Изобретение относится к методу обнаружения метилирования цитозина в образцах ДНК.
Уровни наблюдения, которые были хорошо изучены в молекулярной биологии по разработкам в методах в последние годы, включают сами гены, а также [транскрипцию и] трансляцию этих генов в РНК и далее белки. В процессе развития человека то, какой ген включается и как контролируется активация и ингибирование конкретных генов в некоторых клетках и тканях, можно коррелировать степенью и характером метилирования генов или генома. Патогенные состояния также экспрессируются модифицированной структурой метилирования отдельных генов или генома.
5-Метилцитозин представляет собой основание, которое наиболее часто модифицируется ковалентными связями в ДНК эукариотных клеток. Например, он играет роль в регуляции транскрипции, в генетическом импринтинге и в опухолеобразовании. Таким образом, идентификация 5-метилцитозина как компонента генетической информации представляет значительный интерес. Однако положения 5-метилцитозина нельзя идентифицировать секвенированием, поскольку поведенческая модель 5-метилцитозина в спаривании оснований та же, что и у цитозина. Кроме того, в случае ПЦР-амплификации эпигенетическая информация, которую несет 5-метилцитозин, полностью теряется.
Относительно новый метод, который со временем стал наиболее широко использоваться для исследования ДНК на 5-метилцитозин, основан на специфической реакции бисульфита с цитозином, который после последующего щелочного гидролиза преобразуют в урацил, поведенческая модель которого в спаривании оснований соответствует тимидину. В отличие от него эти условия не влияют на модификацию 5-метилцитозина. Таким образом, первоначальную ДНК преобразуют так, что метилцитозин, который первоначально невозможно отличить от цитозина по его поведению в гибридизации, можно выявить "стандартными" методами молекулярной биологии как единственно остающийся цитозин, например, амплификацией и гибридизацией или секвенированием. Все эти методы основаны на спаривании оснований, которое сейчас обширно используется. Известный уровень, касающийся чувствительности, определяется методом, который включает исследуемую ДНК в агарозную матрицу, что препятствует диффузии и ренатурации ДНК (бисульфит реагирует только на одноцепочечную ДНК) и все стадии осаждения и очистки заменяются быстрым диализом (Olek A., Oswald J., Walter J. A modified and improved method for bisulphate based cytosine methylation analysis. Nucieic Acids Res. 1996, Dec 15; 24 (24): 5064-6). Отдельные клетки можно исследовать этим методом, который иллюстрирует его возможности. Конечно, до настоящего времени были исследованы только отдельные участки длиной приблизительно до 3000 пар оснований; глобальное исследование клеток на тысячи возможных анализов на метилирование невозможно. Конечно, этим методом нельзя надежно анализировать очень малые фрагменты небольших количеств образца. Это приведет к потере несмотря на защиту от диффузии через матрицу.
Представление о других известных возможностях для обнаружения 5-метилцитозинов можно получить из обзорной статьи: Rein, Т., DePamphilis, M.L., Zorbas, H., Identifying 5-methylcytosine and related modifications in DNA genomes. Nucleic Acids Res. 1998 May 15; 26(10): 2255-64.
За некоторыми исключениями бисульфитный метод применялся только в исследованиях (например, Zeschnigk, М, Lich С, Suiting К, Buiting К, Horsthemke В. A single-tube PCR test for the diagnosis of Angetman and Prader-Willi syndrome based on allelic methylation differences at the SNRPN locus. Eur J. Hum. Genet. 1997 Mar-Apr; 5 (2): 94-98). Однако короткие специфические сегменты известного гена всегда амплифицируют после обработки бисульфитом и либо секвенируют полностью (Olek, A. and Walter, J. The preimplantation ontogeny of the H19 methylation imprint. Nat. Genet. 1997 Nov.; 17 (3): 275-6), либо отдельные положения цитозина (Gonzalgo, ML, Jones, PA. Rapid quantitation of methylation differences at specific sites using methylation-sensitive single nucleotide primer extention (Ms-SnuPE) Nucleic. Acids Res. 1997 Jun. 15; 25 (12): 2529-31, WO Patent 95-00669), или ферментную стадию (Xiong Z and Liard PW. COBRA: a sensitive and quantitative DNA methylation assay. Nucleic Acids. Res. 1997 Jun. 15; 25 (12): 2532-4) выявляют реакцией удлинения сегмента праймера. Было также описано обнаружение гибридизацией (Olek et al., WO-A 99/28498).
Мочевина улучшает эффективность обработки бисульфитом до секвенирования 5-метилцитозина в геномной ДНК (Paulin R, Grigg GW, Davey MW, Piper AA. Urea improves efficiency of bi sulphate-mediated sequencing of 5′-methylcytosine in genomic DNA Nucleic Acids Res. 1998 Nov. 1; 26 (21): 5009-10).
Есть другие публикации, связанные с применением бисульфитного метода для обнаружения метилирования в случае отдельных генов, это следующие публикации: Grigg G, Clark S. Sequencing 5-methylcytosine residues in genomic DNA. Bioassays. 1994 Jun.; 16, 431-6, 431; Zeschnik M, Schmitz В, Dittrich В, Buiting К, Horsthemke В, Dorfler W. Imprinted segments in the human genome: different DNA methylation patterns in the Prader-Willi/Angelman syndrome region as determined by the genomic sequencing method. Human Molecular Genetics 1997 Mar; 6 (3): 387-95; Feil R, Charlton J, Bird AP, Walter J, Reik W. Methylation analysis on individual chromosomes: improved protocol for bisulphate genomic sequencing. Nucleic Acids Res. 1994 Feb. 25; 22 (4): 695-6; Martin V, Ribieras S, Song-Wang X, Rio MC, Dante R. Genomic sequencing indicates a correlation between DNA hypomethylation in the 5'region of the pS2 gene and in its expression in human breast cancer cell lines. Gene. 1995 May 19; 157 (1-2); 261-4; WO 97/46705, WO 95/15373 и WO 95/45560.
Другим известным методом является так называемая чувствительная на метилирование ПЦР (Herman JG, Graff JR, Myohanen S, Nelkin BD, Baylin SB. (1996), Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci USA. Sep 3; 93 (18): 9821-6). По этому методу используют праймеры, которые гибридизируются или только с последовательностью, образованной обработкой бисульфитом ДНК, которая не метилирована в соответствующем положении, или наоборот, праймеры, которые связываются только с нуклеиновой кислотой, образованной обработкой ДНК бисульфитом, которая не метилирована(которая метилирована? - Прим. переводчика) в соответствующем положении. Затем с помощью таких праймеров получают амплифицированные продукты, обнаружение которых говорит о наличии метилированного или неметилированного положения в образце, с которым связываются праймеры.
Совсем недавно появившийся метод - выявление метилирования цитозина посредством проведения ПЦР в циклическом реакторе TaqMan, стал известен как "метил-лайт" (WO 00/70090). Таким методом представляется возможным выявить состояние метилирования отдельных положений или нескольких положений непосредственно в ходе ПЦР, что избавляет от необходимости проводить последующий анализ продуктов.
Обзор известного уровня в получении решетки олигомеров проведен в специальном издании Nature Genetics, опубликованном в январе 1999 года (Nature Genetics Supplement, Volume 21, January 1999) и источниках, указанных там, а также патенте США 5994065 по методам получения твердых носителей для молекул-мишеней, таких как олигонуклеотиды со сниженным неспецифичным фоновым сигналом.
Для сканирования решетки иммобилизированной ДНК использовали зонды с множественными флуоресцентными метками. Наиболее приемлемым способом для флуоресцентного мечения является простое введение Сy3 и Сy5 красителей в 5′-OH соответствующего зонда. Флуоресценция гибридизированных зондов обнаруживается, например, посредством конфокального микроскопа. Помимо многих других красители Сy3 и Сy5 имеются в свободной продаже.
Лазерная масс-спектрометрия десорбцией/ионизацией через матрицу (MALDI-TOF) является очень мощным достижением в анализе биомолекул (Karras М, Hillenkamp F., Laser desorption ionization of proteins with molecular masses exceeding 10,000 daltons. Anal. Chem. 1988 Oct. 15; 60 (20): 2299-2301). Анализируемое вещество заливают в светопоглощающую матрицу. Матрицу выпаривают короткой лазерной пульсацией и молекулу анализируемого вещества переносят нефрагментированной в газообразную фазу. Анализируемое вещество ионизируют столкновениями с молекулами матрицы. Приложенное напряжение ускоряет ионы в трубке полета, свободной от поля. Ионы ускоряются с различными скоростями в зависимости от их различных масс. Более мелкие ионы достигают детектора скорее, чем крупные.
MALDI-TOF спектроскопия очень хорошо подходит для анализа пептидов и белков. Анализ нуклеиновых кислот несколько затруднен (Gut, I.G. and Beck, S. (1995), DNA and Matrix Assisted Laser Desorption Ionization Nass Spectrometry. Molecular Biology: Current Innivations and Future Trends 1:147-157). Для нуклеиновых кислот чувствительность приблизительно раз в 100 хуже, чем для пептидов и сверхпропорционально снижается с увеличением размера фрагмента. Для нуклеиновых кислот, у которых основа с множественным отрицательным зарядом, процесс ионизации через матрицу гораздо менее эффективен. В MALDI-TOF спектрометрии выбор матрицы играет особенно важную роль. Для десорбции пептидов было найдено несколько очень мощных матриц, которые дают очень мелкую кристаллизацию. Для ДНК также были разработаны несколько эффективных матриц, но разница в чувствительности не была снижена. Разницу в чувствительности можно снизить химическим модифицированием ДНК таким образом, чтобы она напоминала пептид. Нуклеиновые кислоты фосфоротиоата, в которых обычные фосфаты основы замещены тиофосфатами, можно преобразовать простым алкилированием в ДНК с нейтральным зарядом (Gut I.G. and Beck, S (1995), A procedure for selective DNA alkylation and detection by mass spectrometry. Nucleic Acids Res. 23: 1367-1373). Связывание "метки заряда" с этой модифицированной ДНК обеспечивает повышение чувствительности на тот же порядок величины, которая выявлена для пептидов. Другим преимуществом "мечения зарядом" является повышение стабильности анализа в присутствии примесей, которые делают обнаружение немодифицированных подложек очень трудным.
Геномную ДНК получают из ДНК клеток, тканей или других тестуемых образцов стандартными методами. Эта стандартная методика представлена в ссылках, например, Fritsch and Maniatis, Molecular Cloning: A Laboratory Manual, 1989.
До настоящего времени в известном уровне существовало много методов анализа метилированием. Однако настоящее изобретение имеет своей целью решение задачи, которое недоступно для существующих методов (т.е.) амплифицировать мишенью исследуемую ДНК, присутствующую в жидкости тела или сыворотке человека, при наличии также других гомологичных в последовательности сегментов ДНК различного происхождения.
Исследуемую ДНК, а также другие присутствующие нуклеиновые кислоты, которые далее будут именоваться как фоновые ДНК, амплифицируют одинаково, поскольку используемые праймеры не способны различить исследуемую и фоновую ДНК. Однако существует одна возможность различить эти ДНК. Это посредством отличной структуры метилирования. Применяемый в настоящее время метод представляет собой чувствительную на метилирование ПЦР, сокращенно MSP (methylation-sensitive PCR) (Herman JG, Graff JR, Myohanen S, Nelkin BD, Baylin SB. (1996), Methylation-specific PCR: a novel PCR assay for Methylation status of CpG islands. Proc Natl Acad Sci USA. Sep 3; 93 (18): 9821-6). Этот метод включает несколько стадий. Первая: в соответствии с известным уровнем проводят обработку бисульфитам, в результате которой все основания цитозина преобразуются в урацил, в то время как метилированные основания цитозина (5-метилцитозин) остаются неизмененными. В следующей стадии используют праймеры, которые полностью комплементарны метилированной ДНК, преобразованной бисульфитом, но не соответствующей ДНК, которая присутствовала первоначально в неметилированном состоянии. Когда ПЦР проводят с таким праймером, амплифицируется исключительно изначально метилированная ДНК. Также можно использовать праймер, который, наоборот, амплифицирует только неметилированную ДНК. Таким образом при наличии и исследуемой ДНК и фоновой ДНК, будут получены только селективно и исключительно фрагменты исследуемой ДНК, поскольку они отличны от фоновой ДНК своим состоянием метилирования в CpG-положении. Таким образом, известный уровень не предполагает обнаружение такой молекулы ДНК, исследуемой либо на состояние метилирования, либо на присутствие исследуемой ДНК, что в свою очередь позволяет в принципе проведение диагностики, например, на наличие опухоли у больного, поскольку известно, что, например, у больных онкологией концентрация ДНК в сыворотке значительно возрастает. Помимо фоновой ДНК будет обнаружена только ДНК из опухолей. Такой анализ на ДНК можно проводить и с другими жидкостями в теле.
Однако метод, описанный здесь и считающийся наиболее близким известным уровнем, обладает определенными недостатками. Например, при выявлении амплифицированного фрагмента исследуемой ДНК невозможно определить количество, наличествующее в сыворотке. Даже минимальных количеств такой ДНК достаточно для получения положительного результата, что, с одной стороны, является преимуществом, а с другой, - нет, если необходимо определить, например, насколько удаление опухоли повлияет на уровень ДНК в сыворотке. Наибольшим неудобством является то, что в наличии имеется много положений метилирования, в которых исследуемая ДНК и фоновая ДНК отличаются только степенью. Очевидно, что существующий MSP метод можно проводить только, если известно, что фоновая ДНК точно и на 100% отличается от исследуемой ДНК в конкретном CpG-положении и если исследователь не хочет получить ложные положительные результаты. И наоборот, для опухолевой ткани типично, что специфическое положение присутствует в метилированном состоянии, т.е. в 95% опухолевых клеток, в которых другая присутствующая фоновая ДНК наличествует в метилированном состоянии только максимум в 5%, поэтому методом MSP невозможно получить информативные результаты, учитывая, что количественное определение матричной ДНК посредством ПЦР в принципе невозможно или возможно только при дополнительных затратах. Кроме того, изобретение основано на знании факта, что часто в фрагменте ДНК присутствуют структуры метилирования, которые типичны для специфического типа клетки, например опухолевой клетки.
Кроме того, известный уровень включает метод, разработанный Epigenomics, который одинаково амплифицирует и исследуемую ДНК и фоновую ДНК после обработки бисульфитом и затем методом гибридизации анализирует предшествующие CpG-положения в фрагменте, или посредством минисеквенирования или другим известным методом. Преимущество этого метода в том, что им можно определить количественную структуру исследуемых метилированных положений, т.е. анализ успешно определяет степень метилирования множественных положений, что позволяет, например, очень точную классификацию в случае твердых опухолей. Однако недостаток этого метода заключается в том, что он не дает точной информации в тех случаях, в которых сильно преобладает фоновая ДНК, так как эта информация точно амплифицируется наряду с исследуемой ДНК и обе они анализируются в смеси. Этой проблемы не существует в анализе твердых опухолей, в которых исследуемый материал можно выбрать целевым образом, но анализ, например, сыворотки ДНК затруднен.
Задача настоящего изобретения состоит в том, чтобы избавиться от недостатков известного уровня и соединить преимущества обоих методов обнаружения в жидкостях тела и сыворотке.
Задача решается методом обнаружения степени метилирования цитозина в образцах ДНК, включающим следующие стадии:
образец геномной ДНК, включающий исследуемую ДНК и фоновую ДНК, химически обрабатывают таким образом, что все не метилированные основания цитозина преобразуют в урацил, в то время как основания 5-метилцитозина остаются неизмененными;
химически обработанный образец ДНК амплифицируют посредством по меньшей мере двух праймеров-олигонуклеотидов, а также полимеразы, в результате чего исследуемая ДНК становится матричной по отношению к фоновой ДНК, и
амплифицированные продукты анализируют и делают выводы о состоянии метилирования исследуемой ДНК из наличия амплифицированного продукта и/или из анализа других положений.
Образец ДНК желательно получать из сыворотки или других жидкостей тела человека.
Конкретнее, образец ДНК берут предпочтительно из клеточных линий, крови, мокроты, стула, мочи, сыворотки, цереброспинальной жидкости, ткани, залитой парафином, например ткани глаз, кишечника, почек, мозга, сердца, предстательной железы, легкого, молочной железы или печени, гистологических слайдов или других их возможных комбинаций.
В соответствии с изобретением наиболее предпочтительно проводить химическую обработку бисульфитом ( = дисульфитом, кислым сульфитом). Желательно также, чтобы химическую обработку проводили после заливки ДНК в агарозу. Кроме того, еще желательно, чтобы в химической обработке дополнительно присутствовал реагент, денатурирующий дуплекс ДНК, и/или ловушка для радикалов.
На второй стадии амплификацию желательно проводить в присутствии по меньшей мере одного добавочного олигонуклеотида, который связывается с 5′-CG-3′-динуклеотидом или 5′-TG-3′-динуклеотидом или 5′-CA-3′динуклеотидом, в результате чего добавочный олигонуклеотид связывается с фоновой ДНК и тормозит ее амплификацию.
Желательно, чтобы этот связующий сайт добавочного олигонуклеотида или ПНК олигомера и связующие сайты праймеров на фоновой ДНК перекрывались и добавочный олигонуклеотид замедлял связывание по меньшей мере одного праймера-олигонуклеотида с фоновой ДНК.
Кроме того, желательно применять по меньшей мере два добавочных олигонуклеотида или ПНК олигомера, в результате чего каждый из их связующих сайтов перекрывается в свою очередь со связующим сайтом одного праймера на фоновой ДНК и добавочные олигонуклеотиды и/или ПНК олигомеры замедляют связывание обоих олигонуклеотидов-праймеров с фоновой ДНК.
Особенно предпочтительно, чтобы один из добавочных олигонуклеотидов и/или ПНК олигомеры препятствовали связыванию прямого праймера, в то время как другой препятствовал связыванию обратного праймера.
Желательно также, чтобы концентрация добавочных олигонуклеотидов и/или ПНК олигомеров по меньшей мере в пять раз превышала концентрацию олигонуклеотидов-праймеров.
В другом наиболее предпочтительном варианте метода добавочные олигонуклеотиды и/или ПНК олигомеры связываются с фоновой ДНК и, таким образом, препятствуют полной элонгации олигонуклеотида-праймера в реакции полимеразы. Это тот случай, когда используемая полимераза не обладает активностью 5′-3′-экзонуклеазы. В другом предпочтительном варианте присутствующие добавочные олигонуклеотиды модифицированы на 5′ конце и не могут существенно разрываться полимеразой с 5′-3′ экзонуклеазой.
Кроме того, в соответствии с изобретением желательно амплифицировать химически обработанный ДНК образец на второй стадии с использованием по меньшей мере 2 праймеров-олигонуклеотидов и другого олигонуклеотида, который гибридизируется с 5′-CG-3′-динуклеотидом, или 5′-TG-3′-динуклеотидом, или 5′-CA-3′-динуклеотидом, и по меньшей мере, одного олигонуклеотида-репортера, который гибридизируется с 5′-CG-3′-динуклеотидом, или 5′-TG-3′-динуклеотидом, или 5′-СА-3′-динуклеотидом, а также полимеразой; в результате чего добавочный олигонуклеотид связывается с фоновой ДНК и сдерживает ее амплификацию, и в результате чего олигонуклеотид-репортер связывается с исследуемой ДНК и указывает на ее амплификацию. В дополнение к олигонуклеотиду-репортеру желательно использовать другой олигомер, помеченный флуоресцентным красителем, так чтобы этот другой олигомер гибридизировался с участком, непосредственно прилегающим к олигонукпеотиду-репортеру, и эту гибридизацию можно было обнаружить посредством передачи резонансной энергии флуоресценции. Анализ желательно проводить на циклических реакторах TaqMan и LightCycle.
В соответствии с изобретением предпочтительно, чтобы олигонуклеотиды, используемые в дополнение к праймерам, не имели 3′ функции. Желательно, чтобы олигонуклеотид-репортер имел по меньшей мере одну флуоресцентную метку. И молекулы репортера указывали амплификацию либо усилением, либо ослаблением флуоресценции. Особое преимущество дает то, что усиление или ослабление флуоресценции также непосредственно используется для анализа и состояние метилирования исследуемой ДНК можно определить по флуоресцентному сигналу.
В соответствии с изобретением предпочтительно, чтобы концентрация имеющейся фоновой ДНК в 100 раз превышала концентрацию исследуемой ДНК. Еще лучше, чтобы концентрация имеющейся фоновой ДНК в 1000 раз превышала концентрацию исследуемой ДНК.
Анализ или дополнительный анализ лучше проводить посредством гибридизации с олигомерными решетками, где олигомерами могут быть нуклеиновые кислоты или молекулы, схожие с ними по своим свойствам гибридизации, такие как ПНК.
В соответствии с изобретением желательно, чтобы олигомеры гибридизировались с анализируемой ДНК посредством сегмента длиной в 12-22 основания и включали CG-, TG- или СА-динуклеотид.
Состояние метилирования более 20 положений метилирования анализируемой ДНК лучше выявлять в одном эксперименте.
Более того, состояние метилирования более 60 положений метилирования анализируемой ДНК лучше выявлять в одном эксперименте.
Также желательно анализ или дополнительный анализ проводить измерением длины исследуемой амплифицированной ДНК, и методы измерения длины включают гель-электрофорез, капиллярный гель-электрофорез, хроматографию (например, ЖХВР), масс-спектрометрию и другие подходящие методы. Преимущество также дает использование методов Санджера и Максам-Гилберта для секвенирования и других методов, таких как секвенирование гибридизацией (SBH).
Предпочтителен также метод, в котором секвенирование для каждого CpG-положения или небольшой группы этих положений проводят каждый раз отдельным олигонуклеотидом-праймером и удлинение сегмента праймера включает только одно или несколько оснований, и по типу элонгации праймера можно сделать вывод о состоянии метилирования соответствующих положений в ДНК, предназначенной для исследования.
О наличии расстройства или другого клинического состояния больного можно сделать вывод из степени метилирования различных исследуемых CpG-положений.
Преимущество заключается также в том, что сами амплифицированные продукты для их выявления снабжены обнаруживаемой меткой. И метки являются флуоресцентными или представляют собой радионуклиды или съемные метки массы, которые обнаруживаются масс-спектрометром.
Также желательно, чтобы в амплификации один из праймеров был связан с твердой фазой.
В соответствии с изобретением предпочтительно обнаружение всех амплифицированных продуктов в масс-спектрометре и четкая их характеристика по массе.
Другим объектом настоящего изобретения является использование метода по изобретению для диагностики и/или прогнозирования неблагоприятных явлений для больных, если такие неблагоприятные явления входят по меньшей мере в одну из следующих категорий: нежелательная лекарственная интерференция; раковые заболевания; CNS дисфункции, нарушения или заболевания; симптомы агрессии или поведенческие реакции; клинические, психологические или социальные последствия нарушения мозговой деятельности; психотические расстройства и изменения личности; старческое слабоумие и/или сопутствующие синдромы; сердечно-сосудистые заболевания, расстройства и нарушения; расстройства, нарушения и заболевания желудочно-кишечного тракта; расстройства, нарушения и заболевания дыхательной системы; повреждения, воспаление, инфекция, состояния иммунитета и/или выздоровления; дисфункции, нарушения или заболевания организма как отклонение в развитии; патология, повреждения или заболевания кожи, мышц, соединительной ткани или костей; нарушение, дисфункции или заболевания эндокринной системы и обменных процессов; головные боли или расстройства функции половой сферы.
Метод по изобретению также применяется для различения типов клеток или тканей или для исследования клеточной дифференциации.
Объектом изобретения является также комплект, включающий реагент, содержащий бисульфит, праймеры и другие олигонуклеотиды без 3′-ОН функции для получения амплифицированных продуктов, а также инструкцию для проведения анализов по изобретению.
Таким образом, настоящее изобретение описывает метод обнаружения состояния метилирования образцов геномной ДНК. В отличие от ранее известных методов степень метилирования ряда CpG-положений определяют в выбранной подгруппе фрагментов ДНК, например в сыворотке, так что анализ также возможен в присутствии избытка ненужной для диагностики фоновой ДНК.
Предпочтительный метод состоит из нескольких этапов, которые можно суммировать следующим образом.
Первый. У больного берут сыворотку или другие жидкости тела и при необходимости изолируют выявленную в них ДНК. На второй стадии проводят химическую обработку, предпочтительно бисульфитом ( = кислым сульфитом, дисульфитом), в которой, например, все неметилированные основания цитозина преобразуют в урацил за исключением метилированных оснований цитозина (5-метилцитозин), которые остаются неизмененными. На третьей стадии метода проводят амплификацию, в которой амплифицируют ДНК, предназначенную для исследования, но не фоновую ДНК, или последнюю амплифицируют только в небольшой степени. На следующей четвертой стадии амплифицированные фрагменты анализируют на их сигнатуру метилирования и определяют степень метилирования нескольких предшествующих CpG-положений а амплифицированных продуктах. На пятой стадии метода из степени метилирования в разных исследуемых CpG-положениях делают вывод о наличии расстройства или другого клинического состояния больного.
Суть настоящего изобретения в том, что два типа CpG-положений играют роль и равно участвуют в анализе и ниже именуются как положения "определителя" и положения "классификатора". Цель положений определителя - отличить в амплификации анализируемую ДНК от фоновой ДНК. Технически это можно сделать различными способами, как представлено подробнее ниже. Свойство этих положений состоит в том, что их степень метилирования в исследуемой ДНК очень существенно отличается от степени метилирования фоновой ДНК, и это создает преимущество в амплификации для исследуемой ДНК. Положения же классификатора служат цели экстрагирования информации из амплифицированного продукта важной для диагностики и полученной из исследуемой ДНК посредством соответствующей степени метилирования. До нескольких сот таких положений классификатора можно использовать для одного анализа, если анализ проводить, например, на олигомерных решетках, хотя в этом часто нет необходимости. Однако в этом случае значение для результата исследования имеет не образование специфического амплифицированного продукта, а анализ CpG-положений в том же амплифицированном продукте. Однако в некоторых случаях можно и нужно отнести информацию, полученную из образования амплифицированного продукта, к анализу; в этом случае несколько положений представляют собой и классификатор и определитель.
Первую стадию метода, получение образцов, проводят забором жидкостей тела, таких, как, например, слюны или сыворотки, но очевидно, что метод можно проводить со многими видами образцов из разных источников, которые указаны здесь без претензий на их полноту.
Предпочтительно геномную ДНК, используемую в методе, получают из образца ДНК, источники которого включают, например, клеточные линии, кровь, мокроту, стул, мочу, сыворотку, цереброспинальную жидкость, ткань, залитую парафином, например ткань глаз, кишечника, почки, мозга, сердца, предстательной железы, легкого, молочной железы или печени, гистологические слайды или другие их возможные комбинации.
В некоторых случаях очистку или концентрирование ДНК проводят до обработки бисульфитом, с тем чтобы избежать прерывания реакции с бисульфитом и/или последующей ПЦР из-за слишком высокой концентрации примесей. Однако известно, например, что ПЦР можно начать с тканью после обработки, например, протеиназой К без последующей очистки, и это также относится к обработке бисульфитом и последующей ПЦР.
Химическую обработку предпочтительно проводят обработкой бисульфитом (=кислым сульфитом, дисульфитом), лучше бисульфитом натрия (бисульфит аммония менее пригоден). Реакцию проводят либо в соответствии с известным вариантом, когда ДНК заливают в агарозу, чтобы удерживать ее в одноцепочечном состоянии во время обработки, либо по новому варианту, когда обработку проводят в присутствии ловушки для радикалов и денатурирующего агента, предпочтительно олигоэтиленгликольдиалкилэфира, например диоксана. До ПЦР реакции реагенты удаляют либо промыванием в случае метода с агарозой, либо методом очистки ДНК (известный уровень, осаждение или связывание с твердой фазой, мембрана), или простым разбавлением в концентрации, которая существенно не влияет на ПЦР.
На третьей стадии очень важно выбрать положения определителя и подходящий метод, что позволяет провести селективную амплификацию ДНК, предназначенной для исследования. Выбор положений делается исходя из того, что их функция - максимальное различение фоновой ДНК и исследуемой ДНК по степени метилирования. Для этой цели определяют профили метилирования сегментов генов, которые анализируют каждый раз как для исследуемых опухолей, так и для фоновой ДНК от здоровых людей. Те положения, которые обладают наибольшим различием между опухолевой ДНК и фоновой ДНК (например, в сыворотке), отбирают как положения определителя. Такие положения уже известны для большого числа генов, например, для GSTpi, HIC-1 MQMT (von Wronski МА, Harris LC, Tano К, Mitra S, Bigner DD, Brent TP. (1992) Cytcosine methylation and suppression of 06-methylguanine-DNA. methyltransferase expression in human rhabdomyosarcoma cell lines and xenografts. Qncol Res.; 4 (4-5): 167-74; Esteller M, Toyota M, Sanchez-Cespedes M, Capella G, Peinado МА., Watkins DN, Issa JP, Sidransky D, Baylin SB, Herman JG. (2000), Inactivation of the DNA. repair gene 06-methylguanine-DNA. methyltransferase by promoter hypermethylation is associated with G to A mutations in K-ras in colorectal tumorigenesis. Cancer Res. May 1; 60 (9): 2368-71). Теперь в наличии есть несколько методов, которые целиком предпочтительны и посредством которых исследуемую ДНК амплифицируют с использованием этих положений определителя.
Прежде всего представляется возможным провести реакцию, соответствующую MSP, использованием праймеров, которые полностью гибридизируются с последовательностью, соответствующей исследуемой ДНК после обработки бисульфитом, но не аналогично обработанной фоновой ДНК. Другими словами, праймеры гибридизируются с сегментами ДНК, в которых есть одно или несколько положений определителя, и только если их состояние метилирования в первоначальной ДНК соответствует тому, которое характерно для исследуемой ДНК, амплификация имеет место в значительной степени. По сути это простой вариант, но обладает тем недостатком, что положения определителя каждый раз размещаются на одном или обоих концах фрагмента ДНК, т.е. положения классификатора должны располагаться между положениями определителя (или, в случае только одного положения определителя последний не должен находиться в середине положений классификатора). Хотя по изобретению и желательно проводить такой вариант, его фактически можно применять только в относительно небольшом числе случаев, поскольку распределение положений определителя и классификатора будет идеальным только в нескольких случаях. Однако поскольку он прост в проведении, его приводят здесь как предпочтительный в любом случае.
Однако наиболее предпочтителен вариант, в котором праймеры не перекрываются положением определителя или не гибридизируются с последним, а на амплификацию ПЦР влияет по меньшей мере еще один олигонуклеотид, который не несет функции праймера и который связывается на положении определителя.
Это означает, что в принципе химически обработанная ДНК, как в известном уровне, амплифицируется посредством двух праймеров. Одно или несколько положений определителя находятся внутри сегмента ДНК, связанного двумя праймерами. В отличие от стандартной ПЦР вводятся добавочные олигонуклеотиды, которые связываются с этими положениями определителя, и фактически делают это селективно в зависимости от того, были последние в метилированном или неметилированном состоянии до обработки бисульфитом. Соответственно исследуемая ДНК амплифицируется, если добавочные олигонуклеотиды менее эффективно связываются с положениями определителя, чем с фоновой ДНК. Другими словами, введенные олигонуклеотиды блокируют амплификацию фоновой ДНК.
Предпочтительно эти введенные олигонуклеотиды включают по меньшей мере один CG-, один TG- или один СА-динуклеотид. Кроме того, они должны обладать свойством, не позволяющим им элонгироваться полимеразой, применяемой в ПЦР. С этой целью вводят 3′-деоксиолигонуклеотиды или олигонуклеотиды с другими функциями в положении 3′, например, 3′-O-ацетилолигонуклеотиды. Кроме того, необходимо воспрепятствовать разложению этих олигонуклеотидов полимеразой. Это осуществляют либо применением полимеразы, не обладающей активностью нуклеазы, или применением модифицированных олигонуклеотидов, которые имеют, например, тиоат-мосты на 5′ конце и, таким образом, устойчивы к разложению.
Другой очень подходящий вариант представляет собой применение ПНК-олигомеров (пептидной нуклеиновой кислоты), которые используют в этом эксперименте как олигонуклеотиды. Олигомеры ПНК не подвержены разложению полимеразой и не могут ею элонгироваться, так что они идеально подходят для этого варианта метода. А методы построения и синтеза олигомеров ДНК представляют собой известный уровень.
Как уже упоминалось, в таких методах применяют несколько положений определителя и также несколько олигонуклеотидов, каждый из которых специфичен для состояния метилирования, наличествующего в фоновой ДНК.
После селективной амплификации исследуемой ДНК определяют состояние метилирования нескольких положений классификатора в соответствии с известными методами.
Однако очевидно, что даже в этом случае образование самого определенного фрагмента ПЦР в отдельном случае может представлять достаточную информационную ценность, поскольку так же, как для MSP, в такой ситуации положение определителя находится в неметилированном состоянии практически до 100% в фоновой ДНК, но в метилированном состоянии в исследуемой ДНК. Если в ПЦР использовать олигонуклеотид, который связывается с последовательностью, образующейся от неметилированной фоновой ДНК в обработке бисульфитом, в ПЦР образуется только один продукт, поскольку в общем присутствует по меньшей мере небольшое количество исследуемой ДНК. В отдельном случае даже этого может быть достаточно для проведения диагностики, и это включает метод со свойствами, аналогичными свойствам MSP. Хотя такая процедура не всегда предпочтительна, этот метод до сих пор не был известен и его также можно отнести к объекту данного изобретения.
В ПЦР реакции желательно одновременное получение нескольких фрагментов, т.е. проведение мультиплексной ПЦР. В схеме реакции следует предусмотреть, чтобы не только праймеры, но и добавочные олигонуклеотиды не были комплементарны по отношению один к другому; иначе в этом случае труднее достичь высокой степени мультиплексности. Однако в случае с ДНК, обработанной бисульфитом, есть преимущество, заключающееся в том, что прямой праймер никогда не функционирует как обратный праймер по причине разного G и С содержания двух ДНК нитей, что, в свою очередь, облегчает мультиплексное образование и значительно компенсирует этот недостаток.
В самом простом случае образующиеся фрагменты обнаруживаются. Для такого обнаружения приемлемы все возможные известные методы молекулярной биологии, такие как гель-электрофорез, секвенирование, жидкая хроматография или гибридизация без анализа олигонуклеотидов классификатора. Они приемлемы также для качественного контроля предшествующих стадий метода. Однако, как уже упоминалось ранее, особенно предпочтителен последующий анализ степени метилирования положений классификатора.
Существует ряд возможностей выгодно объединить амплификацию исследуемой ДНК вышеописанный методом с методом обнаружения для олигонуклеотидов классификатора.
Наиболее приемлемые методы обнаружения представляют собой гибридизацию с олигомерными решетками и, например, реакции удлинения сегмента праймера (мини-секвенирование). Гибридизацию с олигомерными решетками применяют без введения дополнительных изменений в протокол ближайшего известного уровня (Olek A, Olek S, Walter J; WO Patent 99 28498). Однако лучше гибридизировать амплифицированные продукты с решеткой олигомеров, которая состоит из пар олигонуклеотидов, иммобилизированных на твердой фазе, один из которых каждый раз гибридизируется с сегментом ДНК, содержащим первоначально не метилированный CpG (положение классификатора), а другой из которой гибридизируется с соответствующим сегментом, в котором содержался первоначально метилированный CpG, при этом каждый раз до обработки бисульфитом и амплификации. В этом случае желательно пометить амплифицированный продукт или амплифицированные продукты флуоресцентными или радиоактивными метками или съемной меткой массы с тем, чтобы после гибридизации на основании этой метки можно было обнаружить и дать количественный анализ фрагментов, связанных с двумя олигонуклеотидами пары. После калибрования эксперимента с полностью метилированной и неметилированной ДНК получают соотношение интенсивности, из которого определяют, например, степень метилирования на соответствующем положении классификатора. На такой олигомерной решетке (фиг.1) одновременно можно определить множественное число фрагментов и положений классификатора. Предпочтительно и существенно то, что решетка также содержит олигомеры, выявляющие положения определителя для мониторинга эксперимента, так как представляется возможным определение соотношения исследуемой ДНК, введенной в анализ, и фоновой ДНК.
К тому же реакции удлинения сегмента праймера можно проводить на олигонуклеотидах, иммобилизированных на твердой фазе. Хотя это совсем не обязательно, лучше эти праймеры иммобилизировать, поскольку придется исследовать очень большое число положений классификатора, включающих несколько амплифицированных продуктов, и проведение исследования на твердой фазе, здесь на олигомерной решетке, будет значительно проще и осуществлено в одном эксперименте. Желательно также, чтобы праймеры были расположены вплотную прилегающими к положению классификатора и чтобы элонгация составляла только один нуклеотид. В качестве нуклеотидов предпочтительнее вводить только дидеокситимидин и дидеоксицитидин, каждый из которых следует помечать разным флуоресцентным красителем, хотя возможно, и даже предпочтительно применение и любых других меток, например, таких, как метки массы. После обработки бисульфитом и амплификации ранее метилированный CG остается CG, а ранее не метилированный CG теперь становится TG. Таким образом, результатом реакции удлинения сегмента праймера будет включение или дидеоксицитидина или дидеокситимидина. Степень метилирования соответствующего положения определяется из соотношения флуоресцентных меток, обнаруженных каждый раз для этих двух терминаторов. В этом случае, кроме того, представляется возможным и желательным провести удлинение сегмента праймера дидеоксицитидином и дидеокситимидином, если не присутствует производное гуанина и, следовательно, удлинение сегмента праймера заканчивается без других дополнений в TG- или CG-последовательности даже после одного основания. Аналогичным образом анализ на контр-нити проводят различением СА и CG посредством дидеокси-АТФ и дидиокси-GTP или их производных.
Однако особенно предпочтителен вариант метода, в котором в одном эксперименте проводят одновременное обнаружение положений определителя и положений классификатора, и который можно осуществить применением вариантов методики TaqMan или LightCycler. Добавочные меченные флуоресцентным красителем олигонуклеотиды вводят в олигонуклеотиды, которые обеспечивают амплификацию исследуемой ДНК, и во время ПЦР замеряют изменения в флуоресценции. В принципе, поскольку исследуемую ДНК амплифицируют, информацию о состоянии метилирования различных CPG-положений классификатора получают непосредственно из этого изменения в флуоресценции. Поскольку каждый из разных олигонуклеотидов снабжен разным флуоресцентным красителем, для каждого отдельного положения во время ПЦР возможно отчетливое изменение флуоресценции.
Такого изменения в флуоресценции в зависимости от состояния метилирования можно добиться разными методами, два из которых описаны ниже на примерах.
Прежде всего применяют зонды олигонуклеотидов, которые специфично связываются либо с последовательностью, возникшей благодаря химической обработке ДНК, не метилированной в соответствующем положении, либо соответствующей последовательности, которая образовалась посредством химической обработки ДНК, метилированной в соответствующем положении. Эти зонды снабжают двумя флуоресцентными красителями, подавляющим красителем и флуоресцентным красителем, служащим в качества маркера. Оба они связаны с одним и тем же зондом-олигонуклеотидом, и если в ПРЦ исследуемая ДНК выступает в роли матричной ДНК, на этот раз реакция будет блокироваться зондом-олигомером с флуоресцентным мечением. Однако поскольку последний неустойчив к активности нуклеазы в полимеразе, во время ПЦР-реакции происходит разложение зонда, связанного с матричной ДНК, что коррелируется со связующей эффективностью зонда с матрицей, так как несвязанный зонд не разлагается полимеразой. Благодаря тому факту, что подавляющий краситель и флуоресцентный краситель, служащий маркером, отделены друг от друга, разложение зонда непосредственно видимо при усилении флуоресценции красителя маркера. Фактически это положено в основу для проведения анализа по варианту так называемого метода TaqMan.
Соответственно измеряется образование ПЦР продукта из исследуемой ДНК, но только если исследуемое положение классификатора также присутствует в метилированном состоянии, которое зонд может обнаружить гибридизацией с химически обработанной ДНК. Таким образом нужен контр-образец, содержащий зонд, который соответственно связывается с положением классификатора в другом состоянии метилирования.
Различные флуоресцентные красители с разными длинами волн излучения применяют на нескольких зондах вместе с подавляющим красителем, что дает способность различить эти зонды и обеспечить мультиплексность.
Даже при таком анализе применяют олигонуклеотиды, связывающиеся с положениями определителя, что препятствует значительной амплификации фоновой ДНК. Амплификацию исследуемой ДНК можно анализировать таким образом, что одно и то же положение исследуется зондам, как описано выше, и амплификацию соответственно обнаруживают зондом, связывавшимся с определителем. В этом случае лучше, если олигонуклеотид, не подверженный разложению, селективно связывается с фоновой ДНК, а зонд, помеченный флуоресцентным красителем, связывается с исследуемой ДНК. В наиболее предпочтительном варианте метода у зонда и олигонуклеотида, не подверженного разложению, одна и та же последовательность за исключением одного нуклеооснования, но в любом случае, не более двух нуклеооснований.
Особенно хорош вариант, в котором несколько способных к метилированию положений определены как положения определителя и по меньшей мере один олигонуклеотид, предпочтительно связанный с фоновой ДНК, а также зонд используют для этих положений. Поскольку амплификация фоновой ДНК в этом случае подавляется несколькими олигонуклеотидами, этот метод особенно пригоден в случаях, когда избыток фоновой ДНК особенно высок в сравнении с исследуемой ДНК. Во многих случаях в случае таких вариантов и при наличии нескольких положений определителя в одном фрагменте дальнейшее исследование положений классификатора будет излишним, поскольку с современным оборудованием очень большое число разных красителей нельзя выявить одновременно (по большей части, это число [разных красителей] составляет 4-5). Поэтому исследование положений добавочных классификаторов лучше проводить одним из способов обнаружения, представленных выше.
Кроме того, лучше, если степень метилирования нескольких положений будет исследоваться одновременно одним зондом. Если необходимо более точное количественное определение степени метилирования положений классификатора, используют два зонда с разными красителями, где один из них связывается в случае неметилированного положения в исследуемой ДНК, а другой связывается в обратном случае, в случае метилированного положения. Из соотношения интенсивности флуоресценции двух красителей делается вывод о степени метилирования исследуемого положения.
Кардинально отличный метод, в котором во время ПЦР также имеет место изменение флуоресценции, известен как техника LightCyclerтм. В нем используют тот факт, что передача резонансной энергии флуоресценции (FRET) может происходить только между двумя красителями, если они располагаются в непосредственной близости друг от друга, т.е. в пределах 1-5 нуклеотидов. Только в этом случае второй краситель возбуждается излучением первого красителя и в свою очередь излучает свет с другой длиной волны, которую выявляют. В данном случае анализа на метилирование гибридизация меченного флуоресцентной меткой зонда с обработанной химически соответствующим образом ДНК имеет место в положении классификатора, и связывание этого зонда в свою очередь, зависит от того, в какой форме, метилированной или неметилированной, исследуемая ДНК присутствовала в этом положении. Другой зонд с другим флуоресцентным красителем связывается непосредственно с частью, прилегающей к этому зонду. Второе связывание происходит также в зависимости от метилирования, если в соответствующем сегменте последовательности имеется другое способное метилироваться положение. Во время амплификации ДНК множатся, в результате чего еще большее количество зондов с флуоресцентным мечением связывается с участком, прилегающим к соответствующему положению, так как последнее имеет состояние метилирования, необходимое для этого, и таким образом измеряют усиление передачи резонансной энергии флуоресценции (FRET).
При этом методе также происходит образование мультиплекса с несколькими зондами, меченными разными флуоресцентными красителями.
При этом желательно и возможно измерить положение определителя. Если предположить, что фоновая ДНК присутствует в неметилированной форме в соответствующем положении и после химической обработки и амплификации в этом положении оказывается TG-динуклиотид, и наоборот, исследуемая метилированная ДНК дает CG-динуклеотид, зонд с флуоресцентной меткой будет связываться с последовательностью, содержащей CG, а конкурентный олигомер без метки связывается с соответствующей TG-последовательностью фоновой ДНК.
Важным представляется то, что неметилированный олигонуклеотид препятствует амплификации благодаря своей более высокой температуре плавления в отличие от более коротких олигонуклеотидов-зондов. В этой связи, поскольку зонды и олигомеры, связывающиеся с химически обработанной фоновой ДНК, не идентичны за исключением нескольких оснований, они существенно длиннее (на 5-15 оснований). Можно и желательно применять модифицированные олигонуклеотиды и/или ПНК. В этом случае все зонды и олигонуклеотиды блокируются за исключением праймеров на своем 3′конце, с тем чтобы избежать элонгации в ПЦР. Это делается, например, фосфатной группой.
Два метода отличаются принципиально тем, что в одном случае замеряют ослабление флуоресценции, а в другом - усиление. В обоих случаях измеряют и положения определителя и положения классификатора.
В общем, наиболее предпочтителен метод обнаружения метилирования цитозина в образцах ДНК, в котором проводят следующие стадии: Первая, образец геномной ДНК, включающий исследуемую ДНК и фоновую ДНК, обрабатывают химически таким образом, что все неметилированные основания цитозина преобразуют в урацил, при этом основания 5-метилцитозина остаются неизмененными; далее обработанный химически образец ДНК амплифицируют применением по меньшей мере двух олигонуклеотидов-праймеров, а также полимеразы, в результате чего исследуемая ДНК становится матричной по отношению к фоновой ДНК, и на следующей стадии амплифицированные продукты анализируют, и из наличия амплифицированного продукта и/или из анализа добавочных положений делают вывод о состоянии метилирования в исследуемой ДНК.
В наиболее предпочтительном варианте изобретения образец ДНК получают из сыворотки или других жидкостей тела человека. Другие источники образцов ДНК включают клеточные линии, кровь, мокроту, стул, мочу, сыворотку, цереброспинальную жидкость, ткань, залитую парафином, например ткань глаз, кишечника, почки, мозга, сердца, предстательной железы, легкого, молочной железы или печени, гистологические слайды или другие их возможные комбинации.
В наиболее предпочтительном варианте метода химическую обработку проводят бисульфитом ( = кислым сульфитом, дисульфитом). Химическую обработку лучше проводить после заливки ДНК агаровой. Лучше, если при химической обработке присутствует реагент, денатурирующий дуплекс ДНК, и/или ловушка для радикалов.
В наиболее предпочтительном варианте метода амплификацию проводят на второй стадии в присутствии по меньшей мере одного добавочного олигонуклеотида, который связывается с 5′-CG-3′-динуклеотидом, или 5′-TG-3′-динуклеотидом, или 5′-СА-3′-динуклеотидом, в то время как другой олигонуклеотид связывается с фоновой ДНК и тормозит ее амплификацию.
В наиболее предпочтительном варианте метода химически обработанный образец ДНК амплифицируют на второй стадии применением по меньшей мере 2 олигонуклеотидов-праймеров и другого олигонуклеотида, который гибридизируется с 5′-CG-3′-динуклеотидом, или 5′-TG-3′-динуклеотидом, или 5′-CA-3′-динуклеотидом, и по меньшей мере одного олигонуклеотида-репортера, который гибридизируется с 5′-CG-3′-динуклеотидом, или 5′-TG-3′-динуклеотидом, или 5′-СА-3′-динуклеотидом, вместе с полимеразой; а добавочный олигонуклеотид предпочтительно связывается с фоновой ДНК и сдерживает ее амплификацию, при этом олигонуклеотид-репортер связывается с исследуемой ДНК и указывает на ее амплификацию.
Предпочтительно также, чтобы помимо олигонуклеотида-репортера применяли другой олигомер, помеченный флуоресцентным красителем, который гибридизируется непосредственно вплотную к олигонуклеотиду-репортеру, и эту гибридизацию можно обнаружить посредством передачи резонансной энергии флуоресценции (FRET).
В анализе желательно воспользоваться методикой TaqMan. Хорошо еще провести анализ и по методике LightCycler (как описано выше).
Применяемые в дополнение к праймерам олигонуклеотиды не имеют 3′-ОН функции. И олигонуклеотид-репортер должен иметь по меньшей мере одну флуоресцентную метку. Особенно желательно, чтобы молекулы репортера указывали амплификацию либо усилением, либо ослаблением флуоресценции и это усиление или ослабление флуоресценции применяли непосредственно для анализа и из флуоресцентного сигнала делали вывод о состоянии метилирования исследуемой ДНК.
В наиболее предпочтительном варианте метода анализ или дополнительный анализ проводят посредством гибридизации с олигомерными решетками, в которых олигомерами могут быть нуклеиновые кислоты или молекулы, аналогичные им по своим свойствам гибридизации, такие как ПНК. При этом олигомеры гибридизируются с исследуемой ДНК сегментом длиной в 12-22 основания и включают CG-, TG- или СА-динуклеотид. Этим методом в одном эксперименте исследуют состояние метилирования более 20 положений метилирования в одном эксперименте, а еще лучше более 60 положений метилирования.
Также особенно предпочтителен метод, в котором дополнительный анализ проводят измерением длины исследуемой амплифицированной ДНК и который включает гель-электрофорез, капиллярный гель-электрофорез, хроматографию (например, ЖХВР), масс-спектрометрию и другие подходящие методы.
Особенно предпочтителен метод, в котором дополнительный анализ проводят секвенированием и методы секвенирования включают метод Санджера, метод Максама-Гилберта и другие, такие как секвенирование гибридизацией (SBH). Предпочтителен метод, в котором секвенирование (по Санджеру) проводят для каждого из CpG-положений или для небольшой группы этих положений и каждый раз отдельным олигонуклеотидом-праймером и удлинение сегмента праймера включает только одно основание или только несколько оснований и состояние метилирования соответствующих положений в исследуемой ДНК определяется типом удлинения сегмента праймера.
В наиболее предпочтительном варианте метода о наличии расстройства или другого клинического состояния больного можно сделать вывод из степени метилирования различных исследуемых CpG-положений.
Преимущество заключается также в том, что сами амплифицированные продукты снабжены обнаруживаемой меткой для их выявления. И метки являются флуоресцентными или представляют собой радионуклиды или съемные метки массы, которые обнаруживаются масс-спектрометром.
Кроме того, предпочтителен метод, в котором в амплификации один из праймеров связан с твердой фазой.
А также метод, в котором амплифицированные продукты обнаруживаются масс-спектрометром в целом и четко характеризуются по массе.
Другим объектом настоящего изобретения является использование одного из описанных методов для диагностики и/или прогнозирования неблагоприятных явлений для больных, если такие неблагоприятные явления входят по меньшей мере в одну из следующих категорий: нежелательная лекарственная интерференция; раковые заболевания; CNS дисфункции, нарушения или заболевания; симптомы агрессии или поведенческие реакции; клинические, психологические или социальные последствия нарушения мозговой деятельности; психотические расстройства и изменения личности; старческое слабоумие и/или сопутствующие синдромы; сердечно-сосудистые заболевания, расстройства и нарушения; расстройства, нарушения и заболевания желудочно-кишечного тракта; расстройства, нарушения и заболевания дыхательной системы; повреждения, воспаление, инфекция, состояния иммунитета и/или выздоровления; дисфункции, нарушения или заболевания организма как отклонение в развитии; патология, повреждения или заболевания кожи, мышц, соединительной ткани или костей; нарушение, дисфункции или заболевания эндокринной системы и обменных процессов; головные боли или расстройства функции половой сферы.
Кроме того, применение одного из описанных методов желательно для различения типов клеток или тканей или для исследования клеточной дифференциации.
Другим объектом настоящего изобретения является комплект, включающий реагент, содержащий бисульфит, праймеры и добавочные олигонуклеотиды без 3′-ОН функции для получения амплифицированных продуктов, а также инструкцию для проведения по меньшей мере одного из описанных вариантов метода.
Изобретение иллюстрируют следующие примеры.
Пример 1:
Чувствительная на метилирование амплификация MDR1 гена с применением блокирующих зондов ПНК (фиксации ПЦР) в MDR-1 гене.
а) Специфическая на метилирование ПЦР с зондами ПНК (блокирующими ПНК зондами), связующие сайты которых не перекрывают сайты праймеров.
Первое, для обработанной бисульфитом ДНК определены условия ПЦР, при которых опознается аллель-специфическое влияние блокирующего зонда ПНК на ПЦР. В этом первом эксперименте связующие сайты между ПНК и праймерами не перекрываются.
Влияние 11-mer ПНК последовательности AAAATGTGTT тестировали в ПЦР с праймерами TAAGTATGTTGAAGAAAGATTATTGTAG и TAAAAACTATCCCATAACTCCCAAC. Воздействие ПНК на ПЦР-реакцию невозможно определить при стандартных условиях ПЦР с температурой отжига 55°С. Стандартная программа циклического реактора, применяемого для амплификации MDR-1 фрагмента, включает следующие стадии программы:
Стадия 1: тем. = 96°С 20 мин
Стадия 2: тем. = 96°С 30 сек
Стадия 3: тем. = 56°С 1.15 мин
Стадия 4: тем. = 72°С 2.00 мин
Стадии 2-4 проводят 40 циклов.
Стадия 5: тем. = 96°С 15 мин
Стадия 6: Охлаждение до 4°С и поддержание температуры
Следовательно, длина праймеров, температура отжига и длина образца ДНК должны быть оптимально подобраны, чтобы достичь аллель-специфического подавления амплификации.
Чтобы достичь оптимального отжига праймеров и ПНК, протестировали более короткие 21-mer праймеры TAAGTATGTTGAAGAAAGATT и AATCCCCATAAACTTACCAAA с более длинной 13-mer ПНК AAAGACGTGTTAT. Дополнительные тесты проводили с 18, 19 и 20-mer праймерами, которые отличаются от указанных последовательностей только тем, что на 3′ конце основания отсутствуют. 21-mer праймеры тестировали с градиентом температуры отжига. В параллельной партии 13-mer ПНК AAAGACGTGTTAT или ПНК AAAGATGTGTTAT, предназначенные для последовательности, полученной из неметилированного аллеля, добавляли в разных концентрациях от 20 до 100 пмоль/мкл.
Четкое влияние на ПЦР наблюдали с температурами отжига 49.4°С и 46.7°С с добавлением ПНК в концентрации 70 и 100 пмоль/мкл. Наиболее видимый эффект был с применением 18-mer праймера.
Далее исследовали, насколько более низкая температура элонгации в 54°С влияет на тормозящую активность ПНК.
С этой целью к партии ПЦР добавляли каждую из вышеуказанных 13-mer ПНК или обе 13-mer ПНК вместе в концентрации 50 и 70 пмоль/мкл. В сравнении с положительным контролем без ПНК можно было наблюдать явное подавление ПЦР (Фиг.2, агароза-гель-электрофорез, концентрации ПНК для 13-mer ПНК зондов; аннотация: MDR 1-5 ЕМ: 13-mer ПНК AAAGATGTGTTAT). Тесты делают очевидным, что используемые в них ДНК присутствуют в основном в неметилированном состоянии в интересующих нас положениях, что подтверждает бисульфитное секвенирование. Поэтому в этом эксперименте видна четкая аллель-специфичность блокирующего ПНК зонда, что обеспечивает нужную амплификацию неметилированных фрагментов.
b) Специфическая на метилирование ПЦР с ПНК зондами (блокирующими ПНК зондами), связующие сайты которых перекрывают сайты праймеров ("исключение праймера").
В вышепроведенных экспериментах праймеры выбирали таким образам, чтобы мишень-последовательность ПНК размещалась приблизительно в центре участка, предназначенного для амплификации. Такое расположение описывалось как более чувствительное, если последовательность праймера и ПНК прилегали друг к другу или перекрывались. В случае связывания ПНК со специфической последовательностью при таком расположении воздействие ПНК наблюдалось даже с меньшими концентрациями ПНК.
Для этого эксперимента выбирали праймер с последовательностью TTATGTGAATTTTGAAAG, так чтобы он перекрывался последовательностью ПНК ("исключение праймера"). Обе 13-mer ПНК AAAGACGTGTTAT и AAAGATGTGTTAT добавляли к загрузкам в реакцию в 3 разных концентрациях.
Полное подавление ПЦР реакции наблюдали даже с концентрацией в 25 пмоль/мкл. В предшествующем эксперименте полного подавления ПЦР можно было достичь добавлением обеих ПНК только в концентрации, достигающей 70 пмоль/мкл.
с) Исключение праймера с фрагментами, соответствующими не метилированной ДНК
Для обнаружения связывания со специфической последовательностью действие ПНК на матрицы с подробными характеристиками по состоянию метилирования исследовали в дополнительных экспериментах. Матричной ДНК в этом эксперименте была полностью неметилированная обработанная бисульфитом ДНК.
Ее использовали как матрицу в ПЦР. Программа включала следующие стадии:
Стадия 1: тем. = 96°С 20 мин
Стадия 2: тем. = 96°С 30 сек
Стадия 3: тем. = 49°С 1.15 мин
Стадия 4: тем. = 54°С 2.00 мин
Стадии 2-4 проводили 36 циклов.
Стадия 5: тем. = 72°С 15 мин
Стадия 6: Охлаждение до 4°С и поддержание температуры.
К партиям реакции, как указывалось выше, 13-mer [ПНКы] добавляли в трех разных концентрациях. В этом случае с не метилированной матрицей предполагалось, что MDR 1-5 FU (3) ПНК, адаптированный к этой матрице, будет явно сильнее влиять, чем MDR 1-5-FM (3). На Фиг.3 показано (для аннотации см. Фиг.2), что это уже имеет место даже в случае сравнительно низких концентраций ПНК.
Эти результаты показывают, что подавление ПЦР-реакции благодаря специфически связывающим ПНК делает возможным получение специфических на метилирование амплификаций. В контрольных экспериментах с ПНК, не комплементарными MDR-1, эти реакции не оказывают сколько-нибудь заметного влияния на ПЦР в указанных концентрациях (не показано).
Пример 2: Чувствительная к метилированию амплификация фрагмента GSTpi-гена.
Специфические CpG-положения GSTPi-гена были определены как маркеры опухоли для рака предстательной железы. В паре праймеров, выбранных для экспериментов, GGAAAGAGGGAAAGGTTTT и TACTAAAAACTCTAAACCCCAT, праймер локализован так, что он точно связывает ПНК-последовательность CCCCGAAAACGCG (или CCCTGAAAATGTG). Здесь ПНК последовательность содержит три релевантные CpG-положения в отличие от положений ПНК, используемых для MDR1 фрагмента. Релевантные CpG фрагментов GSTPi присутствуют в "нормальной" ДНК в неметилированном состоянии, т.е. ДНК от здоровых людей. Таким образом, на ПЦР реакцию должен заметно повлиять присутствующий в партии реакции "GSTP-ниже" ПНК с последовательностью CCCTGAAAATGTG, а не соответствующий "GSTP-выше", CCCCGAAAACGCG.
ПНК "GSTP-ниже" добавляли к тестуемой партии в трех разных концентрациях и тестировали градиент температуры отжига.
Результаты эксперимента показаны на Фиг.4. Наибольшее подавление ПЦР определяли введением ПНК (GSTP-ниже") при температуре отжига 55°С. В сравнении с положительным контролем (без введения ПНК) было выявлено, что ПЦР реакция значительно подавляется введением ПНК в концентрации 20 [р]моль/мкл.
Далее, чтобы обнаружить связывание со специфической последовательностью (и таким образом, в конечном анализе чувствительное к метилированию связывание), сравнивали воздействие "GSTP-выше" и GSTP-ниже" ПНК на амплификацию не метилированной ДНК в первоначальном образце и метилированной ДНК из ткани опухолевой предстательной железы как матрицы.
Неметилированную ДНК и ДНК из больного органа применяли в ПЦР как матрицы. "GSTP-вьше" и GSTP-ниже" ПНК вводили в партии реакции в трех разных концентрациях.
Введение "GSTP-ниже" ПНК имело видимое влияние на ниженеметилированную матрицу: ПЦР полностью подавлялась, когда ПНК вводили в концентрации 70 пмоль/мкл. С другой стороны, в случае введения "GSTP-выше" ПНК было отмечено только слабое действие торможения ПНК на ПЦР.
Однако введение "GSTP-выше" ПНК в тестуемую ДНК ткани предстательной железы оказывало значительное торможение на ПЦР (Фиг.5). Введение ПНК в концентрации 20 пмоль/мкл явно подавляло ПЦР. Полное подавление ПЦР определяли добавлением ПНК в концентрации 50 пмоль/мкл. И наоборот, добавление "GSTP-ниже" к ДНК больной железы оказывало явно меньшее воздействие. Даже добавление ПНК в концентрации 70 пмоль/мкл не подавляло ПЦР полностью.
Эксперименты показывают, что посредством блокирующих зондов олигомера возможно селективное подавление амплификации метилированных или неметилированных аллелей на определенных положениях. В значении этого изобретения соответствующими положениями являются положения определителя, т.е. представляется возможным селективно подавлять амплификацию нежелательных метилированных матриц фоновой ДНК.
Пример 3: Различные возможности для применения зондов, подавляющих ПЦР в зависимости от состояния матилирования на примере GSTPi-гена.
На Фиг.6 представлено несколько вариантов, как расположить праймеры для данной матричной последовательности в применении на чувствительность к метилированию. На Фиг.6а представлены матрицы после обработки бисульфитом: ДНК 1 соответствует первоначально метилированному образцу ДНК и ДНК 2 соответствует первоначально не метилированному образцу ДНК.
На Фиг.6B представлено расположение одного из праймеров в применении аллель-специфической ПЦР или специфической на метилирование ПЦР (MSP). В этом случае с применением показанного праймера имеет место амплификация только метилированной ДНК 1.
На Фиг.6С и 6D показано, как соответственно можно применять праймеры специфические для неметилированного состояния либо использованием дегенерированных положений (6С), либо универсальных оснований (здесь, инозина) на Фиг.6D.
На Фиг.7 представлено несколько вариантов, как праймеры и зонды ("блокаторы") следует располагать для амплификации, реагирующей на метилирование, в случае данной матричной последовательности. В этих примерах не сами применяемые праймеры являются чувствительными к метилированию, а специфичность к метилированию определяется только зондами ("блокаторами"). Специфические зонды применяют каждый раз в качестве блокаторов для ДНК 1 и ДНК 2.
Праймеры и зонды не перекрываются на Фиг.7А, а зонды прямо соединены с 3′ концом праймера. Показанный зонд представляет собой олигонуклеотид, модифицированный на 3′ конце, который сам не может элонгироваться в амплификации. Аналогично можно применять РНК, но они должны быть короче в этом примере и соответствовать своей температуре плавления. Тот же праймер применяют на Фиг.7B, но здесь зонд ДНК перекрывает праймер (исключение праймера).
На Фиг.7С и 7D праймер с дегенерированными положениями и зонд специфичный на метилирование перекрываются. Аналогично в праймерах могут использоваться также универсальные основания.
На Фиг.8 показан пример, аналогичный прямому и обратному праймерам, где используется зонд, который не перекрывает ни один из праймеров.
Эти примеры иллюстрируют многочисленные возможности того, как олигомерные зонды можно применять для специфичной на метилирование амплификации для подавления фоновой ДНК в отличие от анализируемой ДНК. Однако объем изобретения не ограничивается вариантами, описанными здесь в качестве примеров.
Пример 4: Получение неметилированной и метилированной ДНК и обработка бисульфитом.
Для получения метилированной ДНК человеческую геномную ДНК обрабатывали S-аденозилметионином и CpG-метилазой (SssI, New England Biolabs) в соответствии с инструкцией производителя. Для получения неметилированной ДНК фрагмент гена ELK-1 амплифицировали посредством ПЦР праймерами GCTCTATGGTCTTGTCTAACCGTA (Seq. ID: 1) и AGGTGGTGGTGGCGGTGG (Seq. ID: 2) из геномной ДНК человека. Полученную таким образом неметилированную и метилированную ДНК, а также геномную ДНК человека обрабатывали бисульфитом (кислым сульфитом, дисульфитом), так что все цитозины, не метилированные в 5-положении основания, подвергали модификации таким образом, что образовывалось основание с другой поведенческой моделью в спаривании оснований, в то время как цитозины, метилированные в 5-положении, оставались неизмененными. Если для реакции использовать бисульфит в пределах концентрации между 0.1 M и 6 М, присоединение имеет место на неметилированных основаниях цитозина. Кроме того, должны присутствовать денатурирующий реагент или растворитель, а также ловушка для радикалов. Далее последующий щелочной гидролиз ведет к преобразованию оснований нуклеиновых кислот неметилированного цитозина в урацил. Эта преобразованная ДНК служит для обнаружения метилированных цитозинов.
Пример 5: Получение зондов гена, меченных Сy5
Начиная с образцов ДНК, обработанных бисульфитом, амплифицировали взятый фрагмент длиной в 529 пар оснований от участка промотора ELK-1-гена. Амплификацию проводили олигонуклеотидами-праймерами ATGGTTTTGTTTAATYGTAGAGTTGTTT (Seq. ID: 3) и TAAACCCRAAAAAAAAAAACCCAATAT (Seq. ID: 4). При использовании олигонуклеотидов-праймеров, помеченных флуоресцентной краской Сy5, фрагмент непосредственно помечали в ПЦР. В качестве матричной ДНК применяли (1) неметилированную ДНК, (2) метилированную ДНК и (3) геномную ДНК человека, обработанные бисульфитом (кислым сульфитом, дисульфитом). Затем эти три разных фрагмента ДНК исследовали в отдельных гибридизациях на их степень метилирования в специфическом CpG-положении.
Пример 6:
Проведение гибридизации и оценка "чипа" с гибридизированной ДНК.
Зонды гена, полученные в Примере 5, гибридизировали с ДНК на чипе. Сначала олигонуклеотиды иммобилизировали на чипе. Последовательности олигонуклеотидов получали из амплифицированного фрагмента гена ELK-1, указанного в Примере 2, и они представляли собой CG-динуклеотиды, включающие непосредственно прилегающие участки. Длина олигонуклеотидов доходила до 14-22 нуклеотидов; положение CG-динуклеотида внутри олигонуклеотида было разным. После гибридизации ДНК-чип сканировали (см. Фиг.1) и сигналы гибридизации оценивали по номерам (данные не представлены). Результаты гибридизации для олигонуклеотидов CTACTCAACGAAAACAAA (Seq. ID: 5) и CTACTCAACAAAACAAA (Seq. ID: 6) показаны на Фиг.1. Здесь CTACTCAACGAAAACAAA (Seq. ID: 5) гибридизируется, если цитозин ELK-1 фрагмента, который выявлен в положении 103 амплифицированного продукта, метилирован; и CTACTCAACAAAAACAAA (Seq. ID: 6) гибридизируется, если этот цитозин не метилирован.
ДНК-чип показан на Фиг.1 после гибридизации с фрагментом промотора. Псевдо-цветовое изображение показано сразу после его получения сканированием. В отличие от черно-белого изображения, показанного здесь, сканнером получено цветовое изображение. Интенсивность разных цветовых оттенков представляет степень гибридизации, и степень гибридизации уменьшается от красного (это представлено в виде светлых пятен на Фиг.1) до синего (представлено темными пятнами на Фиг.1).
Пример 7: Получение матричной ДНК и проведение GSTpl-ПЦР
В качестве матричной ДНК использовали необработанную ДНК человека из периферической крови (Promega, Madison [WI] USA), метилированную ферментами in vitro, которую подвергли обработке бисульфитом. Для метилирования всех CG-динуклеотидов в соответствии с инструкцией производителя 6 мкг ДНК в реакторе объемом 150 мкл подвергли реакции с SssI (New England Biolabs, Frankfurt/Main). Обработку бисульфитом проводили в соответствии с известным методом (Olek A, Oswald J, Walter J. A modified and improved method for bisulphate based cytosine methylation analysis. Nucleic Acids Res. 1996 Dec 15; 24 (24: 5064-6).
GSTpl-фрагмент длиной в 153 пар оснований (положения 1242-1393) в последовательности №М24485.1) амплифицировали бисульфит-ДНК-специфичными праймерами 2cf GTTTT (СТ) GTTATTAGTGAGT и 2cr TCCTAAATCCCCTAAACC в реакторе объемом в 25 мкл (1 × буфер реакции, Qiagen; 1 U HotstarTaq, Qiagen; 200 мкл каждого dNTP, 500 нМ каждого праймера, 0.05-10 нг обработанной бисульфитом матричной ДНК) при следующих условиях ПЦР (95°С - 15 мин; 46 циклов: 96°С - 0:45 мин, 52°С - 0:45 мин, 72°С - 0:20 мин; 72°С - 10 мин) (см. Фиг.9 и 10). Секвенирование GSTpl-фрагментов показало, что в человеческой ДНК из периферической крови нет метилированных CG-динуклеотидов для этого фрагмента, в то время как все CG-динукпеотиды присутствуют в метилированной форме в ДНК, обработанной SssI (см. Фиг.9). Секвенирование GSTpl-фрагмента подтверждает другие результаты (см. например, WO 99/55905), что в GSTpl-гене в отличие от опубликованной последовательности (Genbank Асс №24485.1) присутствует добавочный G-нуклеотид (между положениями 1273 и 1274 в Genbank Асс. №2485.1; положение 33 в GSTpl-фрагменте ПЦР, см. Фиг.9). Что касается эффективности ПЦР, между CpG-метилированной и CpG-не метилированной матричной ДНК разницы нет. (см. Фиг.10).
Пример 8: Селективная амплификация метилированных GSTpl-фрагментов.
Экспериментальная схема селективной амплификации метилированных GSTpl-фрагментов показана на Фиг.11. Амплификации GSTpl-фрагмента праймерами 2cf GTTTT (СТ) GTTATTAGTGAGT и 2сr ТССТАААТССССТАААСС на неметилированной матричной ДНК препятствуют два блокирующих олигонуклеотида (В5 + 9FT6, GTGAGTATGTGTGGTTTGTGT-P; B15 + 17RT11, TAAACCCCCATCCCAAATCTCA-Р, см. Фиг.11), последовательность которых соответствует неметилированной обработанной бисульфитом ДНК. Эти олигонуклеотиды модифицированы на 3′ конце фосфатной группой, препятствующей их элонгации во время ПЦР. ПЦР проводили в реакторе объемом в 25 мкл со следующей программой циклов (95°С - 15 мин; 46 иклов: 96°С - 0:45 мин, 52°С - 0:45 мин, 72°С - 0:20 мин; 72°С - 10 мин) (см. Фиг.9 и 10). Партия ПЦР включала: (1 × буфер реакции, Qiagen, Hilden) 2 U HotstarTaq, (Qiagen, Hilden); 200 мкл каждого dNTP, 500 нМ каждого праймера, 10 мкМ каждого блокатора (В5 + 9FT6, GTGAGTATGTGTGGTTTGTGT-P и B15 + 17RT11, TAAACCCCCATCCCAAATCTCA-P, 20 нг - 20 пг обработанной бисульфитом матричной ДНК. При таких условиях ПЦР возможно полное подавление амплификации GSTpl-фрагмента в объеме 25 мкг неметилированной матричной ДНК (см. Фиг.12А, ряд 8). Если ПЦР проводить без блокирующих олигонуклеотидов, GSTpl-фрагмент амплифицируется (см. Фиг.12D, ряд 8). В отличие от этого GSTpl-ПЦР продукт можно обнаружить при тех же условиях ПЦР с блокирующими олигонуклеотидами и без них на 100 пг метилированной матричной ДНК (см. Фиг.12С, ряд 7; 12F ряд 7). Таким образом, абсолютная чувствительность ПЦР показывают по меньшей мере 100 пг метилированной матричной ДНК.
Чтобы исследовать относительную чувствительность ПЦР, были подготовлены смеси неметилированной и метилированной матричной ДНК, в которых соотношение неметилированной и метилированной ДНК составляло от 1:1 до 1:1000. Для получения смесей этих ДНК ДНК человека из периферической крови (Promega, Madison [WI] USA) смешивали с Sssl-обработанной ДНК (см. Пример 7) в соответствии с нужным соотношением и подвергали обработке бисульфитом. Результаты ПЦР на этих смесях матричных ДНК (25 мкг общей смеси ДНК), проведенной с блокирующими олигонуклеотидами и без них, показаны на Фиг.12А, В или Фиг.12D, Е. Приведенные результаты показывают, что одну копию метилированного гена GSTpl можно репродуктивно обнаружить на фоне 200 копий неметилированного GSTpl-гена (см. Фиг.12А, ряд 6; В, ряд 6). Относительной чувствительности в соотношении 1:1000 можно достичь дальнейшей оптимизацией условий ПЦР (см. Фиг.12В, ряд 7).
Анализы последовательности продуктов ПЦР, амплифицированных от смеси ДНК (1:200, соотношение неметилированной и метилированной ДНК) с блокаторами (см. Фиг.12А, ряд 6) и без блокаторов (см. Фиг.12D, ряд 6) показали ожидаемые результаты. Продукт ПЦР, полученный в ПЦР без блокатора, соответствовал неметилированному GSTpl-гену, и наоборот, фрагмент GSTpl-гена, полученный в ПЦР с блокирующими олигонуклеотидами, имел метилированное эпигенетическое состояние.
Успешно были протестированы также другие 3′ модификации, такие как ddNTP или добавочные нуклеотиды, которые не могут соответствовать соответствующей последовательности GSTpl-нуклеотида.
Пример 9: Селективная амплификация метилированных GSTpl-фрагментов на LightCycle.
LightCycle (Roche) - устройство, предназначенное для проведения ПЦР и одновременного обнаружения и анализа продуктов ПЦР. Работа на устройстве проводилась по инструкции производителя. Количественный и качественный анализы ПЦР проводили по версии программы LightCycle Software Version 3.5.
Селективную амплификацию метилированных фрагментов GSTpl-гена проводили в 10 мкл объема реакции (1 × буфер реакции (Qiagen Hilden) 5 U HotstarTaq (Qiagen Hilden); 200 мкМ каждого dNTP, 625 нМ каждого праймера (2cf GTTTT(CT) GTTATTAGTGAGT; 2 cr TCCTAAATCCCCTAAACC, 4 мкМ блокатора (В5 + 9 FT16, GTGAGTATGTGTGGTTTGTGTT-P), 0.25 мкг/мкл BSA (Sigma, Munich), 250 нМ якорного олигонуклеотида (GSTpl-Fluo, TTTAGAGTTTTTAGTATGGGGTTAATT-флуоресцеин; TibMolBiol, Berlin), 250 нМ зонда гибридизации (GSTpl-красный 705, красный 705-GTATTAGGTTTGGGTTTTTGGT-P; TibMolBiol, Berlin) и/или GSTpl-красный 650, красный 650-TAGTATTAGGTTCGGGTTTTCGG-P, TibMolBiol, Berlin), 20 нг - 200 пг матричной ДНК) со следующей программой циклического реактора: 95°С-15 мин; 46 циклов: денатурация 96°С - 4 сек, отжиг 52°С - 30 сек; удлинение 72°С - 20 сек. Обнаружение проводили на каждом цикле амплификации генспецифичными и специфичными на метилирование зондами обнаружения LightCycler на стадии отжига после 10 сек. Фрагмент GSTpl ПЦР был обнаружен, когда оба якорный зонд, не специфичный на метилирование, GSTpl Fluo, и один из зондов, специфичных на метилирование GSTpl-красный 705 или GSTpl-красный 650 гибридизировались с фрагментом ПЦР.
С целью проверки специфичности зондов обнаружения на метилирование 15 нг каждой обработанной бисульфитом метилированной и неметилированной матричной ДНК амплифицировали в LightCycler. ПЦР включала якорный зонд GSTpl-Fluo и эквимолярную смесь зондов гибридизации GSTpl-красного 705 и GSTpl-красного 650. Флуоресценцию зонда измеряли в F2/F1 канале обнаружения LightCycler для метилированного GSTpl-гена, GSTpl-650, а зонд для неметилированного GSTpl-гена, GSTpl-красный 705, обнаруживали в канале F3/F1 (см. Фиг.13). Эксперименты показали, что GSTpl-красный 650 специфически обнаруживает метилированный GSTpl-ген, а неметилированный вариант не дает флуоресцентного сигнала (см. Фиг.13А). И наоборот, зонд GSTpl-красный 705 обнаруживает неметилированный и метилированный GSTpl-ген, но обнаруживает последний со значительно сниженной эффективностью (см. Фиг.13В).
Аналогично Примеру 8 исследовали абсолютную и относительную чувствительности амплификации метилированного фрагмента GSTpl. Для определения абсолютной чувствительности GSTpl-ПЦР проводили в LightCycler на различных количествах метилированной обработанной бисульфитом матричной ДНК с блокирующими олигонуклеотидами и без них. Дополнительно с "якорным" зондом для обнаружения применяли зонд гибридизации GSTpl-красный 650. Результаты приведены на Фиг.14. Расчет "точек скрещивания" проводили версией программы 3.5 LightCycler, и он показал количество циклов ПЦР, в которых продукт GSTpl ПЦР обнаруживали впервые более сильным сигналом в отличие от отрицательного на контрольном образце. Это означает, что чем ниже величина "точки скрещивания", тем более эффективно проходила амплификация фрагмента GSTpl. Если "точка скрещивания" не указана, это означает, что ПЦР продукт не обнаружен. В показанном эксперименте GSTpl можно амплифицировать 75 пг метилированной обработанной бисульфитом матричной ДНК, даже при наличии блокатора. (см. Фиг.14).
Для определения относительной чувствительности GSTpl-ПЦР проводили с 20 нг смесей матричных ДНК (см. Пример 8) с блокирующими олигонуклеотидами и без них (см. Фиг.15). Для обнаружения ПЦР включала "якорный" зонд GSTpl-Fluo и эквимолярную смесь зондов гибридизации GSTpl-красного 705 и GSTpl-красного 650. Флуоресценцию зонда измеряли в F2/F1 канале обнаружения LightCycler для метилированного GSTpl-гена, GSTpl-красного 650, а зонд для не метилированного GSTpl-гена, GSTpl-красного 705 обнаруживали в канале F3/F4.
Определенные "точки скрещивания" показали, что в ПЦР с блокирующим олигонуклеотидом одну копию метилированного гена GSTpl можно репродуктивно обнаружить на фоне 500 копий неметилированного GSTpl-гена (см. Фиг.15, "положение ротора" 17, колонка F2/F1). Это соответствует абсолютной чувствительности 40 пг метилированной матричной ДНК. Без блокирующих олигонуклеотидов можно достичь только относительной чувствительности в соотношении 1:10 (см. Фиг.15, "положение ротора" 3, колонка F2/F1). При таких же условиях амплификация GSTpl-гена из 15 нг неметилированной обработанной бисульфитом ДНК подавляется полностью (см. Фиг.15, "положение ротора" 19 и 9, колонка F3/F1).
Пример 10: Селективная амплификация метилированных GSTpl-фрагментов на TaqMan.
(TaqMan (Applied Biosystems, Weiterstadt) - другое устройство, предназначенное для проведения ПЦР и одновременного обнаружения и анализа продуктов ПЦР. Работа на устройстве проводилась по инструкции производителя. Количественный и качественный анализы ПЦР проводили по программ TaqMan Software.
Селективную амплификацию метилированных фрагментов GSTpl-гена проводили в 20 мкл объема реакции (1 х буфер реакции (Applied Biosystems) 2 U AmplitagGold (Applied Biosystems); 3.5 мМ MgCl2, 400 мкМ каждого dNTP, 500 нМ каждого праймера (2cft GTTTT(CT) GTTATTAGTGAGTA; 2 cr TCCTAAATCCCCTAAAACC, блокатор 1, 7.5 мкМ (В5 + 9 FT6, GTGAGTATGTGTGGTTTGTGT-P), блокатор 2, 7.5 мкМ (В15 + 17RT19, TAAACCCCCCATGCCAAATCTC-P), 450 нМ зонда TaqMan (Taql, Blackhole-TAATTCGTAGTATTAGGTTCGGGTTTTCGGTAGGG-FAM; Biosearch Technologies, 10 нг матричной ДНК) со следующей программой циклического реактора: 95°С - 10 мин; 3 цикла: денатурация 96°С - 15 сек, отжиг 60°С - 60 сек; 3 цикла: денатурация 96°С - 15 сек, отжиг 58°С - 30 сек; удлинение 60°С - 30 сек; 3 цикла: денатурация 96°С - 15 сек, отжиг 55°С - 30 сек, удлинение 60°С - 30 сек; 40 циклов: денатурация 96°С - 15 сек, отжиг 52°С - 30 сек, удлинение 60°С - 40 сек). Обнаружение проводили на каждом цикле амплификации генспецифичными зондами TaqMan после стадии удлинения.
Для определения относительной чувствительности GSTpl-ПЦР проводили с 10 нг смесей матричных ДНК {см. Пример 8) с блокирующими олигонуклеотидами и без них (см. Фиг.16). Расчет "порогового цикла" проводили программой TaqMan и он показал количество циклов ПЦР, в которых продукт GSTpl-ПЦР обнаруживали впервые более сильным сигналом в отличие от отрицательного на контрольном образце в сравнении с величинами "точки скрещивания" на LightCycle. Это означает, что чем ниже величина "порогового цикла", тем более эффективной была амплификация фрагмента GSTpl.
Определенные величины "порогового цикла" показали, что в ПЦР с блокирующим олигонуклеотидом одну копию метилированного гена GSTpl можно обнаружить на фоне 200 копий неметилированного GSTpl гена (см. Фиг.16). Это соответствует абсолютной чувствительности 50 пг метилированной матричной ДНК. При таких же условиях амплификациия GSTpl-гена из 10 нг неметилированной обработанной бисульфитам матричной ДНК подавляется полностью (см. Фиг.16).
Описание Фиг.:
Фиг.10: Агарозный гель фрагментов GSTpl-ПЦР. ПЦР проводили с 10 нг, 5 нг, 1 нг, 0.5 нг и 0.1 нг метилированной (А) и неметилированной (В) обработанной бисульфитам матричной ДНК.
Фиг.11: Последовательность фрагмента GSTpl с положениями праймеров и блокирующих олигонуклеотидов.
Фиг.12: Агарозные гели фрагментов GSTpl-ПЦР. Были проанализированы относительная чувствительность амплификации метилированного GSTpl-гена (А, В, D, Е) и абсолютная чувствительность (С, F) амплификации метилированного GSTpl-гена. ПЦР гена GSTpl проводили с блокирующим олигонуклеотидом (А, В, С) и без блокирующего олигонуклеотида (D, Е, F). В качестве матричной ДНК применяли: 20 нг метилированной обработанной бисульфитом ДНК (A1, B1, D1, E1, C1, F1, С2, F2) И 20 нг неметилированной обработанной бисульфитом ДНК (А8, В8, D8, Е8); метилированную и неметилированную обработанную бисульфитом ДНК в соотношении смеси 1:2 (А2, В2, D2, Е2), 1:10 (A3, В3, D3, Е3), 1:20 (А4, В4, D4, Е4), 1:100 (А5, В5, D5, Е5), 1:200 (А6, В6, D6, Е6), 1:1000 (А7, В7, D7, Е7); 10 нг (С3, F3), 2 нг (С4, F4), 1 нг (С5, F5), 0.2 нг (С6, F6), 0.1 нг (С7, F7), 0.02 нг (С8, F8), метилированную обработанную бисульфитом ДНК и без ДНК (А9, D9).
Фиг.13: Анализ специфичности метилированния зондов обнаружения. На графике показано проявление флуоресценции на протяжении GSTpl-ПЦР метилированной обработанной бисульфитом ДНК (сплошная линия) и неметилированной обработанной бисульфитом ДНК (пунктирная линия), обнаруженной зондом гибридизации GSTpl-красным 650 (А) или GSTpl-красным 705 (В).
Фиг.14: Определение абсолютной чувствительности GSTpl-ПЦР на LightCycle. GSTpl-ПЦР проводили с блокирующим нуклеотидом ("положения ротора" 9, 10, 11, 12, 13, 14, 15, 16, 18) и без блокирующего олигонуклеотида ("положения ротора" 1, 2, 3, 4, 5, 6, 7, 8, 17). В качестве матричной ДНК применяли: метилированную обработанную бисульфитом ДНК: 7.5 нг ("положения ротора" 1, 9), 3.7 нг ("положения ротора" 2, 10), 0.75 нг ("положения ротора" 3, 11), 0.37 нг ("положения ротора" 4, 12), 0.075 нг Г положения ротора" 5, 13), 0.037 нг ('"положения ротора" 6, 14), 0.015 нг ('"положения ротора" 1, 15), 0.0075 нг(" положения ротора" 8, 16) и без ДНК ('"положения ротора" 17, 18).
Фиг.15: Определение относительной чувствительности амплификации метилированного GSTpl-гена. Амплификацию метилированного GSTpl-гена проводили на LightCycle. Обнаружение показано гибридизирукдцими зондами GSTpl-красным 650 (F2/F1, точка скрещивания) и GSTpl-красным 705 (F3/F1, точка скрещивания). GSTpl-ПЦР проводили блокирующим олигонуклеотидом ("положения ротора" 11, 12, 13, 14, 15, 17, 18, 19, 20) и без блокирующего олигонуклеотида ("положения ротора" 1, 2, 3, 4, 5, 7, 8, 9, 10). В качестве матричной ДНК применяли: 15 нг метилированной обработанной бисульфитом ДНК ("положения ротора 1, 11) и 15 нг неметилированной обработанной бисульфитом ДНК ("положения ротора" 10, 20), метилированную и неметилированную обработанную бисульфитом ДНК в соотношении смеси 1:2 ("положения ротора" 2, 12), 1:10 ("положения ротора" 3, 13), 1:20 ("положения ротора" 4, 14), 1:100 ("положения ротора" 5, 15), 1:500 ("положения ротора" 7, 17), 1:1000 ("положения ротора" 18), и без ДНК ("положения ротора" 10, 20).
Фиг.16: Определение относительной чувствительности амплификации мэтилированного GSTpl-гена. Амплификацию метилированного GSTpl-гена проводили на циклическом реакторе TaqMan.



| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ АНАЛИЗА НАРУШЕНИЙ, СВЯЗАННЫХ С РАКОМ ЯИЧНИКОВ | 2008 |
|
RU2511408C2 |
| ОБЪЕДИНЕННАЯ ОЧИСТКА И ИЗМЕРЕНИЕ МЕТИЛИРОВАНИЯ ДНК С СОВМЕСТНЫМ ИЗМЕРЕНИЕМ МУТАЦИЙ И/ИЛИ УРОВНЕЙ ЭКСПРЕССИИ мРНК В АВТОМАТИЧЕСКОМ РЕАКЦИОННОМ КАРТРИДЖЕ | 2017 |
|
RU2780660C2 |
| СПОСОБЫ АМПЛИФИКАЦИИ ДНК ДЛЯ СОХРАНЕНИЯ СТАТУСА МЕТИЛИРОВАНИЯ | 2018 |
|
RU2754038C2 |
| СПОСОБ АНАЛИЗА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ МОЛОЧНОЙ ЖЕЛЕЗЫ | 2008 |
|
RU2492243C2 |
| НАБОР ДЛЯ СКРИНИНГА НА РАК ПРЯМОЙ И ОБОДОЧНОЙ КИШКИ И ПРОГРЕССИРУЮЩЕЙ АДЕНОМЫ И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2812023C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ Pu(5mC)GPy В ЗАДАННОМ ПОЛОЖЕНИИ ПРОТЯЖЕННОЙ ДНК | 2013 |
|
RU2525710C1 |
| ДИАГНОСТИЧЕСКИЕ МАРКЕРЫ | 2012 |
|
RU2614254C2 |
| Набор синтетических олигонуклеотидов для проведения метилчувствительной амплификации ДНК | 2017 |
|
RU2662664C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МЕТИЛИРОВАНИЯ САЙТОВ PuCGPy РЕГУЛЯТОРНЫХ ОБЛАСТЕЙ ГЕНОВ-ОНКОМАРКЕРОВ КОЛОРЕКТАЛЬНОГО РАКА МЕТОДОМ GLAD-ПЦР-АНАЛИЗА И ОЛИГОНУКЛЕОТИДНЫЕ ПРАЙМЕРЫ И ФЛУОРЕСЦЕНТНО-МЕЧЕНЫЕ ЗОНДЫ ДЛЯ ОСУЩЕСТВЛЕНИЯ УКАЗАННОГО СПОСОБА | 2015 |
|
RU2596404C1 |
| ПРИМЕНЕНИЕ ВЫСОКОТЕРМОСТОЙКОГО CAS-БЕЛКА И СПОСОБ И НАБОР РЕАКТИВОВ ДЛЯ ОБНАРУЖЕНИЯ ЦЕЛЕВОЙ МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ | 2019 |
|
RU2800778C2 |
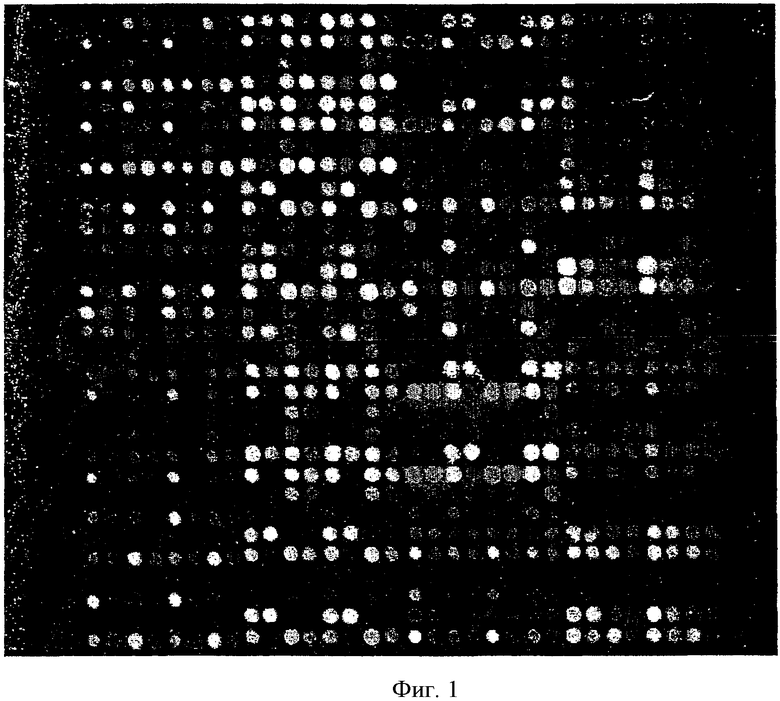
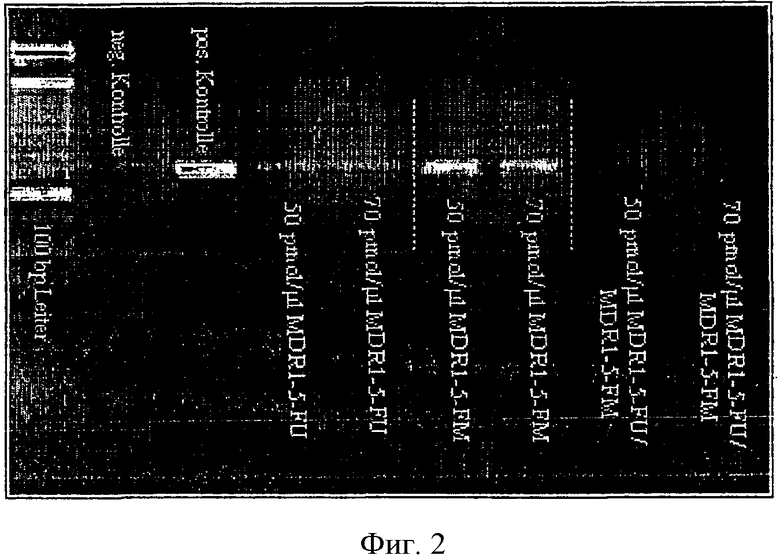
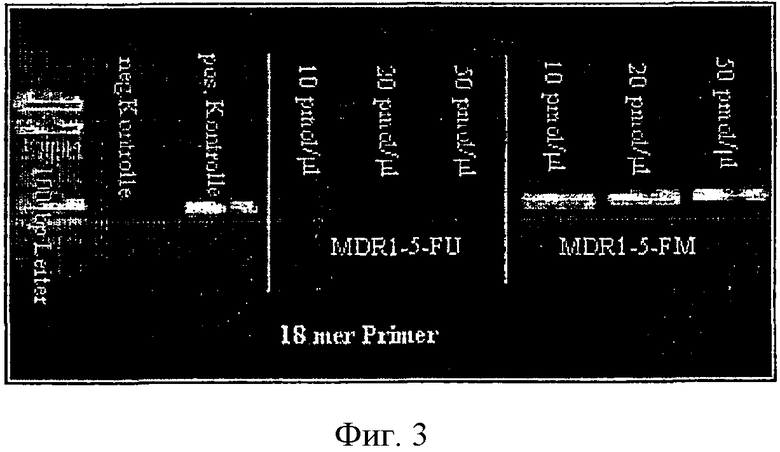
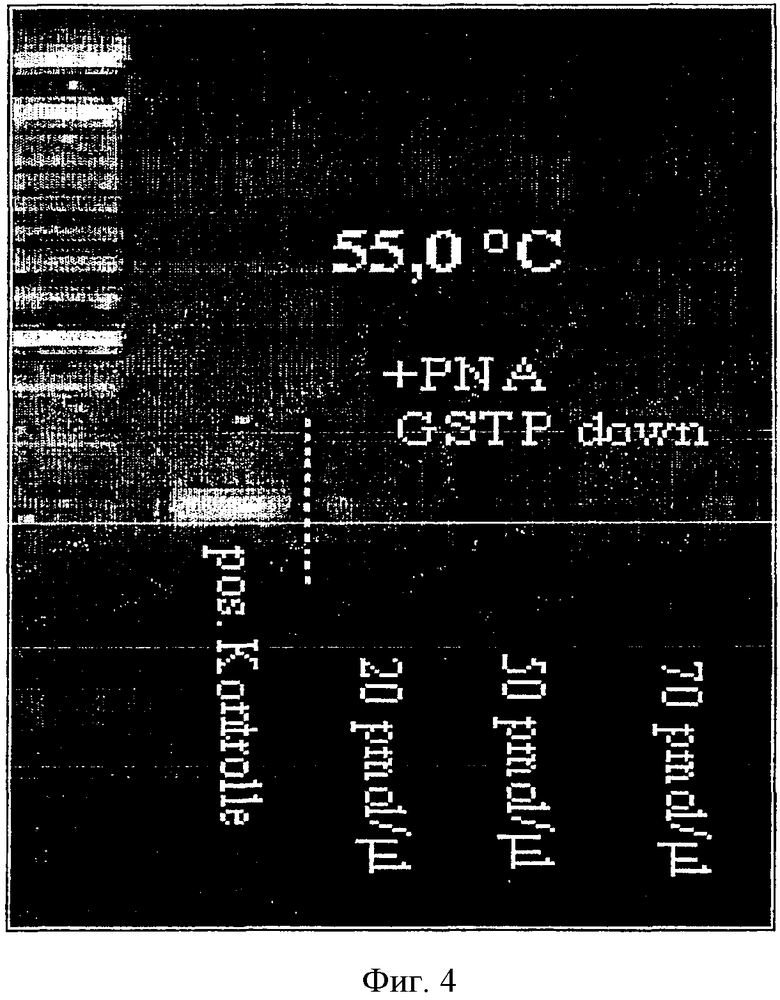
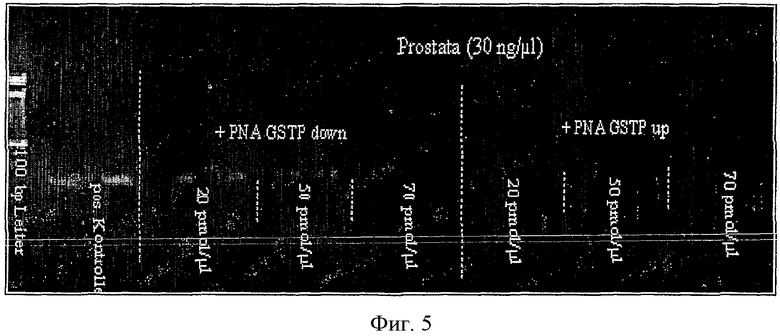


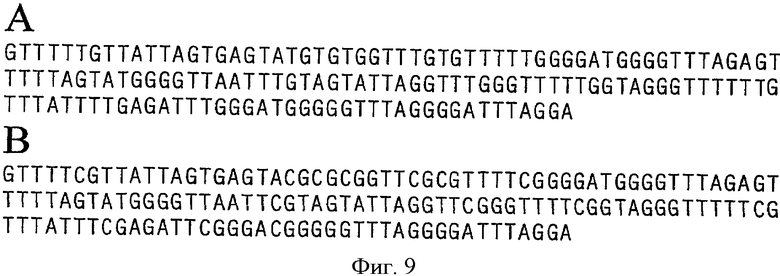
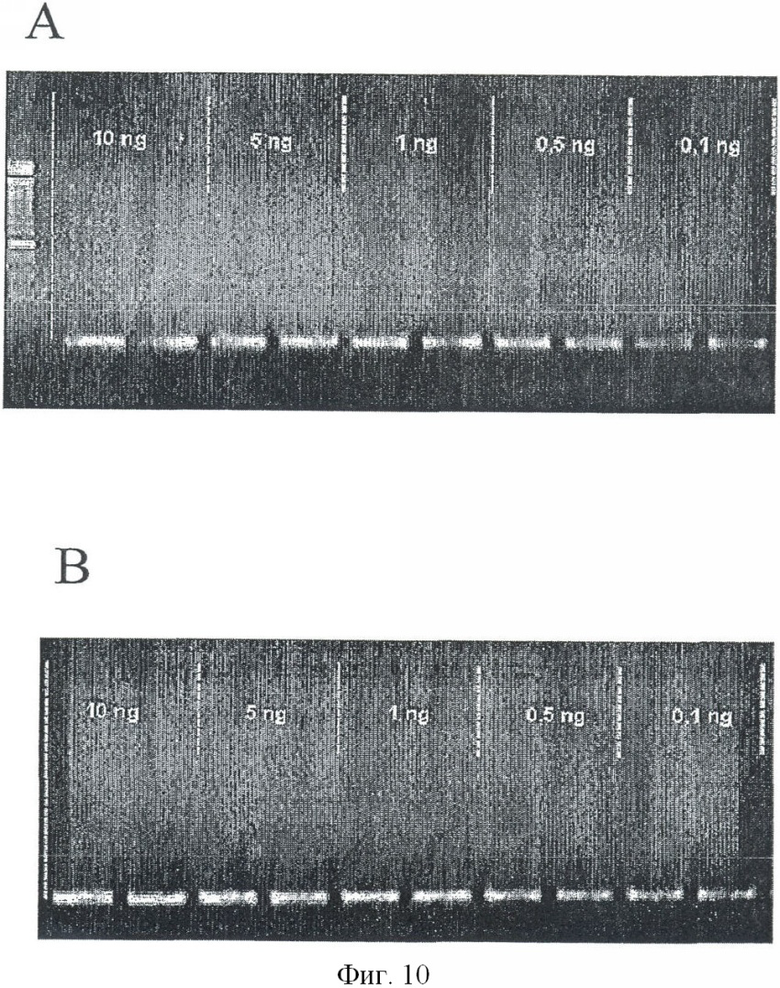
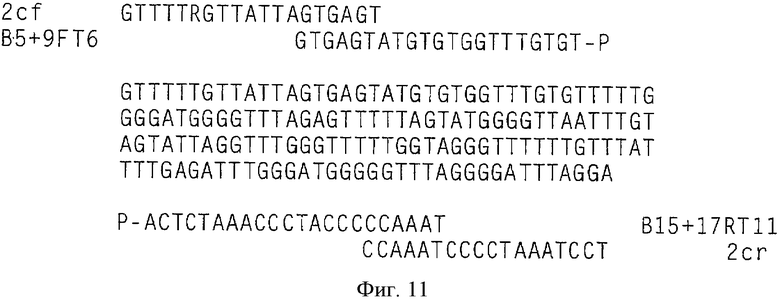
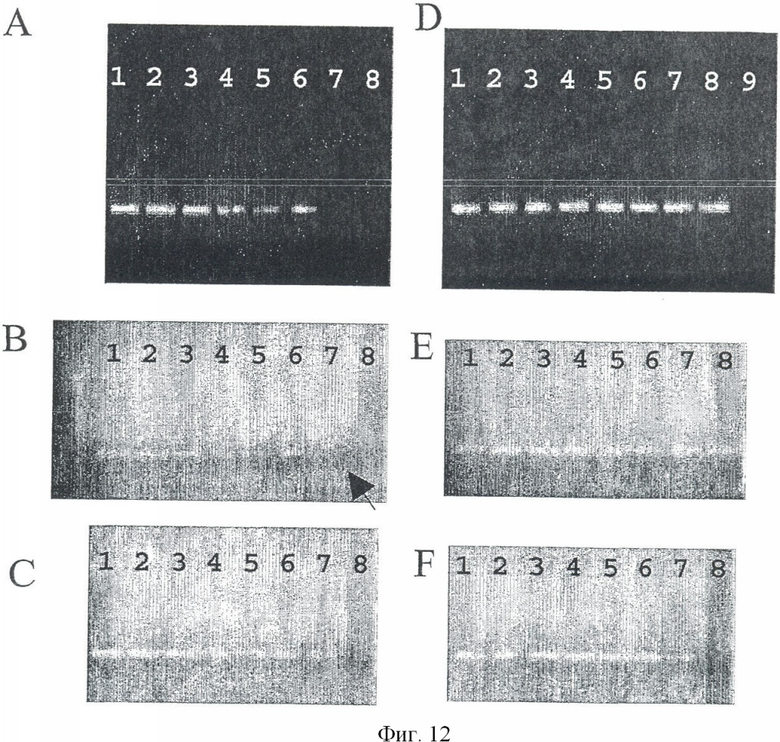
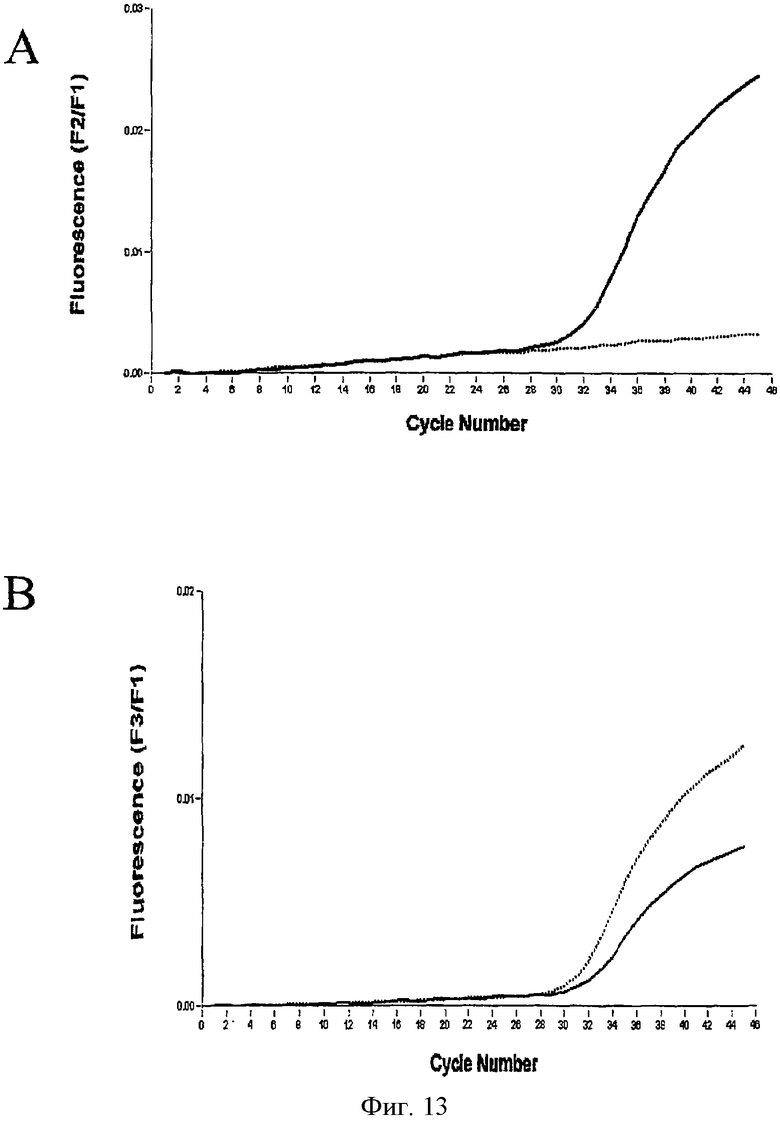
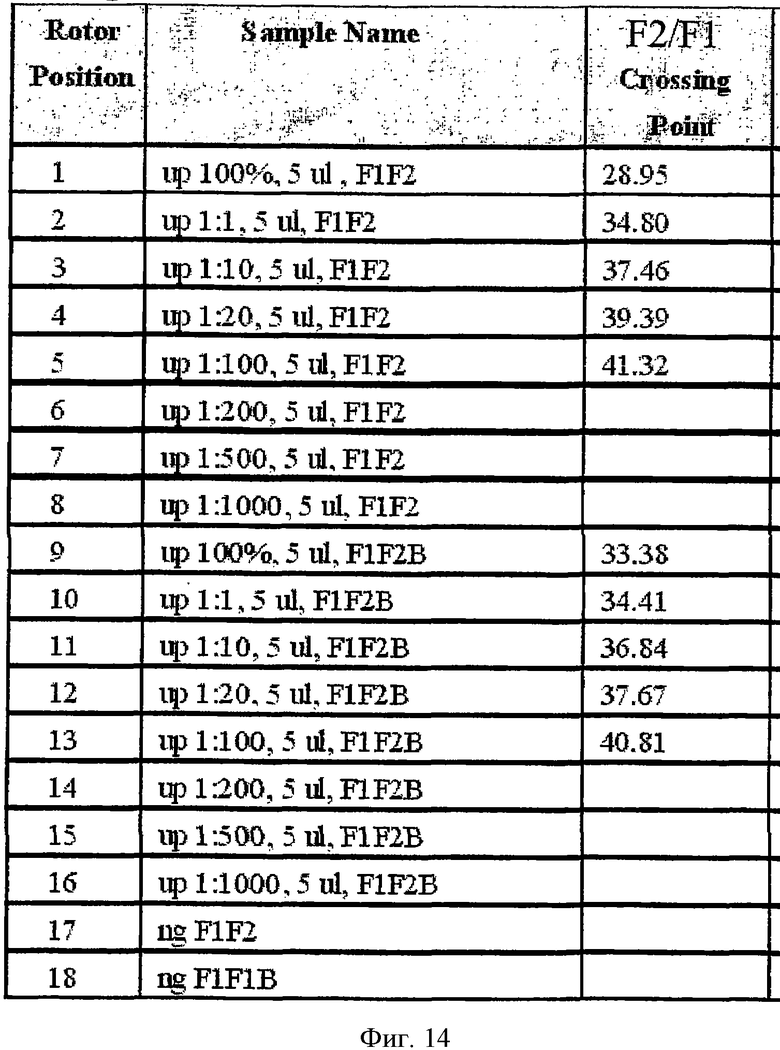
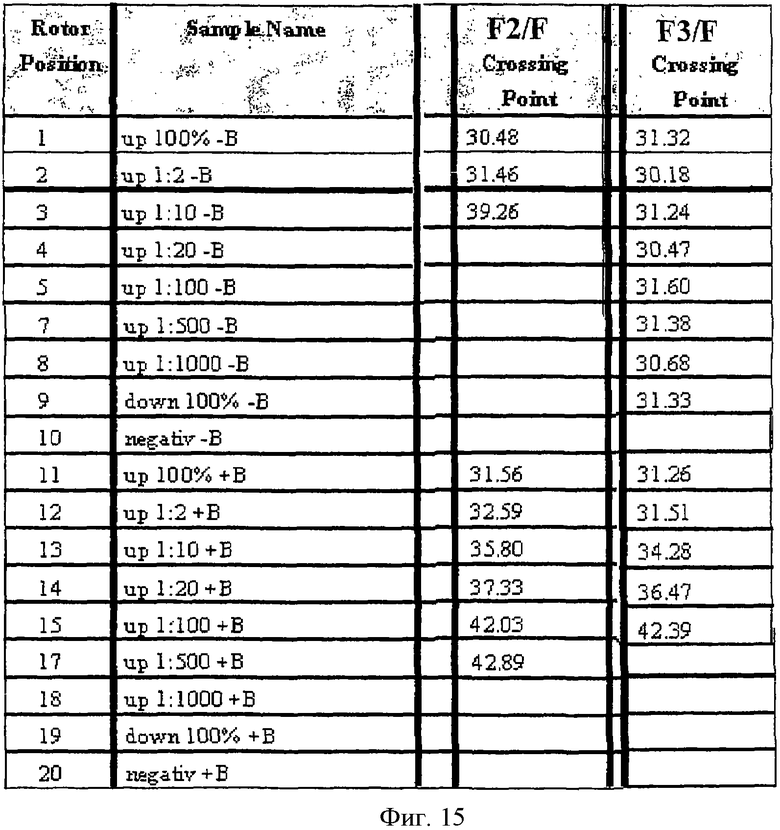
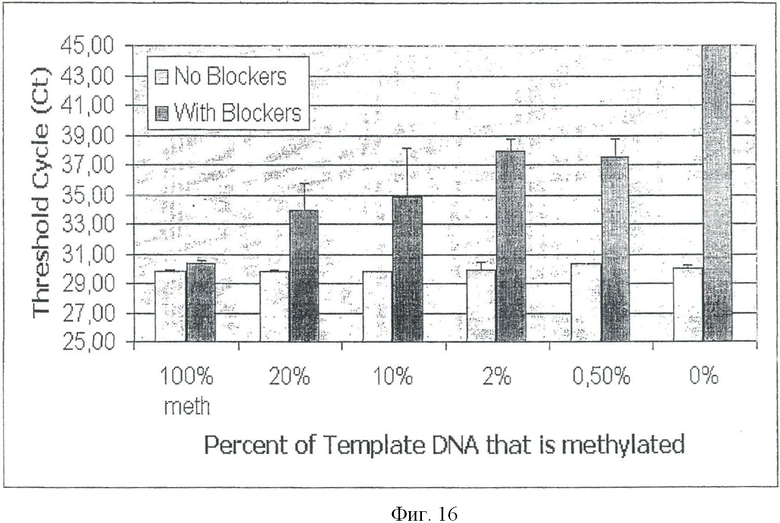
Изобретение относится к области молекулярной биологии и может быть использовано в диагностических методах различного назначения. Предложенный метод предусматривает а) обработку образца геномной ДНК, содержащего наряду с анализируемой ДНК фоновую ДНК, бисульфитом; б) ПЦР-амплификацию обработанного образца с использованием, по меньшей мере, одной пары праймеров и, по меньшей мере, одного дополнительного олигонуклеотида или ПНК-олигомера, который обладает способностью предпочтительно связываться с 5'-CG-3'-, или 5'-TG-3'-, или 5'-СА-3'- сайтом в фоновой ДНК, препятствуя ее амплификации, и в) анализ амплифицированных продуктов, на основании которого делается вывод о состоянии метилирования исследуемой ДНК. Метод по изобретению предлагается использовать для диагностики и/или прогнозирования предрасположенности к некоторым заболеваниям, а также в анализах, предусматривающих определение различных типов клеток и/или различных стадий их дифференцировки. 4 н. и 35 з.п.ф-лы, 16 ил.
а) образец геномной ДНК, включающий исследуемую ДНК и фоновую ДНК, подвергают обработке бисульфитом таким образом, что все неметилированные основания цитозина преобразуются в урацил, а основания 5-метилцитозина остаются неизмененными;
б) обработанный образец ДНК амплифицируют с применением, по меньшей мере, двух олигонуклеотидов-праймеров, а также, по меньшей мере, одного другого олигонуклеотида или ПНК-олигомера, который связывается с 5'-CG-3'-динуклеотидом, или 5'-TG-3'-динуклеотидом, или 5'-СА-3'-динуклеотидом, причем другой олигонуклеотид или ПНК-олигомер связывается предпочтительно в фоновой ДНК и препятствует ее амплификации, и полимеразы;
в) амплифицированные продукты анализируют и делают вывод о состоянии метилирования исследуемой ДНК на основании наличия и/или результатов его анализа.
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| ГВОЗДЕВ В.А | |||
| СОРОС образ | |||
| ж., 1999, №10, с.11-17 | |||
| ORUM ET AL., Curr | |||
| Issues Mol | |||
| Biol., 2000, 2(1), 27-30 | |||
| PARSONS В, HEFLICH R., Mutagenesis, 1998, 13(6), 581-588 | |||
| YU D | |||
| ET AL., Biotechniques, 1997, 714-720. | |||

