Перекрестная ссылка на связанные заявки
По настоящей заявке испрашивается приоритет по предварительной заявке США с серийным номером №60/647911, зарегистрированной 27 января 2005, которая приведена здесь полностью в качестве ссылки.
Заявление относительно финансируемого государством исследования
Это изобретение было осуществлено при правительственной поддержке в рамках грантов Министерства здравоохранения № RO1 AI46464, R21 AI061533 от Национального Института Аллергии и Инфекционных Заболеваний, относящегося к Национальным Институтам Здоровья, и T32-HL007951 от Национального Института Сердца, Легких и Крови, относящегося к Национальным Институтам Здоровья. Правительство может обладать определенными правами на это изобретение.
Область техники, к которой относится изобретение
Это изобретение относится к вакцинам широкого спектра действия для профилактики заболеваний, вызываемых Neisseria meningitidis.
Уровень техники
Neisseria meningitidis представляет собой грамотрицательную бактерию, заселяющую верхние дыхательные пути человека и ответственную за распространенные во всем мире спорадические и циклические вспышки эпидемий, что особенно касается менингита и сепсиса. Показатели поражения и заболеваемости наиболее высоки для детей в возрасте до 2 лет. Подобно другим грамотрицательным бактериям, Neisseria meningitidis, как правило, имеет цитоплазматическую мембрану, пептидогликановый слой, наружную мембрану, которая вместе с полисахаридом капсулы формирует бактериальную стенку, и пили, выступающие во внешнюю среду. Имеющие капсулу штаммы Neisseria meningitidis являются основной причиной бактериального менингита и септицемии у детей и молодежи (Rosenstein et al. J Infect. Dis. 1999; 180:1894-901).
Человек представляет собой единственный известный резервуар видов Neisseria meningitidis. Соответственно, виды Neisserial в ходе эволюции выработали широкое разнообразие высокоэффективных способов уклонения от иммунной системы человека. Эти способы включают экспрессию полисахаридной капсулы, структурно сходной с полисиаловой кислотой организма-хозяина (т.е. серогруппой B), и высокую антигенную изменчивость в отношении иммунодоминантных некапсульных эпитопов, т.е. эпитопов антигенов, которые присутствуют на поверхности в относительно больших количествах, доступны для антител и вызывают значительный гуморальный иммунный ответ.
Широкая распространенность и экономическая значимость инвазивных инфекций Neisseria meningitidis привели к поиску эффективных вакцин, способных приводить к выработке иммунитета в отношении диапазона серотипов, а особенно диапазона серотипов или сероподтипов группы B. Однако многочисленные попытки разработать вакцины широкого спектра действия были затруднены широким разнообразием высокоэффективных способов, используемых видами Neisserial для уклонения от иммунной системы человека.
Вакцины на основе капсул доступны для профилактики заболевания, вызываемого штаммами группы A, C, Y и W-135 (обзор в Granoff et al. Meningococcal Vaccines. In: Plotkin SA, Orenstein WA, eds. Vaccines. 4th ed. Philadelphia: W. B. Saunders Company, 2003). Однако не существует вакцины, утвержденной для использования в США или Европе для профилактики заболевания, вызываемого штаммами группы B, которые являются причиной приблизительно 30% случаев заболевания в Северной Америке (Lingappa et al. Vaccine 2001; 19:4566-75; Raghunathan et al. Annu Rev. Med. 2004; 55:333-5) и более 2/3 случаев в Европе (Cartwright et al. Vaccine 2001; 19:4347-56; Trotter et al. Lancet 2004; 364:365-7). Одной из причин отсутствия вакцин на основе капсулы группы B является то, что капсула группы B может вызывать у человека реакцию аутоантител (Finne et al. Lancet 1983; 2:355-7), а полисахарид является недостаточно иммуногенным, даже при конъюгации с белками-носителями (Jennings et al. J. Immunol. 1981; 127:1011-8). Кроме того, в случае вакцины на основе капсулы группы B существуют возможные проблемы с безопасностью, поскольку вакцина способна вызывать образование аутореактивных антител против капсулы группы B. Поэтому недавнее исследование менингококковой вакцины группы B было сосредоточено на использовании некапсульных антигенов.
Для вакцин на основе везикул наружной мембраны (OMV) было подтверждено, что они вызывают у человека защитный иммунитет против менингококкового заболевания группы B (обзор в Jodar et al. Lancet 2002; 359:1499-1508). Недавно в Новой Зеландии под влиянием воздействия общественного здравоохранения лицензировали и внедрили вакцину OMV для прекращения эпидемического заболевания группы B, которое продолжалось в течение более десяти лет (Thomas et al. N Z Med. J 2004; 117:U1016; Desmond et al. Nurs N Z 2004; 10:2; Baker et al. J Paediatr Child Health 2001; 37:S 13-9). Были описаны другие способы иммунизации на основе везикул (см., например, Cartwright K. et al, 1999, Vaccine; 17:2612-2619; de Kleinjn et al, 2000, Vaccine, 18:1456-1466; Rouupe van der Voort ER, 2000, Vaccine, 18:1334-1343; Tappero et al., 1999, JAMA 281:1520; Rouupe van der Voort ER, 2000, Vaccine, 18:1334-1343; патент США 2002/0110569; WO 02/09643).
Иммунизация детей и взрослых менингококковыми вакцинами на основе везикул наружной мембраны (OMV) индуцирует бактерицидные антитела сыворотки, серологический аналог защиты от заболевания (Goldschneider et al., 1969, J. Exp. Med. 129:1307). Также эффективность вакцин OMV для профилактики менингококкового заболевания B была непосредственно показана для детей старшего возраста и взрослых в рандомизированных проспективных клинических испытаниях и в ретроспективных исследованиях случай-контроль. Таким образом, клиническая эффективность вакцин на основе везикул наружной мембраны не является предметом обсуждения. Такие вакцины лицензированы в Новой Зеландии для использования у детей и близки к лицензированию в Норвегии для использования у детей старшего возраста и взрослых, а также находятся на поздней стадии клинической разработки для лицензирования в других европейских странах. Также является коммерчески доступной и была введена миллионам детей в Южной Америке вакцина OMV, полученная в Finley Institute на Кубе.
Однако для реакции бактерицидных антител сыворотки на вакцины OMV свойственна штаммоспецифичность (Tappero et al., 1999, JAMA 281:1520; и Rouupe van der Voort ER, 2000, Vaccine, 18:1334-1343). Кроме того, доступные в настоящее время вакцины OMV также ограничены тем, что реакции бактерицидных антител в основном направлены против представленных на поверхности петель основного белка порина, PorA (Tappero et al. JAMA 1999; 281:1520-7), который различается в отношении антигенов (Sacchi et al. J Infect. Dis 2000; 182:1169-76). Вследствие иммунодоминантности PorA индуцируемый иммунитет преимущественно специфичен к штаммам, из которых были получены везикулы мембран (Tappero et al., 1999, JAMA 281:1520; Martin SL et al., 2000, Vaccine, 18:2476-2481). Таким образом, вакцины OMV эффективны для профилактики заболевания в случаях эпидемии, вызываемой преобладающим менингококковым штаммом с единственным сероподтипом PorA, например, эпидемическим штаммом P1.4 в Новой Зеландии (Baker et al. 2001, выше). Однако существует значительное разнообразие PorA среди штаммов, вызывающих эндемическое заболевание, например, встречающееся в США (Sacchi et al. 2000, выше). Кроме того, даже незначительные аминокислотные полиморфизмы могут снижать восприимчивость штаммов к бактерицидной активности антител против PorA (Martin et al. Vaccine 2000; 18:2476-81).
Завершение проектов секвенирования геномов для нескольких штаммов Neisseria meningitidis обеспечило список всех возможных менингококковых белковых антигенов. Посредством сочетания биоинформатики, технологии микрочипов, протеомики и иммунологического отбора было выявлено большое количество новых кандидатов на менингококковые вакцины (Pizza et al. Science 2000; 287:1816-20; De Groot et al. Expert Rev Vaccines 2004; 3:59-76). Среди этих многочисленных кандидатов находится полученный из генома антиген Neisserial 1870 (GNA1870). GNA1870, который также известен как NMB 1870 (WO 2004/048404) или LP2086 (см., например, Fletcher et al. Infect. Immun. 2004 72:2088-2100), представляет собой липопротеин приблизительно 27 кДа, экспрессируемый всеми тестируемыми штаммами N. meningitidis (Masignani et al. J. Exp. Med. 2003; 197:789-99; Giuliani et al. Infect. Immun. 2005; 73:1151-60; Welsch et al. J Immunol. 2004; 172:5606-15).
На основе разнообразия аминокислотных последовательностей и иммунологической перекрестной реактивности штаммы N. meningitidis можно подразделять на три группы вариантов GNA1870 (v.1, v.2 и v.3) (Masignani et al. J. Exp. Med. 2003; 197:789-99). Штаммы варианта 1 составляют приблизительно 60% вызывающих заболевание изолятов группы B (Masignani et al. 2003, выше). В пределах каждой группы вариантов степень консервативности аминокислотной последовательности составляет приблизительно 92% или более.
У мышей, иммунизированных рекомбинантным GNA1870, развивались значительные реакции бактерицидных антител сыворотки против штаммов, экспрессирующих белки GNA1870 гомологичной группы вариантов (Masignani et al. 2003, выше; Welsch et al. 2004, выше). Однако множество штаммов, экспрессирующих субварианты соответствующего белка GNA1870, были устойчивы к бактериолизу, опосредуемому комплементом против GNA1870. Хотя причина этого явления неизвестна, предположительно, это может быть обусловлено незначительными полиморфизмами GNA1870 или различиями штаммов в доступности особо важных эпитопов GNA1870 на поверхности бактерий, что приводит к снижению связывания и/или активации комплемента антителами против GNA1870. Вакцину рекомбинантного белка GNA1870, используемую в указанных выше исследованиях иммуногенности, экспрессировали в E. coli в виде белка с меткой His, не содержащего лидерный пептид. Также у рекомбинантного белка отсутствовал фрагмент, необходимый для посттрансляционной липидизации, что может снижать иммуногенность (Fletcher et al. Infect. Immun. 2004; 72:2088-100).
Были исследованы вакцинные возможности сочетания рекомбинантного PorA и рекомбинантного GNA1870 (Fletcher et al. Infect. Immun. 2004, 72:2088-1200). При введении комбинированной вакцины мышам выраженной интерференции реакций антител на два антигена не наблюдали. Однако рекомбинантному сочетанию требовалось восстановление конформации эпитопов PorA, что необходимо для вызывания образования бактерицидных антител против PorA (см., например, Christodoulides et al., Microbiology, 1998; 144:3027-37 и Muttilainen et al., Microb Pathog 1995; 18:423-36). Кроме того, для комбинированной рекомбинантной вакцины не наблюдали увеличения количества бактерицидных антител против GNA1870 против штаммов N. meningitidis, экспрессирующих субварианты белка GNA1870, который используют в вакцине.
В O'Dwyer et al. (Infect. Immun. 2004; 72:6511-80) описано получение вакцины на основе везикул наружной мембраны (OMV) из штамма-комменсала N. flavescens, генетически сконструированного для экспрессии поверхностного белка A Neisserial (NspA), высококонсервативного белка мембраны менингококков, кандидата для вакцины, который в природе не экспрессируется N. flavescens. У иммунизированных мышей развивалась опсонофагоцитирующая активность сыворотки, специфичная к NspA. Кроме того, после абсорбции антител к OMV остаточные антитела против NspA сообщали пассивную защиту мышам, которые получают летальную дозу имеющего капсулу штамма N. meningitidis. Однако в этом исследовании не наблюдали, чтобы антитела, образование которых вызывается модифицированной вакциной OMV N. flavescens, сообщали более хорошую защиту в сравнении с антителами, образование которых вызывается OMV из N. flavescens, не экспрессирующей гетерологичный антиген. Кроме того, модифицированная OMV N. flavescens не вызывала бактерицидных иммунных ответов, тогда как в предыдущих исследованиях у мышей, которых иммунизировали рекомбинантным NspA, экспрессируемым в везикулах E. coli (Moe et al. Infect. Immun. 1999; 67:5664-75; Moe et al. Infect. Immun. 2001; 69:3762-71) или воспроизведенным в липосомах (Martin et al. In: Thirteenth International Pathogenic Neisseria Conference. Oslo: Nordberg Aksidenstrykkeri AS, 2002), образовывались бактерицидные антитела сыворотки. В публикации PCT № WO 02/09746 и публикации США №20040126389 также описана OMV, получаемая из штаммов, которые сконструированы для сверхэкспрессии антигена Neisserial, где в качестве конкретных примеров таких антигенов указаны NspA, Omp85, пили (PilQ, PilC), PorA, PorB, Opa, Tbp2, TbpA, TbpB, Hsf, PldA, HasR, FrpA/C, FrpB, Hap, LbpA/LbpB, FhaB, lipo02, MltA и ctrAi.
Настоящее изобретение решает недостатки предшествующего уровня техники в отношении способов вакцинации и получения защитного иммунитета против широкого спектра штаммов Neisseria meningitidis, в том числе, особенно (но не исключительно), штаммов, которые относятся к серогруппе B.
ЛИТЕРАТУРА
Bjune et al. NIPH Ann 1991; 14:125-30; обсуждение 130-2; Chen et al. In: Thirteenth International Pathogenic Neisseria Conference Nordberg Aksidenstrykkeri AS, 2002; Christodoulides et al. Microbiology 1998; 144 (Pt 11):3027-37; Claassen et al. Vaccine 1996; 14:1001-8; de Kleijn et al. Vaccine 2000; 18:1456-66; Frasch et al. Meningococcal vaccines: methods and protocols. Totowa, New Jersey: Humana Press, 2001:81-107; Fukasawa et al. FEMS Immunol. Med. Microbiol. 2004; 41:205-10; Holst et al. Vaccine 2003; 21:734-7; Humphries Vaccine 2004; 22: 1564-9; Jansen et al. FEMS Immunol. Med. Microbiol. 2000; 27:227-33; Kijet et al. In: Thirteen international Pathogenic Neisseria Conference Nordberg Aksidenstrykkeri, 2002; Martin et al. Vaccine 2000; 18:2476-81; McGuinness et al. Lancet 1991; 337:514-7; Morley et al. Vaccine 2001; 20:666-87; Muttilainen et al. Microb. Pathog. 1995; 18:423-36; Parmar et al. Biochim Biophys Acta 1999; 1421:77-90; Newcombe et al. Infect. Immun. 2004; 72:338-44; O'Dwyer et al. Infect. Immun. 2004; 72:6511-8; Oliver et al. Infect. Immun. 2002; 70:3621-6 Peeters et al. Vaccine 1996; 14:1009-15; Peeters et al. Vaccine 1999; 17:2702-12; Rouppe van der Voort et al. Vaccine 2000; 18:1334-43; Sanchez et al. Vaccine 2002; 20:2964-71; Steeghs et al. EMBO J 2001; 20:6937-45; Steeghs et al. J Endotoxin Res. 2004; 10:113-9; Troncoso et al. FEMS Immunol. Med. Microbiol. 2000; 27:103-9; Vandeputte et al. J. Biol. Chem. 2003; van der Ley P et al. Vaccine 1995; 13:401-7; Claassen et al. Vaccine 1996; 14:1001-8; Peeters et al. Vaccine 1996; 14:1009-15; Cantini et al. J. Biol. Chem. 2005 Dec 31; [электронная публикация, предшествующая печати].
Сущность изобретения
В целом, настоящее изобретение относится к способам и композициям для вызова у индивидуума иммунной реакции против видов бактерий Neisseria, особенно против штамма Neisseria meningitidis серогруппы B.
В одном из аспектов изобретение относится к композициям, содержащим антигенные везикулы, получаемые из первой бактерии вида Neisseria, где бактерия вида Neisseria продуцирует полипептид GNA1870 на уровне, достаточном для обеспечения получения везикулы, которая при введении индивидууму вызывает образование антител против GNA1870; а также фармацевтически приемлемый носитель. Везикула может представлять собой везикулы наружной мембраны (OMV), микровезикулы (MV) или смесь OMV и MV. Бактерия вида Neisseria может представлять собой природную бактерию или генетически модифицированную в отношении продуцирования полипептида GNA1870 (например, для обеспечения экспрессии полипептида GNA1870 с гетерологичного промотора, для экспрессии экзогенного полипептида GNA1870 и т.п.). Полипептид GNA1870 может являться эндогенным для клетки-хозяина. В некоторых вариантах осуществления бактерия вида Neisseria генетически модифицирована для прекращения продуцирования эндогенного полипептида GNA1870, а также генетически модифицирована для продуцирования полипептида GNA1870 с экзогенной для клетки-хозяина нуклеиновой кислоты. В других вариантах осуществления бактерия вида Neisseria генетически модифицирована для продуцирования по крайней мере двух различных полипептидов GNA1870 (например, полипептидов GNA1870 различных групп вариантов (v.1, v.2 и v.3)). В дополнительных связанных вариантах осуществления бактерия вида Neisseria дефектна по продукции полисахарида капсулы.
В одном из вариантов осуществления композиция дополнительно содержит антигенную везикулу, полученную из второй бактерии вида Neisseria, где вторая бактерия вида Neisseria продуцирует полипептид GNA1870 на уровне, достаточном для обеспечения получения везикул, которые при введении индивидууму вызывают образование антител против GNA1870, и где вторая бактерия вида Neisseria генетически отлична от первой бактерии вида Neisseria (например, первая и вторая бактерии различаются по крайней мере по одному из серогруппы, серотипа или сероподтипа). В дополнительных связанных вариантах осуществления полипептид GNA1870 из второй бактерии вида Neisseria отличается от полипептида GNA1870 из первой бактерии вида Neisseria.
В другом варианте осуществления композиция дополнительно содержит антигенную везикулу, полученную из третьей бактерии вида Neisseria, где третья бактерия вида Neisseria продуцирует полипептид GNA1870 на уровне, достаточном для обеспечения получения везикул, которые при введении индивидууму вызывают образование антител против GNA1870, и где третья бактерия вида Neisseria генетически отлична от первой бактерии вида Neisseria (например, отличается по крайней мере по одному из серогруппы, серотипа или сероподтипа). В связанных вариантах осуществления полипептиды GNA1870 из первой, второй и третьей бактерий вида Neisseria являются различными.
В представляющем особый интерес варианте осуществления композиция содержит первую антигенную везикулу, полученную из первой бактерии Neisseria meningitidis, которая генетически модифицирована для сверхэкспрессии полипептида GNA1870; вторую антигенную везикулу, полученную из второй бактерии Neisseria meningitidis, которая генетически модифицирована для сверхэкспрессии полипептида GNA1870; и фармацевтически приемлемый носитель; где полипептиды GNA1870 из первой и второй бактерий относятся к различным группам вариантов полипептида GNA1870, а первая и вторая бактерии продуцируют различные полипептиды PorA. В связанном варианте осуществления композиция дополнительно содержит третью антигенную везикулу, полученную из третьей бактерии Neisseria meningitidis, которая генетически модифицирована для сверхэкспрессии полипептида GNA1870, где полипептид GNA1870 из третьей бактерии относится к группе вариантов полипептида GNA1870, отличной от групп для первой и второй бактерий, и где третья бактерия продуцирует полипептид PorA, отличный от полипептида PorA из первой и второй бактерий. В дополнительно связанных вариантах осуществления везикулы получают без использования детергента.
В другом аспекте изобретение относится к способу получения антигенной композиции посредством культивирования бактерии вида Neisseria, продуцирующей полипептид GNA1870, где полипептид GNA1870 продуцируется на уровне, достаточном для обеспечения получения везикул, которые при введении индивидууму вызывают образование антител против GNA1870; посредством получения везикул из культивируемой бактерии; и смешиванием везикул с фармацевтически приемлемым носителем для получения эффективной для введения индивидууму антигенной композиции. Первая и вторая везикулы могут независимо друг от друга представлять собой везикулу наружной мембраны (OMV) или микровезикулу (MV). Бактерия вида Neisseria может представлять собой природную бактерию и, следовательно, экспрессировать эндогенный GNA1870 или она может быть генетически модифицирована в отношении продуцирования полипептида GNA1870 (например, для обеспечения экспрессии полипептида GNA1870 с гетерологичного промотора, для экспрессии экзогенного полипептида GNA1870 и т.п.). Полипептид GNA1870 может быть эндогенным в отношении клетки-хозяина. В некоторых вариантах осуществления бактерия вида Neisseria генетически модифицирована для прекращения продуцирования эндогенного полипептида GNA1870. В других вариантах осуществления бактерия вида Neisseria генетически модифицирована для продуцирования по крайней мере двух различных полипептидов GNA1870 (например, полипептидов GNA1870 различных групп вариантов (v.1, v.2 и v.3)). В других вариантах осуществления бактерия вида Neisseria генетически модифицирована для прекращения продуцирования полноразмерного эндогенного полипептида GNA1870 и продуцирует полипептид GNA1870 с нуклеиновой кислоты, экзогенной для клетки-хозяина. В дополнительных связанных вариантах осуществления бактерия вида Neisseria дефектна по продукции полисахарида капсулы.
В другом аспекте изобретение относится к способу вызова иммунной реакции против Neisseria посредством введения млекопитающему иммунологически эффективного количества композиции, содержащей первый антигенный препарат, который включает везикулы, полученные из первой бактерии вида Neisseria, где бактерия вида Neisseria продуцирует полипептид GNA1870 на уровне, достаточном для обеспечения получения везикул, которые при введении индивидууму вызывают образование антител против GNA1870; где указанное введение достаточно для вызова иммунной реакции на присутствующий в везикулах полипептид GNA1870. Везикулы могут представлять собой везикулы наружной мембраны (OMV), микровезикулы (MV) или смесь OMV и MV. Бактерия вида Neisseria может представлять собой природную бактерию и, следовательно, экспрессировать эндогенный GNA1870 или она может быть генетически модифицирована в отношении продуцирования полипептида GNA1870 (например, для обеспечения экспрессии полипептида GNA1870 с гетерологичного промотора, для экспрессии экзогенного полипептида GNA1870 и т.п.). Полипептид GNA1870 может быть эндогенным в отношении клетки-хозяина. В некоторых вариантах осуществления бактерия вида Neisseria генетически модифицирована для прекращения продуцирования эндогенного полипептида GNA1870. В других вариантах осуществления бактерия вида Neisseria была сконструирована для сверхэкспрессии GNA1870. В еще одних вариантах осуществления полипептид GNA1870 представляет собой химерный белок (слитый белок), где химерный белок содержит по крайней мере антигенный участок GNA1870 для презентирования на везикулах (например, OMV, MV). В дополнительных связанных вариантах осуществления бактерия вида Neisseria дефектна по продукции полисахарида капсулы.
В других вариантах осуществления бактерия вида Neisseria генетически модифицирована для продуцирования по крайней мере двух различных полипептидов GNA1870 (например, полипептидов GNA1870 различных групп вариантов (v.1, v.2 и v.3)). В других вариантах осуществления бактерия вида Neisseria генетически модифицирована для прекращения продуцирования полноразмерного эндогенного полипептида GNA1870 и продуцирует полипептид GNA1870 с нуклеиновой кислоты, экзогенной для клетки-хозяина.
В связанных вариантах осуществления вводимая по способу композиция содержит иммунологически эффективное количество второго антигенного препарата, включающего везикулы, полученные из второй бактерии вида Neisseria, где вторая бактерия вида Neisseria продуцирует полипептид GNA1870 на уровне, достаточном для обеспечения получения везикул, которые при введении индивидууму вызывают образование антител против GNA1870, и где вторая бактерия вида Neisseria генетически отличается от первой бактерии вида Neisseria (например, первая и вторая бактерии относятся к различным серогруппам, серотипам или сероподтипам). Полипептид GNA1870 из второй бактерии вида Neisseria может отличаться от полипептида GNA1870 из первой бактерии вида Neisseria.
В дополнительных связанных вариантах осуществления композиция дополнительно содержит третий выделенный антигенный препарат, включающий везикулы, полученные из третьей бактерии вида Neisseria, где третья бактерия вида Neisseria продуцирует полипептид GNA1870 на уровне, достаточном для обеспечения получения везикул, которые при введении индивидууму вызывают образование антител против GNA1870, и где третья бактерия вида Neisseria генетически отличается от первой или второй бактерии вида Neisseria (например, первая, вторая и третья бактерии вида Neisseria генетически отличны тем, что они различаются по крайней мере по одному из серогруппы, серотипа или сероподтипа). Полипептиды GNA1870 из первой, второй и третьей бактерий вида Neisseria могут отличаться.
Способ может обеспечивать вызов у индивидуума защитной иммунной реакции против более чем одного штамма Neisseria, особенно N. meningitidis, более конкретно Neisseria meningitidis серогруппы B.
Описанные в настоящей заявке антигенные композиции могут вызывать сочетание оптимальных реакций бактерицидных антител против GNA1870, PorA и/или OpC и, таким образом, сообщать защиту широкого спектра в отношении менингококкового заболевания.
Вакцины, получаемые из сверхэкспрессирующих GNA1870 штаммов, как описано в настоящей заявке, могут вызывать иммунный ответ, который является бактерицидным в отношении штаммов Neisserial, обладающих вариантом GNA1870 и/или PorA из штамма, из которого были получены везикулы, а также иммунный ответ, являющийся бактерицидным в отношении штаммов Neisserial, которые обладают субвариантом GNA1870 и гетерологичным PorA относительно продуцирующего везикулы штамма.
Вакцины, полученные из сверхэкспрессирующих GNA1870 штаммов, также способны снижать вероятность отбора и частоту возникновения вызывающих заболевание штаммов N. meningitidis в популяции со сниженной экспрессией PorA. Эти мутанты представляют особый интерес при широком использовании в популяции общепринятых вакцин OMV. Поскольку экспрессия PorA изменяется в зависимости от фазы (van der Ende et al, J. Bacteriology 1995:177:2475-2480), а мутанты, дефектные по экспрессии PorA, относительно широко распространены и могут быть легко отобраны посредством уничтожения N. meningitidis с использованием антитела против PorA и комплемента. Дефектные по PorA штаммы также являются вирулентными и способны вызывать заболевание.
Также настоящее описание относится к способам, которые могут обладать преимуществами в отношении легкости получения композиции эффективной вакцины, относительно получения вакцины, которая содержит рекомбинантный полипептид, или препарата комбинированной вакцины, включающего множество отдельных антигенов, или рекомбинантного белка, такого как PorA, для которого необходимо восстановление конформационных эпитопов для вызова образования бактерицидных антител.
Из прочтения настоящего описания специалисту в данной области легко понятны аспекты, признаки и преимущества изобретения.
Краткое описание чертежей
На фиг.1A представлены результаты экспериментального измерения проточной цитометрией связывания антител против GNA1870 на поверхности живых, имеющих капсулу клеток N. meningitidis штамма RM1090 и мутантов RM1090, как определено непрямой флуоресценцией. Ряд A. штамм RM1090ΔGNA1870. Ряд B. штамм RM1090 дикого типа. Ряд C. штамм RM1090, трансформированный "челночным" вектором pFP12, который содержит ген GNA1870. Столбец 1. Отрицательный контроль в виде сыворотки (разведение 1:10) от мышей, иммунизированных только фосфатом алюминия. Столбец 2. Положительный контроль в виде моноклонального антитела против полисахарида группы C (10 мкг/мл). Столбец 3. Положительный контроль в виде моноклонального антитела против PorA (против P1.2, разведение 1:500). Столбец 4. Моноклональное антитело против GNA1870 (v. 1) (2 мкг/мл). Столбец 5. Поликлональная антисыворотка против GNA1870, получаемая против рекомбинантных белков v.1, 2 и 3 (разведение 1:10). Столбец 6. Аналогично колонке 5, но при разведении сыворотки 1:250.
На фиг.1B представлено связывание антител с поверхностью живых клеток N. meningitidis группы B, как определено непрямой флуоресцентной цитометрией. Ряд 1: штамм H44/76 дикого типа (серая область); сверхэкспрессирующий GNA1870 мутант H44/76 (черная область). Ряд 2. H44/76ΔGNA1870. Панель A, отрицательный контроль в виде антисыворотки против адъюванта при разведении 1:10; Панель B, моноклональное антитело против PorA (P1.16) при разведении 1:500; Панель C, моноклональное антитело против капсулы в количестве 10 мкг/мл; Панель D, моноклональное антитело JAR3 против rGNA1870 в количестве 10 мкг/мл; Панель E, поликлональная антисыворотка против rGNA1870 при разведении 1:10; Панель F, аналогично панели E при разведении 1:250.
На фиг.2A представлены результаты анализа OMV посредством SDS PAGE и Вестерн-блоттинга. На панели A представлена фотография окрашенного Кумасси SDS PAGE. Дорожки 1-5, препараты OMV (приблизительно 5 мкг белка в каждой дорожке, за исключением дорожки 5, куда загружали 10 мкг). Дорожка 1, штамм RM1090 дикого типа (WT); дорожка 2, штамм WT, который трансформирован "челночным" вектором pFP12, не содержащим ген GNA1870; дорожка 3, нокаутная форма (KO) RM1090ΔGNA1870; дорожка 4, KO RM1090ΔGNA1870, которая трансформирована pFP12, не содержащим гена GNA1870; дорожка 5, KO RM1090ΔGNA1870, которая трансформирована "челночным" вектором pFP12-GNA1870, содержащим ген GNA1870; дорожка 6, rGNA1870 (приблизительно 1 мкг). На панелях B и C представлены фотографии Вестерн-блотов с использованием поликлональной антисыворотки против GNA1870 от мышей, иммунизированных белками rGNA1870 вариантов 1, 2 и 3. Панель B: Чувствительность регистрации этой антисыворотки для варианта 2 (v.2) рекомбинантного белка GNA1870 была незначительно выше в сравнении с вариантом 1 рекомбинантного белка GNA1870 (v.1). Панель C: дорожка 1, рекомбинантный GNA1870 v.1; дорожка 2, OMV из RM1090 WT; дорожка 3, OMV из RM1090ΔGNA1870; дорожка 4, OMV из RM1090, который трансформирован "челночным" вектором pFP12, содержащим ген GNA1870. Сверхэкспрессия GNA1870 v.1 в штамме RM1090ΔGNA1870, трансформированном "челночным" вектором, является более высокой, чем природный уровень экспрессии GNA1870 в штамме дикого типа (дорожка 2).
На фиг.2B представлены результаты анализа Вестерн-блоттингом вакцин OMV, меченных поликлональным антителом против rGNA1870. Дикий тип, OMV, получаемые из штамма H44/76 дикого типа; ΔGNA1870, OMV, получаемые из мутанта H44/76, в котором был инактивирован ген, кодирующий GNA1870; OE GNA1870, OMV из мутанта H44/76, сконструированного для сверхэкспрессии GNA1870; rGNA1870, очищенный GNA1870 с меткой His, экспрессируемый в E. coli.
На фиг.3A представлены графики титров бактерицидной сыворотки мышей, как измерено против четырех иллюстративных штаммов N. meningitidis, имеющих капсулу: Cu385, M6190, Z1092 и NZ98/254. Группы вакцин представляли собой: столбец 1, только адъювант в виде фосфата алюминия; столбец 2, вакцина OMV из RM1090 дикого типа; столбец 3, вакцина OMV из RM1090ΔGNA1870; столбец 4, смесь вакцины OMV из RM1090ΔGNA1870 + рекомбинантный белок GNA1870; столбец 5, вакцина OMV из RM1090, сверхэкспрессирующего GNA1870; столбец 6, рекомбинантный белок GNA1870. Вертикальные черты, представляющие 95% доверительные интервалы для средних геометрических значений, отображают группы вакцин, где анализировали сыворотку от 9-10 отдельных животных. Вертикальные линии со звездочками (*) отображают средние геометрические значения результатов анализа двух совокупностей сывороток из каждой группы вакцин (каждая совокупность из сывороток от 4-5 различных мышей).
На фиг.3B представлены графики бактерицидной активности сыворотки (1/GMT ± SD) для сыворотки от мышей, иммунизированных вакцинами OMV H44/76. Совокупности сывороток получали, как описано в подписи к фиг.3A. Группы мышей иммунизировали с использованием (1) адъюванта, (2) rGNA1870, (3) H44/76 дикого типа (4) H44/76ΔGNA1870 (5) H44/76 OE GNA1870. Хотя на графиках не показано, все штаммы были уничтожены посредством комплемента вместе с положительным контролем в виде моноклональных антител против капсулы и/или против PorA.
На фиг.4A представлены серии графиков, отображающих активацию отложения комплемента C3b и iC3b человека на поверхности живых, имеющих капсулу клеток N. meningitidis, как определено непрямой флуоресцентной проточной цитометрией. Ряд A. штамм NZ98/254. Ряд B. штамм M1390. Столбец 1, комплемент вместе с положительным контролем в виде моноклональных антител (mAb) против капсулы группы B, 25 мкг/мл (открытый участок) или совокупность сывороток в разведении 1:40 от служащих отрицательным контролем мышей, иммунизированных только фосфатом алюминия (закрытый участок). Столбец 2, комплемент вместе с mAb JAR3 против GNA1870, 1 мкг/мл (открытый) или инактивированный нагреванием комплемент + mAb против GNA1870, 5 мкг/мл (закрытый). Столбцы 3, 4 и 5, комплемент вместе с совокупностями сывороток в разведении 1:100 от мышей, иммунизированных с использованием: столбец 3 (вакцина rGNA1870); столбец 4 (вакцина OMV из штамма RM1090 WT); или столбец 5 (смесь вакцины rGNA1870 и вакцины OMV из штамма RM1090ΔGNA1870). Столбец 6, комплемент вместе с разведениями совокупности сывороток от мышей, иммунизированных вакциной OMV из штамма RM1090, сверхэкспрессирующего GNA1870 (открытый участок, разведение 1:100 и промежуточный участок, разведение 1:400). Для сравнения на графиках в столбце 6 также представлены данные для комплемента вместе с совокупностью сывороток в разведении 1:100 от мышей, иммунизированных вакциной OMV из штамма RM1090ΔGNA1870 (закрытый участок).
На фиг.4B представлены серии графиков, отображающих активацию отложения комплемента C3b и iC3b человека на поверхности живых, имеющих капсулу клеток N. meningitidis, как определено непрямой флуоресцентной проточной цитометрией. Штаммы NZ 98/254, BZ198, Z1092 и M6190. Панель A, открытый участок: комплемент вместе с mAb против капсулы (25 мкг/мл для штаммов NZ98/254, BZ198 и M61903 группы B и 1 мкг/мл для штамма Z1092 группы A); закрашенный участок: комплемент вместе с антисывороткой против адъюванта в разведении 1:100. Панель B, открытый участок: комплемент вместе с mAb JAR3 против rGNA1870 в количестве 25 мкг/мл; закрашенный участок: комплемент вместе с поликлональной антисывороткой против rGNA1870 в разведении 1:100. Панель C, комплемент вместе с антисывороткой против OMV из H44/76 дикого типа в разведении 1:100; панель D, открытый участок: комплемент вместе с разведением 1:100 антисыворотки, полученной против OMV со сверхэкспрессируемым GNA1870, которая была абсорбирована с использованием колонки в качестве отрицательного контроля (только Ni-NTA); закрашенный участок: комплемент вместе с разведением 1:25 антисыворотки, полученной против OMV со сверхэкспрессируемым GNA1870 после абсорбции с использованием колонки с твердой фазой GNA1870.
На фиг.5 представлена столбчатая диаграмма, отображающая иммунные ответы сыворотки против GNA1870, как измерено посредством ELISA (GMT ± SD). Антиген на планшете представлял собой вариант 1 rGNA1870. Вторичное антитело представляло собой конъюгированное с щелочной фосфатазой козье антитело против IgG+A+M мыши. Столбцы представляют собой соответствующие средние геометрические значения титров 2 совокупностей антисывороток (4-5 мышей на совокупность) от групп мышей, иммунизированных (1) адъювантом; (2) rGNA1870; (3) OMV H44/76 дикого типа; (4) OMV H44/76ΔGNA1870; (5) OMV H44/76 OE GNA1870.
На фиг.6 представлены графики, отображающие результаты анализа пассивной защиты на модели детенышей крыс с менингококковой бактериемией. В момент времени 0 детенышей крыс обрабатывали внутрибрюшинно (IP) с использованием разведений совокупностей сывороток от иммунизированных мышей (N=9-10 отдельных сывороток в совокупности) и через два часа вызывали иммунную реакцию с использованием штамма NZ98/294 группы B (приблизительно 60000 CFU/крыса, вводимого IP). Через 4-6 часов после стимуляции бактериями получали количественные культуры клеток крови. Панель A: разведения сыворотки 1:15. Панель B: разведения сыворотки 1:60. Столбец 1: сыворотка от мышей, иммунизированных только фосфатом алюминия; столбец 2: mAb против капсулы (10 мкг/крыса); столбец 3: mAb против GNA1870 (10 мкг/крыса); столбец 4: сыворотка от мышей, иммунизированных вакциной OMV из RM1090ΔGNA1870; столбец 5: сыворотка от мышей, иммунизированных смесью вакцины OMV от RM1090ΔGNA вместе с вакциной рекомбинантного белка GNA1870; столбец 6: сыворотка от мышей, иммунизированных вакциной OMV из RM1090, сверхэкспрессирующего GNA1870; столбец 7: сыворотка от мышей, иммунизированных вакциной рекомбинантного белка GNA1870.
На фиг.7 представлено выравнивание типичных аминокислотных последовательностей вариантов 1, 2 и 3 GNA1870 из штаммов N. meningitidis MC58, 951-5945 и M1239 соответственно. "1" указывает на первую аминокислоту зрелого белка, а аминокислоты, являющиеся частью лидерной последовательности, указаны отрицательными числами. Серый и черный фоны указывают консервативные и идентичные аминокислотные остатки соответственно.
На фиг.8A-8H представлены аминокислотные последовательности типичных, эффективных по изобретению полипептидов GNA1870, включая выравнивания аминокислотных последовательностей выбранных типичных полипептидов GNA1870 (фиг.8H).
На фиг.9 представлены выравнивания аминокислотных последовательностей типичного прототипа PorA семейства VR1 (панель A) и аминокислотных последовательностей типичного прототипа PorA семейства VR2 (панель B).
Перед описанием настоящего изобретения и иллюстративных вариантов осуществления изобретения следует понимать, что это изобретение не ограничено конкретными описанными вариантами осуществления, поскольку таковые могут, безусловно, различаться. Также следует понимать, что используемая в настоящей заявке терминология представлена только в целях описания конкретных вариантов осуществления и не предназначена для ограничения, поскольку объем настоящего изобретения ограничен только прилагаемой формулой изобретения.
Если представлен диапазон величин, то следует понимать, что в изобретение входит каждая находящаяся между верхним и нижним пределами этого диапазона промежуточная величина, до десятой доли единицы величины нижнего предела, пока в контексте ясно не указано иначе, а также любая другая указанная или промежуточная величина в этом указанном диапазоне. Верхний и нижний пределы этих меньших диапазонов могут быть независимо включены в меньшие диапазоны, которые также входят в изобретение, допуская любой конкретный исключаемый предел в указанном диапазоне. Если указанный диапазон включает один или оба предела, то диапазоны, в которых исключены оба этих включенных предела, также входят в изобретение.
Если не указано иначе, все используемые в настоящей заявке технические и научные термины имеют то же значение, в котором их обычно понимает специалист в данной области, к которой относится это изобретение. Хотя в осуществлении на практике или тестировании настоящего изобретения можно также использовать любые способы и вещества, сходные или эквивалентные тем, которые описаны в настоящей заявке, далее приведены предпочтительные способы и вещества. Все указанные в настоящей заявке публикации приведены здесь полностью в качестве ссылки для изложения и описания способов и/или веществ, в связи с которыми публикации процитированы.
Следует отметить, что в рамках настоящей заявки и прилагаемой формулы изобретения формы единственного числа "некоторый", "и" и "этот" включают обозначаемые объекты во множественном числе, если в контексте ясно не указано иначе. Таким образом, например, ссылка на "некоторый антиген" включает множество таких антигенов, а ссылка на "эту везикулу" включает ссылку на одну или несколько везикул и их эквивалентов, известных специалисту в данной области, и т.д.
Указанные в настоящей заявке публикации представлены только для их описания до даты регистрации настоящей заявки. Ничто в настоящей заявке не следует рассматривать, как признание того, что настоящее изобретение не правомочно предшествовать такой публикации вследствие свойства предшествующего изобретения. Кроме того, даты представленной публикации могут отличаться от фактических дат публикации, для которых может быть необходимо независимое подтверждение.
Подробное описание изобретения
Настоящее изобретение основано на открытии, что вакцина OMV, которую получают из мутантного штамма N. Meningitidis, сконструированного для сверхэкспрессии GNA1870, вызывает у мышей бактерицидные гуморальные иммунные ответы более широкого спектра, чем вакцина из рекомбинантного белка GNA1870 (rGNA1870) или OMV, полученная из природного штамма, или сочетание вакцины из рекомбинантного белка и вакцины OMV.
Вакцины OMV были безопасно введены миллионам людей, и для них подтверждена эффективность против развивающегося менингококкового заболевания. Как указано во вводном разделе, их основное ограничение заключается в том, что они вызывают штаммоспецифичные бактерицидные гуморальные иммунные ответы. Кроме того, существует проблема, что при широком использовании вакцин OMV в популяции иммунная реакция может служить отбором на предмет возникновения "ускользающих мутантов" штаммов N. meningitidis (т.е. штаммов с мутациями в аминокислотной последовательности петель PorA, доступных на поверхности, или со сниженной экспрессией PorA). Вкратце, изобретение, посредством отбора преобладающего сероподтипа PorA и получения сверхэкспрессирующего GNA1870 мутанта, обеспечивает возможность получения вакцины на основе везикул (например, OMV, MV), вызывающей сочетание оптимальных бактерицидных гуморальных иммунных ответов против GNA1870 и против PorA и, тем самым, сообщающей защиту широкого спектра от менингококкового заболевания. Также для использования такой вакцины свойственен более низкий риск в отношении отбора вызывающих заболевание, дефектных по PorA мутантных штаммов в популяции в сравнении с общепринятой вакциной OMV.
Кроме того, везикулы, получаемые из сверхэкспрессирующего штамма GNA1870, имеют измененный профиль белков в сравнении с везикулами, получаемыми из штамма, который экспрессирует GNA1870 на относительно более низком уровне. Как более подробно описано в примерах, у OMV, получаемых из сверхэкспрессирующих GNA1870 штаммов, наблюдают сниженную экспрессию множества других белков клеточной оболочки в сравнении с OMV, получаемой из вакцины штамма RM1090 дикого типа или нокаутного штамма RM1090ΔGNA1870. Хотя способность антисыворотки от мышей, иммунизированных посредством OMV со сверхэкспрессией GNA1870, вызывать образование бактерицидного антитела в отношении штамма Cu385 или активировать отложение C3b на штамме NZ98/294 возникала в результате образования антител, вызванного посредством GNA1870, снижение в отношении других таких белков наружной клеточной оболочки может служить дополнительному усилению иммуногенности и защитной иммунной реакции, вызываемой везикулами, которые получают из сверхэкспрессирующих GNA1870 штаммов (например, вследствие "раскрытия" других антигенов в везикуле).
Представленные в настоящей заявке примеры иллюстрируют широту защиты, обусловленной иммунизацией с использованием вакцины OMV, которую получают из штамма N. meningitidis, который сверхэкспрессирует (например, генетически сконструирован для сверхэкспрессии) GNA1870. Функциональная активность антител против GNA1870, образование которых вызвано вакциной OMV со сверхэкспрессией GNA1870, была более высокой, чем активность антител, образование которых вызвано вакциной рекомбинантного GNA1870 или сочетанием рекомбинантного GNA1870 и OMV, получаемой из штамма дикого типа. Например, несмотря на более низкую величину иммунного ответа против GNA1870, как измерено посредством ELISA (таблица 2), для сыворотки от мышей, иммунизированных вакциной OMV, которую получают из сконструированных для сверхэкспрессии GNA1870 штаммов, наблюдают более высокую бактерицидную активность в отношении штамма Z1092, чем активность сыворотки от мышей, иммунизированных вакциной рекомбинантного белка GNA1870 или вакцинами OMV, которые получают из штаммов RM1090 дикого типа или нокаутных по GNA1870, или вакциной OMV в смеси с вакциной рекомбинантного белка GNA1870 (фиг.3).
Кроме того, даже в отсутствие значительной бактерицидной активности антитела, образование которых вызвано вакциной OMV со сверхэкспрессией GNA1870, вызывали большее отложение C3b на поверхности штаммов NZ98/254 или M1390 (фиг.4A, столбец 6), чем антитела, вырабатываемые в ответ на другие вакцины, а, кроме того, указанные первыми антитела сообщают более значительную пассивную защиту против бактериемии у детенышей крыс, у которых иммунную реакцию вызывали посредством штамма NZ98/254 (фиг.6, панели A-B). После абсорбции антител против GNA1870 способность активировать отложение C3b на штамме NZ98/254 была утрачена (таблица 3). Вкратце, модифицированная вакцина OMV сообщала защитную активность более широкого спектра, чем рекомбинантный белок GNA1870 или вакцина OMV из используемого для вакцины штамма дикого типа, что является результатом способности модифицированной вакцины OMV вызывать как специфичную к сероподтипу бактерицидную активность против штаммов, экспрессирующих молекулу PorA, гомологичную молекуле из штамма для вакцины, так и вызывать образование антител против GNA1870 с более высокой функциональной активностью в отношении штаммов, экспрессирующих субварианты варианта 1 белка GNA1870, чем вызываемая вакциной рекомбинантного GNA1870.
Модифицированная вакцина OMV, полученная из сверхэкспрессирующего GNA1870 штамма, обладала преимуществом над рекомбинантным GNA1870 в отношении штаммов, которые экспрессируют субварианты варианта 1 белка GNA1870 и/или экспрессируют PorA гомологичного сероподтипа. Интересно, что мыши, иммунизированные вакциной на основе везикул, которую получают из сконструированного для сверхэкспрессии NspA штамма N. meningitidis (RM1090), имели при ELISA более чем в 10 раз большие титры антитела против NspA, но меньшие титры бактерицидной сыворотки против некоторых штаммов N. meningitidis, таких как Cu385 или Z1092, в сравнении с контрольными мышами, которых иммунизировали контрольной вакциной на основе везикул, получаемой из штамма RM1090, в котором был инактивирован ген, кодирующий NspA (таблица 5). O'Dwyer et al. также наблюдали отсутствие бактерицидной активности сыворотки у мышей, вакцинированных с использованием вакцины OMV, которую получают из штамма N. flavescens, сконструированного для сверхэкспрессии NspA (Infect. Immun. 2004; 72:6511-80). Таким образом, настоящие открытия, в которых показаны усиленные реакции бактерицидных и защитных антител в ответ на вакцину OMV со сверхэкспрессией GNA1870, являются неожиданными.
Сверхэкспрессия GNA1870 v.1 в штамме H44/76 приводит к ~3-кратному повышению количества GNA1870 в OMV в сравнении с более высокими природными количествами GNA1870 в OMV, получаемой из штамма H44176 дикого типа. В отличие от проведенного авторами изобретения предыдущего исследования мышей, иммунизированных OMV из штамма RM1090 дикого типа, у мышей, которые иммунизированы посредством OMV, получаемой из H44/76 дикого типа, развивались иммунные ответы против GNA1870, как определено посредством ELISA (фиг.5). Однако группа мышей, которым вводили OMV из штамма со сверхэкспрессией GNA1870, имела ~10-кратно большие титры. Титры, измеряемые посредством ELISA, недостаточно хорошо соответствуют функциональной активности антител. Например, наивысшие титры сыворотки против GNA1870 были у мышей, иммунизированных вакциной рекомбинантного GNA1870, однако бактерицидная активность и активность в отношении отложения C3b сыворотки от мышей, иммунизированных рекомбинантным белком, была ограниченной в отношении штамма H44/76. Восприимчивость этого штамма была ожидаемой, поскольку фактически все штаммы N. meningitidis, генетически происходящие от ET 5, обладают высокой экспрессией канонического белка GNA1870 v.1 (с аминокислотной последовательностью, аналогичной таковой для MC58), и эти штаммы высокочувствительны к опосредуемой комплементом бактерицидной активности антител против GNA1870 (Masignani et al. 2003, выше; Welsch et al. 2004, выше). Остальные пять тестируемых штаммов N. meningitidis в исследовании авторов изобретения экспрессировали более низкие количества GNA1870, чем штамм H44/76, а соответствующие белки представляли собой субварианты GNA1870 v.1. Также пять штаммов имели молекулы PorA, гетерологичные в отношении таких молекул из штамма для вакцины H44/76. Эти пять штаммов были устойчивы к бактерицидной активности и активации комплемента антителами, образование которых вызвано вакциной рекомбинантного GNA1870, или антителами против PorA, образование которых вызвано вакцинами OMV. В отличие от этого, четыре из пяти штаммов были чувствительны к бактерицидной активности и/или активности в отношении отложения комплемента сыворотки от мышей, иммунизированных вакциной OMV H44/76 со сверхэкспрессией GNA1870. Активация C3b на поверхности живых бактерий приводила к ожидаемой пассивной защите детенышей крыс против менингококковой бактериемии (Welsch et al. J. Infect. Dis. 2003; 188:1730-40; Welsch et al., J. Immunol. 2004; 172:5606-15; Hou et al. J Infect. Dis. 2005; 192:580-90; Moe et al. Infect. Immun. 2002; 70:6021-31). Вакцины OMV со сверхэкспрессируемым GNA1870 состоят из сложной смеси антигенов и от них ожидают вызова образования антител против множества антигенных мишеней. Однако в экспериментах по абсорбции функциональная активность антител против этих штаммов была направлена против GNA1870 (таблица 3).
Удивительно, что вакцина OMV, полученная из мутантного штамма только с незначительным повышением уровня GNA1870, вызывала более высокие и более широкого спектра реакции специфичных к GNA1870 бактерицидных антител и/или более высокое отложение C3, чем вакцина OMV, получаемая из штамма дикого типа, который отбирают на предмет обладания относительно высокой экспрессией GNA1870. Таким образом, даже незначительное изменение соотношения GNA1870 и общего белка в препарате вакцины OMV, вероятно, определяет наличие или отсутствие иммунного ответа на GNA1870. Кроме того, качество антител, образование которых вызвано вакциной OMV со сверхэкспрессируемым GNA1870, превосходит качество антител, образование которых вызвано вакциной рекомбинантного GNA1870. Например, рекомбинантная вакцина вызывала в ELISA более высокие титры связывания антител, чем титры, которые вызваны вакциной OMV со сверхэкспрессируемым GNA1870, однако антитела против рекомбинантного белка имели более низкую бактерицидную активность и активность в отношении активации комплемента. Определение механизмов, посредством которых модифицированная вакцина GNA1870-OMV вызывает образование антител сыворотки с более широкой функциональной активностью, чем рекомбинантный белок или контрольная вакцина OMV, требует дальнейшего исследования.
Таким образом, настоящее изобретение относится к способам и композициям для вызова иммунной реакции, широко реагирующей на различные вызывающие заболевание штаммы N. meningitidis. Изобретение решает проблему иммунодоминирования различающихся антигенами доменов PorA в вакцинах, которые основаны на везикулах или PorA, посредством усиления реакции антител на GNA1870 и, возможно, на другие распространенные антигены в штаммах для вакцины. Важно, что способы по изобретению приводят к образованию бактерицидного антитела сыворотки, единственного подтвержденного серологического аналога защите у человека (Goldschneider et al. 1969, выше), против штаммов Neisseria, экспрессирующих эпитопы сероподтипов, которые не использовали в препаратах вакцин. Кроме того, способ приводит к образованию бактерицидного антитела сыворотки против штаммов, которые не были унечтожены антителом против консервативного белка, такого как поверхностный белок A Neisserial, кандидата для менингококковой вакцины (Martin et al., 2000. J. Biotechnol. 83:27-31; Moe et al. (1999 Infect. Immun. 67:5664; Moe et al. Infect. Immun. 2001 69:3762). Не настаивая на теории, вакцина и схема иммунизации по изобретению обеспечивают его неожиданные преимущества в отношении защитного иммунитета широкого спектра посредством вызова образования антител, специфичных как к консервативным, так и неконсервативным антигенам.
Определения
Термин "защитный иммунитет" обозначает то, что вакцина или схема иммунизации, которую используют для млекопитающего, индуцирует иммунную реакцию, предотвращающую, задерживающую развитие или снижающую тяжесть заболевания, которое вызвано Neisseria meningitidis, или снижающую или, в целом, устраняющую симптомы заболевания.
Выражение "заболевание, вызываемое штаммом Neisseria meningitidis серогруппы B" включает любой клинический симптом или сочетание симптомов, присутствующее при заражении членом Neisseria meningitidis серогруппы B. Эти симптомы включают, но ими не ограничиваются: заселение верхних дыхательных путей (например, слизистой оболочки носоглотки и миндалин) патогенным штаммом Neisseria meningitidis серогруппы B, проникновение бактерий в слизистую оболочку и подслизистое сосудистое русло, септицемию, септический шок, воспаление, геморрагические повреждения кожи, активацию фибринолиза и свертывания крови, дисфункцию органов, таких как почки, легкие, а также сердечную недостаточность, кровоизлияние в надпочечниках и мышечный инфаркт, повышенную проницаемость капилляров, отек, периферическую ишемию конечностей, респираторный дистресс-синдром, перикардит и менингит.
Выражение "защитный иммунитет широкого спектра действия" обозначает то, что вакцина или схема иммунизации вызывают "защитный иммунитет" против по крайней мере одного или нескольких (или против по крайней мере двух, по крайней мере трех, по крайней мере четырех, по крайней мере пяти, по крайней мере восьми или по крайней мере более восьми) штаммов Neisseria meningitidis, где каждый из штаммов относится к различному сероподтипу как штаммы, используемые для получения вакцины. Изобретение особо предусматривает и включает вакцину или схему вакцинации, придающую защиту против заболевания, которое вызвано членом Neisseria meningitidis серогруппы B, а также против других серогрупп, особенно серогрупп A, C, Y и W-135.
Выражение "специфически связывается с антителом" или "специфически иммунореактивно в отношении к" при ссылке на антиген, такой как полисахарид, фосфолипид, белок или пептид, относится к реакции связывания, основанной на и/или служащей для подтверждения наличия антигена в образце, который также может содержать гетерогенную популяцию других молекул. Таким образом, при указанных условиях иммунологического анализа конкретное антитело или антитела связываются(ется) с конкретным антигеном или антигенами в образце и в значительной степени не связываются с другими присутствующими в образце молекулами. Для специфичного связывания с антителом при таких условиях может быть необходимо антитело или антисыворотка, отобранные на предмет их специфичности к конкретному антигену или антигенам.
Выражение "в достаточном количестве для вызова иммунной реакции на эпитопы, присутствующие в указанном препарате" обозначает то, что существует поддающееся регистрации различие между показателем иммунной реакции, измеряемым до и после введения конкретного антигенного препарата. Показатели иммунной реакции включают, но ими не ограничиваются: титр или специфичность антител, как определяют посредством анализа, такого как твердофазный иммуноферментный анализ (ELISA), анализ на предмет бактерицидности, проточная цитометрия, иммунопреципитация, иммунодиффузия по Оухтерлони; анализы для регистрации связывания, например, точечные, Вестерн-блоттинг или матрицы антигенов; анализы цитотоксичности и т.д.
"Поверхностный антиген" представляет собой антиген, присутствующий в поверхностной структуре Neisseria meningitidis (например, наружной мембране, внутренней мембране, периплазматическом пространстве, капсуле, пилюлях и т.д.).
Выражение "генетически различные", как использовано в контексте генетически различных штаммов Neisseria meningitidis, относится к штаммам, которые отличаются друг от друга аминокислотной последовательностью по крайней мере одного, а обычно по крайней мере двух, более обычно - по крайней мере трех полипептидов, особенно антигенных полипептидов. Генетическое разнообразие штаммов можно достигать селекцией штаммов, отличающихся по крайней мере по одной или нескольким, предпочтительно - по крайней мере двум или более из серогруппы, серотипах или сероподтипах (например, два штамма, отличающихся по крайней мере одним из белков, выбранных из наружной мембраны, белков PorA и PorB, называют генетически отличными друг от друга). Также генетическое отличие можно определять посредством типирования множества участков последовательности и/или типирования множества участков фермента (см., например, Maiden et al., 1998, Proc. Natl. Acad. Sci USA 95:3140; Pizza et al. 2000 Science 287:1816), электрофорезом множества участков фермента и другими известными в данной области способами.
В рамках настоящей заявки "серогруппа" относится к классификации Neisseria meningitidis по признаку иммунологически выявляемых различий в полисахариде капсулы. Известны приблизительно 12 серогрупп: A, B, C, X, Y, Z, 29-E, W-135, H, I, K и L. Любая серогруппа может включать множество серотипов и множество сероподтипов.
В рамках настоящей заявки "серотип" относится к классификации штаммов Neisseria meningitidis на основе определяемых посредством моноклональных антител антигенных различий в белке наружной мембраны порине B. Во множестве серогрупп и множестве сероподтипов можно наблюдать один серотип.
В рамках настоящей заявки "сероподтип" относится к классификации штаммов Neisseria meningitidis на основе определяемых посредством антител антигенных различий в белке наружной мембраны, который называют порин A, или посредством типирования аминокислотных последовательностей VR, установленных в результате секвенирования ДНК (Sacchi et al., 2000, J. Infect. Dis. 182:1169; см. также веб-сайт Multi Locus Sequence Typing). Наибольшая вариабельность белков PorA возникает в двух (петли I и IV) из восьми предполагаемых петель, представленных на поверхности. Вариабельные петли I и IV были обозначены VR1 и VR2 соответственно. Во множестве серогрупп и множестве серотипов можно наблюдать один сероподтип.
"Обогащенный" обозначает то, что антиген в антигенной композиции обработан экспериментатором или клиницистом таким образом, что он присутствует по крайней мере в трехкратно большей концентрации от общей массы, обычно по крайней мере в 5-кратно большей концентрации, более предпочтительно по крайней мере 10-кратно большей концентрации, более обычно по крайней мере 100-кратно большей концентрации, чем концентрация этого антигена в штамме, из которого получали антигенную композицию. Таким образом, если концентрация конкретного антигена составляет 1 микрограмм на грамм общего бактериального препарата (или общего бактериального белка), то обогащенный препарат содержит по крайней мере 3 микрограмма на грамм общего бактериального препарата (или общего бактериального белка).
Термин "гетерологичный" относится к двум биологическим компонентам, которые в природе вместе не наблюдаются. Компоненты могут представлять собой клетки-хозяева, гены или регуляторные участки, такие как промоторы. Хотя гетерологичные компоненты в природе вместе не наблюдаются, они могут функционировать вместе, например, когда гетерологичный в отношении гена промотор функционально связан с геном. Другим примером является случай, когда последовательность Neisserial гетерологична организму-хозяину Neisserial из другого штамма. "Гетерологичный", как использовано в контексте белков, экспрессируемых двумя различными бактериальными штаммами, например, "гетерологичный PorA" или "гетерологичный GNA1870", указывает на то, что рассматриваемые белки отличаются аминокислотной последовательностью. Например, первый штамм Neisserial, экспрессирующий PorA 1,5-2,10, и второй штамм Neisserial, экспрессирующий PorA 7-2,4, называют имеющими "гетерологичные белки PorA" или "гетерологичными в отношении PorA".
Термин "иммунологически наивный в отношении Neisseria meningitidis" обозначает индивидуума (например, млекопитающее, такое как пациент человек), который никогда не подвергался воздействию (посредством инфицирования или введения) Neisseria meningitidis или антигенной композиции, получаемой из Neisseria meningitidis, в количествах, достаточных для вызова защитного иммунитета, или, если подвергался, то оказался неспособным проявить защитную иммунную реакцию. (Примером последнего является индивидуум, подвергающийся воздействию в слишком раннем возрасте, когда защитные иммунные реакции могут не возникнуть. Molages et al., 1994, Infect. Immun. 62:4419-4424). Кроме того, желательно (но не обязательно), чтобы "иммунологически наивный" индивидуум также не подвергался воздействию вида Neisserial, отличного от Neisseria meningitidis (или антигенной композиции, получаемой из вида Neisserial), в особенности не подвергался воздействию перекрестно реагирующего штамма из вида Neisserial (или антигенной композиции). Индивидуумы, которые подверглись воздействию (посредством инфицирования или введения) вида Neisserial или антигенной композиции, получаемой из этого вида Neisserial, в количествах, достаточных для вызова иммунной реакции на проявляемые у этого вида эпитопы, являются "примированными" в отношении иммунологического ответа на проявляемые у этого вида эпитопы.
Экспрессирующие GNA1870 штаммы Neisserial для использования в получении везикул
В целом, изобретение относится к получению везикул (микровезикул или везикул наружной мембраны) из природного или генетически модифицированного штамма Neisserial, продуцирующего белок GNA1870 на уровне, достаточном для обеспечения везикул, которые при введении индивидууму вызывают образование антител сыворотки против GNA1870. Продуцируемые антитела против GNA1870 способствуют иммунной защите против 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более штаммов Neisserial, где эти штаммы могут быть генетически различными (или "гетерологичными") в отношении, например, серогруппы, серотипа, сероподтипа (например, как определено посредством белка PorA), типа последовательности, электрофоретического типа, варианта GNA1870 и/или субварианта GNA1870.
Любое множество штаммов видов Neisseria, которые продуцируют или могут быть модифицированы для продукции GNA1870, и, необязательно, которые продуцируют или могут быть модифицированы для продукции другого интересующего антигена, такого как PorA, можно использовать в способах по изобретению. Характеристики приемлемых штаммов в отношении продукции GNA1870 более подробно описаны ниже.
Особый интерес представляют собой патогенные виды Neisseria или штаммы, получаемые из патогенных видов Neisseria, особенно штаммы, патогенные для человека или получаемые из штаммов, которые патогенны или являются комменсалами для человека. Примеры видов Nessserial включают N. meningitidis, N. flavescens, N. gonorrhoeae, N. lactamica, N. polysaccharea, N. cinerea, N. mucosa, N. subflava, N. sicca, N. elongata и т.п. "Получаемые из" в контексте бактериальных штаммов означают указание на то, что штамм был получен посредством пересева in vivo или in vitro культуры родительского штамма и/или представляет собой рекомбинантную клетку, получаемую модификацией родительского штамма.
Особый интерес по настоящему изобретению представляют собой штаммы N. meningitidis. Штаммы N. meningitidis можно разделить на серологические группы, серотипы и подтипы на основе реакций с поликлональными (Frasch, C. E. and Chapman, 1973, J. Infect. Dis. 127:149-154) или моноклональными антителами, взаимодействующими с различными поверхностными антигенами. Разделение на серогруппы основано на иммунологически регистрируемых различиях в полисахариде капсулы. Известно приблизительно 12 серогрупп (A, B, C, X, Y, Z, 29-E и W-135). Штаммы серогрупп A, B, C, Y и W-135 ответственны практически за каждое менингококковое заболевание.
Серотипирование основано на определяемых посредством моноклональных антител антигенных различиях в белке наружной мембраны, называемом порин B (PorB). В настоящее время известны антитела, определяющие приблизительно 21 серотип (Sacchi et al, 1998, Clin. Diag. Lab. Immunol. 5:348). Сероподтипирование основано на определяемых посредством антител антигенных различиях в белке наружной мембраны, называемом порин A (PorA). В настоящее время известны антитела, определяющие приблизительно 18 сероподтипов. Типирование сероподтипов особенно важно у штаммов Neisseria meningitidis, когда иммунитет может быть специфичен к сероподтипу. Наибольшая вариабельность белков PorA возникает в двух (петли I и IV) из восьми предполагаемых петель, представленных на поверхности. Вариабельные петли I и IV были обозначены VR1 и VR2 соответственно. Поскольку существует больше вариантов последовательностей VR1 и VR2 PorA, которые не были определены специфическими антителами, то была предложена альтернативная номенклатура на основе типирования аминокислотной последовательности VR, установленной из секвенирования ДНК (Sacchi et al., 2000, J Infect. Dis. 182:1169; см. также веб-сайт Multi Locus Sequence Typing). Также можно использовать липополисахариды в качестве антигенов для типирования, приводящих к возникновению так называемых иммунотипов: L1, L2 и т.д.
Также N. meningitidis можно подразделять на группы или подгруппы клонов с использованием различных способов, прямо или косвенно характеризующих бактериальный геном. Эти способы включают электрофорез множества участков фермента (MLEE), основанный на различии электрофоретической подвижности фермента, которое отражает лежащие в основе этого полиморфизмы в конкретном участке гена. Посредством характеризации вариантов множества таких белков можно из соотношения несоответствий сделать вывод о генетическом "расстоянии" между двумя штаммами. Сходным образом, можно сделать заключение о клональности между двумя изолятами, если два изолята имеют одинаковые наборы электрофоретических вариантов во множестве участков. Позднее MLEE было заменено типированием множества участков последовательности (MLST) в качестве предпочтительного способа характеризации микроорганизмов. С использованием MLST генетическое расстояние между двумя изолятами или клональность, устанавливают из соотношения несоответствий в последовательностях ДНК 11 генов "домашнего хозяйства" в штаммах Neisseria meningitidis (Maiden et al., 1998, Proc. Natl Acad. Sci. USA 95:3140).
Используемый для получения везикул штамм можно отбирать в соответствии с множеством различных характеристик, которые могут быть желательны. Например, в дополнение к селекции в соответствии с уровнем продукции GNA1870 штамм можно отбирать в соответствии с: желаемым типом PorA ("сероподтипом", как указано выше), серогруппой, серотипом и т.п.; сниженной продукцией полисахарида капсулы; и т.п.
Например, штамм для продукции может продуцировать любой желаемый полипептид PorA и может экспрессировать один или несколько полипептидов PorA (природно или вследствие генетического конструирования). Примеры штаммов включают штаммы, которые продуцируют полипептид PorA, придающий сероподтип P1.7,16; P1.19,15; P1.7,1; P1.5,2; P1.22a,14; P1.14; P1.5,10; P1.7,4; P1.12,13; а также варианты таких полипептидов PorA, которые могут сохранять или не сохранять реакционную способность в отношении общепринятых серологических реагентов, используемых в типировании сероподтипов.
Также интересны полипептиды PorA, охарактеризованные в соответствии с типированием вариабельной области (VR) PorA (см., например, Russell et al. Emerging Infect. Dis. 2004 10:674-678; Sacchi CT, et al, Clin. Diagn. Lab. Immunol. 1998; 5:845-55; Sacchi et al, J. Infect. Dis. 2000; 182:1169-1176). Было выявлено значительное число различных типов VR, которые можно классифицировать на "прототипы" семейств VR1 и VR2. Доступная во всемирной сети база данных, в которой описана эта номенклатура и ее связь с предыдущими схемами типирования, находится на neisseria.org/nm/typing/pora. Выравнивания типичных типов VR1 и VR2 PorA представлены в Russell et al. Emerging Infect. Dis. 2004 10:674-678, а также представлены на фиг.9 для удобства читателя.
Типичные полипептиды PorA, как охарактеризовано посредством сероподтипов PorA, включают P1.5,2; P1.5a,2a; P1.5a,2c; P1.5a,2c; P1.5a,2c; P1.5b,10; P1.5b,10; P1.5b,C; P1.7,16; P1.7d,1; P1.7d,1; P1.7d,1; P1.7d,1; P1.7b,3; P1.7b,4; P1.7b,4; P1.12,16; P1.12a,13a; P1.22,9; P1.23,14; P1.23,14; P1.19,15; P1.B,1; P1.C,1; P1.E,A; P1.E,A; P1.E,A; P1.5,2; P1.5,2; P1.5a,10a; P1.5b,10; P1.5b,10; P1.5b,10b; P1.7,16; P1.7,16; P1.7b,1; P1.7b,13e; P1.7b,4; P1.7b,4; P1.7d,1; P1.7d,1; P1.7b,13a; P1.23,3; P1.23,3; P1.23,3; P1.19,15; P1.19,1; P1.19,15; P1.19,15; P1.19,15; P1.19,15; P1.19,15; P1.19,15; P1.19,15; P1.E,A; P1.E,A; P1.E,16a; P1.E,4a; P1.E,4a; P1.Ea,3; P1.Eb,9; P1.Eb,9; P1.Eb,9; P1.Eb,9; P1.Eb,9; P1.F,16; P1.7a,1; P1.7b,3; P1.7d,1; P1.Ea,3; P1.5b,10; P1.5b,10; P1.5b,10; P1.5b,10; P1.5b,10; P1.5b,10; P1.5b,10b; P1.5b,10; P1.22,14a; P1.F,16; P1.D,2d; P1.D,2; P1.D,2d; P1.19c,2c; P1.D,10f; P1.A,10e; P1.A,10g; P1.A,10; P1.19,15; P1.19,15; P1.19,15; P1.19,15; P1.7b,16; P1.7,16b; P1.7,16; P1.19,15; P1.Eb,9; P1.5,2e; P1.E,A; P1.7b,13d; P1.Ea,3; P1.7,16b; P1.Ec,1; P1.7b,4; P1.7b,4; P1.7,9; P1.19,15; P1.19,15; P1.19,15; P1.19,15a; P1.19a,15b; P1.19,15; P1.5b,16; P1.19b,13a; P1.5,16; P1.5,2; P1.5,2b; P1.7b,16; P1.7,16b; P1.7b,3; P1.Ea,3; P1.5a,2c; P1.F,16; P1.5a,9; P1.7c,10c; P1.7b,13a; P1.7,13a; P1.7a,10; P1.20,9; P1.22,B; P1.5b,de1; P1.5b,10; P1.7,13a; P1.12a,13f; P1.12a,13; P1.12a,13a; P1.12a,13a; P1.12a,13; P1.12a,13; P1.E,13b; P1.7b,13a; P1.7b,13; P1.5,2; P1.5,2; P1.Ea,3; P1.22,9; P1.5,2; P1.5,2; P1.19,15; P1.5,2; P1.12b,13a; P1.5c,10a; P1.7e,16e; P1.B,16d; P1.F,16e; P1.F,16e; P1.7b,13e; P1.B,16d; P1.7e,16e; P1.7b,13g; P1.B,16f; P1.7,16c; P1.22,14b; P1.22,14c; P1.7,14; P1.7,14; и P1.23,14.
Аминокислотные последовательности типичных полипептидов PorA имеют уникальные идентификаторы базы данных Gen Bank X57182, X57180, U92941, U92944, U92927, U92931, U92917, U92922, X52995, X57184, U92938, U92920, U92921, U92929, U92925, U92916, X57178, AF051542, X57181, U92919, U92926, X57177, X57179, U92947, U92928, U92915, X57183, U92943, U92942, U92939, U92918, U92946, U92496, U97260, U97259, AF042541, U92923, AF051539, AF051538, U92934, AF029088, U92933, U97263, U97261, U97262, U92945, AF042540, U92935, U92936, U92924, AF029086, AF020983, U94958, U97258, U92940, AF029084, U92930, U94959, U92948, AF016863, AF029089, U92937, AF029087, U92932, AF029090, AF029085, AF051540, AF051536, AF052743, AF054269, U92495, U92497, U92498, U92499, U92500, U92501, U92502, U92503, AF051541, X12899, Z48493, Z48489, Z48485, Z48494, Z48487, Z48488, Z48495, Z48490, Z48486, Z48491, Z48492, X66478, X66479, X66477, X66480, X81110, X79056, X78467, X81111, X78802, Z14281/82, Z14273/74, Z14275/76, Z14261/62, Z14265/66, Z14277/78, Z14283/84, Z14271/72, Z14269/70, Z14263/64, Z14259/60, Z14257/58, Z14293/94, Z14291/92, Z14279/80, Z14289/90, Z14287/88, Z14267/68, Z14285/86, L02929, X77423, X77424, X77433, X77426, X77428, X77430, X77427, X77429, X77425, X77432, X77431, X77422, Z48024/25, Z48032/33, Z48020/21, Z48022/23, Z48028/29, Z48016/17, Z48012/13, Z48014/15, Z48018/19, Z48026/27, U31060, U31061, U31062, U31063, U31064, U31065, U31066, U31067, U93898, U93899, U93900, U93901, U93902, U93903, U93904, U93905, U93906, U93907 и U93908.
Альтернативно или дополнительно, продуцирующий штамм может представлять собой штамм, дефицитный по капсуле. Из штаммов, дефицитных по капсуле можно получать вакцины на основе везикул, обеспечивающие сниженный риск вызова значительной реакции аутоантител у индивидуума, которому вводят вакцину (например, вследствие продукции антител, перекрестно реагирующих с сиаловой кислотой на поверхностях клеток-хозяев).
В рамках настоящей заявки "дефектный по капсуле" или "дефектный по полисахариду капсулы" относится к уровню полисахарида капсулы на бактериальной поверхности, который ниже, чем уровень у природного штамма, или, когда штамм генетически модифицирован, ниже, чем уровень у родительского штамма, из которого получают дефектный по капсуле штамм. Дефектный по капсуле штамм включает штаммы, у которых снижена продукция поверхностного полисахарида капсулы по крайней мере на 10%, 20%, 25%, 30%, 40%, 50%, 60%, 75%, 80%, 85%, 90% или более, а также включает штаммы, в которых полисахарид не поддается выявлению на бактериальной поверхности (например, посредством ELISA с целыми клетками при использовании антитела против полисахарида капсулы).
Дефектные по капсуле штаммы включают штаммы, которые дефектны по капсуле вследствие природной или рекомбинантно полученной генетической модификации. Природные дефектные по капсуле штаммы (см., например, Dolan-Livengood et al. J. Infect. Dis. (2003) 187(10):1616-28), а также способы выявления и/или получения дефектных по капсуле штаммов (см., например, Fisseha et al. (2005) Infect. Immun. 73(7):4070-4080; Stephens et al. (1991) Infect. Immun. 59(11):4097-102; Frosch et al. (1990) Mol. Microbiol. 1990 4(7):1215-1218) известны в данной области.
Модификация клетки-хозяина Neisserial для обеспечения сниженной продукции полисахарида капсулы может включать модификацию одного или нескольких генов, вовлеченных в синтез капсулы, где модификация обеспечивает, например, сниженные уровни капсульного полисахарида в сравнении с родительской клеткой до модификации. Такие генетические модификации могут включать изменения в нуклеотидных и/или аминокислотных последовательностях в одном или нескольких генах биосинтеза капсулы, придавая штамму дефектность по капсуле (например, вследствие одной или нескольких вставок, делеций, замен и т.п. в одном или нескольких генах биосинтеза капсулы). У дефектных по капсуле штаммов могут отсутствовать или являться нефункциональными один или несколько генов капсулы.
Особый интерес представляют штаммы, дефектные по биосинтезу сиаловой кислоты. Такие штаммы способны обеспечивать продукцию везикул, обладающих сниженным риском вызова образования антител против сиаловой кислоты, которые перекрестно реагируют с антигенами сиаловой кислоты человека, а также способны дополнительно обеспечивать повышенную безопасность получения. Штаммы с дефектом в биосинтезе сиаловой кислоты (вследствие природной модификации или конструируемой модификации) могут быть дефектны по любому количеству различных генов в каскаде биосинтеза сиаловой кислоты. Особый интерес представляют штаммы, которые дефектны по продукту гена, кодируемому геном N-ацетилглюкозамин-6-фосфат-2-эпимеразы (известным как synX AAF40537.1 или siaA AAA20475), где особый интерес представляют штаммы, у которых этот ген инактивирован. Например, в одном из вариантов осуществления дефектный по капсуле штамм получают прекращением продукции функционального продукта гена synX (см., например, Swartley et al. (1994) J Bacteriol. 176(5):1530-4).
Также дефектные по капсуле штаммы можно получать из природных штаммов с использованием нерекомбинантных способов, например, с использованием бактерицидных антител против капсулы для отбора штаммов со сниженным количеством капсульного полисахарида.
Если по изобретению предусмотрено использование двух или более штаммов (например, для получения антигенных композиций везикул из различных штаммов, как более подробно описано ниже), то штаммы можно отбирать таким образом, чтобы они отличались по одной или нескольким характеристикам штамма, например, для обеспечения везикул, отличающихся типом PorA и/или группой вариантов GNA1870.
Продукция GNA1870 в клетках-хозяевах Neisserial
В целом, как указано выше, везикулы можно получать по изобретению с использованием природного или модифицированного, не встречающегося в природе штамма Neisserial, который продуцирует везикулы с достаточным количеством белка GNA1870 которые при введении индивидууму обеспечивают продукцию антител против GNA1870.
В одном из вариантов осуществления штамм Neisserial, используемый для получения везикул по изобретению, может представлять собой природные штаммы, экспрессирующие GNA1870 на более высоком уровне в сравнении со штаммами, которые не экспрессируют GNA1870 на поддающемся выявлению уровне или экспрессируют на низком уровне. RM1090 представляет собой пример штамма, продуцирующего GNA1870 на низком уровне. Особый интерес представляют природные штаммы, которые продуцируют GNA1870 на уровне, превышающем в 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 5,5, 6, 6,5, 7, 7,5, 8, 8,5, 9, 9,5 или 10 раз или более продукцию GNA1870 в штамме с низкой продукцией GNA1870, таком как RM1090. Примеры природных штаммов, экспрессирующих GNA1870 на высоком уровне, включают штаммы ET-5, такие как H44/76, Cu385 и MC58. Для описания штаммов, экспрессирующих GNA1870 на низких или не поддающихся выявлению уровнях, промежуточных уровнях GNA1870 или высоких уровнях GNA1870, см. Masignani et al. 2003, J Exp. Med. 197:789-199. В конкретных вариантах осуществления штамм продуцирует GNA1870 на уровне, который превышает уровень продукции в RM1090 и который может быть выше по крайней мере в 1,5, 2, 2,5 3, 3,5, 4, 4,5, 5, 5,5, 6, 6,5, 7, 7,5, 8, 8,5, 9, 9,5 или 10 раз или более, чем уровень в RM1090.
В другом варианте осуществления штамм Neisserial модифицирован рекомбинантными или нерекомбинантными способами для обеспечения продукции GNA1870 на достаточно высоком уровне. Как правило, такие модифицированные штаммы получают таким образом, чтобы обеспечивать увеличение продукции GNA1870 в 1,5, 2, 2,5 3, 3,5, 4, 4,5, 5, 5,5, 6, 6,5, 7, 7,5, 8, 8,5, 9, 9,5 или 10 раз или более в сравнении с продукцией GNA1870 в немодифицированной родительской клетке или в сравнении с продукцией GNA1870 штаммом RM1090. В этом варианте осуществления можно использовать любой приемлемый штамм, в том числе штаммы, продуцирующие GNA1870 на низких или не поддающихся выявлению уровнях до модификации, а также штаммы, природно продуцирующие GNA1870 на высоких уровнях в сравнении со штаммами, которые не экспрессируют GNA1870 на поддающемся выявлению уровне или экспрессируют на низком уровне.
Модифицированные штаммы можно получать нерекомбинантными способами, такими как, например, воздействие химическими веществами, излучением или другим средством, модифицирующим или повреждающим ДНК, и т.п. Модифицированные штаммы с желаемым профилем экспрессии белков, особенно в отношении продукции GNA1870, можно выявлять скринингом на предмет штаммов, продуцирующих GNA1870 на желаемом уровне (например, на повышенном уровне GNA1870 в сравнении с немодифицированным родительским штаммом или с продуцирующим GNA1870 на низком уровне (например, RM1090), или на уровне, сходном с уровнем у штамма, продуцирующего GNA1870 на приемлемо высоких уровнях).
Альтернативно и более обычно, модифицированные штаммы получают с использованием рекомбинантных способов, как правило, посредством введения нуклеиновой кислоты, кодирующей полипептид GNA1870, или обработкой эндогенного гена GNA1870 для обеспечения повышенной экспрессией эндогенного GNA1870.
Способы определения уровней продукции GNA1870 известны в данной области. Например, такие способы включают Вестерн-блоттинг (необязательно, посредством анализа с использованием денсиметрического сканирования), анализ проточной цитометрией (например, FACS) с использованием антитела против GNA1870, регистрацию уровней РНК GNA1870 и т.п. В некоторых случаях штаммы с продукцией GNA1870 на более высоких уровнях, природной или вследствие генетической модификации, в настоящей заявке называют "сверхэкспрессирующими" GNA1870 или указывают, что они "сверхэкспрессируют" GNA1870.
Получение генетически модифицированных штаммов Neisserial
Как указано выше, посредством введения нуклеиновой кислоты, кодирующей полипептид GNA1870, или обработкой эндогенного гена GNA1870 для обеспечения повышенной экспрессии эндогенного GNA1870.
Клетки-хозяева Neisserial, генетически модифицированные для обеспечения повышенной экспрессии эндогенного GNA1870
Экспрессию эндогенного GNA1870 можно повышать изменением in situ регуляторного участка, контролирующего экспрессию GNA1870. Способы обеспечения повышенной экспрессии эндогенного гена Neisserial известны в данной области (см., например, WO 02/09746). Кроме того, известны последовательности нуклеиновых кислот генов, кодирующих геномные варианты и субварианты GNA1870, что обеспечивает быстрое приспособление таких способов для повышения экспрессии эндогенного GNA1870.
Эндогенный GNA1870 может представлять собой любую желаемую группу вариантов (например, v.1, v.2, v.3 и т.п.) или субвариант GNA1870. Особый интерес представляет "канонический" полипептид GNA1870 v.1 из штамма MC58. Также интересен субвариант полипептида GNA1870 из штамма NZ98/294 и полипептид GNA1870 v.2 из штамма 2996.
Модификация клетки-хозяина Neisserial для обеспечения повышенной продукции эндогенного GNA1870 может включать частичную или полную замену всего участка эндогенного гена, контролирующего экспрессию GNA1870, где модификация обеспечивает, например, повышенную транскрипционную активность в сравнении с немодифицированным родительским штаммом. Повышенную транскрипционную активность можно получать посредством вариантов (точечные мутации, делеции и/или вставки) эндогенных контролирующих участков, посредством природных или модифицированных гетерологичных промоторов или посредством их сочетания. Как правило, генетическая модификация приводит к более высокой транскрипционной активности, чем немодифицированная эндогенная транскрипционная активность, (например, посредством введения сильного промотора), что приводит к повышенной экспрессии GNA1870.
Типичные сильные промоторы, которые могут быть эффективны в повышении продукции транскрипта GNA1870, могут включать, например, промоторы для porA, porB, lbpB, tbpB, p110, hpuAB, lgtF, Opa, p110, 1st и hpuAB. PorA, RMp и PorB представляют особый интерес, как конститутивные сильные промоторы. За активность промотора PorB ответственен участок, соответствующий нуклеотидам от -1 до -250 в обратном направлении от стартового кодона porB.
В данной области доступны способы для осуществления введения промотора в геном клетки-хозяина таким образом, чтобы функционально связать промотор с нуклеиновой кислотой, кодирующей эндогенный GNA1870. Например, способ гомологичной рекомбинации с двойным кроссинговером для введения промотора в участок, расположенный в обратном направлении от кодирующей последовательности, например, приблизительно 1000 п.о., от приблизительно 30-970 п.о., приблизительно 200-600 п.о., приблизительно 300-500 п.о. или приблизительно 400 п.о. в обратном направлении (5') от стартового кодона ATG последовательности нуклеиновой кислоты, кодирующей GNA1870, для обеспечения повышения активности. Оптимальное расположение промотора можно определять общепринятым использованием способов, доступных в данной области.
Например, высокоактивный промотор (например, промоторы PorA, PorB или Rmp), расположенный в обратном направлении от заданного гена. В качестве примера, промотор PorA можно оптимизировать для экспрессии, как описано van der Ende et al. Infect. Immun. 2000; 68:6685-90. Вставку промотора можно осуществлять, например, амплификацией посредством PCR расположенного выше участка заданного гена GNA1870, клонируя расположенный выше участок в вектор и вставляя соответствующие участки рестрикции в ходе амплификации посредством PCR или используя природные участки рестрикции для вставки промоторного участка PorA. Например, можно клонировать расположенный выше участок гена GNA1870 величиной приблизительно 700 п.о. с использованием природных участков для ферментов рестрикции, расположенных на соответствующем расстоянии (например, приблизительно 400 п.о.), выше промотора GNA1870, в который вставляют клонированный участок промоторного участка PorA. Еще выше по отношению к промотору PorA в участок можно вставлять кассету устойчивости к антибиотику (например, эритромицину), а конструкт использовали для замены расположенного выше участка GNA1870 дикого типа посредством гомологичной рекомбинации.
Другой способ включает введение последовательности, кодирующей полипептид GNA1870, ниже по отношению к эндогенному промотору, проявляющему высокую транскрипционную активность в геноме клетки-хозяина. Например, кодирующий участок гена Rmp можно заменять кодирующей последовательностью для полипептида GNA1870. В этом способе используют преимущество высокоактивного конститутивного промотора Rmp для управления экспрессией.
Клетки-хозяева Neisserial, генетически модифицированные для экспрессии экзогенного GNA1870
Штаммы Neisserial можно генетически модифицировать для сверхэкспрессии GNA1870 посредством введения в клетку-хозяина Neisserial конструкции, кодирующей полипептид GNA1870. В настоящей заявке вводимый для экспрессии GNA1870 называют "экзогенным" GNA1870. Клетка-хозяин продуцирует эндогенный GNA1870, а экзогенный GNA1870 может иметь такую же или отличную аминокислотную последовательность в сравнении с эндогенным GNA1870.
Штамм, используемый в качестве клетки-хозяина в этом варианте осуществления, может продуцировать GNA1870 на любом уровне (например, продукция GNA1870 на высоком уровне, промежуточном уровне или низком уровне). Особый интерес представляет использование штамма, отобранного для продукции GNA1870 на низком или не поддающемся выявлению уровне или модифицированного для обладания продукцией GNA1870 на не поддающемся выявлению или низком уровне. Например, клетку-хозяина можно генетически модифицировать таким образом, чтобы нарушить эндогенный ген GNA1870, вследствие чего GNA1870 не продуцируется или не присутствует в клеточной оболочке (и таким образом не присутствует на поддающихся выявлению уровнях в везикуле, получаемой из такой модифицированной клетки). В других вариантах осуществления клетка-хозяин продуцирует GNA1870 на промежуточном или высоком уровне GNA1870 (например, относительно уровня, на котором GNA1870 продуцируется, например, RM1090).
Полипептиды GNA1870
Клетку-хозяина можно генетически модифицировать для экспрессии любого приемлемого полипептида GNA1870, в том числе вариантов или субвариантов GNA1870. Как более подробно описано ниже, известны аминокислотные последовательности многих полипептидов GNA1870; выравнивание этих последовательностей обеспечивает указание на остатки, которые являются консервативными среди вариантов, обеспечивая таким образом указание на модификации аминокислот (например, замены, вставки, делеции), которые можно осуществить.
Соответственно, в рамках настоящей заявки "полипептид GNA1870" включает природные и синтетические (не встречающиеся в природе) полипептиды, последовательность которых по крайней мере приблизительно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более совпадает на нуклеотидном или аминокислотном уровне с природным полипептидом GNA1870, и которые способны вызывать образование антител, специфически связывающих природный полипептид GNA1870, который присутствует на целой бактериальной клетке Neisserial. Также "полипептид GNA1870" включает слитые белки, например, полипептид GNA1870 с гетерологичным полипептидом на N- и/или C-конце.
Клетку-хозяина можно генетически модифицировать для экспрессии по крайней мере 1 полипептида GNA1870, а также можно модифицировать для экспрессии 2, 3, 4 или более полипептидов GNA1870 в одной и той же клетке-хозяине. Например, одну клетку-хозяина можно генетически модифицировать для экспрессии по крайней мере одного варианта полипептида GNA1870 1, по крайней мере одного варианта полипептида GNA1870 2 и по крайней мере одного варианта полипептида GNA1870 3.
Если при экспрессии множества полипептидов GNA1870 возникает проблема, обусловленная токсичностью в отношении клетки-хозяина, то различные полипептиды GNA1870 можно экспрессировать с различных промоторов для того, чтобы делать возможным существование диапазона уровня экспрессии. Например, изменение как состава оснований, так и количества оснований между участками -10 и -35 промотора PorA должно приводить к значительному диапазону уровня экспрессии желаемого рекомбинантного белка (van der Ende et al. Infect. Immun. 2000; 68:6685-90).
Нуклеиновые кислоты, кодирующие полипептид GNA1870 для использования по изобретению, известны в данной области. Например, приемлемые полипептиды GNA1870 описаны в WO 2004/048404; Masignani et al. 2003 J Exp. Med. 197:789-799; Fletcher et al. Infect. Immun. 2004 2088-2100; Welsch et al. J Immunol. 2004 172:5606-5615; и WO 99/57280. Также нуклеиновая кислота (и аминокислотные последовательности) для вариантов и субвариантов GNA1870 представлены в банке генов под уникальными идентификаторами: NC_003112, идентификатор гена: 904318 (ссылка NCBI NP_274866) (из штамма N. meningitidis MC58); AY548371 (AAT01290.1) (из штамма N. meningitidis CU385); AY548370 (AAT01289.1) (из штамма N. meningitidis H44/76); AY548377 (AAS56920.1) (из штамма N. meningitidis M4105); AY548376 (AAS56919.1) (из штамма N. meningitidis M1390); AY548375 (AAS56918.1) (из штамма N. meningitidis N98/254); AY548374 (AAS56917.1) (из штамма N. meningitidis M6190); AY548373 (AAS56916.1) (из штамма N. meningitidis 4243); и AY548372 (AAS56915.1) (из штамма N. meningitidis BZ83).
На фиг.7 представлено выравнивание типичных аминокислотных последовательностей вариантов 1, 2 и 3 GNA1870 из штаммов N. meningitidis MC58, 951-5945 и M1239 соответственно (WO 2004/048404). Незрелый белок GNA1870 содержит лидерную последовательность из приблизительно 19 остатков, где каждый вариант обычно включает N-концевой цистеин, к которому можно ковалентно присоединять молекулу липида. Как правило, в природном белке этот остаток цистеина липидизирован. "1" указывает на первую аминокислоту зрелого белка, а аминокислоты, являющиеся частью лидерной последовательности, указаны отрицательными числами. Серый и черный фоны указывают консервативные и одинаковые аминокислотные остатки соответственно. Дополнительные аминокислотные последовательности полипептидов GNA1870, в том числе не встречающиеся в природе варианты, представлены на фиг.8A-8H и 9.
GNA1870 может быть липидизированным или нелипидизированным. Как правило, предпочтительно, чтобы GNA1870 был липидизированным для обеспечения размещения полипептида в мембране. Липидизированный GNA1870 можно получать экспрессией полипептида GNA1870, имеющего N-концевой сигнальный пептид для прямой липидизации посредством диацилглицерилтрансферазы с последующим расщеплением сигнальной пептидазой, специфичной к липопротеину (тип II).
Эффективный по изобретению полипептид GNA1870 включает не встречающиеся в природе (искусственные или мутантные) полипептиды, аминокислотная последовательность которых отличается от природного полипептида GNA1870, но которые присутствуют в мембране организма-хозяина Nesserial таким образом, что везикулы, получаемые из организма-хозяина, содержат GNA1870 в форме, обеспечивающей презентирование интересующих эпитопов, предпочтительно бактериального эпитопа, а также обеспечивающей реакцию антитела против GNA1870. В одном из вариантов осуществления полипептид GNA1870 представляет собой вариант 1 (v.1) или вариант 2 (v.2), или вариант 3 (v.3) полипептида GNA1870 с интересующими субвариантами v.1, v.2 и v.3, в том числе субвариантами v.1 (см., например, Welsch et al. J Immunol. 2004 172:5606-5615). В одном из вариантов осуществления полипептид GNA1870 содержит аминокислотную последовательность полипептида GNA1870, которая является наиболее распространенной среди штаммов, эндемичных в отношении подлежащей вакцинированию популяции.
Эффективные по изобретению полипептиды GNA1870 также содержат слитые белки, где слитый белок включает полипептид GNA1870, содержащий участника слияния на своем N-концевом участке или C-концевом участке. Интересующие участники слияния включают, например, глутатион-S-трансферазу (GST), связывающий мальтозу белок (MBP), метку His и т.п., а также лидерные пептиды из других белков, особенно липопротеинов (например, аминокислотную последовательность перед N-концевым цистеином можно заменять другим интересующим лидерным пептидом).
Другие нуклеиновые кислоты, кодирующие полипептид GNA1870, можно выявлять с использованием хорошо известных в данной области способов, где полипептиды GNA1870 можно устанавливать на основе сходства аминокислотных последовательностей с известным полипептидом GNA1870. Как правило, такие полипептиды GNA1870 обладают по крайней мере приблизительно 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или большим сходством последовательностей на нуклеотидном или аминокислотном уровне. Сходство последовательностей можно определять с использованием способов для выравнивания и сравнения последовательностей нуклеиновых кислот или аминокислот, где эти способы хорошо известны в данной области. Для сравнения более длинных последовательностей могут быть необходимы более сложные способы для достижения оптимального выравнивания двух последовательностей. Оптимальное выравнивание последовательностей для выравнивания рамки сравнения можно осуществлять посредством алгоритма локальной гомологии Smith и Waterman (1981) Adv. Appl. Math. 2:482, алгоритма гомологичного выравнивания Needleman и Wunsch (1970) J. Mol. Biol. 48:443, поиска для способа подобия Pearson и Lipman (1988) Proc. Natl. Acad. Sci. (USA) 85:2444, компьютеризированного осуществления этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, версия 7.0, Genetics Computer Group, 575 Science Dr., Madison, WI) или посредством наблюдения, при этом выбирают наилучшее выравнивание (т.е. приводящее к наибольшему проценту сходства последовательностей в пределах рамки сравнения).
Термины "идентичный" или процент "идентичности" в контексте двух или более последовательностей нуклеиновых кислот или полипептидов относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или имеют определенный процент одинаковых аминокислотных остатков или нуклеотидов при сравнении и выравнивании на предмет максимального соответствия, как определяют с использованием одного из следующих алгоритмов сравнения последовательностей или посредством визуального наблюдения. Интересующие полипептиды включают полипептиды по крайней мере с 60%, 70%, 75%, 80%, 85%, 90%, 95% или большей идентичностью нуклеотидов или остатков аминокислот при сравнении и выравнивании на предмет максимального соответствия, как определяют с использованием одного из следующих алгоритмов сравнения последовательностей или посредством визуального наблюдения. Предпочтительно, участок сходства последовательностей существует на протяжении участка, длиной по крайней мере 10, 20, 30, 40, 50, 60, 70, 80 или 100 смежных остатков. В наиболее предпочтительном варианте осуществления идентичности последовательностей определяют сравнением последовательностей на протяжении полной длины кодирующего участка контрольного полипептида.
Как правило, при сравнении последовательностей одна последовательность служит в качестве контрольной последовательности (например, природная последовательность полипептида GNA1870), с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и контрольные последовательности вводят в компьютер, указывают координаты подпоследовательностей, если необходимо, и указывают параметры программы алгоритма для последовательностей. Затем посредством алгоритма сравнения последовательностей рассчитывают процент сходства последовательностей для тестируемой последовательности(ей) в сравнении с контрольной последовательностью, основываясь на указанных параметрах программы.
Оптимальное выравнивание последовательностей для сравнения можно осуществлять, например, посредством алгоритма локальной гомологии Smith & Waterman, Adv. Appl. Math. 2:482 (1981), алгоритма гомологичного выравнивания Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), поиска для способа подобия Pearson & Lipman, Proc. Natl. Acad. Sci. USA 85:2444 (1988), компьютеризированного осуществления этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, Sjftwart Package, Genetics Computer Group, 575 Science Dr., Madison, WI) или посредством визуального наблюдения (в основном, см. Current Protocols in Molecular Biology, F.M. Ausubel et al., eds., Current Protocols, совместная компания Greene Publishing Associates, Inc. и John Wiley & Sons, Inc., (1995 Supplement) (Ausubel)).
Примерами алгоритмов, эффективных для определения процента тождественности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al. (1990) J. Mol. Biol. 215:403-410 и Altschuel et al. (1977) Nucleic Acids Res. 25:3389-3402 соответственно. Программное обеспечение для проведения анализа BLAST находится в открытом доступе при посредстве Национального Центра для Биотехнологической Информации (http://www.ncbi.nlm.nih.gov/). Этот алгоритм включает первоначальное выявление пар последовательностей с высоким сходством (HSP) посредством выявления в рассматриваемой последовательности коротких "слов" с длиной W, которые совпадают или удовлетворяют некоторому пороговому показателю с положительной величиной T при сравнении со "словом" той же длины в последовательности из базы данных. T называют пороговым показателем для соседнего "слова" (Altschul et al, выше).
Эти первоначальные совпадения соседних "слов" служат в качестве затравок для запуска поиска для нахождения более длинных HSP, которые их содержат. Затем совпадения "слов" удлиняют в обоих направлениях вдоль каждой последовательности до тех пор, пока можно повышать совокупную количественную оценку выравнивания. Совокупные количественные оценки для нуклеотидных последовательностей рассчитывают с использованием параметров M (наградные баллы для пары совпадающих остатков; всегда >0) и N (штрафные баллы для несовпадающих остатков; всегда <0). В случае аминокислотных последовательностей для расчета совокупной количественной оценки используют балльную матрицу. Удлинение совпадений "слов" в каждом направлении прекращают, если: совокупная количественная оценка выравнивания снижается на величину X от своего максимального достигнутого значения; совокупная количественная оценка достигает нуля или менее вследствие накопления одного или нескольких отрицательно оцениваемых выравниваний остатков; или при достижении окончания любой из последовательностей. Параметры алгоритма BLAST W, T и X определяют чувствительность и скорость выравнивания. В программе BLASTN (для последовательностей нуклеотидов) в качестве установленных по умолчанию значений используют длину "слова" (W) 11, ожидание (E) 10, M=5, N=-4 и сравнение обоих цепей. Для аминокислотных последовательностей в программе BLASTP в качестве установленных по умолчанию значений используют длину "слова" (W) 3, ожидание (E) 10 и балльную матрицу BLOSUM62 (см. Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1989)).
В дополнение к расчету процента сходства последовательностей алгоритм BLAST также выполняет статистический анализ сходства двух последовательностей (см., например, Karlin & Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787 (1993)). Одним из показателей сходства, предоставляемых алгоритмом BLAST, является наименьшая вероятность сумм (P(N)), обеспечивающая указание вероятности, с которой происходит случайное совпадение двух нуклеотидных или аминокислотных последовательностей. Например, нуклеиновую кислоту считают сходной с контрольной последовательностью, если наименьшая вероятность сумм при сравнении тестируемой нуклеиновой кислоты с контрольной нуклеиновой кислотой составляет приблизительно менее чем 0,1, более предпочтительно - приблизительно менее чем 0,01, а наиболее предпочтительно - приблизительно менее чем 0,001.
Дополнительно указание на то, что две последовательности нуклеиновых кислот или полипептидов обладают сходством последовательностей, состоит в том, что полипептид, кодируемый первой нуклеиновой кислотой, иммунологически перекрестно реагирует с полипептидом, кодируемым второй нуклеиновой кислотой, как описано выше. Таким образом, как правило, полипептид обладает сходством последовательности со вторым полипептидом, например, если два полипептида отличаются только консервативными заменами. Другим указанием на то, что две последовательности нуклеиновых кислот обладают сходством последовательностей, является то, что две молекулы гибридизуются друг с другом в строгих условиях. Отбор конкретного набора условий гибридизации проводят следующими общепринятыми в данной области способами (см., например, Sambrook, et al., Molecular Cloning: A Laboratory Manual, Second Edition, (1989) Cold Spring Harbor, N.Y.). Примером строгих условий гибридизации является гибридизация при 50°C или выше и 0,1×SSC (15 мМ хлорида натрия/1,5 мМ цитрата натрия). Другим примером строгих условий гибридизации является инкубация в течение ночи при 42°C в растворе: 50% формамида, 5×SSC (150 мМ NaCl, 15 мМ тринатрийцитрата), 50 мМ фосфата натрия (pH 7,6), 5 × раствор Денхардта, 10% декстрансульфата и 20 мг/мл денатурированной, фрагментированной ДНК спермы лосося, с последующей промывкой фильтров в 0,1×SSC приблизительно при 65°C. Строгие условия гибридизации представляют собой условия гибридизации, которые по крайней мере настолько же строгие, насколько указанные выше типичные условия, где условия считают по крайней мере настолько же строгими, если они по крайней мере приблизительно на 80% такие же строгие, как правило, по крайней мере приблизительно на 90% такие же строгие, как указанные выше конкретные строгие условия. В данной области известны другие строгие условия гибридизации и их можно также использовать для выявления нуклеиновых кислот в этом конкретном варианте осуществления изобретения.
Предпочтительно, остатки в несовпадающих положениях отличаются консервативными заменами аминокислот. Консервативные замены аминокислот относятся к взаимозаменяемости остатков, имеющих сходные боковые цепи. Например, группа аминокислот с алифатическими боковыми цепями представляет собой глицин, аланин, валин, лейцин и изолейцин; группа аминокислот с алифатически-гидроксильными боковыми цепями представляет собой серин и треонин; группа аминокислот с амидсодержащими боковыми цепями представляет собой аспарагин и глутамин; группа аминокислот с ароматическими боковыми цепями представляет собой фенилаланин, тирозин и триптофан; группа аминокислот с основными боковыми цепями представляет собой лизин, аргинин и гистидин; а группа аминокислот с серосодержащими боковыми цепями представляет собой цистеин и метионин. Предпочтительные группы консервативных замен аминокислот представляют собой: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин и аспарагин-глутамин.
Вектор и способы введения генетического материала в клетки-хозяева Neisserial
В данной области известны способы и композиции, которые можно легко приспособить для обеспечения генетической модификации клетки-хозяина Neisserial для экспрессии экзогенного полипептида GNA1870. Примеры векторов и способов представлены в WO 02/09746 и O'Dwyer et al. Infect/Immun/ 2004; 72:6511-80.
Например, способы переноса генетического материала в организм-хозяин Neisserial включают конъюгацию, трансформацию, электропорацию, способы на основе фосфата кальция и т.п. Способ переноса должен обеспечивать стабильную экспрессию вводимой нуклеиновой кислоты, кодирующей GNA1870. Кодирующую GNA1870 нуклеиновую кислоту можно предоставлять в виде наследуемого эписомального элемента (например, плазмиды) или ее можно встраивать в геном.
Эффективные векторы различаются в композиции в зависимости от вида события рекомбинации, которое следует осуществить. Векторы для встраивания могут условно представлять собой репликативные плазмиды или плазмиды-"самоубийцы", бактериофаги, транспозоны или фрагменты линейной ДНК, получаемые рестрикционным гидролизом или амплификацией посредством ПЦР. Отбор события рекомбинации можно осуществлять посредством селектируемого генетического маркера, такого как гены, придающие устойчивость к антибиотикам (например, к канамицину, эритромицину, хлорамфениколу или гентамицину), гены, придающие устойчивость к тяжелым металлам и/или токсическим соединениям, или гены, дополняющие ауксотрофные мутации (например, pur, leu, met, aro).
В одном из вариантов осуществления вектор представляет собой вектор экспрессии на основе автономно реплицирующихся в E. coli и N. meningitidis эписомальных плазмид, которые содержат селектируемые маркеры устойчивости к лекарственному средству. Одним из примеров такого "челночного" вектора является плазмида pFP10 (Pagotto et al. Gene 2000 244:13-19).
Получение везикул Neisseria meningitidis
Как правило, антигенные композиции для использования по изобретению включают векторы, получаемые из клеток Neisserial, которые экспрессируют GNA1870 на приемлемом уровне, природно или вследствие генетической модификации (например, вследствие экспрессии рекомбинантного GNA1870). В рамках настоящей заявки подразумевают, что "везикулы" включают везикулы наружной мембраны, а также микровезикулы (которые также называют пузырьками).
В одном из вариантов осуществления антигенная композиция содержит везикулы наружной мембраны (OMV), получаемые из наружной мембраны культивируемого штамма из видов Neisseria meningitidis. OMV можно получать из Neisseria meningitidis, выращиваемых в жидкой или твердой культуральной среде, предпочтительно, посредством отделения бактериальных клеток от культуральной среды (например, фильтрацией или низкоскоростным центрифугированием, приводящим к осаждению клеток, или т.п.), лизиса клеток (например, добавлением детергента, осмотическим шоком, воздействием ультразвука, порообразованием, гомогенизацией или т.п.) и отделения фракции наружной мембраны от цитоплазматических молекул (например, фильтрацией; или дифференциальным осаждением или агрегацией наружных мембран и/или везикул наружной мембраны, или способами аффинного разделения с использованием лигандов, которые специфически распознают молекулы наружной мембраны; или высокоскоростным центрифугированием, которое приводит к осаждению наружных мембран и/или везикул наружной мембраны, или т.п.); где фракции наружной мембраны можно использовать для получения OMV.
В другом варианте осуществления антигенная композиция содержит микровезикулы (MV) или пузырьки, высвобождающиеся в ходе культивирования указанных видов Neisseria meningitidis. MV можно получать культивированием штамма Neisseria meningitidis в жидкой культуральной среде, отделением целых клеток от жидкой культуральной среды (например, фильтрацией или низкоскоростным центрифугированием, которое приводит к осаждению только клеток, а не более мелких пузырьков, или т.п.), а затем сбором MV, которые присутствуют в не содержащей клетки культуральной среде (например, посредством фильтрации, дифференциального осаждения или агрегации MV или посредством высокоскоростного центрифугирования, приводящего к осаждению пузырьков, или т.п.). Как правило, штаммы для использования в получении MV отбирают на основе количества пузырьков, продуцируемых в культуре (например, бактерии можно культивировать в надлежащем количестве для обеспечения продукции пузырьков, эффективных для выделения и введения по описанным в настоящей заявке способам). Пример штамма, продуцирующего пузырьки на высоких уровнях, описан в публикации PCT № WO 01/34642. Кроме продукции пузырьков, штаммы для использования в получении MV можно также отбирать на основе продукции NspA, где могут быть предпочтительны штаммы, которые продуцируют NspA на более высоких уровнях (для примеров штаммов N. meningitidis с различными уровнями продукции NspA см., например, Moe et al. (1999 Infect. Immun. 67:5664)).
В другом варианте осуществления антигенная композиция содержит везикулы из одного штамма или из 2, 3, 4, 5 или более штаммов, где штаммы могут быть гомологичными или гетерологичными, обычно гетерологичными в отношении друг друга относительно одного или обоих GNA1870 или PorA. В одном из вариантов осуществления везикулы получают из штамма, экспрессирующего 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более белков GNA1870, которые могут относиться к различным вариантам (v.1, v.2, v.3) или субвариантам (например, субвариант v.1, v.2 или v.3). В другом варианте осуществления антигенные композиции содержат смесь OMV и MV, которые могут быть из одинаковых или различных штаммов. В таких вариантах осуществления везикулы из различных штаммов можно вводить в виде смеси. Кроме того, в виде смеси можно вводить OMV и MV из одинаковых или различных штаммов. Кроме везикул (OMV и/или MV), в антигенные композиции по изобретению можно включать отдельные антигены или конкретные сочетания.
Снижение токсичности липидов
При желании (например, если используемые для получения везикул штаммы связаны с эндотоксином или особенно высокими уровнями эндотоксина), везикулы, необязательно, обрабатывают для снижения количества эндотоксина, например, для снижения токсичности после введения. Хотя и менее желательно, как описано ниже, снижение количества эндотоксина можно осуществлять экстракцией приемлемым детергентом (например, BRIJ-96, дезоксихолатом натрия, лауроилсаркозинатом натрия, Empigen BB, Triton X-100, TWEEN 20 (монолаурат полиоксиэтиленсорбитана), TWEEN 80 в концентрации 0,1-10%, предпочтительно 0,5-2%, и SDS). При использовании экстракции детергентом предпочтительно применять детергент, отличный от дезоксихолата. В определенном варианте осуществления везикулы получают без использования детергента, например, без использования дезоксихолата или другого детергента.
В представляющих особый интерес вариантах осуществления везикулы для антигенных композиций получают без детергента. Хотя обработка детергентом эффективна для устранения активности эндотоксина, она может приводить к снижению количества природного липопротеина GNA1870 в результате экстракции в ходе получения везикул. Таким образом, может быть особенно желательно снижать активность эндотоксина с использованием способа, для которого не требуется детергент. В одном из способов штаммы, продуцирующие эндотоксин (липополисахарид, LPS) на относительно низком уровне, используют таким образом, чтобы избежать необходимости удалять эндотоксин из конечного препарата перед применением у человека. Например, везикулы можно получать из мутантов Neisseria, в которых снижены или устранены липоолигосахарид или другие антигены, могущие быть нежелательными в вакцинах (например, Rmp).
Например, везикулы можно получать из штаммов N. meningitidis, содержащих генетические модификации, которые приводят к сниженной или не поддающейся выявлению токсической активности липида A. Например, такой штамм может быть генетически модифицирован в отношении биосинтеза липида A (Steeghs et al. Infect. Immun. 1999; 67:4988-93; van der Ley et al. Infect. Immun. 2001; 69:5981-90; Steeghs et al. J Endotoxin. Res. 2004; 10:113-9). Мутации в генах, ответственных за конечные стадии модификации, приводят к термочувствительным (htrB) или пермиссивным (msbB) фенотипам. Мутации, приводящие к сниженной (или отсутствию) экспрессии этих генов (или сниженной или отсутствию активности продукта этих генов), приводят к измененной токсической активности липида A. Нелауроилированный (мутант htrB) или немиристоилированный (мутант msbB) липид A менее токсичен, чем липид A дикого типа. Мутации в гене (lpxK), кодирующем 4'-киназу липида A, также снижают токсическую активность липида A.
Также токсичную активность LPS можно изменять введением мутаций в гены/участки, которые вовлечены в устойчивость к полимиксину B (такая устойчивость была связана с добавлением аминоарабинозы к 4'-фосфату липида A). Эти гены/участки могут представлять собой pmrE, кодирующий UDP-глюкозодегидрогеназу, или участок распространенных у многих энтеробактерий генов устойчивости к противомикробному пептиду, вовлеченных в синтез и перенос аминоарабинозы. Ген pmrF, присутствующий в этом участке, кодирует доликолфосфат-маннозилтрансферазу (Gunn J. S., Kheng, B. L., Krueger J., Kim K., Guo L., Hackett M., Miller S. I. 1998. Mol. Microbiol. 27:1171-1182).
Мутации в регуляторной системе PhoP-PhoQ, представляющей собой двухкомпонентную регуляторную систему с передачей сигнала на основе фосфора (например PhoP, конститутивный фенотип, PhoPc), или окружающая среда с низким количеством Mg++ или условия культивирования (активирующие регуляторную систему PhoP-PhoQ) приводят к добавлению аминоарабинозы к 4'-фосфату и замене миристата 2-гидроксимиристатом (гидроксилирование миристата). Этот модифицированный липид A проявляет сниженную способность стимулировать экспрессию E-селектина эндотелиальными клетками человека и секрецию TNF-α моноцитами человека.
Также эффективны для использования по изобретению штаммы, устойчивые к полимиксину B, поскольку для таких штаммов была показана сниженная токсичность LPS (см., например, van der Ley et al. 1994. In: Proceedings of the ninth international pathogenic Neisseria conference. The Guildhall, Winchester, England). Альтернативно, для снижения токсической активности LPS к антигенным композициям можно добавлять синтетические пептиды, воспроизводящие связывающую активность полимиксина B (см., например, Rustici et al. 1993, Science 259:361-365; Porro et al. Prog. Clin. Biol. Res. 1998; 397:315-25).
Также количество эндотоксина можно снижать посредством отбора условий культивирования. Например, культивирование штамма в среде для роста, содержащей 0,1 мг - 100 мг аминоарабинозы в литре среды, обеспечивает сниженную токсичность липида (см., например, WO 02/097646).
Препараты
Используемые в качестве вакцин иммуногенные композиции содержат иммунологически эффективное количество антигена, особенно иммунологически эффективное количество GNA1870, а также любые другие совместимые компоненты по мере необходимости. "Иммунологически эффективное количество" означает, что введение индивидууму этого количества, в разовой дозе или в виде части серий введений, эффективно для достижения лечения или профилактики. Это количество различается в зависимости от здоровья и физического состояния подлежащего лечению индивидуума, возраста, таксономической группы подлежащего лечению индивидуума (например, не относящийся к человеку примат, примат, человек и т.д.), способности иммунной системы индивидуума синтезировать антитела, степени желаемой защиты, состава вакцины, оценки медицинской ситуации лечащим клиницистом и других соответствующих факторов. Предполагают, что количество входит в относительно широкий диапазон, который можно определять общепринятыми исследованиями.
Схема введения доз может представлять собой схему разовой дозы или схему многократных доз (например, включая ревакцинирующие дозы) с использованием стандартной лекарственной формы антигенной композиции, вводимой в различные моменты времени. В рамках настоящей заявки термин "стандартная лекарственная форма" относится к физически дискретным формам, приемлемым в качестве стандартных доз для человека и животных, где каждая форма содержит заданное количество антигенных композиций по настоящему изобретению в количестве, достаточном для оказания желаемого эффекта, где эти композиции предоставлены совместно с фармацевтически приемлемым эксципиентом (например, фармацевтически приемлемым разбавителем, носителем или инертным веществом). Вакцину можно вводить в сочетании с другими иммунорегуляторными средствами.
Подлежащие введению антигенные композиции представляют в фармацевтически приемлемом разбавителе, таком как водный раствор, часто солевой раствор, в полутвердом виде (например, гель) или в виде порошка. Такие разбавители могут быть инертными, хотя композиции по изобретению могут также содержать адъювант. Примеры известных приемлемых адъювантов, которые можно использовать для человека, включают в себя, но не обязательно ограничиваются ими, алюминий, фосфат алюминия, гидроксид алюминия, MF59 (4,3% мас./об. сквалена, 0,5% мас./об. Tween 80, 0,5% мас./об. Span 85), CpG-содержащую нуклеиновую кислоту (где цитозин не метилирован), QS21, MPL, 3DMPL, экстракты Aquilla, ISCOMS, мутанты LT/CT, микрочастицы из сополимера D,L-лактида и гликолида) (PLG), Quil A, интерлейкины и т.п. Для экспериментальных животных можно использовать адъювант Фрейнда, N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-ацетилнормурамил-L-аланил-D-изоглутамин (CGP 11637, называемый нор-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)этиламин (CGP 19835A, называемый MTP-PE), а также RIBI, содержащий три экстрагируемых из бактерий компонента, монофосфориллипид A, димиколат трегалозы и каркас клеточной стенки (MPL+TDM+CWS) в 2% эмульсии сквален/Tween 80. Эффективность адъюванта можно определять измерением количества антител, направленных против иммуногенного антигена.
Дополнительные примеры адъювантов для повышения эффективности композиции включают, но ими не ограничиваются: (1) препараты эмульсий масло-в-воде (при наличии или отсутствии других специфичных иммуностимулирующих средств, таких как пептиды мурамила (см. ниже) или компоненты бактериальной клеточной стенки), такие как, например, (a) MF59TM (W090/14837; глава 10 в Vaccine design: the subunit and adjuvant approach, eds. Powell & Newman, Plenum Press 1995), содержащий 5% сквалена, 0,5% Tween 80 и 0,5% Span 85 (необязательно, содержащего MTP-PE), который составляют в виде субмикронных частиц с использованием микрофлюидизатора, (b) SAF, содержащий 10% сквалена, 0,4% Tween 80, 5% плюронового блок-полимера L 121 и thr-MDP, микрофлюидизированный в субмикронную эмульсию или встряхиваемый для получения эмульсии с частицами более крупного размера, а также (c) систему адъюванта REBITM (RAS), (Ribi Immunochem, Hamilton, MT), содержащую 2% сквалена, 0,2% Tween 80 и один или несколько компонентов бактериальной клеточной стенки, таких как монофосфориллипид A (MPL), димиколат трегалозы (TDM) и каркас клеточной стенки (CWS), предпочтительно MPL + CWS (DETOXTM); (2) можно использовать сапониновые адъюванты, такие как QS21 или STIMULONTM (Cambridge Bioscience, Worcester, MA), или получаемые из них частицы, такие как ISCOM (иммуностимулирующие комплексы), где эти ISCOM могут не содержать дополнительный детергент, например WO00/07621; (3) полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (IFA); (4) цитокины, такие как интерлейкины (например, IL-1, IL-2, IL-4, IL-5, IL-6, IL-7, IL-12 (WO99/44636) и т.д.), интерфероны (например, гамма-интерферон), макрофагальный колониестимулирующий фактор (M-CSF), фактор некроза опухолей (TNF) и т.д.; (5) монофосфориллипид A (MPL) или 3-O-деацетилированный MPL (3dMPL), например GB-2220221, EP-A-0689454, необязательно, при практическом отсутствии алюминия в случае использования с пневмококковыми сахаридами, например WO00/56358; (6) сочетания 3dMPL с, например, QS21 и/или эмульсиями масло-в-воде, например EP-A-0835318, EP-A-0735898, EP-A-0761231; (7) олигонуклеотиды, содержащие CpG-фрагменты [Krieg Vaccine 2000, 19, 618-622; Krieg Curr. Opin. Mol Ther. 2001 3:15-24; Roman et al., Nat. Med, 1997, 3, 849-854; Weiner et al., PNAS USA, 1997, 94, 10833-10837; Davis et al., J. Immunol., 1998, 160, 870-876; Chu et al., J. Exp. Med, 1997, 186, 1623-1631; Lipford et al., Ear. J. Immunol., 1997, 27, 2340-2344; Moldoveami et al., Vaccine, 1988, 16, 1216-1224, Krieg et al., Nature, 1995, 374, 546-549; Klinman et al., PNAS USA, 1996, 93, 2879-2883; Ballas et al., J. Immunol., 1996, 157, 1840-1845; Cowdery et al., J. Immunol., 1996, 156, 4570-4575; Halpern et al., Cell Immunol., 1996, 167, 72-78; Yamamoto et al., Jpn. J. Cancer Res., 1988, 79, 866-873; Stacey et al., J. Immunol., 1996, 157, 2116-2122; Messina et al., J. Immunol., 1991, 147, 1759-1764; Yi et al., J. Immunol., 1996, 157, 4918-4925; Yi et al., J. Immunol., 1996, 157, 5394-5402; Yi et al., J. Immunol., 1998, 160, 4755-4761; и Yi et al., J. Immunol., 1998, 160, 5898-5906; международные патентные заявки WO96/02555, W098/16247, WO98/18810, WO98/40100, WO98/55495, WO98/37919 и WO98/52581], т.е. содержащие по крайней мере один динуклеотид CG, где цитозин не метилирован; (8) простой эфир полиоксиэтилена или сложный эфир полиоксиэтилена, например WO99/52549; (9) поверхностно-активный сложный эфир полиоксиэтиленсорбитана в комбинации с октоксинолом (WO01/21207) или поверхностно-активный простой алкиловый эфир или сложный эфир полиоксиэтилена в комбинации по крайней мере с одним дополнительным неионным поверхностно-активным средством, таким как октоксинол (WO01/21152); (10) сапонин и иммуностимулирующий олигонуклеотид (например, олигонуклеотид CpG) (WO00/62800); (11) иммуностимулирующее вещество и частицу соли металла, например WO00/23105; (12) сапонин и эмульсию масло-в-воде, например WO99/11241; (13) сапонин (например, QS21) + 3dMPL + IM2 (необязательно, + стерин), например WO98/57659; (14) другие вещества, действующие как иммуностимулирующие средства, для повышения эффективности композиции. Пептиды мурамила включают N-ацетилмурамил-L-треонил-D-изоглутамин (thr-MDP), N-25-ацетилнормурамил-L-аланил-D-изоглутамин (нор-MDP), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)этиламин MIP-PE) и т.д.
Антигенные композиции можно сочетать с общепринятым фармацевтически приемлемым эксципиентом, таким как относящиеся к фармацевтическим категориям маннит, лактоза, крахмал, стеарат магния, сахаринат натрия, тальк, целлюлоза, глюкоза, сахароза, карбонат магния и т.п. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, как необходимо для соответствия физиологическим условиям, например, доводящие pH и буферные вещества, корректирующие токсичность средства и т.п., например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и т.п. Концентрация антигена в этих препаратах может широко различаться и ее исходно выбирают, основываясь на объемах жидкостей, величинах вязкости, массе тела и т.п., в соответствии с конкретным выбранным способом введения и потребностями пациента. Получаемые композиции могут находиться в виде раствора, суспензии, таблетки, пилюли, капсулы, порошка, геля, крема, лосьона, мази, аэрозоли или т.п.
Концентрация белка в антигенных композициях по изобретению в фармацевтических препаратах может широко различаться, т.е. приблизительно от менее чем 0,1%, обычно от или по крайней мере приблизительно от 2%, до 20%-50% или более по массе, и, в основном, ее выбирают исходя из объемов жидкостей, величин вязкости и т.д. в соответствии с конкретным выбранным способом введения.
Иммунизация
В целом, способы по изобретению обеспечивают введение млекопитающему (например, человеку) одной или нескольких антигенных композиций по изобретению таким образом, чтобы вызвать защитную иммунную реакцию в отношении более чем одного штамма бактерий вида Neisseria и, таким образом, защиту против заболевания, вызываемого такими бактериями. В частности, способы по изобретению могут обеспечивать иммунопротективную иммунную реакцию в отношении 1, 2, 3, 4 или более штаммов вида Neisseria meningitidis, где штаммы отличаются по крайней мере по одному из серогруппы, серотипа, сероподтипа или полипептида GNA1870 (например, различные варианты и/или субварианты GNA1870). Особый интерес представляет индукция протективной иммунной реакции против множества штаммов Neisseria meningitidis серогруппы B, особенно, когда штаммы различаются по сероподтипу (например, имеют гетерологичные PorA). Также особый интерес представляет индукция протективной иммунной реакции против штаммов, гетерологичных друг другу в отношении PorA и/или GNA1870.
Антигенные композиции по изобретению можно вводить перорально, назально, через носоглотку, парентерально, энтерально, гастрально, местно, трансдермально, подкожно, внутримышечно, в таблетке, в виде твердого вещества, в виде порошка, жидкости, в форме аэрозоли, местно или системно, с наличием или отсутствием добавления эксципиентов. Существующие способы получения парентерально вводимых композиций известны или понятны специалисту в данной области и более подробно описаны в таких публикациях, как Remington's Pharmaceutical Science, 15th ed., Mack Publishing Company, Easton, Pennsylvania (1980).
Признано, что при пероральном введении может быть необходима защита композиций от переваривания. Как правило, это осуществляют посредством связывания композиции со средством, сообщающим ей устойчивость к кислотному и ферментативному гидролизу, или посредством упаковки композиции в соответствующий устойчивый носитель. Способы защиты от переваривания хорошо известны в данной области.
Композиции вводят животному, подверженному риску возникновения заболевания Neisserial, для профилактики или по крайней мере частичного прекращения развития заболевания и его осложнений. Соответствующее для достижения этого количество определено как "терапевтически эффективная доза". Количества, эффективные для терапевтического использования, зависят, например, от антигенной композиции, способа введения, массы и общего состояния здоровья пациента, а также оценки назначающего лечение врача. Разовые или многократные дозы антигенных композиций можно вводить в зависимости от дозы и частоты, необходимых и переносимых пациентом, а также от способа введения.
Описанные в настоящей заявке антигенные композиции могут содержать смесь везикул (например, OMV и MV), где эти везикулы могут быть из одинаковых или различных штаммов. В другом варианте осуществления антигенные композиции могут содержать смесь везикул из 2, 3, 4, 5 или более штаммов, где везикулы могут представлять собой OMV, MV или и то и другое.
Антигенные композиции вводят в количестве, эффективном для вызова в организме-хозяине иммунной реакции, особенно гуморальной иммунной реакции. Как правило, количества смеси для иммунизации входят в диапазон приблизительно от 0,001 мг до приблизительно 1,0 мг для пациента с массой 70 килограмм, более обычно - приблизительно от 0,001 мг до приблизительно 0,2 мг для пациента с массой 70 килограмм. Можно использовать дозы от 0,001 вплоть до приблизительно 10 мг для пациента в сутки, особенно, если антиген вводят в обособленный участок, а не в кровоток, например, в полость тела или в полость органа. Значительно более высокие дозы (например, от 10 до 100 мг или более) возможны при пероральном, назальном или местном применении. Предпочтительно, за исходным введением смеси может следовать повторная иммунизация той же или отличной смесью с использованием по крайней мере одной ревакцинирующей дозы, более обычно - двух ревакцинирующих доз.
В одном из вариантов осуществления антигенные композиции, используемые для первичной и повторной вакцинации, получают из штаммов Neisseria, имеющих различные иммунодоминантные антигены (основные антигены, которые общепринято определяют антисывороткой из различных животных-хозяев, инфицированных Neisseria; типичные примеры включают порин A, порин B, пилин, NspA, фосфолипиды, полисахариды, липополисахариды, пилины, OmpA, Opa, Opc и т.д.) и/или различные белки GNA1870. Также штаммы могут различаться в отношении молекулы капсулы, что отражено их серогруппой.
В настоящее время классификацию по серотипам и сероподтипам проводят посредством выявления того, какой набор известных моноклональных антител, для которых известно распознавание специфических молекул порина, связывается с неизвестным штаммом (Sacchi et al., 1998, Clin. Diag. Lab. Immunol. 5:348). Существует вероятность выявления других таких моноклональных антител. В изобретении специально предусмотрено использование любых новых серотипов и сероподтипов, которые можно определить любыми новыми моноклональными антителами. Кроме того, серотипы и сероподтипы можно определять не только посредством взаимодействия с моноклональными антителами, но также и структурно, по отсутствию и/или наличию определенных пептидных остатков и пептидных эпитопов (Sacchi et al., 2000, J. Infect. Dis. 182:1169). В изобретении специально предусмотрены схемы классификации по серотипам и сероподтипам на основе структурных особенностей поринов (которые известны или могут быть открыты в дальнейшем).
В другом варианте осуществления вводимые антигенные композиции получают из 2, 3, 4, 5 или более штаммов, где эти штаммы могут быть гомологичными или гетерологичными, обычно гетерологичными друг другу в отношении одного или обоих GNA1870 или PorA. В одном из вариантов осуществления везикулы получают из штаммов, экспрессирующих различные белки GNA1870, где эти белки GNA1870 могут представлять собой различные варианты (v.1, v.2, v.3) или субварианты (например, субвариант v.1, v.2 или v.3). В другом варианте осуществления везикулы получают из штаммов, гетерологичных друг другу в отношении PorA.
В представляющих особый интерес вариантах осуществления везикулы получают из штаммов Neisserial, генетически отличных друг от друга (например, штаммов, относящихся к различным серотипам и/или сероподтипам; экспрессирующих различные белки PorA; экспрессирующих различные варианты или субварианты GNA1870; и/или могущих также, необязательно, относиться к различным серогруппам капсулы). Везикулы можно использовать для получения антигенной композиции, представляющей собой смесь везикул, которые получают по крайней мере из 2, 3, 4 или более таких генетически различных штаммов. Например, белок GNA1870 и/или PorA из второго штамма Neisserial, из которого получают и используют антигенные композиции, является/являются отличными от таковых из первого штамма, используемого для получения везикул.
Необязательно, вторую, третью и дальнейшие вводимые антигенные композиции можно получать из штаммов Neisserial, генетически отличных от второго штамма (например, штаммы относятся к различным серотипам и/или сероподтипам; экспрессируют различные белки GNA1870; экспрессируют различные белки PorA; и/или относятся к различным серогруппам капсулы). Например, третий штамм, используемый для получения третьей антигенной композиции, может генетически отличаться от первого и второго штаммов, используемых для получения первой и второй антигенных композиций, однако в некоторых вариантах осуществления может генетически не отличаться в отношении первого штамма.
Также по изобретению предусмотрено, что антигенные композиции можно получать из одного или нескольких штаммов Neisseria, особенно Neisseria meningitidis, генетически сконструированных известными способами (см., например, патент США №6013267) для экспрессии одной или нескольких нуклеиновых кислот, которые кодируют GNA1870. Клетка-хозяин может экспрессировать эндогенный полипептид GNA1870 или она может быть модифицирована или отобрана таким образом, чтобы не экспрессировать какое-либо поддающееся выявлению количество эндогенного полипептида GNA1870. Полипептид GNA1870, экспрессируемый в клетке-хозяине посредством рекомбинантных способов (т.е. экзогенный полипептид GNA1870), может быть одинакового или отличного типа варианта в сравнении с эндогенным полипептидом GNA1870.
Клетки-хозяева можно дополнительно модифицировать для экспрессии дополнительных интересующих антигенов, таких как порин A, порин B, NspA, пилин или другие белки Neisserial. Кроме того, антигенные композиции по изобретению могут содержать дополнительные антигены Neisserial, такие как антигены, приведенные в качестве примера в публикациях PCT №№ WO 99/24578, WO 99/36544; WO 99/57280, WO 00/22430 и WO 00/66791, а также антигенные участки таких белков.
Как правило, антигенные композиции вводят млекопитающему, иммунологически наивному в отношении Neisseria, особенно в отношении Neisseria meningitidis. В конкретном варианте осуществления млекопитающее представляет собой ребенка человека в возрасте приблизительно пяти лет или младше, а предпочтительно - приблизительно двух лет или младше, а антигенные композиции вводят в любой один или несколько следующих моментов времени: две недели, один месяц, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11 месяцев, или один год, или 15, 18 или 21 месяц после рождения, или в возрасте 2, 3, 4 или 5 лет.
Как правило, введение любому млекопитающему предпочтительно начинают до первого проявления симптомов заболевания или при первом признаке возможного или фактического воздействия Neisseria.
Пассивный иммунитет
Также настоящее изобретение относится к иммунопротективным антителам, получаемым иммунизацией с использованием антигенной композиции по изобретению, и к способам использования. Такие антитела можно вводить индивидууму (например, пациенту человеку) для обеспечения пассивного иммунитета против заболевания Neisserial, или для профилактики возникновения инфекции или заболевания, или в качестве лечения для улучшения исхода заболевания у пациентов с выявленным заболеванием (например, снижения выраженности осложнений, таких как шок, снижения смертности или снижения заболеваемости, например глухотой).
Антитела, которые вводят индивидууму, относящемуся к виду, отличному от видов, в которых образуются антитела, часто являются иммуногенными. Таким образом, например, вводимые человеку мышиные или свиные антитела часто индуцируют иммунологическую реакцию против антитела. Иммуногенные свойства антитела снижают посредством изменения участков или всего антитела в характерные для человека последовательности, тем самым получая химерные антитела или антитела человека соответственно.
Химерные антитела представляют собой молекулы иммуноглобулинов, которые содержат принадлежащие человеку и не принадлежащие человеку участки. Более конкретно, связывающий антиген участок (или вариабельная область) гуманизированного химерного антитела происходит из не относящегося к человеку источника (например, мышиный), а константная область химерного антитела (которая придает иммуноглобулину биологическую эффекторную функцию) происходит из источника, относящегося к человеку. Химерное антитело должно обладать антигенсвязывающей специфичностью не принадлежащей человеку молекулы антитела и эффекторной функцией, которую придает молекула антитела, относящаяся к человеку. Специалистам в данной области хорошо известно большое число способов получения химерных антител (см., например, патенты США №№5502167, 5500362, 5491088, 5482856, 5472693, 5354847, 5292867, 5231026, 5204244, 5202238, 5169939, 5081235, 5075431 и 4975369). Альтернативным способом является получение гуманизированных антител посредством связывания способами рекомбинантной ДНК областей CDR из не принадлежащих человеку антител с константными областями, относящимися к человеку. См. Queen et al, Proc. Natl. Acad. Sci. USA 86:10029-10033 (1989) и WO 90/07861.
В одном из вариантов осуществления вектор рекомбинантной ДНК используют для трансфицирования линии клеток, продуцирующих антитело против пептида по изобретению. Новый вектор рекомбинантной ДНК содержит "ген замены" для замены всего или части гена, кодирующего константную область иммуноглобулина в линии клеток (например, ген замены может кодировать всю или часть константной области иммуноглобулина человека или специфического класса иммуноглобулинов), а также содержит "последовательность-мишень", которая делает возможной заданную гомологичную рекомбинацию последовательностей иммуноглобулинов в клетке, продуцирующей антитела.
В другом варианте осуществления вектор рекомбинантной ДНК используют для трансфекции линии клеток, продуцирующих антитело с желаемой эффекторной функцией (например, константной областью иммуноглобулина человека), где в этом случае ген замены, содержащийся в рекомбинантном векторе, может кодировать всю или часть области антитела, а последовательность-мишень, содержащаяся в рекомбинантном векторе, делает возможной гомологичную рекомбинацию и заданную модификацию гена в клетке, которая продуцирует антитело. В любом варианте осуществления, если заменяют только часть вариабельной или константной области, получаемое в результате химерное антитело может определять тот же антиген и/или иметь ту же эффекторную функцию, хотя и быть измененным или улучшенным таким образом, что химерное антитело может проявлять более высокую антигенную специфичность, более высокую константу аффинности связывания, усиленную эффекторную функцию или повышенную секрецию и продукцию трансфицированной линией клеток, продуцирующих антитело, и т.д.
В другом варианте осуществления это изобретение относится к антителам, полностью относящимся к человеку. Антитела человека полностью состоят из характерных последовательностей полипептидов человека. Антитела человека по этому изобретению можно получать множеством различных способов (см., например, Larrick et al., патент США №5001065). В одном из вариантов осуществления антитела человека по настоящему изобретению исходно получают в клетках-триомах (происходящих из трех клеток, двух человеческих и одной мышиной). Затем кодирующие антитела гены клонируют и экспрессируют в других клетках, особенно в клетках млекопитающих, не относящихся к человеку. Общий подход к получению антител человека способом триом был описан Ostberg et al. (1983), Hybridoma 2:361-367, Ostberg, патент США №4634664 и Engelman et al., патент США № 4634666. Было открыто, что триомы продуцируют антитела более стабильно, чем обычные гибридомы, получаемые из клеток человека.
Способы получения и составления антител, эффективных для введения индивидууму (например, человеку), хорошо известны в данной области. Например, антитела можно предоставлять в фармацевтической композиции, содержащей эффективное количество антитела и фармацевтические эксципиенты (например, солевой раствор). Необязательно, фармацевтическая композиция может содержать другие добавки (например, буферные вещества, стабилизаторы, консерванты и т.п.). Как правило, эффективное количество антитела представляет собой количество, эффективное для обеспечения защиты против заболевания Neisserial или симптомов в течение желаемого периода, например, периода величиной по крайней мере приблизительно от 2 суток до 10 суток или от 1 месяца до 2 месяцев.
Диагностические анализы
Антигенные композиции по изобретению или антитела, получаемые введением таких композиций, также можно использовать в диагностических целях. Например, антигенные композиции можно использовать для скрининга преиммунной и иммунной сывороток для подтверждения того, что вакцинация была эффективной. Также антитела можно использовать в иммуноанализах для выявления присутствия конкретных молекул антигенов, связанных с заболеванием Neisserial.
ПРИМЕРЫ
Понятно, что примеры и варианты осуществления описаны в настоящей заявке только в иллюстративных целях и что специалисты в данной области предлагают в их связи различные модификации или изменения, которые входят в сущность и рамки этой заявки и в объем прилагаемой формулы изобретения. Все публикации, патенты и патентные заявки, цитируемые в настоящей заявке, приведены здесь полностью во всех целях в качестве ссылки.
Вещества и способы
Следующие вещества и способы использовали в приведенных ниже примерах.
Бактериальные штаммы. В таблице 1 перечислены девять штаммов N. meningitidis, используемых в этом исследовании (шесть с капсулой группы B, и один с капсулой группы A, а два с капсулой группы C). Штаммы собирали в течение периода 25 лет от пациентов, госпитализированных на Кубе, в Нидерландах, Германии, Новой Зеландии или США. Штаммы генетически отличались на основе электрофоретических анализов кластеров и/или типирования последовательностей.

Штамм RM1090 (C:2a:5-1,2) группы C и описанные ниже мутанты, происходящие из этого штамма, а также штамм H44/76 группы B и описанные ниже мутанты, происходящие из этого штамма, использовали для получения вакцин на основе везикул наружной мембраны (OMV). В природе штамм RM1090 экспрессирует варианта 2 белка GNA1870 на низком уровне. Штамм, в котором ген GNA1870 инактивирован (описанный ниже RM1090ΔGNA1870), использовали для сверхэкспрессии варианта 1 GNA1870. Штамм H44/76 экспрессирует вариант 1 GNA1870 на относительно высоком уровне. Остальные семь штаммов в природе экспрессируют субварианты варианта 1 белка GNA1870 и были отобраны в качестве тестируемых организмов для определения диапазона индуцируемого вакциной защитного иммунитета против варианта 1 GNA1870. Эти штаммы генетически различны, как определено по электрофоретическому типу и/или типу при секвенировании множества участков, а также они экспрессируют различные типы последовательностей VR PorA. Штаммы варианта 1 были выбраны, поскольку они составляют приблизительно 60% изолятов группы B, вызывающих заболевание (Masignani et al. J Exp. Med. 2003; 197:789-99).
Штамм Cu385 и штамм H44/76 экспрессируют вариант 1 GNA1870 с аминокислотной последовательностью, совпадающей с таковой для штамма MC58 (Welsch et al. J Immunol. 2004; 172:5606-15), где ген используют для экспрессии рекомбинантного варианта 1 белка GNA1870 в E. coli, а также используют в "челночном" векторе для сверхэкспрессии GNA1870 в штамме для вакцины RM1090 N. meningitidis (см. ниже). Остальные семь штаммов экспрессируют субварианты варианта 1 GNA1870 с незначительными отличиями последовательностей от варианта белка GNA1870, кодируемого геном из штамма MC58 (Masignani et al. 2003, выше). В предыдущем исследовании штамм Cu385 был высоко чувствителен к опосредуемой комплементом бактерицидной активности антител, образующихся у мышей, которые иммунизированы вакциной рекомбинантного белка GNA1870 (таблица 1). В отличие от этого штаммы BZ198, M1390, M6190 и NZ98/294 N. meningitidis были выбраны, поскольку они устойчивы к бактерицидной активности антисыворотки, получаемой против рекомбинантной вакцины GNA1870 (бактерицидные титры <1:10).
Конструкция "челночного" вектора pFP12-GNA1870. Сверхэкспрессию GNA1870 в N. meningitidis осуществляли с использованием "челночного" вектора FP12, который имеет участок инициации репликации из природной плазмиды в N. gonorrhoeae, и для которого была показана стабильная трансформация E. coli и N. meningitidis (Pagotto et al. Gene 2000;244:13-9). Ген варианта 1 GNA1870, в том числе предполагаемую промоторную рамку FUR из штамма MC58 N. meningitidis, амплифицировали с геномной ДНК посредством PCR с использованием следующих праймеров: GNA1870FURSphIF 5', 5"-ATCGGCATGCGCCGTTCGGACGACATTTG-3" и GNA1870FURStuIR 3' 5"-AAGAAGGCCTTTATTGCTTGGCGGCAAGGC-3". Затем продукт ПЦР расщепляли эндонуклеазами рестрикции SphI и StuI и лигировали в плазмиду pFP12, расщепляемую посредством SphI и StuI, что устраняло ген GFP. Получаемую плазмиду pFP12-GNA1870 трансформировали и размножали в компетентных клетках штамма TOP10 E. coli (Invitrogen), выращиваемых в среде Лурия-Бертани при 37° при селекции хлорамфениколом (50 мкг/мл).
Трансформация N. meningitidis. Штамм RM1090, в котором инактивирован ген GNA1870, (RM1090ΔGNA1870) получали гомологичной рекомбинацией посредством трансформации плазмидой pBSUDGNA1870ERM с использованием селекции эритромицином (5 мкг/мл). Для получения мутанта, сверхэкспрессирующего GNA1870, отбирали 3-4 колонии нокаутного штамма RM1090ΔGNA1870 с планшета с шоколадным агаром, выращиваемые в течение ночи. Колонии бактерий перемешивали с 3 мкг плазмиды pFP12-GNA1870 в 20 мкл буфера EB (Qiagen), высевали на планшет с шоколадным агаром и инкубировали в течение 5 часов при 37°C. Серийные разведения бактерий повторно культивировали на планшетах с шоколадным агаром, содержащих хлорамфеникол (5 мкг/мл). Культуральные планшеты инкубировали в течение ночи при 37°C и отбирали колонии на предмет экспрессии GNA1870 посредством блот-анализа колоний с использованием мышиного поликлонального антитела против rGNA1870. Отбирали отдельные положительные колонии и повторно культивировали на планшетах с шоколадным агаром, содержащих хлорамфеникол. Менингококковые бактериальные клетки замораживали в 2% снятом молоке (мас./об.) и хранили при -80°C.
Аналогичную процедуру использовали для трансформации штамма H44/76 и его мутанта, сверхэкспрессирующего GNA1870. Хромосомный gnal870 в штамме H44/76 был инактивирован трансформацией посредством pBSUDgnal870erm (Hou et al. Infect. Immun. 2003; 71:6844-49). Затем мутант (H44/76Δgnal870) трансформировали плазмидой pFP12-GNA1870, кодирующей вариант 1 GNA1870 из штамма MC58. Трансформанты отбирали на планшетах с шоколадным агаром, содержащих 5 мкг/мл хлорамфеникола.
Препараты мембран. Пересевали N. meningitidis из замороженного исходного источника на планшеты с шоколадным агаром (Remel, Laztakas, Kans). После инкубации в течение ночи при 37°C в 5% CO2 отбирали несколько колоний и инокулировали приблизительно в 6 мл жидкой питательной среды Mueller-Hinton, включающей 0,25% глюкозы и 0,02 мМ CMP-NANA, в атмосфере, содержащей 5% CO2, до оптической плотности величиной 0,1 при 620 нм (OD620). Все штаммы, содержащие введенный "челночный" вектор, выращивали в присутствии 5 мкг/мл хлорамфеникола. Инокулированную жидкую питательную среду инкубировали при 37°C и 5% CO2 при покачивании до достижения OD620 от 0,6 до 0,7 (от 2 до 3 часов). Для инокулирования 1 л жидкой питательной среды Mueller-Hinton использовали шесть начальных культур объемом 6 мл. Наращивали большее количество культуры при 37°C при энергичном встряхивании до OD620 от 0,8 до 1,0. Добавляли фенол (0,5% мас./об.) и оставляли жидкую питательную среду на ночь при 4°C для уничтожения бактерий. Бактериальные клетки осаждали центрифугированием (10000×g) в течение 30 минут при 4°C и замораживали и хранили при -20°C до использования для получения вакцин на основе везикул наружной мембраны.
Для культур, содержащих штамм H44/76 и его мутант, использовали шесть начальных культур объемом 7 мл. Клетки переносили в 1 л жидкой питательной среды Mueller-Hinton в отсутствие добавления хлорамфеникола и выращивали при энергичном встряхивании до достижения OD620 от 0,8 до 1,0. Добавляли фенол (0,5% мас./об.), оставляли культуру на два часа при 37°C и инкубировали в течение ночи при 4°C для уничтожения бактерий. Клетки осаждали центрифугированием (11000×g) в течение 30 минут при 4°C.
Фракции мембран N. meningitidis для OMV получали, как описано ранее, без использования детергентов для избегания экстракции липопротеина GNA1870 (Moe et al. 2002, выше). Вкратце, замороженные бактериальные клетки суспендировали в 40 мл PBS и подвергали воздействию во льду ультразвуком с использованием ультразвукового дезинтегратора, снабженного микронасадкой (Branson, Danbury, Conn), посредством четырех 15-секундных импульсов, что являлось достаточным для высвобождения мембранных пузырьков, но не вызывало полный лизис бактерий. Бактериальные суспензии между импульсами охлаждали во льду. Продукты разрушения клеток удаляли центрифугированием в течение 15 минут при 5000×g, а фракцию мембран, остающихся в надосадочной жидкости, получали ультрацентрифугированием при 100000×g в течение 1 часа при 4°C и ресуспендировали в 5 мл PBS. Эти препараты были названы OMV. Альтернативно, можно использовать MV, получаемые из пузырьков, которые высвобождаются бактериями в надосадочную жидкость, как описано в (Moe et al. 2002, выше); см. также WO 02/09643.
В случае H44/76 (и его мутанта) замороженные бактериальные клетки ресуспендировали в 20 мл буфера PBS и воздействовали ультразвуком посредством четырех 15-секундных импульсов. Продукты разрушения клеток удаляли центрифугированием (16000×g) в течение 30 минут при 4°C, а клеточные мембраны, обогащенные белками наружной мембраны, собирали из растворимой фракции посредством центрифугирования (100000×g) в течение 2 часов.
Характеризация вакцин. Концентрации белков определяли анализом белков DC (Bio-Rad, Richmond, CA.) и посредством набора для анализа белков BCA (Pierce, Rockford, IL). Препараты OMV анализировали посредством 15% SDS-PAGE (12,5% SDS-PAGE для препаратов H44/76), как описано Laemmli (Nature 1970;227:680-5) с использованием устройства для электрофореза Mini-Protean II (Bio-Rad) и Вестерн-блоттинга. Образцы суспендировали в буфере для образцов (0,06 М Tris·HCl, pH 6,8, 10% (об./об.) глицерина, 2% (мас./об.) SDS, 5% (об./об.) 2-меркаптоэтанола, 10 мкг/мл бромфенолового синего) и нагревали до 100°C в течение 5 минут перед загрузкой непосредственно в гель.
В случае Вестерн-блоттингов гель уравновешивали буфером (48 мМ Tris·HCl, 39 мМ глицина [pH 9,0] 20% (об./об.) метанола) и переносили на нитроцеллюлозную мембрану (Bio-Rad) с использованием полусухого электрофоретического переноса клеток Trans-BlotTM (Bio-Rad). Нитроцеллюлозные мембраны блокировали 2% (мас./об.) обезжиренным молоком в PBS и воздействовали на них антисывороткой против rGNA1870 при разведении 1:20000 в PBS, содержащем 1% (мас./об.) BSA и 1% (мас./об.) Tween-20. Связавшееся антитело выявляли с использованием конъюгированного с пероксидазой хрена кроличьего поликлонального антитела против IgG+A+M мыши (Zymed, South San Francisco, CA) и хемилюминисцентных реагентов "Western Lightning" (PerkinElmer Life Sciences, Inc., Boston, MA). Антисыворотка против GNA1870 для регистрации была от мышей, последовательно иммунизированных разовыми инъекциями, по 10 мкг каждая, рекомбинантного GNA1870 v.1 (ген из штамма MC58 N. meningitidis) с последующей дозой рекомбинантного белка v.3 (ген из штамма M1239) с последующей дозой рекомбинантного белка v.2 (ген из штамма 2996). Каждая инъекция была отделена периодом 3-4 недель.
Иммунизация. Вакцину рекомбинантного белка экспрессировали в E. coli, как описано ранее, с использованием последовательности ДНК GNA1870, кодирующей шесть COOH-концевых гистидинов (метка His) и не содержащей N-концевой последовательности, кодирующей предполагаемый лидерный пептид (Welsch et al. J Immunol. 2004; 172:5606-15). Этот нелипидизированный белок GNA1870 с меткой His использовали, поскольку он обеспечивает большую легкость получения, чем рекомбинантный липопротеин, а данные из предыдущих исследований показывают, что нелипидизированный антиген, вводимый с полным и неполным адъювантами Фрейнда, вызывал у мышей значительные реакции бактерицидных антител против большинства тестируемых штаммов.
Препараты OMV или рекомбинантный белок GNA1870 разводили в PBS и адсорбировали с использованием равного объема адъюванта из фосфата алюминия (1% Alhydrogel в конечной концентрации [масс./об.; Superfos Biosector, Frederikssund, Denmark]), который инкубировали с буфером PBS. Внутрибрюшинно (IP) иммунизировали группы самок мышей CD1 в возрасте 4-6 недель (Charles River Breeding Laboratories, Raleigh, NC) (N=10 на группу). Каждой мыши вводили дозу, содержащую 5 мкг общего белка (для смешанной группы по 2,5 мкг каждого из OMV и rGNA1870). Суммарно проводили три инъекции, где каждая была отделена 3-недельным периодом. Через две недели после третьей дозы у мышей удаляли кровь посредством пункции сердца и их умерщвляли. Отделяли сыворотку и хранили замороженной при -20°C.
В случае препаратов H44/76 каждой мыши вводили дозу 1,25 мкг присутствующего в OMV общего белка и 170 мкг фосфата алюминия. Проводили три инъекции, разделенные тремя неделями. Кровь собирали пункцией сердца через три недели после третьей дозы. Сыворотку отделяли центрифугированием и до использования хранили замороженной при -70°C.
Абсорбция антител против GNA1870. Для теста вклада антител против GNA1870 в функциональную активность антител авторы изобретения абсорбировали совокупности сывороток для удаления антител против GNA1870. Вкратце, 100 мкл совокупностей сывороток, разведенных 1:2 в буфере PBS, содержащем 10 мМ имидазола, добавляли в колонку, которая содержала 250 мкл сефарозы Ni-NTA (Qiagen, Valencia, CA), находящейся в комплексе с 200 мкг рекомбинантного белка GNA1870 с меткой His или, в качестве отрицательного контроля, рекомбинантного белка NadA с меткой His (Comanducci et al. J Exp. Med. 2002; 195:1445-54; Hou et al. 2005, выше). Колонки инкубировали в течение ночи при 4°C и промывали посредством 500 мкл буфера PBS, содержащего 10 мМ имидазола. Объединяли пять прошедших через колонку фракций (100 мкл каждая) и концентрировали до исходных объемов сыворотки 50 мкл посредством фильтрации через мембраны (Microcon YM-10, 10000 MWCO, Millipore Corp., Bedford, MA). Основываясь на ELISA, посредством колонки GNA1870 были удалены более чем 98-99% антител против GNA1870.
Антитело против GNA1870. Для измерения титров антител сыворотки против GNA1870 использовали ELISA, которую проводили, как описано ранее (Welsch et al. J Immunol. 2004; 172:5606-1). Антиген твердой фазы состоял из белков rGNAl870 v.1 или v.2. Вторичное антитело представляло собой конъюгированное с щелочной фосфатазой кроличье антитело против IgM+G+A мыши (Zymed) в разведении 1:2000. Титр сыворотки определяли как разведение, приводящее к OD405 величиной 0,5 после инкубации с субстратом в течение 30 минут.
Опосредуемая комплементом бактерицидная активность антител. Анализ на предмет бактерицидности проводили, как описано ранее (Moe et al. 2002, выше), с использованием бактерий в середине логарифмической фазы роста, выращиваемых в жидкой питательной среде Mueller-Hinton, которая дополнена 0,25% глюкозой. Конечная реакционная смесь содержала различные разведения тестируемой сыворотки, 20% (об./об.) комплемента человека и буфер Gey, содержащий 1% BSA. Источником комплемента являлась сыворотка человека от здорового взрослого, не обладающая поддающейся выявлению собственной бактерицидной активностью (Granoff et al. J. Immunol. 1998; 160:5028-36; Welsch et al. 2003, выше). Бактерицидные титры сыворотки определяли как разведение сыворотки, приводящее к снижению на 50% величины CFU в мл после 60 минут инкубации бактерий в реакционной смеси в сравнении с контрольной величиной CFU в мл в момент времени 0. Как правило, для бактерий, инкубируемых со служащими в качестве отрицательного контроля антителом и комплементом, наблюдают повышение величины CFU/мл от 150 до 200% в ходе инкубации в течение 60 минут.
Связывание антител с поверхностью живых, имеющих капсулу N. meningitidis. Способность антител против GNA1870 связываться с поверхностью живых N. meningitidis определяли анализом непрямой флуоресценции при регистрации проточной цитометрией, как описано ранее (Granoff et al. J Immunol. 2001; 167:3487-3496). Положительные контроли включали в себя моноклональные антитела мыши, специфичные к полисахариду капсулы группы C (1076.1 (Garcia-Ojeda et al. Infect. Immun. 2000; 68:239-46)), PorA P1.2 (Granoff et al. J Immunol. 2001; 167:3487-3496) и к варианту 1 GNA1870 (JAR3) (Welsch et al. J Immunol. 2004; 172:5606-15), а также конъюгированные с FITC козьи антитела против (Fab')2 IgG (H+L) мыши при разведении 1:300 (Jackson Immuno Research Laboratories, West Grove, PA).
Активация отложения комплемента человека на поверхности живых, имеющих капсулу менингококков. Зависимое от антитела против GNA1870 отложение C3b или iC3b на бактериальной поверхности живых бактерий N. meningitidis определяли проточной цитометрией, которую проводили, как описано выше (Welsch et al. J Infect. Dis. 2003; 188:1730-40). Промытые бактерии в фазе логарифмического роста инкубировали в реакционной смеси, содержащей 5% (об./об.) комплемента человека и соответствующие разведения сыворотки в буфере из веранола. Отложение комплемента регистрировали с использованием конъюгированного с FITC козьего антитела против комплемента человека C3c (BioDesign Intl., Saco, ME), которое реагирует с C3b и iC3b. Источник комплемента представлял собой такую же сыворотку человека, как описанная выше для анализа на предмет бактерицидности.
Пассивная защита у детенышей крыс. Способность антисыворотки придавать пассивную защиту против бактериемии, обусловленной N. meningitidis группы B, тестировали у детенышей крыс, у которых иммунную реакцию вызывали IP способом с использованием штамма NZ98/254 группы B (Welsch et al. 2003, выше; Moe et al. Infect. Immun. 1999; 67:5664-75; Moe et al. Infect Immun 2001; 69:3762-71). Вкратце, детенышей крыс в возрасте 4 суток из помета аутбредных крыс Wistar (Charles River, Hollister, CA) перераспределяли случайным образом среди кормящих матерей. В момент времени 0 группам из восьми животных вводили IP антисыворотку или антитела, разведенные в PBS, который содержит 1% BSA. Через два часа у животных вызывали иммунную реакцию IP способом с использованием приблизительно 6Ч104 CFU промытых бактерий в фазе логарифмического роста в Mueller-Hinton, дополненной 0,25% глюкозы и 10 мкМ CMP-NANA (Sigma, St. Louis, MO). Через период от четырех до шести часов после вызова бактериями иммунной реакции получали посредством пункции сердца образцы крови и аликвоты крови 1, 10 и 100 мкл высевали на шоколадный агар для определения CFU/мл.
ПРИМЕР 1
ПОВЕРХНОСТНАЯ ДОСТУПНОСТЬ GNA1870 НА ШТАММЕ RM1090 N. MENINGITIDIS
Для определения того, является ли белок GNA1870, который экспрессируется трансформированным pFP12-GNA1870 штаммом RM1090, составной частью наружной мембраны и представлен ли на клеточной поверхности, а также для определения того, является ли сверхэкспрессируемый GNA1870 в штамме H44/76 прикрепленным и поверхностно доступным в наружной мембране, измеряли проточной цитометрией связывание антител против GNA1870 и контрольных антител с живыми, имеющими капсулу бактериальными клетками (фиг.1).
Как представлено на фиг.1A, для служащих положительным контролем mAb, специфичных к полисахариду капсулы группы C (столбец 2) или к PorA (против P1.2, столбец 3), наблюдали значительное связывание с родительским штаммом RM1090 (ряд B) и двумя мутантными штаммами RM1090: нокаутным по GNA1870, трансформированным "челночным" вектором без гена GNA1870 (ряд A), и нокаутным, трансформированным "челночным" вектором, который кодирует вариант 1 белка GNA1870. Для всех трех штаммов не наблюдали значительного связывания со служащей отрицательным контролем совокупностью сывороток от мышей, иммунизированных только фосфатом алюминия, при разведении 1:10 (столбец 1). Также не наблюдали значительного связывания моноклональных или поликлональных антител против GNA1870 со штаммом, нокаутным по GNA1870 (ряд A, столбцы 5 и 6 соответственно). Штамм RM1090 дикого типа, в природе экспрессирующий белок GNA1870 v.2 на низких уровнях, не проявлял поддающегося выявлению связывания с mAb против GNA1870, специфичным к белку v.1 (ряд B, столбец 4), а также проявлял минимальное связывание, превышающее фоновый уровень, с поликлональной антисывороткой мыши (столбцы 5 и 6), получаемой против рекомбинантных белков GNA1870 v.1, 2 и 3 (см. ниже). В отличие от этого для штамма, трансформированного "челночным" вектором, который кодирует GNA1870 (вариант 1), наблюдали значительное связывание с поликлональными и моноклональными антителами против GNA1870. Таким образом, GNA1870 представлен на поверхности штамма RM1090, трансформированного "челночным" вектором pFP12-GNA1870.
Как представлено на фиг.1B, служащие положительным контролем моноклональные антитела против капсулы и PorA (P1.16) связывались со штаммом H44/76 дикого типа и мутантным штаммом H44/76, сверхэкспрессирующим GNA1870 (оба представлены в ряду 1). Также служащие положительным контролем антитела связывались с H44/76ΔGNA1870 (представлено в ряду 2). В соответствии с ожидаемым не наблюдали связывания моноклональных или поликлональных антител против GNA1870 с мутантным штаммом H44/76, в котором был инактивирован ген, кодирующий GNA1870 (столбцы от D до F). При инкубации штамма дикого типа с антителами против GNA1870 наблюдали значительное связывание, результат, который отражает относительно высокий уровень природной экспрессии GNA1870 в штамме H44/76. Существовало небольшое увеличение связывания с мутантным штаммом, сконструированным для сверхэкспрессии GNA1870, о чем свидетельствует незначительный сдвиг вправо иммунной флуоресценции. Таким образом, сверхэкспрессия GNA1870 приводила к незначительному повышению количества белка в наружной мембране, и белок является представленным на поверхности.
ПРИМЕР 2
АНАЛИЗ ВАКЦИНЫ OMV
Основные белки в препаратах OMV из штамма RM1090 и соответствующих мутантов разделяли посредством SDS-PAGE и наблюдали окрашиванием с использованием Кумасси синего (фиг.2A, панель A). Как характерно для OMV, получаемых из N. meningitidis, наблюдали ограниченное количество основных белков, разделяющихся при наблюдаемых массах от 29 кДа (Opa/Opc) до 43 кДа (PorA). OMV, получаемые из штамма дикого типа (дорожка 1) и нокаутного по GNA1870 штамма (дорожка 3) экспрессировали сходные соответствующие количества каждого из этих белков. В отличие от этого для OMV из штаммов, трансформированных "челночным" вектором pFP12, который не содержал кодирующий GNA1870 ген (дорожки 2 и 4 соответственно), наблюдали сниженную относительную экспрессию трех белков, перемещающихся с наблюдаемыми массами от 38 до 43 кДа. Вероятно, этот результат отчасти отражает сниженную экспрессию белков поринов в результате селекции посредством антибиотика при наличии 5 мкг/мл хлорамфеникола в среде для роста (Tommassen et al. Infect. Immun. 1990; 58:1355-9). На дорожке 5 представлены OMV, получаемые из штамма RM1090, трансформированного кодирующим GNA1870 "челночным" вектором. Для лучшего наблюдения белков эту дорожку загружали в 2 раза большим количеством белка (приблизительно 10 мкг), чем дорожки 1-4. В сравнении с другими препаратами OMV для OMV, получаемой из штамма, который трансформировали содержащим ген GNA1870 "челночным" вектором, наблюдали сниженную экспрессию белков, разделяющихся между 29 и 32 кДа. При SDS PAGE в любом из препаратов OMV, в том числе OMV, получаемой из мутантного штамма, который сверхэкспрессирует GNA1870 (дорожка 5), практически не наблюдают GNA1870. (Для сравнения на дорожке 6 представлен 1 мкг рекомбинантного варианта белка GNA1870).
На фиг.2A для оценки экспрессии GNA1870 в различных препаратах вакцин использовали Вестерн-блоттинг с применением поликлональной мышиной антисыворотки, полученной против рекомбинантных белков GNA1870 v.1, 2 и 3. Как представлено на панели B, антисыворотка была незначительно более реакционноспособна с белком rGNA1870 v.2, чем с рекомбинантным белком v.1. Даже при таком отклонении OMV, получаемая из RM1090, который трансформируют pFP12-GNA1870, проявляла повышенную реакционную способность при Вестерн-блоттинге в сравнении с OMV, получаемой из штамма RM1090 дикого типа, который в природе экспрессирует белок v.2 (фиг.2A, панель C). В отличие от этого служащая отрицательным контролем OMV из нокаутного по GNA1870 мутанта (RM1090ΔGNA1870) не обладала поддающейся выявлению реакционной способностью. Результаты денситометрических измерений указывают на то, что экспрессия белка GNA1870 v.1 в штамме, трансформированном "челночным" вектором, была приблизительно в 10 раз выше, чем таковая для белка v.2, экспрессируемого в природе родительским штаммом RM1090 дикого типа.
Препараты OMV H44/76 анализировали Вестерн-блоттингом с использованием поликлональной антисыворотки против GNA1870 (фиг.2B). Загружаемое в гель количество OMV было стандартизировано на основе общего содержания белка в препаратах. В соответствии с ожидаемым GNA1870 экспрессировался в препаратах мембран из штамма дикого типа и его количество было повышено в соответствующем препарате из мутанта, сконструированного для сверхэкспрессии этого белка. Однако повышение количества GNA1870 было незначительным (приблизительно в 3 раза).
ПРИМЕР 3
АНАЛИЗ ИММУННОГО ОТВЕТА СЫВОРОТКИ
В таблице 2 и на фиг.5 обобщены данные для иммунного ответа сыворотки против GNA1870 для различных групп мышей, как определено посредством ELISA. Наиболее высокие иммунные ответы в отношении белка варианта 1 в таблице 2 были у мышей, иммунизированных только вакциной рекомбинантного GNA1870 v.1 или вакциной рекомбинантного GNA1870 v.1, вводимой в виде смеси с вакциной OMV (титры против белка варианта 1 составляют 1:120000 и 1:300000 соответственно). Мыши, иммунизированные OMV, которую получают из сверхэкспрессирующего вариант 1 GNA1870 штамма RM1090, имели меньший в 4-10 раз титр антител против GNA1870 (1:32000). Интересно, что у мышей, иммунизированных OMV, которую получают из штамма RM1090 дикого типа, не наблюдали или наблюдали пренебрежимо малые иммунные ответы против GNA1870, как измерено против белков варианта 1 или 2. Этот результат позволяет предположить, что GNA1870 в OMV из штамма дикого типа является недостаточно иммуногенным при отсутствии сверхэкспрессии.
Группы мышей иммунизировали с использованием OMV H44/76 (1,25 мкг общего белка) или 5 мкг rGNA1870, вводимых с фосфатом алюминия. Образцы сыворотки получали через 3 недели после третьей дозы и объединяли в совокупность (2 совокупности на группу вакцин, каждую совокупность получают от 4-5 мышей). Как представлено на фиг.5, контрольные мыши, иммунизированные только адъювантом из алюминия, не имели поддающихся выявлению антител против GNA1870 (GMT <1:10, столбец 1), тогда как мыши, иммунизированные посредством rGNA1870, проявляли наиболее сильные реакции (GMT 1:23500, столбец 2). Мыши, иммунизированные OMV, которую получают из сверхэкспрессирующего GNA1870 H44/76, имели в 10 раз более высокие реакции антител против GNA1870, чем соответствующая группа, иммунизированная OMV из штамма дикого типа (для сравнения столбцы 5 и 3). Мыши, иммунизированные OMV, которую получают из H44/76ΔGNA1870, обладали пренебрежимо малыми реакциями антител (GMT <1:10, столбец 4).
Реакции мышиных антител против GNA1870, как измерено посредством ELISA
bРазведение сыворотки при ELISA, приводящее к OD 0,5 через 30 минут к инкубации с субстратом. Представленные данные являются соответствующими средними геометрическими значениями титров, измеряемых в 2 совокупностях сывороток из каждой группы вакцин. Каждая совокупность содержала равные объемы сыворотки от 4-5 иммунизированных мышей.
На фиг.3A обобщены данные для реакций бактерицидных антител сыворотки для различных групп мышей, как измерено против четырех тестируемых штаммов. У мышей, иммунизированных только вакциной рекомбинантного белка GNA1870 или вакциной рекомбинантного GNA1870 в сочетании с вакциной OMV, или OMV, сверхэкспрессирующей GNA1870, образовывались высокие бактерицидные титры против штамма Cu385, которые друг от друга значительно не отличались (для сравнения столбцы 4, 5 и 6 на верхней панели). В отличие от этого не наблюдали поддающейся выявлению бактерицидной активности против штамма Cu385 для сыворотки от контрольных мышей, иммунизированных вакцинами OMV, которые получают из штамма RM1090 дикого типа или нокаутного по GNA1870 штамма (столбцы 2 и 3 соответственно; титры <1:10). Следует отметить, что штамм Cu385 экспрессирует канонический белок NA1870 v.1 (аминокислотная последовательность совпадает с таковой для штамма MC58, где ген используют для экспрессии рекомбинантного белка GNA1870), и для этого штамма из предыдущих исследований авторов изобретения была известна высокая чувствительность к бактерицидной активности антитела, образование которого у мышей вызвано вакциной рекомбинантного GNA1870 (таблица 1). Кроме того, Cu385 имел сероподтип PorA (P1.19,15), гетерологичный таковому для штамма для вакцины RM1090 (P1.5,2), и поэтому от штамма Cu385 ожидали устойчивости к бактерицидной активности антител, образованных против контрольной вакцины OMV, которая не экспрессировала вариант 1 GNA1870 (Tappero et al. JAMA 1999; 281:1520-7; Moe et al. 2002, выше).
На фиг.3A также представлены соответствующие бактерицидные титры сыворотки, измеряемые против штамма M6190 (вторая панель сверху), который экспрессирует субвариант белка v.1 GNA1870, в сравнении с таковыми для сконструированного штамма для вакцины. Поддающейся выявлению бактерицидной активности не наблюдали для сыворотки от мышей, иммунизированных рекомбинантным белком варианта 1 GNA1870 (столбец 6, среднее геометрическое значение титра <1:10), результат, аналогичный результату предыдущего исследования авторов изобретения (таблица 1). Однако, поскольку сероподтип PorA (P1.5,2) штамма M6190 гомологичен сероподтипу штамма для вакцины RM1090, сыворотка от мышей, иммунизированных любой из содержащих OMV вакцин, являлась высоко бактерицидной (столбцы 2, 3, 4 или 5).
На фиг.3A (третья и четвертая панели сверху) представлены соответствующие бактерицидные реакции против штаммов Z1092 и NZ98/254 соответственно. Оба штамма экспрессируют молекулы PorA, гетерологичные молекулам из штамма для вакцины RM1090 (таблица 1), и они не уничтожались сывороткой от мышей, иммунизированных вакцинами OMV, которые получают из штамма RM1090 дикого типа или нокаутного по GNA1870 штамма (столбцы 2 и 3, средние геометрические значения титров <l:10). Однако мыши, иммунизированные вакциной OMV, которую получают из сверхэкспрессирующего GNA1870 штамма RM1090 (столбец 5), обладали значительно более высоким средним геометрическим значением титра бактерицидных антител против штамма Z1092, чем таковой для мышей, иммунизированных рекомбинантным GNA1870 (столбец 6, P<0,02) или смесью рекомбинантного белка GNA1870 и вакцины OMV (столбец 4, P<0,04). Сходные тенденции наблюдали для соответствующих бактерицидных реакций сыворотки, измеряемых против штамма NZ98/254 (нижняя панель) или против штаммов BZ198 и M1390 (данные не представлены). Однако для этих последних трех штаммов величины бактерицидных реакций сыворотки от мышей, иммунизированных вакциной OMV со сверхэкспрессируемым GNA1870, были ниже, чем величины, измеряемые против штамма Z1092. Кроме того, средние геометрические значения бактерицидных титров сыворотки против штаммов NZ98/294, BZ198 и M1390 у мышей, иммунизированных посредством OMV, которая сверхэкспрессировала GNA1870, статистически значимо не отличались при сравнении с соответствующими средними геометрическими значениями титров для мышей из других групп вакцин (P>0,10).
На фиг.3B обобщены данные для реакций бактерицидных антител сыворотки против шести штаммов, в том числе H44/76, которые использовали для получения вакцины OMV. Пять из шести штаммов имели сероподтипы PorA, гомологичные таковым для H44/76. Штамм H44/76 также экспрессировал белок варианта 1 GNA1870 с последовательностью, совпадающей с таковой для штамма MC58, который содержит ген, используемый для клонирования и экспрессии рекомбинантного белка GNA1870 для вакцины. Все препараты вакцин, за исключением служащего отрицательным контролем адъюванта из алюминия, вызывали значительные реакции бактерицидных антител сыворотки при измерении против штамма для вакцины H44/76 (фиг.3B, панель A). В отличие от этого при измерении против гетерологичных штаммов 4243 (панель B), Z1092, NZ98/254 и BZ198 (панель C) или M6190 (панель D) сыворотка от мышей, иммунизированных вакциной rGNA1870 или вакцинами OMV, которые получают из штаммов H44/76 дикого типа или H4476ΔGNA1870, имела низкие или не поддающиеся выявлению бактерицидные титры (столбцы 2, 3 и 4 соответственно). Мыши, иммунизированные вакциной OMV со сверхэкспрессируемым GNA1870 (столбец 5), обладали значительными реакциями бактерицидных антител против штамма 4243, низкими, но поддающимися выявлению бактерицидными реакциями против штаммов NZ98/254, BZ198 и Z1092, и не поддающейся выявлению бактерицидной активностью против M6190 (титр <1:10). Хотя на фиг.3B не представлено, все штаммы легко уничтожались комплементом вместе со служащими положительным контролем антителами против соответствующего PorA и/или полисахарида капсул.
ПРИМЕР 4
АКТИВАЦИЯ ОТЛОЖЕНИЯ КОМПЛЕМЕНТА C3B НА ПОВЕРХНОСТИ ЖИВЫХ, ИМЕЮЩИХ КАПСУЛУ КЛЕТОК N. MENINGITIDIS
В предыдущих исследованиях авторами изобретения было открыто, что определенные мышиные антитела против менингококков, у которых отсутствует бактерицидная активность, придавали пассивную защиту против менингококковой бактериемии при отсутствии бактерицидной активности (Welsch et al. J Immunol. 2004; 172:5606-15; Welsch et al. 2003, выше). Защита соотносилась со способностью антител активировать отложение компонентов комплемента C3 на поверхности живых, имеющих капсулу менингококков, как измерено посредством проточной цитометрии. Присутствие C3b обеспечивает лиганд для опсонизации, которая является наиболее вероятным механизмом, сообщающим защиту в отсутствие бактерицидной активности. Таким образом, проводили исследование способности антисыворотки от мышей, иммунизированных различными вакцинами OMV, активировать отложение C3b человека (фиг.4). Для этих экспериментов использовали два тестируемых штамма N. meningitidis, NZ98/254 (фиг.4A, ряд A) и M1390 (фиг.4A, ряд B), и четыре тестируемых штамма N. meningitidis, NZ98/254, BZ198, Z1092 и M6190 (фиг.4B). Это были штаммы, для которых антисыворотка от мышей, иммунизированных вакциной OMV со сверхэкспрессией GNA1870, не обладала статистически значимо более высокими бактерицидными титрами, чем другие группы вакцин.
Признаков отложения комплемента не наблюдали, если бактериальные клетки какого-либо из тестируемых штаммов инкубировали с источником комплемента человека вместе с разведением 1:40 служащей в качестве отрицательного контроля совокупности сывороток от мышей, иммунизированных только фосфатом алюминия (закрашенные области панелей на фиг.4A, столбец 1). Сходным образом, не наблюдали поддающегося выявлению отложения C3b с инактивированным посредством нагревания комплементом вместе с 5 мкг/мл мышиного моноклонального антитела против GNA1870 (JAR 3) (закрашенные области панелей на фиг.4A, столбец 2). В отличие от этого добавление активного комплемента к 25 мкг/мл служащего положительным контролем моноклонального антитела против капсулы группы B (открытые области панелей на фиг.4A, столбец 1) или к 1 мкг/мл моноклонального антитела против GNA1870 (открытые области панелей на фиг.4A, столбец 2) вызывало значительное отложение C3b на бактериальной поверхности обоих тестируемых штаммов, о чем свидетельствует увеличение процентной доли бактерий, у которых наблюдают высокую иммунофлуоресценцию с антителом против C3c, распознающим C3b и iC3bi.
На панелях в столбцах 3-6 на фиг.4A представлен эффект добавления комплемента к разведениям совокупностей сывороток, полученных от групп иммунизированных мышей, которых иммунизировали различными вакцинами. Добавление комплемента к разведению 1:100 сыворотки от мышей, иммунизированных рекомбинантным GNA1870 (столбец 3) или OMV, получаемой из штамма RM1090 дикого типа (столбец 4), или OMV, смешанной с рекомбинантным GNA1870 (столбец 5), не активировало отложение C3b на каком-либо из тестируемых штаммов. В отличие от этого разведения 1:100 или 1:400 совокупности сывороток от мышей, иммунизированных OMV, которую получают из штамма RM1090 со сверхэкспрессией GNA1870, активировали значительное отложение C3b в отношении обоих тестируемых штаммов (столбец 6).
Как представлено на фиг.4B, служащие положительным контролем mAb против капсулы вызывали отложение комплемента на каждом из штаммов (открытые области на столбце A), тогда как разведение 1:100 служащей отрицательным контролем антисыворотки от мышей, иммунизированных только адъювантом из алюминия, приводило к отрицательному результату (закрашенные области на столбце A). Разведение 1:100 сыворотки от мышей, иммунизированных вакциной rGNA1870 (закрашенные области на столбце B) или вакциной OMV H44/76, получаемой из штамма дикого типа (закрашенные области на столбце C), также не вызывало значительного отложения комплемента на каком-либо из штаммов. В отличие от этого mAb против rGNA1870 вызывало отложение комплемента на штаммах NZ98/254, BZ198 и Z1092, но не на штамме M6190 (открытые области на столбце B). Сходным образом, разведение 1:100 антисыворотки от мышей, иммунизированных вакциной H44/76 со сверхэкспрессией GNA1870, активировало отложение C3 в случае штаммов Z1092, NZ98/254 и BZ198 (открытые области на столбце D), но не в случае штамма M6190.
ПРИМЕР 5
ОПРЕДЕЛЕНИЕ АНТИГЕННОЙ МИШЕНИ АНТИТЕЛ, КОТОРЫЕ ЯВЛЯЮТСЯ БАКТЕРИЦИДНЫМИ ИЛИ АКТИВИРУЮТ ОТЛОЖЕНИЕ C3B НА ГЕТЕРОЛОГИЧНЫХ ШТАММАХ
Колонку для аффинной хроматографии Ni-NTA, загруженную рекомбинантным GNA1870 с меткой His, использовали для абсорбции антител против GNA1870 из совокупности сывороток, получаемых от мышей, которых иммунизировали OMV H44/76 со сверхэкспрессией GNA1870. Как представлено в таблице 3, при ELISA на этой колонке удалялось 98% антител против GNA1870 в сравнении с сывороткой, абсорбируемой на служащей отрицательным контролем колонке, которая содержит только матрицу Ni-NTA. После абсорбции на колонке, служащей отрицательным контролем, бактерицидная активность против штамма 4243 была сходной с таковой для исходной неабсорбированной совокупности сывороток, тогда как абсорбция антител против GNA1870 приводила к полной утрате бактерицидной активности.
Эффект абсорбции антител против GNA1870 на отложение C3 анализировали в отношении штаммов Z1092, NZ98/254, BZ198 и M6190 (таблица 3 и фиг.4B, ряд 4). Как представлено на фиг.4B, столбец D, удаление антител против GNA1870 из сывороток от мышей, иммунизированных OMV H44/76 со сверхэкспрессией GNA1870, приводило к полной утрате способности антисывороток активировать отложение комплемента в трех штаммах, чувствительных к активации и отложению iC3b/C3b (закрашенные области на фиг.4B). Эти результаты, а также данные по бактерицидности для абсорбированных сывороток, обобщенные выше, указывают на то, что в случае штаммов с белками PorA, гетерологичными таковым из штамма для вакцины H44/76, активация отложения C3 и бактерицидная активность антисывороток, получаемых против OMV H44/76, содержащей сверхэкспрессируемый GNA1870, опосредуются антителами против GNA1870.
Активность сывороток от мышей, иммунизированных OMV со сверхэкспрессируемым GNA1870, после абсорбции антител против GNA1870
Совокупность, получаемую из сывороток от мышей, которых иммунизировали OMV H44/76 со сверхэкспрессируемым GNA1870, абсорбировали на колонке, содержащей матрицу Ni-NTA (Qiagen), которую инкубировали в течение ночи с 50 мкг/мл рекомбинантного GNA1870 с меткой His. Проходящую жидкость собирали и концентрировали до исходного объема. Контрольная колонка содержала матрицу Ni-NTA без белка с меткой His.
ПРИМЕР 6
РОЛЬ АНТИТЕЛА ПРОТИВ GNA1870 В ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ
Для вакцины OMV, получаемой из штамма RM1090 N. meningitidis, который сконструирован для сверхэкспрессии GNA1870, наблюдали сниженную экспрессию нескольких других белков клеточной оболочки в сравнении с соответствующими белками в OMV, получаемой из штамма для вакцины RM1090 дикого типа или нокаутного штамма RM1090ΔGNA1870 (фиг.2A, панель A). Таким образом, существовала возможность, что более высокая функциональная активность антисывороток от мышей, иммунизированных OMV со сверхэкспрессией GNA1870, была обусловлена антителами, образование которых вызвано антигенами, отличными от GNA1870. Для исследования этой возможности совокупность сывороток от мышей, иммунизированных OMV RM1090 со сверхэкспрессией GNA1870, абсорбировали с использованием колонки для аффинной хроматографии против GNA1870. Посредством ELISA было удалено 99% антител против GNA1870. Получаемая абсорбированная антисыворотка также утрачивала всю способность активировать отложение C3b человека на штамме NZ98/294 N. meningitidis (таблица 4). В отличие от этого не наблюдали эффекта на отложение C3b в результате абсорбции совокупности сывороток на колонке для аффинной хроматографии против NadA, служащей в качестве отрицательного контроля (таблица 4).
Функциональная активность антисыворотки от мышей, иммунизированных OMV со сверхэкспрессией GNA1870, после извлечения антител против GNA1870a
bРазведение сыворотки в анализе проточной цитометрией на предмет активации комплемента, которое вызывало 10-кратное повышение иммунофлуоресценции в сравнении с сывороткой, служащей отрицательным контролем (см. фиг.4).
Также в таблице 4 обобщены бактерицидные титры абсорбированных совокупностей сывороток, как измерено против штаммов Cu385 и M6190. Абсорбция антител против GNA1870 полностью устраняла бактерицидную активность против штамма Cu385, но не оказывала значительный эффект на титр против штамма M6190. Этот последний результат был ожидаем, поскольку штамм M6190 экспрессирует PorA с сероподтипом, гомологичным PorA, экспрессируемому штаммом для вакцины RM1090, а бактерицидные антитела против PorA не устраняются посредством колонок для аффинной хроматографии с GNA1870 или NadA.
ПРИМЕР 8
ПАССИВНАЯ ЗАЩИТА НА МОДЕЛИ МЕНИНГОКОККОВОЙ БАКТЕРИЕМИИ У ДЕТЕНЫШЕЙ КРЫС
Детенышей крыс предварительно обрабатывали совокупностями сывороток от различных групп мышей и через 2 часа вызывали иммунную реакцию посредством штамма NZ98/254 N. meningitidis. На фиг.6 представлены средние геометрические значения CFU/мл в крови, получаемой через 4-6 часов после вызова иммунной реакции. У всех 10 крыс, обработанных разведением 1:15 совокупности сывороток от служащих отрицательным контролем мышей, которых иммунизировали только фосфатом алюминия, наблюдали бактериемию со средним геометрическим значением CFU/мл ~105 (панель A, столбец 1). В отличие от этого предварительная обработка посредством 10 мкг/крыса служащего положительным контролем антитела против капсулы группы B (столбец 2) или моноклонального антитела против GNA1870 (столбец 3) приводила к 3-4-кратно более низкому в логарифмическом масштабе среднему геометрическому значению CFU/мл (P<0,0001). При сравнении с животными, обрабатываемыми служащей отрицательным контролем сывороткой, не наблюдали значительной пассивной защитной активности совокупностей сывороток от мышей, иммунизированных вакциной OMV, которую получали из нокаутного штамма RM1090ΔGNA1870 (столбец 4), или вакциной OMV, смешанной с рекомбинантным GNA1870 (столбец 5). В отличие от этого защиту придавала совокупность сывороток от мышей, иммунизированных вакциной OMV со сверхэкспрессируемым GNA1870 (столбец 6) (снижение среднего геометрического значения CFU/мл в 4 раза в логарифмическом масштабе, P<0,0001). Совокупность сывороток от мышей, иммунизированных только вакциной рекомбинантного GNA1870 (столбец 7), придавала незначительную защиту (снижение в ~2 раза в логарифмическом масштабе, P<0,0001), однако защитная активность была ниже таковой для совокупности сывороток от мышей, иммунизированных OMV со сверхэкспрессируемым GNA1870 (P<0,0001, в сравнении с соответствующими средними геометрическими значениями CFU/мл).
На фиг.6, панели B, представлены соответствующие средние геометрические значения CFU/мл для крыс, которых предварительно обрабатывали совокупностями сывороток при разведениях 1:60. При этом более высоком разведении совокупность сывороток от мышей, иммунизированных вакциной OMV со сверхэкспрессией GNA1870 (столбец 6), сообщала защиту (P<0,0002 в сравнении со средним геометрическим значением для крыс, которых обрабатывали служащей отрицательным контролем сывороткой при разведении 1:15), однако значительная защитная активность в результате более высокого разведения совокупностей сывороток от мышей, иммунизированных какой-либо из других 3 тестируемых групп вакцин, включая сыворотку от мышей, которым вводили вакцину рекомбинантного GNA1870 (столбец 7, P>0,10), отсутствовала.
ПРИМЕР 9
ИММУНИЗАЦИЯ МЫШЕЙ ВАКЦИНОЙ НА ОСНОВЕ ВЕЗИКУЛ, ПОЛУЧАЕМОЙ ИЗ ШТАММА RM1090, КОТОРЫЙ СВЕРХЭКСПРЕССИРУЕТ ПОВЕРХНОСТНЫЙ БЕЛОК A (NspA) NEISSERIAL, НЕ СВЯЗАНА С УСИЛЕНИЕМ РЕАКЦИЙ БАКТЕРИЦИДНЫХ АНТИТЕЛ СЫВОРОТКИ
Представляло интерес определить, являлась ли специфичной к GNA1870 усиленная защита, индуцируемая вакциной на основе везикул, которую получают из сконструированного для сверхэкспрессии GNA1870 штамма RM1090, или она также возникает при использовании вакцин на основе везикул, получаемых из штаммов, которые сконструированы для сверхэкспрессии другой мишени для вакцины. Таким образом, вакцину на основе микровезикул получали из штамма RM1090, в котором был инактивирован ген для NspA в штамме дикого типа. Вторую вакцину на основе везикул получали из нокаутного по NspA штамма RM1090, трансформированного "челночным" вектором pFP12, который содержит ген NspA из штамма 8047. В соответствии с SDS PAGE получаемые везикулы из штамма, трансформированного "челночным" вектором, характеризовались экспрессией белка NspA, повышенной в 10 раз в сравнении со штаммом RM1090 дикого типа (данные не представлены).
Группы мышей иммунизировали посредством 3 доз вакцин на основе везикул, вводимых вместе с фосфатом алюминия, а сыворотку собирали через 3 недели после последней иммунизации. Вакцина со сверхэкспрессией NspA вызывала высокие титры антител против NspA, как измерено посредством ELISA (1:19000 в сравнении с титром 1:700 у мышей, иммунизированных вакциной на основе везикул, которую получают из нокаутного по NspA штамма, и с титром <1:50 у мышей, иммунизированных только фосфатом алюминия). В таблице 5 обобщены данные по реакциям бактерицидных антител сыворотки, как измерено против четырех тестируемых штаммов, BZ198, NZ98/254, Cu385 и Z1090.
Иммунизация мышей вакцинами на основе везикул, получаемыми из мутантного штамма RM1090, который генетически сконструирован для сверхэкспрессии поверхностного белка A Neisserial (NspA);
(1/Титр)с
(1/Титр)с
bНаиболее низкая концентрация, которая приводит к 50% уничтожению бактерий после инкубации с комплементом человека в течение 1 часа.
cНаиболее высокое разведение сыворотки, которое приводит к 50% уничтожению бактерий после инкубации с комплементом человека в течение 1 часа.
dЭкспрессия на фоне нокаута по NspA.
Все четыре штамма экспрессировали PorA, гетерологичным в сравнении с PorA из штамма для вакцины RM1090. Для штамма BZ198, который выбрали в этом эксперименте для тестирования бактерицидной активности, основываясь на предшествующих данных, в которых показана высокая чувствительность к бактерицидной активности антисыворотки против NspA, получаемой против экспрессируемого в везикулах E. coli рекомбинантного NspA (Moe et al., Infection and Immunity 1999; 67:5664-5675), не наблюдали признаков повышенной бактерицидной активности в антисыворотке от мышей, иммунизированных вакциной на основе везикул, которую получают из штамма со сверхэкспрессией NspA. При этом повышение бактерицидной активности отсутствовало против штамма NZ98/254, а для штаммов Cu385 и Z1090 наблюдали признаки того, что иммунизация вакциной на основе везикул, сверхэкспрессирующей NspA, индуцировала в 3-10 раз более низкие реакции бактерицидных антител сыворотки, чем реакции, индуцируемые контрольной вакциной на основе везикул, которую получают из соответствующего штамма с нокаутом по NspA. Таким образом, в отличие от вакцин на основе везикул, сверхэкспрессирующих GNA1870, вакцина на основе везикул, сверхэкспрессирующая NspA, не обеспечивала постоянно повышенные реакции бактерицидных антител и, вероятно, обладала подавленными реакциями бактерицидных антител на некоторые штаммы.
| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛОК NMB1125 И ЕГО ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ | 2004 |
|
RU2336900C2 |
| МУЛЬТИВАЛЕНТНАЯ ВАКЦИНА ИЗ НАТИВНЫХ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ МЕНИНГОКОККОВ, СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2009 |
|
RU2477145C2 |
| БЕЛОК NMB0928 И ЕГО ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ | 2004 |
|
RU2335505C2 |
| МЕНИНГОКОККОВЫЕ ПОЛИПЕПТИДЫ fHBP | 2009 |
|
RU2475496C2 |
| УСОВЕРШЕНСТВОВАНИЯ, КАСАЮЩИЕСЯ ВЕЗИКУЛ ВНЕШНЕЙ МЕМБРАНЫ МЕНИНГОКОККОВ | 2005 |
|
RU2420312C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТРАНСФЕРРИН-СВЯЗЫВАЮЩИЙ БЕЛОК И HSF ИЗ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ | 2003 |
|
RU2359696C2 |
| МЕНИНГОКОККОВЫЕ ПОЛИПЕПТИДЫ fHBP | 2012 |
|
RU2567003C2 |
| НАБОР ДЛЯ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ ПРОТИВ Neisseria meningitidis СЕРОЛОГИЧЕСКОЙ ГРУППЫ В | 2008 |
|
RU2498815C2 |
| МОЛЕКУЛЯРНЫЕ МИМЕТИКИ ЭПИТОПОВ МЕНИНГОКОККА В, КОТОРЫЕ ВЫЗЫВАЮТ ВЫРАБОТКУ ФУНКЦИОНАЛЬНО АКТИВНЫХ АНТИТЕЛ | 2002 |
|
RU2322451C2 |
| СПОСОБ БЕЗДЕТЕРГЕНТНОГО ПОЛУЧЕНИЯ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ ГРАМОТРИЦАТЕЛЬНОЙ БАКТЕРИИ | 2012 |
|
RU2607021C2 |
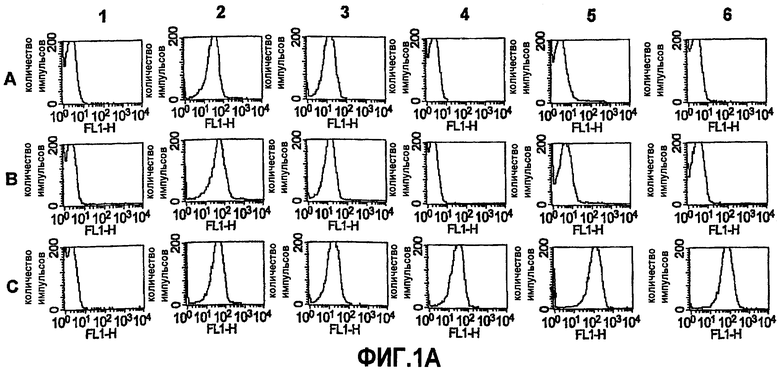
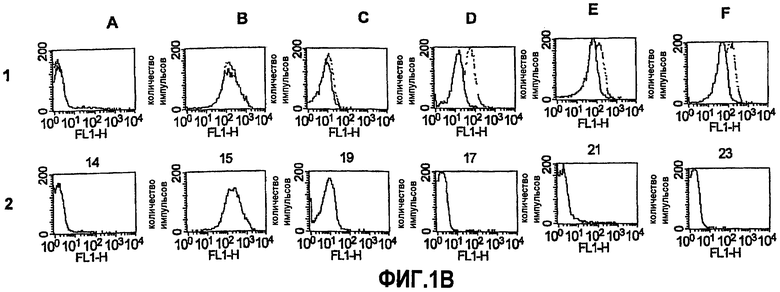
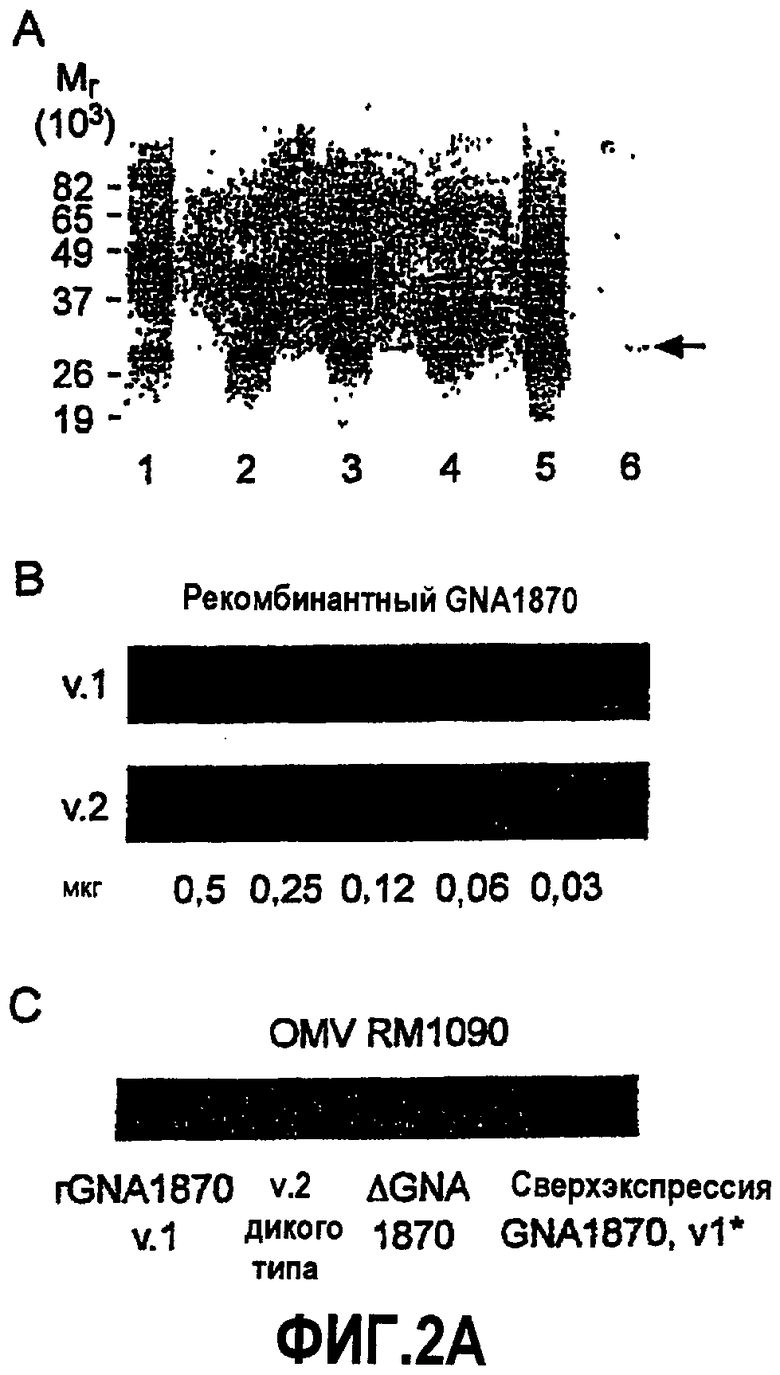

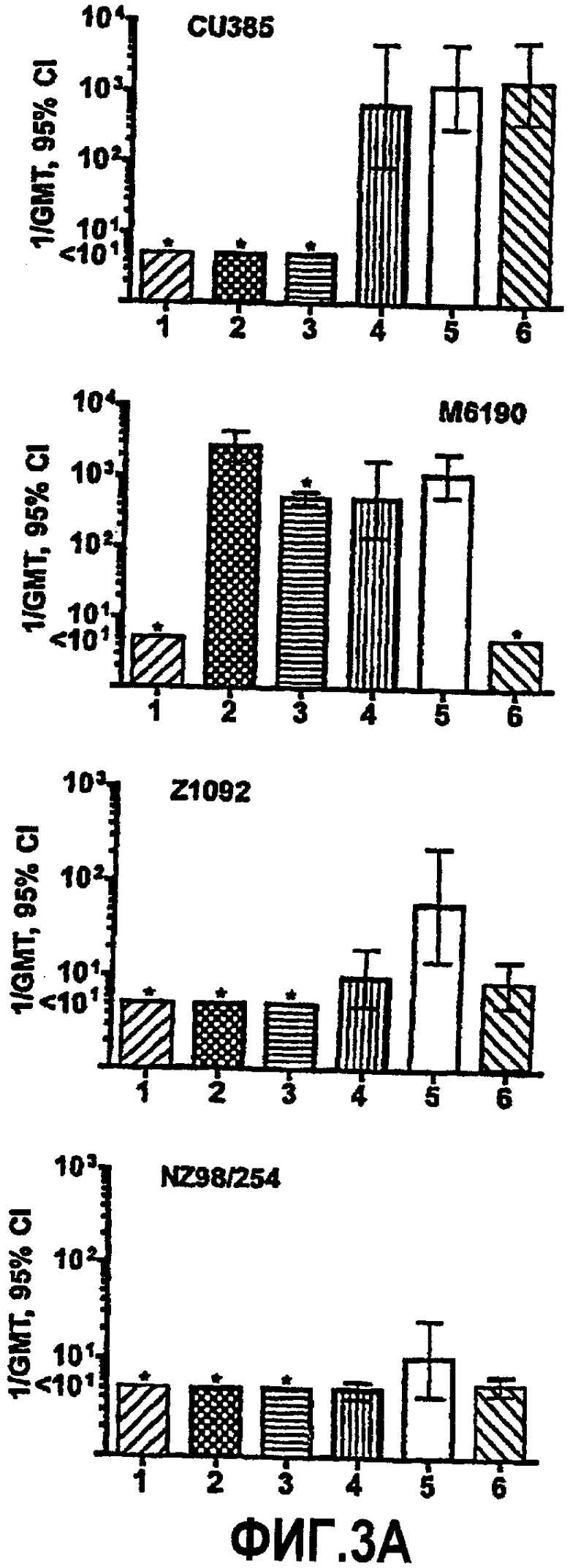
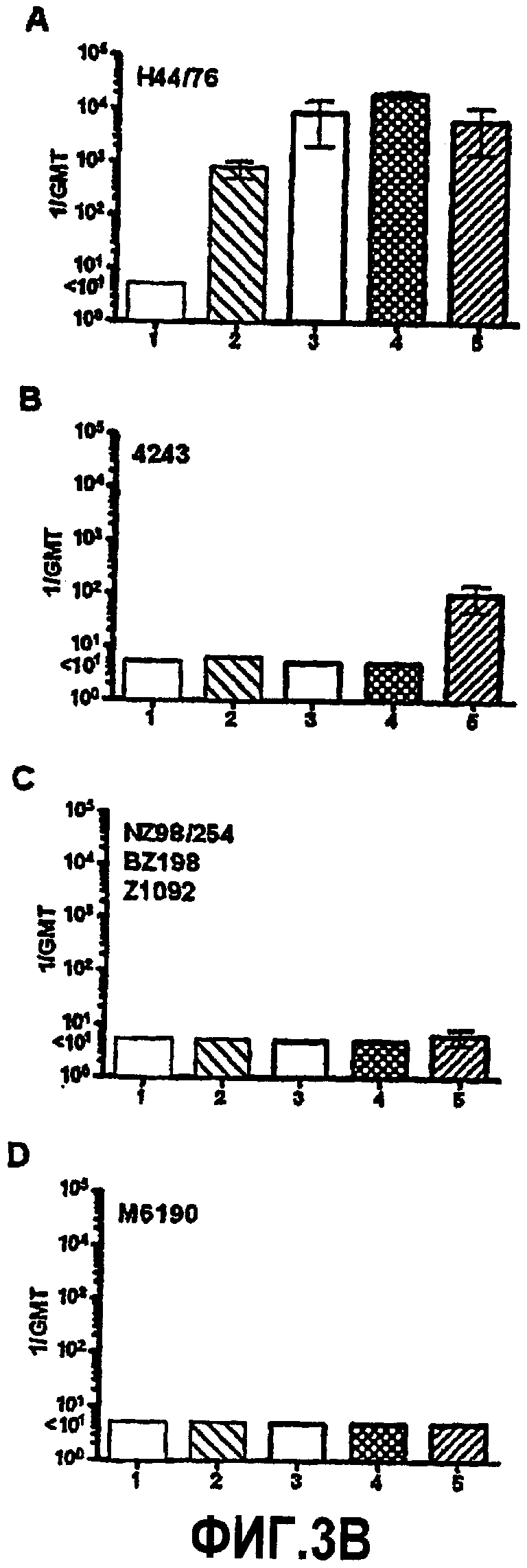
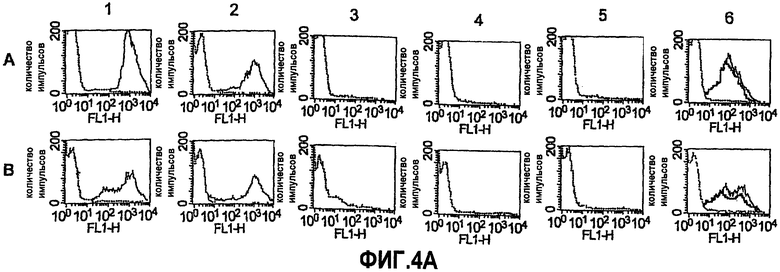
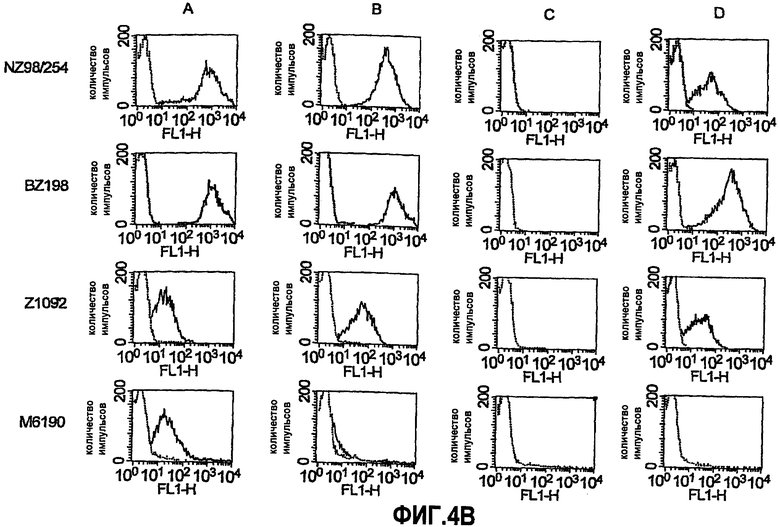
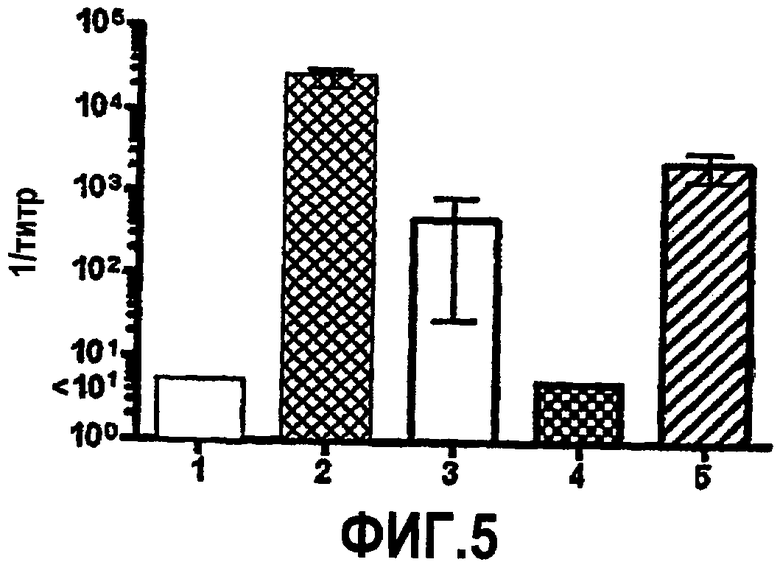
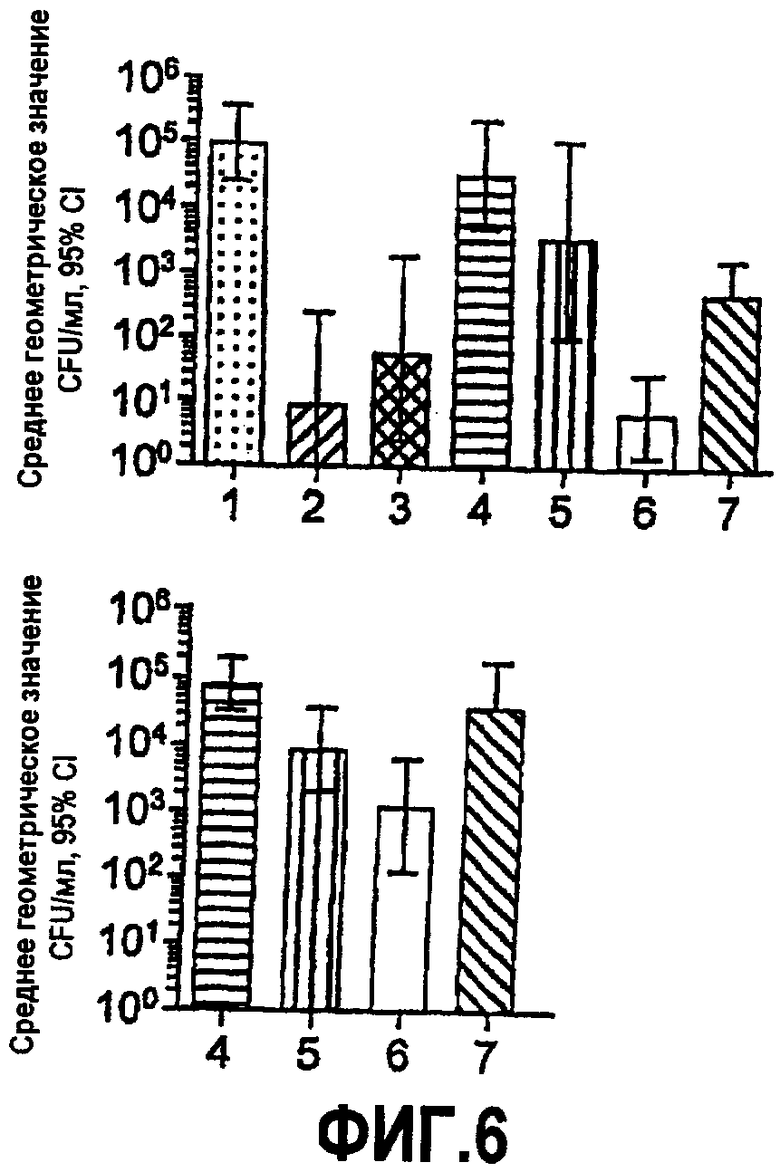








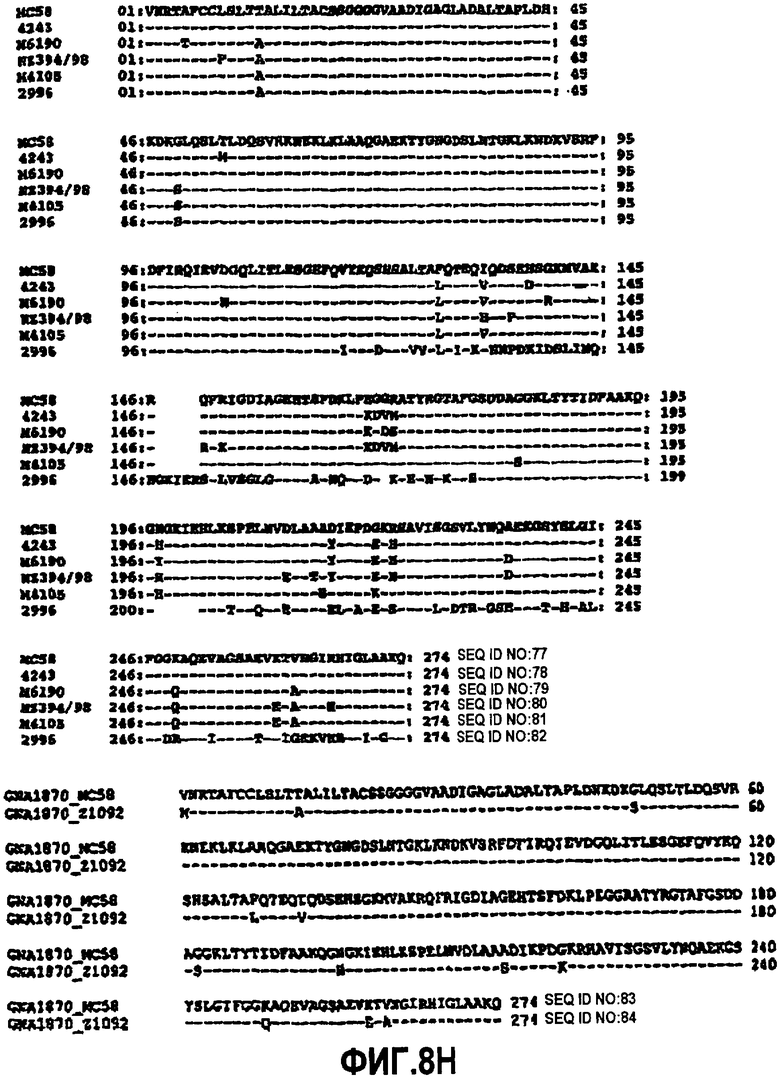

В изобретении раскрывается композиция для вызова иммунной реакции против Neisseria meningitidis на основе везикул, выделенных из бактерии N. meningitidis, у которой инактивирован эндогенный ген GNA 1870 и которая трансформирована генной конструкцией, кодирующей полипептид GNA 1870. Такая бактерия обеспечивает сверхэкспрессию полипептида GNA 1870 в везикулах N. meningitidis. Композиция может дополнительно содержать антигенную везикулу из второй, третьей бактерии N. meningitidis, при условии, что обе бактерии генетически отличаются друг от друга и от вышеуказанной первой бактерии. Полипептиды GNA 1870, экспрессируемые первой, второй и третьей бактериями также генетически отличаются друг от друга. Изобретение описывает способ вызова иммунной реакции против бактерий видов Neisseria путем введения млекопитающему композиции на основе везикул, содержащих полипептид GNA 1870, а также способ получения композиции на основе везикул, полученных в результате культивирования соответствующим образом подготовленных бактерий, путем их смешения с фармацевтически приемлемым носителем. Изобретение обеспечивает защитный иммунитет против широкого спектра штаммов N. meningitidis, в том числе против штаммов N. meningitidis серогруппы В. 3 н. и 42 з.п. ф-лы, 20 ил., 5 табл.
1. Композиция для вызова иммунной реакции против Neisseria meningitidis, содержащая:
выделенные антигенные везикулы, полученные из первой бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, и фармацевтически приемлемый носитель.
2. Композиция по п.1, где везикулы представляют собой везикулы наружной мембраны (OMV), микровезикулы (MV) или смесь OMV и MV.
3. Композиция по п.1, где бактерия Neisseria meningitidis представляет собой Н44/76.
4. Композиция по п.1, где сверхэкспрессируемый полипептид GNA1870 идентичен эндогенному полипептиду GNA1870 бактерии Neisseria meningitidis.
5. Композиция по п.1, где полипептид GNA1870 сверхэкспрессируется с гетерологичного промотора.
6. Композиция по п.1, где сверхэкспрессируемый полипептид GNA1870 является гетерологичным для бактерии Neisseria meningitidis.
7. Композиция по п.1, где бактерия Neisseria meningitidis дефектна по полисахариду капсулы.
8. Композиция по п.1, которая дополнительно содержит:
выделенную антигенную везикулу, полученную из второй бактерии Neisseria meningitidis, где вторая бактерия Neisseria meningitidis генетически отличается от первой бактерии Neisseria meningitidis.
9. Композиция по п.1, которая дополнительно содержит:
выделенную антигенную везикулу, полученную из второй бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная вторая бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, и где вторая бактерия Neisseria meningitidis генетически отличается от первой бактерии Neisseria meningitidis.
10. Композиция по п.8 или 9, где первая и вторая бактерии Neisseria meningitidis генетически отличны тем, что они различаются по крайней мере по одному из серогруппы, серотипа или сероподтипа.
11. Композиция по п.8 или 9, где полипептид GNA1870 из второй бактерии Neisseria meningitidis отличается от полипептида GNA1870, сверхэкспрессируемого в первой бактерии Neisseria meningitidis.
12. Композиция по п.10, которая дополнительно содержит:
выделенную антигенную везикулу, полученную из третьей бактерии Neisseria meningitidis, где третья бактерия Neisseria meningitidis генетически отличается от первой бактерии Neisseria meningitidis.
13. Композиция по п.10, которая дополнительно содержит:
выделенную антигенную везикулу, полученную из третьей бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная третья бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, и где третья бактерия Neisseria meningitidis генетически отличается от первой бактерии Neisseria meningitidis.
14. Композиция по п.12 или 13, где первая, вторая и третья бактерии Neisseria meningitidis генетически отличны в том, что они различаются по крайней мере по одному из серогруппы, серотипа или сероподтипа.
15. Композиция по п.12 или 13, где полипептиды GNA1870, экспрессируемые в первой, второй и третьей бактериях Neisseria meningitidis являются различными.
16. Композиция по п.1, где первая бактерия Neisseria meningitidis генетически модифицирована для продуцирования по крайней мере двух различных полипептидов GNA1870.
17. Композиция по п.16, где полипептиды GNA1870 относятся к различным группам вариантов полипептида GNA1870, где группы вариантов представляют собой v.1, v.2 и v.3.
18. Композиция по п.1, дополнительно содержащая:
выделенные антигенные везикулы, полученные из второй бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная вторая бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitides и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, где полипептиды GNA1870, сверхэкспрессируемые в первой и второй бактериях, относятся к различным группам вариантов полипептида GNA1870, и первая и вторая бактерии продуцируют различные полипептиды PorA.
19. Композиция по п.18, где композиция дополнительно содержит выделенные антигенные везикулы, полученные из третьей бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная третья бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningit idis, где полипептид GNA1870 из третьей бактерии относится к группе вариантов полипептида GNA1870, отличной от полипептидов из первой и второй бактерий, и где третья бактерия продуцирует полипептид PorA, отличный от полипептида PorA из первой и второй бактерий.
20. Композиция по п.18, где везикулы, относящиеся к первой и второй антигенным везикулам, получают без использования детергента.
21. Композиция по п.18 или 19, где различные группы вариантов полипептида GNA1870 представляют собой v.1, v.2 и v.3.
22. Способ вызова иммунной реакции против Neisseria, включающий стадии:
введение млекопитающему иммунологически эффективного количества композиции по п.1, где указанное введение достаточно для вызова иммунной реакции на присутствующий в везикулах полипептид GNA1870.
23. Способ по п.22, где везикулы представляют собой везикулы наружной мембраны (OMV), микровезикулы (MV) или смесь OMV и MV.
24. Способ по п.22, где бактерия Neisseria meningitidis представляет собой Н44/76.
25. Способ по п.22, где сверхэкспрессируемый полипептид GNA1870 идентичен эндогенному полипептиду GNA1870 бактерии Neisseria meningitidis.
26. Способ по п.22, где полипептид GNA1870 сверхэкспрессируется с гетерологичного промотора.
27. Способ по п.22, где композиция дополнительно содержит иммунологически эффективное количество второго антигенного препарата, содержащего везикулы, полученные из второй бактерии Neisseria meningitidis, где вторая бактерия Neisseria meningitidis генетически отличается от первой бактерии Neisseria meningitidis.
28. Способ по п.22, где композиция дополнительно содержит иммунологически эффективное количество второго антигенного препарата, содержащего везикулы, полученные из второй бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная вторая бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, и где вторая бактерия Neisseria meningitidis генетически отличается от первой бактерии Neisseria meningitidis.
29. Способ по п.27 или 28, где полипептид GNA1870 из второй бактерии Neisseria meningitidis отличается от полипептида GNA1870, сверхэкспрессируемого в первой бактерии Neisseria meningitidis.
30. Способ по п.27 или 28, где композиция дополнительно содержит третий выделенный антигенный препарат, включающий везикулы, полученные из третьей бактерии Neisseria meningitidis, где третья бактерия Neisseria meningitidis генетически отличается от первой или второй бактерии Neisseria meningitidis.
31. Способ по п.27 или 28, где композиция дополнительно содержит третий выделенный антигенный препарат, включающий везикулы, полученные из третьей бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная третья бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, и где третья бактерия Neisseria meningitidis генетически отличается от первой или второй бактерии Neisseria meningitidis.
32. Способ по п.30 или 31, где первая, вторая и третья бактерии Neisseria meningitidis генетически отличны в том, что они различаются по крайней мере по одному из серогруппы, серотипа или сероподтипа.
33. Способ по п.30 или 31, где полипептиды GNA1870 из первой, второй и третьей бактерий Neisseria meningitidis являются различными.
34. Способ по п.22, где первая бактерия Neisseria meningitidis генетически модифицирована для продуцирования по крайней мере двух различных полипептидов GNA1870.
35. Способ по п.34, где полипептиды GNA1870 относятся к различным группам вариантов полипептида GNA1870, где группы вариантов представляют собой v.1, v.2 и v.3.
36. Способ по п.22, где указанное введение вызывает у индивидуума защитную иммунную реакцию против более чем одного штамма Neisseria meningitidis.
37. Способ по п.22, где бактерия Neisseria meningitidis представляет собой Neisseria meningitidis серогруппы В.
38. Способ получения композиции по п.1, включающий:
культивирование бактерии Neisseria meningitidis, у которой инактивирован эндогенный ген GNA1870, где указанная бактерия трансформирована кодирующей полипептид GNA1870 экспрессирующей конструкцией, которая стабильно трансформирует Neisseria meningitidis и которая обеспечивает сверхэкспрессию полипептида GNA1870 в везикулах Neisseria meningitidis, получение везикул из культивируемой бактерии и объединение везикул с фармацевтически приемлемым носителем для получения антигенной композиции, приемлемой для введения индивидууму.
39. Способ по п.38, где везикула представляет собой везикулу наружной мембраны (OMV) или микровезикулу (MV).
40. Способ по п.38, где бактерия Neisseria meningitidis представляет собой Н44/76.
41. Способ по п.38, где полипептид GNA1870 идентичен эндогенному полипептиду GNA1870 из первой бактерии Neisseria meningitidis.
42. Способ по п.38, где сверхэкспрессируемый полипептид GNA1870 является гетерологичным для первой бактерии Neisseria meningitidis.
43. Способ по п.38, где бактерия Neisseria meningitidis генетически модифицирована для продуцирования по крайней мере двух различных полипептидов GNA1870.
44. Способ по п.43, где полипептиды GNA1870 относятся к различным группам вариантов полипептида GNA1870, где группы вариантов представляют собой v.1, v.2 и v.3.
45. Способ по п.38, где бактерия Neisseria meningitidis дефектна по полисахариду капсулы.

