Изобретение относится к области медицины, конкретно экспериментальной медицине (патологической физиологии), и может быть использовано для изучения индивидуальных реакций системы крови у инбредных животных с разным типом высшей нервной деятельности при патологии различного генеза.
Известна роль нервной системы в регуляции кроветворения при чрезвычайных воздействиях различной природы [1]. Однако при этом не проводят сопоставления изменений гемопоэза в зависимости от индивидуальных особенностей высшей нервной деятельности. Особенно это актуально при неврозах с учетом того, что у человека реакция организма зависит, прежде всего, от индивидуальной значимости невротизирующего фактора.
Известен способ изучения реакций системы крови при экспериментальных невротических воздействиях (конфликтной ситуации, депривации парадоксального сна и их сочетания), где у инбредных животных после невротического воздействия проводится изучение кроветворения [2, 3].
Данный способ является близким к заявляемому по технической сущности и достигаемому результату и выбран в качестве прототипа.
Недостатком прототипа является усреднение экспериментальных данных. Это делает не возможным изучение индивидуальных реакций системы при невротических воздействиях.
Задачей, решаемой данным изобретением, является возможность определения индивидуальных реакций системы крови при патологии (в том числе при невротических воздействиях), что поможет выявить и предопределить индивидуальную устойчивость к патологии различного генеза.
Поставленная задача достигается тем, что инбредных мышей разделяют на группы хорошо и плохо обучаемых животных путем типирования при выработке условного питьевого рефлекса со сложной пространственной ориентировкой в 3-звенном Т-образном лабиринте, далее животных подвергают экспериментальным невротическим воздействиям, после которых в течение 1-6 сут у них исследуют сдвиги со стороны системы крови.
Новым в предлагаемом изобретении является предварительное разделение инбредных мышей на типологические группы с учетом индивидуальных особенностей высшей нервной деятельности и изучение изменений гемопоэза в каждой группе животных раздельно.
Проблема индивидуальности психоэмоционального стресса занимает ведущее место в современной медицине. Различные системы организма обладают разной генетически обусловленной резистентностью к стрессорным повреждениям: сердечно-сосудистая система, желудочно-кишечный тракт, гуморальный и клеточный иммунный ответ, гормональная система, соединительнотканные структуры в органах пищеварения и стенок дуги аорты.
Индивидуальная устойчивость к стрессу может значительно варьировать между представителями одного вида животных. В основе центральных механизмов, определяющих устойчивость органов и функциональных систем к эмоциональному стрессу, лежит специфическая организация молекулярных и нейрохимических свойств нейронов головного мозга, в частности нейромедиаторов, особенности NO-системы, а также гипофизарно-адренокортикальной системы [4].
Известно, что система крови одной из первых среди прочих систем гомеостаза реагирует на любое стрессовое воздействие [5]. Показано, что невротические расстройства в эксперименте приводят к нарушениям в системе крови [1].
Вместе с тем, остаются совершенно не изученными основные формы ответа системы крови на эмоционально-стрессовые воздействия у животных с различными типами поведенческого статуса, а также механизмы, лежащие в их основе.
Отличительные признаки проявили в заявляемой совокупности новые свойства явным образом не вытекающие из уровня техники в данной области и неочевидные для специалиста. Идентичной совокупности признаков не обнаружено в проанализированной патентной и научно-медицинской литературе.
Предлагаемый способ может быть использован на практике для индивидуализации изучения патологий различного генеза и создания новых методов их фармакологической коррекции.
Исходя из вышеизложенного, заявляемое изобретение соответствует критериям патентоспособности изобретения "Новизна" и "Изобретательский уровень" и "Промышленная применимость".
Способ осуществляют следующим образом: предварительно животных разделяют по итогам обучения условному питьевому навыку со сложной пространственной ориентировкой в Т-образном лабиринте в течение 2 дней по 2 побежки с интервалом в 1 ч. К группе хорошо обучаемых животных относят особей, у которых средние значения 4 побежек вертикальной активности равны или ниже 12 условных единиц, количества ошибок, совершаемых животным при поиске поилки, равны или ниже 12 условных единиц, времени достижения поилки равны или менее 100 с, времени неподвижности равны или менее 3 с и отсутствуют невротические реакции. Животных, у которых 60% и более этих показателей были выше данных показателей и присутствуют невротические реакции, относят к плохо обучаемым. Затем моделируют экспериментальные невротические воздействия (конфликтная ситуация и депривация парадоксального сна), далее ежедневно в течение 1-6 сут после воздействия у животных исследуют показатели системы крови.
Следует отметить, что в Т-лабиринте выявлены и амбивалентные животные, занимающие среднее положение между хорошо и плохо обучаемыми особями, которые были исключены из дальнейших исследований.
Предлагаемый способ изучен в экспериментах на мышах-самцах линии CBA/CaLac в количестве 1200 штук, массой 20-22 г. Животные первой категории, конвенциональные линейные мыши, получены из питомника отдела экспериментального биомедицинского моделирования ГУ НИИ фармакологии ТНЦ СО РАМН (сертификат имеется).
Выбор вышеуказанных животных (линейных мышей) был продиктован следующими обстоятельствами. Так, современные методы изучения механизмов регуляции кроветворения (в первую очередь культуральные) не могут проводиться на неинбредных животных. В связи с этим перед нами встала задача выбора адекватной модели (линии мышей), которая, с одной стороны, позволяла бы оценивать тип приспособительного поведения мышей, а, с другой, широко использовалась в экспериментальной гематологии. При этом необходимо иметь в виду, что в лабораторных линиях, прошедших процесс селективного отбора, животные реагируют более или менее единообразно, обнаруживая либо низкую, либо высокую способность к обучению. Вместе с тем, в литературе имеются сведения о внутрипопуляционных различиях животных инбредных линий, в частности мышей линии СВА, как в уровне поведенческой активности [6, 7], так и в генотипе [8], которые, даже при высоком уровне генетического единообразия, могут приводить к существенным фенотипическим различиям между особями внутри линии. В связи с вышесказанным для проведения экспериментов нами была выбрана линия мышей СВА.
Выработку и воспроизведение сложного условного питьевого рефлекса у здоровых животных проводят в Т-образном лабиринте в течение 2 дней по 2 побежки с интервалом в 1 ч каждый день. Далее по итогам выработки условного питьевого рефлекса производят разделение животных на хорошо и плохо обучаемых. Затем через неделю после типирования животных подвергают экспериментальным невротическим воздействиям (конфликтная ситуация, депривация парадоксального сна), после которых ежедневно в течение 6 сут исследуют показатели костного мозга и периферической крови.
Изучение общего количества лейкоцитов, содержания ретикулоцитов, подсчет лейкоцитарных формул проводят стандартными гематологическими методами [9]. Мазки крови фиксируют в метаноле в течение 3-5 минут и после высушивания окрашивают по Нохту-Максимову.
Для изучения клеточности костного мозга выделяют бедренную кость мыши, очищают ее от мягких тканей и промывают центральный канал 1 мл 3% раствора уксусной кислоты. Костный мозг суспендируют шприцем через иглы уменьшающегося диаметра. Общее количество миелокариоцитов подсчитывают в камере Горяева. Для приготовления мазков костного мозга содержимое бедренной кости выдавливают на обезжиренное стекло, затем разводят изологичной сывороткой и делают мазок с помощью шлифованного стекла. Высохшие мазки фиксируют в метаноле 3-5 минут и окрашивают азур II-эозином. Миелограмму подсчитывают на 400 клеток, после чего определяют абсолютное содержание клеточных элементов отдельных ростков гемопоэза [9].
Моделями невротических состояний являлись конфликтная ситуация и депривация парадоксального сна [10].
Обработку результатов проводили методом вариационной статистики с использованием t-критерия Стьюдента.
Пример 1.
Первоначально животных типируют в Т-образном лабиринте, затем у интактных животных изучают показатели костного мозга и периферической крови.
Проведенные эксперименты не выявили статистически достоверной разницы между общим количеством кариоцитов (ОКК), содержанием эозинофильных и зрелых форм нейтрофильных гранулоцитов, эритрокариоцитов и моноцитов в костном мозге у хорошо обучаемых и плохо обучаемых мышей в условиях оптимальной жизнедеятельности (табл.1, 2). В то же время у плохо обучаемых животных количество лимфоцитов превосходило (на 54%, Р≤0,05), а число незрелых форм нейтрофильных гранулоцитов, напротив, уступало (на 36%, Р≤0,05) таковым у хорошо обучаемых особей.
При сравнении картины периферической крови мышей с разным типом поведения выявлено, что число палочкоядерных и сегментоядерных нейтрофильных лейкоцитов у хорошо обучаемых животных статистически достоверно превышает таковое у плохо обучаемых особей (на 101% и 43% соответственно, Р=0,043) (табл.3, 4). Межгрупповых различий в содержании эозинофилов, моноцитов, лимфоцитов и ретикулоцитов в циркулирующей крови не обнаружено (табл.3, 4, 5).
Таким образом, в условиях оптимальной жизнедеятельности у пассивных мышей содержание лимфоцитов в костном мозге превосходит, а число нейтрофильных гранулоцитов, напротив, уступает таковым у активных животных. Межгрупповых различий в содержании остальных зрелых и незрелых клеточных элементов в костном мозге и периферической крови не обнаружено. В то же время по уровню поведенческой активности при выработке условного питьевого рефлекса хорошо и плохо обучающиеся особи между собой достоверно различались.
Пример 2.
Первоначально животных типируют в Т-образном лабиринте, затем у них моделируют конфликтную ситуацию, далее производят забор экспериментального материала ежедневно в течение 6 сут после конфликтной ситуации.
Проведенные исследования позволили установить, что конфликтная ситуация оказывает неоднозначное действие на процессы костномозгового кроветворения у хорошо обучаемых мышей линии CBA/CaLac. Так, 1-е сут опыта характеризовались значительным уменьшением общего количества кариоцитов (до 43%, Р=0,043), что являлось следствием статистически достоверного снижения всех типов морфологически распознаваемых нуклеаров в костном мозге в указанный срок (табл.1). Однако в дальнейшем - на 4, 5-е сут исследования наблюдалось накопление ОКК (до 135%, 126%, Р=0,043 соответственно). Увеличение показателя было сопряжено с ростом представительства зрелых форм нейтрофильных (до 130%, Р=0,043) гранулоцитов на 4-е сут и эозинофильных гранулоцитов (до 177%, 230%, Р=0,043 соответственно) на 3, 4-е сут, а также лимфоцитов (до 138%, 147%, 155%, Р=0,043 соответственно) на 3, 4, 5-е сут и эритрокариоцитов (до 173%, 176%, Р=0,043 соответственно) на 4, 5-е сут. Наряду с этим выявлено, что число эозинофильных гранулоцитов превосходило фоновые значения и на 2-е сут опыта. На 6-е сут эксперимента исследованные показатели костного мозга хорошо обучаемых особей были на уровне интактных животных, за исключением сниженного количества незрелых нейтрофильных гранулоцитов (до 70%, Р=0,043).
Плохо обучаемые мыши в условиях конфликтной ситуации характеризовались снижением общего количества кариоцитов на 1, 2, 3 и 5-е сут опыта (до 64%, 57%, Р=0,01, 71% Р=0,028 и 78%, Р=0,043 соответственно ниже интактного контроля) и их увеличением на 6-е сут (до 118%, Р=0,043) (табл.2). Анализ миелограмм позволил установить, что динамика содержания лимфоцитов и эритрокариоцитов в костном мозге в целом повторяла таковую для ОКК. В то же время уменьшение количества незрелых нейтрофильных гранулоцитов отмечалось только на 1, 2-е сут исследования (до 79-65%, Р=0,043 соответственно), а зрелых форм - на 1, 2 и 3-й сут (до 65%, 59%, 76%, Р=0,043 соответственно). В противоположность этому на 4-е сут опыта содержание зрелых форм нейтрофильных гранулоцитов превышало исходный уровень на 32% (Р=0,043), а эозинофильных гранулоцитов - на 77% (Р=0,043).
Сопоставление показателей костномозгового кроветворения у животных, разных по типу поведения, позволило установить, что в условиях конфликтной ситуации у плохо обучаемых мышей клеточность гранулоцитарного, лимфоидного и эритроидного ростков кроветворения статистически достоверно превосходила таковую в аналогичных ростках у хорошо обучаемых особей на 1-е сут опыта (табл.1, 2). На 2, 3, 4, 5-е сут исследования содержание нейтрофильных гранулоцитов, лимфоцитов и эритрокариоцитов в костном мозге плохо обучаемых животных было меньше, чем у хорошо обучаемых мышей. В то время как на 6-е сут опыта ОКК за счет увеличенного числа лимфоцитов было достоверно выше у плохо обучаемых особей.
Результаты проведенных экспериментов показали, что у хорошо обучаемых мышей в условиях конфликтной ситуации на 2, 4, 5-е сут исследования имело место увеличение общего количества лейкоцитов (ОКЛ) (до 129%, 124%, 133%, Р=0,043 соответственно) по сравнению с интактным контролем (табл.3). Обнаруженный лейкоцитоз был сопряжен с ростом представительства сегментоядерных нейтрофильных гранулоцитов на 5-е сут опыта (до 125%, Р=0,043), лимфоцитов на 2, 4, 5-е сут (до 158%, 154%, 157%, Р=0,043 соответственно) и моноцитов на 2, 4-е сут (до 300%, 220%, Р=0,043 соответственно). На 3-й сут наблюдалась лейкопения за счет снижения числа палочкоядерных и сегментоядерных нейтрофильных гранулоцитов и лимфоцитов. Для динамики содержания моноцитов характерно увеличение их числа и на 1-е сут эксперимента (до 210%, Р=0,043) выше интактного контроля. Количество палочкоядерных нейтрофилов было ниже фоновых значений на 2, 4-е сут. Одновременно с этим в крови развивался ретикулоцитоз (2, 3, 4, 5-е сут) с последующим развитием ретикулоцитопении (6-е сут) (табл.5).
У плохо обучаемых мышей (как и у хорошо обучаемых особей) конфликтная ситуация вызывала увеличение ОКЛ в периферической крови на 1, 2, 5-е сут опыта (до 124%, Р=0,01; до 117%, Р<0,05; до 129%, Р=0,037 соответственно), однако на 6-е сут отмечалось снижение ОКЛ (до 77%, Р=0,047) (табл.4). Выявленный лейкоцитоз связан с ростом числа палочкоядерных и сегментоядерных нейтрофилов на 1-е сут исследования (до 163% и 252%, Р=0,043 соответственно) и лимфоцитов на 2, 5-е сут (до 125%, 133%, Р=0,043 соответственно). В противоположность этому на 1, 3, 4 и 6-е сут опыта содержание лимфоцитов статистически достоверно уменьшалось (на 14%, 38,5%, 15% и 42% соответственно ниже интактного контроля). Количество палочкоядерных нейтрофилов на 2, 4-е сут было ниже, а на 3 и 6-е сут выше, чем в интактном контроле. Ретикулоцитопению на 1, 2-е сут исследования сменял ретикулоцитоз на 3, 4, 5-е сут (табл.5).
В дополнение к вышеизложенному следует добавить, что у хорошо обучаемых мышей содержание лейкоцитов и ретикулоцитов в периферической крови статистически достоверно превосходило таковое у плохо обучаемых особей на протяжении всего периода наблюдения (табл.3, 4, 5).
Таким образом, конфликтная ситуация у хорошо обучаемых мышей линии CBA/CaLac в целом приводит к развитию продолжительной гиперплазии костномозгового гемопоэза, а так же лимфоцитозу, нейтрофильному лейкоцитозу и ретикулоцитозу в периферической крови. В то же время у плохо обучаемых животных гранулоцитарный (в меньшей степени), эритроидный и лимфоидный (в большей степени) ростки кроветворения, напротив, угнетаются. При этом в циркулирующей крови периоды уменьшения содержания лейкоцитов и ретикулоцитов сменяются на периоды их увеличения.
Пример 3.
Первоначально животных типируют в Т-образном лабиринте, затем у них моделируют депривацию парадоксального сна, далее производят забор экспериментального материала ежедневно в течение 6 сут после депривации парадоксального сна.
В ходе исследований установлено, что депривация парадоксального сна оказывала в основном угнетающее действие на костномозговое кроветворение у хорошо обучаемых мышей линии CBA/CaLac. Так, на 1, 2, 3, 5-е сут опыта наблюдалось снижение ОКК (до 78%, Р=0,043; 64%, Р=0,028; 70%, Р=0,028; 62%, Р=0,043 соответственно) (табл.6). Динамика содержания отдельных форменных элементов изучаемых ростков кроветворения во многом повторяла таковую для ОКК. В частности, количество незрелых и зрелых форм нейтрофильных гранулоцитов в костном мозге значительно было ниже исходного уровня на 2, 3, 5-е сут (на 44-45%) и 1, 2, 3, 5-е сут (на 28-32%) эксперимента соответственно. На 2, 3, 4, 5-е сут исследования отмечалось уменьшение числа эозинофильных гранулоцитов (до 45-62%) и моноцитов (до 0-25%). В свою очередь, представительство лимфоидных клеток уступало фоновым значениям на 1,2, 3, 5-е сут (до 58-73%), а содержание эритроидных клеток - на протяжении всего периода исследования (до 48-81%). Вместе с тем следует отметить, что число зрелых форм нейтрофильных гранулоцитов на 4-е сут исследования составляло 8,39±0,44×106 клеток/бедро при (6,46±0,62)×106 клеток/бедро в интактном контроле.
Депривация парадоксального сна вызывала статистически достоверное уменьшение количества эритрокариоцитов на 2, 3, 4, 6-е сут опыта (до 64-42%), лимфоцитов на 2, 3, 4, 5, 6-е сут (до 51%, 54%, 28%, 60%, 57% соответственно), эозинофильных гранулоцитов на 3, 4, 5, 6-е сут (до 35%, 18%, 48%, 42% соответственно) и зрелых форм нейтрофилов на 4-е сут (до 41%) в костном мозге плохо обучаемых мышей (табл.7). Наряду с этим наблюдалось увеличение количества незрелых на 1, 5-е сут исследования (на 50% выше исходного) и на 3-й сут (до 50% выше исходного) зрелых форм нейтрофильных гранулоцитов. На 1-е сут эксперимента представительство эозинофильных гранулоцитов составляло 0,844:0,08×106 клеток/бедро при 0,62±0,09×106 клеток/бедро в интактном контроле (Р=0,046).
При сопоставлении показателей костномозгового кроветворения у мышей с разным типом поведения при выработке условного питьевого рефлекса было выявлено, что депривация парадоксального сна оказывала наиболее выраженное ингибирующее действие на гемопоэз у хорошо обучаемых мышей, чем у плохо обучаемых особей (табл.6, 7).
У хорошо обучаемых мышей при депривации парадоксального сна наблюдалось на 1, 4, 5 и 6-е сут эксперимента (до 62%, 71%, Р=0,043 и 73%, Р=0,028; 71%, Р=0,043 соответственно) значительное снижение, а на 3-и сут (до 121%) увеличение ОКЛ в периферической крови (табл.8). Обнаруженная лейкопения была связана с падением содержания всех форм лейкоцитов: лимфоцитов на 1, 4, 5 и 6-е сут исследования, палочкоядерных нейтрофилов на 1, 2, 4 и 5-е сут (до 24%, 37%, 18%, Р=0,028 и 19%, Р=0,043 соответственно) и сегментоядерных нейтрофилов на 1, 4-е сут (до 76%, Р=0,046 и 73%, Р=0,028 соответственно). Выраженная ретикулоцитопения (1, 2-е сут) сменялась на ретикулоцитоз (5-е сут) (табл.5).
У плохо обучаемых мышей, перенесших депривацию парадоксального сна (как и у хорошо обучаемых особей) наблюдалось уменьшение ОКЛ в периферической крови на 3, 4, 5-е сут исследования (до 64%, Р=0,028; 71%, Р=0,043 и 73%, Р=0,028 соответственно) (табл.9). Однако в описываемой группе лейкопения была сопряжена, прежде всего, со статистически достоверным падением содержания лимфоцитов весь период исследования, вклад палочкоядерных нейтрофилов (2, 4, 5-е сут) в ее развитие не значителен. Количество ретикулоцитов на 2-е сут опыта было сниженным, а на 6-е сут, наоборот, повышенным (табл.5). В то же время в циркулирующей крови наблюдалось увеличение числа палочкоядерных (6-е сут) и сегментоядерных нейтрофилов на 1, 5-е сут эксперимента (до 193-123%, Р=0,028), а эозинофилов - на 1-е сут (до 231%, Р=0,028).
Итак, депривация парадоксального сна вызывает развитие гипоплазии костномозгового гемопоэза, а также лейкопении и ретикулоцитопении в периферической крови у хорошо обучаемых мышей. В свою очередь, у плохо обучаемых животных выраженность депрессии эритроидного и лимфоидного ростков кроветворения уступает таковой у хорошо обучаемых особей, но содержание нейтрофильных гранулоцитов в системе крови, напротив, возрастает.
Проведенные исследования показали, что предлагаемый способ позволяет определить, реакции изучаемых ростков кроветворения: разнонаправленные у хорошо обучаемых мышей и во многом однозначные у плохо обучаемых особей и дает возможность установить корреляцию между типом поведения и индивидуальной устойчивостью к экспериментальным невротическим воздействиям.
Литература
1. Гольдберг Е.Д., Дыгай A.M., Провалова Н.В. и др. Роль нервной системы в регуляции кроветворения. - Томск: Изд-во Том. Унта, 2004. - С.43-49, 84-111.
2. Дыгай A.M., Суслов Н.И., Скурихин Е.Г., Чурин А.А. Реакции эритроидного ростка кроветворения при различных типах невротических воздействий // Бюл. эксперим. и медицины. - 1997. - Т.123, №2. - С.158-161.
3. Дыгай A.M., Скурихин Е.Г., Суслов Н.И. и др. Реакции гранулоцитарного ростка кроветворения при различных типах невротических воздействий // Бюл. эксперим. и медицины. - 1998. - Т.126, №12. - С.628-631.
4. Пшенникова М.Г. Феномен стресса. Эмоциональный стресс и его роль в патологии // Патол. физиология и эксперим. терапия. - 2001. - №3. - С.28-32.
5. Горизонтов П.Д. Система крови как основа резистентности и адаптации организма // Патол. физиология и эксперим. терапия. - 1981. - №2. - С.55-63.
6. Августинович Д.Ф., Липина Т.В., Бондарь Н.П., Кудрявцева Н.Н. Особенности проявления наследственно обусловленной тревожности у самцов мышей линии C57BL/6J и CBA/Lac // Журнал высшей нервной деятельности им. И.П. Павлова. - 1999. - Т.49, №6. - С.1008-1017.
7. Кудрявцева Н.Н. Агонистическое поведение: модель, эксперимент, перспективы // Российский физиол. журнал им. И.М. Сеченова. - 1999. - Т.85, №1. - С.67-83.
8. Вейко Н.Н., Шубаева Н.О., Малашенко A.M. и др. Сравнение копийности активности рибосомных генов в четырех инбредных линиях мышей // Генетика и селекция животных. - 2002. - С.11.
9. Гольдберг Е.Д., Дыгай A.M., Шахов В.П. Методы культуры ткани в гематологии. - Томск, 1992. - С.208, 214.
10. Скурихин Е.Г. Механизмы регуляции кроветворения при экспериментальных неврозах. - Дис.... докт. мед. наук. - 2004. - С.78.
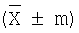
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕКАРСТВЕННЫЕ СРЕДСТВА ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ АНЕМИИ ПРИ НЕВРОТИЧЕСКИХ ВОЗДЕЙСТВИЯХ | 2006 |
|
RU2329035C1 |
| СПОСОБ РАЗДЕЛЕНИЯ ИНБРЕДНЫХ МЫШЕЙ НА ТИПОЛОГИЧЕСКИЕ ГРУППЫ С УЧЕТОМ ОСОБЕННОСТЕЙ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ | 2006 |
|
RU2316057C1 |
| СПОСОБ ТЕРАПИИ ИНДИВИДУАЛЬНЫХ НАРУШЕНИЙ В СИСТЕМЕ КРОВИ ПРИ ЭКСПЕРИМЕНТАЛЬНЫХ НЕВРОТИЧЕСКИХ ВОЗДЕЙСТВИЯХ | 2006 |
|
RU2329806C2 |
| СПОСОБ КОРРЕКЦИИ ДЕПРЕССИИ ГРАНУЛОЦИТОПОЭЗА ПРИ ЦИТОСТАТИЧЕСКОМ ВОЗДЕЙСТВИИ | 2008 |
|
RU2366422C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЭФФЕКТИВНОСТИ ГЕМОСТИМУЛЯТОРОВ ПРИ ЦИТОСТАТИЧЕСКОЙ МИЕЛОСУПРЕССИИ | 2009 |
|
RU2421720C2 |
| СПОСОБ СТИМУЛЯЦИИ МИЕЛОПОЭЗА | 2007 |
|
RU2336899C1 |
| СПОСОБ ОЦЕНКИ ГРАНУЛОЦИТОПОЭЗСТИМУЛИРУЮЩЕЙ АКТИВНОСТИ ФАРМАКОЛОГИЧЕСКИХ ВЕЩЕСТВ | 2006 |
|
RU2332667C1 |
| СПОСОБ СТИМУЛЯЦИИ МИЕЛОПОЭЗА | 2008 |
|
RU2366452C1 |
| СРЕДСТВО, ПРЕПЯТСТВУЮЩЕЕ РАЗВИТИЮ ДЕПРЕССИИ ЭРИТРОИДНОГО РОСТКА КРОВЕТВОРЕНИЯ ПРИ ВВЕДЕНИИ ЦИТОСТАТИКА | 2007 |
|
RU2354376C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ГЕМОСТИМУЛИРУЮЩИМ, АНТИМУТАГЕННЫМ, ПРОТИВООПУХОЛЕВЫМ, ЦЕРЕБРОПРОТЕКТОРНЫМ, АНТИГИПОКСИЧЕСКИМ, НООТРОПНЫМ, АНКСИОЛИТИЧЕСКИМ И ПРОТИВОНЕВРОТИЧЕСКИМ ДЕЙСТВИЕМ | 2010 |
|
RU2438691C1 |
Изобретение относится к области экспериментальной медицины, а именно к патологической физиологии. Сущность способа: инбредных мышей предварительно разделяют на группы хорошо и плохо обучаемых животных путем типирования при выработке условного питьевого рефлекса со сложной пространственной ориентировкой в 3-звенном Т-образном лабиринте в течение 2 дней по 2 побежки с интервалом в 1 час. К группе хорошо обучаемых животных относят особей, у которых средние значения 4 побежек вертикальной активности не выше 12 условных единиц, количества ошибок при поиске поилки не выше 12 условных единиц, время достижения поилки не более 100 с, время неподвижности не более 3 с, отсутствуют невротические реакции; к группе плохо обучающихся относят животных, у которых не менее 60% указанных показателей превышают указанные значения, и присутствуют невротические реакции. Использование способа позволяет определить реакции изучаемых ростков кроветворения и установить корреляцию между типом поведения и индивидуальной устойчивостью к экспериментальным невротическим воздействиям. 9 табл.
Способ изучения гемопоэза с учетом индивидуальных особенностей высшей нервной деятельности, заключающийся в изучении реакций системы крови при экспериментальных невротических воздействиях, при котором инбредных мышей предварительно разделяют на группы хорошо и плохо обучаемых животных путем типирования при выработке условного питьевого рефлекса со сложной пространственной ориентировкой в 3-звенном Т-образном лабиринте, отличающийся тем, что типирование проводят в течение 2 дней по 2 побежки с интервалом в 1 ч, при этом к группе хорошо обучаемых животных относят особей, у которых средние значения 4 побежек вертикальной активности не выше 12 условных единиц, количества ошибок при поиске поилки не выше 12 условных единиц, время достижения поилки не более 100 с, время неподвижности не более 3 с, отсутствуют невротические реакции, к группе плохо обучающихся относят животных, у которых не менее 60% указанных показателей превышают указанные значения, и присутствуют невротические реакции.
| ПЕРШИНА О.В | |||
| и др | |||
| Особенности реакций эритроидного ростка кроветворения у хорошо и плохо обучающихся мышей линии CBA/CALAC при невротических воздействиях | |||
| Бюл | |||
| эксперим | |||
| биол | |||
| и медицины, 2004, т | |||
| Прибор для определения всасывающей силы почвы | 1921 |
|
SU138A1 |
| СКУРИХИН Е.Г | |||
| и др | |||
| Особенности реакций гранулоцитарного ростка кроветворения у мышей с разной способностью к обучению при | |||

