Предпосылки изобретения
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к области иммунологии и пролиферативным заболеваниям, таким как злокачественная опухоль. В частности, оно относится к композициям и способам их использования, где указанные композиции включают суперантигены, модифицированные для уменьшения серореактивности.
Описание близкого уровня техники
Суперантигены (SAg) представляют собой группу бактериальных и вирусных белков, которые чрезвычайно эффективны для активации большой части Т-клеточной популяции. Суперантигены непосредственно связываются с главным комплексом гистосовместимости (MHC) без процессирования. Фактически, суперантигены связываются с непроцессированной внешней антигенсвязывающей бороздкой молекул MHC класса II, избегая тем самым большой части полиморфизма в обычном пептидсвязывающем сайте. Механизм связывания зависит от суперантигена, связывающегося с Т-клеточным рецептором (TCR) в Vβ-цепи, вместо связывания с гипервариабельными петлями Т-клеточного рецептора (TCR).
Стафилококковые энтеротоксины (SE) представляют собой гомологичную по структуре и функции группу суперантигенов (Papageorgiou и соавт., 2000). Известно, что они являются основной причиной пищевого отравления и токсического шока у людей.
На основе SAg разработан новый способ противоопухолевой терапии для адъювантного лечения солидных опухолей. В нем используются оба основных звена иммунной системы благодаря включению Fab-участка опухолеспецифичного моноклонального антитела и Sag, активирующей Т-клетки, в одном рекомбинантном слитом белке. Fab-SAg-белки, связавшиеся с клетками опухоли, могут инициировать уничтожение клеток данной опухоли цитотоксическими SAg-активированными Т-клетками непосредственно с помощью суперантигенной антителозависимой клеточноопосредованной цитотоксичности, SADCC. Кроме того, активированные Т-клетки продуцируют уничтожающие опухолевые клетки и провоспалительные цитокины, позволяющие преодолеть проблемы, связанные, соответственно, с гетерогенностью злокачественной опухоли и высокомолекулярным поглощением.
Противоопухолевые лекарственные агенты, основанные на суперантигене, оказывали некоторое благоприятное действие, однако одна из клинических проблем, которую следует рассмотреть, заключается в активации системной иммунной системы. Слияние белков с SEA дикого типа изучали при клинических испытаниях пациентов со злокачественной опухолью прямой кишки и поджелудочной железы (Alpaugh и соавт., 1998). Хотя были получены обнадеживающие результаты, но отмечены и ограничения. Во-первых, данный продукт оказался весьма токсичным. Во-вторых, образовавшиеся ранее у пациентов антитела против суперантигенов делают дозирование сложным. Кроме того, данный продукт был иммуногенным. Поэтому повторные циклы терапии возможны лишь для ограниченного числа пациентов.
До появления настоящего изобретения лечение с применением SAg было дозолимитирующим. Настоящее изобретение является первым, в котором модификация суперантигена приводит к снижению серореактивности при сохранении активности суперантигена; таким образом, настоящее изобретение является новым и удовлетворяет требованию неочевидности.
Краткое описание настоящего изобретения
Вышеизложенное довольно широко обрисовывает особенности и технические преимущества настоящего изобретения, чтобы лучше понять нижеследующее подробное описание настоящего изобретения. Дополнительные особенности и преимущества настоящего изобретения описываются ниже, составляют предмет формулы изобретения настоящего изобретения. Специалистам в данной области техники должно быть понятным, что концепцию и раскрытие конкретного варианта можно легко использовать в качестве основы для модификации или создания других конструкций для достижения тех же целей настоящего изобретения. Специалистам в данной области техники должно быть понятным, что такие эквивалентные конструкции не выходят за рамки существа и объема настоящего изобретения, сформулированных в прилагаемой формуле изобретения. Новые признаки, которые, как полагают, характерны для настоящего изобретения, что касается его составления и способа использования, наряду с целями и преимуществами окажутся в большей мере очевидными исходя из нижеследующего описания при рассмотрении его вместе с прилагаемыми чертежами. Вместе с тем, следует ясно понимать, что каждый из рисунков, представленный исключительно с иллюстративной и описательной целью, не следует рассматривать в качестве ограничивающего рамками настоящего изобретения.
В настоящем изобретении создан конъюгат, включающий бактериальный суперантиген и антительный компонент, в котором суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и ДНК-последовательность, кодирующая данный суперантиген, является измененной таким образом, что в А-области не более 15 аминокислотных остатков являются замещенными на другие аминокислоты так, что измененный суперантиген обладает пониженной серореактивностью по сравнению с суперантигеном, из которого он получен; и в котором указанный антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью. Примеры суперантигенов включают, но не ограничиваются указанным, стафилококковый энтеротоксин (SE), экзотоксин Streptococcus pyogenes (SPE), токсин Staphylococcus aureus, вызывающий токсический шоковый синдром (TSST-1), стрептококковый митогенный экзотоксин (SME) и стрептококковый суперантиген (SSA). В конкретных вариантах осуществления настоящего изобретения стафилококковый энтеротоксин представляет собой стафилококковый энтеротоксин А (SEA) или стафилококковый энтеротоксин Е (SEE).
В конкретных вариантах осуществления настоящего изобретения положения заменяемых аминокислотных остатков в области А выбирают из группы, состоящей из позиций 20, 21, 24, 27, 173 и 204. Предполагается также, что область С может включать замены не более 15 аминокислотных остатков. Эти замены могут происходить в позициях аминокислотных остатков 79, 81, 83 и 84. Кроме того, область Е может включать замены не более 15 аминокислотных остатков, в которой замены могут происходить по аминокислотному остатку в положении 227.
В другом варианте осуществления настоящего изобретения создан конъюгат, включающий бактериальный суперантиген и антительный компонент, в котором указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность данного суперантигена является замещенной таким образом, что в области В не более 15 аминокислотных остатков являются замещенными другими аминокислотами так, что замещенный суперантиген обладает пониженной серореактивностью по сравнению с суперантигеном, из которого он получен; и в котором антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированные со злокачественной опухолью. В частности, позиции заменяемых аминокислотных остатков в области В можно выбрать из группы, состоящей из позиций 34, 35, 39, 40, 41, 42, 44, 45 и 49.
В другом варианте осуществления настоящего изобретения создан конъюгат, включающий бактериальный суперантиген и антительный компонент, в котором указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность суперантигена является замещенной таким образом, что в области С не более 15 аминокислотных остатков являются замещенными другими аминокислотами так, что полученный замещенный суперантиген обладает пониженной серореактивностью по сравнению с суперантигеном, из которого он получен; и в котором указанный антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которое нацелено против структуры на поверхности клетки, ассоциированной со злокачественной опухолью. В конкретных вариантах осуществления настоящего изобретения данная злокачественная опухоль выбрана из группы, состоящей из злокачественной опухоли легкого, молочной железы, прямой кишки, почки, поджелудочной железы, яичника, желудка, шейки матки и предстательной железы. Позиции заменяемых аминокислотных остатков в области С выбирают из группы, состоящей из позиций 74, 75, 78, 79, 81, 83 и 84.
Примеры суперантигенов включают, но не ограничиваются указанным, стафилококковый энтеротоксин (SE), экзотоксин Streptococcus pyogenes (SPE), токсин Staphylococcus aureus, вызывающий токсический шоковый синдром (TSST-1), стрептококковый митогенный экзотоксин (SME) и стрептококковый суперантиген (SSA). В конкретных вариантах осуществления настоящего изобретения стафилококковый энтеротоксин представляет собой стафилококковый энтеротоксин А (SEA) или стафилококковый энтеротоксин Е (SEE).
В конкретных вариантах осуществления настоящего изобретения данный конъюгат может дополнительно включать в области А замены не более чем 15 аминокислотных остатков. Замены в области А могут происходить в позициях аминокислотных остатков 20, 21, 24, 27, 173 или 204. Кроме того, данный конъюгат может включать в области Е замены не более чем 15 аминокислотных остатков. В частности, такая замена может происходить в области Е в положении аминокислотного остатка 227.
В дополнительном конкретном варианте осуществления настоящего изобретения данный конъюгат может включать аминокислотную последовательность SEE, включающую замены R20G, N21T, S24G, R27K, K79E, K81E, K83S, K84S и D227S, или аминокислотную последовательность SEE, включающую замены R20G, N21T, S24G, R27K, K79E, K81E, K83S, K84S и D227A. Кроме того, данный конъюгат может включать аминокислотную последовательность SEQ ID NO:2.
В дополнительных вариантах осуществления настоящего изобретения данный конъюгат может включать антительный компонент, например, но не ограничиваясь указанным, Fab-фрагмент. Специфичные Fab-фрагменты могут включать C215Fab или 5T4Fab.
Далее, данный конъюгат может также включать цитокин, такой как интерлейкин. В конкретных вариантах осуществления настоящего изобретения интерлейкин представляет собой IL2 или его производное, обладающее, по существу, такой же биологической активностью, что и нативный IL2.
Другой вариант осуществления настоящего изобретения включает конъюгат, включающий бактериальный суперантиген и антительный компонент, в котором указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность данного суперантигена является замещенной таким образом, что в области D не более 15 аминокислотных остатков являются замещенными другими аминокислотами так, что данный замещенный суперантиген, по сравнению с суперантигеном из которого он получен, обладает сниженной серореактивностью; и в котором данный антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью. Позиции замещаемых аминокислотных остатков в области D выбраны из группы, состоящей из позиции 187, 188, 189 и 190.
В другом варианте осуществления настоящего изобретения создан конъюгат, включающий бактериальный суперантиген и антительный компонент, в котором указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность данного суперантигена является замещенной таким образом, что в области Е не более 15 аминокислот являются замещенными другими аминокислотами так, что указанный замещенный суперантиген обладает сниженной серореактивностью по сравнению с суперантигеном, из которого он получен; и в котором антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью. В конкретных вариантах осуществления настоящего изобретения стафилококковый энтеротоксин представляет собой стафилококковый энтеротоксин А (SEA) или стафилококковый энтеротоксин Е (SEE). Кроме того, замещаемые позиции аминокислотных остатков в области Е выбирают из группы, состоящей из позиций 217, 220, 222, 223, 225 и 227.
В конкретном варианте осуществления настоящего изобретения конъюгат дополнительно включает замены в области А не более чем для 15 аминокислотных остатков. В частности, замены в области А могут происходить по аминокислотным остаткам в позициях 20, 21, 24, 27, 173 и 204.
В другом конкретном варианте осуществления настоящего изобретения конъюгат дополнительно включает замены не более чем 15 аминокислотных остатков в области В, в которой замены могут происходить по аминокислотным остаткам в позициях 34, 35, 39, 40, 41, 42, 44, 45 и 49.
Еще в одном конкретном варианте осуществления настоящего изобретения данный конъюгат может включать замены не более чем 15 аминокислотных остатков в области С. В частности, в области С замены происходят по аминокислотным остаткам в позициях 74, 75, 78, 79, 81, 83 и 84. Кроме того, данный конъюгат может дополнительно включать замены не более чем 15 аминокислотных остатков в области D, в которой замены могут происходить по аминокислотным остаткам в позициях 187, 188, 189 и 190.
В другом конкретном варианте осуществления настоящего изобретения создана фармацевтическая композиция, включающая терапевтически эффективное количество конъюгата, который включает бактериальный суперантиген и антительный компонент, где указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность данного суперантигена является замещенной таким образом, что не более 15 аминокислотных остатков в области С являются замещенными другими аминокислотамитак, что данный замещенный суперантиген обладает сниженной серореактивностью по сравнению с суперантигеном, из которого он получен; и где антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью. В частности, позиции замещаемых аминокислотных остатков в области С выбирают из группы, состоящей из позиции 74, 75, 78, 79, 81, 83 и 84.
В дополнительных вариантах осуществления настоящего изобретения данная фармацевтическая композиция может включать конъюгат, содержащий замены не более чем 15 аминокислотных остатков в области А, в которой замены происходят по аминокислотным остаткам в позициях 20, 21, 24, 27, 173 и 204. Еще в одном дополнительном варианте осуществления настоящего изобретения фармацевтическая композиция может также включать замены не более чем 15 аминокислотных остатков в области Е. В частности, замена в области Е может происходить по аминокислотному остатку в положении 227.
В конкретных вариантах осуществления настоящего изобретения фармацевтическая композиция может содержать конъюгат, включающий аминокислотную последовательность SEE (SEQ ID NO: ID NO:7), а также дополнительные замены R20G, N21T, S24G, R27K, K79E, K81E, K83S, K84S и D227S.
В другом конкретном варианте осуществления настоящего изобретения фармацевтическая композиция может включать аминокислотную последовательность SEE (SEQ ID NO: ID NO:7), а также дополнительные замены R20G, N21T, S24G, R27K, K79E, K81E, K83S, K84S и D227A. Еще в одном варианте осуществления настоящего изобретения фармацевтическая композиция включает конъюгат, который обладает аминокислотной последовательностью SEQ ID NO:1.
В дополнительных конкретных вариантах осуществления настоящего изобретения фармацевтическая композиция включает антительный компонент, например Fab-фрагмент. В частности, Fab-фрагмент представляет собой C215Fab или 5T4Fab. Фармацевтическая композиция может также включать цитокин, такой как интерлейкин. Интерлейкин может представлять собой IL2 или его производное, обладающее по существу той же биологической активностью, что и нативный IL2.
Другой вариант осуществления настоящего изобретения включает способ лечения млекопитающего со злокачественной опухолью путем активации иммунной системы указанного млекопитающего, предусматривающий введение указанному млекопитающему терапевтически эффективного количества конъюгата, который включает бактериальный суперантиген и антительный компонент, где указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность суперантигена является замещенной таким образом, что не более чем 15 аминокислотных остатков в области С являются замененными другими аминокислотами так, что замещенный суперантиген обладает сниженной серореактивностью по сравнению с суперантигеном, из которого он получен; где антительный компонент представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью. Примеры злокачественной опухоли включают, но не ограничиваются указанным, злокачественную опухоль легкого, молочной железы, прямой кишки, почки, поджелудочной железы, яичника, желудка, шейки матки и предстательной железы. В частности, позиции замещаемых аминокислотных остатков в области С выбирают из группы, состоящей из позиций 74, 75, 78, 79, 81, 83 и 84.
В дополнительных вариантах осуществления настоящего изобретения область А может также включать замены не более чем 15 аминокислотных остатков, где замены происходят по аминокислотным остаткам в позициях 20, 21, 24, 27, 173 и 204. Кроме того, область Е может дополнительно включать замены не более чем 15 аминокислотных остатков. В частности, замена в области Е может происходить по аминокислотному остатку в положении 227. Указанный конъюгат может включать аминокислотную последовательность SEE (SEQ ID NO: ID NO:7), а также дополнительные замены R20G, N21T, S24G, R27K, K79E, K81E, K83S, K84S и D227A или замены R20G, N21T, S24G, R27K, K79E, K81E, K84S и D227A. Еще в одном дополнительном варианте осуществления настоящего изобретения конъюгат имеет аминокислотную последовательность SEQ ID NO:1.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Нижеследующие чертежи составляют часть описания настоящего изобретения и включены для дополнительной демонстрации некоторых аспектов настоящего изобретения. Настоящее изобретение можно лучше понять путем обращения к одному или нескольким из этих чертежей в сочетании с подробным описанием представленных здесь конкретных вариантов осуществления настоящего изобретения.
На фиг.1 представлены пептидные фрагменты, распознаваемые антитела человека против SEA, которые идентифицировали в обработанном пепсином SEA/E-18, элюированного с анти-SEA-колонки. Данные фрагменты идентифицировали перед и после выделения очисткой с использованием HPLC в обращенной фазе, совмещенного с масс-спектрометрией (MS). Фрагменты, обнаруживаемые в данном гидролизате, при одном и том же времени удержания до и после выделения очисткой по сродству, рассматривались в качестве позитивных.
На фиг.2 представлены семь разных идентифицированных пептидов, воспроизведенных в виде линий выше аминокислотной последовательности SEA/E-120. Символы, отмеченные светло-серой линией, указывают на остатки, которые были изменены в SEA/E-120, по сравнению с SEA/E-18.
На фиг.3 представлены выравненные структурные последовательности SEA, SED и SHE, использованные в качестве матриц для построения сравнительной компьютерной модели SEA/E-18. Структурные консервативные области отмечены с помощью черных рамок.
На фиг.4 показано множественное выравнивание последовательностей SEA, SEE, SEA/E-18 и SEA/E-120. Участки, показанные в виде линий над выравниванием, соответствуют пяти разным областям А-Е, внутри которых имеют место все замены в SEA/E-120.
На фиг.5 представлена модель (выделенная черным) SEA/E-18, наложенная на SEA (1SXT, выделенная серым).
На фиг.6 представлены области SEA/E-18, которые соответствуют идентифицированным серореактивным пептидам.
На фиг.7 представлен сцинтилляционный анализ близкого расстояния (SPA), который использовали для измерения специфичного связывания меченного 125Iчеловеческого антитела против SEA, связанного с C215FabSEA, C215FabSEA/E-18, -65, -97, -109, -110, -113 или -120, с анти-мышиным F(ab)2, конъюгированным с биотином, на стрептавидиновых PVT-шариках.
На фиг.8А и фиг.8В иллюстрируется способность к опосредованию направленной на опухоль цитотоксичности. На фиг.8А иллюстрируется цитотоксичность, которую измеряют в отношении суперантигена при анализе антителозависимой клеточноопосредованной цитотоксичности, SADCC. На фиг.8В показано, что эффективность суперантигенов опосредовать Т-клеточное уничтожение клеток, экспрессирующих MHC класс II приводит к системной цитотоксичности, которая могла бы обусловить побочные эффекты, измеряемые в анализе суперантигензависимой клеточной цитотоксичности, SDCC. Все новые химеры снижали их действие в SDCC, по меньшей мере, на 1log и максимум на 3log, что касается C215FabSEA/E-120.
На фиг.9 представлена ленточная диаграмма модели SEA/E-120. Боковые цепи остатков G20, T21, G24 и K27 помечены темно-серым цветом, боковые цепи остатков S34, S39, S40, E41, K42, A44, T49, T74, A75, S78, E79, E81, S83 и S84 помечены серым цветом, боковые цепи остатков T217, S220, T222, S223, S225 помечены черным цветом, а боковая цепь остатка S227 помечена светло-серым цветом.
На фиг.10 представлена аминокислотная последовательность 5T4FabSEA/E-120 (SEQ ID NO:1) с вариабельными частями мышиного антитела 5Т4 и константными частями мышиного антитела С242. Позиции 1-458 соответствуют цепи А, а позиции 459-672 соответствуют цепи В.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Специалистам в данной области техники должно быть очевидно, что в настоящем изобретении могут быть осуществлены различные варианты и модификации, не выходящие за рамки существа и объема настоящего изобретения.
Используемое здесь обозначение, "а" или "an", может означать одно или несколько. Используемые здесь в формуле изобретения, в сочетании со словом "включающий (comprising)", слова "а" или "an" могут означать один или более чем один. Используемое здесь слово "другой (another)" может означать, по крайней мере, второй или несколько других.
Используемый здесь термин "антитело" относится к иммуноглобулиновой молекуле, которая способна специфически связываться со специфичным эпитопом на антигене. Подразумевается, что используемое здесь антитело, в широком смысле, относится к любому иммунологическому связывающему агенту, такому как IgG, IgM, IgA, IgD и IgE. Антитела могут представлять собой интактные иммуноглобулины, полученные из природных источников или из рекомбинантных источников, и могут представлять собой иммуноактивные части интактных иммуноглобулинов. Антитела в настоящем изобретении могут существовать в разнообразных формах, включая, например, поликлональные антитела, моноклональные антитела, Fv, Fab и F(ab)2, а также одноцепочечные антитела и гуманизированные антитела (Harlow и соавт., 1988; Bird и соавт., 1988).
Используемый здесь термин "антиген" определен в качестве молекулы, которая вызывает иммунный ответ. Этот иммунный ответ может влечь за собой образование антител, активацию специфичных иммунологически компетентных клеток, либо и то и другое. Антиген можно получить из организмов, белковых/антигенных субъединиц, убитых или инактивированных целых клеток или лизатов. Поэтому квалифицированные специалисты понимают, что любая макромолекула, включая практически все белки, может служить в качестве антигенов. Кроме того, антигены можно получать из рекомбинантной ДНК.
Используемый здесь термин "злокачественная опухоль" определен в качестве пролиферативного заболевания или злокачественного новообразования (опухоли). Примеры включают, но не ограничиваются указанным, злокачественную опухоль молочной железы, злокачественную опухоль предстательной железы, злокачественную опухоль яичника, злокачественную опухоль шейки матки, злокачественную опухоль кожи, злокачественную опухоль поджелудочной железы, злокачественную опухоль прямой кишки и злокачественную опухоль легких.
Используемый здесь термин "конъюгат" определен в качестве слитого белка из суперантигена или варианта суперантигена, слитого или конъюгированного с антителом или с фрагментом антитела.
Используемый здесь термин "иммуногенный" или "иммуногенность" определен в качестве вещества или молекулы, которые вызывает иммунный ответ.
Используемый здесь термин "главный комплекс гистосовместимости", или "MHC", определен в качестве специфичного кластера генов, многие из которых кодируют эволюционно родственные белки клеточной поверхности, участвующие в антигенной презентации, среди которых наиболее существенными являются детерминанты гистосовместимости. MHC класса I, или MHC-I, функционируют, главным образом, в антигенной презентации Т-лимфоцитов CD8. MHC класса II, MHC-II, функционируют, главным образом, в антигенной презентации Т-лимфоцитов CD4.
Используемый здесь термин "серореактивный", "серореакция" или "серореактивность" определен в качестве реакции или действия, обусловленного сывороткой или сыворотками. Специалистам в данной области техники понятно, что сыворотка или сыворотки пациента или животного содержат нейтрализующие антитела или заранее созданные антитела или эндогенные антитела к различным антигенам или молекулам. Таким образом, серореактивность относится к реакции нейтрализующих антител в сыворотке.
Используемый здесь термин "суперантиген" определен в качестве класса молекул, которые стимулируют подгруппу Т-клеток путем связывания с молекулами MHC класса II и Vβ-доменами Т-клеточных рецепторов, стимулируя активацию Т-клеток, экспрессирующих специфические Vβ-сегменты V-гена.
Используемый здесь термин "Т-клеточный рецептор" определен в качестве рецептора, который состоит из соединенного дисульфидной связью гетеродимера с высоковариабельными α- или β-цепями, экспрессируемого в клеточной мембране в виде комплекса с инвариантными CD3-цепями. Т-клетки, несущие этот тип рецептора, часто называют α:β-Т-клетками. Альтернативный рецептор, создаваемый из вариабельных γ- и δ-цепей, экспрессирует CD3 в подгруппе Т-клеток.
Используемый здесь термин "терапевтически эффективный" определен в качестве количества фармацевтической композиции, которая эффективна в лечении заболевания или состояния.
Используемый здесь термин "вариант" или "варианты" относится к белкам или пептидам, которые отличаются, соответственно, от исходного белка или пептида. Варианты в этом смысле описываются ниже и более подробно в другом месте настоящего изобретения. Например, изменения в нуклеотидной последовательности данного варианта могут быть молчащими, т.е., они могут не изменять аминокислоты, кодируемые данной нуклеотидной последовательностью. Если изменения ограничиваются молчащими изменениями этого типа, то вариант будет кодировать пептид с той же аминокислотной последовательностью, что и исходный пептид. Изменения в нуклеотидной последовательности варианта могут изменять аминокислотную последовательность пептида, кодируемого исходной нуклеотидной последовательностью. Такие изменения нуклеиновой кислоты могут приводить к аминокислотным заменам, добавкам, делециям, слияниям и укорочениям в пептиде, кодируемом исходной последовательностью, как указано ниже. Вообще, различия в аминокислотных последовательностях ограничиваются так, что исходная и вариантная последовательности оказываются в итоге очень сходными и идентичными по многим участкам. Аминокислотные последовательности вариантного и исходного пептидов могут отличаться одной или несколькими заменами, добавками, делециями, слияниями и укорочениями, которые могут быть представлены в любом сочетании. Вариант может также представлять собой фрагмент пептида настоящего изобретения, который отличается от исходной пептидной последовательности, являясь более коротким, чем исходная последовательность, например, в результате концевой или внутренней делеции. Другой вариант пептида настоящего изобретения включает также пептид, который сохраняет по существу ту же функцию или активность, что и исходный пептид. Вариант может также представлять собой (i) вариант, в котором один или несколько аминокислотных остатков заменены консервативными или неконсервативными остатками, и такой замененный аминокислотный остаток может или может не быть кодируемым с помощью генетического кода, или (ii) вариант, в котором один или несколько аминокислотных остатков включают замещающую группу, или (iii) вариант, в котором зрелый пептид сливается с другим соединением, таким, например, как соединением, которое увеличивает время полужизни пептида (например, полиэтиленгликоль)) или (iv) вариант, в котором дополнительные аминокислоты сливаются со зрелым пептидом, такие, например, как лидерная или секреторная последовательность или последовательность, которая используется для выделения очисткой зрелого пептида. Варианты могут быть созданы методами мутагенеза, в том числе методами, применяемыми для нуклеиновых кислот, аминокислот, клеток или организмов или могут быть созданы рекомбинантными способами. Предполагается, что все указанные выше варианты такого рода не выходят за рамки, принимаемые специалистами в данной области техники, представленных здесь отличительных особенностей и от данной области техники.
Используемый здесь термин "биологическая активность" относится к внутреннему свойству конкретной молекулы, например, к активации некоторых клеток или к связыванию с некоторыми рецепторами. Используемое здесь определение исходно является скорее качественным, нежели количественным.
Модификация суперантигенов
Настоящее изобретение относится к модификации суперантигенов путем уменьшения их иммуногенности в результате снижения их серореактивности. Специалисты в данной области техники знают, что серореактивность относится к взаимодействию молекул или антигенов с нейтрализующими антителами в сыворотке. В частности, настоящее изобретение относится к конъюгату, включающему бактериальный суперантиген и антительную составляющую, в котором указанный суперантиген представляет собой суперантиген с низким титром, включающий области А-Е, причем область А представляет собой TCR-связывающий сайт, а области В-Е определяют связывание с молекулами MHC класса II; и аминокислотная последовательность указанного суперантигена является замещенной таким образом, что не более 15 аминокислотных остатков в области А-Е являются замещенными отличными аминокислотами так, что замещенный суперантиген обладает сниженной серореактивностью по сравнению с суперантигеном, из которого он получен; и в котором антительная составляющая представляет собой полноразмерное антитело или любой другой связывающий молекулу активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью.
А. Суперантигены
Бактериальные суперантигены, которые рассматриваются для использования в настоящем изобретении, включают, но не ограничиваются указанным, стафилококковый энтеротоксин (SE), экзотоксин Staphylococcus aureus (SPE), токсин Staphylococcus aureus, вызывающий токсический шок (TSST-1), стрептококковый митогенный экзотоксин (SME) и стрептококковый суперантиген (SSA). Специалисты в данной области техники знают, что сведения о трехмерной структуре вышеперечисленных суперантигенов можно получить в Банке Данных Белков (PDB, www.rcsb.org). Кроме того, специалисты в данной области техники могут получить нуклеотидные последовательности и аминокислотные последовательности вышеперечисленных суперантигенов и других суперантигенов из GenBank (http://www.ncbi.nlm.nih.gov/Genbank/GenbankSearch.html).
В конкретных вариантах осуществления настоящего изобретения суперантиген представляет собой суперантиген с низким титром. Специалистам в данной области техники известно и очевидно, что сыворотка человека обычно содержит высокие титры антител против суперантигенов. Например, для стафилококковых суперантигенов соотношение титров дает ряд TSST-1 > SEB > SEC-1 > SEC2 > SEA > SED > SEE. Специалисты в данной области знают, что эти соотношения титров отражают трудности, связанные с иммуногенностью, и трудности, связанные с серореактивностью, или трудности, связанные с нейтрализующими антителами. Поэтому в настоящем изобретении рассматривается использование суперантигена с низким титром, такого как SEA или SEE, чтобы исключить серореактивность парентерально вводимого суперантигена.
Далее, хорошо известно и очевидно, что белковые последовательности и иммунологическая перекрестная реактивность суперантигенов или стафилококковых энтеротоксинов делятся на две родственные группы. Одна группа состоит из SEA, SEE, SED и SHE. Во вторую группу входят SPEA, SEC, SEB и SSA. Поэтому в настоящем изобретении рассматривается также использование суперантигенов с низким титром для уменьшения или исключения из настоящего изобретения перекрестной реактивности антител с высоким титром или эндогенных антител против стафилококковых энтеротоксинов.
В. Варианты суперантигенов
Варианты аминокислотной последовательности суперантигенных белков могут представлять собой замещенные, встроенные или делеционные варианты. Эти варианты можно выделить очисткой известными способами, такими как осаждение (например, сульфатом аммония), ВЭЖХ (HPLC), ионообменная хроматография, аффинная хроматография (в том числе, иммуноаффинная хроматография) или разного вида разделения по размеру (седиментация, гель-электрофорез, гель-фильтрация).
Варианты замещения или замещенные варианты содержат, как правило, замену одной аминокислоты на другую по одному или нескольким сайтам в рамках данного белка. Замены могут быть консервативными, то есть одна аминокислота замещается другой сходной формы и заряда. Консервативные замены хорошо известны в данной области техники и включают, например, следующие замены: аланин на серин; аргинин на лизин; аспарагин на глутамин или гистидин; аспартат на глутамат; цистеин на серин; глутамин на аспарагин; глутамат на аспартат; глицин на пролин; гистидин на аспарагин или глутамин; изолейцин на лейцин или валин; лейцин на валин или изолейцин; лизин на аргинин; метионин на лейцин или изолейцин; фенилаланин на тирозин, лейцин или метионин; серин на треонин; треонин на серин; триптофан на тирозин; тирозин на триптофан или фенилаланин; и валин на изолейцин или лейцин.
Поэтому заявители считают, что в ДНК-последовательностях генов можно осуществить различные изменения без существенной потери биологической ценности или активности данных белков, что и рассматривается ниже. Активность, которая индуцирует Т-клеточный ответ, имеет следствием цитотоксичность для клеток злокачественной опухоли. И еще, сродство суперантигена и молекул MHC класса II уменьшают с минимальными эффектами цитотоксичность суперантигена.
При создании таких изменений рассматривают гидропатический индекс аминокислот. Значимость гидропатического аминокислотного индекса в придании интерактивной биологической функции белку, в целом, в данной области техники ясна (Kyte and Doolittle, 1982). Принято считать, что относительная гидропатическая характеристика аминокислоты определяет вторичную структуру результирующего белка, которая в свою очередь определяет взаимодействие белка с другими молекулами, например, с ферментами, субстратами, рецепторами, ДНК, антителами, антигенами, и т.п.
Каждой аминокислоте приписан гидропатический индекс исходя из характеристик их гидрофобности и заряда (Kyte and Doolittle, 1982): изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серин (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин
(-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9); и аргинин (-4,5).
В данной области техники известно, что определенные аминокислоты могут быть замещены другими аминокислотами, обладающими подобным гидропатическим индексом или показателем, и, тем не менее, получить белок с аналогичной биологической активностью, т.е., получить все же биологически функциональный эквивалентный белок. Для создания таких изменений замещение аминокислот, гидропатические индексы которых находятся в пределах ±2, является предпочтительным, замещение аминокислот, гидропатические индексы которых находятся в пределах ±1, является особенно предпочтительными, а замещение аминокислот, гидропатические индексы которых находятся в пределах ±0,5, является наиболее предпочтительными.
Очевидно также, что замещение подобных аминокислот можно эффективно осуществить исходя из их гидрофильности. В патенте США №4554101, включенном здесь путем ссылки, указывается, что наибольшая локальная средняя гидрофильность белка, которая регулируется гидрофильностью его следующих друг за другом аминокислот, коррелирует с биологической характеристикой данного белка. В патенте США №4554101 подробно изложены номинальные величины гидрофильности следующих аминокислотных остатков: аргинин (+3,0); лизин (3,0); аспартат (+3,0 ± 1); глутамат (+3,0 ± 1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5 ± 1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); триптофан (-3,4).
Очевидно, что аминокислоту можно заменить другой, обладающей сходной величиной гидрофильности и все же получить биологически эквивалентный и иммунологически эквивалентный белок. При таких изменениях, замена аминокислот, величины гидрофильности которых колеблются в пределах ±2, является предпочтительной, замена аминокислот, величины гидрофильности которых колеблются в пределах ±1, является особенно предпочтительной, а в пределах ±0,5 - является наиболее предпочтительной.
С. Слитые белки
Специализированный вид инсерционного варианта представляет собой слитый белок. Его молекула является целой нативной белковой молекулой или ее существенной частью, присоединенной по N- или С-концу к целому второму полипептиду или к его части. Например, слитый белок настоящего изобретения включает присоединение иммунологически активного домена, такого как фрагмент антитела, для нацеливания специфичных клеток опухоли.
Кроме того, включение сайта расщепления по месту слияния, или вблизи него, облегчает удаление постороннего полипептида после выделения очисткой. Другие полезные слияния включают присоединение функциональных доменов, таких как активные сайты ферментов, домены гликозилирования, другие клеточно-мишенирующие сигналы или трансмембранные области.
D. Доменное переключение
Путем замещения гомологичных областей различных белков можно создать ряд интересных вариантов. Для некоторых случаев это известно как "доменное переключение".
Доменное переключение влечет за собойобразование химерных молекул с использованием разных, но, в данном случае, родственных полипептидов. При сравнении разнообразных SAg-белков можно предсказать, насколько функционально значимы области этих молекул. Далее, представляется возможным переключать родственные домены в этих молекулах в попытке определить критичность этих областей для SAg-функции. Данные молекулы могут обладать дополнительной ценностью в том, что эти "химеры" можно отличить от встречающихся в природе молекул, несмотря на возможное приобретение той же функции.
Е. Выделение белков очисткой
Представляется желательным выделить очисткой SAg или его варианты. Методы выделения белков очисткой хорошо известны специалистам в данной области техники. Эти методы включают, на первом уровне, грубое фракционирование клеточного содержимого на пептидные и непептидные фракции. После отделения данного белка от других белков представляющий интерес белок можно подвергнуть дальнейшему выделению очисткой с использованием хроматографических и электрофоретических методов для достижения частичной или полной очистки (или очистки до гомогенности). Аналитические способы, особенно пригодные для получения чистого пептида, представляют собой ионообменную хроматографию, эксклюзионную хроматографию; электрофорез в полиакриламидном геле; изоэлектрофокусирование. Особенно эффективным способом выделения пептидов очисткой является быстрая жидкостная хроматография белков или даже ВЭЖХ.
Некоторые аспекты настоящего изобретения касаются выделения очисткой и, в конкретных вариантах осуществления настоящего изобретения, существенного выделения очисткой кодируемого белка или пептида. Подразумевается, что используемый здесь термин "выделенный очисткой белок или пептид" относится к любой композиции, отделяемой от других компонентов, в которой данный белок или пептид выделен очисткой до любой степени, относительно его естественно получаемого состояния. Поэтому выделенный очисткой белок или пептид относится также к белку или пептиду, свободному от окружающей среды, в которой он возникает естественным образом.
Как правило, термин "выделенный очисткой" относится к белковой или пептидной композиции, подвергнутой фракционированию для удаления различных других компонентов и состав которой практически сохраняет свою выраженную биологическую активность. Если используется термин "по существу очищенный", то это обозначение должно относиться к композиции, в которой данный белок или пептид образует главный компонент данной композиции, составляя, например, около 50%, около 60%, около 70%, около 80%, около 90%, около 95%, или более от белков данной композиции.
Специалистам в данной области техники, в свете настоящего описания, должны быть известны разные способы количественного определения степени выделения очисткой белка или пептида. Они включают, например, определение специфической активности активной фракции, или оценку количества полипептидов во фракции с помощью SDS/PAGE-анализа. Предпочтительный способ оценки степени очистки фракции заключается в вычислении специфической активности фракции, для сравнения ее со специфической активностью первоначального экстракта, и в вычислении, таким образом, степени очистки в ней с помощью "показателя кратности очистки". Понятно, что реальные единицы, используемые для отображения величины активности, должны зависеть от конкретного метода анализа, выбранного для того, чтобы следовать за очисткой и от того, проявляет ли экспрессируемый белок или пептид детектируемую активность.
Специалистам в данной области техники должны быть хорошо известны различные методики, пригодные для использования их в очистке белка. Они включают, например, осаждение с помощью сульфата аммония, PEG, антител и т.п. или путем тепловой денатурации с последующим центрифугированием; хроматографические стадии, такие как ионообменная хроматография, хроматография гель-фильтрацией, хроматография в обращенной фазе, хроматография на гидроксилапатите, и аффинная хроматография; изоэлектрическое фокусирование; гелевый электрофорез; сочетание подобных и иных методов. В данной области техники принято, что порядок осуществления различных стадий выделения очисткой можно менять, или что некоторые стадии можно отбросить и все же иметь возможность подходящим способом получить по существу очищенный белок или пептид.
Известно, что миграция полипептида в разных условиях SDS/PAGE может изменяться, иногда значительно (Capaldi и соавт., 1977). Поэтому следует иметь в виду, что в отличающихся условиях электрофореза средняя молекулярная масса очищенных или частично очищенных экспрессируемых продуктов может меняться.
Высокоэффективная жидкостная хроматография (ВЭЖХ) характеризуется очень быстрым разделением и чрезвычайным разрешением пиков. Это достигается благодаря использованию сверхтонкоизмельченных частиц и высокому давлению для поддержания адекватной скорости потока. Разделение можно осуществить очень быстро, в считанные минуты, или самое большое за 1 час. Кроме того, необходим лишь малый объем образца, поскольку указанные частицы столь малы и столь плотно упакованы, что свободный объем составляет очень маленькую часть чистого объема вещества. Далее, требуется не очень большая концентрация анализируемого образца, потому что хроматографические полосы настолько узкие, что существует очень небольшое разведение данного образца.
Гель-хроматография или "молекулярно-ситовая" хроматография представляет собой специальный вид распределительной хроматографии, которая основана на размере молекул. Теория, лежащая в основе гель-хроматографии, состоит в том, что колонка, которую заполняют мельчайшими частицами инертного вещества, содержащими маленькие поры, отделяет большие молекулы от более маленьких, так как они проходят через или вокруг пор, в зависимости от их размера. Поскольку материал, из которого изготовлены частицы, не адсорбируют данные молекулы, то единственным фактором, определяющим скорость потока, является их размер. Поэтому, молекулы элюируются из данной колонки с уменьшением размера, поскольку их форма остается относительно константной. Гель-хроматография является непревзойденной для разделения молекул разного размера потому, что разделение не зависит от всех других факторов, таких как рН, ионная сила, температура, и т.д. Кроме того, практически отсутствует адсорбция, меньше зональное расширение, а объем элюирования прямо связан с молекулярной массой.
Аффинная хроматография представляет собой хроматографическую процедуру, которая зависит от специфического сродства между веществом, которое выделяют, и молекулой, которая может специфически связываться с ним. Это соответствует взаимодействию типа рецептор-лиганд. Материал колонки синтезируется в результате ковалентного связывания одного из связывающихся партнеров с нерастворимой матрицей. Затем материал колонки способен специфически адсорбировать искомое вещество из данного раствора. Элюция происходит путем изменения условий на условия, при которых не будет происходить связывания (изменение рН, ионной силы, температуры, и т.д.).
F. Мутагенез вариантов
Настоящее изобретение предполагает, что модификация сродства суперантигена к молекулам MHC класса II может уменьшать токсичность данного суперантигена. Следовательно, уменьшенное сродство к молекулам MHC класса II приведет к уменьшению серореактивности или уменьшению взаимодействия с нейтрализующими антителами либо эндогенными или заранее сформированными антителами.
В конкретных вариантах осуществления настоящего изобретения мутагенез следует использовать для модификации области суперантигена, которая определяет связывание с молекулами MHC класса II. Мутагенез следует осуществлять с помощью разнообразных стандартных мутагенных методик. Мутация представляет собой процесс, в соответствии с которым в организме происходят количественные или структурные изменения. Мутация может включать модификацию нуклеотидной последовательности единственного гена, блока генов или целой хромосомы. Изменения в одиночных генах могут являться следствием точковых мутаций, которые включают удаление, добавление или замену основания единственного нуклеотида в ДНК-последовательности, либо они могут быть следствием изменений, включающих вставку или делецию большого числа нуклеотидов.
Одним из самых успешных методов мутагенеза является аланиновый сканирующий мутагенез, при котором ряд остатков индивидуально замещается аминокислотой аланином так, что можно определить эффекты от утраты взаимодействий, связанных с боковой цепью, при минимизации риска нарушений в конформации белка (Cunningham и соавт., 1989).
В последние годы разработаны методы оценки константы равновесия лигандного связывания с использованием очень маленьких количеств белка (Патенты США №№5221605 и 5238808). Способность осуществлять функциональные анализы с небольшим количеством материала можно использовать для создания высокоэффективных in vitro-методов насыщающего мутагенеза антител. Заявители обходили стадии клонирования, сочетая ПЦР-мутагенез с объединенной in vitro транскрипцией/трансляцией для высокомасштабного создания белковых мутантов. При этом полученные ПЦР-продукты используют непосредственно в виде матрицы для транскрипции/трансляции in vitro мутантных одноцепочечных антител. Из-за высокой эффективности, с которой можно получить и проанализировать таким путем все 19 аминокислотных замен, представляется возможным осуществить насыщающий мутагенез по многочисленным представляющим интерес остаткам, процесс который можно описать или сканирование in vitro насыщающего мутагенеза (Burks и соавт., 1997).
In vitro-сканирование насыщающего мутагенеза создает быстрый способ получения большого количества структурно-функциональной информации, включающей: (i) идентификацию остатков, которые модулируют лиганд-связывающую специфичность, (ii) лучшее осмысление лигандного связывания на основе идентификации тех аминокислот, которые сохраняют активность, и тех, которые упраздняют активность по данному местоположению, (iii) оценку общей пластичности активного сайта или белкового субдомена, (iv) идентификацию аминокислотных замен, которые повышают связывание.
Структурно-направленный сайт-специфический мутагенез представляет мощный инструмент для анализа и конструирования взаимодействий белок-лиганд (Wells, 1996, Braisted и соавт., 1996). Данный метод разработан для получения и тестирования вариантов последовательностей путем введения одного или нескольких изменений в нуклеотидную последовательность выбранной ДНК.
Сайт-специфический мутагенез использует специфические олигонуклеотидные последовательности, которые кодируют заданную мутацию в определенной ДНК-последовательности, а также достаточное количество примыкающих немодифицированных нуклеотидов. Таким путем создается достаточного размера и сложности праймерная последовательность для образования стабильного дуплекса по обеим сторонам перемещаемого делеционного стыка. Предпочтителен праймер длиной около 17-25 нуклеотидов и приблизительно 5-10 остатками по обеим сторонам стыка в данной измененной последовательности.
В данном методе используют, как правило, бактериофаговый вектор, который существует как в одноцепочечной, так и в двухцепочечной форме. Векторы, используемые в сайт-направленном мутагенезе, включают такие векторы как фаг М13. Эти фаговые векторы коммерчески доступны и их использование вообще хорошо известно специалистам в данной области техники. Двухцепочечные плазмиды также регулярно используются в сайт-направленном мутагенезе, что исключает стадию переноса интересующего гена из фага в плазмиду.
В общих чертах, вначале получают одноцепочечный вектор или плавят две цепи двухцепочечного вектора, который включает, в рамках своей последовательности ДНК-последовательность, кодирующую требуемый белок или генетический элемент. Синтетически получают олигонуклеотидный праймер, несущий требуемую мутантную последовательность, после чего отжигают его вместе с одноцепочечным ДНК-препаратом, учитывая степень несоответствия при выборе условий гибридизации. Для завершения синтеза цепи, несущей мутацию, полученный гибридизационный продукт подвергают действию ДНК-полимеризующих ферментов, таких как полимераза I E.coli (фрагмент Кленова). Таким образом, образуется гетеродуплекс, в котором одна цепь кодирует исходную немутантную последовательность, а вторая цепь несет требуемую мутацию. Затем этот гетеродуплексный вектор используют для трансформации соответствующих клеток-мишеней, таких как клетки E.coli, и отбирают клоны, которые включают рекомбинантные векторы, несущие конструкцию мутантной последовательности.
Исчерпывающуюинформацию о функциональной значимости и содержании информации данного остатка можно получить наилучшим образом с помощью насыщающего мутагенеза, в котором анализируются все 19 аминокислотных замен. Недостаток данного подхода заключается в том, что математическая логика анализа по многим остаткам при насыщающем мутагенезе отпугивает (Warren и соавт., 1996, Brown и соавт., 1996; Zeng и соавт., 1996; Burton and Barbas, 1994; Yelton и соавт., 1995; Jackson и соавт., 1995; Short и соавт., 1995; Wong и соавт., 1996; Hilton и соавт., 1996). Необходимо исследовать сотни, а возможно даже тысячи сайт-специфических мутантов. Впрочем, усовершенствованные методы делают получение и быстрый скрининг мутантов гораздо более простыми. Смотрите также патенты США №№5798208 и 5830650 для описания "прогонного" ("walk-through") мутагенеза.
Другие методы сайт-направленного мутагенеза раскрыты в патентах США №№520007; 5284760; 5354670; 5366878; 5389514; 5635377; и 5789166.
Помимо биологических функциональных эквивалентов, которые получают с использованием вышеуказанных методов мутагенеза, авторы настоящего изобретения считают также, что могут быть созданы структурно похожие соединения, которые имитируют ключевые части суперантигена или конъюгата настоящего изобретения. Такие соединения, которые можно назвать пептидомиметики, можно использовать таким же образом, что и конъюгаты настоящего изобретения, и они, следовательно, также являются функциональными эквивалентами.
Некоторые миметики, которые имитируют элементы вторичной и третичной структуры белка, описаны у Johnson и соавт. (1993). Причина, лежащая в основе использования пептидных миметиков, заключается в том, что основная пептидная цепь белков существует преимущественно для ориентирования аминокислотных боковых цепей таким образом, чтобы способствовать молекулярному взаимодействию, такому, например, как взаимодействие антитела и/или антигена. Следовательно, пептидный миметик создается для того, чтобы сделать возможными молекулярные взаимодействия, присущие природной молекуле.
Несколько успешных применений идеи о пептидном миметике сосредоточено на миметиках β-витков в белках, которые, как известно, высокоантигенны. Вероятную β-витковую структуру в полипептиде можно предсказать с помощью компьютерных алгоритмов, что и рассматривается здесь. После того как определены аминокислоты, составляющие данный виток, можно создавать миметики, чтобы добиться похожей пространственной ориентации существенных элементов боковых аминокислотных цепей.
Другие подходы сосредоточены на использовании небольших белков со множеством дисульфидных связей в качестве привлекательных структурных матриц для получения биологически активных конформаций, которые имитируют связывающие сайты больших белков, Vita и соавт. (1998). Структурный мотив, который в некоторых токсинах выглядит эволюционно консервативным, является небольшим (30-40 аминокислот), стабильным и весьма рекомендуемым для мутации. Данный мотив состоит из бета-складки и альфа-спирали, соединенных мостиком во внутренней коре с помощью трех дисульфидов.
Бета-II-витки удачно имитировали с использованием циклических L-пентапептидов и циклических пентапептидов с D-аминокислотами (Weisshoff и соавт., 1999). Кроме того, Johanhesson и соавт. (1999) сообщают о бициклических трипептидах со свойствами, индуцирующими обратный виток.
В данной области техники раскрыты способы создания специфических структур. Например, миметики альфа-спирали раскрыты в патентах США №№5446128; 5710245; 5840833; и 5859184. Эти структуры придают данному пептиду или белку больше термостабильности, а также увеличивают резистентность к протеолитической деградации. Описаны шести-, семи-, одиннадцати-, двенадцати-, тринадцати- и четырнадцатичленные кольцевые структуры.
Способы создания конформационно ограниченных бета-витков и бета-горбов описаны, например, в патентах США №№5440013; 5618914; и 5670155. Бета-витки позволяют изменяться боковым заместителям, без изменений в соответствующей скелетной конформации и обладают соответствующими концами для включения в пептиды с помощью стандартных методов синтеза. Другие виды витков миметиков включают обратные и гамма-витки. Миметики с обратными витками раскрыты в патентах США №№5475085 и 5929237, а миметики с гамма-витками описаны в патентах США №№5672681 и 5674976.
G. Экспрессия суперантигенов
Настоящее изобретение включает также использование экспрессионных векторов и клеток-хозяев. Эти экспрессионные векторы, которые были созданы геноинженерным способом, и содержат нуклеотидную последовательность конъюгатов, вводят или трансформируют в клетки-хозяева для получения конъюгатов настоящего изобретения.
Клетки-хозяева можно создать биотехнологически, чтобы ввести в них нуклеотидные последовательности и экспрессировать пептиды настоящего изобретения. Введение нуклеотидных последовательностей в клетку-хозяина можно осуществить с помощью трансфекции кальций-фосфатом, трансфекции, опосредованной DEAE-декстраном, трансвекции, микроинъекции, катионной липид-опосредованной трансфекции, электропорации, трансдукции, соскоба (scrape loading), бомбардировки микрочастицами, инфицированием или иными способами. Такие способы описаны во многих типовых лабораторных руководствах, таких как Davis, и соавт., BASIC METHODS IN MOLECULAR BIOLOGY (Основные методы молекулярной биологии), (1986) и Sambrook, и соавт., MOLECULAR CLONING: A LABORATORY MANUAL, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989).
Типичные примеры подходящих клеток-хозяев включают бактериальные клетки, такие как стрептококки, стафилококки, E.coli, стрептомицеты и клетки Bacillus subtilis; клетки грибов, такие как дрожжевые клетки и клетки из рода Aspergillus, клетки насекомых, такие как клетки Drosophila S2 и Spodoptera Sf9; клетки животных, такие как CHO, COS, HeLa, C127, 3T3, BHK, 293 и клетки меланомы Bowes.
Лечение злокачественных опухолей
В настоящем изобретении суперантиген конъюгируют с антителом или с фрагментом антитела для нацеливания и разрушения клеток злокачественной опухоли. Примеры злокачественных опухолей включают, но не ограничиваются указанным, злокачественную опухоль легких, молочных желез, прямой кишки, почек, поджелудочной железы, яичника, желудка, шейки матки и предстательной железы.
В одном аспекте настоящего изобретения злокачественная клетка должна нести некий маркер, который поддается нацеливанию, т.е., он не представлен в большинстве других клеток. Существует множество опухолевых маркеров и любой из них может оказаться пригодным для нацеливания в контексте настоящего изобретения. Специфические мишени настоящего изобретения включают антитела. Антитела, которые рассматриваются в настоящем изобретении, включают, но не ограничиваются указанным, Fab-фрагмент. Примеры Fab-фрагмента включают C215Fab или 5T4Fab. Кроме Fab, другие общие опухолевые маркеры включают карциноэмбриональный антиген, специфичный для предстательной железы антиген, ассоциированный со злокачественной опухолью мочевой системы антиген, фетальный антиген, тирозиназу (p97), gp68, TAG-72, HMFG, сиалилантиген Льюиса, MucA, MucB, PLAP, рецептор эстрогена, рецептор ламинина, erb B и p155.
Другим аспектом настоящего изобретения является использование иммуностимулирующей молекулы в качестве агента или, более предпочтительно, вместе с другим агентом, такой, например, как цитокины, такие, например, как IL-2, IL-4, IL-12, GM-CSF, фактор некроза опухоли; интерфероны альфа, бета, и гамма; F42K и другие цитокиновые аналоги; хемокин, такой, например, как MIP-1, MIP-1бета, MCP-1, RANTES, IL-8; или фактор роста, такой, например, как FLT3-лиганд. Данная стимулирующая молекула может быть сопряжена с конъюгатом настоящего изобретения или введена в виде адъюванта в сочетании с конъюгатом настоящего изобретения.
Одним из цитокинов, рассматриваемых для использования в настоящем изобретении, является IL2 или производное, обладающее по существу такой же биологической активностью, что и нативный IL2. Интерлейкин-2 (IL-2), первоначально обозначенный как фактор I Т-клеточного роста, является весьма совершенным индуктором Т-клеточной пролиферации и является фактором роста всех субпопуляций Т-лимфоцитов. IL-2 представляет собой антиген-независимый фактор пролиферации, который стимулирует прохождение клеточного цикла покоящимися клетками, и таким образом делает возможной клональную экспансию активированных Т-лимфоцитов. Поскольку свежевыделенные лейкозные клетки также секретируют IL2 и отвечают на него, IL2 может функционировать в качестве аутокринного модулятора роста для этих клеток, способного ухудшать ATL. IL2 способствует также пролиферации активированных В-клеток, хотя это требует присутствия дополнительных факторов, например, IL4. In vitro IL2 также стимулирует рост олигодендроглиальных клеток. В связи с его действием на Т-клетки и В-клетки, IL2 является центральным регулятором иммунных ответов. Он также играет роль в противовоспалительных реакциях, в гемопоэзе и в надзоре за злокачественной опухолью. IL2 стимулирует синтез γ-IFN в периферических лейкоцитах и также индуцирует секрецию IL-1, α-TNF и β-TNF. Индукция секреции уничтожающих опухолевые клетки цитокинов, помимо активности в экспансии LAK-клеток (лимфокинактивированные киллерные клетки), является главным фактором, ответственным за противоопухолевую активность IL2.
Предполагается, что настоящее изобретение может помочь пациенту, страдающему от злокачественной опухоли или от пролиферативного заболевания. Количество прописанного данному пациенту лекарства представляет собой терапевтически эффективное количество или количество, которое поможет в лечении злокачественной опухоли или заболевания. Введение конъюгата может быть парентеральным или пищевым. Типичные пищевые способы введения включают, но не ограничиваются указанным, пероральный, ректальный, подъязычный или буккальный. Типичные парентеральные способы введения включают, но не ограничиваются указанным, внутрибрюшинный, внутривенный, подкожный, внутримышечный, интрадермальный, внутриопухолевый, и внутрисосудистый.
III. Фармацевтические композиции
Соединения настоящего изобретения можно использовать отдельно или в сочетании с другими соединениями, такими как терапевтические соединения.
Фармацевтические формы, пригодные для инъецирования, включают стерильные водные растворы и/или дисперсии; композиции, включающие кунжутное масло, арахисовое масло и/или водный пропиленгликоль; и/или стерильные порошки для немедленного приготовления инъекционных растворов и/или дисперсий. Во всех случаях фармацевтическая форма должна быть стерильной и/или должна быть жидкой за такой степени, чтобы можно было легко заполнить шприц. Она должна быть стабильной в условиях производства и/или при хранении и/или должна быть защищена от действия контаминирующих микроорганизмов, таких как бактерии и/или грибы.
Растворы активных соединений в виде свободного основания и/или фармакологически приемлемых солей можно приготовить в воде, смешав надлежащим образом с поверхностно-активным соединением, таким как гидроксипропилцеллюлоза. Дисперсии можно также приготовить в глицерине, жидких полиэтиленгликолях, и/или в их смесях, и/или в маслах. В соответствии с обычными условиями хранения и/или использования эти препараты содержат консервант для предотвращения роста микроорганизмов.
Конъюгат настоящего изобретения можно составить в композицию в нейтральной и/или солевой форме. Фармацевтически приемлемые соли включают кислотно-аддитивные соли (образуемые со свободными аминогруппами белка) и/или соли, образуемые неорганическими кислотами, такими, например, как хлористоводородная и/или фосфорная кислоты, и/или такими органическими кислотами как уксусная, щавелевая, виннокаменная, миндальная, и/или им подобным. Соли, образуемые свободными карбоксильными группами, можно также получить из неорганических оснований, таких, например, как натриевая, калиевая, аммонийная, кальциевая, и/или железистая гидроокиси, а также/или из таких органических оснований как изопропиламин, триметиламин, гистидин, прокаин и/или им подобным. С точки зрения использования пептидных терапевтических средств в качестве активных ингредиентов можно применить технологию, изложенную в патентах США №№4608251; 4601903; 4599231; 4599230; 4596792 и/или 4578770, включенных здесь, каждый путем ссылки.
Носитель может также представлять собой растворитель и/или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, и/или жидкий полиэтиленгликоль, и/или им подобные), соответствующие их смеси, и/или растительные масла. Надлежащую текучесть можно поддерживать, например, путем использования покровного материала, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и/или путем использования поверхностно-активных соединений. Предотвратить действие микроорганизмов можно с помощью различных антибактериальных и противогрибковых агентов, например парабензоатов, хлорбутанола, фенола, сорбиновой кислоты, тиомерсала, и/или им подобных. Во многих случаях предпочтительно включать изотоничные агенты, например сахара и/или хлорид натрия. Пролонгированную абсорбцию инъецируемых композиций можно осуществить путем использования в данных композициях агентов, задерживающих поглощение, например, алюминиймоностеарата и/или желатина.
Стерильные инъекционные растворы готовят путем введения активных соединений в нужное количество соответствующего растворителя с различными, если потребуется, другими вышеперечисленными ингредиентами, с последующей стерилизацией фильтрованием. Обычно дисперсии готовят путем введения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и/или другие необходимые ингредиенты из вышеперечисленных. Что касается стерильных порошков, то для получения стерильных инъекционных растворов предпочтительными способами их приготовления являются методы вакуумной сушки и/или сушки сублимацией, с помощью которых производят порошок активного ингредиента и любого дополнительного требуемого ингредиента из их раствора, ранее простерилизованного фильтрацией. Предполагается также для прямой инъекции приготовление растворов с большой или высокой концентрацией, где, как считают, использование в качестве растворителя DMSO имеет своим результатом чрезвычайно быстрое проникновение, что приводит к доставке высоких концентраций активных агентов в небольшую область.
После приготовления растворы вводят способом, совместимым с дозированной композицией и/или в таком количестве, которое является терапевтически эффективным. Данные композиции легко вводятся в разнообразных дозированных формах, таких как вышеописанные типы инъекционных растворов, хотя можно также использовать капсулы и/или им подобное для высвобождения лекарственного средства.
Для парентерального введения, например, в водном растворе, раствор, если необходимо, должен быть должным образом забуферен и/или жидкому разбавителю вначале сообщают изотоничность с помощью достаточного количества соли и/или глюкозы. Эти индивидуальные водные растворы особенно пригодны для внутривенного, внутримышечного, подкожного и/или внутрибрюшинного введения. В этой связи стерильная водная среда, которая может быть использована, должна быть хорошо известна специалистам в данной области техники в свете представленного описания. Например, одну дозу можно было бы растворить в 1 мл изотоничного раствора NaCl, и/или либо довести до 1000 мл жидкостью для введения в подкожную клетчатку, и/или инъецировать в предполагаемый участок введения, (смотрите, например, "Remington's Pharmaceutical Sciences" 15th Edition, pages 1035-1038 и/или 1570-1580). В зависимости от состояния пациента, которого лечат, неизбежно происходит некоторое изменение дозировки. Лицо, ответственное за введение, должно в любом случае определить надлежащую дозу для отдельного пациента.
Активный конъюгат и/или агенты можно составить в терапевтическую смесь, которая включает около 0,0001-1,0 миллиграмма, и/или около 0,001-0,1 миллиграмма, и/или около 0,1-1,0 и/или даже около 10 миллиграмм на дозу и/или и так далее. Можно также вводить многократные дозы.
Кроме соединений, составленных для парентерального введения, такого как для внутривенного, внутрисуставного и/или внутримышечного инъецирования, другие фармацевтически приемлемые формы включают, например, таблетки и/или иные твердые частицы для перорального введения; липосомные композиции; капсулы с замедленным высвобождением данной композиции; и/или любую иную ныне используемую форму, в том числе и кремы.
Можно также использовать растворы в нос и/или распыляемые растворы, аэрозоли и/или ингалируемые средства настоящего изобретения. Назальные растворы представляют собой, как правило, водные растворы, созданные для введения в носовые ходы по каплям и/или в виде распыляемых растворов. Назальные растворы готовят таким образом, чтобы они, во многих отношениях, были подобны назальным секретам для поддержания нормального цилиарного действия. Поэтому водные назальные растворы, как правило, являются изотоничными и/или слегка забуференными для поддержания рН 5,5-6,5. Кроме того, в данную композицию могут быть включены, если понадобится, противомикробные антисептики, аналогичные используемым в офтальмологических препаратах, и/или соответствующие стабилизаторы лекарственных средств. Известны различные коммерческие назальные препараты, и они включают, например, антибиотики, и/или антигистамины и/или используемые для профилактики астмы.
Дополнительные композиции, которые пригодны для других способов введения, включают вагинальные суппозитории и/или пессарии. Можно также использовать прямокишечный пессарий и/или суппозиторий. Суппозитории представляют собой твердые дозированные формы разного веса и/или формы, обычно - лекарственные свечи, для введения в прямую кишку, влагалище и/или уретру. После введения суппозитории размягчаются, плавятся и/или растворяются в полостных жидкостях. В целом, суппозитории могут включать традиционные связующие вещества и/или носители, например, полиалкиленгликоли и/или триглицериды; такие суппозитории могут быть созданы из смесей, содержащих активный ингредиент в диапазоне от 0,5% до 10%, предпочтительно 1%-2%.
Пероральные композиции включают такие используемые в норме наполнители как, например, фармацевтически чистые маннит, лактозу, крахмал, магнийстеарат, натрийсахарин, целлюлозу, углекислый магний и/или тому подобное. Эти композиции принимают форму растворов, суспензий, таблеток, пилюль, капсул, композиций с замедленным высвобождением и/или порошков. В некоторых определенных вариантах осуществления настоящего изобретения пероральные фармацевтические композиции включают инертный разбавитель и/или усвояемый пищевой носитель, и/или они могут быть включены в твердый и/или мягкий желатиновый корпус капсулы, и/или они могут быть сжаты в таблетки, и/или они могут быть включены непосредственно в пищу. Для перорального терапевтического введения активные соединения могут быть введены с наполнителями и/или использоваться в форме проглатываемых таблеток, защечных таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток и/или тому подобное. Такие композиции и/или препараты должны содержать, по меньшей мере, 0,1% активного соединения. Разумеется, процентные доли данных композиций и/или препаратов могут варьировать и/или могут находиться в пределах от около 2 до около 75% от веса дозированной единицы, и/или предпочтительно между 25-60%. Количество активных соединений в таких терапевтически используемых композициях является таким, что можно получить соответствующую дозу.
Таблетки, пастилки, пилюли, капсулы и/или им подобные могут также содержать: связывающее вещество в виде трагакантовой камеди, камеди акации, кукурузного крахмала, и/или желатины; наполнители, такие как дикальцийфосфат; разрыхляющие агенты, такие как кукурузный крахмал, картофельный крахмал, альгиновая кислота и/или им подобные; смазывающее вещество, такое как магнийстеарат; и/или подслащивающий агент, такой как сахароза, лактоза и/или сахарин; может быть добавлен и/или ароматизирующий агент, такой как мята перечная, масло грушанки, и/или вишневый ароматизатор. Если форма дозированной единицы представляет собой капсулу, то она может содержать, кроме материалов вышеуказанного типа, жидкий носитель. Различные иные материалы могут быть представлены в виде покрытий и/или иным образом модифицировать физическую форму дозированной единицы. Например, таблетки, пилюли и/или капсулы могут быть покрыты шеллаком, сахаром и/или тем и другим. Сироп эликсира может содержать активные соединения, сахарозу в качестве подслащивающего агента, метил и/или пропилапарабензоаты в качестве антисептиков, краситель и/или ароматизатор, такой как вишневый и/или апельсиновый ароматизатор.
В некоторых вариантах осуществления настоящего изобретения липидные композиции, и/или нанокапсулы предполагают использовать для введения в клетки-хозяева конъюгата/или агентов, и/или для введения векторов генотерапии, включая векторы дикого типа и/или антисмысловые векторы.
Как правило, нанокапсулы могут включать соединения в стабильном и/или воспроизводимом состоянии. Чтобы исключить побочные эффекты, связанные с внутриклеточной полимерной перегрузкой, такие ультратонкие частицы (размером около 0,1 мкм) должны создаваться с использованием полимеров, способных к деградации in vivo. Биодеградирующиеся полиалкилцианоакрилатные наночастицы, которые удовлетворяют этим требованиям, предполагается использовать в настоящем изобретении и/или такие частицы можно легко изготовить.
В любом варианте осуществления настоящего изобретения данный конъюгат может быть ассоциирован с липидом. Такие конъюгаты, ассоциированные с липидом, можно инкапсулировать в водное ядро любой липосомы, распределить в липидном бислое липосомы, прикрепить к липосоме с помощью линкерной молекулы, которая связывается и с липосомой и с олигонуклеотидом, заключить в липосому, связать в комплекс с липосомой, диспергировать в растворе, содержащем липид, смешать с липидом, объединить с липидом, они также могут содержать в виде суспензии в липиде, содержаться или образовывать комплекс с мицеллой, или по-другому ассоциироваться с липидом. Липидная или липид/конъюгат-ассоциированная композиции настоящего изобретения не ограничиваются какой-либо конкретной структурой в растворе. Например, они могут быть представлены бислойной структурой, в виде мицелл, или свернутой структурой. Они могут быть также просто рассеяны в растворе, возможно образуя агрегаты, которые неоднородны по размеру или форме.
Липиды представляют собой жировые вещества, которые могут быть встречаемыми в природе или синтетическими липидами. К примеру, липиды включают жировые капельки, которые естественным образом находятся в цитоплазме, а также класс соединений, хорошо известные специалистам в данной области техники, которые содержат длинноцепочечные алифатические углеводороды и их производные, такие как жирные кислоты, спирты, амины, аминоспирты и альдегиды.
В соответствии с настоящим изобретением для получения липосом можно использовать фосфолипиды, и они могут нести суммарный положительный, отрицательный или нейтральный заряд. Для придания липосомам отрицательного заряда можно использовать диацетилфосфат, а стеариламин можно использовать для придания липосомам положительного заряда. Липосомы можно создать из одного или нескольких фосфолипидов.
Нейтрально заряженный липид может включать липид без заряда, почти незаряженный липид или липидную смесь с равным количеством положительных и отрицательных зарядов. Подходящие фосфолипиды включают фосфатидилхолины и другие, которые хорошо известны специалистам в данной области техники.
Липиды, пригодные для использования в соответствии с настоящим изобретением, могут быть получены из коммерческих источников. Например, димиристилфосфатидилхолин ("DMPC") можно получить от Sigma Chemical Co., диацетилфосфат ("DCP") получают от K & K Laboratories (Plainview, NY); холестерин ("Chol") получают от Calbiochem-Behring; димиристилфосфатидилглицерин ("DMPG") и другие липиды можно получить от Avanti Polar Lipids, Inc. (Birmingham, Ala.). Маточные растворы липидов в хлороформе или в хлороформ/метаноле можно хранить при около -20°С. Предпочтительно, хлороформ используется только в качестве растворителя, так как он быстрее испаряется, чем метанол.
Фосфолипиды природного происхождения, такие как фосфатидилхолин яйца или из соевых бобов, фосфатидная кислота мозга, фосфатидилинозитол мозга или растений, кардиолипин сердца и фосфатидилэтаноламин растений или бактерий предпочтительно не используются в качестве первичного фосфатида, т.е., составляющего 50% и больше от общего фосфатидного состава, из-за нестабильности и негерметичности полученных липосом.
Фосфолипиды при диспергировании в воде могут образовывать различные структуры, отличные от липосом, в зависимости от молярного соотношения липида и воды. При низком соотношении предпочтительной структурой является липосома. Физические характеристики липосом зависят от рН, ионной силы и/или присутствия двухвалентных катионов. Липосомы могут демонстрировать низкую проницаемость для ионов и/или полярных веществ, но при повышенной температуре подвергаются фазовому переходу, что явно изменяет их проницаемость. Такой фазовый переход влечет за собой изменение плотно упакованной, упорядоченной структуры, известной в качестве гелеобразного состояния, в неплотно упакованную, менее упорядоченную структуру, известную под именем жидкое состояние. Это происходит при характерной для фазового перехода температуре и/или приводит к повышению проницаемости для ионов, сахаров и/или лекарственных средств.
Липосомы взаимодействуют с клетками с помощью четырех разных механизмов: эндоцитоза, осуществляемого фагоцитарными клетками ретикулоэндотелиальной системы, такими как макрофаги и/или нейтрофилы; адсорбции на клеточной поверхности либо с помощью неспецифических слабых гидрофобных и/или электростатических сил, и/или с помощью специфических взаимодействий с компонентами клеточной поверхности; слияния с плазматической мембраной клетки, путем встраивания липидного бислоя липосомы в плазматическую мембрану, и одновременного высвобождение содержимого липосом в цитоплазму; и/или путем переноса липосомных липидов в клеточную и/или субклеточную мембраны, или наоборот, без какой-либо ассоциации содержимого липосом. Изменяя липосомный состав, можно видоизменять действующий механизм, хотя и не более чем на одно действие в одно и то же время.
Липосомопосредованная доставка олигонуклеотида и экспрессия чужеродной ДНК in vitro оказывалась очень успешной. Wong и соавт., (1980) продемонстрировали осуществимость липосом-опосредованной доставки и экспрессии чужеродной ДНК в культивируемых клетках куриного эмбриона, HeLa и гепатомы. Nicolau и соавт., (1987) осуществили успешный липосом-опосредованный генный перенос у крыс после внутривенной инъекции.
В некоторых вариантах осуществления настоящего изобретения липид может быть ассоциирован с гемагглютинирующим вирусом (HVJ). Показано, что при этом облегчается слияние с клеточной мембраной и активизируется введение в клетки инкапсулированной в липосому ДНК (Kaneda и соавт., 1989). В других вариантах осуществления настоящего изобретения липид может быть связан в комплекс или быть использован в соединении с ядерными негистоновыми белками хромосом (HMG-1) (Kato и соавт., 1991). Еще в одних вариантах осуществления настоящего изобретения липид может быть связан в комплекс или использован в соединении и с HVJ, и с HMG-1. Поскольку такие экспрессионные векторы успешно использовались для переноса и экспрессии олигонуклеотида in vitro и in vivo, они применимы для настоящего изобретения. Если в ДНК-конструкции используют бактериальный промотор, также желательно включить в данную липосому соответствующую бактериальную полимеразу.
Липосомы, используемые в соответствии с настоящим изобретением, можно создать разными способами. Размер липосом варьирует в зависимости от способа синтеза. Липосома, суспендированная в водном растворе, как правило, принимает форму сферической везикулы, обладающей одним или несколькими концентрическими слоями молекул липидного бислоя. Каждый слой состоит из молекул в параллельном порядке, представленных формулой XY, где Х представляет собой гидрофильную составляющую, Y представляет собой гидрофобную составляющую. В водной суспензии концентрические слои располагаются таким образом, что гидрофильные компоненты стремятся остаться в контакте с водной фазой, а гидрофобные участки стремятся самоассоциировать. Например, если водные фазы представлены и внутри липосомы и без нее, то липидная молекула может образовать бислой, известный под именем ламелла, со структурой XY-YX. Агрегаты липидов могут образоваться, если гидрофильная и гидрофобная части более чем одной липидной молекулы становятся ассоциированными друг с другом. Размер и форма этих агрегатов зависит от многих разных переменных, таких как природа растворителя и наличие других соединений в данном растворе.
Липосомы в рамках настоящего изобретения можно получить в соответствии с известными лабораторными методами. В одном из предпочтительных вариантов осуществления настоящего изобретения липосомы получают путем смешивания липосомных липидов в растворителе в контейнере, например, стеклянной грушевидной колбе. Данный контейнер должен обладать объемом, в десять раз превышающем объем рассчитанной суспензии липосом. Используя роторный испаритель растворитель удаляют при температуре приблизительно 40°С под отрицательным давлением. Обычно растворитель удаляют в пределах около 5 мин - 2 часов, в зависимости от требуемого объема для данных липосом. Полученную композицию в дальнейшем можно обезводить в эксикаторе при отрицательном давлении. Эти обезвоженные липиды, как правило, отбрасывают примерно через 1 неделю из-за тенденции деградировать со временем.
Обезвоженные липиды можно гидратировать до концентрации примерно 25-50 мМ фосфолипидов в стерильной апирогенной воде путем встряхивания до тех пор, пока вся данная липидная пленка не ресуспендируется. Полученные водные липосомы можно затем разделить на аликвоты, каждую аликвоту поместить в ампулу, лиофилизировать и запаять под отрицательным давлении.
В качестве альтернативы липосомы можно получить в соответствии с другими известными лабораторными методами: методом Bangham и соавт., (1965), содержание которого включено здесь путем ссылки; методом Gregoriadis, как описано в DRUG CARRIERS IN BIOLOGY AND MEDICINE, G. Gregoriadis ed. (1979) pp. 287-341, содержание которого включено здесь путем ссылки; и методом обращено-фазового испарения, который описан у Szoka and Papahadjopoulos (1978). Вышеуказанные методы отличаются, соответственно, по их способностям включать водорастворимый материал и, соответственно, по их соотношениям водное пространство-липид.
Полученные, как указано выше, обезвоженные липиды или лиофилизованные липосомы можно дегидратировать и воссоздать в растворе ингибирующего пептида и разбавить до соответствующей концентрации с помощью подходящего растворителя, например, DPBS. Затем данную смесь энергично перемешивают в вихревом смесителе. Неинкапсулированную нуклеиновую кислоту удаляют центрифугированием при 29000 х g и полученный осадок липосом промывают. Промытые липосомы ресуспендируют до соответствующей общей фосфолипидной концентрации, например, около 50-200 мМ. Количество инкапсулированной нуклеиновой кислоты можно определить с помощью стандартных способов. После определения количества нуклеиновой кислоты инкапсулированной в липосомном препарате, эти липосомы можно разбавить до соответствующей концентрации и хранить при 4°С до использования.
Фармацевтические композиции, включающие липосомы будут, как правило, включать стерильный, фармацевтически приемлемый носитель или разбавитель, такой как вода или физиологический раствор.
ПРИМЕРЫ
Нижеследующие примеры включены для демонстрации предпочтительных вариантов осуществления настоящего изобретения. Специалистам в данной области техники следует иметь в виду, что методы, раскрытые в данных примерах, описываются автором настоящего изобретения как хорошо зарекомендовавшие себя при осуществлении данного изобретения и поэтому могут рассматриваться в качестве предпочтительных практических методов. Тем не менее, специалисты в данной области техники должны, в свете настоящего описания, понимать, что в конкретных вариантах осуществления настоящего изобретения, которые раскрыты, могут быть внесены многочисленные изменения, не выходящие за рамки и суть настоящего изобретения, и все же получить близкий или аналогичный результат.
Пример 1
Мутагенез in vitro
С использованием метода, основанного на полимеразной цепной реакции (PCR) созданы различные варианты суперантигена.
Вкратце, ПЦР-продукты, содержали два уникальных сайта фермента рестрикции, один из которых концевой. Для субклонирования использовали pUC19 (GIBCO BRL Life Technologies, Middlesex, Соединенное Королевство), приготовленную в соответствии с протоколом QIAprep Spin Miniprep Kit (QIAGEN, Hilden, Германия). Для облегчения последующего анализа были включены точковые мутации, не влияющие на аминокислотную последовательность. ПЦР-реакцию осуществляли на Perkin Elmer Gene Amp PCR system 2400 с Taq-ДНК-полимеразой и соответствующим ПЦР-буфером, содержащем 15 мМ MgCl2 (Roche Molecular Biochemicals, Basel, Швейцария). ПЦР-продукты и векторы обрабатывали в течение ночи с помощью соответствующих ферментов рестрикции. Их выделяли очисткой с помощью электрофореза в 1%-ном агарозном геле (GIBCO BRL Life Technologies), содержащем 0,5 мкг/мл этидийбромида (Sigma-Aldrich, Steinheim, Германия) в TAE-буфере (Sigma-Aldrich). Фрагмент, содержащий ДНК, вырезали из геля и экстрагировали с использованием системы CONSERTTM Rapid Gel Extraction System (GIBCO BRL Life Technologies). Вектор и вставку лигировали (Т4-ДНК-лигаза, Roche Molecular Biochemicals) при комнатной температуре в течение 3-4 часов. Смесь, подвергнутую лигированию, трансформировали в клетки Escherichia coli, штамма DH5α (GIBCO BRL Life Technologies) в соответствии с инструкциями, прилагаемыми к данному штамму клеток. Позитивные клоны подтверждали с использованием ДНК-секвенирования. Правильные последовательности обрабатывали с помощью RsrII/HindIII при 37°С в течение ночи и лигировали в экспрессионный вектор (Dohlstein и соавт., 1994). Вариабельные участки Fab меняли на С215, чтобы соответствовать моделям домашних животных. Наконец, данную конструкцию электропорировали в клетки штамма K12 UL635 Escherichia coli (xyl-7, ara-14, T4R, Δompt).
Пример 2
Идентификация областей, связывающих человеческие антитела против SEA
Области распознаваемые человеческими антителом против SEA, идентифицировали в обработанном пепсином SEA или химерном варианте SEA и SEE, SEA/E-18, ранее описанного в качестве SEE/A-A (Antonsson и соавт., 1997), с заменой D227A.
Каждый суперантиген инкубировали с 0,5% пепсина, 10 мМ HCl, 150 мМ NaCl (w/w) в течение 60 минут при 37°С. Полученную пептидную смесь нейтрализовали с помощью 2 М Трис-HCl, рН 8,0, и нагружали на HiTrap-колонку размером 1 мл (Amersham Pharmacia Biotech, Uppsala, Швеция) с иммобилизованными человеческими антителами к SEA.В качестве промывочного буфера используютPBS, 8,1 мМ Na2HPO4, 1,5 мМ KH2PO4, 137 мМ NaCl, 2,7 мМ KCl, рН 7,3, а связывающие антитела фрагменты элюировали с использованием 0,1 М уксусной кислоты, рН 3,0. Фрагменты идентифицировали до и после выделения очисткой с использованием ВЭЖХ, совмещенной с масс-спектрометрией (MS) (фиг.1). Хроматографию осуществляли на колонке С18 (2х250 мм) (VYDACTM, Hesperia, California, США) с использованием линейного градиента от 10 до 60% ацетонитрила в 0,1 % трифторуксусной кислоте в течение 30 мин. при 40°С. Определение массы осуществляли с использованием MS с электрораспылительной ионизацией (Finnigan LCQ, Thermoquest, San Jose, California, США). Фрагменты, обнаруживаемые в гидролизате с одним и тем же временем удержания до и после аффинной очистки, рассматривались в качестве позитивных (фиг.2).
Пример 3
Молекулярное моделирование
Химерный суперантиген SEA/E-18 основывался на последовательности SEE, за исключением четырех аминокислотных остатков вблизи N-конца, которые происходили из SEA, и одного замещения на С-концевом участке D227A (фиг.3 и фиг.4) (Antonsson и соавт., 1997)
Вкратце, трехмерные структуры суперантигенов, с наибольшей идентичностью с SEE, которые доступны из PBD, использовали в качестве матриц для создания гомологичной модели SEAE-18, т.е., SEA (1ESF, Shard и соавт., 1SXT, Sundstrom и соавт., 1996 А), SED (Sundstrom и соавт., 1996 В) и SHE (1ENF) (Hakansson и соавт., 2000). Последовательность SEA была наиболее близка SEE и идентична ей на 80%. Последовательность SED была идентична SEE на 60%, а SEH была идентична SEE на 50%.
Создание модели осуществляли с использованием модуля HOMOLOGY в программе INSIGHTII (MSI, San Diego). Структуры трех суперантигенов SEA, SED и SHE выравнивали и определяли структурные консервативные области (SCR) (фиг.3). Эти области обычно картировали в данных молекулах как регулярные [периодические] вторичные структуры. Необработанную последовательность SEA/E-18 нагружали и скрепляли с SCR SEA-структуры (фиг.5). Использовали гомологичную модель 1SXT для SEA, за исключением первых девяти остатков на N-конце, где использовали 1ESF. Области между SCR в большинстве случаев представляли собой гибкие выпетливающие области, и были построены из SEA и SED. Большинство этих петлей были построены из SEA за исключением остатков Gln19, Ile140, Asp141, Lys142, Ser189, Gly191, Asp200, Pro206, Asp207 и Leu224, которые были построены из SED. Некоторые области в SCR продемонстрировали большее сходство последовательности с SED и были поэтому выстроены с использованием SED в качестве структурной матрицы (Ile37, Glu49, Asn50, Thr51, Leu52, Ser195, и Thr218) (фиг.3).
Вследствие того факта, что SEA использовали в качестве структурной матрицы для большинства остатков в SEA/E-18 трудности не возникали в случае происходящего перекрывания боковых цепей. Точки сплайсинга перед и после SCR были репарированы. Сначала были ослаблены замещенные боковые цепи, и затем моделирование энергетической минимизации и молекулярной динамики, с использованием стандартных протоколов в HOMOLOGY, ослабило все боковые цепи в SCR. Выпетливающие области ослабляли по одной во время использования первых 1000 стадий энергетической минимизации с последующими 1000 стадиями молекулярной динамики. Этот протокол усовершенствования был применен сначала к выпетливающим боковым цепям, а затем - ко всем атомам в данной петле. Во всех моделях использовали силовое поле CVFF с силовой постоянной 100 ккал/Е2 с применением временного шага в 2 fs.
Конечную модель тестировали на наличие непригодных областей с использованием модуля PROSTAT в INSIGHTII. Устанавливали отсутствие непригодных областей. При сравнении с SEA внутреннее устройство упакованного белка существенно не отличалось. Все остатки оставались в разрешенных областях на ramachandran-графике. Совмещение 1SXT с данной моделью дает RMSD в 0.4Е, если сравнивали Сα-атомы. Главное отличие между данными двумя структурами просматривается для петли β9-β10 (остатки His187-Thr193) (фиг.5).
Используя в качестве матрицы модель SEA/E-18 создали новые модели новых суперантигенных вариантов. Специфичные аминокислотные остатки заменяли непосредственно в данной модели. Наиболее подходящую конформацию боковой цепи выбирали с использованием простого поиска пространственных затруднений с последующей короткой энергетической минимизацией.
Пример 4
Культивирование и выделение очисткой
Химеры C215FabSEA/E экспрессировали в виде слитых белков в клетках E.coli штамма K12 UL635 с использованием плазмиды с промотором Lac UV-5, индуцируемым IPTG, и геном устойчивости к канамицину.
Вкратце, бактерии из замороженного (-70°) стокового раствора в 20%-и глицерине инкубировали при 25°С в течение 22-24 ч. во встряхиваемых колбах, содержащих (на 1 литр) 2,5 г (NH4)2SO4, 3 г KH2PO4, 2 г K2HPO4, 0,5 г натрийцитрата, 1 г MgSO4•H2O, 0,05 г канамицина, 12 г моногидрата глюкозы и 1 мл раствора редких элементов, но без Na2MoO4•2H2O. Клетки выращивали до ОП620 2-3, и для инокуляции 1-литрового ферментера (Belach Bioteknik, Швеция) со стартовым объемом 800 мл использовали 10 мл прокультивированной среды. Среда в ферментере содержала (на 1 литр) 2,5 г (NH4)2SO4, 9 г KH2PO4, 6 г K2HPO4, 0,5 г натрийцитрата, 1 г MgSO4•7H2O, 0,05 г канамицина, 23,1 г моногидрата глюкозы и 1 мл раствора редких элементов, как указано выше. В период ферментации рН поддерживали на постоянном уровне 7,0 путем титрования 25%-м NH3, аэрация составляла 1 литр/минуту, а температура равнялась 25°С. На стадии периодического культивирования насыщение O2 поддерживали на уровне 30%, регулируя перемешивание от 400 об./мин. до 2000 об./мин., а во время периодического культивирования с подпиткой - регулируя подачу глюкозы (60% w/v). Образование продукта индуцировали при ОП620 нм=45 путем добавления 0,1 мМ изопропил-β-D-тиогалактопиранозида (IPTG). По окончании ферментации клетки удаляли центрифугированием при -20°С перед выделением очисткой.
Процедура выделения очисткой была разделена на три стадии. Вначале из культурального супернатанта удаляли ДНК с помощью 0,19% полиэтиленимина (w/v) в 0,2 М NaCl, рН 7,4, применяя для этого перистальтический насос со скоростью потока 12 мл/мин. После центрифугирования при 7500 x g в течение 30 мин., собирали супернатант.
Его загружали в 60 мл-овую быстропроточную колонку с белок-G-сефарозой 4 (Amersham Pharmacia Biotech) со скоростью потока 14 мл/мин. Загруженную колонку промывали с использованием PBS, и элюирование осуществляли с помощью 100 мМ уксусной кислоты, 0,25% Твина 20, рН 3,0. Собирали элюируемый продукт, и доводили рН, составляющей на 1,5 единиц ниже до предполагаемой изоэлектрической точки, с помощью 1 М NaOH, фильтровали (0,2 мкм) и разбавляли в четыре раза 0,25%-м Твином 20. Удаляли деградированные варианты с использованием ионообменной хроматографии. Ионную силу в полученном образце доводили до 2 mS/см, а примененная колонка представляла собой SP-Sepharose-HP, Hiload 16/10 (Amersham Pharmacia Biotech). Элюцию осуществляли со скоростью потока 4,0 мл/мин в течение 50 мин с использованием линейного градиента 0-55% буфера В, 100 мМ NaAc, 400 мМ NaCl, 0,025 % Твина 20, рН 5,0 в буфере А, 10 мМ NaAc, 0,25% Твина 20, рН 5,0.
Пример 5
Серореактивность
Реактивность между полученными суперантигенными вариантами и человеческими антителами против SEA измеряли в сцинтилляционном анализе близкого расстояния (SPA).
В планшете для микротитрования (OptiPlate, Packard Instruments) PVT-шарики, покрытые стрептавидином, 150 мкг шариков/лунку (Amersham Pharmacia Biotech), инкубировали в течение 30 мин при комнатной температуре с биотинконъюгированными F(ab)2-фрагментами антимышиных IgG, 3 мкг/мг шариков. Указанные шарики прединкубировали с суперантигенами, конъюгированными с С215Fab, в ряду разведений 1:2, где наивысшая конечная концентрация в лунках составляла 40 нМ. В конечном итоге их инкубировали с 1 нМ 125I-конъюгированных аффинно очищенных человеческих антител против SEA, а уровень 0-сцинтилляции измеряли в Top-счетчике (Packard Instruments).
Реактивность суперантигенных вариантов с человеческими антителами против SEA измеряли также в твердофазном иммуноферментном анализе, ELISA (Cavallin и соавт., 2000). Полученные результаты были аналогичны результатам, полученным в SPA.
Пример 6
Биологическая функция
Способность индуцировать суперантигенную антителозависимую клеточную цитотоксичность, SADCC, и суперантигензависимую клеточную цитотоксичность, SDCC, сравнивали в стандартном 4 ч анализе по 51Cr-высвобождению.
Вкратце, мишени, которые использовали для SDCC, представляли собой Raji-клетки В-клеточной лимфомы человека, а мишени для SADCC представляли собой Colo205-клетки колоректальной карциномы человека. Эти клетки метили с помощью 51Cr и разбавляли до концентрации 50000 клеток/мл в V-образных лунках планшета для микротитрования. В качестве эффекторных клеток использовали реагирующую SEA линию Т-клеток человека при соотношении эффектора и мишени 45:1 для SADCC и 30:1 - для SDCC. Sag-варианты добавляли в концентрациях 10-9-10-16 для SADCC и 10-7-10-14 для SDCC. Собирали супернатанты и измеряли высвобождение 51Cr в Top-счетчике (Packard Instruments). Процент специфической цитотоксичности вычисляли как 100 х [(cpm при высвобождении в опыте - cpm при фоновом высвобождении)/(cpm при суммарном высвобождении - cpm при фоновом высвобождении)].
Пример 7
Идентификация эпитопов для антител
У пациентов с уже существующими антителами против суперантигенов клиническое их применение осложняется, требуя корректировки их доз в лечении (Alpaugh и соавт., 1998). Другой подход к ограничению воздействия уже существующих антител заключался в модификации области суперантигена, ответственной за связывание с Т-клеточным рецепторном (Antonsson, и др., 1997). Однако настоящее изобретение дополнительно улучшало терапевтический потенциал суперантигенов путем использования генной инженерии для удаления эпитопов данного суперантигена для антител.
Установлено, что SEE, по сравнению с SEA, проявляет значительно сниженную реактивность с антителами (Antonsson и соавт., 1997). К сожалению, при этом снижении происходит также явное уменьшение способности к уничтожению злокачественной опухоли, в том случае, если SEE слит с реагирующим со злокачественной опухолью Fab (Antonsson и соавт., 1997). Поэтому были исследованы химерные конструкции SEA и SEE. При введении соответствующих аминокислот из SEA по четырем позициям TCR-связывающей области SEE, получали требуемые свойства. Эти замены: Arg20Gly, Asn21Thr, Ser24Gly и Arg27Lys (область А) в SEE, приводили к химере SEA/E-18 (фиг.4) (Antonsson и соавт., 1997). Для данной химеры обнаруживается более чем 50%-е снижение реактивности с антителами, как и в SEE, несмотря на сохранение эффективного уровня цитотоксичности как и в SEA. Кроме того, для уменьшения сродства между данным суперантигеном и MHC класса II, которое снижает SDCC и тем самым улучшает терапевтическое окно, SEA/E-18 содержит также замену Asp227Ala (Abrahmsen и соавт., 1995).
Для дальнейшего снижения способности человеческих антител против SEA распознавать SEA/E-18 в суперантигенах определяли антителосвязывающие эпитопы. Используя иммобилизованные антитела против SEA, улавливали пептид/фрагменты при частичной обработке пепсином SEAwt или SEA/E-18. После выделения очисткой полученные пептидные последовательности идентифицировали с использованием LC-MS (фиг.1). Таким образом, в аминокислотной последовательности локализовали потенциальные области, участвующие в распознавании антител. Примечательно, что большинство извлекаемых пептидов оказывались локализованными вокруг областей, известных по взаимодействию с MHC класс II (Abrahmsen и соавт., 1995) (фиг.2 и фиг.6). Для местонахождения остатков в идентифицированных пептидах, расположенных на поверхности, использовали трехмерную структуру SEA (Schad и соавт., 1995; Sundström и соавт., 1996) и компьютерную модель SEA/E-18 (фиг.5), основанных на кристаллографических характеристиках SEA (Schad и соавт., 1995; Sundström и соавт., 1996 А). Были идентифицированы следующие остатки, которые являются вероятными кандидатами в качестве образующих связывающие антитела эпитопы: Glu34, Lys35, Glu39, Asn40, Lys41, Glu42, Asp44, Asp45, Glu49, Lys74, Asp75, Asn78, Lys79, Lys81, Lys83, Lys84, Asp173, His187, Ser189, Glu190, Gln204, Lys217, Asn220, Glu222, Asn223, His225 и Asp227 (Таблица 1).
Эти остатки были последовательно заменены для уменьшения связывания с антителами. Непрерывно создавались новые компьютерные модели с улучшенными в дальнейшем Sag-вариантами, чтобы подтвердить и сравнить полученные последние результаты. В частности, изучалось влияние боковых цепей и идентифицировались изменения, воздействующие на стабильность белка.
Пример 8
Модификация суперантигена для уменьшения серореактивности
Уровни антителосвязывающих идентифицируемых остатков были первоначально охарактеризованы в SEA/E-18 по двум-шести одновременным заменам. Таким образом, получали SAg-варианты SEA/E-62 (Lys217Thr, Asn220Ala, Glu222Thr, Asn223Ala, His225Ala) (область Е), SEA/E-63 (Ser189Asp, Glu190Ala, (область D), SEA/E-64 (Glu34Lys, Lys35Glu, Glu39Lys, Asn40Ser, Lys41Glu, Glu42Lys) (область В), SEA/E-65 (Lys79Glu, Lys81Glu, Lys83Glu, Lys84Glu) (область С), SEA/E-74 (Asp44Ala, Asp45Ala, Glu49Thr) (область В) и SEA/E-75 (Lys74Thr, Asp75Ala, Asn78Ser) (область С) (Таблица 1, фиг.4).
Для изучения возможности распознавания антителами против SEA из пула человеческих IgG, различных SAg-вариантов был разработан сцинтилляционный анализ близкого расстояния (SPA). По сравнению с SEA/E-18 все модифицированные варианты оказались распознаваемыми в меньшей степени (Таблица 1). Наиболее существенное уменьшение в связывании было вызвано заменами, произведенными в SEA/E-65. В SPA-анализе наблюдалось снижение более чем на 40% (фиг.7). Однако многие замещения также вызывали снижение в уровне продукции в E.coli и, кроме того, уменьшалось и биологическая активность. При тщательном изучении замен авторы настоящего изобретения смогли идентифицировать ответственные остатки в каждом варианте и исключить или модифицировать их для получения лучших характеристик. Вообще, уровень продукции повышается при гидрофильных заменах по сравнению с более гидрофобными заменами.
Уменьшение в связывании антител синергически повышалось при объединении вариантов, таких как SEA/E-91, составленного из SEA/E-63, SEA/E-65, и модифицированного SEA/E-74 (с Asp45 дикого типа) (Таблица 1). Вариантом с наиболее выделяющимся результатом в SPA-анализе и сниженным почти на 70% связыванием, по сравнению с SEA/E-18, оказался SEA/E-110, сочетание SEA/E-63, SEA/E-75 и модифицированного SEA/E-62 (SEA/E-97), SEA/E-64 (SEA/E-108), SEA/E-65 (SEA/E-84), и SEA/E-74 (Asp45 дикого типа) (фиг.7, таблица 1). Модификациями, ответственными за наибольшее снижение связывания антител, оказались в SEA/E-109 (Glu34Ser, Glu39Ser, Asn40Ser, Lys41Glu, Glu42Lys, Asp44Ala, Glu49Thr, Lys74Thr, Asp75Ala, Asn78Ser, Lys79Glu, Lys81Glu, Lys83Ser, Lys84Ser), сочетания SEA/E-75 и модифицированного SEA/E-64 (SEA/E-108), SEA/E-65 (SEA/E-84) и SEA/E-74 (Asp45 дикого типа). Это обусловлено тем, что все суперантигенные варианты, содержащие такие замены, обнаруживали хорошее снижение в SPA-анализе.
Таким образом, остатки, замещаемые в SEA/E-62, SEA/E-64, SEA/E-65 и SEA/E-74, по сравнению с SEA/E-18, приводили к снижению реактивности с антителами между 20 и 40%, (Таблица 1).
Пример 9
Замены, влияющие на уровень продукции
Как указано выше, некоторые из замен на поверхности суперантигена приводили к пониженному уровню продукции в E.coli. Многие сочетания таких замен было даже невозможно создать. Поэтому было решено исследовать альтернативные модификации остатков, по-видимому вызывающие снижение продукции. Замены, которые влияли на продукцию без уменьшения связывания с антителами, далее не исследовались. Вместо них использовались аминокислотные остатки дикого типа.
В исходной группе суперантигенных вариантов остаток Lys35 в SEA/E-64 влиял на уровень экспрессии отрицательно. При использовании остатка дикого типа в положении 35, наряду с сериновыми заменами Glu34 и Glu39, дающими SAg-вариант SEA/E-108, происходило увеличение синтеза с 23 мг/л до 30 мг/л. Однако сниженная реактивность с антителами сохранялась. Замены на глутаминовую кислоту остатков Lys79, Lys81, Lys83 и Lys84 в SEA/E-65 приводили к уровню синтеза лишь 1,5 мг/л. Из-за того, что эффект реактивности с антителами снижался на 43%, по сравнению с SEA/E-18, была предпринята попытка выявить лучшие замены. Наилучшее сочетание с точки зрения продукции и сниженной реактивности с антителами, обнаружено для SEA/E-18 с сериновыми остатками в положении 83 и 84 и сохраненной глутаминовой кислотой в положениях 79 и 81 (Таблица 1). По сравнению с SEA/E-18 уровень синтеза оказывался выше в десять раз, а реактивность с антителами снижалась на 41% (Таблица 1). Уровень синтеза оказывается также сниженным более чем в десять раз при заменах Lys217Thr, Asn220Ala, Glu222Thr, Asn223Ala, His225Ala и Asp227Ala в SEA/E-62 до 1,0 мг/л. Вместе с тем, заменив аланиновые замены на сериновые остатки, что приводило к SEA/E-97, получали уровень синтеза в 48 мг/мл (Таблица 1).
Интересно отметить, что при объединении SEA/E-65 с большинством вариантов, такими как SEA/E-63 и модифицированным SEA/E-74, также как и SEA/E-91, возвращался низкий уровень продукции до 12 мг/л (Таблица 1). С другой стороны, уровень экспрессии суперантигенного варианта SEA/E-110 составлял, соответственно, лишь 0,5 мг/л и 14 мг/л. Но уровень продукции SEA/E-110 возрастал до 30 мг/л при удалении замен Asp174Ala, His87Ala, Ser188Thr, Ser189Asp, Glu190Ala и Gln204Thr, создающих SEA/E-120 (Таблица 1).
Введение большого числа замен в суперантиген может создать проблемы для экспрессии E.coli. Для этого существует, по меньшей мере, три различных механизма; сниженная термодинамика, нарушение структуры естественной укладки или вновь вводимые протеолитические сайты. Хотя целью данного исследования было удаление антигенных эпитопов с поверхности, которые бы с большой вероятностью не мешали каким-либо структурным основным цепям, всегда существует вероятность того, что для поддержания стабильности новые структуры будут зависеть от других остатков, а не от конструкции дикого типа. Поэтому постоянно создавались новые компьютерные модели для прогнозирования или подтверждения местонахождения замененных остатков в данной новой структуре. Таким образом, авторы настоящего изобретения могли идентифицировать ответственные остатки в ранних суперантигенных вариантах, обусловивших трудности, в отношении, например, экспрессионных уровней, и создать улучшенные варианты либо с остатками дикого типа, либо с лучшими заменами (Таблица 1).
В заключение, для осуществления более высокого уровня синтеза необходимо остатки Lys83, Lys84, Asn220, Asn223, His225 и Asp227 заменить на серин, но не на аланин. Кроме того, чтобы исключить снижение экспрессионных уровней, необходимо сохранить остатки Lys35, Asp173, His187, Ser188, Ser189, Glu190 и Gln204.
Пример 10
Оценка биологической функции у разных Sac-вариантов
Поскольку суперантигены первоначально создавались для противоопухолевой терапии (Dohlsten и соавт., 1994), было важно исключить в новых суперантигенных вариантах замены, снижающие противоопухолевую цитотоксичность. Поэтому, у всех новых суперантигенных вариантов в SADCC-анализе измеряли способность, которая опосредует эту противоопухолевую цитоксичность (фиг.3). Кроме того, в SDCC-анализе измеряли эффективность суперантигенов опосредовать уничтожение Т-клетками гибель клеток, экспрессирующих молекулы MHC класса II, имеющую своим результатом системную цитотоксичность, которая могла бы вызвать побочные эффекты (фиг.3). Для клинического использования SDCC следует, вероятно, снизить, чтобы увеличить терапевтическое окно.
Самая первая группа SAg-вариантов обладало тем же уровнем специфической противоопухолевой цитотоксической потенции, что и SEA/E-18 (Таблица 1). Исключением были SEA/E-75 с заменами Lys74Thr, Asp75Ala и Asn78Ser, активность которого оказывалась десятикратно сниженной, и SEA/E-64, с заменами Glu34Lys, Lys35Glu, Glu39Lys, Asn40Ser, Lys41Glu и Glu42Lys, активность которого оказывалась пятикратно сниженной по сравнению с SEA/E-18 (Таблица 1). Интересно, что сниженная активность у SEA/E-75 наблюдалась лишь в данном варианте, но в сочетании с последующими заменами, например, у SEA/E-109, обнаруживалась полная активность (Таблица 1). Кроме того, SDCC-активность неизменна у SEA/E-75 при сравнении с SEA/E-18. Поэтому вероятно замены Lys74Thr, Asp75Ala и Asn78Ser нарушают взаимодействия, существенные исключительно для антителозависимой цитотоксичности.
Большинство описанных здесь суперантигенных вариантов демонстрировали отчетливое снижение SDCC. По сравнению с SEA/E-18 у исходных вариантов SEA/E-62, SEA/E-63, SEA/E-64, SEA/E-65 и SEA/E-74 наблюдалось несущественное уменьшение SDCC-активности.
Все суперантигенные варианты содержали замещенный остаток Asp227Ala или Ser. Известно, что данное замещение снижает сродство к молекулам MHC класса II в 100 раз, а следовательно, и SDCC-активность (Abrahmsen и соавт., 1995). Однако, поскольку SAg-вариант SEA/E-109 c N-концевыми заменами демонстрировал большее уменьшение по сравнению с SEA/E-18, чем SEA/E-113 с С-концевыми заменами, это свидетельствовало о том, что в SEA/E-109 дополнительные остатки были заменены, которые существенны для SDCC, и, вероятно, связаны с MHC класса II (фиг.8).
Таким образом, остатки, которые обусловливали наибольшее снижение, являлись Lys79Ser и Lys81Ser в SEA/E-83, и замена Asp45Ala в SEA/E-74. Большинство этих замен располагается вокруг остатков, которые, как было показано раньше, взаимодействуют с MHC класса II (Abrahmsen и соавт., 1995).
Пример 11
Создание нового суперантигенного варианта
С целью создания оптимального суперантигенного варианта были объединены все благоприятные замены, приводящие к наилучшему SEA/E-120 (фиг.4 и фиг.9).
Во-первых, были собраны все подходящие С-концевые модификации, т.е., Asp173Ala, Ser189Thr, Glu190Ala, Lys217Thr, Asn220Ser, Glu222Thr, Asn223Ser, His225Ser и Asp227Ser, образующие вместе с Gln204Thr SAg-вариант SEA/E-113. Данный вариант обнаруживал ожидаемое снижение реактивности с антителами против SEA и приемлемый уровень экспрессии, но с несколько уменьшенной биологической активностью (Таблица 1, фиг.7, и фиг.8А, и фиг.8В). Все подходящие N-концевые замены, т.е., Glu34Ser, Glu39Ser, Asn40Ser, Lys41Glu, Glu42Lys, Asp44Ala, Glu49T, Lys74T, Asn78Ser, Lys79Glu, Lys81Glu, Lys83Ser и Lys84Ser были собраны в SEA/E-109. В этом суперантигенном варианте наблюдалось явное снижение анти-SEA-реактивности, наряду с высоким уровнем экспрессии и даже улучшенным биологическим профилем (Таблица 1, фиг.7 и фиг.8А и фиг.8В). Однако при создании объединения этих двух вариантов SEA/E-113 и SEA/E-109 в SEA/E-110, происходила существенная утрата и продукции и биологической функции (Таблица 1). Биологическая эффективность полностью восстанавливалась, если в SEA/E-115 вновь использовали остатки дикого типа Ser189, Glu190 и Gln204 (Таблица 1), но уровень продукции все же оставался низким. Молекулярное моделирование данного варианта дает основание полагать, что остатки Asp173, His187 и Ser188 могли бы быть существенными для стабилизации укладки, а впоследствии приводить и к более высокой продукции.
Для оценки этих остатков было создано несколько различных сочетаний, проводящих к SEA/E-118, SEA/E-119, SEA/E-120, SEA/E-121 и SEA/E-122 (Таблица 1). Самый высокий синтез получали для SEA/E-120 с остатками дикого типа во всех трех позициях. Вместе с ранее созданными SEA/E-21, SEA/E-74, SEA/E-97, SEA/E-108 и SEA/E-109 они представляют собой SAg-варианты с уровнями экспрессии, превышающими 20 мг/л (Таблица 1). Существенных различий в биологической активности или реактивности с антителами между этими вариантами не отмечено.
Создание нового конъюгата
SEA/E-120 генетически сливали с Fab-компонентом антитела, реагирующего с опухолью, которое представляет собой 5Т4 (Dohlsten и соавт., 1994) (фиг.10).
Антиген 5Т4 экспрессируется во множестве разнообразных опухолей, таких как немелкоклеточная злокачественная опухоль легкого, злокачественная опухоль молочной железы, гипернефроидная злокачественная опухоль, злокачественная опухоль поджелудочной железы, злокачественная опухоль яичника и злокачественная опухоль прямой кишки. Замены в последовательности дикого типа 5Т4 также осуществляли для получения более высокого выхода. В тяжелой цепи: His41Pro, Ser44Gly, Ile69Thr и Val113Gly; в легкой цепи: Phe10Ser, Thr45Lys, Ile63Ser, Phe73Leu, Thr77Ser, Leu78Val и Leu83Ala.
Несмотря на подробное описание настоящего изобретения и его преимуществ, следует иметь в виду, что различные изменения, замены и вариации могут быть внесены в него, не выходя за рамки существа и объема настоящего изобретения, которые определены прилагаемой формулой изобретения. Кроме того, объем настоящей заявки не предполагается ограничить конкретными вариантами осуществления процесса, устройства, изготовления, композиции, средств, методов и стадий, представленных в настоящем описании. Каждому специалисту в данной области техники из представленного описания настоящего изобретения очевидно, что процессы, устройства, производство, композиции, средства, методы или стадии, ныне существующие либо созданные позже, которые, в основном, выполняют одно и то же назначение или по существу достигают одного и того же результата, в качестве описанных здесь соответствующих вариантов осуществления, могут быть использованы в соответствии с настоящим изобретением. Следовательно, прилагаемую формулу изобретения предполагается включить в объем таких процессов, устройств, производства, композиций, средств, методов или стадий.
Каждому специалисту в данной области ясно, что настоящее изобретение хорошо адаптировано для осуществления поставленных целей и получения указанных, а также присущих ему результатов и преимуществ. Композиции, способы, операции и методики, описанные здесь, представляют предпочтительные варианты осуществления настоящего изобретения и, как подразумевается, являются иллюстративными, а не ограничивающими объем изобретения. В этом отношении изменения и иные использования будут иметь место у специалистов в данной области техники, что и включено в существо настоящего изобретения или определяются пунктами патентной формулы.
Все патенты и публикации, указанные в описании, свидетельствуют об уровне специалистов в данной области техники, которые имеют отношение к настоящему изобретению. Все патенты и публикации, включенные здесь путем ссылки, так же как и индивидуальные публикации, были в связи с этим и индивидуально указаны для включения путем ссылки.
U.S. Patent 4554101.
U.S. Patent 5221605.
U.S. Patent 5238808.
U.S. Patent 5798208.
U.S. Patent 5830650.
U.S. Patent 5220007.
U.S. Patent 5284760.
U.S. Patent 5354670.
U.S. Patent 5366878.
U.S. Patent 5389514.
U.S. Patent 5635377.
U.S. Patent 5789166.
U.S. Patent 5446128.
U.S. Patent 5710245.
U.S. Patent 5840833.
U.S. Patent 5859184.
U.S. Patents 5440013.
U.S. Patent 5618914.
U.S. Patent 5670155.
U.S. Patent 5475085.
U.S. Patent 5929237.
U.S. Patent 5672681.
U.S. Patent 5674976.
U.S. Patents 4608251.
U.S. Patent 4601903.
U.S. Patent 4599231.
U.S. Patent 4599230.
U.S. Patent 4596792.
U.S. Patent 4578770.
Abrahmsen L., et al. EMBO J. 14:2978-86, 1995.
Alpaugh R. K., et al. Clin Cancer Res. 4:1904-14, 1998.
Antonsson P., et al. J. Immunol 158:4245-51, 1997.
Bangham et al., J. Mol. Biol., 13:238-252, 1965.
Bird et al., Science. 242:423-6, 1988.
Braisted et al., Proc Natl Acad Sci USA. 93(12):5688-92, 1996
Burks et al., Proc Natl Acad Sci USA. 94(2):412-7, 1997.
Capaldi et al., Biochem. Biophys. Res. Comm., 76:425, 1977.
Cavallin А., et al. J. Biol Chem. 275:1665-72, 2000.
Cunningham et al., Science. 244(4908):1081-5, 1989.
Davis et al., Basic Methods in Molecular Biology, 1986.
Dohlsten М., et al. Proc Natl Acad Sci USA. 91:8945-9, 1994.
DRUG CARRIERS IN BIOLOGY AND MEDICINE, G. Gregoriadis ed. (1979) рр.287-341.
Hakansson, М. et al., J. Mol Biol. 302:527-37, 2000.
Harlow, et al. Antibodies: А Laboratory Manual, 1988.
Johannesson et al., J. Med. Chem. 42:601-608, 1999.
Kaneda et al., J. Biol Chem., 264 (21):12126-12129, 1989.
Kato et al., J. Biol Chem., 266(6):3361-3364, 1991.
Nicolau et al., Methods Enzymol., 149:157-176, 1987.
Papageorgiou А. С. et al., Trends in Microbiology 8:369-375, 2000.
Remington's Pharmaceutical Sciences, 15th Edition, Chapter 61, pages 1035-1038 and 1570-1580.
Sambrook et. al., In: Molecular Cloning: А Laboratory Manual, 2d Ed., 1989.
Schad E. М., et al., EMBO J. 14:3292-3301, 1995.
Short et al., J. Biol Chem. 270(48):28541-50, 1995.
Sundstrom М, et al., EMBO J. 15:6832-40, 1996 А.
Sundstrom М., et al., J. Biol Chem 271:32212-16, 1996 В.
Szoka and Papahadjopoulos, Proc. Natl. Acah. Sci., 75:4194-4198, 1978.
Vita et al., Biopolymers 47:93-100, 1998.
Warren et al., Biochemistry 35 (27):8855-62, 1996.
Weisshoff et al., Eur. J. Biochem. 259:776-788, 1999.
Wells et al., Methods. 10(1): 126-34, 1996.
Wong et al., Gene, 10: 87-94, 1980.
Yelton et al., J. Immunol. 155(4):1994-2004, 1995.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНЪЮГАТ, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ АКТИВИРОВАТЬ ИММУННУЮ СИСТЕМУ, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ВКЛЮЧАЮЩАЯ УКАЗАННЫЙ КОНЪЮГАТ | 1997 |
|
RU2198895C2 |
| КОНЪЮГАТ ДЛЯ СТИМУЛИРОВАНИЯ ИММУННОЙ РЕАКЦИИ ПРОТИВ КЛЕТКИ-МИШЕНИ, СПОСОБ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ У МЛЕКОПИТАЮЩЕГО | 1995 |
|
RU2183215C2 |
| РАСТВОРИМЫЙ КОНЪЮГАТ АНТИТЕЛА, СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКОГО ЗАБОЛЕВАНИЯ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1991 |
|
RU2125889C1 |
| АНТИТЕЛА К АНТИГЕНУ ЭПИТЕЛИАЛЬНЫХ ОПУХОЛЕЙ ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА ЧЕЛОВЕКА, РОДСТВЕННОМУ АЛЬФА 6 БЕТА 4 ИНТЕГРИНУ | 2000 |
|
RU2266298C2 |
| АНТИТЕЛА ПРОТИВ KIT И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2681730C2 |
| КОНСТРУКЦИИ, НАПРАВЛЕННЫЕ НА КОМПЛЕКСЫ ПЕПТИДА AFP/МНС, И ВИДЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2754041C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ У ПАЦИЕНТА С НЕАДЕКВАТНЫМ ОТВЕТОМ НА ИНГИБИТОР TNF-АЛЬФА | 2008 |
|
RU2489166C2 |
| КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО (ADC), КОТОРЫЕ СВЯЗЫВАЮТСЯ С БЕЛКАМИ FLT3 | 2016 |
|
RU2739617C2 |
| КОМПОЗИЦИЯ АНТИТЕЛ К HER2 | 2005 |
|
RU2361880C2 |
| НОВОЕ АНТИТЕЛО СО СПЕЦИФИЧНОСТЬЮ К ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ ТОЛСТОЙ КИШКИ | 2001 |
|
RU2268068C2 |
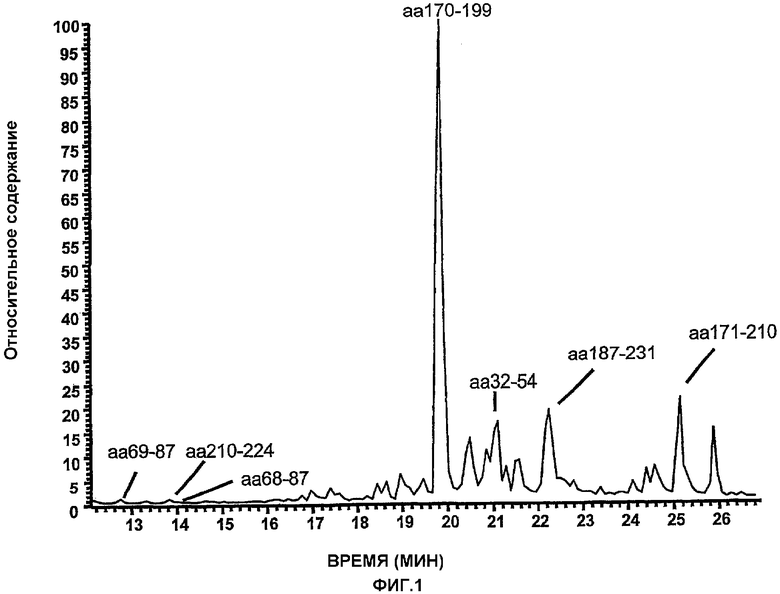



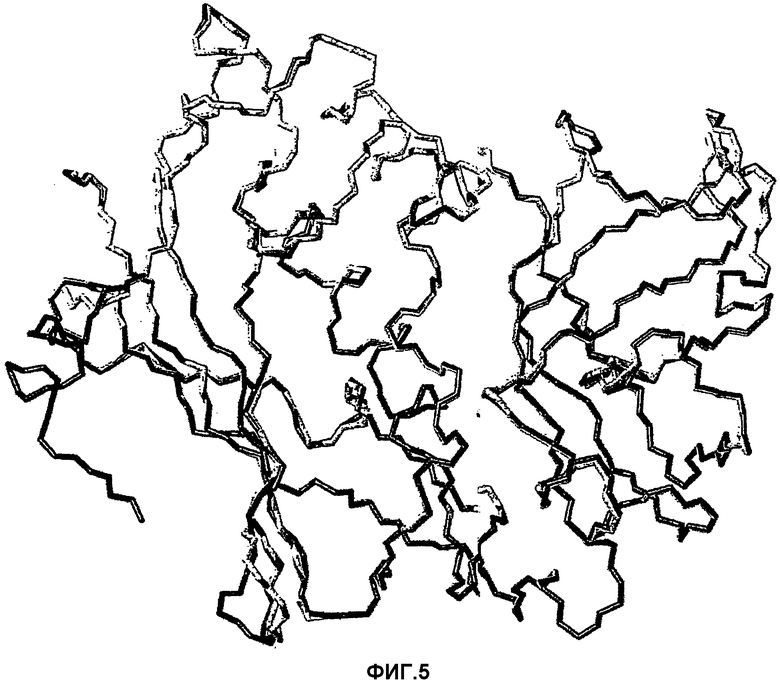
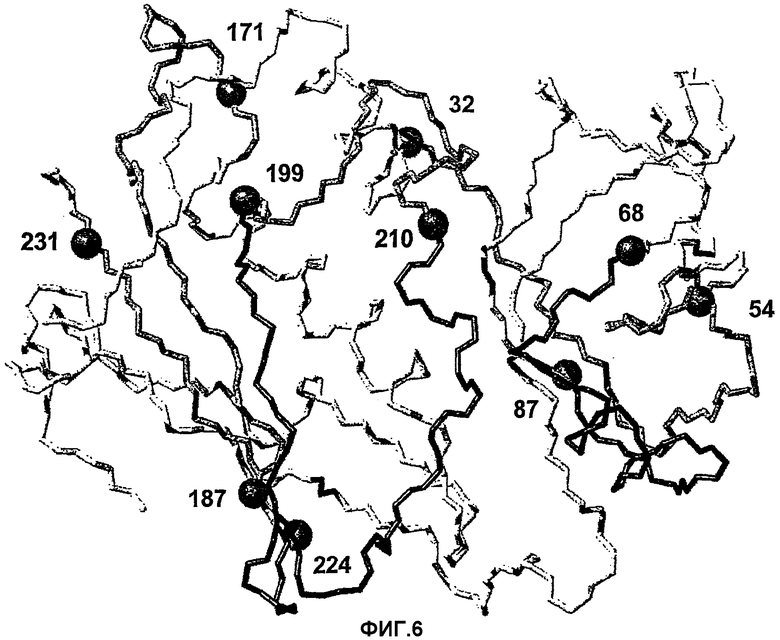
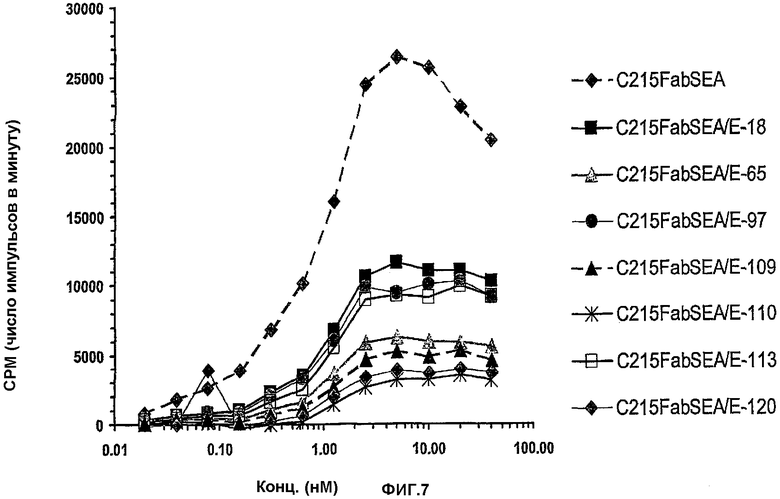
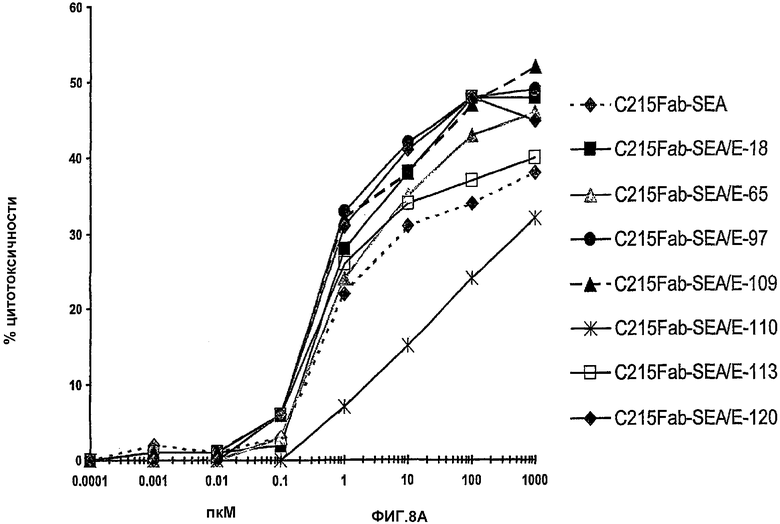
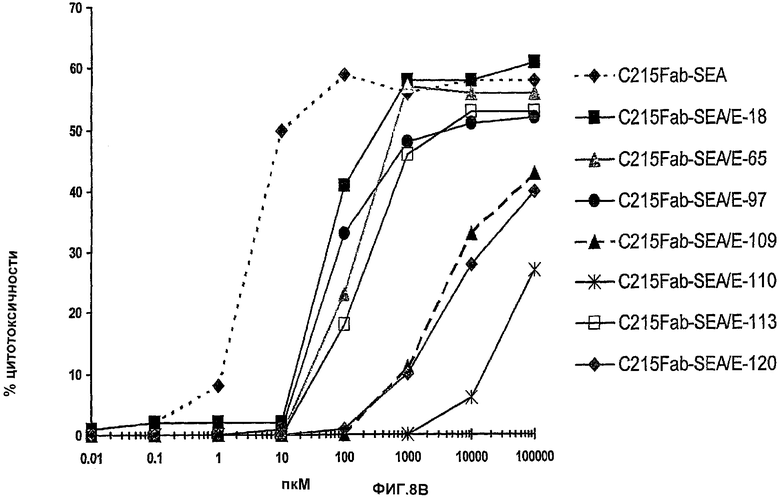

Изобретение относится к конъюгатам, используемым для лечения злокачественной опухоли. Конъюгат включает стафилококковый или стрептококковый суперантиген дикого типа или модифицированный суперантиген и антительную составляющую. Бактериальный суперантиген модифицирован для уменьшения серореактивности при сохранении его антигенной активности. Аминокислотная последовательность суперантигена включает области А-Е, определяющие связывание с TCR и молекулами МНС класса II. Изобретение касается применения конъюгата для получения лекарственного противоопухолевого средства, а также для получения фармацевтической композиции. Использование конъюгата по изобретению обеспечивает активацию иммунной системы, следствием которой обеспечивается лечение млекопитающего со злокачественной опухолью. 8 н. и 59 з.п. ф-лы, 11 ил., 1 табл.
аминокислотная последовательность указанного суперантигена замещена по меньшей мере таким образом, что один или несколько аминокислотных остатков в положениях, выбранных из группы, состоящей из положений 74, 75, 78, 79, 81, 83 и 84 области С в SEA или SEE, замещены одним или несколькими отличными аминокислотными остатками так, что замещенный суперантиген обладает пониженной серореактивностью по сравнению с суперантигеном, из которого он получен,
и в котором указанная антительная составляющая представляет собой полноразмерное антитело или любой другой антигенсвязывающий активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью.
и в котором указанная антительная составляющая представляет собой полноразмерное антитело или любой другой антигенсвязывающий активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью.
аминокислотная последовательность указанного суперантигена по меньшей мере замещена таким образом, что один или несколько аминокислотных остатков в положениях, выбранных из группы, состоящей из положений 34, 35, 39, 40, 41, 42, 44, 49, 188, 189, 190, 217, 220, 222 и 223 области В, D и/или Е в SEA или SEE, замещены одним или несколькими отличными аминокислотными остатками таким образом, что замещенный суперантиген обладает сниженной серореактивностью по сравнению с суперантигеном, из которого он получен,
и в котором указанная антительная составляющая представляет собой полноразмерное антитело или любой другой антигенсвязывающии активный фрагмент антитела, которые нацелены против структуры на поверхности клетки, ассоциированной со злокачественной опухолью.
| W09736932 A1, 09.10.1997 | |||
| WO 9601650 A1, 25.01.1996 | |||
| RU 97111749 А, 22.07.1999. |

