Область техники, к которой относится изобретение
Настоящее изобретение относится к области IgE-опосредованной аллергии, в частности, к профессиональной астме, такой как астма пекарей. Более конкретно изобретение относится к идентификации нового аллергена пшеницы и его применению при терапии и диагностики. Изобретение также относится к способам диагностики и лечения IgE-опосредованной аллергии у млекопитающих.
Уровень техники
Аллергены пшеницы (Triticum aestivum) могут вызывать три разных типа IgE-опосредованной аллергии, респираторную аллергию в результате вдыхания пшеничной муки, пищевую аллергию в результате употребления в пищу продуктов из пшеницы и аллергию на пыльцу пшеницы, которая относится к группе аллергий на пыльцу растений. Аллергическая сенсибилизация к компонентам пшеничной муки является одной из наиболее частых причин профессиональной астмы, и поражает примерно 1-10% работников пекарен, поэтому заболевание называют астмой пекарей. У таких работников пекарни вырабатываются IgE-антитела против аллергенов пшеничной муки и индуцированная действием муки астма и/или ринит. Подтверждение предположения о том, что астма пекарей является настоящим профессиональным заболеванием, получено в результате обнаружения того, что распространенность сенсибилизации к ассоциированным с производством выпечки аллергенов примерно в десять раз выше у людей, подвергающихся воздействию муки, по сравнению с контрольными популяциями, не контактирующими с мукой. Кроме того, оказалось возможным установить пороговые значения в случае воздействия пшеничной муки, которые, как известно, вызывают бронхиальную астму у сенсибилизированных людей. Первое систематическое исследование аллергии на муку было осуществлено еще в 1933 году Baggoe. Другие более ранние сообщения сфокусированы на описании случаев астмы пекарей после механистических исследований, показывающих важность IgE-опосредованных механизмов в развитии астмы пекарей. Затем было предпринято несколько попыток охарактеризовать вызывающие заболевание аллергены муки иммунохимическими способами, с использованием методики RAST, иммуноблоттинга, а недавно с использованием методики молекулярного клонирования.
Triticum aestivum является важным представителем семейства злаковых трав. До 40% всех пациентов с аллергией несут сывороточные IgE-антитела, взаимодействующие с аллергенами пыльцы растений. В нескольких исследованиях сообщалось о перекрестной реактивности между пшеничной мукой и пыльцой растений вследствие наличия общих эпитопов для IgE в белках пшеничной муки и пыльцы растений. Описана перекрестная реактивность между аллергенами пыльцы растений и аллергенами зерна пшеницы, и существует доказательство того, что пациенты, страдающие астмой пекарей и IgE-опосредованной пищевой аллергией на пшеницу, могут узнавать разные аллергены, которые можно использовать для дифференциальной диагностики астмы пекарей, пищевой аллергии на пшеничную муку и аллергии на пыльцу растений. Однако огромное количество растворимых аллергенов пшеничной муки еще не идентифицировано. В настоящее время идентифицировано и охарактеризовано только небольшое количеств аллергенов пшеничной муки, и к ним относятся представители семейства ингибиторов альфа-амилазы, ацил-CoA-оксидаза, пероксидаза, фруктозодифосфатальдолаза, и в последнее время тиоредоксины.
Диагностика, основанная на использовании экстракта пшеничной муки, не позволяет обнаружить различие между пациентами, страдающими респираторной аллергией или пищевой аллергией на пшеницу. Поэтому точная диагностика все еще основана на специальной ингаляционной провокации в случае респираторной аллергии на пшеничную муку и двойной слепой плацебо-контролируемой пищевой провокации (DBPCFC) в случае предполагаемой пищевой аллергии, и остается открытым вопрос, можно ли идентифицировать аллергены, которые могут быть использованы для селективной диагностики и лечения различных индуцированных пшеницей проявлений аллергии, таких как астма пекарей, пищевая аллергия и поллинозы. Таким образом, существует необходимость в идентификации новых аллергенов пшеницы и разработки способов и диагностических тестов, чтобы более точно выявлять пациентов, страдающих IgE-опосредованной аллергией, такой как респираторная аллергия, например, астма пекарей, чтобы отличать их от пациентов, страдающих пищевой аллергией и/или аллергией на пыльцу. Кроме того, существует необходимость в применении таких новых аллергенов пшеницы для лечения IgE-опосредованной аллергии. Настоящее изобретение обращено к проблеме, связанной с необходимостью получения новых аллергенов пшеницы и их применением для терапии и диагностики. Кроме того, настоящее изобретение относится к применению известных пептидов и белков в терапии и диагностике. Некоторые из таких пептидов известны: Gennaro S. D. et al., Biological Chemistry, April 2005, 386: 383-389; UNIPROT, номер доступа P82977; UNIPROT, номер доступа Q6W8Q2; US 7214786 и US2006/0107345.
Сущность изобретения
Как указано выше, несколько аллергенов пшеничной муки, которые были идентифицированы и охарактеризованы, включают представителей семейства ингибиторов альфа-амилазы, ацил-CoA-оксидазу, пероксидазу, фруктозодифосфатальдолазу и тиоредоксины. До настоящего времени не были идентифицированы аллергены, которые можно применять для селективной диагностики и лечения различных индуцированных пшеницей проявлений аллергии, таких как астма пекарей, пищевая аллергия и поллиноз. Это привело авторов настоящего изобретения к поиску дополнительных, еще не идентифицированных аллергенов пшеницы.
Настоящее изобретение, по меньшей мере, частично удовлетворяет потребность предшествующего уровня техники, так как предлагает новые аллергены пшеницы и способы диагностики и лечения IgE-опосредованной аллергии у млекопитающих.
В первом аспекте изобретение относится к полипептидам, представляющим собой новые аллергены пшеницы, выделенным из пшеницы или полученным рекомбинантно, и их фрагментам или вариантам, имеющим общие эпитопы для антител. Изолированные полипептиды содержат аминокислотную последовательность клона №10, №112 или №126.
Во втором аспекте изобретение относится к нуклеиновым кислотам, кодирующим полипептиды согласно изобретению, нуклеиновым кислотам, имеющим нуклеотидную последовательность клона №10, №112 или №126.
В другом аспекте изобретение относится к полипептиду, обладающему идентичностью с клонами №10, №38, №112 или №126, для применения в терапии или диагностике, предпочтительно терапии и диагностике IgE-опосредованной аллергии, такой как респираторная аллергия на пшеничную муку, например, астма пекарей. Кроме того, изобретение относится к изолированному полипептиду, содержащему аминокислотную последовательность клона №37, для применения в терапии или диагностике, предпочтительно терапии и диагностике IgE-опосредованной аллергии, такой как респираторная аллергия на пшеничную муку, например, астма пекарей.
В еще одном аспекте изобретение относится к фармацевтической композиции, содержащей полипептид, обладающий идентичностью с клонами №10, №38, №112, №126 или №37, или к его гипоаллергенной форме, модифицированной с тем, чтобы отменить или ослабить IgE-связывающий ответ, и необязательно фармацевтически приемлемые эксципиенты, носители, буферы и/или разбавители. Гипоаллергенная форма полипептида, обладающего идентичностью с клонами №10, №38, №112, №126 или №37, может быть модифицирована посредством фрагментации, укорочения или образования тандема молекулы, делеции внутренних участков, перестановки доменов, замены аминокислотных остатков, разрушения дисульфидных мостиков.
В следующем аспекте изобретение относится к композиции аллергенов, «обогащенной» полипептидом, обладающим идентичностью с клонами №10, №38, №112, №126 или №37. Такая композиция аллергенов может представлять собой экстракт аллергенов или смесь очищенных или рекомбинантных аллергенных компонентов, которые не содержат или содержат небольшое количество полипептида согласно изобретению, при этом полипептид добавляют для того, чтобы связать IgE у пациентов, у которых IgE может быть не связан или слабо связан с другими аллергенными компонентами в композиции. Данный аспект изобретения также относится к способу получения такой композиции, и такой способ включает стадию добавления полипептида, обладающего идентичностью с клонами №10, №38, №112, №126 или №37, к композиции аллергенов, такой как экстракт аллергенов (необязательно обогащенный другими компонентами) или смесь очищенных нативных или рекомбинантных аллергенных компонентов.
В следующем аспекте изобретение относится к композиции аллергенов, получаемой описанным выше способом.
Изобретение, кроме того, относится к способу диагностики in vitro IgE-опосредованной аллергии, включающему стадии контактирования образца жидкости, такого как образец крови, плазмы или сыворотки, из организма млекопитающего, у которого предполагается наличие IgE-опосредованной аллергии, такой как респираторная аллергия на пшеничную муку, например, астма пекарей, с полипептидом, обладающим идентичностью с клонами №10, №38, №112, №126 или №37, и выявления присутствия в образце IgE-антител, которые специфично связываются с полипептидами по изобретению. Присутствие таких антител, специфично связывающихся с полипептидом, обладающим идентичностью с клонами №10, №38, №112, №126 или №37, является показателем IgE-опосредованной аллергии. Один вариант такого способа включает осуществление способа с использованием анализа на микроматрице.
В следующем аспекте изобретения предлагается диагностический набор для осуществления способа диагностики in vitro IgE-опосредованной аллергии, такой как респираторная аллергия на пшеничную муку, например, астма пекарей, при этом указанный набор содержит полипептид, обладающий идентичностью с клонами №10, №38, №112, №126 или №37, и средства для выявления связывания IgE с указанным полипептидом, такие как твердая подложка, например, нитроцеллюлозная мембрана или микроматрица, содержащая связанный с ней полипептид, обладающий идентичностью с клонами №10, №38, №112, №126 или №37.
В следующем аспекте изобретение относится к способу лечения IgE-опосредованной аллергии у млекопитающего, такой как респираторная аллергия, например, астма пекарей. В одном варианте способ включает введение пациенту, чувствительному к такому лечению, полипептида, обладающего идентичностью с клонами №10, №38, №112, №126 или №37, или его фрагмента или варианта, имеющих общие эпитопы для антител. В другом варианте способ включает введение пациенту, чувствительному к такому лечению, фармацевтической композиции согласно предыдущему аспекту.
Определения
Определения клонов
Следует считать, что варианты и фрагменты полипептида по изобретению означают белки или пептиды длиной, по меньшей мере, 10 аминокислот, более предпочтительно, по меньшей мере 25, еще более предпочтительно, по меньшей мере, 50 или 75 аминокислотных остатков, и идентичность последовательности с указанным полипептидом, составляющую, по меньшей мере, 50 %, предпочтительно более 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99%.
Модифицированный полипептид в контексте настоящего изобретения следует толковать как полипептид, который был химически или генетически модифицирован для того, чтобы изменить его иммунологические свойства, например, как проиллюстрировано в отношении аспекта изобретения, связанного с иммунотерапией.
Варианты и фрагменты полипептида, имеющего общие с указанным полипептидом эпитопы для антител, следует толковать как такие фрагменты и варианты, связывание которых с IgE-антителами из образца сыворотки типичного пациента, сенсибилизированного полипептидом согласно изобретению, может быть в значительной степени ингибировано полипептидом. Такой анализ ингибирования может быть, например, осуществлен согласно протоколу, описанному в примере 4.
Гипоаллергенный модифицированный полипептид или вариант или фрагмент полипептида следует толковать как модифицированный полипептид или вариант или фрагмент полипептида, который не способен связывать IgE-антитела, реактивные к указанному полипептиду, из образца сыворотки типичного сенсибилизированного полипептидом пациента, который определен, например, в протоколе согласно примеру 1 или который не проявляет или проявляет в значительной степени сниженную биологическую аллергенную активность, которую определяют с использованием анализа активации клеток, такого как анализ высвобождения гистамина базофилами.
Краткое описание таблиц и фигур
В таблице I показаны демографические, клинические и серологические характеристики пациентов, страдающих астмой пекарей.
В таблице II показаны демографические, клинические и серологические характеристики пациентов, страдающих пищевой аллергией на пшеницу (F1-F4) и аллергией на пыльцу растений (G1-G4).
В таблице III показаны идентичности аминокислотных последовательностей в процентах между полученным из клона 10 аллергеном и гомологичными белками, где следующие белки пронумерованы 1-30: 1. gi|122065237 (Triticum aestivum), 2. gi|66356278 (Triticum aestivum), 3. gi|124122 (Hordeum vulgare подвид vulgare), 4. gi|48093360 (Zea diploperennis), 5. gi|48093418 (Tripsacum dactyloides), 6. gi|75994161 (Zea mays подвид parviglumis), 7. gi|58396945 (Oryza sativa [группа японских сортов]), 8. gi|115649132 (Strongylocentrotus purpuratus), 9. gi|37904392 (Brachypodium distachyon), 10. gi|26224744 (Citrus x paradise), 11. gi|224447 (Vicia faba), 12. gi|124395862 (Paramecium tetraurelia), 13. gi|50262213 (Cucurbita maxima), 14. gi|547743 (Nicotiana sylvestris), 15. gi|54610713 (Lumbricus terrestris), 16. gi|169491 (Solarium tuberosum), 17. gi|218290 (Nicotiana glauca x Nicotiana langsdorffiij, 18. gi|124121 (Vigna angularis), 19. gi|603890 (Sambucus nigra), 20. gi|14718445 (Ipomoea batatas), 21. gi|114950 (Momordica charantia), 22. gi|109138554 (Fagopyrum esculentum), 23. gi|18404883 (Arabidopsis thaliana), 24. gi|27734408 (Canavalia lineate), 25. gi|37901103 (Hevea brasiliensis), 26. gi|92874842 (Medicago truncatula), 27. gi|13959383 (Linum usitatissimum), 28. gi|22759723 (Zinnia elegans), 29. gi|37359345 (Vitis vinifera) и 30. gi|6453287 (Amaranthus hypochondriacus).
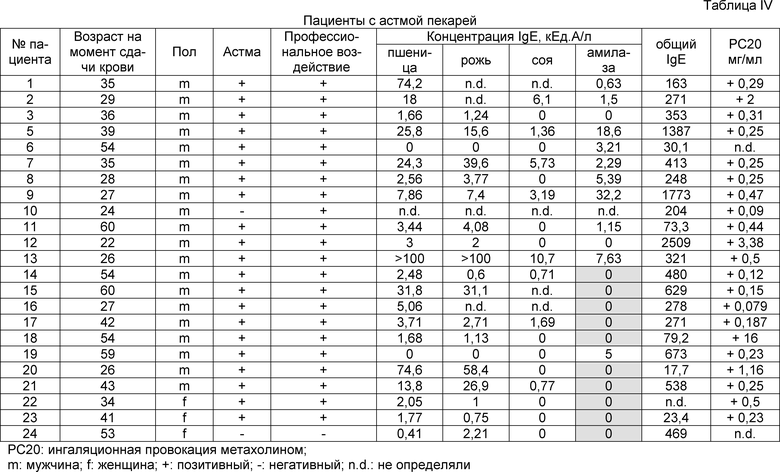
В таблице IV показаны клинические данные для пациентов с астмой пекарей из примера 2.
В таблице V показаны клинические данные для пациентов с пищевой аллергией и аллергией на пыльцу растений из примера 2.
В таблице VI показаны ПЦР-праймеры, используемые для амплификации кДНК клонов №10, №38, №112, №123, №126 и №37.
Таблица VII. Демографические, клинические и серологические характеристики пациентов, страдающих астмой пекарей.
Таблица VIII. Демографические, клинические и серологические характеристики пациентов, страдающих аллергией на пыльцу растений.
Таблица IX. Демографические, клинические и серологические характеристики пациентов, страдающих пищевой аллергией на пшеницу.
Фигура 1 иллюстрирует нуклеотидную и рассчитанную аминокислотную последовательность полученного из клона 10 аллергена (SEQ ID NO: 1). Кодирующая и не кодирующая области показаны заглавными буквами и строчными буквами, соответственно, стартовый кодон (ATG) и стоп-кодон подчеркнуты. Надписи аминокислот семейства ингибиторов I картофеля напечатаны на сером фоне. Номера слева указаны для нуклеотидов, а номера справа указаны для аминокислот. Последовательность представлена в GenBank с номером доступа (EU051824).
Фигура 2 иллюстрирует определение параметров очищенного аллергена, подобного ингибитору сериновой протеиназы. A: Окрашенный Кумасси бриллиантовым синим SDS-ПААГ, содержащий очищенный полученный из клона 10 аллерген. Маркер молекулярной массы (кД) показан слева. B: Масс-спектрометрия (МС) очищенного полученного из клона 10 аллергена. Отношение масса/заряд показано на оси x, а интенсивность показана на оси y в виде процента от наиболее интенсивного сигнала, полученного в исследованном диапазоне масс. C: Анализ кругового дихроизма (КД) в дальней ультрафиолетовой области очищенного полученного из клона 10 аллергена. Спектры представлены в виде средней эллиптичности остатков (θ) (ось y), регистрируемой при 25°C (жирная линия), 95°C (обычная линия) и 25°C после охлаждения (пунктирная линия) при заданных длинах волн (ось x).
Фигура 3 иллюстрирует реактивность IgE у пациентов, страдающих астмой пекарей, аллергией на пыльцу растений и пищевой аллергией. Очищенный полученный из клона 10 аллерген, HSA, rPh1 p 1, rPh1 p 5, rPh1 p 7, rPh1 p 12, экстракт пыльцы пшеницы и экстракт зерна пшеницы в виде пятен наносили на полоски нитроцеллюлозной мембраны и инкубировали с сыворотками 22 пациентов с астмой пекарей (1-22), четырьмя сыворотками пациентов с аллергией на пыльцу растений (G1-G4), четырьмя сыворотками пациентов, страдающих пищевой аллергией на пшеницу (F1-F4), одной сывороткой человека, не страдающего аллергией (NC) и с буфером без добавления сыворотки (B). Связанные IgE-антитела выявляли с использованием 125I-меченых антител против IgE человека и визуализировали с помощью авторадиографии.
На фигуре 4 представлена коробчатая диаграмма реактивностей подклассов IgG по отношению к полученному из клона 10 аллергену. Реактивность подклассов IgG1-4 к полученному из клона 10 аллергену определяли в ELISA для пациентов, страдающих астмой пекарей (n=22) и представляли в виде коробчатых диаграмм, где 50% значений находятся в пределах прямоугольников и не являются выбросами за пределы линий, указывающих величины ошибки. Линии внутри прямоугольников указывают медианные значения, кружками указаны выбросы и звездочками указаны экстремальные значения.
Фигура 5 иллюстрирует аллергенную активность аллергена, полученного из клона 10. Клетки RBL нагружали IgE сыворотки от трех пациентов с астмой пекарей (№2, №4, №12) или сывороткой не страдающего аллергией пациента (NC) и затем провоцировали рекомбинантным полученным из клона 10 аллергеном или аллергеном пыльцы тимофеевки луговой rPh1 p 1. Средние значения высвобождения β-гексозаминидазы показаны по оси y в виде процента от общего высвобождения после вычитания процента спонтанного высвобождения.
На фигуре 6 показана экспрессия аллергена, подобного ингибитору сериновых протеиназ, в семенах во время созревания семян. Нитроцеллюлозные блоты экстракта пшеницы из незрелых (7, 10, 15, 20, 25, 30, 35 день) и зрелых (M) семян пшеницы анализировали, используя кроличьи антитела, специфичные для полученного из клона 10 аллергена, и в целях контроля используя соответствующую сыворотку до иммунизации. Молекулярные массы указаны слева в килодальтонах (кД).
На фигуре 7 показана идентификация полученного из клона 10 аллергена в пыльце пшеницы и экстрактах семян. Экстракты на нитроцеллюлозных блотах анализировали, используя кроличьи антитела, специфичные для полученного из клона 10 аллергена (20), антитела, специфичные для профилина пшеницы (I №123), для клещевого аллергена (NC), или используя буфер без добавления кроличьих антител (B). Соответствующие сыворотки, полученные до иммунизации, указаны как P №10 и P №123, соответственно. Связанные IgG-антитела выявляли с использованием 125I-меченых антител осла против Ig кролика и визуализировали с помощью авторадиографии. Маркеры молекулярной массы (в кД) указаны слева.
Фигура 8 иллюстрирует выявление аллергена, подобного ингибитору сериновых протеиназ, в экстрактах семян пшеницы, риса, кукурузы, фасоли и картофеля. A: Окрашенный Кумасси синим гель, содержащий экстракты пшеницы (W), риса (R) и кукурузы (M), фасоли обыкновенной (B) и картофеля (P). B: Экстракты на нитроцеллюлозных блотах подвергали воздействию сыворотки кролика до иммунизации и C: воздействию антител, специфичных для полученного из клона 10 аллергена. Молекулярные массы (кД) указаны слева.
На фигуре 9 показана локализация полученного из клона 10 аллергена в семенах пшеницы с помощью трансмиссионной электронной микроскопии с иммунологическим мечением частицами золота. A и B: Поперечный срез зерна пшеницы при малом (A) и большом (B) увеличении. На фигуре A показана плодовая оболочка и семенная оболочка (C), алейроновый слой (AL) и начало крахмалистого эндосперма (SE). Прямоугольником на фигуре A показана область, сопоставимая с областью, показанной на фигуре B, т.е. граница между алейроновым слоем и крахмалистым эндоспермом. Прямоугольниками на фигуре B очерчены области, показанные при большом увеличении на фигурах C, D и E, F, соответственно. C и D: Детальное изображение алейроновой клетки зерна пшеницы, исследованной с использованием Ig кролика, полученного против клона 10 (C) или Ig, полученного до иммунизации (D). E и F: Микрофотография при большом увеличении крахмалистого эндосперма после локализации на основе иммунологического мечения частицами золота белка пшеницы 10 с использованием Ig кролика, полученного против Ig клона 10 (E) или Ig до иммунизации (F). Связанные кроличьи антитела выявляли с помощью конъюгированной с золотом антисыворотки козы против Ig кролика (частицы золота = черные точки). Стрелки указывают частицы коллоидного золота. Линиями указано: A, 20 мкм; B, 5 мкм; C-F, 0,5 мкм. AG, алейроновое зерно; AL, алейроновый слой; C, многослойная плодовая и семенная оболочка; CY, вещества цитоплазмы; L, липидное тельце; M, митохондрия; SE, крахмалистый эндосперм; SG, крахмально зерно; W, клеточная стенка.
Фигура 10. Дот-блот-анализ IgE пациентов, страдающих астмой пекарей.
Фигура 11. Дот-блот-анализ IgE пациентов, страдающих пищевой аллергией на пшеницу и аллергии на пыльцу растений.
Фигура 12. Микроматрица аллергенов. A: Схема нанесения на микроматрицу белков и экстракта пыльцы пшеницы. Рекомбинантные белки пшеницы обозначены: 10, 37, 38, 112, 123, 126; WP: экстракт пыльцы пшеницы; рекомбинантные аллергены пыльцы тимофеевки луговой: Ph1 p 1, Ph1 p 5, Ph1 p 7 и Ph1 p 12. Числами в находящемся снизу прямоугольнике указаны маркеры положения. B и C: Изображения микроматриц после инкубации с сывороткой и выявления IgE-реактивных пятен с использованием конъюгированных с флуорофором анти-IgE-антител. B: Изображение после инкубации с сывороткой пациента, не страдающего аллергией. C: Изображения после инкубации с сывороткой типичного пациента, страдающего астмой пекарей (1:B4), индуцированной пшеницей пищевой аллергией (2:F26), аллергией на пыльцу растений (3:G16). Пятнами снизу стекол указаны маркеры положения, которые представляли собой очищенный IgE-антитела, выявляемые с использованием конъюгированных с флуорофором анти-IgE-антител.
Фигура 13. Распространенность IgE-реактивности на белки семян пшеницы, экстракт пыльцы пшеницы и аллергены пыльцы растений. Процент пациентов с IgE-реактивностью показан для случаев астмы пекарей (A) (n=23), пищевой аллергии на пшеницу (B) (n=38) и аллергии на пыльцу растений (C) (n= 17) на оси y. На оси x показаны тестируемые рекомбинантные белки пшеницы №10, №37, №38, №112, №123 и №126, экстракт пыльцы пшеницы, рекомбинантные аллергены пыльцы растений Ph1 p 1 , Ph1 p 5, Ph1 p 7 и Ph1 p 12, «пшеничная смесь», содержащая все рекомбинантные белки пшеницы, CAP пшеничной муки и тимофеевки луговой, используемые для измерения IgE-реактивности.
Подробное описание изобретения
Приведенные ниже примеры иллюстрируют настоящее изобретение в отношении выделения и применения полипептида согласно изобретению. Примеры являются только иллюстративными, и их не следует рассматривать как ограничивающие изобретение, которое определено объемом прилагаемой формулы изобретения. Клон №123, упоминаемый в примерах, является профилином, известным, например, из US 7214786.
ПРИМЕР 1: Клон №10 нового аллергена пшеницы
В примере 1 показана идентификация и характеристика нового аллергена семян пшеницы, клона №10, относящегося к семейству ингибиторов картофеля, семейству ингибиторов сериновых протеаз, которые вместе с другими ингибиторами протеаз называют белками, связанными с патогенезом (PR), семейству PR6. Клон №10 является первым аллергеном, идентифицированным и описанным для семейства PR6. Кроме того, в примере 1 показана экспрессия и очистка рекомбинантного полученного из клона 10 аллергена.
Клон №10 специфично распознавался IgE сыворотки пациентов с астмой пекарей, но не проявлял реактивности по отношению к IgE при тестировании с сыворотками от пациентов, страдающих пищевой аллергией на пшеницу, целиакией или аллергией на пыльцу растений. Поэтому полученный из клона 10 аллерген вместе с другими аллергенами пшеницы можно использовать для разработки диагностических тестов, которые позволяют специфично идентифицировать пациентов, страдающих от IgE-опосредованной астмы пекарей, и отличать таких пациентов от пациентов с пищевой аллергией или аллергией на пыльцу.
Способы
Биологические материалы, сыворотки пациентов и антитела
Семена пшеницы Triticum aestivum сорта Michael получали из Osterreichische Agentur fur Gesundheit und Ernahrungssicherheit GmbH и высевали в теплицу. Незрелые семена собирали на 7, 10, 15, 20, 25, 30 и 35 день после начала опыления сразу в жидкий азот и хранили при -80°C вплоть до использования. Пыльцу пшеницы получали из Allergon (Valinge, Sweden). Рис, кукурузу, фасоль и картофель покупали на местном рынке. Рекомбинантные Ph1 p 1, Ph1 p 5, Ph1 p 7 и Ph1 p 12 приобретали из BIOMAY (Vienna, Austria) и сывороточный альбумин человека (HSA) приобретали из Behring (Marburg, Germany). Сыворотки получали от 22 пациентов, страдающих астмой пекарей. Астму пекарей диагностировали на основании положительного анамнеза, IgE, специфичного по отношению к пшеничной и ржаной муке, выявляемого с помощью системы CAP-FEIA (Phadia, Uppsala, Sweden) и специальных провокационных ингаляционных тестов для подтверждения клинически значимой сенсибилизации (1). Демографические, клинические и серологические данные для таких пациентов суммированы в таблице I. Кроме того, в эксперименты включали сыворотку человека, не страдающего аллергией, сыворотки 4 пациентов, страдающих пищевой аллергией на пшеницу и 4 пациентов с аллергией на пыльцу растений без признаков астмы пекарей, но с IgE-реактивностью сыворотки на пшеничную и ржаную муку (таблица II). Сыворотки пациентов с аллергией на пыльцу растений анализировали в отношении общих уровней IgE в сыворотке и IgE, специфичного в отношении пыльцы тимофеевки луговой в системе CAP-FEIA (Phadia), а пациентов с пищевой аллергией на пшеницу характеризовали как описано ранее (2). Специфичность полученного из клона 10 аллергена для астмы пекарей подтверждали в результате тестирования дополнительных 20 сывороток пациентов с целиакией, 119 пациентов с пищевой аллергией, 23 сывороток пациентов с аллергией на пыльцу растений и 25 пациентов с астмой пекарей с использованием анализа чипов (Constantin et al, не опубликовано).
Специфичные кроличьи антитела против полученного из клона 10 аллергена были получены при иммунизации кролика с месячными интервалами очищенным полученным из клона 10 аллергеном (200 мкг на инъекцию) с использованием полного адъюванта Фрейнда и дважды с использованием IFA (Charles River, Kisslegg, Germany). Сыворотку до иммунизации получали от кролика перед иммунизацией. В целях контроля использовали иммунную сыворотку кролика, специфичную для клещевого аллергена домашней пыли, и кроличью антисыворотку, специфичную для профилина пшеницы.
Конструирование библиотеки кДНК λgt11 из семян пшеницы
Суммарную РНК экстрагировали согласно способу Yeh (3) из семян пшеницы, собранных через двадцать пять дней после начала опыления, и хранили при -80°C. Затем осадок РНК растворяли в буфере, содержащем изотиоцианат гуанидиния (4М изотиоцианат гуанидиния, 0,83% об./об. 3М ацетата натрия, pH 6, 11 мМ β-меркаптоэтанол), и очищали ультрацентрифугированием в градиенте плотности хлорида цезия (4). Поли-A+-РНК выделяли аффинной хроматографией на олиго-dT-целлюлозе (Nucleo Trap mRNA; Machery-Nagel) и синтезировали двунитевую кДНК, используя набор для синтеза кДНК (Система для синтеза кДНК; Roche Diagnostics, Mannheim, Germany). После метилирования EcoRI-метилазой (New England Biolabs, Beverly, MA), к кДНК добавляли EcoRI-линкеры (New England Biolabs). Связанную линкерами кДНК расщепляли EcoRI (Roche Diagnostics). Отщепленные линкеры удаляли на колонке Nick (Pharmacia Biotech, Uppsala, Sweden) и кДНК лигировали в плечи λgt11 (Stratagene, La Jolla, CA, USA). Продукт лигирования подвергали упаковке in vitro (набор для клонирования Gigapack III Gold, Stratagene), получая экспрессирующую библиотеку кДНК в λgt11 с 2,43×106 БОЕ.
Выделение и характеристика IgE-реактивных клонов из библиотеки кДНК семян пшеницы
E. coli Y1090 инфицировали, используя 7×105 БОЕ рекомбинантных фагов, и проводили иммунный скрининг, используя IgE сыворотки четырех пациентов (№1, №2, №4, №12), страдающих астмой пекарей, как описано (5). Пятнадцать IgE-реактивных фаговых клонов были отобраны для дальнейшего повторного клонирования, и их ДНК была амплифицирована в ПЦР с использованием смеси для ПЦР Platinum PCR Supermix (Invitrogen, Life Technolgies) и праймеров λgt11 и секвенирована (MWG, Ebersberg, Germany). Полученные последовательности сравнивали с последовательностями, представленными в базе данных GenBank в Национальном центре биотехнологической информации (NCBI). Осуществляли множественное выравнивание последовательностей, используя базу данных GenBank в NCBI. Для определения идентичности аминокислотных последовательностей использовали программу множественных выравниваний Clustal W. Поиск мотивов осуществляли с помощью программы PROSITE, имеющейся на сервере ExPASy proteomics, для аминокислотного состава использовали программу ProtParam ExPASy. Прогнозирование доступности растворителю и вторичной структуры осуществляли, используя компьютерную программу PROT из центра биоинформатики Колумбийского университета. Филогенетическое древо реконструировали на основе аминокислотной последовательности полученного из клона 10 аллергена и гомологичных белков, используя компьютерную программу «Множественное выравнивание и реконструкция филогенетического древа», предоставленную Институтом молекулярной генетики Макса Планка.
Экспрессия и очистка полученного из клона 10 рекомбинантного аллергена
Кодирующую область кДНК клона 10 амплифицировали в ПЦР, используя следующую пару праймеров: прямой 5'-CATATGAGCCCTGTGGTGAAGAAGCCGGAGGGA-3' и обратный 5'-GAATTCTTAGTGATGGTGATGGTGATGGCCGACCCTGGGGAC-3' (MWG). Продукт ПЦР содержат сайты рестрикции NdeI (курсивом), EcoRI (подчеркнут) и последовательность, кодирующую гексагистидиновую метку (жирным шрифтом). Продукт ПЦР субклонировали в векторе AccepTor (Novagen, Madison, WI) и снова секвенировали (MWG). Затем вставку вырезали из вектора AccepTor, используя NdeI и EcoRI (Roche Diagnostics), очищали в геле (Promega, Madison, WI, USA) и субклонировали в экспрессирующей плазмиде pET 17b (Novagen). Последовательность ДНК подтверждали секвенированием обеих нитей ДНК (Microsynth, Balgach, CH). Конструкцией pET 17b-клон 10 трансформировали клетки E. coli BL21 (DE3) (Stratagene) и выращивали в среде Луриа (17), содержащей 100 мг/л ампициллина, при 37°C до OD (600 нм) 0,8-1. Экспрессию белка индуцировали добавлением изопропил-β-D-тиогалактопиранозида (IPTG) до конечной концентрации 0,5 мМ и выращивали бактерии еще в течение 3 часов. Бактерии собирали центрифугированием и гомогенизировали в 25 мМ имидазоле, pH 7,5, 0,1% (об./об.) тритоне X-100, используя Ultraturrax (IKA, Stauffen, Germany). ДНК расщепляли, добавляя ДНКазу I, дополнительно перемешивали еще в течение 10 минут при 20°C, реакцию останавливали, используя 200 мкл 5 М NaCl, и затем центрифугировали при 4°C (6000×g, 20 минут). Большую часть полученного из клона 10 аллергена обнаружили в нерастворимой фракции бактериального экстракта. Полученный из клона 10 аллерген очищали из осадка, содержащего тельца включения, в денатурирующих условиях, используя аффинные колонки с Ni-NTA-смолой, согласно руководству QIAexpressionist (QIAGEN, Hilden, Germany). Фракции, содержащие рекомбинантный аллерген, объединяли и диализовали против 10 мМ NaH2PO4, pH 7,5. Концентрацию белка определяли с помощью набора для анализа белка Micro BCA (Pierce, Rockford, IL).
ELISA
Полученный из клона 10 аллерген растворяли в PBS в концентрации 5 мкг/мл и покрывали планшеты для ELISA (Nunc Maxisorb, Roskilde, Denkmark). После блокирования 1% (масс./об.) БСА в PBS, 0,05% (об./об.) твин-20 (PBST), планшеты инкубировали с сыворотками в разведении 1:50 в PBST с 0,5% (масс./об.) БСА для измерения IgG1, IgG2, IgG3 и IgG4, как описано (6). Связанные антитела выявляли посредством инкубации сначала с моноклональными антителами мыши против подкласса IgG человека (BD Biosciences, Franklin Lakes, NJ) в разведении 1:1000 в PBST, 0,5% (масс./об.) БСА и затем со связанной с пероксидазой хрена антисывороткой овцы против Ig мыши (GE Healthcare, Little Chalfont, UK) в разведении 1:2000 в PBST, 0,5% (масс./об.) БСА, как описано ранее (2). Все определения осуществляли в двух повторах, и результаты выражали в виде средних значений.
Экстракты белков, SDS-ПААГ и иммуноблоты
Экстракты белков с SDS из зрелых и незрелых семян пшеницы, риса (Oryza sativa), кукурузы (Zea mays), фасоли (Phaseolus vulgaris) и картофеля (Solarium tuberosum) готовили посредством гомогенизации 3 грамм ткани в 32 мл буфера для образцов (6) и затем кипятили в течение 10 минут. Чтобы удалить нерастворимые частицы экстракты центрифугировали при 10000×g в течение 10 минут при 4°C и надосадки хранили в виде аликвот при -20°C. Кроме того, готовили экстракт белка в PBS из семян пшеницы, как описано ранее (2). Пыльцу Triticum aestivum (500 мг) экстрагировали при 4°C в течение ночи в 5 мл PBS, 2 мМ EDTA, 1 мМ PMSF. После центрифугирования в течение 1 часа при 13000×g и 4°C определяли концентрацию белка в надосадке, используя набор для анализа белка Micro BCA (Pierce), и аликвоты хранили при -20°C вплоть до использования. Равные количества экстрактов белка с SDS разделяли в 14% препаративных SDS-полиакриламидных гелях (7). В качестве стандарта использовали маркер молекулярной массы (маркер Rainbow, GE Healthcare; стандарт белка Precision Plus, BioRad, Herkules, CA; предварительно окрашенный лэддер белков Page Ruler, Fermentas, Burlington, Ontario). После электрофоретического разделения белки либо окрашивали Кумасси бриллиантовым синим, либо переносили на нитроцеллюлозные мембраны (Schleich and Schuell, Dassel, Germany) (8). Мембраны блокировали в буфере A (50 мМ натрий-фосфатный буфер, pH 7,4, 0,5% масс./об. БСА, 0,5% об./об. твин-20, 0,05% масс./об. NaN3) два раза по 10 минут и один раз в течение 30 минут и инкубировали в течение ночи при 4°C с кроличьей антисывороткой, специфичной для полученного из клона 10 аллергена, с соответствующий сывороткой, полученной до иммунизации, и в целях контроля с кроличьей антисывороткой, специфичной для неродственного антигена, или только с буфером. Сыворотки кролика разводили 1:50000 в буфере A. Связанные антитела выявляли с использованием разведенных 1:2000 в буфере A 125I-меченых антител осла против Ig кролика (GE Healthcare) в течение 2 часов при комнатной температуре и визуализировали на пленках Kodak XOMAT с использованием усиливающих экранов (Kodak, Heidelberg, Germany) при -70°C. В случае дот-блот-экспериментов для анализа IgE 100 нг рекомбинантного полученного из клона 10 аллергена и рекомбинантных аллергенов пыльцы растений, Ph1 p 1 , Ph1 p 5, Ph1 p 7 и Ph1 p 12, а также 3 мкг экстракта пыльцы пшеницы и 2 мкг PBS-экстракта зрелых семян пшеницы в виде пятен наносили на нитроцеллюлозную мембрану. Полоски нитроцеллюлозы блокировали буфером A и подвергали воздействию сыворотки пациентов в разведении 1:10 в буфере A в течение ночи при 4°C. Связанные IgE-антитела выявляли с использованием 125I-меченых антител против IgE человека (RAST RIA, Demeditec Diagnostics, Germany) в разведении 1:20 в буфере A в течение ночи при комнатной температуре и визуализировали с помощью авторадиографии, используя пленки Kodak XOMAT с усиливающими экранами (Kodak) при -70°C.
Анализы МС и КД рекомбинантного клона 10
Масс-спектры с лазерной десорбцией получали в линейном режиме, используя прибор TOF Compact MALDI II (Kratos, Manchester, UK; piCHEM, Research and Development, Graz, Austria). Образцы растворяли в 10% ацетонитриле (0,1% трифторуксусная кислота), и α-циано-4-гидроксикоричную кислоту (растворенную в 60% ацетонитриле с добавлением 0,1% трифторуксусной кислоты) использовали в качестве матрицы. Для получения образцов смесь 1/1 белка и раствора матрицы помещали на мишень и сушили на воздухе.
Измерения КД осуществляли с использованием очищенного полученного из клона 10 аллергена (в H2O) при концентрации белка 0,1 мг/мл на спектрополяриметре Jasco J-810 (Tokyo, Japan), используя прямоугольную кварцевую кювету, длина пути 0,2 см. КД-спектры в дальней ультрафиолетовой области регистрировали от 190 нм до 260 нм с разрешением 0,5 нм и скоростью сканирования 50 нм/мин и получали среднее значение из трех сканирований. Результаты выражены в виде средней остаточной эллиптичности (θ) при данной длине волны. Температурное сканирование осуществляли согласно способу пошагового сканирования, при котором образец нагревали от 25°C до 95°C со скоростью нагревания 2°C/мин и снова охлаждали до 25°C с такой же скоростью. Через каждые 5°C регистрировали спектры с непрерывным изменением длины волны с использованием конкретных параметров. Кроме того, температурное сканирование осуществляли при 215 нм с разрешением шагов 0,5°C. Результаты выражены в виде средней остаточной молярной эллиптичности (θMRE) при данной длине волны. Конечные спектры корректировали, вычитая соответствующий спектр исходного уровня, полученный в идентичных условиях. Информацию о вторичной структуре полученного из клона 10 аллергена рассчитывали с использованием программы оценки вторичной структуры CDSSTR (9).
Анализ клеток базофильного лейкоза крыс, несущих рецептор человека
Для количественной оценки опосредованных IgE-Ат реакций немедленного типа осуществляли анализ высвобождения медиаторов клетками huRBL. Клетки RBL (clone RBL-703/21), трансфицированные FcεRI человека (10), культивировали в RPMI 1640 с добавлением 5% FCS, 4 мМ L-глутамина и 1 мг/мл сульфата G418. Клетки собирали после инкубации с трипсином/EDTA, промывали, ресуспендировали в культуральной среде и концентрацию клеток доводили до 2×106 клеток/мл. Аликвоты суспензии клеток по пятьдесят микролитров добавляли в лунки 96-луночного микропланшета с плоским дном (плотность клеток/лунку составляла 1×105 клеток). Сыворотку человека разбавляли 1:10 в культуральной среде, добавляли к клеткам и инкубировали в течение ночи при 37°C, 7% CO2, относительной влажности 95%. Среду удаляли и планшеты 3 раза промывали 200 мкл/лунку буфера Тайрода +0,1% БСА. Для перекрестного связывания IgE 100 мкл полученного из клона 10 аллергена или rPh1 p 1, (0,3 мкг/мл), разбавленного в буфере Тайрода, содержащем 50% D2O и 0,1% (масс./об.) БСА, добавляли к клеткам. Для спонтанного высвобождения в лунки добавляли буфер Тайрода без белка. Общее высвобождение определяли при добавлении буфера Тайрода, содержащего 10% тритон X-100. После инкубации при 37°C, 7% CO2, относительной влажности 95% в течение 1 часа клетки собирали центрифугированием и 50 мкл надосадка переносили в новый планшет и добавляли 50 мкл раствора для анализа (0,1М лимонная кислота или цитрат натрия, pH 4,5 и 160 мкМ 4-метилумбеллиферил-N-ацетил-β-D-глюкозаминида) на лунку. После дополнительной инкубации еще в течение одного часа реакцию останавливали добавлением 100 мкл глицинового буфера (0,2 М глицин, 0,2% NaCl, pH 10,7) в каждую лунку. Флуоресценцию измеряли при λex:360/λem:465 в устройстве для регистрации флуоресценции в микропланшетах. Спонтанное высвобождение определяли в контрольных лунках, которые не подвергали лизису тритоном X-100. Специфичное высвобождение вычисляли с использованием формулы: (образец-спонтанное/общее-спонтанное) ×100.
Электронная микроскопия с иммунологическим мечением частицами золота
Сухие зерна пшеницы нарезали на небольшие кусочки (кубики размером примерно 0,5 мм), используя острое лезвие бритвы. Чтобы сохранить клетки в сухом состоянии, кубики фиксировали в безводных условиях в парах акролеина в течение 5 суток при комнатной температуре. Кубики переносили при комнатной температуре на 1 сутки в диметоксипропан (DMP) для удаления остаточной воды и заливали в смолу Lowicryl K4M, используя по восходящей серию смеси DMP:этанол и смеси этанол:мономерный Lowicryl K4M в качестве промежуточных стадий. Полимеризацию осуществляли при -35°C.
Нарезали ультратонкие срезы периферических и центральных тканей зерна и помещали на серебряную сетку для иммунологического мечения.
Мечение полученного из клона 10 аллергена осуществляли в камере с увлажнением при комнатной температуре (буфер PBS +1% (масс./об.) БСА, pH 7,4, трис-буфер +1% (масс./об.) БСА, pH 8,2) следующим образом: 1) 5% (масс./об.) БСА в буфере PBS, 15 минут; 2) антитела кролика против белка 10 пшеницы и антитела, полученные до иммунизации в разведении 1:35 в буфере PBS, 2 часа; 3) буфер PBS, 5 минут, трис-буфер, 2×5 минут; 4) антитела козы против IgG кролика, связанные с частицами коллоидного золота размером 10 нм (BioCell, Piano, Wetzlar, Germany) в разведении 1:20 в трис-буфере; 5) трис-буфер, 1×5 минут, дистиллированная вода, 2×5 минут.
Срезы окрашивали, используя уранилацетат (5 минут) и цитрат свинца (10 секунд).
Образцы анализировали в трансмиссионном электронном микроскопе EM 410 (FEI, Eindhoven, The Netherlands).
Результаты
Выделение и характеристика кДНК пшеницы, кодирующей аллерген подобный ингибитору сериновой протеиназы
Экспрессирующую библиотеку кДНК семян пшеницы подвергали скринингу с использованием IgE-антител четырех пациентов, страдающих астмой пекарей. Открытая рамка считывания кДНК IgE-реактивного клона 10 содержала 262 нуклеотидов, кодирующих 84-аминокислотный полипептид (фиг. 1). Молекулярную массу 9,4 кД и изоэлектрическую точку (pI) 6,08 вычислили на основании рассчитанной аминокислотной последовательности полученного из клона 10 аллергена. Анализ аминокислотного состава показал высокое содержание остатков валина (15,5%) и отсутствие остатков цистеина. Согласно компьютеризованному анализу вторичной структуры полученный из клона 10 аллерген, главным образом, состоит из случайных клубков и бета-слоев и одного альфа-спирального домена. Согласно расчетам доступности растворителям почти 80% аминокислот подвергаются воздействию растворителя. Поиск мотивов последовательности показал наличие одного потенциального участка фосфорилирования казеинкиназой II (аминокислота 32), двух N-концевых участков миристоилирования (аминокислоты 11, 55) и признак семейства ингибиторов I картофеля (фиг. 1: аминокислоты 25-36 указаны внутри прямоугольника). Сравнение полученной из клона 10 аминокислотной последовательности с последовательностями, депонированными в базе данных NCBI, показало, что аллерген почти идентичен предшественнику ингибитора WSCI субтилизина-химотрипсина Triticum aestivum (номер доступа gi 122065237) и ингибитору протеиназы WSCI T. aestivum (номер доступа gi 66356278) и проявляет значительную гомологию по последовательности с группой ингибиторов сериновых протеиназ, встречающихся у растений и животных. Такие ингибиторы сериновых протеиназ составляют семейство, названное семейством ингибиторов I картофеля, и характеризуются типичной консенсусной картиной последовательностей, которая консервативна у всех белков. Ингибиторы сериновых протеинах, относящиеся к семейству ингибиторов I картофеля, представляют собой небольшие белки из 60-90 аминокислот, в которых отсутствуют дисульфидные связи, и содержат только один сайт ингибирования. Последовательности ингибиторов сериновых протеиназ пшеницы имеют значительную степень консервативности последовательностей с ингибиторами протеиназ других однодольных растений, подобных ячменю (Hordeum vulgare), кукурузе (Zea mays, Zea diploperennis), трипсакуму (Tripsacum dactyloides) и рису (Oryza sativa), двудольных растений и червей (таблица III). В таблице III показаны проценты идентичности последовательностей между отдельными ингибиторами сериновых протеиназ.
Экспрессия, очистка и физико-химическая характеристика аллергена, подобного ингибиторам сериновых протеиназ
Полученный из клона 10 аллерген экспрессировали в E. coli BL21 (DE3) с C-концевой гексагистидиновой меткой. С использованием хроматографии на никеле из литра жидкой культуры можно очистить примерно 25 мг аллергена, подобного ингибиторам сериновых протеиназ (фиг. 2A). При MALDI-TOF-анализе очищенного рекомбинантного белка 10 получен пик массы 9970,8 Д (фиг. 2B). Спектр КД в дальнем ультрафиолете очищенного рекомбинантного аллергена, полученного из клона 10, (фиг. 2C) показал, что белок подвергается фолдингу и содержит значительное количество β-слоев и небольшое содержание α-спиралей. Спектр характеризуется минимумом при 204 нм и максимумом при 190 нм. При анализе вторичной структуры с использованием программы CDSSTR со ссылкой на набор данных 7 получили 8% α-спирали, 23% β-слоев, 14% β-поворотов и 53% случайных клубков. Значение NRMSD 0,033 показало хорошее соответствие между рассчитанными и экспериментально полученными спектрами. При нагревании до 95°C наблюдали небольшой сдвиг минимума КД-спектра (от 204 нм до 201 нм), свидетельствующий о частичной денатурации белка. При охлаждении до 25°C белок подвергался рефолдингу (фиг. 2C). Однако минимум при 204 нм был ниже, чем до нагревания, что свидетельствует о перестройке β-слоев. Итак, спектры, наблюдаемые при температурном сканировании, свидетельствуют о высокой температурной устойчивости полученного из клона 10 аллергена.
Рекомбинантный полученный из клона 10 аллерген представляет собой новый аллерген пшеницы, подобный ингибиторам сериновых протеиназ
Очищенный полученный из клона 10 аллерген тестировали в отношении IgE-реактивности, используя сыворотки пациентов, страдающих астмой пекарей, аллергией на пыльцу растений и пищевой аллергией на пшеницу, с помощью дот-блот-анализа (фиг. 3). Рекомбинантный полученный из клона 10 аллерген взаимодействовал с IgE-антителами 3 (№1, №4, №12) из 22 пациентов, страдающих астмой пекарей (13,6%). У пациента №1, который имел только низкие уровни IgE, специфичного к основному аллергену пшеницы, ингибитора альфа-амилазы, обнаружили высокую IgE-реактивность к аллергену, подобному ингибитору сериновых протеиназ. Интересно, что полученный из клона 10 аллерген узнавали только IgE-антитела пациентов с астмой пекарей, но не пациентов, страдающих от IgE-опосредованной пищевой аллергии на пшеницу, или пациентов, страдающих аллергией на пыльцу растений. Несколько сывороток из каждой из трех групп пациентов проявляли IgE-реактивность на рекомбинантные аллергены пыльцы тимофеевки луговой вследствие косенсибилизации на пыльцу растений. Специфичная IgE-реактивность на полученный из клона 10 аллерген у пациентов с астмой пекарей была подтверждена в анализе на чипах с использованием дополнительно 20 сывороток пациентов с целиакией, 119 пациентов с пищевой аллергией, 23 сывороток пациентов с аллергией на пыльцу растений и 25 пациентов с астмой пекарей (Constantin et al., не опубликовано, данные не показаны).
Анализ реактивности подклассов IgG на полученный из клона 10 аллерген в группе пациентов с астмой пекарей показал наличие специфичного для аллергена IgG1 и более низкие уровни специфичного для аллергена IgG4, при этом оба факта являются показателями Th2-ответа, тогда как существенной реактивности IgG2 и IgG3, специфичной для полученного из клона 10 аллергена не выявлено (фиг. 4).
Чтобы исследовать аллергическую активность IgE-антител, специфичных для аллергена, подобного ингибитору сериновых протеиназ, клетки RBL, экспрессирующие FcεRI, нагружали IgE сыворотки пациентов, имеющих и не имеющих специфичных IgE-антител, и затем подвергали воздействию аллергена (фиг. 5). В клетках RBL, нагруженных IgE сыворотки пациента №1, наблюдали наиболее сильную дегрануляцию при воздействии аллергена (51% общего высвобождения β-гексозаминидазы). Низкую дегрануляцию наблюдали в случае клеток RBL, которые были нагружены IgE пациентов №4 и №12 (22% и 19%, соответственно), что соответствовало интенсивности распознавания IgE на дот-блотах (фиг. 3). Почти не наблюдали дегрануляции в том случае, когда клетки RBL нагружали сывороткой человека, не страдающего аллергией (фиг. 5: NC). Основной аллерген пыльцы тимофеевки луговой rPh1 p 1 индуцировал сильную дегрануляцию клеток RBL, нагруженных IgE из сыворотки пациента № 12, и умеренную дегрануляцию в том случае, когда клетки RBL нагружали сывороткой пациентов №1 и №4 (фиг. 5). Действительно, пациенты №1, №4 и №12 также страдали аллергией на пыльцу растений (таблица I).
Аллерген, подобный ингибитору сериновых протеиназ, накапливается в семенах пшеницы по время созревания
Кроличьи антитела, специфичные для полученного из клона 10 аллергена, использовали для исследования экспрессии белка во время созревания семян пшеницы (фиг. 6). Нитроцеллюлозные листы, содержащие экстракты семян пшеницы, собранных в разных временных точках созревания семян, исследовали, используя в качестве зондов специфичные кроличьи антитела и Ig, полученный до иммунизации. Специфичные для клона 10 антитела взаимодействовали с белком 40 кД, представляющим собой тетрамер аллергенов, подобных ингибиторам сериновых протеиназ (фиг. 6) (11). Экспрессия белка становилась регистрируемой в 15-дневных семенах и продолжала возрастать в ходе дальнейшего созревания семян (фиг. 6). Иммунореактивность не выявляли, когда блоты инкубировали с Ig, полученными до иммунизации из того же кролика (фиг. 6).
Аллерген, подобный ингибитору сериновых протеиназ, преимущественно выявляется в семенах пшеницы.
Когда авторы сравнили пыльцу и семена, они обнаружили, что аллерген, подобный ингибитору сериновых протеиназ, преимущественно экспрессировался в семенах (фиг. 7), тогда как только слабый сигнал в области примерно 65 кД был получен в случае экстракта пыльцы пшеницы. Паналлерген профилин был выявлен с использованием антител кролика против профилина семян пшеницы в семенах и пыльце пшеницы (фиг. 7: №123). Не обнаружено реактивности в случае использования Ig, полученного до иммунизации (фиг. 7).
Затем авторы использовали антитела, специфичные к аллергену, подобному ингибитору сериновых протеиназ, для поиска перекрестно реагирующих структур риса, кукурузы, фасоли обыкновенной и картофеля, для которых были описаны гомологичные белки с идентичностью последовательностей 50% (фасоль), 49% (кукуруза, рис) и 33% (картофель) (таблица III).
Аллерген, подобный ингибитору сериновых протеиназ снова выявляли в виде тетрамера в семенах пшеницы, полосу в области примерно 23 кД выявляли в рисе, но не выявлено реактивности в кукурузе, фасоли обыкновенной или картофеле (фиг. 8C), несмотря на то, что сравнимые количества каждого экстракта подвергали SDS-ПААГ (фиг. 8A). Реактивности не наблюдали в том случае, когда блоты инкубировали с Ig, полученным до иммунизации (фиг. 8B).
Локализация аллергена, подобного ингибитору сериновых протеиназ, в алейроновом слое и между крахмальными гранулами зерна пшеницы с помощью электронной микроскопии с иммунологическим мечением частицами золота
На фигуре 9A показан ультратонкий срез зерна пшеницы при небольшом увеличении в трансмиссионном электронном микроскопе. Видны три основных морфологических компонента зерна: наружная многослойная плодовая и семенная оболочка (C), алейроновый слой (AL) и начало объемного внутреннего содержимого зерна, крахмалистого эндосперма (SE). Прямоугольником отмечена область, сопоставимая с областью, показанной на фигуре B. На фигуре 9B показана граница между алейроновой клеткой и примыкающим крахмалистым эндоспермом при большем увеличении. Алейроновая клетка заполнена алейроновыми зернами (AG) (белковыми вакуолями), которые окружены небольшими липидными везикулами (L). Оба компонента погружены в цитоплазматический матрикс клетки. Крахмалистый эндосперм состоит из гранул крахмала разного размера, плотно упакованных так, что остаются лишь небольшие промежутки, заполненные аморфным цитоплазматическим веществом. Прямоугольниками указаны области, показанные при большом увеличении на фигурах C, D и E, F, соответственно. На фигуре 9C показана локализация полученного из клона 10 аллергена в алейроновой клетке с использованием Ат, полученных против белка 10 пшеницы. Частицы золота (стрелки) свидетельствуют о присутствии белка 10 пшеницы преимущественно в цитоплазматическом матриксе между органоидами клетки, а также в периферических частях липидных везикул (L). В крахмалистой области белок 10 пшеницы ассоциирован с аморфным цитоплазматическим веществом (CY) между гранулами крахмала (SG) (фигура 9E). Контрольные эксперименты с использованием Ат, полученных до иммунизации, показали очень низкую степень неспецифичного мечения (фиг. 9, D и F).
ПРИМЕР 2: Экспрессия и очистка рекомбинантных аллергенов
В примере 2 показана идентификация и характеристика шести реактивных по отношению к IgE аллергенов семян пшеницы, названных клонами №10, №37, №38, №112, №123 и №126. Кроме того, в примере 2 показан способ экспрессии и очистки рекомбинантных аллергенов указанных клонов.
Биологические материалы, сыворотки пациентов
Семена пшеницы Triticum aestivum сорта Michael получали из Osterreichische Agentur fur Gesundheit und Ernahrungssicherheit GmbH и высевали в теплицу. Незрелые семена собирали непосредственно в жидкий азот через 25 дней после начала опыления и хранили при -80°C вплоть до использования. Сыворотки получали от 24 пациентов, страдающих астмой пекарей (таблица IV). Астму пекарей диагностировали на основе истории болезни, общих уровней IgE в сыворотке, IgE, специфичного для пшеницы и ржи, выявляемого с помощью системы CAP-FEIA (Phadia, Uppsala, Sweden) и после специальных провокационных ингаляционных тестов (1) (фиг. 10). Сыворотку пациентов с аллергией на пыльцу растений анализировали в отношении общих уровней IgE в сыворотке и IgE, специфичного для пыльцы тимофеевки луговой, с использованием системы CAP-FEIA (Phadia), и пациентов с пищевой аллергией на пшеницу характеризовали, как описано ранее (2) (фиг. 11). Клинические данные для таких пациентов суммированы в таблице V. В целях контроля в эксперименты включали сыворотку человека, не страдающего аллергией.
Конструирование библиотеки кДНК λgt11 из семян пшеницы
Суммарную РНК экстрагировали из семян пшеницы, которые хранились при -80°C, согласно способу Yeh [3]. Затем осадок РНК растворяли в буфере, содержащем изотиоцианат гуанидиния (4М изотиоцианат гуанидиния, 0,83% об./об. 3М ацетат натрия, pH 6, 11 мМ β-меркаптоэтанол), и очищали ультрацентрифугированием в градиенте плотности хлорида цезия. Поли-A+-РНК выделяли аффинной хроматографией на олиго-dT-целлюлозе (Nucleo Trap mRNA; Machery-Nagel) и синтезировали двунитевую кДНК, используя набор для синтеза кДНК (Система для синтеза кДНК; Roche Diagnostics, Mannheim, Germany). После метилирования EcoRI-метилазой (New England Biolabs, Beverly, MA), к кДНК добавляли EcoRI-линкеры (New England Biolabs). Связанную линкерами кДНК расщепляли EcoRI (Roche Diagnostics). Отщепленные линкеры удаляли на колонке Nick (Pharmacia Biotech, Uppsala, Sweden) и кДНК лигировали в плечи λgt11 (Stratagene, La Jolla, CA, USA). Продукт лигирования подвергали упаковке in vitro (набор для клонирования Gigapack III Gold, Stratagene), получая экспрессирующую библиотеку кДНК в λgt11 с 2,43×106 БОЕ (бляшкообразующих единиц).
Выделение и характеристика IgE-реактивных клонов из библиотеки кДНК семян пшеницы
E. coli Y1090 инфицировали 7×105 БОЕ рекомбинантных фагов и подвергали иммунологическому скринингу с использованием IgE сыворотки четырех пациентов (№1, №2, №5, №13), страдающих астмой пекарей, как описано [5]. Шесть IgE-реактивных фаговых клонов отбирали для дальнейшего повторного клонирования и их ДНК амплифицировали в ПЦР, используя смесь для ПЦР Platinum PCR Supermix (Invitrogen, Life Technolgies) с праймерами λgt11 и секвенировали (MWG, Ebersberg, Germany). Полученные последовательности сравнивали с последовательностями, представленными в базе данных GenBank в Национальном центре биотехнологической информации (NCBI).
Экспрессия и очистка рекомбинантных аллергенов
Кодирующую область клонов амплифицировали в ПЦР, используя праймеры, перечисленные в таблице VI. Продукты ПЦР субклонировали в векторе AccepTor (Novagen, Madison, WI) и снова секвенировали (MWG). Затем содержащие вставки векторы AccepTor расщепляли, используя NdeI и EcoRI (Roche Diagnostics), вставки очищали в геле (Promega, Madison, WI, USA) и субклонировали в экспрессирующей плазмиде pET 17b (Novagen). Последовательности ДНК подтверждали секвенированием обеих нитей ДНК (Microsynth, Balgach, CH). Конструкциями со вставками pET 17b трансформировали клетки E. coli BL21 (DE3) (Stratagene) и выращивали в среде Луриа (17), содержащей 100 мг/л ампициллина, при 37°C до OD (600 нм) 0,8-1. Экспрессию белка индуцировали добавлением изопропил-β-D-тиогалактопиранозида (IPTG) до конечной концентрации 0,5 мМ и выращивали бактерии еще в течение 3 часов. Затем бактерии собирали центрифугированием и гомогенизировали в 25 мМ имидазоле, pH 7,5, 0,1% (об./об.) тритоне X-100, используя Ultraturrax (IKA, Stauffen, Germany). ДНК расщепляли, добавляя ДНКазу I, дополнительно перемешивали еще в течение 10 минут при 20°C, реакцию останавливали, используя 200 мкл 5 М NaCl, и затем центрифугировали при 4°C (6000×g, 20 минут). Полученный из клона 10 аллерген очищали из содержащего тельца включения осадка в денатурирующих условиях, используя аффинные колонки с Ni-NTA-смолой, согласно руководству QIAexpressionist, протокол 17 (QIAGEN, Hilden, Germany). Фракции, содержащие рекомбинантный аллерген, объединяли и диализовали против 10 мМ NaH2PO4, pH 7,5. Рекомбинантные белки №37, №38, №112, №123 и №126 очищали из осадка лизата бактериальных клеток в обычных условиях на аффинных колонках со смолой Ni-NTA согласно руководству QIAexpressionist, протокол 12 (QIAGEN). Фракции, содержащие рекомбинантные белки, объединяли и диализовали против 10 мМ NaH2PO4 pH 7,5. Образцы белка анализировали в отношении чистоты в 14% SDS-ПААГ-геле и визуализировали, используя окрашивание Кумасси бриллиантовым синим. Концентрации белка определяли с использованием набора для анализа белков Micro BCA (Pierce, Rockford, IL).
ПРИМЕР 3: Матрица аллергенов
В примере 3 показана матрица аллергенов, с помощью которой обнаружено, что рекомбинантные аллергены семян пшеницы, полученные как описано в примере 2, специфично узнаются IgE сыворотки пациентов с астмой пекарей, но не пациентов с пищевой аллергией на пшеницу или аллергией на пыльцу растений. Следовательно, рекомбинантные аллергены специфичны для респираторной аллергии на пшеничную муку. Кроме того, в примере 3 показано применение маркерных аллергенов пыльцы тимофеевки луговой ph1 p 1 и Ph1 p 5 и пыльцы пшеницы для матрицы с целью усовершенствования диагностики респираторной аллергии in vitro.
Методика
Пациенты и сыворотки
Сыворотки получали от пациентов испанцев, страдающих астмой пекарей (B 1-B23), австрийцев (F1-F26) и немцев (F27-F38), страдающих индуцированной пшеницей пищевой аллергией и пациентов австрийцев (G1-G17), страдающих аллергией на пыльцу растений. Пациентов отбирали на основании позитивной истории болезни, специальных ингаляционных провокационных тестов для подтверждения клинически значимой сенсибилизации в случае астмы пекарей (1) и двойной слепой плацебо-контролируемой пищевой провокации (DBPCFC) у детей с пищевой аллергией на пшеницу. Сыворотки всех пациентов анализировали в отношении суммарных уровней IgE в сыворотке и IgE, специфичного для пшеничной муки и пыльцы тимофеевки луговой, используя систему CAP-FEIA (Phadia, Uppsala, Sweden). Демографические, клинические и серологические данные таких пациентов суммированы в таблице VII, VIII и IX. В целях контроля во все эксперименты включали сыворотки здоровых людей.
Биологические материалы
Рекомбинантные белки пшеницы №10, №37, №38, №112, №123 и №126, полученные из библиотеки кДНК с помощью скрининга с использованием сывороток пациентов с астмой пекарей, экспрессировали в E. coli и очищали как описано (12). Пыльцу пшеницы приобретали из Allergon (Vallinge, Sweden), а рекомбинантные Ph1 p 1, Ph1 p 5, Ph1 p 7, Ph1 p 12 из BIOMAY (Vienna, Austria).
Экстракт белков
Пыльцу Triticum aestivum (500 мг) экстрагировали при 4°C в течение ночи в 5 мл PBS, 2 мМ EDTA, 1 мМ PMSF. После центрифугирования в течение 1 часа при 13000×g и 4°C, концентрацию белка в надосадке определяли с использованием набора для анализа белков BCA (Pierce, Rockford, IL) и аликвоты хранили при -20°C вплоть до использования.
Анализ матрицы аллергенов
Экстракт пыльцы пшеницы, 1-1,5 нг/пятно, рекомбинантные и очищенные белки пшеницы и рекомбинантные аллергены пыльцы растений, 0,1-0,15 нг/пятно, наносили в виде пятен с использованием нано-плоттера NP2 (Gesellschaft fur Silizium-Mikrosysteme mbH, Groβerkmannsdorf, Germany) на нитроцеллюлозные мембраны, которые закрепляли на предметных стеклах для микроскопа как описано (13). Очищенные IgE человека использовали в качестве маркера положения (14). Микроматрицы с нанесенными пятнами предварительно промывали 30 мкл буфера для анализа (слабый фосфатный буфер, pH 7,5) и инкубировали с 30 мкл неразбавленной сыворотки пациентов с аллергией или контролями. После промывки 30 мкл буфера для анализа связанные IgE-антитела выявляли, используя 20 мкл конъюгированного с флуорофором анти-IgE-антитела, и интенсивности флуоресценции (FI) измеряли при длине волны 635 нм (GenePix 4000B fran Axon). Уровень отсечения устанавливали на FI=300 на основе значений, полученных с сывороточным альбумином человека.
Результаты
Описание пациентов
Авторы анализировали пациентов, страдающих астмой пекарей, индуцированной пшеницей пищевой аллергией или аллергией на пыльцу растений. Группа пациентов с астмой пекарей состояла из 23 человек (4 женщины, 19 мужчин: средний возраст 39 лет, диапазон 22-60 лет), по роду профессии, подвергающихся воздействию пшеничной муки. Девяносто один процент пациентов с астмой пекарей страдали астмой вследствие вдыхания пшеничной муки и 94% жаловались на симптомы риноконъюнктивита. Сенсибилизация к другим источникам респираторных аллергенов (например, кошка, таракан, собака, пыльца растений, лошадь, клеши домашней пыли, плесневые грибы и/или пыльца оливы) обнаружена у 70% пациентов с астмой пекарей и 48% пациентов, страдающих аллергией на пыльцу растений (таблица VII). Интересно, что ни у одного из пациентов с астмой пекарей не проявлялась аллергия на пищу, и симптомы были ограничены респираторными проявлениями.
Группа пациентов, страдающих индуцированной пшеницей пищевой аллергией, состояла из 38 человек (25 женского пола, 13 мужского пола: средний возраст 13 лет, диапазон 0,5-65 лет) (таблица VIII). Авторы снова обнаружили, что 71% были сенсибилизированы к другим источникам респираторных аллергенов (например, пыльце березы, пыльце злаковых трав, клещам домашней пыли и пыльце полыни), и 58% также были сенсибилизированы на пищевые аллергены (например, морковь, коровье молоко, фундук, куриное яйцо, солод, орехи, апельсин, слива, рис, соя, сельдерей, морепродукты, креветки и/или специи) (таблица VIII). IgE-опосредованная сенсибилизация на пыльцу растений обнаружена у 55% таких пациентов. Симптомы у таких пациентов варьировали от респираторных симптомов (например, астма, бронхит, кашель, конъюнктивит, диспноэ, закупорка носового канала, ринорея, риноконъюнктивит) до желудочно-кишечных симптомов (например, боль в животе, диарея, задержка газов, боль в горле и рвота) и кожных симптомов (например, экзема, зуд и крапивница) (таблица VIII).
Группа пациентов с аллергией на пыльцу растений состояла из 17 пациентов (5 женского пола, 12 мужского пола: средний возраст 13 лет, диапазон 0,5-65 лет). Семьдесят один процент таких пациентов также страдали аллергией на другие источники респираторных аллергенов (например, пыльцу березы, кошек, собак, клещей домашней пыли, кроликов и пыльцу полыни) (таблица IX). Согласно серологии у 65% наблюдали IgE-реактивность на пшеничную муку, и 23% имели IgE против других пищевых аллергенов. Однако только один пациент страдал от крапивницы, и у таких пациентов не регистрировали симптомов пищевой аллергии. Респираторные симптомы, такие как астма, конъюнктивит, диспноэ, риноконъюнктивит и ринит преобладали у пациентов с аллергией на пыльцу растений.
Композиция матрицы аллергенов
Шесть рекомбинантных аллергенов семян пшеницы, названных №10, №37, №38, №112, №123 и №126, в виде пятен наносили на покрытые нитроцеллюлозой предметные стекла (фиг. 12, A). Рекомбинантные белки пшеницы экспрессировали в Escherichia coli на основе кДНК, которые были выделены из библиотеки кДНК семян пшеницы с использованием сывороток пациентов с астмой пекарей, как описано (15). В соответствии с гомологией с последовательностями, депонированными в базе данных NCBI, аллергены пшеницы могут быть описаны следующим образом: №10 имел 96% гомологию предшественником ингибитора WSCI субтилизина-химотрипсина Triticum aestivum (номер доступа gi 122065237), №37 представлял собой тиоредоксин H T. aestivum (номер доступа gi 27461 140), №38 представлял собой глутатионтрансферазу T. aestivum (номер доступа gi 20067419), №112 имел 99% гомологию с 1-Cys-пероксиредоксином T. aestivum (номер доступа gi 34539782), №123 имеет 96% гомологию с профилином T. aestivum (номер доступа gi 1346803) и №126 имеет 69% гомологию с дегидрином 11 Hordeum vulgare (номер доступа gi 4105101).
Кроме рекомбинантных аллергенов пшеницы экстракт пыльцы пшеницы, рекомбинантные аллергены пыльцы тимофеевки луговой (rPh1 p 1, rPh1 p 5, rPh1 p 7 и rPh1 p 12) (16-19) и IgE также наносили в виде пятен на предметные стекла (фиг. 12, A). Типичные изображения микроматриц аллергенов после инкубации с сыворотками показаны на фиг. 12, B-C. На чипе, который анализировали с использованием в качестве зонда сыворотки человека, не страдающего аллергией, не выявляли реактивности ни к одному из аллергенов и выявляли только нанесенные в виде пятен маркеры IgE на основе анти-IgE-конъюгата (фиг. 12, B). На фигуре 12C показаны типичные изображения микроматриц на чипах, инкубированных с сыворотками пациента с астмой пекарей (1), пациента, страдающего индуцированной пшеницей пищевой аллергией (2) и пациента с аллергией на пыльцу растений (3). У пациента с астмой пекарей (1: таблица I: B4) показана высокая IgE-реактивность на рекомбинантные аллергены пшеницы №10 и слабая реактивность на №126 и №112. В случае пациента с пищевой аллергией (2: таблица VIII: F26) наблюдали сильное связывание IgE с рекомбинантным профилином №123 пшеницы и rPh1 p 12 пыльцы тимофеевки луговой и слабое связывание с экстрактом пыльцы пшеницы и rPh1 p 1. На изображении чипа для пациента с аллергией на пыльцу растений (3: G 16) видны сильные сигналы на rPh1 p 5 и rPh1 p 12 и более слабые сигналы на профилин №123 пшеницы, экстракт пыльцы пшеницы и rPh1 p 1.
Идентификация аллергенов семян пшеницы, специфично узнаваемых IgE-антителами пациентом с астмой пекарей
Девяносто один процент пациентов с астмой пекарей были позитивными в CAP на пшеничную муку (таблица VII). Используя нанесенные в виде пятен аллергены пшеницы можно установить профиль IgE-реактивности для 62% пациентов с астмой пекарей. Аллергены №126 (30%), №10 (26%), №123 (22%) и №122 (17%) были наиболее часто выявляемыми компонентами, тогда как №37 и №38 взаимодействовали только с 4% сывороток (фиг. 13, A). Интересно, что два пациента, которые были оценены негативно в CAP на пшеничную муку (B 5, B 18: таблица VII), реагировали на аллерген №126. Сорок восемь процентов пациентов с астмой пекарей проявляли IgE-реактивность на экстракт пыльцы тимофеевки луговой, и каждого из таких пациентов диагностировали с использованием сочетания пятен rPh1 p 1 и rPh1 p 5. Рекомбинантный Ph1 p 12 (35%) всегда был более сильным и более часто узнаваемым, чем профилин пшеницы №123 (22%). Перекрестно реактивный аллерген пыльцы rPh1 p 7 взаимодействовал с 13% сывороток.
Рекомбинантные аллергены семян пшеницы, распознаваемые пациентами с астмой пекарей, не являются мишенями для IgE-антител пациентов, страдающих индуцированной пшеницей пищевой аллергией
Все пациенты с пищевой аллергией на пшеницу были позитивными в CAP в отношении пшеничной муки, но почти не распознавали рекомбинантные белки пшеницы (5%) (фиг. 13, B). Два пациента, которые был позитивными в отношении перекрестно реактивного профилина пыльцы тимофеевки луговой, также имели положительный сигнал на профилин пшеницы №123 на микроматрице. Пятьдесят пять процентов пациентов с индуцированной пшеницей пищевой аллергией, имели IgE-реактивность в CAP в отношении Phleum, но только 18% проявляли реактивность на экстракт пыльцы пшеницы на чипе, и 26% на сочетание пятен rPh1 p 1 и rPh1 p 5. Нанесенные в виде пятен rPh1 p 1 и rPh1 p 5 отдельно распознавались 18% и 16% сывороток, соответственно, перекрестно реактивные аллергены rPh1 p 7 и rPh1 p 12 взаимодействовали с 13% и 26% сывороток.
rPh1 p 1 и rPh1 p 5 являются диагностическими маркерными аллергенами в случае аллергии на пыльцу растений, тогда как профилин является узнаваемым пациентами с астмой пекарей и пищевой аллергией на пшеницу
Шестьдесят пять процентов пациентов с аллергией на пыльцу растений были позитивными в CAP в отношении пшеничной муки. Из рекомбинантных белков пшеницы, тестированных на микроматрице, только рекомбинантный белок пшеницы №123 был узнаваемым 23,5% пациентов (фиг. 13, C). Каждый из таких пациентов также был позитивным на перекрестно реактивный профилин пыльцы тимофеевки луговой rPh1 p 12. Но rPh1 p 12 всегда был сильнее и чаще узнаваемым (35%), чем №123. Все пациенты, страдающие аллергией на пыльцу растений, были позитивными в CAP в отношении пальцы тимофеевки луговой и в отношении сочетания нанесенных в виде пятен rPh1 p 1 и rPh1 p 5. Рекомбинантный Ph1 p 1 отдельно был более часто узнаваемым (94%), чем rPh1 p 5 (88%) пациентами с аллергией на пыльцу растений. Экстракт пыльцы пшеницы взаимодействовал с 82% сывороток, а перекрестно реактивный rPh1 p 7 взаимодействовал с 29% сывороток. Все пациенты с аллергией на пыльцу растений и пациенты с астмой пекарей и аллергией на пыльцу растений были позитивными в отношении сочетания rPh1 p 1 и rPh1 p 5 на микроматрице и в CAP в отношении Phleum (фиг. 13, A; C). Полученные данные согласуются с ранее опубликованными данными, показывающими, что аллергия на пыльцу растений может быть диагностирована по IgE-реактивности на rPh1 p 1 и rPh1 p 5 (20, 21). Однако среди пациентов с индуцированной пшеницей пищевой аллергией только 26% реагировало на rPh1 p 1 и rPh1 p 5 в анализе на микроматрице, но более чем в два раза больше (55%) были позитивными в CAP в отношении Phleum (фиг. 13, B). Кроме того, пациенты, реагирующие на экстракт пыльцы пшеницы в системе CAP и профилин пыльцы тимофеевки луговой (rPh1 p 12) на микроматрице, также были позитивными в отношении профилина пшеницы (22) вследствие перекрестной реактивности профилинов растений (23). Тем не менее, частота и интенсивность узнавания IgE всегда была сильнее на rPh1 p 12, чем на профилин пшеницы (22).
ПРИМЕР 4
Аналит иммобилизовали на твердой подложке, такой как ImmunoCAP (Phadia, Uppsala, Sweden). Образцы сыворотки, по меньшей мере, трех типичных пациентов, сенсибилизированных на аллерген и проявляющих IgE-реактивность на такой аллерген, инкубировали в течение 3 часов при комнатной температуре с аллергеном с конечной концентрации 100 мкг/мл и параллельно в качестве негативных контролей отдельно с буфером и с неаллергенным белком, связывающим мальтозу (MBP), E. coli. Затем образцы анализировали в отношении связывания IgE с использованием тестов ImmunoCAP (Phadia, Uppsala, Sweden), несущих иммобилизованный аналит, чтобы исследовать вызывает ли предварительная инкубация с аллергеном специфичное ингибирование или значительное снижение связывания IgE.
Демографические, клинические и серологические характеристики пациентов, страдающих астмой пекарей
Демографические, клинические и серологические характеристики пациентов, страдающих пищевой аллергией на пшеницу и аллергией на пыльцу растений
-: негативный; n.d.: не определяли; кЕд. А/л: килоединиц антигена на литр.

Пациенты с пищевой аллергией и аллергией на пыльцу растений
Праймеры ПЦР, используемые для амплификации кДНК
Демографические, клинические и серологические характеристики пациентов, страдающих астмой пекарей
мы
мг/мл
цы,
мг/кг
Демографические, клинические и серологические характеристики пациентов, страдающих пищевой аллергией
онный тест
Демографические, клинические и серологические характеристики пациентов, страдающих аллергией на пыльцу растений
Ссылки
1. Quirce, S., M. Fernandez-Nieto, C. Escudero, J. Cuesta, M. de Las Heras, and J. Sastre. 2006. Bronchial responsiveness to bakery-derived allergens is strongly dependent on specific skin sensitivity. Allergy 61:1202-1208.
2. Constantin, C, W. D. Huber, G. Granditsch, M. Weghofer, and R.Valenta. 2005. Different profiles of wheat antigens are recognised by patients suffering from coeliac disease and IgE-mediated food allergy. Int Arch Allergy Immunol 138:257-266.
3. Yeh, K., R. Juang, and J. Su. 1991. A rapid and efficient method for a RNA isolation from plants with high carbohydrate content. Focus 13:102-103.
4. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular Cloning: A laboratory Manual. Cold Spring Harbor Laboratory Press.
5. Breiteneder, H., K. Pettenburger, A. Bito, R. Valenta, D. Kraft, H.Rumpold, O. Scheiner, and M. Breitenbach. 1989. The gene coding for the major birch pollen allergen Betvl, is highly homologous to a pea disease resistance response gene. Embo J8:1935-1938.
6. Stern, D. A., J. Riedler, D. Nowak, C. Braun-Fahrlander, I. Swoboda, N.Balic, K. W. Chen, S. Vrtala, H. Gronlund, M. van Hage, R. Valenta, S.Spitzauer, E. Von Mutius, and D. Vercelli. 2007. Exposure to a farming environment has allergen-specific protective effects on TH2-dependent isotype switching in response to common inhalants. J Allergy Clin Immunol 119:351-358.
7. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685.
8. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. ProcNatlAcad Sci US A 76:4350-4354.
9. Whitmore, L., and B. A. Wallace. 2004. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res 32:W668-673.
10. Kaul, S., D. Luttkopf, B. Kastner, L. Vogel, G. Holtz, S. Vieths, and A. Hoffmann. 2007. Mediator release assays based on human or murine immunoglobulin E in allergen standardization. Clin Bxp Allergy 37:141-150.
11. Richardson, M., and L. Cossins. 1974. Chymotryptic inhibitor I from potatoes: the amino acid sequences of subunits B, C, and D. FEBS Lett 45:11-13.
12. Vrtala S, Fischer S, Grote M, Vangelista L, Pastore A, Sperr WR, et al. 1999. Molecular, immunological, and structural characterization of Phi p6, a major allergen and P-particle-associated protein from Timothy grass (Phleum pratense) pollen. J Immunol 163:5489-96.
13. Nystrand M. 2006. A multiplexed immunoassay for the rapid detection of specific IgE in allergy diagnosis. TVDT 2006:61.
14. Nilsson K, Bennich H, Johansson SG, Ponten J. 1970. Established immunoglobulin producing myeloma (IgE) and lymphoblastoid (IgG) cell lines from an IgE myeloma patient. Clin Exp Immunol 7:477-89.
15. Constantin C, Quirce S, Grote M, Touraev A, Swoboda I, Stoecklinger A, et al. Molecular and immunological characterization of a wheat serine proteinase-inhibitor as a novel allergen in baker's asthma. Unpublished.
16. Laffer S, Valenta R, Vrtala S, Susani M, van Ree R, Kraft D, et al. 1994. Complementary DNA cloning of the major allergen Phi p I from timothy grass (Phleum pratense); recombinant Phi p I inhibits IgE binding to group I allergens from eight different grass species. J Allergy Clin Immunol 94:689-98.
17. Vrtala S, Sperr WR, Reimitzer I, van Ree R, Laffer S, Muller WD, et al. 1993. cDNA cloning of a major allergen from timothy grass (Phleumpratense) pollen; characterization of the recombinant Phi pV allergen. J Immunol 151:4773-81.
18. Niederberger V, Hayek B, Vrtala S, Laffer S, Twardosz A, Vangelista L, et al. 1999, Calcium-dependent immunoglobulin E recognition of the apo-and calcium-bound form of a cross-reactive two EF-hand timothy grass pollen allergen, Phi p 7. Faseb J 13:843-56.
19. Valenta R, Ball T, Vrtala S, Duchene M, Kraft D, Scheiner O. 1994. cDNA cloning and expression of timothy grass (Phleum pratense) pollen profiling in Escherichia coli: comparison with birch pollen profilin. Biochem BiophysRes Commun 199:106-18.
20. Kazemi-Shirazi L, Niederberger V, Linhart B, Lidholm J, Kraft D, Valenta R. 2002. Recombinant marker allergens: diagnostic gatekeepers for the treatment of allergy. Int Arch Allergy Immunol 127:259-68.
21. Andersson K, Lidholm J. 2003. Characteristics and immunobiology of grass pollen allergens. Int Arch Allergy Immunol 130:87-107.
22. Sampson HA. 1999. Food allergy. Part 2: diagnosis and management. J Allergy CUn Immunol 103:981-9.
23. Radauer C, Willerroider M, Fuchs H, Hoffmann-Sommergruber K, Thalhamer J, Ferreira F, et al. 2006. Cross-reactive and species-specific immunoglobulin E epitopes of plant profilins: an experimental and structure-based analysis. Clin Exp Allergy 36:920-9.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ ОСНОВНЫХ АЛЛЕРГЕНОВ ГРУППЫ 4 ИЗ ЗЛАКОВЫХ | 2004 |
|
RU2373283C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И ПОЛУЧЕНИЕ АЛЛЕРГЕНА ПЫЛЬЦЫ ТРАВ Phl p 4 С ПОМОЩЬЮ РЕКОМБИНАНТНЫХ СПОСОБОВ | 2003 |
|
RU2327739C2 |
| ГИБРИДНЫЕ БЕЛКИ-НОСИТЕЛИ ПЕПТИДОВ В КАЧЕСТВЕ ВАКЦИН ОТ АЛЛЕРГИИ | 2012 |
|
RU2630652C2 |
| ВАРИАНТ АЛЛЕРГЕНА ГРУППЫ I ИЗ Роасеае, ХАРАКТЕРИЗУЮЩИЙСЯ СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ И СОХРАНЕННОЙ Т-КЛЕТОЧНОЙ РЕАКТИВНОСТЬЮ (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО МОЛЕКУЛА ДНК И ЕГО ПРИМЕНЕНИЕ | 2005 |
|
RU2409589C2 |
| АЛЛЕРГЕНЫ, ПОЛУЧЕННЫЕ РЕКОМБИНАНТНЫМИ СПОСОБАМИ | 2010 |
|
RU2592674C2 |
| КЛЕЩЕВОЙ АЛЛЕРГЕН ДОМАШНЕЙ ПЫЛИ | 2007 |
|
RU2570563C2 |
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2658767C1 |
| Рекомбинантный полипептид на основе аллергена пыльцы березы и аллергена яблока в качестве вакцины от аллергии | 2020 |
|
RU2761431C1 |
| ГИПОАЛЛЕРГИЧЕСКИЕ ВАРИАНТЫ Mal d 1, ГЛАВНОГО АЛЛЕРГЕНА Malus domectica | 2012 |
|
RU2624030C2 |
| ВАРИАНТЫ ГРУППЫ 5 АЛЛЕРГЕНОВ ЗЛАКОВЫХ СО СНИЖЕННОЙ АЛЛЕРГЕННОСТЬЮ ВСЛЕДСТВИЕ МУТАГЕНЕЗА ОСТАТКОВ ПРОЛИНА | 2010 |
|
RU2575606C2 |











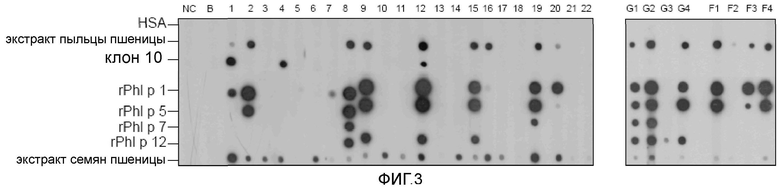
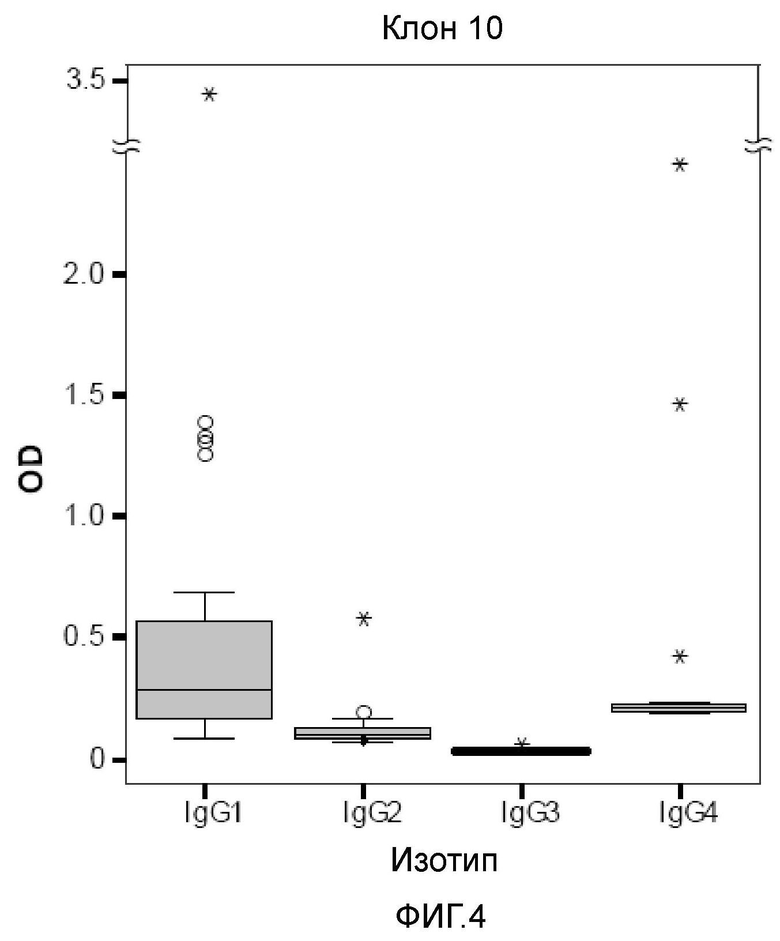
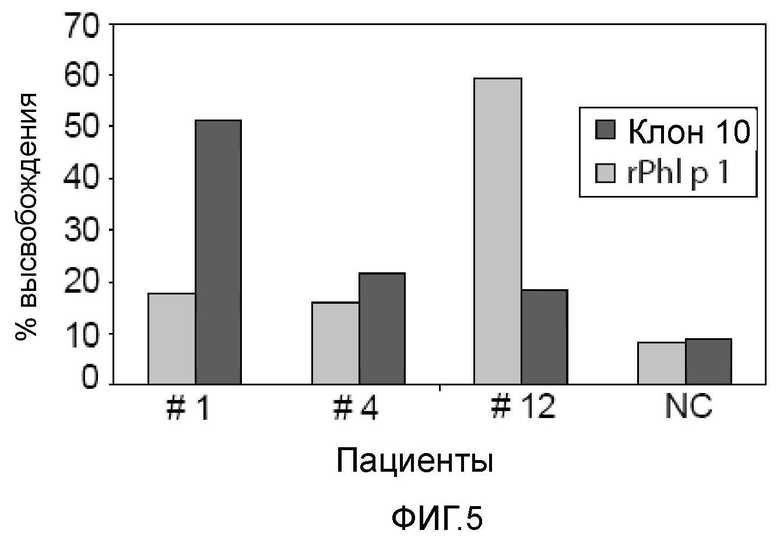
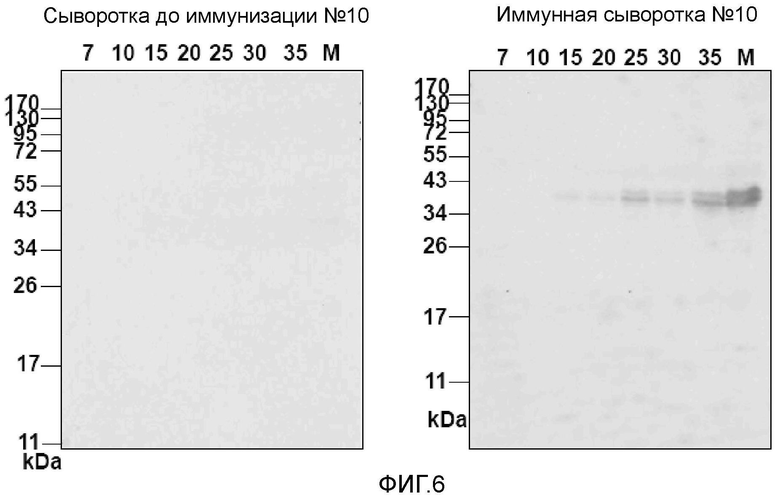

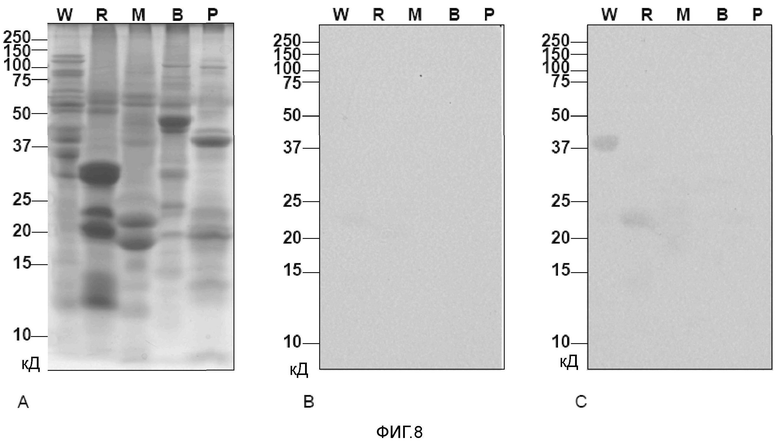

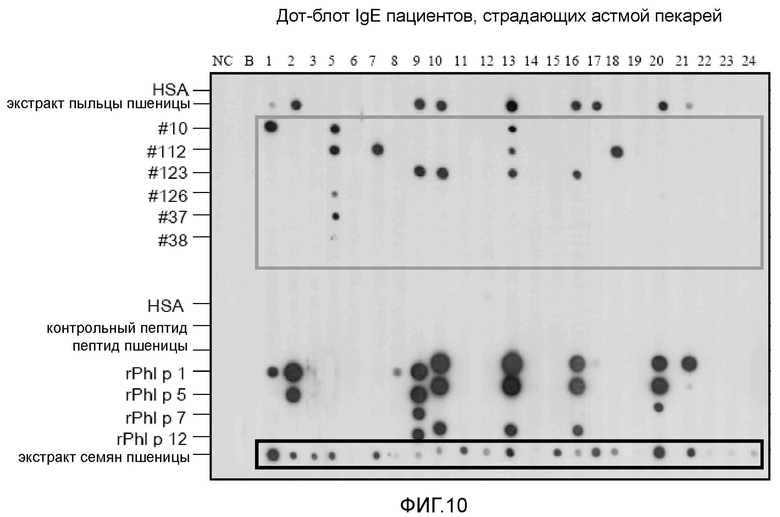

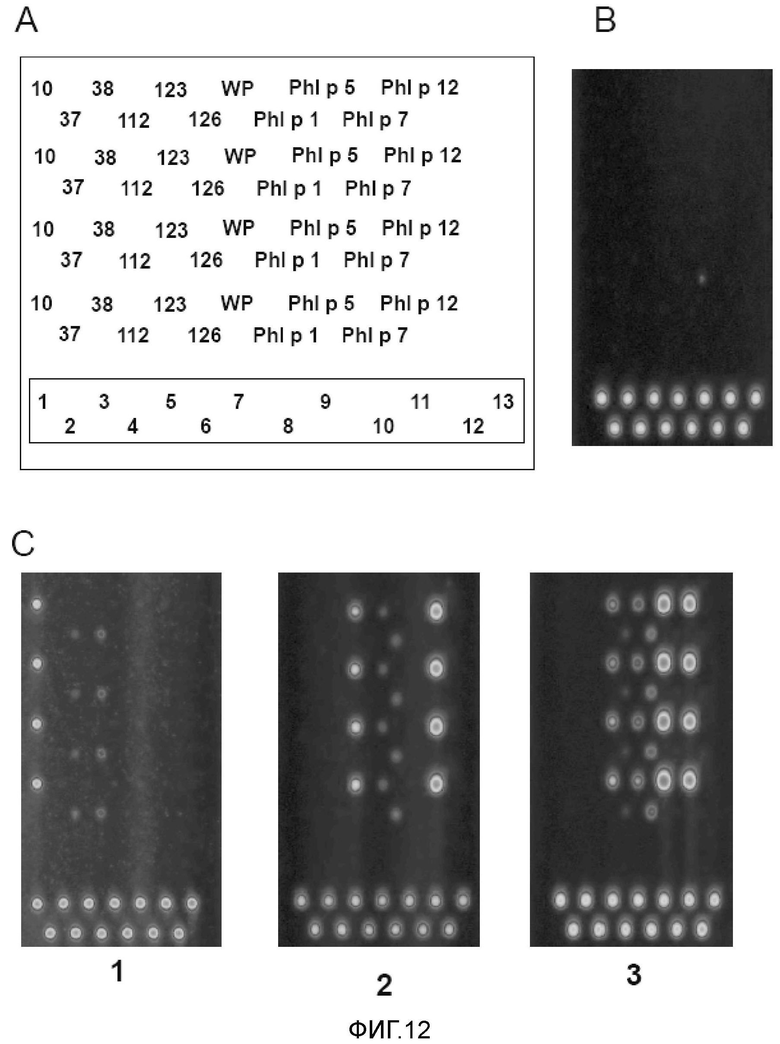
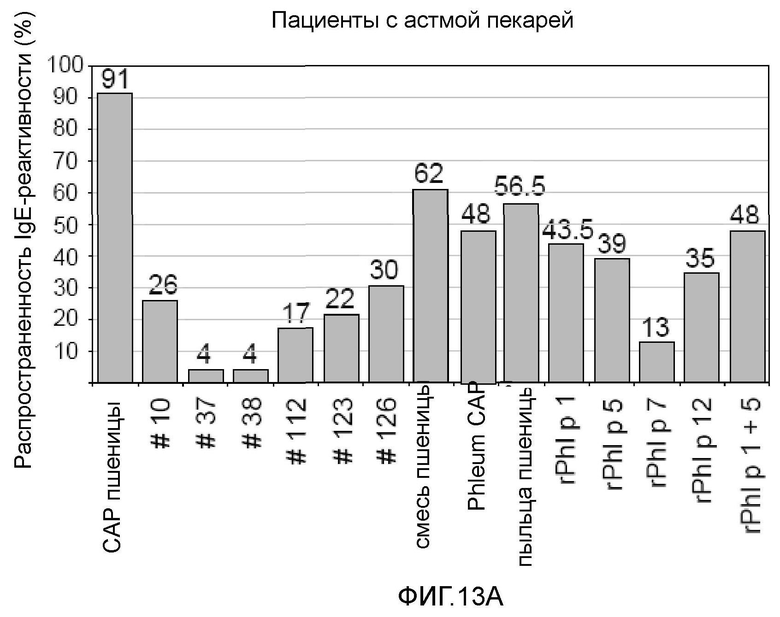
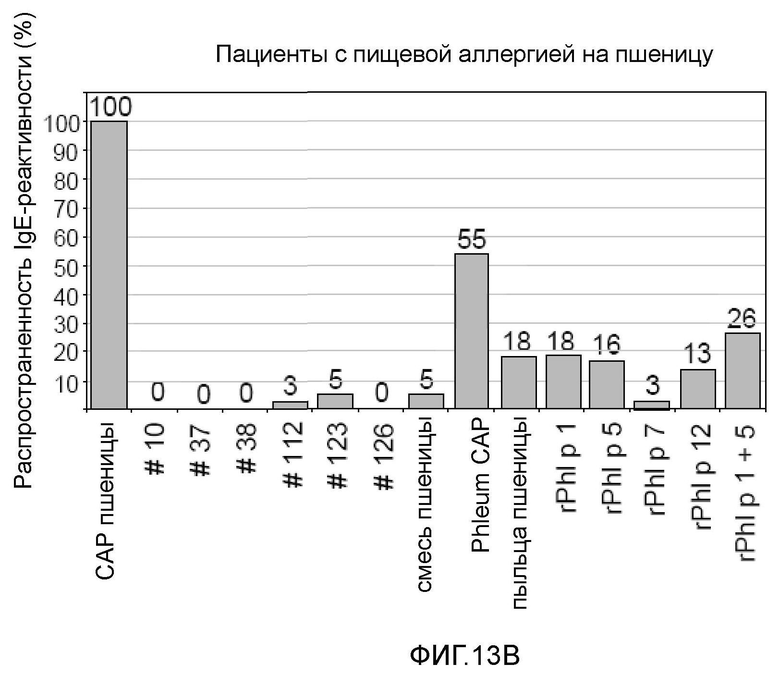
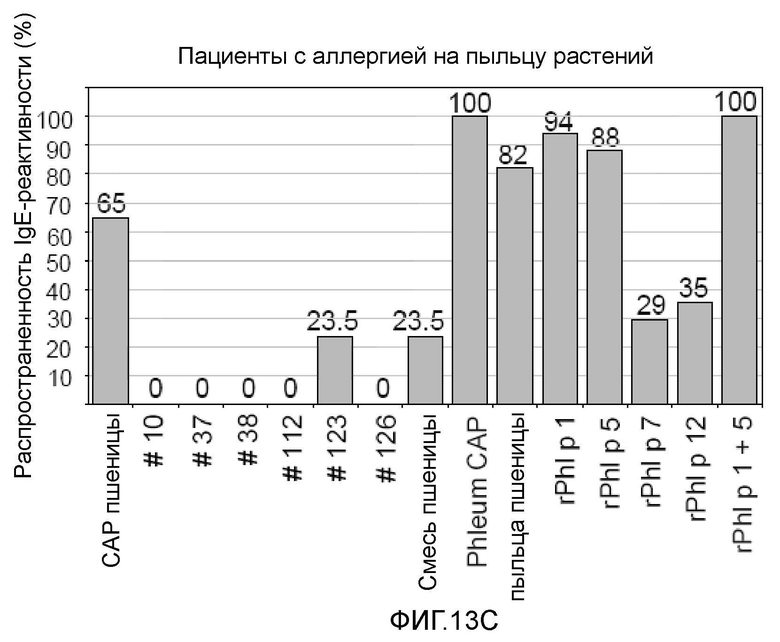
Настоящее изобретение относится к биотехнологии, конкретно к новым аллергенам, и может быть использовано в диагностике аллергии. Новые аллергены используют в способе диагностики IgE-опосредованной аллергии in vitro на пшеницу, включающем контактирование образца жидкости организма из млекопитающего, у которого предполагается наличие IgE-опосредованной аллергии на пшеницу, по меньшей мере, с одним полипептидом с SEQ ID NO: 2 или 8, или с его фрагментом или вариантом, имеющим общие эпитопы для антител с указанным полипептидом и имеющим последовательность, идентичную SEQ ID NO: 2 или 8 по крайней мере на 95%; и выявление присутствия в образце IgE-антител, специфично связывающихся с указанным полипептидом или полипептидами. Присутствие в образце таких антител является показателем IgE-опосредованной аллергии на пшеницу. Диагностический набор включает указанные полипептиды и средства для выявления связывания IgE с указанным полипептидом, такие как твердая подложка, например нитроцеллюлозная мембрана или микроматрица, содержащая связанный с ней полипептид. Изобретение позволяет эффективно диагностировать IgE-опосредованную аллергию на пшеницу. 5 н. и 3 з.п. ф-лы, 13 ил., 9 табл., 4 пр.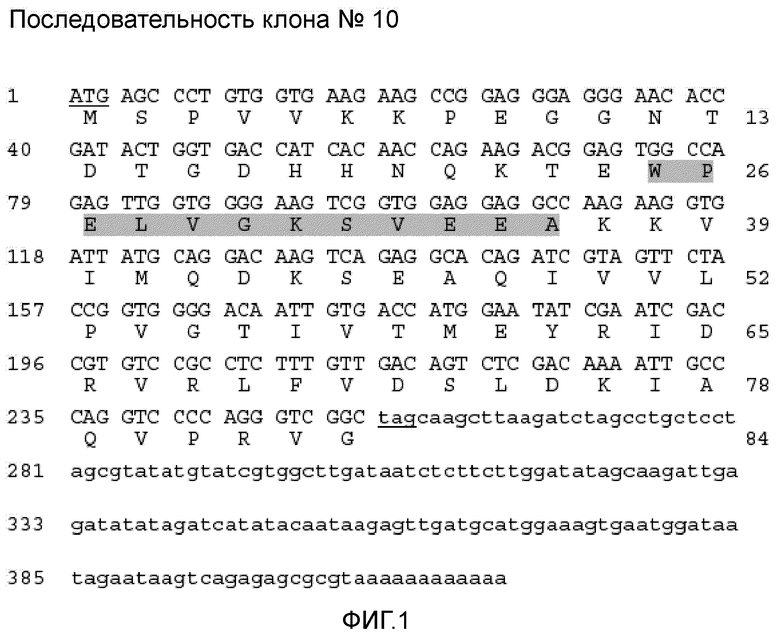
1. Изолированный полипептид, имеющий аминокислотную последовательность согласно SEQ ID NO: 2 или 8, для применения в диагностике IgE-опосредованной аллергии на пшеницу.
2. Полипептид по п.1, отличающийся тем, что он выделен из пшеницы или получен рекомбинантно.
3. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид по п.1 или 2.
4. Нуклеиновая кислота по п.3, имеющая нуклеотидную последовательность, согласно SEQ ID NO: 1,7.
5. Применение изолированного полипептида, имеющего аминокислотную последовательность согласно SEQ ID NO: 2 или 8, или его фрагмента или варианта, имеющего общие эпитопы для антител с указанным полипептидом и имеющего последовательность идентичную SEQ ID NO: 2 или 8 по крайней мере на 95%, для диагностики IgE-опосредованной аллергии на пшеницу.
6. Способ диагностики IgE-опосредованной аллергии in vitro на пшеницу, включающий стадии
- контактирования образца жидкости организма из млекопитающего, у которого предполагается наличие IgE-опосредованной аллергии на пшеницу, по меньшей мере, с одним полипептидом, имеющим аминокислотную последовательность согласно SEQ ID NO: 2 или 8, или с его фрагментом или вариантом, имеющим общие эпитопы для антител с указанным полипептидом и имеющим последовательность идентичную SEQ ID NO: 2 или 8 по крайней мере на 95%; и
- выявления присутствия в образце IgE-антител, специфично связывающихся с указанным полипептидом или полипептидами;
при этом присутствие таких антител, специфично связывающихся с указанным полипептидом или полипептидами, является показателем IgE-опосредованной аллергии на пшеницу.
7. Способ по п.6, в котором IgE-опосредованной аллергией является респираторная аллергия на пшеничную муку.
8. Диагностический набор для осуществления способа по п.6 или 7, содержащий полипептид, имеющий аминокислотную последовательность согласно SEQ ID NO: 2 или 8, или его фрагмент или вариант, имеющий общие эпитопы для антител с указанным полипептидом и имеющий последовательность идентичную SEQ ID NO: 2 или 8 по крайней мере на 95%, и средства для выявления связывания IgE с указанным полипептидом, такие как твердая подложка, например, нитроцеллюлозная мембрана или микроматрица, содержащая связанный с ней полипептид.
| BITTNER ЕТ AL., Identification of wheat gliadins as an allergen family related to baker's asthma, JOURNAL OF ALLERGY AND CLINICAL IMMUNOLOGY, 2007, v.121, n.3, p.744-749 | |||
| Приспособление для счесывания шишек с деревьев | 1974 |
|
SU556745A1 |
| МИКРОАНАЛИЗ НА АЛЛЕРГЕНЫ | 2001 |
|
RU2276790C2 |
| Схема включения дополнительной обмотки линейного реле в дуплексной телеграфной связи | 1948 |
|
SU82977A1 |

