Настоящее изобретение относится к композициям и способам для уменьшения или подавления воспаления, и для лечения воспалительных процессов и других патологических нарушений, в которых участвует межклеточная адгезия.
Эндотелиальные клетки сосудов и кровяные пластинки (тромбоциты) играют ключевую роль в нескольких биологических ответах при помощи селективного связывания некоторых клеток, например, фагоцитных лейкоцитов, в кровяном потоке. Например, эндотелиальные клетки в предпочтительном варианте связывают моноциты и гранулоциты перед их миграцией через стенку кровеносного сосуда и в окружающую ткань в воспалительной реакции. Известно, что некоторые, подавляющие воспаление, соединения действуют непосредственно на эндотелий сосудов, чтобы промотировать адгезию лейкоцитов к стенкам сосудов. Далее клетки проходят через стенки в область поражения или инфекции. Предполагается, что клеточная адгезия к эндотелию сосудов существует также в метастазах опухолей. Циркулирующие раковые клетки очевидно используют преимущество нормальных воспалительных механизмов организма и связываются с областями стенок кровеносных сосудов, где эндотелий активирован.
Кровяные пластинки также принимают участие в тех же реакциях. Известно, что кровяные пластинки становятся активированным и во время инициирования гемостаза и подвергаются основным морфологическим, биохимическим и функциональным изменениям (например, быстрый экзоцитоз гранул или дегрануляция), в которых мембрана альфа-гранулы пластинки сливается с внешней мембраной плазмы. В результате начинают экспрессироваться новые протеины клеточной поверхности, которые наделяют активированные пластинки новыми функциями, такими как способность связываться как с другими активированными пластинками, так и другими клетками. Активированные пластинки (тромбоциты) включаются в растущие тромбы или быстро освобождают циркулирующую кровь. Активированные тромбоциты, как известно, связываются с фагоцитными лейкоцитами, включая моноциты и нейтрофилы. Примеры патологических и других биологических процессов, в которых видимо участвует в качестве промежуточного последний процесс, включают атеросклероз, образование сгустков крови и воспаление.
В результате недавних работ было обнаружено, что специлизированные рецепторы клеточной поверхности на эндотелиальных клетках и тромбоцитах, именуемые как эндотелиальная молекула-1 адгезии лейкоцитов (ELAM-1) и протеин-140 мембраны гранул (GMP-140) соответственно, включаются в узнавание различных циркулирующих клеток эндотелием и тромбоцитами. Эти рецепторы являются поверхностными гликопротеинами с областью, напоминающей лектин, зоной с гомологией относительно эпидермального фактора роста и зоной с гомологией относительно дополнительных регуляторных протеинов /см. Bevilacgua и др., Science т. 243, стр. 1160 /1989/, которая включена здесь в качестве ссылки/. Например, было показано, что ELAM-1 участвует в эндотелиальной адгезии лейкоцитов, что является первой стадией во многих воспалительных реакциях. В частности, ELAM-1 связывается с человеческими нейтрофилами, моноцитами, эозинофилами, некоторыми Т-лимфоцитами /N. Graber и др., J. Jmmunol, т. 145, стр. 819 /1990//, NK-клетками и линией промиэлоцитных клеток HL-60.
Термин "селектин" был предложен для общего класса рецепторов, который включает ELAM-1 и GMP-140, ввиду их области, напоминающей лектин, и селективной природы их адгезионных функций. Эти рецепторы клеточной поверхности экспрессируются на самых разнообразных клетках. GMP-140 (известный также под обозначением PADGEM) содержится на поверхности тромбоцитов и эндотелиальных клеток, где он участвует во взаимодействиях тромбоцит-лейкоцит и эндотелий-лейкоцит. Еще одним членом класса селектина является антиген MEL-14 и его человеческий аналог LAM-1, которые являются клеточными поверхностными рецепторами лимфоцитов и действуют как "домашние" рецепторы лимфатических узлов. Точная природа лиганда, узнаваемого рецепторами селектина, остается неизвестной.
Ранее были разработаны различные другие приемы блокировки действия селектинов и, таким образом, ингибирования клеточной адгезии. Например, использование моноклональных антител, направленных на ELAM-1, было предложено в качестве процедуры ингибирования адгезии эндотелий-лейкоцит с целью лечения патологических реакций, таких как воспаление. Эндотелиальный интерлеукин-8, как было установлено, также является ингибитором взаимодействий лейкоцит-эндотелий.
После того, как будет выяснено взаимодействие лиганд-рецептор, станет возможным разрабатывать высокоспецифические, эффективные ингибиторы клеточной адгезии с участием селектина, которые были бы весьма полезны при терапевтическом лечении. Лиганд (лиганды) также можно использовать для синтеза других фармацевтических соединений, таких как противовоспалительные агенты или антиоксиданты, для применения к местам поражений. До настоящего времени недостаточное понимание взаимодействия лигандов и молекул рецепторов на соответствующих клетках осложняло эти усилия. Настоящее изобретение удовлетворяет эти и некоторые другие нужды.
Новые композиции, которые селективно связываются с рецептором селектина и которые содержат по крайней мере одну связывающую селектин составляющую, являются предметом настоящего изобретения. Связующие селектин составляющие имеют общую формулу:
R1-Gal β 1,4(Fue α 1,3)GlcNAc - (R2)a,
в которой
R1 является олигосахаридом или R3-R4-C/CO2H/-, в которой R3 и R4, которые могут быть как одинаковыми, так и различными, являются -H, C1-C8 алкилом, -гидроксил C1-C8 алкилом, -арил C1-C8 алкилом или -алкокси C1-C8 алкилом;
в которой R2 является β 1, 3Gal, α 1,2 Man, или α 1,6 GalNAc и a равно 0 или 1.
В некоторых предпочтительных вариантах осуществления R1 является сиаловой кислотой, в общем случае, NeuAc или NeuGc.
Если R1 является олигосахаридом, им в предпочтительном варианте является Neu Ac α 2, 3 Gal β 1, 4GlcNAc β 1 или Neu Gc α 2, 3 Gal β 1, 4GlcNAc β 1,3. Селектинсвязывающими составляющими являются в общем случае олигосахариды, содержащие минимальный тетрасахарид (именуемый SLX), узнаваемый рецептором селектина.
Это соединение в общем случае получают из полисахарида, имеющего повторяющийся блок, содержащий структуру ядра нефукозилированного SLX. При фукозилировании получают поливалентный, несущий SLX, полисахарид. Предпочтительными полисахаридами для этой цели являются полисахариды типа 1a, типа II и типа III из стрептококка Группы B. Поливалентные, связывающие селектин, соединения получают также при помощи соединения составляющих, связывающих селектин, с различными составляющими линкера.
Предлагаемые композиции ингибируют межклеточную адгезию, в которой участвует клеточный поверхностный рецептор селектина, и при этом они способны, например, ингибировать воспаление и другие патологические реакции, связанные с клеточной адгезией. В соответствующих вариантах осуществления композицией, которая связывает селектин, может быть полисахарид, гликопротеин, гликолипид или олигосахарид.
В соответствии с настоящим изобретением, в частности, предлагаются вышеупомянутые соединения в фармацевтических композициях. Эти фармацевтические композиции могут содержать, например, липосомы, которые содержат составляющую, способную селективно связываться с рецептором селектина, и приемлемый с фармацевтической точки зрения носитель. Липосома, содержащая такую составляющую, может также служить в качестве "переносчика к цели" для известного хемотерапевтического агента, который заключен внутри липосомы, и доставляется к намеченным клеткам, которые экспрессируют рецептор селектина. В общем случае хемотерапевтическим агентом является противовоспалительный агент или антиоксидант. Использование составляющих, описанных здесь, для доставки к цели химических агентов, заключенных внутри липосом, является известным и эффективным приемом для снижения терапевтических концентраций медикамента и минимизации побочных эффектов.
Фармацевтические композиции, являющиеся предметом настоящего изобретения, могут также содержать иммуноглобулины, способные селективно связываться с лигандом олигосахарида, узнаваемым рецептором селектина. Соответствующими иммуноглобулинами для этой цели являются CSLEX-1, FH6, SNH3, SNH4 и УТМ-2.
В других аспектах, в соответствии с настоящим изобретением предлагаются способы ингибирования межклеточной адгезии у пациента с таким заболеванием, как воспаление, при помощи применения к пациенту терапевтически эффективной дозы соединения, содержащего составляющую, способную связываться с рецептором селектина. Рецептор селектина, такой как ELAM-1 или GMP-140, может экспрессироваться на клетках эндотелия сосудов или тромбоцитах. Воспалительным процессом может быть, например, септический шок, рана, связанная с сепсисом, острое респираторное заболевание.
Фиг. 1 иллюстрирует способность клеток, которые экспрессируют SLX (LEC 11), связываться с IL-1β -активированными эндотелиальными клетками в сравнении с теми клетками, которые экспрессируют несиалированный Lex (CHO-K1 и LEC 12).
Фиг. 2 иллюстрирует способность моноклональных антител, специфических для SLX, блокировать связывание в участием селектина клеток HL-60 при температуре 37oC (фиг. 2A) и 4oC (фиг. 2B) по сравнению с моноклональными антителами, которые не связывают SLX-детерминанты.
Фиг. 3 иллюстрирует эффекты инкубирования LEC 11- (фиг. 3A) и LEC 12- фиг. 3B) клеток с SLX и не-SLX специфическими моноклональными антителами на связывание с активированными эендотелиальными клетками.
Фиг. 4 иллюстрирует результаты, полученные при помощи обработки клеток HL-60, LEC11 и LEC12 сиалидазой перед связыванием с активированными эндотелиальными клетками.
Фиг. 5 сравнивает способность липосом, которые содержат гликолипиды с SLX, Lex или аналогичными углеводными структурами, ингибировать связывание клеток HL-60 с активированными эндотелиальными клетками.
Фиг. 6 сравнивает ингибирование адгезии тромбоцитов с участием GMP-140 при помощи моноклональных антител, специфических относительно SLX и Lex детерминантов.
Фиг. 7 сравнивает способность липосом, которые содержат гликолипиды с SLX, Lex или аналогичными углеводными структурами, ингибировать связывание клеток HL-60 с активированными тромбоцитами.
Фиг. 8 сравнивает способность липосом, которые содержат гликолипиды с SLX, Lex или аналогичными углеводными структурами, ингибировать связывание PMN с активированными тромбоцитами.
Фиг. 9 показывает ингибирование адгезии с участием GMP-140 при помощи гликолипидов с концевой сиаловой кислотой, либо NeuAc, либо NeuGc.
Фиг. 10 демонстрирует профилактически применимые моноклональные антитела против ELAM-1, чтобы предотвратить вызываемую липополисахаридами гибель крыс.
Фиг. 11 демонстрирует терапевтически примененные моноклональные антитела против ELAM-1, чтобы предотвратить вызванную липополисахаридами гибель крыс.
Предлагаются композиции и способы ингибирования воспалительных и других болезненных реакций, в которых участвует клеточная адгезия. В соответствии с настоящим изобретением предлагаются также соединения (например, гликоконъюгаты и моноклональные антитела), которые обладают способностью блокировать или ингибировать адгезию клеток с участием клеточных поверхностных рецепторов селектина. Предложены также способы получения и "просеивания" таких соединений. Предложено также диагностическое и терапевтическое использование таких соединений.
Основу настоящего изобретения составляет открытие углеводной составляющей, узнаваемой поверхностными клеточными рецепторами селектина. Как уже было указано выше, селектины, известные также как семейство "LEC-CAM" молекул в клеточной адгезии, являются уникальными гликопротеинами, экспрессируемыми на поверхности самых разнообразных клеток. Например, ELAM-1 индуцированно экспрессируется на эндотелиальных клетках сосудов /Bevilacgua и др., см. выше и Hession и др., Proc. Natl. Acad. Sci., т. 87, стр. 1673-1677 /1990/; обе эти работы включены здесь в качестве ссылок/. Как было установлено, этот рецептор индуцируется воспалительными цитокинами, такими как интерлеукин 1β\ (IL-1 β) и фактор α некроза опухоли (TNF α , а также бактериальным эндотоксином /липополисахаридом/ /см. Bevilacgua и др., Proc. Natl. Acad. Sci., т. 84, стр. 9238-9242 /1987/, которая включена здесь в качестве ссылки/. Эти соединения действуют непосредственно на эндотелиальные клетки in vitro, чтобы существенно увеличить адгезию полиморфоядерных лейкоцитов (нейтрофило) и моноцитов (Bevilacgua и др., Proc. Natl. Acad. Sci., см. выше).
Как уже было указано выше, GMP-140 является мембранным гликопротеином тромбоцитов и эндотелиальных секреторных гранул /Geng и др., Nature т. 343, стр. 757-760 /1990/, которая включена здесь в качестве ссылки/. Активированные тромбоциты, которые экспрессируют GMP-140 на своей поверхности, как известно, связываются с моноцитами и нейтрофилами /Jungi и др., Blood, т. 67, стр. 629-636 /1986//, а также с линиями клеток, похожими на моноциты, например HL60 и U937 /Jungi и др., см. выше; Silvestein и др., J. Clin Invest, т. 79, стр. 867-874 /1987//; все эти работы включены здесь в качестве ссылок. GMP-140 является мембранным протеином гранул альфа с молекулярной массой 140 000, который экспрессируется на поверхности активированных тромбоцитов при стимуляции тромбоцитов и секретировании гранул /HSU-Lin и др., J. Biol. Chem. т. 259, стр. 9121-9126 /1984/; Sternberg и др., J. Cell. Biol т. 101, стр. 880-886 /1985/; Berman и др., J. Clin. Invest т. 78, стр. 130-137 /1986//. Его также обнаружили в мегакариотах /Beckstead и др., Blood, т. 67, стр. 285-293 /1986// и в эндотелиальных клетках /Mc Ever и др., Blood т. 70, стр. 355а /1987// внутри тел Вейбела-Палада /Bonfanti и др., Blood, т. 73, стр. 1109-1112 /1989//. Фури и др. в Патенте США N 783330 описывают моноклональные антитела, активные относительно GPM-140. Все вышеупомянутые работы включены здесь в качестве ссылок.
Третьим рецептором селектина является "домашний" рецептор лимфоцитов, MEL-14-антиген или LAM-1 /Gallatin и др., Nature, т. 304, стр. 30-34 /1983/; Siegellman и др. , Science, т. 243, стр. 1165-1172 /1989/; Rosen Cell Biology, т. 1, стр. 913-919 /1989/; и Lasky и др., Cell, т. 56, стр. 1045-1055 /1989/; все эти работы включены здесь в качестве ссылок. В дополнение к лимфоцитам, "домашний" антиген MEL-14/LAM-1 видимо функционирует ранее при связывании нейтрофила с эндотелим.
Структура и функция рецепторов селектина была выяснена при помощи клонирования и экспрессии кДНК полной длины, кодирующей каждый из вышеупомянутых рецепторов /см. , например, Bevilacgua и др., Science, см. выше, (ELAM-1), Geng и др., см. выше (GMP-140), и Lasky и др., см. выше /MEL-14 антиген//. Внеклеточная порция селектинов может быть разделена на три сегмента на основе гомолоний с ранее описанными протеинами. N-концевая зона (примерно 120 аминокислот) связана с семейством протеинов лектина млекопитающего типа C, как это описано в Drickamer. J. Biol. Chem. т. 263, стр. 9557-9560, /1988/ /которая включена здесь в качестве ссылки/, которое включает рецептор IgE слабого сродства CD23. Далее следует полипептидный сегмент, который содержит последовательность, которая связана с протеинами, содержащими мотив фактора эпидермального роста (EGF). Наконец, после области EGF имеются один или несколько повторяющихся тандемом мотивов в примерно 60 аминокислот каждый, связанные с теми, что содержатся в семействе дополнительных регулирующих протеинов.
Так как рецепторы селектина содержат область, напоминающую лектин, специфичность этих молекул аналогичным образом основана на взаимодействиях протеина-углевода. Факты, приводимые здесь, указывают на то, что сиалилированная, фукозилированная N-ацетиллактозаминовая составляющая антигена Льюиса X, обозначенная здесь как SLX, является составляющей, узнаваемой зоной лектина рецептора селектина. В частности, анализ показывает узнавание этой составляющей как ELAM-1, так и GMP-140. Соединения настоящего изобретения содержат этот фукозилированный, сиалилированный N-ацетиллактозаминовый блок в различных конфигурациях.
Термин "селективное связывание", как он здесь используется, относится к специфическому узнаванию одной молекулой (в общем случае, именуемой рецептором) другой молекулы (именуемой в общем случае, лигандом) при помощи пространственной или полярной организации детерминантного сайта на второй молекуле. Селективное связывание между этими двумя молекулами имеет место там, где сродство является достаточно сильным. Связывающее сродство в общем случае представляется константой сродства (Kа) для равновесных концентраций ассоциированных и диссоциированных конфигураций, а именно, Kа = [R-L]/[R][L], где [R], [L] и [R-L] являются концентрациями в равновесном состоянии рецептора [R], лиганда [L] и комплекса рецептора-лиганда [R-L] соответственно.
Взаимодействия специфического связывания рецептора и молекул лиганда в общем случае включают обратимые нековалентные ассоциации, такие как электростатическое притяжение, силы Ван дер Ваальса и водородные связи. Смотри в общем случае, Stryer, Biochemistry /W.H. Freeman and Company, N.Y. 3rd. Ed. 1988/, которая включена здесь в качестве ссылки. Примеры взаимодействий селективного связывания включают узнавание антитело-антиген, узнавание фермент-субстрат и т.п.
Номенклатура, используемая для описания олигосахаридных составляющих в соответствии с настоящим изобретением, следует стандартной номенклатуре. Используются стандартные сокращения для отдельных моносахаридов. Например, 2-N-ацетилглюкозамин обозначается через GlcNAc, фукоза - через Fuc, галактоза - через Gal и глюкоза - через Glc. Двумя сиаловыми кислотами, которые могут содержаться на олигосахаридах, являющихся предметом настоящего изобретения, являются 5-N-ацетилнейраминовая кислота (NeuAc) и 5-N-гликолилнейраминовая кислота (NeuGc). Если не указано противное, все сахара, за исключением фукозы (L-изомера) являются D-изомерами в циклической конфигурации (например, пираноза или фураноза). Два аномера циклических форм представляются α и β.
Моносахариды в общем случае соединены гликозидными связями с образованием олиго- и полисахаридов. Ориентация связи относительно пласкости колец указывается при помощи α и β. Отмечаются также определенные атомы углерода, которые образуют связь между этими двумя моносахаридами. Так β -гликозидная связь между C-1 галактозы и C-4 глюкозы представляется при помощи Gal β 1,4 Glc. Для D-сахаров (например, D-GlcNAc, D-Gal и D-NeuAc) обозначение α обозначает гидроксил, присоединенный к C-1 (C-2 в NeuAc) ниже пласкости кольца, а β -выше кольца. В случае L-фукозы обозначение α означает гидроксил, расположенный выше кольца, а β означает, что он ниже.
После того, как SLX идентифицирован как углеводный лиганд, который является "посредником" в адгезии лейкоцит-эндотелий и лейкоцит-клетка тромбоцита, соединения, содержащие SLX и близкие структуры, могут быть очищены или синтезированы de novo. Как подробно описано ниже, в соответствии с настоящим изобретением предлагается семейство соединений, содержащих связывающие селектин составляющие, являющиеся предметом настоящего изобретения. Например, биомолекулы можно использовать в качестве несущего эту составляющую соединения. Биомолекулы, как этот термин здесь используется, включают, но ими не исчерпывается полный список биологически значимых молекул, такие как аминокислоты (и их заменители), олигопептиды, протеины (например, гликопротеины и протеиновые гормоны, жирные кислоты), липиды (например, гликолипиды, фосфолипиды, сфинголипиды и ганглиозиды), стероидные гормоны, олигосахариды, полисахариды и нуклеиновые кислоты (например, деоксирибонуклеиновые кислоты и рибонуклеиновые кислоты). Эти соединения могут быть очищены и/или синтезированы в соответствии со стандартными приемами, известными каждому специалисту в этой области техники. Кроме того, самые разнообразные соединения, несущие эту составляющую, можно синтезировать de novo, как это описано ниже.
После того, как получены, такие соединения можно использовать для различных целей, включая, например, конкурирующее ингибирование связывания SLX-несущих клеток с клетками, которые экспрессируют рецепторы селектина. При помощи связывания соединений, являющихся предметом настоящего изобретения, с клеточным поверхностным селективном будет предотвращаться взаимодействие селектина с нативным SLX-лигандом на мигрирующих клетках, которое препятствует нормальному и патологическому связыванию лейкоцитов и других клеток с эндотелием или тромбоцитами. Таким образом, соединения, которые содержат одну или несколько связывающих селектин составляющих, могут служить в качестве эффективных ингибиторов, например, воспаления, атеросклероза, образования сгустков и других патологий с участием эндотелия или тромбоцитов.
Остаток сиаловой кислоты в SLX может находится в различных формах, лишь бы связывание селектина существенно не нарушалось. В общем случае, сиаловая кислота является 5-N-ацетилнейраминовой кислотой (NeuAc) или 5-N-ацетилнейраминовой кислотой (NeuGc), Однако при осуществлении настоящего изобретения можно использовать другие сиаловые кислоты. По поводу обзора различных форм сиаловой кислоты, пригодных для использования в соответствии с настоящим изобретением см. в общем случае, R. Schauer, Methods in Enzymology, т. 50, стр. 64-89 /1987/ и Schaur, Advances in Carbohydrate Chem. Biochem, т. 40, стр. 131-234; обе эти работы здесь включены в качестве ссылки. Как подтверждается в Примере IX ниже, сродство для рецепторов селектина является одним и тем же, если олигосахарид заканчивается в NeuAc или NeuGc. Таким образом, термин "SLX", как он здесь используется, относится к минимальному тетрасахаридному блоку (сиаловая кислота α 1,3 Gal β 1,4 [Fuc α 1,3] GlcNAc β 1,3), в которой сиаловой кислотой является NeuAc, NeuGc или другие эквивалентные формы сиаловой кислоты. Структуры, представленные здесь, которые имеют остаток сиаловой кислоты в виде NeuAc, следует понимать, как включающие и другие формы, в частности, NeuGc.
Анализ, представленный ниже, показывает, что пентасахарид, содержащий формулу
NeuAcα2, 3 Gal β 1, 4[Fuc α 1,3] GlcNAc β 1,3 Gal β, является минимальной структурой, обладающей существенно большим ингибиторным действием, чем тетрасахарид. Таким образом, в некоторых предпочтительных вариантах осуществления олигосахарид будет содержать эту пентасахаридную структуру.
Другие вариации на основе базисного SLX-блока также узнаются рецепторами селектина. Например, анализ, приведенный в Примере VIII ниже, показывает, что олигосахаридная составляющая, именуемая SY2 (известная также под названием антигена VIM-2), имеющая структуру
Neu Gc α 2, 3 Gal β 1, 4 Glc NAc β 1, 3 Gal β 1, 4 (Fuc α 1,3) Glc NAc β 1, 3 Gal β 1, 4 Glc β
связывает рецепторы селектина в той же степени, что и SLX. SY2-составляющая содержит два сиалилированных N-ацетиллактозаминовых блока, одним из которых является SLX. Таким образом, олигосахариды, узнаваемые рецепторами селектина, могут содержать несколько сиалилированных N-ацетиллактозамидовых блоков, по крайней мере один из которых фукозилирован /см., Teimeyer и др., Proc. Natl. Acad. Sci (USA), т. 88, стр. 1138-1142 /1991/, которая здесь включена в качестве ссылки.
Составляющая олигосахарида в соответствии с настоящим изобретением в предпочтительном варианте заканчивается остатком сиаловой кислоты. В некоторых вариантах осуществления остаток сиаловой кислоты может быть, кроме того, соединен с другими остатками сахаридов, такими как вторая сиаловая кислота в α 2,8-связи.
В качестве альтернативы концевой остаток сиаловой кислоты может быть заменен самыми различными радикалами. Таким образом, некоторые составляющие, связывающие селектин, в соответствии с настоящим изобретением имеют общую формулу:
R1 - NeuAc α 2,3 Gal β 1,4 Glc NAc β 1 ,
в которой R1 является R2R3C/CO2H/-, в которой R2 и R3, которые могут быть как одинаковыми, так и различными, являются H, низшим алкилом (C1-C8), гидроксил низшим алкилом (C1-C8), арилалкилом, алкоксиалкилом. Кроме того, R2 и R3 могут быть соединены, чтобы образовать 4-8 элементное карбоциклическое или гетероциклическое кольцо.
Соединения, содержащие SLX и близкие структуры, могут быть получены из клеточных поверхностных гликопротеинов или гликолипидов из нескольких типов клеток. Например, антиген SLX содержится на соединенных при помощи N углеводных группах клеточных поверхностных гликопротеинов клеток LEC11, мутанта гликозилирования CHO-клеток. LEC11 экспрессируют этот уникальный гликопептид, который содержит концевую структуру, несущую как сиаловую кислоту, так и фукозу в последовательности SLX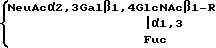
в которой
R является:
/Смотри, Stanley и др., J. Biol. Chem, т. 263, стр. 11374 /1988/, которая включена в качестве ссылки/.
Используя процедуру, описанную ниже, было показано, что мутант LEC11связывается с активированными эндотелиальными клетками сосудов человека. Ни CHO-клетки дикого типа, ни другие близкие линии мутантных CHO-клеток гликозилирования без специальной структуры гликозилирования /SLX/ не демонстрировали такого же уровня связывания.
Другие источники, которые можно использовать для получения SLX-блока, включают любую клетку, которая естественно экспрессирует эту составляющую на углеводных группах гликолипидов или гликопротеинов. Так, полиморфоядерные нейтрофилы, лимфоциты, клетка опухоли или клетки HL-60 использовали для того, чтобы очистить этот блок. Другие клетки, которые связываются с активированным эндотелием сосудов, можно также использовать для того, чтобы выделить лиганд /см. , Symington и др. , J. Jmmunol, т. 134, стр. 2498-2506 /1985/, Mizoguchi и др. , J. Biol Chem., т. 259, стр. 11949-11957 /1984/, Mizoguchi и др., J. Biol Chem., т. 259, стр. 11943-11948 /1984/, Paietta и др. , Cancer Res. т. 48, стр. 28-287 /1988/, которые включены здесь в качестве ссылки/.
Соединения, содержащие SLX или его аналоги, могут быть получены из натуральных источников, используя хорошо известные в этой области техники приемы изоляции поверхностных гликопропиенов, гликопептидов, олигосахаридов и гликолипидов из клеток /Смотри, например, Gerard, "Purification of glycoproteins" и Thomas и др. "Purification of membrane proteins", обе работы в книге Guide to Protein Purification, том 182, Methods in Enzimology /ред. Deutscher, 1990/, которые включены здесь в качестве ссылки/. Например, клетки LEC11 можно использовать для того, чтобы получить гликопротеин или гликолипид, который содержит SLX-блок, используя, например, прием, описанный у Stanley и др., см. выше. Если коротко, то LEC-11 клетки инфицируют вирусом стоматита сосудов. Затем структурные углеводные изменения, вызванные в LEC11, приводят в экспрессии на N-связанных, снабженных двумя "усами", углеводах гликопротеина G этого вируса. Вирус очищают равновесным градиентным центрифугированием, а гликопептиды очищают с использованием переваривания протеиназой, как этой описано у Stanley и др.
Для того чтобы выделить связывающую селектин составляющую из HL-60, HT-29, colo 205, нейтрофилов и линий других клеток, которые содержат лиганд, узнаваемый селектинами, используют несколько подходов. Так как лиганд в общем случае экспрессируется на поверхности клеток этих типов, один из подходов заключается в изоляции фракции мембран плазмы, обогащенной этим лигандом. После того, как мембраны плазмы изолированы, лиганды могут быть выделены, а затем идентифицированы с использованием моноклональных антител, в частности, тех, что являются химически активными относительно SLX-олихосахарида и близких структур, таких как моноклональные антитела FH6, SNH3 и CSLEX-1.
Чтобы охарактеризовать лиганд селектина на гликопротеине, высвобождение олигосахарида является в общем случае первым шагом в структурном анализе цепи олигосахарида. Этот добиваются при помощи химического расщепления протеин-углеводной связи или при помощи специфического высвобождения олигосахарида эндогликозидазой. В большинстве случаев, различные процедуры можно использовать для того, чтобы установить "правильные" условия для каждого отдельного гликопротеина. Связанные аспарагином олигосахариды высвобождают при помощи гидразинолиза, эндогликозидаз, интенсивного щелочного гидролиза и трифторацетолиза. O-связанные углеводные блоки высвобождают при помощи щелочного β -исключения. Олигосахариды отделяют от гликопептидов при помощи гелевой фильтрации. Полученные в результате олигосахариды затем отделяют друг от друга, используя комбинацию гелевой фильтрации, ВЭЖХ, тонкослойной хроматографии и ионообменной хроматографии. Изолированные олигосахариды затем целиком анализируют. Полный структурный анализ очищенных олигосахаридных блоков требует определения моносахаридных блоков, их кольцевой формы, конфигурации /D или L/, аномерной связи / α или β /, позиций связей между сахарами и их последовательности. Кроме того, устанавливают позицию всех групп-заместителей. Для того чтобы определить позиции гликозидных связей между моносахаридами, используют анализ на основе метилирования. Аномерная конфигурация остатков сахаров может быть определена с использованием 500-МГц 1H-ЯМР-спектроскопии. Условия и приемы, используемые для осуществления полного структурного углеводного анализа, описаны в общем случае в книге: Beeley, Laboratory Technigues in Biochemistry and Molecycar Biology, ред. Burdon и Knippenberg, Elsevier, Amsterdam /1985/, которая включена здесь в качестве ссылки.
Современные приемы для осуществления полной характеризации сахаров олигосахарида включают использование нескольких аналитических процедур, таких как FAB-MS /масс-спектроскопия при бомбардировке быстрыми атомами/, HPAE /хроматография анионного обмена при высоких pH/ и 1H-ЯМР. Эти приемы являются /взаимно/ дополнительными. Последние примеры того, как эти приемы используют для полной характеризации структуры олигосахарида, можно найти в анализе по Спеллмэну и др. , J. Biol. Chem., т. 264, стр. 14100 /1989/ и Стенли и др. , см. выше. Другие приемы включают масс-спектроскопию бомбардировки быстрыми атомами положительных ионов /FAB-MS/ и анализ метилирования при помощи газовой хроматографии-масс-спектроскопии соударения электронов /GC/EI-MS/ /см., Патентная заявка EPO N 893055153.2, которая здесь включена в качестве ссылки/.
Один и подходов в характеризации лиганда селектина на гликолипидах состоит в разрушении клеток, используя органические растворители, выделении гликолипидов и идентификации гликолипидов, активных относительно моноклональных антител к SLX, таких, например, как FH6, SNH3, SNH4, CSLEX-I или УТМ-2, и последующем определении структуры цепей олигосахаридов. Чтобы получить гликолипиды и ганглиозиды, которые содержат SLX, можно использовать стандартные приемы для получения гликолипидов /смотри, например, Ledeen и др., J. Neurochem т. 21, стр. 829 /1973/, которая здесь включена в качестве ссылки/. Например, гликолипиды экстрагируют из HL-60, HT-29, PMN, человеческих лейкоцитов и других линий клеток, экспрессирующих лиганд селектина при помощи приемов, в общем случае, известных каждому специалисту в этой области техники /см., например, Symington и др., J. Jmmunol. т. 134, стр. 2498 /1985/ и Macher и Beckstead, Leukemia Res, т. 14, стр. 119-130 /1990/, которые здесь включены в качестве ссылки/. Клетки выращивают в суспензии и собирают центрифугированием. Гликолипиды экстрагируют из таблетки клеток при помощи хлороформа/метанола 2: 1 и изопропилового спирта/гексана/воды 55:25:20, как это описано в Kannagi и др., J. Biol. Chem. т. 257, стр. 14865 /1982/, которая здесь включена в качестве ссылки. Полученные в результате экстракты разделяют при помощи хлороформа/метанола/воды /3:2:1/, разделение Фолха. Полученная в результате верхняя фаза экстрагирования содержит ганглиозиды, а нижняя фаза содержит гликолипиды.
Верхнюю фазу, содержащую ганглиозиды /гликосфинголипиды, которые содержат по крайней мере одну составляющую сиаловой кислоты, выделяют и отделяют в нейтральных и кислых фракциях, используя хроматографию на ДЭАЭ-Сефадексе, как это описано подробно у Ledeen и Yu, Methods Enzymol, т. 83, стр. 139 /1982/, которая здесь включена в качестве ссылки. Полученные в результате ганглиозиды собирали, подвергали лиофилизации и растворяли в хлороформе/метаноле /2: 1/. Нижняя фаза разделения Фолха содержит гликолипиды. Они разделяются и изолируются при помощи препаративной тонкослойной хроматографии, используя хлороформ/метанол/воду /60:35:8/ в качестве системы растворителей, как это описано Саймингтоном.
Чтобы идентифицировать эти ганглиозиды и гликолипиды, которые содержат лиганд селектина, осуществляли иммунохимический анализ гликолипидов в соответствии с процедурой, предложенной Magnani и др., Anal. Biochem. т. 109, стр. 399 /1980/, которая включена здесь в качестве ссылки. Если коротко, то порцию ганглиозида, описанную выше, подвергали хроматографии на тонкослойной хроматографии. Тонкослойную пластинку затем инкубировали с меченным 125I CSLEX-I или другим моноклональным антителом, которое связывается специфически с SLX или близкими структурами. После инкубирования с меченым антителом пластинку экспонировали на радиографическую пленку и проявляли. Черные точки на рентгеновской пленке соответствуют ганглиозидам, которые связываются с моноклональным антителом, и эти ганглиозиды извлекали при помощи соскабливания соответствующих областей с пластинки из окиси кремния и элюирования ганглиозидов смесью хлороформ/метанол/вода. Гликолипиды также сушат и снова суспендируют в хлороформе и проявляются в аналогичной тонкослойной системе, и зондируют радиомеченным антителом. Структурный анализ олигосахаридов, полученных из гликолипидов, осуществляли по существу, как это описано для гликопротеинов.
Олигосахариды, содержащие SLX-блок, могут быть получены из гликопротеинов с использованием приемов, известных в этой области техники /см., например. Gerard, выше, на стр. 537-539/. В общем случае, N-гликозидазу F /N-гликаназу/ используют для того, чтобы расщепить N-связанные олигосахариды в то время, как O-связанные группы расщепляются эндо- и ацетилгалактозаминадазой.
Синтетические соединения, содержащие SLX или его аналоги, присоединенные к различным составляющим, могут быть получены в зависимости от конкретного предполагаемого применения. Например, SLX можно превратить в ганглиозид при помощи связывания составляющей керамида с С-1 восстанавливающего концевого GlcNAc-блока. SLX-структуры и близкие структуры могут быть также связаны с самыми разнообразными составляющими, такими как различным образом замещенные аминогруппы, гетероциклические соединения, эфирные связи с разветвленными или неразветвленными углеводородными цепями и эфирные связи с арильными или алкиларильными составляющими. Связывающая селектин составляющая может быть также связана с различными полисахаридами, аминокислотами, аналогами аминокислот, олигопептидами или протеинами, используя приемы, хорошо известные в этой области техники.
Термин "алкил", как он здесь используется, обозначает разветвленную или неразветвленную, насыщенную или ненасыщенную углеводородную цепь, включая низшие алкилы с 1-8 атомами углерода, такие как метил, этил, н-пропил, бутил, н-гексил и т. п. , циклоалкилы /3-7 углеродов, циклоалкилметилы /4-8 атомов углерода/ и арилалкилы. Термин "арил" относится к радикалу, полученному из ароматического углеводорода при помощи удаления одного атома, например, фенила из бензола. Ароматический углеводород может иметь более одного ненасыщенного углеродного кольца, например, нафтил. Термин "алкоксил" относится к алкильному радикалу, присоединенному к остатку молекулы при помощи кислорода, например, этоксил. Термин "гетероциклическое соединение" относится к кольцевому соединению, содержащему три и более атомов, в котором по крайней мере один из атомов отличен от углерода /например, N, O, S, Se, P или As/. Примеры таких соединений включают фураны, пиримидины, пурины, пиразины и т.п. Термин "олиго" относится к полимерной молекуле, состоящей из от 2 до приблизительно 10 остатков, например, аминокислот /олигопептид/, моносахаридов /олигосахариды/ и нуклеиновых кислот /олигонуклеотид/. Термин "поли" относится к полимерной молекуле, содержащей более 10 остатков.
Для синтеза поливалентных форм связывающих селектин составляющих мономерные блоки, содержащие SLX или другие структуры, могут быть соединены с образованием молекул, имеющих от одной до примерно четырех и более связывающих селектин составляющих. Примером такой поливалентной формы является форма, в которой блоки связаны при помощи следующих составляющих: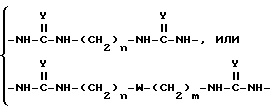
в которой
n и m, которые могут быть как одинаковыми, так и различными, являются целыми числами от 2 до 12; Y является O или S; а W является O, S или NH.
В качестве альтернативы такой составляющей является 5- - 14-элементное кольцо, содержащее два заместителя, причем каждый заместитель имеет формулы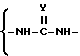
в которой
Y является O или S, заместители находятся в цис- или транс-отношении.
SLX и близкие структуры могут быть также присоединены к различным гетероциклическим соединениям /например, соединению, содержащему атомы азота/. В этом случае составляющие в предпочтительном варианте связаны с атомами азота на кольце, причем каждый азот связан с оной составляющей. Примерами гетероциклических соединений, которые пригодны для этой цели, являются пиперазин и гомопиперазин.
В качестве альтернативы поливалентные формы SLX или его аналогов могут быть созданы при помощи присоединения искомой составляющей к предварительно полученным составляющим носителя с несколькими точками для присоединения. Примеры включают присоединение SLX к аминогруппам лизина и содержащих лизин пептидов, протеинов, гликопротеинов или аспарагиновой боковой цепи таких соединений.
Еще один прием получения поливалентных, связывающих селектин соединений заключается в присоединении целевых моносахаридных остатков к полисахаридам. Например, полисахарид, который содержит повторяющийся блок, имеющий линейную структуру ядра SLX /т.е. без остатка фукозы/ может быть превращен в поливалентный SLX, содержащий полисахарид, при помощи ферментативного фукозилирования. Для этой цели используют в предпочтительном варианте типа Ia, II или III нативных полисахаридов, полученных из Streptococcus Группы B. Эти полисахариды могут быть изолированы в соответствии со стандартными приемами ил линий клеток, сданных на хранение в Американское Собрание Типов Культур /Тип Ia - АТСС N 12400 и N 31574; Тип II - АТСС N 12973 и N 31576; и Тип III - АТСС N 31577/. Смотри, например, Jennings и др. Biochem, т. 22, стр. 1258-1263 /1983/ и Патентную Публикацию PCT N 8706267; оба они здесь используются в качестве ссылки.
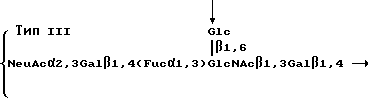
Эти полисахариды содержат повторяющиеся блоки, имеющие формулы
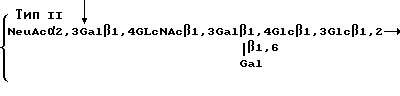
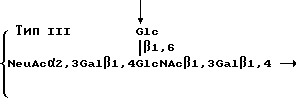
Стрелки в приведенных выше структурах указывают на основную цепь в молекуле каждого полисахарида. Как можно видеть, полисахарид типа Ia содержит повторяющийся блок, содержащий блоковую цепь, которая соответствует линейной структуре ядра SLX. В других двух полисахаридах основная цепь содержит структуру ядра SLX. Ферментативное фукозилирование этих полисахаридов, используя α 1,3 фукозилтрансферазу в соответствии со стандартными приемами, описанными ниже, дает поливалентное SLX-соединение. После фукозилирования повторяющиеся блоки имеют следующие формулы:
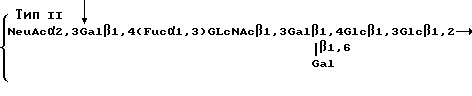
Для этой цели можно использовать целый полисахарид, а также его фрагменты. Так, можно использовать полисахариды, имеющие молекулярную массу от примерно 5000 до примерно 300000. В предпочтительном варианте молекулярная масса изменяется в области от примерно 25000 до примерно 100000. Любое число боковых цепей на полисахариде типа Ia может быть фокузилировано, чтобы полисахарид обладал активностью. В общем случае, фокузилируют от примерно 5 до примерно 200 боковых цепей, в предпочтительном варианте от примерно 50 до примерно 150.
Синтез связывающей селектин составляющей может быть осуществлен с использованием химических, ферментативных или комбинированных химических и ферментативных стратегий /смотри, например, Публикацию EPO N 319253, которая здесь включена в качестве ссылки/. В предпочтительном варианте осуществления /схема 1 ниже/ соединение, содержащее один или несколько N-ацетилглюкозаминовых блоков /GlcNAc-R/, может взаимодействовать последовательно с галактозилтрансферазой /N-ацетилглюкозамин β 1,4 галактозилтрансферазой /E.C. 2.4.1.90//, сиалилтрансферазой /Gal β 1,4 GlcNAc α 2,3 сиалилтрансферазой /E. C. 2.4.99.6/ или Gal β 1,3 GalNAc α 2,3 сиалилтрансферазой /E.C. 2.4.99.4/ и фукозилтрансферазой /N-ацетилглюкозаминид α 1,3 фукозилтрансферазой /E.C. 2.4.1.152//, чтобы получить финальные, содержащие SLX, структуры. В этом случае, R может быть несущей составляющей или активируемой промежуточной составляющей, которая будет допускать присоединение к соответствующей несущей составляющей. Каждая ферментативная реакция использует соответствующий нуклеотидный сахар в качестве донорного субстрата, чтобы получить последующие промежуточные соединения при синтезе SLX. Реакции переноса гликозила могут быть оптимальным образом осуществлены с добавленной щелочной фосфатазой /например, из кишечника коровы, CIAP/, чтобы "поглотить" побочный продукт, нуклеотид фосфат, который может ингибировать эту реакцию.
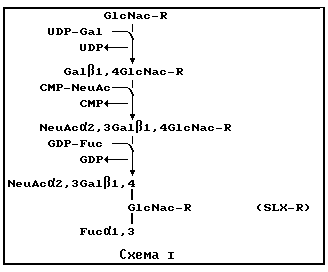
Общие условия для препаративного ферментативного синтеза углеводных групп, аналогичных SLX, известны /см., например, Toone и др., Tetrahedron т. 45, стр. 5365-5422 /1989/; Wong и др., J. Amer. Chem. Soc. т. 47, стр. 5416-5418 /1982/; Unverragt и др., J. Amer. Chem. Soc. т. 112, стр. 9308-9309 /1990/; Prieels и др., J. Biol. Chem. т. 256, стр. 10456-10463 /1981/; все эти работы включены здесь в качестве ссылки/. Каждая из ключевых ферментативных реакций известна /Beyer и др., Adv. Enzymol, т. 52, стр. 23-176 /1981/; Toone и др., см. выше; и Howard и др., J. Biol. Chem. т. 262, стр. 16830-16837 /1981/; все эти работы включены здесь в качестве ссылки/. Для препаративных реакций галактозилтрансферазу и сиалилтрансферазу очищали из натуральных источников /Beyer и др., см. выше, и Weinstein и др., J. Biol. Chem. т. 257, стр. 13835-13844 /1982/, которые здесь включены в качестве ссылки/. Фукозилтрансферазы можно также идентифицировать из натуральных источников, как это в общем случае описано Grawley и Hidsgaul, Carbohyd. Res, т. 193, стр. 249-256 /1989/, которая здесь включена в качестве ссылки. Клонировали кДНК галактозилтрансферазы и сиалилтрансферазы /Paulson и Colley, J. Biol. Chem. , т. 264, стр. 17615-17618 /1989/, которая здесь включена в качестве ссылки/, допуская продуцирование растворимых рекомбинантных ферментов для крупномасштабного препаративного синтеза /Colley и др., J. Biol. Chem. т. 264, стр. 17619-17622 /1989//.
Чтобы получить достаточные количества фукозилтрансферазы для крупно-масштабных реакций, фермент может быть клонирован и экспрессирован в форме рекомбинантного растворимого фермента каждым специалистом в этой области техники. В качестве предпочтительной процедуры РНК может быть выделена из клеток CHO дикого типа и клеток LEC11, как это описано в Chirgwin и др., Biochemistry, т. 18, стр. 5214-5299 /1979/, а поли А + РНК может быть изолирована при помощи хроматографии на олиго/dT/-целлюлозе. Далее кДНК из клеток LEC-11 может быть получена, как это описано в Sambrook и др., Molecular Cloning: A Laboratory Manual, 2-е издание /1989/, изд. Cold. Spring Harbor Press, New York, которая здесь включена в качестве ссылки. kДНК может быть выделена при помощи процедуры Дэвиса /Handbook of Experimental Immunology, т. 2, стр. 1-13 /1986//, используя избыток поли А + РНК из клеток CHO дикого типа, которые не экспрессируют целевую фукозилтрансферазу, но, тем не менее, содержат большинство иРНК-видов клеток LEC11, кДНК-библиотека может быть затем построена в векторе экспрессии CDM8, используя выделенную кДНК /Seed, Nature, т. 329, стр. 840-842 /1987//. Клоны, экспрессирующие фукозилтрансферазу, могут быть изолированы с использованием процедуры клонирования экспрессии, описанной в Larsen и др., Proc. Natl. Acad. Sci USA, т. 86, стр. 8227-8231 /1989/, применяя трансфекцию клеток COS-1 и просеивая клетки, экспрессирующие антиген SLX с антителом CSLEX или другим антителом со специфичностью для антигена SLX. Клон полной длины фукозилтрансферазы может быть затем использован для продуцирования растворимого рекомбинантного фермента, как это описано у Colley и др., см. выше.
Другим источником SLX вляется гликопротеин α1- кислоты, который является гликопротеином плазмы, углеводные составляющие которого могут быть фукозилированы, чтобы получить SLX /см., Alpha-Acidglycoprotein: Genetics, Biochemistry, Physiological Functions and Pharmacology, ред. Bauman и др. /изд. Wiley 1989/, и Walz и др., Science т. 250, стр. 1132-1135 /1990/; обе эти работы здесь включены в качестве ссылки/.
Хотя ферментативный или комбинированный химический и ферментативный синтез SLX-соединений является предпочтительным, возможен также и химический синтез, как это показано на Схемах II и IIa в конце текста описания.
Предварительно синтезировали отдельные "куски" SLX-структуры. Например, получение сиаловой кислоты, содержащей гликозиды, включая SLX, описано в Европейской Патентной Заявке N 88311312.8, которая здесь включена в качестве ссылки. Nicolaou и др. /J. Amer. Chem. Soc., т. 112, стр. 3693 /1990// описал общий синтез связанного с опухолью семейства Lex гликозфинтолипидов. Там предположен синтез защищенного трисахарида Gal β 1,4/ Fuc α 1,3/ GlcNAc /A/, как это показано на схеме II. Реакция этого промежуточного материала с соответствующим акцептором гликозила /например, составляющей спирта/ приводит к соединению /B/. Селективную депротекцию и ацетилирование глюкозаминовой составляющей осуществляли по существу, как это описано у Nicolaou и др., чтобы получить соединение /C/. Реакция /C/ с сиалилтрансферазой, как она описана выше, дает целевой продукт SLX-P, хотя он может быть получен с относительно низким выходом, используя схему II.
Модифицированные фукозиды могут быть включены в синтетические схемы, чтобы получить SLX-аналоги, которые варьируются в этой составляющей. Например, α -D-арабинозил гликозиды могут быть синтезированы, следуя известным процедурам, Nicolaou и др. , J. Amer. Chem. Soc. т. 112, стр. 3693-3695 /1990/ через использование три-О-бензил арабинозил галидов. Другие C-5 арил или алкил замещенные арабинозиловые составляющие могут быть также синтезированы, Danishefsky и др. , J. Amer. Chem. Soc. т. 107, стр. 1274 /1985/, Danishefsky, Aldrichimica Acta т. 19, стр. 59-68 /1986/ и ссылки, приведенные там, и введены в дисахариды при помощи той же процедуры. Все эти работы включены здесь в качестве ссылки.
В соответствии с альтернативной схемой IIa, с трисахарида /A/ частично снимается защита, чтобы получить /D/, который взаимодействует последовательно со сложным метиловым эфиром перацетилированной сиаловой кислоты /E/, следуя процедуре, описанной в Kameyama и др., XV Intl. Carbohyd. Symp. Abst N A096 /1990/ и Carbohydrate Res, т. 209, стр. c1-c4 /1991/ /которые здесь включены в качестве ссылки/, что дает /F/ после хроматографической очистки. Последовательная обработка /F/ метилгидразином, N-ацетилированием, O-деацетилированием и гидролиз с использованием сложного эфира дает SLX-P.
Предпочтительные примеры R для схем II и IIa включают алкил /с линейной цепью, разветвленный, насыщенный, моно- и полиненасыщенный/; серин /D или L/; полипептиды, содержащие серин; ди- и три-алканоламины /например, [HO/CH2/n] 2NH, [HO/CH2/n]3N; где n = C2-C20 в форме линейной цепи, разветвленной, ненасыщенной, моно- и поли-ненасыщенной/. R может быть также арилом, замещенным арилом /например, Me, OH, I; отдельно или в комбинации, включающей 125I/, алкиларилом, арилалкилом или другой составляющей, которую может включить любой специалист в этой области техники в зависимости от применения. Введение иода в феноловое соединение, такое как тирозин, известно в этой области техники. Радикальные группы, содержащие фенолы, используют для введения радиоизотопа 125I, чтобы получить соединения, которые используют в диагностике.
Соединения, содержащие SLX и близкие структуры, можно также использовать для того, чтобы осуществить анализ на присутствие других соединений, которые способны ингибировать межклеточную адгезию с участием селектинов. Несколько приемов можно использовать для того, чтобы проанализировать биологическую активность испытуемых соединений на способность ингибировать реакцию, передаваемую с участием селектина. В идеале анализы в соответствии с настоящим изобретением допускают крупно-масштабное in vitro или in vivo просеивание самых разных соединений.
Агентом или испытуемым соединением, подлежащим просеиванию, будет в общем случае синтетическая или естественно продуцируемая биомолекула, такая как пептид, полипептид, протеин /например, моноклональное антитело/, углевод /например, олигосахарид/, гликоконъюгат, нуклеиновая кислота и т.п. Эти соединения продуцируют синтетически, используя, например, приемы для синтеза олигосахаридов, описанные выше /см., также Khadem, Carbohydrate Chemistry /Academic Press, San Diego CA, 1988/, которая здесь включена в качестве ссылки/. Различные приемы для синтеза полипептидов определенного состава хорошо известны в этой области техники /см., Atherton и др., Solid Phase Peptide Synthesis /IRL Press, Oxford 1989/, которая здесь включена в качестве ссылки/. Если синтетические испытуемые соединения являются полимерными /например, полипептиды или полисахариды/, они в предпочтительном варианте модифицируются систематическим путем с тем, чтобы идентифицировать последовательность мономеров, которые имеют целевой эффект /см., например, Патент США N 4833092, который здесь используется в качестве ссылки/. Испытуемые соединения могут быть также выделены из любого натурального источника, такого как животное, растение, грибы или бактериальные клетки в соответствии со стандартными процедурами, как это описано выше. Потенциально полезные моноклональные антитела могут быть получены в соответствии со стандартными приемами, описанными более подробно ниже.
Анализы в соответствии с настоящим изобретением особенно полезны при идентификации соединений, которые действуют в качестве антагонистов или агонистов молекулы лиганда. Антагонистами являются соединения, которые обращают физиологический эффект лиганда или исключают связывание лиганда с рецептором. Антагонист в общем случае конкурирует непосредственно или косвенно с лигандом за счет связывания рецептора и, таким образом, снижает долю молекул лиганда, связывающихся с рецептором. В общем случае, антагонист будет топографически эквивалентным натуральному лиганду и будет конкурировать непосредственно с лигандом за сайт связывания на селектине. Такие соединения именуются здесь "аналогами" /"заменителями"/. Аналогом SLX является молекула, которая структурно и функционально служит в качестве заменителя SLX-составляющей в том, что он узнаваем рецептором селектина. В качестве альтернативы, если лиганд и испытуемое соединение могут связывать рецептор одновременно, это соединение может действовать неконкурирующим образом. Неконкурирующий ингибитор действует при помощи уменьшения или ингибирования последующих физиологических эффектов взаимодействия рецептор-лиганд, а не как агент, снижающий долю молекул лиганда, связывающихся с рецептором. Наконец, анализы в соответствии с настоящим изобретением могут быть использованы для того, чтобы идентифицировать синтетически или естественно встречающиеся агонисты, то есть, соединения, которые связывают рецептор и инициируют физиологическую реакцию, аналогичную натуральному лиганду.
Многочисленные прямые и непрямые способы для in vitro просеивания ингибиторов взаимодействий лиганд-рецептор доступны и известны каждому специалисту в этой области техники. Например, можно определить способность ингибировать адгезию SLXX-несущих клеток с клетками, экспрессирующими специфический селектин. Как уже было указано выше, гены рецептора селектина предварительно клонировали, поэтому эти гены могут быть вставлены и экспрессированы в самых разнообразных клетках, таких как COS-клетки, CHO-клетки и т. п. Кроме того, клетки, которые обычно не экспрессируют SLX, способны трансформироваться одним или несколькими генами гликозилтрансферазы, которые наделяют трансформированные клетки способностью синтезировать лиганд. /См., например, Lowe и др. , Cell, т.63, стр. 475-484 /1990/, которая включена здесь в качестве ссылки/. В общем случае, испытуемое соединение или агент инкубируют с клетками, несущими меченый SLX, и активированными эндотелиальными клетками, иммобилизованными из твердой поверхности. Ингибирование клеточной адгезии далее определяют при помощи обнаружения метки, связанной с этой поверхностью после соответствующей промывки. В анализах, приведенных в примерах, помещенных ниже, использовали промиелоцитные клетки HL-80 и активированные эндотелиальные клетки человека или активированные тромбоциты.
Так как в настоящее время идентифицирован лиганд, специфический относительно рецепторов селектина, молекулы изолированного лиганда можно также использовать в анализах. Термины "изолированный агент связывания селектина" или "составляющая изолированного SLX", как они здесь используются, относятся к связывающему селектин соединению, которое находится в состоянии, отличном от своего нативного состояния, например, оно не ассоциировано с клеточной мембраной клетки, которая обычно экспрессирует лиганд. Таким образом, составляющая изолированного SLX может быть компонентной изолированной молекулы, такой как олигосахарид или гликоконъюгат. Изолированная молекула может быть синтезирована или получена из мембран несущих SLX клеток. В качестве альтернативы, изолированный, связывающий селектин, агент или SLX-составляющая может быть ассоциирована с липосомой или присоединена к твердой поверхности перед использованием в этом анализе. Приемы для получения связывающих селектин липосом и для иммобилизации различных биомолекул подробно обсуждаются ниже.
В общем случае in vitro анализы в соответствии с настоящим изобретением являются конкурирующим анализами, которые обнаруживают способность испытуемого соединения к конкурирующему ингибированию связывания соединения, о котором известно, что оно связывает либо рецептор, либо лиганд. Ингибирование связывания между SLX и рецептором селектина испытывают в общем случае. Можно также испытывать ингибирование других связывающих взаимодействий, например, ингибирование связывания между моноклональным антителом /например, CSLEX-I/ и SLX или между SLX-заменителем и ингибитором селектина. Известны многочисленные типы конкурирующих анализов /см., например, Патенты США NN 3 376 110, 4 016 043, и Harlow и Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Publications, N.Y. /1988/. которые здесь включены в качестве ссылки/.
Анализы в соответствии с настоящим изобретением пригодны также для измерения связывание испытуемого соединения только с одной компонентой, а не используя конкурирующий анализ. Например, для идентификации соединений, которые содержат SLX-составляющую, можно использовать иммуноглобулины. Можно использовать стандартные процедуры для анализа на моноклональные антитела, такие как ELISA /см. , Harlow и Lane выше/. Когда осуществляют анализ на гликолипиды, содержащие SLX-антиген, химическую активность моноклонального антитела с антигеном можно анализировать при помощи TXC-иммуно- окрашивания при помощи приемов, первоначально описанных в Magnani и др., Anal. Biochem. т. 109, стр. 339-402 /1980/, или радиоиммуноанализа в твердой фазе, как это описано Kanagi и др. , Cancer Res, т.43, стр. 4997-5005 /1983/; которые включены здесь в качестве ссылки. Гликопротеины можно анализировать при помощи стандартных приемов иммуноблотирования, как это описано в статье Harlow и Lane, см. выше. Пригодны также форматы анализов типа "сэндвич" /см., например, Патенты США NN 4 642 285, 4 299 916 и 4 391 904; и Harlow и Lane, см. выше; все эти материалы включены здесь в качестве ссылки/. В общем случае, соединения, которые были идентифицированы в анализе на связывание, могут быть далее испытаны на определение их способности ингибировать взаимодействия рецептор-лиганд.
Другие форматы анализов включают обнаружение присутствия или отсутствия различных физиологических изменений либо в несущих лиганд, либо несущих селектин клетках, которые являются результатом взаимодействия. Примеры таких анализов включают измерение изменений в активности по транскрипции, вызванной связыванием /смотри, например, публикацию EPO N 3712820/, обнаружение различных внеклеточных эффектов с участием клеток, см., например, Публикацию PCT N 90/00503/ и обнаружение изменений в потенциале мембран отдельных клеток /см., например, Патент США N 4 343 782/; все эти работы включены здесь в качестве ссылок. В качестве альтернативы, можно обнаруживать структурные изменения в изолированных рецепторах или лигандах; см., например, Патент США N 4 859 609, который здесь включен в качестве ссылки.
Любую компоненту анализа, включая лиганд, рецептор или испытуемое соединение, можно связать с твердой поверхностью. В этой области техники известны многочисленные приемы иммобилизации биомолекул на твердых поверхностях. Например, твердой поверхностью может быть мембрана /например, нитроцеллюлоза/, микротитрованная чашка /например, ПВХ или полистирол/ или шарик. Целевая компонента может быть ковалентно связана или нековалентно присоединена через неспецифическое связывание.
В качестве материала для твердой поверхности можно использовать самые разнообразные органические и неорганические полимеры, как натуральные, так и синтетические. Примеры таких полимеров включают полиэтилен, полипропилен, поли/4-метилбутен/, полистирол, полиметакрилат, поли/этилен терфталат/, искусственный шелк, найлон, поли/винил бутират/, силиконы, полиформальдегид, целлюлозу, ацетат целлюлозы, нитроцеллюлозу и т.д. Другие материалы, которые можно использовать, включают бумагу, стекло, керамику, металлы, металлоиды, полупроводниковые материалы, металлокерамику и т.п. Кроме того, они включают материалы, которые образуют гели, такие как протеины, например, желатины, липополисахариды, силикаты, агарозу и полиакриламиды и полимеры, которые образуют несколько водных фаз, такие как декстраны, полиалкилен гликоли /алкилен с 2-3 атомами углерода/ или поверхностно-активные агенты, например, амфифильные соединения, такие как фосфолипиды, аммониевые соли алкила с длинной цепью /12-24 атома углерода/ и т.п. Так, где твердая поверхность пористая, можно использовать различные размеры пор, в зависимости от природы системы.
При получении поверхности можно использовать самые разнообразные материалы, в частности, ламинаты, чтобы получить различные свойства. Например, можно использовать протеиновые покрытия, такие как желатин, с тем, чтобы избежать неспецифического связывания, упростить ковалентную конъюгацию, усилить сигнал обнаружения и т.п..
Если желательно получить ковалентное связывание между соединением и поверхностью, поверхность может быть в общем случае полифункциональной или способной быть полифункциональной. Функциональные группы, которые могут содержаться на поверхности и использованы для связывания, могут включать карбоновые кислоты, альдегиды, аминогруппы, циано группы, этиленовые группы, гидроксильные группы, меркапто группы и т.п. Механизм связывания разнообразных соединений с различными поверхностями хорошо известен и в литературе имеются многочисленные его иллюстрации. Смотри, например, "Immobilized Enzymes "Ichiro Chibata Halsted Press, New York, 1978, и Cuatrecasas, J. Biol. Chem т.245, стр. 3059 /1970/, которые здесь включены в качестве ссылки.
В дополнение к ковалентному связыванию, можно использовать различные приемы для нековалентного связывания аналитической компоненты. Нековалентное связывание является в общем случае неспецифическим поглощением соединения на поверхности. В общем случае, поверхность блокируется вторым соединением, чтобы предотвратить неспецифическое связывание меченых компонент анализа. В качестве альтернативы, поверхность создается такой, что она неспецифически связывается с одной компонентой, но по существу не связывается с другой. Например, поверхность, несущая лектин, такой как Конканавалин A будет связываться с содержащим углевод соединением, но не с меченым протеином, который не был подвергнут гликозилированию. Различные твердые поверхности для использования в нековалентном присоединении аналитических компонент описаны в Патентах США NN 4 447 576 и 4 254 082, которые включены здесь в качестве ссылки.
Форматы многих анализов используют меченые компоненты анализа, такие как SLX-лиганды, SLX-аналоги, иммуноглобулины, рецепторы или испытуемые соединения. Эта метка может быть связана непосредственно или косвенно с целевой компонентной анализа в соответствии с приемами, хорошо известными в этой области техники. Можно использовать самые различные метки. Эта компонента может быть мечена любым из нескольких известных приемов. Наиболее известный прием обнаружения состоит в использовании авторадиографии с меченными 3H, 125I, 35S, 14C или 32P соединениями и т.п. Выбор радиоактивного изотопа зависит от предпочтений исследователя в простоте синтеза, вариации стабильности и периоде полураспада выбранных изотопов. Другие нерадиоактивные метки включают лиганды, которые связываются с мечеными антителами, флуорофорами, хемилюминесцентными агентами, ферментами и антителами, которые служат в качестве элементов специфической связывающей пары для меченого лиганда. Выбор метки зависит от необходимой чувствительности, простоты конъюгации с соединением, требований стабильности и имеющегося в распоряжении оборудования.
Нерадиоактивные метки часто присоединяют при помощи непрямых средств. В общем случае молекула лиганда /например, биотин/ ковалентно связана с молекулой. Лиганд далее связывается с антилигандом /например, стрептавидином/, который является либо наследственно обнаруживаем, либо ковалентно связывается с сигнальной системой, такой как обнаруживаемый фермент, флуоресцентное соединение или хемилюминесцентное соединение. Лиганды и антилиганды могут быть самыми разнообразными. Так, где лиганд содержит натуральный антилиганд, например биотин, тироксин и кортизол, его можно использовать в конъюгации с мечеными, встречающимися в природе, антилигандами. В качестве альтернативы, в комбинации с антителом можно использовать любое гаптеновое или антигенное соединение.
Эти молекулы могут быть конъюгированы непосредственно с формирующими сигнал соединениями, например, при помощи конъюгации с ферментом или флуорофором. Ферментами, представляющими интерес в качестве меток, являются главным образом гидролазы, в частности фосфатазы, эстеразы и глюкозидазы, или оксидоредуктазы, в частности пероксидазы. Флуоресцентные соединения включают флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон и т. д. Хемилюминесцентные соединения включают люциферин и 2,3-дигидрофталазиндионы, например, луминол. По поводу обзора различных, продуцирующих сигнал, систем, которые могут быть использованы, смотри Патент США N 4 391 904, который здесь включен в качестве ссылки.
Как уже обсуждалось выше, в дополнение к соединениям различных ингибиторов, которые содержат доступный SLX-блок или SLX-аналог, в соответствии с настоящим изобретением также предлагаются моноклональные антитела, способные ингибировать межклеточную адгезию, с участием селектинов, а также способы идентификации таких антител. Моноклональные антитела связываются с лигандом селектина или рецептором, и блокируют клеточную адгезию. Таким образом, самые разнообразные приемы, известные каждому специалисту в этой области техники, для продуцирования и манипулирования с молекулами различных иммуноглобулинов могут быть применены для ингибирования межклеточной адгезии.
Термин "иммуноглобулин", как он здесь используется, относится к прототипу, состоящему из одного или нескольких полипептидов, которые по существу закодированы генами иммуноглобулина. Гены "узнаваемого" иммуноглобулина включают гены постоянных зон каппа, лямбда, альфа, гамма, дельта, эпсилон и мю, а также мириады генов вариабельных зон иммуноглобулина. Иммуноглобулины могут существовать в самых разнообразных формах помимо антител, включая, например Fv, FAB и F(ab)2, а также одинарные цепи /например, Huston и др., Proc. Nat. Acad. Sci. USA, т. 85, стр. 5879 - 5883 /1988/ и Bird и др., Science, т. 242, стр. 423 - 426 /1988/, которые включены здесь в качестве ссылки/. /см., в общем случае, Hood и др., Immunology, 2-е изд., изд. Benjamin, N.Y. /1984/ и Hunkapiller и Hood Nature т. 323, стр. 15 - 16 /1986/, которые здесь включены в качестве ссылки/.
Антитела, которые связывают SLX-антиген, могут продуцироваться самыми разнообразными средствами. Продуцирование нечеловеческих моноклональных антител, например, мышиных, lagomorpha, лошадиных и т.д. хорошо известно и может быть осуществлено при помощи, например, иммунизации животного SLX-антигеном или препарацией, содержащей гликопротеин или гликолипид, содержащий антиген. Продуцирующие антитела клетки, полученные из иммунизированных животных, выделяли и просеивали или сначала просеивали на продуцирование антитела, которое ингибирует взаимодействие протеина вирусной поверхности с молекулой рецептора, а затем выделяли. За обсуждением общей процедуры продуцирования моноклональных антител см. Harlow и Lane, Antibodies, A Laboratory Manual /1988/, см. выше.
Образование человеческих моноклональных антител относительно человеческого антигена /в случае SLX-блока, выделенного из человеческой ткани/ может оказаться трудным делом при использовании известных приемов. Поэтому может оказаться желательным перенести зоны связывания антигена нечеловеческих антител, например, F/ab'/2 или гипервариабельные зоны в человеческие постоянные зоны /Fc/, или оснащенные зоны при помощи приемов рекомбинантной ДНК, чтобы продуцировать по существу человеческие молекулы. Такие приемы в общем случае известны в этой области техники и описаны, например, в Патенте США NN 4 816 397, Публикациях EP NN 173 494 и 239 400, которые здесь включены в качестве ссылки. В качестве альтернативы, можно выделить ДНК-последовательности, которые индуцируют человеческое моноклональное антитело или его доли, которое специфически связывается с человеческим SLX, при помощи просеивания ДНК-библиотеки из клеток B человека в соответствии с общим описанием, кратко приведенным в Huse и др., Science т. 246, стр. 1275 - 1281 /1989/, которая здесь включена в качестве ссылки, а затем клонирования и амплификации последовательностей, которые кодируют антитело /или связывающий фрагмент/ с целевой специфичностью.
Несколько доступных в настоящее время моноклональных антител можно использовать в соответствии с настоящим изобретением для того, чтобы ингибировать межклеточную адгезию с участием селектинов. Например, CSLEX-I /см., Campbell и др., J. Biol. Chem. т. 259, стр. 11208 - 11214 /1984//, YIM-2, который узнает последовательность, немного отличающуюся от SLX /см., Macher и др. , см. выше/, FH6 /описанный в патенте США N 4 904 596/ /все эти работы включены здесь в качестве ссылки/ или SH3 и SH4, полученные д-ром S. Hakomori из Института Биомембран в Сиэттле, Вашингтон.
Соединения, являющиеся предметом настоящего изобретения, включая иммуноглобулины, можно использовать при получении фармацевтических форм, как это подробно обсуждается ниже. Если соединением является олигосахарид или гликоконъюгат, SLX или SLX-заменяющая составляющая может быть представлена самыми разнообразными формами, но она должна быть способна эффективно связываться с рецептором селектина, таким как ELAM-1, GMP-140 или MEL-14 антиген и при этом ингибировать межклеточную адгезию.
Фармацевтические композиции, являющиеся предметом настоящего изобретения, можно использовать для того, чтобы блокировать или ингибировать клеточную адгезию, ассоциированную с несколькими из заболеваний. Например, несколько воспалительных заболеваний связаны с селектинами, экспрессируемыми на эндотелиальных клетках сосудов и тромбоцитах. Термин "воспаление" используется здесь для того, чтобы именовать реакции обеих, специфической и неспецифической, систем защиты. Реакцией специфической системы защиты является специфическая реакция иммунной системы на антиген. Пример специфических реакций системы защиты включает реакцию антитела на антигены, такие как вирусы и гиперчувствительность запаздывающего типа. Неспецифическая реакция системы защиты является воспалительной реакцией, передающейся лейкоцитами, которые в общем случае лишены иммунологической памяти. Такие клетки включают макрофаги, эозинофилы и нейтрофилы. Примеры неспецифических реакций включают немедленное вздутие после укуса и накопление лейкоцитов PMN в местах бактериальных инфекций /например, легочные инфильтраты при бактериальной пневмонии и образование гноя при абсцессах/.
Другие нарушения, которые можно лечить в соответствии с настоящим изобретением, включают ревматоидные артриты, послеишемические разрушения ткани с участием лейкоцитов /нарушения при повторяющейся перфузии/, поражения или шок при обморожениях, острое /с участием лейкоцитов/ заболевание легких /например, синдром дистресса дыхания у взрослых/, астму, травматический шок, септический шок, нефриты и острые и хронические воспаления, включая атопические дерматиты, псориазы и воспалительное заболевание кишечника. В соответствии с настоящим изобретением можно лечить также различные патологии с участием тромбоцитов, такие как атеросклероз и образование сгустков. Кроме того, можно ингибировать или предотвратить образование метастаз опухоли при помощи ингибирования адгезии циркулирующих клеток рака. Примеры включают карциному толстой кишки и меланому.
При помощи примера было установлено, что поражения в результате повторной перфузии особенно восприимчивы к лечению композициями, являющимися предметом настоящего изобретения. Композиции, которые ингибируют взаимодействие селектин GMP-140-лиганд, могут быть особенно полезными при лечении или предотвращении поражений при повторной перфузии. Настоящее изобретение может быть использовано профилактически перед хирургией сердца, чтобы ускорить постхирургическое восстановление.
Ввиду того, что GMP-140 хранится в телах Вейбела-Палады тромбоцитов и эндотелиальных клетках, и высвобождается при активации тромбином, чтобы участвовать в адгезии нейтрофилов и моноцитов, ингибиторы взаимодействия с GMP-140-лиганд могут быть особенно эффективными при минимизации поражений ткани, которые часто сопровождаются тромботическими нарушениями. Например, такие ингибиторы могут представлять терапевтическую ценность для пациентов, которые недавно перенесли удар, инфаркт миокарда, глубокий тромбоз вен, эмболию легких и т.д. Эти соединения особенно полезны в профилактической терапии.
Композиции, являющиеся предметом настоящего изобретения, находят, в частности, использование при лечении вторичных эффектов септического шока или рассеянной внутрисосудистой коагуляции /DIC/. Миграция лейкоцитов в ткани во время септического шока или DIC часто приводит к патологическому разрушению ткани. Кроме того, эти пациенты могут иметь широко распространенное воспаление от микроциркулирующего тромба и диффузии. Терапевтические композиции, предлагаемые в соответствии с настоящим изобретением, ингибируют миграцию лейкоцитов в эти места и смягчают нарушение ткани.
Ингибиторы взаимодействия селектин-лиганд также полезны при лечении травмартического шока и острых тканевых поражений, которые связаны с ним. Ввиду того, что селектины играют роль пополнения лейкоцитов в места поражения, в частности, ELAM-I в случаях острых поражений и воспалений, их ингибиторы могут быть применены локально или систематически, чтобы контролировать повреждение ткани, ассоциированное с такими поражениями. Кроме того, ввиду специфичности таких ингибиторов для мест воспаления, например, где экспрессируются рецепторы ELAM-I, эти композиции будут более эффективными и маловероятно, что они вызовут осложнения при сравнении с известными противовоспалительными агентами.
Таким образом, в соответствии с настоящим изобретением предлагаются также фармацевтические композиции, которые могут быть использованы при лечении вышеупомянутых заболеваний. Эти фармацевтические композиции состоят из биомолекул или других соединений, которые содержат SLX-блок, антитела, которые связываются с SLX, или другие соединения, которые ингибируют взаимодействие между SLX-лигандом и рецепторами селектина, вместе с приемлемыми с фармацевтической точки зрения носителями. Биомолекула в соответствии с настоящим изобретением может быть пептидом, полипептидом, протеином /например, иммуноглобулином/, углеводом /например, олигосахаридом или полисахаридом/, гликоконъюгатом /например, гликолипидом или гликопротеином/, нуклеиновой кислотой и т. п. Эти фармацевтические композиции пригодны для использования в самых разнообразных системах для доставки лекарственных препаратов. По поводу краткого обзора современных способов получения средств доставки препаратов, см. , Langer, Science т. 249, стр. 1527 - 1533 /1990/, которая здесь включена в качестве ссылки.
Ввиду сложности воспалительной реакции у млекопитающих, каждый специалист в этой области техники должен понять, что фармацевтические композиции, являющиеся предметом настоящего изобретения, могут содержать несущие SLX соединения в смеси с другими соединениями, о которых известно, что они препятствуют реализации функции других молекул клеточной адгезии. Например, члены семейства интегрина молекул адгезии видимо играют роль в экстравазате лейкоцитов в местах инфекции. По поводу обзора рецепторов межклеточной адгезии, включающего рецепторы селектина, и их роли в иммунной функции, смотри Springer, Nature, т. 3463, стр. 425 - 434 /1990/, которая здесь включена в качестве ссылки. Кроме того, последовательное лечение с использованием фармацевтических композиций, являющихся предметом настоящего изобретения, может быть также определено состоянием развития заболевания, подлежащего лечению. Так как различные молекулы адгезии могут быть отрегулированы сверху или снизу в том, что касается реакции на различные факторы во время заболевания или симптомов, каждый специалист в этой области техники должен знать, что различные фармацевтические композиции могут потребоваться для лечения различных воспалительных состояний.
В одном из вариантов осуществления SLX-лиганд фармацевтической композици можно использовать для "нацеливания" известных антивоспалительных препаратов или других агентов на определенные места поражений ткани. При помощи использования связывающей селектин составляющей, такой как SLX-лиганд или SLX-заменитель, чтобы "нацелить" лекарственный препарат на рецептор селектина, например, на эндотелиальных клетках сосудов, можно добиться более высоких концентраций таких препаратов в местах поражений. Побочные эффекты от применения известных антивоспалительных хемотерапевтических агентов могут быть существенно смягчены при помощи более низких доз, локализацией агента в местах повреждений и/или заключением агента в капсулу перед его доставкой.
"Целевая" компонента, т.е. SLX-лиганд или SLX-заменитель, который ввязывается с целевым селектином, может быть непосредственно или косвенно "спарен" с хемотерапевтическим агентом. Спаривание, которое может быть осуществлено при помощи средств, в общем случае известных в этой области техники, не должно по существу ингибировать способность лиганда связываться с рецептором и не должно по существу снижать активность хемотерапевтического агента. Самые разнообразные хемотерапевтические агенты могут быть спарены для "нацеливания". Например, противовоспалительные агенты, которые могут быть спарены, включают SLX-несущие соединения, являющиеся предметом настоящего изобретения, иммуномодуляторы, антагонисты фактора, активирующего тромбоциты /PAF/, ингибиторы циклооксигеназы, ингибиторы липоксигеназы и антагонисты лейкотриена. Некоторые предпочтительные составляющие включают циклоспорин A, индометацин, напроксен, FK-506, микофеноловую кислоту и т.д. Аналогичным образом антиоксиданты, например дисмутазу суперокиси, используют для лечения поражений при повторных перфузиях, когда "нацеливание" осуществляют при помощи SLX-лиганда или заменителя. Аналогично, противораковые агенты можно нацеливать при помощи спаривания SLX-лиганда или заменителя с хемотерапевтическим агентом. Примеры агентов, которые могут быть спарены, включают дауномицин, доксорубицин, винбластин, блеомицин и т.д.
"Нацеливание" рецептора селектина может быть также осуществлено через двойственные пути или молекулы двойственного характера /полярный : неполярный/, которые существуют в виде агрегатов в водном растворе. Эти двойственные пути включают неполярные липиды, полярные липиды, моно- и диглицериды, сульфатиды, лизолецитин, фосфолипиды, сепонин, желчные кислоты и соли. Эти молекулы могут существовать как эмульсии и пены, мицеллы, нерастворимые монослои, жидкие кристаллы, фосфолипидные дисперсии и пластинчатые слои. Они здесь в общем случае именуются как липосомы. В эти препарации лекарственный препарат, подлежащий доставке, включается в качестве части липосомы вместе с SLX-лигандом или заменителем, который связывается с рецептором селектина. Таким образом, липосомы, заполненные необходимым хемотерапевтическим агентом, могут быть направлены на место повреждения ткани при помощи взаимодействия селектин-SLX-лиганд. Когда липосомы попадают в области, близкие к поврежденным клеткам, они высвобождают выбранные терапевтические композиции.
Липосомы, являющиеся предметом настоящего изобретения, образуются из стандартных, образующих носитель, липидов, которые в общем случае включают нейтральные и отрицательно заряженные фосфолипиды и стерол, такой как холестерин. Выбор липидов в общем случае диктуется такими факторами, как размер липосомы и стабильность липосом в кровяном потоке.
В общем случае основной компонентой липида в липосомах является фосфатидилхолин. Фосфатидилхолины, содержащие самые разнообразные группы ациловых цепей с различными длинами цепей и степенью насыщения, доступны или могут быть изолированы или синтезированы при помощи хорошо известных приемов. В общем случае, более насыщенным фосфатидилхолинам более просто придать необходимые размеры, в частности, соли липосомы должны иметь размеры меньше примерно 0,3 мкм для целей стерилизации фильтром. Приемы, используемые для придания необходимых размеров липосомам и свойств стерилизации фильтрами, описаны ниже. Состав ациловых цепей фосфолипида может также нарушать стабильность липосом в крови. Одним предпочтительным фосфатидилхолином является частично гидрогенизированный фосфатидилхолин яиц.
"Нацеливание" липосом с использованием самых разнообразных агентов "нацеливания" /например, лигандов, рецепторов и моноклональных антител/ хорошо известно в этой области техники /см., например, Патенты США NN 4 957 773 и 4 603 044, которые здесь включены в качестве ссылки/. В качестве "нацеливающих" агентов можно использовать гликопротеины и гликолипиды с самой различной молекулярной массой. В общем случае используют гликопротеины, имеющие молекулярную массу менее примерно 300000 дальтон, в предпочтительном варианте от примерно 40000 до примерно 250000, в более предпочтительном варианте от примерно 75000 до примерно 150000. Используют гликолипиды с молекулярной массой менее примерно 10000 дальтон, в предпочтительном варианте от примерно 600 до примерно 4000 дальтон.
Можно использовать стандартные приемы для "спаривания" нацеливающих агентов с липосомами. Эти приемы в общем случае включают введение в липосомы жидких компонентов, таких как фосфатидилэтаноламин, которые могут быть активированы для присоединения нацеливающих агентов или полученных липофильных соединений, таких как липид, из которого получают блеомицин. Липосомы, нацеленные на антитела, могут быть сконструированы с использованием, например, липосом, которые содержат протеин A /см., Renneisen и др., J. Biol. Chem. т. 65, стр. 16337-16342 /1990/ и Leonetti и др., Proc. Natl. Acad. Sci. USA т. 87, стр. 2448 - 2451 /1990/, которые здесь включены в качестве ссылки/.
Механизмы прицеливания в общем случае требуют, чтобы "нацеливающие" агенты были размещены на поверхности липосомы таким образом, чтобы "целевые" агенты были доступны для взаимодействия с рецептором селектина. Липосома в общем случае конструируется таким образом, что часть соединительного средства сначала включается в мембрану во время образования мембраны. Часть соединительного средства должна иметь липофильную часть, которая прочно вложена и закреплена в мембране. Оно должно также иметь гидрофильную часть, которая химически доступна на водной поверхности липосомы. Гидрофильную часть выбирают так, чтобы она была химически пригодной для образования стабильной химической связи с нацеливающим агентом, который добавляют позже. Таким образом, молекула соединительного средства /коннектора/ должна иметь и липофильное закрепление и гидрофильную химически активную группу, пригодную для взаимодействия с агентом-целью и удержания агента-цели в его правильном положении, начиная с поверхности липосомы. В некоторых случаях имеется возможность присоединить агент-гель непосредственно к молекуле коннектора, но в большинстве случаев более удобно использовать третью молекулу, которая действует в качестве химического мостика, который, таким образом, связывает молекулу коннектора, которая находится в мембране, с агентом-целью, который выступает в трехмерном пространстве на поверхности пузырька.
Заряд липосомы является важной детерминантой при очистке липосом от крови, причем отрицательно заряженные липосомы отбираются быстрее при помощи ретикулоэндотелиальной системы /Juliano, Biochem. Biophys. Res. Commun. т. 63, стр. 651 /1975// и, таким образом, имеют более короткий период полураспада в кровяном потоке. Липосомы с более продолжительной циркуляцией в общем случае желательны для терапевтического или диагностического использования. Липосомы, которые могут поддерживаться от 8, 12 или до 24 часов в кровяном потоке обеспечивают устойчивое высвобождение ингибиторов селектина-лиганда, являющихся предметов настоящего изобретения, или могут облегчить нацеливание ингибиторов /в которые могут быть введены метки, чтобы обеспечить in vivo диагностическое представление/ на целевые места перед тем, как они будут удалены при помощи ретикулоэндотелиальной системы.
В общем случае липосомы получают с примерно 5-15 молярными процентами отрицательно заряженных фосфолипидов, таких как фосфатидилглицерин, фосфатидилсерин или фосфатидилинозитол. Добавляемые отрицательно заряженные фосфолипиды, такие как фосфатидилглицерин служат также для того, чтобы предотвратить спонтанное агрегирование липосом и, таким образом, минимизировать риск нежелательного образования агрегатов липосом. Отверждающие мембраны агенты, такие как сфингомиелин и насыщенный нейтральный фосфолипид в концентрации не менее примерно 50 молярных процентов и 5-15 молярных процентов монозиалилганглиозида могут обеспечить более высокое циркулирование препарации липосомы в кровяном потоке, как это в общем случае описано в Патенте США N 4837028, который здесь включен в качестве ссылки.
Кроме того, суспензия липосом может включать защищающие липид агенты, которые защищают липиды и компоненты лекарственного препарата против повреждений свободными радикалами и в результате окисления жиров при хранении. Предпочтительными являются липофильные свободно-радикальные прерыватели реакций, такие как альфатокоферол и растворимые в воде, специфические относительно железа, соединения, образующие хелаты, такие как ферриоксианин.
Самые разнообразные приемы можно использовать для приготовления липосом, как это описано, например, в Szoka и др., Ann. Rev. Biophys. Bioeng т.9, стр.467 /1980/, Патент США N 4235871, 4501728 и 4837028, которые здесь включены в качестве ссылки. Один такой прием дает возможность получить многослойные пузырьки гетерогенных размеров. В этом приеме липиды, образующие пузырек, растворяют в соответствующем органическом растворителе или системе растворителей и сушат под вакуумом или инертным газом, чтобы образовать тонкую липидную пленку. Если желательно, то пленку можно снова растворить в соответствующем растворителе, таком как третичный бутанол, а затем лиофилизовать, чтобы получить более гомогенную липидную смесь, которая находится в более легко гидратируемой форме, напоминающей порошок. Эту пленку покрывали водным раствором "нацеливаемого" лекарственного препарата и "нацеливающей" компоненты и давали возможность гидратироваться в общем случае в течение 15-60 минут при перемешивании. Распределения по размеру полученных в результате многослойных пузырьков может быть сдвинуто в направлении меньших размеров при помощи гидратирования липидов при условиях более энергичного перемешивания или при помощи добавления солюбилизирующих детергентов, таких как деоксихолат.
Среда для гидратирования содержит нацеливаемый лекарственный препарат в концентрации, которая желательна во внутреннем объеме липосом в финальной суспензии липосом. В общем случае раствор лекарственного препарата содержит 10-100 мг/мл в буферированном соляном растворе. Концентрация нацеливающей SLX-молекулы или заменителя, которая связывает с селектином, в общем случае составляет примерно 0,1-20 мг/мл.
Следуя процедуре получения липосомы, липосомы могут иметь необходимую область размеров и относительно узкое распределение размеров липосом. Одна предпочтительная область размеров составляет примерно 0,2-0,4 мкм, которые позволяют суспензию липосом стерилизовать фильтрацией через известный фильтр, в общем случае фильтр в 0,22 мкм. Процедура стерилизации фильтром может быть осуществлена на основе высокой проницаемости, если липосомы должны иметь размер примерно 0,2-0,4 мкм.
Имеется несколько приемов, которые обеспечивают липосомы необходимого размера. Один такой способ обеспечения заданного размера описан в Патенте США N 4737323, который здесь включен в качестве ссылки. Разрушение суспензии липосом ультразвуком либо в ванне, либо при помощи ультразвукового зонда дает постепенное снижение размеров до небольших однослойных пузырьков размером менее примерно 0,05 мкм. Гомогенизация является еще одним приемом, который основан на энергии среза, чтобы расщепить большие липосомы на липосомы меньшего размера. При осуществлении общей процедуры гомогенизации многослойные пузырьки рециркулировали через стандартный эмульсионный гомогенизатор до тех пор, пока не будут получены выбранные размеры липосом, в общем случае, от примерно 0,1 до 0,5 мкм. В обеих процедурах распределение размеров частиц может быть проанализировано при помощи известного разделения частиц по размеру при помощи лазерного луча.
Экструзия липосомы через поликарбонатную мембрану с небольшими порами или асимметрические керамические мембраны также является эффективным приемом для уменьшения размеров липосом до относительно хорошего распределения по размеру. В общем случае суспензия проходит через мембрану один или несколько раз до тех пор, пока не будет получено целевое распределение липосом по размеру. Липосомы могут быть подвергнуты экструзии через мембраны с последовательно уменьшающимися размерами пор с тем, чтобы добиться постепенно снижения размера липосом.
Даже при самых эффективных процедурах заключения в капсулу суспензия липосом по размеру в начальный момент может содержать до примерно 50% и более лекарственного препарата и нацеливающего агента в свободной /не включенной в капсулу/ форме. Следовательно, чтобы оптимизировать преимущества липосомального "нацеливаемого" лекарственного препарата, важно удалить свободный препарат и нацеливающий агент из финальной инъектируемой суспензии.
Имеется несколько приемов, которые можно использовать для удаления не захваченного соединения из суспензии липосом. В соответствии с одним их этих приемов, липосомы в суспензии превращают в таблетки при помощи высокоскоростного центрифугирования, которое переводит свободное соединение и очень маленькие липосомы в верхний слой. Другой такой прием включает концентрирование суспензии при помощи ультра-фильтрации с последующим суспендированием концентрированных липосом в среду без лекарственного препарата. В качестве альтернативы, гель-фильтрацию можно использовать для того чтобы отделить частицы больших липосом от молекул растворимого вещества.
После обработки с тем, чтобы удалить свободный лекарственный препарат и/или "нацеливающий" агент, суспензию липосом доводят до целевой концентрации для использования с целью внутреннего применения. Это может включать повторное суспендирование липосом в соответствующем объеме инъектируемой среды, где липосомы концентрировали, например, при помощи центрифугирования или ультрафильтрации, или концентрирования суспензии, где стадия удаления препарата увеличивала общий объем суспензии. Затем суспензию подвергали стерилизации при помощи фильтрации, как это описано выше. Препарацию липосома-лиганд можно применять парентерально или местным способом в дозе, которая варьируется в зависимости, например, от способа применения, доставляемого лекарственного препарата и типа заболевания, подлежащего лечению, и т.д.
Для фармацевтических композиций, которые содержат SLX-лиганд и/или SLX-заменитель, которые связываются с рецепторами селектина, доза этого соединения будет варьироваться в зависимости, например, от конкретного соединения, способа применения, конкретного заболевания, подлежащего лечению, и его серьезности, общего состояния и самочувствия пациента и предписаний лечащего врача. Например, для лечения повреждений, вызванных повторной перфузией, доза изменяется в области от примерно 50 мкг до 2000 мг/день пациента массой 70 кг. В идеальном случае терапевтическое применение должно начинаться настолько быстро, насколько это возможно после инфаркта миокарда или других повреждений. Эти фармацевтические композиции предназначены для парентерального, местного, стоматического или локального применения, такого как аэрозоли или через кожу, для профилактического и/или терапевтического лечения. Фармацевтические композиции могут быть применены в самых разнообразных формах единичных доз, в зависимости от способа применения. Например, формы единичных доз, предназначенные для стоматического применения, включают порошки, таблетки, пилюли, капсулы и драже.
В предпочтительном варианте фармацевтические композиции применяют внутривенным способом. Таким образом, в соответствии с настоящим изобретением предлагаются композиции для внутривенного применения, которые содержат раствор соединения, растворенного или суспендированного в приемлемом с фармацевтической точки зрения носителе, в предпочтительном варианте в водном носителе. Самые разнообразные носители можно использовать, например, воду, буферированную воду, 0,4% соляной раствор и т.п. Эти композиции можно стерилизовать при помощи хорошо известных приемов стерилизации или их можно стерилизовать на фильтре. Полученные в результате водные растворы можно упаковать для использования или их можно подвергнуть лиофилизации, причем лиофилизованные препарации соединяют со стерильным водным раствором перед применением. Эти композиции могут содержать приемлемые с фармацевтической точки зрения вспомогательные материалы, которые требуются для аппроксимации физиологических условий, такие как регуляторы pH и буферирующие агенты, агенты, регулирующие тонус, смачивающие агенты и т.п., например, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, монолаурат сорбитана, олеат триэтаноламина и т.д.
Концентрация SLX-лиганда или заменителя, который может быть скомбинирован с другими SLX-лигандами или заменителями, чтобы образовать "коктейль" для увеличения эффективности фармацевтической формы, может широко варьироваться, а именно, от менее примерно 0,05%, в общем случае не менее примерно 1% до 10-30 массовых %, и она может быть выбрана главным образом на основании объемом жидкости, вязкостей и т.д., в соответствии с конкретно выбранным способом применения. Такой коктейль может содержать моноклональное антитело, которое связывается с рецептором селектина, например, моноклональное антитело к ELAM-I или GMP-140, соединенное с SLX-лигандом, заменителем лиганда или моноклональным антителом к этому лиганду с тем, чтобы эффективно ингибировать взаимодействие лиганд-рецептор. Как было описано выше, компоненты коктейля могут быть доставлены при помощи липосомальных препараций.
Таким образом, общая фармацевтическая композиция для внутривенной инфузии может содержать 250 мл стерильного раствора Рингера и 25 мг соединения. Реальные примеры для получения парентерально применяемых соединений известны или очевидны каждому специалисту в этой области техники и описаны более подробно, например, в Remington's Pharmaceutical Sciences 17-е издание, изд. Mack Publishing Company, Easton PA /1985/, которая здесь включена в качестве ссылки.
Для твердых композиций можно использовать известные нетоксичные твердые носители, которые включают, например, фармацевтические сорта маннита, лектозы, крахмала, стеарата магния, сахарина натрия, талька, целлюлозы, глюкозы, сахарозы, карбоната магния и т.д. Для стоматического применения приемлемая с фармацевтической точки зрения нетоксичная композиция образуется при помощи введения любого из известных наполнителей, таких как те носители, которые были перечислены выше, и в общем случае 10-95% активного ингредиента, то есть одного или нескольких SLX-лигандов или заменителей, являющихся предметом настоящего изобретения, в предпочтительном варианте примерно 20%, (см. Remington's, выше).
Для аэрозольного применения соединения в предпочтительном варианте используются в тонко измельченном виде вместе с поверхностно-активным агентом и распыляющих агентом. Общее процентное содержание SLX-олигосахаридного лиганда или заменителя составляет 0,05-30 массовых %, в предпочтительном варианте 1-10%. Поверхностно-активный агент должен быть, разумеется, нетоксичным и в предпочтительном варианте растворимым в распыляющем агенте. Представителем таких агентов являются сложные эфиры или частичные сложные эфиры жирных кислот, содержащих от 6 до 22 атомов углерода, таких как капроновая, октановая, лауриновая, пальмитиновая, стеариновая, линолевая, линоленовая, олестеровая и олеиновая кислоты с алифатическим многоатомным спиртом или его циклическим ангидридом, таким как, например, этилен гликоль, глицерин, эритрит, арабит, маннит, сорбит, ангидриды гексита, полученные из сорбита, и производные полиоксиэтилена и полиоксипропилена этих сложных эфиров. Можно использовать смешанные сложные эфиры, такие как смешанные или натуральные глицериды. Поверхностно-активный агент может составлять 0,1-20 массовых % от композиции, в предпочтительном варианте 0,25-5%. Баланс композиции в общем случае составляет распыляющий агент. Ожиженные распыляющие агенты в общем случае являются газами при окружающих условиях и конденсируются под давлением. Среди соответствующих ожиженных распыляющих агентов находятся низшие алканы, содержащие до 5 атомов углерода, такие как бутан и пропан; а в предпочтительном варианте - фторированные или фторхлорированные алканы. Можно также использовать смеси вышеуказанных материалов. При получении аэрозоля контейнер, снабженный соответствующим клапаном, заполняют соответствующим распыляющим агентом, содержащим тонко измельченные соединения и поверхностно-активный агент. Ингредиенты таким образом поддерживаются при повышенном давлении до тех пор, пока не освобождаются под действием клапана.
Композиции, содержащие эти соединения, могут быть применены при помощи профилактического и/или терапевтического введения. При терапевтических приложениях композиции применяют к пациенту, уже страдающему заболеванием, как это описано выше, в количестве, достаточном для того, чтобы излечить или по крайней мере частично остановить симптомы заболевания и его осложнения. Количество, адекватное для достижения этой цели, определяется как "терапевтически эффективная доза". Количества, эффективные для такого использования, будут зависеть от серьезности заболевания, массы и общего состояния больного, но в общем случае изменяется в области от примерно 0,5 мг до примерно 2 000 мг SLX-олигосахарида или SLX-заменителя в день для пациента массой 70 кг с дозировкой от примерно 5 мг до примерно 200 мг соединения в день в предпочтительном варианте.
При профилактических применениях композиции, содержащие соединения, являющиеся предметом настоящего изобретения, применяют к пациенту, восприимчивому или, другими словами, подвергающемуся риску конкретного заболевания. Такое количество определяется, как "профилактически эффективная доза". При таком применении точные количества снова зависят от состояния пациента и его массы, но в общем случае изменяется в области от примерно 0,5 мг до примерно 1 000 мг на пациента массой 70 килограмм, в более предпочтительном варианте от примерно 5 мг до примерно 200 мг для пациента массой 70 кг.
Однократное или многократное применение композиций может быть осуществлено с уровнями доз и периодичностью, которые выбирает лечащий врач. В любом случае, фармацевтическая форма должна обеспечивать количество SLX-олигосахарида или SLX-заменителя, являющихся предметом настоящего изобретения, достаточное для эффективного лечения пациента.
Эти соединения могут также найти использование в качестве диагностических реагентов. Например, меченые соединения можно использовать для локализации областей воспаления или метастаз опухоли у пациента, который подозревается на воспалительное заболевание. Для такого использования соединения можно "пометить" при помощи 125I, 14C или трития.
Приводимые ниже примеры служат целям иллюстрации и не следует рассматривать в качестве ограничения настоящего изобретения.
Пример I. Изоляция α 1,3,-фукозилтрансферазы 1 из аппарата Голджи.
LEC11, HL-60, HT-29, некоторые аденокарциномы /в частности, клетки colo 205/ и полиморфоядерные лейкоциты /PMN, нейтрофилы/ содержат весьма специфическую α 1,3-фукозилтрансферазу 1, которая способна переносить фукозу из GDP-фукозы в сиалилированные субстраты Neu Ac α 2,3Gal β 1,4GlcNAc или NeuGc α 2,3Gal β 1,4GlcNAc.
В этой области техники хорошо известно, что фукоза переносится в олигосахаридные цепи в полости аппарата Голджи через специфическую фукозилтрансферазы, обзор которых содержится в книге Schacter и Roseman "The Biochemistry of Hycoproteins and Proteoglycans" ред. W. Lennary изд. Plenum Press, New York, стр. 85-160 /1980/, которая здесь включена в качестве ссылки. Так как субклеточная локализация фукозилтрансферазы находится в аппарате Голджи, первый шаг в изоляции этих ферментов состоит в том, чтобы изолировать фракцию аппарата Голджи из линии клеток, которая экспрессирует эту новую и специфическую α 1,3-фукозилтрансферазу.
Пузырьковые фракции, полученные в аппарате Голджи, обрабатывали модифицированной процедурой, описанной в Balch и др., Cell, т. 39, стр. 405 /1984/, которая здесь включена в качестве ссылки. Линии клеток LEC11, HL-60, HT-29, PMN, colo 205 и других, содержащих α 1,3-фукозилтрансферазу 1, выращивали в суспензии до плотности приблизительно 5 • 105 клеток/мл. Клетки собирали из суспензионной культуры центрифугированием со скоростью 2000 х g. Полученную в результате таблетку клеток из 12 литров суспензии /6 • 109 клеток/ снова суспендировали в 3 объемах /упакованный объем клеток/ охлажденного льдом 0,25М раствора сахарозы /м/о/, содержащего Трис-Cl /10 мМ/, pH 7,0, дезактивированной нагреванием сыворотки зародыша коровы /7%/ и Апротинина /100 μг/мл, фирма Sigma Chemical, Co., St. Louis, Mo./.
Клетки разрушали /при помощи приблизительно 60 ударов/ с использованием стеклянного гомогенизатора Уитона с плотной пригонкой и пестика A. Гомогенат центрифугировали 5 мин со скоростью 500 х g с использованием настольной клинической центрифуги. Липид и нерастворимый материал, оставшиеся в верхней части раствора в пробирке для центрифуги, сбрасывали. Мутный верхний слой переносили в чистую пробирку, а затем концентрацию сахарозы во фракции верхнего слоя обеспечивали на уровне 40% /м/о/ сахарозы в Трис-Cl /20 мМ/, pH 7,0, при помощи рефрактометра. Пять миллилитров этой суспензии переносили в пробирку для ультрацентрифугирования и наносили слоем последовательно 2,5 мл 35% /м/о/ сахарозы в Трис-Cl /10 мМ, pH 7,0/ и 2,0 мл 29% /м/о/ сахарозы в 10 мМ буфере Трис-Cl. Градиент центрифугирования 1 час со скоростью 110 000 х g, используя двигатель SW-41 /Бекмэн/, при температуре 5oC. Пузырьки, обогащенные при помощи аппарата Голджи, собирали при помощи граничной фазы 29-35% сахарозы. В других граничных фазах градиента обнаруживали другие субклеточные фракции, например, пузырьки, полученные из полосы грубой и гладкой эндоплазматической ретикулярной ткани, ниже пузырьков, полученных при помощи аппарата Голджи и т.д. Полосу, извлеченную из граничной фазы 29-35%, анализировали на присутствие и количество активности сиалилтрансферазы.
О ферменте сиалилтрансферазы известно только, что он находится внутри пузырьков, полученных из аппарата Голджи, и что он используется специалистами в этой области техники в качестве маркера, чтобы обеспечить аутентичность полосы, собранной из граничной фазы 29-35%.
Анализы на сиалилтрансферазу осуществляли с использованием азиалофетуина в качестве акцептора, как это описано в Briles и др., J. Biol. Chem., т. 252, стр. 1107 /1977/. Хорошая препарация пузырьков, получаемых на аппарате Голджи из LEC-клеток, в общем случае имеет специфическую активность сиалилтрансферазы в 3,0 нмоль/мг протеина/час.
Полученную в результате препарацию на аппарате Голджи затем использовали в качестве источника α 1,3-фукозилтрансферазы 1, используемой в ферментативном синтезе, описанном выше.
Пример II. Доказательство межклеточной адгезии при помощи клеток, экспрессирующих SLX.
Способность клеток LEC11 /которые экспрессируют SLX/ связываться с активированными эндотелиальными клетками, экспрессирующими ELAM-I, сравнивали с той же способностью CHO-клеток и еще одним мутантом гликозилирования, LEC12, который экспрессирует структуру Lex, несиалилированную форму SLX.
Материалы.
Пассаж 5 эндотелиальных клеток человеческой умбиликальной вены /HUYEC/ /фирма Clonetics/, который выращивали на покрытом желатином планшете с 48 углублениями, использовали в качестве источника эндотелиальных клеток. Клетки стимулировали при помощи IL-1 β /фирма Genzyme/ в концентрации 30 μг/мл. Клетки стимулировали в точности в течение 4 часов. В качестве источника контрольных, несущих лиганд, клеток использовали клетки HL-60, полученные из Американского Собрания Типов Культур /АТСС N CCL 240/. Их собирали из объема культуры в RPM1 1640 /фирма Gibco/, содержащей Пенициллин /100 единиц/мл/ /Стрептомицин /100 мкг/мл/ /фирма Irvine Scientific/, L-глютамин /2 мМ/ /фирма Irvine Scientific/ и 10% Сыворотки зародыша коровы /фирма Hazleton/ /в дальнейшем именуемой CPPMI/. LEC11, LEC12 и CHO-KI представлены д-ром Стенли /P. Stanley/. Их выращивали в суспензионной культуре в полной среде альфа MEM, содержащей рибонуклеотиды и деоксирибонуклеотиды /фирма Gibco/, Пенициллин /100 единиц/мл/ /Стрептомицин /100 μг/мл/ /фирма Irvine Scientific/, L-Глютамин /2 мМ/ /фирма Irvine Scientific/, L-Глютамин /2 мМ/ /фирма Jrvine Scientific/ и 10% Сыворотку Зародыша коровы /фирма Hazelton/.
Процедура.
1. HL-60, LEC11, LEC12 и CHO-K1 клетки собирали и промывали в CRPMI. Подсчет живых клеток делали с использованием трипановой сини. Из 3 • 106 клеток каждого типа изготавливали таблетки в пробирке емкостью 10 мл и в каждую таблетку добавляли 300 μл 51Cr /450 μCi/ /фирма New England Nuclear/. Пробирки инкубировали 1 час при 37oC при спокойном перемешивании.
2. Меченые клетки промывали 3Х в среде и снова суспендировали в 2 • 105 /400 μл /6 мл/. Затем пробирки помещали в ледяную ванну при температуре 4oC.
3. После 4 часов инкубирования с IL-1β аналитическую пластинку, содержащую активированный HOYEK, удаляли из инкубатора, и этот процесс быстро прекращали в течение 15 минут, выдерживали пластинку в ледяной ванне при 4oC.
4. Когда температура в обоих образцах выравнивалась, среду удаляли из углублений пипеткой Пастера, из нескольких углублений одновременно.
5. Меченые клетки добавляли в углубления в объеме 400 μл, эквивалентные 2 • 105 клеткам/углубление. Три порции по 400 μл каждой суспензии клеток помещали в стеклянные трубки для определения ЧОМ /числа отсчетов/минуту/.
6. Пластинку инкубировали в ванне лед/вода в течение 30 минут.
7. Несвязанные клетки удаляли из углублений аналитической пластины при помощи систематического ресуспендирования, используя пипетку Пастера, затем при помощи добавления и удаления 0,7 мл среды.
8. Всю среду удаляли из углублений и добавляли раствор 0,125 М Трис, 2% ДСН /додецил сульфата натрия/ и 10% глицерина /0,3 мл/. Пластину выдерживали 30 минут, затем в каждое углубление добавляли 0,5 мл d H2O.
9. Жидкость в каждом углублении снова суспендировали при помощи пипетки P1000 и переносили в стеклянную пробирку. Наконечник P1000 вставляли в пробирку.
10. Пробирки, включая те, что содержат входные ЧОМ-образцы /ЧОМ - число отсчетов/минуту/, анализировали на гамма-счетчике.
11. ЧОМ, связанный с каждым углублением, делили на входные ЧОМ для каждого образца, чтобы определить связывание в %. Точки тройного анализа для среднего и стандартного отклонения наносили на координаты.
Результаты, полученные в этом эксперименте, приведенные на фиг. 1, указывают на то, что клетки, экспрессирующие SLX, обладают способностью связываться эффективно с активированными эндотелиальными клетками сосудов, экспрессирующими ELAM-1. Эти данные показывают, что LEC11-клетки, которые экспрессируют высокие концентрации единственного углевода SLX, связываются исключительно хорошо с активированными при помощи 1L-1 β HUYEC, в то время, как LEC12 и CHO-K1, которые не дают существенных количеств этого углевода, являются слабыми связывающими агентами активированного HUYEC. Этот вывод, кроме того, подтверждается наблюдением, что такое связывание имеет место при температуре 4oC, специфической для связывания с участием ELAM-1.
Пример III. Ингибирование межклеточной адгезии при помощи моноклональных антител, специфических для SLX.
Описаны две серии экспериментов, которые подтверждают, что лиганд нейтрофилов для ELAM-1 содержит олигосахарид, в котором концевыми сахарами являются Neu Ac α 2,3 Gal β 1,4 /Fuc α 1,3/ Glc NAc /SLX/.
Эти эксперименты осуществляли при помощи анализа способности моноклональных антител, специфических относительно сиалилированного Lex и несиалилированного Lex, блокировать адгезию с участием ELAM-1 клеток HL-60 со стимулированными при помощи 1L-1 β HUYEC.
A. Полоса 1 моноклонального антитела.
Материалы: пассаж 3 HUYEC, из культур, инициированных для настоящих экспериментов, использовали, как это описано выше. Две серии тройных углублений оставляли нестимулированными в качестве контрольных. Четыре тройных углубления стимулировали при помощи 1L-1 β /фирма Genzyme/ в концентрации 10 μг/мл и 4 в концентрации 20 μг/мл. Клетки стимулировали в точности 4 часа. Клетки HL-60, полученные из Американского Собрания Типов Культур /АТСС/, использовали в качестве источника несущих лиганд клеток. Их собирали из объема культуры в RPMI-1640 /фирма Gibco/, содержащей пенициллин /100 единиц/мл/, стрептомицин /100 μг/мл/ /фирма Irvine Scientific, L-Глютамин /2 мМ/ /фирма Irvine Scientific/ и 10% сыворотки зародыша коровы /фирма Hazleton/ / именуемая в дальнейшем, как cRPM1/.
Препарации моноклональных антител включали SNH3 /IgM/ в концентрации примерно 20 μг/мл и SH1 /IgG3/ в концентрации примерно 10 μг/мл. Специфичность SNH3 направлена на SLX в то время, как SH1 "узнает" несиалилированную структуру.
Процедура.
1. Клетки HL-60 собирали и промывали в CRPMI. Подсчет живых клеток осуществляли с использованием триптановой сини. 3 • 106 клеток помещали в каждую из двух 10 мл пробирок и в каждую пробирку добавляли 300 μл 51Cr /450 μCi/ /фирма New England Nuclear/. Пробирки инкубировали 1 час при температуре 37oC при спокойном перемешивании.
2. Антитела использовали в виде верхних слоев культуры гибридомы и они содержали 0,01% NaN3 и 0,05% тимерозала. Чтобы удалить эти консерванты, 5 мл каждого антитела подвергали анализу против 3 замен в 500 мл каждая старой среды культуры ткани в течение 72 часов.
3. Антитела собирали после диализа и 3,5 мл каждого помещали в 10 мл пробирки. Остаток сохраняли для использования в ELISA-анализе на HL-60-связывание. 7 мл RPMI 1640 5% сыворотки зародыша коровы помещали в четвертую пробирку для использования в качестве контрольного анализа.
4. Меченые HL-60 клетки промывали 3Х в CRPMI и собирали в одну пробирку. Затем их подвергали центрифугированию и снова суспендировали в 1 мл среды.
5. 200 μл клеточной суспензии добавляли в каждое из антител, содержащихся в пробирке, и 400 μл в контрольную пробирку. Пробирки инкубировали 20 минут при 37oC при спокойном перемешивании.
6. Пластинку с анализом стимулированного HUYEK удаляли из инкубатора и среду удаляли из углубления пипеткой Пастера, из нескольких углублений одновременно.
7. 0,5 мл суспензии клеток добавляли в каждую тройку углублений. Контрольные клетки помещали на пластинки на стимулированные и нестимулированные HUYEC при обеих концентрациях 1L-1β. Испытуемые клетки добавляли только в стимулированные углубления.
8. Порции в 0,5 мл каждой клеточной суспензии добавляли в стеклянные пробирки, чтобы затем использовать для определения входных ЧОМ.
9. Аналитические пластинки возвращали в инкубатор /5% CO2, 37oC/ на 30 минут.
10. Порцию каждой клеточной суспензии смешивали с равным объемом трипановой сини и клетки анализировали под микроскопом на живучесть. Получали такие результаты: Контрольный = 98%, SH1 = 92% и SNH3 = 99%.
11. Несвязанные клетки удаляли из углублений аналитической пластины при помощи систематического ресуспендирования, используя пипетку Пастера с последующим добавлением и удалением 0,7 мл среды.
12. Всю среду удаляли из углублений и добавляли раствор 0,125 М Трис, 2 % ДСН /фирма Bio-Rad/ и 10% глицерина /фирма Fisher/ /0,3 мл/. Пластинки выдерживали в течение 30 минут, а затем в каждое углубление добавляли 0,6 мл d H2O.
13. Жидкость в каждом углублении снова суспендировали с использованием пипетки P1000 и переносили в стеклянную пробирку. Наконечник P1000 вставляли в пробирку.
14. Пробирки, включая те, что содержат входные отсчеты в минуту /ОМ/, анализировали на гамма-счетчике.
15. ЧОМ, связанное с каждым углублением, делили на входное ЧОМ для каждого образца, чтобы определить % связывания. Точки среднего и стандартного отклонения для тройного анализа наносили на координаты.
Репликаты рассматривали, как наилучшие в эксперименте, если в этом эксперименте использовали высокие 1L-1 β , чтобы индуцировать эндотелиальные клетки.
Полученные результаты показывали, что моноклональное антитело SNH3 блокировало связывание HL-60 клеток со стимулированными при помощи 1L-1 β HUYEC через рецептор ELAM-1. Контрольное антитело SH1, которое не связывает SLX-детерминант, не блокировало связывание клеток HL-60 с ELAM-1. Это наводит на мысль, что концевая сиаловая кислота в лиганде необходима для связывания с ELAM-1.
B. Полоса 2 моноклональных антител
Материалы: пассаж 3 HUYEC, которые выращивали на покрытых желатином аналитических планшетах с 48 углублениями /фирма Costar/, использовали в качестве источника эндотелиальных клеток. Планшеты подготавливали так, как это было описано ранее. Две серии по три углубления оставляли нестимулированными в качестве контрольных. Семь троек на каждой планшетке стимулировали при помощи IL-1 β в концентрации 30 μг/мл в 0,5 мл EGM-UY. Клетки стимулировали ровно 4 часа. HL-60 клетки /ATCC/ использовали в качестве клеток, несущих лиганд. Их собирали из объемной культуры в CRPMI. Верхние слои свежей гибридомы, содержащие моноклональные антитела, включали: FH6 /IgM/, mAb с более низким сродством; SNH-3 /IgM/ /20 μг/мл/; SH-1 /IgG3/ /10 μг/мл/; FH-2 /IgM/, mAb, активное относительно Lex; SNH-4 /IgG3/, антитело с высоким сродством; и CSLEX-1 /IgM/ /предоставленное д-ром П.Терасаки, UCLA в виде очищенного иммуноглобулина в концентрации 2,8 мг/мл, разбавленного до 9 μ г/мл в Модифицированной Среде Иглса Далбекко /DMEM/, содержащей 5% СЗК для использования в этом анализе/. Специфичности этих антител были следующие: FH6, SNH-4, SNH3 и CSLEX-1 были специфичны для SLX; FH2 и SH1 были специфичны для несиалилированного Lex.
Процедура.
1. HL-60 клетки собирали и промывали в CRPMI. Подсчет живых клеток осуществляли с использованием триптановой сини. 3•106 клеток помещали в каждую из двух 10 мл пробирок и в каждую пробирку добавляли 300 μл 51Cr /450 μCi/ /фирма New England Nuclear/. Пробирки инкубировали 1 час при 37oC при спокойном перемешивании.
2. Меченые клетки HL-60 промывали 3Х в DMEM, содержащем 5% СЭК /именуемом в дальнейшем, как cDMEM/, и собирали в одну пробирку. Затем их центрифугировали и снова суспендировали в 4•106 клеток на мл в ту же среду.
3. 3,2 мл верхнего слоя каждой культуры моноклонального антитела и 3,2 мл очищенного CSLEX-1 /29 μг/ добавляли в отдельные пробирки; в контрольную пробирку добавляли 6,4 мл среды.
4. 200 μл клеточной суспензии /эквивалентной примерно 8•105 клеток/ добавляли в пробирки, содержащие эти моноклональные антитела, и 400 μл в контрольную пробирку. Затем пробирки инкубировали 20 минут при 37oC при спокойном перемешивании.
5. Пластинку с анализами на стимулированные HUYEC удаляли из инкубатора и углубления промывали один раз при помощи сDMEM и среду удаляли из углубления при помощи пипетки Пастера, из нескольких углублений одновременно.
6. 0,4 мл клеточной суспензии добавляли в каждое углубление одной из двух пластин. Контрольные клетки наносили на нестимулированные и стимулированные HUYEC. Клетки, обработанные антителом, добавляли только в стимулированные углубления.
7. Порции в 0,4 мл каждой клеточной суспензии добавляли в стеклянные трубки, чтобы использовать для определения входных ЧОМ.
8. Аналитическую пластину инкубировали при температуре 37oC в течение 30 минут.
9. Остаток каждой клеточной суспензии и аналитические пластинки помещали в ледяную ванну для быстрого охлаждения на 20 минут.
10. Клеточные суспензии наносили слоем на охлажденную пластину, как это делали выше для пластинок при 37oC. Эту пластину инкубировали в течение 30 минут при 40oC.
Оставшиеся стадии анализа осуществляли, как это описано на стадиях 11-15 раздела A выше, за тем исключением, что на стадии 12 пластины выдерживали в течение 15 минут, а не 30 минут.
Результаты, приведенные на фиг. 2A, показывают, что моноклональные антитела SNH-3, FH6, SNH-4 и CSLEX-1, причем все они являются специфическими для SLX, существенно блокировали связывание клеток HL-60 со стимулированными при помощи IL-1β HUYEC через рецептор ELAM-1, когда их инкубировали при 37oC. Моноклональные антитела, специфические для Lex /FH2 и SH1/ не были эффективными ингибиторами. Таким образом, лиганд для ELAM-1 содержит сиалилированный антиген Lex или аналогичную структуру обнаруживли в гликопротеинах или гликолипидах клеточной поверхности.
После инкубирования при 4oC /фиг. 2B/ антитела FH6 и SNH-3 /оба IgM/ усиливали связывание. В этих испытаниях проявлялась существенная агглютинация HL-60 клеток в углублениях, которая может оказываться существенной для этого наблюдения.
C. Моноклональные антитела блокируют адгезию клеток LEC11 с клетками, которые экспрессируют ELAM-1.
В этой серии экспериментов анализировали способность моноклональных антител, специфичных для SLX и для несиалилированной формы, Lex, блокировать адгезию с участием ELAM-1 клеток LEC11 /которые эксперссируют SLX/ и клеток LEC12 /которые экспрессируют Lex/ со стимулированными IL-1 β HUYEC.
Материалы: Пассаж 4 HUYEC служил в качестве источника эндотелиальных клеток. Пластины подготавливали в точности так же, как это было описано ранее. Две серии тройных углублений оставляли нестимулированными в качестве контрольных. 7 троек на каждой пластине стимулировали при помощи IL-1 β в концентрации 30 μг/мл в 0,5 мл объеме EGM-UV. Клетки стимулировали в точности 4 часа. LEC11 и LEC12 клетки, описанные в общем случае в Stanley и др., J. Biol. Chem. т. 263, стр. 11374 /1988/, см. выше, были предоставлены д-ром П. Стенли. Их выращивали в суспензионной культуре в полной альфа-МEМ, содержащей рибонуклеотиды и деоксирибонуклеотиды /фирма Gibco/, пенициллин /100 единиц/мл/ /стрептомицин /100 мкг/мл/ /фирма Irvine Scientific/, L-глютамин /2 мМ/ /фирма Irvine Scientific/ и 10% СЗК /фирма Hazelton/. Моноклональные антитела, используемые в этих экспериментах, описаны в Разделе B, выше. Они включают: FH6, SNH-3, SH-1, FH-2, SNH-4 и CSLEX-1.
Процедура.
1. LEC11 и LEC12 клетки собирали и промывали в CRPMI. Живые клетки подсчитывали с использованием триптановой сини. 3•106 клеток каждой линии клеток помещали в каждую из двух 10 мл пробирок и в каждую пробирку добавляли 300 μл 51Cr /450 μCi/ /фирма New England Nuclear/. Пробирки инкубировали 1 час при температуре 37oC при спокойном перемешивании.
2. Радиомеченные клетки промывали X3 в cDMEM и собирали в одну трубку. Их затем центрифугировали и снова суспендировали в 4•106 клеток на мл в той же среде.
3. 1,6 мл верхнего слоя каждого моноклонального антитела и 1,6 мл очищенного CSLEX-1 /15 мкг/ добавляли в отдельные пробирки; в контрольные пробирки помещали 3,2 мл среды.
4. 200 мкл клеточной суспензии, эквивалентной 4 х 105 клеток LEC11 или LEC12 добавляли а пробирки, содержащие моноклональные антитела, и 400 μ в контрольную пробирку. Пробирки инкубировали 20 минут при температуре 37oC при спокойном перемешивании.
5. Пластинку с анализом для стимулированных HUYEC удаляли из инкубатора и углубления промывали один раз cDMEM и среду удаляли из углублений при помощи пипетки Пастера, из нескольких углублений одновременно.
6. Клеточные суспензии и аналитическую пластину помещали в ледяную ванну, чтобы быстро охладить на 20 минут.
7. 0,4 мл клеточной суспензии добавляли в каждое углубление описанной ранее аналитической пластины. Контрольные клетки наносили на пластины на нестимулированные и стимулированные HUYEC. Клетки, обработанные антителом, добавляли только в стимулированные углубления. Каждый анализ осуществляли в трех повторах.
8. Порции в 0,4 мл каждой клеточной суспензии добавляли в стеклянные пробирки, чтобы использовать для определения входных ЧОМ.
9. Аналитическую пластину инкубировали 30 минут при температуре 4oC.
Оставшиеся стадии анализа осуществляли, как это описано в стадиях 11-15 Раздела А выше, за тем исключением, что на стадии 12 пластины выдерживали в течение 15 минут.
Результаты, показанные на фиг. 3А и 3В, указывают на то, что моноклональные антитела SNH-3, FH6, SNH-4 и SCLEX-1 /все специфичны для SLX/ существенно блокировали связывание LEC11 клеток со стимулированными IL-I β HUYEC через рецептор ELAM-1. Клетки LEC12, которые не экспрессируют SLX-эпитоп, не связывали активированный эндотелий. Моноклональные антитела, специфические для Lex/НГ2 и SH1/, вызвали слабое ингибирование связывания LEC11.
Дальнейшее подтверждение того, что SLX является основным лигандом для рецептора ELAM-1, получали при помощи удаления сиаловой кислоты из клеток LEC11 и HL-60. В этих экспериментах обработку клеток LEC 11 и HL-60 перед адгезией анализируют при помощи нейраминидазы /сиалидазы/ Clostridium perfringens, 1,6 ед/мл /Типа X, фирма Sigma Chem., Co/ в течение 90 минут при 37oC во время 51Cr-мечения. Результаты, приведенные на фиг.4, подтверждают, что сиалидаза существенно снижала адгезию клеток LEC 11 и HL-60 на 70-85% с активированными эндотелиальными клетками.
Пример IV. Липосомы гликосфинголипидов блокируют связывание SLX-клеток с активированными эндотелиальными клетками.
В этом Примере описано получение липосом, которые содержат различные биосинтетически продуцированные гликосфинголипиды, на которых концевой углевод является либо SLX, либо Lex, либо похожие, но не идентичные соединения. Показана способность липосом, которые содержат SLX или SLX-заменители, блокировать связывание SLX-экспрессирующих HL-60 клеток и LEC11 клеток с эндотелиальными клетками, которые были предварительно стимулированы с целью экспрессии ELAM-1 при помощи обработки с использованием IL-1 β .
Материалы.
Гликосфинголипиды, используемые в этом эксперименте, приведены в табл. 1; они были получены из Института Биомембран, Сиэттл, BA, и были либо очищены, либо биосинтетически продуцированы и охарактеризованы при помощи ЯМР- и масс-спектрометрии, как это описано в общем случае в Hakomori, S.I. и др. , J. Biol. Chem., т.259, стр.4672 /1984/ и Fukushi, J. и др., J/ Biol. Chem. , т.259, стр. 10511 /1984/, которые здесь включены в качестве ссылок. S-diLex /SLX/ синтезированы ферментативно при помощи добавления фукозиловых остатков, используя линию клеток Colo 205 в качестве источника фермента, а SH в качестве субстрата. Несиалилированный diLex аналогичным образом синтезировали с использованием nLc6 в качестве субстрата и линии клеток NC1 H-69. Смотри Holmes и др., J. Biol Chem. т.260, стр.7619 /1985/, которая здесь включена в качестве ссылки. SPG и SH подвергали очистке из бычьих красных кровяных клеток ми nLc6 продуцировали при помощи химического удаления концевого сиалозилового остатка из SH.
Липосомы, содержащие гликосфинголипиды, получали следующим образом: 100 μг глинолипида добавляли в 300 μг фосфатидилхолина /фирма Sigma, желток яйца/ и 500 μг холестерина /фирма Sigma/ в хлороформе-метаноле /2:1/ и весь раствор выпаривали до сухого состояния с использованием N2 в 15 мл пробирках с завинчивающейся пробкой.
Пассаж 3 HUYEC, которые выращивали до слияния на покрытой желатином аналитической пластине с 48 углублениями /фирма Costar/, использовали в качестве источника эндотелиальных клеток. Пластинки подготавливали, как это было описано ранее. Две серии тройных углублений оставляли нестимулированными в качестве контрольных. 14 троек стимулирования при помощи IL- 1β в концентрации 30 μг/мл в 0,5 мл объеме EGM-UV. Клетки стимулировали в точности в течение 4 часов. HL-60 клетки и LEC11 клетки культивировали так, как это описано выше.
Процедура.
1. Одну чашку с 48 углублениями типа Костар, содержащую HUYEC, выращиваемые до слияния на желатине, удаляли из инкубатора, а среду в каждом углублении удаляли при помощи пипетки Пастера и заменяли либо 0,5 мл свежей среды EGM-UY, либо той же средой, содержащей 30 μг/мл IL-1β , и затем пластину возвращали в инкубатор на 4 часа.
2. Клетки HL-60 и LEC11 собирали и промывали в CRPMI. Подсчет живых клеток осуществляли с использованием трипановой сини. 6• 106 клеток каждого клеточного типа метили радиометкой следующим образом: 3•106 клеток каждого типа помещали в каждую из двух 10 мл пробирок и в каждую пробирку добавляли 300 μл 51Cr /450 μCi/ /фирма Нью Инглэнд Ньюклеа/. Пробирки инкубировали 1 час при температуре 37oC при спокойном перемешивании.
3. Радиомеченные HL-60 и LEC11 клетки промывали 3X в CRPMI и собирали в одну пробирку. Затем их центрифугировали и снова суспендировали до 2•106 клеток на мл в той же среде.
4. Аналитическую пластину со стимулированными HUYEC удаляли из инкубатора и углубления промывали два раза средой RPMI 1640, содержащей 5 мг/мл альбумина бычьей сыворотки /АБС/.
5. Липосомы получали следующим образом. Выпаренные таблетки растворяли в 100 μл абсолютного этанола и обрабатывали ультразвуком в течение 2 мин. В пробирки в течение 2 минут медленно добавляли 2 мл PBS, одновременно продолжая обработку ультразвуком. Это сырье разбавляли в отношении 1:10 в RPMI 1640-среде непосредственно перед использованиеv и 50 μл сырьевого раствора АБС в концентрации 100 мг/мл добавляли в каждый 1 мл разбавленных липосом, чтобы обеспечить финальную концентрацию 5 мг/мл АБС.
6. Среду удаляли из углублений аналитической пластины пипеткой Пастера, из нескольких углублений одновременно, и 0,3 мл суспензии липосом добавляли в каждое из 6 стимулированных при помощи IL-1β углублений. В контрольные углубления помещали буфер липосом, содержащий этанол, RPMI 1640 и АБС в тех же концентрациях, что в углублениях, содержащих липосомы. Контрольный буфер наносили на пластину на стимулированные и нестимулированные HUYEC. В стимулированные углубления добавляли образцы, содержащие липосомы.
7. Пластины инкубировали в течение 40 минут при 37oC, а затем в углубления добавляли 50 μл 51Cr - меченных HL-60 или LEC11 клеток. Каждую линию клеток анализировали в тех повторах для каждой препарации липосом. Финальная концентрация клеток составляла 106 /350 μл/ углубление. Три порции по 50 μл каждой суспензии клеток добавляли в стеклянные пробирки, которые подлежат использованию при определении входных ЧОМ, и аналитическую пластину инкубировали при температуре 37oC в течение 30 мин.
8. Несвязанные клетки удаляли из углублений аналитических пластин при помощи систематического повторного суспендирования с использованием пипетки Пастера и последующим добавлением и удалением 0,7 мл среды. Всю среду удаляли из углублений и добавляли раствор 0,125 М Трис, 2% ДСН и 10% глицирина /0,3 мл/. Пластины выдерживали в течение 15 минут, а затем в каждое углубление добавляли 0,5 мл dH2O.
9. Жидкость в каждом углублении снова суспендировали и переносили в стеклянную пробирку. Наконечник пипетки вставляли в пробирку. Пробирки, включая те, что содержат образца с входными ЧОМ, анализировали в гамма-счетчике. ЧОМ, связанное с каждым углублением, делили на входные ЧОМ для каждого образца, чтобы определить % связывания. Точки средних и стандартной ошибки анализа тройных повторов наносили на координаты.
Как показано на фиг.5, липосомы, содержащие выбранные гликолипиды, содержащие концевые последовательности, которые содержали SLX/S-diLex, таблица 1/, значительно подавляли адгезию HL-60 клеток с активированными эндотелиальными клетками при температуре 4oC. Липосомы, содержащие гликолипиды с Lex /di-Lex/ или другие близкие углеводные структуры /Таблица 1/, проявляли минимальное ингибирование, которое не зависело от структуры углеводной группы. Аналогичные результаты получали с адгезией LEC 11 -клеток. Когда осуществляли эксперименты при температуре 37oC, адгезия HL-60 клеток снижается при помощи липосом, содержащих гликолипиды с SLX-структцурой /S-diLex, 70%/, а также в меньшей степени липосомами, содержащими Lex /di-Lex, 40%/, что наводит на мысль, что Lex может также взаимодействовать с ELAM-1, но с более низким сродством. Эти эксперименты показывают, что биосинтетически продуцируемый SLX или аналогичные соединения заменителей SLX, когда они включены в композиции липосом, могут служить в качестве терапевтических соединений, например, для снижения инфильтрации лейкоцитов в месте воспалений.
Клетки Джуркат связываются с активированными IL-1 эндотелиальными клетками главным образом через пару адгезии Y-CAM /эндотелиальная клетка/ - YLA-4/ клетка Джуркат/ /Wayner и др., J. Cell. Biol. т.109, стр.1321/ в контрасте с тем, что адгезия HL-60 и LEC 11 клеток с активированными эндотелиальными клетками осуществляется через рецептор ELAM-1. Адгезия клеток Джуркат не ингибировалась при использовании липосом, которые содержали SLX, но полностью ингибировалась моноклональным антителом с α-субъединицей молекулы интегрина YLA-4. Этот результат подтверждает, что ингибирование SLX-липосомами HL-60 и LEC 11 клеток не является стеариновым эффектом, приписываемым связыванию липосом с эндотелиальными клетками, но подтверждает вывод, что SLX-липосом ингибируют адгезию через прямую конкуренцию с точкой связывания лиганда ELAM-1.
Пример V.Антитела для SLX-ингибирования связывания с участием GMP-140 на активированных человеческих тромбоцитах.
В этом примере изучали способность моноклональных антител, специфических для SLX и для несиалилированного Lex, блокировать адгезию с участием GMP-140 клеток HL-60 с активированными человеческими тромбоцитами.
Материалы:
HL-60 клетки описаны выше и их использовали в качестве источника клеток, несущих лиганд. Клетки Джуркат использовали в качестве контрольных, не несущих лиганд. Использовали те же моноклональные антитела SH-1, FH-2, SNH-4 и CSLEX-1, что были описаны выше.
Процедура:
1. Кровь брали у нормального человеческого донора через шприц, содержащий антикоагулянт ACD /декстроза, 2,0 г; цитрат натрия 2,49 г; и лимонная кислота 1,25 г; до 100 мл при помощи dH2O/ в отношении 6 частей крови на 1 часть антикоагулянта.
Тромбоциты выделяли при помощи дифференциального центрифугирования следующим образом: Кровь центрифугировали со скоростью 800 об/мин /приблизительно 90 x g/ в течение 15 минут при комнатной температуре. Верхний слой собирали и центрифугировали со скоростью 1200 об/мин /приблизительно 400 x g/ 6 минут. Верхний слой удаляли и центрифугировали со скоростью 2 000 об/мин /1200 x g/ в течение 10 минут, чтобы таблетировать тромбоциты. Эту таблетку промыли 2 раза буфером Tyrode - HEPES, pH 6,5 / NaCl 8,0 г; KCl 0,2 г; NaH2PO4•H2O 0,057 г; MgCl2•6H2O 0,184 г; NaHCO30,1 г; Декстроза, 1,0 г; и HEPES, 2,383 г; доводили до 1 л ДИ-водой, обеспечивали pH 6,5 при помощи IN раствора NaOH/, затем один раз промывали СЗК. Тромбоциты снова суспендировали в концентрации 108/ мл в СЗК.
2. Приблизительно за 20 минут до того как тромбоциты, наконец, снова суспендировали, планшеты с 48 углублениями покрывали 0,1% желатином и инкубировали 15 мин при температуре 37oC. Избыток желатина удаляли пипеткой непосредственно перед добавлением суспензии тромбоцитов. Тромбоциты активировали добавлением 0,25 единиц тромбина/мл/ фирма Sigma, T-6759/ суспензии тромбоцитов. Тромбоциты выдерживали при комнатной температуре 20 минут.
3. Чтобы получить связывание активированных тромбоцитов, 300 μл суспензии тромбоцитов добавляли в каждое углубление покрытой гелем планшете. Планшет инкубировали при температуре 37oC в течение 15 минут, затем обрабатывали со скоростью 800 об/мин/90 х g/ в течение 2 минут. Несвязанные тромбоциты удаляли промывкой планшета 3 раза с использованием PBS.
4. Так как тромбоциты обладают высоко активными Fc-рецепторами, чтобы предотвратить поглощение агрегированного IgG из препарации антител, Fc-рецепторы тромбоцитов блокировали следующим образом. Очищенный мышиный IgG W6/32/IgG2a/ в концентрации 27 мг/мл агрегировали при помощи нагревания до 63oC в течение 5 минут, 300 μл нагретой препарации в концентрации 20 μг/мл в PBS добавляли в каждое углубление планшета, покрытого тромбоцитами. Планшет инкубировали при 37oC в течение 15 минут, затем промывали PBS.
5. HL-60 и клетки Джуркат собирали и промывали в CRPMI. Подсчет живых клеток осуществляли с использованием трипановой сини, 3•106 клеток каждого типа помещали в каждую из двух 10 мл пробирок и в каждую пробирку добавляли 300 μл 51Cr /450 μCi// фирма Нью Инглэнд Ньюклеа/. Пробирки инкубировали 1 час при температуре 37oC при спокойном перемешивании.
6. Радиомеченные клетки промывали 3Х в CRPMI и собирали в одну пробирку. Затем их центрифугировали и снова суспендировали до концентрации 4•106 клеток на мл в той же среде.
7. 1,6 мл верхнего слоя культуры каждого моноклонального антитела и 1,6 мл очищенного CSLEX-1 /15 μг/ добавляли в отдельные пробирки; контрольные пробирки получали 1,6 мл среды.
8. 100 μл суспензии меченых клеток HL-60 или клеток Джуркат /содержащей 4•105 клеток/ добавляли в каждую из пробирок, которая содержала моноклональное антитело. Их инкубировали в течение 20 минут при 37oC при спокойном перемешивании. После этого периода инкубирования 0,3 мл каждой клеточной суспензии /содержащей 7,5 •104 клеток/ добавляли в каждое углубление ранее описанной аналитической пластины, содержащей связанные активированные тромбоциты. Каждый анализ осуществляли в трех повторах.
9. Аналитическую пластину центрифугировали со скоростью 90 х g в течение 2 мин, а затем инкубировали 5 минут при комнатной температуре. Несвязанные клетки удаляли из углублений аналитической пластины при помощи переворачивания пластины во вместилище для радиоактивных сбросов и блоттирования пластины на полотенцах. Углубления промывали 3Х при помощи аккуратного добавления 300 μл PBS в каждое углубление, и перевертывания и блоттирования пластины. Всю среду удаляли из углублений и добавляли 0,3 мл раствора 0,125 М Трис, 2% ДСН и 10% глицерина. Пластины выдерживали 15 минут, а затем в каждое углубление добавляли 0,6 мл dH2O.
10. Жидкость в каждом углублении снова суспендировали пипеткой и переносили в стеклянную пробирку. Наконечник вставляли в пробирку. Пробирки, включая те, что содержат входные образцы, анализировали в гамма-счетчике. ЧОМ, связанные с каждым углублением, делили на входные ЧОМ для каждого образца, чтобы определить % связывания. Входные ЧОМ определяли при помощи анализа порции в 0,3 мл каждой клеточной суспензии, описанной на стадии 8.
Результаты, приведенные на фиг.6, показывают, что моноклональные антитела SNH-4 и CSLEX-1, специфические относительно SLX, блокировали связывание клеток HL-60 c GMP-140 на активированных тромбоцитах. Моноклональные антитела, специфические для Lex /FH2 и SH-1/, также блокировали это связывание, но в меньшей степени. Этот Пример наводит на мысль, что и SLX и Lex могут быть лигандами для GMP-140, но что SLX-структура может иметь более высокое сродство для GMP-140, чем Leх структура.
Пример VI. Липосомы гликосфинголипидов блокируют связывание SLX-клеток с активированными тромбоцитами.
Этот пример демонстрирует способность липосом, которые содержат SLX или SLX-аналоги, блокировать связывание SLX-экспрессирующих HL-60 клеток и PMN с тромбоцитами, которые предварительно стимулировали с целью экспрессии GMP-140 при помощи обработки Тромбином. При выполнении анализов в общем случае следовали описанию, данному в Larsen и др. Cell, т.63, стр.467-474 /1990/, которая здесь включена в качестве ссылки.
Материалы.
Гликосфинголипиды получали, как это описано в Примере IV. Тромбоциты получали, как это описано в Примере V за тем исключением, что блокировку Fc-рецептеров не производили. HL-60 клетки получали, как это описано выше.
PMN получали из 50 мл цельной крови, извлеченной из доноров-добровольцев, в обработанных гепарином пробирках Vacutainer, которые переворачивали, чтобы перемешать кровь. Все стадии осуществляли при температуре 22-24oC. Каждые 25 мл крови наносили слоем на 15 мл Моно-Поли растворяющей Среды /фирма Tlow Labs/. Пробирки центрифугировали со скоростью 800 х g в течение 25 мин, затем со скоростью 1300 х g еще в течение 25 минут. PMN-слой удаляли и помещали в чистую пробирку для центрифуги 50 см3. Тридцать мл сбалансированного раствора солей Хэнкса /фирма Gibco/, содержащего 20 мМ HEPE /фирма Gibco/ и 0,2% глюкозы /Фирма Fisher/, добавляли в каждую пробирку, которую затем центрифугировали со скоростью 1900 х g в течение 3 мин. PMN промывали 3Х в том же буфере при помощи центрифугирования со скоростью 1900 х g в течение 3 минут. ЧОМ подсчитывали с использованием гемацитометра и снова суспендировали до концентрации 2•106/мл, затем хранили при комнатной температуре до использования.
Процедура.
1. 20 ул препарации активированных тромбоцитов помещали в каждую из 28 1,5 мл пробирок Эппендорфа /14 двойных образцов/.
2. 20 ул разбавленных липосом в 10 уг, 5 уг или 2 уг, или контрольных буферов добавляли в соответствующую пробирку каждой двойки.
3. Тромбоциты инкубировали с препарациями липосом в течение 20 мин при комнатной температуре.
4. Нейтрофилы или HL-60 клетки в концентрации 2•106 клеток/мл каждые добавляли в одну серию тромбоцитов, обработанных липосомами. В каждую пробирку добавляли 20 ул клеточной суспензии.
5. Пробирки смешивали и выдерживали при комнатной температуре в течение 20 минут. Затем наносили их в гемацитометр и клетки оценивали как положительные /2 и больше тромбоцитов присоединилось/клетку/ или отрицательные /менее 2 тромбоцитов присоединилось /клетку/.
Как показано на фиг.7, липосомы, содержащие выбранные гликолипиды, имеющие концевые последовательности, которые содержали SLX/S-diLex, Таблица 1/, значительно ингибировали адгезию клеток HL-60 с активированными тромбоцитами. Липосомы, содержащие гликолипиды с Lex/di-Lex/ или другие близкие углеводные структуры /Таблица 1/, проявляли минимальное ингибирование, которое не зависило от структуры углеводной группы. Аналогичные результаты получали с адгезией PMN-клеток /фиг.8/. Эти эксперименты показывают, что биосинтетически продуцированный SLX или похожие SLX-заменители, когда их включают в композиции липосом, могут служить в качестве терапевтических соединений, например, для снижения связывания лейкоцитов с тромбоцитами в местах воспаления.
Пример VII. Олигосахариды, содержащие SLX, блокируют связывание нейтрофилов с тромбоцитами.
В этом примере способность минимального тетрасахарида SLX ингибировать GMP-140-адгезию сравнивали с таким же свойством пента- и гексасахаридов, содержащих SLX. Если коротко, то тромбоциты и нейрофилы выделяли при помощи приемов, описанных ранее. Тромбоциты активировали тромбином, а затем инкубировали при разбавлениях различных олигосахаридов. Нейтрофилы добавляли и определяли эффект сахаридов на адгезию нейтрофилов с активированными тромбоцитами. Использовали следующие олигосахариды: SLX/гекса/, Neu Ac α 2,3 Gal β 1,4 (Fucα1,3) GlcNAc β 1,3 Gal β 1,4 Glc-O-CH2CH2-SiMe3 /неоценимый подарок профессора Хасегава, Университет Гифу, Япония/, SLX /пента/ Neu Ac α 2,3 Gal β 1,4 (Fuc α 1,3) Glc NAc β 1,3 Gal β и SLX /тетра/, Neu Ac α 2,3 Gal β 1,4 (Fuc α 1,3) Glc NAc.
Процедура.
1. Тромбоциты выделяли так, как это описано выше, и активировали /2•108/ мл/ при помощи инкубирования в течение 20 минут при комнатной температуре с тромбином при финальной концентрации 0,25 ед/мл.
2. Нейтрофилы выделяли при помощи нанесения слоя обработанной гепарином крови на Моно-Поли растворяющую среду /фирма Ficoll-Hypaque-Flow Laboratories, затем центрифугировали в течение 25 минут со скоростью 2000 об/мин, а затем еще в течение 25 минут со скоростью 2500 об/мин, как это было описано выше.
3. Для анализа 20 μл суспензии тромбоцитов /2 •108/мл/ помещали в пробирку для центрифуги Эппендорфа. Добавляли равный объем препараций олигосахарида в концентрациях от 200 μг/мл до 0,3 μг/мл, или препараций липосомы-гликолипида /полученных, как это описано выше/ в концентрациях от 2 μг/мл до 0,25 μг/мл, и пробирки выдерживали при комнатной температуре в течение 20 минут. Добавляли 20 μл препарации нейтрофилов /2•106/мл/ и пробирки выдерживали еще в течение 20 минут при комнатной температуре.
4. Адгезию активированных тромбоцитов к нейтрофилам анализировали под микроскопом. Анализировали одну сотню нейтрофилов. Их оценивали как положительные, если 2 и больше тромбоцитов присоединились, и отрицательные, если были связаны менее 2 тромбоцитов. Вычисляли процент клеток с 2 и более связанными тромбоцитами.
Средние значения результатов трех идентичных экспериментов приведены в табл. 2.
Как показано в табл. 2 выше, приблизительно в 20 раз требуется SLX-тетрасахарида больше для 50% ингибирования связывания с участием GMP-140 нейтрофилов с активированными тромбином тромбоцитами, чем SLX-гексахарида. Количество требующегося тетрасахарида приблизительно равно количеству, которое необходимо для аналогичной степени ингибирования, когда используют несиалилированный Lex. Пентасахарид дает 50% ингибирование при концентрациях, аналогичных тем, что необходимы для 50% ингибирования при помощи гексасахарида, что указывает на то, что минимальная структура для максимального ингибирования более близка к пентасахариду.
Пример VIII. Блокирование адгезии с использованием варианта SLX-структур.
Этот пример описывает эксперименты по испытанию различных структур гликолипидов. В частности, испытывали SV2, сиалилированный полисахарид, в котором фукоза вместо того, чтобы быть присоединенной к последнему GlcNAc, как в SLX, присоединена к предпоследнему GlcNAc. Тромбоциты и нейтрофилы выделяли при помощи приемов, описанных выше. Тромбоциты активировали тромбином, а затем инкубировали с разбавлениями различных гликолипидов, вложенных в липосомы, полученные так, как это было описано выше. Добавляли нейтрофилы и определяли эффект гликолипидов на адгезию нейтрофилов с активированными тромбоцитами.
Структуры различных анализируемых гликолипидов приведены ниже:
SDIY2, NeuGc α 2, 3Gal β 1,4 (Fuc α 1,3) GlcNac β 1,3Gal β 1,4(Fuc α 1,3) GlcNAc β 1,3Gal β 1,4Glc β 1,1Cer; SLX, NeuGc α 2,3Gal β 1,4(Fuc α 1,3)GlcNAc β 1,3Gal β 1,4Glc β 1,1Cer; SY2, NeuGc α 2,3Gal β 1,4GlcNAc β 1,3Gal β 1,4(Fuc α 1,3)GlcNAc β 1,3Gal β 1,4Glc β 1,1Cer; SH, NeuGc α 2,3Gal β 1,4GlcNAc β 1,3Gal β 1,4GlcNAc β 1,3Gal β 1,4Glc β 1,1Cer; SPG, NeuGc α 2,3Gal β 1,4GlcNAc β 1,3Gal β 1,4Glc β 1,1Cer;
Результаты двух идентичных экспериментов приведены в табл. 3.
Эти результаты показывают, что SY2 ингибировал адгезию с участием GMT-140 нейтрофилов с активированными тромбином тромбоцитами в той же степени, что это делали SLX и SDiY2.
Пример IX. Блокирование адгезии, используя другие варианты SLX.
Этот пример подтверждает, что сродство сиалилированного Lex/SLX/ относительно GMP-140 одно и то же независимо от того, находится ли концевая сиаловая кислота в форме N-ацетил нейрамината /NeuAc/ или N-гликоль нейрамината /NeuGc/. Все материалы получали в соответствии с описанием, приведенным выше. Тромбоциты и нейтрофилы выделяли в соответствии с описанными приемами. Тромбоциты активировали тромбином, а затем инкубировали с разбавлениями различных гликолипидов, содержащимися в липосомах. Добавляли нейтрофилы и определяли эффект гликолипидов на адгезию нейтрофилов с активированными тромбоцитами.
Результаты эксперимента, в котором синтетический SLX /Neu Ac/ и препарацию SLX, полученную при помощи ферментативного фукозилирования сиалилипараглобозида, очищенного из бычьих эритроцитов SLX /Neu Gc/, непосредственно сравнивали, приведены в табл. 4.
Эти результаты показывают, что SLX-гексасахариды ингибировали адгезию с участием GMP-140 нейтрофилов с активированными тромбином тромбоцитами одинаково, независимо от того является ли сиаловая кислота Neu Ac или NeuGc. Этот результат указывает на то, что либо N-ацетиловое, либо N-гликолоиловое производное сиаловой кислоты узнаются при помощи GMP-140. Аналогичные результаты получали при помощи ELAM-1.
В одном и том же анализе испытывали также различные гликолипиды. Структуры испытуемых гликолипидов представлены ниже:
SLX(hexa), NeuGc α 2,3Gal β 1,4(Fuc α 1,3)GlcNAac β 1,3Gal β 1,4 Glс β 1,1Ceramide; α 2,3 SLX cer, NeuAc α 2,3Gal β 1,4(Fuc α 1,3) GlcNac β 1,3Gal β 1,4Glc β 1,1 Ceramide; α 2,6 SLX cer, NeuAc α 2,6Gal α 1,4 (Fuc β 1,3)GlcNAc α 1,3Gal β 1,4Glc β 1,1Ceramide; SH, NeuGc β 2,3Gal α 1,4 GlcNAc β 1,3Gal β 1,4GlcNAc β 1,3Gal β 1,4Glc β 1,1Ceramide.
Пример X Блокирование адгезии, используя синтетический SLX.
Этот пример подтверждает, что синтетический SLX связывает ELAM-1 и ингибирует адгезию нейтрофилов к активированному эндотелию. Этот пример показывает также, что химическая связь в сиаловой кислоте нарушает связывание с ELAM-1.
Получали два синтетических соединения. Одно содержало сиаловую кислоту в β 2,3 связи, как в естественно встречающемся SLX. Второе содержало сиаловую кислоту в α 2,6 связи, чтобы исследовать важность природы связи со связыванием рецептора.
Липосомы получали при помощи добавления 12 αл абсолютного ETOH в каждую пробирку, быстро нагревая в водяной ванне при 50oC и обрабатывая ультразвуком 2 минуты. В каждую пробирку медленно добавляли 238 μл теплого, буферированного фосфатом, соляного раствора /PBS/, одновременно осуществляя обработку ультразвуком, которую продолжали затем еще в течение 10 минут. Финальная концентрация сырьевых липосом составила 400 μг гликолипидов/мл в 5% ETOH/PBS.
Процедура.
1. HUYEC, PMN и липосомы получали так, как это описано выше.
2. Аналитический планшет со стимулированными HUYEC удаляли из инкубатора и углубления промывали два раза средой RPMI 1640, содержащей 5 мг/мл альбумина бычьей сыворотки /АБС/.
3. Липосомное сырье разбавляли в буфере HBSS/АБС, чтобы получить растворы, эквивалентные: 40 μг/мл, 30 μг/мл, 15 μг/мл, 7,5 μг/мл, 3,75 μг/мл и 1,87 μг/мл. Аналогичные разбавления получали из контрольного сырья, состоящего из PBS-5% ETOH.
4. Среду удаляли из углублений аналитического планшета при помощи пипетки Пастера, из нескольких углублений одновременно.
5. 0,05 мл суспензии каждой липосомы добавляли в двойные углубления на стимулированных аналитических планшетах. В контрольные углубления вводили липосомальный буфер, содержащий этаноловый HBSS и АБС в тех же концентрациях, что и в углублениях, содержащих липосомы. Контрольный буфер наносили на планшет на нестимулированные и стимулированные HUYEC. Образцы, содержащие липосомы, добавляли только в стимулированные углубления.
6. Планшеты инкубировали в течение 40 мин при температуре 37oC, а затем в углубления анализа добавляли 50 μл PMN. Финальная концентрация клеток составляла 5•105 на углубление в 100 μл.
7. Аналитический планшет возвращали в инкубатор /5% CO2, 37oC/ в течение 8 мин.
8. Несвязанные клетки удаляли из углублений планшетов при помощи систематического повторного суспендирования, используя многоканальную пипетку p200, с последующим добавлением и удалением 0,2 мл среды.
9. Всю среду удаляли из углублений и добавляли 50 μл буфера солюбилизации. Он состоял из цитратного буфера /24,3 мл 0,1 М лимонной кислоты, 10,5 г/500 мл + 25,7 мл 0,2 М двухосновного фосфата натрия, 14,2 г/500 мл и затем воды до 100 мл/, содержащего 0,1% детергента NO-40.
10. Планшеты инкубировали на роторном вибраторе в течение 10 мин, а затем в каждое углубление добавляли 0,05 мл раствора OPDA [8 мг орто-фенилен-диамина, фирма Sigma cat P-1528, 8 μл 30% H2O2 и 10 мл цитратного буфера (как выше)] . Реакции давали возможность развиваться в течение 15 минут, а затем в каждое углубление, чтобы прекратить реакцию, добавляли 25 μл 4N раствора серной кислоты.
11. Объем реагентов получали при помощи смещения объемов в 100 μл буфера солюбилизации и раствора OPDA с 50 μ л 4N раствора серной кислоты.
12. 100 μл верхнего слоя удаляли из каждого из 2 углублений и переносили на эластичный аналитический планшет ELISA /фирма Falcon/. Планшет сканировали на спектрофотометре в области 492 нм в течение 30 мин.
Полученные результаты этих двух экспериментов представлены в табл. 5.
Эти результаты показывают, что липосомы, содержащие синтетический α /2, 3/ Сиалил Lex, но не α /2, 6/ Сиалил Lex, ингибируют адгезию нейтрофилов с активированным эндотелием в анализе на связывание, зависящем от ELAM-1. Таким образом, α 2, 3 связь сиаловой кислоты оказывается необходимой для узнавания при помощи ELAM-1. Кроме того, эти результаты показывают, что синтетически продуцированный олигосахарид, α /2, 3/ Сиалил Lex, связывается с ELAM-1 и блокирует связывание нейтрофилов с активированным эндотелием. Аналогичные результаты получали для GMP-140 и они представлены на фиг. 9. Аналогичные результаты обнаружили с GMP-140. См. фиг. 9. Это соединение или производные этого соединения, таким образом, становятся потенциальными кандидатами на противовоспалительный лекарственный препарат.
Пример XI. Обработка клеток HL-60 эндо- β -галактозидазой.
В этом примере описаны эксперименты, позволяющие определить, доступна ли внутренняя сахарная связь основной цепи β -галактозы сиалилированного Lex клеток HL-60 расщеплению при помощи Эндо- β -галактозидазы, фермента, известного для использования с целью расщепления внутренней связи β -галактозы в полилактозаминиловых структурах, но не β -gal, когда GlaNAc присоединена к маннозе /структурам типа ядра/.
Процедура.
Тромбоциты выделяли и активировали при помощи тромбина с использованием приемов, описанных выше. Активированные при помощи 1l-1 β HUYEC получали, как это было описано выше. Культивируемые клетки HL-60 обрабатывали эндо β -галактозидазой, как это описано ниже, и определяли эффект обработки фермента на адгезию с участием GMP-140 клеток HL-60 с активированными тромбоцитами.
Обработку ферментом клеток HL-60 осуществляли следующим образом: 12,4 • 106 клеток дважды промывали сбалансированным раствором солей Хэнкса, содержащим 20 мМ HEPES и 0,2% глюкозы, затем осуществляли стадию однократной промывки в нормальном соляном растворе. Эндо- β -галактозидазу /0,1 единицу, фирма ICN Chemicals Inc., Irvine, CA/ растворяли в 200 мкл нормального соляного раствора и 200 μл буфера ацетата натрия, pH 6,01. 200 μл /содержащих 0,05 единиц фермента/ добавляли в 3 • 106 клеток HL60, и в такое же число клеток добавляли 200 μл ацетатного буфера, последнее использовали в качестве буферного контроля. Обе пробирки инкубировали при температуре 37oC в течение 60 мин при осторожном встряхивании. Затем пробирки охлаждали во льду и клетки промывали три раза в HBSS, содержащем HEPES и глюкозу, далее осуществляли подсчет и суспендировали в 2 • 106/мл.
Для анализа с GMP-140 20 μл буфера Tyrode-HEPES, pH 7,2 помещали в пробирку Эппендорфа. Добавляли такой же объем активированных тромбоцитов /2 • 108/ мл/ и клеток HL-60 /2 • 106/мл/ и после перемешивания пробирки выдерживали при комнатной температуре в течение 20 мин. Адгезию тромбоцитов с клетками HL60 анализировали под микроскопом, как это было описано ранее для адгезии активированных тромбоцитов с нейтрофилами.
Для ELAM-1 анализа обработку ферментом клеток HL-60 осуществляли, как это было описано выше, за тем исключением, что эти клетки одновременно метили при помощи 51Cr, как это было описано ранее. Адгезию с участием ELAM-1 прекращали при помощи инкубирования 2 • 105 обработанных или необработанных клеток с активированными IL-1HUYEC в течение 30 минут при температуре 4oC, а затем промывали пластинку при помощи пипетки Пастера.
Результаты этих экспериментов показывали, что обработка клеток HL-60 эндо-β-галактозидазой ингибировали их способность связываться с /1/ активированными тромбином тромбоцитами на 87,5% и /2/ активированными 1L-1 β HUYEC при температуре 4oC на 70%. Таким образом, минимальный SLX-содержащий тетрасахаридный лиганд для GMP-140 вероятно присоединяется с лактозе или полилактозаминиловой структуре, а не к маннозе.
Пример XII. Фукозилированный полисахарид блокирует связывание нейтрофилов с тромбоцитами.
В этом примере способность фукозилированного полисахарида ингибировать адгезию с участием GMP-140 сравнивали с такими же свойствами нефукозилированного полисахарида, гексасахарида SLX и Lex. Если коротко, то тромбоциты и нейтрофилы выделяли при помощи процедур, описанных выше. Тромбоциты активировали тромбином, а затем инкубировали при помощи разбавлений различных олигосахаридов. Добавляли нейтрофилы и определяли эффект сахаридов на адгезию нейтрофилов с активированными тромбоцитами. Использовали олигосахариды: Нативный полисахарид и его фукозилированное производное /получение обеих описано ниже/; SLX-гексасахарид, LNF III /Lex/ и LNF1 /структуры описаны выше/.
Превращение полисахарида, который содержит структуру линейного ядра SLX, в многовалентный, содержащий SLX, полисахарид осуществляли при помощи ферментативного фукозилирования. Нативный полисахарид типа 1a получали из Streptococcus Группы B, как это описано в Iennings и др., Biochem. т.22, стр. 1258-1263 /1983/, которая здесь включена в качестве ссылки. Соответствующие бактериальные штаммы сданы на хранение в Американское Собрание Типов Культур /АТСС/ под номерами хранения NN 12400, 31574, 12401 и 31575.
Чтобы получить фукозилированный полисахарид, нативный полисахарид типа 1a /1 мг/ растворяли в смеси 6 μл 1 М хлорида марганца, гуанозин 5'-дифосфата β -L-фукозы с радиомеченным индикатором /удельная активность 1,82 • 106 отсчетов μмоль/, 0,9 μмоль в воде 90 μл и воды 137 μл. В эту смесь добавляли 100 μ л раствора 3/4 фукозил трансферазы, выделенной из человеческого молока, как это ранее было описано Prieels и др., J. Biol. Chem. т.256, стр. 10456-10463 /1981/, которая здесь включена в качестве ссылки.
Реакционную смесь концентрировали против мембраны /срезающей 100 К/ несколько раз с водой, а то, что оставалось, подвергали лиофилизации, чтобы получить порошок. Это твердое вещество растворяли в воде и пропускали через слабую катионообменную колонну, чтобы удалить любой оставшийся протеин. Радиоактивные фракции, содержащие фукозилированный полисахарид, собирали и подвергали лиофилизации. Приблизительно пятьдесят из доступных боковых цепей фукозилировали, что устанавливали при помощи введения радиометки.
Процедура.
Тромбоциты выделены, как это описано выше, и активировали /2 • 108/мл/ при помощи инкубирования в течение 20 минут при комнатной температуре с тромбином при финальной концентрации 0,25 Ед/мл.
Нейтрофилы выделяли при помощи образования слоя обработанной гепарином крови на Моно-Поли растворяющей среде /фирма Ficoll-Hypague, Flow Zaboratories/, а затем центрифугировали в течение 25 мин со скоростью 2000 об/мин, а затем еще 25 минут со скоростью 2500 об/мин, как это описано выше.
Для этого анализа 20 μл суспензии тромбоцитов /2 • 108/мл/ помещали в пробирку для центрифуги Эппендорфа. Добавляли равный объем препараций олигосахаридов в концентрациях от 500 μг/мл до 2,0 μг/мл, и пробирки выдерживали при комнатной температуре 20 мин. Затем добавляли двадцать μл препарации нейтрофилов /2 • 106/мл/ и пробирки выдерживали еще в течение 20 минут при комнатной температуре.
Адгезию активированных тромбоцитов с нейтрофилами анализировали под микроскопом. Изучали сто нейтрофилов. Их оценивали как положительные, если 2 и более тромбоцитов были присоединены, и как отрицательные, если менее 2 тромбоцитов при этом связывалось. Рассчитывали процент клеток с 2 и более связанными тромбоцитами.
Как показано в таблице 6, фукозилированные полисахариды весьма эффективно ингибировали связывание нейтрофилов с активированными тромбином тромбоцитами с участием GMP-140-50% ингибирования достигали при помощи менее 1 μг/мл. Это сравнивали с 20 μг/мл - в такой концентрации был необходим нативный полисахарид, и с 8 μг/мл - в такой концентрации был необходим SLX-гексасахарид для аналогичной степени ингибирования.
Пример XIII. Защита крыс от эндотоксического шока /гибели/ при помощи моноклонального антитела относительно LAM-2.
Этот пример подтверждает эффективность mAb P6E2 /мышиного 1gG-3k, функционального анти-человеческого ELAM-1 mAb, описанного в одновременно находящейся в стадии рассмотрения патентной заявке U.S.S.N 07/645878, которая здесь включена в качестве ссылки/ в животной модели вызванной липополисахаридом гибели. Выбирали систему крысы ввиду того, что P6E2, как было показано, осуществляет перекрестное взаимодействие с крысиным эквивалентом ELAM-1.
Материалы и методы.
LPS из E.coli 0111:B4 /фирма Sigma/ получили свежим из одной партии за один день до использования при помощи растворения стерильного, не содержащего пирогенов, соляного раствора при концентрации 5 мг/мл. Раствор обрабатывали ультразвуком на льду в течение 30 секунд, используя средство для разрушения клеток ультразвуком типа Текмарк /Tekmark/. Непосредственно перед использованием материалы обрабатывали ультразвуком второй раз в течение 30 секунд.
Самок крысы Льюиса весом 200 г (± 10 г/ получали от фирмы Charles River Breeding Labs и содержали в течение по крайней мере 7 дней после получения /для адаптации/. Использовали группы /по 10 животных/, если не указано явно противное. Все реагенты инъектировали парентерально через хвостовую вену. В качестве отрицательных контрольных животных использовали животных, которым давали либо стерильный, не содержащий LPS, соляной раствор, либо мышиный протеин миеломы IgG 3k / J 606, низкий проген - < 2 нг/мг протеина/.
Результаты.
Протоколы доза P6E2/график получали эмпирически из фармакокинетических данных, которые получали при применении с профилактическими целями P6E2 к крысам. Минимальную ЛД100 дозу определяли эмпирически и она составляла 7,5 мг/кг для этих крыс.
В одном эксперименте крыс обрабатывали дозой 10 мг/кг за один час до введения LPS. Препарат вводили через 3 часа после LPS. 4/10 обработанных животных выживало. Напротив, все 10 контрольных животных /которым инъектировали соляной раствор/ погибали. За период наблюдения в двадцать четыре часа у выживших животных можно было наблюдать некоторые клинические признаки, характерные для обработанных LPS животных.
Заявитель в дальнейшем испытывал дозы P6E2, которые /1/ в два раза превышали дозу из первого эксперимента, и /2/ порядок величины был ниже чем 10 мг/кг, которая использовалась в первом эксперименте. Результаты показывают, что P6E2 имел значительный эффект: 80% животных выживало при дозе 10 мг-кг /фиг. 10/. Заметим, что одно животное выжило в контрольной группе соляного раствора - что наводило на мысль, что эта группа животных была не "самой удачной" в смысле тяжести исхода.
Осуществляли еще одно исследование для того, чтобы подтвердить терапевтическую ценность P6E2. Животным давали с кормом дозу 10 мг/кг P6E2 за 1 час до или через 2, 4 или 6 часов после введения LPS.
Снова 1 из 10 животных выживало в группе, обработанной соляным раствором, а также в группе, обработанной в дозе 10 мг/кг протеина миеломы J 606 при T = -60 минут /фиг. 11/. P6E2 обладал защитным действием, даже когда его применяли через 2 или 4 часа после LPS.
Заключение.
Защита, отмеченная для Cytel mAb P6E2, подтверждает важность ELAM-1 в животных моделях смертельных заболеваний.
Пример XIV. "Прокатка" липосом SLEX на активированных 1L-1эндотелиальных клетках кролика.
Цель.
Этот эксперимент подтверждает, что сиалилированный Льюис /SLX/ и сиалопараглобозид /SPG/, выделенные из биологических источников и включенные в липосомы, будут "кататься" вдоль активированных венул кролика.
"Прокатка" представляет собой раннее межклеточное взаимодействие между лейкоцитами и стенками эндотелиальных клеток. Лейкоцит будет "кататься" на эндотелиальных клетках. Далее лейкоцит будет либо /1/ высвобождаться снова в циркуляцию, либо /2/ "прилипать" к эндотелиальной клетке, и начинаются ранние события, которые завершаются воспалением. Селектины являлись следствием взаимодействий типа клетка-клетка при "качении", см. von Adrian и др., Proc. Natl. Acad. Sci. USA, 1991.
Материалы.
Липосомы, содержащие гликосфинголипиды, получали следующим образом: 50 уг гликолипида добавляли в 150 уг фосфатидилхолина /фирма Sigma, яичный желток/, 250 уг холестерина /фирма Сигма/ и 1 мМ карбоксифторсцеина /фирма Сигма/ в смеси хлороформ: метанол /2:1/, и весь раствор выпаривали до сухого состояния в стеклянных пробирках с завинчивающимися пробками. Липосомы получали при помощи добавления 12,5 ул абсолютного этанола в каждую пробирку, краткого нагревания и водяной ванне и обработки ультразвуком в течение 2 минут. 238 ул теплого PBS медленно добавляли в каждую пробирку при одновременной обработке ультразвуком и такую обработку продолжали всего в течение 10 минут. Липосомы переносили в 1 мл с PBS и подвергали центрифугированию со скоростью 14000 об/мин в течение 2 минут для того, чтобы удалить избыток EtOH и карбоксифторсцеина. Верхний слой сбрасывали, а липосомы снова суспендировали в 1 мл PBS, анализировали на гемацитометре и регулировали с целью обеспечения 5 •106 липосом на мл. Один мл инъектировали кролику, "активированному" 4 часами раньше при помощи 1L-1.
"Качение" наблюдали с использованием витального микроскопа, как это описано у фон Адриана и др. Брыжейку активированного при помощи 1L-1 кролика временно извлекали и раскладывали на нагретом до 37oC стеклянном окне на столике микроскопа. Брыжейку опускали в ванну при температуре 36, 5oC с соляным раствором, уравновешенным 5% CO2 в N2. Взаимодействия липосом-эндотелия наблюдали при помощи витального микроскопа с 50Х объективом при погружении в соленую воду. Липосомы делали наблюдаемыми при освещения флюоресцентным светом. За одну минуту подсчитывали липосомы в сегменте выбранной венулы брыжейки /20-40 μм в диаметре/.
Результаты. Качение липосом по активированным при помощи 1L-1 клеткам кролика
Инъектируемый материал - качение
человеческие нейтрофилы - 4 /максимальное количество качений/
SLex липосомы - 2 \\\SPG липосомы - 0 /качение отсутствует/
Заключение.
В дополнение к LAM-1 /фон Андриан и др./, ELAM-1 и его лиганды, как оказалось, включены в процессе скопления лейкоцитов по краям или "качения".
Хотя вышеупомянутое изобретение было описано в некоторых деталях при помощи иллюстрации и примера с целью большей ясности, должно быть также ясно, что могут быть осуществлены некоторые изменения и модификации, оставаясь в области, охватываемой приложенной формулой изобретения.
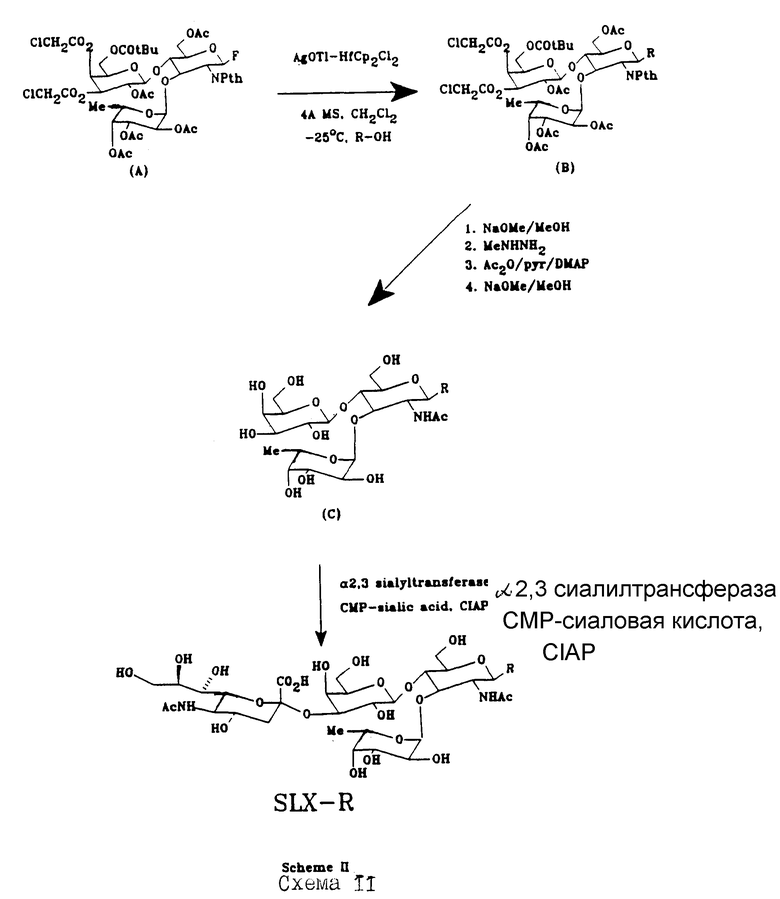
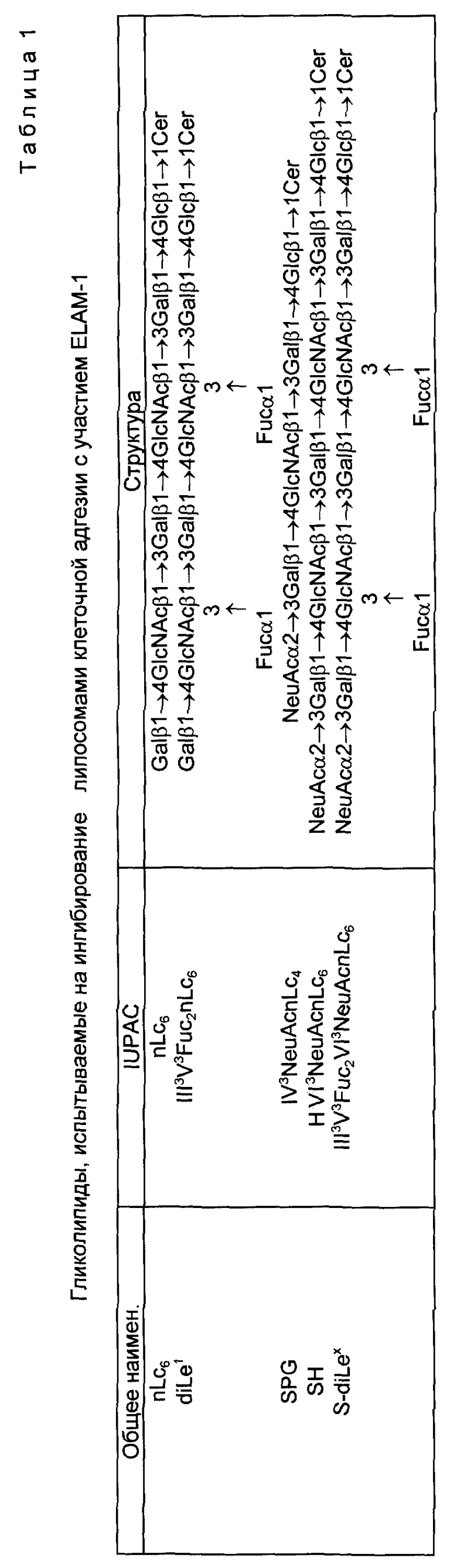
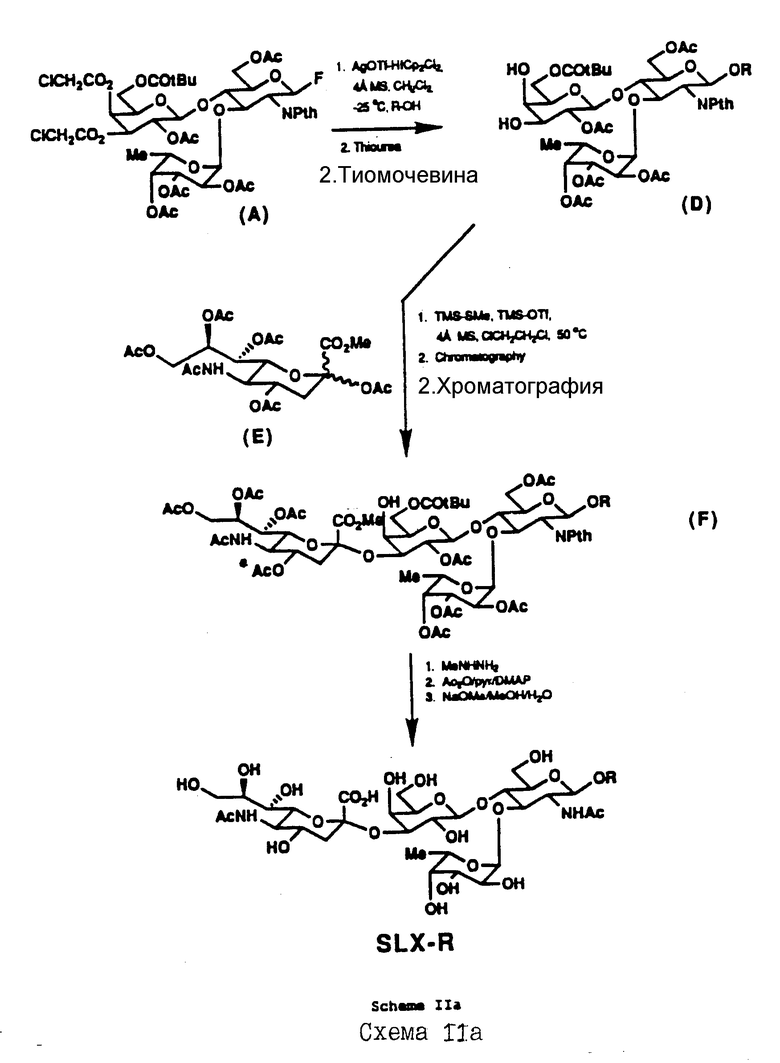
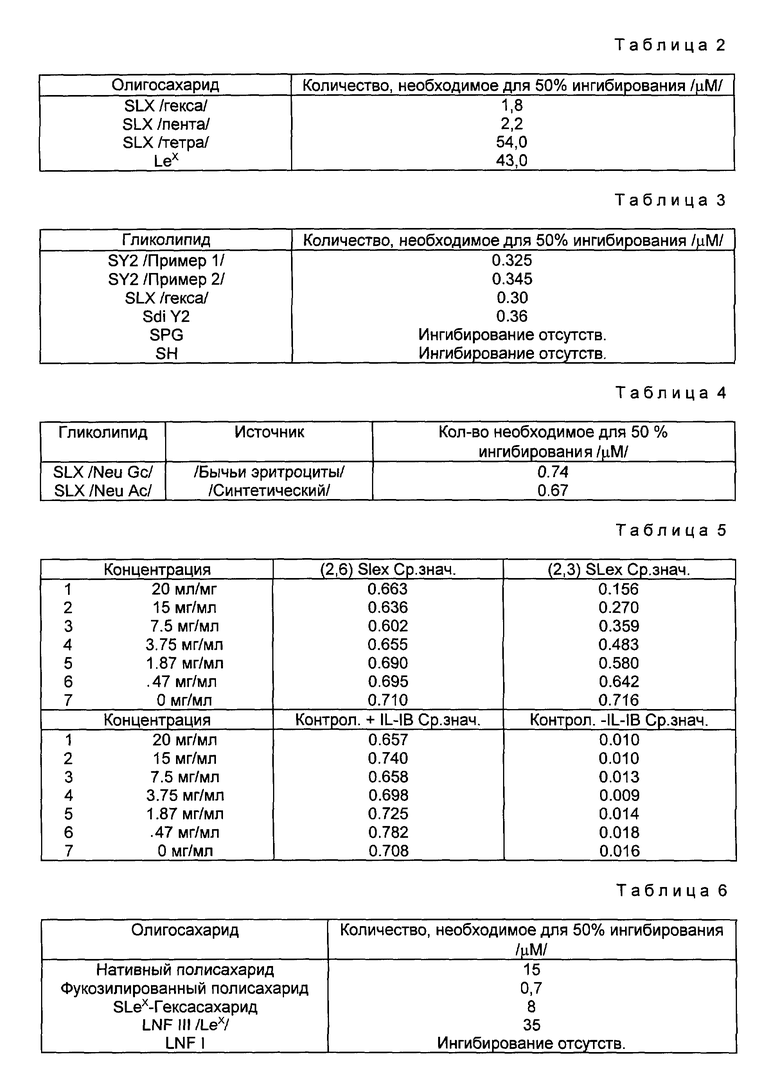
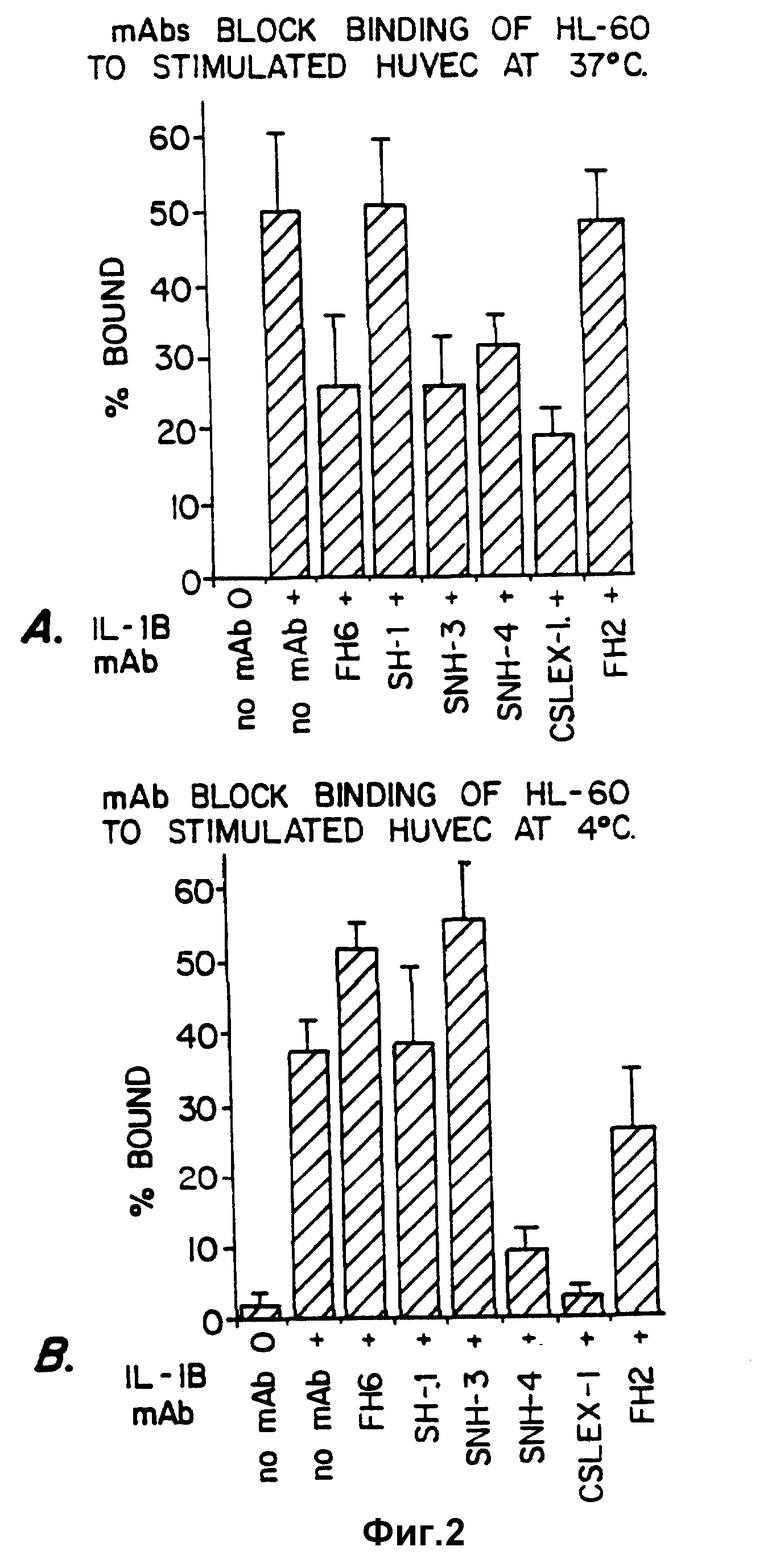
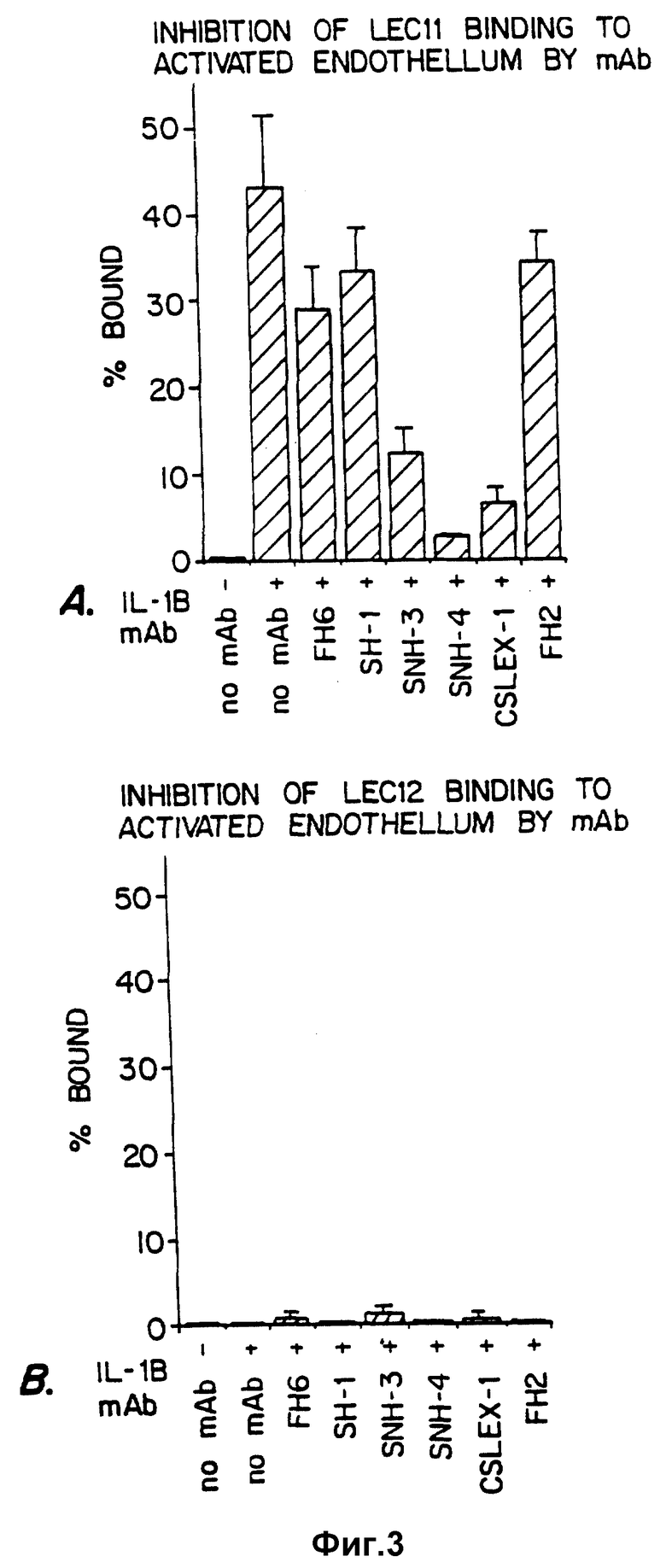
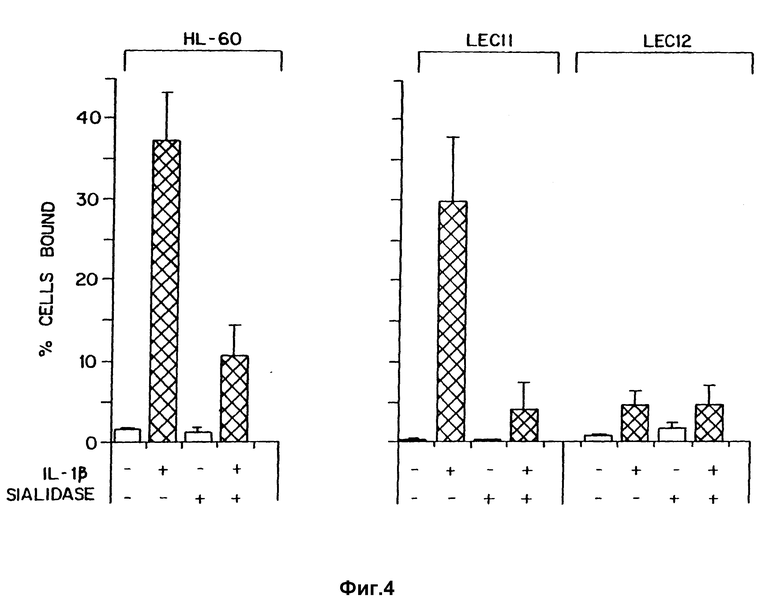
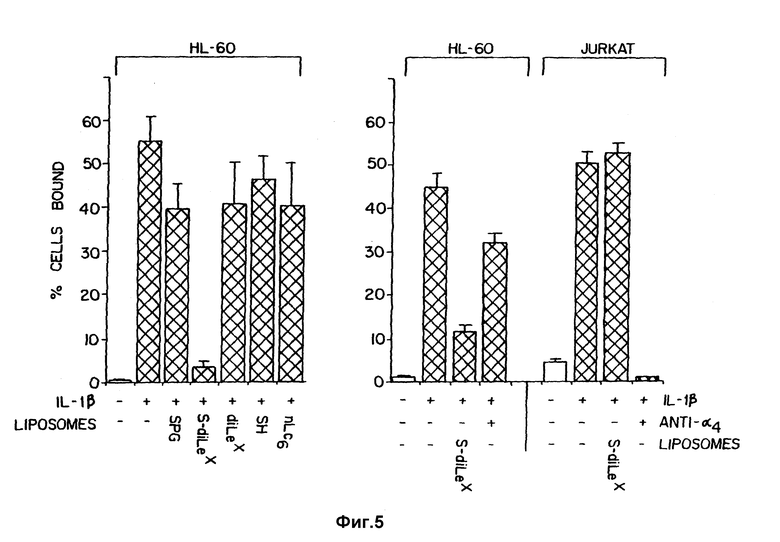
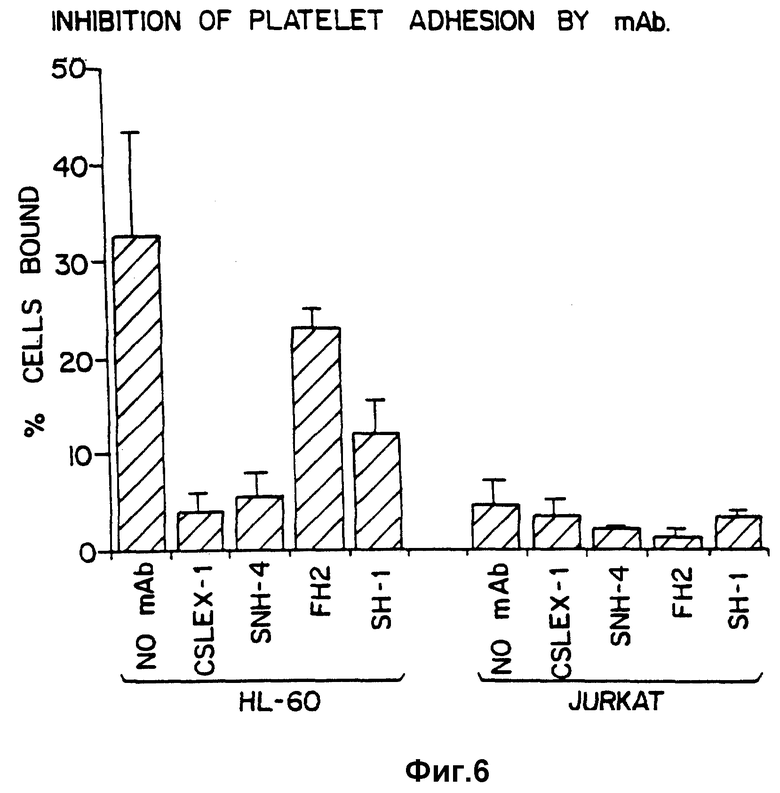
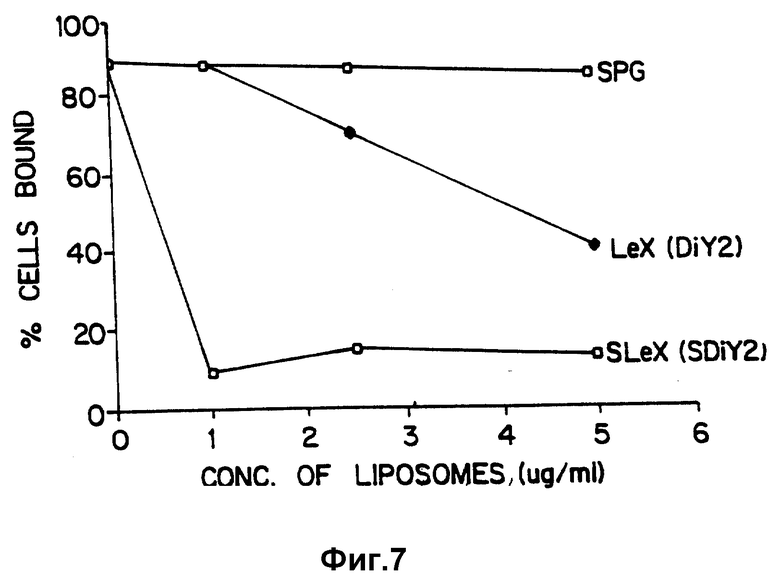
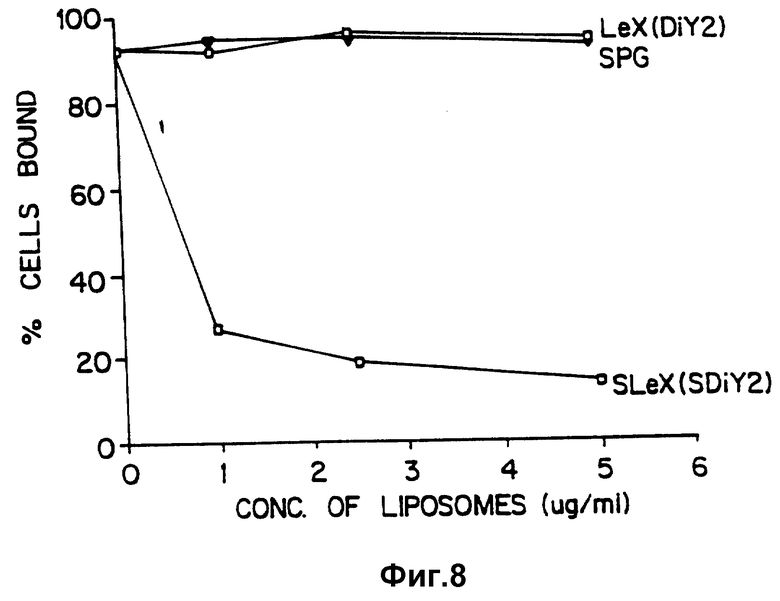
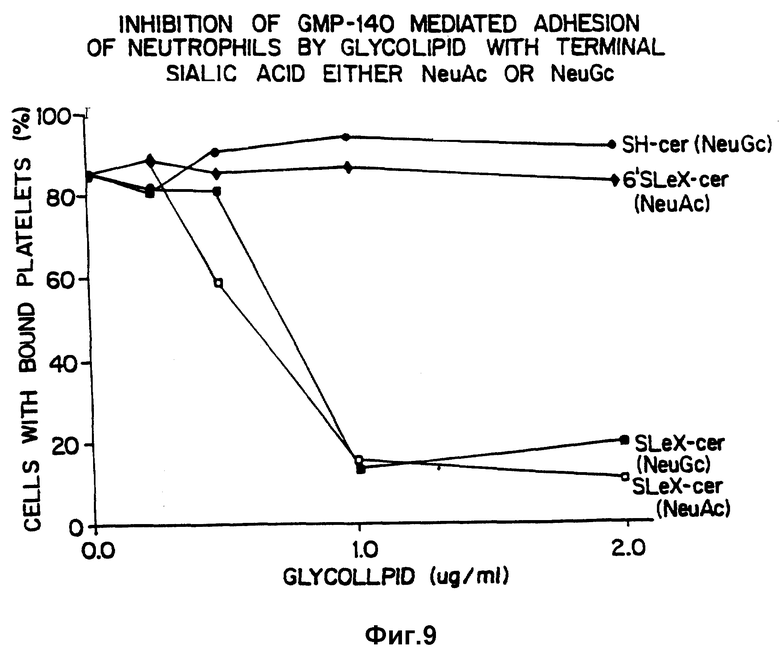
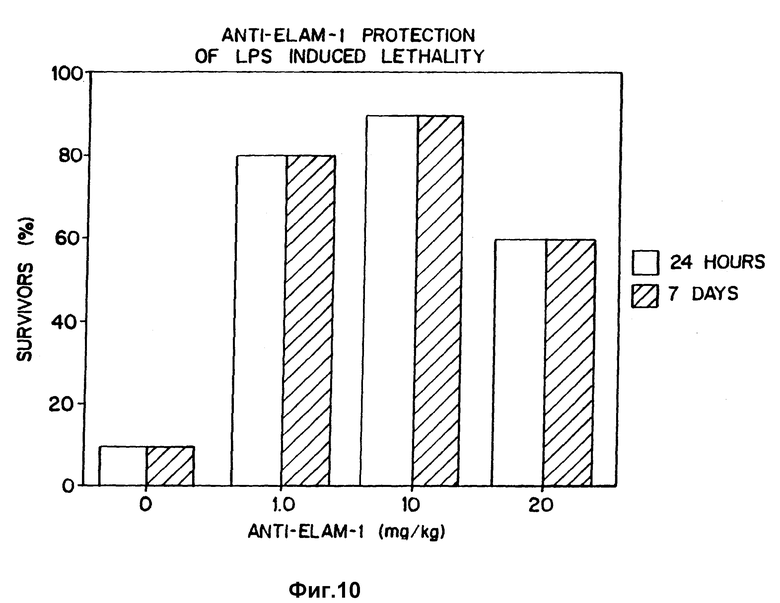
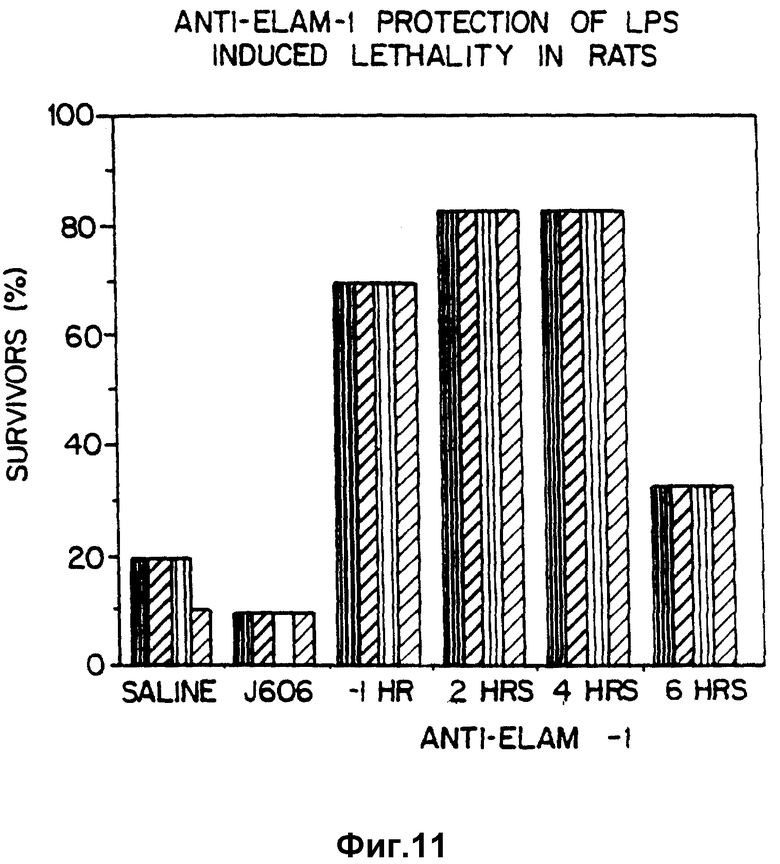
Изобретение относится к композициям и способам для уменьшения или регулирования воспаления и для лечения воспалительных заболеваний и других патологических нарушений, в которых участвует межклеточная адгезия. В соответствии с настоящим изобретением предлагаются композиции, которые селективно связываются с рецептором селектина и которые содержат по крайней мере одну связывающуюся с селектином составляющую. Связывающие селектин составляющие имеют общую формулу: R1 -Galβ1,4 (Fucα1,3)GlcNAc-(R2)a-X, в которой R1 является олигосахаридом или R3 - R4 - C (CO2H)-, в которой R3 и R4, которые могут быть одинаковыми, так и различными, являются - H, - C1 - C8 алкилом, - гидрокси C1 - C8 алкилом, -арил C1 - C8 алкилом или - алкокси C1 - C8 алкилом, в которой R2 является β1,3Gal, α1,2Man или α1,6GalNAc, a = 0 или 1. Изобретение расширяет арсенал средств для регулирования патологических нарушений, в которых участвует межклеточная адгезия. 13 с. и 66 з.п. ф-лы, 11 ил., 6 табл.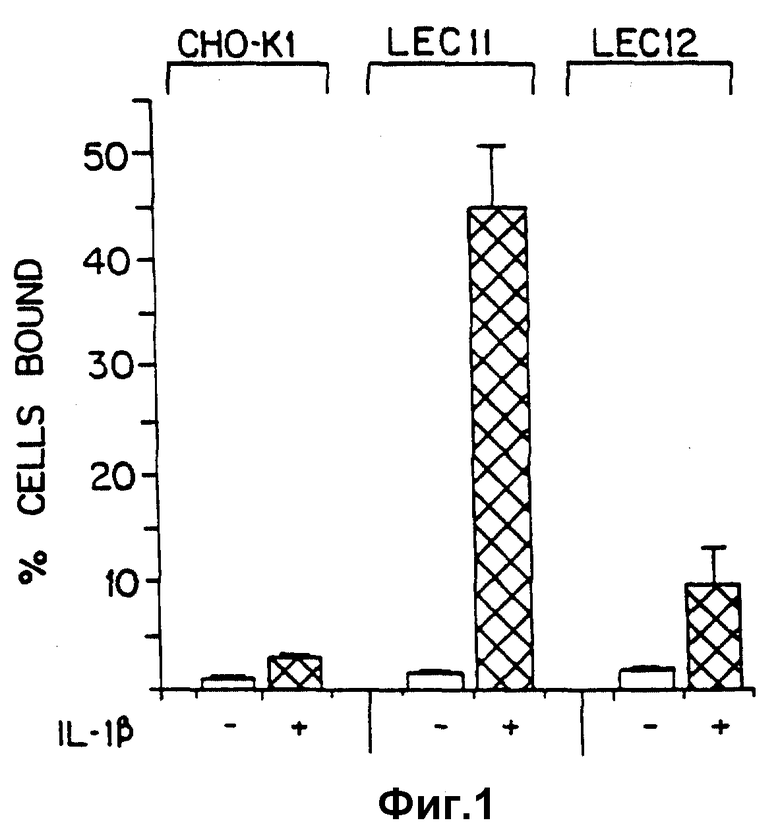
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a-X,
в которой R1 является элементом, выбранным из группы, состоящей из сиаловой кислоты, олигосахарида, содержащего сиаловую кислоту и группы, имеющей формулу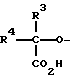
в которой R3 и R4, взяты отдельно, которые могут быть одинаковыми или различными, выбирают из группы, состоящей из H, C1-C8 алкила, окси (C1-C8 алкила), арил-(C1-C8 алкила) и (C1-C8 алкокси) - (C1-C8 алкила); или
R3 и R4, взятые вместе, образуют 4 - 8 членное кольцо;
R2 является β1, 3Gal, α1,2Man или α1,6GalNAc и a равно 0 или 1,
X является H, низшим (C1-C8) алкилом, арилалкилом, окси (C1-C8) алкилом или (C1-C8) алкокси-(C1-C8) алкилом.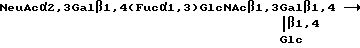
10. Композиция по п.8, отличающаяся тем, что полисахарид содержит повторяющуюся единицу, имеющую формулу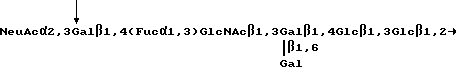
11. Композиция по п.8, отличающаяся тем, что полисахарид представляет собой фукозилированный полисахарид Streptococcus Группы B, типа Ia.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a-Y
в которой R1 выбирают из группы, состоящей из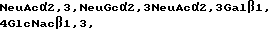
NeuGcα2,3Galβ1,4GlcNAcβ1,3,
R2 является β 1,3Gal, α 1,2 Man или α 1,6GalNAc, и a равно 0 или 1;
Y является протеином или липидом.
NeuAcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a-X,
NeuGcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a-X,
NeuGcα2,3Galβ1,4GlcNAcβ1,3Galβ1,4(Fucα1,3)GlcNAc-R)a-X,
в которой R является β 1,3 Gal, α 1,2 Man или α 1,6 Gal A и a равно 0 или 1,
X является H, низшим алкилом (C1-C8), арилалкилом, гидрокси-(C1-C8)алкилом или (C1-C8) алкокси-(C1-C8) алкилом.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a-X,
в которой R1 является сиаловой кислотой, R3(R4)C(CO2H)-, или трисахаридом, содержащим сиаловую кислоту;
R3 и R4, которые могут быть одинаковыми или различными, являются H, C1-C8 алкилом, оксизамещенным C1-C8 алкилом, арилзамещенным C1-C8 алкилом или алкоксизамещенным C1-C8 алкилом;
R2 является β 1,3Gal, α 1,2 Man или α 1,6GalNAc и a равно 0 или 1;
X выбирают из группы, состоящей из H, низшего (C1-C8) алкила, окси(C1-C8) алкила, (C1-C8) алкокси (C1-C8)алкила, арила, алкиларила и арилалкила.
NeuAcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a,
NeuGcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a,
NeuGcα2,3Galβ1,4GlcNAcβ1,3Galβ1,4(Fucα1,3)GlcNAc-R)a,
в которой R является β1,3 Gal, α1,2 Man или α1,6 GalNAc и a равно 0 или 1.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a-X,
в которой R1 является частью, выбранной из группы, состоящей из сиаловой кислоты, олигосахарида, содержащего сиаловую кислоту, и группы, имеющей формулу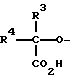
в которой R3 и R4, взятые отдельно, которые могут быть одинаковыми или различными, выбирают из группы, состоящей из H, C1-C8 алкила, окси-(C1-C8 алкила), арил-(C1-C8 алкила) и (C1-C8 алкокси) - (C1-C8) алкила; или
R3 и R4, взятые вместе, образуют 4 - 8-членное кольцо,
R2 является β1,3 Gal, α1,2 Man или α1,6 GalNAc, и a равно 0 или 1;
X является H, низшим (C1-C8) алкилом, арилалкилом, окси-(C1-C8) алкилом или (C1-C8) алкокси (C1-C8) алкилом.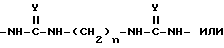
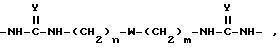
в которой n и m, которые могут быть одинаковыми или различными, являются целыми числами от 2 до 12; Y является O или S; а W является O, S или NH.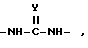
в которой Y является O или S; a заместители находятся в цис- или транс- положении.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a,
в которой R1 является сиаловой кислотой, R3(R4)C(CO2H)-, или трисахаридом, содержащим сиаловую кислоту,
где R3 и R4, которые могут быть одинаковыми или различными, являются H, C1-C8 алкилом, гидроксилзамещенным C1-C8 алкилом, арилзамещенным C1-C8 алкилом или алкоксизамещенным C1-C8 алкилом;
R2 является β 1,3Gal, α1,2 Man или α1,6GalNAc и a равно 0 или 1.
NeuAcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a-,
NeuGcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a-,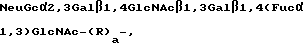
в которой R является β 1,3Gal, α 1,2 Man или α 1,6 GalNAc и a равно 0 или 1.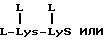
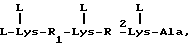
в которой R1 и R2, которые могут быть одинаковыми или различными, являются любым аминокислотным остатком, а L является олигосахаридной частью.
NeuAcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a-,
NeuGcα2,3Galβ1,4(Fucα1,3)GlcNAc-(R)a-,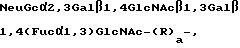
в которой R является β 1,3 Gal, α 1,2 Man или α 1,6 GalNAc и a равно 0 или 1.
R1-Galβ1,4(R2)GlcNac-(R3)a-X,
в которой R1 является NeuAc α2,3, NeuGc α2,3, NeuAcα 2,3, Galα 1,4 GlcNAc β 1,3, или NeuGc β2,3 Gal α1,4 Glc A β1,3;
R2 является Fuc β 1,3, Aragal 1,3, /R,S/-5-алкил-Ara α 1,3 и /R,S/-5-арил-Ara α1,3;
R3 является 1,3 β Gal, 1,2 αMan или 1,6 αGalNAc и a равно 0 или 1;
X является H, низшим (C1-C8) алкилом, арилалкилом, окси-(C1-C8) алкилом или (C1-C8) алкокси (C1-C8) алкилом.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a-X,
в которой R1 является сиаловой кислотой, R3(R4)C(CO2H)- или трисахаридом, содержащим сиаловую кислоту;
где R3 и R4, которые могут быть одинаковыми или различными, являются H, C1-C8 алкилом, гидроксизамещенным C1-C8 алкилом, арилзамещенным C1-C8 алкилом или алкоксизамещенным C1-C8 алкилом; и
R2 является β 1,3Gal, α 1,2 Man или α 1,6GalNAc и a равно 0 или 1;
X является H, низшим (C1-C8) алкилом, арилалкилом, окси-(C1-C8) алкилом или (C1-C8) алкокси (C1-C8) алкилом.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a-X,\
в которой R1 является сиаловой кислотой, R3(R4)C(CO2H) или трисахаридом, содержащим сиаловую кислоту;
где R3 и R4, которые могут быть одинаковыми или различными, являются H, C1-C8 алкилом, гидроксизамещенным C1-C8 алкилом, арилзамещенным C1-C8 алкилом или алкоксизамещенным C1-C8 алкилом; и
R2 является ββ 1,3Gal, α 1,2 Man или α 1,6 GalNAc и a равно 0 или 1;
X является H, низшим (C1-C8) алкилом, арилалкилом, окси-(C1-C8) алкилом или (C1-C8) алкокси-(C1-C8) алкилом.
R1-Galβ1,4GlcNAc-,,
в котором R является сиаловой кислотой.
R1-Galβ1,4(Fucα1,3)GlcNAc-(R2)a,
в которой R1 является сиаловой кислотой, R3(R4)C(CO2H)- или трисахаридом, содержащим сиаловую кислоту;
где R3 и R4, которые могут быть одинаковыми или различными, являются H, C1-C8 алкилом, гидроксизамещенным C1-C8 алкилом, арилзамещенным C1-C8 алкилом или алкоксизамещенным C1-C8 алкилом;
R2 является β1,3Gal, α1,2 Man или α1,6 Gal NAc и a равно 0 или 1.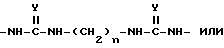
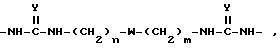
в которой n и m, которые могут быть одинаковыми или различными, являются целыми числами от 2 до 12; Y является 0 или S; а W является О, S или NH.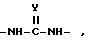
в которой Y является O или S; а заместители находятся в цис- или транс- положении.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| EP заявка 0213072, 04.03.87 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| US 5061699, 29.10.91 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| M.P | |||
| Bevilacgua et al | |||
| Endothelial Liucocyte Adhesion Molecule., Science, v.243, pp.1160, 1989. | |||
