Область, к которой относится изобретение
Настоящее изобретение относится к препарату и к применению рекомбинантных вариантов группы 5 аллергенов Роасеае (злаковые), которые отличаются сниженной IgE реактивностью по сравнению с известными аллергенами дикого типа и в то же время по существу сохраненной реактивностью с Т-лимфоцитами.
Сниженная IgE реактивность главным образом достигается посредством замещения или делеции некоторых остатков пролина, которые прочно сохраняются в группе 5 аллергенов. К тому же, аминокислота пролин в положении 211 (Phl р 5.0109 нумерация) признавалась в качестве ключевой позиции для целенаправленного влияния на свойства растворимости вариантов в водных композициях.
Такие гипоаллергенные варианты аллергенов могут использоваться для специфической иммунотерапии (гипосенсибилизация) пациентов с аллергией на пыльцу трав или для профилактического лечения для предотвращения проявления аллергий на пыльцу трав.
Предпочтительная форма осуществления изобретения относится к вариантам основного аллергена Phl р 5а из пыльцы тимофеевки луговой (Phleum pratense).
Предпосылки к созданию изобретения
Аллергии типа 1 имеют значение мирового масштаба. Вплоть до 20% населения в промышленно развитых странах страдают от осложнений, таких как аллергический ринит, конъюнктивит или бронхиальная астма. Эти аллергии вызываются источниками различного происхождения, такими как деревья и трава (пыльца), грибы (споры), клещи (экскременты), коты или собаки. Источники аллергии высвобождаются непосредственно в воздух (пыльца, споры) или могут попадать в воздух связанными с частицами дизельной сажи (пыльца) или домашней пыли (экскременты клещей, частички кожи, волосы). Так как вызывающие аллергию вещества находятся в воздухе, то также используют понятие аэроаллергенов.
Вещества, которые вызывают аллергию типа 1, представляют собой белки, гликопротеины или полипептиды. После поглощения через слизистые оболочки эти аллергены вступают в реакцию с молекулами IgE, связанными с поверхностью тучных клеток у чувствительных индивидуумов. Если эти молекулы IgE сшиваются друг с другом с помощью аллергена, то это приводит к высвобождению медиаторов (например, гистамина, простагландинов) и цитокинов эффекторными клетками и, таким образом, к соответствующим клиническим симптомам.
Вплоть до 40% людей, страдающих аллергией типа 1, проявляют специфическую IgE реактивность с экстрактами пыльцы злаковых (Burney et al., 1997, J. Allergy Clin. Immunol. 99:314-322; D'Amato et al., 1998, Allergy 53: 567-578; Freidhoff et al., 1986, J. Allergy Clin Immunology, 78, 1190-2002). Семейство злаковых (Poaceae) охватывает более чем 10000 видов, более чем 20 из которых до сих пор известны как пусковые механизмы аллергических симптомов (Andersson & Lidholm, 2003, Int. Arch. Allergy Immunol. 130:87-107; Esch, 2008, Allergens и Allergen Immunotherapy, Clinical Allergy и Immunology Series, 107-126).
Большая часть злаковых, вызывающих аллергию, принадлежит к подсемейству Pooideae. Наряду с встречающимися дикими формами видов трав, такими как, например, Holcus lanatus (бухарник шерстистый), Phalaris aquatica (канареечник Канарский), Anthoxanthum odoratum (душистый колосок), Dactylis glomerata (ежа сборная), Festuca pratensis (овсяница луговая), Роа pratensis (мятлик луговой) или Lolium регеппе (плевел многолетний), культурные зерновые, такие как Triticum aestivum (пшеница), Secale cereale (рожь) и Hordeum vulgare (ячмень), также являются известными членами этого подсемейства.
Одним из видов Pooideae, который был лучше всего исследован относительно его аллергенов, является тимофеевка луговая (Phleum pratense), которая представляет собой широко распространенное во всем мире дикорастущее растение, а также имеет экономическое значение в качестве пастбищного растения и зимостойкой кормовой травы.
В зависимости от относительной частоты у населения, с которой молекулы индивидуального аллергена реагируют с IgE антителами страдающих аллергией пациентов, делается различие между основными и второстепенными аллергенами.
Шесть аллергенов тимофеевки луговой можно рассматривать как основные аллергены: Phl р 1 (Petersen et al., 1993, J. Allergy Clin. Immunol. 92: 789-796), Phl p 5 (Matthiesen и Lowenstein, 1991, Clin. Exp. Allergy 21: 297-307; Petersen et al., 1992, Int. Arch. Allergy Immunol. 98: 105-109), Phl p 6 (Petersen et al., 1995, Int. Arch. Allergy Immunol. 108, 49-54), Phl p 2/3 (Dolecek et al., 1993, FEBS 335 (3): 299-304), Phl p 4 (Haavik et al., 1985, Int. Arch. Allergy Appl. Immunol. 78: 260-268; Valenta et al., 1992, Int. Arch. Allergy Immunol. 97: 287-294; Nandy et al., Biochem. Biophys. Res. Commun., 2005, 337(2): 563-70) и Phl p 13 (Suck et al., 2000, Clin. Exp.Allergy 30: 1395-1402).
Доминантные основные аллергены тимофеевки луговой представляет собой Phl р 1 и Phl р 5 (Andersson & Lidholm, 2003, Int. Arch. Allergy Immunol. 130:87-107), причем Phl p 5 встречаются в двух формах 5а и 5b, которые кодируются независимыми генами и отличаются в отношении своего молекулярного веса. Следуя официальной номенклатуре аллергенов, Phl р 5а обозначается как Phl р 5.01 и Phl р 5b обозначается как Phl р 5.02 (WHO/IUIS Allergen Nomenclature Subcommittee, www.allergen.org). Аминокислотные последовательности как Phl р 5а, так и Phl р 5b были выведены из клонированных «ДНК последовательностей. Были установлены природные варианты обеих изоформ, которые отличаются одна от другой точечными мутациями и соответствуют различным аллельным формам (Vrtala et al., 1993, J. Immunol., 151: 4773-4781; Gelharetal., 1997, Eur. J. Biochem., 247: 217-23). Эти варианты записаны в базе данных WHO/IUIS как Phl р 5.01хх и Phl р 5.02хх.
Природный Phl р 5а (nPhl р 5а) представляет собой белок прибл. 32 кДа и реагирует с антителами IgE у 85-90% страдающих от аллергии на пыльцу трав (Rossi et al., 2000, Allergy Int., 49: 93-97).
Пыльца родственных видов злаковых из семейства Роасеае и в частности подсемейства Pooideae, таких как Lolium perenne или Роа pratensis, содержит аллергены, которые являются гомологичными с Phl р 5 и вместе они известны как группа 5 аллергенов. Высокая структурная гомология этой группы 5 аллергенов соответственно служит причиной высокой перекрестной реактивности молекул с IgE антителами (Lorenz et al., 2009, Int. Arch. Immunol. 148:1-17). В конечном счете, по причине этой перекрестной реактивности может быть достаточно сенсибилизации одним видом травы, чтобы вызвать аллергическую реакцию другими родственными травами.
В конце концов, высокая перекрестная реактивность группы 5 аллергенов основывается на подобной первичной последовательности гомологичны) аллергенов. Это показывает сравнение аминокислотной последовательности группы 5 аллергенов выбранных видов Pooideae (фиг. 1).
Наряду с перекрестной реактивностью группы 5 аллергенов друг с другом, также известна перекрестная реактивность Phl р 5 с другим основным аллергеном тимофеевки луговой ( 1978, Allergy 33: 30-41; Petersen et al., 1995, Int. Arch. Allergy Immunol. 108: 55-59; Blume et al., 2004, Proteomics 4:1366-71). Полипептидная цепь аллергена Phl p 6 проявляет большое сходство с N-концевой половиной различных Phl р 5 последовательностей (фиг. 1). Предполагается, что аллергены могут восходить к общему предковому гену. Сходство между аллергенами двух групп является причиной того, что некоторые из Phl р 5-реактивных IgE антител также связываются с Phl р 6 (Petersen et al., 1995, Int. Arch. Allergy Immunol. 108:49-54; Andersson & Lidholm, 2003, Int. Arch. Allergy Immunol. 130:87-107).
1978, Allergy 33: 30-41; Petersen et al., 1995, Int. Arch. Allergy Immunol. 108: 55-59; Blume et al., 2004, Proteomics 4:1366-71). Полипептидная цепь аллергена Phl p 6 проявляет большое сходство с N-концевой половиной различных Phl р 5 последовательностей (фиг. 1). Предполагается, что аллергены могут восходить к общему предковому гену. Сходство между аллергенами двух групп является причиной того, что некоторые из Phl р 5-реактивных IgE антител также связываются с Phl р 6 (Petersen et al., 1995, Int. Arch. Allergy Immunol. 108:49-54; Andersson & Lidholm, 2003, Int. Arch. Allergy Immunol. 130:87-107).
В прошлом 3D структура многих аллергенов объяснялась ЯМР спектроскопией или рентгеновским структурным анализом и среди прочего служила основой для локализации IgE-связывающих антигенных детерминантов на поверхности белка. В случае группы 5 аллергенов пыльцы трав, до настоящего времени было невозможно создать модель, которая охватывает целую полипептидную цепь (Rajashankar et al., 2002, Acta Cryst. D58:1175-1181; Maglio et al., 2002, Protein Engineering 15: 635-642).
На основе 3D структур аллергенов Phl p 6 (RCSB банк данных белков запись: 1NLX) и Phl р 5b полумолекулы (RCSB банк данных белков запись: 1L3P), можно было создать гомологичную модель Phl р 5а (Wald et al., 2007, Clin. Exp.Allergy 37:441-450). Согласно этой модели Phl p 5a создается из двух спиральных пучков, но точное положение двух пучков друг к другу не может быть объяснено гомологичной моделью (фиг. 2).
Специфическая иммунотерапия (СИТ) или гипосенсибилизация рассматривается как эффективный подход к терапевтическому лечению аллергий (Fiebig 1995 Allergo J. 4 (6):336-339, Bousquetet al., 1998, J. Allergy Clin. Immunol. 102 (4): 558-562); Cox et al., 2007, J. Allergy Clin. Immunol. 120:S25-85; James & Durham, 2008, Clin. Exp.Allergy 38: 1074-1088).
Классическая терапевтическая форма инъекционного лечения (SCIT), при которой пациенту впрыскивают подкожно природные экстракты аллергенов в увеличивающихся дозах, с успехом применялась в течение приблизительно 100 лет. При этой форме, иммунная система страдающего от аллергии человека неоднократно сталкивается с аллергенами, вследствие чего достигается перепрограммирование иммунной системы вместе с устойчивостью к аллергенам. После поглощения антигенов из аллергенных препаратов антиген-представляющими клетками, пептиды презентируются антигенам на поверхности клеток. Некоторые определенные пептиды, содержащие так называемые Т-клеточные эпитопы, распознаются антиген-специфическими Т-клетками. Среди прочего это связывание приводит к проявлению различных типов Т-клеток, обладающих регулятивной функцией. В ходе СИТ ответ регулирующей Т-клетки приводит к устойчивости в отношении аллергенов, понижающей регуляции Тн2 цитокинов, восстановлению равновесия TH1/TH2, подавлению аллерген-специфических IgE, индукции lgG4, lgG1 и IgA антител, подавлению эффекторных клеток (тучных клеток, базофилов и эозинофилов), а также к обновлению воспаленных тканей (Akdis et al., 2007, J. Allergy Clin. Immunol. 119 (4):780-789; Larche et al., 2008, Nature Reviews 6:761-771). Таким образом, Т-клеточные эпитопы имеют ключевое значение для терапевтического действия аллергенных препаратов в случае гипосенсибилизации.
Вследствие перекрестной реактивности основных аллергенов злаковых, которая присутствует на уровне IgE, а также Т-клеточном уровне, чаще всего достаточно успешной терапии экстрактами аллергенов единственного представительного вида трав (Mailing et al., 1993, EAACl Position Paper: Immunotherapy, Allergy 48: 9-35; Cox et al., 2007, J Allergy Clin Immunol 120: 25-85).
Наряду с подкожной иммунотерапией, подъязычная терапевтическая форма, при которой аллергены или производные аллергенов поглощаются через ротовую слизистую оболочку, подвергается клиническим исследованиям и используется в качестве альтернативы инъекционной терапии (James & Durham, 2008, Clin. Exp.Allergy 38: 1074-1088).
Другую возможность представляет лечение с помощью способной к экспрессии ДНК, которая кодирует релевантные аллергены (иммунотерапевтическая вакцинация). Экспериментальное подтверждение специфического для аллергена влияния на иммунный ответ было продемонстрировано у грызунов с помощью инъекции ДНК, кодирующей аллерген. (Hsu et al. 1996, Nature Medicine 2 (5):540-544, Weiss et al., 2006, Int. Arch. Allergy Immunol. 139: 332-345).
Во всех этих терапевтических формах, существует значительный риск аллергических реакций или даже анафилактического шока (Kleine-Tebbe, 2006, Allergologie, 4:135-156). Для того чтобы минимизировать этот риск, применяют передовые препараты в форме аллергоидов. Они представляют собой химически модифицированные экстракты аллергенов, которые обладают существенно сниженной IgE реактивностью, но идентичной Т-клеточной реактивностью по сравнению с необработанным экстрактом (Fiebig 1995 Allergo J. 4 (6):336-339, Kahlert et al., 1999, Int. Arch. Allergy Immunol, 120: 146-157).
Оптимизация терапии возможна с аллергенами, полученными с помощью рекомбинантных способов. Определенные смеси аллергенов с высокой степенью чистоты, полученные с помощью рекомбинантных способов и необязательно подобранные к индивидуальным моделям сенсибилизации пациентов, могут заменить экстракты из источников природных аллергенов, поскольку кроме различных аллергенов они содержат относительно высокое количество иммуногенных, но неаллергенных сопутствующих белков. Первые клинические исследования с рекомбинантными аллергенами уже были осуществлены с успехом (Jutel et al., 2005, J. Allergy Clin. Immunol., 116: 608-613; Valenta & Niederberger, 2007, J. Allergy Clin. Immunol. 119: 826-830).
Реальные перспективы, которые могут приводить к безопасной гипосенсибилизации рекомбинантными продуктами экспрессии, в особенности предлагаются при использовании мутированных рекомбинантных аллергенов, в которых эпитопы IgE изменены без повреждения Т-клеточных эпитопов, которые являются существенными для терапии (Schramm et al. 1999, J. Immunol. 162: 2406-2414). Эти гипоаллергенные белки могли быть использованы в относительно высоких дозах во время СИТ, при этом не увеличивая вероятность нежелательных IgE-способствующих побочных эффектов.
В прошлом, были опубликованы такие "гипоаллергенные" варианты со сниженным IgE связыванием для многих аэроаллергенов (в частности аллергены пыльцы и аллергены клещей домашней пыли) и пищевых аллергенов. На основе ДНК немодифицированных аллергенов, можно было получить и экспрессировать рекомбинантную ДНК, среди прочего посредством фрагментации, олигомеризации, делеций, точечных мутаций или рекомбинации индивидуальных участков аллергена (перестановка ДНК) (Ferreira et al., 2006, Inflamm. & Allergy - Drug Targets 5: 5-14; Bhalla & Singh, 2008, Trends in Biotechnology 26: 153-161).
Что касается аллергенов пыльцы трав, то были описаны гипоаллергенные варианты групп 1, 2, 5а, 5b, 6, 7 и 12 (Ferreira et al., 2006, Inflamm. & Allergy - Drug Targets 5: 5-14; Westritschnig et al., 2007, J. Immunol. 179: 7624-7634).
До настоящего времени во множестве публикаций описаны подходы к развитию гипоаллергенной группы 5 аллергенов. Для Phl р 5а и Phl р 5b, можно было показать, что комбинированная делеция двух участков последовательности влечет за собой значительное уменьшение IgE связывания, а также уменьшенную способность к стимулированию базофильных эффекторных клеток. Тем не менее, Т-клеточная реактивность мутантов с делецией была незначительно модифицирована по сравнению с таковой немодифицированного аллергена (Schramm et al., 1999, J. Immunol. 162: 2406-2414; Wald et al., 2007, Clin. Exp. Allergy 37: 441-450).
В другом документе группа 5 аллергенов из Lolium perenne (Lol р 5) была изменена аминокислотными замещениями и/или короткими делециями у С-конца (Swoboda et al., 2002, Eur. J. Immunol. 32: 270-280). Мутации не ограничивались определенным типом аминокислоты. Замещенные аминокислоты представляли собой лизин, фенилаланин, треонин, валин или аланин. Равным образом был описан концептуальный подход создания гипоаллергенных мутантов посредством целенаправленной мутации некоторого количества остатков отдельной аминокислоты. Gelhar et al. описывали создание рекомбинантного Phl р 5b фрагмента посредством замещения аланином десяти остатков лизина, локализованных на поверхности белка (Gelhar et al., 2006, Int. Arch. Allergy Immunol. 140:285-294). Однако мутационная стратегия, основанная на точечных мутациях в остатках пролина, для группы 5 аллергенов пыльцы трав до сих пор не была опубликована.
Задача, лежащая в основе настоящего изобретения, заключается в предоставлении новых рекомбинантных вариантов группы 5 аллергенов Роасеае на белковом уровне и уровне ДНК, которые отличаются сниженной IgE реактивностью при одновременно существенном сохранении Т-клеточной реактивности, и поэтому они пригодны для лечебной и превентивной специфической иммунотерапии и иммунотерапевтической ДНК вакцинации.
Описание изобретения
Неожиданно было обнаружено, что варианты группы 5 аллергенов семейства злаковых (Роасеае), в которых пролины, которые соответствуют в одном выравнивании пролинам в положениях 57, 58, 117,146, 155, 211, 229 в аминокислотной последовательности дикого типа Phl р 5.0109 мутировали отдельно или в комбинации, обладают сниженной IgE реактивностью по сравнению с аллергенами дикого типа и в то же время обладают существенно сохраненной реактивностью с Т-лимфоцитами и соответственно являются гипоаллергенными.
Соответственно изобретение относится к гипоаллергенным вариантам группы 5 аллергенов семейства злаковых {Роасеае), в которых пролины которые соответствуют в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 211, 229 в аминокислотной последовательности дикого типа Phl р 5.0109 мутировали отдельно или в комбинации.
Особое предпочтение отдается вариантам аллергенов в соответствии с изобретением, отличающимся тем, что пролины делетированы или замещены.
В соответствии с изобретением предпочтение отдается гипоаллергенным вариантам группы 5 аллергенов из подсемейства Pooideae, предпочтительно из групп Poodae и Triticodae, предпочтительно представленных Phleum pratense, Holcus lanatus, Phalaris aquatica, Anthoxanthum odoratum, Dactylis glomerata, Lolium perenne, Poa pratensis, Festuca pratensis, Hordeum vulgare, Secale cereale и Triticum aestivum. Предпочтительно речь идет о гипоаллергенных вариантах в соответствии с изобретением Tri а 5, Sec с 5 и Hor v 5 из Triticum aestivum, Secale cereale и Hordeum vulgare. Особое предпочтение отдается гипоаллергенным вариантам в соответствии с изобретением группы 5 аллергенов Poodae. Эта группа 5 аллергенов предпочтительно включает Роа р 5, Dac g 5, Hol р 5, Lol р 5 и Pha а 5 из Роа pratensis, Dactylis glomerata, Holcus lanatus, Lolium perenne и Phalaris aquatica и в высшей степени предпочтительным является Phl р 5 из Phleum pratense. Согласно изобретению предпочтительными являются все природно встречающиеся изомеры, полиморфы и варианты указанных выше аллергенов, а также их белки-предшественники.
В гипоаллергенных вариантах в соответствии с изобретением, мутированные пролины предпочтительно являются теми, которые соответствуют в одном выравнивании (фиг. 1а) пролинам в положениях 57, 58, 117, 146, 155, 211 или 229 в аминокислотной последовательности зрелого Phl р 5.0109 (Phl р 5 a, UniProtKB запись: Q84UI2) фиг. 3 и 4, SEQ ID NO: 1, SEQ ID NO: 2) или их варианты (фиг. 1b) или зрелого Phl р 5.0201 (Phl р5 b; Swiss-Prot: Q40963.2; фиг. 24, SEQ ID NO: 5) или их варианты (фиг. 1b), особенно предпочтительно зрелого Phl р 5.0109.
Несмотря на то, что было известно, что пролины могут оказывать влияние на структуру белка, целенаправленные точечные мутации остатков пролина в качестве отправной точки для генерации гипоаллергенных мутантов аллергенов были исследованы только для группы 2 основного аллергена клеща домашней пыли Dermatophaogides farinae (Derf 2, замещение остатков пролина аланином) (Takai et al., 2000, Eur. J. Biochem. 267: 6650-6656). Тем не менее, IgE связывающая способность и способность стимулировать базофильные клетки была снижена всего лишь незначительно в случае трех точечных мутантов, в то время как три остальные проявляли себя как немодифицированный аллерген. Таким образом, мутации пролина в случае Der f 2 не проявляли вообще или проявляли только очень слабое уменьшение аллергенности. Другие стратегии получения гипоаллергенных мутантов посредством мутаций обмена пролина не были опубликованы. Таким образом, для специалиста в данной области техники не было очевидным, что мутации пролина были бы успешными в качестве отправной точки для генерации гипоаллергенных мутантов аллергенов.
Кроме того, до настоящего времени ни для одного аллергена не было исследовано то, каким образом специфическая делеция остатков пролина воздействует на общую IgE связывающую способность продукта экспрессии и какие влияния осуществляются на активацию аллерго-релевантных эффекторных клеток.
Аминокислотная последовательность Phl р 5.0109 (UniProtKB: Q84UI2) содержит 16 остатков пролина (фиг. 1). Шесть остатков пролина 97, Р10, Р13, Р19, Р22 и Р27 локализуются в N-концевой области, которая сохраняется незначительно в пределах группы 5 аллергенов. Пролины в аминокислотных положениях 57, 58, 85, 117, 146, 155, 211, 229 и 256 располагаются в начале или в конце ос-спиралей или в петлях, которые связывают α-спирали и прочно сохраняются за исключением аминокислоты Р85 (фиг. 1, 2).
Исходя из аминокислотной последовательности Phl р 5а изоформ Phl р 5.0109, получают рекомбинантный немодифицированный аллерген дикого типа (rPhl р 5а wt; фиг. 4, SEQ ID N0:2), а также варианты, модифицированные при помощи генной инженерии. Аналогично представленным в дальнейшем способам получения, также могут быть получены белки дикого типа и предлагаемые в изобретении гипоаллергенные варианты другой группы 5 аллергенов злаковых согласно изобретению, например, Lol р 5; Роа р 5, Pha а 5, Dac g 5; Hol I 5, Tri a 5 и Hor v 5. С этой целью пролины, которые в одном выравнивании соответствуют пролинам в положениях 57, 58, 117, 146, 155, 211, 229 в аминокислотной последовательности дикого типа Phl р 5, мутировали отдельно или в комбинациях, предпочтительно путем замещения или делеции.
Вдобавок к описанным вариациям группы 5 аллергенов, естественно также возможны другие модификации в других положениях - например, для того, чтобы повысить гипоаллергенность. Эти модификации могут представлять собой, например, аминокислотные внедрения, делеции, замены и расщепления белка на фрагменты, а также слияния белка или его фрагментов с другими белками или пептидами, и мультимеры путем слияний идентичных белков или фрагментов.
Фрагменты в соответствии с изобретением предпочтительно охватывают 20-109 аминокислоты, предпочтительно 30-100 аминокислоты, особенно предпочтительно 40-90 аминокислоты. Кроме того, варианты в соответствии с изобретением включают белки-предшественники, такие как, например, ProPhl р 5, с предшествующей природной или искусственной сигнальной последовательностью. К тому же, в соответствии с изобретением включены белки слияния, имеющие N- или С-концевые метки слияния (например, His tag, как на фиг. 5 и 6, MBP tag, последовательности, контролирующие экспрессию и т.п.), гибридные молекулы, такие как, например, слияния с другими аллергенами или их гипоаллергенные варианты или слияния фрагментов в любой последовательности. Кроме того, варианты в соответствии с изобретением также включают гомологичные последовательности (полиморфы (SNPs), изоформы), обладающие идентичностью аминокислотной последовательности по меньшей мере в 80% с соответствующей группой 5 аллергена дикого типа, предпочтительно, по меньшей мере, в 90% с соответствующей группой 5 аллергена дикого типа, особенно предпочтительно по меньшей мере в 95% с соответствующей группой 5 аллергена дикого типа. В этих вариантах одна или несколько аминокислот предпочтительно замещены консервативно, например, одна полярная аминокислота замещена другой полярной аминокислотой или одна нейтральная аминокислота замещена другой нейтральной аминокислотой, однако согласно изобретению также существуют варианты, обусловленные неконсервативным замещением. Предпочтительно мультимеры включают димеры или тримеры гипоаллергенных вариантов в соответствии с изобретением, связанные линкерной последовательностью или которые подверглись прямому слиянию.
Примерами изоформ являются аллергены Phl р 5а и Phl р 5b, в которых отдельные аминокислоты, которые не являются релевантными для действия в соответствии с изобретением, замещены, или в которых области в аминокислотной последовательности отсутствуют или добавлены (см. фиг. 1а). Эти изоформы аллергенов дикого типа обладают, например, идентичностью аминокислотной последовательности в 63-71%. Другие примеры вариантов в соответствии с изобретением представляют собой варианты изоформ аллергенов дикого типа Phl р 5а и Phl р 5b, такие как, например, Phl р 5.0109, Phl р 5.0201, Phl р 5.0204, Phl р 5.0206, Phl р 5.0207 и т.п., и другие варианты с замещением одной или нескольких аминокислот, отсутствием одной или нескольких аминокислот в N- и/или С-конце или с соответствующими пробелами делеции в пределах аминокислотной последовательности. Равным образом в изобретении предлагаются варианты с внедрениями отдельной или нескольких аминокислот индивидуально в различные положения или как группы в положение в пределах аминокислотной последовательности или в N- и/или С-конец.
Также объектом изобретения являются гипоаллергенные варианты группы 5 аллергенов злаковых (Роасеае), отличающиеся тем, что они представляют собой фрагмент или вариант гипоаллергенного варианта в соответствии с изобретением или мультимер одного или нескольких гипоаллергенных вариантов в соответствии с изобретением или отличающиеся тем, что один или несколько гипоаллергенных вариантов в соответствии с изобретением или их фрагменты, варианты или мультимеры представляют собой составную часть рекомбинантного белка слияния.
Кроме того, объектом изобретение является молекула ДНК, которая кодирует гипоаллергенный вариант в соответствии с изобретением.
Другим объектом изобретения является рекомбинантный вектор экспрессии, содержащий такую молекулу ДНК в соответствии с изобретением, функционально связанный с последовательностью, контролирующей экспрессию. Под последовательностью, контролирующей экспрессию, понимают, например, промотор или участок последовательности, с помощью которого оказывается влияние на экспрессию белка-мишени и который функционально связан с геном-мишенью, однако не обязательно должен локализоваться в непосредственной близости к гену-мишени.
Также изобретение относится к нечеловеческому организму-хозяину, трансформированному посредством молекулы ДНК в соответствии с изобретением или вектора экспрессии согласно изобретению.
Изобретение относится к способу получения гипоаллергенного варианта в соответствии с изобретением путем культивирования нечеловеческого организма-хозяина в соответствии с изобретением и выделения соответствующего варианта аллергена из культуры.
Пригодными нечеловеческими организмами-хозяинами могут быть прокариотические или эукариотические, одноклеточные или многоклеточные организмы, такие как бактерии или дрожжи. Предпочтительным организмом-хозяином в соответствии с изобретением является Е. coli.
Влияние делеции отдельных или двух тесно прилегающих пролинов на IgE связывающую способность Phl р 5а может быть исследовано посредством делеции пролинов 57+58, пролина 85, пролина 117, пролина 146+155, пролина 180, пролина 221, пролина 229 и пролина 256. Эти пролины локализуются в Phl р 5 протеине дикого типа в областях петли в начале или в конце α-спиралей (фиг. 3). Аналогично этому можно исследовать влияние мутаций пролина в соответствующих гомологичных положениях другой предлагаемой в изобретении группы 5 аллергенов злаковых, например, Роа р 5 и Lol р 5 на IgE связывающую способность.
Для более быстрой очистки с высоким выходом, для этих исследований предусматривается кодирующая ДНК с последовательностью, кодирующей N-концевой гексагистидиновый компонент слияния (+6His) (фиг. 5, SEQ ID NO: 3; фиг. 6, SEQ ID NO: 4). Варианты без меток в соответствии с изобретением и белки дикого типа, которые могут использоваться для фармацевтических целей, равным образом очищают стандартными методами и подтверждают результаты His-tag белков.
Соответственно этому получают последовательности, кодирующие белки rPhl p 5а d[P57, Р58] + 6His, rPhl p 5a d[P85] + 6His, rPhl p 5a d[P117] + 6His, rPhl p 5a d[P146, P155] + 6His, rPhl p 5a d[P180] + 6His, rPhl p 5a d[P211] + 6His, rPhl p 5a d[P229] + 6His и rPhl p 5a d[P256] + 6His. Последовательности могут экспрессироваться во всех известных эукариотических и прокариотических экспрессирующих системах, предпочтительно в Е. coil. Затем белки очищают как растворимые мономеры стандартными методами. В конце концов, чистота может быть проверена путем анализа в денатурирующем полиакриламидном геле (НДС-ПАГЭ).
Аналитическая гель-фильтрация (SEC) с соединением рефрактометра (RI детектор) и многоракурсного светорассеивающего детектора (MALS детектор) дает онлайн определение молекулярного веса элюированных белков (метод SEC/MALS/RI).
Кроме того, рекомбинантные варианты в соответствии с изобретением могут быть исследования относительно их связывающей способности с человеческими IgE антителами посредством тестов ингибирования IgE (EAST). В этом способе, взаимодействие аллерген/IgE может быть исследовано в растворе, чем могут быть исключены мешающие маскировки эпитопов тестируемого вещества, например, посредством иммобилизации на мембране.
Сниженное IgE связывание мутантов rPhl p 5а d[P57, Р58] + 6His, rPhl p 5a d[P117] + 6His, rPhl p 5a d[P146, P155] + 6His, rPhl p 5a d[P180] + 6His, rPhl p 5a d[P211] + 6His и rPhl p 5a d[P229] + 6His может быть установлено по сравнению с немодифицированным rPhl p 5а wt (фиг. 7).
Это доказывает по существу, что делеция остатков пролина из группы 5 аллергенов снижает IgE связывающую способность этих аллергенов. С другой стороны только делеция определенных пролинов влечет за собой пониженное IgE связывание.
Поэтому, кроме того, настоящее изобретение относится к гипоаллергенным вариантам группы 5 аллергенов семейства злаковых (Роасеае), в которых пролины, которые соответствуют в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 211 или 229 в аминокислотной последовательности дикого типа Phl p 5.0109, мутировали отдельно.
Настоящее изобретение предпочтительно относится к гипоаллергенным вариантам Phl p 5, Роа p 5, Lol p 5, Dac g 5, Hol Ι 5, Pha a 5, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 211 или 229 в аминокислотной последовательности дикого типа Phl p 5.0109, мутировали отдельно.
Настоящее изобретение особенно предпочтительно относится к гипоаллергенным вариантам Phl p 5, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 211 или 229 в аминокислотной последовательности дикого типа Phl p 5.0109 мутировали отдельно.
Другим предпочтительным объектом настоящего изобретения являются гипоаллергенные варианты в соответствии с изобретением группы 5 аллергенов семейства злаковых (Роасеае), в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 211 или 229 в аминокислотной последовательности дикого типа Phl p 5.0109, были удалены отдельно.
Кроме того, другим объектом настоящего изобретения являются гипоаллергенные варианты в соответствии с изобретением группы 5 аллергенов семейства злаковых (Роасеае), в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 211 или 229 в аминокислотной последовательности дикого типа Phl p 5.0109, были замещены индивидуально. Здесь, пролин заменен, к примеру, лейцином (L). Однако согласно изобретению пролины, предлагаемые в изобретении, могут быть заменены любой аминокислотой.
В частности, гипоаллергенные варианты rPhl p 5а d[P57], rPhl p 5a d[P58], rPhl p 5a d[P57, P58], rPhl p 5a d[P117], rPhl p 5a d[P146], rPhl p 5a d[P155], rPhl p 5a d[P146, P155], rPhl p 5a d[P180], rPhl p 5a d[P211], rPhl p 5a d[P229], rPhl p 5a P57L, rPhl p 5a P58L, rPhl p 5a P57, P58L, rPhl p 5a P117L, rPhl p 5a P146L, rPhl p 5a P155L, rPhl p 5a P146L, P155L, rPhl p 5a P180L, rPhl p 5a P211L и rPhl p 5a P229L, и т.п., включая все гипоаллергенные варианты в соответствии с изобретением, описанные в дальнейшем, соответствуют изобретению, причем нумерация последовательности Phl p 5.0109 следует.
К тому же, эти примеры не ограничиваются вариантами Phl p 5а, а также относятся, в частности, к Phl p 5b, Роа p 5, Hol I 5, Pha a 5, Ant о 5, Dac g 5, Lol p 5, Fes p 5, Hor v 5, Sec с 5, Tri a 5 и группе 5 аллергенов всех других злаковых. Особенно следует отметить соответствующие варианты Роа p 5, Lol p 5, Dac g 5, Hol I 5, Pha a 5 и Phl p 5b.
Неожиданно варианты с комбинациями делеций из группы мутаций d[P57, Р58], d[P117], d[P146, Р155], d[P180], d[P211] и d[P229] проявляют существенно более сниженную IgE связывающую способность в EAST способе, как показано на примере варианта rPhl p 5а d[P117, 180] + 6His (фиг. 8).
Поэтому другим объектом настоящего изобретения являются гипоаллергенные варианты группы 5 аллергенов семейства злаковых (Роасеае), в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 180, 211, 229 в аминокислотной последовательности дикого типа Phl p 5.0109, мутировали в комбинациях. Предпочтение отдается мутациям путем делеции и путем замещения другими аминокислотами. При этом любая аминокислота может быть выбрана для замены пролином.
Предпочтительно настоящее изобретение относится к гипоаллергенным вариантам Phl p 5, Роа p 5, Lol p 5, Dac g 5, Hol I 5, Pha a 5, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 180, 211, 229 в аминокислотной последовательности дикого типа Phl p 5.0109, мутировали в комбинациях.
Особенно предпочтительно настоящее изобретение относится к гипоаллергенным вариантам Phl p 5, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 146, 155, 180, 211, 229 в аминокислотной последовательности дикого типа Phl p 5.0109, мутировали в комбинациях.
Более того, эти примеры не ограничиваются вариантами Phl p 5, а также относятся, в частности, к Роа p 5, Hol I 5, Pha a 5, Ant о 5, Dac g 5, Lol p 5, Fes p 5, Hor v 5, Sec с 5, Tri a 5 и группе 5 аллергенов всех других злаковых. Особенно следует отметить соответствующие варианты Phl p 5а, Phl p 5b, Роа p 5, Lol p 5, Dac g 5, Hol I 5, Pha a 5.
Особое предпочтение отдается гипоаллергенным вариантам в соответствии с изобретением, в которых пролины 57, 58, 117, 146, 155, 180, 211, 229 были удалены в комбинациях. Также особое предпочтение отдается гипоаллергенным вариантам в соответствии с изобретением, в которых пролины 57, 58, 117, 146, 155, 180, 211, 229 были замещены в комбинациях.
Кроме того, особое предпочтение отдается гипоаллергенным вариантам в соответствии с изобретением, в которых пролины 57, 58, 117, 146, 155, 180, 211, 229 были удалены и/или замещены в комбинациях. Поэтому гипоаллергенные варианты rPhl p 5а d[57, 58, 117], rPhl p 5a d[57, 58, 146], rPhl p 5a d[57, 58, 150], rPhl p 5a d[57, 58, 180], rPhl p 5a d[57, 58, 211], rPhl p 5a d[57, 58, 229], rPhl p 5a d[117, 146, 155], rPhl p 5a d[117, 146, 155, 180], rPhl p 5a d[117,146, 155, 229], rPhl pa 5 P117L, P146L, P155L, P211L, rPhl p 5a d[117, 180, 229] P211L, rPhl p 5a d[117, 180, 229] P211L, rPhl p 5ad[57, 58, 117, 180, 229] P211L и т.п., включая все гипоаллергенные варианты в соответствии с изобретением, описанные ниже, например, соответствуют изобретению, причем нумерация последовательности Phl p 5.0109 следует.
Здесь, пролин заменен, к примеру, лейцином (L). Однако согласно изобретению пролины в соответствии с изобретением могут быть заменены любой аминокислотой. Таким образом, все упомянутые или возможные гипоаллергенные варианты, в которых один или несколько пролинов в соответствии с изобретением замещены другой аминокислотой, соответствуют изобретению. Далее, эти примеры не ограничиваются вариантами Phl p 5, а также относятся, в частности к Phl p 5b, Роа p 5, Hol Ι 5, Pha a 5, Ant о 5, Dac g 5, Lol p 5, Fes p 5, Hor v 5, Sec с 5, Tri a 5 и группе 5 аллергенов всех других злаковых. Тем не менее, особое предпочтение отдается всем упомянутым гипоаллергенным вариантам в соответствии с изобретением Phleum pratense Phl p 5а и Phl p 5b, в частности, основывающимся на Phl p 5.0109.
На основе этого знания, могут быть получены варианты rPhl p 5а d[P57, Р58, Р85, Р117, Р146, Р155, Р180, Р211, Р229, Р256] + 6His (краткая форма: MPV.1 + 6His) и rPhl p 5а d[P57, Р58, Р117, Р146, Р155, Р180, Р211, Р229] + 6His (краткая форма: MPV.2 + 6His), которые содержат как можно большее количество комбинированных делеций.
Эти белки при их экспрессии осаждаются в клетках Е. coli как нерастворимые агрегаты (тельца включений). Тельца включений в большинстве случаев растворяются в денатурирующих агентах, таких как 6-8 молярный раствор гидрохлорида гуанидиния или раствор мочевины, и позже преобразовываются в неденатурирующий водный раствор. Это преобразование в неденатурирующую растворяющую среду представляет собой решающий шаг в очистке белка. Как правило, только такие белки, которые после этого процесса сохраняются растворенными, могут включаться в конечные композиции терапевтических средств. Наиболее распространенным промышленно применяемым способом преобразования денатурированно растворенных белков в водный раствор является способ "быстрого разведения". При этом растворенные в денатурирующем агенте белки добавляют в большой объем неденатурирующего растворителя, причем денатурирующий агент сильно разбавляют.
Свойство растворимости описанных здесь мутантов может быть исследовано систематически. При этом тельца включений, полученные в Е. coli и растворенные в гидрохлориде гуанидиния, разбавляют в десяти разных водных буферных растворах, и затем определяют степень растворимости посредством ультрафиолетовой и видимой спектроскопии.
В ультрафиолетовой и видимой спектроскопии спектр поглощения белкового раствора регистрируется в диапазоне длин волн в 240-800 нм. Нерастворимые агрегаты в белковых растворах абсорбируют в диапазоне длин волн >300 нм, в то время как высокорастворимые белки в этом диапазоне не абсорбируют.
Испытуемые растворы охватывают широкий спектр рН (4.5-9.0) и частично содержат стабилизирующие добавки. Добавки глицерин, Tween 80 и L-аргинина моногидрохлорид очень часто используются в качестве вспомогательных средств при растворении труднополучаемых рекомбинантных белков. Они представляют три наиболее важные группы таких сорастворителей: глицерин как полиспирт, Tween как неионогенное поверхностно-активное вещество и L-аргинина моногидрохлорид как производное аминокислоты.
Хотя варианты MPV.1 + 6His и MPV.2 + 6His могут быть растворены посредством использования денатурирующего агента гуанидиния гидрохлорида, однако систематическое исследование растворимости показывает, что последующее преобразование белков в композицию, не содержащую гидрохлорид гуанидиния, постоянно сопровождается образованием нерастворимых белковых агрегатов и, таким образом, не является возможным (таблица 1, 2).
Во время ПЦР экспериментов, осуществленных для синтеза ДНК, в настоящем примере вследствие многочисленных полимеразных ошибок образуется ДНК, кодирующая белок rPhl p 5а d[P57, Р58, Р229] К61Е, Е205К, P211L + 6His (краткая форма: MPV.3 + 6His). Эта ДНК равным образом экспрессируется в Е. coli. Как и варианты MPV.1 и MPV.2, белок осаждается как тельца включений в клетке Е. coli.
Однако неожиданно белок MPV.3 + 6His может быть легко преобразован в неденатурирующий буферный раствор (таблица 3).
Тесты ингибирования EAST-IgE показывают явно сниженную связывающую способность белка с IgE антителами по сравнению с вариантами rPhl p 5а d[P57, Р58] + 6His и rPhl p 5a d[P229]+6His, которые обладают делециями пролина в только одной петле (фиг. 9).
Для другой характеристики мутаций К61Е, Е205К и P211L варианта MPV.3 + 6His, получают ДНК трех вариантов rPhl p 5а К61Ε + 6His, rPhl p 5a E205K + 6His и rPhl p 5a P211L + 6His и исследуют в EAST. Все три мутанта проявляют слегка сниженную IgE связывающую способность, что указывает на синергическое влияние всех мутаций на сниженную аллергенность MPV.3 + 6His (фиг. 10). При этом как делеция, так и замена пролина 211 показывает явно более сильный отдельный эффект, чем мутации К61Е или Е205К (фиг. 10).
С помощью теста с базофильными гранулоцитами, которые были выделены из цельной крови пациентов, страдающих аллергией на пыльцу, действие сниженной IgE связывающей способности MPV.3 + 6His на активацию человеческих эффекторных клеток исследуется in vitro. При одинаковой концентрации rPhl p 5а wt и MPV.3 + 6His, последний показывает меньшую активацию базофильных гранулоцитов. Это указывает на функционально сниженную аллергенность MPV.3 + 6His (фиг. 11).
Случайно генерированная ДНК варианта MPV.3 + 6His теперь служит отправной точкой для целенаправленной конструкции других вариантов. При этом должна сохраняться высокая растворимость MPV.3 + 6His и должна быть далее снижена IgE связывающая способность посредством вставки дополнительных мутаций пролина.
Прежде всего, получают ДНК вариантов rPhl p 5а d[P57, Р58, Р117, Р146, Р155, Р180, Р229] К61Е, Е205К, P211L (краткая форма: MPV.4) и rPhl p 5а d[P57, Р58, Р117, Р180, Р229] К61Е, Е205К, P211L (краткая форма: MPV.5). Эта ДНК создается как с, так и без гистидинового компонента слияния и экспрессируется в Е. coli. Белки могут быть очищены как растворимые белки из телец включения клеток Е. coli посредством простой процедуры очистки.
Потому что как белки с, так и без компонента слияния могут быть очищены растворимыми, то растворимость не зависит от наличия гистидинового компонента слияния. Гистидиновый компонент слияния равным образом не оказывает влияния на IgE связывающую способность, как показано посредством IgE тестов ингибирования с белками с и без компонента слияния (фиг. 12).
ДНК вариантов rPhl p 5а d[P57, Р58, Р117, Р146, Р155, Р180, Р229] P211L (краткая форма: MPV.6) и rPhl p 5а d[P57, Р58, Р117, Р180, Р229] P211L (краткая форма: MPV.7) создаются исключительно без компонентов слияния и экспрессируются в Е. coli. Они могут равным образом быть очищены как высокорастворимые белки из телец включения посредством простой процедуры очистки.
Вариант MPV.6 отличается от варианта MPV.2 только изменением в положении 211. Вместо делеции Р211, в MPV.6 имеется замена аминокислоты на лейцин. Предельно отличающееся свойство растворимости, таким образом, зависит непосредственно от положения 211. Все мутанты, которые несут в себе мутацию P211L (MPV.3-MPV.7), отличаются высокой растворимостью в различных водных композициях, тогда как мутанты с делецией в положении 211 остаются нерастворимыми (таблица 1 - таблица 5).
Поэтому объектом настоящего изобретения являются гипоаллергенные варианты в соответствии с изобретением группы 5 аллергенов злаковых, в которых остаток пролина аминокислотного положения 211, соответствующий зрелому Phl p 5.0109, не делетируется, а заменяется другой аминокислотой.
Поэтому объектом настоящего изобретения являются гипоаллергенные варианты в соответствии с изобретением группы 5 аллергенов злаковых в которых остаток пролина, соответствующий в одном выравнивании пролину 211 в аминокислотной последовательности дикого типа Phl p 5 или зрелого Phl p 5.0109, не делетируется, а заменяется лейцином.
В аналитической гель-фильтрации (SEC), виды белков не разделены исключительно соответственно их молекулярному весу (молярной массе); здесь равным образом важную роль играют конформация, специфический гидродинамический радиус и возможные взаимодействия с матрицей. Правильное онлайн определение молекулярного веса может быть достигнуто посредством присоединения рефрактометра (RI детектор) и многоракурсного светорассеивающего детектора (MALS детектор) к SEC хроматографической системе (способ SEC/MALS/RI). При этом определяется заданная к моменту измерения концентрация частицы посредством RI детектора, и светорассеяние частиц записывается MALS детектором (Wen et al., 1996, Anal. Biochem. 240:155-166). Мономеры, димеры, мультимеры и агрегаты могут быть определены однозначно посредством SEC/MALS/RI.
Результат от SEC/ MALS/ RI MPV.5 и MPV.7 показывает, что эти варианты существуют в чистом виде и в виде растворимых мономеров (фиг. 13; 14; таблица 6). Два варианта MPV.4 и MPV.6 в каждом случае обнаруживаются как смесь растворимых мономеров и димеров (фиг. 15; 16; таблица 6). Варианты MPV.5 и MPV.7 отличаются от вариантов MPV.4 и MPV.6 в каждом случае только положением 146 и 155. Пролины Р146 и Р155 присутствуют в MPV.5 и MPV.7. Это показывает, что делеция d[P146, Р155] вызывает сильную тенденцию к димеризаци соответствующих вариантов.
IgE связывающая способность оптимизированных вариантов от MPV.4 до MPV.7 сначала исследуется способом, при котором белки иммобилизуют на нитроцеллюлозной мембране и инкубируют с содержащими IgE антитела сыворотками отдельных пациентов, страдающих аллергией на пыльцу трав. Затем комплексы вариант аллергена / антитело окрашивают ферментативной реакцией. В этом способе проявляется очень сильно сниженная IgE связывающая способность всех четырех белков (фиг. 17).
Этот результат подтверждается тестом EAST IgE-ингибирования с репрезентативной сборной сывороткой пациентов, страдающих аллергией, причем варианты MPV.4 и MPV.6 обладают IgE связыванием, которое в каждом случае немного ниже, чем соответствующие мутанты MPV.5 и MPV.7 без мутации d[P146, Р155] (фиг. 18; 19). Меньшее IgE связывание у имеющейся мутации d[P146, Р155] вероятно может быть приписано тенденции к димеризации этих вариантов, которая, возможно, приводит к маскировке IgE эпитопов на контактной поверхности участников димеризации.
Результаты тестирования для активации базофильных гранулоцитов показывают функционально значительно сниженную аллергенность для всех четырех вариантов (фиг. 20; 21; 22; 23).
Таким образом, объектом настоящего изобретения являются гипоаллергенные варианты в соответствии с изобретением группы 5 аллергенов злаковых, отличающиеся тем, что пролин, который в одном выравнивании соответствует пролину 211 дикого типа Phl p 5.0109, не делетирован, а заменен другой аминокислотой.
Предпочтительно изобретение также относится к гипоаллергенным вариантам в соответствии с изобретением группы 5 аллергенов злаковых, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58, 117, 180 и 229 в аминокислотной последовательности дикого типа Phl p 5.0109, мутировали отдельно или в комбинации, предпочтительно делетированы, и пролин 211 заменен лейцином.
Соответственно этому предпочтительно изобретение также относится к гипоаллергенным вариантам в соответствии с изобретением группы 5 аллергенов злаковых, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 57, 58 и 229 в аминокислотной последовательности дикого типа Phl p 5.0109, делетированы и пролин 211 заменен лейцином.
Таким образом, настоящее изобретение также относится к гипоаллергенным вариантам в соответствии с изобретением, в которых пролины, соответствующие в одном выравнивании пролинам в положениях 146 и 155 в аминокислотной последовательности дикого типа Phl p 5.0109, не мутировали. Предпочтительно они существуют в виде мономера.
Также соответствуют изобретению все указанные выше гипоаллергенные варианты согласно изобретению, в которых дополнительно аминокислоты, соответствующие в одном выравнивании лизину в положении 61 и глутаминовая кислота в положении 205 в аминокислотной последовательности дикого типа Phl p 5, мутировали отдельно или в комбинации. Предпочтительно эти аминокислоты замещены.
Т-клеточная реактивность, лежащая в основе действенности специфической иммунотерапии, проверяется in vitro посредством пролиферативного теста с Phl p 5-специфическими Т-лимфоцитами пациентов, страдающих аллергией на пыльцу трав. Например, здесь описаны результаты вариантов аллергенов MPV.4 и MPV.7. Димеризирующий вариант MPV.4 несет все исследуемые здесь мутации (d[P57, Р58, Р117, Р146, Р155, Р180, Р229] К61Е, Е205К, P211L) и таким образом, представляет молекулу с максимально измененной аминокислотной последовательностью. Мутант MPV.7 выбирается в качестве примера мономерной формы с чистыми мутациями пролина. Оба мутанта проявляют хорошую Т-клеточную реактивность, сравнимую с неизмененным аллергеном (таблица 7; таблица 8). Удерживание решающих Т-клеточных эпитопов делает возможным иммунотерапевтическое применение описанных вариантов.
Т-хелперные лимфоциты вступают в реакцию с пептидными фрагментами аллергенов, которые образуются посредством процессов распада в антигенпредставляющих клетках (АПК) и презентируются связанными с МНС классом II молекул на поверхности АПК. Как правило, пептиды имеют длину 13-18 аминокислот, но по причине открытых сбоку участков связывания МНС класса 2 могут быть еще длиннее. Главные точки контакта пептидов с молекулой МНС класса можно найти в сердцевинной последовательности приблизительно 7-10 аминокислот. Аллерген-специфическая активация Т-хелперных лимфоцитов является предпосылкой для их пролиферации и функциональной дифференциации (например, Treg, Тн1 и Тн2). Способность аллергена или варианта аллергена к стимулированию аллерген-специфических Т-лимфоцитов рассматривается как ключ к его действенности.
Все варианты аллергенов, полученные на основе Phl p 5 или Phl p 5.0109 и описанные здесь в экспериментах, показывают существенное удерживание решающих Т-клеточных эпитопов.
Таким образом, впервые описаны варианты группы 5 аллергенов Роасеае, которые обладают новыми белковыми свойствами благодаря изменению остатков пролина. Рассматриваемые остатки пролина локализуются в петлевых областях. Только изменение определенных остатков пролина приводит к новым вариантам, которые отличаются сниженной IgE реактивностью с существенным удержанием Т-клеточной реактивности, и поэтому пригодны для лечебной и профилактической специфической иммунотерапии. Соответствующие молекулы ДНК пригодны для иммунотерапевтической вакцинации.
Вследствие этого объектом настоящего изобретения являются описанные варианты аллергенов, молекулы ДНК и рекомбинантные векторы экспрессии в соответствии с изобретением в качестве лекарственных средств.
Гипоаллергенные варианты, молекулы ДНК и рекомбинантные векторы экспрессии в соответствии с изобретением или лекарственные средства в соответствии с изобретением могут использоваться, в частности, для профилактики и/или для лечения заболеваний и болезненных состояний. Лекарственные средства в соответствии с изобретением особенно пригодны для лечения и/или профилактики аллергий типа 1, т.е. для специфической иммунотерапии (гипосенсибилизации) пациентов с аллергией на пыльцу трав или для профилактической иммунотерапии аллергий на пыльцу трав, которые вызываются группой 5 аллергенов видов Роасеае. Молекулы ДНК и рекомбинантные векторы экспрессии в соответствии с изобретением могут использоваться для соответствующей иммунотерапевтической и иммунопрофилактической ДНК вакцинации.
Изобретение также относится к применению, по меньшей мере, одного гипоаллергенного варианта в соответствии с изобретением для получения лекарственного средства для профилактики и/или терапевтического лечения аллергий типа 1, которые причинно вызываются группой 5 аллергенов злаковых.
Также соответствует изобретению применение, по меньшей мере, одной молекулы ДНК в соответствии с изобретением и/или рекомбинантного вектора экспрессии в соответствии с изобретением, включая их смеси во всех соотношениях, для получения лекарственного средства для иммунотерапевтической ДНК вакцинации.
Кроме того, изобретение относится к фармацевтическому препарату, содержащему, по меньшей мере, один гипоаллергенный вариант в соответствии с изобретением, по меньшей мере, одну молекулу ДНК в соответствии с изобретением и/или, по меньшей мере, один рекомбинантный вектор экспрессии в соответствии с изобретением, включая их смеси во всех соотношениях, и необязательно другие носители и/или вспомогательные вещества для профилактики и/или терапевтического лечения аллергий типа 1.
В особенности, фармацевтические препараты в соответствии с изобретением пригодны для профилактики и/или терапевтического лечения аллергий типа 1 которые причинно вызываются группой 5 аллергенов злаковых.
Фармацевтические препараты в смысле данного изобретения могут применяться как терапевтические агенты в медицине человека или в ветеринарной медицине и, следовательно, могут вводиться людям и животным, в частности млекопитающим, таким как обезьяны, собаки, коты, крысы или мыши, и могут применяться при терапевтическом лечении человеческого или животного организма, а также при борьбе с указанными выше заболеваниями. Далее они могут применяться в качестве диагностических средств или как реагенты.
При применении препаратов или лекарственных средств в соответствии с изобретением, гипоаллергенные варианты, молекулы ДНК или рекомбинантные векторы экспрессии в соответствии с изобретением, как правило, применяют аналогично известным, коммерчески доступным препаратам, предпочтительно в дозах между 0.001 и 500 мг, в случае гипоаллергенных вариантов приблизительно 1-500 μг, предпочтительно 5-200 μг, на дозу в фазе поддержания. Препарат может вводиться один раз или несколько раз в день, например, дважды, три раза или четыре раза в день. Типично дозы в фазе увеличения дозы увеличивают для поддержания дозы. С этой целью возможны схемы увеличения и поддержания дозы. В случае подкожной иммунотерапии (SCIT), например, они могут включать краткосрочные терапии (ограниченное количество инъекций перед началом сезонных жалоб, обычно 4-7 инъекции), предсезонные терапии (начало терапии перед сезоном пыльцы, обычно еженедельными инъекциями во время фазы увеличения и ежемесячными инъекциями с поддержанием дозы до начала сезона пыльцы) или терапии в течение всего года (фаза увеличения дозы типично с до 16 еженедельными инъекциями с последующими ежемесячными инъекциями с поддержанием дозы, при необходимости сниженная доза во время сезона пыльцы). В случае подъязычной иммунотерапии водными или твердыми препаратами (таблетки, пластинки и т.д.), терапия может вводиться с или без фазы увеличения дозы. Предпочтительно терапию осуществляют с помощью ежедневных доз в течение года, но также можно проводить предсезонные или другие схемы употребления (например, каждый второй день, еженедельно, ежемесячно).
Выражение "эффективное количество" означает количество лекарственного средства или фармацевтического действующего вещества, которое вызывает биологический или медицинский ответ в ткани, системе, животном или человеке, который ищет или которого добивается, например, исследователь или доктор.
Кроме того, выражение "терапевтически эффективное количество" означает количество, которое по сравнению с соответствующим субъектом, который это количество не получил, влечет следующие последствия: улучшенное лечение, излечение, предотвращение или ликвидация заболевания, синдрома, состояния, жалобы, расстройства или предотвращение побочных эффектов или также снижение развития болезни, жалобы или расстройства. Понятие "терапевтически эффективное количество" также охватывает количества, которые являются эффективными, чтобы повысить нормальную физиологическую функцию.
Лекарственные средства могут быть приспособлены к введению любым приемлемым путем, например, оральным (включая буккальный или подъязычный), ректальным, пульмональным, назальным, местным (включая буккальный, подъязычный или чрескожный), вагинальным или парентеральным (включая подкожный, внутримышечный, внутривенный или внутрикожный) путем. Такие лекарственные средства могут быть получены всеми известными в фармацевтической области способами, например, посредством соединения действующего вещества с носителем(носителями) или вспомогательным веществом(веществами).
Для парентерального применения служат, в частности, растворы, предпочтительно масляные или водные растворы, далее суспензии, эмульсии или имплантаты. Варианты аллергенов в соответствии с изобретением также могут быть лиофилизированы, и полученные лиофилизаты применяют, например, для получения инъекционных препаратов. Указанные препараты могут быть стерилизованы и/или содержать вспомогательные вещества, такие как смазывающие вещества, консерванты, стабилизаторы и/или смачивающие агенты, эмульгаторы, соли для влияния на осмотическое давление, буферные вещества и/или многие другие действующие вещества. Далее благодаря соответствующей технологии приготовления могут быть получены препараты пролонгированного действия вариантов аллергенов в соответствии с изобретением, например, посредством адсорбции на гидроксиде алюминия, фосфате кальция или тирозине.
В качестве носителей принимают в расчет органические или неорганические вещества, которые пригодны для парентерального введения и не вступают в реакцию с группой 5 аллергенов вариантов в соответствии с изобретением. Примерами таковых являются носители, такие как вода, растительные масла, бензиловые спирты, полиэтиленгликоли, триацетат глицерина, желатины, углеводы, такие как лактоза или крахмал, стеарат магния, тальк, ланолин или вазелин.
Для применения лекарственного средства в соответствии с изобретением преимущественно пригодно парентеральное введение. В случае парентерального введения особенно предпочтительными являются внутривенное, подкожное, внутрикожное или внутрилимфатическое введение. В случае внутривенного введения инъекция может осуществляться непосредственно или как добавка к инфузионным растворам.
К лекарственным средствам, которые приспособлены к парентеральному введению, относятся водные и неводные стерильные растворы для инъекций, которые включают антиоксиданты, буферы, бактериостатики и вещества, способствующие растворению посредством которых композиция становится изотонической с кровью реципиента, подлежащего лечению, и водные и неводные стерильные суспензии, которые могут содержать суспендирующие агенты и загустители. Композиции могут применяться в емкостях для разовой дозы или для многократной дозы, например, запечатанных ампулах и флаконах, и храниться в сублимированном (лиофилизированном) состоянии, так что необходимо только добавление стерильной жидкости-носителя, например, воды для инъекционных целей непосредственно перед применением. Растворы для инъекций и суспензии, приготовленные согласно рецептуре, могут быть получены из стерильных порошков, гранул и таблеток.
По желанию препараты или лекарственные средства в соответствии с изобретением могут содержать одно или несколько других действующих веществ и/или один или несколько агентов, усиливающих действие (адъювантов).
Таким образом, настоящее изобретение далее относится к фармацевтическим препаратам, содержащим другие действующие вещества и/или вспомогательные вещества. Изобретение предпочтительно относится к фармацевтическим препаратам в соответствии с изобретением, отличающимся тем, что другие действующие вещества представляют собой аллергены или их варианты. Примерами других пригодных действующих веществ являются другие аллергены, в частности аллергены злаковых, особенно предпочтительно аллергены из подсемейства Pooideae, предпочтительно из групп Poodae и Triticodae, предпочтительно представленные такими как Phleum pratense, Holcus lanatus, Phalaris aquatica, Dactylis glomerate, Lolium perenne, Poa pratensis, Hordeum vulgare, Secale cereale и Tríticum aestivum, например, группа 1, 2, 3, 4, 5, 6, 7, 10, 12 или 13 аллергенов и их варианты, например, гипоаллергенные варианты, фрагменты, мультимеры, гибридные молекулы или рекомбинантные белки слияния. Кроме того, настоящее изобретение относится к фармацевтическим препаратам, содержащим, по меньшей мере, одно другое вспомогательное вещество, особенно предпочтительно так называемые агенты, усиливающие действие. Примерами агентов, усиливающих действие, являются гидроксид алюминия, монофосфорил липид А, активаторы Toll-подобных рецепторов, такие как, например, липополисахариды и CpG-олигонуклеотиды, витамин D3, микобактериальные антигены и молекулы из паразитов (например, шистосом или филарий), такие как, например, цистатин или ES-62.
Изобретение также относится к комплектам (наборам), состоящим из отдельных упаковок:
а) фармацевтического препарата в соответствии с изобретением, содержащего эффективное количество гипоаллергенного варианта, молекулы ДНК или рекомбинантного вектора экспрессии в соответствии с изобретением;
б) фармацевтического препарата, содержащего эффективное количество другого фармацевтически действующего вещества и/или агента, усиливающего действие.
Комплект содержит пригодные емкости, такие как коробки или картонные коробки, отдельные бутылки, пакеты или ампулы. Комплект может содержать, например, обособленные ампулы, в которых каждый раз находится композиция в соответствии с изобретением, включающая эффективное количество гипоаллергенного варианта, ДНК молекулы или рекомбинантного вектора экспрессии в соответствии с изобретением и композиции другого лекарственного действующего вещества в растворенном или лиофилизованном виде.
Пояснения фигур:
Фиг. 1а: Выравнивание выведенной аминокислотной последовательности группы 5 аллергенов Роасеае:
Выравнивание зрелой группы 5 последовательностей разных видов.
Рамки показывают положение α-спиралей Phl p 6 (PDB запись 1NLX) и С-концевой половины Phl p 5.02 фрагмента (PDB запись 1L3P). Аминокислотные положения остатков пролина надписаны в соответствии с их положением в зрелом Phl p 5.0109.
Ссылки последовательностей: Phl p 5.0101 (Phleum pratense lUIS последовательность, UniProtKB Q40960), Phl p 5.0104 (Phleum pratense lUIS последовательность, UniProtKB P93467), Phl p 5.0109 (Phleum pratense lUIS последовательность, UniProtKB Q84UI2), Phl p 5.0201 (Phleum pratense lUIS последовательность, UniProtKB Q40963), Phl p 6.0101 (Phleum pratense lUIS последовательность, UniProtKB P43215), Lol p 5.0101 (Lolium perenne lUIS последовательность, UniProtKB Q40237), Pha a 5.0101 (Phalaris aquatica lUIS последовательность, UniProtKB Р56164), Dacg 5 (Dactylis glomerata, UniProtKB Q93XD9), Hol I 5.0101 (Holcus lanatus IUIS последовательность, UniProtKB 023972), Hol I 5.0201 (Holcus lanatus IUIS последовательность, UniProtKB 23971), Роа p 5.0101 (Роа pratensis IUIS последовательность, UniProtKB Q9FPR0), Tri a 5 (Triticum aestivum, UniProtKB Q70JP9), Hor v 5 (Hordeum vulgare, EST TC190653).
Фиг. 1b: Сохранение остатков пролина
Фиг. 2: Рабочая модель положения остатков пролина в 3D структуре Phl p 5а
Модель 3D гомологии N-концевого (аминокислоты 31-139 Phl p 5.0109; модельная молекула: Phl p 6, PDB запись 1NLX; изображенная на левой стороне) и С-концевого 4-спирального пучка (аминокислоты 155-255; модельная молекула: Phl p 5b фрагмент, PDB запись 1L3P; справа).
a) Сильно упрощенная модель двух 4-спиральных пучков. Н1-Н8: спирали 1-8. Пролины показаны с их обозначением положения и атомной структуры. Остатки пролина Р146 и Р256 находятся в областях, которые по причине отсутствующих структурных данных в 3D моделях модельных молекул не могли быть показаны (пунктирные линии). Петля, представленная между спиралями 2 и 3, является спекулятивной, так как модельная молекула Phl p 6 не имеет области, гомологичной к этому Phl p 5а диапазону.
b) Модель поверхности. Все изображаемые остатки пролина экспонированы на поверхности (окрашены в черный цвет).
Фиг. 3: rPhl p 5а wt (IUIS запись Phl p 5.0109); кДНК последовательность (GenBank запись: AJ555152; 855 bp), SEQ ID NO: 1
Фиг. 4: rPhl p 5a wt (IUIS запись Phl p 5.0109); выведенная аминокислотная последовательность (UniProtKB запись: Q84UI2; 284 аа), SEQ ID NO: 2
Фиг.5: N-концевой гистидиновый компонент слияния; ДНК последовательность (57 bp), SEQ ΙΟ ΝΟ: 3
Фиг. 6: N-концевой гистидиновый компонент слияния; аминокислотная последовательность (19 аа) SEQ ID: 4
Фиг. 7: Результаты теста IgE ингибирования с Phl p 5а вариантами с делециями пролина в отдельных петлях при использовании сборной сыворотки
(а)+(b): Данные из одного индивидуального эксперимента в каждом случае с двойным определением. Символы представляют собой средние значения дубликатов при измерении восьми концентраций ингибиторов в каждом случае. Горизонтальные линии планок погрешности показывают отдельные значения двойного определения.
(с): Обобщение результатов нескольких индивидуальных экспериментов. Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов. Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 8: Сравнение IgE ингибирования rPhl p 5а d[P117,180] + 6His с однопозиционными мутантами пролина rPhl p 5а d[P117] + 6His и d[P180] + 6His
Обобщение результатов нескольких индивидуальных экспериментов: Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов. Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 9: Сравнение IgE ингибирования MPV.3 + 6His с однопозиционными мутантами пролина rPhl p 5а d[P57, 58] + 6His и d[P229] + 6His
Обобщение результатов нескольких индивидуальных экспериментов:
Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов.
Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 10: Сравнение IgE ингибирования MPV.3 + 6His с однопозиционными мутантами пролина rPhl p 5а d[P211] + 6His, P211L + 6His, К61Е + 6His и E205K + 6His
Обобщение результатов нескольких индивидуальных экспериментов: Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов.
Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 11: Тест на функциональную аллергенность MPV.3 + 6His
Доказательство сниженной функциональной аллергенности MPV.3 + 6His с помощью теста активации базофилов с базофилами четырех клинически определенных пациентов, страдающих аллергией на пыльцу трав (Р).
Горизонтальная линия: уровень стимуляции негативного контроля.
Фиг. 12: тест IgE ингибирования с MPV.4 + 6His и MPV.4
Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов.
Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 13: Определение молекулярного веса MPV.5
Хроматограмма аналитического SEC с онлайн определением молекулярного веса.
Фигура показывает относительный УФ-сигнал при 280 нм (правая ось Y) и молекулярный вес (левая ось Y; линия точки измерения в области пика), нанесенный против времени элюирования (ось X).
Для онлайн определения белковой концентрации использовали рефрактометрический детектор OptilabrEX (Wyatt, Santa Barbara, США). Светорассеяние частиц определяли с использованием многоракурсного детектора MiniDAWN Treos (Wyatt). Вес частиц подсчитывали с помощью программного обеспечения ASTRA 5.3.2.17 (Wyatt) посредством формализма Debeye с инкрементом показателя преломления в 0.180 мл/г.
Колонка: Superdex 200 GL 10/ 300 (GE Healthcare, Uppsala, Швеция). Исключение размеров (t0) находится при 20.45 мин. (соответствует ~670 кД).
Элюент: 20 мМ Tris 8.0, 150 мМ NaCl
Фиг. 14: Определение молекулярного веса MPV.7
Хроматограмма аналитической SEC с онлайн определением молекулярного веса.
Фигура показывает относительный УФ-сигнал при 280 нм (правая ось Y) и молекулярный вес (левая ось Y; линия точки измерения в области пика), нанесенный против времени элюирования (ось X).
Для онлайн определения белковой концентрации, использовали рефрактометрический детектор OptilabrEX (Wyatt, Santa Barbara, США). Светорассеяние частиц определяли с использованием многоракурсного детектора MiniDAWN Treos (Wyatt). Вес частиц подсчитывали с помощью программного обеспечения ASTRA 5.3.2.17 (Wyatt) посредством формализма Debeye с инкрементом показателя преломления в 0.180 мл/г.
Колонка: Superdex 200 GL 10/300 (GE Healthcare, Uppsala, Швеция). Исключение размеров (t0) находится при 20.45 мин. (соответствует ~670 кД).
Элюент: 20 мМ Tris 8.0, 150 мМ NaCl
Фиг. 15: Определение молекулярного веса MPV.4
Хроматограмма аналитического SEC с онлайн определением молекулярного веса.
Фигура показывает относительный УФ-сигнал при 280 нм (правая ось Y) и молекулярный вес (левая ось Y; линия точки измерения в области пика), нанесенный против времени элюирования (ось X).
Для онлайн определения белковой концентрации, использовали рефрактометрический детектор OptilabrEX (Wyatt, Santa Barbara, США). Светорассеяние частиц определяли с использованием многоракурсного детектора MiniDAWN Treos (Wyatt). Вес частиц подсчитывали с помощью программного обеспечения ASTRA 5.3.2.17 (Wyatt) посредством формализма Debeye с инкрементом показателя преломления в 0.180 мл/г.
Колонка: Superdex 200 GL 10/300 (GE Healthcare, Uppsala, Швеция).
Исключение размеров (t0) находится при 20.45 мин. (соответствует ~670 кД).
Элюент: 20 мМ Tris 8.0, 150 мМ NaCl
Фиг. 16: Определение молекулярного веса MPV.6
Хроматограмма аналитического SEC с онлайн определением молекулярного веса.
Фигура показывает относительный УФ-сигнал при 280 нм (правая ось Y) и молекулярный вес (левая ось Y; линия точки измерения в области пика), нанесенный против времени элюирования (ось X).
Для онлайн определения белковой концентрации, использовали рефрактометрический детектор OptilabrEX (Wyatt, Santa Barbara, США). Светорассеяние частиц определяли с использованием многоракурсного детектора MiniDAWN Treos (Wyatt). Вес частиц подсчитывали с помощью программного обеспечения ASTRA 5.3.2.17 (Wyatt) посредством формализма Debeye с инкрементом показателя преломления в 0.180 мл/г.
Колонка: Superdex 200 GL 10/ 300 (GE Healthcare, Uppsala, Швеция).
Исключение размеров (t0) находится при 20.45 мин. (соответствует ~670 кД).
Элюент: 20 мМ Tris 8.0, 150 мМ NaCl
Фиг.17: IgE связывание с иммобилизованным MPV.4, MPV.5, MPV.6 и MPV.7
Тест на связывание IgE с иммобилизованными на полосках нитроцеллюлозной мембраны Phl p 5 белками.
rPhl p 5b wt: рекомбинантный Phl p 5b изоформа
rPhl p 5a wt: рекомбинантный Phl p 5a изоформа
nPhl p 5a/b: Phl p 5 аллерген природного происхождения, состоящий из белка изоформы а и b (Allergopharma)
MPV.4: d[P57, P58, P117, P146, P155, P180, P229] K61E E205K P211L
MPV.5: d[P57, P58, P117, P180, P229] K61E E205K P211L
MPV.6: d[P57, P58, P117, P146, P155, P180, P229] P211L
MPV.7: d[P57, P58, P117, P180, P229] P211L
HSA: альбумин из человеческой сыворотки (негативный контроль).
Всего: контроль для равномерной нагрузки полос. Окрашивание реагентом DB71 (Sigma, США)
SP: сборная сыворотка Bor18/100 (Allergopharma)
Фиг. 18: тест IgE ингибирования с MPV.5 и MPV.4
Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов.
Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 19: тест IgE ингибирования с MPV.6 и MPV.7
Левая полоса (темно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 5 μг/мл.
Правая полоса (бледно-серая): IgE ингибирование по отношению к rPhl p 5а wt при концентрации ингибитора 1.25 μг/мл.
Высота полосы показывает среднее значение, полученное из числа (n) индивидуальных экспериментов.
Планки погрешности показывают максимальные (горизонтальная линия верхней планки погрешности) и минимальные (горизонтальная линия нижней планки погрешности) значения, полученные в числе (n) оцененных индивидуальных экспериментов.
Твердая фаза: rPhl p 5а wt. сборная сыворотка: Bor 18/100, Allergopharma.
Фиг. 20: Тест на функциональную аллергенность MPV.4
Доказательство сниженной функциональной аллергенности MPV.4 с помощью теста активации базофилов с базофилами четырех клинически определенных пациентов, страдающих аллергией на пыльцу трав (Р). Горизонтальная линия: уровень стимуляции негативного контроля.
Фиг. 21: Тест на функциональную аллергенность MPV.5
Доказательство сниженной функциональной аллергенности MPV.5 с помощью теста активации базофилов с базофилами четырех клинически определенных пациентов, страдающих аллергией на пыльцу трав (Р). Горизонтальная линия: уровень стимуляции негативного контроля.
Фиг. 22: Тест на функциональную аллергенность MPV.6
Доказательство сниженной функциональной аллергенности MPV.6 с помощью теста активации базофилов с базофилами четырех клинически определенных пациентов, страдающих аллергией на пыльцу трав (Р). Горизонтальная линия: уровень стимуляции негативного контроля.
Фиг. 23: Тест на функциональную аллергенность; MPV.7
Доказательство сниженной функциональной аллергенности MPV.7 с помощью теста активации базофилов с базофилами четырех клинически определенных пациентов, страдающих аллергией на пыльцу трав (Р). Горизонтальная линия: уровень стимуляции негативного контроля.
Фиг.24: rPhl p 5b wt предшественник (lUIS запись Phl p 5.0201); выведенная аминокислотная последовательность 284 аа (Swiss prot: Q40963.2), SEQ ID NO: 5
Даже без других форм осуществления исходят из того, что специалист в данной области техники может использовать описание выше в самом широком объеме. Поэтому предпочтительные формы осуществления должны рассматриваться только как описательное, а ни в коем случае не как каким-либо образом ограничивающее раскрытие.
Таким образом, следующие примеры должны пояснить изобретение, не ограничивая его. Если не указано другое, то процентные данные означают весовой процент. Все температуры указаны в градусах Цельсия. "Обычная переработка": вода добавляется при необходимости, рН устанавливается, если необходимо, до значений между 2 и 10, в зависимости от строения конечного продукта.
Следующие гипоаллергенные варианты в соответствии с изобретением получали и характеризовали с помощью биотехнологических методов. Тем не менее, получение и характеристика веществ может также проводиться специалистом в данной области техники другими методами. Например, гипоаллергенные варианты в соответствии с изобретением также могут быть синтезированы химически. Изобретение равным образом относится к гипоаллергенным вариантам в соответствии с изобретением, описанным ниже.
Пример 1: Варианты Phl p 5а с одной или двумя смежными делециями пролина
Получение вариантов rPhl p 5а d[P57, Р58] + 6His, rPhl p 5a d[P85] + 6His, rPhl p 5a d[P117] + 6His, rPhl p 5a d[P146, P155] + 6His, rPhl p 5a d[P180] + 6His, rPhl p 5a d[P211] + 6His, rPhl p 5a d[P229] + 6His и rPhl p 5a d[P256]+6His и их иммунологическая характеристика описаны в дальнейшем. Аналогично получается и исследуется рекомбинантный неизмененный аллегрен (rPhl p 5 wt + 6His), и гипоаллергенные варианты другой группы 5 аллергенов злаковых в соответствии с изобретением, а также их белки дикого типа, в частности Lol p 5 и Роа p 5, могут также быть получены и исследованы аналогично.
Конструкция генной инженерией:
Синтез ДНК вариантов осуществляют посредством соединения длинных перекрывающихся ДНК олигонуклеотидов и амплификации ДНК стандартным методом ПЦР. Кодоны выбирают таким образом, что выведенная аминокислотная последовательность основывается на последовательности Phl p 5.0109 (фиг. 3, 4). Мутации для делеций пролина вводят тем, что в реакциях ПЦР используют специфические олигонуклеотиды, которым не хватает соответствующих кодонов для пролина. Олигонуклеотиды выбирают таким образом, что выведенный белок несет гексагистидиновый компонент слияния в 5'-конце (фиг. 5, 6). кДНК лигируют в вектор экспрессии pTrcHis2 Topo (Invitrogen, Carlsbad, США). Правильность ДНК подтверждается последовательностью.
Экспрессия, очищение и биохимический анализ:
Экспрессия осуществляется в Escherichia coli(Тор10 штамм; Invitrogen). rPhl p 5а wt, а также варианты очищают специфическим связыванием N-концевых остатков гистидина с Ni2+ хелатной матрицей (аффинная хроматография с иммобилизованным ионом металла, IMAC; материал: HiTrap, GE Healthcare, Uppsala, Швеция). Отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией.
Доказательство сниженного IgE связывания:
Исследование IgE связывающей способности тестируемых веществ осуществляется с использованием EAST-IgE теста ингибирования (энзимоаллергосорбентный тест). В этом способе, взаимодействие аллерген/IgE может быть исследовано в растворе, чем может быть исключена маскировка эпитопов тестируемого вещества, например, посредством иммобилизации на мембране.
Тест ингибирования EAST осуществляется, как приведено ниже. Микротитрационные планшеты покрывают аллергенами, в данном случае рекомбинантный дикий тип Phl p 5.0109 (rPhl p 5a wt). После удаления несвязанной молекулы аллергена промыванием, планшет блокируется бычьим сывороточным альбумином, чтобы предотвратить более позднее неспецифическое связывание. Антитела IgE пациентов, страдающих аллергией, в виде репрезентативной смеси сывороток пациентов, страдающих аллергией (сборная сыворотка), в приемлемом разведении инкубируют с покрытыми аллергенами микротитрационными планшетами. Количество аллергенсвязанных IgE антител количественно определяется фотометрически через анти-человеческий- IgE/щелочная фосфатаза конъюгат путем взаимодействия субстрата до получения цветного конечного продукта.
Связывание IgE антител ингибируется специфически для вещества посредством растворимого аллергена или проверяемого вещества (рекомбинантный модифицированный аллерген) в зависимости от концентрации. Изображенные на фиг. 7. результаты теста ингибирования IgE с рекомбинантными вариантами аллергена Phl p 5а показывают, что сниженная IgE связывающая способность Phl p 5а вызывается делецией остатков пролина Р57 и Р58; Р117; Р146 и Р155; Р180; Р211 или Р229, а не делецией Р85 или Р256. Сниженное ингибирующее действие указывает на потерю IgE эпитопов.
Пример 2: Варианты Phl p 5а с комбинациями делеций пролина
Получение и иммунологическая характеристика варианта rPhl p 5а d[P117, Р180] + 6His представлены ниже на примере гипоаллергенных вариантов в соответствии с изобретением группы 5 аллергенов Роасеае с комбинациями нескольких делеций пролина, которые соответствуют аминокислотным положениям 57 и 58, 117, 146, и 155, 180, 211 и 229, в пересчете на Phl p 5.0109. Аналогично получают и исследуют рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты другой группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, могут быть получены и исследованы аналогичным образом.
Конструкция генной инженерией:
Синтез вариантов осуществляли посредством соединения длинных перекрывающихся ДНК олигонуклеотидов и амплификации ДНК посредством ПЦР. Кодоны выбирают таким образом, что выведенная аминокислотная последовательность основывается на последовательности Phl p 5.0109. Мутации для делеций пролина вводят тем, что в ПЦР реакциях используют специфические олигонуклеотиды, которым не хватает соответствующих кодонов для пролина. Олигонуклеотиды выбирают таким образом, что выведенный белок несет гексагистидиновый компонент слияния в 5'-конце (фиг. 5, 6). кДНК трансформируют в вектор экспрессии pTrcHis2 Topo (Invitrogen) и в Escherichia coli. Правильность ДНК подтверждается последовательностью.
Экспрессия, очищение и биохимический анализ:
Экспрессия осуществляется в Escherichia coli (Тор10 штамм; Invitrogen). Варианты очищают посредством IMAC. Чистота элюированных белков контролируется посредством НДС-ПАГЭ, и отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией.
Доказательство сниженного IgE связывания:
Исследование IgE связывающей способности тестируемых веществ осуществляется с использованием EAST теста ингибирования с IgE антителами пациентов, страдающих аллергией, которые были использованы в виде репрезентативной сборной сыворотки. Представленные на фиг. 8 результаты показывают, что IgE ингибирование варианта rPhl p 5а d[P117, Р180] + 6His является существенно ниже, чем ингибирование вариантов с только одной из делеций пролина rPhl p 5а d[P117] + 6His и rPhl p 5a d[P180] + 6His. Этот результат показывает, что комбинация отдельных делеций пролина может приводить к повышенной редукции IgE связывающей способности Phl p 5а.
Пример 3: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, Р85, Р117, Р146, Р155, Р180, Р211, Р229, Р256] (MPV.1 + 6His)
Вариант аллергена MPV.1+6His имеет комбинацию делеций всех исследованных остатков пролина Phl 5а. Целью получения является максимально сниженная IgE связывающая способность с приемлемой белковой растворимостью.
Конструкция генной инженерией:
Синтез вариантов осуществляют посредством соединения длинных перекрывающихся ДНК олигонуклеотидов и амплификации ДНК посредством ПЦР. Кодоны выбирают таким образом, что выведенная аминокислотная последовательность основывается на последовательности Phl p 5.0109. Мутации для делеций пролина вводят тем, что в реакциях ПЦР используют специфические олигонуклеотиды, которым не хватает соответствующих кодонов для пролина.
Олигонуклеотиды выбирают таким образом, что выведенный белок несет гексагистидиновый компонент слияния в 5'-конце. кДНК трансформируют в вектор экспрессии pTrcHis2 Topo (Invitrogen) и в Escherichia coli. Правильность ДНК подтверждается последовательностью.
Экспрессия:
Экспрессия осуществляется в Escherichia coli (Тор10 штамм; Invitrogen). Белки осаждаются клеткой-хозяином как нерастворимые тельца включений.
Тест на растворимость:
Агрегаты белков выделяют с чистотой примерно в 80% после клеточного расщепления с использованием стандартного метода и растворяют денатурирующим образом путем обработки 6 молярным раствором гидрохлорида гуанидиния.
Денатурированные белки для этого испытания разбавляют 1:50 при 4°С в объеме 25 мл и в течение ночи выдерживают при 4°С. На следующий день возможно видимое образование осадка перепроверяют органолептически. После парциальной концентрации образцы центрифугируют, и светлые надосадочные жидкости исследуют на наличие нерастворимых микроагрегатов с помощью ультрафиолетовой и видимой спектроскопии.
Систематическое исследование свойств растворимости варианта MPV.1 + 6His показывает, что последующее превращение белков в композицию, не содержащую гидрохлорид гуанидиния, всегда сопровождается образованием белковых агрегатов и таким образом является невозможным (таблица 1)
Пример 4: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, Р117, Р146, Р155, Р180, Р211, Р229] (MPV.2 + 6His)
Получение и иммунологическая характеристика варианта MPV.2+6His представлены ниже на примере гипоаллергенных вариантов группы 5 аллергенов Роасеае с комбинациями белковых делеций, соответствующих аминокислотным положениям 57, 58, 117, 146, 155, 180, 211 и 229, в пересчете на Phl p 5.0109. Аналогично получается и исследуется рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, также могут быть получены и исследованы аналогичным образом.
Конструкция генной инженерией:
Синтез вариантов осуществляют посредством соединения длинных перекрывающихся ДНК олигонуклеотидов и амплификации ДНК посредством ПЦР. Кодоны выбирают таким образом, что выведенная аминокислотная последовательность основывается на последовательности Phl p 5.0109.
Мутации для делеций пролина вводят тем, что в ПЦР реакциях используют специфические олигонуклеотиды, которым не хватает соответствующих кодонов для пролина.
Олигонуклеотиды выбирают таким образом, что выведенный белок несет гексагистидиновый компонент слияния в 5'-конце. кДНК лигируют в вектор экспрессии pTrcHis2 Topo (Invitrogen) и трансформируют в Escherichia coli. Правильность ДНК подтверждается последовательностью.
Экспрессия:
Экспрессия осуществляется в Escherichia coli (ТорЮ штамм; Invitrogen). Белки осаждаются клеткой-хозяином как нерастворимые тельца включений.
Тест на растворимость:
MPV.2 + 6His проявляет плохую растворимость в общем исследованном рН пределе в 4.5-9.0. Образование видимых осадков может быть предотвращено только с раствором, содержащим Tween 80 и L-аргинина моногидрохлорид, однако в дальнейшем также в этой партии обнаруживаются микроагрегаты (таблица 2).
Пример 5: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, Р229] К61Е, Е205К, P211L (MPV.3 + 6His)
Получение и иммунологическая характеристика варианта MPV.3 + 6His представлены ниже на примере гипоаллергенных вариантов группы 5 аллергенов Роасеае с комбинациями делеций остатков пролина 57, 58, 117, 146, 155, 180 или 229 соответственно аминокислотным положениям в Phl p 5 5.0109, в которых остаток пролина 211 мутируется в любую другую аминокислоту или в которых лизин 61 дополнительно преобразуется в глутамат, или глутамат 205 преобразуется в лизин. Аналогично получается и исследуется рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, также могут быть получены и исследованы аналогичным образом.
Воздействие мутаций К61Е, Е205К и P211L, которые содержатся в аминокислотной последовательности MPV.3+6His на IgE связывающую способность исследуется тем, что получают и характеризуют иммунологически варианты rPhl p 5а К61Е+6His, rPhl p 5a Ε205Κ + 6His и rPhl p 5a P211L + 6His.
Конструкция генной инженерией:
Нуклеотидная последовательность, кодирующая MPV.3 + 6His, в этом примере неожиданно создается вследствие нескольких полимеразных ошибок при синтезе кДНК. ДНК лигируется в вектор экспрессии pTrcHis2 Topo (Invitrogen). ДНК, кодирующие варианты rPhl p 5а К61Е + 6His, rPhl p 5a Ε205Κ + 6His и rPhl p 5a P211L + 6His получают целенаправленно и лигируют в вектор экспрессии pTrcHis2 Topo. Правильность созданных последовательностей кДНК проверяется посредством ДНК секвенирования.
Экспрессия:
Экспрессия осуществляется в Escherichia coli (Тор10 штамм; Invitrogen). Белки осаждаются клеткой-хозяином как нерастворимые тельца включений.
Тест на растворимость MPV.3 + 6His:
Белковые агрегаты выделяют с чистотой примерно в 80% после клеточного расщепления и растворяют денатурирующим образом путем обработки 6 молярным раствором гидрохлорида гуанидиния. Серийное испытание разных растворов с целью проверить свойства растворимости MPV.3 + 6His в неденатурирующей среде осуществляется аналогично экспериментам с вариантами MPV.1 + 6His и MPV.2 + 6His.
Неожиданно устанавливается, что вариант MPV.3 + 6His обладает высокой растворимостью при слегка щелочном рН во время процесса разбавления (таблица 3). Это свойство представляет собой противоположность исследованным ранее вариантам MPV.1 + 6His и MPV.2 + и отображает решающее преимущество для приготовления белка в неденатурирующей среде.
Очищение MPV.3 + 6His, а также мутантов rPhl p 5а К61Е + 6His, rPhl p 5a Ε205Κ + 6His и rPhl p 5a P211L + 6His:
Денатурированные белки разбавляют 1:50 в 20 мМ Tris, 150 мМ NaCl, рН 9.0, и удерживают в течение ночи при 8°С. Хроматографическая очистка осуществляется посредством IMAC (HiTrap материал, GE Healthcare) и SEC (Superdex 75 материал, GE Healthcare). В заключение белки находятся в стабильном и растворимом виде в 25 мМ буфера фосфата натрия с 150 мМ NaCl, рН 7.5.
Биохимический анализ:
Чистота контролируется посредством НДС-ПАГЭ. Отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией.
Доказательство сниженного IgE связывания MPV.3 + 6His:
Исследование IgE связывающей способности MPV.3 + 6His осуществляли с использованием EAST теста ингибирования с IgE антителами пациентов, страдающих аллергией, которые используют в виде репрезентативной сборной сыворотки. Представленные на фиг. 9 результаты показывают, что IgE ингибирование MPV.3 + 6His является существенно ниже, чем ингибирование вариантов rPhl p 5а d[P57, Р58] + 6His и rPhl p 5a d[P229] + 6His.
Для другой характеристики мутаций К61Е, Е205К и P211L варианта MPV.3 + 6His с помощью EAST исследуют три варианта rPhl p 5а К61Е + 6His, rPhl p 5а Е205К + 6His и rPhl p 5а P211L + 6His.
Вариант rPhl p 5a P211L + 6His проявляет сравнительно низкое свойство ингибирования, чем протестированный ранее точечный мутант rPhl p 5а d[P211] + 6His. Варианты rPhl p 5a Κ61Ε + 6His и rPhl p 5a E205K + 6His показывают слегка сниженное IgE связывание (фиг. 10).
Можно сделать заключение, что три точечные мутации К61Е, Е205К и P211L способствуют явно пониженному IgE связыванию MPV.3 + 6His.
Доказательство снижения функциональной аллергенности MPV.3 + 6His: Функциональное действие MPV.3 + 6His при сшивании мембраносвязанного IgE эффекторных клеток и их активация исследуют in vitro.
Для теста активации базофилов, гепаринизированную цельную кровь пациентов, страдающих аллергией на пыльцу трав, инкубируют с разными концентрациями тестируемых веществ. При этом аллергические вещества могут связывать специфические IgE антитела, которые ассоциируются с высокоаффинными IgE рецепторами базофильных гранулоцитов. Сшивание комплексов IgE/рецептор, инициированное молекулами аллергенов, приводит к сигнальной трансдукции, которая приводит к дегрануляции эффекторных клеток и тем самым вызываются аллергические реакции in vivo.
Вызванная аллергеном активация базофильных иммуноцитов может быть подтверждена in vitro посредством количественного выражения экспрессии поверхностного белка (CD203c), спаренной с сигнальной трансдукцией сшивания IgE-рецептора (Kahlert et al., Clinical Immunology и Allergy in Medicine Proceedings of the EAACI 2002 (2003) Naples, Italy 739-744). Число поверхностных белков, экспрессирующихся на клетке и процентное значение активированных клеток популяции клеток измеряется высокочувствительно посредством связывания флуоресцентно-меченого моноклонального антитела с поверхностным белком и последующего анализа с помощью флуоресцентно активированной проточной цитометрии.
MPV.3 + 6His здесь показывает сниженную активацию базофильных гранулоцитов пациентов, страдающих аллергией на пыльцу трав, по сравнению с rPhl p 5а wt и тем самым функционально сниженную аллергенность (фиг. 11)
Пример 6: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, 117, 146, 155, 180, Р229] К61Е, Е205К, P211L (MPV.4)
Получение и иммунологическая характеристика варианта MPV.4 представлены ниже на примере гипоаллергенных вариантов группы 5 аллергенов Роасеае с комбинациями делеций остатков пролина 57, 58, 117, 146, 155, 180 или 229 соответственно аминокислотным положениям в Phl p 5.0109), в которых остаток пролина 211 мутирован в любую другую аминокислоту или в которых лизин 61 дополнительно преобразуется в глутамат, или глутамат 205 преобразуется в лизин. Аналогично получается и исследуется рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, также могут быть получены и исследованы аналогичным образом.
Конструкция MPV.4 (+ 6His) генной инженерией:
Для получения кДНК гистидинового белка слияния MPV.4 (MPV.4 + 6His) фрагмент уже клонированной кДНК MPV.1 + 6His переклонируют в уже существующий плазмид MPV.3 + 6His/ pTrcHis2 Topo.
Для получения белка, не содержащего компонент слияния (MPV.4), ДНК лигируется в вектор рТМР (Allergopharma, Reinbek) без участка, кодирующего гистидиновый компонент слияния. Правильность последовательности перепроверяется ДНК секвенированием.
Экспрессия MPV.4 (+ 6His):
Экспрессия осуществляется или как гистидиновый белок слияния (вектор экспрессии pTrcHis2Topo; Invitrogen) в Escherichia coli (Тор10 штамм; Invitrogen), или без компонента слияния (вектор экспрессии рТМР) в Escherichia coli (BL21 штамм; Merck, Дармштадт). В обоих случаях, рекомбинантные белки осаждаются как тельца включений. Белки растворяют, используя 6 молярный раствор гидрохлорида гуанидиния.
Тест на растворимость MPV.4 + 6His:
Серийное испытание разных растворов с целью проверить свойства растворимости MPV.4 + 6His в неденатурирующей среде осуществляют аналогично экспериментам с вариантом MPV.1 + 6His. Установлено, что вариант MPV.4 + 6His обладает растворимостью при слегка щелочном рН (таблица 4)
Очищение MPV.4 + 6His:
Белок слияния очищают в препаративном масштабе посредством IMAC (HiTrap материал, GE Healthcare) и SEC (Superdex 75 материал, GE Healthcare) и в заключение он находится в 25 мМ буфере фосфат натрия с150 мМ NaCl, рН 7.5.
Очищение неслитого белка MPV.4:
Раствор денатурированного неслитого белка сначала разбавляют 1:50 с 20 мМ Tris, 150 мМ NaCl, рН 9.0, аналогично схеме очищения белка слияния и выдерживают при 4° в течение ночи. В первой стадии очищения белок-мишень обогащают посредством хроматографии с гидрофобным взаимодействием (HIC).
При высокой ионной силе белки связываются с HIC колонкой в зависимости от их поверхностной гидрофобности с особой силой, так что они могут быть селективно элюированы путем постепенного снижения концентрации соли.
Разбавленный белковый раствор далее разбавляют сначала 1:2 с 20 мМ Tris, 2 M сульфата аммония, 150 мМ NaCl, рН 9.0, чтобы достичь конечной концентрации сульфата аммония 1 М. Затем этот белковый раствор пропускают через HiTrap бутил-S FF HIC колонку (GE Healthcare, Uppsala, Швеция) и, после связывания белка-мишени постепенно элюируют с 20 мМ Tris, 150 мМ NaCl, рН 9.0.
В качестве подготовки для второй стадии очищения, НIC элюаты перебуферизуют в 20 мМ Tris, 50 мМ NaCl, рН 8.0, через Sephadex G25 column (GE Healthcare).
В качестве второй стадии очищения элюаты наносят на колонку анионообменной хроматографии (HiTrap Q HP, GE Healthcare), и проходящий через колонку материал собирают. Белок-мишень при этом находится в материале, проходящем через колонку, в то время как чужеродные белки связываются с колонкой и эффективно удаляются.
В способе AIEX, белки, имеющие отрицательный поверхностный заряд связываются с колоночным материалом при незначительной ионной силе, в то время как незараженные или положительно заряженные белки не связываются.
В последней стадии AIEX продукт дополнительно очищают через SEC колонку (Superdex 75, GE Healthcare), так что в заключение белок-мишень находится в 20 мМ Tris, 150 мМ NaCl, рН 8.0.
Биохимический анализ MPV.4 (+ 6His):
Чистоту белков проверяют посредством НДС-ПАГЭ. Идентичность MPV.4 без компонента слияния подтверждают определением молекулярного веса с помощью масс-спектроскопии (MALDI-TOF) и секвенирования N-концевых аминокислот (последовательность: A-D-L-G-Y-G-P-A-T) (таблица 6). Отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией.
Анализ молекулярного веса MPV.4 с помощью SEC/MALS/RI неожиданно показывает, что масса элюированных белков находится между 28.2 и 49.3 кД (средняя масса 36.9 кД). Так как теоретическая масса мономера составляет 27.6 кД, то можно исходить из того, что имеется смесь из мономеров и димеров (фиг. 15, таблица 6). Качественно такой же результат получают с белком слияния.
Доказательство сниженного IgE связывания MPV.4 (+ 6His): Исследование IgE связывающей способности осуществляют с использованием EAST теста ингибирования с IgE антителами пациентов, страдающих аллергией, которые используют в виде репрезентативной сборной сыворотки.
Представленные на фиг. 12 результаты показывают, что MPV.4 имеет одинаково сильно сниженное IgE связывание как с, так и без компонента слияния. Таким образом, показано, что сниженная IgE реактивность не зависит от наличия гистидиновой метки.
Простым методом тестирования для определения IgE реактивности специфического IgE из сыворотки пациентов, страдающих аллергией, на мембраносвязанных тестируемых белках является тест-полоска. Для этой цели тестируемые вещества в одинаковой концентрации и количестве связывают рядом с полоской нитроцеллюлозной мембраны при неденатурирующих условиях. Ряд таких мембранных полосок может быть инкубирован параллельно с разными сыворотками пациентов, страдающих аллергией. После стадии промывания специфически связанные IgE антитела становятся видимыми на мембране посредством цветной реакции, активированной посредством конъюгата античеловеческий IgE/щелочная фосфатаза.
Результаты вариантов MPV.4 с использованием индивидуальных сывороток пациентов, страдающих аллергией на пыльцу трав, представлены на фиг. 17. Применяют сыворотки пациентов, страдающих аллергией на пыльцу трав, с антителами против природного Phl p 5 (nPhl p 5a/b, смесь из Phl p 5а и b изоформа). IgE антитела подобным образом вступают в реакцию с рекомбинантным rPhl p 5а wt и равным образом, исследуемым рекомбинантным диким типом b изоформы (rPhl p 5b wt).
Является очевидным, что Phl p 5-специфические IgE антитела всех сывороток пациентов, страдающих аллергией, связывают вариант MPV.4 в сильно сниженной степени, в то время как рекомбинантные белки дикого типа rPhl p 5а wt и rPhl p 5b wt связываются так же сильно как и nPhl p 5a/b.
Доказательство снижения функциональной аллергенности MPV.4:
Функциональное действие MPV.4 при сшивании мембраносвязанного IgE эффекторных клеток и его активирование исследуется как с белками слияния, так и без неслитых белков in vitro. Как белок слияния, так и не содержащий компонент слияния MPV.4 здесь показывают сильно сниженное активирование базофильных гранулоцитов пациентов, страдающих аллергией на пыльцу трав, по сравнению с rPhl p 5а wt и тем самым сильно функционально сниженную аллергенность (фиг. 20).
Пример 7: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, Р117, Р180, Р229] К61Е, Е205К, P211L (MPV.5)
Получение и иммунологическая характеристика варианта MPV.5 представлены ниже на примере гипоаллергенных вариантов группы 5 аллергенов Роасеае с комбинациями делеций остатков пролина 57, 58, 117, 180 или 229 соответственно аминокислотным положениям в Phl p 5.0109, в которых остаток пролина 211 мутирован в любую другую аминокислоту или в которых лизин 61 дополнительно преобразуется в глутамат или глутамат 205 преобразуется в лизин. Аналогично получается и исследуется рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, также могут быть получены и исследованы аналогичным образом.
Конструкция MPV.5 (+ 6His) генной инженерией:
Для получения «ДНК гистидинового белка слияния (MPV.5 + 6His), фрагмент уже клонированной кДНК rPhl p 5а d[P117, Р180] + 6His переклонируют в уже имеющийся плазмид MPV.3 + 6His/ pTrcHis2 Topo. Для получения белка, не содержащего компонент слияния (MPV.5), ДНК лигируется в вектор рТМР (Allergopharma, Reinbek) без участка, кодирующего гистидиновый компонент слияния. Правильность последовательности перепроверяется посредством ДНК секвенирования.
Экспрессия MPV.5 (+ 6His):
Экспрессия осуществляется или как гистидиновый белок слияния (вектор экспрессии pTrcHis2Topo; Invitrogen) в Escherichia coli (Topi0 штамм; Invitrogen), или без компонента слияния (вектор экспрессии рТМР; Allergopharma) в Escherichia coli (BL21 штамм; Merck, Дармштадт).
В обоих случаях рекомбинантные белки осаждаются как тельца включения. Белки растворяют с помощью 6 молярного раствора гидрохлорида гуанидиния.
Очищение MPV.5 + 6His:
Белок слияния очищают в препаративном масштабе посредством IMAC (HiTrap материал, GE Healthcare) и SEC (Superdex 75 материал, GE Healthcare) и в заключение он находится в 25 мМ буфере фосфата натрия с 150 мМ NaCl, рН 7.5.
Очищение неслитого белка:
Раствор ТВ денатурированного неслитого белка сначала разбавляют 1:50 с 20 мМ Tris, 150 мМ NaCl, рН 9.0, аналогично схеме очищения белка слияния и выдерживают при 4° в течение ночи.
В первой стадии очищения, белок-мишень обогащают посредством хроматографии с гидрофобным взаимодействием (HIC). Разбавленный белковый раствор разбавляют далее сначала 1:2 с 20 мМ Tris, 2 M сульфата аммония, 150 мМ NaCl, рН 9.0, чтобы достичь конечной концентрации сульфата аммония в 1 М. Затем этот белковый раствор пропускают через HiTrap бутил-S FF HIC колонку (GE Healthcare, Uppsala, Швеция) и, после связывания белка-мишени постепенно элюируют с 20 мМ Tris, 150 мМ NaCl, рН 9.0.
В качестве подготовки для второй стадии очищения, HIC элюаты перебуферизуют в 20 мМ Tris, 50 мМ NaCl, рН 8.0, через колонку Sephadex-G25 (GE Healthcare). В качестве второй стадии очищения, элюаты наносят на колонку анионообменной хроматографии (HiTrap Q HP, GE Healthcare), и проходящий через колонку материал, который содержит белок-мишень, собирают.
В последней стадии AIEX продукт дополнительно очищают через SEC колонку (Superdex 75, GE Healthcare), так что в заключение белок-мишень находится в 20 мМ Tris, 150 мМ NaCl, рН 8.0.
Биохимический анализ MPV.5 (+ 6His):
Чистоту белков контролируют посредством НДС-ПАГЭ. Отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией. Идентичность не содержащего компонент слияния MPV.5 подтверждается определением молекулярного веса с помощью масс-спектроскопии (MALDI-TOF) и секвенирования N-концевой аминокислотной последовательности (последовательность: A-D-L-G-Y-G-Р-А-Т) (таблица 6).
Анализ MPV.5 посредством SEC/MALS/RI показывает, что белок находится исключительно в виде мономера (фиг. 13, таблица 6). Сопоставимый результат получается с белком слияния. Высокая склонность к димеризации, которая установлена в случае варианта MPV.4, таким образом, зависит от наличия мутации d[P146, Р155].
Доказательство сниженного IgE связывания MPV.5 (+ 6His):
Исследование IgE связывающей способности осуществляется с использованием EAST теста ингибирования с IgE антителами пациентов, страдающих аллергией, которые применяли в виде репрезентативной сборной сыворотки. MPV.5 проявляет одинаково сильно сниженную IgE связывающую способность как с, так и без компонента слияния. IgE связывающая способность является отчасти выше, чем таковая варианта MPV.4, которая может основываться на лучшей доступности IgE эпитопов в случае MPV.5, который находится исключительно в мономерной форме (фиг. 18).
Результаты варианта MPV.5 в тесте-полоске с использованием индивидуальных сывороток пациентов, страдающих аллергией на пыльцу трав, представлены на фиг. 17. Становится очевидным, что Phl p 5-специфические IgE антитела всех сывороток пациентов, страдающих аллергией, связывают вариант MPV.5 в сильно сниженном объеме.
Доказательство снижения функциональной аллергенности MPV.5 (+ 6His):
Функциональное действие MPV.5 при сшивании мембраносвязанного IgE эффекторных клеток и его активация исследуется как с белками слияния, так и с неслитыми белками in vitro. MPV.5 здесь проявляет сильно сниженную активацию базофильных гранулоцитов пациентов, страдающих аллергией на пыльцу трав, по сравнению с rPhl p 5а wt и тем самым сильно функционально сниженную аллергенность (фиг. 21).
Пример 8: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, Р117, Р146, Р155, Р180, Р229] P211L (MPV.6)
Получение и иммунологическая характеристика варианта MPV.6 представлены ниже на примере гипоаллергенных вариантов группы 5 аллергенов Роасеае с комбинациями делеций остатков пролина 57, 58, 117, 146, 155, 180 или 229 соответственно аминокислотным положениям в Phl p 5.0109, в которых остаток пролина 211 мутирован в любую другую аминокислоту. Аналогично получается и исследуется рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, также могут быть получены и исследованы аналогичным образом.
Конструкция генной инженерией:
Для получения ДНК, лигируется ДНК фрагмент MPV.2 + 6His в имеющийся к этому моменту времени вектор rPhl p 5а d[57, 58, 117, 180, 229] P211L/ рТМР (Allergopharma). Правильность последовательности перепроверяется посредством ДНК секвенирования.
Экспрессия:
Экспрессия осуществляется исключительно как белок, не содержащий компонент слияния, в вектор экспрессии рТМР (Allergopharma) в Escherichia coli (штамм BL21; Merck, Дармштадт). Рекомбинантные белки осаждаются как тельца включения (ТВ).
Очищение:
Раствор ТВ денатурированного неслитого белка сначала разбавляют 1:50 с 20 мМ Tris, 150 мМ NaCl, рН 9.0, и выдерживают при 4° в течение ночи.
В первой стадии очищения, белок-мишень обогащают посредством хроматографии с гидрофобным взаимодействием (HIC).
Разбавленный белковый раствор разбавляют далее сначала 1:2 с 20 мМ Tris, 2 M сульфата аммония, 150 мМ NaCl, рН 9.0, чтобы достичь конечной концентрации сульфата аммония в 1 М. Затем этот белковый раствор пропускают через HiTrap бутил-S FF НIC колонку (GE Healthcare, Uppsala, Швеция), и после связывания белка-мишени постепенно элюируют с 20 мМ Tris, 150 мМ NaCl, рН 9.0.
В качестве подготовки для второй стадии очищения, HIC элюаты перебуферизуют в 20 мМ Tris, 50 мМ NaCl, рН 8.0, через колонку Sephadex G25 (GE Healthcare). В качестве второй стадии очищения, элюаты наносят на колонку анионообменной хроматографии (HiTrap Q HP, GE Healthcare), и проходящий через колонку материал, который содержит белок-мишень, собирают.В последней стадии AIEX продукт дополнительно очищают через SEC колонку (Superdex 75, GE Healthcare), так что в заключение белок-мишень находится в 20 мМ Tris, 150 мМ NaCl, рН 8.0.
Биохимический анализ:
Чистоту белков контролируют посредством НДС-ПАГЭ. Идентичность не содержащего компонент слияния MPV.6 подтверждается определением молекулярного веса с помощью масс-спектроскопии (MALDI-TOF) и секвенирования N-концевой аминокислотной последовательности (последовательность: A-D-L-G-Y-G-P-A-T) (таблица 6).
Отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией. При анализе MPV.6 с помощью SEC/MALS/RI очищенные белки элюируют в два пика. Пик 1 представляет собой мономерную форму и пик 2 является димерной формой (фиг. 16; таблица 6).
Способность к димеризации, как также показано при помощи вариантов MPV.4 и MPV.5, сводится к наличию мутации d[P146, 155].
Доказательство сниженного IgE связывания:
Исследование IgE связывания осуществляется с использованием EAST теста ингибирования с IgE антителами пациентов, страдающих аллергией, которые применяли в виде репрезентативной сборной сыворотки. Представленные на фиг. 19 результаты показывают, что MPV.6 обладает сильно сниженным IgE связыванием. Результаты в тесте-полоске с использованием индивидуальных сывороток пациентов, страдающих аллергией на пыльцу, представлены на фиг. 17. Становится очевидным, что Phl p 5-специфические IgE антитела сывороток пациентов, страдающих аллергией, связывают вариант MPV.6 в сильно сниженном объеме.
Доказательство снижения функциональной аллергенности:
Функциональное действие MPV.6 при сшивании мембраносвязанного IgE и активирование базофильных гранулоцитов сильно снижено по сравнению с rPhl p 5а wt, как показывают результаты с цельной кровью двух пациентов, страдающих аллергией на пыльцу (фиг. 22).
Пример 9: Гипоаллергенный вариант rPhl p 5а d[P57, Р58, Р117, Р180, Р229] P211L(MPV.7)
Получение и иммунологическая характеристика варианта MPV.7 представлены ниже на примере гипоаллергенных вариантов группы 5 аллергенов Роасеае с комбинациями делеций остатков пролина 57, 58, 117,180 или 229 соответственно аминокислотным положениям в Phl p 5а дикого типа или Phl p 5.0109, в которых остаток пролина 211 мутирован в любую другую аминокислоту. Аналогично получается и исследуется рекомбинантный немодифицированный аллерген (rPhl p 5 wt + 6His), и гипоаллергенные варианты группы 5 аллергенов злаковых в соответствии с изобретением и их белки дикого типа, в частности Lol p 5 и Роа p 5, также могут быть получены и исследованы аналогичным образом.
Конструкция генной инженерией:
Получение ДНК осуществляется тем, что получают ДНК фрагмент с помощью специфических олигонуклеотидов посредством метода ПЦР и затем переклонируют в уже имеющийся к этому моменту времени вектор rPhl p 5а d[P57, Р58, Р117, Р180, Р211, Р229]/рТМР (Allergopharma). Правильность последовательности перепроверяется посредством ДНК секвенирования.
Экспрессия:
Экспрессия осуществляется как белок, не содержащий компонент слияния, в векторе экспрессии рТМР (Allergopharma) в Escherichia coli (штамм BL21; Merck, Дармштадт). Рекомбинантные белки осаждаются как тельца включений (ТВ).
Тест на растворимость:
Серийное испытание с целью перепроверки свойств растворимости рекомбинантного белка в неденатурирующей среде показывает высокую растворимость белка в слегка щелочном рН диапазоне (таблица 5).
Очищение:
Раствор ТВ денатурированного неслитого белка сначала разбавляют 1:50 с 20 мМ Tris, 150 мМ NaCl, рН 9.0, и выдерживают при 4° в течение ночи. В первой стадии очищения, белок-мишень обогащают посредством хроматографии с гидрофобным взаимодействием (HIC). Разбавленный белковый раствор разбавляют далее сначала 1:2 с 20 мМ Tris, 2 M сульфата аммония, 150 мМ NaCl, рН 9.0, чтобы достичь конечной концентрации сульфата аммония в 1 М. Затем этот белковый раствор пропускают через HiTrap бутил-S FF HIC колонку (GE Healthcare, Uppsala, Швеция) и, после связывания белка-мишени постепенно элюируют с 20 мМ Tris, 150 мМ NaCl, рН 9.0.
В качестве подготовки для второй стадии очищения, HIC элюаты перебуферизуют в 20 мМ Tris, 50 мМ NaCl, рН 8.0, через Sephadex G25 колонку (GE Healthcare). В качестве второй стадии очищения, элюаты наносят на колонку анионообменной хроматографии (HiTrap Q HP, GE Healthcare), и проходящий через колонку материал, который содержит белок-мишень, собирают.В последней стадии AIEX продукт дополнительно очищают через SEC колонку (Superdex 75, GE Healthcare), так что в заключение белок-мишень находится в 20 мМ Tris, 150 мМ NaCl, рН 8.0.
Биохимический анализ:
Чистоту белков контролируют посредством НДС-ПАГЭ. Идентичность не содержащего компонент слияния MPV.7 подтверждается определением молекулярного веса с помощью масс-спектроскопии (MALDI-TOF) и секвенирования N-концевой аминокислотной последовательности (последовательность: A-D-L-G-Y-G-P-A-T) (таблица 6). Отсутствие нерастворимых белковых агрегатов подтверждается ультрафиолетовой и видимой спектроскопией. Анализ посредством SEC/MALS/RI показывает, что элюированные белки находятся исключительно в виде мономеров (фиг. 14; таблица 6). Это сводится к отсутствию мутации d[P146, Р155].
Доказательство сниженного IgE связывания:
Исследование IgE связывания осуществляется с использованием EAST теста ингибирования с IgE антителами пациентов, страдающих аллергией, которые использовали в виде репрезентативной сборной сыворотки. Представленные на фиг. 19 результаты показывают, что MPV.7 имеет сильно сниженное IgE связывание. Результаты в тесте-полоске с использованием индивидуальных сывороток пациентов, страдающих аллергией на пыльцу, представлены на фиг. 17. Становится очевидным, что Phl p 5-специфические IgE антитела сывороток пациентов, страдающих аллергией, связывают вариант MPV.7 в сильно сниженном объеме.
Доказательство снижения функциональной аллергенности: Функциональное действие MPV.7 при сшивании мембраносвязанного IgE и активирование базофильных гранулоцитов пациентов, страдающих аллергией на пыльцу, является сильно сниженным по сравнению с rPhl p 5а wt (фиг. 23).
Т-клеточная реактивность гипоаллергенных Phl p 5а вариантов:
Для исследования Т-клеточной реактивности олигоклональные Т-клеточные линии пациентов, страдающих аллергией на пыльцу трав, устанавливали обычными методами при стимуляции природными nPhl p 5а или rPhl p 5а wt молекулами. В тесте пролиферации разные Т-клеточные линии стимулировали образцовыми аллергенами rPhl p 5а wt, а также модифицированными рекомбинантными вариантами аллергенов. Скорость пролиферация определяли обычными методами посредством встраивания [3Н]-тимидина.
Результаты тестов пролиферации MPV.4 и MPV.7 с Т-клеточными линиями 12 пациентов, страдающих аллергией на пыльцу, представлены здесь на примере для описанных модифицированных вариантов аллергенов.
Т-клеточная реактивность варианта MPV.4, которая несет большую часть мутаций всех исследуемых молекул не является сниженной, несмотря на модификации аминокислотной последовательности по сравнению с неизмененным rPhl р 5а wt, что демонстрирует удержание решающих Т-клеточных эпитопов (таблица 7).
Мутант MPV.7 содержит минимальное число измененных аминокислотных положений всех исследуемых мутантов мультипролина. Как предполагалось молекула стимулирует человеческие Т-лимфоциты сравнительно хорошо, как немодифицированный аллерген rPhl р 5а (таблица 8).








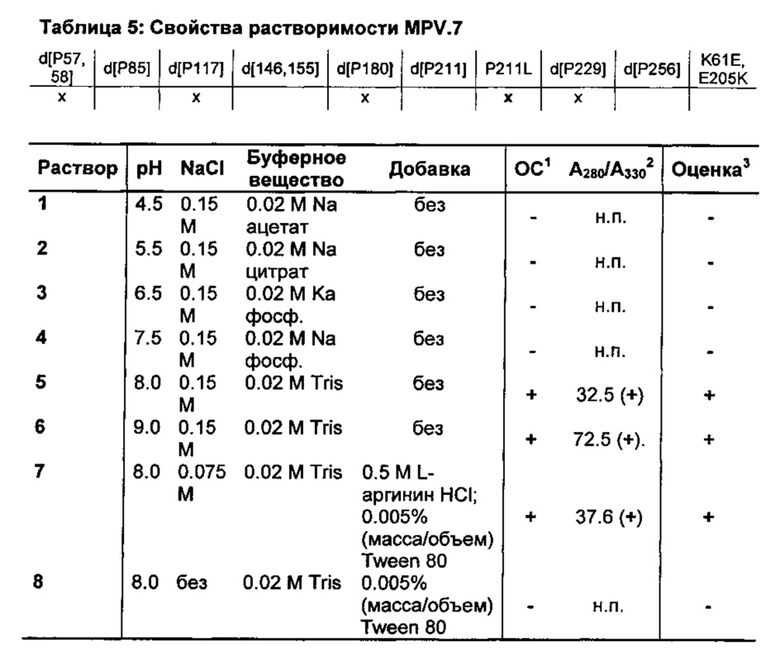

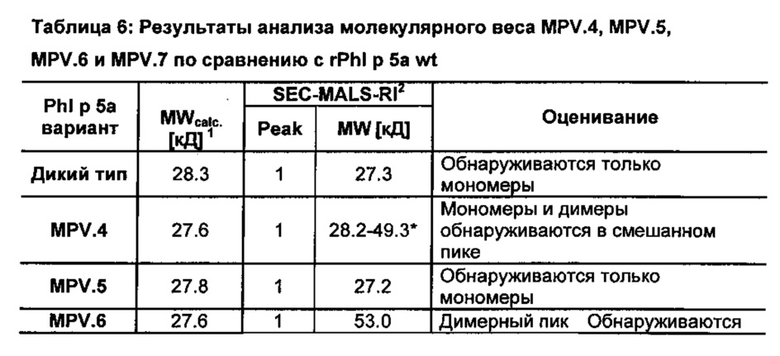

Для онлайн определения белковой концентрации использовали OptilabrEX детектор показателя преломления (RI) (Wyatt, Santa Barbara, США). Светорассеяние частиц определяли с использованием многоракурсного детектора MiniDAWN Treos (Wyatt). Массу частиц рассчитывали с использованием ASTRA 5.3.2.17 программного обеспечения (Wyatt) посредством формализма Debeye с принятым инкрементом показателя преломления в 0,180 мл/г.
Колонка: Superdex 200 GL 10/ 300 (GE Healthcare, Uppsala, Швеция). Исключение размеров (t0) находится при 20.45 мин (соответствует ~670 кД).
Элюент: 20 мМ Tris 8.0, 150 мМ NaCl

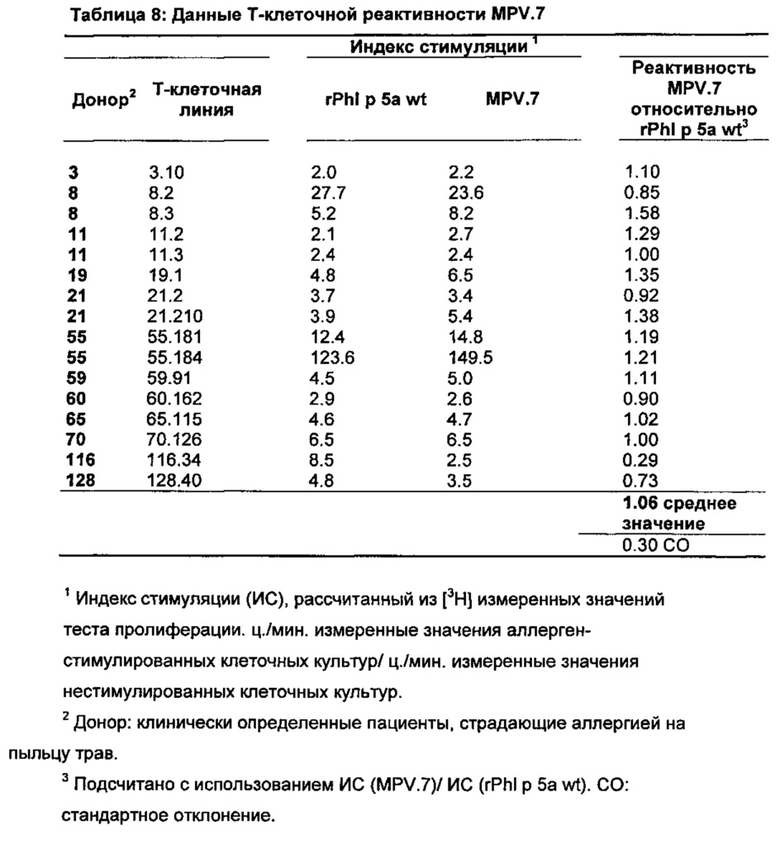
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2575606C2 |
| МОДИФИКАЦИИ АЛЛЕРГЕНОВ ГРУППЫ 6 POACEAE (МЯТЛИКОВЫХ), ИМЕЮЩИХ ПОНИЖЕННУЮ АЛЛЕРГЕННОСТЬ БЛАГОДАРЯ МУТАГЕНЕЗУ ПРОЛИНОВЫХ ОСТАТКОВ | 2010 |
|
RU2607373C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И ПОЛУЧЕНИЕ АЛЛЕРГЕНА ПЫЛЬЦЫ ТРАВ Phl p 4 С ПОМОЩЬЮ РЕКОМБИНАНТНЫХ СПОСОБОВ | 2003 |
|
RU2327739C2 |
| ВАРИАНТЫ ОСНОВНОГО АЛЛЕРГЕНА Phl p 1 ИЗ ТИМОФЕЕВКИ ЛУГОВОЙ | 2003 |
|
RU2323942C2 |
| ПРОИЗВОДНЫЕ PhI p 5а, ОБЛАДАЮЩИЕ СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ И СОХРАНЕННОЙ Т-КЛЕТОЧНОЙ РЕАКТИВНОСТЬЮ | 2004 |
|
RU2368620C2 |
| ГИБРИДНЫЕ БЕЛКИ-НОСИТЕЛИ ПЕПТИДОВ В КАЧЕСТВЕ ВАКЦИН ОТ АЛЛЕРГИИ | 2012 |
|
RU2630652C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ ОСНОВНЫХ АЛЛЕРГЕНОВ ГРУППЫ 4 ИЗ ЗЛАКОВЫХ | 2004 |
|
RU2373283C2 |
| ВАРИАНТ АЛЛЕРГЕНА ГРУППЫ I ИЗ Роасеае, ХАРАКТЕРИЗУЮЩИЙСЯ СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ И СОХРАНЕННОЙ Т-КЛЕТОЧНОЙ РЕАКТИВНОСТЬЮ (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА ДНК И ЕГО ПРИМЕНЕНИЕ | 2005 |
|
RU2409589C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И ПОЛУЧЕНИЕ АЛЛЕРГЕНА ЗЛАКОВЫХ РЕКОМБИНАНТНЫМ СПОСОБОМ | 2000 |
|
RU2238321C2 |
| МИКРОЧАСТИЦЫ, ВКЛЮЧАЮЩИЕ УГЛЕВОДНЫЕ СФЕРЫ, КОВАЛЕНТНО СВЯЗАННЫЕ С АЛЛЕРГЕНОМ | 2003 |
|
RU2329830C2 |










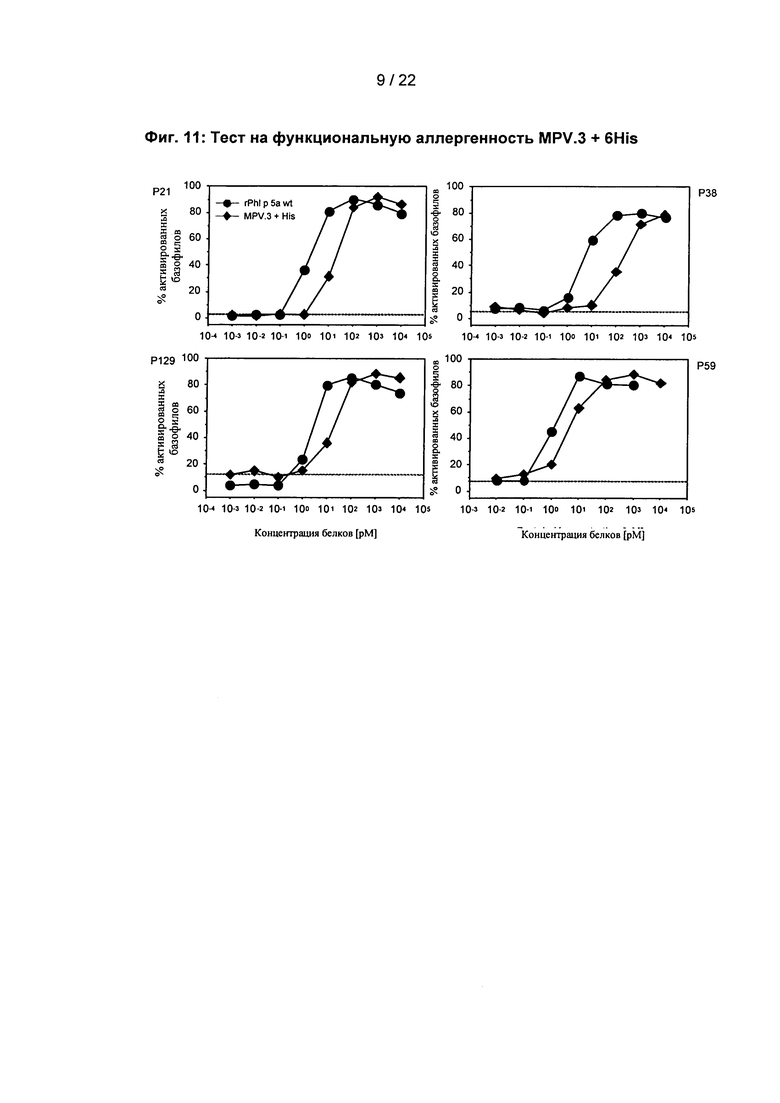
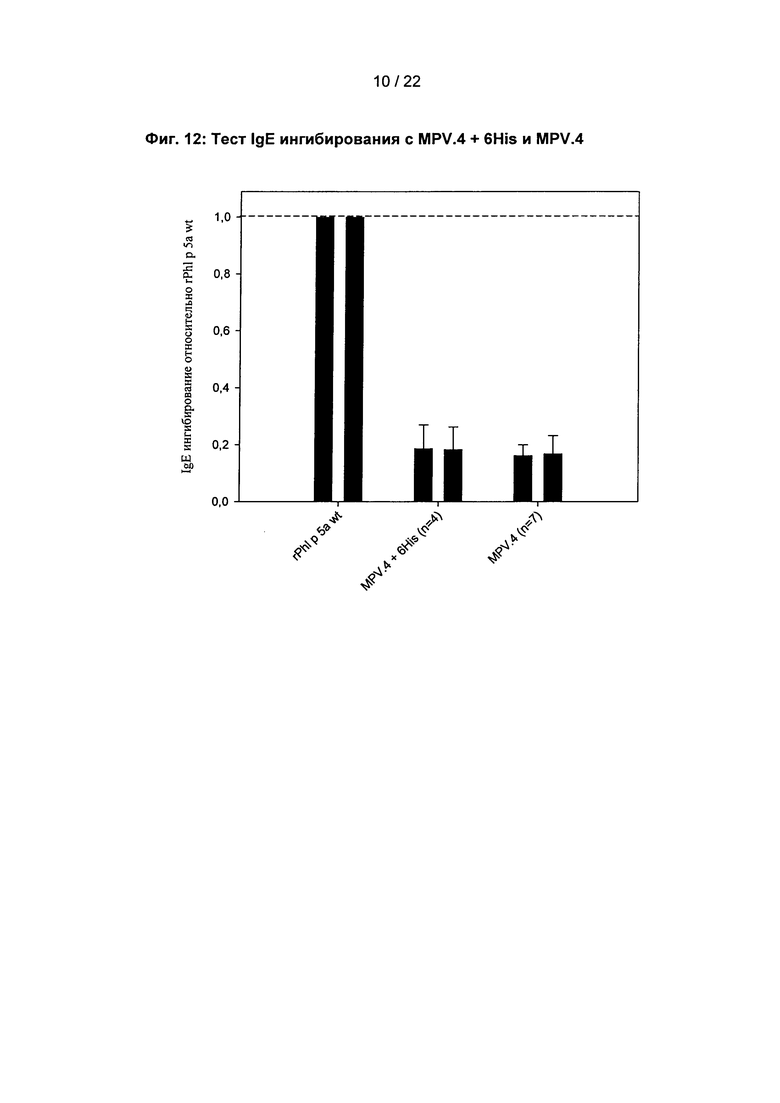







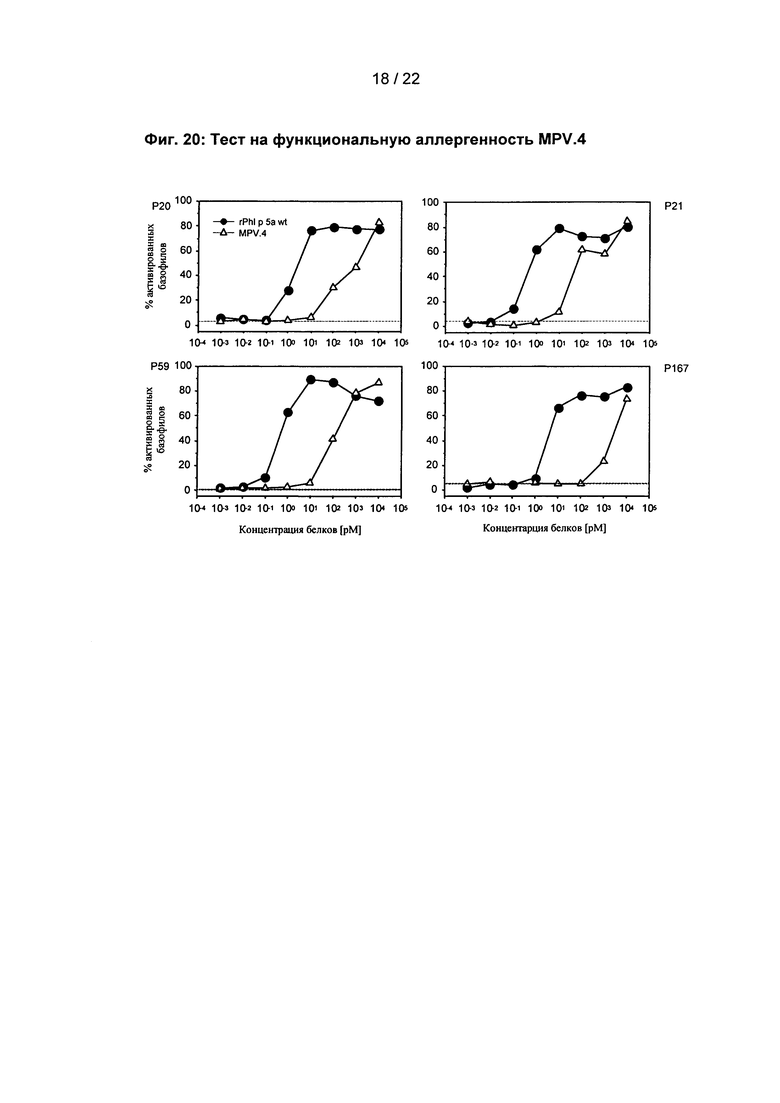




Изобретение относится к области биотехнологии, конкретно к гипоаллергенным полипептидам аллергена злаковых Phl p 5, и может быть использовано в медицине для терапии аллергии типа 1, в инициирование которой вовлечены аллергены пятой группы семейства злаковых (Poaceae). Мутантный полипептид аллергена Phl p 5 получают путем мутаций одного или нескольких пролинов в положениях 57, 58, 117, 146, 155, 180, 211 и 229 аминокислотной последовательности дикого типа Phl р5.0109. Изобретение позволяет получить полипептиды аллергена, которые отличаются сниженной IgE реактивной способностью по сравнению с известными аллергенами дикого типа и одновременно существенно сохраняющейся реактивностью с Т-лимфоцитами. 11 н. и 4 з.п. ф-лы, 24 ил., 8 табл., 9 пр.
1. Гипоаллергенный полипептид аллергена Phl p 5, в котором только пролины, соответствующие при выравнивании пролинам в положениях 57, 58, 117, 146, 155, 180, 211, 229 в аминокислотной последовательности дикого типа Phl р5.0109, представленной в SEQ ID NO. 2, мутировали по отдельности или в комбинации путем точечных мутаций, состоящих из делеций и аминокислотных замен, где точечная мутация представляет собой:
a) делецию одного пролина, выбранного из группы, состоящей из Р57, Р58, Р117, Р146, Р155, Р211 и Р229,
b) комбинацию делеций двух или более пролинов, выбранных из группы, состоящей из Р57, Р58, Р117, Р146, Р155, Р180, Р211 и Р229,
c) замену отдельного пролина, выбранного из группы, состоящей из Р117, Р146 и Р155,
d) комбинацию замен двух или более пролинов, выбранных из группы, состоящей из Р57, Р58, Р117, Р146, Р155, Р180, Р211 и Р229, и
e) комбинацию делеций и замен двух или более пролинов, выбранных из группы, состоящей из Р57, Р58, Р117, Р146, Р155, Р180, Р211 и Р229,
и необязательно дополнительно содержащий по меньшей мере одну мутацию, состоящую из делеций или аминокислотных замен аминокислот, которые при выравнивании соответствуют лизину в положении 61 и/или глутаминовой кислоте в положении 205 в аминокислотной последовательности SEQ ID NO. 2.
2. Гипоаллергенный полипептид по п. 1, отличающийся тем, что пролин, соответствующий в одном выравнивании пролину 211 дикого типа Phl p 5.0109, не делетирован, а заменен другой аминокислотой.
3. Гипоаллергенный полипептид по п. 1 или 2, в котором пролины, соответствующие в одном выравнивании пролинам в положениях 146 и 155 в аминокислотной последовательности дикого типа Phl p 5.0109, не мутированы.
4. Гипоаллергенный полипептид по п. 1 или 2, отличающийся тем, что группа 5 аллергенов представляет собой группу 5 аллергенов подсемейства Pooideae.
5. Гипоаллергенный полипептид по п. 1 или 2, отличающийся тем, что группа 5 аллергенов выбрана из группы, состоящей из Phi p 5, Lol p 5, Poa p 5, Hoi l 5, Dac g 5, Pha a 5, Ant o 5, Fes p 5, Ног v 5, Sec с 5 и Tri а 5.
6. Молекула ДНК, кодирующая гипоаллергенный полипептид по одному из пп. 1-5.
7. Рекомбинантный вектор экспрессии, содержащий молекулу ДНК по п. 6, функционально связанную с последовательностью, контролирующей экспрессию.
8. Прокариотическая клетка-хозяин, трансформированная посредством молекулы ДНК по п. 6, функционально связанной с последовательностью, контролирующей экспрессию, предназначенная для получения гипоаллергенного полипептида по п. 1.
9. Способ получения гипоаллергенного полипептида по одному из пп. 1-5 путем культивирования прокариотической клетки-хозяина по п. 8 и выделения соответствующего полипептида из культуры.
10. Применение гипоаллергенного полипептида по любому из пп. 1-5 в качестве лекарственного средства для лечения или профилактики аллергии типа 1, в инициирование которой вовлечены аллергены пятой группы семейства злаковых (Poaceae).
11. Применение молекулы ДНК по п. 6 в качестве лекарственного средства для лечения или профилактики аллергии типа 1, в инициирование которой вовлечены аллергены пятой группы семейства злаковых (Poaceae).
12. Применение рекомбинантного вектора экспрессии по п. 7 в качестве лекарственного средства для лечения или профилактики аллергии типа 1, в инициирование которой вовлечены аллергены пятой группы семейства злаковых (Poaceae).
13. Применение по меньшей мере одного гипоаллергенного полипептида по одному из пп. 1-5 для получения лекарственного средства для профилактики или терапевтического лечения аллергий типа 1, в инициирование которой вовлечены аллергены пятой группы семейства злаковых (Poaceae).
14. Применение по меньшей мере одной молекулы ДНК по п. 6 и/или рекомбинантного вектора экспрессии по п. 7 для получения лекарственного средства для иммунотерапевтической ДНК вакцинации против аллергенов пятой группы семейства злаковых (Poaceae).
15. Фармацевтическая композиция для лечения или профилактики аллергии типа 1, в инициирование которой вовлечены аллергены пятой группы семейства злаковых (Poaceae), содержащая эффективное количество по меньшей мере одного гипоаллергенного полипептида по любому из пп. 1-5, или по меньшей мере одной молекулы ДНК по п. 6, или по меньшей мере одного рекомбинантного вектора экспрессии по п. 7, и при необходимости другие действующие и/или вспомогательные вещества, где другие действующие вещества представляют собой аллергены злаковых или их варианты.
| GEHLHAR K | |||
| ET AL., Investigation of different recombinant isoforms of grass group-V allergens (timothy grass pollen) isolated by low-stringency cDNA hybridization--antibody binding capacity and allergenic activity, Eur | |||
| J | |||
| Biochem., 1997, v.247, n.1,p.217-223 | |||
| ТЕПЛОИЗОЛЯЦИОННАЯ СМЕСЬ ДЛЯ УТЕПЛЕНИЯ ПРИБЫЛЕЙ ОТЛИВОК | 2011 |
|
RU2455108C1 |
| 0 |
|
SU240676A1 | |
| WO 2009022155 A2, 19.02.2009 | |||
| WO 9934826 A1, 15.07.1999 | |||
| VRTALA S | |||
| cDNA cloning of a major allergen from timothy grass (Phleum pratense) pollen; characterization of the recombinant Phl pV allergen, J | |||
| Immunol., 1993, v.151, n.9, p.4773-4781 | |||
| WO 9843657 A2, 08.10.1998 | |||
| WALD M | |||
| ET AL., Generation of a low immunoglobulin E-binding mutant of the timothy grass pollen major allergen Phl p 5a, Clin | |||
| Exp | |||
| Allergy, 2007, v.37,n.3,p.441-450 | |||
| СПОСОБ ПОЛУЧЕНИЯ ГИПОАЛЛЕРГЕННОГО ОСНОВНОГО АЛЛЕРГЕНА ПЫЛЬЦЫ БЕРЕЗЫ r Bet v 1 | 2003 |
|
RU2311424C2 |

