Предлагаемое изобретение относится к медицине, а именно к неврологии, психопатологии, нейрохирургии, нейрофизиологии и экспериментальной нейробиологии, а также психофизиологии и предназначено для определения функционального и метаболического состояния нервной ткани в норме и при патологии.
Известен способ определения функционального и метаболического состояния нервной ткани путем проведения позитронно-эмиссионной томографии мозга /Buchsbaum MS, Gillin JC, Wu J, Hazlett E, Sicotte N, Dupont RM, Bunney WE Jr. Regional cerebral glucose metabolic rate in human sleep assessed by positron emission tomography // Life Sci 1989; 45(15): 1349-56/.
Недостатком этого способа является сложность реализации, высокие экономические затраты и использование дорогостоящего оборудования для проведения исследования, необходимость предварительной подготовки пациентов к исследованию. Недостатками способа являются также невозможность проведения динамического контроля над состоянием мозга во время медицинских манипуляций в клинике, сложность и неудобство для реализации в экспериментальных исследованиях на мелких лабораторных животных.
Известен способ регистрации суммарной медленной электрической активности мозга с поверхности головы (ЭЭГ) /Биопотенциалы мозга человека. Математический анализ. // Под ред. Русинова B.C., М.: Медицина, 1987, 254 с./. Известно, что активационные процессы в нервной системе сопровождается депрессией альфа-активности. Развитие патологических состояний в связи с нарушением метаболизма, как при ишемии мозга, связано с появлением медленноволновой активности тета- и дельта-диапазонов. Угнетение функционального состояния при углублении гипоксии и ишемии приводит к депрессии ЭЭ.
Несмотря на наличие у данного способа ряда положительных свойств, он не позволяет тонко дифференцировать многие физиологические и патологические ФС.
Ближайшим аналогом является способ определения функционального и метаболического состояния нервной ткани /Патент РФ №2245673, А61В 5/04, А61В 5/0476, 2005 г./, включающий регистрацию электроэнцефалограммы (ЭЭГ), одновременно с ЭЭГ регистрируют уровень постоянного потенциала (УПП) и при негативизации УПП и увеличении мощности ЭЭГ определяют деполяризационную активность нейронов и усиление метаболизма; при негативизации УПП и уменьшении мощности ЭЭГ - деполяризационное торможение нейронов и угнетение метаболизма; при позитивации УПП и увеличении мощности ЭЭГ - реполяризационную или гиперполяризационную активизацию нейронов и усиление метаболизма; при позитивации УПП и уменьшении мощности ЭЭГ - гиперполяризационное торможение нейронов и снижение метаболизма нервной ткани.
Однако известный способ определяет лишь изменение потребности клеток нервной ткани с количественной точки зрения, не характеризуя функциональное и метаболическое состояние клеток с качественной стороны. Так в данном способе усиление метаболизма наблюдают как при негативизации УПП и увеличении мощности ритмов ЭЭГ, так и при позитивизации УПП и увеличении мощности ритмов ЭЭГ. В одинаковом ли функциональном состоянии (ФС) при этом находится нервная ткань не ясно. Понятие ФС требует качественной характеристики текущей деятельности /Медведев В.И. Функциональные состояния оператора // Эргономика: Принципы и рекомендации. М., 1970. Вып.1. С.127-160; Доскин В.А., Лаврентьева Н.А., Мирошников М.П., Шарай В.Б. Тест дифференциальной самооценки функционального состояния. // Вопросы психологии, 1973, №6, с.141-145; Медведев В.И., Леонова А.Б. Функциональные состояния человека. В кн.: Физиология трудовой деятельности. Санкт-Петербург: Наука, 1993, с.25-61/. Характеризуя метаболическое и ФС нервных клеток с этих позиций, необходимо описать в хорошем состоянии они находятся или плохом. В известном способе это не определяется.
Задачей предлагаемого изобретения является создание способа, позволяющего характеризовать текущее метаболическое и ФС нервной ткани с качественной точки зрения, за счет регистрации и дифференцирования характера изменений УПП и ЭЭГ.
Поставленная задача достигается тем, что в известном способе определения функционального и метаболического состояния нервной ткани, включающем одновременную регистрацию электроэнцефалограммы (ЭЭГ) и уровня постоянного потенциала (УПП), поляризационные и активационные процессы, наблюдаемые по характеру изменений УПП и ЭЭГ сопоставляют с состоянием лабильности и резистентности нервных клеток и их биохимических систем и дают качественную характеристику метаболического и ФС нервной ткани.
Позитивизацию УПП и уменьшение мощности ритмов ЭЭГ рассматривают либо как формирование гиперполяризационного торможения нейронов и развитие превосходного метаболического и ФС нервной ткани, либо как реполяризацию мембраны после периода деполяризационного возбуждения и улучшение метаболического и ФС нервной ткани.
Позитивизацию УПП и увеличение мощности ритмов ЭЭГ рассматривают либо как постгиперполяризационную активации нейронов и развитие у них хорошего метаболического и ФС, либо как развитие реполяризационной активации нейронов и улучшение у них метаболического и ФС после деполяризационного торможения (парабиоза).
Негативизацию УПП и увеличение мощности ритмов ЭЭГ рассматривают как развитие деполяризационной активации нейронов и плохого метаболического и ФС нервной ткани.
Еще большую негативизацию УПП и уменьшение мощности ритмов ЭЭГ рассматривают как развитие деполяризационного торможения и очень плохого метаболического и ФС нервной ткани.
Сопоставление поляризационных и активационных процессов, наблюдаемых по характеру изменений УПП и ЭЭГ с состоянием лабильности и резистентности нервных клеток и их биохимических систем позволяет дать качественную характеристику метаболического и ФС нервной ткани.
Способ осуществляется следующим образом.
У объекта исследования неполяризующимися (хлорсеребряными) электродами с помощью усилителя постоянного тока проводят одновременную регистрацию уровня постоянного потенциала и суммарной медленной электрической активности (ЭЭГ) в исследуемой нервной ткани.
При позитивных сдвигах УПП и угнетении мощности ЭЭГ, сопровождающих возрастание потенциала покоя клеток (ре- или гиперполяризацию) и торможение нейрональной активности, определяют либо улучшение метаболического и ФС нервной ткани в случае возвращения мембранного потенциала к уровню потенциала покоя после деполяризационной экзальтации возбудимости, либо развитие превосходного метаболического и ФС в случае гиперполяризационного торможения клеток.
При позитивных сдвигах УПП и увеличении мощности ЭЭГ, сопровождающих возрастание потенциала покоя клеток (ре- или гиперполяризацию) и активацию нейрональной активности, определяют улучшение метаболического и ФС в связи с выходом клеток из парабиоза, либо хорошего метаболического и ФС в случае гиперполяризационной экзальтации возбудимости.
При негативных сдвигах УПП и увеличении мощности ЭЭГ, сопровождающих снижение потенциала покоя (деполяризацию) и активацию нервных клеток, определяют развитие плохого метаболического и ФС клеток нервной ткани.
При негативных сдвигах УПП и угнетении мощности ЭЭГ, сопровождающих сильное снижение потенциала покоя (сильную деполяризацию) клеток и торможение нейрональной активности (по типу парабиоза), определяют развитие очень плохого метаболического и ФС клеток мозга.
Пример
Давно известно, что наркозные препараты барбитуратового ряда при действии на нервные клетки вызывают их гиперполяризацию /Sato P., Austin G., Yai H. Increase in permeability of the postsynaptic membrane to potassium produced by "nembutal" // Nature, 1967, 215, р.1506-1509/ и торможение нейрональной активности. Вместе с тем, показано, что эти вещества обладают нейропротекторным действием /Самушия О.Ш. Ультраструктурные проявления эффекта лечения ишемического поражения головного мозга оксибутиратом натрия // Вопросы современной неврологии. Сборник научных трудов. Тбилиси, 1985, с.184-190/. Гиперполяризационное торможение и нейропротекторное действие оказывают также аденозин и его аналоги /Кулинский В.И., Усов Л.А., Суфианова Г.З., Суфианов А.А. Защитный эффект интерцеребровентрикулярного введения А-Агонистов при полной ишемии головного мозга // Бюллетень экспериментальной биологии и медицины, 1994, №6, с.622-624/. Исследование функциональной подвижности (лабильности) элементов нервной ткани /Сологуб М.И. Функциональные характеристики клеток при их гиперполяризации и деполяризации // Физиологические механизмы основных нервных процессов. Труды Ленингр. об-ва естествоисп. Л., 1985, т.75, вып.5, с.31-40/ показало, что возрастание потенциала покоя (ПП) клеток (гиперполяризация) сопровождается увеличением их функциональной подвижности, т.е. лабильности. Наконец, имеются данные о том, что гиперполяризированные нейроны обладают в целом более высокой резистентностью к неблагоприятным факторам /Насонов Д.Н. Местная реакция протоплазмы и распространяющееся возбуждение. Л., 1959.; Голиков Н.В. Проблема местного и распространяющегося возбуждения в современной нейрофизиологии // Механизмы местной реакции и распространяющегося возбуждения. Л.: Наука, 1970, с.5-12/. Положительное биологическое действие гиперполяризации может быть связано с мобилизацией в этот период энергетических, а соответственно и гомеостатических механизмов клетки /Горбань Е.И. Взаимосвязь мембранного потенциала адренокортикоцитов и функциональной активности надпочечников у взрослых и старых/.
При деполяризации ПП клеток их лабильность снижается /Сологуб М.И. Функциональные характеристики клеток при их гиперполяризации и деполяризации // Физиологические механизмы основных нервных процессов. Труды Ленингр. о-ва естествоисп. Л., 1985, т.75, вып.5, с.31-40., Голиков Н.В. Физиологическая лабильность и ее изменения при основных нервных процессах. Л., 1950, 240 с./. Одновременно с этим при устойчивой деполяризации клеток в них развивается целый ряд патогенетических процессов: вход ионов Са2+, активация перекисного окисления липидов / Дупин А.М., Барсков И.В., Викторов И.В., Ерин А.Н. Уровень перикисного окисления липидов в очаге компрессионной ишемии коры головного мохга крыс // Бюллетень экспериментальной биологии и медицины, 1994, №12, с.589-590. Голубев А.Г. Окись азота (NO) в центральной нервной системе // Бюллетень экспериментальной биологии и медицины, 1994, №2, с.201/, нарушение энергетического обмена /Гаевская М.С. Углеводный обмен головного мозга и его связь с функциональным состоянием центральной нервной системы при угасании и восстановлении жизненных функций организма // Биохимия; Шапот B.C., Громова К.Г. Энергетический обмен головного мозга и проблема нервной системы, Киев: Изд-во АН Укр ССР, 1954, с.151-161; Мак-Ильвейн Г. Биохимия и центральная нервная система. М.: Изд-во иностр. лит-ра, 1962/, приводящих в конечном итоге к гибели клеток / Исаев Н.К., Зоров Д.Б., Лыжин А.А. и др. Глутамат вызывает понижение мембранного потенциала митохондрий в культивированных клетках-зернах мозжечка // Бюллетень экспериментальной биологии и медицины, 1994, №2, с.208; Глушков С.И., и др. Острые отравления. М.: Медицина, 1989, 432 с.; Пузырев В.П., Голубенке Н.В., Фрейдин М.Б. Сфера компетенции митохондриальной генетики // Вестник РАМН, 2001, №10, с.31-37; Kristian Т., Siesjo B.K. Calcium in ischemic cell death // Stroke, 1998, v.29, 3, p.705-718, Mattson M.P., Culmsee C., Yu Z.F. Apoptotic and antiapoptotuc mechanisms in stroke // Cell Tissue Res., 2000, v.301, 1, p.173-187/.
Таким образом, в настоящее время, есть все основания говорить о том, что возрастание ПП (гиперполяризация) сопровождается развитием хорошего метаболического и ФС, тогда как снижение ПП (устойчивая (стационарная) деполяризация) сопровождается развитием плохого метаболического и ФС нервных клеток.
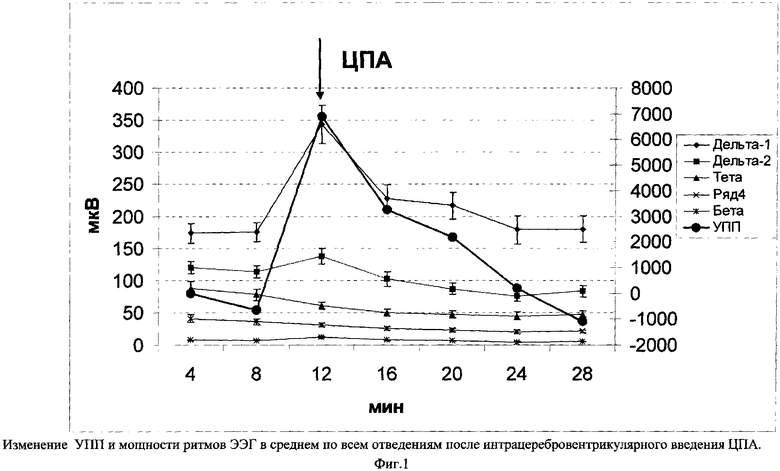
Для оценки функционального и метаболического состояния нервной ткани по-нашему способу было проведено исследование УПП и ЭЭГ при интрацеребровентрикулярном введении циклопентиладенозина (ЦПА). На фиг.1 показано изменение УПП и ЭЭГ у крыс (n=10) после интрацеребровентрикулярного введения ЦПА в неокортексе. Видно, что инъекция ЦПА вызывала позитивное отклонение УПП с первичной активацией мощности большинства (дельта-1, дельта-2 и бета) ритмов ЭЭГ, которая затем (через 5-10 минут) сменилась их депрессией (см. Таблица).
Звездочками обозначены значения амплитуды ритмов, достоверно отличающиеся от периода, предшествовавшего введению препарата: *-р<.05, **-р<.01, ***-р<.001.
В среднем позитивное отклонение УПП через 4 минуты после введения ЦПА равнялось 6890±2418 мкВ.
Спустя 40 минут после инъекции ЦПА, мощность ритмов ЭЭГ снизилась на 34,56±1,96% (р<0,001). Уменьшение мощности ЭЭГ на фоне ЦПА было примерно одинаковым для всех ритмов и колебалось в пределах от 32,77±5,37% (для альфа-) до 37,19±2,64% (для бета-ритма).
Таким образом, введение ЦПА с течением времени вызывало на фоне позитивного отклонения УПП разнонаправленные изменения мощности ритмов ЭЭГ. Сразу после введения препарата мощность ритмов увеличивалась, а затем, спустя 5-10 минут, происходило ее угнетение.
По литературным данным /Kostopoulos G.К., Phillis J.W. Purinergic depression of neurons in different areas of the rat brain // Exp. Neurol. 1977. Vol.55. P.719-724, Shefner S.A. Chiui R.H. Adenosine inhibits locus coereleus neurons: an intracellular study in a rat brain slices preparation //Brain Res. 1986. VoL 366, №1-2. P.364-368/, аденозин и его аналоги (к которым относится и ЦПА) угнетают импульсную активность нейронов и вызывают гиперполяризацию мембраны. Позитивизация УПП, полученная в нашем эксперименте, также свидетельствует о гиперполяризации нейронов. Тот факт, что гиперполяризующие воздействия повышают устойчивость нервных клеток к действию неблагоприятных факторов /Самушия О.Ш. Ультраструктурные проявления эффекта лечения ишемического поражения головного мозга оксибутиратом натрия // Вопросы современной неврологии. Сборник научных трудов. Тбилиси, 1985, с.184-190; Кулинский В.И., Усов Л.А., Суфианова Г.З., Суфианов А.А. Защитный эффект интерцеребровентрикулярного введения А-Агонистов при полной ишемии головного мозга // Бюллетень экспериментальной биологии и медицины, 1994, №6, с.622-624; Насонов Д.Н. Местная реакция протоплазмы и распространяющееся возбуждение. Л., 1959 г./, а также увеличивают их лабильность, позволяет говорить, что при действии аденозина в нервной ткани развилось хорошее метаболическое и ФС. Однако гиперполяризация могла сочетаться как с увеличением мощности ритмов, так и ее угнетением. Первичная активация ЭЭГ в дельта- и бета-диапазонах (фиг.1) при позитивном сдвиге УПП отражает, по всей видимости, развитие хорошего функционального состояния, подобного анодной экзальтации с некоторым увеличением метаболической потребности на фоне сохранения ионного гомеостаза, которое постепенно сменяется на полное гиперполяризационное торможение и развитие еще лучшего (превосходного) метаболического и ФС. Развившееся после введения ЦПА угнетение мощности ритмов было устойчивым и имело место на протяжении 60 минут наблюдения.
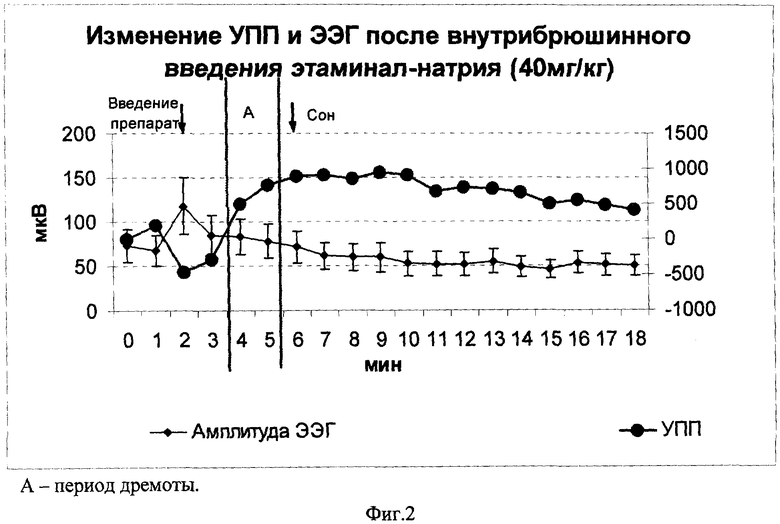
На фиг.2 показано изменение регистрируемых биопотенциалов при внутрибрюшинном введении крысам (n=22) этаминал-натрия (нембутала). Видно, что вначале после инъекции препарата наблюдалось негативное отклонение УПП (р<0,001) и увеличение мощности ЭЭГ (р<0,001). Негативизация потенциала составила в среднем 638,8±124,1 мкВ, а увеличение мощности ритмов 28,0±2,3%. Спустя 1-2 минуты, появилось позитивное смещение постоянного потенциала при сохранении повышенной мощности ритмов (фиг.2, А), животное при этом находилось в дремотном состоянии. Потеря болевой чувствительности и засыпание животного происходили на фоне дальнейшей позитивизации УПП при одновременном снижении амплитуды ритмов ЭЭГ, угнетение которых достигало максимума при углублении сна. Среднее значение позитивного сдвига потенциала при этаминаловом сне составило 1043±283 мкВ. Характер изменения диапазонов ЭЭГ-ритмов при наркотизации друг от друга существенно не отличался.
В целом результаты последнего исследования показали, что сложная картина взаимоотношений УПП и ритмов ЭЭГ, выявленная на модели интрацеребровентикулярного введении ЦПА, наблюдается и после внутрибрюшинной инъекции нембутала. Предсонное (дремотное) состояние сочеталось позитивным отклонением УПП и относительно высокой мощностью ритмов ЭЭГ, тогда как засыпание - еще большей позитивизацией УПП, сочетающейся, однако, со снижением мощности ритмов ЭЭГ. Полученные данные указывают на формирование под действием нембутала гиперполяризационных процессов в ткани головного мозга. Характер изменения регистрируемых показателей говорит о том, что нембутал, так же, как и ЦПА, вызывает развитие в целом хорошего метаболического и ФС нервной ткани мозга. Содружественные сдвиги УПП и ЭЭГ позволяют также видеть динамику данного состояния и развитие превосходного метаболического и ФС при переходе от дремоты ко сну.
Данное исследование позволило выявить также период, когда наблюдался негативный сдвиг УПП сочетающийся с большой мощностью ритмов ЭЭГ. Данные изменения биопотенциалов наблюдались сразу после внутрибрюшинной инъекции нембутала. Очевидно, что процедура введения наркоза (фиксация животного в руках, прокалывание кожных покровов инъекционной иглой и введение препарата) сильно мотивировала животных, и первые 1-2 минуты крысы были сильно возбуждены. В данном состоянии наблюдалась негативизация УПП и увеличение мощности ритмов ЭЭГ. Характер изменения, указанных биопотенциалов говорит, о том, что в нервной ткани коры больших полушарий головного мозга в это время развиваются деполяризационные процессы, сочетающиеся с высокой нейрональной активностью, т.е. состояние типа катодической экзальтации. В подобном состоянии в нервной ткани регистрируется снижение содержания макроэргов, а в целом метаболическое состояние нервных клеток при этом подобно тому, которое развивается при гипоксии. Кроме этого, устойчивая деполяризация ПП клеток активирует целый каскад внутриклеточных патогенетических процессов /Исаев Н.К., Зоров Д.Б., Лыжин А.А. и др. Глутамат вызывает понижение мембранного потенциала митохондрий в культивированных клетках-зернах мозжечка // Бюллетень экспериментальной биологии и медицины, 1994, №2, с.208, Дупин А.М., Барсков И.В., Викторов И.В., Ерин А.Н. Уровень перикисного окисления липидов в очаге компрессионной ишемии коры головного мозга крыс // Бюллетень экспериментальной биологии и медицины, 1994, №12, с.589-590, Пузырев В.П., Голубенко Н.В., Фрейдин М.Б. Сфера компетенции митохондриальной генетики // Вестник РАМН, 2001, №10, с.31-37.; Kristian Т., Siesjo B.K. Calcium in ischemic cell death // Stroke, 1998, v.29, 3, p.705-718; Mattson M.P., Culmsee C., Yu Z.F. Apoptotic and antiapoptotuc mechanisms in stroke // Cell Tissue Res., 2000, v.301, 1, p.173-187 и др./. Факт снижения у деполяризованных нейронов лабильности /Сологуб М.И. Функциональные характеристики клеток при их гиперполяризации и деполяризации // Физиологические механизмы основных нервных процессов. Труды Ленингр. об-ва естествоисп. Л., 1985, т.75, вып.5, с.31-40/ также говорит о том, что устойчивая деполяризация мембранного потенциала отражает развитие относительно плохого метаболического и ФС нервных клеток.
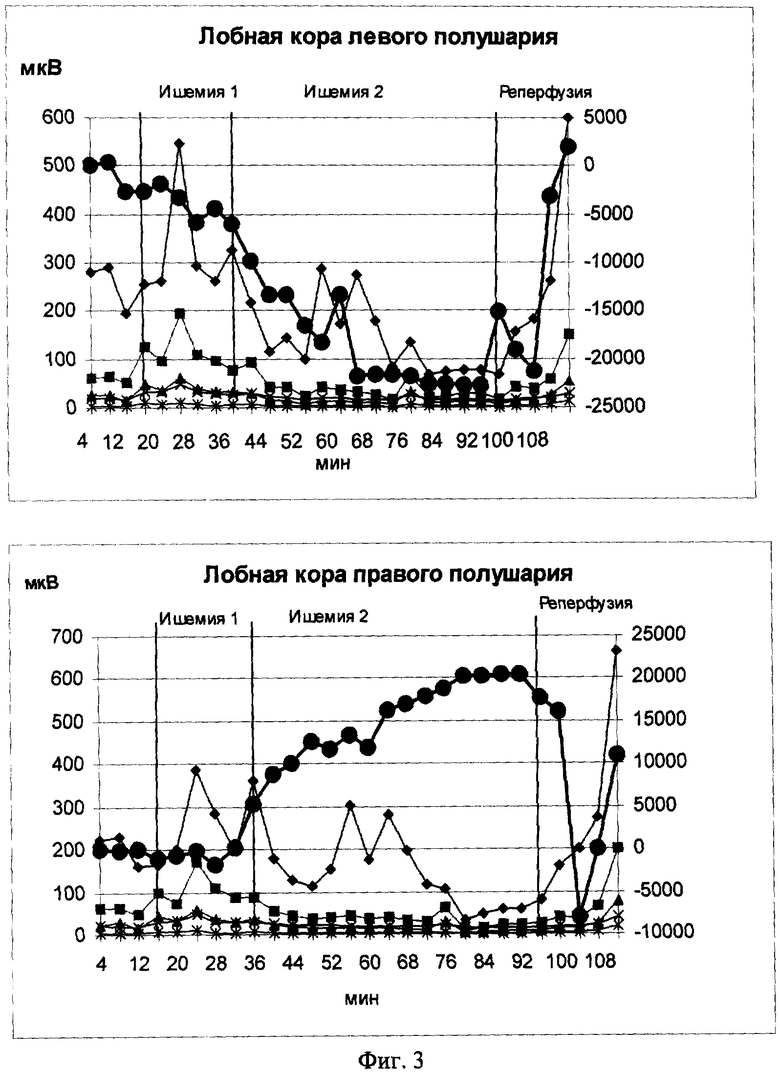
На фиг.3 и 3а показаны содружественные изменения УПП и ЭЭГ при моделировании у крыс ишемии мозга разной тяжести. Моделирование ишемии проводилось двумя способами на одних и тех же крысах последовательно в течение одного эксперимента. Предварительно за 2-3 дня до опыта беспородным белым крысам массой 150-200 г обоего пола (n=12) под нембуталовым наркозом под кости черепа над лобной корой правого и левого полушарий вживлялись хлорсеребряные электроды. Индифферентный серебряный хлорированный электрод размещался в костях над лобными пазухами. Выводы электродов крепились к черепу быстрозатвердевающей пластмассой.
Первая модель ишемии ("Ишемия-1") заключалась в перевязывании обеих общих сонных артерий. Длительность изолированного действия этой модели ишемии составляла 20 минут, после чего проводилось дополнительное введение крысам окклюдера в среднюю мозговую артерию (СМА) левого полушария ("Ишемия-2") /Суфианова Г.З., Усов Л.А., Суфианов А.А., Шапкин А.Г., Раевская Л.Ю., Голубев С.С., Мурик С.Э. Малоинва-зивная модель фокальной ишемии головного мозга у крыс// Экспериментальная и клиническая фармакология. 2001, т.64, №4, с.63-67/. Сочетанное моделирование двух видов циркуляторной ишемии осуществлялось в течение 60 минут, после чего окклюдер извлекался и проводилась регистрация УПП и ЭЭГ еще на протяжении 16 минут.
На фиг.3 и 3а показано изменение мощности ЭЭГ и УПП во время моделирования ишемии двумя способами. Видно, что перевязывание общих сонных артерий ("Ишемия 1") привело к увеличению мощности ЭЭГ в лобной и теменной коре правого и левого полушарий. Увеличение мощности спектра ритмов во всех отведениях в целом у всей выборки крыс равнялось 14,04±1,75% (р<0,001). Наибольшее увеличение амплитуды (20,04±3,2%) наблюдалось в альфа-диапазоне, где она выросла с 23,71±0,86 до 28,46±1,09 мкВ (р<0,001). Для тета-ритма увеличение составляло 16,31±3,15%, и он изменился с 46,52±2,03 до 54,01±2,08 мкВ (р<0,01). Повышение амплитуды дельта-ритма равнялось 10,61±2,79%: с 106,85±3,96 до 118,38±3,74 мкВ (р<0,01). Меньше всего изменился бета-ритм. Его мощность увеличилась только на 9,21±4,33% с 7,31±0,34 до 7,98±0,33 мкВ. Тем не менее, и это увеличение было статистически достоверно (парный t-критерий Стьюдента показал различия при р<0,01). Одновременно с увеличением мощности ритмов ЭЭГ наблюдалось небольшое негативное отклонение УПП, которое к концу периода составило 1222,51±290,1 мкВ (р<0,01).
Характер изменения УПП и ЭЭГ при моделировании "Ишемии-1" свидетельствует о том, что в нервной ткани развиваются деполяризационные процессы, сочетающиеся с нейрональной активацией, т.е. состояние типа католической экзальтации.
Дополнительное введение окклюдера в СМА левого полушария на фоне развития общего угнетения ритмов ЭЭГ привело к более значительным изменениям УПП в неокортексе, которые имели при этом как позитивную, так и негативную направленность (Фиг.3, "Ишемия-2"). В левом полушарии как в лобной, так и теменной коре наблюдалось значительное негативное отклонение УПП, которое составило соответственно 20929±8177 и 12880±4175 мкВ. В отдельных случаях в лобной коре негативизация достигала около 40 мВ.
Среднее угнетение ритмов ЭЭГ в левом полушарии в целом по выборке составило в лобной коре 25,62±2,41, в теменной - 22,74±1,94%. Больше всего изменения затронули медленные частоты. В правом полушарии угнетение ритмов было достоверно меньше, чем в левом полушарии, и составило для лобной коры 14,28±2,49% и 13,03±2,19 - для теменной. Анализ изменения ЭЭГ в правом полушарии, отдельно по частотам, что угнетение затрагивало только дельта-ритм. В остальных частотных диапазонах введение окклюдера в СМА левого полушария не привело к достоверным изменениям ритмов по сравнению с периодом, предшествовавшим ишемии мозга. Отличия в изменении биопотенциалов в правом полушарии касались также УПП. В теменной коре правого полушария негативное отклонение было почти в 3 раза меньше, чем в левом полушарии и составляло 7705±1373 мкВ. В лобной коре правого полушария отклонение постоянного потенциала носило вообще позитивную направленность и равнялось 6183±2605 мкВ.
Таким образом, развитие циркуляторной ишемии, по модели 1, сопровождалось относительно небольшим негативным отклонением УПП и увеличением мощности ритмов ЭЭГ во всех отведениях. Использование, в модели 2, окклюзии СМА левого полушария привело к дополнительной значительной негативизации УПП левого полушария и угнетению в нем, на этом фоне, ЭЭГ.
Характер изменения регистрируемых потенциалов в левом полушарии после введения окклюдера в СМА левого полушария указывает на развитие в неокортексе сильных деполяризационных процессов, сочетающихся с угнетением нейрональной активности. Иначе говоря, в нервной ткани левого полушария в это время развивается состояние типа парабиоза или католической депрессии Вериго. Метаболическое и ФС нейронов, сопровождающее данное явление, очевидно еще хуже, чем при "Ишемии-1".
Изменения ЭЭГ и УПП, наблюдаемые в правом полушарии во время "Ишемии-2", связаны, по всей видимости, с перераспределением кровотока за счет активации механизмов коллатерального кровообращения. В литературе имеются данные об увеличении кровотока в бассейне симметричных артерий противоположного инсульту полушария. Наблюдаемые нами одновременно с негативизацией потенциала в левом полушарии позитивные сдвиги УПП в лобной коре правого полушария свидетельствуют об увеличении кровотока в этой части мозга и развитии в нем реполяризационных процессов, а снижение мощности ритмов ЭЭГ отражает улучшение метаболического и функционального состояния нервной ткани, ухудшенное в период "Ишемии-1". Иначе говоря, в лобной коре правого полушария после введения окклюдера, по всей видимости, сформировалось состояние близкое к дооперационному. Об этом говорит и гистологический анализ состояния нервной ткани правого полушария /Суфианова Г.З., Усов Л.А., Суфианов А.А., Шапкин А.Г., Раевская Л.Ю., Голубев С.С., Мурик С.Э. Малоинвазивная модель фокальной ишемии головного мозга у крыс // Экспериментальная и клиническая фармакология. 2001, т.64, №4, с.63-67/. За счет усиления кровоснабжения лобной коры правого полушария, по всей видимости, произошло частичное "обкрадывание" теменной коры этого же полушария. Негативный сдвиг УПП в теменной коре правого полушария вкупе с некоторым уменьшением в нем мощности ритмов указывает на то, что здесь в период "Ишемии-2" в нервной ткани также сформировалось относительно неблагоприятное метаболическое и функциональное состояние.
На фиг.3 и 3а также видно, что после извлечения окклюдера в отведениях регистрировались, как правило, изменения постоянного потенциала, противоположные тем, которые имели место при его введении в СМА. В среднем по всей группе, в левом полушарии в лобной и теменной коре за период в 16 минут потенциал позитивизировал соответственно на 42,7±10% (Р<0,05) и 40,0±22% (Р<0,05). В правом полушарии в лобной коре извлечение окклюдера вызвало снижение постоянного потенциала на 23,8±12,7% (Р<0,05). В теменной коре этого же полушария наблюдалась небольшая позитивизация УПП, равная 17,2±11,2%.
Анализ происходящих при этом изменений ритма показал, что реперфузия привела к увеличению мощности ЭЭГ по всем каналам и всем частотам в среднем на 11,6±2,03% (р<0,001). Сравнение изменений ЭЭГ в правом и левом полушариях выявило увеличение мощности ритмов преимущественно в теменной коре правого и лобной коре левого полушарий. Наибольшее увеличение средней амплитуды всех ритмов наблюдалось в теменной коре правого полушария и составило 29,08±2,86% (р<0,001), при этом увеличение больше всего затрагивало альфа- и бета-диапазоны (соответственно 31,47±5,14% (р<0,002) и 36,13±9,3% (р<0,004) против 20,25±3,05% (р<0,01) для дельта - и 18,98±2,86% (р<0,01) для тета-ритмов).
Увеличение мощности ритмов в лобной коре левого полушария равнялось 16,59±2,89% (р<0,001) без явного преобладания в каком-то из диапазонов. В лобной коре правого полушария при этом наблюдалась лишь тенденция увеличения амплитуды в области тета-ритма. В теменной коре левого полушария на фоне увеличения активности в соответствующей области правого полушария не было найдено достоверных изменений ни по одной частоте.
Таким образом, по ЭЭГ данным реперфузионные изменения затрагивали преимущественно теменную кору правого полушария, в которой мощность всех ритмов увеличилась. В левом полушарии некоторое увеличение амплитуды ритмов наблюдалось в лобной коре. Сопоставление ЭЭГ, установившейся в течение 16 мин после реперфузии, с ЭЭГ, наблюдавшейся у этих животных до и во время "Ишемии 1", показало, что правое полушарие по характеристике основных ритмов приблизилось к периоду "Ишемия I", хотя амплитуда и была несколько ниже. В левом же полушарии в целом мощность ритмов, как и во время ишемии, осталась в среднем ниже даже в сравнении с дооперационным периодом на 18,89±2,63% (р<0,001).
Таким образом, извлечение окклюдера и реперфузия мозга по СМА в целом вызвало увеличение мощности ритмов ЭЭГ по всем отведениям. В лобной и теменной коре левого полушария и теменной коре правого полушария при этом наблюдалось позитивное отклонение УПП. В лобной коре правого полушария реперфузия СМА левого полушария привела к увеличению мощности ЭЭГ на фоне негативизации УПП. Иначе говоря, восстановление режима кровотока, соответствующего периоду "Ишемии-1" в левом полушарии по электрофизиологическим данным, отражало выход из парабиотического состояния, то есть улучшение метаболического и функционального состояния нервной ткани. Сходные процессы имели место и в теменной коре правого полушария в то время, как в лобной коре извлечение окклюдера, снизив коллатеральное кровоснабжение, по всей видимости, ухудшило его метаболическое состояние, что вновь его активировало (как при моделировании "Ишемии-1").
Данные реперфузии указывают на то, что активация мозга, наблюдаемая по показателю ЭЭГ, возможна на фоне как негативного, так и позитивного отклонения УПП. Если в первом случае это свидетельствует об ухудшении функционального и метаболического состояния (также как и при "Ишемии-1"), то во втором случае - об его улучшении в связи с выходом из состояния катодической депрессии при усилении кровотока.
Анализ литературных данных и характера изменений комплекса биоэлектрических показателей (ЭЭГ и УПП), полученных в наших экспериментах с ишемией позволяет сделать заключение о функциональных и метаболических процессах в нервной ткани головного мозга, сопровождающих эти изменения:
- негативные сдвиги УПП и увеличение мощности ЭЭГ происходят при деполяризации мембраны нейронов, сопровождающейся повышением возбудимости и отражают развитие плохого метаболического и ФС нервной ткани;
- негативные сдвиги УПП и угнетение мощности ЭЭГ имеют место при еще большей деполяризации нейронов, развитии деполяризационного торможения (по парабиотическому или католическому типу) и отражают развитие еще более плохого метаболического и ФС нервной ткани;
- обратные изменения УПП и мощности ЭЭГ отражают улучшение функционального и метаболического состояния нервной ткани мозга.
Результаты представленных экспериментов продемонстрировали высокие диагностические возможности предлагаемого способа. Наш способ впервые позволил по данным биоэлектрической активности с качественной точки зрения дифференцировать изменение метаболического и ФС нервной ткани мозга при действии на мозг ЦПА, нембутала и ишемии разной тяжести. Данный способ может быть использован не только для выявления особенности действия лекарственных препаратов и разнообразных условий среды на клетки мозга, но и роли изменений метаболического и ФС нервных клеток в механизме различных психических феноменов, в частности, в мотивационно-эмоциональных реакциях. Предлагаемый способ позволяет регистрировать и дифференцировать переход одного функционального и метаболического состояния в другое, тем самым повышает точность диагностики патологических и нормальных состояний.
Таким образом, предложенный способ дает возможность на основе качественного анализа точно определить текущее функциональное и метаболическое состояние нервной ткани и адекватно дифференцировать физиологические и патологические явления в мозге, а также позволяет проводить разработку новых нейропротекторных препаратов и изучать механизмы патологических и физиологических состояний в эксперименте.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ ФУНКЦИОНАЛЬНОГО И МЕТАБОЛИЧЕСКОГО СОСТОЯНИЯ НЕРВНОЙ ТКАНИ | 2002 |
|
RU2245673C2 |
| СПОСОБ ОЦЕНКИ ФУНКЦИОНАЛЬНОГО И МЕТАБОЛИЧЕСКОГО СОСТОЯНИЯ НЕРВНОЙ ТКАНИ | 2014 |
|
RU2562230C1 |
| СПОСОБ ДИАГНОСТИКИ НАРУШЕНИЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ МОЗГА У БОЛЬНЫХ С ОПУХОЛЯМИ ГОЛОВНОГО МОЗГА | 2011 |
|
RU2473302C1 |
| Способ нейрокоррекции легких когнитивных нарушений у лиц пожилого возраста | 2022 |
|
RU2806235C1 |
| Способ диагностики нарушений функциональной активности мозга при вибрационной болезни, связанной с комбинированным воздействием локальной и общей вибрации | 2024 |
|
RU2837526C1 |
| СПОСОБ ПРОГНОЗА НЕЙРОИММУННЫХ НАРУШЕНИЙ ПРИ ИНСУЛИННЕЗАВИСИМОМ САХАРНОМ ДИАБЕТЕ | 2011 |
|
RU2459577C1 |
| СПОСОБ ДИАГНОСТИКИ НЕЙРОИММУННЫХ НАРУШЕНИЙ | 2006 |
|
RU2329762C2 |
| СПОСОБ ДИАГНОСТИКИ ТОКСИЧЕСКОЙ ЭНЦЕФАЛОПАТИИ У МЕЛКИХ ЛАБОРАТОРНЫХ ЖИВОТНЫХ ПРИ ХРОНИЧЕСКОМ ВОЗДЕЙСТВИИ ПАРОВ МЕТАЛЛИЧЕСКОЙ РТУТИ | 2010 |
|
RU2461893C2 |
| Способ неинвазивной слаботочной электростимуляции структур головного мозга | 2023 |
|
RU2820133C1 |
| Способ прогнозирования неблагоприятного течения эпилепсии у детей на первом году жизни | 2022 |
|
RU2787512C1 |

Изобретение относится к области медицины, а именно к нейрофизиологии. Одновременно регистрируют электроэнцефалограмму (ЭЭГ) и уровень постоянного потенциала (УПП), сравнивают их и по характеру изменения измеряемых параметров за один и тот же промежуток времени определяют функциональное и метаболическое состояние нервной ткани. При негативизации УПП и увеличении мощности ЭЭГ, сопровождающих деполяризационную активацию нейронов, определяют развитие плохого метаболического и функционального состояния (ФС) нервных клеток. При негативизации УПП и уменьшении мощности ЭЭГ, сопровождающих деполяризационное торможение нейронов, определяют развитие еще более плохого метаболического и ФС. При позитивизации УПП и увеличении мощности ЭЭГ, сопровождающих реполяризационную активацию нейронов после деполяризационного торможения, либо гиперполяризационную активацию, определяют развитие хорошего метаболического и ФС нервной ткани. При позитивизации УПП и уменьшении мощности ЭЭГ, сопровождающих либо реполяризационное торможение нейронов после деполяризационной активации, либо гиперполяризационное торможение, определяют развитие очень хорошего метаболического и ФС клеток нервной ткани. Изобретение обеспечивает новую качественную трактовку состояния нервной ткани. 1 табл., 3 ил.
Способ определения функционального и метаболического состояния нервной ткани, включающий одновременную регистрацию электроэнцефалограммы (ЭЭГ) и уровня постоянного потенциала (УПП), сравнение их и по характеру изменения измеряемых параметров за один и тот же промежуток времени определение функционального и метаболического состояния нервной ткани, отличающийся тем, что при негативации УПП и увеличении мощности ЭЭГ, сопровождающих деполяризационную активацию нейронов определяют развитие плохого метаболического и функционального состояния (ФС) нервных клеток, при негативизации УПП и уменьшении мощности ЭЭГ, сопровождающих деполяризационное торможение нейронов, определяют развитие еще более плохого метаболического и ФС, при позитивизации УПП и увеличении мощности ЭЭГ, сопровождающих реполяризационную активацию нейронов после деполяризационного торможения, либо гиперполяризационную активацию, определяют развитие хорошего метаболического и ФС нервной ткани, а при позитивизации УПП и уменьшении мощности ЭЭГ, сопровождающих либо реполяризационное торможение нейронов после деполяризационной активации, либо гиперполяризационное торможение,определяют развитие очень хорошего метаболического и ФС клеток нервной ткани.
| СПОСОБ ОПРЕДЕЛЕНИЯ ФУНКЦИОНАЛЬНОГО И МЕТАБОЛИЧЕСКОГО СОСТОЯНИЯ НЕРВНОЙ ТКАНИ | 2002 |
|
RU2245673C2 |
| СПОСОБ ИССЛЕДОВАНИЯ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ГОЛОВНОГО МОЗГА, УСТРОЙСТВО ДЛЯ ИССЛЕДОВАНИЯ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ГОЛОВНОГО МОЗГА И СПОСОБ ИЗМЕРЕНИЯ ПОДЭЛЕКТРОДНОГО СОПРОТИВЛЕНИЯ | 2003 |
|
RU2252692C2 |
| Щитковый успокоитель уровня воды в паровом котле | 1935 |
|
SU48344A1 |
| БАРАШКОВА А.Б | |||
| Клинико-функциональная и метаболическая характеристика формирования и прогнозирования уровня здоровья подростков, автореф | |||
| дисс, 2004, с.3-19 | |||
| MURIK S.E., SHAPKIN A.G | |||
| Simultaneous recording of EEG and direct current (DC) potential makes it possible to | |||

