Область изобретения
Настоящее изобретение относится к новым веществам, связывающимся с Helicobacter pylori, и их применению, например, в фармацевтических композициях и способах лечения состояний, вызванных Helicobacter pylori.
Предпосылки к созданию изобретения
Адгезию микроорганизмов считают первым этапом патогенеза инфекций, когда специфичность адгезинов инфекционного агента, с одной стороны, и рецепторные структуры, экспрессируемые эпителиальными клетками органа-мишени хозяина, с другой, представляют собой важные определяющие факторы ряда хозяев и тканевого тропизма патогена [1].
Человеческий желудочный патоген Helicobacter pylori представляет собой этиологический агент хронического поверхностного гастрита [2] и также связан с развитием язвы двенадцатиперстной кишки, язвы желудка и аденокарциномы желудка [3-7]. Указанный микроорганизм поражает отчетливо определенный ряд хозяев и имеет выраженный тканевый тропизм, т.е. он требует наличия человеческого эпителия желудочного типа для колонизации [8]. В желудке человека большинство бактерий находят в слое слизи, но показана также избирательная ассоциация бактерий с поверхностными мукоцитами [8, 9].
Ранее было показано несколько различных связывающих специфичностей Helicobacter pylori. Так, сообщалось о связывании бактерии с такими разнообразными соединениями, как фосфатидилэтаноламин и ганглиотетраозилцерамид [10, 11], детерминанта группы крови Leb [12], гепарансульфат [13], ганглиозид GM3 [14], сульфатид [14, 15] и лактозилцерамид [16]. Зависящее от сиаловой кислоты связывание Helicobacter pylori с большим комплексом гликосфинголипидов (полигликозилцерамидов) человеческих эритроцитов, гранулоцитов и плаценты, также подтверждена документально [17, 18].
Помимо своей связи с желудочно-кишечными заболеваниями Helicobacter pylori связана с множеством заболеваний, также поражающих другие органы кроме органов желудочно-кишечного тракта [74]. Например, были показаны связи с заболеваниями сердца, особенно атеросклерозом [75], заболеваниями печени, включая аденокарциному печени [76, 77], заболеваниями кожи [78] и синдромом внезапной младенческой смерти [79, US 6083756].
Краткое описание существа изобретения
Главной целью настоящего изобретения является создание новых путей для лечения состояний, вызываемых Helicobacter pylori.
Настоящее изобретение основано на открытии специфичного рецептора Helicobacter pylori в эпителии желудка человека. Во многих случаях указанный рецептор представляет собой гликолипид, лактотетраозилцерамид, который обнаруживают исключительно в желудочно-кишечном тракте, и во время исследовательской работы было показано, что эпитоп минимального связывания представляет собой Galβ3GlcNAc или очень сходную структуру Galβ3GalNAc.
Настоящее изобретение, таким образом, относится к веществам, связывающимся с Helicobacter pylori, которые включают указанный связывающий эпитоп, или к их аналогам или производным.
Одной целью настоящего изобретения является создание фармацевтических композиций для лечения состояний, вызванных Helicobacter pylori.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, для производства фармацевтических композиций для лечения состояния, обусловленного присутствием Helicobacter pylori.
Другой целью настоящего изобретения является создание способа лечения состояния, обусловленного присутствием Helicobacter pylori.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, для идентификации бактериальных адгезинов.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, для ингибирования связывания с Helicobacter pylori как для терапевтических целей, так и для немедицинских целей, таких как анализы in vitro.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, как ведущих соединений для идентификации других веществ, связывающихся с Helicobacter pylori.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, в пищевых продуктах или пищевых добавках.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, или упомянутых выше бактериальных адгезинов для производства вакцин против Helicobacter pylori.
Другой целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, для диагностики инфекций, вызванных Helicobacter pylori.
Еще одной целью настоящего изобретения является применение упомянутых выше веществ, связывающихся с Helicobacter pylori, для типирования Helicobacter pylori.
Подробное описание изобретения
Как упоминалось выше, настоящее изобретение относится к специфичному веществу, которое связывается с Helicobacter pylori. В работе, результатом которой стало настоящее изобретение, множество различных штаммов Helicobacter pylori метаболически пометили 35S-метионином и исследовали их связывание с рядом различных натуральных гликосфинголипидов, разделенных на пластинках для тонкослойной хроматографии. С помощью ауторадиографии были выявлены в повторностях две отличающихся друг от друга специфические формы. Как было подробно описано ранее, Helicobacter pylori связывается с лактозилцерамидом, ганглиотриаозилцерамидом и ганглиотетраозилцерамидом [16]. Единственная связывающая активность, первоначально выявленная на человеческом материале, относилась к соединению тетрагликозилцерамидного ряда в некислотной фракции мекония человека.
Гликосфинголипидный состав человеческого желудочного эпителия изучен не слишком хорошо. Однако в недавнем исследовании гликосфинголипидов клеток слизистых оболочек и подслизистой ткани желудочно-кишечного тракта человека [55] сообщалось об обогащении сульфатидами слизистой оболочки дна и антрального отдела желудка. Большая часть некислотных гликосфинголипидов на пластинках для тонкослойной хроматографии мигрировала как галактозилцерамид, лактозилцерамид, глоботриаозилцерамид и глобозид, в то время как главные ганглиозиды мигрировали как GM3, GM1 и GD3. Лактозилцерамид, связывающий Helicobacter pylori, с фитосфингозином и гидроксильными жирными кислотами, в эпителиальных клетках желудка человека также был описан [16].
Помимо этого, Cad-активный ганглиозид группы крови (GalNAcβ4 (NeuAcα3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer) был идентифицирован в области дна желудка человека [56], в то время как он не был обнаружен в пилорическом отделе [57], что указывает на дифференциальную экспрессию гликосфинголипидов в различных отделах желудка человека.
В силу ограниченного доступа к ткани желудка человека авторы настоящего изобретения первоначально сосредоточились на гликосфинголипиде, связывающемся с Helicobacter pylori, выявленном в меконии человека, который представляет собой первый стерильный кал новорожденного и состоит, главным образом, из отторгнутых клеток слизистой оболочки развивающегося желудочно-кишечного тракта. После выделения указанный гликосфинголипид, связывающийся с Helicobacter pylori, был охарактеризован посредством масс-спектрометрии, протонной ЯМР-спектроскопии и анализа метилирования как Galβ3GlcNAcβ3Galβ4Glcβ1Cer (лактотетраозилцерамид). Тканевое распределение лактотетраозилцерамида весьма ограничено. До недавних пор лактотетраозилцерамид был выделен только из мекония человека [45], из тонкого кишечника индивидуума, перенесшего ранее резекцию по Бильроту II [46], из нормальной слизистой оболочки желудка человека и из ткани рака желудка человека [58]. Однако "нормальная" слизистая оболочка в 4 из 5 случаев, описанных в последнем сообщении, была получена при антрэктомии по поводу язвы двенадцатиперстной кишки или желудка. Иммуногистохимические исследования с использованием моноклонального антитела К-21 показало избирательную экспрессию последовательности Galβ3GlcNAc в поверхностных участках слизистой оболочки желудка человека (фовеолярном эпителии) у индивидуумов-несекреторов [59], которая совпадала с локализацией связывания Helicobacter pylori со срезами ткани [8, 9]. Иммуногистохимическое исследование с использованием поликлональных антител, связывающихся с последовательностью Galβ3GlcNAc, показало наличие лактотетраозилцерамида в каемочных клетках тощей и подвздошной кишки человека у индивидуумов-несекреторов, с группой крови OLe(a-b-), а также у одного индивидуума-несекретора, с группой крови OLe(a+b+) [60].
Среди 66 изолятов Helicobacter pylori, проанализированных в данном исследовании, 57 штаммов (86%), как было установлено, экспрессируют лактотетраозилцерамидсвязывающую специфичность, в то время как 9 штаммов были отрицательными. Большое преобладание свойства связывать лактотетраоэилцерамид, которое наблюдалось среди изолятов Helicobacter pylori, показывает, что оно представляет собой преобразованное свойство указанного желудочного патогена и может, таким образом, представлять важный фактор вирулентности.
Биологическая уместность лактотетраозилцерамидсвязывающей специфичности была дополнительно подтверждена связыванием Helicobacter pylori с тетрагликозилцерамидной областью некислотных гликосфинголипидов, выделенных из эпителиальных клеток-мишеней человеческого желудка. С помощью протонной ЯМР-спектроскопии и газовой хроматографии - масс-спектрометрии перметилированных тетрасахаридов, полученных гидролизом церамидгликаназой, было показано, что активная в отношении связывания фракция содержит лактотетраозилцерамид. Активный в отношении связывания лактотетраозилцерамид был выявлен только у одного из семи изученных индивидуумов, что наводит на мысль о том, что, хотя инфекция Helicobacter pylori и связанный с ней хронический гастрит является очень распространенным явлением, но только у малой части инфицированных индивидуумов развивается какое-либо из дальнейших последствий, таких как пептическая язва или аденокарцинома желудка [7]. Теоретическая предпосылка, таким образом, такова: присутствие лактотетраозилцерамида на клетках желудочного эпителия представляет собой один из кофакторов, необходимых для развития тяжелых последствий инфекции, таких как пептическая язвенная болезнь или рак желудка.
Активная в отношении связывания лактотетраозилцерамидная фракция, выделенная из мекония человека, содержала как гидроксильные, так и негидроксильные виды церамидов. Теоретически связывание может, таким образом, ограничиваться видами с гидроксицерамидами, как это описано для лактозилцерамидсвязывающей специфичности [16]. Однако лактотетраозилцерамид, выделенный из вилочковой железы кролика, с церамидом, состоящим исключительно из сфингозина и негидроксильных 16:0 и 24:0 жирных кислот (В.Lanne et al., готовится к печати), был также активен, как лактотетраозилцерамид из мекония человека (не показано), что показывает, что связывание с лактотетраозилцерамидом не зависит от состава церамида.
Профиль связывания, полученный с 125I-меченными поверхностными бактериальными белками, был идентичен профилю, полученному с 35S-меченными интактными бактериальными клетками, что наводит на мысль о том, что указанные препараты поверхностных белков можно использовать для выделения и изучения свойств адгезинов, связывающихся с углеводами.
Подводя итоги, можно сказать, что адгезия Helicobacter pylori к клеткам слизистой оболочки человеческого желудка, как представляется, является многоэлементной системой, в которой несколько бактериальных адгезинов распознают и связываются с различными рецепторами в ткани-мишени. Указанное исследование идентифицирует еще одно активное в отношении связывания соединение, т.е. лактотетраозилцерамид, выявленное по связыванию с гликосфинголипидами на пластинках для тонкослойной хроматографии. Распределение данного гликосфинголипида ограничено, и до настоящего времени он был обнаружен только в желудочно-кишечном тракте человека. В других человеческих тканях лактотетраозилцерамид замещен фукозой или сиаловой кислотой и, следовательно, при условиях применявшегося анализа не связывался.
Выделение и описание структуры указанного связывающегося с Helicobacter pylori гликосфинголипида, а также идентификация указанного соединения в клетках слизистой оболочки человеческого желудка привели к идентификации эпитопа минимального связывания, а именно Galβ3GlcNAc. Эпитоп Galβ3GalNAc является очень сходным как по структуре, так и по функции, с Galβ3GlcNAc, и они, таким образом, являются практически взаимозаменяемыми.
Настоящее изобретение, таким образом, относится к веществам, связывающимся с Helicobacter pylori, которые включают по меньшей мере одно соединение, имеющее формулу 1:
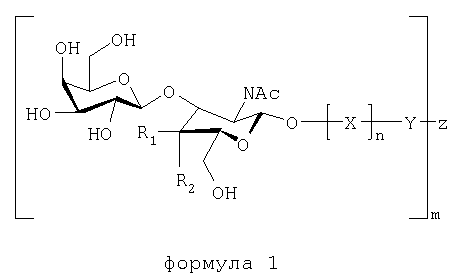
в которой:
R1 и R2 представляют собой Н или ОН, при условии, что когда R1 представляет собой Н, R2 представляет собой ОН, а когда R1 представляет собой ОН, R2 представляет собой Н;
Х представляет собой остаток моносахарида или олигосахарида, и предпочтительно Х представляет собой лактозил-, галактозил-, поли-N-ацетиллактозаминил, или образует часть олигосахаридной последовательности O-гликана или N-гликана;
Y не является ничем, либо представляет собой пространственную группу или терминальный конъюгат, такой как липидная часть церамида или связь (-O-) с Z;
Z представляет собой олиговалентный или многовалентный носитель или -Н;
n равно 0 или 1;
m представляет собой целое число, равное или превышающее 1, и m может представляет собой число до нескольких тысяч или нескольких миллионов, в зависимости от вещества,
или к их аналогам или производным, имеющим такую же или более высокую связывающую активность, как и соединение, имеющее формулу 1, в отношении Helicobacter pylori.
Когда в формуле 1 R1 представляет собой ОН, a R2 представляет собой Н, получают соединение формулы 2, а когда R1 представляет собой Н, а R2 представляет собой ОН, получают соединение формулы 3.


Настоящее изобретение также включает вещества, соответствующие формулам 1, 2 и 3, в которых -O-Х заменена на -S-X, -N-X или -С-Х, поскольку специалистам известно, что они являются взаимозаменяемыми.
Настоящее изобретение относится также к веществам, связывающимся с Helicobacter pylori, которые включают или состоят из Galβ3GlcNAc (соответствует формуле 1, в которой R1=ОН, a R2=Н) или Galβ3GalNAc (соответствует формуле 1, в которой R1=Н, а R2=ОН), или их аналогам или производным, имеющим такую же или более высокую связывающую активность, как и Galβ3GlcNAc или Galβ3GalNAc, в отношении Helicobacter pylori.
Согласно настоящему изобретению возможно использование Galβ3GlcNAc или Galβ3GalNAc per se или любого их натурального или синтезированного аналога или производного, имеющего такую же или более высокую связывающую активность в отношении Helicobacter pylori. Возможно также использование вещества, такого как лактотетраоза, лактотетраозилцерамид (Galβ3GlcNAcβ3Galβ4Glcβ1Cer) или ганглиотетраозилцерамид (Galβ3GalNAcβ4Galβ4Glcβ1Cer), включающего сайт связывания Galβ3GlcNAc или Galβ3GalNAc, или его аналога или производного, имеющего такую же или более высокую связывающую активность в отношении Helicobacter pylori. Может быть предпочтительным расположение указанного эпитопа минимального связывания или его аналога или производного на терминальном, невосстанавливающемся конце указанного вещества.
Может быть предпочтительным использование лактотетраозы или ганглиотетраозы как по одиночке, так и в поливалентной форме.
Вещество, связывающееся с Helicobacter pylori, no настоящему изобретению может также состоять из или включать носитель, к которому присоединено одно или более упомянутых выше веществ.
Вещество, связывающееся с Helicobacter pylori, no настоящему изобретению может также состоять из или включать мицеллу, включающую одно или более упомянутых выше веществ. Примером указанной мицеллы является липосома, содержащая, например, несколько молекул лактотетраозы.
Вещество, связывающееся с Helicobacter pylori, no настоящему изобретению может также быть конъюгировано с полисахаридом, таким как полилактозаминовая цепочка или ее конъюгат, или с антибиотиком, предпочтительно антибиотиком, действующим против Helicobacter pylori.
Вещества по настоящему изобретению могут, таким образом, представлять собой часть сахаридной цепочки или гликоконъюгата или смеси гликосоединений, содержащих другие известные эпитопы связывания с Helicobacter pylori, имеющих различающиеся сахаридные последовательности и конформации, как Lewis b [Fucα2Galβ3(Fucα4)GlcNAc] или NeuNAcα3Galβ4Glc/GlcNAc. Использование сразу нескольких связывающихся веществ может быть благоприятным для лечения.
Вещество по настоящему изобретению может быть конъюгировано с антибиотическим веществом, предпочтительно с антибиотиком пенициллинового ряда. Вещество по настоящему изобретению нацеливает антибиотик на Helicobacter pylori. Указанный конъюгат выгоден для лечения, поскольку для лечения или терапии против Helicobacter pylori требуется меньшее количество антибиотика, в результате чего меньше выражены побочные эффекты антибиотика. Антибиотическая часть конъюгата нацелена на уничтожение или ослабление бактерий, но конъюгат также может оказывать антиадгезионное действие, как описано ниже.
Известно, что Helicobacter pylori может связываться с несколькими видами олигосахаридных последовательностей. Некоторая часть связывания специфичными штаммами может представлять более симбиотические взаимодействия, которые не ведут к развитию злокачественной опухоли или тяжелых состояний. Настоящие данные по связыванию с эпитопами сахаридов ракового типа показывают, что вещество по настоящему изобретению может предотвращать более патологические взаимодействия, оставляя, таким образом некоторые из менее патогенных бактерий/штаммов Helicobacter pylori связанными с другими рецепторными структурами. Таким образом, для предупреждения заболеваний, связанных с Helicobacter pylori, тотальное удаление бактерий может быть необязательным. Менее патогенные бактерии могут даже оказывать пробиотическое действие в предотвращении воздействия более патогенных штаммов Helicobacter pylori.
Понятно также, что Helicobacter pylori содержит на своей поверхности последовательности Galβ3GlcNAc, которые, по меньшей мере, у некоторых штаммов находятся в нефукозилированной форме, которая может быть связана бактерией, как описано в настоящем изобретении. Вещество по настоящему изобретению может также предотвращать связывание между бактериями Helicobacter pylori и таким образом ингибировать бактерии, например, в процессе колонизации.
Вещество, связывающееся с Helicobacter pylori, no настоящему изобретению может представлять собой, например, гликолипид, гликопротеин или неогликопротеин. Оно также может представлять собой олигомерную молекулу, включающую, по меньшей мере, две олисахаридные цепочки.
Для лечения заболевания или состояния, вызванного присутствием Helicobacter pylori в желудочно-кишечном тракте пациента, возможно применение вещества по настоящему изобретению для противодействия адгезии, т.е. для ингибирования связывания Helicobacter pylori с рецепторами эпителия желудка пациента. После введения вещества или фармацевтической композиции по настоящему изобретению они будут конкурировать с рецептором за связыванием с бактериями, и все или некоторая часть бактерий, которые присутствуют в желудочно-кишечном тракте, будут связываться с веществом по настоящему изобретению, а не с рецептором на эпителии желудка. Затем бактерии будут проходить через кишечник и из организма пациента, будучи связанными с веществом по настоящему изобретению, в результате чего воздействие бактерий на организм пациента уменьшится. Предпочтительно используемое вещество представляет собой растворимое соединение, включающее сайт связывания Galβ3GlcNAc или Galβ3GalNAc, такое как растворимый аналог лактотетраозы, лактотетраозилцерамида, ганглиотетраозы или ганглиотетраозилцерамида. Возможно также и часто предпочтительно присоединять вещество по настоящему изобретению к подходящему носителю. В случае использования носителя к одному носителю можно присоединять несколько молекул вещества по настоящему изобретению, что улучшает ингибирующую эффективность.
Согласно настоящему изобретению возможно также лечение других заболеваний, вызванных присутствием Helicobacter pylori, таких как заболевания печени, сердечные заболевания или синдром внезапной младенческой смерти.
Согласно настоящему изобретению возможно инкорпорирование вещества по настоящему изобретению необязательно вместе с носителем в фармацевтическую композицию, подходящую для лечения состояния, вызванного присутствием Helicobacter pylori в желудочно-кишечном тракте пациента, или использовать вещество по настоящему изобретению в способе лечения указанного состояния. Примерами состояний, которые можно лечить по настоящему изобретению, являются хронический поверхностный гастрит, язва двенадцатиперстной кишки, язва желудка и аденокарцинома желудка.
Фармацевтическая композиция по настоящему изобретению может также включать другие вещества, такие как инертный наполнитель или фармацевтически приемлемые вспомогательные вещества, носители, консерванты и т.п., которые хорошо известны специалистам.
Помимо этого вещество по настоящему изобретению можно вводить вместе с другими лекарственными средствами, такими как лекарственные средства, применяемые для лечения заболеваний желудка, включая ингибиторы протонного насоса или лекарственные средства, регулирующие рН в желудке (омепразол, ланзопразол, ранитидин и т.п.) и антибиотики, применяемые против Helicobacter pylori.
Вещество или фармацевтическую композицию по настоящему изобретению можно вводить любым удобным путем, хотя предпочтительно использовать пероральное введение.
Термин «лечение», используемый в настоящем документе, относится как к лечению с целью излечения или облегчения заболевания или состояния, так и к лечению с целью предотвращения развития заболевания или состояния. Лечение может осуществляться как кратковременно, так и продолжительно.
Термин «пациент», используемый в настоящем документе, относится к любому человеку или млекопитающему, которое не является человеком, нуждающемуся в лечении по настоящему изобретению.
Кроме того, возможно применение вещества по настоящему изобретению с целью идентификации одного или более адгезинов путем скрининга для обнаружения последовательностей, которые связываются с веществом по настоящему изобретению. Указанные последовательности могут представлять собой, например, белки или углеводы. Белок, связывающийся с углеводами, может представлять собой лектин или фермент, связывающийся с углеводами. Скрининг можно осуществлять, например, аффинной хроматографией или способами аффинного перекрестного связывания.
Помимо этого возможно применение веществ, которые специфичным образом связываются с Galβ3GlcNAc или Galβ3GalNAc, присутствующими на тканях человека, и, таким образом, предотвращают связывание с Helicobacter pylori. Примеры указанных веществ включают моноклональное антитело К-21, специфичное в отношении Galβ3GlcNAc, и другие антитела или лектины, связывающиеся с указанной структурой, или фермент β-галактозидазу, способный расщеплять галактозы с β3-связью, или лакто-N-биозидазу, фермент эндогликозидазу, - который высвобождает терминальную часть Galβ3GlcNAc из олигосахаридных цепочек. Кроме того, для ингибирования связывания Helicobacter pylori с рецептором Galβ3GlcNAc можно использовать адгезин, связывающий GalβGlcNAc, или особенно его связывающую часть. В случае использования для человека связывающее вещество должно быть удобным для указанного использования, таким как гуманизированное антитело или рекомбинантная гликозидаза человеческого происхождения, т.е. должно быть неиммуногенным и способным отщеплять остаток/остатки терминального моносахарида от веществ по настоящему изобретению. Однако при введении через желудочно-кишечный тракт многие натуральные лектины и гликозидазы, происходящие, например, из пищи, являются толерантными.
Помимо этого возможно применение вещества по настоящему изобретению в качестве матрицы для производства вакцины, подходящей для вакцинации против Helicobacter pylori при упомянутых выше состояниях.
Помимо этого возможно применение вещества по настоящему изобретению для диагностики состояния, вызванного инфекцией Helicobacter pylori.
Помимо этого возможно применение вещества по настоящему изобретению для ингибирования связывания Helicobacter pylori в немедицинских целях, таких как система анализа in vitro, которую, например, можно использовать для идентификации других веществ, связывающихся с Helicobacter pylori.
Помимо этого возможно применение вещества по настоящему изобретению в качестве ведущего соединения для идентификации других веществ, связывающихся с Helicobacter pylori.
Помимо этого возможно применение вещества по настоящему изобретению для типирования Helicobacter pylori.
И, наконец, также возможно применение вещества по настоящему изобретению в пищевых продуктах или питательной смеси как для человека, так и для животных, например, в кормах, молоке, йогурте или других молочных продуктах, напитках и продуктах детского питания. Описанные в настоящем документе питательные смеси или продукты питания не являются натуральным человеческим молоком. Предпочтительно использовать вещество по настоящему изобретению как часть так называемой функциональной или функционализированной пищи. Указанная функционализированная пища оказывает положительное действие на здоровье человека или животного путем ингибирования или предотвращения связывания Helicobacter pylori с клетками или тканями-мишенями. Вещество по настоящему изобретению может являться частью определенной композиции пищи или функциональной пищи. Функциональная пища может содержать другие известные пищевые ингредиенты, разрешенные учреждениями, которые осуществляют контроль за продуктами питания, такими как Управление по продуктам питания и лекарственным средствам США. Вещество по настоящему изобретению может также применяться в качестве пищевой добавки, предпочтительно в качестве добавки в пищу или напитки для производства функциональной пищи или функционального напитка. Пищу или пищевую добавку можно производить также, если коровы или другие животные будут вырабатывать натуральным образом, через свое молоко, вещество по настоящему изобретению в больших количествах. Это можно осуществить, если добиться повышенной экспрессии подходящих гликозилтрансфераз в молоко животного. Можно выбрать и осуществлять селекционную работу конкретной линии или вида домашнего животного на предмет повышенной выработки вещества по настоящему изобретению. Вещество по настоящему изобретению и особенно вещество по настоящему изобретению для питательной смеси или пищевой добавки можно также производить с использованием микроорганизма/микроорганизмов, таких как бактерии или дрожжи.
Особенно полезно, если вещество по настоящему изобретению является частью продуктов питания или питательных смесей для младенцев или детей младшего возраста, предпочтительно частью смесей для младенцев. «Смеси для младенцев» в настоящем документе относятся к специальным составам для кормления младенцев, таким как состав с белковыми гидролизатами, состав для младенцев, родившихся с недостаточным весом, или дополняющий состав. Многих младенцев кормят специальными составами, которые заменяют натуральное человеческое молоко. Указанные составы могут не содержать особых олигосахаридов человеческого молока на основе лактозы, особенно удлиненных, таких как лакто-N-тетраоза, Galβ3GlcNAcβ3Galβ4Glc и его производные. Состав для кормления младенцев может быть сухим порошком, которые нужно разводить водой для получения конечного продукта, используемого для кормления младенца или ребенка младшего возраста. В предпочтительном варианте осуществления настоящего изобретения детское питание для младенца должно содержать сходную с натуральным человеческим молоком концентрацию лакто-N-тетраозы, около 0,05-5 г на литр, более предпочтительно 0,1-0,5 г на литр.
Известно, что лакто-N-неотетраоза и пара-лакто-N-гексаоза Galβ3GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc имеются в человеческом молоке и, таким образом, могут считаться безопасными добавками или ингредиентами смесей для кормления младенцев. Helicobacter pylori является особенно инфективной для младенцев и детей младшего возраста и, учитывая, какие заболевания она может впоследствии вызвать, имеет смысл предотвращать инфекцию. Известно также, что Helicobacter pylori является причиной синдрома внезапной младенческой смерти, но лечение мощными антибиотиками, которое используется для уничтожения данной бактерии, может быть особенно неподходящим для детей младшего возраста или младенцев.
Когда вещество по настоящему изобретению предназначается для диагностики или типирования, его, например, можно включать, например, в зонд или тест-карандаш, необязательно составляющими частями тест-набора. Когда указанный зонд или тест-карандаш приводят в контакт с образцом, содержащим Helicobacter pylori, бактерии связываются с зондом или тест-карандашом и, таким образом, могут быть изъяты из образца и подвергнуты дальнейшему анализу.
Номенклатура гликосфинголипидов следует рекомендациям комиссии по биохимической номенклатуре IUPAC-IUB (CBN for Lipids: Eur. J. Biochem. (1977) 79, 1121, J. Biol. Chem. (1982), 257, 3347-3351, J. Biol. Chem. (1987) 262, 13-18).
Предполагают, что Gal, Glc, ClcNAc, GalNAc, NeuAc и NeuGc имеют D-конфигурацию, Fuc-L-конфигурацию, а все сахара присутствуют в пиранозной форме.
Кроме того, лактотетраоза Galβ3GlcNAcβ3Galβ4Glc известна также как лакто-N-тетраоза.
В краткой номенклатуре жирных кислот и оснований число перед колонкой относится к длине углеводородной цепочки, а число после колонки дает общее количество двойных связей в молекуле. Жирные кислоты с 2-гидроксильной группой обозначены префиксом h перед аббревиатурой, например, h16:0. Для длинноцепочечных оснований d обозначает дигидроксил, а t - тригидроксил. Таким образом, d18:1 обозначает сфингозин(1,3-дигидрокси-2-аминооктадецен), a t18:0 - фитосфингозин(1,3,4-тригидрокси-2-аминооктадецен).
Несмотря на то что в описании, примерах и формуле изобретения упоминается только Helicobacter pylori, другие весьма сходные виды Helicobacter также включены в объем настоящего изобретения.
Настоящее изобретение далее иллюстрируется примерами, которые никоим образом не предназначены для ограничения объема настоящего изобретения.
Краткое описание фигур
В примерах, которые приведены ниже, имеются ссылки на прилагаемые фигуры, на которых:
Фиг.1 иллюстрирует связывание 35S-меченной Helicobacter pylori с гликосфинголипидами, разделенными тонкослойной хроматографией. Фиг.1А иллюстрирует гликосфинголипиды, выявленные анизальдегидным реагентом. Фиг.1В и фиг.1С иллюстрируют гликосфинголипиды, выявленные ауторадиографией после связывания меченного радиоактивными изотопами штамма 17875 Helicobacter pylori. Полоса 1 = некислотные гликосфинголипиды эритроцитов человеческой крови группы А, полоса 2 = некислотные гликосфинголипиды тонкого кишечника собаки, полоса 3 = некислотные гликосфинголипиды тонкого кишечника морской свинки, полоса 4 = некислотные гликосфинголипиды мышиных фекалий, полоса 5 = некислотные гликосфинголипиды эпителиальных клеток тонкого кишечника черно-белой крысы, полоса 6 = некислотные гликосфинголипиды человеческого мекония, полоса 7 = кислые гликосфинголипиды эритроцитов человеческой крови группы О, полоса 8 = кислые гликосфинголипиды вилочковой железы кролика, полоса 9 = ганглиозиды головного мозга теленка, полоса 10 = кислые гликосфинголипиды из гипернефромы человека. Обозначения слева от (А) указывают число углеводных остатков в полосах.
Фиг.2 иллюстрирует масс-спектр перметилированного гликосфинголипида, связывающегося с Helicobacter pylori, который был выделен из человеческого мекония. Выше спектра находится упрощенная формула, представляющая виды церамидов с сфингозином и гидроксильной 24:0 жирной кислотой.
Фиг.3 иллюстрирует аномерную область протонной ЯМР-спектрограммы гликосфинголипида из человеческого мекония. Было снято 4000 сканирований при температуре датчика 30°С. Большой дисперсионноподобный сигнал на 5,04 м.д. является инструментальным артефактом, и имеется также неидентифицированная примесь на 4,93 м.д.
Фиг.4 иллюстрирует связывание Helicobacter pylori с чистыми гликосфинголипидами, разделенными на пластинках для тонкослойной хроматографии. Полоса 1 = лактотриаозилцерамид, полоса 2 = лактотетраозилцерамид, полоса 3 = гликосфинголипид Н5 типа 1, полоса 4 = гликосфинголипид Lea-5,
полоса 5 = гликосфинголипид Leb-6,
полоса 6 = гликосфинголипид Х-5,
полоса 7 = гликосфинголипид Y-6,
полоса 8 = гликосфинголипид В6 типа 1. Фиг.4А показывает химическое детектирование с помощью анизальдегида, а фиг.4В представляет собой ауторадиограмму, полученную с помощью связывания с 35S-меченной Helicobacter pylori.
Фиг.5 иллюстрирует эффект предварительной инкубации Helicobacter pylori с олигосахаридами лактозой и лактотетраозой. Фиг.5А представляет собой тонкослойную хроматограмму, окрашенную анизальдегидом, фиг.5В показывает связывание Helicobacter pylori, инкубированной с лактозой, а фиг.5С показывает связывание Helicobacter pylori, инкубированной с лактотетраозой.
Полоса 1 = ганглиотетраозилцерамид,
полоса 2 = лактотетраозилцерамид,
полоса 3 = неолактотетраозилцерамид.
Фиг.6 иллюстрирует тонкослойную хроматограмму разделенных гликосфинголипидов, выявленных с помощью анизальдегида (фиг.6А), и ауторадиограмму, полученную с помощью связывания с 35S-меченным штаммом 002 Helicobacter pylori (фиг.6В). Полоса 1 = лактотетраозилцерамид человеческого мекония, полоса 2 = некислотные гликосфинголипиды человеческого мекония, полоса 3 = некислотные гликосфинголипиды желудка человека с группой крови A(Rh+)p, полоса 4 = некислотные гликосфинголипиды желудка человека с группой крови A(Rh+)P. Количество углеводных остатков в полосах указано обозначениями слева.
Фиг.7 иллюстрирует связывание Helicobacter pylori с некислотными гликосфинголипидами из эпителиальной клетки человеческого желудка. Полоса 1 = стандарт некислотных гликосфинголипидов тонкого кишечника собаки, полоса 2 = стандарт некислотных гликосфинголипидов мышиных фекалий, полоса 3 = стандарт некислотных гликосфинголипидов человеческого мекония, полосы 4-8 = некислотные гликосфинголипиды (80 мкг на полосу) эпителиальной клетки человеческого желудка от пяти индивидуумов (случаи 1-5 в таблице III). Фиг.7А иллюстрирует химическое обнаружение с помощью анизальдегида, а фиг.7В представляет собой ауторадиограмму, полученную путем связывания 35S-меченной Helicobacter pylori. Количество углеводных остатков в полосах указаны обозначениями слева.
Фиг.8 представляет собой тонкослойную хроматограмму, показывающую тетрагликозилцерамидсодержащие фракции, полученные из эпителиальных клеток желудка случая 4 и 5 в таблице III (А), и аномерные области протонных ЯМР-спектрограмм при 500 МГц фракции 4-II (В) и 5-II (С). Полоса 1 = общие некислотные гликосфинголипиды желудочного эпителия случая А, полоса 2 = фракция 4-I из случая 4, полоса 3 = фракция 4-II из случая 4, полоса 4 = общие некислотные гликосфинголипиды желудочного эпителия случая 5, полоса 5 = фракция 5-I из случая 5, полоса 6 = фракция 5-II из случая 5. Количество углеводных остатков в полосах указано обозначениями слева.
Фиг.9 показывает реконструированные ионные хроматограммы перметилированных олигосахаридов, высвобожденных с помощью церамидгликаназы. Прогон А = стандартная смесь глобозида, лактотетраозилцерамида и лактонеотетраозилцерамида, прогон В = тетрагликозилцерамиды из желудочного эпителия случая 4 в таблице III, прогон С = тетрагликозилцерамиды из желудочного эпителия случая 5 в таблице III. Олигосахариды стандартной смеси (прогон А) помечены.
Фиг.10 показывает масс-спектрограммы, полученные высокотемпературной газовой хроматографией и EI масс-спектрометрией перметилированных олигосахаридов, высвобожденных с помощью церамидгликаназы из стандартных гликосфинголипидов (I и II), тетрагликозилцерамидной фракции из желудочного эпителия случая 4 в таблице III (III), и тетрагликозилцерамидной фракции из желудочного эпителия случая 5 в таблице III (IV).
Фиг.11 иллюстрирует распознавание лактотетраозилцерамидов с помощью связывающегося с сиаловой кислотой штамма CCUG 17874 Helicobacter pylori (В) и штамма CCUG 17875 Helicobacter pylori, который не имеет способности связываться с сиаловой кислотой (С).
Фиг.12 показывает конформеры минимальной энергии лактотетраозилцерамида, связывающегося с Helicobacter pylori (фиг.12А) и несвязывающегося Lea-5 гликосфинголипида (В), Leb-6 гликосфинголипида (С) и дефукозилированного гликосфинголипида В6 типа 1(D).
Фиг.13 показывает молекулярные модели конформеров минимальной энергии лактотетраозилцерамида и ганглиотетраозилцерамида, показывающие, что концевой дисахарид может быть представлен идентично путем варьирования только двугранных углов Glcβ1Cer. Генерация всех возможных конформаций низкой энергии, имеющих вариантные двугранные углы над связью Glcβ1Cer (Ф, Ψ и θ), для лактотетраозилцерамида и ганглиотетраозилцерамида с последующим попарным сравнением соответствующих конформеров, показала, что были получены две пары, в которых концевой дисахарид имеет одинаковую ориентацию для указанных двух гликосфинголипидов. В первой паре двугранные углы для лактотетраозилцерамида (А) над связью GlcβlCer равны 51, -179 и 67, в то время как для ганглиотетраозилцерамида (В) те же углы равны 51, 180 и 177. Конформация в (А) стабилизирована внутримолекулярной водородной связью между 2-ОН в Glc и 3-О в длинноцепочечном основании, в то время как конформация в (В) была расценена как удлиненная. Во второй паре двугранные углы Glcβ1Cer для лактотетраозилцерамида (С) равны 13, -90 и -59, а для ганглиотетраозилцерамида (D) равны 53, -173 и -64. В случае лактотетраозилцерамида 2-ОН в Glc образует водородную связь с 2-ОН в жирной кислоте и NH в длинноцепочечном основании, в то время как ганглиотетраозилцерамид имеет такую же конформацию Glcβ1Cer, которая была обнаружена в кристаллической структуре Glcβ1Cer. Метильный углерод ацетамидных групп GlcNAc/GalNAc показан черным.
Примеры
В примерах использованы следующие аббревиатуры:
CFU = колониеобразующие единицы;
Hex = гексоза;
HexN = N-ацетилгексозамин;
EI = электронная ионизация.
В данных примерах связывание Helicobacter pylori с гликосфинголипидами изучали с помощью связывания 35S-меченных бактерий с гликосфинголипидами на тонкослойных хроматограммах. Часто выявлялись две отдельные специфичности связывания; с одной стороны, связывание Helicobacter pylori с лактозилцерамидом, ганглиотриаозилцерамидом и ганглиотетраозилцерамидом, а с другой - избирательное связывание с некислотным тетрагликозилцерамидом из человеческого мекония. Последний связывающийся с Helicobacter pylori гликосфинголипид был выделен и на основе масс-спектрометрии, протонной ЯМР-спектроскопии и исследований по разложению идентифицирован как Galβ3GlcNAcβ3Galβ4Glcβ1Cer (лактотетраозилцерамид). Связывание Helicobacter pylori с тетрагликозилцерамидной областью фракции некислотных гликосфинголипидов из эпителиальных клеток желудка было получено для одного из семи человек, и наличие лактотетраозилцерамида в данной фракции было подтверждено протонной ЯМР-спектроскопией и газовой хроматографией - EI масс-спектрометрией перметилированных тетрасахаридов, полученных после воздействия церамидгликаназы. Экспрессия свойства связывать лактотетраозилцерамиды была выявлена в 57 из 66 изолятов Helicobacter pylori (86%).
МАТЕРИАЛЫ И МЕТОДЫ
Штаммы бактерий, условия культивирования и мечение
Использовавшиеся бактерии и их источники описаны в таблице I в конце описательной части. В большинстве экспериментов параллельно использовали четыре штамма, типовой штамм 17875 (полученный из коллекции культур университета Гетеборга, (CCUG), Швеция) и клинические изоляты 002, 032 и 306.
Штаммы хранили при -80°С в триптическом соевом бульоне, содержавшем 15% глицерина (по объему), и сначала выращивали на агаре GAB-CAMP [19] во влажной (98%) микроаэрофильной атмосфере (5-7% О2, 8-10% CO2, 85% N2) при 37°С в течение 48-72 ч. Для мечения колонии инокулировали на GAB-CAMP, или Brucella, чашки с агаром и 50 мкКи [35]S-метионин (Amershain, UK), разведенный в 0,5 мл физиологического раствора с фосфатным буфером (PBS), рН 7,3, разбрызгивали по чашкам. После инкубации в течение 12-36 ч при 37°С в микроаэрофильных условиях клетки соскабливали, промывали три раза PBS и ресуспендировали в PBS до концентрации 1×108 CFU/мл.
Альтернативно, колонии инокулировали (1×105 CFU/мл) в среду Хэма F 12 (Gibco BRL, UK), дополненную 10% термоинактивированной фетальной бычьей сыворотки (Sera-lab, Goteborgs Termometerfabric, Sweden) и 50 мкКи [35]S-метионина. Культуральные флаконы инкубировали при встряхивании и микроаэрофильных условиях при 37°С в течение 24 ч. Аликвоты из культуральных флаконов анализировали на активность уреазы, оксидазы и каталазы и изучали посредством фазово-контрастной микроскопии, чтобы убедиться в низком содержании коккоподобных форм. Бактериальные клетки собирали центрифугированием и после двух промывок с PBS ресуспендировали в PBS до концентрации 1×108 CFU/мл.
В результате обеих процедур мечения получали суспензии со специфической активностью приблизительно 1 имп./мин на 100 организмов Helicobacter pylori.
Экстрагирование бактериальных поверхностных белков
Перед процедурой экстрагирования штаммы Helicobacter pylori (обозначенные знаком* в таблице I) культивировали на агаре с 5% лошадиной крови в микроаэрофильных условиях при 37°С в течение 2-3 дней, собирали и один раз промывали PBS. Сырые экстракты получали инкубированием бактериальных клеток с 1 М LiCl в PBS при 45°С в течение 2 ч [20]. После центрифугирования надосадки диализировали против PBS в течение ночи. Концентрации белков в экстрактах составляли 300-1500 мкг/мл по результатам определения с использованием кислого раствора красителя кумасси бриллиантового голубого G-250 (Bio-Rad, Richmond, UK). Из каждого экстракта брали аликвоты приблизительно 100 мкг белка и метили [125]I способом иодогена [21], до специфической активности 2-5×103 имп./мин на мкг.
Тонкослойная хроматография
Тонкослойную хроматографию осуществляли на пластинах с силикагелем 60 HPTLC со стеклянной или алюминиевой основой (Merck, Darmstadt, Germany) с использованием в качестве систем растворителей смесей хлороформ/метанол/вода (60:35:8 по объему) или хлороформ/метанол/вода с 0,02% CaCl2 (60:40:9 по объему). Химическое обнаружение осуществляли анизальдегидным [22] или резорциноловым реагентом [23].
Анализ связывания на хроматограмм
Анализы связывания на хроматограммах осуществляли, как описано [24]. Смеси гликосфинголипидов (20-80 мкг на полосу) или чистые соединения (1-4 мкг на полосу) разделяли на пластинах с силикагелем 60 HPTLC с алюминиевой основой. Высушенные хроматограммы пропитывали в течение 1 мин диэтиловым эфиром/н-гексаном (1:5 по объему), с содержанием 0,5% (мас./об.) полиизобутилметакрилата (Aldrich Chem. Сотр. Inc., Milwaukee, WI). После высушивания на хроматограммы наносили покрытие, чтобы блокировать участки неспецифичного связывания. Сначала испытывали несколько различных условий покрытия, например, 1% поливинилпирролидон (мас./об.) в PBS (раствор 1), 2% желатин (мас./об.) в PBS (раствор 2), 2% бычий сывороточный альбумин (мас./об.) в PBS (раствор 3), 2% бычий сывороточный альбумин (мас./об.) и 0,1% (мас./об.) Твин 20 в PBS (раствор 4) или 2% бычий сывороточный альбумин (мас./об.) и 0,2% дезоксихолевую кислоту в PBS (раствор 5). Наиболее стабильные результаты были получены с раствором 4, который впоследствии использовали в качестве стандартного условия. Покрытие было завершено в течение 2 ч при комнатной температуре. Затем суспензию 35S-меченных бактерий (разведенных в PBS до 1×10 g CFU/мл и 1-5×106 имп./мин на мл) или 125I-меченных бактериальных поверхностных белков (разведенных в растворе 4 до концентрации приблизительно 2×106 имп./мин на мл) осторожно разбрызгивали по хроматограммам и инкубировали в течение 2 ч при комнатной температуре. После шестикратного промывания PBS и высушивания пластинки для тонкослойной хроматографии ауторадиографировали в течение 3-120 ч при комнатной температуре или при -70°С с использованием рентгеновской пленки XAR-5 (Eastman Kodak, Rochester, NY).
Гликосфинголипидные препараты
Стандартные гликосфинголипиды
Фракции кислых и некислотных гликосфинголипидов из источников, перечисленных в таблице II в конце описательной части, получали с помощью стандартных процедур [25]. Отдельные гликосфинголипиды выделяли ацетилированием общих гликосфинголипидных фракций и повторной хроматографией на колонках. с силикагелем. Идентичность очищенных гликосфинголипидов подтверждали масс-спектрометрией [26], протонной ЯМР-спектроскопией [27-30] и исследованиями по разложению [31, 32].
Galβ3GlcNH2β3Calβ4Glcβ1Cer (№3 в таблице II) получали из Galβ3GlcNAcβ3Galβ4Glcβ1Cer (№2 в таблице II) действием безводного гидразина, как описано в [16].
Человеческий меконий
Образцы мекония от 17 новорожденных доношенных детей, родившихся в родильном доме Sahlgrenska University Hospital, Göteborg, объединяли. Собирали только первую порцию, выделенную в течение 24 ч после рождения, и после лиофилизации хранили при -70°С. Некислотные гликосфинголипиды выделяли из объединенного материала (сухой вес 23,3 г), как описано в [25]. Кратко, лиофилизированный материал экстрагировали в два этапа на аппарате Soxhiet смесями хлороформа и метанола (2:1 и 1:9 по объему соответственно). Объединенные экстракты подвергали мягкому щелочному метанолизу и диализу, после чего разделяли на колонке с кремневой кислотой (Mallinckrodt Chem. Work, St. Louis). Фракции кислых и некислотных гликолипидов получали хроматографией на колонке с DEAE-целлюлозой (DE-23, Whatman). С целью удаления щелочно-стабильных фосфолипидов из некислотных гликолипидов указанную фракцию ацетилировали [24] и разделяли на второй колонке с кремневой кислотой, после чего дезацетилировали и подвергали диализу. После окончательной очистки на колонках с DEAE-целлюлозой и кремневой кислотой получали 262 мг некислотных гликосфинголипидов.
Выделение гликосфинголипидов, связывающихся с Helicobacter pylori, осуществляли в два этапа. Сначала 240-мг фракцию некислотных гликосфинголипидов разделяли с помощью ВЭЖХ на колонке 2,2×30 см с диоксидом кремния (YMC SH-044-10, частицы 10 мкм; Skandinavska Genetec, Kungsbacka, Sweden). Колонку уравновешивали в смеси хлороформ/метанол/вода (65:25:4, по объему) (растворитель А) и элюировали (2 мл/мин) линейными градиентами хлороформ/метанол/ вода (40:40:12, по объему, растворитель В) в растворителе А. Колонку сначала элюировали растворителем А в течение 2 мин, затем процентную долю растворителя В в растворителе А увеличивали от 0 до 50% в течение 5 мин, от 50 до 80% - в течение 140 мин, от 80 до 100% в течение 10 мин, и держали на 100% в течение 23 мин. Аликвоты каждой 2 мл фракции анализировали тонкослойной хроматографией и фракции, положительные по результатам окрашивания анизальдегидом, далее изучали на предмет связывания с Helicobacter pylori с использованием анализа связывания на хроматограммах. Фракции, связывающиеся с Helicobacter pylori, собирали в пробирки 78-88 и после объединения этих фракций получали 14,2 мг.
Указанный материал ацетилировали и далее разделяли с помощью ВЭЖХ на колонке YMC SH-044-10. Колонку, уравновешенную в хлороформе, элюировали со скоростью потока 2 мл/мин линейными градиентами хлороформ/метанол (95:5, по объему) (растворитель С) в хлороформе. Процентную долю растворителя С в хлороформе увеличивали от 0 до 20% в течение 10 мин, от 20 до 100% - в течение 70 мин, и держали на 100% в течение 10 мин. После дезацетилирования аликвоты каждой 1 мл фракции анализировали окрашиванием анизальдегидом на тонкослойных хроматограммах, и фракции, содержавшие гликосфинголипиды, изучали на предмет связывания с Helicobacter pylori. Большую часть гликосфинголипида, связывающегося с Helicobacter pylori, собирали в пробирку 62 и указанную фракцию (2,4 мг) использовали для изучения структуры.
Эпителиальные клетки желудка человека
Ткань желудка (кусочки 10×10 см) получали из области дна от пациентов, подвергавшихся факультативному оперативному вмешательству по поводу патологического ожирения. После промывания 0,9% NaCl (мас./об.) клетки слизистой оболочки осторожно соскабливали и хранили при -70°С. Материал лиофилизировали и выделяли кислые и некислотные гликосфинголипиды, как описано [25]. В двух случаях гликосфинголипиды выделяли также из источников, не являвшихся слизистой оболочкой. Группа крови пациентов, а также количества гликосфинголипидов, выделенных из каждого образца ткани, представлены в таблице III в конце описательной части.
Некислотные гликосфинголипиды из случая 4 в таблице III (2,9 мг) разделяли с помощью ВЭЖХ на колонке с силикагелем 1,0×25 см (Kromasil-Sil, частицы 10 мкм, Skandinavska Genetec) с использованием градиента хлороформ/метанол/вода (от 65:25:4 до 40:40:12, по объему) в течение 180 мин при скорости потока 2 мл/мин. Аликвоты из каждой фракции анализировали тонкослойной хроматографией с использованием анизальдегида в качестве окрашивающего реагента. Тетрагликозилцерамиды собирали в пробирки 12-17. Пробирки 12-14 также содержали соединение с подвижностью в области тригликозилцерамидов на тонкослойных хроматограммах и после объединения данных трех фракций получали 0,2 мг (обозначенная как фракция 4-I). Фракции в пробирках 15-17 объединяли отдельно с получением 0,5 мг тетрагликозилцерамидов (обозначенная как фракция 4-II).
Разделение 10,0 мг фракции некислотных гликосфинголипидов из случая 5 осуществляли с использованием такой же системы, которая описана выше, градиентом, образованным из смеси хлороформ/метанол/вода (от 60:35:8 до 40:40:12 по объему). Фракция, собранная в пробирку 11 (обозначенная как фракция 5-I), содержала тригликоэилцерамиды и тетрагликозилцерамиды (0,1 мг), в то время как в пробирке 12 и 13 получали только тетрагликозилцерамиды. Объединение последних двух фракций дало 0,3 мг (обозначенная как фракция 5-II).
EI масс-спектрометрия
Перед масс-спектрометрией гликосфинголипиды перметилировали с использованием твердого NaOH в диметилсульфоксиде и иодметане, как описано [33]. Тетрагликозилцерамид, выделенный из человеческого мекония, анализировали на масс-спектрометре VG ZAB 2F/I-IF (VG Analytical, Manchester UK), с использованием методики луча [34]. Условия анализа приведены в описании воспроизведенного спектра. Тетрагликозилцерамиды из клеток слизистой оболочки желудка человека анализировали с использованием той же методики на масс-спектрометре JEOL SX102A (JEOL, Tokyo, Japan). Условия анализа были следующими: энергия электрона 70 эВ, ток захвата 300 мкА и напряжение ускорения 10 кВ. Температуру повышали на 15°С в мин, начиная со 150°С.
Исследования по разложению
Перметилированный гликосфинголипид из человеческого мекония подвергали гидролизу, восстанавливали и ацетилировали [31, 32], и полученные частично метилированные ацетаты альдитол- и гексозаминитолов анализировали посредством газовой хроматографии - EI масс-спектрометрии на четырехполюсном масс-спектрометре Trio-2 (VG Masslab, Altrincham, UK). Газовый хроматограф Hewlett Packard 5890A был оборудован колоночным инжектором и 15 м×0,25 мм капиллярной колонкой с диоксидом кремния, DB-5 (J&W Scientific, Ranco Cordova, CA), с толщиной пленки 0,25 мкм. Образцы инъецировали в колонки при 70°С (1 мин) и температуру нагревателя повышали с 70 до 170°С со скоростью 50°С/мин, и с 170 до 260°С со скоростью 8°С/мин. Условия масс-спектрометрии были следующими: энергия электрона 40 эВ, ток захвата 200 мкА. Компоненты идентифицировали сравнением времени удерживания и масс-спектров частично метилированных ацетатов альдитола, полученных из стандартных гликосфинголипидов.
Протонная ЯМР-спектроскопия
Протонные ЯМР-спектры получали при 7,05 Т (300 МГц) на Varian VXR 300 (Varian, Palo Alto, CA) и при 11,75 Т (500 МГц) на JEOL Alpha-500 (JEOL, Tokyo, Japan). Данные обрабатывали в автономном режиме с использованием ЯМР1 (NMRi, Syracuse, NY). Дейтерийобменные гликосфинголипидные фракции растворяли в диметилсульфоксиде-d6/D2O (98:2, по объему) и записывали спектры при 30°С с цифровым разрешением 0,4 Гц. Химические сдвиги даны относительно тетраметилсилана.
Ингибирование растворимыми олигосахаридами
В качестве теста на возможное ингибирование связывания растворимыми сахарами 35S-меченные штаммы Helicobacter pylori 002 и 032 инкубировали при комнатной температуре в течение 1 ч с различными концентрациями (0,05, 0,1 и 0,2 мг/мл) лактотетраозы (Accurate Chem. & Sci. Corp., Westbury, NY) или лактозы (J.Т. Baker Chem. Co., Phillipsburg, NJ) в PBS. Затем проводили анализ связывания на хроматограммах, как описано выше, с использованием хроматограмм с разделенными ганглиотетраозилцерамидом, лактотетраозилцерамидом и стандартными гликосфинголипидами.
Молекулярное моделирование
Конформации различных гликосфинголипидов минимальной энергии, перечисленные в таблице II, были рассчитаны по программе молекулярного моделирования Biograf (Molecular Simulations Inc., Waltham, MA) с использованием силового поля Deriding-II (35) на терминале Silicon Graphics4D/3STG. Заряды генерировали с использованием методики уравновешивания зарядов [36], а для кулоновских взаимодействий использовали зависящую от расстояния диэлектрическую постоянную ε=3,5 г. Помимо этого, использовали специальное условие связывания водорода, при котором Dhb было установлено как -4 ккал/моль [35].
Действие, на тетрагликозилцерамиды из эпителия желудка человека церамидгликаназы
Для ферментативного гидролиза применяли методику Hansson et al. [37]. Кратко, 100 мкг фракции 4-II из случая 4, фракции 5-II из случая 5, стандартного глобозида из эритроцитов человека [38], стандартного лактотетраозилцерамида из человеческого мекония и стандартного лактонеотетраозилцерамида (полученного действием сиалидазы на сиалиллактонеотетраозилцерамид из эритроцитов человека; ссылка [39]) растворяли в 100 мкл 0,05 М буфера ацетата натрия, рН 5,0, содержавшего 120 мкг холата натрия, и подвергали кратковременному воздействию ультразвука. Затем добавляли 1 мЕд церамидгликаназы из пиявок Macrobdella decora (Boehringer Mannheim, Mannheim, Germany) и смеси инкубировали при 37°С в течение 24 ч. Реакцию останавливали добавлением смеси хлороформ/метанол/вода до окончательной пропорции 8:4:3 (по объему). Содержавшую олигосахариды верхнюю фазу, полученную таким способом, отделяли от церамидов и детергента на картридже Sep-Pak C18 (Waters, Milford, MA). Элюат, содержавший олигосахариды, высушивали в атмосфере азота и в условиях вакуума, а затем перметилировали, как описано [33].
Высокотемпературная газовая хроматография и газовая хроматография - EI масс-спектрометрия перметилированных олигосахаридов
Условия анализа были практически такими же, как описано в [40]. Капиллярную газовую хроматографию выполняли на газовом хроматографе.Hewlett-Packard 5890A с использованием колонки плавленной двуокиси кремния (10 м×0,25 мм (внутренний диаметр)), покрытой 0,03 мкм PS с поперечными связями 264 (Fluka, Buchs, Switzerland) и с водородом в качестве газа-носителя. Перметилированные олигосахариды растворяли в этилацетате и 1-мкл образец инъецировали в колонку при 70°С (1 мин). Использовали двухстадийную температурную программу: от 70 до 200°С со скоростью 50°С/мин, затем - со скоростью 10°С/мин до 350°С.
Газовую хроматографию - EI масс-спектрометрию выполняли на газовом хроматографе Hewlett-Packard 5890-II с присоединенным масс-спектрометром JEOL SX-102A. Условия хроматографии, а также капиллярная колонка были такими же, как для анализов с помощью газовой хроматографии, а условия масс-спектрометрии были следующими: температура поверхности 350°С, температура источника ионов 330°С, энергия электрона 70 эВ, ток захвата 300 мкА, напряжение ускорения 10 кВ, пределы сканируемой массы 100-1600, общее время цикла 1,4 сек, разрешение 1000 и давление на участке источника ионов 10-5 Па.
РЕЗУЛЬТАТЫ
Связывание со смесями стандартных гликосфинголипидов
Ряд хорошо охарактеризованных гликосфинголипидных смесей, представляющих большое разнообразие углеводных последовательностей, был разделен посредством тонкослойной хроматографии.
Результаты представлены на фиг.1, которая показывает связывание 35S-меченной Helicobacter pylori или 125I-меченных бактериальных поверхностных белков с гликосфинголипидами, разделенными посредством тонкослойной хроматографии. Фиг.1А иллюстрирует гликосфинголипиды, выявленные анизальдегидным реагентом. Посредством ауторадиографии после связывания меченного радиоактивными изотопами штамма 17875 Helicobacter pylori визуализировалось только небольшое количество избирательных полос, как показано на фиг.1В и фиг.1С. Такой же профиль связывания получали с меченными радиоактивными изотопами белками бактериальной поверхности (не показано). Гликосфинголипиды разделяли на пластинах для HPTLC с силикагелем 60 на алюминиевой основе, используя в качестве системы растворителей смесь хлороформ/метанол/вода (60:35:8, по объему), а анализ связывания выполняли, как описано в разделе «Материалы и методы». Ауторадиограмму на фиг.1В получали после нанесения покрытия на тонкослойную хроматограмму с использованием 2% BSA и 0,1% Твин 20 в PBS, в то время как ауторадиограмму на фиг.1С получали, когда покрывающий буфер содержал только 2% BSA в PBS. Полосы содержали следующие гликосфинголипиды: некислотные гликосфинголипиды эритроцитов человека с группой крови А, 40 мкг (полоса 1); некислотные гликосфинголипиды тонкого кишечника собаки, 40 мкг (полоса 2); некислотные гликосфинголипиды тонкого кишечника морской свинки, 20 мкг (полоса 3); некислотные гликосфинголипиды мышиных фекалий, 20 мкг (полоса 4); некислотные гликосфинголипиды эпителиальных клеток тонкого кишечника черно-белой крысы, 40 мкг (полоса 5); некислотные гликосфинголипиды человеческого мекония, 40 мкг (полоса 6); кислые гликосфинголипиды эритроцитов человека с - группой крови О, 40 мкг (полоса 7); кислые гликосфинголипиды вилочковой железы кролика, 20 мкг (полоса 8); ганглиозиды головного мозга теленка, 40 мкг (полоса 9); кислые гликосфинголипиды из гипернефромы человека, 40 мкг (полоса 10). Ауторадиография продолжалась 12 ч.
Связывание на полосе 2 (лактозилцерамид), полосе 3 (ганглиотриаозилцерамид) и полосе 4 (ганглиотетраозилцерамид) было расценено как соответствующее «лактозилцерамидсвязывающей специфичности» и «ганглиосвязывающей специфичности» Helicobacter pylori, подробно описанных ранее [16].
Кроме того, было выявлено избирательное связывание Helicobacter pylori с компонентом, обладающим подвижностью в тетрагликозилцерамидной области фракции некислотных гликосфинголипидов из человеческого мекония (фиг.1В, полоса 6). Последняя связывающая активность была выявлена, только когда покрывающий буфер содержал детергент (Твин 20 или дезоксихолевую кислоту), как показано на фиг.1. Раствор 4 (2% бычий сывороточный альбумин и 0,1% Твин 20 в PBS) впоследствии использовали в качестве стандарта процедуры нанесения покрытия. Активный в отношении связывания тетрагликозилцерамид из человеческого мекония выделяли посредством ВЭЖХ и охарактеризовывали посредством масс-спектрометрии, протонной ЯМР-спектроскопии и газовой хроматографии - EI масс-спектрометрии после разложения, как указано ниже.
Химическое строение связывающегося с Helicobacter pylori гликосфинголипида из человеческого мекония
Активный в отношении связывания тетрагликозилцерамид выделяли из 240 мг общих некислотных гликосфинголипидов. Посредством ВЭЖХ нативной гликосфинголипидной смеси получали 14,2 мг тетрагликозилцерамидов. Указанная тетрагликозилцерамидная фракция представляла собой сложную смесь, и помимо связывающегося с Helicobacter pylori соединения она содержала по меньшей мере три других гликосфинголипида. Тетрагликозилцерамидную фракцию ацетилировали и в дальнейшем разделяли посредством ВЭЖХ с получением 2,4 мг чистого активного в отношении связывания гликосфинголипида. Каждый этап по время препаративной процедуры контролировали по связыванию меченной радиоактивными изотопами Helicobacter pylori на тонкослойных хроматограммах.
EI масс-спектрометрия
Масс-спектр перметилированного активного в отношении связывания гликосфинголипида из человеческого мекония также изучался совместно с упрощенной формулой для интерпретации, представляющей вид с церамидом d18:1 - h24:0. Результаты представлены на фиг.2. Выше спектра находится упрощенная формула для интерпретации, представляющая вид церамида с сфингозином и гидроксильной 24:0 жирной кислотой. Условия анализа были следующими: масса образца 16 мкг, энергия электрона 45 эВ, ток захвата 500 мкА и напряжение ускорения 8 кВ. Начиная с 250°С, температуру повышали со скоростью 6°С/мин. Воспроизведенный спектр был получен при 300°С.
Спектр перметилированного гликосфинголипида доминировался ионами оксония, что давало углеводную последовательность, и фрагментными ионами из-за индуктивного расщепления церамида. Относительное содержание других фрагментных ионов было весьма низким, но ионы иммония и в случае фитосфингозина в виде ионов длинноцепочечного основания из-за α-расщепленйя основания присутствовали.
Ионы иммония, образованные утратой части длинноцепочечного основания, были выявлены при m/z 1298 и 1326. Указанные ионы дают информацию о количестве и типе сахаров и составе жирных кислот, и в настоящем случае продемонстрировали присутствие одного N-ацетилгексозамина, трех гексоз, комбинированных с h22:0 и h24:0 жирными кислотами.
Ионы углеводной последовательности, наблюдавшиеся при m/z 219 и 187 (219 минус 32), 464, 668 и 872, показали, что указанный гликосфинголипид представлял собой тетрагликозилцерамид с углеводной последовательностью Hex-HexN-Hex-Hex. Это подтверждалось фрагментным ионом при m/z 945 (944+1), который состоял из целой углеводной цепочки и части жирной кислоты. Цепочка типа 1 (Нехβ-3НехМ) была обозначена отсутствием фрагментного иона при m/z 182, который представляет собой доминирующий ион в случае 4-замещенной HexN [41, 42]. Интенсивный фрагментный ион при m/z 228 представлял собой вторичный фрагмент из внутренней HexN, поскольку при m/z 260 концевая HexN не была обнаружена.
Молекулярная область была слабой. Однако ионы [М-Н]+, соответствующие частным случаям с d18:1-24:0, d18:1 - h22:0 и d18:1 - h24:0 церамидами, были обнаружены при m/z 1548, 1550 и 1578 соответственно. Наблюдалась также утрата концевых частей углеводной цепочки молекулярными ионами (объясняется ниже формулы для случая с d18:1 - h24:0 церамидом). Ионы при m/z 1342 и 1370 наблюдались, вероятно, вследствие расщепления между двумя гидроксильными группами t18:0 длинноцепочечного основания, t18:0 - h22:0 и t18:0 - h24:0 церамидных видов соответственно.
Дополнительная информация о церамидном составе была предоставлена сериями фрагментных ионов при m/z 548-722, которая демонстрировала смесь видов, варьировавших от d18:1-16:0 до t18:0-h24:0. Доминирующие церамидные виды представляли собой d18:1-24:0, d18:1 - h24:0, t18:0 - h22:0 и t18:0 - h24:0, как можно было судить по относительным интенсивностям церамидных ионов, ионов иммония и молекулярных ионов.
Таким образом, масс-спектрометрия перметилированного гликосфинголипида показала углеводную цепочку с последовательностью Hex-HexN-Hex-Hex и d18:1 и t18:0 длинноцепочечные основания, комбинированные как с гидроксильными, так и негидроксильными жирными кислотами, содержавшими в основном 22 и 24 атома углерода.
Исследования по разложению
Позиции связывания между углеводными остатками были получены разложением перметилированного тетрагликозилцерамида, т.е. образец подвергали кислотному гидролизу, а затем восстановлению и ацетилированию. Полученные частично метилированные ацетаты альдитола анализировали посредством газовой хроматографии - EI масс-спектрометрии. Хроматограмма реконструированного иона, полученная таким способом, имела четыре углеводных пика (не показано). Ацетат 2,3,4,6-тетраметилгалактитола обозначал концевую галактозу, в то время как присутствие ацетата 4,6-диметил-2Н-метилацетамидглюцитола (3-замещенного N-ацетилглюкозамина) обозначал цепочку типа 1. Два оставшихся пика, ацетаты 2,4,6-триметилгалактитола и 2,3,6-триметилглюцитола, идентифицировали как 3-замещенную галактозу и 4-замещенную глюкозу соответственно.
В сочетании с данными масс-спектрометрии можно, таким образом, вывести углеводную цепочку с последовательностью Gal1-3GlcNac1-3Gal1-4Glc1.
Протонная ЯМР-спектроскопия
Затем осуществляли протонное ЯМР-спектроскопическое исследование при 300 МГц гликосфинголипида из человеческого мекония, и его результаты представлены на фиг.3. Было получено 4000 сканирований при температуре датчика 30°С. Большой дисперсионноподобный сигнал на 5,04 м.д. является инструментальным артефактом. Имеется также неидентифицированная примесь на 4,93 м.д.
Аномерная область протонного ЯМР-спектра содержала пять больших 3-дублета (J1,2≈8 Гц). Аномерный протонный сигнал глюкозы (4,20 м.д., J1,2=7,2 Гц) был расщеплен на два сигнала, как это часто случается из-за различий головной группы церамида. На 4,28 м.д. (J1,2=7,2 Гц) Gal появился аномерный протон i4, что указывает на замещение по 3 позиции. Внутренний аномер GlcNacβ наблюдался на 4,79 м.д. (J1,2=8 Гц), а протоны его N-ацетамидметила резонировали на 1,82 м.д. И, наконец, концевая Gal (3 сигнал был зафиксирован на 4,15 м.д. (J1,2=6,6 Гц)) указывала на связь 1-3. Все аномерные химические сдвиги, таким образом, согласовывались с опубликованными результатами для лактотетраозилцерамида [45]. Помимо основного соединения была выявлена небольшая примесь по β-дублетам на 4,67 и 4,47 м.д., как представляется, соответствующая лактоганглиотетраозилцерамидной гибридной структуре, описанной в недифференцированных клетках мышиного лейкоза [44].
После объединения всех данных была установлена структура связывающегося с Helicobacter pylori гликосфинголипида из человеческого мекония - Galβ3GlcNAcβ3Galβ4Glcβ1Cer, т.е. лактотетраозилцерамид, который ранее был идентифицирован из того же источника [45]. Преобладающий вид церамида в настоящем случае (в основном, d18:1-24:0, d18:1 - h24:0, t18:0 - h22:0 и t18:0 - h24:0) отличался от ранее описанного, в котором были обнаружены только гидроксильные жирные кислоты.
Сравнение с изорецепторами на тонкослойных хроматограммах
Ряд чистых гликосфинголипидов, структурно родственных лактотетраозилцерамиду, исследовали на предмет активности связывания с Helicobacter pylori с помощью анализа связывания на хроматограммах. Результаты представлены в таблице II и показаны на фиг.4. Полосы на фиг.4 следующие: GlcNAcβ3Galβ4Glcβ1Cer (лактотриаозилцерамид), 4 мкг (полоса 1); Galβ3GlcNAcβ3Galβ4Glcβ1Cer (лактотетраозилцерамид), 4 мкг (полоса 2); Fucα2Galβ3GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид Н5 типа 1), 4 мкг (полоса 3); Galβ3(Fucα4)GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид Lea-5), 4 мкг (полоса 4); Fucα2Galβ3(Fucα4)GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид Leb-6), 4 мкг (полоса 5); Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид Х-5), 4 мкг (полоса 6); Fucα2Galβ4 (Fucα3) GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид Y-6), 4 мкг (полоса 7); Galα3(Fucα2)Galβ3GlcNAcβ3Galβ4Glcβ1Cer (гликосфинголипид В6 типа 1), 4 мкг (полоса 8).
Фиг.4А показывает химическое обнаружение с помощью анизальдегида, в то время как фиг.4В показывает ауторадиограмму, полученную с помощью связывания с 35S-меченным штаммом Helicobacter pylori 032. Гликосфинголипиды разделяли на пластинах для HPTLC с силикагелем 60 на алюминиевой основе, используя в качестве системы растворителей смесь хлороформ/метанол/вода (60:35:8, по объему), а анализ связывания выполняли, как описано в разделе «Материалы и методы», с использованием 2% BSA и 0,1% Твина 20 в PBS в качестве покрывающего буфера. Ауторадиографию проводили в течение 12 ч.
Единственным активным в отношении связывания гликосфинголипидом был лактотетраозилцерамид (№2), в то время как все изученные замены отменяли связывание. Так, добавление α-фукозы на 2 позиции (№4 в таблице II), α-N-гликолилнейраминовой кислоты (№11) или α-галактозы (№8) на 3 позиции концевой галактозы, или α-фукозы на 4 позиции N-ацетилглюкозамина (№5) были неприемлемы. Не наблюдалось связывания с гликосфинголипидом GlcNAcβ3Galβ4GlcβlCer (№1), что показывает важность части Galβ3GlcNAcβ. Ацетамидная группа на 2 позиции предпоследнего N-ацетилглюкозамина вносила существенный вклад в данное взаимодействие, поскольку удаление этой части (№3) полностью отменяло связывание.
Ингибирование связывания на тонкослойных хроматограммах
Способность растворимых олигосахаридов препятствовать связыванию Helicobacter pylori с гликосфинголипидами на пластинках для тонкослойной хроматографии изучали посредством инкубирования меченного радиоактивными изотопами штамма Helicobacter pylori 17875 со свободной лактотетраозой (0,1 мг/мл) или лактозой (0,2 мг/мл) в PBS в течение 1 ч при комнатной температуре перед анализом связывания на хроматограммах указанных суспензий. Результаты показаны на фиг.5. Фиг.5А показывает тонкослойную хроматограмму, окрашенную анизальдегидом, фиг.5В показывает связывание Helicobacter pylori, инкубированной с лактозой, а фиг.5С показывает связывание Helicobacter pylori, инкубированной с лактотетраозой. Полосы были следующими:
Galβ3GalNAcβ4Galβ4Glcβ1Cer (ганглиотетраозилцерамид), 4 мкг (полоса 1); Galβ3GlcNAcβ3Galβ4Glcβ1Cer (лактотетраозилцерамид), 4 мкг (полоса 2); Galβ4GlcNAcβ3Galβ4Glcβ1Cer (неолактотетраозилцерамид), 4 мкг (полоса 3). Гликосфинголипиды разделяли на пластинах для HPTLC с силикагелем 60 на алюминиевой основе, используя в качестве системы растворителей смесь хлороформ/метанол/вода (60:35:8, по объему), а анализ связывания выполняли, как описано в разделе «Материалы и методы», с использованием 2% BSA и 0,1% Твина 20 в PBS в качестве покрывающего буфера. Ауторадиографию проводили в течение 12 ч.
Так, инкубирование с лактотетраозой (0,1 мг/мл) ингибировало связывание Helicobacter pylori с лактотетраозилцерамидом, в то время как инкубирование с лактозой не имело ингибирующего эффекта.
Связывание Helicobacter pylori с гликосфинголипидами желудка человека
Некислотные гликосфинголипиды цельной стенки желудка человека
С целью изучения экспрессии активных в отношении связывания гликосфинголипидов в ткани-мишени бактерий исследовали связывание Helicobacter pylori с гликосфинголипидами, выделенными из цельной стенки желудка человека, и результаты показаны на фиг.6, на которой представлены тонкослойная хроматограмма разделенных гликосфинголипидов, выявленных анизальдегидом (фиг.6А) и ауторадиограмма, полученная связыванием 35S-меченного штамма Helicobacter pylori 002 (фиг.6В). Полосы были следующие: лактотетраозилцерамид человеческого мекония, 4 мкг (полоса 1); некислотные гликосфинголипиды человеческого мекония, 40 мкг (полоса 2); некислотные гликосфинголипиды желудка человека с группой крови A(Rh+)p, 40 мкг (полоса 3); некислотные гликосфинголипиды желудка человека с группой крови A(Rh+)P, 40 мкг (полоса 4). Гликосфинголипиды разделяли на пластинах для HPTLC с силикагелем 60 на алюминиевой основе, используя в качестве системы растворителей смесь хлороформ/метанол/вода (60:35:8, по объему), а анализ связывания выполняли, как описано в разделе «Материалы и методы». Покрывающий буфер содержал 2% BSA и 0,1% Твина 20 в PBS. Ауторадиографию проводили в течение 5 ч. Число углеводных остатков в полосах указано обозначениями слева.
Над тетрагликозилцерамидной областью указанных некислотных фракций доминировал глобозид (пример на полосе 4 фиг.6А), который, по крайней мере, для тонкого кишечника [46] и толстого кишечника [47] человека, получали из неэпителиальной стромы. Связывание указанных фракций получено не было (пример на фиг.6В, полоса 4). Однако при использовании фракции некислотных гликосфинголипидов, выделенных из желудка человека с группой крови A(Rh+)p [48], у которых отсутствовала реакция на действие галактозилтрансферазы по превращению лактозилцерамида в глоботриаозилцерамид [49] и которые впоследствии были лишены глобозида (фиг.6А, полоса 3), было выявлено связывание Helicobacter pylori в тетрагликозилцерамидной области (фиг.6В, полоса 3). В данном случае ткань получали в результате оперативного вмешательства по поводу пептической язвенной болезни. Поскольку было получено ограниченное количество указанного активного в отношении связывания тетрагликозилцерамида, изучение его химических свойств было невозможно.
Гликосфинголипиды эпителиальных клеток желудка человека. Далее авторы настоящего изобретения изучали связывание Helicobacter pylori с гликосфинголипидами, выделенными из эпителиальных клеток желудка человека. Поскольку ткань пилорического отдела желудка, которая не является неопластической, редко удаляется во время обычных оперативных вмешательств, гликосфинголипиды выделяли из образцов области дна, полученных от пациентов, подвергавшихся оперативному вмешательству по поводу ожирения, хотя указанная область желудка гистологически отличается от пилорической области, где наиболее часто находят Helicobacter pylori [50, 51].
В целом гликосфинголипиды выделяли из соскобов слизистой оболочки от семи пациентов, а в двух случаях - также из остатков тканей, не являвшихся слизистой оболочкой. Поскольку количество указанного материала было ограничено, связывание указанных фракций изучалось только на штаммах Helicobacter pylori 002 и 032.
Наибольшая часть соединений во фракциях кислых гликосфинголипидов мигрировала на тонкослойных хроматограммах как сульфатид и GM3. Связывание Helicobacter pylori с указанными главными кислыми гликосфинголипидами получено не было (не показано). Связывания бактерий с гликосфинголипидами из неэпителиальной стромы не наблюдалось.
Затем изучали связывание Helicobacter pylori с фракциями некислотных гликосфинголипидов, выделенных из эпителиальных клеток желудка человека, от пяти из семи случаев, и результаты представлены на фиг.7А, которая иллюстрирует химическое выявление анизальдегидом. У одного из семи индивидуумов было выявлено связывание Helicobacter pylori в тетрагликозилцерамидной области, как показано на фиг.7В. Полосы 1-3 на данной фигуре представляют собой стандартные некислотные гликосфинголипиды тонкого кишечника собаки, 40 мкг (полоса 1); мышиных фекалий, 20 мкг (полоса 2); человеческого мекония, 40 мкг (полоса 3), в то время как полосы 4-8 представляли собой некислотные гликосфинголипиды (80 мкг на полосу) эпителиальной клетки желудка человека от пяти индивидуумов (случаи 1-5 в таблице III). Ауторадиограмма, полученная связыванием 35S-меченного штамма Helicobacter pylori 032 (фиг.7В). Гликосфинголипиды разделяли на пластинах для HPTLC с силикагелем 60 на алюминиевой основе, используя в качестве системы растворителей смесь хлороформ/метанол/вода (60:35:8, по объему), а анализ связывания выполняли, как описано в разделе «Материалы и методы», с использованием 2% BSA и 0,1% Твина 20 в PBS в качестве покрывающего буфера. Ауторадиографию проводили в течение 12 ч. Число углеводных остатков в полосах указано обозначениями слева.
Помимо этого в одном случае было выявлено активное в отношении связывания соединение с подвижностью в дигликозилцерамидной области, как описано ранее [16]. Фракцию, содержавшую активный в отношении связывания тетрагликозилцерамид (случай 4), и одну несвязывающуюся фракцию (случай 5) разделяли посредством ВЭЖХ и выделенные тетрагликозилцерамиды от каждого случая характеризовали посредством 1H-ЯНР-спектроскопии, EI масс-спектрометрии и газовой хроматографии - EI масс-спектрометрии перметилированных тетрасахаридов, полученных гидролизом с использованием церамидгликаназы. Результаты показаны на фиг.8, которая представляет собой тонкослойную хроматограмму, показывающую тетрагликозилцерамидсодержащие фракции, полученные из эпителиальных клеток желудка от случаев 4 и 5 в таблице III (А), и аномерные области протонных ЯМР-спектров при 500 МГц фракций 4-II (В) и 5-II (С). Полосы на тонкослойной хроматограмме были следующие: общие некислотные гликосфинголипиды желудочного эпителия от случая 4, 80 мкг (полоса 1); фракция 4-I от случая 4, 4 мкг (полоса 2); фракция 4-II от случая 4, 4 мкг (полоса 3); общие некислотные гликосфинголипиды желудочного эпителия от случая 5, 80 мкг (полоса 4); фракция 5-I от случая 5, 4 мкг (полоса 5); фракция 5-II от случая 5, 4 мкг (полоса 6). Гликосфинголипиды разделяли на пластинах для HPTLC с силикагелем 60 на стеклянной основе, используя в качестве системы растворителей смесь хлороформ/метанол/вода (60:35:8, по объему), и окрашивали анизальдегидом. Число углеводных остатков в полосах указано обозначениями слева. Для протонной ЯМР-спектроскопии было получено 4000 сканирований от 0,5 мг (4-II) и 0,3 мг (5-II) образца соответственно при температуре датчика 30°С.
Протонная ЯМР-спектроскопия тетрагликозилцерамидных фракций из эпителиальных клеток желудка человека
В протонном ЯМР-спектре фракции 4-II, выделенной от случая 4 (фиг.8В), доминировал глобозид с аномерными сигналами, которые появлялись на 4,81 м.д. (Galα), 4,52 м.д. (GalNAcβ), 4,26 м.д. (Galβ) и 4,20/4,17 м.д. (Glcβ). Однако малый пик на основе сигнала Galα H1 выявил присутствие еще одного гликосфинголипида в данной фракции. Этот сигнал согласовывался с GalNAcβ H1 лактотетраоэилцерамида; другие потенциальные сигналы перекрывались резонансами глобозида. Однако Galα H1 глоботриаозилцерамида также должны иметь очень сходный химический сдвиг. Точные сдвиги варьируют в зависимости от температуры и других факторов. Для решения этого вопроса авторы настоящего изобретения сравнивали стандартные спектры лактотетраозил-, глоботетраозил- и глоботриаозилцерамида, полученные при сходных условиях на 400 МГц. Изготавливали также стандартную смесь лактотетраозилцерамида и глоботетраозилцерамида и пропускали ее через данную процедуру на 500 МГц. Указанные сравнения ясно показали, что сигнал на 4,79 м.д. принадлежит β-аномерному протону из N-ацетилглюкозамина лактотетраозилцерамида. Далее это подтвердилось при анализе более ранней элюированной тетрагликозилцерамидсодержащей фракции (4-I) из случая 4. Здесь было выявлено два неперекрывающихся α-аномерных сигнала с галактозы, один, соответствующий внутреннему Galα H1 глоботетраозилцерамида (4,81 м.д.), и другой, соответствующий концевому Galα H1 глоботриаозилцерамида (4,78 м.д.) (не показаны).
Присутствие лактотетраозилцерамида также должно было способствовать появлению метального сигнала с N-ацетамидоглюкозы [52], отличающегося от N-ацетамидогалактозы глоботриа- и глоботетраозилцерамида. GalNAc метильный сигнал наблюдался на 1,85 м.д., а метильный сигнал GlcNAc в лактотетраозилцерамиде - на 1,82 м.д., что идентично полученным авторами стандартным спектрам и близко соответствует ранее сообщавшимся величинам [53]. По интенсивностям метильных сигналов было установлено, что фракция 4-II содержит приблизительно 5% лактотетраозилцерамида.
Элюированная на ранней стадии тетрагликозилцерамидсодержащая фракция (5-I) из случая 5 содержала как глоботриа-, так и глоботетраозилцерамид, о чем свидетельствуют α-аномерные сигналы на 4,81 и 4,78 м.д. соответственно (не показаны). Элюированная на более поздней стадии тетрагликозилцерамидсодержашая фракция (5-II), показанная на фиг.8С, также содержала β-дублет на 4,65 м.д., соответствующий GlcNAcβ лактонеотетраозилцерамида (53). N-ацетамидоглюкоза указанного гликосфинголипида имела метильный сигнал на 1,82 м.д., что соответствовало более ранним данным по лактонеотетраозилцерамиду [52].
EI масс-спектрометрия тетрагликозилцерамидных фракций из эпителиальных клеток желудка человека
Масс-спектры (не показаны), полученные EI масс-спектрометрией перметилированных производных фракции 4-II и 5-II из случая 4 и 5, соответственно, были очень сходными. В обоих спектрах ионы при m/z 260 и 228 (260 минус 32) были выражены, что показывает концевую HexN, в то время как не было выявлено иона, обозначающего концевую Hex при m/z 219. Концевая HexN-Hex была показана ионом при m/z 464. Фрагментный ион при m/z 945 (944+1), содержавший цельную углеводную цепочку и часть жирной кислоты, показал углеводную последовательность HexN-HexHex-Hex.
По относительным интенсивностям фрагментных ионов из церамидной части, ионов иммония и молекулярных ионов было показано, что преобладающим видом церамида фракции 4-II был d18:1-16:0, d18:1 - h24:0 и d18:1 - h24:1, в то время как фракция 5-II имела, главным образом, d18:1-16:0, d18:1-22:0, d18:1-24:0, d18:1-22:1 и d18:1 - h24:0 церамиды.
Таким образом, посредством масс-спектрометрии было идентифицировано только главное соединение двух образцов, т.е. глобозид, в то время как второстепенные соединения фракций, выявленные экспериментами с протонным ЯМР, распознаны не были. Однако повышенное разрешение, полученное комбинированием хроматографических методов и масс-спектрометрии, позволило идентифицировать указанные второстепенные соединения, как описано в следующей части.
Высокотемпературная газовая хроматография - EI масс-спектрометрия перметилированных тетрасахаридов из эпителиальных клеток желудка человека
Фракцию 4-II из случая 4 и фракцию 5-II из случая 5 подвергали гидролизу церамидгликаназой, а высвобожденные тетрасахариды перметилировали и анализировали посредством газовой хроматографии и газовой хроматографии - EI масс-спектрометрии. Результаты представлены на фиг.9 и 10. Каждый хроматографический пик был размешен в α- и β-конформере.
Фиг.9 показывает реконструированные ионные хроматограммы перметилированных олигосахаридов, высвобожденных церамидгликаназой. Прогон А представлял собой стандартную смесь глобозида, лактотетраозилцерамида и лактонеотетраозилцерамида, в то время как прогон В представлял собой тетрагликозилцерамиды из желудочного эпителия случая 4 в таблице III, а прогон С представлял собой тетрагликозилцерамиды из желудочного эпителия случая 5 в таблице III. Аналитические условия были теми же, которые описаны в разделе «Материалы и методы». Олигосахариды стандартной смеси (прогон А) помечены.
Фиг.10 показывает масс-спектрограммы, полученные высокотемпературной газовой хроматографией и EI масс-спектрометрией перметилированных олигосахаридов, высвобожденных с помощью церамидгликаназы из стандартных гликосфинголипидов (I и II), тетрагликозилцерамидной фракции из желудочного эпителия случая 4 в таблице III (III) и тетрагликозилцерамидной фракции из желудочного эпителия случая 5 в таблице III (IV). Аналитические условия см. в разделе «Материалы и методы». Обозначения прогон А-С относятся к частичным общим ионным хроматограммам, показанным на фиг.10. Формулы для интерпретации показаны вместе со стандартными спектрами.
Тетрасахариды желудочного эпителия связывающегося с Helicobacter pylori случая 4 разлагались на два пика, как показано на фиг.9, прогон В. Доминирующий пик элюировался с тем же временем удерживания, что и сахарид из стандартного глобозида, в то время как второстепенный пик элюировался со временем удерживания сахарида из стандартного лактотетраозилцерамида.
Тетрасахариды желудочного эпителия несвязывающегося случая 5 (фиг.9, прогон С) также разлагались на два пика со временем удерживания главного пика, как у сахарида из стандартного глобозида. Второстепенный пик в данном случае элюировался со временем удерживания сахарида из стандартного лактонеотетраозилцерамида.
Для дальнейшей конкретизации различий между тетрагликозилцерамидными фракциями из связывающегося с Helicobacter pylori случая 4 и несвязывающегося случая 5 были получены масс-спектры перметилированных олигосахаридов (фиг.10).
Спектры доминантных пиков в обоих случаях соответствовали спектрам стандартных глобозидов (не показаны). Однако спектры второстепенных тетрасахаридов связывающегося с Helicobacter pylori случая 4 (фиг.10, III) и несвязывающегося случая 5 (фиг.10, IV) продемонстрировали некоторые различия.
Фрагментные ионы, демонстрирующие концевую углеводную последовательность Hex-HexN-Hex, наблюдались при m/z 187 (219 минус 32), 219, 432 (464 минус 32), 464 и 668 в обоих спектрах. Однако в спектре элюированного на поздней стадии пика случая 5 был выражен фрагментный ион при m/z 182, в то время как данный ион отсутствовал в спектре элюированного на поздней стадии пика случая 4. Фрагментный ион при m/z 182 характерен для углеводных цепочек типа 2, Galβ4GlcNAcβ [41, 42], хотя недавно было показано, что его обнаруживают, только когда температура источника составляет более 280°С [54].
Фрагментный ион при m/z 432 (464 минус 32) также был выражен в спектре сахарида из случая 5, как и в спектре стандартного лактонеотетраозилцерамида (фиг.10, II), указывая на то, что метанол более легко элиминируется из цепочек Galβ4GlcNAcβ, чем из цепочек Galβ3GlcNAcβ, наиболее вероятно, из С2-С3.
Сахарид из случая 4 давал сильный фрагментный ион при m/z 228. Данный ион был также выражен в спектре стандартного лактотетраозилцерамида (фиг.10, I), и, вероятно, происходил из внутреннего GlcNAc, поскольку при m/z 260 иона не было.
В заключение можно сказать, что посредством газовой хроматографии и газовой хроматографии - EI масс-спектрометрии перметилированных олигосахаридов из тетрагликозилцерамидов случая 4 и 5 были подтверждены результаты протонной ЯМР-спектроскопии указанных фракций. Преобладающее соединение обеих фракций было идентифицировано как глоботетраоза, в то время как второстепенные компоненты различались. В случае связывающегося с Helicobacter pylori случая 4 второстепенное соединение идентифицировали как лактотетраозу, в то время как несвязывающийся случай 5 имел неолактотетраозу.
Частота связывания с лактотетраозилцерамидом среди изолятов Helicobacter pylori
Частоту экспрессии свойства связываться с лактотетраозилцерамидом оценивали посредством анализа связывания 66 изолятов Helicobacter pylori, перечисленных в таблице I, с гликосфинголипидами на тонкослойных хроматограммах. Для анализов связывания бактерии выращивали из исходных культур и изучали их на предмет связывания с лактотетраозилцерамидом из человеческого мекония посредством анализа связывания на хроматограммах. На положительное связывание указывал образец, идентичный тому, который наблюдался на полосе 6 фиг.1В. Штаммы, которые не связывались, дважды рекультивировали из хранившейся культуры и повторно изучали посредством анализа связывания на хроматограммах, т.е. несвязывающимися признавались те штаммы, которые не показывали связывания с лактотетраозилцерамидом в трех последовательных анализах. По данным критериям 9 из 66 изученных изолятов (штаммы 15, 65, 176, 198, 239, 269, 271, 272 и ВН000334 в таблице I) представляли собой несвязывающиеся штаммы, в то время как 57 изолятов (86%) экспрессировали свойство связывания с лактотетраозилцерамидом.
ОБСУЖДЕНИЕ
Серологическое типирование с использованием эритроцитов и слюны показало, что группа крови в случае 4 представляла собой несекретор ALe(a+b-), что совпадало с присутствием связывающегося с Helicobacter pylori незамещенного лактотетраозилцерамида в слизистой оболочке желудка указанного индивидуума. Однако с помощью связывания моноклональных антител, направленных против детерминанты Leb, было обнаружено значительное количество гликосфинголипида Leb-G во фракции некислотных гликосфинголипидов, выделенных от данного индивидуума, а также в некислотных фракциях из образцов желудка других людей (не показано). Это показывает, что экспрессия антигенов группы крови Льюиса в слизистой оболочке желудка человека не коррелирует с экспрессией антигенов Льюиса на эритроцитах или в слюне, как было показано ранее для других человеческих тканей, например, для эпителиальной ткани мочевыводящий путей [61, 62] и толстой кишки [47, 63].
Открытие того факта, что указанный индивидуум является несекретором, интересно с точки зрения повышенной частоты распространенности язвы двенадцатиперстной кишки среди несекреторов [64-66]. Недавние исследования [67] показали, что несекреция не связана с повышенной чувствительностью к инфекции Helicobacter pylori. Однако статус секретера может определять результаты колонизации, т.е. повышенную склонность несекреторов к развитию пептической язвенной болезни, может быть, в силу присутствия на эпителиальных клетках желудка указанных индивидуумов лактотетраозилцерамида, связывающегося с Helicobacter pylori.
В условиях эксперимента настоящего изобретения Helicobacter pylori распознавала лактотетраозилцерамид, в то время как связывания с гликосфинголипидами, экспериментально идентифицированными как сульфатид и ганглиозид GM3, в кислых фракциях, выделенных из эпителиальных клеток желудка человека, не было. На связывание Helicobacter pylori с лактотетраозилцерамидом не влияло изменение условий роста, поскольку данное связывание было получено как в случае, когда бактерии выращивали на агаре, так и в случае, когда бактерии выращивали на бульоне. Кроме того, связывание с лактотетраозилцерамидом было выявлено как в случае, когда бактерии- выращивали в течение 12 ч, так и в случае, когда бактерии выращивали в течение 120 ч.
Связывание с лактотетраозилцерамидом было получено также с мутантным штаммом bаbА1А2, в котором ген, кодирующий адгезин, связывающийся с Leb, был инактивирован (данные не показаны; см. [68]). Таким образом, связывание Helicobacter pylori с детерминантой Leb и лактотетраозилцерамидом представляет две отдельные специфичности связывания.
Детерминанта Leb (Fucα2Galβ3(Fucα4)GlcNAcβ) имеет в своей основе дисахаридную единицу типа 1, которая представляет собой концевую часть лактотетраозилцерамида. Взаимодействие Helicobacter pylori с Leb зависело, однако, от фукозы, как минимум от α-фукозы в 2 позиции концевой галактозы, а улучшение взаимодействия за счет замещения α-фукозы в 4 позиции N-ацетилглюкозамина [9]. Напротив, связывание с лактотетраозилцерамидом требовало незамещенной углеводной цепочки, поскольку все изучавшиеся замещения основной последовательности рецептора устраняли взаимодействие.
Фиг.12 показывает молекулярную модель лактотетраозилцерамида минимальной энергии (№2 в таблице II, фиг.12А) в сравнении с Leb-6 гликосфинголипидом (№6 в таблице II, фиг.12С) и двумя другими несвязывающимися соединениями, а именно, гликосфинголипидом Lea-5 (№5 в таблице II, фиг.12В) и дефукозилированным гликосфинголипидом В6 типа 1 (№8 в таблице II, фиг.12D). Верхние схемы показывают те же структуры, вид сверху. Связь GlcβCer показана в продольном виде. Замещения основной структуры лактотетраозилцерамида в (B)-(D) обозначены пунктиром, а метильные углероды фукоз и ацетамидной группы GlcNAc обозначены черным цветом. При попытке различить важные части, составляющие эпитоп связывания лактотетраозилцерамида, было сделано два наблюдения: отсутствие связывания лактотриаозилцерамида (№1) и лактотетраозилцерамида, в котором ацетамидная часть восстановлена до амина (№3), которые показывают, что эпитоп составляет концевой дисахарид Galβ3GlcNAcβ3. Отсутствие связывания последней структуры (№3) также показывает, или что интактная ацетамидная группа является существенной для связывания, или что происходит изменение конформации, поскольку амин больше не может принимать участие во взаимодействиях водородной связи с 2-ОН группой внутренней Galβ4. Возможно также сочетание двух указанных эффектов. Кроме того, удлинение концевой Gal лактотетраозилцерамида с помощью Galα3 (№8) или Fucα2 (№4) или замещение предпоследней GlcNAc с помощью Fucα4 (№5) дают неактивные структуры, что наводит на мысль о том, что главная часть концевого дисахарида Galβ3GlcNAcβ3 непосредственно вовлечена во взаимодействия с адгезином, отвечающим за связывание.
Сообщалось о связывании Н.pylori с гликосфинголипидами с сиаловой кислотой (ганглиозидами) [80]. Однако лактотетраозилцерамид распознается штаммами, не имеющими способности связываться с сиаловой кислотой, такими как, например, штамм CCUG 17875, и штаммами, которые связываются с участием сиаловой кислоты, такие как, например, штамм CCUG 17874 (см. фиг.11). Кроме того, замещение концевой Gal лактотетраозилцерамида α3-связанной сиаловой кислотой устраняло связывание Н.pylori, см. таблицу II, №11. Таким образом, способность Н.pylori связываться с лактотетраозилцерамидом не связана с распознаванием ганглиозида.
В Leb структуре остаток GlcNAcβ3 является недоступным, а предпоследняя Galβ3 - частично недоступной, поскольку они перекрыты двумя фукозами, как следует из вида сверху (в левой части страницы) на фиг.12С. Кроме того, поскольку связывание Helicobacter pylori с Leb ингибировано изоструктурой Ley [9], остаток GlcNAcβ3 в структуре Leb не является существенным для связывания с данным соединением. Синхронизация структур минимальной энергии концевой тетрасахаридной части Leb-6 и Ley-6 показывает, что единственным различием является поворот остатка GlcNAcβ3 приблизительно на 180°, что подтверждает отсутствие необходимости наличия ацетамидной части остатка GlcNAcβ3 (или даже, более вероятно, всего остатка) в структуре Leb, в то время как для лактотетраозилцерамида верно обратное. Можно также заметить, что угол между плоскостью кольца концевой Galβ3 в лактотетраозилцерамиде и соответствующей плоскостью в структуре Leb приближается к 40° из-за скученности, вызванной двумя дополнительными фукозными единицами, что представляет дополнительную причину, по которой данные структуры следует рассматривать как отдельные рецепторы для Helicobacter pylori.
Фиг.11 иллюстрирует распознавание лактотетраозилцерамидов как связывающимся с сиаловой кислотой штаммом Н.pylori CCUG 17874 (В), так и штаммом CCUG 17875, который не имеет способности связываться с сиаловой кислотой (С). Хроматограмма (А) окрашена анизальдегидом. Полоса 1 = глобозид человеческих эритроцитов (GalNAcβ3Galα4Galβ4Glcβ1Cer), полоса 2 = лактотетраозилцерамид человеческого мекония (Galβ3GlcNAcβ3Galβ4Glcβ1Cer), полоса 3 = ганглиоэид GM3 (NeuAcα3Galβ4Glcβ1Cer), полоса 4 = ганглиозид человеческих гранулоцитов. Лактотетраозилцерамид (полоса 2) распознается обоими штаммами, в то время как способность штамма CCUG 17874 связываться с сиаловой кислотой продемонстрирована связыванием с ганглиозидами человеческих гранулоцитов (полоса 4).
Эксперимент по молекулярному моделированию - Перекрестное связывание лактотетраозилцерамидных и ганглиотетраозилцерамидных структур
Выше было показано, что Н.pylori специфичным образом связывается с концевым дисахаридом лактотетраозилцерамида. Это имеет отношение к интерпретации ганглиотетраозилцерамидсвязывающего эпитопа, поскольку две данные структуры, главное отличие между которыми заключается в связи между остатками сахаров два и три, оканчиваются одной и той же дисахаридной последовательностью, если пренебречь различием по позиции четыре GalNAc (GlcNAc). Сочетание с наблюдающимся отсутствием связывания де-N-ацилированных видов ганглиотриаозилцерамида и ганглиотетраозилцерамида, а также с резким повышением аффинности от первого до последнего гликосфинголипида [16] представляет убедительные доводы в пользу случая, при котором концевой дисахарид также ганглиотетраозилцерамида составляет эпитоп связывания. Генерация и попарное сравнение всех возможных конформеров минимальной энергии для указанных лактотетраозилцерамидной и ганглиотетраозилцерамидной структур показывает, что по меньшей мере две пары конформеров имеют идентичную презентацию соответствующих связывающих эпитопов из концевого дисахарида (фиг.13), что наводит на мысль о том, что в связывании двух гликосфинголипидов может принимать участие один и тот же бактериальный адгезин. Различие в конформациях связи Glcβ1Cer подтверждается также тем наблюдением, что Н.pylori связывается с лактотетраозилцерамидом, только когда другие липиды или желчные соли так же, как Твин 20 или дезоксихолат, используются в анализе тонкослойных хроматограмм (фиг.1), в то время как указанные добавления лишь слегка влияют на связывание с ганглиотетраозилцерамидом. Таким образом, в отсутствии других липидов или желчных кислот лактотетраозилцерамид, наиболее вероятно, имеет конформацию связи Glcβ1Cer, которая отличается от конформаций, показанных на фиг.13, в которых эпитоп связывания некорректно представлен для наблюдающегося связывания Н.pylori. Недавно был сделан обзор сходных влияний других липидов на конформацию гликосфинголипидов [73]. Кроме того, прямая демонстрация того, что лактотетраоза способна блокировать связывание Н.pylori с ганглиотетраозилцерамидом (фиг.5), сделанная выше, подтверждает правильность доводов в этом направлении.
Таким образом, можно сделать заключение о том, что связывание Н.pylori с эпитопом Galβ3GalNAcβ4 ганглиотетраозилцерамида на самом деле должно рассматриваться как специфичность в отношении лактотетраозилцерамида с допустимостью 4-замещения внутренней Gal и осевой ориентации на позиции четыре у остатка GlcNAcβ3.
Пример аналога
Тетрасахарид Galβ3GlcNAcβ3Galβ4Glc (Isosep, Tullinge, Sweden) и мальтогептаоза (Sigma, Saint Louis, USA) были подвергнуты восстановительному аминированию 4-гексадециланилином (сокращение HDA, от компании Aldrich, Stockholm, Sweden) цианборогидридом (Halina Miller-Podraza, будет опубликовано позже). Указанные продукты были охарактеризованы масс-спектрометрией и, как было подтверждено, представляли собой Galβ3GlcNAcβ3Galβ4Glc (восст.)-HDA и мальтогептаозу (восст.)-HDA [где «(восст.)» означает структуру аминной связи, образованной восстановительным аминированием с восстанавливающего конца глюкозы сахаридов и аминогруппой гексадециланилина (HDA)]. Соединение Galβ3GlcNAcβ3Galβ4Glc(восст.)-HDA обладает активностью связывания в отношении Helicobacter pylori, подобной активности лактотетраозилцерамидного гликосфинголипида в анализе покрытия ТСХ, описанном выше, в то время как контрольный конъюгат мальтогептаоза(восст.) - HDA был абсолютно неактивным. Указанный пример показывает синтетическое производное последовательности Galβ3GlcNAc. Он показывает также, что трисахарид Galβ3GlcNAcβ3Gal представляет собой структуру, связывающуюся с Helicobacter pylori, и что для связывания глюкоза на восстанавливающем конце не требуется (восстановление разрушает структуру пиранозного кольца восстанавливающего конца Glc).








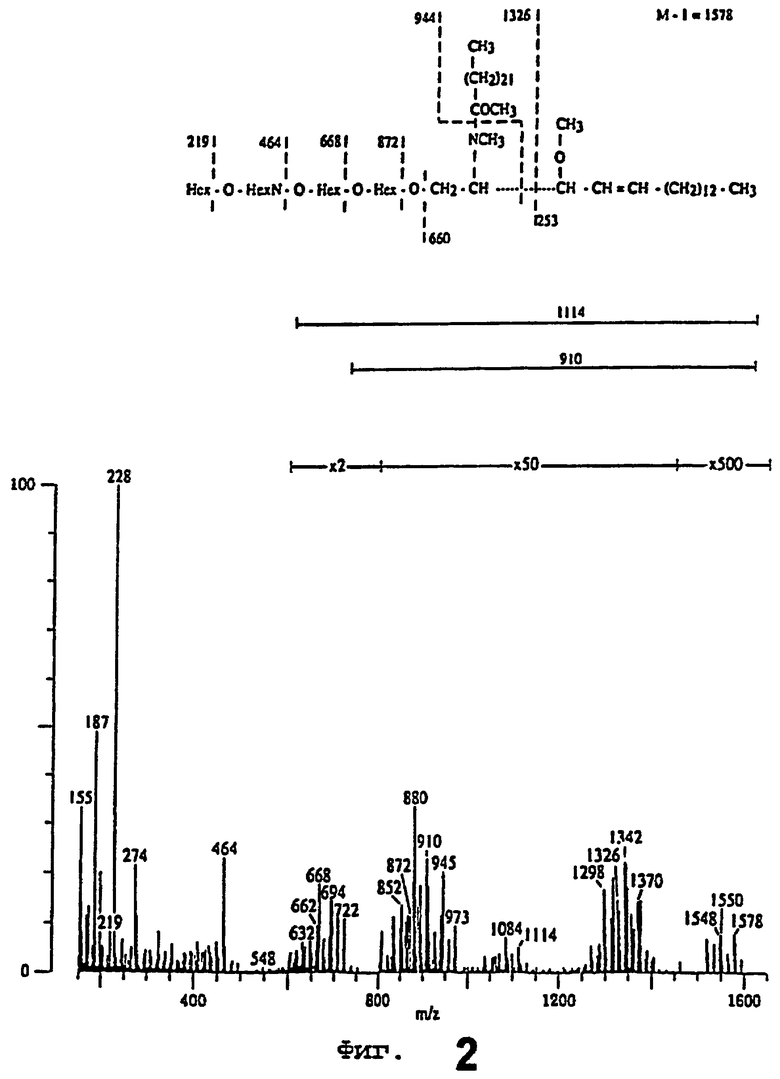
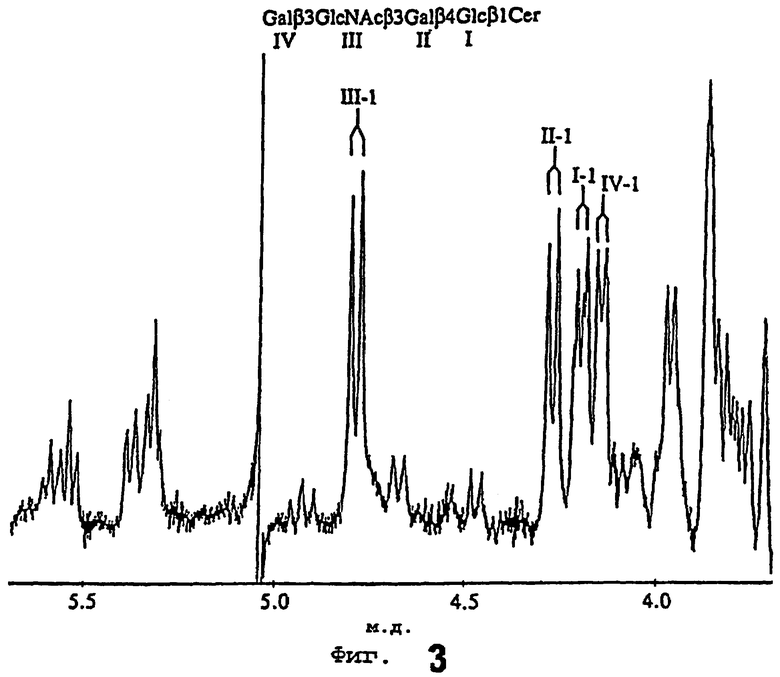
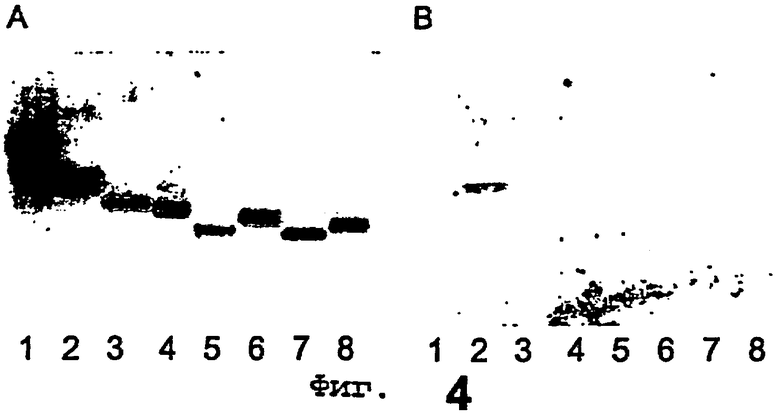
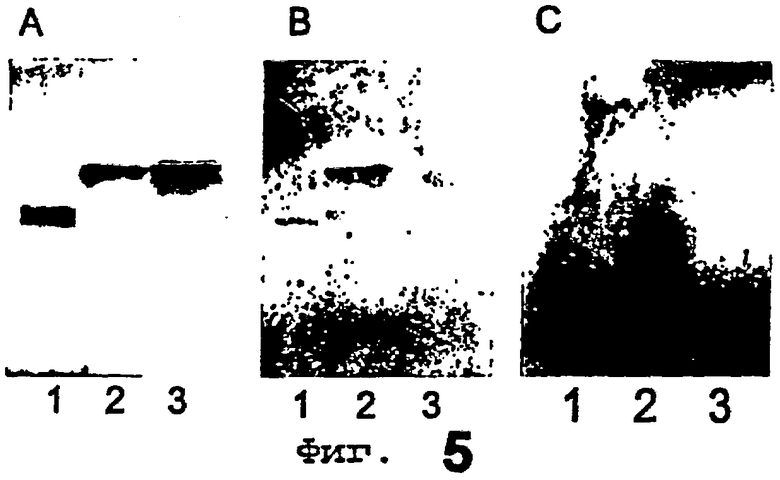

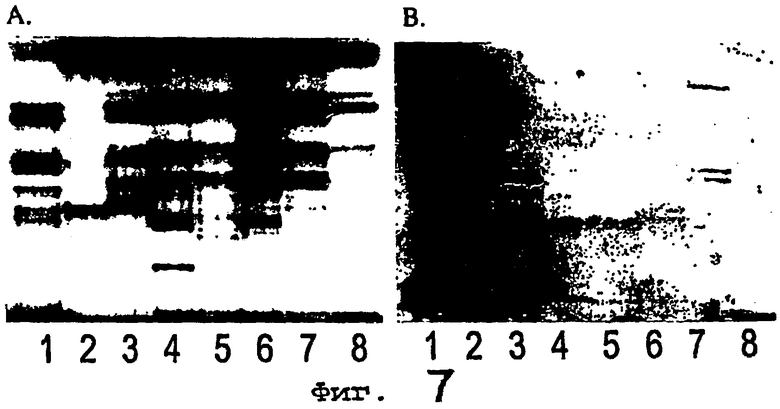




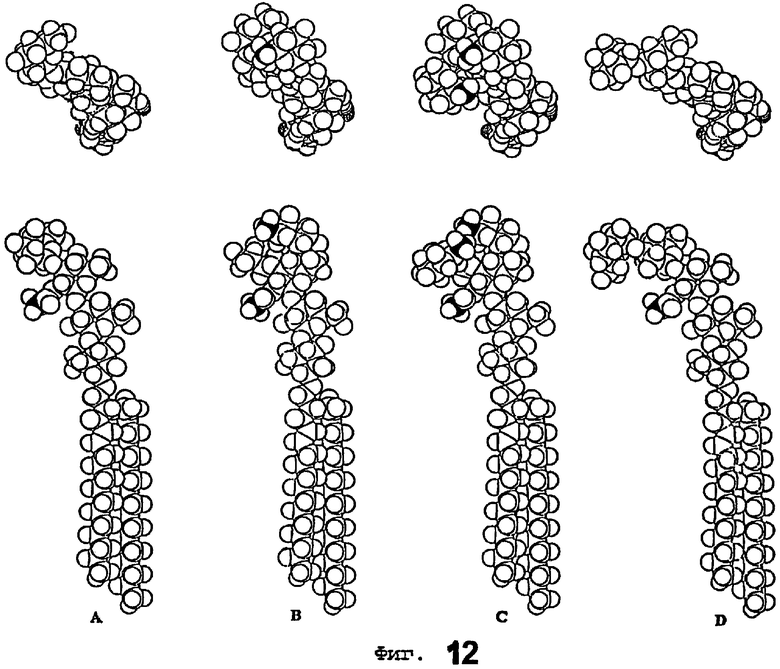

Предложено средство формулы (1) для связывания с Helicobacter pylori ex vivo и вещество на его основе, а также фармацевтическая композиция, пищевая добавка и способ идентификации бактериального адгезина. Вещество состоит из средства формулы (1) и носителя. В частности, средство включает Galβ3GlcNAc. Средство может быть конъюгировано с олигосахаридом или с церамидом. Заявленное средство представляет собой специфически связывающий эпитоп лактотетраозы или лактотетраозилцерамид (Galβ3GlcNAcβ3Galβ-4Glcβ1Cer) с активностью, подобной таковой лактотетраозилцерамидного сфинголипида и может применяться при типировании Helicobacter pylori, для ингибирования связывания Helicobacter pylori, в немедицинских целях, для идентификации бактериального адгезина. 6 н. и 12 з.п. ф-лы, 13 ил., 3 табл.
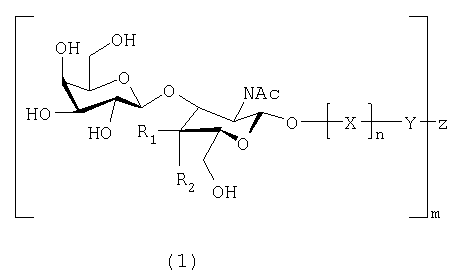

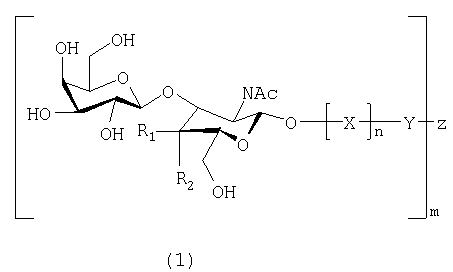
в которой R1 представляет собой Н или ОН;
R2 представляет собой Н или ОН при условии, что когда R1 представляет собой Н, то R2 представляет собой ОН, а когда R1 представляет собой ОН, то R2 представляет собой Н;
Х представляет собой остаток моносахарида или олигосахарида при условии, что когда R2 представляет собой ОН, Х не является лактозой или лактозилом;
Z представляет собой олигосахарид, церамид или -Н;
n равно 0 или 1;
m представляет собой целое число ≥1.
| УСТРОЙСТВО ДЛЯ ХРОМАТОГРАФИЧЕСКОГО АНАЛИЗА ДЛЯ ОБНАРУЖЕНИЯ И/ИЛИ ОПРЕДЕЛЕНИЯ АНАЛИТА В ОБРАЗЦЕ (ВАРИАНТЫ) | 1994 |
|
RU2124729C1 |
| реферат из АБД Medline: Aspinall GO et al | |||
| A structural comparison of lipopolysaccharides from two strains of Helicobacter pylori, of which one strain (442) does and the other strain (471) does not stimulate pepsinogen secretion. | |||

