Настоящее изобретение относится к гипоаллергическим последовательностям вариантов белка Mal d 1, молекулам нуклеиновых кислот, кодирующих их, фармацевтическим композициям, содержащим их, и их применению в иммунотерапии аллергических болезней, вызванных видами Malus domectica (Яблоня домашняя).
Уровень техники, предшествующий изобретению
Аллергии вызываются дисфункцией в иммунной системе, которая реагирует с безвредными белками, в основном содержащимся в пыльце, клещах, эпителии и пищевых продуктах, посредством выработки антител класса IgE.
Последние оценки указывают на то, что более 25% популяции индустриализированных стран страдает от этих заболеваний, которые, если будут продолжаться, могут привести к ухудшению симптомов (напр., возникновение астмы) и сенситизации к другим аллергенам, таким образом, осложняя выбор наиболее подходящего лечения.
Около трети всех подверженных аллергии пациентов в мире подвержены аллергии к пыльце деревьев. В умеренных районах пыльца деревьев, принадлежащих к порядку Fagales (Букоцветные: береза, ольха, лесной орех, дуб и граб) являются одной из главных причин аллергической астмы и ринита. Около 95% пациентов, подверженных аллергии на березовую пыльцу вырабатывают антитела IgE против Bet v 1, и 60% этих пациентов реагируют только на Bet v 1 (кДНК, депонированная в кДНК GenBank с номером доступа X15877), главный аллерген березовой пыльцы (1).
У высокого процента индивидуумов, подверженных аллергии к пыльце березы (50-93%), развиваются аллергические реакции после съедания некоторых продуктов (яблоки, морковь, лесные орехи и сельдерей). Этот тип гиперчувствительности у пациентов с аллергией пыльцу березы, характеризуемый аллергией на фрукт, орехи и овощи, описан как "перекрестный пыльцевой и пищевой синдром" (PFS). Симптомы варьируются от реакции слизистой оболочки верхнего пищеварительного тракта (синдром оральной аллергии) и желудочно-кишечного тракта до уртикарии и астмы, и в некоторых случаях могут вызвать анафилактический шок. Нежелательные реакции на фрукт происходят наиболее часто с яблоком. В случае поллиноза, вызванного березовой пыльцой этот синдром в основном опосредован перекрестно-реагирующими антителами IgE первоначально индуцированными Bet v 1(2).
Лечение таких пациентов главным образом основано на исключении пищевого аллергена из диеты. Было доказано, что для лечения аллергии на пыльцу, животных и клещей понижающая восприимчивость специфическая иммунотерапия (SIT) в отличие от фармакологического лечения является эффективной формой этиологического лечения, которое положительно воздействует на некоторые иммунологические параметры, которые лежат в основе заболевания. SIT содержит введение возрастающих доз стандартизированных экстрактов (вакцины), полученных из вещества, которое вызывает заболевание (3). В этом пути у пациента индуцируется вид "иммунологической устойчивости" по отношению к этому веществу постепенно, сопровождаемый снижением, если не исчезновением симптомов аллергии. С другой стороны специфическая иммунотерапия не является широко используемой для лечения пищевых аллергий в связи с высоким риском индуцирования серьезных побочных эффектов, включая анафилактический шок.
Хотя перекрестная реактивность между Bet v 1 и гомологичными пищевыми белками лежит в основе реакций, индуцированных PFS, было обнаружено, что SIT с березовой пыльцой, не индуцирует какое-либо улучшение симптомов пищевой аллергии.
Такая неэффективность наблюдалась в случае аллергии на морковь, в основном вызванной аллергеном Dau с 1, гомологом Bet v 1, в которой возникает первичная сенситизация по отношению к березовому аллергену и последующая сенситизация, после съедания моркови, по отношению к новым эпитопам Dau с 1, которые не дают перекрестную реакцию с Bet v 1 (4).
В лечении яблочной аллергии SIT с экстрактом березовой пыльцы привело к неоднозначным, иногда благоприятным результатам, вероятно из-за большей идентичности последовательности между Bet v 1 и Mal d 1 (60% аминокислотной последовательности, по сравнению с 40% для Dau с 1).
В исследовании 1998 года у 84% пациентов, подверженных аллергии к березе и яблоку, которых лечили с SIT, основанной на экстракте березовой пыльцы, значительное снижение (50-95%) или полное исчезновение симптомов OAS (синдром оральной аллергии) после съедания яблока происходило уже после первого года лечения (5). В более позднем исследовании, опубликованном в 2004, SIT с экстрактом березовой пыльцы привело к значительному снижению SPT реактивности к Bet v 1 и Mal d 1 после лишь трех месяцев лечения. Антитела IgG4 сильно индуцированные против Bet v 1, проявляли перекрестную реактивность к Mal d 1, таким образом, подтверждая гипотезу, что SIT против аллергии на пыльцу березы может также снижать аллергию на продукты, содержащие гомологичные Bet v 1 аллергены (6). Другое исследование, опубликованное в том же году (2004) показало, что у пациентов, подверженных аллергии к березе и яблоку, SIT, основанная на экстракте березовой пыльцы, значительно улучшала аллергические симптомы, вызванные пыльцой, но не снижала тяжесть аллергии на яблоко (7).
Вследствие этого на основании современного уровня знаний невозможно достоверно утверждать, что иммунотерапия с аллергенами березовой пыльцы предоставляет значительные эффекты в случае аллергии на продукты, содержащие гомологи Bet v 1, которую вследствие этого необходимо лечить как отдельное заболевание.
Одним из аллергенов, наиболее часто вовлеченных в перекрестную реактивность PFS к Bet v 1, является Mal d 1, главный аллерген яблока, фрукта, принадлежащего к семейству розоцветных. Mal d 1 разделяет приблизительно 60% его аминокислотной последовательности с Bet v 1 (идентичность), и эксперименты перекрестной реактивности продемонстрировали наличие общих эпитопов IgE и T (8, 9).
Mal d 1 (номер доступа Q9SYW3 или номер AJ417551) представляет собой белок из 159 аминокислот с молекулярной массой 17,7 кДа. (10). Этот аллерген принадлежит к семейству PR-10 "связанные с патогенезом белки", т.е. убиквитарные белки, вырабатываемые растениями в ответ на стресс окружающей среды и патологический стресс, функция которых, как предполагается, связана со стероидным транспортом. Mal d 1 находится как кожуре, так и в мякоти яблок. Измерение содержания Mal d 1 в белковых экстрактах из многочисленных сортов яблок демонстрирует, что этот аллерген присутствует в значительно различающихся концентрациях в различных сортах; изменчивость в содержании Mal d 1 также была обнаружена в одном и том же сорте, выращенном в различных местах. Более того, даже в сортах с низким содержанием Mal d 1, наблюдалась то, что концентрация аллергена значительно возрастала во время созревания и хранения фрукта.
Mal d 1 представлен семейством генов с по меньшей мере восемнадцатью представителями, характеризуемым наличием или отсутствием интронов в гене. Некоторые представители высоко консервативны в различных сортах яблок (Mal d 1.01, Mal d 1.02), тогда как другие представлены более вариабельно (Mal d 1.04, Mal d 1.05, Mal d 1.06 А, В, C) (11). При проведении анализа SPT пациентов с перекрестным пыльцевым и пищевым синдромом наблюдалась связь между белковыми вариантами, кодируемыми генами Mal d 1.04 и Mal d 1.06A, и большая аллергенность сортов яблок. Результаты этого типа исследования могут иметь применения в идентификации и культивировании сортов яблока с более низкой аллергенностью для применения в качестве пищи или в качестве сырьевого материала для общепринятой иммунотерапии.
Разработка гипоаллергических пищевых аллергенов для применения в специфической иммунотерапии может являться хорошей альтернативой исключению пищевых аллергенов и позволяет лечение заболевания, которое в дополнение к наличию высоких рисков для здоровья пациентов имеет значительный негативный вклад в качество жизни и может вызвать инвалидизирующие нарушения пищевого баланса.
В последние годы большее внимание было сосредоточено на развитии более безопасных, более эффективных вакцин, состоящих из рекомбинантных белков, мутагенизированных на уровне аминокислот, важных для связывания IgE, а именно гипоаллергических вариантов, которые благоприятно влияют на естественное прогрессирование заболевания, не вызывая нежелательные эффекты (12).
Некоторые исследования гипоаллергических вариантов Mal d 1 или гомологичных белков из другого фрукта, принадлежащих к семейству розоцветных, доступны в литературе.
На основании перекрестной реактивности между Bet v 1 и Mal d 1 выработан мутант Mal d 1, главного аллергена яблока, сайт-направленным мутагенезом пяти аминокислотных остатков изоформы Mal d 1.0108 (T10P; I30V, T57N, T112C и I113V), выбранных по аналогии с гипоаллергическим мутантом аллергена Bet v 1 (9). Замена указанных аминокислот снижала аллергеннную активность Mal d 1 на 90%, как продемонстрировано посредством SPT и анализа DBPCFC (двойная слепая плацебо-контролируемая пищевая провокация). Тот же мутант с пятью аминокислотными заменами тестировали параллельно с аналогичным мутантом Bet v 1 и с соответствующими молекулами дикого типа (13). Хотя сыворотки пациентов, подверженных аллергии к березе и яблоку, проявляли большую IgE реактивность против Bet v 1 в тестах методом иммуноблоттинга, наблюдалось снижение связывания с обоими тестируемыми мутантами. В методе ELISA (твердофазный иммуносорбентный анализ) в большей части тестируемых сывороток (10/14) наблюдалось снижение от 30 до 88% в IgE-связывании мутанта Mal d 1 по сравнению с аллергеном дикого типа. Однако, мутагенез пяти выбранных аминокислот, по-видимому, не наносит вред IgE реактивности основных эпитопов Mal d 1, так как не наблюдалось изменения в специфическом связывании IgE в некоторых из анализируемых сывороток (3/14).
Было доказано, что в Pru av 1, главном аллергене вишни, гомологичном Mal d 1, точечная замена серина 112 является критичной для распознавания молекулы посредством IgE. Мутагенез той же аминокислоты в пролин в гомологе Bet v 1 подтвердил важность серина 112 в сохранении структуры перекрестнореагирующего эпитопа IgE. Замена Glu45 на триптофан в последовательности P-петли Pru av 1 продемонстрировала, что эта область представляет собой эпитоп IgE, перекрестнореагирующий с Bet v 1 (14). Три других варианта Pru av 1 получали мутагенезом аминокислот в положении 28 (Asn28Lys), 108 (Pro 108Ala) или в обоих. Наблюдалось уменьшенное связывание IgE в вплоть до 80% сывороток пациентов, подверженных аллергии на березу и вишню, для одиночного мутанта (Asn28Lys) и двойного мутанта (Asn28Lys, Pro108Ala), тогда как было получено снижение только на 12% для одиночного мутанта по Pro 108, что предполагает, что аминокислота Asn28 в Pru av 1 вовлечена в эпитоп IgE. Этот аспарагин экспонирован к растворителю, и является частью одной из областей, являющихся, как предполагается, основными антигенными сайтами в гомологичном белке Bet v 1 (15).
Исследование трехмерной структуры Bet v 1 посредством ЯМР анализа и дифракции рентгеновых лучей привело к идентификации трех зон с площадью, превышающей 600 Å, которые могут быть задействованы в связывании IgE (16). Такие три зоны, экспонированные на поверхность, состоят из высококонсервативных аминокислот в гомологичных аллергенах, эспрессируемых в видах, принадлежащих к порядку Букоцветные, и было предположено, что они являются потенциальным эпитопами IgE, ответственными за перекрестную реактивность между Bet v 1 и гомологичными белками растительной пыльцы. Сайт-направленный мутагенез аминокислот в таких областях подтвердил их задействование в связывании IgE. Мутации в Bet v 1, которые характеризуют два множественных мутанта (T28, Q32, S45, G108) и (V5, S42, S45, K78, V103,1123, E134, H156, N160), например, модифицируют вплоть до пяти различных областей, рассредоточенных на молекулярной поверхности, включая три зоны, описанные выше, и вызывают снижение аллергенности (17). Аминокислотные последовательности, которые составляют три эпитопа IgE, предположенные Gajhede, также являются консервативными в пищевых аллергенах, гомологичных Bet v 1. Для того чтобы снизить сходство поверхности с Bet v 1, консервативные аминокислоты Thr 28, Gin 32 и Ser 45 Mal d 1, расположенные в одной из областей, описанных выше, были заменены аминокислотами, не представленными в Bet v 1 в соответствующих положениях. Замена таких остатков не изменяла способность мутанта Mal d 1 ингибировать связывание IgE-Bet v 1 по сравнению с аналогом дикого типа (дт), в то время как выброс гистамина был прекращен в базофилах крови тех же пациентов с аллергией на березу только в одном из пяти тестируемых случаев (24).
Для того чтобы идентифицировать эпитопы IgE, задействованные в клинических симптомах перекрестного пыльцевого и пищевого синдрома, был синтезирован химерный белок трансплантацией четырех коротких пептидных участков Mal d 1 в Bet v 1 последовательность (18). Трансплантированные области содержали аминокислотные остатки, которые, как предварительно было доказано, являлись критическими для связывания IgE пациентов с обоими аллергенами Bet v 1 и Mal d 1: T10, F30, S57, S112, I113 и D125. IgE реактивность химерного белка тестировали с применением сывороток двух групп пациентов с аллергией на березу, без PFS или демонстрирующих PFS симптомы после съедания яблока. Химерная молекула распознавалась IgE обеих групп, но связывание было значительно ниже в группе без PFS по сравнению с пациентами с аллергией на яблоко, что предполагает, что трансплантированные последовательности Bet v 1 были задействованы в Bet v 1/Mal d 1 перекрестной IgE реактивности.
Аллергия на пыльцу березы, характеризуемая PFS, представляет отличную модель исследования для идентифицирования перекрестно-реагирующих эпитопов IgE и обеспечения производства рекомбинантных аллергенов с уменьшенной способностью связывания с IgE.
Возможность использования сконструированных мультимерных молекул, состоящих из различных аллергенов из одного и того же организма или различных организмов для специфической понижающей восприимчивость терапии долгое время считалась чрезвычайно интересной. Этот подход позволяет собрать несколько аллергенов в одиночную молекулу с преимуществом, заключающемся в продуцировании одного препарата, содержащего аллергены в точном молярном соотношении. Объединение Bet v 1 и Mal d 1 в одиночную гибридную молекулу должно представлять T-эпитоп репертуар обеих молекул, и может индуцировать сильный IgG защитный ответ (17) против обоих аллергенов. Индукция антител IgG, специфических для сенсибилизирующего аллергена, является одним из факторов, коррелирующих с эффектом, индуцированным SIT. Такие (защитные) антитела могут ингибировать связывание IgE с антигеном, изменяя трехмерную конформацию молекулы.
Были осуществлены некоторые исследования для изучения клинических эффектов на аллергию на пыльцу березы SIT с тримерными производными аллергена Bet v 1 (19). Лечение индуцировало сильный IgG1- и IgG4-специфический к аллергену гуморальный иммунный ответ и уменьшенную назальную и кожную реактивность, но не приводило к значительному улучшению клинических симптомов.
Значительный IgG иммунный ответ также наблюдался в экспериментах с мышами, обработанных димерной дт или мутантной формой в положении 112 аллергена моркови Dau с 1, гомологичного аллергену Bet v 1 (20). Было доказано, что оба димерных варианта были более антигенными, чем смесь соответствующих мономерных форм. Кроме того, все мышиные сыворотки, выработанные против Dau с 1 (в мономерной, димерной, дт и мутантной форме), содержали специфические антитела, перекрестно-реагирующие с эпитопами, распознаваемыми человеческим IgE, что указывает на то, что структурно измененные антигены, подобные димерами, способны индуцировать выработку антител, специфических для конформационных эпитопов.
Применение гипоаллергического гибридного варианта, такого как гибридный вариант Bet v 1 - Mal d 1, может устранить сложности, вызванные сильными нежелательными реакциями на пищевые аллергены, и улучшает толерогенный ответ не только на Bet v 1, как происходит, когда применяется SIT с экстрактом березовой пыльцы, но также и на главный аллерген яблока.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Было обнаружено, что связывание аллергена Mal d 1 с IgE может быть уменьшено модифицированием его последовательности посредством замены специфических аминокислотных остатков.
В соответствии с первым аспектом изобретение предоставляет вариант последовательности Mal d 1, полученный из главного аллергена Malus domectica, Mal d 1 (последовательность дикого типа SEQ ID NO:1) или ее изоформу, обладающую по меньшей мере 95% идентичности последовательности с SEQ ID NO:1, предпочтительно по меньшей мере 97%, при этом указанный вариант характеризуется:
a) уменьшенной реактивностью к IgE по сравнению с Mal d 1 дт SEQ ID NO:1;
b) аминокислотная последовательность, которая при выравнивании с SEQ ID NO:1, имеет по меньшей мере одну, и предпочтительно две замены в Asp и/или Asn остатках в положениях 25 и 78 SEQ ID NO:1 или в соответствующих положениях указанных изоформ Mal d 1.
Изоформы Mal d 1, обладающие по меньшей мере 95% идентичности последовательности c SEQ ID NO:1, содержат природные последовательности, депонированные с номерами доступа Mal d 1.0201 (Uniprot Q40280) и Mal d 1.0204 (Uniprot Q9SYV4).
Предпочтительными вариантами аллергена Mal d 1 являются те, в которых остаток Asp 25 замещен нейтральной, полярной или основной аминокислотой, которая предпочтительно выбрана из Ala, Thr, Gly, Pro, Leu, Ile, Ser, Phe, Lys и Arg, и более предпочтительно из Ala, Thr, Ser, Gly, Lys и Arg, при этом остаток Asn 78 замещен нейтральной, кислой или основной аминокислотой, которая предпочтительно выбрана из Ala, Gly, Pro, Leu, Ile, Phe, Lys, Arg, Asp и Glu, и более предпочтительно из Ala, Gly, Lys, Arg, Asp и Glu.
В предпочтительном варианте осуществления вариант в соответствии с изобретением, несущий 2 замены, имеет последовательность, определяемую SEQ ID NO:2.
Замененный вариант аллергена Mal d 1 в соответствии с настоящим изобретением демонстрирует снижение реактивности к IgE по меньшей мере на 10% по сравнению с молекулой дикого типа, предпочтительно по меньшей мере на 50%, и более предпочтительно по меньшей мере на 80%, в сыворотке пациентов, подверженных аллергии к пыльце березы бородавчатой (Betula verrucosa) и яблоку.
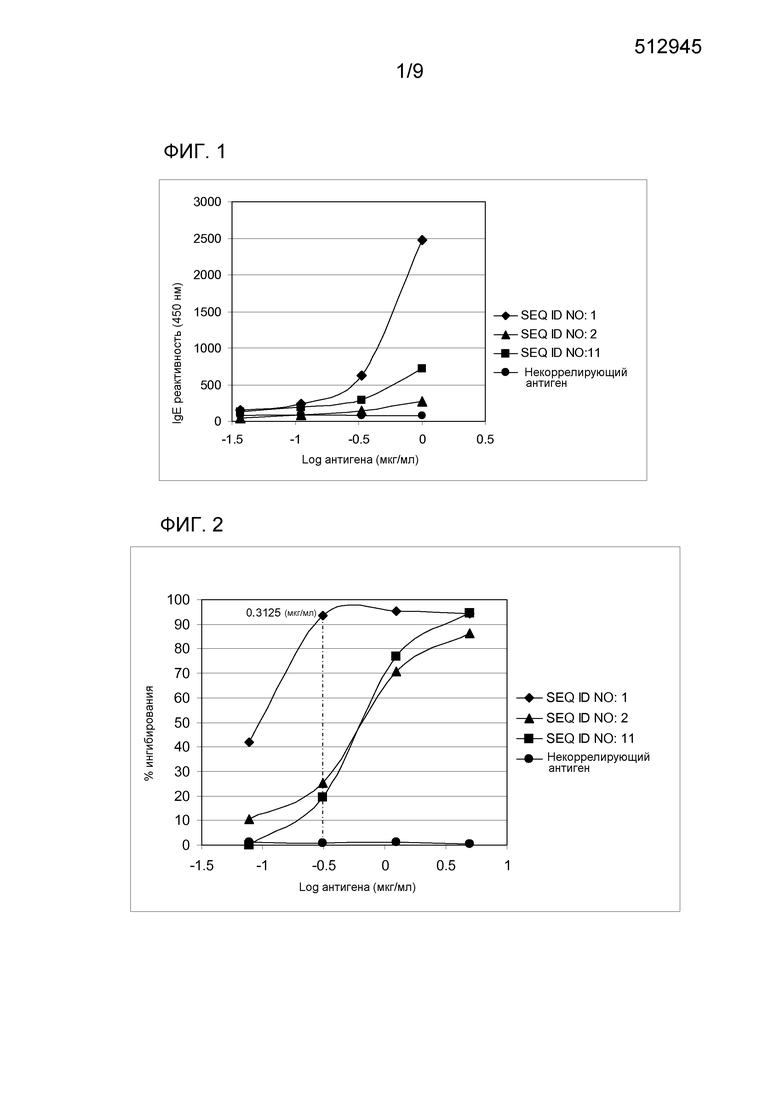
IgE реактивность варианта SEQ ID NO:2 анализировали в пуле сывороток аллергических индивидуумов методом ELISA (фиг. 1). При инкубировании с пулом сывороток пациентов, подверженных аллергии к березовой пыльце и яблоку, указанный вариант демонстрировал снижение IgE реактивности (до 1 мкг/мл) по сравнению с аллергеном Mal d 1 дт (SEQ ID NO:1) на 89% (SEQ ID NO:2). Другой пример варианта аллергена Mal d 1 с заменой Asn 78 на Ala (SEQ ID NO:11) демонстрировал снижение связывания IgE 70,9% в тех же условиях (фиг. 1).
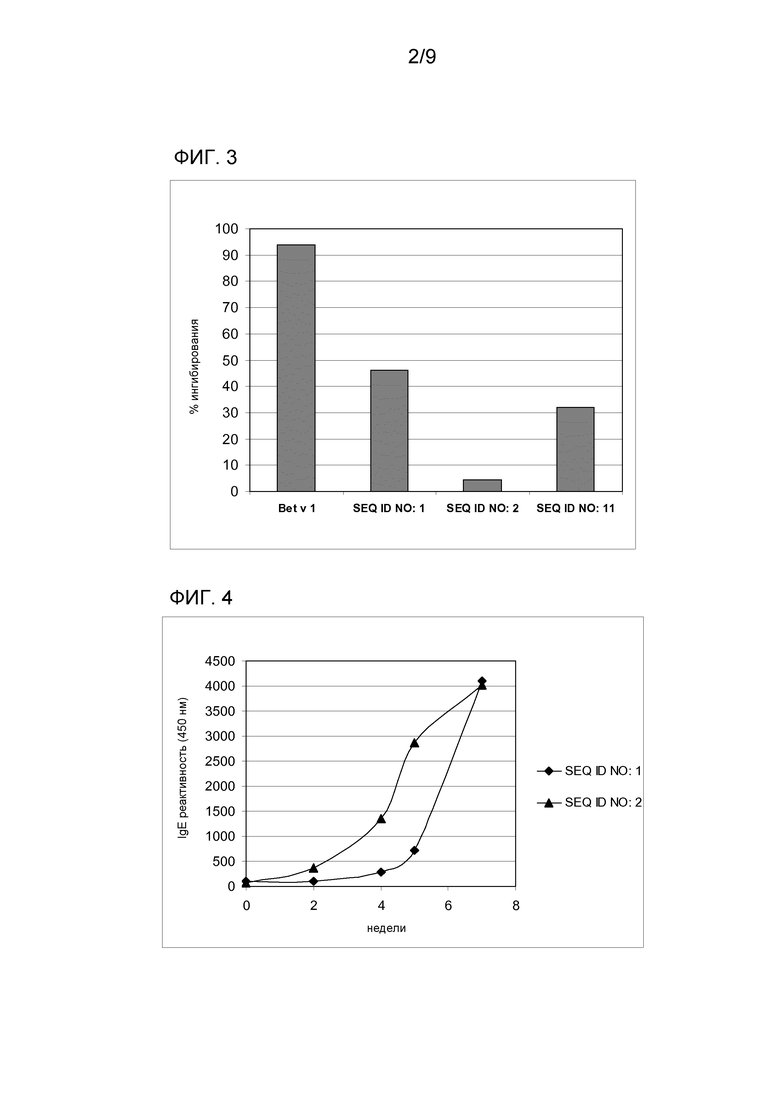
Эти результаты были подтверждены экспериментами ELISA-ингибирования, которые позволяют оценивать реактивность гомологичных эпитопов различных белков. Было обнаружено, что с 0,3125 мкг/мл ингибитора связывание белка Mal d 1 дт (SEQ ID NO: 1) с IgE из пула сывороток ингибируется на 93,3%, когда сыворотку предварительно обрабатывали тем же белком, и на 25,4%, когда ее предварительно инкубировали с вариантом SEQ ID NO: 2 (фиг. 2). В тех же условиях вариант с единственной заменой в Asn78 ингибирует связывание SEQ ID NO:1-IgE на 19,7% (фиг. 2, SEQ ID NO: 11).
Такие результаты однозначно указывают на то, что замена аминокислот в положениях 25 и/или 78 SEQ ID NO:1 препятствует распознаванию аллергена Mal d 1 IgE.
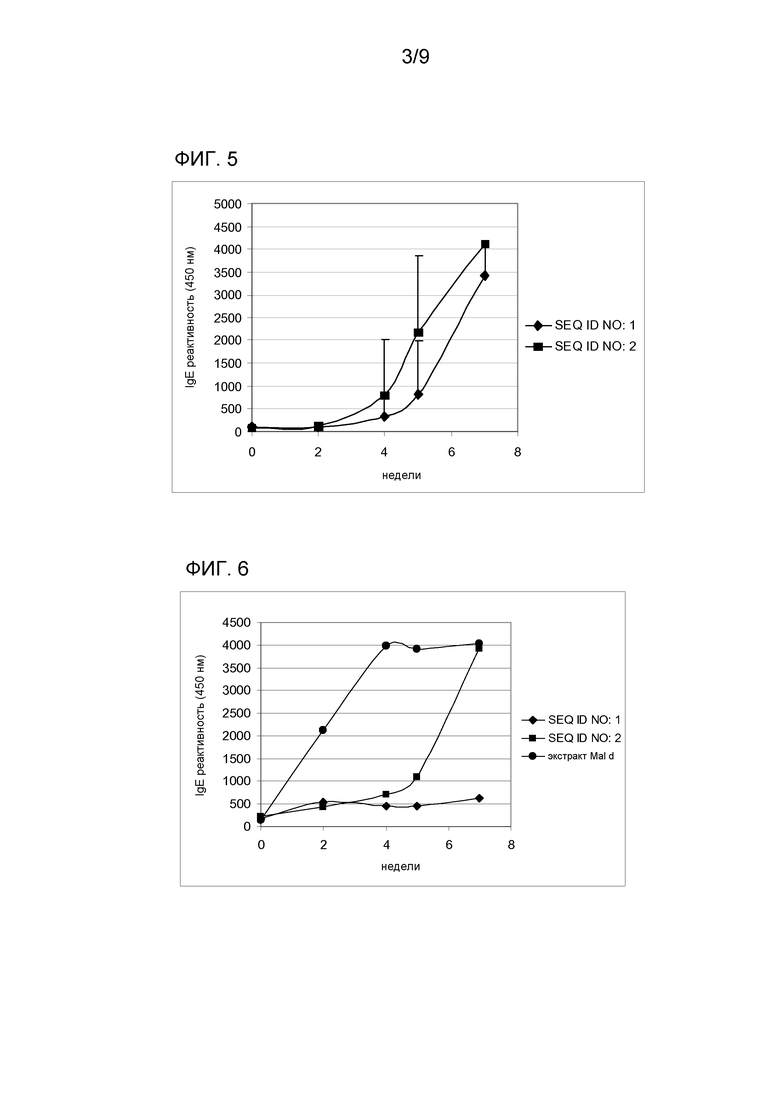
Также наблюдали то, что аллерген Mal d 1 дт SEQ ID NO:1 и гипоаллергический вариант SEQ ID NO:2, применяемые с концентрацией 0,625 мкг/мл, ингибируют связывание Bet v 1 дт с IgE из пула позитивных в отношении березы и яблока сывороток с различными степенями эффективности. Ингибирование составляет до 94%, когда сыворотку предварительно обрабатывали тем же белком Bet v 1 дт, 46% когда предварительно инкубировали с белком SEQ ID NO:1, 4,2% с SEQ ID NO:2 и 32,1%, когда сыворотку предварительно обрабатывали вариантом с единственной заменой SEQ ID NO:11 (фиг. 3).
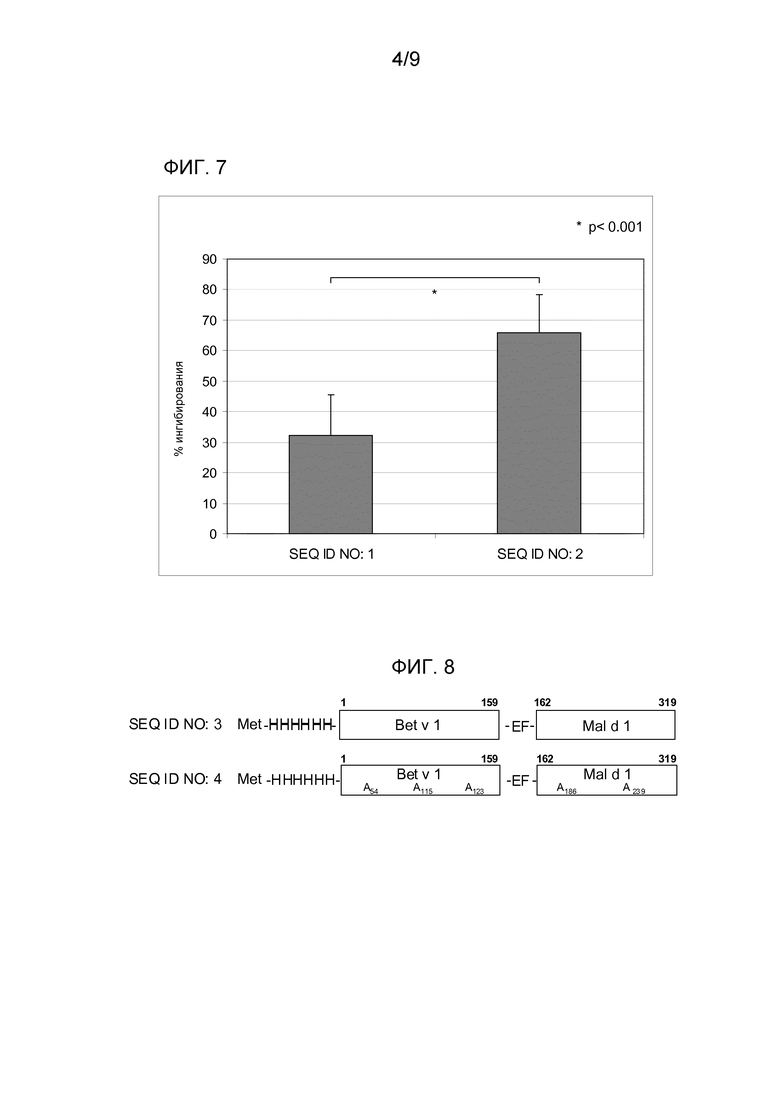
Аллерген Mal d 1 дт SEQ ID NO:1 и гипоаллергический вариант SEQ ID NO:2, применяемый для иммунизации мышей Balb/c, индуцирует специфический IgG-ответ (фиг. 4). В частности, антитела, выработанные против SEQ ID NO:2, также распознают аналог дт SEQ ID NO:1 (фиг. 5), что указывает на то, что модификация остатков в положении 25 и 78 не вызывает значимого изменения в IgG эпитопах молекулы. Более того, иммунизация с вариантом SEQ ID NO:2 индуцирует более высокий и более быстрый иммунный ответ, чем полученный с аналогом дт SEQ ID NO:1. С другой стороны, антитела, представленные в сыворотке животных, иммунизированных не имеющим отношения антигеном, не способны распознавать Mal d 1 дт и SEQ ID NO:2.
Реактивность по отношению к яблочному экстракту антител, индуцированных иммунизацией мышей SEQ ID NO:2, является детектируемой с пятой недели, и достигает пика на седьмую неделю, когда наблюдается очень слабое распознавание антителами, выработанными иммунизацией SEQ ID NO:1, хотя титр антител по отношению к Mal d 1 дт в семь недель сопоставим, с титром, полученным для мышей, иммунизированных SEQ ID NO:2 (фиг. 6).
Что касается индукции защитных антител, способных участвовать в связывании между аллергеном и IgE, наблюдалась, что антитела IgG, выработанные против SEQ ID NO:2, ингибируют связывание Mal d 1 (SEQ ID No. 1) с IgE пациентов, подверженных аллергии к березе и яблоку, более эффективно, чем против белка дт (SEQ ID NO:1) (p<0,001) (фиг. 7). Эксперименты ELISA ингибирования продемонстрировали, что IgG мышей, иммунизированных SEQ ID NO:2, ингибируют IgE реактивность сывороток семи пациентов в среднем на 66% (со значениями, варьирующимися от 49,6 до 82,2%), и IgG, выработанные против SEQ ID NO:1, на 32,3% (11,5-53%). Сыворотка неиммунизированных животных, использованная в качестве контроля, не демонстрировала роста какого-либо ингибирования специфического связывания IgE-Mal d 1.
Следующий аспект настоящего изобретения относится к иммунологически активному пептиду, соответствующему фрагменту Mal d 1, содержащему по меньшей мере одну из замен, описанных выше. Указанный пептид предпочтительно содержит от 15 до 35, и более предпочтительно от 15 до 20 аминокислотных остатков. В используемом в данном документе значении выражение "иммунологически активный" означает, что пептид должен быть способен стимулировать IgE-независимый иммунный ответ.
Другой аспект изобретения относится к гибридному белку, содержащему вариант последовательности главного аллергена Malus domectica, как описано в данном документе, и гипоаллергический вариант Bet v 1, главного аллергена пыльцы Betula verrucosa, возможно разделенные линкером.
В гибридном белке в соответствии с изобретением указанные гипоаллергические варианты Mal d 1 и Bet v 1 независимо расположены на амино- или карбокси-конце с ориентацией «голова к хвосту»; другими словами, когда амино-конец гибридного белка совпадает с амино-концом Bet v 1 или Mal d 1, карбокси-конец гибридного белка совпадает с карбокси-концом белка Mal d 1 или Bet v 1 соответственно. В соответствии с предпочтительным вариантом осуществления амино-конец гибридного белка занят Bet v 1 и карбокси-конец - Mal d 1 (фиг. 8).
Линкер, который разделяет мутированные последовательности Bet v 1 и Mal d 1, предпочтительно состоит из цепи из 8 аминокислот, более предпочтительно цепи из двух аминокислот, и даже более предпочтительно из дипептида EF (Glu-Phe).
В предпочтительном варианте осуществления изобретения гибридный белок содержит гипоаллергический вариант Bet v 1, описанный в международной заявке на патент W02007/073907 и заявке на европейский патент EP2172215, зарегистрированных тем же заявителем, и полностью включенных в данный документ посредством ссылки. В частности, гипоаллергический вариант Bet v 1, содержащийся в гибридном белке в соответствии с изобретением, получен из белковой последовательности SEQ ID NO:5 или ее изоформы, которая по меньшей мере на 94%, и предпочтительно по меньшей мере на 97% идентична с указанной последовательностью SEQ ID NO:5, с заменой остатков Lys в положении 54, 115 и/или 123 (в случае SEQ ID NO:5) или в соответствующих положениях указанной изоформы, нейтральными или полярными аминокислотами, выбранными из Ala, Thr, Gly, Pro, Leu, Ile, Phe и Ser. В предпочтительном варианте осуществления указанный гипоаллергический вариант Bet v 1 представляет собой SEQ ID NO:6. Гипоаллергические варианты Bet v 1, упоминаемые в данном документе, описаны в двух вышеприведенных заявках на патент.
Изоформы Bet v 1, имеющие более 94% идентичности последовательности с SEQ ID NO:5, содержат природные молекулы, депонированные с номерами доступа Bet V 1-a (Uniprot PI5494), Bet v 1-j (Uniprot P43184) и Bet v 1-f (Uniprot P43179).
Гибридный белок последовательности SEQ ID NO:4, в котором гипоаллергический вариант Bet v 1 (SEQ ID NO:6) связывается с гипоаллергическим вариантом Mal d 1 (SEQ ID NO:2) с ориентацией «голова к хвосту» (Bet v 1 → Mal d 1) посредством дипептидного линкера EF, является особенно предпочтительным.
Гибридный вариант в соответствии с настоящим изобретением демонстрирует снижение IgE-связывания на по меньшей мере на 10%, предпочтительно на 50% и более предпочтительно на 70% по сравнению с гибридом дикого типа.
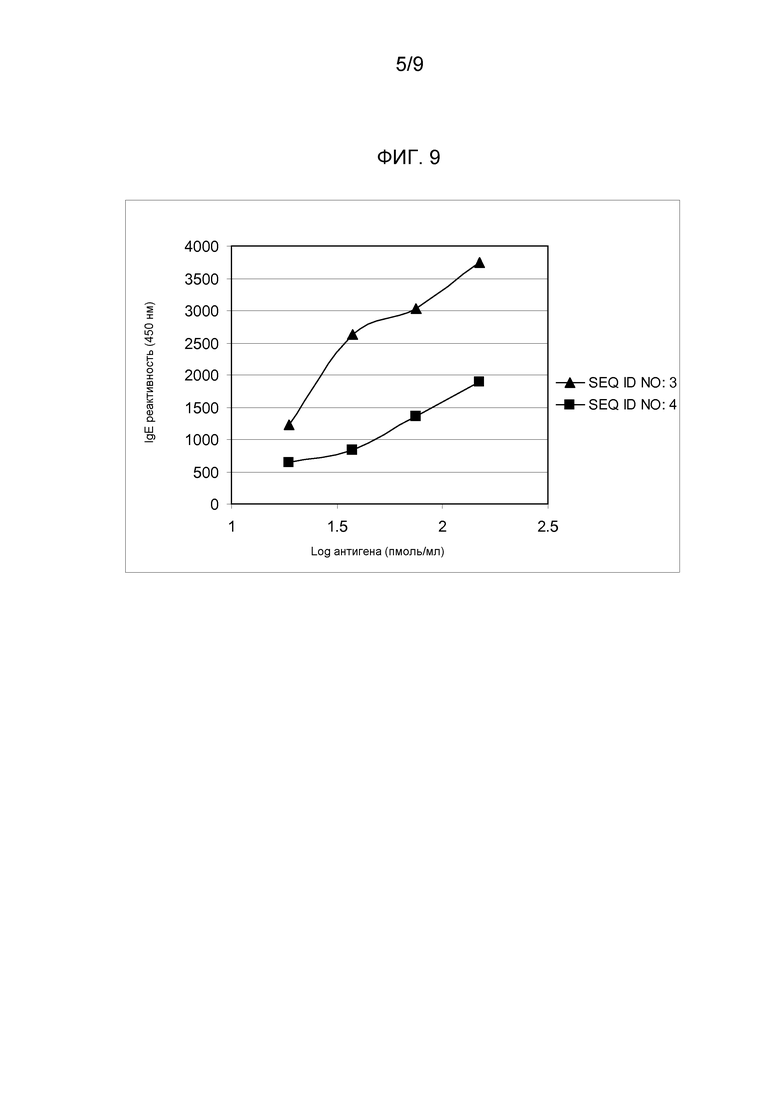
IgE реактивность варианта SEQ ID NO:4 анализировали в пуле сывороток индивидуумов, подверженных аллергии к березовой пыльце и яблоку методом ELISA (фиг. 9). При инкубировании с сыворотками указанный вариант демонстрировал среднее снижение на 55,1% IgE реактивности по сравнению с гибридом дикого типа (SEQ ID NO:3).
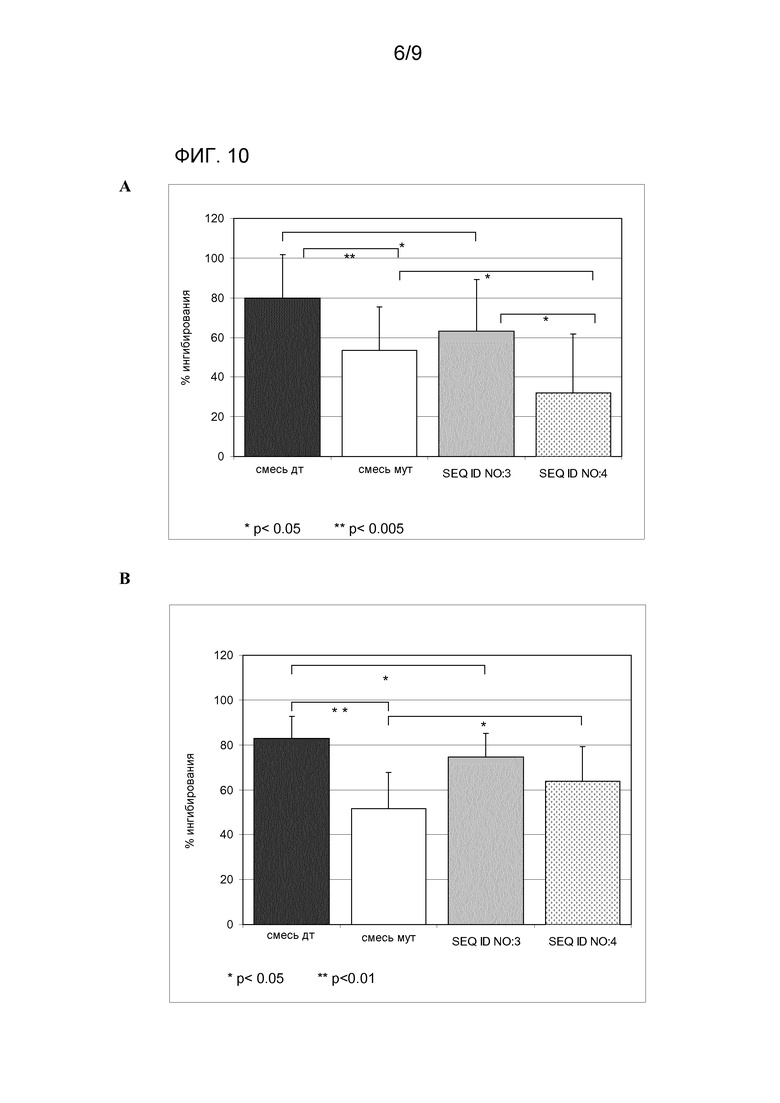
Эти результаты были подтверждены экспериментами ELISA-ингибирования. Наблюдали, что при равных концентрациях (1,25 мкг/мл) ингибитора, связывание между аллергеном Bet v 1 дт (SEQ ID NO:5), адсорбированном на лунках, и специфическим IgE, содержащемся в сыворотках 7 пациентов, ингибируется в среднем на 79,9%, когда сыворотку предварительно обрабатывали смесью отдельных аллергенов дт, на 53,3%, когда предварительно инкубировали со смесью двух мутагенизированных компонентов, на 63,3%, когда предварительно обрабатывали SEQ ID NO:3, и на 32,1%, когда сыворотку предварительно инкубировали c SEQ ID NO:4 (таблица 2 и фиг. 10A). Способность мутантного гибрида SEQ ID NO:4 участвовать в связывании Bet v 1-IgE значительно уменьшается (p<0,05) по сравнению и с SEQ ID NO:3, и со смесью двух мутагенизированных аллергенов (mut Bet v 1 - SEQ ID NO:6 и mut Mal d 1 - SEQ ID NO:2).
С таким же количеством (1,25 мкг/мл) ингибитора связывание между аллергеном Mal d 1 дт (SEQ ID NO:1), адсорбированным на лунках, и специфическим IgE, содержащимся в сыворотках 7 пациентов, ингибируется в среднем на 82,8%, когда сыворотку предварительно обрабатывали смесью одиночных аллергенов дт, на 51,6%, когда предварительно инкубировали со смесью двух мутагенизированных компонентов, на 74,6%, когда предварительно обрабатывали SEQ ID NO:3, и на 63,6%, когда сыворотку предварительно инкубировали с SEQ ID NO:4 (таблица 3 и фиг.10B). Результаты, полученные для Mal d 1, сопоставимы с результатами, измеренными для Bet v 1, за исключением SEQ ID NO:4, который оказался более реактивным против Mal d 1.
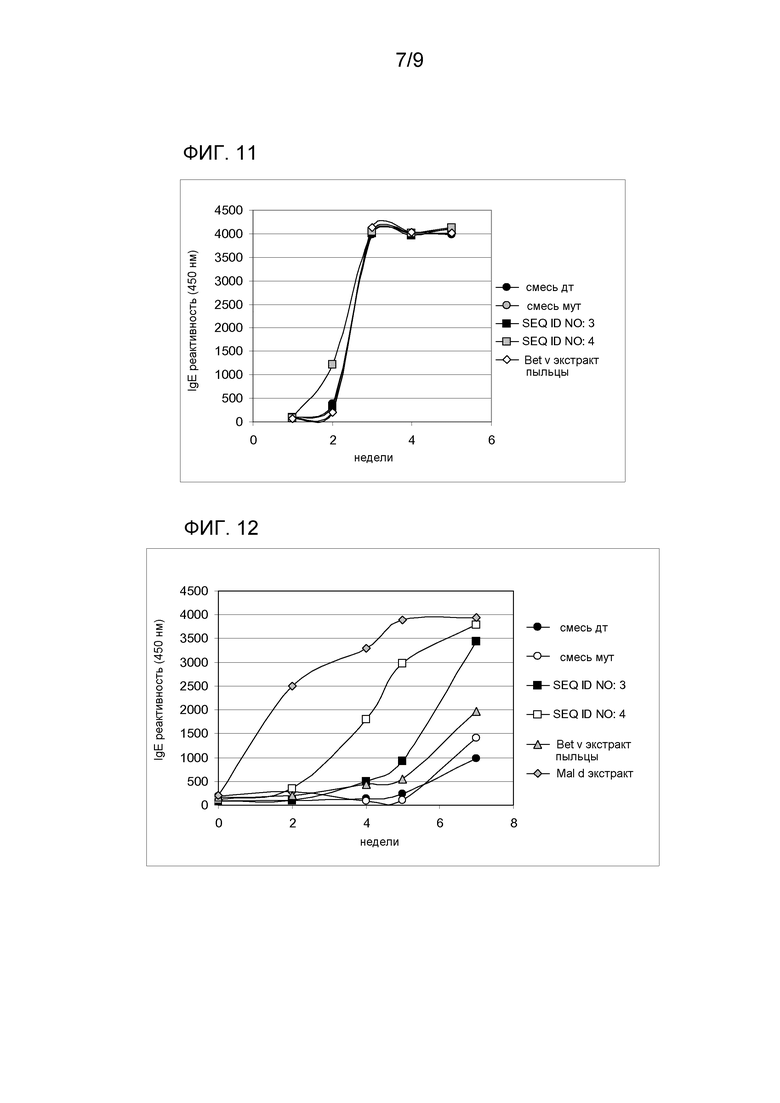
Также наблюдалось то, что гипоаллергический вариант SEQ ID NO:4, применяемый для иммунизации мышей Balb/c, индуцирует выработку специфических IgG, способных распознавать аллерген Bet v 1, представленный в экстракте Betula verrucosa (фиг. 11). Гибридная молекула SEQ ID NO:4 индуцирует специфический IgG-ответ, подобный индуцированному у мышей экстрактом березы, SEQ ID NO:3 или смесью соответствующего аллергена дикого типа или мутагенизированного аллергена. С другой стороны антитела, представленные в сыворотках животных, иммунизированных не имеющим отношения антигеном, не способны распознавать SEQ ID NO:3 и 4 и смеси аллергена дикого типа и мутантного аллергена.
Гипоаллергический гибридный вариант SEQ ID NO:4 также способен индуцировать специфический IgG-ответ на компоненты экстракта мякоти Malus domestica(фиг. 12). Через пять недель после первой иммунизации IgG-ответ, индуцированный иммунизацией с SEQ ID NO:4, в 30 раз больше, чем ответ полученный иммунизацией эквимолярными количествами смеси двух одиночных мутагенизированных вариантов (мут. смесь), и в 3,2 раз больше, чем гибридом аллергена дикого типа SEQ ID NO:3. Иммунизация экстрактом березы индуцирует выработку IgG, способных распознавать Mal d 1, содержащийся в экстракте яблока начиная с пятой недели.
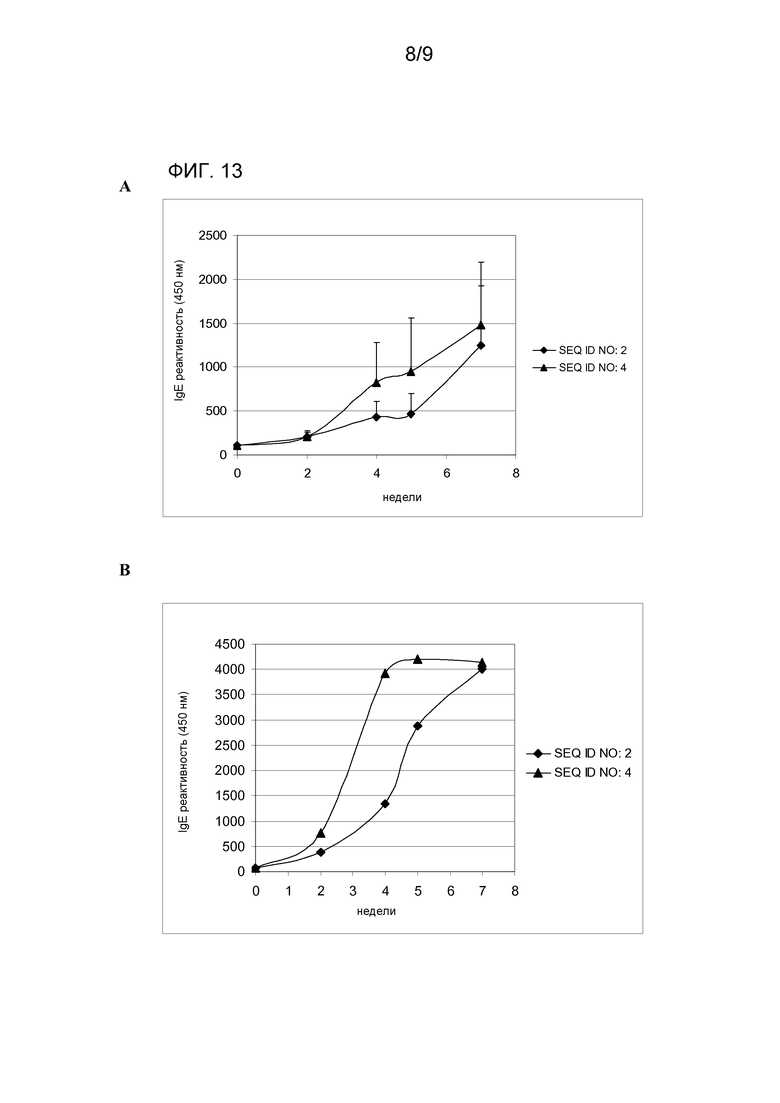
Кроме того иммуногенность гипоаллергического варианта SEQ ID NO:2 увеличивается, когда он является частью гибридной гипоаллергической молекулы SEQ ID NO:4. Индукция антител IgG, специфических к Mal d 1 дт (SEQ ID NO:1) и мутантному Mal d 1 (SEQ ID NO:2), является более высокой и более ранней у мышей, иммунизированных гибридным вариантом SEQ ID NO:4, чем у мышей, иммунизированных одиночным аллергеном SEQ ID NO:2. Существенное увеличение индукции SEQ ID NO:4 наблюдается уже на четвертой неделе лечения, тогда как проходит по меньшей мере семь недель для получения такого же ответа иммунизацией животных белком SEQ ID NO:2 (фиг. 13-A,B).
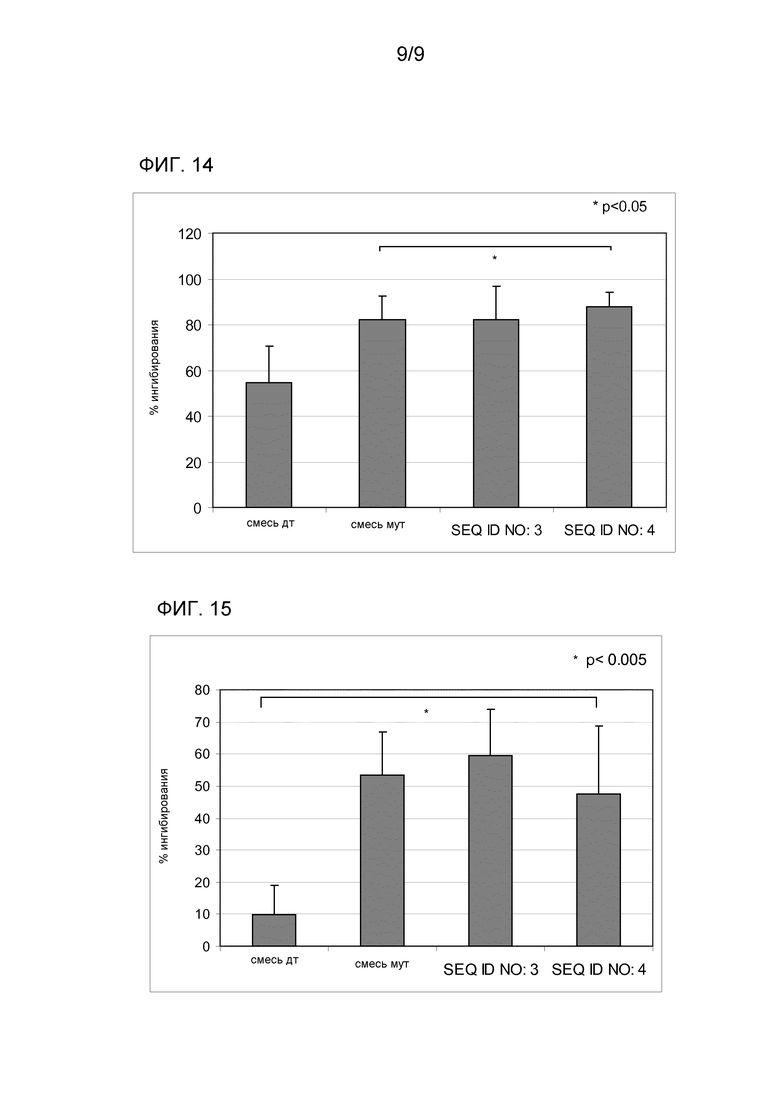
Что касается индукции защитных антител, способных ингибировать связывание между аллергеном и IgE, наблюдали то, что антитела IgG, выработанные против SEQ ID NO:4, ингибируют связывание Bet v 1 (SEQ ID NO:5) с IgE пациентов, подверженных аллергии к березе и яблоку более эффективно, чем антитела, индуцированные смесью двух одиночных мутированных вариантов (мут. смесь) (p<0,05) (фиг. 14). Эксперименты ELISA ингибирования продемонстрировали, что IgG, выработанные у мышей, иммунизированных SEQ ID NO:4, ингибируют IgE реактивность сыворотки семи пациентов в среднем на 87,7% (со значениями, варьирующимися от 77,4 до 94,7%), и IgG, выработанные против смеси двух мутагенизированных вариантов (мут. смесь), на 82% (63,5-92,3%); антитела IgG, выработанные против смеси белки дикого типа (смесь дикого типа), ингибируют связывание в среднем на 54,7% (32,4-68,6%), и антитела, полученные иммунизацией гибридом дикого типа SEQ ID NO:3, на 82,2% (51,9-93,5%).
Антитела IgG, индуцированные в мышах, иммунизированных SEQ ID NO:4, также ингибируют связывание между Mal d 1 (SEQ ID NO:1) и IgE из сывороток пациентов, подверженных аллергии к березе и яблоку, в среднем на 47,5%, на 9,9%, если они иммунизированы со смесью белков дикого типа, на 53,5% со смесью мутированных вариантов, и на 59,4% с иммуногеном SEQ ID NO:3 (фиг. 15).
Сыворотка неиммунизированных животных, применяемая в качестве контроля, не вызывает какое-либо ингибирования специфического связывания IgE с Bet v 1 и Mal d 1.
Варианты с заменами в соответствии с изобретением могут быть легко приготовлены мутагенезом последовательности кДНК Mal d 1 (SEQ ID NO:7), Bet v 1 (SEQ ID NO:8), их изоформ или природных вариантов или последовательности кДНК гибрида дикого типа (SEQ ID NO:9) с применением технологий, известных специалисту в данной области (21).
SEQ ID NO:10 и 19 представляют последовательности кДНК, кодирующие (мономерный) вариант с двумя заменами или гибридный вариант, определяемые SEQ ID NO:2 и 4 соответственно.
Дополнительные аспекты изобретения вследствие этого относятся к молекуле нуклеиновой кислоты, кодирующей вариант аллергена Mal d 1, описанный в данном документе, для получения пептида из нее или гибридного белка Mal d 1-Bet v 1, и экспрессионному вектору, содержащему указанные молекулы вместе с элементами для контроля экспрессии в эукариотических или прокариотических клетках, таких как транскрипционные промоторы или энхансеры, сигнальные последовательности или другие регулирующие транскрипцию последовательности. Вектор может являться плазмидой, вирусом, фагом или любым другим вектором, широко используемым в генетической инженерии.
Изобретение также содержит прокариотическую или эукариотическую клетку хозяина, трансформированную или трансфицированную вектором в соответствии с изобретением. Прокариотические клетки, такие как Escherichia coli и Bacillus subtilis, или эукариотические клетки, такие как Saccharomyces cerevisiae, обычно применяются для клонирования вектора и экспрессии кДНК.
Гипоаллергические варианты в соответствии с изобретением могут также быть получены в виде слитых белков.
В связи с их уменьшенной IgE реактивностью, Mal d 1 варианты в соответствии с настоящим изобретением могут быть удобно применены для приготовления фармацевтических композиций (напр., таблетки) для применения в иммунотерапии пациентов, подверженных аллергии к яблоку и/или пыльце Betula verrucosa.
Следующий аспект изобретения вследствие этого относится к фармацевтической композиции, содержащей эффективное количество гипоаллергического варианта Mal d 1, при желании в комбинации с другими аллергенами Betula verrucosa, вместе с фармацевтически приемлемыми носителями, наполнителями или адъювантами. В предпочтительном варианте осуществления указанная фармацевтическая композиция представлена в виде подходящей вакцины для профилактического или терапевтического лечения аллергических заболеваний, таких как бронхиальная астма и ринит, конъюнктивит и синдром оральной аллергии. Подъязычные, подкожные и чрескожные формы введения являются наиболее предпочтительными.
Принципы вакцинации и способы известны специалисту и описаны, например, в (22).
Примеры далее иллюстрируют изобретение более подробно.
ПРИМЕРЫ
Если не указано иное, способы, использованные в следующих примерах, описаны в Sambrook, Fritsch ET Maniatis "Molecular Cloning: A Laboratory Manual" II Ed. Vol. 1-2-3 CSH Lab Press 1989.
ПРИМЕР 1 - Сайт-специфический мутагенез кДНК, кодирующей аллерген Mal d 1
Сайт-специфический мутагенез кДНК, кодирующей аллерген Mal d 1(SEQ ID NO:7, с предшествующей на 5' последовательностью, кодирующей шесть гистидинов), осуществляли клонированием кДНК в прокариотическом векторе (pBluescript, GenBank acc. n. X52327) с последующей амплификацией ПЦР. Олигонуклеотиды, используемые в качестве праймеров в ПЦР (таблица 1), несли соответствующие замены оснований. Для каждого мутагенеза использовали комплементарное олигонуклеотидное связывание с соответствующей областью нити ДНК (21). После амплификации неизмененную исходную матрицу селективно разрушали ферментативным расщеплением, катализируемым ферментом рестрикции Dpn I. Затем клетки Escherichia coli трансформировали мутагенезированными молекулами. Клоны, полученные из отдельных бактериальных колоний, секвенировали в соответствии с методом Сэнгера для определения корректной модификации оснований и отсутствия неспецифических мутации в кДНК.
Последовательности олигонуклеотидов, используемые в
качестве праймеров в сайт-специфическом мутагенезе.
Мутированные основания выделены жирным шрифтом
ПРИМЕР 2 - Конструирование плазмиды, кодирующей гибридную молекулу Bet v 1 дт-Mal d 1 (wtHybrid)
Гибридная молекула, содержащая генетическую информацию для гибрида Bet v 1 дт-Mal d 1, было получена слиянием кДНК, кодирующих одиночные аллергены.
кДНК, кодирующие зрелые белки Bet v 1 и Mal d 1, получали отдельно ПЦР с применением Bet v 1 DIM FW (ggt gtt ttc aat tac gaa act g - SEQ ID NO:14) и Bet v 1 DIM Eco RV (Gc gaa ttc gtt gta ggc ate gga g - SEQ ID NO:15), олигонуклеотидные праймеры для Bet v 1, и Mal d 1 DIM Eco FW (cgc gaa ttc ggt gtc tac аса ttt gag aac g - SEQ ID NO:16) и Mal d 1 DIM Bam RV (Gcg gga tcc tta gtt gta tgc gtc ggg gtg - SEQ ID NO:17), олигонуклеотидные праймеры для Mal d 1. Клоны Bet v 1 SEQ ID NO:8 и Mal d 1 SEQ ID NO:7 применяли в качестве матриц.
Продукт амплификации, полученный из Bet v 1, реамплифицировали замещением праймера Bet v 1 DIM FW на Bet v 1 DIM Kpn FW (gcg ggt acc cat atg cat cac cat cac cat cac ggt gtt ttc aat tac gaa act g - SEQ ID NO:18), посредством чего вводили последовательность, кодирующую шесть гистидинов перед последовательностью Bet v 1. Сайт Kpn I и сайт Nde I (содержащий ATG) вводили на 5' продукта амплификации Bet v 1, и сайт Eco R I вводили на 3' вместо стоп-кодона. Сайт Eco R I вводили вместо ATG на 5' Mal d 1, и сайт Bam H I вводили на его 3' после стоп-кодона. Амплифицированные продукты очищали, расщепляли рестрикционными ферментами Kpn I и Eco R I (Bet v 1) или Eco R I и Bam H I (Mal d 1) (сайты рестрикции в праймерах подчеркнуты), и затем вводили в Kpn I/Bam H I сайты вектора pEt 3c (Stratagene, La Jolla, CA) для получения конструкции, способной экспрессировать слитый белок Bet v 1-Mal d 1 с предшествующей последовательностью из шести гистидинов. Введение сайта рестрикции Eco R I, который необходим для клонирования фрагментов, позволяло встраивание двух аминокислот (глутаминовая кислота и фенилаланин) в участок соединения двух белков без изменения рамки считывания (фиг. 8).
Клоны, полученные из одиночных бактериальных колоний, секвенировали по методу Сэнгера для подтверждения того, что изменение оснований было корректным и отсутствовали неспецифические мутации в кДНК.
ПРИМЕР 3 - Конструирование плазмиды, кодирующей мутантную гибридную молекулу Bet v 1-Mal d 1 (MutHybrid)
Гибридная молекула, кодирующая мутантный гибрид Bet v 1-Mal d 1, была получена по способу, описанному в примере 2 для гибридного варианта дикого типа.
Олигонуклеотидные пары, используемые в ПЦР, были идентичными, при этом кДНК, используемые в качестве матриц, кодировали два гипоаллергических варианта, последовательности которых в данном документе определяются SEQ ID NO:20 (для мутанта Bet v 1) и SEQ ID NO:10 (мутанта Mal d 1).
Клоны, полученные из одиночных бактериальных колоний, секвенировали по методу Сэнгера для подтверждения того, что изменение оснований было корректным и отсутствовали неспецифические мутации в кДНК.
ПРИМЕР 4 - Выработка белков Mal d 1 и Bet v 1, соответствующих мутантам, wtHybrid и MutHybrid
кДНК Bet v 1 дт (SEQ ID NO:8) и Mal d 1 (SEQ ID NO:7), мутагенезированные кДНК (SEQ ID NO:20 и SEQ ID NO:10), и сконструированные кДНК дикого типа и гибрида Mut (SEQ ID NO:9 и 19), с предшествующей последовательностью, кодирующей шесть гистидинов, клонировали в экспрессионный вектор и эспрессировали в клетках Escherichia coli в соответствии со стандартными протоколами (23). Клетки собирали центрифугированием и ресуспендировали в 100 мМ буфере NaH2PO4, pH 8 и лизировали озвучиванием. Рекомбинантные белки разделяли центрифугированием. Осадок, содержащий нерастворимый белковый агрегат, ресуспендировали в денатурирующем буфере (мочевина 6 M, NaH2PO4 100 мМ,Tris 10 мМ pH 8) и перемешивали в течение 60 мин при 20°C. Солюбилизированные рекомбинантные белки отделяли от нерастворимого дебриса центрифугированием и очищали от супернатанта афинной хроматографией с применением колонок с агарозой, к которой была присоединена нитрилотриуксусная кислота хелатирующая ионы никеля, которые взаимодействуют с участком из шести гистидинов, слитым с аллергеном. Очищенные белки рефолдировали диализом в течение 18 часов при 4°C в 5 мМ растворе (NH4)HCO3.
ПРИМЕР 5 - Свойства сывороток аллергических пациентов
Сыворотки собирали у пациентов с клинической историей сезонной аллергии к пыльце Betula verrucosa, и RAST 3+ реактивностью, специфической к аллергенам Bet v 1 и Mal d, и использовали в одиночной или объединенной форме. Пул сывороток неподверженных аллергии пациентов использовали в качестве контроля.
ПРИМЕР 6 - ELISA анализ реактивности вариантов Mal d 1 к IgE из пула сывороток
Серийно разбавленные в три раза аликвоты в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, аллергена дикого типа (SEQ ID NO:1) и мутагенезированных вариантов (SEQ ID NO:2, SEQ ID NO:11) адсорбировали на лунках полистирольных планшетов для метода ELISA инкубированием при 4°C в течение 16 часов. Лунки промывали промывочным раствором (60 мМ фосфатный буфер, pH 6,5, содержащим 0,05% Твин-20) и блокировали разбавляющим раствором (25% козья сыворотка, 1 мМ ЭДТК, 0,05% Твин 20, 0,01% Тиомерсал в 150 мМ фосфатном буфере, pH 7,4). Аликвоты по 60 мкл в разбавляющем буфере пула человеческих сывороток RAST 3+ или неподверженных аллергии пациентов добавляли в каждый образец и инкубировали при 25°C в течение 2 часов. После трех промывок добавляли коньюгированную с пероксидазой сыворотку против IgE человека (1:4000 в разбавляющем буфере) с последующей инкубацией при 25°C в течение 1,5 часа. После трех промываний проводили колориметрическую реакцию добавлением 100 мкл реагента TMB (BioFX Laboratories, Owings Mills, MD) и инкубированием в течение 15 минут при 25°C. Реакцию останавливали добавлением 100 мкл 1 Н HCl и считывали при 450 нм с применением микропланшет-ридер спектрофотометра. Результаты были подтверждены тремя независимыми экспериментами. Такой же протокол применяли с некоторыми модификациями для тестирования IgE реактивности сконструированных гибридов дт и Mut.
Серийное разведение гибридных аллергенов (SEQ ID NO:3 и 4) готовили в соотношении 1:2 в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, начиная от 150 нм и адсорбировали на лунках полистирольных планшетов. Пул сывороток, позитивных к Bet v 1 и Mal d 1, или неподверженных аллергии пациентов в качестве отрицательного контроля разбавляли 1:2,5 в разбавляющем растворе, добавляли (70 мкл) в каждую лунку и инкубировали при 25°C в течение 3 часов.
ПРИМЕР 7 - ELISA исследование ингибирования - Мономерные варианты Mal d 1 ингибируют связывание Mal d 1 с IgE в сыворотке
Равные количества (0,1 мкг) Mal d 1 дт в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, адсорбировали на лунки полистирольных планшетов для метода ELISA инкубацией при 4°C в течение 16 часов. Лунки затем промывали промывочным раствором (60 мМ фосфатный буфер, pH 6,5, содержащий 0,05% Твин-20), и свободные сайты блокировали разбавляющим раствором (15% козья сыворотка, 1 мМ ЭДТК, 0,05% Твин-20, в 150 мМ фосфатном буфере, pH 7,4). Аликвоты (100 мкл) разбавления 1:3 объединенных человеческих сывороток, позитивных к Bet v 1 и Mal d 1, предварительно инкубировали с четырехкратными серийными разведениями дт или мутагенезированных аллергенов, начиная от 5 мкг/мл при 25°C в течение 2 часов. Смеси затем добавляли в каждую лунку и инкубировали при 4°C в течение 16 часов. После трех промываний буфером (0,06 M фосфат, pH 6,5, 0,05% Твин-20) добавляли сыворотку антител против человеческих IgE, коньюгированных с пероксидазой, разбавляли 1:4000 в разбавляющем буфере, и инкубировали при 25°C в течение 1,5 часа. После трех промываний, осуществляли колориметрическую реакцию добавлением 100 мкл реагента ТМБ (BioFX Laboratories, Owings Mills, MD), и инкубацией в течение 15 минут при 25°C. Реакцию останавливали добавлением 100 мкл 1Н HCl и оценивали считыванием при 450 нм спектрофотометром. Процент ингибирования рассчитывали с применением следующей формулы: 100 x [(A-B)/A], где A является абсорбцией при 450 нм в отсутствии ингибитора, и В является абсорбцией при наличии ингибитора. Данные являются характеризующими три независимых эксперимента.
ПРИМЕР 8 - ELISA исследование ингибирования - Мономерные варианты Mal d 1 ингибируют связывание Bet v 1 с IgE в сыворотке
Равные количества (0,1 мкг) Bet v 1 дт в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, адсорбировали на лунки полистирольных планшетов для метода ELISA инкубацией при 4°C в течение 16 часов. Лунки затем промывали промывочным раствором (60 мМ фосфатный буфер, pH 6,5, содержащий 0,05% Твин-20), и свободные сайты блокировали разбавляющим раствором (15% козья сыворотка, 1 мМ ЭДТК, 0,05% Твин-20, в 150 мМ фосфатном буфере, pH 7,4). Аликвоты (100 мкл) разбавления 1:3 объединенных человеческих сывороток, позитивных к Bet v 1 и Mal d 1, предварительно инкубировали с серийными четырехкратными разведениями Bet v 1 дт, Mal d 1 дт или мутагенезированных вариантов, начиная от 10 мкг/мл при 25°C в течение 2 часов. Смеси затем добавляли в каждую лунку и инкубировали при 4°C в течение 16 часов. После трех промываний буфером: 0,06 M фосфат, pH 6,5, 0,05% Твин-20, добавляли сыворотку атител против человеческих IgE, коньюгированных с пероксидазой, разбавленную 1:4000 в разбавляющем буфере, и инкубировали при 25°C в течение 1,5 часа. После трех промываний проводили колориметрическую реакцию добавлением 100 мкл реагента ТМБ (BioFX Laboratories, Owings Mills, MD) и инкубацией в течение 15 минут при 25°C. Реакцию останавливали добавлением 100 мкл, 1Н N HCl, и оценивали считыванием при 450 нм спектрофотометром. Процент ингибирования рассчитывали с применением следующей формулы: 100 x [(A-B)/A], где A является абсорбцией при 450 нм в отсутствии ингибитора, и В является абсорбцией при наличии ингибитора. Значения были подтверждены тремя независимыми экспериментами.
ПРИМЕР 9 - ELISA исследование ингибирования - SEQ ID NO:3 и SEQ ID NO:4 ингибируют связывание Bet v 1 или Mal d 1 с IgE в сыворотке
Равные количества (0,1 мкг) Bet v 1 дт или Mal d 1 дт в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, адсорбировали на лунки полистирольных планшетов для метода ELISA инкубацией при 4°C в течение 16 часов. Лунки затем промывали промывочным раствором (60 мМ фосфатный буфер, pH 6,5, содержащий 0,05% Твин-20) и свободные сайты блокировали разбавляющим раствором (15% козья сыворотка, 1 мМ ЭДТК, 0,05% Твин-20 в 150 мМ фосфатном буфере, pH 7,4). Аликвоты (70 мкл) разбавления 1:3 пула человеческих сывороток, позитивных к Bet v 1 и Mal d 1, предварительно инкубировали с равными количествами (1,25 мкг/мл) аллергена дикого типа, мутагенезированных или сконструированных вариантов (гибридов) при 25°C в течение 2 часов. Смеси затем добавляли в каждую лунку и инкубировали при 4°C в течение 16 часов. После трех промываний буфером (0,06 M фосфат, pH 6,5, 0,05% Твин-20) добавляли сыворотку антител против IgE человека, коньюгированных с пероксидазой, разбавленную 1:4000 в разбавляющем буфере и инкубировали при 25°C в течение 1,5 часа. После трех промываний проводили колориметрическую реакцию добавлением 100 мкл реагента ТМБ (BioFX Laboratories, Owings Mills, MD) и инкубацией в течение 15 минут при 25°C. Реакцию останавливали добавлением 100 мкл 1Н HCl, и оценивали считыванием при 450 нм спектрофотометром. Процент ингибирования рассчитывали с применением следующей формулы: 100 x [(A-B)/A], где A является абсорбцией при 450 нм в отсутствии ингибитора, и В является абсорбцией при наличии ингибитора.
Гибрид дикого типа и мутагенезированные молекулы ингибируют Bet v 1 - связывание IgE
Гибрид дикого типа и мутагенезированные молекулы ингибируют связывание Mal d 1 - IgE
ПРИМЕР 10 - Иммунизация мышей Balb/c
Десять групп мышей, каждая из которых состояла из пяти самок линии Balb/c (Charles River), подкожно иммунизировали 150 пмоль дикого типа, мутагенезированного или синтезированного (гибридного) аллергена или 10 мкг экстракта пыльцы Betula verrucosa или Malus domestica, смешанного с 2 мг Al(OH3) в 200 мкл солевого раствора. Другие две инъекции выполняли после 21 и 42 дней. В качестве контроля пять мышей получали то же лечение не имеющим отношения антигеном (данные не приведены). Через две, четыре, пять и семь недель после первой иммунизации осуществляли отбор крови из яремной вены мыши, и проверяли гуморальный иммунный ответ на соответствующий иммуноген посредством ELISA. У мышей, иммунизированных SEQ ID NO:2 и 4, также анализировали способность распознавать белок дт или экстракты пыльцы Betula verrucosa и Malus domectica. Сыворотки мышей объединяли на основании иммуногенетического типа и времени, прошедшего после первой иммунизации.
ПРИМЕР 11 - Анализ специфического IgG ответа у иммунизированных мышей методом ELISA
Равные количества экстракта пыльцы Betula verrucosa и Malus domectica (20 мкг/мл) или Bet v 1 дт или Mal d 1 или SEQ ID NO:2 (2 мкг/мл), в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, адсорбировали на лунки полистирольных планшетов для метода ELISA инкубацией при 4°C в течение 16 часов. Лунки затем промывали промывочным раствором (60 мМ фосфатный буфер, pH 6,5, содержащий 0,05% Твин-20), и свободные сайты блокировали разбавляющим раствором (15% козья сыворотка, 1 мМ ЭДТК, 0,05% Твин-20 в 150 мМ фосфатном буфере, pH 7,4). Равные аликвоты (100 мкл) каждой мышиной сыворотки или объединенные сыворотки добавляли в каждую лунку при разбавлении 1:1000 в разбавляющем буфере, и инкубировали при 25°C в течение 2 часов. После трех промываний добавляли сыворотку антител против тотальных мышиных IgG, коньюгированных с пероксидазой, разбавленную 1:2000 в разбавляющем буфере, и инкубировали при 25°C в течение 1,5 часа. После трех промываний осуществляли колориметрическую реакцию добавлением 100 мкл реагента ТМБ (BioFX Laboratories, Owings Mills, MD), и инкубацией в течение 20 минут при 25°C. Реакцию останавливали добавлением 100 мкл 1Н HCL и считывали при 450 нм спектрофотометром. Данные демонстрируют среднюю реактивность, полученную анализом сывороток мыши для каждой группы.
ПРИМЕР 12 - ELISA исследование ингибирования. IgG против SEP ID NO:2 и SEQ ID NO:4 ингибируют связывание между Bet v 1 или Mal d 1 дт и IgE в сыворотках аллергических пациентов, позитивных к Bet v 1 и Mal d 1
Равные количества Bet v 1 (0,1 мкг) или Mal d 1 (0,2 мкг) в 50 мМ карбонатно/бикарбонатном буфере, pH 9,6, адсорбировали на лунки полистирольных планшетов для метода ELISA инкубацией при 4°C в течение 16 часов. Лунки затем промывали промывочным раствором (60 мМ фосфатный буфер, pH 6,5, содержащий 0,05% Твин-20) и свободные сайты блокировали разбавляющим раствором (15% козья сыворотка, 1 мМ ЭДТК, 0,05% Твин-20 в 150 мМ фосфатном буфере, pH 7,4). Аликвоты (100 мкл) разбавленных 1:10 пулов мышиных сывороток, собранных после семи недель после первой иммунизации, инкубировали при 4°C в течение 16 часов. После трех промываний 0,06 M фосфатным буфером, pH 6,5, 0,05% Твин-20 добавляли семь разбавленных 1:3 человеческих сывороток, позитивных к Bet v 1 и Mal d 1, при 25°C в течение 3 часов. После трех промываний 0,06 M фосфатным буфером, pH 6,5, 0,05% Твин-20, добавляли сыворотку антител против человеческих IgE, коньюгированных с пероксидазой, разбавленную 1:4000 в разбавляющем буфере, и инкубировали при 25°C в течение 1,5 часа. После трех промываний проводили колориметрическую реакцию добавлением 100 мкл реагента ТМБ (BioFX Laboratories, Owings Mills, MD) и инкубацией в течение 20 минут при 25°C. Реакцию останавливали добавлением 100 мкл 1Н HCL и считывали при 450 нм спектрофотометром. Процент ингибирования рассчитывали с применением следующей формулы: 100 x [(A-B)/A], где A является абсорбцией при 450 нм в отсутствии ингибирующей человеческой сыворотки, и В является абсорбцией при наличии ингибирующей человеческой сыворотки.
ПРИМЕР 13 - Статистический анализ
На фигурах результаты представлены в виде средних значений плюс соответствующие стандартные отклонения.
Для статистических анализов применяли UNISTAT 5.5 Light для программного обеспечения Excel. Данные анализировали посредством парного t-критерия.
Источники информации




| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный полипептид на основе аллергена пыльцы березы и аллергена яблока в качестве вакцины от аллергии | 2020 |
|
RU2761431C1 |
| ГИБРИДНЫЕ БЕЛКИ-НОСИТЕЛИ ПЕПТИДОВ В КАЧЕСТВЕ ВАКЦИН ОТ АЛЛЕРГИИ | 2012 |
|
RU2630652C2 |
| НОВЫЕ МУТАНТНЫЕ АЛЛЕРГЕНЫ | 2001 |
|
RU2285042C2 |
| ПРОИЗВОДНЫЕ PhI p 5а, ОБЛАДАЮЩИЕ СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ И СОХРАНЕННОЙ Т-КЛЕТОЧНОЙ РЕАКТИВНОСТЬЮ | 2004 |
|
RU2368620C2 |
| ВАРИАНТ АЛЛЕРГЕНА ГРУППЫ I ИЗ Роасеае, ХАРАКТЕРИЗУЮЩИЙСЯ СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ И СОХРАНЕННОЙ Т-КЛЕТОЧНОЙ РЕАКТИВНОСТЬЮ (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА ДНК И ЕГО ПРИМЕНЕНИЕ | 2005 |
|
RU2409589C2 |
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2658767C1 |
| МОДИФИКАЦИИ АЛЛЕРГЕНОВ ГРУППЫ 6 POACEAE (МЯТЛИКОВЫХ), ИМЕЮЩИХ ПОНИЖЕННУЮ АЛЛЕРГЕННОСТЬ БЛАГОДАРЯ МУТАГЕНЕЗУ ПРОЛИНОВЫХ ОСТАТКОВ | 2010 |
|
RU2607373C2 |
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2575606C2 |
| НОВЫЕ АЛЛЕРГЕНЫ ПШЕНИЦЫ | 2008 |
|
RU2502742C2 |
| АЛЛЕРГЕНЫ, ПОЛУЧЕННЫЕ РЕКОМБИНАНТНЫМИ СПОСОБАМИ | 2010 |
|
RU2592674C2 |











Изобретение относится к области биотехнологии, конкретно к гипоаллергенным вариантам главного аллергена Malus domectica, и может быть использовано в медицине для лечения аллергических заболеваний. Получают белок - гипоаллергенный мутант главного аллергена Malus domectica Mal d 1 дт, имеющего SEQ ID NO: 1, для применения в иммунотерапии пациентов, подверженных аллергии к пыльце Malus domectica и/или Betula verrucosa. Полученный белок имеет аминокислотную последовательность, в которой при выравнивании с SEQ ID NO: 1 присутствуют либо две замены, включающие замену остатка Asp в положении 25 и замену остатка Asn в положении 78, либо одна замена остатка Asn в положении 78 последовательности SEQ ID NO: 1, где указанные остатки Asp в положении 25 и/или Asn в положении 78 заменены Ala остатком. Изобретение позволяет получить гипоаллергенный мутант главного аллергена Malus domectica Mal d 1, который демонстрирует уменьшенную IgE реактивность по сравнению с указанным аллергеном Mal d 1 с SEQ ID NO: 1. 8 н. и 6 з.п. ф-лы, 15 ил., 3 табл., 13 пр.
1. Белок, который представляет собой гипоаллергенный мутант главного аллергена Malus domectica Mal d 1 дт, имеющего SEQ ID NO: 1, для применения в иммунотерапии пациентов, подверженных аллергии к пыльце Malus domectica и/или Betula verrucosa, где указанный белок при этом характеризуется тем, что:
a) демонстрирует уменьшенную IgE реактивность по сравнению с указанным аллергеном Mal d 1 SEQ ID NO: 1;
b) имеет аминокислотную последовательность, в которой при выравнивании с SEQ ID NO: 1 присутствуют либо две замены, включающие замену остатка Asp в положении 25 и замену остатка Asn в положении 78, либо одна замена остатка Asn в положении 78 последовательности SEQ ID NO: 1,
где указанные остатки Asp в положении 25 и/или Asn в положении 78 заменены Ala остатком.
2. Белок по п. 1, в котором IgE реактивность сывороток пациентов, подверженных аллергии к Malus domectica и/или пыльце Betula verrucosa, уменьшена по меньшей мере на 10% по сравнению с аллергеном Mal d 1 SEQ ID NO: 1.
3. Белок по пп. 1-2, состоящий из SEQ ID NO: 2.
4. Гибридный белок для применения в иммунотерапии пациентов, подверженных аллергии к пыльце Malus domectica и/или Betula verrucosa, содержащий белок, который представляет собой гипоаллергенный мутант главного аллергена Malus domectica, как определено в пп. 1-3, и белок, который представляет собой гипоаллергенный мутант главного аллергена Bet v 1 из пыльцы Betula verrucosa, разделенные линкером, где
(i) указанный мутант Bet v 1 получен из белка SEQ ID NO: 5 заменой остатков Lys в положениях 54, 115 и 123 SEQ ID NO: 5 аминокислотой Ala;
(ii) указанные Bet v 1 и Mal d 1 мутанты локализуются на амино- и карбоксил-конце гибридного белка, соответственно;
(iii) указанный линкер состоит из дипептида EF.
5. Гибридный белок по п. 4, в котором указанный мутант Bet v 1 состоит из SEQ ID NO: 6.
6. Гибридный белок по п. 4, состоящий из SEQ ID NO: 4.
7. Молекула нуклеиновой кислоты, кодирующая мутант Mal d 1 по пп. 1-3.
8. Молекула нуклеиновой кислоты по п. 7, последовательность которой выбрана из SEQ ID NO: 10 и 19.
9. Молекула нуклеиновой кислоты, кодирующая гибридный белок по пп. 4-6.
10. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п. 7.
11. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п. 9.
12. Фармацевтическая композиция для применения в иммунотерапии пациентов, подверженных аллергии к пыльце Malus domectica и/или Betula verrucosa, содержащая эффективное количество гипоаллергенного мутанта Mal d 1 по пп. 1-3 или гибридного белка в соответствии с пп. 4-6, и фармацевтически приемлемые носители, наполнители или адъюванты.
13. Композиция по п. 12, которая представлена в форме, подходящей для подъязычного, подкожного или чрескожного введения.
14. Применение белка, который представляет собой гипоаллергенный мутант главного аллергена Malus domectica Mal d 1 по пп. 1-3 или белка по пп. 4-6 или композиции по пп. 12-13 в иммунотерапии пациентов, подверженных аллергии к пыльце Malus domectica и/или Betula verrucosa, в котором указанные пациенты страдают от бронхиальной астмы, ринита, конъюнктивита или синдрома оральной аллергии.
| WO 2004039834 A2, 13.05.2004 | |||
| HOLM J., Allergy vaccine engineering: epitope modulation of recombinant Bet v 1 reduces IgE binding but retains protein folding pattern for induction of protective blocking-antibody responses, J | |||
| Immunol., 2004, v.173, n.8, p.5258-5267 | |||
| ПНЕВМАТИЧЕСКАЯ ФЛОТАЦИОННАЯ МАШИНА | 1990 |
|
RU2028188C1 |
| EA 200801463 A1, 30.12.2008. | |||

