Область изобретения
Это изобретение относится в основном к области биологии и химии. В частности, изобретение направлено на флуоресцирующие белки.
Уровень техники
Введение меток в белки, клетки или организмы, представляющие интерес, играет важную роль во многих биохимических, молекулярных биологических и медицинских диагностических применениях. Множество различных меток было разработано и использовалось в данной области, включая мечение радиоизотопами, красителями, флуоресцирующими метками, хемилюминесцентными метками и т.п., с различными свойствами и оптимальными применениями. Однако существует постоянный интерес в разработке новых меток. Особым интересом является разработка новых белковых меток, включая флуоресцирующие белковые метки.
Зеленый флуоресцирующий белок (GFP), его мутанты и гомологи, широко известные сегодня вследствие их интенсивного использования как флуоресцирующие маркеры in vivo в биомедицине, рассмотрены подробно Lippincott-Schwartz и Patterson в Science (2003) 300 (5616):87-91). GFP из гидромедузы Aequorea aequorea (синоним A. victoria), обнаруженный Johnson et ai. в J Cell Comp Physiol. (1962), 60:85-104, был обнаружен как часть биолюминесцентной системы медузы, где GFP играл роль вторичного источника, преобразовывающего синий свет от фотобелка экворина в зеленый свет. Затем сходные белки были выделены из нескольких биолюминесцентных кишечнополостных, включая гидроидную медузу Phialidium gregarium, морских перьев Renilla (класс Anthozoa) и других (см. Ward et al. in Photochem. Photobiol. (1982), 35: 803-808; Levine et al. in Comp. Biochem. Physiol. (1982), 72B: 77-85; Chalfie in Photochem. Photobiol. (1995), 62:651-656). Все эти белки демонстрируют зеленую фуюоресценцию, (излучение при 497-509 нм) и действовали как вторичные источники в биолюминесценции. Флуоресцирующие белки были также выделены из видов Physalia и их N-концевые аминокислотные последовательности были определены. (WO 03/017937).
кДНК, кодирующая GFP A. victoria, была клонирована Prasher et al. (Gene (1992), 111(2):229-33). Оказалось, что этот ген может быть гетерологично экспрессирован в практически любом организме благодаря уникальной способности GFP самостоятельно образовывать флуорофор (Chalfie et al., Gene (1992), 111(2):229-233). Эти сведения открывают широкие перспективы для использования GFP в клеточной биологии в качестве генетически кодируемой флуоресцирующей метки.
GFP широко использовался в различных областях применений, включая изучение экспрессии гена и локализации белка (Chalfie et al., Science 263 (1994), 802-805, и Heim et al. in Proc. Nat Acad. Sci. (1994), 91: 12501-12504), как средство для того, чтобы визуализировать внутриклеточное распределение органелл в клетках (Rizzuto et al, Curr. Biology (1995), 5: 635-642), для визуализации белкового переноса по секреторному пути (Kaether and Gerdes, FEES Letters (1995), 369: 267-271).
Проводились многие исследования для улучшения свойств GFP и для производства GFP-реактивов, пригодных и оптимизированных для ряда исследовательских целей. Были разработаны новые варианты GFP, такие как ДНК "гуманизированного" GFP, белковый продукт которой имеет повышенный синтез в клетках млекопитающих (Haas, et al., Current Biology (1996), 6: 315-324; Yang, et al., Nucleic Acids Research (1996), 24: 4592-4593). Один такой гуманизированный белок представляет собой "усиленный зеленый флуоресцирующий белок" (EGFP). Другие мутации к GFP привели к вариантам, излучающим сине-, сине-зеленый и желто-зеленый свет. Однако несмотря на большую полезность GFP, другие флуоресцирующие белки со свойствами, сходными или отличными от GFP, были бы полезны в данной области. В частности, преимущества от новых флуоресцирующих белков включают возможности флуоресцентного резонансного переноса энергии (FRET), возможности которого основаны на новых спектрах и лучшей пригодности при широком спектре возбуждения. В 1999 году гомологи GFP были клонированы из небиолюминесцентных видов Anthozoa (Matz et al., Nature Biotechnol. (1999), 17: 969-973). Это открытие показало, что эти белки не являются необходимыми компонентами биолюминисцентного механизма. Извлеченные из Anthozoa GFP-сходые белки показали большое спектральное разнообразие, включая сине-зеленые, зеленые, желтые, красные флуоресцирующие белки и фиолетово-синие нефлуоресцирующие хромопротеины (CPs) (Matz et al., Bioessays (2002), 24(10):953-959).
Главный недостаток извлеченных из Anthozoa GFP-сходых белков представляет собой сильную олигомеризацию, которая препятствует использованию этих белков во многих применениях (Lauf et al., FEBS Lett. (2001), 498: 11-15; Campbell et al., Proc. Natl. Acad. Sci. USA (2002), 99: 7877-7882; Mizuno et al., Biochemistry (2001), 40: 2502-2510). Соответственно, целью является обеспечение новых мономерных флуоресцирующих белков различных цветов, так же, как и ДНК, кодирующих их, которые не имеют недостатков известного GFP.
Вид Hydrozoa представляет собой потенциальный источник таких белков. Кроме GFP из Aequorea victoria и гомологов GFP из других видов Aequorea, подобно очень близким гомологам GFP из Aequorea macrodactyla (GenBank, инвентарные номера AF435427-AF435433) и Aequorea coerulescens (Gurskaya et al., Biochem J. (2003), 373(Pt 2): 403-408), никакие другие гены, кодирующие флуоресцирующие белки из Hydrozoa, до настоящего времени не клонировались, хотя некоторые из них были охарактеризованы на белковом уровне очень давно. Клонирование и мутагенез флуоресцирующих белков из других источников, не относящихся к Aequorea Hydrozoa, представляет собой перспективный способ получения новых флуоресцирующих меток с улучшенными свойствами.
Сущность изобретения
Настоящее изобретение обеспечивает молекулы нуклеиновых кислот, кодирующих флуоресцирующие или хромобелки и их мутанты, и их производные. Указанная нуклеиновая кислота может быть выделена, синтезирована или присутствовать в своей искусственной среде.
В некоторых воплощениях нуклеиновая кислота настоящего изобретения выделена из вида, не относящегося к Aequorea Hydrozoa, включая Phialidium sp. и двух флуоресцирующих медуз или гидроидной медузы 1 и 2 (гидромедузы 1 и 2) подотряда Anthomedusae, или мутантов или их производных.
В некоторых воплощениях нуклеиновая кислота настоящего изобретения кодирует белок, который имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 или 22. В некоторых воплощениях нуклеиновая кислота кодирует гомолог, мутант, производное соединение, миметик или фрагмент указанного белка.
В некоторых воплощениях, нуклеиновая кислота настоящего изобретения имеет нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 или 21 или которая является гомологичной, по существу такой же, или идентичной к указанным. Последовательности нуклеиновой кислоты, которые отличаются от представленных последовательностей нуклеиновой кислоты, вследствие вырожденности генетического кода или гибридизуемые с ними, также входят в рамки настоящего изобретения.
В других воплощениях изобретение направлено на белки, которые кодируются заявленными нуклеиновыми кислотами или по существу сходными с ними, или гомологами, производными или их мутантами, или направлено на слитые белки, включающие белки настоящего изобретения.
Также обеспечиваются фрагменты нуклеиновых кислот настоящего изобретения и нуклеиновых кислот, которые гибридизуются в условиях высокой жесткости с нуклеиновыми кислотами настоящего изобретения.
В других воплощениях также обеспечиваются векторы, включающие нуклеиновую кислоту настоящего изобретения. Кроме того, настоящее изобретение обеспечивает кассеты экспрессии, включающие нуклеиновую кислоту настоящего изобретения и регуляторные элементы, необходимые для экспрессии нуклеиновой кислоты в клетке.
В еще одном воплощении обеспечиваются способы получения хромогенного и/или флуоресцентного белка, включающие экспрессию белка в соответствующей клетке-хозяине и выделение из нее белка. Указанный способ включает (а) обеспечение молекулы нуклеиновой кислоты настоящего изобретения, кодирующей флуоресцентный или хромо-белок, соединенной с соответствующими последовательностями, регулирующими экспрессию, (b) экспрессию белка из указанной молекулы нуклеиновой кислоты, и (с) выделение белка, по существу свободного от других белков.
Кроме того, обеспечиваются антитела, специфические к белкам настоящего изобретения или их фрагментам.
Дополнительно, обеспечиваются клетки-хозяева, стабильные клеточные линии, трансгенные животные и трансгенные растения, содержащие нуклеиновые кислоты, векторы или кассеты экспрессии настоящего изобретения.
В еще одном воплощении обеспечиваются олигонуклеотиды или зонды, включающие нуклеотидные последовательности, способные к гибридизации с заявленными нуклеиновыми кислотами.
Также обеспечиваются способы, в которых используется хромо- или флуоресцирующий белок настоящего изобретения или кодирующая его нуклеиновая кислота.
В предпочтительном воплощении обеспечивается способ введения меток в биологическую молекулу, где указанный способ включает связывание указанной биологической молекулы с белком настоящего изобретения.
В другом предпочтительном воплощении обеспечивается способ введения меток в клетку, где указанный способ включает продуцирование белка настоящего изобретения в клетке.
В другом предпочтительном воплощении обеспечивается способ введения меток в органеллу клетки, где указанный способ включает получение белка настоящего изобретения, слитого с подходящим внутриклеточным сигналом локализации в клетке.
В еще одном предпочтительном воплощении обеспечивается способ анализа биологической молекулы, клетки или органеллы клетки, где указанный способ включает обнаружение флуоресцирующего сигнала от белка настоящего изобретения.
В еще одном предпочтительном воплощении обеспечивается способ анализа биологической молекулы, клетки или органеллы клетки, где указанный способ включает экспрессию молекулы нуклеиновой кислоты настоящего изобретения в клетке.
Дополнительно обеспечиваются наборы, включающие нуклеиновые кислоты или векторы или кассеты экспрессии, содержащие в себе указанные нуклеиновые кислоты, или белок настоящего изобретения.
Краткое описание фигур
На фиг.1 проиллюстрировано выравнивание GFP, phiYFP, hydr1GFP и hm2CP аминокислотных последовательностей. Введенные разрывы показаны точками. Остатки, идентичные соответствующим аминокислотам в GFP, представлены штриховыми линиями.
Фиг.2 показывает спектры возбуждения (пунктирная линия) и излучения (сплошная линия) для дикого типа phiYFP (A) и его мутантов: phiYFP-Y1 (B), phiYFP-M0 (C), и phiYFP-М1 (D).
Фиг.3 показывает спектры возбуждения-излучения для белков phiYFP-M1G1 (A) и phiYFP-M1C1 (B).
Фиг.4 представляет зарисовку гидромедузы 1 (A) и гидромедузы 2 (B) подотряда Anthomedusae.
Фиг.5 показывает спектры возбуждения-излучения для дикого типа hydr1GFP.
Фиг.6 показывает спектр поглощения для дикого типа hm2CP.
Фиг.7 показывает спектры возбуждения-излучения для дикого типа hm2CP.
Фиг.8 показывает спектры возбуждения-излучения для красного флуоресцирующего мутанта S3-2 белка hm2CP.
Подробное описание изобретения
Как здесь используется, термин "флуоресцирующий белок" или "флуоропротеин" означает белок, который является флуоресцирующим; например, он может проявлять низкую, среднюю или интенсивную флюоресценцию при облучении светом с подходящей для возбуждения длиной волны. Флуоресцентное свойство этих белков представляет собой такое свойство, которое является результатом взаимодействия двух или более аминокислотных остатков белка, а не одного аминокислотного остатка. Также флуоресцирующие белки настоящего изобретения не включают белки, которые проявляют флюоресценцию только от остатков, которые флуоресцируют сами как внутренние флуорофоры, то есть триптофан, тирозин и фенилаланин.
Как здесь используется, термин "хромопротеин" или "хромогенный белок" означает цветной белок, который может быть флуоресцирующим, слабо- или нефлуоресцирующим. Как здесь используется, термины "хромопротеин" и "флуоресцирующий белок" не включают люциферазы, такие как люцифераза Renilla.
Как здесь используется, термин "GFP" относится к зеленому флуоресцирующему белку из Aequorea victoria, включая варианты GFP, известные из уровня техники, сконструированные для обеспечения большей флюоресценции или флуоресценции в других цветовых областях. Последовательность дикого типа GFP была раскрыта в Prasher et al., Gene 111 (1992), 229-33.
Как здесь используется, термин "EGFP" относится к мутантному варианту GFP, имеющего две аминокислотные замены: F64L и S65T (Heim et al., Nature 373 (1995), 663-664).
Как здесь используется, термин "выделенный" означает молекулу или клетку, которые находятся в среде, отличной от среды, в которой молекула или клетка находятся в естественных условиях.
Как здесь используется, термин "фрагмент" предназначен, чтобы, включить, например, альтернативно сплайсированную или усеченную, или иначе расщепленную молекулу нуклеиновой кислоты или белка.
Как здесь используется, термин "производное" относится к мутанту или измененной за счет РНК, или химически модифицированной или иначе измененной молекуле нуклеиновой кислоты или к мутанту, или химически модифицированному или иначе измененному белку.
Как здесь используется, термин "мутант" относится к белку, раскрытому в настоящем изобретении, в котором одна или более аминокислот добавлены и/или замещены и/или удалены (делетированы) и/или вставлены (инсертированы) в N-конец и/или C-конец, и/или в пределах нативных аминокислотных последовательностей белков настоящего изобретения.
Как здесь используется, термин "мутант" относится к молекуле нуклеиновой кислоты, которая кодирует мутантный белок. Кроме того, термин "мутант" здесь относится к любому варианту, который короче или длиннее белка или нуклеиновой кислоты.
Как здесь используется, "гомолог или гомология" являются термином, использованным в данной области для описания связанности нуклеотида или пептидной последовательности с другим нуклеотидом или пептидной последовательностью, которая определена степенью идентичности и/или сходства между указанными сравниваемыми последовательностями.
Если подвести итог вышеуказанному, настоящее изобретение направлено на молекулы нуклеиновой кислоты, кодирующие флуоресцирующие и хромобелки и мутанты, варианты и их производные, так же, как белки и пептиды, кодируемые этими нуклеиновыми кислотами. Молекулы нуклеиновой кислоты и белки, представляющие интерес, выделены из вида, не относящегося к Aequorea Hydrozoa. Целевые белки, представляющие интерес, включают желтый флуоресцирующий белок, phiYFP, из Phialidium sp., зеленый флуоресцирующий белок hydr1GFP из гидроидной медузы 1 (гидромедуза 1) подотряда Anthomedusae, и фиолетового хромопротеина, hm2CP из гидроидной медузы 2 (гидромедуза 2) подотряда Anthomedusae. Также белками, представляющими интерес, являются те, которые по существу являются сходными или производными, или гомологами, или мутантами вышеупомянутых специфических белков. Также обеспечиваются фрагменты нуклеиновых кислот и пептиды, кодируемые ими, так же, как и антитела, специфические к белкам и пептидам изобретения. Кроме того, обеспечиваются клетки-хозяева, стабильные клеточные линии и трансгенные организмы, включающие вышеупомянутые молекулы нуклеиновой кислоты. Заявленные композиции белка и нуклеиновых кислот находят использование в ряде различных применений и способов, в частности, для применения при мечении белков. В итоге, обеспечиваются наборы для использования в таких способах и применениях.
Молекулы нуклеиновой кислоты
Настоящее изобретение обеспечивает молекулы нуклеиновой кислоты, кодирующие флуоресцирующие/хромобелки из вида Hydrozoa, отличного от рода Aequorea, производные, мутанты и гомологи этих белков, так же, как их фрагменты. Молекула нуклеиновой кислоты, как здесь используется, представляет собой молекулы ДНК, такие как геномные молекулы ДНК или молекулы кДНК, или РНК молекулы, такие как молекулы мРНК. В частности, указанные молекулы нуклеиновой кислоты представляют собой молекулы кДНК, имеющие открытую рамку считывания, которая кодирует хромо/флуоресцирующий белок изобретения из Hydrozoa или его фрагмент и способна, при подходящих условиях, к тому, чтобы обеспечить экспрессию как флуоресцирующего/хромобелка или белкового фрагмента (пептида) по изобретению.
Изобретение также охватывает нуклеиновые кислоты, которые являются гомологичными, по существу сходными, идентичными, производными или миметиками нуклеиновых кислот, кодирующих белки или белковые фрагменты настоящего изобретения. Заявленные нуклеиновые кислоты присутствуют в среде, отличной от их естественной среды; например, они выделены, присутствуют в обогащенных количествах, или присутствуют или экспрессированы in vitro или в клетке, или в организме, другом чем их естественно встречающаяся среда окружения.
Специфические молекулы нуклеиновой кислоты, представляющие интерес, представляют собой такие молекулы, которые кодируют следующие хромо/флуоропротеины из Hydrozoa (и их гомологи/производные/мутанты): желтый флуоресцирующий белок, phiYFP из Phialidium sp., зеленый флуоресцирующий белок, hydr1GFP из гидроидной медузы 1 из подотряда Anthomedusae, и фиолетовый хромопротеин, hm2CP из гидроидной медузы 2 из подотряда Anthomedusae. Каждый из этих специфических типов молекул нуклеиновой кислоты, представляющей интерес, теперь индивидуально раскрывается более детально.
phiYFP
Молекулы нуклеиновой кислоты, кодирующие флуоресцирующие/хромопротеины могут быть выделены из организма класса Hydrozoa, предпочтительно отряда Hydroida, более предпочтительно подотряда Leptomedusae, более предпочтительно из семейства Campanulariidae, и даже более предпочтительно из рода Phialidium. В особенно предпочтительном воплощении молекула нуклеиновой кислоты, выделенная из Phialidiumsp., кодирует специфический белок, называемый PhiYFP. Гомологи/мутанты/производные этого белка, такие как phiYFP-Y1, phiYFP-М1, phiYFP-M0, phiYFP-M1G1 (то есть phiYFP-G1 или phiGFPl), и phiYFP-MlCl (то есть phiYFP-G1 или phiGFP1), описанные ниже более детально в экспериментальной части, также представляющие интерес. Выведенная кодирующая последовательность кДНК дикого типа для PhiYFP, показана на SEQ ID NO: 01.
hydr1GFP
Молекулы нуклеиновой кислоты, кодирующие ф флуоресцирующие/хромопротеины могут быть выделены из организма класса Hydrozoa, предпочтительно отряда Hydroida, более предпочтительно подотряда Anthomedusae. Специфический белок, кодируемый такой молекулой нуклеиновой кислоты, называют hydr1GFP (то есть anmGFP1). Гомологи/мутанты/производные этого белка также представляют интерес. Выведенная последовательность кодирующая кДНК дикого типа для hydr1GFP, показана на SEQ ID NO: 11.
hm2CP
Молекулы нуклеиновой кислоты, кодирующие флуоресцирующие/хромопротеины могут быть выделены из организма класса Hydrozoa, предпочтительно отряда Hydroida, более предпочтительно подотряда Anthomedusae. Специфический белок, кодируемый такой молекулой нуклеиновой кислоты, называют hm2CP (то есть anm2CP). Гомологи/мутанты этого белка, такие как S3-2 красный флуоресцирующий мутант hm2CP, описанный ниже более детально в экспериментальной части, также представляющий интерес. Выведенная последовательность кодирующая кДНК дикого типа для hm2CP, показана на SEQ ID NO: 13.
Гомологи вышеописанных молекул нуклеиновой кислоты также являются объектом интереса. Источник гомологичных нуклеиновых кислот может быть любым видом растения или животного или последовательность, может быть полностью или частично синтетическим, включая миметики нуклеиновой кислоты. В некоторых воплощениях нуклеиновая кислота настоящего изобретения имеет идентичность последовательности с соответствующими гомологами на нуклеотидных или аминокислотных уровнях по меньшей мере приблизительно 40% и, предпочтительно приблизительно 50%, 55%, 60%, 65%, 70% или больше, включая 75%, 80%, 85%, 90% и 95% или больше. Референсная последовательность обычно будет по меньшей мере размером приблизительно в 60 нуклеотидов, чаще по меньшей мере размером приблизительно в 80 нуклеотидов, и может простираться на полную последовательность, с которой сравнивается. Идентичность последовательности рассчитывается исходя из референсной последовательности. Алгоритмы для анализа последовательности известны в данной области, такие как BLAST, описанный в Altschul et al., J. Mol. Biol., 215, pp. 403-10 (1990) (например, с использованием установок по умолчанию, то есть, с параметрами w=4 и T=17).
Гомологи идентифицируются любыми из множества способов. Фрагмент кДНК настоящего изобретения может использоваться как гибридизационный зонд против библиотеки кДНК от целевого организма, с использованием условий низкой жесткости. Зонд может быть большим фрагментом или одним или более коротким вырожденным праймером. Нуклеиновые кислоты, имеющие сходство последовательности, детектируются гибридизацией в условиях низкой жесткости, например, при 50°С и 6×SSC (0,9М хлористого натрия /0,09М цитрата натрия) с последующей промывкой при 55×С в 1×SSC (01,15М хлористого натрия/0,015 М цитрата натрия). Идентичность последовательности может быть определена гибридизацией в условиях высокой жесткости, например, при 50°С или выше и 0,1×SSC (15 мМ хлористого натрия/ 1,5 мМ цитрата натрия). Нуклеиновые кислоты, имеющие область, по существу идентичную представленным последовательностям, например аллельным вариантам, генетически-измененным вариантам нуклеиновой кислоты, и т.д., связываются с представленными последовательностями в условиях высокой жесткости гибридизации. С использованием зондов, в частности меченых зондов последовательностей ДНК, можно выделить гомологичные или схожие гены.
Также обеспечиваются нуклеиновые кислоты, которые гибридизуются с вышеописанными нуклеиновыми кислотами в жестких условиях, предпочтительно в условиях высокой жесткости (то есть, комплементарные предварительно описанным нуклеиновым кислотам). Примером гибридизации в жестких условиях является при 50°С или выше и 0,1×SSC (15 мМ хлористого натрия/ 1,5 мМ цитрата натрия). Другим примером гибридизации в условиях высокой жесткости является инкубация в течение ночи при 42°С в 50% растворе формамида, 5×SSC (150 мМ NaCl, 15 мМ тринатрий цитрат), 50 мМ фосфата натрия (рН 7,6), 5 × раствора Денхардта, 10% сульфата декстрана, и 20 мкг/мл денатурированной, разрезанной ДНК семенной жидкости лосося, с предварительной промывкой в 0,1×SSC при приблизительно 65°С. Другие условия высокой жесткости гибридизации известны в данной области и могут также использоваться для определения нуклеиновых кислот изобретения.
Также обеспечиваются нуклеиновые кислоты, кодирующие варианты, мутанты или производные белков изобретения. Мутанты или производные могут быть получены на матричной нуклеиновой кислоте, выбранной из вышеописанных нуклеиновых кислот, путем модификации, делеции или добавления одного или более нуклеотидов в матричной последовательности или их комбинации, для получения варианта матричной нуклеиновой кислоты. Модификации, добавления или делеции могут быть выполнены любым способом, известным в данной области (см. например Gustin et al., Biotechniques (1993) 14: 22; Barany, Gene (1985) 37: 111-123; and Colicelli et al., Mol. Gen. Genet. (1985) 199:537-539, Sambrook et al., Molecular Cloning: A Laboratory Manual, (1989), CSH Press, pp.15.3-15.108), включая подверженный ошибкам ПЦР, перестановку, олигонуклеотид-направленный мутагенез, ПЦР со сборкой, парный ПЦР мутагенез, мутагенез in vivo, кассетный мутагенез, рекурсивный ensemble мутагенез, экспоненциальный множественный мутагенез, сайт-специфический мутагенез, неспецифический мутагенез, генное реассемблирование, генный сайт-насыщающий мутагенез (GSSM), искусственное реассемблирование с лигацией (SLR) или их комбинации. Модификации, добавления или делеции могут быть также выполнены способом, включающим рекомбинацию, рекурсивную рекомбинацию последовательности, фосфотиоат-модифицированный мутагенез ДНК, мутагенез на урацил-содержащей матрице, мутагенез с двойным пропуском, точечный восстановительный по рассогласованию мутагенез, мутагенез штамма, дефицитного по восстановлениям, химический мутагенез, радоактивный мутагенез, делетационный мутагенез, рестрикционно-избирательный мутагенез, рестрикционный мутагенез с очисткой, синтез с искусственными генами, множественный мутагенез, создание химерных множественных нуклеиновых кислот и их комбинации. В некоторых воплощениях флуоресцирующие белки, кодируемые мутантом или производными нуклеиновых кислот, имеют те же самые флуоресцентные свойства как флуоресцирующий белок дикого типа. В других воплощениях мутант или производные нуклеиновых кислот кодируют флуоресцирующие белки с измененными спектральными свойствами, как здесь описано более подробно для мутантов phiYFP-Y1, phiYFP-М1, phiYFP-M1G1, phiYFP-M1С1, S3-2.
Кроме того, также обеспечиваются вырожденные варианты нуклеиновых кислот, которые кодируют белки настоящего изобретения. Вырожденные варианты нуклеиновых кислот включают замены кодонов нуклеиновой кислоты на другие кодоны, кодирующие те же самые аминокислоты. В частности, вырожденные варианты нуклеиновых кислот создаются, чтобы увеличить экспрессию в клетке-хозяине. В этом воплощении, кодоны нуклеиновой кислоты, которые не являются предпочтительными или являются менее предпочтительными в генах в клетке-хозяине, заменены кодонами, которые излишне представлены в кодирующих последовательностях в генах в клетке-хозяине, где указанные замененные кодоны кодируют ту же самую аминокислоту. Особенным интересом является гуманизированные версии нуклеиновых кислот настоящего изобретения. Как здесь используется, термин "гуманизированный" относится к заменам, сделанным в последовательности нуклеиновой кислоты для оптимизации кодонов для экспрессии белка в клетках млекопитающих (Yang et al., Nucleic Acids Research (1996) 24: 4592-4593). См. также Патент США № 5795737, который описывает гуманизацию белков, раскрытие которого здесь включено ссылкой.
Термин "кДНК", как здесь используется, предназначен для включения нуклеиновых кислот, которые отражают расположение элементов последовательности, находящихся в нативной зрелой разновидности мРНК, где элементы последовательности представляют собой экзоны и 5-' и 3'-некодирующие области. Обычно разновидность мРНК имеет смежные экзоны, с промежуточными интронами, которые, если присутствуют, удаляются ядерным РНК сплайсингом, чтобы создавать непрерывное кодирование белка открытой рамки считывания.
Геномная последовательность, представляющая интерес, может включать нуклеиновую кислоту, присутствующую между инициирующим кодоном и терминирующим кодоном, как определено в перечисленных последовательностях, включая все интроны, которые обычно присутствуют в нативной хромосоме. Геномная последовательность, представляющая интерес, дополнительно может включать 5'- и 3'-нетранслируемые области, находящиеся в зрелой мРНК, так же как специфические транскрипционные и трансляционные регулирующие последовательности, такие как промоторы, энхансеры, и т.д., включающие фланкирующую геномную ДНК размером приблизительно 1 т.п.н., но возможно и больше, у 5'- или 3'-конца транскрибированной области.
Молекулы нуклеиновой кислоты изобретения могут кодировать весь или часть заявленных белков. Двух- и одноцепочечные фрагменты могут быть получены из последовательности ДНК путем химического синтеза олигонуклеотидов в соответствии со стандартными способами, рестрикционной ферментной дигестией, амплификацией ПЦР и т.д. В основном, фрагменты ДНК будут иметь размер по меньшей мере приблизительно 15 нуклеотидов, обычно по меньшей мере приблизительно 18 нуклеотидов или приблизительно 25 нуклеотидов и могут быть по меньшей мере приблизительно 50 нуклеотидов. В некоторых воплощениях заявленные молекулы нуклеиновой кислоты могут иметь размер приблизительно 100, приблизительно 200, приблизительно 300, приблизительно 400, приблизительно 500, приблизительно 600, приблизительно 700 нуклеотидов или более. Заявленные нуклеиновые кислоты могут кодировать фрагменты заявленных белков или полных белков; например, заявленные нуклеиновые кислоты могут кодировать полипептиды приблизительно из 25 аминокислот, приблизительно из 50, приблизительно из 75, приблизительно из 100, приблизительно из 125, приблизительно из 150, приблизительно из 200 аминокислот вплоть до полной длины белка.
Заявленные нуклеиновые кислоты могут быть выделены и получены по существу в очищенной форме. По существу очищенная форма означает, что нуклеиновые кислоты являются по меньшей мере приблизительно на 50% чистыми, обычно по меньшей мере приблизительно на 90% чистыми и являются обычно "рекомбинантными", то есть фланкированы одним или более нуклеотидов, с которыми она обычно не связывается в хромосоме, встречающейся в природе в ее естественном организме-хозяине.
Нуклеиновые кислоты настоящего изобретения, например, имеющие последовательность SEQ ID NO: 01, 03, 05, 07, 09, 11, 13, 15, 17, 19 или 21, соответствующие кДНК, гены полной длины и конструкции, могут быть получены искусственно в соответствии со множеством различных методик, известных специалистам в данной области. Подходящие конструкции нуклеиновой кислоты очищаются, с использованием стандартных способов рекомбинантных ДНК, как описано, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., (1989) Cold Spring Harbor Press, Cold Spring Harbor, NY, и согласно инструкциям, описанным, например, в United States Dept. of HHS, National Institute of Health (NIH) Guidelines for Recombinant DNA Research.
Также обеспечиваются нуклеиновые кислоты, которые кодируют слитые белки, включающие белок настоящего изобретения, или их фрагменты, которые ниже рассматриваются более детально.
Также обеспечиваются вектор и другие конструкции нуклеиновой кислоты, содержащие заявленные нуклеиновые кислоты. Соответствующие векторы включают вирусные и невирусные векторы, плазмиды, космиды, фаги и т.д., предпочтительно плазмиды, и используются для клонирования, амплификации, экспрессии, переноса и т.д., последовательности нуклеиновой кислоты настоящего изобретения в подходящего хозяина. Выбор подходящего вектора является понятным для квалифицированного специалиста в данной области, и много таких векторов доступны коммерчески. Для приготовления конструкции частичная нуклеиновая кислота или полной длины обычно вставляются в вектор посредством прикрепления ДНК-лигазой к расщепленному ферментами рестрикции сайту в векторе. Альтернативно, желательная нуклеотидная последовательность может быть вставлена гомологичной рекомбинацией in vivo, обычно, присоединением гомологичных участков к вектору на флангах желательной нуклеотидной последовательности. Гомологичные участки добавляются лигированием олигонуклеотидов или полимеразной цепной реакцией с использованием праймеров, включающих, например, как гомологичные участки, так и часть желательной нуклеотидной последовательности.
Также обеспечиваются кассеты экспрессии или системы, использованные inter alia для получения заявленных хромогенных или флуоресцирующих белков или их белков-вставок или для снятия реплики заявленных молекул нуклеиновой кислоты. Кассета экспрессии может существовать как внехромосомный элемент или может быть включена в геном клетки в результате введения указанной кассеты экспрессии в клетку. Для экспрессии генный продукт, кодируемый нуклеиновой кислотой изобретения, экспрессируется в любой удобной системе экспрессии, включая, например, бактериальные системы, дрожжи, насекомое, амфибию или млекопитающее. В векторе экспрессии указанная нуклеиновая кислота является функционально связанной с регуляторной последовательностью, которая может включить промоторы, энхансеры, терминаторы, операторы, репрессоры и индукторы. Способы получения кассеты экспрессии или систем, способных к экспрессии желательного продукта, известны специалистом, квалифицированным в данной области.
Клеточные линии, которые устойчиво экспрессируют белки настоящего изобретения, могут быть выбраны способами, известными в данной области (например, котрансфекция с селектируемым маркером, таким как dhfr, gpt, неомицин, гигромицин, что делает возможным определение и выделение транфицированных клеток, которые содержат ген, включенный в геном).
Вышеописанные системы экспрессии могут использоваться в прокариотических или эукариотических хозяевах. Для получения белка могут использоваться клетки-хозяева, такие как E. coli, B. subtilis, S. cerevisiae, клетки насекомого в комбинации с бакуловирусными векторами, или клети высшего организма, такого как позвоночные, например COS 7 клетки, HEK 293, CHO, ооциты Xenopus и т.д.
Если используется любая вышеупомянутая клетка-хозяин или другие подходящие клетки-хозяева или организмы для репликации и/или экспрессии нуклеиновых кислот изобретения, то полученная реплицированная нуклеиновая кислота, экспрессированный белок или полипептид находятся в рамках притязания изобретения как продукт клетки-хозяина или организма. Продукт может быть выделен подходящим способом, известным в данной области.
Также представляют интерес промоторные последовательности геномных последовательностей настоящего изобретения, где последовательность 5'-концевых участков может использоваться для промоторных элементов, включая энхансерные связывающие сайты, которые, например, обеспечивают регуляцию экспрессии в клетках/тканях, где экспрессирован ген заявленных белков.
Также обеспечиваются малые фрагменты ДНК заявленных нуклеиновых кислот, которые применяются как праймеры для ПЦР, гибридизации скрининговых проб и т.д. Большие фрагменты ДНК применяются для получения кодируемых полипептидов, как ранее описано. Однако для использования в геометрических реакциях амплификации, таких как геометрическая ПЦР, используется пара малых фрагментов ДНК, то есть праймеров. Точная композиция последовательностей праймера не является критической для изобретения, но для большинства применений праймеры будут гибридизоваться с заявленной последовательностью в жестких условиях, как известно в данной области. Предпочтительно выбрать пару праймеров, которые дадут продукт амплификации по меньшей мере приблизительно из 50 нуклеотидов, предпочтительно по меньшей мере приблизительно из 100 нуклеотидов и могут простираться на полную последовательность нуклеиновой кислоты. Алгоритмы отбора последовательностей праймера обычно известны, и доступны в коммерческих пакетах программ. Праймеры амплификации гибридизуются с комплементарными цепочками ДНК и будут примировать навстречу друг к другу.
Молекулы нуклеиновой кислоты настоящего изобретения также могут применяться для определения экспрессии гена в биологическом образце. Метод, в котором исследуются клетки на наличие специфических последовательностей нуклеотида, таких как геномная ДНК или РНК, хорошо отработан в данной области. Кратко, ДНК или мРНК выделяются из образца клетки. мРНК может быть амплифицирована ОТ-ПЦР с использованием обратной транскриптазы для формирования комплементарной цепочки ДНК, с предварительной полимеразной цепной реакцией амплификации, с использованием праймеров, специфических для заявленных последовательностей ДНК. Альтернативно, образец мРНК отделяется гель-электрофорезом, переносится на подходящий носитель, например нитроцеллюлозу, нейлон и т.д., и затем зондируется фрагментом заявленной ДНК в качестве зонда. Могут также использоваться другие способы, такие как анализы сшивания олигонуклеотида, гибридизация in situ и гибридизация ДНК-зондами, прикрепленными на твердый чип. Обнаружение мРНК, гибридизованной с заявленной последовательностью, показательно из экспрессии гена в образце.
Заявленные нуклеиновые кислоты, включая фланкирующие участки промотора и кодирующих участков, могут мутироваться различными способами, известными в данной области для получения целенаправленных изменений в силе промотора или изменить последовательность кодируемого белка или свойства кодируемого белка, включая флуоресцентные свойства кодируемого белка.
Во многих воплощениях нуклеиновые кислоты, найденные в виде Aequorea, не включены в объем изобретения. В некоторых воплощениях, гомолог GFP и нуклеиновые кислоты, кодирующие его, из Aequorea victoria, Aequorea macrodactyla, и Aequorea coeridscens не включены в объем заявленного изобретения.
Белки
Также обеспечиваются в соответствии с заявленным изобретением хромо- и флуоресцирующие белки из других видов, не относящихся к Aequorea Hydrozoa, и их мутанты, включая белки полной длины, так же, как их части или фрагменты. Также обеспечиваются вариации встречающегося в природе белка, где такие вариации являются гомологичными или по существу сходными встречающемуся в природе белку и мутантам, встречающимся в природе белков, как описано в более подробном описании ниже.
Во многих воплощениях заявленные белки имеют максимум поглощения в диапазоне приблизительно от 300 до 700, обычно приблизительно от 350 до 650 и чаще приблизительно от 400 до 600 нм. Если заявленные белки являются флуоресцирующими белками, что означает, что они могут быть возбуждены светом одной длины волны, после чего они излучают свет другой длины волны, при этом спектры возбуждения заявленных белков обычно находятся приблизительно от 300 до 700 нм. Заявленные белки в общем случае имеют максимум коэффициента экстинкции, который находится приблизительно от 25000 до 150000 и обычно приблизительно от 45000 до 129000. Размер заявленных белков обычно находится в диапазоне приблизительно от 150 до 300 аминокислот и обычно приблизительно от 200 до 300 аминокислотных остатков и в общем случае они имеют молекулярный вес в пределах приблизительно от 15 до 35 кДа, обычно приблизительно от 17,5 до 32,5 кДа.
В некоторых воплощениях заявленные белки являются яркими, где под яркими понимается то, что хромо- и флуоресцирующие белки могут быть обнаружены обычными способами (например, визуальный скрининг, спектрофотометрия, спектрофлуориметрия, флуоресцентная микроскопия, FACS приборами, и т.д.) Флуоресцентная яркость специфических флуоресцирующих белков определяется их квантовым выходом, умноженным на максимальный коэффициент экстинкции. Яркость хромопротеинов может быть выражена ее максимальным коэффициентом экстинкции.
В некоторых воплощениях заявленные белки укладываются быстро после экспрессии в клетке-хозяине. Под быстрым укладыванием понимается то, что белки достигают своей третичной структуры, которая обеспечивает их хромо- или флуоресцентное свойство, через короткий период времени. В этих воплощениях, белки укладываются в течение периода времени, который в общем случае не превышает приблизительно 3 дня, обычно не превышает приблизительно 2 дня и чаще не превышает приблизительно 1 день.
Специфические белки, представляющие интерес, являются хромо/флуоропротеинами (и гомологами, мутантами, и их производными) из видов, не относящихся к Aequorea Hydrozoa: phiYFP из Phialidiumsp., зеленого флуоресцирующего белка, hydr1GFP из гидроидной медузы 1 (гидромедуза 1) подотряда Anthomedusae, и фиолетового хромопротеина, hm2CP из гидроидной медузы 2 (гидромедуза 2) подотряда Anthomedusae. Каждый из этих специфических типов полипептидных структуры, представляющих интерес, далее будут рассмотрены индивидуально более подробно.
phiYFP (и его производные/мутанты)
Белки этого воплощения имеют максимум поглощения света в диапазоне приблизительно от 350 до 550, обычно приблизительно от 450 до 550 и часто приблизительно от 435 до 540 нм, например, от 515 до 530 нм или от 480 до 490, тогда как максимум излучения обычно находится в диапазоне приблизительно от 400 нм до 650 нм и чаще приблизительно от 450 до 600 нм, тогда как во многих воплощениях спектры излучения находятся в диапазоне приблизительно от 470 до 550 нм, например, от 505 до 515 или от 520 до 530 нм, или от 530 до 540 нм. Размер заявленных белков типично составляет приблизительно от 200 до 250, обычно приблизительно от 210 до 240 аминокислотных остатков, и в общем случае имеют молекулярный вес в пределах приблизительно от 20 до 30, обычно приблизительно от 22,50 до 27,50 кДа. Специфическим интересом во многих воплощениях является phiYFP, который имеет аминокислотную последовательность показанную в SEQ ID NO: 02. Также представляющими интерес являются мутанты и производные этой последовательности, например, phiYFP-Y1, phiYFP-М1, phiYFP-M0, phiYFP-M1G1 и phiYFP-M1C1, как представлено в SEQ ID No: 04, 06, 08, 18 и 20, соответственно.
hydr1GFP (и его производные/мутанты)
Во многих воплощениях заявленные белки имеют максимум поглощения света в пределах приблизительно от 400 до 600 и чаще приблизительно от 450 до 550 нм, и часто приблизительно от 460 до 500 нм, например, от 470 до 480 нм, тогда как спектры излучения заявленных белков обычно находятся в пределах приблизительно от 450 до 650, обычно приблизительно от 460 до 600 нм и чаще приблизительно от 480 до 550 нм, например, от 480 до 500 нм и иногда от 490 до 500 нм. Размер заявленных белков обычно находится в пределах приблизительно от 200 до 300 аминокислот и обычно приблизительно от 220 до 290 аминокислотных остатков, и в общем случае имеют молекулярный вес в пределах приблизительно от 25 до 35 кДа, обычно приблизительно от 26,5 до 32,5 кДа. Специфическим интересом во многих воплощениях является дикий тип флуоресцирующего белка hydr1GFP, который имеет аминокислотную последовательность показанную в SEQ ID No: 12, мутанты и его производные.
hm2CP (и его мутанты)
Во многих воплощениях заявленные белки имеют максимум поглощения света в пределах приблизительно от 350 до 650, обычно приблизительно от 450 до 600 и чаще приблизительно от 490 до 595 нм, например, 560 до 590 нм, тогда как спектры поглощения заявленных белков обычно находятся в пределах приблизительно от 450 до 650, обычно приблизительно от 500 до 640 нм и чаще приблизительно от 580 до 620 нм, например, от 590 до 620 нм. Размер заявленных белков обычно находится в пределах приблизительно от 200 до 250, обычно приблизительно от 210 до 240 аминокислотных остатков, и в общем случае они имеют молекулярный вес в пределах приблизительно от 20 до 30 кДа, обычно приблизительно от 22,50 до 27,50 кДа. Специфическим, представляющим интерес, во многих воплощениях является hm2CP (anm2CP), который имеет аминокислотную последовательность показанную в SEQ ID No: 14. Также представляющим интерес является мутант этой последовательности, например красный флуоресцирующий белок S3-2, и т.п., как представлено, например, в SEQ ID NO: 16.
Также обеспечиваются гомологи или белки, последовательность которых отличаются от описанных выше указанных специфических аминокислотных последовательностей заявленных в изобретении, то есть SEQ ID NO: 02, 04, 06, 08, 10, 12, 14, 16, 18, 20 или 22. Под гомологом понимается белок, имеющий по меньшей мере приблизительно 55%, обычно по меньшей мере приблизительно 60% и чаще по меньшей мере приблизительно 65% идентичность аминокислотной последовательности по отношению к аминокислотным последовательностям SEQ ID NO: 02, 04, 06, 08, 10, 12, 14, 16, 18, 20 или 22 как определено с использованием MegAlign, DNAstar clustal алгоритма, как описано в D.G. Higgins and P.M. Sharp, "Fast and Sensitive multiple Sequence Alignments on a Microcomputer," CABIOS, 5 pp. 151-3 (1989) (с использованием параметров: ktuple 1, gap penalty 3, window 5 и diagonals saved 5). Во многих воплощениях гомологи, представляющие интерес, имеют намного более высокую идентичность последовательности, например 70%, 75%, 80%, 85%, 90% (например, 92%, 93%, 94%) или выше, например 95%, 96%, 97%, 98%, 99%, 99,5%, особенно для последовательности аминокислот, которые обеспечивают функциональные области белка.
Также обеспечиваются белки, которые являются по существу идентичными белку дикого типа, где по существу идентичный означает, что белок имеет идентичность аминокислотной последовательности по отношению к последовательности белка дикого типа по меньшей мере приблизительно 60%, обычно по меньшей мере приблизительно 65% и чаще по меньшей мере приблизительно 70%, причем в некоторых образцах идентичность может быть намного более высокой, например 75%, 80%, 85%, 90%, 95% или выше.
Также обеспечиваются белки, которые являются производными или мутантами вышеописанных белков, встречающихся в природе. Мутанты и производные могут сохранять биологические свойства белков дикого типа (например, встречающихся в природе), или могут иметь биологические свойства, которые отличаются от белков дикого типа. Термин "биологическое свойство" белков настоящего изобретения относится, но без ограничения, к спектральным свойствам, таким как максимум поглощения, максимум излучения, максимальный коэффициент экстинкции, яркость (например, как по сравнению с белком дикого типа или другим референсным белком, таким как зеленый флуоресцирующий белок (GFP) из A.victoria), и т.п.; биохимическим свойствам, таким как стабильность in vivo и/или in vitro (например, период полураспада); скорости созревания, склонности к агрегации и склонности к олигомеризации и другим таким свойствам. Мутации включают замены одной аминокислоты, делецию или инсерцию одной или более аминокислот, усечений или удлинений N-конца, усечений удлинений C-конца и т.п.
Мутанты и производные могут быть получены с использованием стандартных методов молекулярной биологии, как подробно описано в разделе "молекулы нуклеиновой кислоты" выше. Здесь описаны несколько мутантов. Примеры обеспечивают общие приемы и использование стандартных способов, так что специалисты, квалифицированные в данной области, могут легко получить большой ряд дополнительных мутантов и проверить, было ли изменено биологическое (например биохимическое, спектральное, и т.д.) свойство. Например, флуоресцентная интенсивность может быть измерена с использованием спектрофотометра при различных длинах волн возбуждения.
Производные могут быть также получены с использованием стандартных методов и включают изменение при помощи РНК, химические модификации, модификации после трансляции и после транскрипции и т.п. Например, производные могут быть получены методами, такими как измененное фосфорилирование или гликозилирование, или ацетилирование, или липидирование, или различными типами расщепления при созревании и т.п.
Белки, заявленные в изобретении, которые являются встречающимися в природе белками, присутствуют в окружающей среде, не являющейся для них обычной, например, они отделены от обычной среды. Например, обеспечивается очищенный белок, где "очищенный" означает, что белок присутствует в смеси, которая является по существу не содержащей нехромогенные или флуоресцирующие белки, представляющие интерес, где "по существу не содержит" означает, что меньше чем 90%, обычно меньше чем 60% и чаще меньше чем 50% содержимого смеси является нехромогенными или флуоресцирующими белками или их мутантами. Белки настоящего изобретения также могут присутствовать в выделенной форме, что означает, что белок является по существу не содержащим другие белки и других биологических молекул, встречающихся в естественном окружении, таких как олигосахариды, нуклеиновые кислоты и их фрагменты, и т.п., где термин "по существу не содержит" в этом случае означает, что меньше, чем 70%, обычно меньше, чем 60% и чаще меньше, чем 50% композиции, содержащей выделенный белок, представляет собой некоторую другую биологическую молекулу, встречающуюся в природе. В некоторых воплощениях белки присутствуют в по существу очищенной форме, где "по существу очищенная форма" означает очищенная по меньшей мере на 95%, обычно по меньшей мере на 97% и чаще по меньшей мере на 99%.
Также обеспечиваются фрагменты белков, встречающихся в природе, так же, как мутанты и производные белков, описанные выше. Биологически активные фрагменты и/или фрагменты, соответствующие функциональным доменам, и т.п. представляют частный интерес. Фрагментами, представляющими интерес, являются полипептиды, которые обычно имеют размер по меньшей мере приблизительно 30 аминокислот, обычно по меньшей мере приблизительно 50 аминокислот, предпочтительно по меньшей мере приблизительно 75 или 100 аминокислот, и могут быть размером 300 аминокислот или более, но обычно не будут превышать размера приблизительно 250 аминокислот, где фрагмент будет иметь участок аминокислот, который является одинаковым с заявленным белком, размером по меньшей мере приблизительно 25 аминокислот, и обычно по меньшей мере приблизительно 45 аминокислот, и во многих воплощениях по меньшей мере приблизительно 50 аминокислот. В некоторых воплощениях заявленные полипептиды имеют размер приблизительно 25 аминокислот, приблизительно 50, приблизительно 75, приблизительно 100, приблизительно 125, приблизительно 150, приблизительно 200, или приблизительно 250 аминокислот, вплоть до полного размера белка. В некоторых воплощениях белковый фрагмент сохраняет все или по существу все специфическое свойство белка дикого типа.
Заявленные белки и полипептиды могут быть получены из природных источников или искусственно синтезированы. Например, белки дикого типа могут быть получены из биологических источников, которые экспрессируют белки, например виды Hydrozoa, такие как некоторые вышеупомянутые. Заявленные белки могут также быть получены искусственным путем, например экспрессией рекомбинантной нуклеиновой кислоты, кодирующей последовательность белка, представляющего интерес, в соответствующем хозяине, как описано выше. Любые обычные методики очистки белка могут быть применяться, где подходящие методы очистки белка описаны в Guide to Protein Purification, (Deuthser ed.) (Academic Press, 1990). Например, лизат может быть приготовлен из исходного источника и очищен с использованием ВЭЖХ, эксклюзионной хроматографий гель-электрофореза, афинной хроматографии и т.п.
Также обеспечиваются белки слияния, включающие белок настоящего изобретения, или его фрагменты, слитые, например, с вырожденной последовательностью, последовательностью внутриклеточной локализации (например, ядерный сигнал локализации, пероксимальный нацеливающий сигнал, последовательность нацеливающая аппарат Гольджи, последовательность, нацеливающая митохондрию и т.д.), сигнальный пептид или любой белок или полипептид, представляющий интерес. Белки слияния могут включать например, флуоро/хромобелок, заявленный в изобретении, полипептид и второй полипептид ("партнер слияния"), соединенный в рамках в N-конца и/или C-конца флуоро/хромополипептида. Партнеры слияния включают, но не ограничены, полипептидами, которые могут связывать антитела, специфические к партнеру слияния (например, эпитопные метки), антитела или их связывающие фрагменты, полипептиды, которые обеспечивают каталитическую функцию или вызывают клеточный ответ, лиганды или рецепторы или их миметики, и т.п. В таких белках слияния в общем случае партнер слияния естественно не связывается с флуоро/хромобелковой частью белка слияния и обычно является флуоро/хромобелками не из вида Hydrozoa, заявленными в изобретении, или их производным/фрагментом; то есть, они не обнаруживаются в виде Hydrozoa.
Также обеспечиваются антитела, которые специфически связываются с флуоресцирующим или хромопротеином настоящего изобретения. Соответствующие антитела могут быть получены с использованием способов, известных в данной области. Например, поликлональные антитела могут быть получены как описано в (Harlow and Lane Antibodies: A Laboratory Manual, (1988) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York), и моноклональные антитела могут быть получены как описано в (Coding Monoclonal Antibodies: Principles and Practice: Production and Application of Monoclonal Antibodies in Cell Biology, Biochemistry and Immunology; 3rd edition, (1996) Academic Press). Химерные антитела, включая гуманизированные антитела, так же, как одноцепочечные антитела и фрагменты антител, такие как Fv, F(ab')2 и Fab также представляют интерес.
Трансгеники
Нуклеиновые кислоты настоящего изобретения могут использоваться для получения трансгенных организмов или сайт-специфичные генных изменений в клеточных линиях. Трансгенные клетки, заявленные в изобретении, содержат одну или более нуклеиновых кислот, заявленных по настоящему изобретению, в качестве трансгена. Для целей изобретения любая приемлемая клетка-хозяин может быть использована, включая прокариотические (например, Escherichia coli, Streptomyces sp., Bacillus subtilis, Lactobacillus acidophilus, и т.д.) или эукариотические клетки-хозяева. Трансгенный организм, заявленный по изобретению, может быть прокариотическим или эукариотическим организмом, включая бактерии, цианобактерии, грибы, растения и животные, в которых одна или больше клеток организма содержат гетерогенную нуклеиновую кислоту, заявленную по изобретению, введенную посредством вмешательства человека, такими трансгенными способами, которые известны в данной области.
Выделенная нуклеиновая кислота настоящего изобретения может быть введена в хозяина способами, известными в данной области, например инфицированием, трансфектированием, трансформацией или трансконъюгацией. Способы переноса молекулы нуклеиновой кислоты (то есть, ДНК) в такие организмы широко известны и обеспечивается в ссылках, таких как Sambrook et al. (Molecular Cloning: A Laboratory Manual, 3nd Ed., (2001) Cold Spring Harbor Press, Cold Spring Harbor, NY).
В одном воплощении, трансгенный организм может быть прокариотическим организмом. Способы трансформации прокариотических хозяев хорошо описаны в данной области (например, см. Sambrook et al. Molecular Cloning: A Laboratory Manual, 2nd edition (1989) Cold Spring Harbor Laboratory Press и Ausubel et al., Current Protocols in Molecular Biology (1995) John Wiley & Sons, Inc).
В другом воплощении, трансгенный организм может быть грибами, например дрожжами. Дрожжи широко используются как носитель для экспрессии гетерогенного гена (например см. Goodey et al Yeast biotechnology, D R Berry et al, eds, (1987) Allen and Unwin, London, pp 401-429) и King et al Molecular and Cell Biology of Yeasts, E F Walton and G T Yarronton, eds, Blackie, Glasgow (1989) pp 107-133). Несколько типов дрожжевых векторов доступны, включая интегративные векторы, которые требуют рекомбинации с геномом хозяина для их поддержки, и автономно реплицируются плазмидных векторах.
Другой организм хозяина является животным. Трансгенные животные могут быть получены трансгенными способами, известными в данной области и обеспечиваются в ссылках, таких как Pinkert, Transgenic Animal Technology: a Laboratory Handbook, 2nd edition (2203) San Diego: Academic Press; Gersenstein and Vintersten, Manipulating the Mouse Embryo: A Laboratory Manual, 3rd ed, (2002) Nagy A. (Ed), Cold Spring Harbor Laboratory; Blau et al., Laboratory Animal Medicine, 2nd Ed., (2002) Fox J.G., Anderson L.C., Loew F.M., Quimby F.W. (Eds), American Medical Association, American Psychological Association; Gene Targeting: A Practical Approach by Alexandra L. Joyner (Ed.) Oxford University Press; 2nd edition (2000). Например, трансгенные животные могут быть получены гомологичной рекомбинацией, где эндогенный локус изменен. Альтернативно, конструкция нуклеиновой кислоты беспорядочно включается в геном. Векторы для устойчивого включения включают плазмиды, ретровирусы и другие животные вирусы, YAC, и т.п.
Нуклеиновые кислоты могут быть введены в клетку, непосредственно или опосредованно, введением в прекурсор клетки, путем намеренной генетической манипуляции, такой как микроинъекция или инфицирование рекомбинантным вирусом или рекомбинантным вирусным вектором и т.п. Термин «генетическая манипуляция» не включает классическое скрещивание или оплодотворение in vitro, а предпочтительно является направленным на введение рекомбинантных молекул нуклеиновых кислот. Эти молекулы нуклеиновой кислоты могут быть включены в хромосому или они могут быть внехромосомными реплицирующими ДНК.
Конструкции ДНК для гомологичной рекомбинации будут содержать по меньшей мере часть нуклеиновой кислоты настоящего изобретения, где ген имеет желательную генетическую модификацую(ции) и включает области гомологии с целевым локусом. Конструкциям ДНК для произвольного включения не обязательно содержать область гомологии с медиатором рекомбинации. Легко могут быть включены маркеры для положительной и отрицательной селекции. Способы получения клеток, имеющих целевые генные модификации, через гомологическую комбинацию известны в данной области. Для различных способов трансфекции клеток млекопитающих, см. Keown et al., Meth. Enzymol. (1990) 185:527-537.
Для эмбриональных стволовых (ES) клеток могут быть использованы клеточные линии ES, или эмбриональные клетки могут быть получены непосредственно от хозяина, такого как мышь, крыса, морская свинка и т.д. Такие клетки выращиваются на подходящем фибропласт-питающем слое или расти в присутствии фактора ингибирования лейкемии (LIF). Трансформированные ES или эмбриональные клетки могут использоваться для получения трансгенных животных с использованием подходящего способа, описанного в данной области.
Трансгенные животные могут быть любыми животными, не относящимися к человеку, включая млекопитающее, не относящееся к человеку, (например мышь, крыса), птица или амфибия и т.д., и использованы в функциональном исследовании, скрининге лекарственного средства и т.п. Характерные примеры использования трансгенных животных включают те, которые описанные ниже.
Также могут быть получены трансгенные растения. Способы получения трансгенных растительных клеток и растений описаны в патентах США №№ 5767367; 5750870; 5739409; 5689049; 5689045; 5674731; 5656466; 5633155; 5629470; 5595896; 5576198; 5538879; 5484956; раскрытия которых включены сюда ссылкой. Способы получения трансгенных растений также рассмотрены в Plant Biochemistry and Molecular Biology (eds. Lea and Leegood, John Wiley & Sons) (1993) pp. 275-295 и в Plant Biotechnology and Transgenic Plants (eds. Oksman-Caldentey and Barz), (2002) 719 p.
Например, эмбриогенные эксплантаты, содержащие соматические клетки, могут использоваться для получения трансгенного хозяина. После сбора клеток или ткани экзогенная ДНК, представляющая интерес, вводится в растительные клетки, при этом известен для такого введения ряд различных способов. При наличии выделенных протопластов возникает возможность для введения через ДНК-опосредованные протоколы передачи гена, включая инкубацию протопластов с выделенной ДНК, такой как плазмиды, включающие целевую экзогенную последовательность кодирования в присутствии поливалентных катионов (например, PEG или PLO); или электропорации протопластов в присутствии выделенной ДНК, включающей целевую экзогенную последовательность. Протопласты, которые успешно включили экзогенную ДНК, затем отбираются, выращиваются в каллус, и в конечном счете в трансгенное растение при контакте с подходящими количествами и отношениями стимулирующих факторов, таких как ауксины и цитокины.
Другие соответствующие способы получения растения могут использоваться, такие как применение "генной пушки", или Agrobacterium-опосредованная трансформация, которые доступны для квалифицированных специалистов в данной области.
Способы применения
Флуоресцирующие белки настоящего изобретения (так же, как другие, описанные выше компоненты, заявленные в изобретении), находят использование в ряде различных применений. Например, они могут использоваться в способах введения меток, при анализе или детекции биологической молекулы, клетки или органеллы клетки. Характерные применения для каждого из этих типов белков будут описаны ниже, причем применения, описанные здесь, являются только иллюстративными и никоим образом не предназначены для ограничений применения белков настоящего изобретения по данному описанию.
В предпочтительном воплощении относящемуся к способу введения меток в биологическую молекулу, клетку или органеллу клетки, заявленные белки находят применения как метки in vivo (или молекулы-репортеры) в анализах клеточной и молекулярной биологии. Анализы, представляющие интерес, включают, но не ограничены, анализами экспрессии гена, белковой локализацией и совместной локализацией, белок-белковыми взаимодействиями, взаимодействиями белка и нуклеиновой кислоты, взаимодействиями нуклеиновая кислота - нуклеиновая кислота, клеточной локализацией и локализацией органелл клетки и взаимодействиями органелл клетки и т.д. Флуоресцирующие белки настоящего изобретения находят применение как метки биомолекулы или метки органелл клетки в живых и закрепленных клетках; как маркеры в клетке или органелл слияния, как маркеры интеграции клетки или органелл, как маркеры трансфекции (например, как метки для отбора трансфицированных клеток, содержащих вектора экспрессии, кодирующих по меньшей мере один флуоресцирующий белок изобретения), как зонд реального времени, работающий при околофизиологических концентрациях и т.д.
Кроме того, заявленные белки могут применяться в способе анализа биологической молекулы. Например, они находят применение для идентификации и/или измерения экспрессии целевого белка или полипептида в биологическом материале. Этот способ включает: i) введение в клетку молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую флуоресцирующий белок по настоящему изобретению, где указанная молекула нуклеиновой кислоты оперативно связана и находится под контролем последовательности, регулирующей экспрессию, которая управляет экспрессией указанного целевого белка или полипептида; ii) экспрессию упомянутой нуклеиновой кислоты при соответствующих условиях; и iii) обнаружение флуоресцирующего излучения флуоресцирующего белка как средства для измерения экспрессии белка, представляющего интерес.
В частности, заявленные белки находят применение для идентификации и/или измерения экспрессии и/или локализации целевого белка или полипептида в биологическом материале. Этот способ включает: i) введение в клетку молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую флуоресцирующий белок по настоящему изобретению, где указанная молекула нуклеиновой кислоты слита с последовательностью, кодирующей целевой белок или полипептид и оперативно связана и находится под контролем последовательности, которая управляет экспрессией, указанного целевого белка; ii) культивирование клетки в условиях, подходящих для экспрессии целевого белка; и iii) обнаружение флуоресцирующего излучения флуоресцирующего белка как способа измерения экспрессии целевого белка.
Целевые применения включают использование заявленных белков в способах флуоресцентного резонансного переноса энергии (FRET). В этих способах заявленные белки служат донором и/или акцепторами в комбинации со вторым флуоресцирующим белком или красителем, например другим флуоресцирующим белком, заявленным в изобретении, или флуоресцирующим белком, как описано в Matz et al., Nature Biotechnology 17:969-973 (1999); зеленым флуоресцирующим белком из Aequorea victoria или его флуоресцирующим мутантом, например, как описано в патентах США N 6066476; 6020192; 5985577; 5976796; 5968750; 5968738; 5958713; 5919445; 5874304, раскрытия которых включены сюда ссылкой; другими флуоресцентными красителями, такими как кумарин и его производные, 7-амино-4-метилкумарин и аминокумарин; красителями для живых тканей; каскадный голубой; или флуоресцеином и его производными, такими как флуоресцеинизотиоцианат и Орегонский зеленый; родаминовыми красителями, такими как Техасский красный, тетраметилродамином, эозинами и эритрозинами; циановыми красителями, такими как Cy3 и Cy5; макроциклическими хелатами лантоноидных ионов, такими как квантовый краситель; и хемилюминесцентными красителями, такими как люцеферазы, включая описанные в патентах США №№ 5843746; 5700673; 5674713; 5618722; 5418155; 5330906; 5229285; 5221623; 5182202; раскрытия которых включены сюда ссылкой.
Отдельные примеры того, что в FRET-анализах используются заявленные флуоресцирующие белки, включают, но не ограничены, обнаружение белок-белковых взаимодействий, таких как в двухгибридной системе млекопитающих, фактора транскрипции димеризации, мембранной белковой мультимиризации, мультибелковое комплексообразование; как биосенсор для множества различных событий, где пептид или белок ковалентно связывают FRET флуоресцентную комбинацию, включая заявленные флуоресцирующие белки, и связанный пептид или белок представляет собой, например, протеаза-специфический субстрат для опосредованного каспазом расщепления, пептид, который подвергается конформационному изменению после получения сигнала, который увеличивает или уменьшает FRET, такого как PKA регулирующий домен (цАМФ-сенсор), сайт фосфорилирования (например, где находится сайт фосфорилирования в пептиде, или пептид имеет специфичность связывания фосфорилированного/дефосфорилированного домена другого белка), или пептид имеет Ca2+ связывающий домен. Кроме того, применения флуоресцентного резонансного переноса энергии или FRET, в которых белки настоящего изобретения находят применение, включают, но не ограничены, описанные в патентах США №№ 6008373; 5998146; 5981200; 5945526; 5945283; 5911952; 5869255; 5866336; 5863727; 5728528; 5707804; 5688648; 5439797, раскрытия которых включены сюда ссылкой.
Флуоресцирующие белки настоящего изобретения находят применение в способе обнаружения влияния испытуемого материала на регулирование экспрессии и/или транслокации одного или более целевых белков в клетке. Альтернативно, они находят применение в способе обнаружения экспрессии целевого белка и одновременной активности последовательности, регулирующей экспрессию, в ответ на испытуемый материал. Флуоресцирующие белки находят также применение в способе сравнения активности двух или более последовательностей, регулирующих экспрессию в клетке, в ответ на испытательный материал. Такие способы могут быть выполнены при наличии и при отсутствии испытуемого материала, влияние которого на способ должно быть измерено.
Флуоресцирующие белки настоящего изобретения также находят применение в приложениях, включающих автоматизированный скрининг матрицы клеток, экспрессирующих флуоресцентные группы-репортеры, с использованием микроскопического отображения и электронного анализа. Скрининг может использоваться для открытия лекарственного средства и в области функциональной геномики, где заявленные белки используются как маркеры целых клеток для обнаружения изменений в многоклеточном преобразовании и перемещении, например, при формировании многоклеточных канальцев (формирование кровеносного сосуда) эндотелиальными клетками, перемещение клеток сквозь систему Fluoroblok Insert (Becton Dickinson Co.), заживление раны, или отрастании нейрона. Скрининг может также быть использован, если белки настоящего изобретения используются как маркеры, слитые с пептидами (такими как целевые последовательности) или белками, которые обнаруживают изменения во внутриклеточной локализации как индикатор клеточной активности, например, в сигнальной трансдукции, такие как факторы киназы и транслокации транскрипции при стимуляции. Примеры включают протеинкиназу C, протеинкиназу A, фактор транскрипции NFkB и NFAT; клеточные цикличные белки, такие как циклин A, циклин В1 и циклин E; расщепление протеазой с последовательным перемещением расщепленного субстрата; фосфолипиды с маркерами для внутриклеточных структур, такие как эндоплазматическая сеть, аппарат Гольджи, митохондрия, пероксисомы, ядро, нуклеоли, плазматическая мембрана, гистоны, эндосомы, лизосомы или микротрубки.
Белки настоящего изобретения также могут использоваться в скрининге с высоким содержанием для обнаружения совместной локализации других флуоресцирующих слитых белков с маркерами локализации как индикаторы перемещений внутриклеточных флуоресцирующих белков/пептидов или только как маркеры. Примеры применений, включающих автоматизированный скрининг множеств клеток, в которых заявленные флуоресцирующие белки находят применение, включают патенты США № 5989835; так же, как WO 00/17624, WO 00/26408, WO 00/17643 и WO 00/03246, раскрытия которых включено сюда ссылкой.
Флуоресцирующие белки настоящего изобретения также находят применение в скрининговых анализах с высокой производительностью. Заявленные флуоресцирующие белки представляют собой стабильные белки с периодами полураспада больше, чем 24 часа. Также обеспечиваются дестабилизированные варианты заявленных флуоресцирующих белков с уменьшенными периодами полураспада, которые могут использоваться как репортеры транскрипции для открытия лекарственного средства. Например, белок, заявленный в изобретении, может быть слит с предполагаемой протеолитической сигнальной последовательностью, производной из белка с более коротким периодом полураспада, таким как PEST-последовательность из гена орнитиндекарбоксилазы мыши, с боксом разложения циклина B1 из мыши или убиквитина, и т.д. Для описания дестабилизированных белков и векторов, которые могут быть использованы для получения того же, см. например, патент США N 6130313, раскрытие которого включено сюда ссылкой. Промоторы в сигнальных путях трансдукции могут быть обнаружены с использованием дестабилизированных вариантов заявленных флуоресцирующих белков для скрининга лекарственных средств, такие как, например, AP1, NFAT, NFkB, Smad, STAT, p53, E2F, Rb, myc, CRE, ER, GR и TRE и т.п.
Заявленные белки могут использоваться как вторые мессенджерные детекторы путем слияния заявленных белов со специфичными доменами, такими как PKC-гамма Ca связывающий домен, PKC-гамма DAG связывающий домен, SH2 домен или SH3 домен и т.д.
Секретируемые формы заявленных белков, которые в свою очередь могут использоваться в ряде различных применений, могут быть получены слиянием секретируемых лидирующих последовательностей с заявленным белком.
Заявленные белки также находят использование в применении при сортировке клеток, активированных флюоресценцией, (FACS). В таких применениях заявленный флуоресцирующий белок используется для мечения популяции клеток, и полученная меченная популяция клеток затем подвергается сортировке в устройстве для сортировки клеток, активированных флуоресценцией, как известно в данной области. Способы FACS описаны в патентах США №№ 5968738 и 5804387; раскрытия которых включены сюда ссылкой.
Заявленные белки также находят применение как метки in vivo для трансгенных животных. Например, экспрессия заявленного белка может сопровождаться тканеспецифичными промоторами, где такие способы находят применение в исследованиях для генной терапии, таких как тестирование эффективности трансгенной экспрессии, среди других применений. Типичное применение флуоресцирующих белков у трансгенных животных, которое поясняет такие применения, находится в WO 00/02997, раскрытие которого включено сюда ссылкой.
Дополнительные применения белков настоящего изобретения включают использование в качестве маркеров для введения в клетки или животных и при калибровке в количественных измерениях; в качестве маркеров или репортеров в кислородных биосенсорных приборах для контроля клеточной жизнеспособности; в качестве маркеров или меток для животных, домашних животных, игрушек, пищи, и т.п.
Заявленные флуоресцирующие белки также находят применение в анализах протеазного расщепления. Например, инактивированные расщеплением флуоресцентные анализы могут быть осуществлены, с использованием заявленных белков, где заявленные белки подготовлены для включения в протеазоспецифичную последовательность расщепления, не разрушая флуоресцирующих свойств белка. При расщеплении флуоресцирующего белка активированной протеазой флюоресценция резко уменьшается вследствие разрушения функционального хромофора. Альтернативно, активированная расщеплением флюоресценция может быть применена с использованием белов настоящего изобретения, когда получают белки, содержащие дополнительную спейсерную последовательность вблизи/или внутри хромофора. Этот вариант значительно уменьшает его флуоресцентную активность, потому что части функционального хромофора разделены спейсером. Спейсер заключен в рамку двумя идентичными специфическими протеазоспецифическими сайтами расщепления. При расщеплении активированной протеазой спейсер вырезался бы и две остаточные "субъединицы" флуоресцирующего белка могли бы повторно быть реассемблированы для получения функционального флуоресцирующего белка. Оба из описанных выше применений могли бы быть развиты в анализах для ряда различных типов протеаз, таких как каспазы и другие.
Заявленные белки также могут использоваться в анализах для определения фосфолипидного состава в биологических мембранах. Например, белки слияния целевых белков (или любой другой вид ковалентной или нековалентной модификации заявленных белков), которые допускают связывание со специфическими фосфолипидами для локализации/визуализации в модели фосфолипидного распределения в биологических мембранах, тогда как допускающие солокализацию мембранные белки в специфических фосфолипидных очагах могут быть ассоциированы с заявленными белками. Например, PH домен GRP1 имеет высокое сродство к фосфатидил-инозитолтрифосфату (PIP3), но не к PIP2. Также белок слияния между PH доменом GRP1 и заявленными белками может быть создан для специфического мечения PIP3-обогащенных участков в биологических мембранах.
Заявленные флуоресцирующие белки также находят применение как биосенсоры в прокариотических и эукариотических клетках, такое как Ca2+ ионный индикатор; pH индикатор; индикатор фосфорилирования; или как индикатор других, таких как магний, натрий, калий, хлорид и галогенид ионов. Способы использования флуоресцирующих белков в качестве биосенсоров также включают таковые, описанные в патентах США №№ 5972638; 5824485 и 5650135 (так же, как и ссылки, цитированные там), раскрытие которых включены сюда ссылкой.
Антитела заявленные в изобретении, описанные выше, также находят использование во множестве применений, включая дифференцирование заявленных белков от других флуоресцирующих белков.
Наборы
Также обеспечиваются в соответствии с настоящим изобретением наборы для использования при осуществлении одного или более вышеописанных применений. В предпочтительных воплощениях наборы могут использоваться для мечения биологической молекулы. Наборы обычно включают белок изобретения или нуклеиновую кислоту, его кодирующую, предпочтительно с элементами для экспрессии заявленных белов, например, конструкция, такая как вектор, включающий нуклеиновую кислоту, кодирующую заявленный белок. Изобретение также охватывает средства для получения таких компонентов наборов. Указанный способ может включить кДНК из медузы Hydrozoa и пары олигонуклеотидных праймеров для получения нуклеиновой кислоты, заявленной в изобретении, например, с помощью ПЦР, или указанные средства могут включать множество фрагментов нуклеиновой кислоты, которые при лигировании могут давать нуклеиновую кислоту, кодирующую флуоресцирующий белок настоящего изобретения, и т.д. Компоненты наборов обычно присутствуют в соответствующей среде для хранения, такой как забуференный раствор, обычно в соответствующей емкости. Также в наборах могут присутствовать антитела, специфические к указанному белку. В отдельных воплощениях набор включают множество различных векторов, каждый их которых кодирует заявленный белок, где векторы предназначены для экспрессии в различных средах и/или при различных условиях, например, для конститутивной экспрессии, где вектор включает сильный промотор для экспрессии в клетках млекопитающих или вектор с ослабленным промотором с множественными клонированными сайтами для выборочной вставки промотора и нестандартной экспрессии, и т.д.
В дополнение к описанным выше компонентам заявленные наборы будут дополнительно включать инструкции для осуществления заявленных способов. Эти инструкции могут присутствовать в заявленных наборах в различных формах в количестве одна или более, которые могут присутствовать в наборах.
Следующие примеры предлагается в качестве иллюстративных, но не ограничивающих.
ПРИМЕРЫ
Пример 1
Клонирование, секвенирование и получение рекомбинтного белка phiYFP
Яркая желтая флюоресценция в гидромедузе Phialidium Sp. (Cnidaria; Hydrozoa; Hydroida; Leptomedusae; Campanulariidae) была детектирована с использованием флуоресцентного микроскопа. Для нахождения белка, отвечающего за флюоресценцию в этой медузе, была выбрана стратегия, основанная на скрининге экспрессионной библиотеки кДНК в E. coli. Амплифицированные образцы кДНК были приготовлены с использованием набора для амплификации кДНК SMART (Clontech) и клонировались в PCR-Script вектор (Stratagene). Приблизительно 105 рекомбинантных клонов были скринированы визуально с использованием флуоресцентного стереоскопического микроскопа. Два флуоресцентных клона, кодирующие такие же желтые флуоресцирующие белки, были найдены и были названы phiYFP. Нуклеиновая кислота и аминокислотные последовательности для phiYFP показаны в SEQ NO: 01, 02 и 23. Сравнение phiYFP с GFP A. victoria показано на фиг.1. Как оказалось, phiYFP более сходный с GFP (50% идентичности), чем с флуоресцирующими белками из производных кораллов.
Для облегчения очистки белков кодирующий участок phiYFP гена был клонирован в экспрессирующий вектор pQE30 (Qiagen) так, чтобы рекомбинантный белок содержал шесть-гистидиновую метку в его N-конце. После экспрессии в E. coli, phiYFP белок был очищен с помощью металл-аффинной смолы TALON (Clontech). Спектры возбуждения-излучения для phiYFP имели пики при 525 нм и 537 нм (фиг.2A), соответственно. В отличие от дикого типа GFP A. victoria новый белок обладает только одним пиком поглощения-излучения, вероятно в соответствии с депротонированным состоянием хромофора.
Пример 2
Мутагенез PhiYFP
Последовательность PhiYFP, кодирующая нуклеиновую кислоту, была получена, как описано выше в Примере 1. Заявителем был модифицирован неспецифическим мутагенезом кодирующий белок дикого типа. Неспецифический мутагенез phiYFP привел к получению более яркого мутанта, названного phiYFP-Y1, с немного измененным спектром возбуждения-излучения. Этот мутант содержал три аминокислотных замены, в частности S2P, E174G, I201M (SEQ ID NO: 03, 04, и 24). phiYFP-Yl показал яркость в от 1,5 до 2 раз выше, чем дикий тип phiYFP при двупольном визуальном сравнении колоний E. coli, экспрессирующих эти флуоресцирующие белки. Кроме того, phiYFP-Yl демонстрирует спектр излучения, немного смещенный в красную область, с пиком при 542 нм (см. фиг.2B).
Белки как phiYFP, так и phiYFP-Yl, как было найдено, представляют собой димеры. Это было показано белковым гель-электрофорезом не нагретых белковых образцов (см. Baird et al., выше, 2000). При этих условиях эти FPs перемещались как желтая флуоресцентная полоса приблизительно в 50 кДа. Гель-фильтрационные испытания показали димерное состояние phiYFP и phiYFP-Yl. Очищенные белковые образцы (˜1 мг/мл) были загружены на колонку Sephadex-100 (0,7×60 см) и элюировались раствором 50 мМ фосфатного буфера (рН 7,0) и 100 мМ NaCl. EGFP, HcRedl и DsRed2 (Clontech) использовались в качестве мономерных, димерных и тетрамерных стандартов, соответственно.
Сайт-направленный мутагенез использовался для увеличения степени мономерности варианта phiYFP-Y1. Было введено шесть аминокислотных замен, в частности V103N, M166R, Y198N, T202S, Т206К, V221K. Полностью этот мутант phiYFP-MO нес 9 замещений: S2P, V103N, M166R, E174G, Y198N, I201M, T202S, Т206К, V221K (SEQ ID NOs: 05, 06, и 25). phiYFP-MO показывал низкое сворачивание белка и низкую яркость, когда он был экспрессирован в Е. coll. Его спектры возбуждения-излучения были смещены в синюю область по сравнению материнским мутантом (максимумы при 517 и 529 нм, соответственно; фиг.2С). phiYFP-MO являлся более мономерным белком в соответствии с гель-фильтрационными тестами.
Для улучшения phiYFP-MO заявитель применял неспецифический мутагенез. Diversity PCR Random Mutagenesis kit (CLONTECH) использовался в условиях, оптимальных для 5-6 мутаций на 1000 п.о. Колонии Е. coli, экспрессирующие мутантные белки, визуально скринировались флуоресцентным стереоскопическим микроскопом SZX-12 (Olympus). Самый яркий клон со спектром, очевидно смещенным в красную область (по сравнению с исходным phiYFP-MO), был охарактеризован дополнительно. Этот мутант, обозначенный как phiYFP-M1, содержал следующие аминокислотные замены: E88D, V103N, М166С, E174G, I201M, T202S, Т206К, V221K (SEQ ID NOs: 07, 08, и 26). Спектры возбуждения-излучения для этого белка имели пики при 524 и 539 нм, соответственно, аналогично пикам phiYFP дикого типа (фиг.2). Очищенный phiYFP-M1 имел молярный коэффициент экстинкции 130000 М-1см-1 и флуоресцентный квантовый выход 0,40. При определении молярного коэффициента экстинкции заявитель полагался на оценку концентрации зрелого хромофора. Белок денатурировался равным объемом щелочи 2М NaOH. При этих условиях GFP-подобный хромофор поглощает при 446 нм, и его молярный коэффициент экстинкции составляет 44000 M-1см-1 (Ward, W. W. Properties of the coelentrate green-fluorescent protein, in Bioluminescence and Chemiluminescence. Academic Press (1981), 235-242). Были измерены спектры поглощения нативного и денатурированного щелочью phiYFP-M1. Молярный коэффициент экстинкции для нативного состояния белка были оценены, основываясь на поглощении денатурируемого белка. Для определения квантового выхода флюоресценция phiYFP-M1 сравнивалась с одинаково поглощающим EGFP (квантовый выход 0,60 ((Patterson et al., J. Cell. Sci. (2001), 114: 837-838)). phiYFP-M1 представлял собой слабодимерный белок в соответствии с гель-фильтрационными тестами.
Для усиления экспрессии в клетках млекопитающих заявитель синтезировал "гуманизированную" версию phiYFP-M1 с использованием кодонов, оптимизированных для млекопитающих (SEQ ID NOs: 09, 10, и 27). "Гуманизированная" версия phiYFP-M1 была подвергнута сайт-направленному и неспецифическому мутагенезу для получения версий белка, излучающих свет в зеленой и голубой области. Были получены мутантные флуоресцентные белки с зеленой и голубой флюоресценцией. Зеленый мутант гуманизированного phiYFP-M1, названный phiYFP-M1G1, содержит следующие аминокислотные замены (по сравнению с phiYFP-M1) T65S, L148Q, Y203T, K231T, T232A (SEQ ID NO: 17, 18 и 31). Голубой мутант гуманизированного phiYFP-M1, названный phiYFP-M1C1, содержит следующие аминокислотные замены (по сравнению с phiYFP-M1): L6Q, T65S, Y66W, N124K, C147Y, L148Q, Y203T, V224L (SEQ ID NO: 19, 20 и 32). Спектры возбуждения-излучения для этого белка показаны на фиг.3A, B.
Пример 3
Клонирование, секвенирование и получение рекомбинтного белка hydr1GFP
Ярко-зеленая флюоресценция была обнаружена в гидромедузе 1 (длиной приблизительно 1 мм, фиг.4) подотряда Anthomedusae (Cnidaria, Hydrozoa, Anthomedusae) с использованием флуоресцентного микроскопа. Для поиска гена, отвечающего за флюоресценцию в этой медузе, была реализована стратегия, основанная на скрининге экспрессионной библиотеки кДНК в E. coli. Амплификация проб кДНК осуществлялась с использованием набора кДНК SMART (Clontech) и клонировались в вектор PCR-Script (Stratagene). Приблизительно 105 рекомбинантных клонов были визуально скринированы с использованием флуоресцентного стереоскопического микроскопа. Три флуоресцентных клона были идентифицированы, где каждый кодировал такой же зеленый флуоресцирующий белок, который назван hydr1GFP. Нуклеотидная и аминокислотная последовательности для этого белка показаны в SEQ ID NO: 11, 12 и 28. Сравнение hydr1GFP с GFP A. victoria показано на фиг.1. hydr1GFP, как оказалось, имеет большее сходство с GFP (37% идентичности), чем с флуоресцирующими белками из кораллов.
Для облегчения очистки белков кодирующий участок hydr1GFP гена был клонирован в экспрессирующий вектор pQE30 (Qiagen) так, чтобы рекомбинантный белок содержал шесть-гистидиновую метку в его N-конце. После экспрессии в E. coli, hydr1GFP белок был очищен с помощью металл-аффинной смолы TALON (Clontech). Спектры возбуждения-излучения для phiYFP снимались при 474 нм и 494 нм (фиг.5), соответственно. В отличие от дикого типа GFP A. victoria новый белок обладает только одним пиком поглощения-излучения, вероятно в соответствии с депротонированным состоянием хромофора.
Пример 4
Клонирование, секвенирование и получение рекомбинтного белка hm2CP
Ярко-зеленая флюоресценция была обнаружена в малой гидромедузе 2 из подотряда Anthomedusae (Cnidaria, Hydrozoa, Anthomedusae) c использованием флуоресцентного микроскопа. Для поиска FP из этой медузы заявитель выбрал стратегию, основанную на скрининге экспрессионной библиотеки кДНК в E. coli. Амплификация проб кДНК осуществлялась с использованием набора кДНК SMART (Clontech) и клонировалась в вектор PCR-Script (Stratagene). Приблизительно 105 рекомбинантных клонов были визуально скринированы с использованием флуоресцентного стереоскопического микроскопа или невооруженным глазом. Неожиданно заявитель не обнаружил флуоресцирующих клонов. Вместо этого был идентифицирован фиолетовый нефлуоресцирующий СР (hm2CP). Нуклеотидные и аминокислотные последовательности этого белка показаны в SEQ ID NO: 13, 14 и 29. Сравнение hm2CP с GFP показано на фиг.1. hm2CP, как оказалось, представляет собой относительно отдаленный гомолог GFP (до 24% идентичности).
Для облегчения очистки белков кодирующий участок hm2CP был клонирован в экспрессирующий вектор pQE30 (Qiagen) так, чтобы рекомбинантный белок содержал шесть-гистидиновую метку в его N-конце. После экспрессии в E. coli, hm2CP белок был очищен с помощью металл-аффинной смолы TALON (Clontech). Спектр поглощения очищенного hm2CP обладал единственным максимумом при 568 нм (фиг.6). Может быть обнаружена очень слабая красная флюоресценция (максимумы возбуждения при 569 и 597 нм, соответственно) hm2CP (фиг.7).
Пример 5
Мутагенез hm2CP
Последовательность нуклеиновой кислоты, кодирующая hm2CP, была получена как описано выше в Примере 4. Для получения флуоресцирующих мутантов hm2CP заявители использовали неспецифический мутагенез. Набор Diversity PCR Random Mutagenesis (Clontech) использовался для неспецифического мутагенеза hm2CP в условиях, оптимальных для 5-6 мутаций на 1000 п.н. Колонии E. coli, экспрессирующие мутантные белки, визуально скринировались флуоресцентным стереоскопическим микроскопом SZX-12 (Olympus). Самые яркие варианты были выбраны и подвергнуты другому циклу неспецифического мутагенеза. Полные четыре цикла мутагенеза привели к яркому и быстро созревающему красному флуоресцентному мутанту, обозначенному как S3-2. По сравнению с исходным хромопротеином S3-2 имел 13 аминокислотных замены, в частности D24G, I30V, K73R, T91S, II18V, K136R, T145N, S154P, С161А, Y162F, L181M, V199T, I201T (SEQ ID NO: 15, 16, и 30). Спектры возбуждения и излучения для этого мутанта обладали максимумами при 585 и 611 нм, соответственно (фиг.8). Красный флуоресцентный белок S3-2 имеет слабодимерную природу, показанную данными гель-фильтрации. Для усиления экспрессии в клетках млекопитающих заявители синтезировали "гуманизированную" версию S3-2 с использованием кодонов, оптимизированных для млекопитающих (SEQ ID NO: 21, 22, и 33).
Пример 6
Приготовление поликлональных антител
Кодирующие участки нуклеиновых кислот красного флуоресцентного белка S3-2 и желтого флуоресцентного белка Phi-YFP-M1, полученные как описано выше в примерах 2 и 5, соответственно, клонировались в экспрессирующий вектор pQE30 (Qiagen) так, чтобы рекомбинантные белки содержали шесть-гистидиновую метку в N-конце. После экспрессии в Е. Coli, hm2CP был очищен с помощью металл-аффинной смолы TALON (Clontech) в денатурирнующих условиях. Кролики были иммунизированы и дополнительно обрабатывались четыре раза с месячным периодами рекомбинантным полипептидом DSN, эмульгированным в адъюванте Фрейнда. Через десять или 11 дней после каждой дополнительной обработки у животных была взята кровь. Поликлональная антисыворотка была проверена на рекомбинантном белке с помощью ELISA и вестерн-иммуноблотингом.
Пример 7
Мечение клетки млекопитающих, с использованием белков PhiYFP и S3-2.
Для флуоресцентного мечения эукариотических клеток гуманизированные версии белков phiYFP-M1 и S3-2, полученных как описано выше в Примерах 2 и 5, соответственно, клонировались в вектор pEGFP-C1 (CLONTECH) между Agel и BglII рестрикционными участками (вместо EGFP-кодирующего участка). Были использованы следующие клеточные линии: почечные эпителиальные клетки человека 293Т, фибробласты эмбриона мыши ЗТЗ, подкожные фибробласты мыши L929, почечные эпителиальные клетки африканской зеленой обезьяны Vero и фибробласты из почки африканской зеленой обезьяны COS1. Клетки были трансфицированы с использованием реактива LipofectAMINE (Invitrogen) и были протестированы через 20 часов после трансфекции. Флуоресцентный микроскоп Olympus CK40, оснащенный CCD камерой (DP-50, Olympus), использовался для снятия изображений клетки. Экспрессия phiYFP-M1 или S3-2 на различных клеточных линиях привела к ярким желтым или красным сигналам без агрегации. Флюоресценция была ясно обнаружена через 24 часа после трансфекции. Токсичность для клеток не наблюдалась.
Пример 8
Мечение белка и анализ белковой локализации с использованием белков PhiYFP и S3-2.
Гуманизированные версии phiYFP-M1 (содержащего дополнительные замены N21D, Т170А, К221Т) и S3-2 белков, полученных как описано выше в Примерах 2 и 5, соответственно, были слиты с цитоплазматическим бета-актином человека. Трансфекция почечных эпителиальных клеток человека 293Т плазмидами, экспрессирующими phiYFP-M1 или 33-2-меченные слитые конструкции, привела к яркой флюоресценции, которая давала картину, очень похожую с таковой при слияниях с EGFP.
Гуманизированная версия phiYFP-M1 (содержащего дополнительные замены N21D, Т170А, К221Т) была дополнительно слита с ядрышковым белком, фибрилларином. Трансфекция почечных эпителиальных клеток человека 293Т плазмидой, экспрессирующей phiYFP-M1-меченные слитые конструкции, привела к яркой флюоресценции со структурой, соответствующей ожидаемой.
Пример 9
Мечение митохондрий с использованием PhiYFP
Кодирующие последовательности гуманизированной версии phiYFP-M1, полученные как описано выше в Примере 2, были слиты с митохондриальной целевой последовательностью (MTS) из субъединицы VIII цитохромоксидазы С человека. Трансфекция почечных эпителиальных клеток человека 293Т плазмидами, экспрессирующими phiYFP-M1-MTS слитую конструкцию, привела к эффективной транслокации белка в митохондрий клеток-хозяев. Флюоресценция была ясно обнаружена спустя 24 часа после трансфекции.
Пример 10
Мечение аппарата Гольджи с использованием PhiYFP
Кодирующие последовательности гуманизированной версии phiYFP-M1, полученные как описано выше в Примере 2, были слиты с последовательностью, кодирующей 81 аминокислоту на N-конце бета 1,4-галактозилтрансферазы человека (GT; Watzele & Berger (1990) Nucleic Acids. Res. 18:7174). Эта область бета-1,4-GT человека содержит сигнальный пептид связывания с мембраной, которая нацеливает слитые белки на трансоболочечную область аппарата Гольджи (Llopis et al. Proc. Natl. Acad. Sci. USA (1998) 95: 6803-6808; Yamaguchi & Fukuda J. Biol. Chem. (1995)270: 12170-12176; Gleeson et al. Glycoconjugate J. (1994) 11: 381-394). Трансфекция почечных эпителиальных клеток человека 293T плазмидами, экспрессирующими phiYFP-M1-меченную слитую конструкцию, привела к флуоресцирующему мечению трансоболочечного участка аппарата Гольджи в клетках.
Пример 11
Мечение пероксисомы с использованием PhiYFP
Кодирующие последовательности гуманизированной версии phiYFP-M1, полученные как описано выше в Примере 2, были слиты с пероксисомальным нацеливающим сигналом 1 (PTS1). Последовательность PTS1, кодирующая трипептид SKL, который нацеливает белок слияния к матрице пероксисомы (Gould et al. J. Biol. Chem. (1989) 108: 1657-1664; Gould et al. EMBO J. (1990) 9: 85-90; Monosov et al. J. Histo. Cytochem. (1996) 44: 581-589)). Трансфекция почечных эпителиальных клеток человека 293T плазмидами, экспрессирующими phiYFP-M1-меченную слитую конструкцию, привела к флуоресцирующему мечению пероксисом.
Пример 12
Мечение ядра с использованием PhiYFP
Кодирующие последовательности гуманизированной версии phiYFP-M1, полученные как описано выше в Примере 2, были слиты с тремя копиями сигнала ядерной локализации (NLS) больших T-антигенов вируса обезьяны 40, на его C-конце (Kalderon et al. Cell (1984) 39: 499-509; Lanford et al. Cell (1986) 46: 575-582). Трансфекция человеческих почечных эпителиальных клеток 293T плазмидами, экспрессирующими phiYFP-M1-меченную слитую конструкцию, привела к флуоресцирующему мечению ядер.
Все публикации и патентные заявки, цитируемые в настоящем описании, вводятся в настоящее описание посредством ссылки, как если бы каждая отдельная публикация или патентная заявка была конкретно и отдельно введена посредством ссылки. Цитирование любой публикации приводится в соответствии с контекстом и интерпретацией по настоящему изобретению и не должно истолковываться как признание любой такой публикации прототипом данного изобретения.


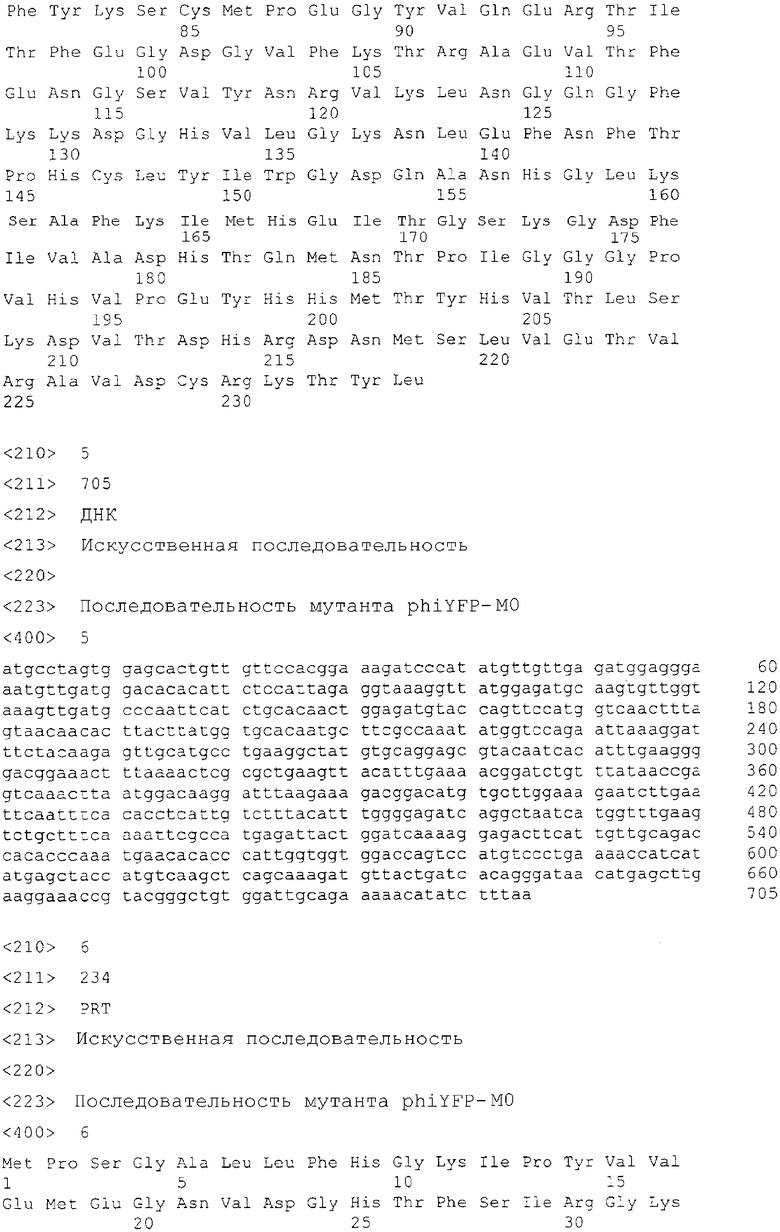













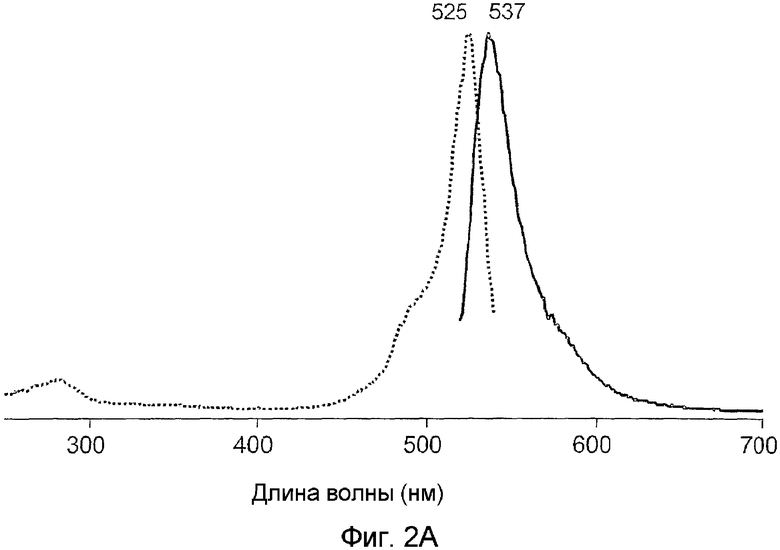
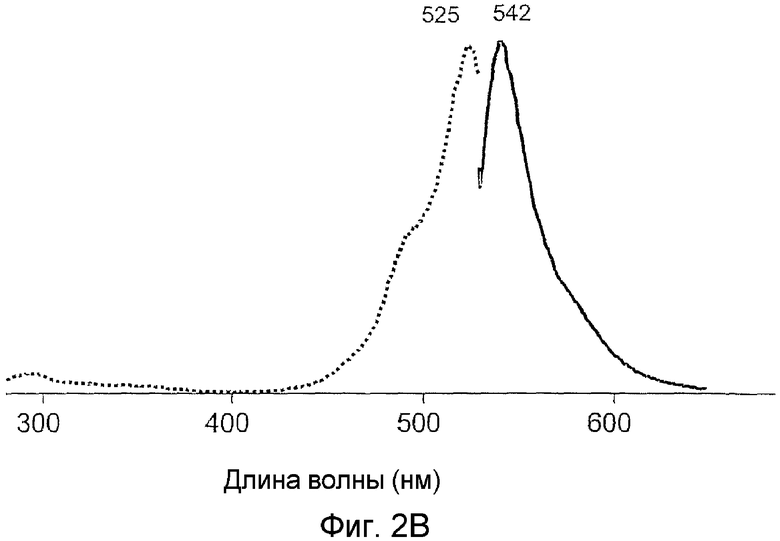
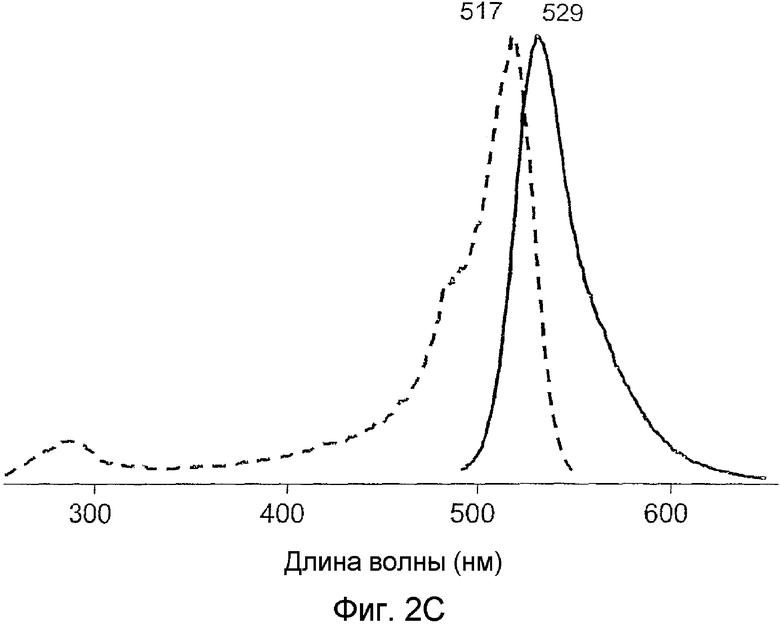
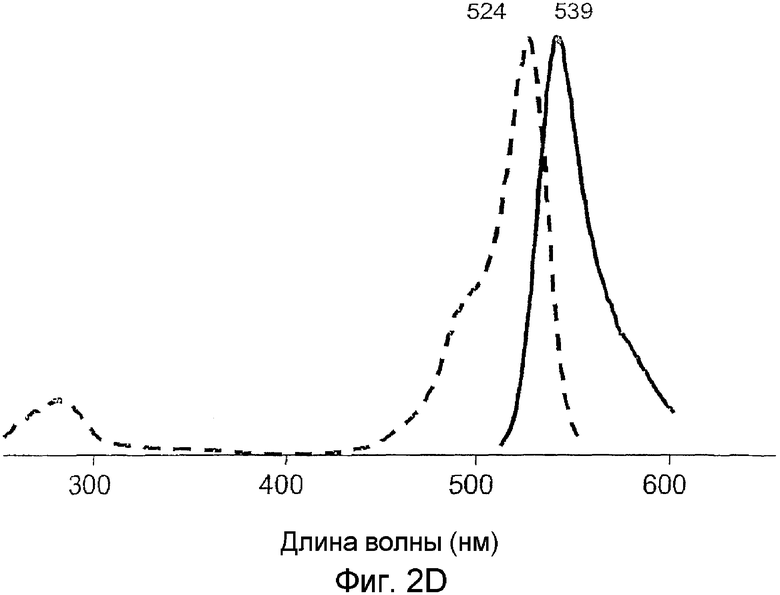
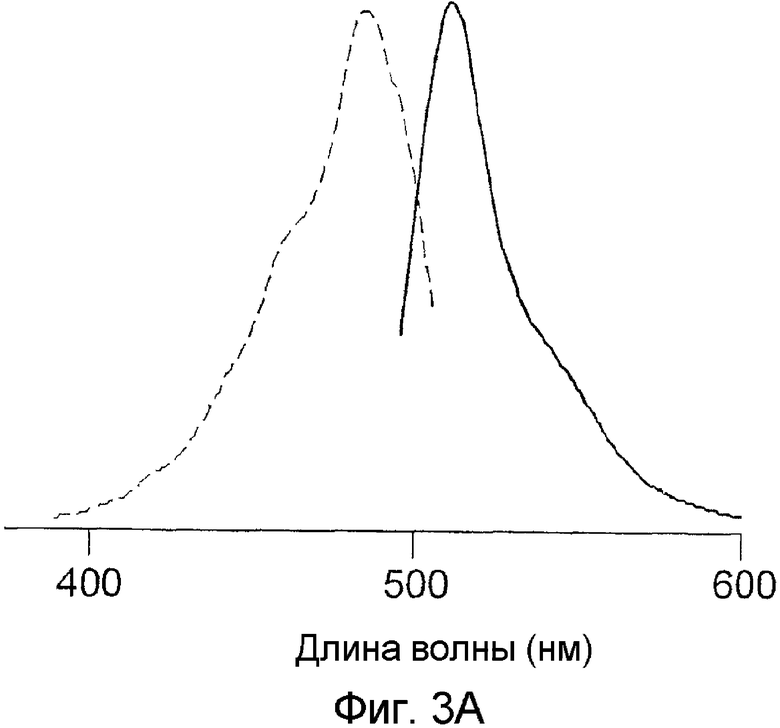
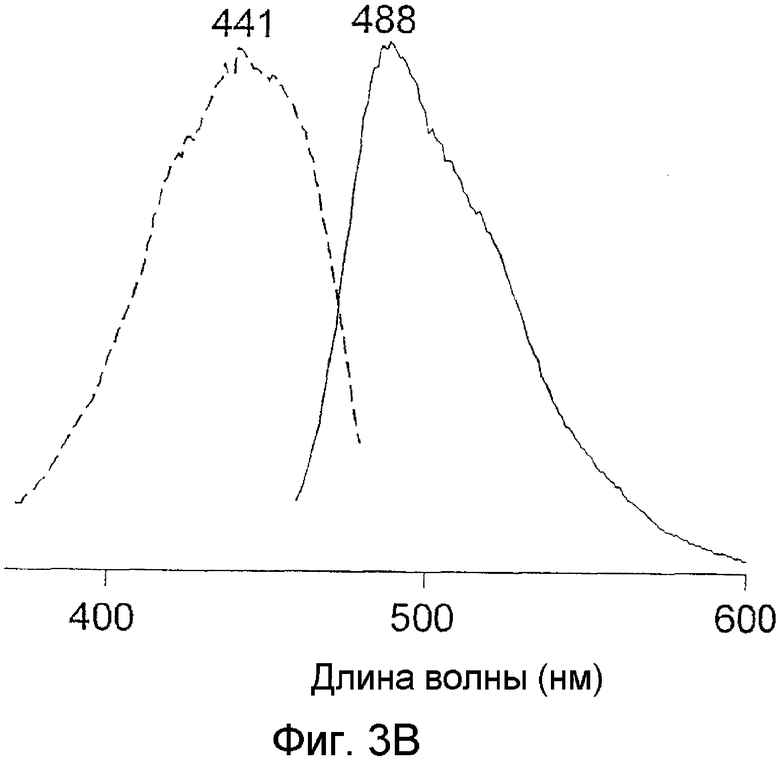
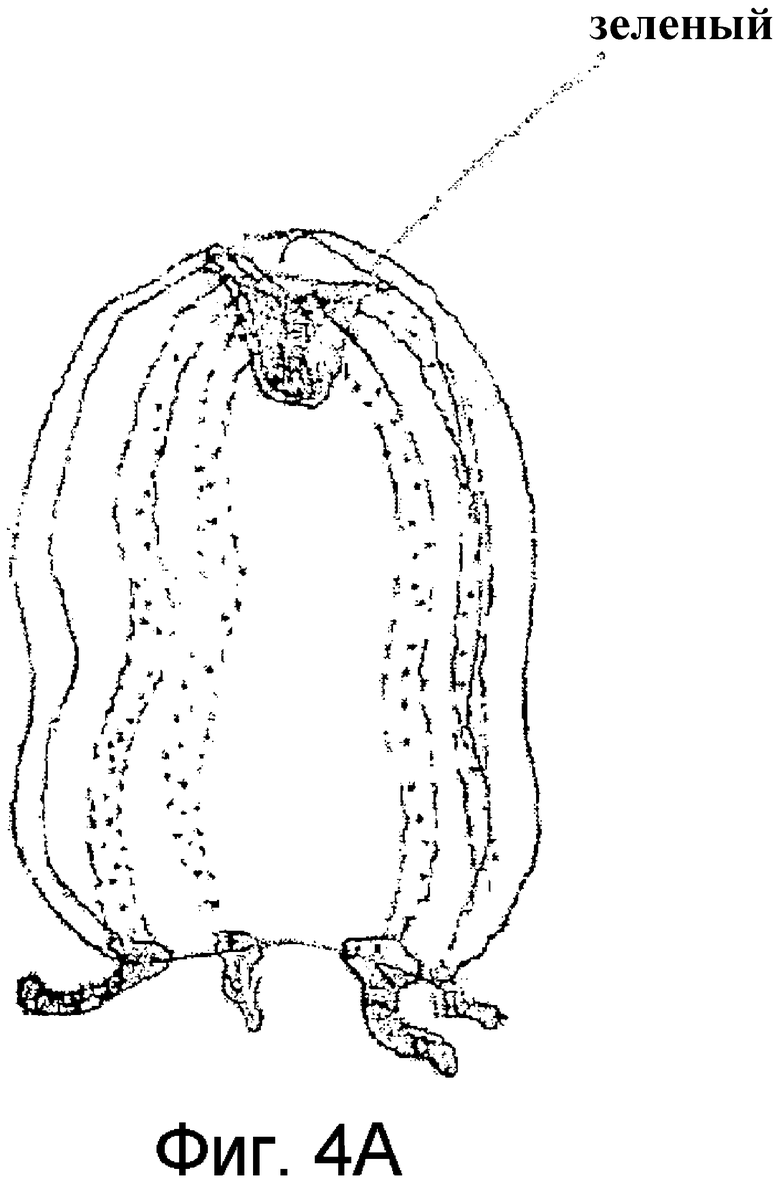
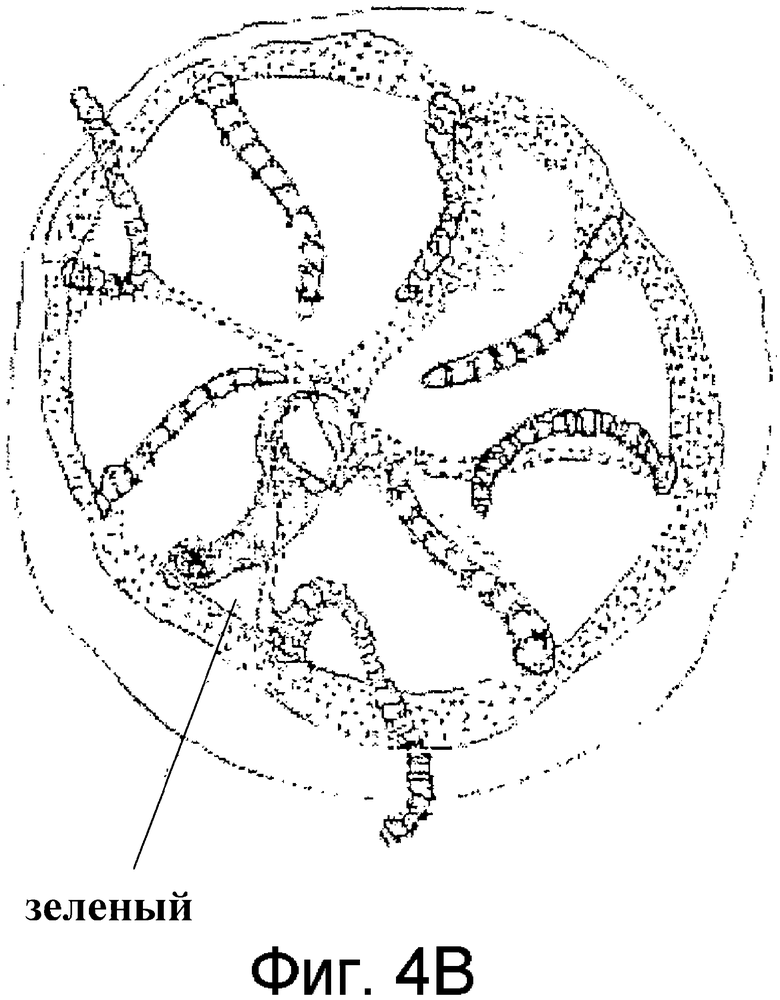
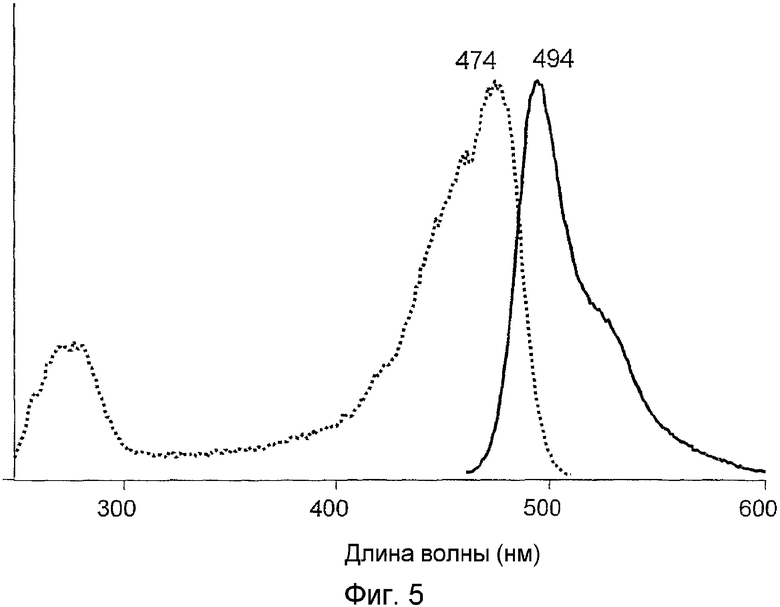
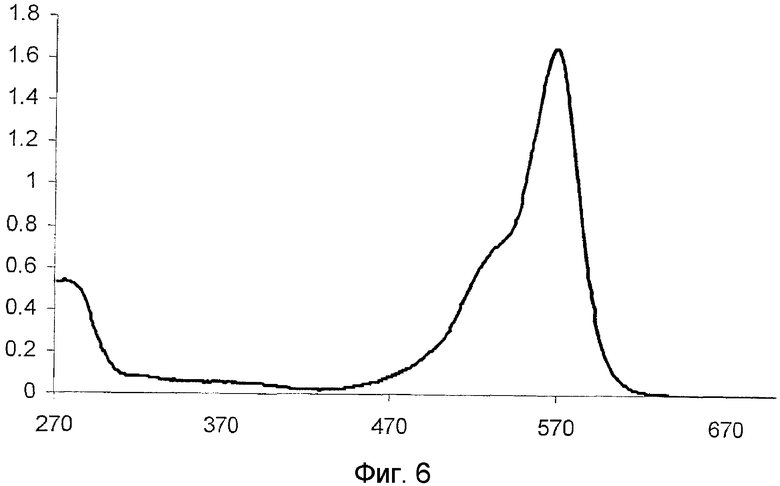
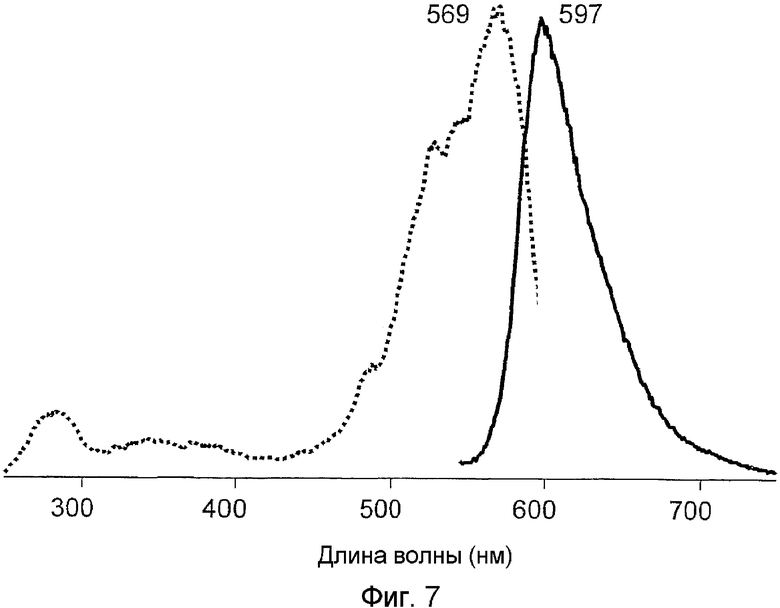
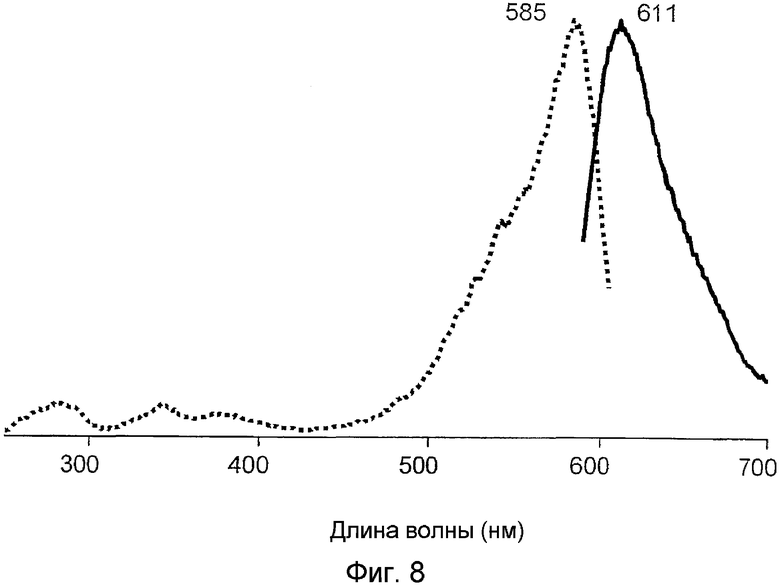
Изобретение относится к области генной инженерии и биотехнологии и может быть использовано для мечения биологических объектов. Выделена молекула нуклеиновой кислоты, которая кодирует флуоресцирующий белок, выбранный из флуоресцирующих белков представителей вида Phialidium sp.и подотряда Anthomedusae и флуоресцирующих мутантов указанных белков. С помощью выделенной нуклеиновой кислоты получены клонирующий и экспрессирующий вектора, флуоресцирующий белок, белок слияния, способный к флуоресценции, а также экспрессирующая кассета. Клетка и стабильная клеточная линия, содержащие такую экспрессионную кассету, продуцируют флуоресцирующий белок. Флуоресцирующий белок, кодирующую его нуклеиновую кислоту и экспрессионные генетические конструкции, содержащие эту нуклеиновую кислоту, используют в наборе для мечения биологической молекулы. Флуоресцентный белок используют также в способах мечения биологической молекулы, клетки или клеточной органеллы. Применение изобретения позволяет расширить арсенал средств для мечения биологических объектов. 13 н.п. ф-лы, 12 ил.
(a) нуклеиновой кислоты, которая кодирует флуоресцирующий белок, характеризующийся аминокислотной последовательностью, по существу соответствующей аминокислотной последовательности, показанной в SEQ ID NO:2, 4, 6, 8, 12, 16, 18 или 20;
(b) нуклеиновой кислоты, характеризующейся нуклеотидной последовательностью, по существу соответствующей нуклеотидной последовательности, показанной в SEQ ID NO: 1, 3, 5, 7, 9, 11, 15, 17, 19 или 21;
(c) нуклеиновой кислоты, которая гибридизуется в жестких условиях с нуклеиновой кислотой (а) или (b), указанных выше;
(d) нуклеиновой кислоты, которая кодирует белок, который имеет по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью (а), указанной выше;
(e) нуклеиновой кислоты, которая имеет по меньшей мере 85% идентичность последовательности с нуклеотидной последовательностью (b), указанной выше.
(a) белка, характеризующегося аминокислотной последовательностью, по существу соответствующей аминокислотной последовательности, показанной в SEQ ID NO: 2, 4, 6, 8, 12, 16, 18 или 20;
(b) белка, кодируемого молекулой нуклеиновой кислоты, по существу соответствующей нуклеотиднои последовательности, показанной в SEQ ID NO: 1, 3, 5, 7, 9, 11, 15, 17, 19 или 21;
(с) белка, который имеет по меньшей мере 95% идентичность последовательности с аминокислотной последовательностью (а) или (b), указанных выше.
Пп.1-13 в части SEQ ID NO: 1, 2, 3, 4 имеют приоритет от 12.11.2002 (по дате подачи заявки №60425570 в Патентное Ведомство США), в части SEQ ID NO: 5, 6, 7, 8, 9 имеют приоритет от 21.04.2003 (по дате подачи заявки №60464258 в Патентное Ведомство США), в части SEQ ID NO: 11, 12 имеют приоритет от 27.11.2002 (по дате подачи заявки №60429795 в Патентное Ведомство США), в части SEQ ID NO: 15, 16 имеют приоритет от 20.06.2003 (по дате подачи заявки №60480080 в Патентное Ведомство США), в части SEQ ID NO: 17, 18, 19, 20 имеют приоритет от 05.11.2003 (по дате подачи заявки PCT/RU03/00474).
| CUBITT et al | |||
| Understanding, improving and using green fluorescent proteins | |||
| Trends Biochem Sci | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| IGFL A | |||
| CHAIN A, Structure of Green Fluorescent Protein, Aug | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| US 5976796, 02.11.1999 | |||
| СПОСОБ ОЦЕНКИ ТЯЖЕСТИ ЭНДОГЕННОЙ ИНТОКСИКАЦИИ ПРИ НЕОТЛОЖНОЙ АБДОМИНАЛЬНОЙ ПАТОЛОГИИ В РАННЕМ ПОСЛЕОПЕРАЦИОННОМ ПЕРИОДЕ | 2006 |
|
RU2331882C1 |
| RU 2000131219 A, 27.10.2002. | |||

