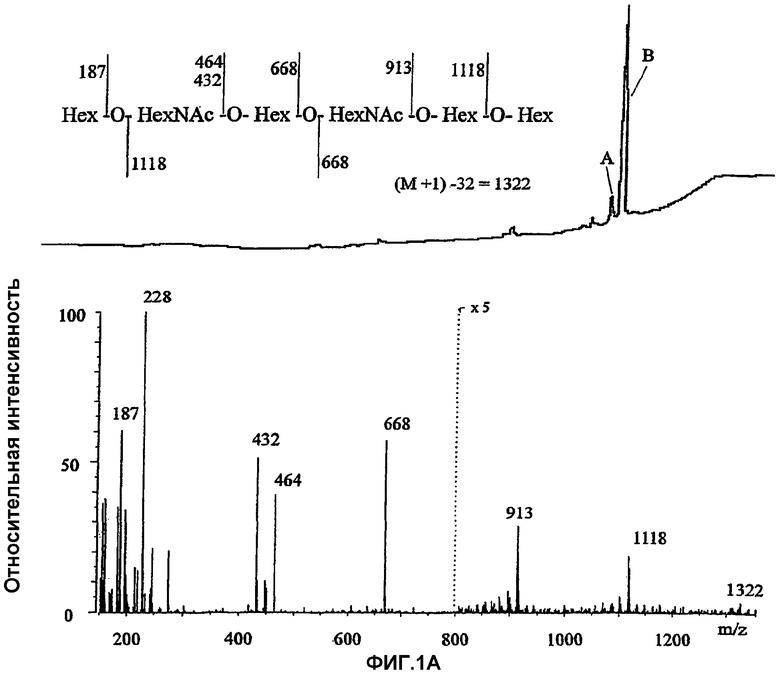
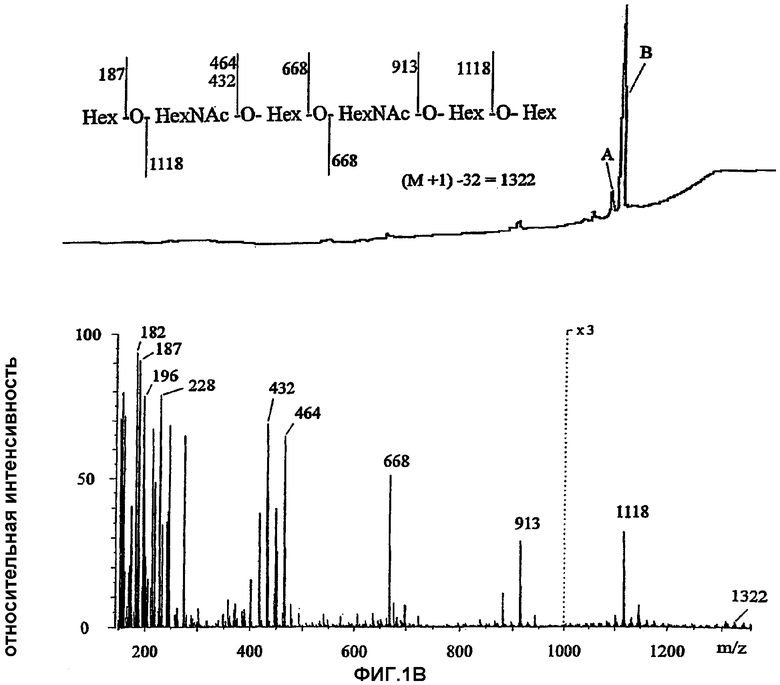
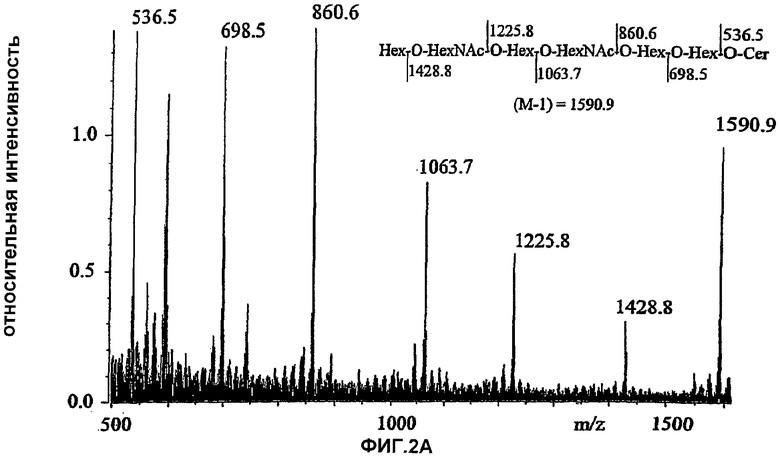
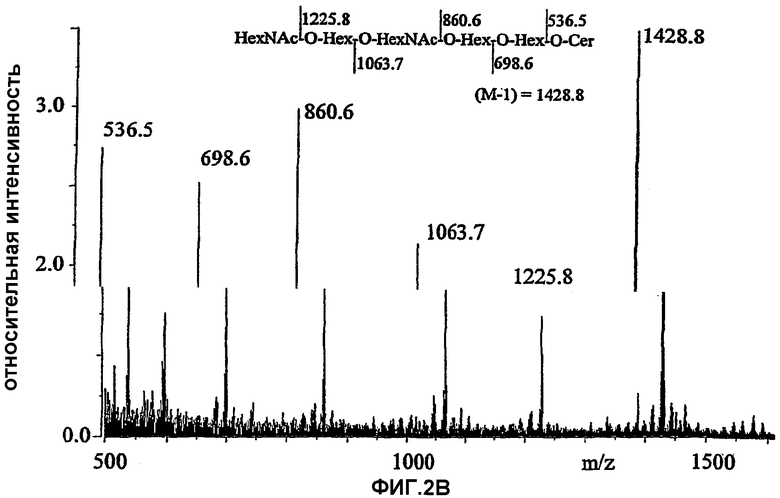
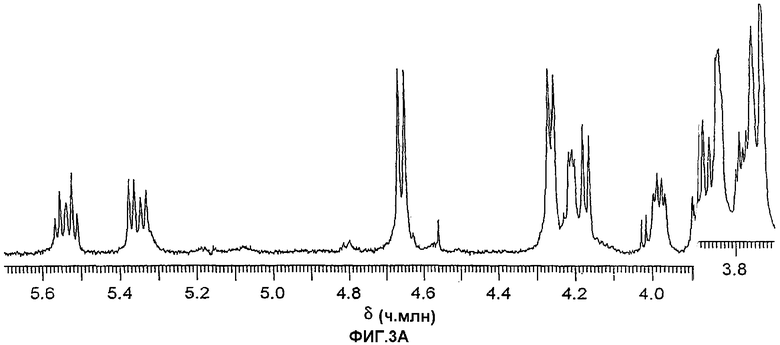
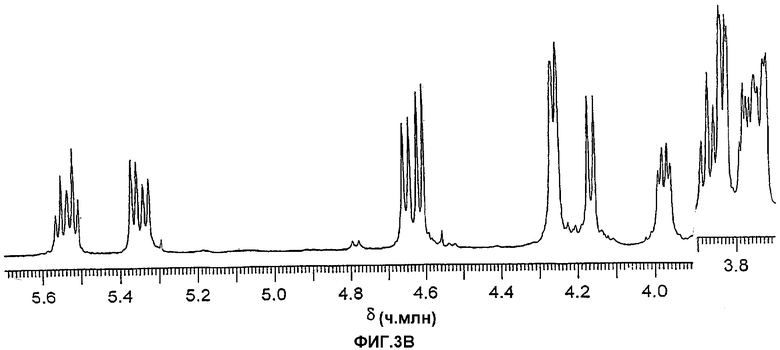


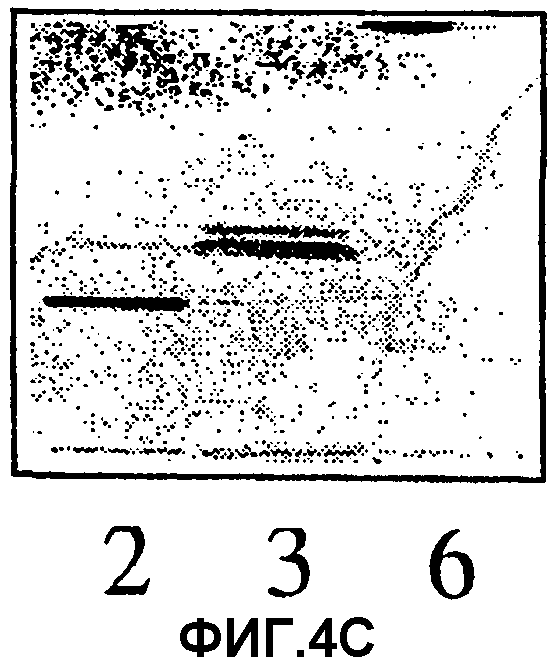
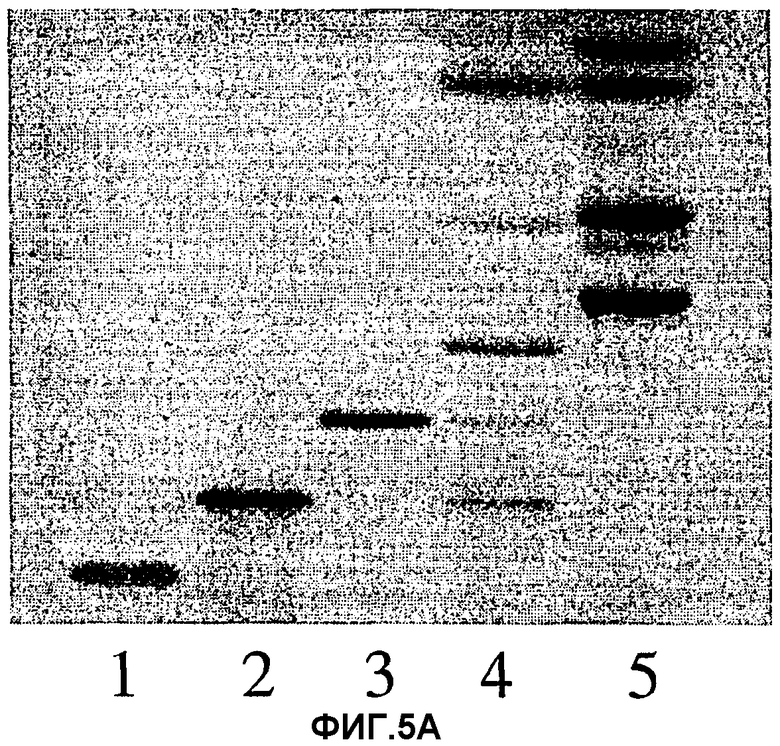

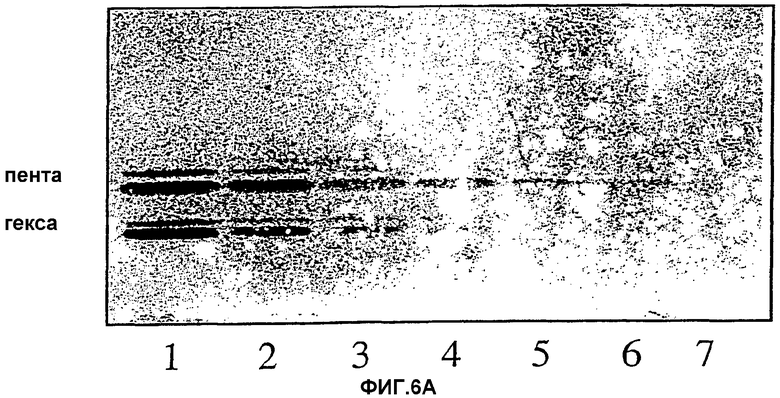
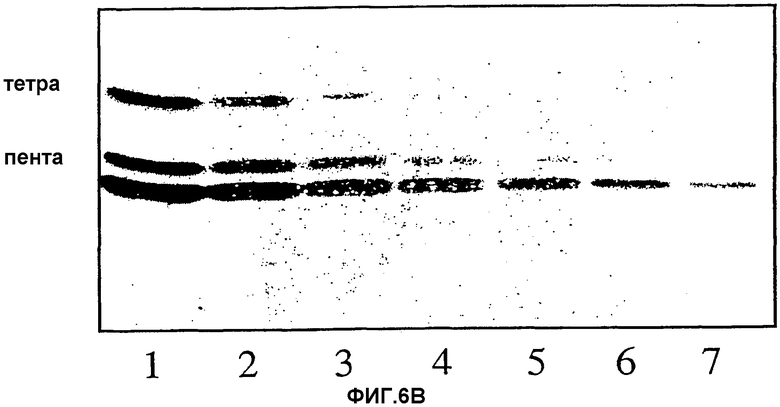
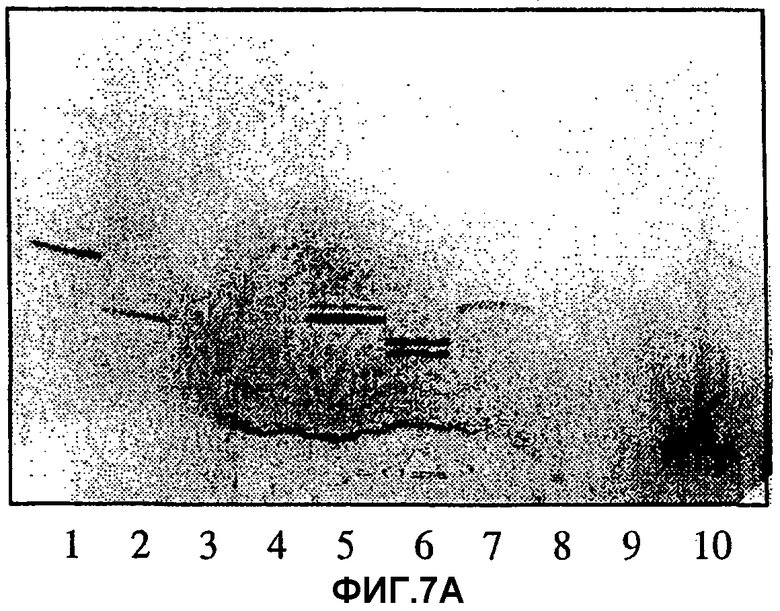
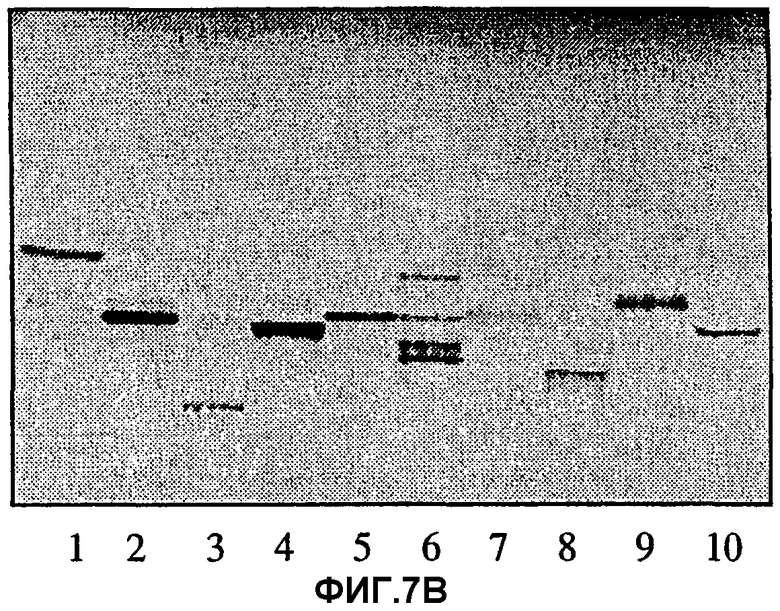
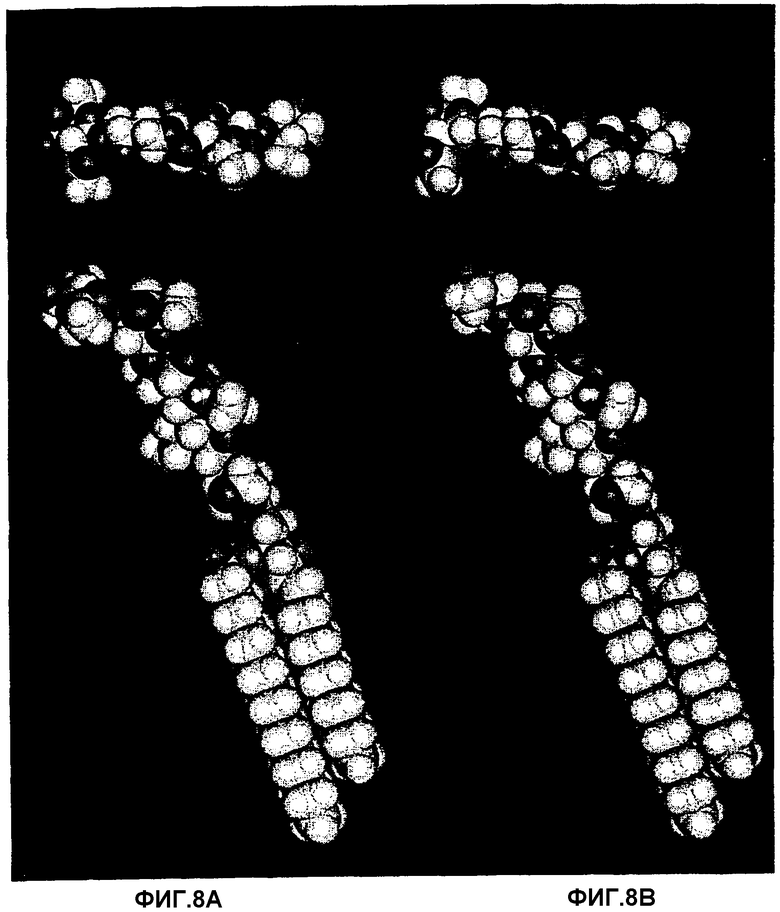
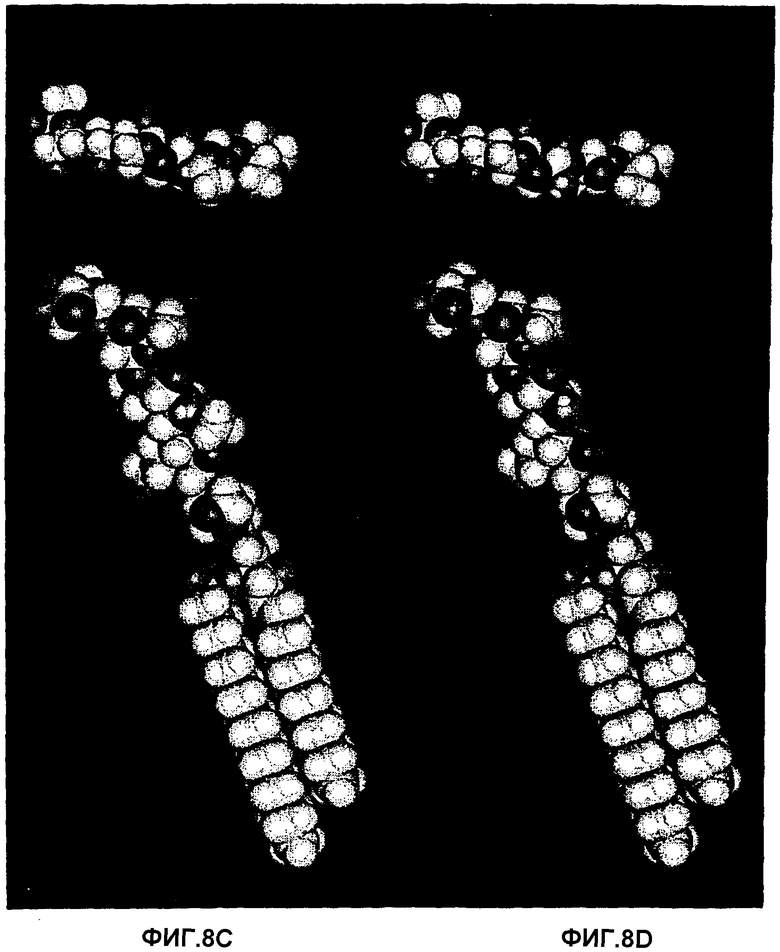
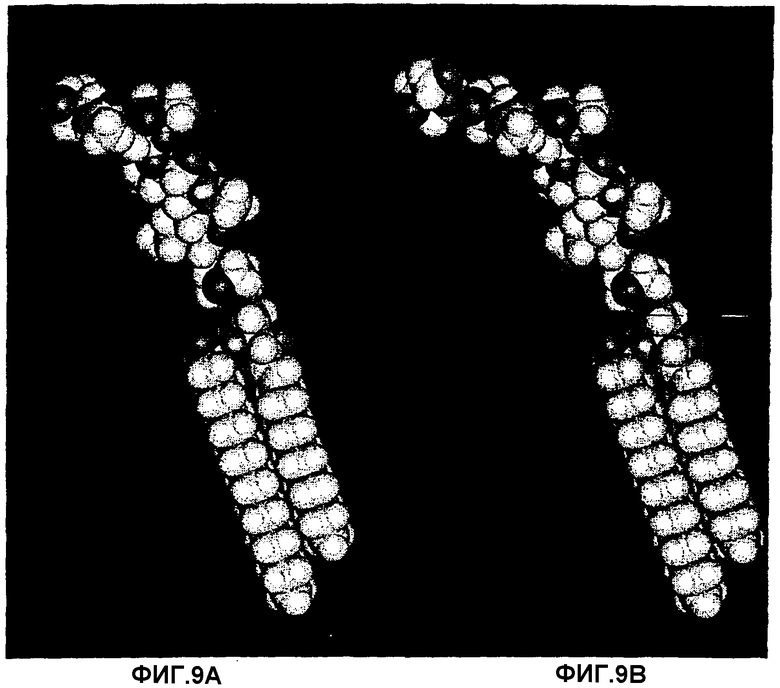
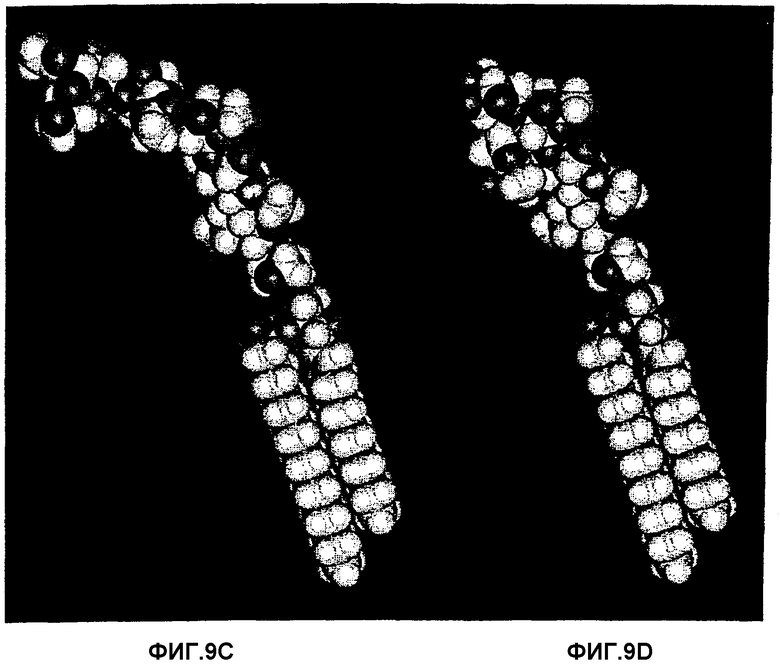
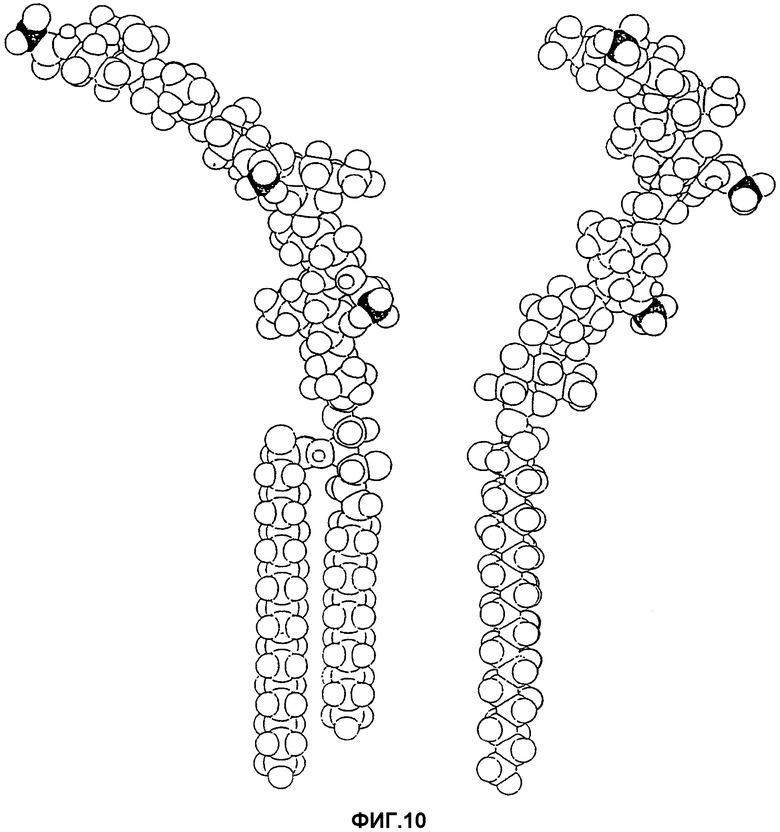
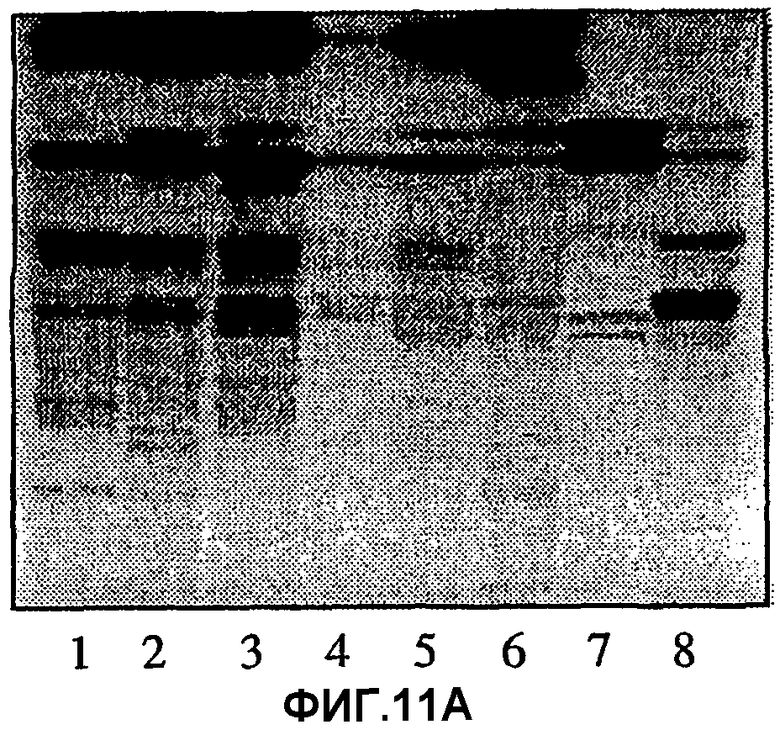
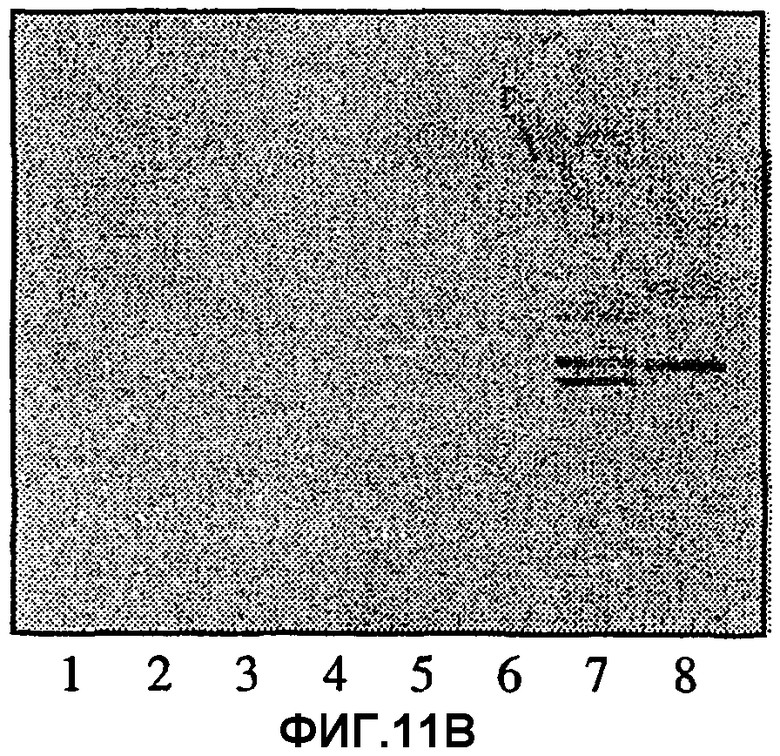
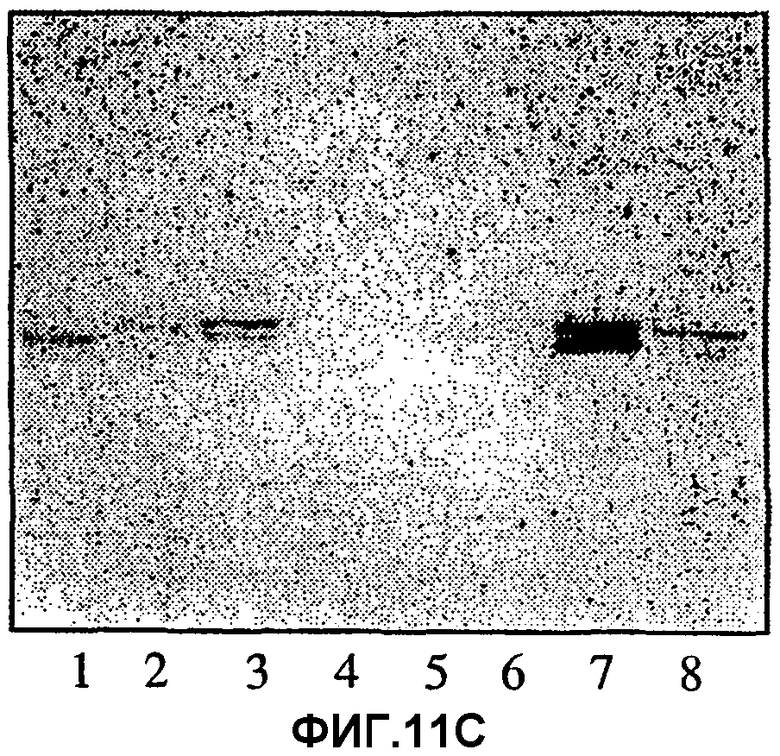
Настоящее изобретение относится к медицине и фармации и описывает соединение или рецептор, включающий связывающую Helicobacter pylori олигосахаридную последовательность [Gal(A)q(Nac)r/Gal(A)q(NAc)rα3/β3]s[Galβ4GlcNAcβ3]tGalβ4Glc(NAc)u, в которой q, r, s, t и u каждый независимо равен 0 или 1, и аналогов или производных указанной олигосахаридной последовательности в качестве эпитопа, связывающегося с Helicobacter pylori, их применение, например, в фармацевтической и пищевой композициях для лечения состояний, вызванных присутствием Helicobacter pylori. Изобретение обеспечивает повышение эффективности лечения. 4 н. и 14 з.п. ф-лы, 11 ил., 2 табл.
[Gal(A)q(NAc)r/Glc(A)q(NAc)rα3/β3]s[Galβ4GlcNAcβ3]tGalβ4Glc(NAc)u,
в которой каждый из q, r, s, t и u независимо равен 0 или 1,
необязательно β1-6-связанной от восстанавливающего конца с GalNAc, GlcNAc, Gal или Glc, и аналогов или производных указанной олигосахаридной последовательности в качестве эпитопа, связывающегося с Helicobacter pylori, для производства композиции, обладающей активностью связывать или ингибировать Helicobacter pylori, так, что когда t=0 и u=0, олигосахаридная последовательность связана с поливалентным носителем или находится в виде свободного олигосахарида в высокой концентрации в указанной композиции.
GlcNAcβ3Galβ4GlcNAc или GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc,
в которой положение С4 концевого GlcNAcβ3 необязательно
связано с Galβ1- или олигосахаридной цепью гликозидной связью.
Galβ4GlcNAc,
GalNAcα3Galβ4GlcNAc, GalNAcβ3Galβ4GlcNAc, GlcNAcα3Galβ4GlcNAc,
GlcNAcβ3Galβ4GlcNAc, Galβ3Galβ4GlcNAc, Glcα3Galβ4GlcNAc, Glcβ3Galβ4GlcNAc,
Galβ4GlcNAcβ3Galβ4GlcNAc, Galβ4GlcNAcβ3Galβ4Glc,
GalNAcα3Galβ4GlcNAcβ3Galβ4Glc,GalNAcβ3Galβ4GlcNAcβ3Galβ4Glc,
GlcNAcα3Galβ4GlcNAcβ3Galβ4Glc, GlcNAcβ3Galβ4GlcNAβp3Galβ4Glc,
Galβ3Galβ4GlcNAcβ3Galβ4Glc, Glcα3Galβ4GlcNAcβ3Galβ4Glc,
Glcβ3Galβ4GlcNAcβ3Galβ4Glc,
GalANAcβ3Galβ4GlcNAc, GalANAcα3Galβ4GlcNAc, GalAβ3Galβ4GlcNAc,
GalAα3Galβ4GlcNAc, GalANAcβ3Galβ4Glc, GalANAcα3Galβ4Glc, GalAβ3Galβ4Glc,
GalAα3Galβ4Glc,
GlcANAcβ3Galβ4GlcNAc, GlcANAcα3Galβ4GlcNAc,GlcAβ3Galβ4GlcNAc,
GlcAα3Galβ4GlcNAc, GlcANAcβ3Galβ4Glc, GlcANAcα3Galβ4Glc, GlcAβ3Galβ4Glc,
GlcAα3Galβ4Glc,
Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc,
GalNAcα3Galβ4Glc, GalNAcβ3Galβ4Glc, GlcNAcα3Galβ4Glc,
GlcNAcβ3Galβ4Glc, Galβ3Galβ4Glc, Glcα3Galβ4Glc, Glcβ3Galβ4Glc
и их поливалентные конъюгаты по восстанавливающему концу.
[OS-O-(X)n-Y]m-Z,
где целые числа n и m имеют значения m≥1, a n независимо равно 0 или 1; Х предпочтительно представляет собой лактозил-, галактозил-, поли-N-ацетиллактозаминил или часть O-гликановой или N-гликановой олигосахаридной последовательности, Y представляет собой спейсерную группу или концевой конъюгат, такой, как липидная группа керамида или связь с Z или гидрофильный спейсер, более предпочтительно гибкий гидрофильный спейсер;
когда m>1, Y необязательно может иметь структуру
L1-CH(Н/{CH1-2OH}p1)-{CH1OH}р2-{СН(NH-R)}р3-{CH1OH }p4-L2,
в которой L1 и L2 представляют собой связывающие группы, независимо содержащие связывающие атомы кислорода, азота, серы или углерода, или два связывающих атома группы, образующей связи, такие, как -О-, -S-, -CH2-, -N, -N(СОСН3)-, амидные группы -CO-NH- или -NH-CO-, или -N-N- (производное гидразина), или аминооксисвязи -O-N- и -N-O-; L1 представляет связь из 1 углерода моносахарида восстанавливающего конца Х или, когда n=0, L1 замещает -О- и связи непосредственно от восстанавливающего конца C1 в OS; p1, р2, р3 и р4 независимо представляют собой целые числа 0-7 при условии, что по меньшей мере один из р1, р2, р3 и р4 равен по меньшей мере 1; CH1-2OH в ответвлении {CH1-2OH}p1 означает, что концевая группа цепи представляет СН2ОН, и когда р1 больше 1, то имеются вторичные спиртовые группы -СНОН-, связывающие концевую группу с остальной частью спейсера; R предпочтительно представляет собой ацетильную группу (-СОСН3) или R представляет собой альтернативную связь с Z и тогда L2 представляет собой концевую группу цепи из одного или двух атомов, в другом воплощении R является образующей аналог группой, содержащей C1-4-ацильную группу, содержащую амидоструктуру, или Н, или C1-4-алкил, образующий амин.
[Gal(A)q(NAc)r/Glc(A)q(NAc)rα3/β3]s[Galβ4GlcNAcβ3]tGalβ4Glc(NAc)u,
в которой каждый из q, r, s, t и u независимо равен 0 или 1,
необязательно β1-6-связанной от восстанавливающего конца с GalNAc, GlcNAc, Gal или Glc, в качестве эпитопа, связывающегося с Helicobacter pylori, для производства композиции, обладающей активностью связывать или ингибировать Helicobacter pylori, где указанная олигосахаридная последовательность конъюгирована с полисахаридом, предпочтительно полилактозаминовой цепью или ее конъюгатом, или указанный конъюгат олигосахаридной последовательности представляет собой гликолипид, олигомерную молекулу, содержащую по меньшей мере две или три олигосахаридных цепи, мицеллу, или указанная олигосахаридная последовательность ковалентно конъюгирована с антибиотиком, эффективным против Helicobacter pylori.
Gal(A)q(NAc)r/Glc(A)q(NAc)rα3/β3Galβ4Glc(NAc)u,
в которой q, r и u независимо равны 0 или 1, при условии, что указанная олигосахаридная последовательность не является Galα3Galβ4Glc/GlcNAc, Galβ3Galβ4Glc или GlcNAcβ3Galβ4GlcNac,
в качестве невосстанавливающей последовательности концевой области, и его связывающие Helicobacter pylori аналоги и производные.
Glc(A)q(NAc)rα3/β3Galβ4Glc(NAc)u,
в которой q, r и u независимо равны 0 или 1,
при условии, что когда указанная олигосахаридная последовательность содержит β3-связь, то оба q и r равны 0 или 1; или
GalA(NAc)rα3/β3Galβ4Glc(NAc)u,
в которой r и u независимо равны 0 или 1,
и его связывающие Helicobacter pylori аналоги и производные.
[Gal(A)q(NAc)r/Glc(A)q(NAc)rα3/β3]s[Galβ4GlcNAcβ3]tGalβ4Glc(NAc)u,
в которой каждый из q, r, s, t и u независимо равен 0 или 1,
необязательно β1-6-связанную от восстанавливающего конца с GalNAc, GlcNAc, Gal или Glc, или ее аналоги или производные, так, что когда t=0 и u=0, олигосахаридная последовательность связана с поливалентным носителем или находится в виде свободного олигосахарида в высокой концентрации в указанной композиции.
| WO 8604065 А1, 17.07.1986 | |||
| АМИНООЛИГОСАХАРИД СК-4416, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ИНГИБИТОР САХАРИДГИДРОЛАЗЫ И АНТИБАКТЕРИАЛЬНЫЙ АГЕНТ | 1995 |
|
RU2134694C1 |
| HU Y | |||
| et al | |||
| Glycobiology, 1994, Jun 4 (3): 251-7. | |||

