Настоящее изобретение относится к получению и применению вариантов группы 5 аллергенов Pooideae, которые характеризуются сниженной IgE реактивностью по сравнению с известными аллергенами дикого типа и в то же время существенно сохраненной реактивностью с Т-лимфоцитами.
Такие гипоаллергенные варианты аллергенов могут использоваться для специфической иммунотерапии (гипосенсибилизация) пациентов с аллергией на пыльцу трав или для превентивной иммунотерапии аллергий на пыльцу трав.
Предпочтительное воплощение данного изобретения относится к вариантам основного аллергена PhI р 5а пыльцы тимофеевки луговой (Phleum pratense).
Предпосылки изобретения
Аллергии типа 1 имеют значение во всем мире. Вплоть до 20% населения в промышленно развитых странах страдают от осложнений, таких как аллергический ринит, конъюнктивит или бронхиальная астма. Эти аллергии вызываются аллергенами, которые присутствуют в воздухе (аэроаллергенами) и которые высвобождаются из источников различного происхождения, таких как пыльца растений, клещи, кошки или собаки. Вплоть до 40% таких людей, страдающих аллергией типа 1, в свою очередь, демонстрируют специфическую IgE реактивность с аллергенами пыльцы трав (Freidhoff и др., 1986, J. Allergy Clin. Immunol. 78, 1190-2002).
Вещества, которые запускают аллергию типа 1, представляют собой белки, гликопротеины или полипептиды. После поглощения через слизистые оболочки эти аллергены реагируют с молекулами IgE, связанными с поверхностью тучных клеток у чувствительных индивидуумов. Если две молекулы IgE сшиваются друг с другом с помощью аллергена, то это приводит к высвобождению медиаторов (например, гистамина, простагландинов) и цитокинов эффекторными клетками и, таким образом, к соответствующим клиническим симптомам.
Делается различие между основными и второстепенными аллергенами в зависимости от относительной частоты, с которой молекулы индивидуального аллергена реагируют с IgE антителами пациентов, страдающих аллергией.
В случае тимофеевки луговой (Phleum pratense), PhI р 1 (Petersen и др., 1993, J. Allergy Clin. Immunol. 92: 789-796), PhI p 5 (Matthiesen и Iowenstein, 1991, Clin. Exp.Allergy 21: 297-307; Petersen и др., 1992, Int. Arch. Allergy Immunol. 98: 105-109), PhI p 6 (Petersen и др., 1995, Int. Arch. Allergy Immunol. 108, 49-54). PhI p 2/3 (Dolecek и др., 1993, FEBS 335 (3), 299-304), PhI p 4 (Haavik и др., 1985, Int. Arch. Allergy Appl. Immunol. 78: 260-268; Valenta и др., 1992, Int. Arch. Allergy Immunol. 97: 287-294, Fischer и др., 1996, J. Allergy Ciln. Immunol. 98: 189-198) и PhI p 13 (Suck и др., 2000, Clin. Exp. Allergy 30: 324-332; Suck и др., 2000, Clin. Exp. Allergy 30: 1395-1402) были к настоящему времени идентифицированы в качестве основных аллергенов.
Доминантные основные аллергены тимофеевки луговой (Phleum pratense) представляют собой PhI р 1 и PhI р 5, при этом PhI р 5 существует в двух формах 5а и 5b, которые отличаются в отношении своего молекулярного веса и кодируются независимыми генами. Дедуцированные аминокислотные последовательности как PhI р 5а, так и PhI р 5b были определены с помощью техники рекомбинантной ДНК. PhI р 5а представляет собой белок с массой приблизительно 32 кДа, который реагирует с IgE антителами у 85-90% пациентов, страдающих аллергией на пыльцу трав. PhI р 5а существует в серии гомологичных вариантов, которые отличаются друг от друга точкой мутаций и, вероятно, соответствуют различным аллельным формам. Пыльца родственных видов трав, таких как, например, Lollum perenne, Роа pratensis среди прочих, содержит аллергены, которые являются гомологичными с таковыми PhI р 5а, и вместе они являются известными как группа 5 аллергенов. Высокая структурная гомология этой группы 5 аллергенов различных видов трав соответственно служит причиной перекрестной реактивности этих молекул с IgE антителами пациентов, страдающих аллергией на пыльцу трав.
Классический подход к эффективному терапевтическому лечению аллергий представляет собой специфическую иммунотерапию или гипосенсибилизацию (Fiebig, 1995, Allergo J. 4 (6): 336-339, Bousquet и др., 1998, J. Allergy Clin. Immunol. 102 (4): 558-562). В этом способе пациенту вводят подкожно экстракты природного аллергена в увеличивающихся дозах. Однако в этом способе существует риск аллергических реакций или даже анафилактического шока. Для того чтобы минимизировать этот риск, используются прогрессивные препараты в форме аллергоидов. Они представляют собой экстракты химически модифицированных аллергенов, которые обладают значительно сниженной IgE реактивностью, но идентичной Т-клеточной реактивностью по сравнению с необработанным экстрактом (Fiebig, 1995, Allergo J. 4 (7): 377-382).
Еще более существенная оптимизация терапии будет возможной с аллергенами, полученными с помощью рекомбинантных способов. Определенные смеси аллергенов с высокой степенью очистки, полученные с помощью рекомбинантных способов и необязательно подобранные к индивидуальным моделям сенсибилизации пациентов, могут заменить экстракты, полученные из источников природных аллергенов, поскольку они в дополнение к различным аллергенам содержат относительно высокое количество иммуногенных, но неаллергогенных, вторичных белков.
Реальные перспективы, которые могут приводить к достоверной гипосенсибилизации при использовании рекомбинантных продуктов экспрессии, предлагаются при использовании специфическим образом мутированных рекомбинантных аллергенов, в которых эпитопы IgE являются специфически делетированными при отсутствии повреждения Т-клеточных эпитопов, которые являются существенными для терапии (Schramm и др., 1999, J. Immunol. 162: 2406-2414).
Дополнительная возможность для терапевтического влияния на нарушенное равновесие Т-хелперных клеток у индивидуумов, страдающих аллергией, представляет собой лечение с помощью способной к экспрессии ДНК, которая кодирует релевантные аллергены (иммунотерапевтическая вакцинация ДНК). Исходное экспериментальное подтверждение специфического для аллергена влияния на иммунный ответ с помощью вакцины ДНК этого типа было продемонстрировано у грызунов с помощью инъекции ДНК, кодирующей аллерген (Hsu и др., 1996, Nature Medicine 2 (5): 540-544).
Задача, на которой основывается настоящее изобретение, основывается на том, что обеспечиваются новые варианты группы 5 аллергенов Pooideae на белковом уровне и на уровне ДНК, которые отличаются пониженной IgE активностью и в то же время существенно сохраняют Т-клеточную реактивность и, таким образом, являются приемлемыми для специфической иммунотерапии и иммунотерапевтической вакцинации ДНК.
Фигуры
Фигура 1: Выравнивание релевантных участков гомологичных последовательностей кДНК PhI р 5а видов Pooideae: Lolium perenne (LoI p), Poa pratensis (Poa p) Triticum aestivum (TrI а) и Hordeum vulgare (Hor v).
Нумерация: положения нуклеотидов вставок ДНК.
Последовательности PhI р 5а, Poa р 5 и LoI р 5: последовательности кДНК из базы данных "GenBank" Национального Центра биотехнологической информации (NCBI), Бетесда, США.
Последовательности Hor v и Tri a: EST последовательности из базы данных EST Института геномных исследований (TIGR), Роквилл, США.
Сплошные границы: идентичность последовательности с PhI р 5а (на основе GenBank AJ 555152).
Пунктирные границы: делеция, соответствующая аминокислотам 94-113 (на основе GenBank AJ 555152).
Штриховые границы: делеция, соответствующая аминокислотам 175-198 (на основе GenBank AJ 555152).
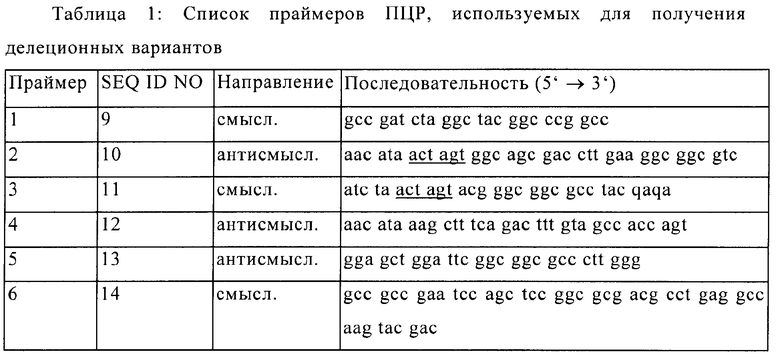
Фигура 2: Выравнивание гомологичных аминокислотных последовательностей PhI р 5а (релевантные участки последовательности, дедуцированной из последовательностей ДНК) видов Pooideae: Solium perenne (LoI p), Poa pratensis (Poa p) Triticum aestivum (TrI а) и Hordeum vulgare (Hor v).
Нумерация: положения нуклеотидов вставок ДНК.
Последовательности PhI р 5а, Роа р 5 и LoI р 5: последовательности кДНК из базы данных "GenBank" Национального Центра биотехнологической информации (NCBI), Бетесда, США.
Последовательности Hor v и Tri a: EST последовательности из базы данных EST Института геномных исследований (TIGR), Роквилл, США.
Сплошные границы: идентичность последовательности с PhI р 5а (на основе GenBank AJ 555152).
Пунктирные границы: делеция, соответствующая аминокислотам 94-113 (на основе GenBank AJ 555152).
Штриховые границы: делеция, соответствующая аминокислотам 175-198 (на основе GenBank AJ 555152).
Фигура 3: SDS-ПАГЭ очищенных делеционных мутантов в форме слитых с гистидином белков
1) Маркер
2) rPhI р 5а дикого типа (His)
3) PhI р 5а DM-Δ94-113 (His)
4) PhI р 5а DM-Δ94-113, 175-198 (His)
5) PhI р 5а DM-Δ175-198 (His)
6) Маркер
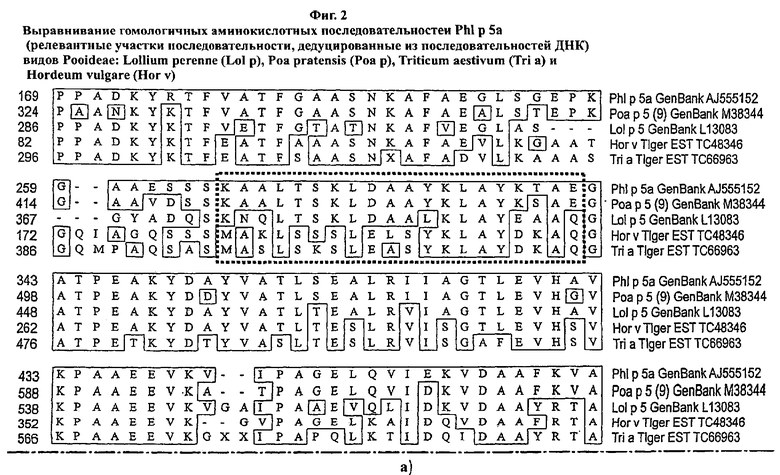
Фигура 4: SDS-ПАГЭ очищенных неслитых белков PhI р 5а DM-Δ94-113, 175-198 и rPhI р 5а дикого типа (вершина) и анализ на идентичность с αPhI р 5 антителами (нижняя часть)
αPhI р 5 монАТ альфа-1D11 связывает участок 175-198 (только rPhI р 5а дикого типа является позитивным)
αPhI р 5а монАТ альфа-3 В2 связывает совместный эпитоп двух молекул PhIp 5а (оба белка являются позитивными)
(монАТ: моноклональное антитело)
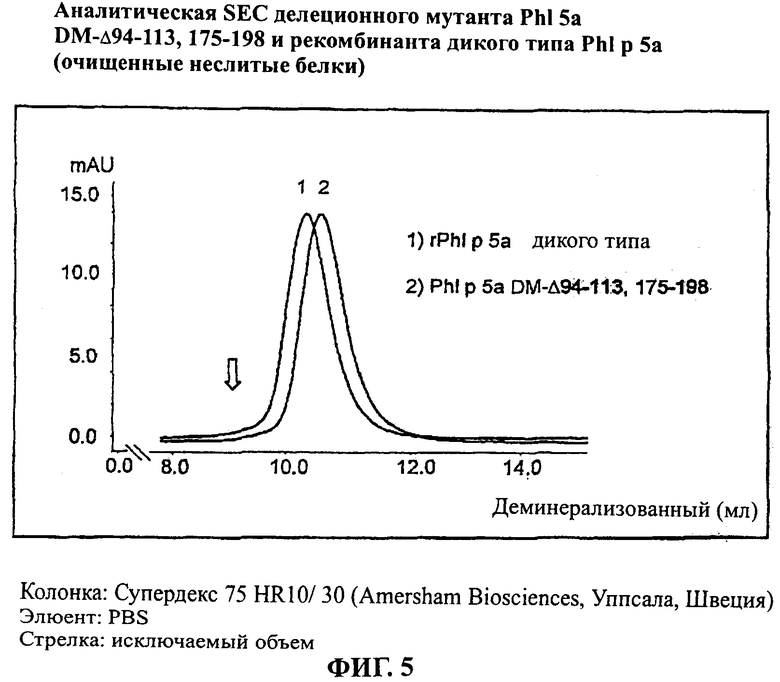
Фигура 5: аналитический анализ на основе SEC делеционного мутанта PhI р 5а DM-Δ94-113, 175-198 и рекомбинанта дикого типа PhI р 5а (очищенные неслитые белки).
Колонка: Супердекс 75 HR10/ 30 (Amersham Biosciences, Уппсала, Швеция)
Элюент: PBS
Стрелка: исключаемый объем
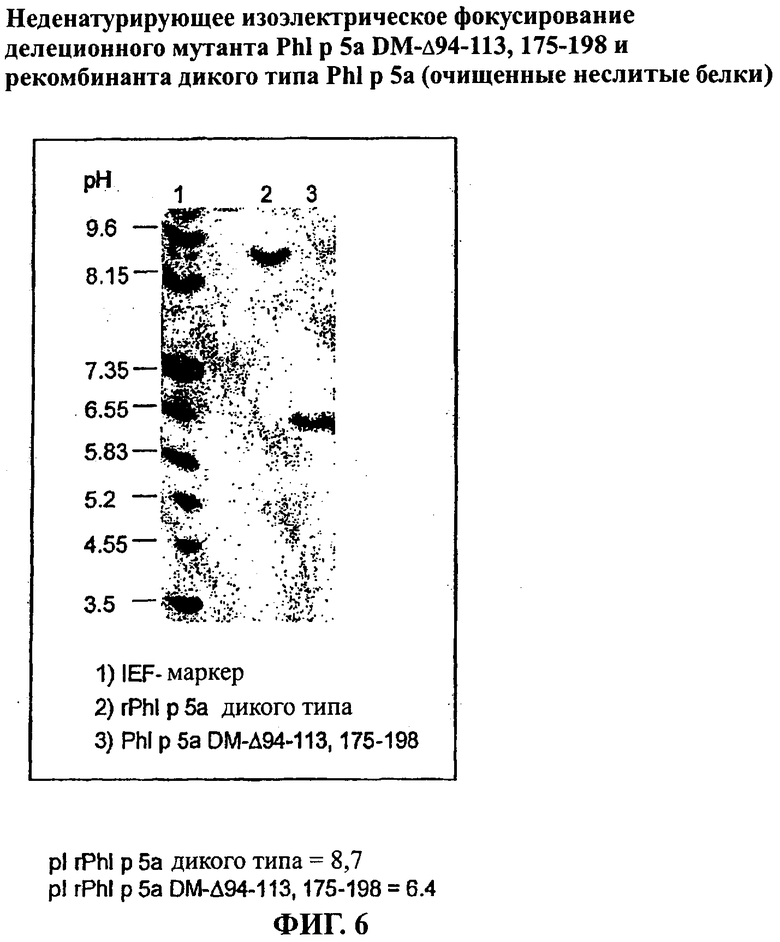
Фигура 6: Неденатурирующее изоэлектрическое фокусирование делеционного мутанта PhI р 5а DM-Δ94-113, 175-198 и рекомбинанта дикого типа PhI р 5а (очищенные неслитые белки)
1) маркер для изоэлектрического фокусирования
2) rPhI р 5а дикого типа
3)PhI p 5a DM-Δ94-113, 175-198
pI rPhI р 5а дикого типа=8,7
pI rPhI р 5а DM-Δ94-113, 175-198=6,4
Фигура 7: Тест-метод для проверки IgE связывающей способности делеционных мутантов PhI р 5а (неденатурирующий)
Р: сыворотка клинически определенных индивидуумов, страдающих аллергией на пыльцу трав
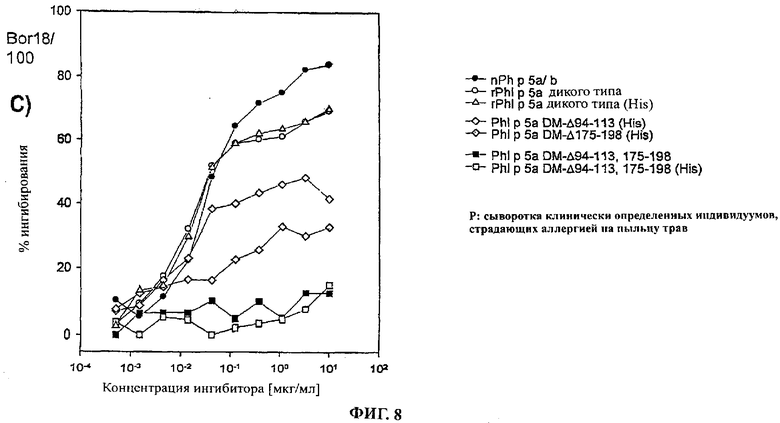
Фигура 8: Определение сниженной IgE реактивности делеционных мутантов PhI р 5а с помощью анализа EAST ингибирования с двумя репрезентативными одиночными сыворотками (а и b) и сывороточным пулом (с)

Р: сыворотка клинически определенных индивидуумов, страдающих аллергией на пыльцу трав
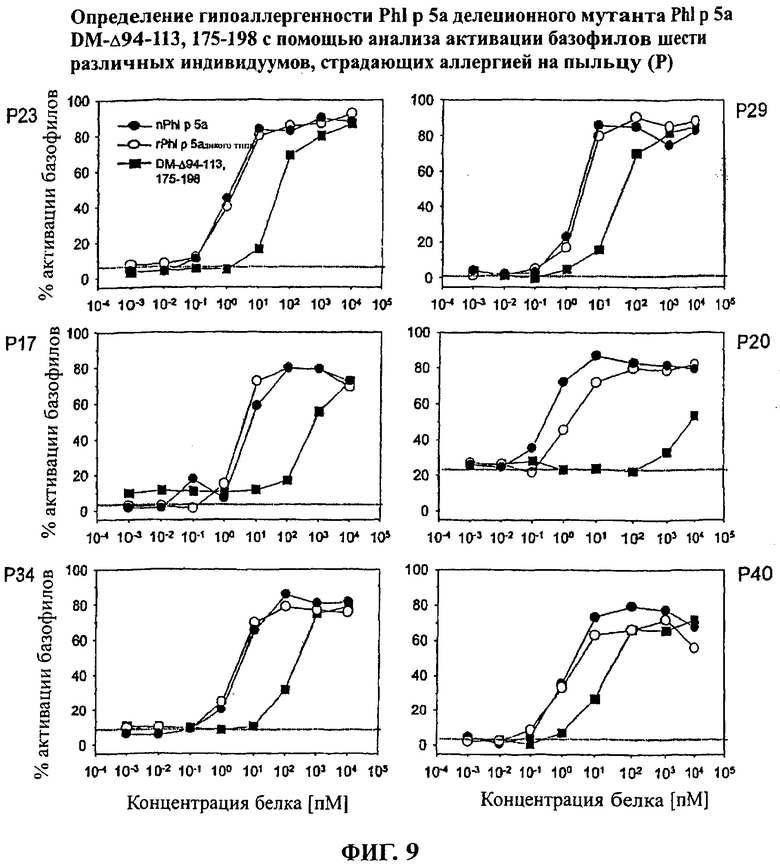
Фигура 9: Определение гипоаллергенности PhI р 5а делеционного мутанта PhI р 5а DM-Δ94-113, 175-198 с помощью анализа активации базофилов шести различных пациентов, страдающих аллергией на пыльцу трав (Р)
Подробное описание изобретения
Мутагенез и клонирование последовательностей кДНК
Отправной точкой для особенно предпочтительных в соответствии с настоящим изобретением гипоаллергенных вариантов PhI р 5а является кДНК изоформы дикого типа PhI р 5а, которая была изолирована с помощью специфических праймеров при использовании полимеразной цепной реакции (ПЦР) из общей кДНК пыльцы тимофеевки луговой (Phleum pratense) (NCBI (Национальный центр биотехнологической информации, Бесезда, США) GenBank номер доступа AJ 555152) (SEQ ID NO 1). Аминокислотная последовательность согласно SEQ ID NO 2 была дедуцирована из последовательности кДНК. PhI р 5а, который состоит из 284 аминокислот, экспрессировали в цитозоле в виде растворимого белка в Е.coli и в дальнейшем подвергали очистке. Эта рекомбинантная форма дикого типа PhI р 5а (rPhI р 5а дикого типа) вступает в реакцию с моноклональными анти-PhI р 5 антителами и с IgE антителами индивидуумов, страдающих аллергией на пыльцу трав, которые обладают реактивностью на природный очищенный PhI p 5а (nPhI р 5а).
Исходя из описанной кДНК rPhI р 5а дикого типа, были получены серии различных делеционных вариантов (делеционных мутантов) с помощью способов рестрикции/лигирования и ПЦР, они были лигированы в экспрессионный вектор pProExHTa (Invitrogen, Карлсбад, США). Участки длиной от 6 до 72 п.о., распределенные по всей длине последовательности молекулы кДНК, были делетированы, что вызвало индукцию соответствующих делеций в полипептидных цепях белков, которые экспрессировались в Е.coli.
Делеционные варианты PhI р 5а оценивали с помощью анализа иммуноблоттинга на их способность связывания с IgE антителами репрезентативного сывороточного пула индивидуумов, страдающих аллергией на пыльцу трав.
В этом способе были неожиданно обнаружены два делеционных мутанта PhI р 5а (PhI р 5а DM-Δ94-113, делеция аминокислот 94-113 и PhI р 5а DM-Δ175-198, делеция аминокислот 175-198 rPhI р 5а дикого типа), которые обладали сниженной способностью связываться с IgE антителами (репрезентативный сывороточный пул). Эти два делеционных варианта PhI р 5а служили в качестве исходной точки для конструирования двойного делеционного мутанта, содержащего обе эффективные делеций (PhI р 5а DM-Δ94-113, 175-198).
Конструирование PhI р 5а DM-Δ94-113, PhI p 5а DM-Δ175-198 и PhI р 5а DM-Δ94-113, 175-198 с помощью способов генетического инжиниринга, а также их биохимическая и иммунологическая характеристика описаны ниже.
Для конструирования делеционного варианта PhI р 5а DM-Δ94-113 (SEQ ID NO 3, последовательность кДНК (795 п.о.) и SEQ ID NO 4, аминокислотная последовательность (264 аминокислоты)), сначала исходя из кДНК rPhI р 5а дикого типа, получали два фрагмента. Фрагмент "F1-93", кодирующий аминокислоты 1-93 rPhI р 5а дикого типа, получали при использовании ПЦР с помощью праймеров 1 и 5, а фрагмент "F114-284" получали с помощью праймеров 4 и 6 (последовательности праймеров смотри в Таблице 1). Фрагменты "F1-93" и "F114-284" использовали в качестве матрицы в дальнейшей ПЦР с помощью праймеров 1 и 4, что приводило к амплификации полной кДНК, кодирующей делеционный вариант PhI р 5а DM-Δ94-113. Основой соединения фрагментов "F1-93" и "F114-284" с помощью ПЦР служил участок последовательности, общий с обоими фрагментами. Этот участок последовательности был получен с помощью амплификации фрагмента "F114-284" посредством ПЦР при использовании индивидуального смыслового олигонуклеотида, который содержит дополнительную последовательность ДНК, кодирующую аминокислоты 88-93 на 5'-участке (Таблица 1).
Последовательность кДНК, кодирующую делеционный вариант PhI р 5а DM-Δ175-198 (SEQ ID NO 5, последовательность кДНК (783 п.о.), и SEQ ID NO 6, аминокислотная последовательность (260 аминокислот)) получали путем рестрикции и последующего лигирования двух отдельно приготовленных фрагментов кДНК. 5'-терминальный фрагмент "F1-174" получали при использовании ПЦР с помощью праймеров 1 и 2, а 3'-терминальный фрагмент "F199-284" с помощью праймеров 3 и 4. Фрагменты кДНК подвергали перевариванию с помощью фермента рестрикции SpeI и последовательно лигировали (смотри Таблицу 1). Продукт лигирования амплифицировали при использовании ПЦР с помощью праймеров 1 и 4.
кДНК делеционного варианта PhI р 5а DM-Δ94-113, 175-198 (SEQ ID NO 7, последовательность кДНК (723 п.о.), и SEQ ID NO 8, аминокислотная последовательность (240 аминокислот)) подобным образом получали из двух фрагментов кДНК. 5'-терминальный фрагмент получали при использовании праймеров 1 и 5, а также кДНК rPhI р 5а дикого типа в качестве матрицы, а 3'-терминальный фрагмент получали при использовании праймеров 4 и 6, а также кДНК PhI р 5а DM-Δ175-198 в качестве матрицы. С помощью общего участка последовательности, соответствующей аминокислотам 88-93 белка rPhI р 5а дикого типа, фрагменты соединяли с помощью третьей ПЦР, используя праймеры 1 и 4, полученный продукт подвергали амплификации.
кДНК, кодирующую модифицированные аллергены, лигировали в экспрессионный вектор pProExHT (Invitrogen, Карлсбад, США) через EheI и HindIII рестрикционные сайты и последовательно подвергали полному секвенированию.
Иммунологическая перекрестная реактивность группы 5 аллергенов Pooideae, таких как, например, Роа pratensis и Lolium perenne, базируется на очень подобной аминокислотной последовательности. Она может быть взята в качестве определенной последовательности, которая соответствует генам, идущим в обратную сторону до общего исходного гена. Гомологичные участки последовательности в группе 5 Pooideae существуют как для последовательностей с делециями Δ94-113 и Δ175-198 белковой последовательности PhI р 5а дикого типа (ссылка: GenBank AJ 555152), а также для участков, которые их фланкируют. Высокая степень гомологии данных участков последовательности может быть продемонстрирована как на уровне ДНК, так и на уровне аминокислотной последовательности (Фиг.1 и Фиг.2).
SpeI сайты рестрикции обозначены с помощью подчеркивания
Экспрессия и очистка рекомбинантных молекул PhI p 5
Рекомбинантные белки экспрессировали в виде белков слияния с гистидином при наличии интегрированного сайта расщепления протеазой (экспрессионный вектор pProExHT; Invitrogen, Карлсбад, США) для необязательного удаления гистидинового компонента слияния (His) в Escherichia coli (штамм JM109). rPhI р 5а дикого типа и делеционные мутанты сначала очищали с помощью специфического связывания N-терминальных остатков гистидина с Ni2+ хелатным матриксом (аффинная хроматография на основе иммобилизованного иона металла, IMAC) при последующем использовании препаративной гель-фильтрации (хроматография на основе исключения по размерам, SEC).
Чистота элюированных белков была проверена с помощью SDS-ПАГЭ и аналитической SEC. Результаты показали, что rPhI р 5а дикого типа (His), PhI р 5а DM-Δ94-113 (His); PhI р 5а DM-Δ175-198 (His) и PhI р 5а DM-Δ94-113, 175-198 (His) могут быть получены с высокой степенью чистоты и в мономерной форме (Фиг.3). Идентичность белков была продемонстрирована с помощью специфических для PhI р 5а моноклональных антител.
В дополнение осуществляли проверку IgE реактивности с помощью методики связывания IgE (иммуноблоттинг, тест-метод, анализ EAST ингибирования и анализ активации базофилов), а также оценку Т-клеточной реактивности с исследуемыми веществами при отсутствии гистидинового компонента слияния.
С этой целью делеционные варианты получали параллельно с белком сравнения rPhI р 5а дикого типа сначала как слитые белки. Однако гистидиновый компонент слияния последовательно отщепляли ферментативно (TEV протеаза, Invitrogen, Карлсбад, США), оставляя только глицин как остаток последовательности отщепления с помощью протеазы на N-терминальном конце белка, представляющего интерес. Как отщепленный гистидиновый компонент, так и протеазу, использованную для отщепления, полностью отделяли с помощью IMAC. После осуществления препаративной SEC чистоту и конформационную структуру элюированных белков проверяли с помощью SDS-ПАГЭ и аналитической SEC, как показано на Фигурах 4 и 5 для rPhl р 5а дикого типа и мутанта PhI р 5а DM-Δ94-113, 175-198 соответственно. Все белки были получены в чистой и мономерной форме. Исследования при использовании неденатурирующего изоэлектрического фокусирования (IEF) неслитых белков показало высокую степень гомогенности в отношении поверхностного заряда (смотри Фиг.6, иллюстративную для PhI р 5а DM-Δ94-113, 175-198).
Идентичность рекомбинантных белков была продемонстрирована с помощью моноклональных анти-PhI p 5 антител (Allergopharma, Рейнбек, Германия) альфа-1D11 или альфа-3В2 (смотри Фиг.4, иллюстративную для PhI p 5а DM-Δ94-113, 175-198) и N-терминального секвенирования.
Определение пониженного связывания с IgE делеционных вариантов PhI р 5а
Простой способ определения IgE реактивности аллергенных молекул представляет собой исследование связывания специфического IgE из сыворотки, полученной от пациентов, страдающих аллергией, со связанными с мембраной исследуемыми белками с помощью тест-метода.
С этой целью исследуемые вещества связывали в одинаковой концентрации, близком количестве с нитроцеллюлозной мембраной в неденатурирующих условиях. Серии тест-методов можно инкубировать параллельно с различными сыворотками индивидуумов, страдающих аллергией. После осуществления этапа промывания специфично связанные IgE антитела визуализировали на мембране с помощью цветной реакции с помощью конъюгата анти-hlgE / щелочная фосфатаза.
IgE реактивность рекомбинантных белков PhI p 5а дикого типа (His), PhI р 5а DM-Δ94-113 (His), PhI p 5а DM-Δ175-198 (His) и PhI р 5а DM-Δ94-113, 175-198 (His) исследовали в сравнении с тест-методом при использовании 43 индивидуальных сывороток, полученных от больных, страдающих аллергией на пыльцу трав (Фиг.7). Все 43 сыворотки, полученные от больных, страдающих аллергией, содержали специфические для PhI p 5а IgE антитела, которые в значительной степени реагировали с природным PhI p 5а (nPhI р 5а, не представлено в данной заявке) и рекомбинантным эквивалентом rPhI р 5а дикого типа (His).
Неожиданно было обнаружено, что специфические для PhI р 5а IgE антитела всех 43 сывороток пациентов вообще не связывались с делеционным вариантом PhI р 5а DM-Δ94-113, 175-198 (His) либо связывались в очень незначительной степени. Пониженная способность к связыванию IgE была также присуща как делеции Δ94-113, так и делеции Δ175-198. Делеционный вариант PhI p 5a DM-Δ175-198 (His) продемонстрировал четко распознаваемую сниженную способность к связыванию IgE в этом анализе в 35 из 43 сывороток, полученных от пациентов, страдающих аллергией. В некоторых анализах влияние делеции аминокислот 175-198 было настолько большим, что связывание IgE фактически полностью предотвращалось (образцы Р3, Р20, Р28)
Влияние делеции Δ94-113 на связывающую реактивность IgE было менее выраженным, но также хорошо видимым. Делеционный вариант PhI р 5а DM-Δ94-113 (His) связывался значительно слабее с IgE в 19 из 43 индивидуальных сывороток, полученных от пациентов, страдающих аллергией, чем стандартный rPhI р 5а дикого типа (His) (образцы: Р31, Р37, Р42). Однако сниженное связывание с IgE было не таким значительно выраженным во многих индивидуальных анализах, чем снижение, вызванное Δ175-198.
Таким образом, понятно, что обе делеции вносят свой вклад в снижение общей связывающей реактивности IgE делеционного мутанта PhI р 5а DM-Δ94-113, 175-198 (His).
В противовес тест-методу анализ EAST ингибирования (ферментативный аллергосорбентный анализ) позволил исследовать взаимодействия аллерген/IgE в растворе, позволяя существенным образом исключить препятствующее маскирование эпитопов исследуемого вещества путем иммобилизации на мембране. Анализ EAST ингибирования осуществляли так, как описано ниже. Микротитровальные планшеты покрывали аллергенами, при этом использовали природный PhI р 5 (nPhI р 5а/b, смесь PhI р 5а и PhI р 5b). После удаления несвязанных молекул аллергена путем промывания планшет блокировали с помощью бычьего сывороточного альбумина для того, чтобы предотвратить неспецифическое связывание. IgE антитела пациентов, страдающих аллергией, либо как репрезентативный пул индивидуальных сывороток (сывороточный пул), либо как одну сыворотку, инкубировали в приемлемом разведении с микротитровальными планшетами, покрытыми аллергеном. Количество связанных с аллергеном IgE антител количественно оценивали фотометрически с помощью фермента, слитого со вторым антителом (конъюгат анти-hlgE/щелочная фосфатаза) посредством превращения субстрата в окрашенный конечный продукт.
Связывание IgE антител ингибировали специфично по отношению к веществу с помощью растворимого аллергена или исследуемого вещества (рекомбинантный модифицированный аллерген) в зависимости от концентрации. Иммунохимически идентичные вещества показали идентичные кривые ингибирования.
Стандартные молекулы, используемые в этой работе, представляли собой nPhI р 5, rPhI р 5а дикого типа и белок слияния с гистидином rPhI р 5а дикого типа (His). Кроме прочих молекул исследовали связывание с IgE белков слияния с гистидином PhI р 5а DM-Δ94-113 (His), PhI р 5а DM-Δ175-198 (His) и PhI р 5а DM-Δ94-113, 175-198 (His) и таковое для неслитого белка PhI p 5а DM-Δ94-113, 175-198 путем сравнения с этими стандартами.
Фигуры 8 а-с репрезентативно демонстрируют кривые специфического ингибирования исследуемых веществ с двумя индивидуальными сыворотками и сывороточным пулом пациентов, страдающих аллергией на пыльцу трав. nPhI р 5а/b показал наибольший ингибиторный эффект во всех анализах (приблизительно 80-95% ингибиторного эффекта при концентрации 10 мкг/мл). Ингибиторный эффект rPhI р 5а был значительно ниже с максимальным ингибированием 70-80%. Этот эффект вызывался композицией nPhI р 5а/b, которая также содержит изоформу PhI р 5b в дополнение к изоформе PhI р 5а. Специфические IgE антитела к PhI р 5b не могут ингибироваться с помощью rPhI р 5а дикого типа.
Гистидиновый компонент слияния не продемонстрировал никакого эффекта на связывание с IgE. Это следует из всех анализов, поскольку кривые ингибирования для rPhI р 5а дикого типа (His) и rPhI р 5а дикого типа являлись идентичными. Это демонстрирует достоверность анализов, проведенных с белками, слитыми с гистидиновым компонентом.
В общем случае две группы сывороток пациентов отличались в отношении качественной стороны связывания с IgE.
Первая группа представлена индивидуальной сывороткой Р15 (Фиг.8а). Эти сыворотки, полученные от больных, страдающих аллергией, содержали IgE антитела, связывание которых с PhI р 5а было снижено при обеих делециях Δ94-113 и Δ175-198. При этом делеционный мутант PhI р 5а ΔDM-94-113 (His) продемонстрировал максимальный ингибиторный эффект только на уровне приблизительно 50%, а делеционный мутант PhI р 5а DM-175-198 (His) показал ингибиторный эффект только на уровне 20-30%.
Двойной делеционный мутант PhI р 5а DM-Δ94-113,175-198 (His) являлся способным ингибировать связывание с IgE антителами только на 0-10% при самой высокой из используемых концентраций. Применение неслитого белка PhI р 5а DM-Δ94-113, 175-198 подтвердило этот результат (0-10% максимального ингибирования IgE).
Вторая группа сывороток, полученных от пациентов, страдающих аллергией, представленная индивидуальной сывороткой Р44 (Фиг.8 b), отличалась от первой группы тем фактом, что IgE антитела, представленные в сыворотке, в равной степени хорошо реагировали с PhI р 5а DM-Δ94-113 (His), а также со стандартом rPhI р 5а дикого типа (His) (70-80% максимального ингибирования), несмотря на то, что не было обнаружено IgE антител или не было выявлено их способного к обнаружению количества, прореагировавшего с PhI р 5а DM-Δ175-198 (His) (0-10% максимального ингибирования). Двойной делеционный мутант PhI р 5а DM-Δ94-113, 175-198 также продемонстрировал в значительной мере сниженный ингибиторный эффект (0-10%) с этой группой сыворотки, полученной от больных, страдающих аллергией, что было продемонстрировано как для слитого белка, так и для белка, в котором отсутствовал компонент слияния. Сыворотка этих больных, страдающих аллергией, предположительно содержала IgE антитела, направленные непосредственно против эпитопов С-терминальной части молекулы.
Данные, полученные при измерении IgE связывающей реактивности IgE антитела сывороточного пула 20 пациентов, страдающих аллергией, подчеркнули важность делеций Δ94-113 и Δ175-198 для снижения IgE связывания PhI р 5а (Фиг.8 с). Оба индивидуальных делеционных мутанта, PhI р 5а DM-Δ94-113 (His) и PhI р 5а DM-Δ175-198 (His) продемонстрировали более низкий максимальный ингибиторный эффект, 40-50% и приблизительно 30% соответственно, чем rPhI р 5а дикого типа (приблизительно 70%). Двойной делеционный мутант PhI р 5а DM-Δ94-113, 175-198 очень слабо связывался с IgE антителами сывороточного пула (10-15% максимального ингибирования), что пребывает в соответствии с анализом 43 больных страдающих аллергией, в тест-методе, что свидетельствует о значительно сниженной IgE связывающей реактивности этого варианта PhI р 5а у очень многих, если не у всех пациентов, страдающих аллергией на пыльцу трав.
Определение гипоаллергенности делеционных мутантов с помощью анализа активации базофилов
С помощью анализа активации базофилов оценивали влияние сниженной способности к связыванию IgE делеционных мутантов на функциональный эффект при перекрестном связывании IgE, связанного с мембраной эффекторных клеток, и на его активацию. Функциональное снижение аллергенности, таким образом, измеряли в чувствительном анализе in vitro.
Для анализа активации базофилов гепаринизированную цельную кровь, полученную от пациентов, страдающих аллергией на пыльцу, инкубировали с различными концентрациями исследуемых веществ. Аллергенные вещества способны связываться со специфическими IgE антителами, при этом они ассоциируются с высоко аффинными рецепторами IgE базофильных гранулоцитов.
Перекрестное связывание комплексов IgE/рецептор, инициированное аллергенными молекулами, приводит к сигнальной трансдукции, что, в свою очередь, приводит к дегрануляции эффекторных клеток и, таким образом, к инициации аллергических реакций in vivo.
В условиях in vitro индуцированная аллергеном активация базофильных иммуноцитов может быть определена с помощью количественной оценки экспрессии поверхностного белка (CD203c), спаренной с сигнальной трансдукцией перекрестного связывания рецептора IgE (Kahlert и др., ClinicaI Immunology and Allergy In Medicine Proceedings of the EAACI 2002 (2003) NapIes, Italy 739-744). Количество экспрессированных поверхностных белков на клетке и процент активированных клеток клеточного пула измеряется с высокой степенью чувствительности посредством связывания флуоресцентно меченного моноклонального антитела с поверхностью белка и последующего анализа при использовании флуоресцентно активированной проточной цитометрии.
Стандартные вещества, используемые при этом, представляют собой как очищенный природный PhI p 5а (nPhI р 5а), так и rPhI р 5а дикого типа, которые исследуются параллельно с исследуемыми веществами. Результаты анализа с двойным делеционным мутантом PhI р 5а DM Δ94-113, 175-198 при использовании базофилов, полученных от шести исследуемых особей, показаны в виде кривых на Фигуре 9. Результаты анализа с базофилами, полученными в целом от 10 пациентов, которые клинически определены как страдающие аллергией, показаны в Таблице 2. Значения А50 (А50: концентрация аллергена при 50% количества базофилов, активированных до максимума) стандартных молекул индивидуально варьировали от 1,3 до 15 пМ для rPhI р 5а дикого типа и от 0,3 до 10 пМ для nPhI р 5а (Таблица 2). В противовес этому значения А50 делеционного мутанта PhI р 5а DM Δ94-113, 175-198 составляли -18-8400 пМ. Значения А50, определенные для трех используемых веществ, применяли для определения алергенной эффективности делеционного варианта PhI р 5а DM Д94-113, 175-198 относительно неизмененных стандартных молекул nPhI р 5а и rPhI р 5а дикого типа для каждого индивидуума, подвергшегося анализу (Таблица 2). Относительная аллергенная эффективность (Pr, относительная эффективность) делеционного варианта PhI р 5а DM А94-113, 175-198 была пониженной и составляла от 12 до 5000 раз по сравнению с rPhI р 5а дикого типа или -16-32000 раз по сравнению со стандартом nPhI р 5а (Таблица 2).

Т-клеточная реактивность
Т-хелперные лимфоциты реагируют с пептидными фрагментами аллергенов (приблизительно 12-25 аминокислот), полученными путем ферментативного переваривания в антиген-презентирующих клетках (АРС), они презентируются для Т-клеток после включения приемлемых пептидов в индивидуальный класс молекул II МНС на поверхности АРС. Такая специфическая для аллергена активация Т-хелперных лимфоцитов представляет собой предпосылку для последующих реакций (пролиферации, анергии, апоптоза) и для функциональной дифференциации (ТН1 и ТН2). Влияние специфических для аллергена Т-лимфоцитов путем обработки аллергеном или вариантом аллергена при гипосенсибилизации рассматривается как ключевой момент терапевтической эффективности.
Для того чтобы оценить Т-клеточную реактивность, олигоклональные линии Т-клеток (TCL), полученные от больных, страдающих аллергией на пыльцу Graminae, применяются при использовании традиционных способов со стимуляцией с помощью молекул PhI р5 или rPhI р 5. В анализе пролиферации различные линии Т-клеток стимулируются с помощью стандартных аллергенов nPhI р5а и rPhI р5а дикого типа и двойного делеционного мутанта PhI р 5а DM Δ94-113, 175-198. Степень пролиферации определяют путем встраивания [3Н] тимидина при использовании традиционных способов.

Результаты с десятью TCL от шести пациентов, страдающих аллергией, показали, что эти TCL были стимулированы к пролиферации с помощью PhI р 5а DM Δ94-113, 175-198 по сравнению с неизмененным природным или рекомбинантным аллергеном дикого типа (Таблица 3).
Настоящее изобретение, таким образом, относится к вариантам группы 5 аллергенов Pooideae, которые характеризуются сниженной IgE реактивностью по сравнению с известными аллергенами дикого типа и сохраненной реактивностью по отношению к Т-лимфоцитам. Эти аллергены группы 5 предпочтительно представляют собой PhI р 5а, Poa р 5 b LoI р 5, особое предпочтение отдается PhI р 5а.
Как было доказано, наиболее благоприятным в отношении участков аминокислотной последовательности для целей данного изобретения является отсутствие или удаление участков аминокислотной последовательности, которые соответствуют положениям 94-113 и 175-198 PhI р 5а в группе 5 аллергенов. Данное изобретение, в частности, относится к таким вариантам аллергенов. Первый упомянутый или второй упомянутый участок может отсутствовать индивидуально, однако оба упомянутых участка могут отсутствовать одновременно, при этом последнее воплощение является особенно предпочтительным. Благодаря высокой гомологии последовательностей в пределах группы 5 аллергенов Pooideae эти участки могут быть однозначно идентифицированы при сличении последовательностей PhI р 5а с другими последовательностями группы 5 аллергенов. Описанные выше варианты аллергенов имеют происхождение от PhI р 5а или соответствуют последовательностям в соответствии с SEQ ID NO 4, 6 или 8.
Варианты аллергенов в соответствии с изобретением могут быть получены при использовании в качестве исходного материала клонированной последовательности ДНК с помощью методов генетической инженерии. В принципе, однако, химические модификации природного экстракта аллергена также являются возможными (FiebIg, 1995, Allergo J. 4 (7), 377-382).
Естественно дальнейшие модификации в других положениях, например, для того, чтобы увеличить гипоаллергенность, являются также возможными посредством вариаций группы 5 аллергенов, описанных в данной патентной заявке. Такие модификации могут быть, например, инсерциями аминокислот, делециями и заменами, расщеплением белка на фрагменты и слиянием белка или его фрагментов с другими белками или пептидами.
Во время получения вариантов аллергена, описанных более подробно в данной заявке, His метка была введена с помощью методов генетической инженерии с целью улучшения очистки сверхэкспрессированных белков.
Изобретение, кроме того, относится к молекуле ДНК, кодирующей вариант аллергена, описанный выше, в частности, соответствующей последовательности в соответствии с SEQ ID NO 3, 5 или 7, к рекомбинантному экспрессионному вектору, содержащему эту молекулу ДНК, и к хозяйскому организму, трансформированному с помощью указанной молекулы ДНК или указанного экспрессионного вектора. Приемлемый хозяйский организм может представлять собой прокариотический или эукариотический, одно- или многоклеточный организм, такой как бактерии или дрожжи. Хозяйский организм, который является предпочтительным в соответствии с изобретением, представляет собой Е.coli.
Кроме того, изобретение относится к способу получения варианта аллергена в соответствии с изобретением путем культивирования указанного хозяйского организма и изоляции соответствующего варианта аллергена из культуры.
Настоящее изобретение дополнительно относится к вариантам аллергена, молекулам ДНК и экспрессионным векторам, описанным выше в качестве лекарственных средств.
Настоящее изобретение, кроме того, относится к фармацевтическим композициям, содержащим, по крайней мере, один из этих вариантов аллергена, или соответствующую молекулу ДНК, или соответствующий экспрессионный вектор и необязательно также активные ингредиенты и/или вспомогательные вещества, для лечения аллергий, в запуск которых вовлечена группа 5 аллергенов Pooideae, или для иммунотерапевтической вакцинации пациентов с аллергиями, в запуск которых вовлечена группа 5 аллергенов Pooideae, и/или для предотвращения таких аллергий.
Если таковые представляют собой фармацевтические композиции второго типа (содержащие, по крайней мере, одну молекулу ДНК или экспрессионный вектор), то эти композиции предпочтительно дополнительно включают гидроокись алюминия, иммуностимуляторный олигонуклеотид, содержащий CpG, или комбинацию обоих в качестве адъювантов.
Для целей настоящего изобретения фармацевтические композиции могут использоваться как терапевтические агенты для лечения человека и в ветеринарной медицине. Приемлемые наполнители представляют собой органические или неорганические вещества, которые являются приемлемыми для парентерального введения и которые не реагируют с группой 5 вариантов аллергена в соответствии с изобретением. Приемлемыми для парентерального введения являются, в частности, растворы, предпочтительно масляные или водные растворы, а также суспензии, эмульсии или имплантаты. Варианты аллергена в соответствии с изобретением могут также быть лиофилизированы и полученные лиофилизаты используют, например, для получения препаратов для инъекций. Назначаемые композиции могут также быть лиофилизированы и полученные лиофилизаты используют, например, для получения инъекционных препаратов. Назначаемые композиции могут быть стерильными и/или содержать вспомогательные вещества, такие как лубриканты, консерванты, стабилизаторы и/или смачивающие агенты, эмульгаторы, соли для модификации осмотического давления, буферные вещества и/или множество дополнительных активных ингредиентов.
Кроме того, приемлемая композиция вариантов аллергена в соответствии с изобретением позволяет получать "депо"-препараты, например, с помощью адсорбции на гидроокиси алюминия.
В завершение, настоящее изобретение относится к применению, по крайней мере, одного варианта аллергена в соответствии с изобретением, или молекулы ДНК в соответствии с изобретением, или экспрессионного вектора в соответствии с изобретением для получения лекарственного средства для лечения аллергий, в запуск которых вовлечена группа 5 аллергенов Pooideae, или для иммунотерапевтической вакцинации пациентов с аллергиями, в запуск которых вовлечена группа 5 аллергенов Pooideae, и/или для предотвращения таких аллергий.










| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТ АЛЛЕРГЕНА ГРУППЫ I ИЗ Роасеае, ХАРАКТЕРИЗУЮЩИЙСЯ СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ И СОХРАНЕННОЙ Т-КЛЕТОЧНОЙ РЕАКТИВНОСТЬЮ (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА ДНК И ЕГО ПРИМЕНЕНИЕ | 2005 |
|
RU2409589C2 |
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2575606C2 |
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2658767C1 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ ОСНОВНЫХ АЛЛЕРГЕНОВ ГРУППЫ 4 ИЗ ЗЛАКОВЫХ | 2004 |
|
RU2373283C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И ПОЛУЧЕНИЕ АЛЛЕРГЕНА ПЫЛЬЦЫ ТРАВ Phl p 4 С ПОМОЩЬЮ РЕКОМБИНАНТНЫХ СПОСОБОВ | 2003 |
|
RU2327739C2 |
| ГИПОАЛЛЕРГИЧЕСКИЕ ВАРИАНТЫ Mal d 1, ГЛАВНОГО АЛЛЕРГЕНА Malus domectica | 2012 |
|
RU2624030C2 |
| МИКРОЧАСТИЦЫ, ВКЛЮЧАЮЩИЕ УГЛЕВОДНЫЕ СФЕРЫ, КОВАЛЕНТНО СВЯЗАННЫЕ С АЛЛЕРГЕНОМ | 2003 |
|
RU2329830C2 |
| НОВЫЕ АЛЛЕРГЕНЫ ПШЕНИЦЫ | 2008 |
|
RU2502742C2 |
| МОДИФИКАЦИИ АЛЛЕРГЕНОВ ГРУППЫ 6 POACEAE (МЯТЛИКОВЫХ), ИМЕЮЩИХ ПОНИЖЕННУЮ АЛЛЕРГЕННОСТЬ БЛАГОДАРЯ МУТАГЕНЕЗУ ПРОЛИНОВЫХ ОСТАТКОВ | 2010 |
|
RU2607373C2 |
| ВАРИАНТЫ ОСНОВНОГО АЛЛЕРГЕНА Phl p 1 ИЗ ТИМОФЕЕВКИ ЛУГОВОЙ | 2003 |
|
RU2323942C2 |





Изобретение раскрывает получение и применение вариантов аллергенов группы 5 Pooideae, которые характеризуются сниженной IgE реактивностью по сравнению с известными аллергенами дикого типа и в то же время существенно сохраненной реактивностью по отношению к Т-лимфоцитам. У вариантов аллергенов по изобретению отсутствуют по меньшей мере один участок или комбинации участков, соответствующие аминокислотной последовательности участков 94-113 или 175-198 PhI р 5а, представленной в описании. Варианты аллергенов получены генно-инженерными способами. В изобретении раскрыты молекула ДНК, кодирующая вариант аллергена, рекомбинантный вектор экспрессии, организм-хозяин, экспрессирующий вариант аллергена и способ получения варианта аллергена с использованием перечисленных средств. Такие гипоаллергенные варианты аллергена могут быть использованы в качестве лекарственного средства против аллергий, обусловленных аллергенами группы 5 Pooideae для специфической иммунотерапии (гипосенсибилизации) пациентов, имеющих аллергию на пыльцу трав, или для превентивной иммунотерапии аллергий на пыльцу трав с использованием фармацевтической композиции. Препараты на основе вариантов аллергенов по изобретению обладают сниженной IgE реактивностью, сохраненной реактивностью по отношению к Т-лимфоцитам. 8 н. и 4 з.п. ф-лы, 9 ил., 3 табл.
1. Вариант группы 5 аллергенов PhI р 5а, Роа р 5 b и LoI р 5, характеризующийся сниженной IgE реактивностью по сравнению с известными аллергенами дикого типа и существенно сохраненной реактивностью по отношению к Т-лимфоцитам, и у которого по меньшей мере один участок или комбинации участков, которые соответствуют аминокислотной последовательности участков 94-113 или 175 -198 PhI р 5а, отсутствуют по сравнению с известными аллергенами дикого типа.
2. Вариант аллергена по п.1, имеющий происхождение от PhI р 5а.
3. Вариант аллергена по п.1, характеризующийся тем, что он получен с помощью способов рекомбинантной генетической инженерии.
4. Вариант аллергена по п.2, характеризующийся тем, что он получен с помощью способов рекомбинантной генетической инженерии.
5. Вариант аллергена PhI р 5а, характеризующийся сниженной IgE реактивностью по сравнению с известными аллергенами дикого типа и существенно сохраненной реактивностью по отношению к Т-лимфоцитам, и который имеет аминокислотную последовательность, выбранную из последовательностей в соответствии с SEQ ID NO 4, 6 и 8.
6. Вариант аллергена по п.5, характеризующийся тем, что он получен с помощью способов рекомбинантной генетической инженерии.
7. Молекула ДНК, кодирующая вариант аллергена по п.2 и имеющая последовательность в соответствии SEQ ID NO 3, 5 и 7.
8. Рекомбинантный вектор экспрессии, содержащий молекулу ДНК по п.7, функционально связанную с последовательностью, которая контролирует экспрессию.
9. Организм-хозяин, экспрессирующий вариант аллергена по п.2, трансформированный вектором экспрессии по п.8.
10. Способ получения варианта аллергена по п.6, путем культивирования организма-хозяина по п.9 и выделения соответствующего варианта аллергена из культуры.
11. Лекарственное средство против аллергий, обусловленных аллергенами группы 5 Pooideae, представляющими собой вариант аллергена по п.6.
12. Фармацевтическая композиция, включающая по меньшей мере один вариант аллергена по п.6 и необязательно дополнительные активные ингредиенты и/или вспомогательные вещества, для лечения аллергий, обусловленных аллергенами группы 5 Pooideae.
| DE 19713001 А, 01.10.1998 | |||
| WO 03025009 А, 27.03.2003 | |||
| ФРАДКИН В.А | |||
| Аллергены, Изд | |||
| Медицина, М., 1978, с.125-139. |

