Настоящее изобретение относится к области медицины, в частности к созданию новых профилактических или терапевтических вакцинных препаратов, что позволяет повысить качество иммунного ответа против вакцинных антигенов заболеваний различной этиологии.
Neisseria meningitidis, грамотрицательный диплококк, единственным известным хозяином которого является организм человека, представляет собой агент-возбудитель менингококкового менингита. Как правило, в нормальной популяции данный вид бактерий находят среди бессимптомных носителей, являющихся в данной нише наиболее обычным источником для их микробиологического выделения.
Во всем мире дети в возрасте до двух лет составляют наиболее чувствительную группу, заболевающую менингококковым менингитом, хотя представители молодежи и люди старшего возраста также могут быть поражены этим заболеванием.
Для большинства пациентов, не подвергающихся лечению, менингококковое заболевание заканчивается смертельным исходом, и вакцинация могла бы предотвратить такой поворот событий, прерывая течение неблагоприятных явлений еще на стадии бактериальной колонизации.
Разработано несколько подходов с целью получения вакцины, способной полностью удовлетворить биологические требования по защите популяций человека против данного заболевания. И в этой связи принимают в расчет антигены капсулы, поскольку их иммунологическая специфичность позволяет классифицировать данный микроорганизм на серогруппы. Установлено, что пять из этих серогрупп ответственны за большинство клинических случаев менингококкового заболевания, зарегистрированных во всем мире. Серогруппа А является главной причиной возникновения эпидемий среди населения стран южной Африки. Серогруппы В и С в большинстве случаев связаны с распространением данного заболевания в развитых странах. Для серогруппы Y и W135 обычно характерно большинство рецидивных случаев данного заболевания, и они преобладают в некоторых районах США с отчетливым увеличением их частоты встречаемости в последние несколько лет. В соответствии с этими данными очевидна причина использования, изучения и анализа капсульных полисахаридов в качестве кандидатов для приготовления вакцины. Тетравалентная вакцина на основе капсульных полисахаридов, придающая способность защищать от серогрупп A, C, Y и W135, была лицензирована в Соединенных Штатах. Выявленные после вакцинации антитела являются серогруппоспецифичными (Rosenstein N. et al. 2001. Meningococcal disease. N. Engl. J. Med., 344, 1378-1388).
Отличающаяся от остальных серогруппа В продолжает оставаться существенной причиной эндемического и эпидемического менингококкового заболевания, что связано главным образом с полным отсутствием эффективных вакцин против нее. Отмечено, что капсульный полисахарид В недостаточно иммуногенен, и, кроме того, теоретически существует риск получения вакцины на основе данного соединения, вызывающего иммунотолерантность и аутоиммунитет из-за его структурного сходства с олигосахаридными цепями, которые присутствуют в структурах нервной системы плода человека (Finne J. et al. 1987. An IgG monoclonal antibody to group B meningococci cross-reacts with developmentally regulated polysialic acid units of glycoproteins in neural and extraneural tissues. J. Immunol., 138: 4402-4407). Поэтому разработка вакцины против серогрупп В сосредоточена на использовании субкапсульных антигенов.
Белки наружной мембраны и везикулярные вакцины
Первые попытки по созданию вакцин на основе белков наружной мембраны, предпринятые в 70-е годы, были основаны на LPS-истощении препаратов белков наружной мембраны с помощью детергента (Frasch C.E. and Robbins J.D. 1978. Protection against group B meningococcal disease. III. Immunogenicity of serotype 2 vaccines and specificity of protection in a guinea pig model. J. Exp. Med. 147(3):629-44). Затем белки наружной мембраны, OMP, осаждали, с получением агрегатов, суспендированных в растворе хлорида натрия. Несмотря на многообещающие результаты в исследованиях на животных, такие вакцины оказались неудачными для индукции бактерицидных антител у взрослых или у детей (Zollinger W.D. et al. 1978. Safety and immunogenicity of a Neisseria meningitidis type 2 protein vaccine in animals and humans. J. Infect. Dis. 137(6):728-39), слабая эффективность таких вакцин большей частью была связана с утратой третичной структуры, что сопровождалось их преципитацией. Поэтому следующим логическим шагом было получение вакцины с белками, представленными в их нативной конформации в виде пузырьков наружной мембраны (Zollinger W.D. et al. 1979. Complex of meningococcal group B polysaccharide and type 2 outer membrane protein immunogenic in man. J. Clin. Invest. 63(5):836-48, Wang L.Y. and Frasch C.E. 1984. Development of a Neisseria meningitidis group B serotype 2b protein vaccine and evaluation in a mouse model. Infect. Immun. 46(2):408-14136).
Такие вакцины из везикул наружной мембраны оказались существенно более иммуногенными, чем OMP-агрегаты, и было также показано, что иммуногенность дополнительно усиливается благодаря адсорбции с адъювантом гидроокиси алюминия (Wang L.Y. and Frasch C.E. 1984. Neisseria meningitidis group B serotype 2b protein vaccine and evaluation in mouse model. Infect. Immun. 46(2):408-14136).
Осуществлен ряд эффективных экспериментов с применением вакцин из растворимых везикул наружной мембраны разного состава. Две наиболее полно изученные вакцины разработаны в 1980 гг. при вспышках заболевания на Кубе (Sierra G.V. et al. 1991. Vaccine against group B Neisseria meningitidis: protection trial and mass vaccination results in Cuba. NIPH Ann. Dis. 14(2):195-210) и в Норвегии (Bjune G. et al. 1991. Effect of outer membrane vesicle vaccine against group B meningococcal disease in Norway. Lancet. 338(8775):1093-6). OMV-вакцину, производимую на Кубе в Институте Finlay (торговая марка VA-MENGOC-BC), получают из штамма B:4P1.19,15 с полисахаридом серогруппы С и препаратом высокомолекулярных OMP и адсорбируют гидроокисью алюминия (Sierra G.V. et al. 1991. Vaccine against group B Neisseria meningitidis: protection trial and mass vaccination results in Cuba. NIPH Ann. Dis. 14(2):195-210). Указанная вакцина способствовала быстрому снижению эпидемии на Кубе (Rodriguez A.P. et al. The epidemiological impact of antimeningococcal B vaccination in Cuba. 1999. Mem. Inst. Oswaldo Cruz. 94(4):433-40).
Соответствующая вакцина, производимая в Национальном Институте Здравоохранения Норвегии (NIPH), первоначально также предназначалась для использования в период гиперэндемического заболевания, вызванного другим микроорганизмом из клона ET-5 (B:15:P1.7,16). Она также представляет собой моновалентную вакцину, получаемую из очищенных везикул наружной мембраны, адсорбированных на гидроокиси алюминия (Bjune G. et al. 1991. Effect of outer membrane vesicle vaccine against group B meningococcal disease in Norway. Lancet. 338(8775):1093-6).
Везикулярные вакцины наружной мембраны, по-видимому, эффективно представляют белки наружной мембраны по существу в нативной конформации, результатом чего является выработка функциональных бактерицидных антител по меньшей мере у подростков и взрослых. Уровень вызываемого антителогенеза свидетельствует также об увеличении опсонофагоцитоза менингококков. Точный состав указанных вакцин (а именно содержание OMP, содержание LPS, а также наличие или отсутствие адъюванта) оказывает существенное влияние на иммуногенность (Lehmann A.K. et al. 1991. Immunization against serogroup B meningococci. Opsonin response in vaccines as measured by chemiluminiscence. APMIS. 99(8):769-72, Gomez J.A. et al. 1998. Effect of adjuvants in the isotypes and bactericidal activity of antibodies against the transferring-binding proteins of Neisseria meningitidis. Vaccine. 16(17):1633-9, Steeghs L. et al. 1999. Immunogenicity of Outer Membrane Proteins in a Lipopolysaccharide-Deficient Mutant of Neisseria meningitidis: Influence of Adjuvants on the Immune Response. Infect. Immun. 67(10):4988-93).
Антигенный профиль изолятов заболевания также быстро изменяется, и вакцина, охватывающая лишь ограниченное число отобранных штаммов, по-видимому, становится неэффективной через несколько лет, пока состав вакцины не изменят в соответствии с местной эпидемиологической ситуацией.
В настоящее время OMV-вакцины используют чаще любой другой вакцины серогруппы В, и потенциально они применимы в случае вспышек заболевания, вызванных единственным PorA-типом.
Соответствующие иммуногены, которые порождают перекрестную реактивность между штаммами, все еще полностью не определены. Исследование поствакцинных сывороток из Института Finlay и вакцинных экспериментов в NIPH дают основание полагать, что антитела к PorA (P1, белок серологического подтипа класс 1) и OpcA (другой основной OMP, раннее известный как Opc) (Wedege E., et al. 1998. Immune Response against Major Outer Membrane Antigens of Neisseria meningitidis in Vaccinees and Controls Who Contracted Meningococcal Disease during the Norwegian Serogroup B Protection Trial. Infect. Immun. 66(7):3223-31), оба существенно важны для опосредования сывороточной бактерицидной активности (иммуногенным PorA mosl), причем у обоих этих антигенов обнаружена значительная изменчивость от штамма к штамму.
Особое положение PorA-белка и существенный уровень изменчивости данного белка, который подвергается непрерывной генетической изменчивости в период между вспышками и во время вспышек (Jelfs J., et al. 2000. Sequence Variation in the porA Gene of a Clone Neisseria meningitidis during Epidemic Spread. Clin. Dagn. Lab. Immunol. 7(3):390-5), в эпитопах, на которые направлена большая часть поствакцинной (а также в период после заболевания) бактерицидной активности, усиливает интерес к тому, что предлагаемая защита на основе OMV-вакцин по единственному штамму (моновалентные) может быть ограничена серологическим подтипом (то есть зависеть от PorA-типа).
Для преодоления данной возможной проблемы в Нидерландах в RIVM создали OMV-вакцину, которая содержит PorA-белки из шести разных широко распространенных патогенных изолятов (Van Der Ley P. and Poolman JT. 1992. Construction of a multivalent meningococcal vaccine strain based on the class 1 outer membrane protein. Infect. Immun. 60(8):3156-61, Claasen I., et al. 1996. Production, characterization and control of a Neisseria meningitidis hexavalent class 1 outer membrane protein containing vesicle vaccine. Vaccine. 14(10):1001-8). В этом случае вакцинные везикулы экстрагировали из двух вариантов достаточно хорошо охарактеризованного штамма H44/76, который был модифицирован с помощью генной инженерии для экспрессии трех отельных PorA-белков.
Поиск универсального антигена
Очевидно, что белки наружной мембраны (OMP) могут индуцировать функциональный иммунный ответ против вызывающих заболевание микроорганизмов серогруппы В, но что ни одна из созданных к настоящему времени указанных вакцин не обеспечивает универсальную защиту из-за высокой гетерогенности белков, экспонированных на поверхностных участках наружной мембраны. Такой умеренный перекрестно-реактивный иммунитет, индуцированный вакцинами из везикул наружной мембраны (OMV), побуждает искать антиген (или группы антигенов) наружной мембраны, который стимулирует образование функциональных антител и который присутствует во всех менингококковых штаммах. Такие антигены, если они представлены во всех штаммах, независимо от серогруппы, могли бы создать основу действительно универсальной менингококковой вакцины, которая исключит возможную проблему капсулярного запуска патогенных штаммов после полисахаридной вакцинации.
После того как стало очевидно, что изменчивость иммунодоминантного PorA-белка будет лимитировать его применение в качестве универсальной вакцины, был рассмотрен ряд других основных белков наружной мембраны в качестве возможной вакцины и некоторые из них взяты в дальнейшую разработку. Те из них, которые были рассмотрены, включают в себя белки класса 5 (OpsA), NspA, а также регулируемые железом белки (TbpA и B, FbpA, FetA). TbpB образует с TbpA часть комплекса, связывающего трансферрин. В современных исследованиях предполагается, что TbpA и играет более существенную функциональную роль в связывании железа и является более эффективным иммуногеном, чем TbpB (Pintor M. et al. 1998. Analysis of TbpA and TbpB functionality in defective mutants of Neisseria meningitidis. J. Med. Microbiol. 47(9):757-60).
Высококонсервативный минорный белок наружной мембраны был обнаружен благодаря новой методике с применением объединенных белковых препаратов разных менингококковых штаммов для иммунизации мышей (Martin D. et al. 1997. Highly Conserved Neisseria meningitidis Surface Protein Confers Protection against Experimental Infection. J. Exp. Med. 185(7):1173-83). В-клетки этих мышей используют для получения гибридом, которые впоследствии подвергают скринингу на перекрестную реактивность против различных штаммов менингококков. Обнаружено, что одно из интенсивно перекрестно-реагирующих моноклональных антител связывается с белком наружной мембраны в 22 кДа, который обозначили NspA. Показано, что иммунизация рекомбинантным NspA-белком индуцирует у мышей перекрестную бактерицидную реакцию против штаммов с серогруппами А-С. Вакцинация также защищает мышей от летального менингококкового заражения (Martin D. et al. 1997. Highly Conserved Neisseria meningitidis Surface Protein Confers Protection against Experimental Infection. J. Exp. Med. 185(7):1173-83). Сравнение NspA-последовательностей у генетически отличающихся менингококковых штаммов свидетельствует о том, что данный белок является высококонсервативным (97% гомологии) (Cadieux N. et al. 1999. Bactericidal and Cross-Protective Activities of a Monoclonal Antibody Directed against Neisseria meningitidis NspA Outer Membrane Protein. Infect Immun. 67(9):4955-9). Присутствие NspA детектируют с помощью ИФА у 99,2% тестируемых штаммов по серогруппам А-С с применением анти-NspA моноклональных антител (Martin D. et al. 1997. Highly Conserved Neisseria meningitidis Surface Protein Confers Protection against Experimental Infection. J. Exp. Med. 185(7):1173-83). Оказалось, что данные антитела являются бактерицидными для многочисленных штаммов менингококков и в мышиной модели способны снижать менингококковую бактериемию (Cadieux N. et al. 1999. Bactericidal and Cross-Protective Activities of a Monoclonal Antibody Directed against Neisseria meningitidis NspA Outer Membrane Protein. Infect Immun. 67(9):4955-9). Хотя эти данные и позволяют рассматривать NspA в качестве многообещающего вакцинного кандидата, способного обеспечить защиту в пределах серогрупп, поликлональная антисыворотка рекомбинантного NspA мышей не взаимодействует с поверхностью примерно 35% менингококковых штаммов с патогенной серогруппой В, несмотря на присутствие nspA-гена у этих организмов (Moe G.R. et al. 1999. Differences in Surface Expression of NspA among Neisseria meningitidis Group B Strains. Infect Immun. 67(11):5664-75).
Презентация антигена и вакцинная композиция
В более ранней работе предполагалось, что форма, в которой представлены данные антигены, является решающей. Соответствующие эпитопы мембраносвязывающих белков часто зависят от сохранения правильной третичной структуры, а она, в свою очередь, часто зависит от мембраносвязывающих гидрофобных доменов. Показано, что препараты белков наружной мембраны активируют иммунитет у человека лишь в том случае, если они представлены в везикулярной форме (Zollinger W.D. et al. 1979. Complex of meningococcal group B polysaccharide and type 2 outer membrane protein immunogenic in man. J. Clin. Invest. 63(5):836-48, Zollinger W.D. et al. 1978. Safety and immunogenicity of a Neisseria meningitidis type 2 protein vaccine in animals and humans. J. Infect. Dis. 137(6):728-39).
Вакцины, представленные единичным белком и используемые в данной области на протяжении десятилетий, демонстрируют, как правило, хорошую стабильность. Если же они представлены в форме везикул, что позволяет соответствующим антигенам оставаться связанными с мембраной, тогда, по-видимому, трудно гарантировать стабильность и воспроизводимость. Иммуногенность и реактогенность везикул наружной мембраны могут меняться вместе с изменением количества белка и LPS, удаляемых в процессе очистки. Вместе с тем, при производстве OMV-вакцины и получаемых в настоящее время вакцин накоплен опыт получения везикул, которые подвергаются тщательному контролю качества. Создание полностью синтетических липосомных везикул может позволить осуществлять дальнейшую оптимизацию и стандартизацию таких вакцин (Christodoulides M. et al. 1998. Immunization with recombinant class 1 outer-membrane protein from Nesseria meningitidis: influence of liposomes and adjuvants on antibody avidity, recognition of native protein and the induction of a bactericidal immune response against meningococci. Microbiology 144(Pt 11):3027-37). Иными словами, белки наружной мембраны представлены одновременно и в везикулах, и в виде экспрессируемых без примесей белков, при этом антительный ответ оказывается достаточно умеренным. До сих пор основные попытки сосредоточивались на внутримышечной инъекции менингококковой вакцины, приводя к системному образованию IgG. Однако при менингококковом заболевании, при котором инвазия хозяина осуществляется через назальный эпителий, существенной может быть также и выработка секреторного IgA.
Геномная последовательность N. meningitidis
В течение 2000 г. были выяснены и опубликованы геномные последовательности MC58 (серогруппа В менингококка) (Tettelin H. et al. 2000. Complete Genome Sequence of Neisseria meningitidis Serogroup B Strain MC58. Science 287 (5459):1809-15172) и Z2491 (серогруппа А штамма) (Parkhill J. et al. 2000. Complete DNA sequence of a serogroup A strain of Neisseria meningitidis Z2491. Nature 404 (6777):502-6173). Доступность аннотированных генных последовательностей должна ощутимо повлиять на изучение менингококковой вакцины. Пока осуществлялось секвенирование генома MC58, Pizza et al. приступили к идентификации предполагаемых открытых рамок считывания, которые кодируют мембраносвязанные, поверхностно экспонированные или экспортируемые белки. Они идентифицировали 570 таких ОРС, амплифицировали их с помощью полимеразной цепной реакции и клонировали их в Escherichia coli для возможной экспрессии указанных кодируемых белков либо в виде белков с гистидиновой меткой, либо в виде слитых с глутатион-S-трансферазой белков (Pizza M. et al. 2000. Identification of Vaccine Candidates Against Serogroup B Meningococcus by Whole-Genome Sequencing. Science 287 (5459):1816-20). Из отобранных ОРС 350 (61%) были успешно экспрессированы, а те из них, которые не экспрессировались, часто содержали более одного гидрофобного трансмембранного домена (возможно, за исключением нескольких белков, связанных с наружной мембраной). Рекомбинантные белки очищали и использовали для вакцинации мышей. Впоследствии у полученных иммунных сывороток определяли способность к связыванию с поверхностью многочисленных менингококковых штаммов с помощью иммуноферментного анализа (ИФА) и проточной цитометрии, а для определения бактерицидной активности в отношении двух штаммов применяли бактерицидный анализ сыворотки. В конечном итоге на основании позитивной реакции во всех трех видах анализа для дальнейшего изучения были отобраны семь белков. При исследование состава вакцин с применением ряда указанных белков в сочетании с адъювантами у мышей была продемонстрирована существенная бактерицидная защита против гомологичного менингококкового штамма (MC58), но ни один из этих белков не индуцировала таких высоких SBA-титров, как везикулярная вакцина наружной мембраны MC58 (Giuliani M.M. et al. 2000. Proceedings 12th IPNC. P.22). С другой стороны, есть определенные основания полагать, что сочетания этих белков могут создавать у мышей более высокую иммуногенность, чем одиночные белки (Santini L. et al. 2000. Proceedings 12th IPNC. P.25). Многочисленные открытые рамки считывания, которые были исключены в период осуществления данной работы, предположительно из-за неудачной экспрессии белка или же вследствие модификации их иммунологических характеристик, могут также обладать вакцинным потенциалом и требуют дальнейшего изучения.
Вакцинные компоненты можно отобрать более эффективно в том случае, если понять и воспользоваться вкладом отдельных антигенов в патогенез N. meningitidis. Сами по себе отдельные антигены могут оказаться эффективными вакцинными кандидатами или - альтернативно - соответствующие ослабленные мутанты могли бы рассматриваться в качестве составляющих вакцин.
В этом отношении применение вакцинных кандидатов с высокой степенью консерватизма последовательности из числа отдельных патогенных микроорганизмов могло бы явиться решением проблемы с многочисленными заболеваниями, причиной которых они могли быть в случае, если бы эти кандидаты индуцировали подходящий ответ на уровне иммунной системы.
Технический замысел, который преследует настоящее изобретение, заключается в создании вакцинных композиций по увеличению и/или расширению индуцируемого иммунного ответа против разных патогенов или против обширного разнообразия вариантов индивидуальных патогенов, будь то патогены злокачественных опухолей, бактерий, вирусов или любого иного происхождения.
Описание изобретения
В настоящем изобретении впервые сообщается об использовании NMB1125-белка в качестве компонента вакцинной композиции терапевтического или профилактического действия против определенного менингококкового заболевания или против любой инфекции, вызванной представителем из рода Neisseria.
Новый отличительный признак настоящего изобретения заключается в использовании, ранее не сообщаемом, белка NMB1125 в композициях с новыми свойствами, способных индуцировать системный иммунный ответ и иммунный ответ слизистой для защиты широкого спектра действия благодаря консервативному характеру данного белка в различных изолятах Neisseria meningitidis и Neisseria gonorrhoeae.
Краткое описание чертежей
Фигура 1. Клонирующий вектор pM100, используемый для клонирования и экспрессии белка NMB1125. pTrip, триптофановый промотор; N-term P64k, N-концевой фрагмент P64k; T4 Terminator, Терминатор транскрипции фага T4.
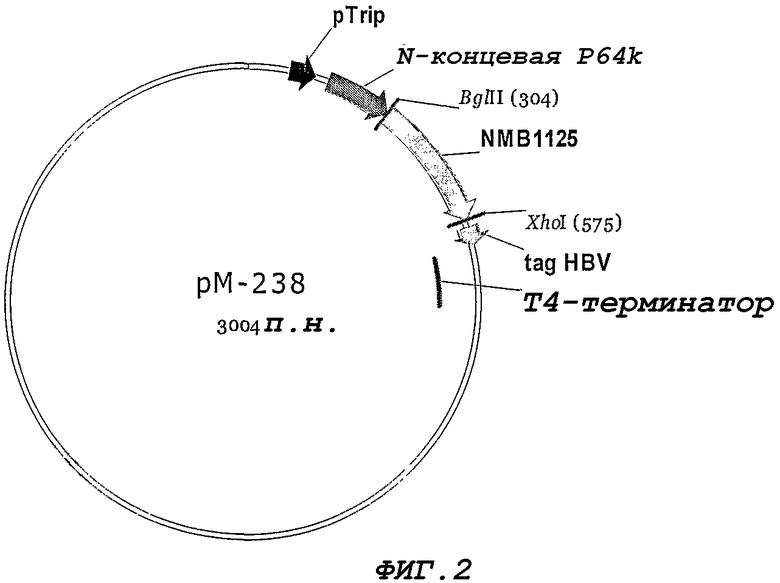
Фигура 2. Окончательная конструкция нуклеотидной последовательности гена NMB1125 в векторе pM100.
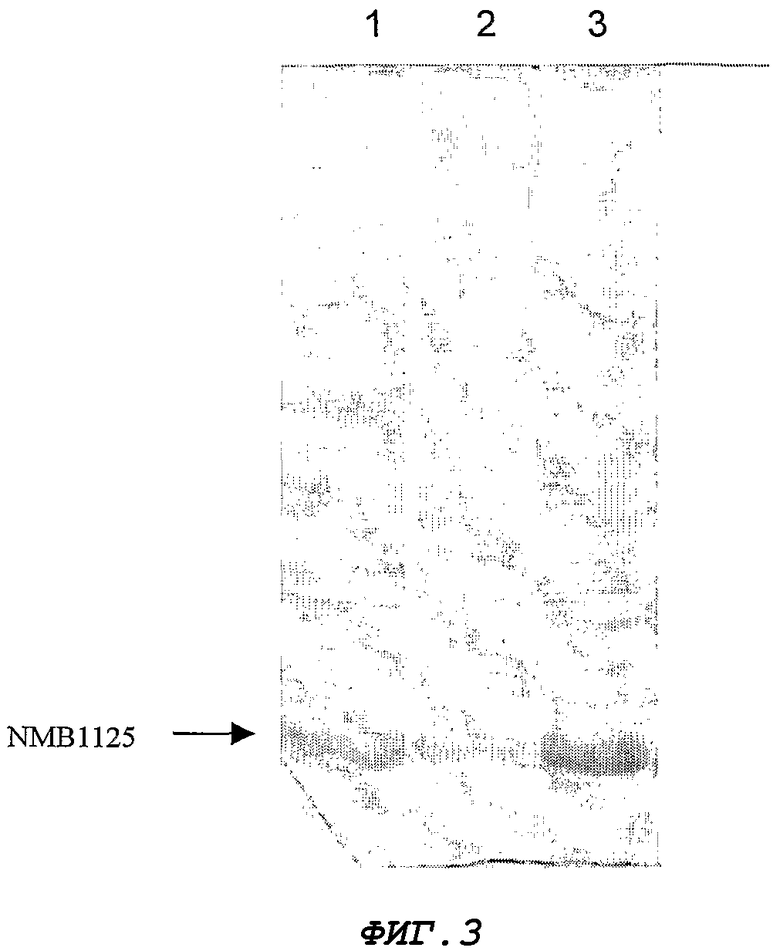
Фигура 3. ДСН-ПААГ-анализ фракций, полученных при разрушении клеток. Дорожка 1, все клетки; Дорожка 2, осадок клеток; Дорожка 3, супернатант.
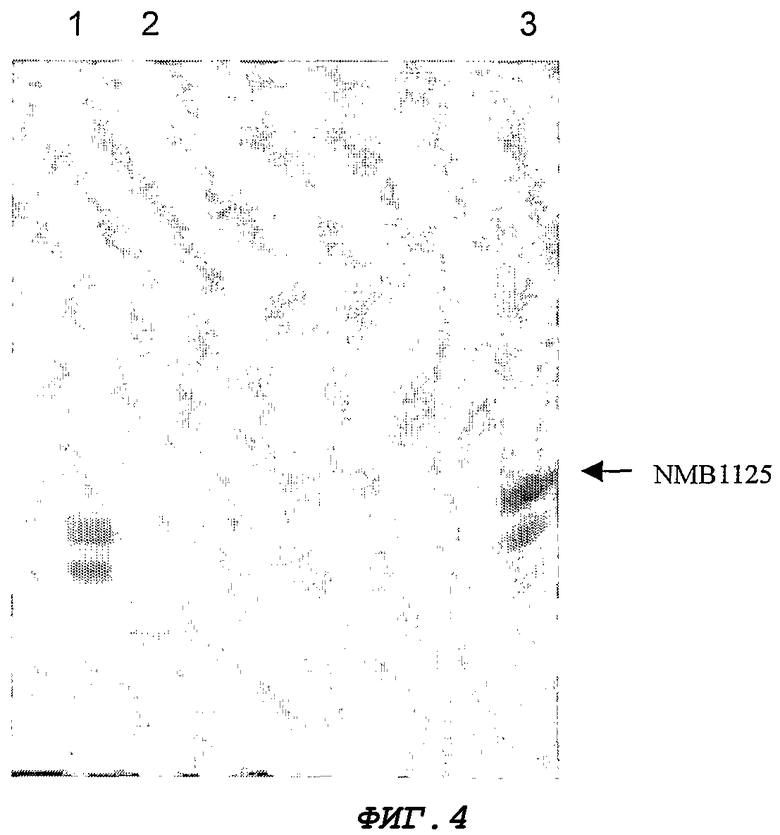
Фигура 4. ДСН-ПААГ-анализ NMB1125 в процессе его очистки из супернатанта разрушенных клеток. Дорожка 1, суммарный белок; Дорожка 2, примесной белок с низкой молекулярной массой, обнаруживаемый в разных хроматографических фракциях. Дорожка 3, образец до применения.
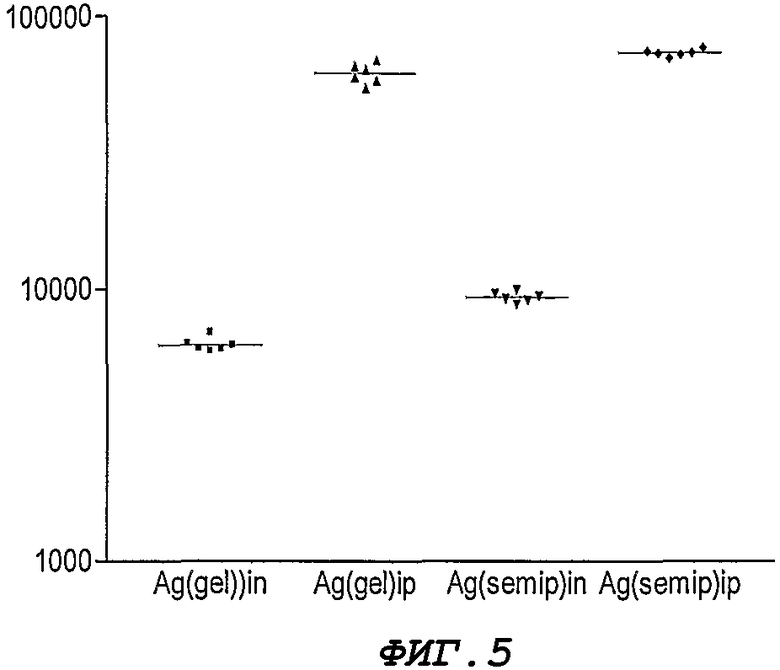
Фигура 5. Уровни антител (IgG) против рекомбинантного белка NMB1125, полученные после иммунизации мышей интраназальным или внутриперитонеальным путем. Представлены результаты ИФА, выраженные обратно пропорционально максимальному разведению, которые дублируют показатели сывороток, полученных после первичной иммунизации.
Фигура 6. Вестерн-блоттинг NMB1125-белка, представленного в OMV N. meningitidis, с применением сыворотки от мышей, иммунизированных указанным рекомбинантным белком. Изображение иммуноидентифицированного NMB1125 выделено графически.
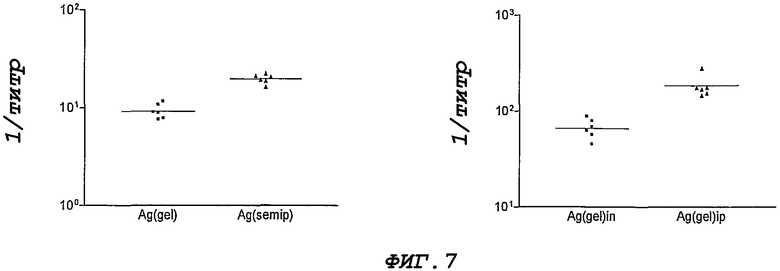
Фигура 7. IgA-антительный ответ против рекомбинантного белка NMB1125 на уровне слизистой у мышей, иммунизированных интраназальным путем. Результаты представлены в виде значений, обратных максимальному разведению, при котором удваиваются показатели преиммунных сывороток. (А) IgA-антительный ответ в слюне. (В) IgA-антительный ответ в смывах легких.
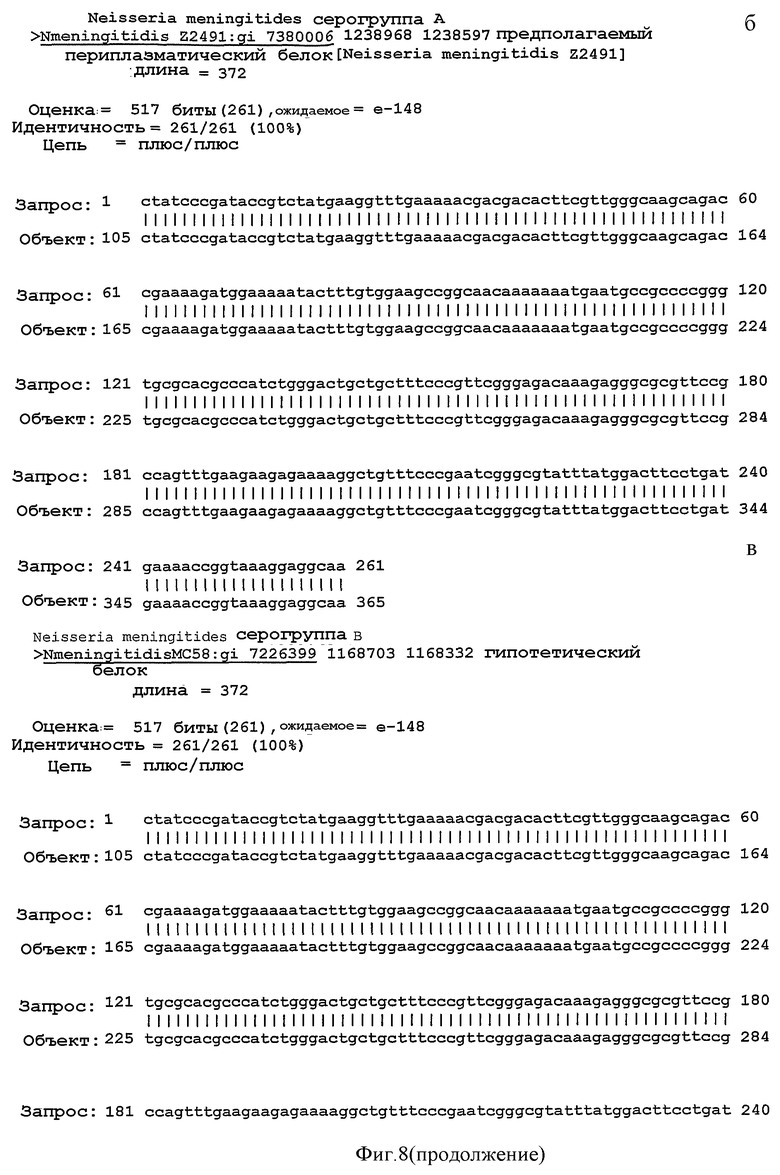
Фигура 8. Результаты поиска гомологии между NMB1125-белком (query, запрос) и аннотированными последовательностями в геномах разных серогрупп Neisseria meningitidis ("Sbjct", объект) с помощью BLAST.
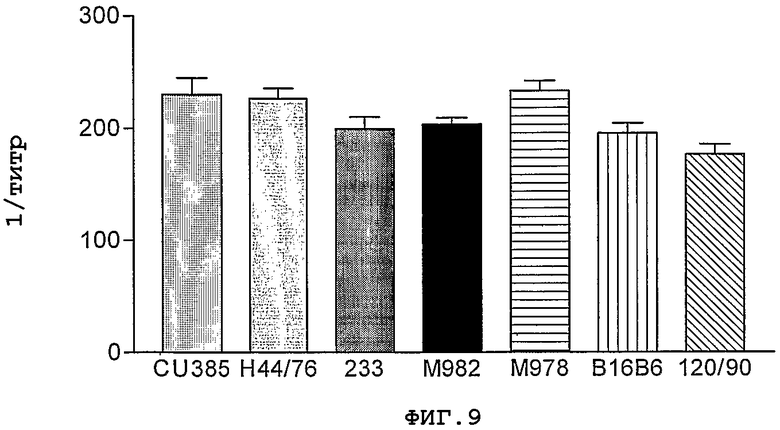
Фигура 9. Распознавание NMB1125-белка в разных штаммах Neisseria memingitidis с помощью сывороток, полученных против указанного рекомбинантного антигена. На данном графике представлены только результаты, полученные при использовании наполовину очищенного белка внутрибрюшинным путем, однако аналогичный характер изменений наблюдается и в остальных случаях. Результаты представлены в виде величины, обратной максимальному разведению, при котором показатели преиммунных сывороток удваиваются.
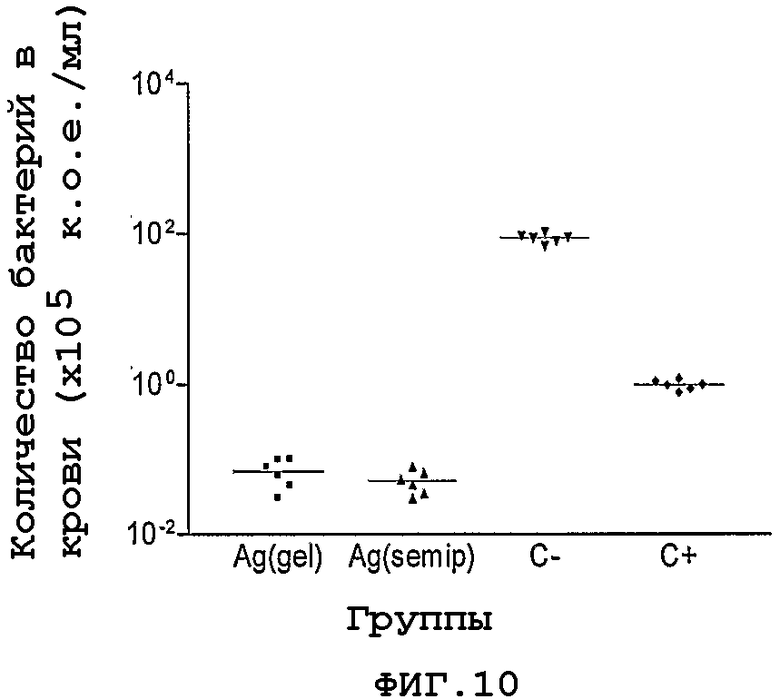
Фигура 10. Сравнение сывороток, полученных двумя способами в результате иммунизации указанным белком, вводимых с помощью интраназального способа, в экспериментах с пассивной защитой от менингококковой инфекции на модели новорожденных крыс.
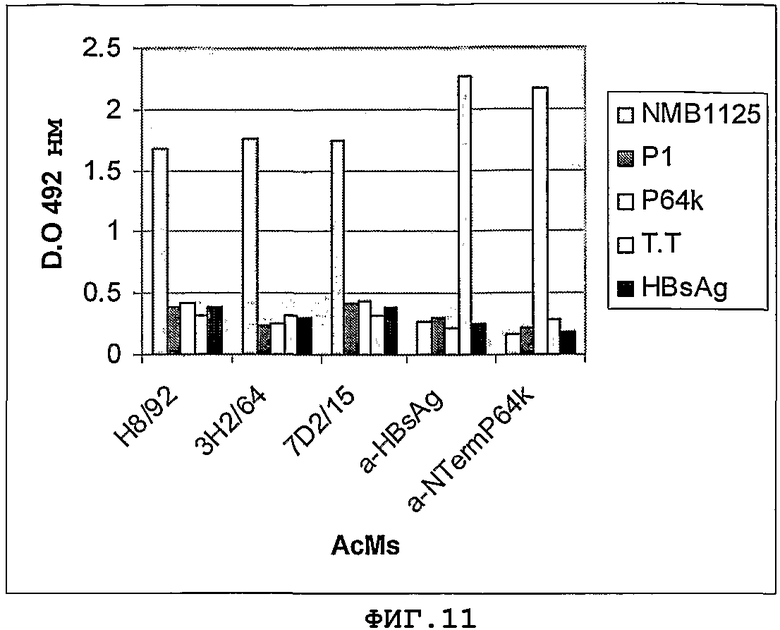
Фигура 11: Распознавание NMB1125-белка и панель неродственных антигенов для получения mAb (mAb H8/92, 3H2764 и 7D2/15). P1-белок класса 1 штамма Neisseria meningiditis B:4:P1.15; P64k, E3-субъединица пируватдегидрогеназы Neisseria meningitidis; T.T, столбнячный анатоксин; HBsAg, поверхностный антиген гепатита В.
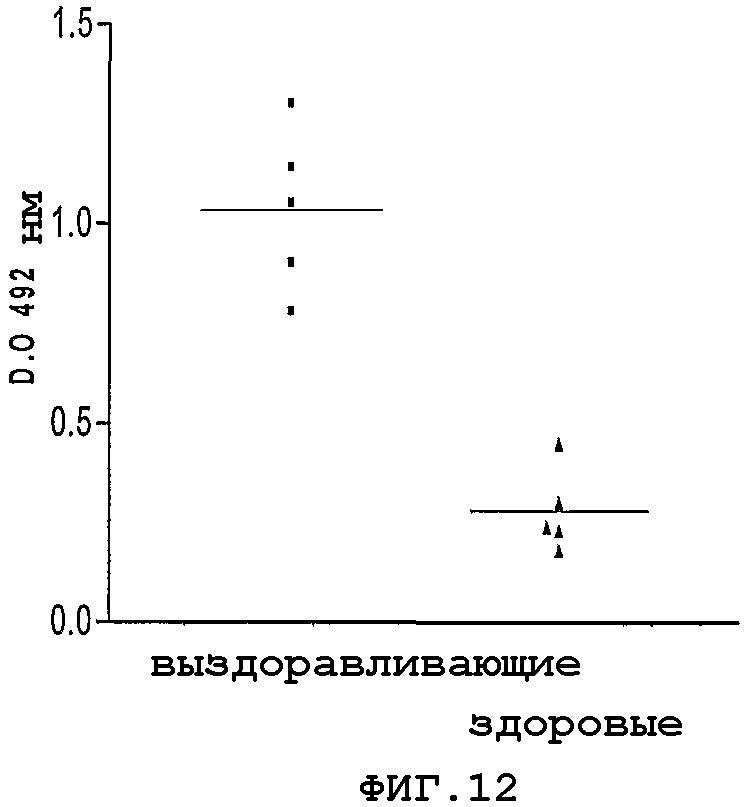
Фигура 12. Распознавание NMB1125-белка с помощью человеческих конвалесцентных сывороток от выживших пациентов, перенесших менингококковое заболевание. В качестве негативного контроля используют сыворотки здоровых доноров. Представлены результаты поглощения при 492 нм в пробах на ИФА.
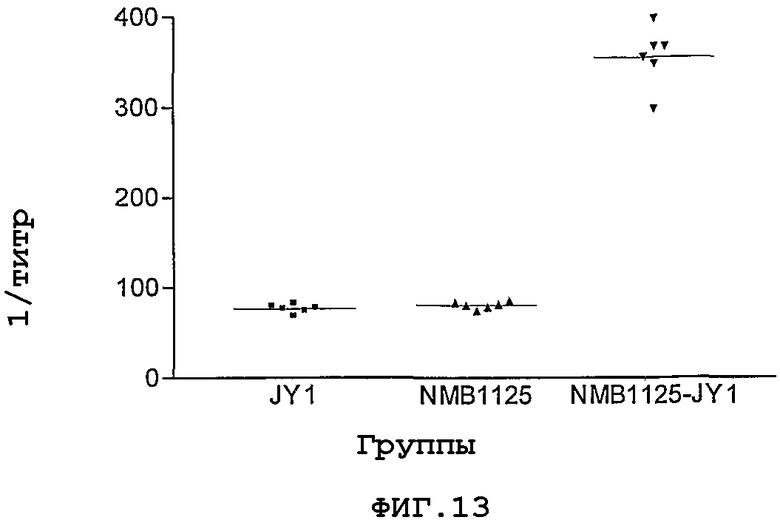
Фигура 13. JY1-противопептидные титры сывороток животных, иммунизированных свободным пептидом (JY1) или рекомбинантным белком (NMB1125), или их конъюгатом JY1-NMB1125.
Фигура 14. Иммуноблоттинг цельноклеточных антигенов из различных штаммов Neisseria.
Примеры
Настоящее изобретение описано здесь при помощи примеров, которые, несмотря на представленные в них сведения о сущности данного изобретения, сами по себе никоим образом не ограничивают объем указанного изобретения.
Пример 1: Детекция NMB1125-белка серогруппы B Neisseria meningitidis в препаратах везикул наружной мембраны
С целью изучения белков серогруппы В Neisseria meningitidis (штамм B:4:P1.19,15), которые представлены в везикулах наружной мембраны, осуществляют двумерный электрофорез в соответствии с описанным раннее методом (Görg A., et al. 1985. Electrophoresis 6:599-604). Затем осуществляют ферментативное расщепление извлекаемых из геля белков с помощью трипсина (Promega, Madison, WI, США). Полученные после расщепления пептиды экстрагируют в раствор с применением микроколонок (ZipTips, Millipore, MA, США). Для масс-спектрометрического анализа пептиды элюируют из микроколонок с помощью 60% ацетонитрила, 1% муравьиной кислоты, с последующим немедленным внесением элюатов в нанонаконечники (Protana, Дания).
Измерения осуществляют в гибридном масс-спектрометре с квадруполем и временем пролета (QTof-2TM, Manchester, Inited Kingdom), снабженном источником ионизации (nanoESI). Масс-спектрометрические данные получают в w/z-диапазоне 400-2000 за 0,98 секунд с промежутком в 0,02 секунды между сканированием. Получение данных и обработку данных осуществляют с применением программы MassLynx (version 3.5, Micromass).
Идентификацию белка по данным его масс-спектра осуществляют с применением программа ProFound (Zhang W. and Chait B.T. 2000. ProFound: an expert system for protein identification using mass spectrometric peptide mapping information. Anal. Chem. 72:2482-2489. http://prowl.rockefeller.edu/cgi-bin/ProFound). Поиск осуществляют по генам и по производимым белковым последовательностям, содержащимся в базе данных SwissProt (http://www.ebi.ac.uk/swissprot/) и NCBI (http://www.ncbi.nlm.nih.gov/), рассматривая окисление метионинов, дезамидирование и карбоксиамидометилирование цистеинов в качестве возможных модификаций.
Идентификацию белков по их масс-спектрам осуществляют с помощью программы MASCOT (Perkins D.N. et al. 1999. Probability-based protein identification by searching sequence database using mass spectrometry data. Electrophoresis 20:3551-3567. http://www.matrixscience.com/). Критерии поиска включают в себя модификации цистеина, а также окисление и деамидирование.
Исходя из анализа результатов, получаемых при идентификации белков, присутствующих в препаратах везикул наружной мембраны, выбирают белок NM1125, рассматривая его в качестве возможного вакцинного кандидата, у которого один из пептидов идентифицируют с помощью масс-спектрометрии.
Пример 2: Анализ гомологии белка NMB1125 с продуктами описываемого гена из доступных баз данных
Для анализа гомологии белка NM1125 с другими генными продуктами осуществляют поиск гомологии c последовательностями из базы данных NCBI с применением программы BLAST (Altschul S.F. et al. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410, http://www.ncbi.nlm.nih.gov/BLAST/). Результаты, полученные в ходе данного исследования, регистрируют в качестве гомологов, и, помимо белков, представленных в опубликованных геномах нейссерий, находят несколько генных продуктов, которые признают в качестве гипотетических белков других организмов, таких как виды Ralstonia, Yersinia и Pseudomonas.
Высокая степень консерватизма данных белков в данных геномах дает основание причислить их к ортологичной группе с консервативным доменом, представленным в базе данных NCBI [(gnl|CDD|13507, COG4259, неохарактеризованный консервативный бактериальный белок с неизвестной функцией], что указывает на существование между ними возможной филогенетической связи и общего предка.
Анализ геномного окружения гена, который кодирует NM1125, осуществляют с применением базы данных MBGD (Uchiyama, I. 2003. MBGD: microbial genome database for comparative analysis. Nucleic Acids Res. 31, 58-62), а обнаруживаемая у раннее указанных микроорганизмов консервативная генетическая организация данных генов, в сочетании с предшествующими данными, побуждает заключить, что они являются эффективными гомологами в соответствующих своих геномах.
Пример 3. Клонирование и экспрессия гена NMB1125, кодирующего белок NMB1125 N. meningitidis в Escherichia coli
Для того, чтобы клонировать и экспрессировать ген NM1125, используют клонирующий вектор pM-100. Указанный вектор позволяет осуществлять клонирование с применением разных ферментов рестрикции и обеспечивать высокие уровни экспрессии гетерологичных белков в форме телец включения у E. coli.
Вектор pM-100 (Фигура 1) включает в себя следующие элементы: триптофановый промотор, генный сегмент, кодирующий 47 аминокислот стабилизирующей последовательности в 64 кДа Nt-фрагменте из штамма N. meningitidis B:4:P1.19,15, последовательность терминатора транскрипции из бактериофага Т4, а также, в качестве селективного маркера, генную последовательность, которая придает резистентность к ампициллину.
Из нуклеотидной последовательности, кодирующей белок NMB1125 (Пример 1), конструируют два праймера (7738 и 7739) для амплификации соответствующего сегмента данного гена, без последовательности, которая кодирует предсказанный сигнальный пептид из геномной ДНК штамма B:4:P1.19,15
BglII_
7738: 5' TTAGATCTCTATCCCGATACCGTCTATGAAGG '3
(SEQ ID NO: 1)
7739: 5' AAGCTCGAGTCGTTTGCCTCCTTTACC 3'
XhoI
(SEQ ID NO: 2)
Для предсказания сигнального пептида используют сервер SignalP World Wide Web (http://www.cbs.dtu.dk/services/SignalP-2.0). После ПЦР-амплификации гена NMB1125 (Randall K., et al. 1988. Science 42394:487-491) с применением праймеров 7738 и 7739, полученный ПЦР-продукт обрабатывают с применением ферментов рестрикции BglII и XhoI и клонируют в заранее обработанный клонирующий вектор pM-100. Конечная конструкция представлена на Фигуре 2, и белок NMB1125 экспрессируется в виде белка, слитого с 64 кДа Nt-сегментом белка Р. Секвенирование клонируемого гена NMB1125 осуществляют с применением автоматического секвенатора ALFexpress (Termo SequenaseTM CyTM 5 Dye Terminator Kit, Amersham Biosciences) и олигонуклеотидов 1573 (SEQ ID NO: 8) и 6795 (SEQ ID NO: 9), которые связывают, соответственно, последовательность стабилизатора P64 и терминатор транскрипции Т4. Полученную плазмиду обозначают pM-238 для ее последующего использования.
Для экспрессии гена NMB1125 штамм E. coli GC366 трансформируют химическим способом с помощью плазмиды pM238 (Фигура 2). Экспрессию осуществляют в минимальной среде (М9) (Miller J.H. 1972. Experiments in Molecular Genetics, Cold Spring Harbor Laboratory Press, NEW York, США), с добавлением 1% глицерина, 1% казеинового гидролизата, 0,1 мМ CaCl2, 1 мМ MgSO4 и 50 мкг/мл ампициллина. Бактериальные культуры инкубируют 12 часов при 37°С и 250 об./мин. Выращенные культуры центрифугируют, а клеточный осадок разрушают, обрабатывая его ультразвуком (IKA LABORTECHNIK). Фракции из осадка и супернатанта анализируют с помощью ДСН-ПААГ (Laemmli U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 277:680) и окрашивания Кумасси бриллиантовым голубым R-250. Процентную долю экспрессии определяют путем денситометрии геля (LKB Bromma 2202 Ultrascan laser densitometr; Amersham Pharmacia Biotech, United Kingdom). Белок NMB1125 получают из супернатантной фракции, составляющей около 60% от содержания общего белка данной фракции (Фигура 3). Содержащую белок фракцию диализуют против буфера А (25 мМ трис-гидроксиметиламинометан), и из нее с помощью ионообменной хроматографии очищают белок NMB1125, используя колонку monoQ 5/5 (Amersham Biosciences) и градиент NaCl от 0 до 100% в течение 1 ч [буфер А в качестве уравновешивающего буфера и буфер В (буфер А + 1 М NaCl) в качестве градиентного буфера], в результате чего получают на 80% очищенный белок, как показано на Фигуре 4.
Пример 4: Оценка иммунного ответа, индуцируемого путем внутрибрюшинной и интраназальной иммунизации белка NMB1125.
Чтобы оценить иммуногенность белка NMB1125, планируют и осуществляют эксперимент по иммунизации мышей, в котором один и тот же белок вводят двумя разными способами. Первый способ заключается в экстрагировании соответствующей полосы из полиакриламидного геля (Castellanos L. et al. 1996. A procedure for protein elution from reverse-stained polyacrylamide gels applicable at the low picomole level: An alternative route to the preparation of low abundance proteins for microanalysis. Electrophoresis 17:1564-1572), а второй способ указан в Примере 3, в котором полученный продукт обозначен как наполовину очищенный белок.
Данными препаратами иммунизируют самок мышей Balb/C (в возрасте 8-10 недель), ранее разделенных на 4 группы по 8 мышей в каждой. Осуществляют три иммунизации интраназальным или внутрибрюшинным путем с 15-дневным интервалом между иммунизациями. При внутрибрюшинном пути введения белок эмульгируют адъювантом Фрейнда. В Таблице 1 описывается состав соответствующих иммуногенов:
Группы мышей Balb/C, используемых для иммунизации
Титры антител (IgG) против рекомбинантного белка и к соответствующего гомологичного белка, представленного у бактерии, определяют с помощью ИФА в образцах сыворотки, взятых после третьей инокуляции. На Фигуре 5 показан титр антител против рекомбинантного белка у индивидуальных животных. Уровень антител определяют после второй инокуляции, хотя он оказывается выше после третьей инокуляции. Кроме того, с помощью Вестерн-блоттинга осуществляют иммуноидентификацию, в которой распознают полосу соответствующего белка. Группы, иммунизированные внутрибрюшинным путем, обладают существенно более высокими титрами антител, чем группы, иммунизированные интраназальным способом. Для статистического анализа полученных результатов используют непараметрический анализ дисперсии по Крускал-Уоллесу, что обусловлено, в соответствии с критерием Бартлетта, негомогенностью дисперсии в данных группах. Для сравнения средних значений при каждой обработке используют критерий Dunn для множественного сравнения.
Сыворотки, полученные после иммунизации с помощью рекомбинантного белка, распознают природный белок, присутствующий в препарате белка наружной мембраны из штамма CU385. Результаты представлены на Фигуре 6. Для исследования ответа слизистой анализируют образцы слюны и легочные смывы. На Фигуре 7 представлены только группы, иммунизированные интраназальным путем. Повышение титра IgA наблюдают в группе, которая получает полуочищенный белок.
Пример 5: Характеристика последовательности гена, кодирующего белок NMB1125 в разных штаммах N. meningitidis
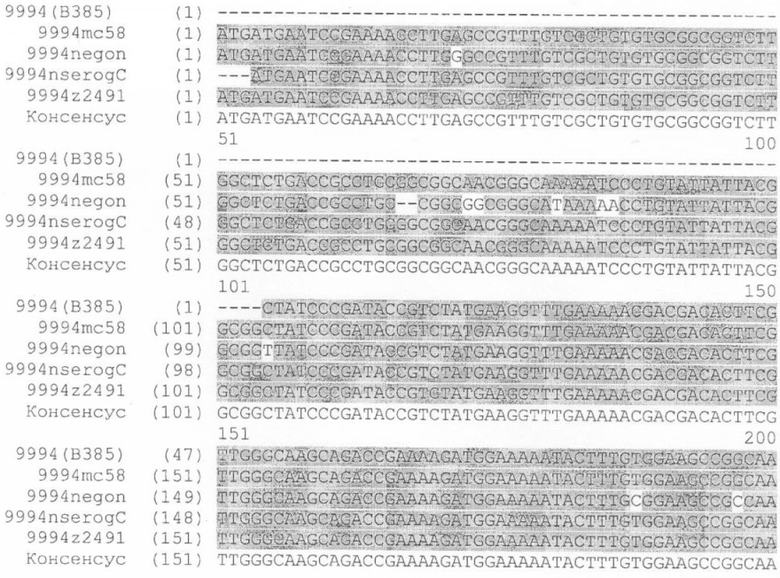
Для исследования консервативной последовательности гена, который кодирует белок NMB1125 в разных штаммах N. meningitidis, осуществляют сходный поиск в отношении геномов Neisseria meningitidis (серогруппы А, В и С), аннотированные в базе данных NCBI (NC 003116.1, NC 003112.1, NC 003221, SANGER 135720|Contig1), используя программу BLAST (Altschul S.F. et al. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. http://www.ncbi.nlm.nih.gov/BLAST/). На Фигуре 8 представлены результаты сравнения последовательности для тех последовательностей, которые подверглись существенному выравниванию в каждом из анализируемых геномов. Последовательности в группах А и В обладают 100%-ной идентичностью с последовательностью, полученной для гена, кодирующего белок NMB1125 (SEQ ID NO: 3), и 99%-ной идентичностью в серогруппе С. Кроме того, последовательность указанного гена, определяемую в 3-х Кубинских изолятах (SEQ ID NO: 5-7), относят к серогруппе В (B:4:P1.19,15), а выравнивание последовательности осуществляют с применением программы ClustalX (http://www.ebi.ac.uk/clustalw/). Полученные результаты выравнивания свидетельствуют о большом консерватизме нуклеотидной последовательности гена NMB1125 у анализируемых штаммов.
Применение белка NMB1125 в качестве вакцинного кандидата, с учетом высокой степени сходства, существующей у ранее упомянутых последовательностей, должно создавать эффективный иммунный ответ (вследствие перекрестной реактивности) с широким защитным спектром действия против менингококкового заболевания.
Пример 6. Характеристика иммунного ответа широкого спектра действия, индуцируемого иммунизацией мышей Balb/C белком NMB1125
Чтобы оценить, индуцирует ли иммунизация белком NMB1125 перекрестную реактивность широкого спектра действия с другими штаммами Neisseria, осуществляют ИФА. Полистироловые планшеты покрывают цельными клетками от 7 штаммов Neisseria, которые принадлежат разным серотипам или серологическим подтипам. Планшеты инкубируют вместе со смешанными сыворотками, полученными против белка NMB1125 с помощью двух способов иммунизации, как описано в Примере 4.
На Фигуре 9 представлены результаты, полученные для сывороток, полученных против вводимого внутрибрюшинным путем наполовину очищенного белка. Отмечается, что иммунные сыворотки узнают представленный в разных штаммах белок на уровнях, сравнимых с уровнем, обнаруживаемым в случае штамма CU385. Остальные сыворотки имели сопоставимую реакцию в данном анализе.
Пример 7. Защита против гомологичных и гетерологичных штаммов, индуцированная специфичной в отношении белка NMB1125 мышиной сывороткой на модели детенышей крыс.
Чтобы определить функциональную активность полученных антисывороток, анализ защиты осуществляют на модели детенышей крыс с менингококковой инфекцией. Двадцать четыре крысы (в возрасте 5-6 дней) распределяют по группам, по 6 крыс в каждой группе.
Было установлено, что если крысам вводят сыворотку внутрибрюшинным путем, предохраняющим от инфекции, вызванной бактериями (штамм CU385), то инокуляцию осуществляют спустя один час таким же способом. Полученные в каждой группе сыворотки объединяют и разбавляют 1/10 (в стерильном PBS) перед тем, как их инокулируют детенышам крыс. Через четыре часа у инокулированных животных берут образцы крови, и в них подсчитывают количество жизнеспособных бактерий.
Для интерпретации полученных результатов осуществляют анализ дисперсии (Anova), с последующим тестом множественных сравнений Дюннета, в котором тестируемые группы сравнивают с негативным контролем. Как можно видеть на Фигуре 10, группы, которые получают антисыворотки против белка NMB1125, демонстрируют статистически значимые различия с негативным контролем, и поэтому они считаются в этой модели защитными.
Аналогичный анализ осуществляют, заражая детенышей крыс штаммами H44/48 и 120/90, выделенными из организма кубинских пациентов, серологическая классификация которых гомологична штамму CU385. Кроме того, осуществляют эксперименты по контрольному заражению штаммом 233 (C:2a:P1.5) из серогруппы С и штаммом H44/76 (B:15:P1.7,16) из серогруппы В. Во всех случаях указанные антисыворотки защищают детенышей крыс от менингококковой инфекции.
Пример 8: Образование моноклональных антител против белка NMB1125, способных опосредовать бактерицидную активность против Neisseria meningitidis
Для создания моноклональных антител (mAb), специфичных в отношении белка NMB1125, и для изучения функциональной способности опосредовать бактерицидную активность против гомологичного и гетерологичного штамма N. meningitidis осуществляют схему иммунизации препаратом белка NMB1125 со степенью очистки, превышающей 80% (Пример 3). Такую иммунизацию осуществляют на мышах Balb/C (H-2d, самка, возраст 5-6 недель) 4-мя порциями по следующей схеме: в дни 0, 15 и 30 при стандартной иммунизации подкожно вводят эмульгированный адъювантом Фрейнда антиген NMB1125 из расчета 10 мкг на мышь (общий объем 100 мкл); на 50-й день внутрибрюшинным путем вводят антиген в фосфатно-солевом растворе (140 мМ NaCl, 270 мМ KCl, 1,5 мМ KH2PO4, 6,5 мМ Na2PO4×2H2O, рН 7,2) из расчета 10 мкг на мышь. Кровь берут в дни 0 и 45.
Спленоциты от животного с наивысшим титром, измеряемые с помощью непрямого ИФА с применением белка NMB1125 в качестве покрывающего антигена (Пример 3), сливают с мышиными миеломными клетками X63 Ag8 653. Полученные гибридомы выделяют и подвергают скринингу в соответствии со стандартной методикой (Gavilondo J.V. 1995. Anticuerpos Monoclonales: Teoria y Práctica, Elfos Scientiae, La Habana, Куба).
Реакционную способность секретируемых гибридомами антител, специфичных в отношении белка NMB1125, а также перекрестно-реагирующих с ними неродственных антигенов, тестируют с помощью непрямого ИФА с применением 5 мкг/мл каждого антигена и такой же концентрации каждого анализируемого mAb. На Фигуре 11 представлены результаты, полученные в данном эксперименте. Получено всего 3 позитивных клона (mAb H8/92, 3H2/64 и 7D2/15), которые специфически распознают белок NMB1125 и не реагируют ни с аминокислотной последовательностью, соответствующей N-концевой области P64k, ни с остальными анализируемыми неродственными антигенами.
Чтобы определить способность mAb, полученных против белка NMB1125, опосредовать бактерицидный ответ против гомологичного и гетерологичного штаммов Neisseria meningitidis, осуществляют бактерицидный тест. Титр бактерицидных антител выражают в виде реципрокного максимального разведения тестируемых антител, которые способны убить 50% или более бактерий, два вида образованных mAb (3H2/64 и 7D2/15) обладают бактерицидными титрами, превышающими разведение 1:128 против гомологичного штамма B:4:P1.19,15, а один вид mAb (H8/92) превышает разведение 1:80. Более того, они обладают титрами, превышающими разведение 1:64, против гетерологичных штаммов B:15:P1.7,16 и C:2a:P1.5.
Пример 9. Характеристика участков-мишеней иммунного ответа против белка NMB1125 у мышей
Для того, чтобы идентифицировать участки в белке, которые чаще всего распознаются мышиными антисыворотками, полученными против рекомбинантного антигена, осуществляют SPOTScan-анализ. Совокупность перекрывающихся пептидов, которые охватывают последовательность белка, синтезируют на целлюлозной мембране, которую инкубируют с объединенными сыворотками с разведением 1:100. Реакцию антиген-антитело детектируют путем инкубирования с конъюгатом антимышиный иммуноглобулин G-щелочная фосфатаза, с последующим добавлением раствора, который содержит субстрат бром-хлор-индолилфосфат.
Отмечено несколько антигенных участков, присущих, независимо от препарата, белку, который используют для иммунизации. Наряду с этим в группах, иммунизированных данным белком, вместе с адъювантом Фрейнда, наблюдается гораздо более полный тип распознавания.
Пример 10: Распознавание белка NMB1125 с помощью человеческих сывороток
В данном исследовании, в котором осуществляют ИФА, используют набор человеческих сывороток, собранных от выздоравливающих пациентов. Планшеты покрывают белком NMB1125, который получают с помощью препаративного электрофореза (5 мкг/мл). После блокирования данных планшетов 3%-ным обезжиренным порошковым молоком в PBS, содержащем твин-20, указанные сыворотки разбавляют (1:50) в этом же растворе и инкубируют в планшетах. Иммуноанализ продолжает далее известным способом. В качестве негативного контроля используют сыворотки здоровых доноров. Кроме того, объединенные сыворотки от пациентов, вакцинированных с помощью рекомбинантной вакцины против гепатита В, используют в независимом контроле (данные не представлены).
На Фигуре 12 представлены результаты, полученные в данном исследовании с сыворотками от 5 выздоравливающих пациентов. Можно видеть, что человеческие сыворотки распознают данный белок, что свидетельствует о том, что он экспрессируется в течение менингококковой инфекции и что он является иммуногенным.
Пример 11: Белок NMB1125 в качестве носителя для пептида и полисахарида
Чтобы продемонстрировать функциональную активность рекомбинантного белка NMB1125 в качестве переносчика, его конъюгировали с 15-членным синтетическим пептидом, полученным из V3-области белка др120 ВИЧ-1, изолят JY1. Конъюгирование осуществляли, используя глутаральдегидный способ.
Свободный пептид JY1, рекомбинантный белок NMB1125 и конъюгат JY1-NMB1125 вводили взрослым мышам по схеме, насчитывающей 3-кратное введение в составе дозировок, в которых иммуногены были эмульгировали с адъювантом Фрейнда. Через две недели после введения третьей дозы у иммунизированных животных получали образцы сыворотки и их анализировали, используя метод ELISA, с целью определения титров антител против пептидов. Для этого планшеты покрывали свободным пептидом (20 мкг/мл), и осуществляли иммунный анализ, как это было описано прежде. Полученные в данном эксперименте результаты (Фигура 13) свидетельствуют о способности белка NMB1125 функционировать в качестве носителя, способного, после его конъюгации с пептидом JY1, существенно усиливать антительный ответ против указанного пептида.
Кроме того, белок NMB1125 конъюгировали с полисахаридным антигеном, выбранным в качестве модельного вакцинного антигена. Капсулярный полисахарид N. meningitidis серогруппы С (MenC) был конъюгирован с NMB1125, с использованием дигидразина адипиновой кислоты и карбодиимидного метода. Сразу же по завершении процедуры конъюгирования конъюгат очищали путем гель-фильтрации на колонке, набитой Сефарозой CL-4B (Amersham Pharmacia Biotech). В полученном в результате конъюгате отношение полисахарида к белку составляло порядка 1,25 мг полисахарида MenC / мг белка NMB1125.
С целью иммунизации мышам подкожным путем вводили три дозы MenC (2,5 мкг), NMB1125 (3 мкг) или MenC-NMB1125 (2,5 мкг MenC/дозу), абсорбированные на гидроокиси алюминия. Во всех случаях иммунизацию производили в объеме 100 мкл в дни 0, 14 и 28. Образцы сыворотки отбирали вначале эксперимента и на 15 день после каждой из иммунизации. Эти образцы хранили, а затем подвергали анализу на предмет определения титров антител против MenC. Результаты, которые были получены, суммированы в Таблице 2. Из таблицы следует, что уже после введения первой дозы в группе, иммунизированной конъюгатом, достигались более высокие титры. Уровни антител продолжали возрастать после введения второй дозы, и уровень этот стабильно удерживался после введения третьей дозы.
Титры антител против MenC в суммарных машиных сыворотках, полученных у иммунизированных мышей.
Таким образом, суммируя результаты экспериментов по иммунизации, можно смело заключить, что белок NMB1125 существенно повышал титры антител как против пептидного антигена, так и против полисахаридного антигена.
Пример 12: Присутствие антигена NMB1125 в нескольких бактериях из рода Neisseria и демонстрация их перекрестной реактивности.
Прежде всего было выполнено выравнивание генов nmb1125 neisserial. Последовательности были взяты из опубликованных данных по геномам Neisseria. Выравнивали следующие последовательности: Neisseria meningitidis MC58:gi_7226399, Neisseria meningitidis_Z2491:gi_7380006, Neisseria_meningitidis_FAM18:orf2, Neisseria_gonorrhoeae_FA1090:orf824. Ниже представлены результаты выравнивания, из которых следует, что высокая степень гомологии наблюдается не только между генами, обнаруживаемыми в нескольких штаммах N. meningitidis, но также и между генами, обнаруживаемыми в штаммах N. meningitidis и N. gonococci. Соответственно, антиген NMB1125 можно расценивать - и, соответственно, исследовать - в качестве защитного антигена для борьбы с заболеваниями, вызываемыми целым рядом видов штаммов, относящихся к одному и тому же роду Neisseria.

Кроме того, для подтверждения наличия указанного антигена в нескольких штаммах Neisseria было проведено два вида экспериментов - цельноклеточный нализ ELISA [Whole Cell ELISA, (WCE)] и эксперименты по иммуноблоттингу.
Для экспериментов WCE различные изоляты Neisseria культивировали в системе Brain Heart Infusion agar plates и собранные в результате культивирования цельные клетки были использованы для покрытия планшетов полученной клеточной суспензией. После запаивания планшеты инкубировали в присутствии суммарной антисыворотки мышей, полученной в результате иммунизации мышей тремя дозами рекомбинантного антигена в сочетании с тремя различными адъювантами. После завершения экспериментов по ELISA результаты были представлены в виде титров антител, и эти результаты суммированы в представленной ниже Таблице 3. Как видно из таблицы, антисыворотка реагировала с антигенами во всех исследуемых штаммах, включая штамм N. lactamica.
В экспериментах по иммуноблоттингу из различных штаммов выделяли антигены интактных цельных клеток и проводили электроблоттинг на нитроцеллюлозных мембранах. Мембранные полоски инкубировали в присутствии суммарных антисывороток (разбавленных в соотношении 1:100), полученных у мышей, иммунизированных рекомбинантным NMB1125. Техника проведенного эксперимента ничем не отличается от таковой, широко применимой в данной области. Результаты проведенного эксперимента представлены на Фиг.14. Антитела, полученные против рекомбинантного белка, реагировали с антигеном, представленным во всех исследованных штаммах, включая штамм N. lactamica, выбранный для данного экспериментального исследования.
Результаты, полученные в исследованиях, проведенных двумя указанными методами, WCE и иммуноблоттинга, дают возможность сделать заключение, что антиген NMB1125 является консервативным во всех исследованных штаммах Neisseria, которые относятся к роду Neisseria meningitidis и Neisseria lactamica. Более того, антитела против антигена, присутствующего у одного вида, способны перекрестно реагировать с антигеном, присутствующим не только у различных изолятов, но даже и с различными видами - представителями Neisseria.
Пример 13: Приготовление композиций, содержащих рекомбинантный полипептид NMB1125 отдельно или в сочетании с несколькими антигенами
Для указанной цели было получено несколько композиций. На основе фармацевтически активного ингредиента были получены две жидкие композиции, содержащие соответственно 20 и 50 мкг NMB1125 на дозу. Эти композиции содержали также 0,5 мг/мл гидроокиси алюминия, 1,12 мг/мл гидрофосфата натрия, 1,10 мг/мл моногидратдигидрофосфата натрия, 8,0 мг/мл хлористого натрия и 0,05 мг/мл тиомерсала (эксципиенты). Третью композицию получали, включая 0,5 мг/мл алюминийфосфата вместо гидроокиси алюминия. Четвертую композицию получали, включая указанные выше компоненты, а кроме того, менингококковый капсулярный полисахарид из серогруппы С (MenC) в концентрации 50 мкг/мл. Пятую композицию получали, полностью заменяя полисахаридный С-компонент тем же самым сахаридом, конъюгированным с белком Р64к, менингококковым антигеном, о котором ранее было сообщалось как об эффективном носителе слабых антигенов. MenC конъюгировали с Р64k через дигидразид адипиновой кислоты, используя карбодиимидный метод. Концентрацию конъюгата доводили до 2 мкг Р64k/дозу. Шестая композиция содержала, помимо рекомбинантного белка и указанных эксципиентов, пептид с последовательностью IPGVAYTSPEVAWVGETELSAKASA, который содержит Т-клеточный эпитоп белка Р64k. Концентрацию пептида поддерживали на уровне 10 мкг на дозу.
В результате получали стабильную лиофилизированную композицию. Композиция содержала NMB1125 (2-5 мг/мл). Композиция содержала также буферную систему на основе фосфата, ацетата, цитрата или цитрат-фосфата с ионной силой от 0,1 до 100 мМ для поддержания рН в интервале от 4,5 до 7,4. Композиция включает в себя эффективное количество ЭДТА в качестве хелатобразующего агента в концентрации от 0,1 до 10 мг/мл, а также количество сахара, сахарного спирта, неионного детергента или аминокислоты или их смеси для стабилизации белков в плане поддержания степени их чистоты и биологической активности. Такими ингредиентами могут быть махароза, лактоза, глюкоза, фруктоза, маннит, сорбит, полисорбат 20, полисорбат 80, цистеин, аланин, глицин и гистидин в концентрации от 0,01 до 100 мг/мл. В конце в композицию добавляли эффективное количество наполнителя, выбранного из полиэтиленгликоля (ПЭГ), маннита, декстрана и глицина, в концентрации от 1 до 100 мг/моль. В составе этой композиции белок, описанный в настоящем изобретении, обладал стабильностью, определяемой на основании анализа высокоэффективной жидкостной хроматографии, в течение по меньшей мере 12 месяцев как при 4°С, так и при 28°С.
Один из вариантов лиофилизированной композиции:
Белок NMB1125: 3 мг/мл
ЭДТА Na2×2Н2O: 0,5 мг/мл
Сорбит: 10 мг/мл
ПЭГ 6000: 5 мг/мл
Полисорбат 80: 0,02 мг/мл
Na2HPO4: 6.34 мг/мл
NaH2PO4×2Н2O: 1,72 мг/мл



| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛОК NMB0928 И ЕГО ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ | 2004 |
|
RU2335505C2 |
| ВАКЦИНЫ С ИСПОЛЬЗОВАНИЕМ ВЕЗИКУЛ НА ОСНОВЕ GNA 1870 ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ДЛЯ ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ NEISSERIA MENINGITIDIS | 2006 |
|
RU2432962C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТРАНСФЕРРИН-СВЯЗЫВАЮЩИЙ БЕЛОК И HSF ИЗ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ | 2003 |
|
RU2359696C2 |
| МУЛЬТИВАЛЕНТНАЯ ВАКЦИНА ИЗ НАТИВНЫХ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ МЕНИНГОКОККОВ, СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2009 |
|
RU2477145C2 |
| УСОВЕРШЕНСТВОВАНИЯ, КАСАЮЩИЕСЯ ВЕЗИКУЛ ВНЕШНЕЙ МЕМБРАНЫ МЕНИНГОКОККОВ | 2005 |
|
RU2420312C2 |
| СПОСОБ БЕЗДЕТЕРГЕНТНОГО ПОЛУЧЕНИЯ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ ГРАМОТРИЦАТЕЛЬНОЙ БАКТЕРИИ | 2012 |
|
RU2607021C2 |
| МЕНИНГОКОККОВЫЕ ПОЛИПЕПТИДЫ fHBP | 2012 |
|
RU2567003C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ ИНФЕКЦИИ, ВЫЗВАННОЙ БАКТЕРИЕЙ NEISSERIA | 2000 |
|
RU2244749C2 |
| КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ АНТИГЕНЫ NEISSERIA MENINGITIDIS ИЗ СЕРОГРУПП В И С И ДОПОЛНИТЕЛЬНЫЙ АНТИГЕН | 2000 |
|
RU2361609C2 |




Изобретение относится к биотехнологии. Описан белок из N.meningitidis, названный NMB1125, представляющий собой антиген, способный вызывать в организме реципиента защитный ответ против инфекций, вызываемых бактериями из рода Neisseria, и имеющий установленную аминокислотную последовательность. Представлен ген, кодирующий описанный белок. Настоящее изобретение может использоваться для получения композиций, обладающих расширенным иммунным диапазоном, и, тем самым, расширить диапазон их действия против различных патогенов. 4 н. и 8 з.п. ф-лы, 14 ил., 3табл.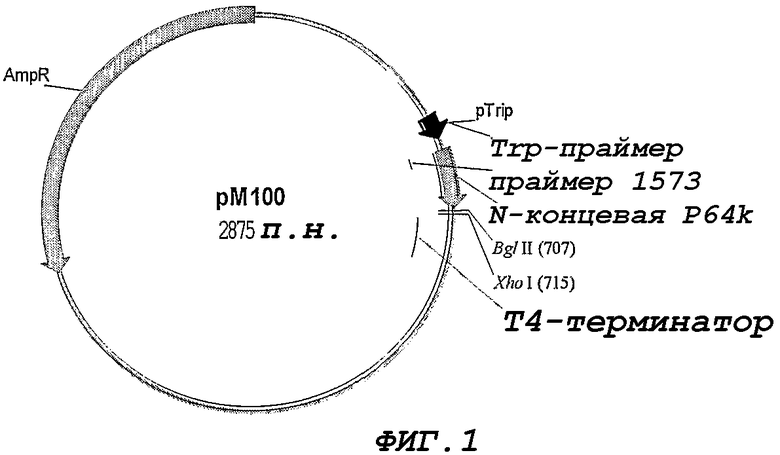
| RU 2001134170 А, 19.05.2000 | |||
| RU 2001134171 А, 19.05.2000 | |||
| JP 2003189879, 08.07.2003. |

