Изобретение относится к генной инженерии, биохимии и биотехнологии.
Лабораторная диагностика вирусного гепатита. В на территории России основана на выявлении специфических антигенов и антител в крови, а также вирусных нуклеиновых кислот: HBsAg - анти-HBs; анти-НВс класса IgM и IgG; HBeAg - анти-НВе; ДНК. Наиболее широко в диагностике используется определение специфического для каждого субтипа вируса поверхностного HBs антигена (HBV surface antigen, HBsAg) (Lieven Stuyver, Sija De Gendt, Caroline Van Geyt, Fabien Zoulim, Michael Fried, Raymond F. Schinazi and Rudi Rossau A new genotype of hepatitis В virus: complete genome аnd phylogenic relatedness Journal of General Virology (2000), 81, 67-74). Антиген выявляется при остром (через 3-5 недель с момента инфицирования) течении болезни задолго до появления клинических признаков и, является в этих случаях единственным серологическим маркером. Циркуляция HBsAg в крови 6 и более месяцев указывает на затяжное или хроническое течение болезни, и позволяет предположить хроническое носительство вируса. Элиминация HBsAg и выявление анти-HBs в крови происходит у больных острым HBV на стадии разрешения инфекционного процесса и является показателем выработки иммунитета к HBV (J.LE SEYEC, P. CHOUTEAU, I. CANNIE, С.GUGUEN-GUILLOUZO, and P. GRIPON Infection Process of Hepatitis B viruses Depends of the Presence of a Define Sequence in the Pre-Sl Domain Journal of Virology, Mar. 1999, p.2052-2057). Основными и наиболее чувствительными методами обнаружения HBsAg и анти-HBs в настоящий момент является качественный и количественный иммуноферментный анализ (ИФА) в разных модификациях.
По природе антиген является липопротеином; закреплен в наружной вирусной оболочке. Состоит из трех частей (чертеж): основного белка S (small) размером 226 аминокислотных остатка (аа) и двух протеинов, на N-конце: белка М (middle), состоящего из белка S и области pre-S2 (55 аа); белка L (large), состоящего из белка М и области pre-S1 (119/108 аа). Области pre-S имеют независимые от основного белка S функциональные домены; их рассматривают как самостоятельные единицы, так и в составе HBsAg. Удаление областей не влияет на структуру и иммунохимические свойства HBsAg (Sue DELOS, Maria Teresa VILLAR, Peisheng HU L. PETERSON Cloning, isolation and characterization of the pre-S domains of hepatitis В surface antigen, devoid of the S protein Biochem J. (1991) 276, 411-416).
Основным компонентом современной вакцины против гепатита В является S белок. Специфические поствакцинальные антитела, вырабатываемые организмом, синтезируются к антигенным детерминантам именно этой области. При заражении организма специфические антитела вырабатываются ко всем антигенным детерминантам HBsAg; так как в составе инфекционных частиц присутствуют три модификации поверхностного антигена. В N-концевой области белка L закодированы белковые последовательности для распознавания и взаимодействия с В и Т клетками организма-хозяина; сайты связывания с гепатоцитами, альбумином человека и многие другие (Wei-Guo Нu, Jun Wei, Heng-Chuan Xia, Xin-Xiu, Feng Li, Guahg-Di Li, Yuan Wang, Zu-Chuan Zhang Identification of the immunogenic domains in HBsAg preSI region using overlapping preSI fragment fusion proteins World J Gastroenterol 2005; 11 (14): 2088-2094). Исследование сывороток в иммунологических реакциях с рекомбинантными белками pre-S области в случае положительного ответа позволит классифицировать поствакцинальные и сыворотки, полученные от больных и переболевших пациентов.
Производство биологических компонентов традиционным способом представляет многостадийный и дорогостоящий процесс, требующий особых условий работы. Актуальным является упрощение схемы производства компонентов, разработка субъединичных препаратов с использованием методов генной инженерии. Один из методов получения и очистки биологических компонентов (в частности, рекомбинантных белков) заключается в иммобилизации их на сорбенте. Такую способность белкам придает один из составляющих белок доменов, который аффинно взаимодействует с полимерными сорбентами с высокой константой связывания. При этом исходные антигенные свойства второго функционального белкового домена не нарушаются.
Используя этот подход, получили препараты двухкомпонентных рекомбинантных белков. Первый белковый домен содержит антигенную детерминанту из N-концевой области белка L субтип ayw (детерминанта L1 21-47 аа или L2 83-108 аа), он определяет функциональные (в данном случае антигенные) свойства белка. Далее следует соединяющая (спейсерная) аминокислотная последовательность из нескольких чередующихся остатков глицина и серина. Второй белковый домен содержит фрагмент последовательности гена эндоглюканазы СеlD из термофильного микроорганизма A. thermophilum, он определяет способность белка взаимодействовать с целлюлозным сорбентом. Конечные белковые продукты нарабатываются в непатогенных лабораторных штаммах Е.coli Иммобилизированные на целлюлозе белки представляют собой суспензию целлюлозосодержащего сорбента с адсорбированными на нем белками. Конечные препараты представляют собой растворы белков в буфере с определенной ионной силой, свободные от примесей липополисахаридов и ДНК штамма-продуцента
Иммунологические исследования подтвердили, что сохраняются антигенные свойства рекомбинантных белков и их способность взаимодействовать со специфическими антителами сыворотки крови.
Одним из перспективных направлений применения очищенных субъединичных препаратов является создание на их основе иммунологических диагностикумов, например белковых биочипов. Компоненты биочипа (рекомбинантные белки, антитела, лиганды и др.), как и в иммуноферментном анализе, иммобилизированы на носителе (подложка, микросферы). Но отличие от 96-луночных планшет (где в одной лунке размещается только один аналит) на поверхности носителя возможно иммобилизовать более одного анализируемого образца. Это позволяет проводить мультиплексный анализ образца, то есть исследование его одновременно по нескольким параметрам (Keij JF, Stenkamp JA. Flow cytometric characterization and classification of multiple dual-color fluorescent microspheres using fluorescence lifetime Cytometry 1998 Nov 1; 33(3):318-23).
Задачей заявляемой группы изобретений является создание штаммов Е. coli, обеспечивающих высокий уровень продукции рекомбинантных доменов белка L HBsAg, а также создание простой и эффективной схемы очистки рекомбинантных белков с последующим их использованием для разработки на их основе мультиплексного метода исследования сывороток крови, кроме того, получение иммобилизованных на полистирольном носителе рекомбинантных белков, сохраняющих способность специфически эффективно взаимодействовать с антителами сыворотки и возможность использования этих конъюгатов для разработки метода мультиплексного анализа.
Техническим результатом является 10% уровень синтеза белков L1spCBD и L2spCBD рекомбинантными штаммами Е.coli M15 от общего синтеза белка [pREP4, pL1spCBD] или Е.coli М15 [pREP4, pL2spCBD], несущими рекомбинантную плазмиду pL1spCBD или pL2spCBD соответственно, а также специфическое взаимодействие рекомбинантных белков L1spCBD и L2spCBD со специфическими антителами сыворотки крови от больных и вакцинированных пациентов и отсутствие взаимодействия с сыворотками от здоровых людей и иммунизированных пациентов.
Поставленная в заявляемой группе задача с достижением упомянутого выше технического результата решена следующим образом.
В части плазмида заявленный технический результат решается за счет того, что заявляемые плазмиды содержат следующие существенные для ее функционирования структурные элементы:
- искусственный бактериальный оперон химерных белков, включающий: промоторную область раннего промотора бактериофага Т5 (7-87 п.н.), обеспечивающий эффективную транскрипцию контролируемой мРНК; рекомбинантный ген, кодирующий целевой химерный белок (домены белка L HBsAg - спейсер - целлюлозосвязывающий домен CBD) (L1spCBD 115-817 п.н. и L2spCBD 115-814 п.н.); нетранслируемую область терминации транскрипции бактериального оперона, обеспечивающую эффективное окончание транскрипции (L1spCBD 857-951 п.н. и L2spCBD 854-948 п.н.);
- бактериальный оперон bla, кодирующий белок бета-лактамазу, являющуюся селективным маркером для трансформации штаммов Е coli (L1spCBD 3045-3905 п.н. и L2spCBD 3042-3902 п.н. комплементарной цепи);
- бактериальный участок инициации репликации типа ColE1, обеспечивающий репликацию плазмиды в штаммах Е.coli (L1spCBD 2286-2292 п.н. и L2spCBD 2283-2289 п.н.).
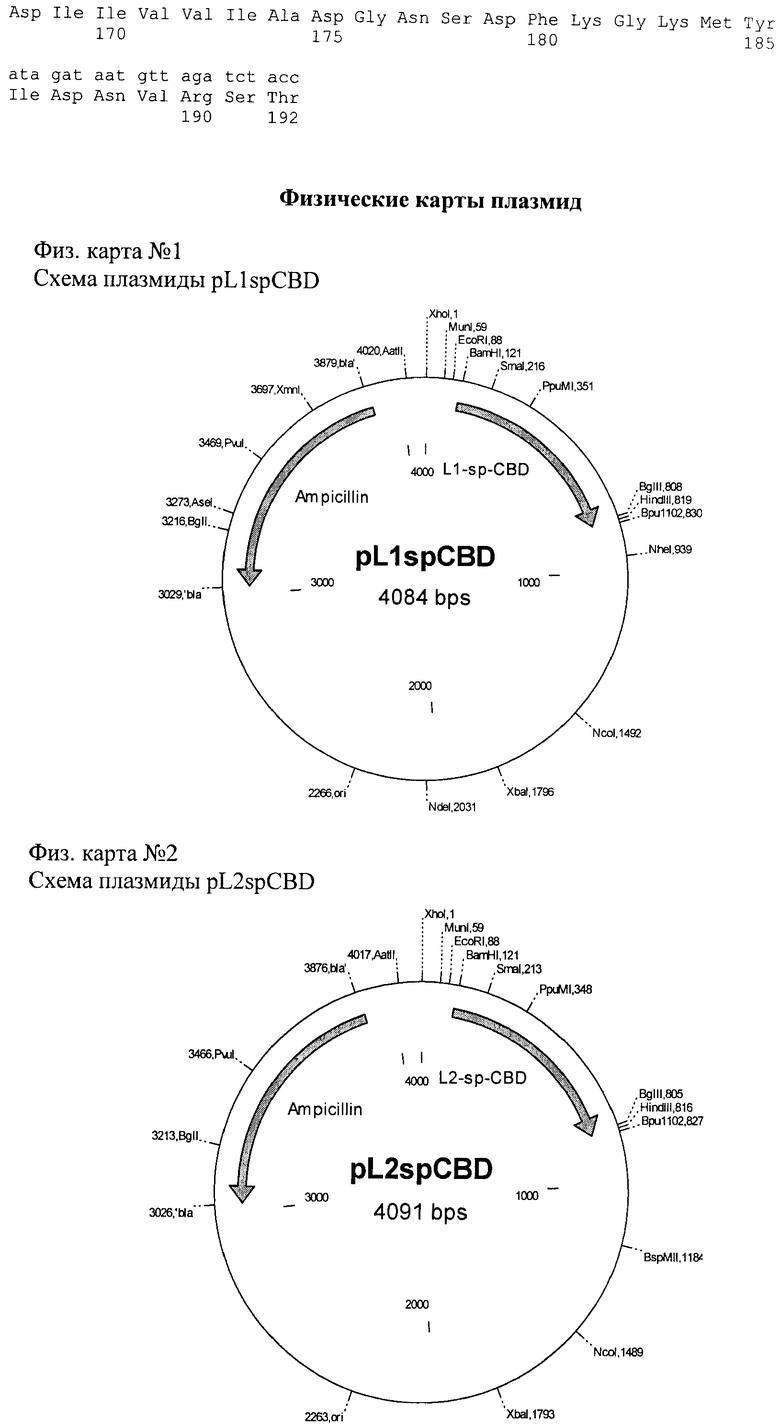
Таким образом, созданы рекомбинантные плазмиды, кодирующие бифункциональные рекомбинантные белки L1spCBD и L2spCBD, обладающие способностью специфически взаимодействовать с антителами сыворотки и способностью самопроизвольно связываться с целлюлозосодержащими сорбентами; размером pL1spCBD 4084 п.н. и pL2spCBD 4091 п.н., содержащие следующие структурные элементы: - искусственный бактериальный оперон химерного белка, включающий промоторную область раннего промотора бактериофага Т5 (80 п.н.), ген химерного белка L1spCBD (702 п.н.) и L2spCBD (699 п.н.) и терминатор транскрипции (94 п.н.); бактериальный оперон бета-лактамазы (860 п.н.) и бактериальный участок инициации репликации типа ColEl (11 п.н.).
Штаммы-продуценты рекомбинантных белков L1spCBD и L2spCBD получают трансформацией штамма Е.coli M15 [pREP4] плазмидами pL1spCBD или pL2spCBD соответственно. Штаммы обеспечивают продукцию рекомбинантных белков L1spCBD и L2pCBD после проведения процедуры индукции (изопропил-β-D-тио-галактопиранозидом) (ИПТГ) до 7-10% от тотального белка клетки.
Штаммы Е.coli M15 [pREP4], несущие плазмиду pL1spCBD и pL2spCBD, - продуценты бифункциональных рекомбинантных белков L1spCBD и L2spCBD характеризуются следующими признаками.
Культурально-морфологические признаки. Клетки прямые, палочковидные, неподвижные, грамотрицательные. При рассеве на чашке с 1,5% LB-агаром рост в виде отдельных колоний, иногда в R-форме с неровными краями. Хорошо растет на плотных и жидких питательных средах (LB-бульон, LB-агар, МПА, МПБ).
Физиолого-биохимические признаки. Клетки растут в пределах +4 - 42°С при оптимуме рН 6,8-7,5. Штаммы разлагают глюкозу, маннит с образованием кислоты, не разлагают сахарозу, арабинозу, галактозу, сбраживают мальтозу, ксилозу, сорбит, рамнозу. Существенным при использовании данных штаммов является их чувствительность к налидиксовой кислоте (25 мг/мл), стрептомицину (20 мг/мл) и рифампицину (25 мг/мл). Проявляют устойчивость к ампициллину (до 100 мкг/мл), обусловленную наличием плазмид pL1spCBD и pL2spCBD, и к канамицину (до 25 мкг/мл), обусловленную наличием плазмиды pREP4.
Рекомбинантные белки L1spCBD и L2spCBD, включающие в себя белковую последовательность, определяющую способность данных белков связываться с целлюлозой, что позволяет проводить в одну стадию концентрирование, очистку и иммобилизацию белкового продукта на целлюлозе. Иммобилизация на целлюлозе обеспечивается за счет присутствия в рекомбинантных белках целлюлозосвязывающего домена из эндоглюконазы CelD из штамма термофильного микроорганизма A. thermophilum, который обладает высоким сродством к кристаллической и аморфной целлюлозам и обеспечивает необратимое связывание с носителем в широком диапазоне значений рН 4-11, температуры 0-75°С и концентраций соли 0-5 М NaCl.
Поскольку в Е.coli отсутствуют белки, связывающиеся с целлюлозой, то синтезируемые в клетках Е.coli, трансформированных плазмидами pL1spCBD и pL2spCBD, рекомбинантные белки L1spCBD и L2spCBD являются единственными белками штаммов-продуцентов, прочно связывающимися с целлюлозой. Это обеспечивает возможность одностадийного получения высокоочищенных препаратов белков, иммобилизованных на сорбенте.
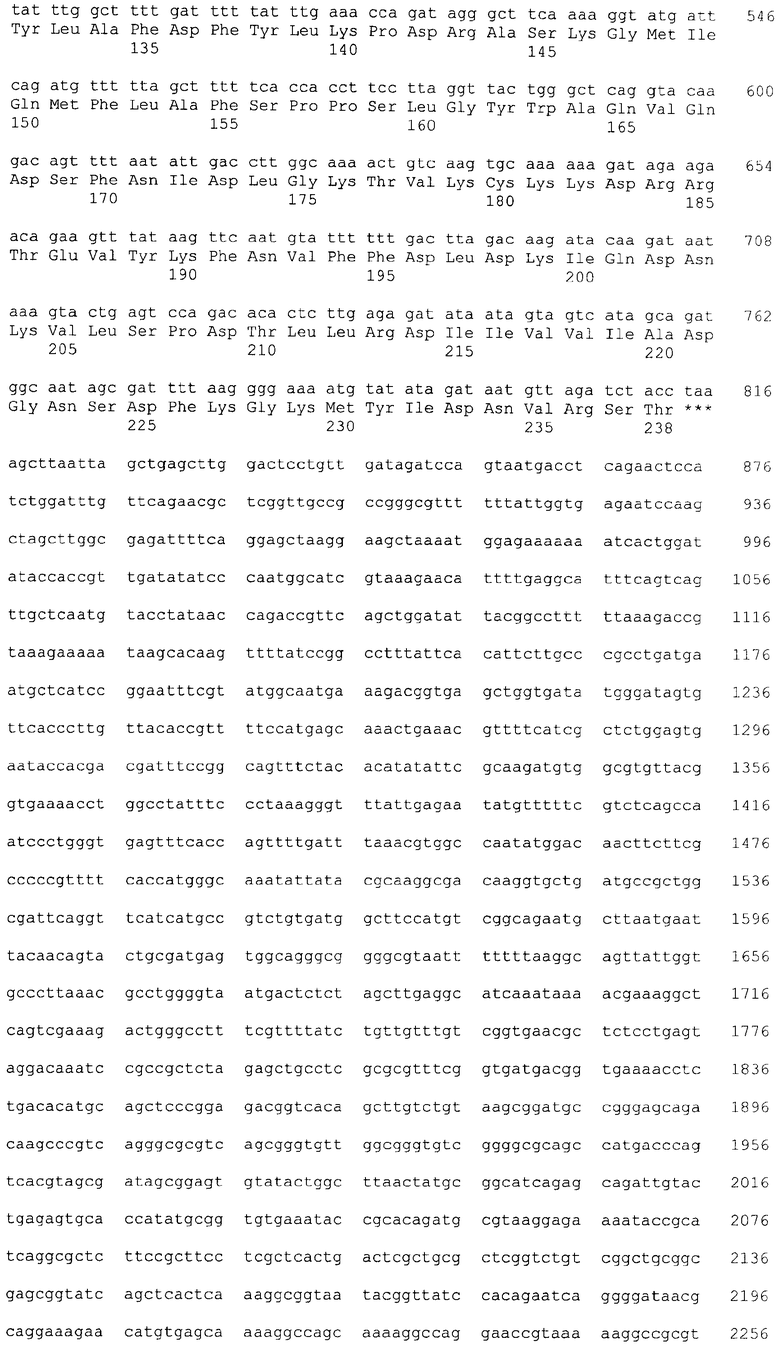
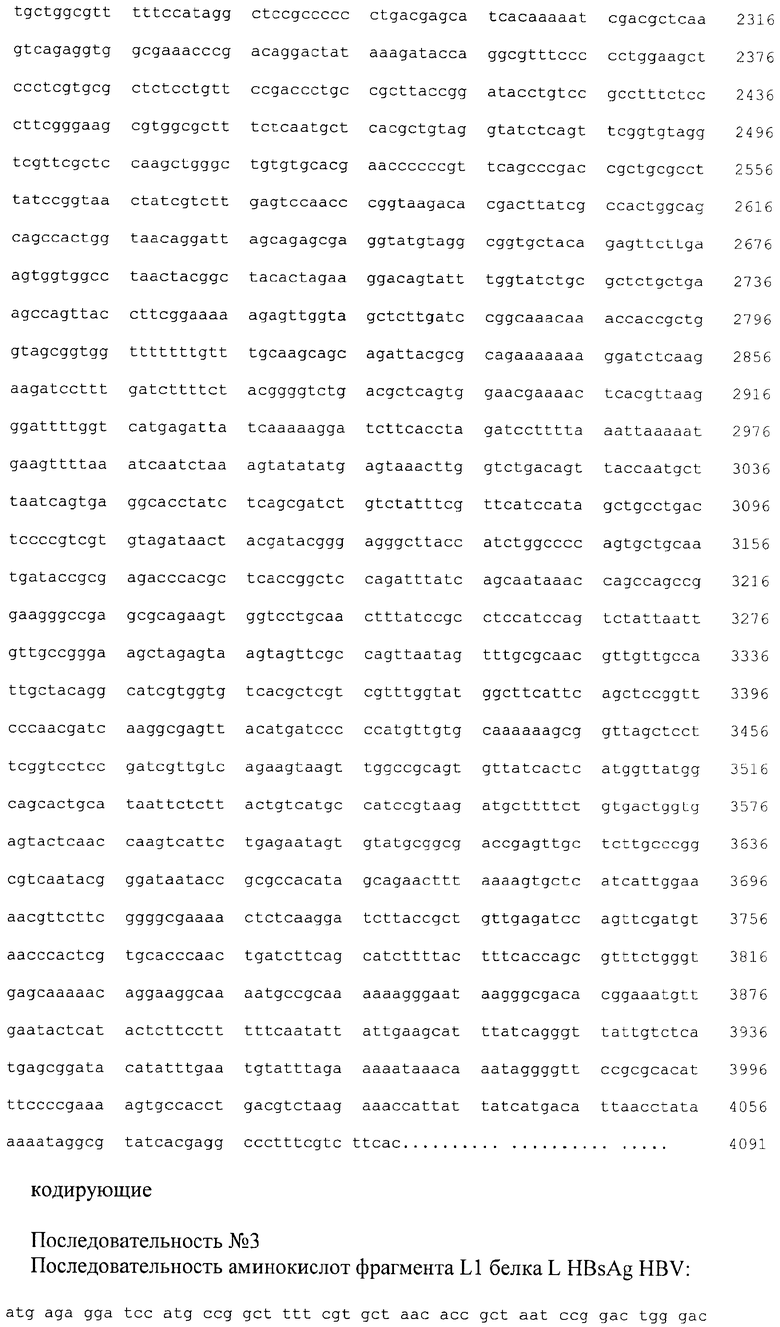
На фиг.1 и 2 приведены схемы плазмид pL1spCBD и pL2spCBD; последовательности плазмид №1 - pL1spCBD, №2 - pL1spCBD.
Изобретение проиллюстрированно следующими примерами, приведенными ниже. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам (Маниатис Т., Фрич Э, Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д.Гловера, Пер. с англ., Москва, Мир, 1988; Saiki R.K., Gelfand D.H., Stoffel S., Sharf S.J., Higuehi R., Horn G.T., Mullis K.B., Eriich H.A. Science, 1988. V.239, №4839, p.487-491; Sanger F., Nicklen S., Coulson A.R. Proc. Nat. Acad. Sci. USA, 1977, V.74, p.5463-5467).
Пример 1. Получение плазмид LIspCBD и L2spCBD.
а) Химический синтез олигодезоксирибонуклеотидов.
Олигодезоксирибонуклеотиды были синтезированы твердофазным амидофосфитным методом и очищены методом электрофореза в 12%-ном полиакриламидном геле (ПААГ).
б) Получение фрагментов гена L белка L (HBsAg Hepatitis В viruses (HBV)) с последующим его клонированием.
Фрагменты гена L получали методом сборки двухцепочечных дуплексов ДНК из синтетических олигонуклеотидов (ЗАО «Синтол», Москва). Олигонуклеотиды соответствовали областям ДНК с 21 по 47 (L1) и с 83 по 108 (L2) аминокислоты гена L. Для клонирования фрагментов ДНК, в последовательности олигонуклетидов запланировали сайты эндонуклеаз рестрикции BamHI, BglII, Xbal (подчеркнут сайт BglII):
Олионуклеотиды для синтетической сборки фрагмента L1:
1 HBL1F1 5'-ga tсc atg ccg get ttt cgt get aac acс get aat ccg gac tgg gac ttt aac ccg aac aaa-3'
2 HBL1F2 5'-P-gaс acс tgg ccg gac get aac aaa gtt aga tct taa t-3'
3 HBL1R1 5'-ct aga tta aga tct aac ttt gtt agс gtс сgg cca ggt gtc ttt gtt сgg gtt aaa gtc cca-3'
4 HBL1R2 5'-P-gtc сgg att age ggt gtt agс acg aaa agс сgg cat g-3'
Олигонуклеотиды для синтетической сборки фрагмента L2:
1 HBL2F1 5'-ga tсc atg ccg get agс acс aac cgt cag tct ggt cgt cag ccg acс ccg ctg tct ccg-3'
2 HBL2F2 5'-P-ccg ctg cgt aat acс cac ccg cag gсt aga tct taa t-3'
3 HBL2R1 5'-ct aga tta aga tct age ctg сgg gtg ggt att acg cag сgg сgg aga cag сgg ggt сgg ctg-3'
4 HBL2R2 5'-P-acg acс aga ctg acg gtt ggt gсt agс сgg cat g-3'
Для получения двухцепочечных фрагментов ДНК смешивали по 50 пкМ каждого олигонуклеотида, прогревали смесь 20 мин при 95°С и охлаждали в течение 4 часов до 25°С. Для клонирования двуцепочечных фрагментов ДНК, плазмидный вектор pQE16 гидролизовали эндонуклеазами рестрикции BamHI и Xbal при 37°С в буфере, содержащем 33 мМ Трис-ацетата (рН 7,9 при 37°С), 10 мМ ацетата магния, 66 мМ ацетата калия и 0,1 мг/мл BSA в течение 1 ч. В 1,2% агарозном геле проводили разделение продуктов рестрикции. Выделенный из агарозного геля фрагмент вектора pQE16 (размером 2419 нт) объединяли с фрагментами L1 (размером 95 п.н.) и L2 (101 п.н.). Лигировали при 25°С в буфере, содержащем 40 мМ Трис-HCl, 10 мМ магния хлорид, 10 мМ DTT, 0,5 мМ АТФ (рН 7,8 при 37°С) с помощью ДНК лигазы бактериофага Т4 в течение 1 ч. Продукты реакции переосаждали смесью 96% этиловый спирт+5М ацетат аммония при -20°С в течение 15 ч. После переосаждения смесь растворяли в деионизированной воде (mQ Н2О). Методом электропорации полученными лигазными смесями трансформировали компетентные клетки штамма Ml 5 Eschenchia coli (E. coh) [pREP4] (Nals, Strs, rifs, lac-, ara-, gal-, mtl-, F-, recA+, uvr+). После трансформации клоны отбирали на агаризованнной среде LB, содержащей антибиотики ампициллин (50 мг/мл) и канамицин (25 мг/мл). Плазмидную ДНК из колоний выделяли методом щелочного лизиса; анализировали методом рестрикционного картирования эндонуклеазами рестрикции BamHI, BspMII, XbaI, сайты которых находятся внутри клонируемых фрагментов. Отбирали клоны pL1 и pL2, плазмидная ДНК которых (размером 2508 п.н. - pL1 и 2515 п.н. - pL2) содержит последовательности гена L. Первичную структуру полученных плазмид подтверждали секвенированием.
в) Получение последовательности, кодирующей Gly-Ser спейсер, с последующим ее клонированием.
Для получения последовательности спейсера, состоящего из повторов аминокислотных остатков Gly-Ser, были синтезированы два комплиментарных олигонуклеотида следующего состава (подчеркнут сайт эндонуклеазы рестрикции BglII; под нуклеотидной последовательностью первого олигонуклеотида приведена кодируемая последовательность аминокислот):
1 5'-ga tсc ccg ggt tct ggc tсc ggc tсt ggt tсc ggt tct ggc gcc aga tct a-3'
S P G S G S G S G S G S G A R S
2 5'-t aga tct ggc gcc aga acс gga acс aga gcc gga gcc aga acс сgg gga tc-3'
Для получения двухцепочечного фрагмента ДНК смешивали по 25 пкМ каждого олигонуклеотида, прогревали смесь 15 мин при 95°С и охлаждали в течение 4 часов до 25°С. Для клонирования двухцепочечного фрагмента ДНК плазмидный вектор pQE13 гидролизовали эндонуклеазами рестрикции BamHI и HindIII при 37°С в буфере, содержащем 66 мМ Трис-ацетата (рН 7,9 при 37°С), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. В 1,2% агарозном геле проводили разделение продуктов рестрикции. Выделенный из агарозного геля фрагмент вектора pQE13 (размером 3420 п.н.) объединяли с двухцепочечным фрагментом ДНК, кодирующим спейсер, и дотировали при 25°С с помощью ДНК-лигазы бактериофага Т4 в течение 1 ч. Методом электропорации полученными лигазными смесями трансформировали компетентные клетки Е.coli шт. M15 [pREP4]. После трансформации клоны отбирали на агаризованнной среде LB, содержащей антибиотики ампициллин и канамицин. Плазмидную ДНК из колоний выделяли методом щелочного лизиса, анализировали методом рестрикционного картирования эндонуклеазами рестрикции BamHI, HindIII, а также эндонуклеазами рестрикции, находящихся внутри клонируемого фрагмента. Отбирали клоны pRsp, плазмидная ДНК которых (размером 3471 п.н.) содержит последовательность Gly-Ser спейсера. Первичную структуру полученной плазмиды подтверждали секвенированием.
г) Получение участка гена CelD (Anaerocellum thermophilum), содержащего последовательность целлюлозосвязывающего домена (ЦСД, CBD), с последующим его клонированном в плазмидную конструкцию pRsp.
Копию фрагмента гена СеlD получали методом полимеразной цепной реакции (ПЦР), для чего были выбраны праймеры, соответствующие началу и концу данной области ДНК следующего состава (подчеркнуты сайты эндонуклеаз рестрикции BamHI, HindIII и BglII):
1 5'-ааа gaa gga tсc aat gca cct tta ggc g-3'
2 5'-cct caa aaa agс ttt agg tag atс taa cat tat cta tat ac-3'
В качестве ДНК-матрицы использовали хромосомную ДНК из Anaerocellum thermophilum (А. thermophilum) Методом ПЦР с использованием олигонуклеотидных праймеров, соответствующих начальной и концевой части ДНК, амплифицировали последовательность нуклеотидов. Реакцию проводили в 25 мкл реакционной смеси, содержащей 2 мкг ДНК, 10 пкМ каждого праймера, 67 мМ трис-НСl (рН 8,8), 15 мМ сульфата аммония, 2,5 мМ хлористого магния, 0,01% Твин-20, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. Taq-ДНК-полимеразы. Реакцию амплификации проводили под вазелиновым маслом: 5 циклов: 93°С - 2 мин, 57°С - 5 мин и 72°С - 30 с; 30 циклов: 93°С - 1 мин, 63°С - 1 мин и 72°С - 30 с; затем 63°С - 5 мин и 72°С - 10 мин. Продукт амплификации размером 583 п.н. обрабатывали хлороформом, переосаждали смесью 96% этиловый спирт +5М ацетат аммония и растворяли в mQ H2O. Для клонирования ПЦР-продукта полученную ДНК гидролизовали эндонуклеазами рестрикции BamHI и HindIII при 37°С в буфере, содержащем 66 мМ Трис-ацетата (рН 7,9 при 37°С), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Плазмидную ДНК pRsp гидролизовали эндонуклеазами рестрикции BglII и HindIII в аналогичных условиях. В 1,2% агарозном геле проводили разделение продуктов рестрикции. Выделенные из агарозного геля фрагменты рестрикции: фрагмент ДНК, соответствующий участку гена CelD (размером 563 п.н.) и фрагмент плазмиды pRsp (размером 3465 п.н.), содержащий селективный маркер, участок инициации репликации, промоторную область и последовательность, кодирующую Gly-Ser спейсер, лигировали при 25°С с помощью ДНК-лигазы бактериофага Т4 в течение 1 ч. Методом электропорации полученными лигазными смесями трансформировали компетентные клетки Е.coli шт. M15 [pREP4]. После трансформации клоны отбирали на агаризованнной среде LB, содержащей антибиотики ампициллин и канамицин. Плазмидную ДНК из колоний выделяли методом щелочного лизиса, анализировали методом рестрикционного картирования эндонуклеазами рестрикции BamHI, HindIII, а также эндонуклеазами рестрикции, находящихся внутри клонируемого фрагмента. Отбирали клоны pspCBD, плазмидная ДНК которых (размером 4028 п.н.) содержит последовательность участка гена CelD со спейсером на N-конце, состоящим из Gly-Ser повторов. Первичную структуру полученной плазмиды подтверждали секвенированием.
е) Получение плазмид, содержащих последовательности гена L, Gly-Ser спейсера, и целлюлозосвязывающего домена CBD.
Для получения плазмид, содержащих последовательности гена L, Gly-Ser спейсера, и целлюлозосвязывающего домена CBD, плазмиды pL1 и pL2 гидролизовали эндонуклеазами рестрикции BglI и BglII при 37°С в буфере, содержащем 50 мМ Трис-НСl (рН 7,5), 10 мМ хлорида магния, 100 мМ хлорида натрия и 0,1 мг/мл BSA, в течение 1 часа. Плазмиду pspCBD гидролизовали эндонуклеазами рестрикции BamHI и BglI при 37°С в буфере, содержащем 66 мМ Трис-ацетат (рН 7,9 при 37°С), 20 мМ ацетат магния, 132 мМ ацетат калия и 0,2 мг/мл BSA в течение 1 часа. В 1,2% агарозном геле проводили разделение продуктов рестрикции. Выделенные из агарозного геля фрагменты рестрикции: фрагменты плазмидной ДНК pLI (размером 1073 п.н.), фрагменты плазмидной ДНК pL2 (размером 1080 п.н.), а также фрагмент плазмидной ДНК pspCBD, включающий Gly-Ser спейсер и последовательность целлюлозосвязывающего домена CBD (размером 3011 п.н.) лигировали с помощью ДНК-лигазы бактериофага Т4. Методом электропорации полученными лигазными смесями трансформировали компетентные клетки Е.coli шт. M15 [pREP4]. После трансформации клоны отбирали на агаризованнной среде LB, содержащей антибиотики ампициллин и канамицин. Плазмидную ДНК из колоний выделяли методом щелочного лизиса, анализировали методом рестрикционного картирования эндонуклеазами рестрикции BamHI, BglI, BgLII, а также эндонуклеазами рестрикции, находящихся внутри клонируемого фрагмента. Отбирали клоны pL1spCBD (размером 4084 п.н.) и pL2spCBD (размером 4091 п.н.), плазмидная ДНК которых содержит последовательности участков гена L, Gly-Ser спейсера, и целлюлозосвязывающего домена CBD. Первичную структуру полученной плазмиды подтверждали секвенированием.
Пример 2. Получение штамма Е coli - продуцента рекомбинантного белка L1spCBD, состоящего из фрагмента L1 белка L, Gly-Ser спейсера, целлюлозосвязывающего домена.
Для получения штамма Е.coli - продуцента рекомбинантного белка L1spCBD, клетки штамма Е.coli M15 [pREP4] трансформировали плазмвдэй pL1spCBD. Трансформированные клетки выращивали в 3,5 мл среды LB с ампициллином и канамицином при 37°С до оптической плотности, соответствующей 1 ед. поглощения при длине волны 550 нм (около 2,5 ч), добавляли 3 мкл 0,1 М раствора изопропил-бета-D-тиогалактопиранозида (ИПТГ, IPTG) и выращивали в течение 3 ч. Для контроля продукции рекомбинантного белка L1spCBD в Е.coli M15 [pREP4, pL1spCBD] применяли метод электрофореза в полиакриламидном геле (ПААГ) в присутствии додецилсульфата натрия (ДСН). Разделение белков проводили в 12% полиакриламипном геле в стандартной системе буферов (электродный буфер. 25 мМ Tris-HCl, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 83; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). По окончании электрофореза гели окрашивали 0,15% раствором Кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывали в 10% уксусной кислоте. При сравнении спектра белков у штаммов Е.coli M15 [pREP4] и Е.coli М15 [pREP4, pL1spCBD] обнаруживали появление дополнительных белковых полос с молекулярной массой 25,97 кДа, что соответствует молекулярной массе рекомбинантного белка L1spCBD. Уровень синтеза белка L1spCBD определяли, сравнивая интенсивность окрашивания полосы рекомбинашного белка с полосой соответствующего белка - стандарта молекулярной массы 23,48 кДа.
Пример 3. Получение штамма Е.coli - продуцента рекомбинантного белка L2spCBD, состоящего из фрагмента L2 белка L, Gly-Ser спейсера, целлюлозосвязывающего домена.
Для получения штамма Е.coli - продуцента рекомбинантного белка L2spCBD, клетки штамма Е.coli М15 [pREP4] трансформировали плазмидой pL2spCBD. Трансформированные клетки выращивали в 3,5 мл среды LB с ампициллином и канамицином при 37°С до ошической плотности, соответствующей 1 ед. поглощения при длине волны 550 нм (около 2,5 ч), добавляли 3 мкл 0,1 М раствора изопропил-бета-D-тиогалактопиранозида (ИПТГ, IPTG) и выращивали в течение 3 ч. Для контроля продукции рекомбинантного белка L2spCBD в Е.coli М15 [pREP4, pL2spCBD] применяли метод электрофореза в полиакриламидном геле (ПААГ) в присутствии додецилсульфата натрия (ДСН). Разделение белков проводили в 12% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ Tris-HCl, 192 мМ глицин, 0,1% додецилсульфаг натрия, рН 83; буфер для геля: 375 мМ трис-хлорицный буфер, рН 8,8). По окончании электрофореза гели окрашивали 0,15% раствором Кумасси G250 в 25% иэопропаноле и 10% уксусной кислоте и отмывали в 10% уксусной кислоте. При сравнении спектра белков у штаммов Е.coli М15 [pREP4] и Е.coli М15 [pREP4, pL2spCBD] обнаруживали появление дополнительной белковой полосы с молекулярной массой 25,68 кДа, что соответствует молекулярной массе рекомбинантного белка L2spCBD. Уровень синтеза белка L2spCBD определяли, сравнивая интенсивность окрашивания полос рекомбинантного белка с полосой соответствующего белка - стандарта молекулярной массы 23,48 кДа.
Пример 4. Получение рекомбинантных белков L1spCBD и L2spCBD, иммобилизированных на целлюлозе и в растворе.
Для получения рекомбинантных белков pL1spCBD, pL2spCBD культуры Е.coli М15 [pREP4, pL1spCBD] и Е.coli М15 [pREP4, pL2spCBD] выращивали в 1000 мл среды LB с ампициллином и канамицином при 37°С. До оптической плотности, соответствующей 1 ед. поглощения при длине волны 550 нм, добавляли 15 мкл 0,1 М раствора ИПТГ и выращивали в течение 3 ч. Клетки осаждали центрифугированием при 5500 об/мин в течение 15 мин. Осадок ресуспендировали в лизирующем буфере, содержащем 50 мМ Трис-НСl (рН 8,0), 0,25 мМ хлорид натрия, 5 мМ хлорид магния, 0,15 мМ PMSF, 0,1% Тритон-Х100, 0,5 мг/ мл лизоцим, 20 мг/ мл ДНКазы; из расчета 1 г биомассы на 4 мл буфера. Дополнительно суспензию обрабатывали ультразвуком 3 раза по 20 с. Клеточный дебрис удаляли центрифугированием при 16000 об/мин в течение 30 мин. Для иммобилизации рекомбинантных белков на целлюлозе, к супернатанту добавляли 1/3 объема 30% суспензии сферической целлюлозы, и буфер, содержащий 0,5% Тригон-X100, 0,5 М хлорид натрия; выдерживали при 25°С 1 час. Центрифугировали (ц/ф) при 8000 об/мин в течение 10 мин. Осадок ресуспендировали в том же буфере. Повторили отмывку целлюлозы 2 раза. Для получения раствора белка суспензию целлюлоза+иммобилизированные белки вносили в колонку. Дополнительно проводили отмывку целлюлозы раствором 4 М мочевины и 50 мМ Трис-НСl (рН 8,0). Градиентное снятие белков с целлюлозы проводили раствором 8 М мочевины и 50 мМ Трис-НСl (рН 8,0). Диализ проводили в при 25°С течение ночи против PBS буфера. Концентрацию белков определяли с помощью спектрофотометра при длине волны 280/ 260 нм, она составила 1,37 мг/мл L1spCBD и 1,22 мг/мл L2spCBD соответственно. Степень чистоты всех препаратов составляла не менее 95%.
Пример 5. Иммобилизация рекомбинантных белков L1spCBD и L2spCBD на полистирольных носителях.
Для работы использовали полистирольные носители - микросферы (Bio-Rad) в концентрациии 1,25×106. Всю работу проводили при 25°С.
Суспензию микросфер ц/ф при 14,700 об/мин (16.000g) в течение 4 мин. Осадок промывали буфером для отмывки (Bead wash buffer (Amine Coupling Kit), Bio-Rad); ц/ф при 14,700 об/мин (16.000g) в течение 4 мин. К осадкам добавляли реакционный буфер (Bead activation buffer), S-NHS (50 мг/мл), EDC (50 мг/мл); инкубировали в темноте с покачиванием 300 об/мин в течение 20 мин. Отмывку проводили PBS буфером 2 раза. К осадкам в разные пробирки добавляли рекомбинантные белки L1spCBD (1,37 мг/мл) и L2spCBD (1,22 мг/мл); инкубировали в течение 2 ч. Отмывку проводили PBS буфером 2 раза. Осадки ресуспендировали в блокирующем буфере (Bead blocking buffer); инкубировали в течение 10 мин. Ц/ф при 14,700 об/мин (16.000g) в течение 4 мин, осадки ресуспендировали в буфере для хранения (Bead storage buffer). Хранят препараты в темноте при +4°С.
Пример 6. Использование микросфер с иммобилизированными рекомбинантными белками L1spCBD и L2spCBD для работы на приборе Bio-Plex System.
Для работы использовали микросферы двух видов, сенсибилизированные рекомбинантными белками L1spCBD и L2spCBD, аналитический планшет MulttiScreen-BV (Millipore, USA); прибор Bio-Plex System (Luminex X-map Technology, USA); работу проводили при 25°С.
Перед началом работы лунки рабочего планшета промывали PBS буфером; жидкость отбирали под вакуумом. Добавляли суспензию меченных рекомбинантными белками микросфер; инкубировали в течение 2 мин. Жидкость отбирали под вакуумом; планшет промывали 2 раза PBS буфером. Добавляли образцы сывороток; инкубировали в течение 30 мин. Жидкость отбирали под вакуумом; планшет промывали 2 раза PBS буфером. В лунки добавляли меченные TAMRA антивидовые антитела; инкубировали в течение 30 мин. Жидкость отбирали под вакуумом; планшет промывали 2 раза PBS буфером. В лунки добавляли PBS буфер и анализировали образцы на приборе. Результаты исследования сывороток, полученные при работе на приборе Bio-Plex System, соответствуют результатам, полученным от проверки сывороток другими методами.
L1spCBD и L2spCBD взаимодействуют только со специфическими антителами сыворотки крови от этих больных.




| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422524C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422525C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| РЕКОМБИНАНТНЫЙ БЕЛОК LACspCBD, ОБЛАДАЮЩИЙ БЕТА-ГАЛАКТОЗИДАЗНОЙ АКТИВНОСТЬЮ И СПОСОБНОСТЬЮ САМОПРОИЗВОЛЬНО СВЯЗЫВАТЬСЯ С ЦЕЛЛЮЛОЗОСОДЕРЖАЩИМИ СОРБЕНТАМИ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА LACspCBD, ШТАММ Escherichia coli M15 [pREP4, pLACspCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА LACspCBD, СПОСОБ ПОЛУЧЕНИЯ ИММОБИЛИЗОВАННОГО РЕКОМБИНАНТНОГО БЕЛКА LACspCBD НА ЦЕЛЛЮЛОЗЕ И СПОСОБ ФЕРМЕНТАТИВНОГО РАСЩЕПЛЕНИЯ ЛАКТОЗЫ | 2004 |
|
RU2278160C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-CBD, ШТАММ Escherichia coli [pREP4, pAg85A-CBD], ХИМЕРНЫЙ БЕЛОК Ag85A-CBD И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2429292C1 |
| РЕКОМБИНАНТНЫЙ БЕЛОК Collbd-CBD, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pОС-Collbd, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА Collbd-CBD, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА Collbd-CBD | 2009 |
|
RU2408726C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-CFP10-DBD, РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli M15 [pREP4, pESAT6-CFP10-DBD], СПОСОБ ПОЛУЧЕНИЯ, ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ESAT6-CFP10-DBD НА ДЕКСТРАНЕ, РЕКОМБИНАНТНЫЙ БЕЛОК ESAT6-CFP10-DBD И ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК ESAT6-CFP10-DBD | 2013 |
|
RU2539026C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD1spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D1-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D1-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D1-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D1-GBD | 2008 |
|
RU2401302C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD4spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D4-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D4-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D4-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D4-GBD | 2008 |
|
RU2401305C2 |
Изобретение относится к биотехнологии. Описаны рекомбинантные плазмиды pL1spCBD и pL2spCBD. Предложены штаммы Escherichia coli - продуценты химерных белков L1spCBD и L2spCBD. Описаны рекомбинантные белки L1spCBD и L2spCBD. Раскрыт способ иммобилизации, концентрирования и очистки белков L1spCBD и L2spCBD на целлюлозе и на полистирольных носителях. Техническим результатом является специфическое взаимодействие рекомбинантных белков L1spCBD и L2spCBD со специфическими антителами сыворотки крови от больных и вакцинированных пациентов и отсутствие взаимодействия с сыворотками от здоровых людей и иммунизированных пациентов. 8 н.п. ф-лы, 1 ил.
1. Рекомбинантная плазмида pL1spСВО, характеризующаяся нуклеотидной последовательностью №1, обеспечивающая экспрессию рекомбинантного белка L1spCBD, состоящего из фрагмента L1 белка L HBsAg Hepatitis В Viruses (HBV), Gly-Ser спейсера, целлюлозосвязывающего домена гена эндоглюканазы СеlD из Anaerocellum thermophilum (A. thermophilum) размером 4084 п.н., состоящая из следующих структурных элементов:
искусственный бактериальный оперон рекомбинантного белка LIspCBD, включающий:
промоторную область раннего промотора бактериофага Т5 (7-87 п.н.), ген рекомбинантного белка L1spCBD (115-817 п.н.), нетранслируемую область терминации транскрипции (857-951 п.н.);
бактериальный оперон бета-лактамазы, обеспечивающий устойчивость к ампицилину (3045-3905 п.н.);
бактериальный участок инициации репликации типа ColEl (2286-2292 п.н.), обеспечивающий репликацию плазмиды в штаммах Е coli.
2. Рекомбинантная плазмида pL2spCBD, характеризующаяся нуклеотидной последовательностью №2, обеспечивающая экспрессию рекомбинантного белка L2spCBD, состоящего из фрагмента L2 белка L HBsAg HBV, Gly-Ser спейсера, целлюлозосвязывающего домена гена эндоглюканазы CelD из А. thermophilum, размером 4091 п.н., состоящая из следующих структурных элементов:
искусственный бактериальный оперон рекомбинантного белка L2spCBD: включающий:
промоторную область раннего промотора бактериофага Т5 (7-87 п.н.), ген рекомбинантного белка L2spCBD (115-814 п.н.), нетранслируемую область терминации транскрипции (854-948 п.н.);
бактериальный оперон бета-лактамазы, обеспечивающий устойчивость к ампицилину (3042-3902 п.н.);
бактериальный участок инициации репликации типа ColEl (2283-2289 п.н.), обеспечивающий репликацию плазмиды в штаммах Е.coli.
3. Рекомбинантный штамм Е.coli M15 [pREP4, pL1spCBD], полученный трансформацией плазмидой по п.1, депонированный во Всероссийской Коллекции Промышленных Микроорганизмов под номером ВКПМ 9897.
4. Рекомбинантный штамм Е.coli M15 [pREP4, pL2spCBD], полученный трансформацией плазмидой по п.2, депонированный во Всероссийской Коллекции Промышленных Микроорганизмов под номером ВКПМ 9898.
5. Способ иммобилизации, концентрирования и очистки рекомбинантных белков L1spCBD и L2spCBD на целлюлозе, включающий:
выращивание клеток штаммов Е.coli M15 [pREP4, pL1spCBD] по п.3 или Е.coli M15 [pREP4, pL2spCBD] по п.4;
связывание белка L1spCBD или L2spCBD в составе соответствующих клеточных экстрактов штамма Е.coli M15 [pREP4, pL1spCBD] или Е.coli M15 [pREP4, pL2spCBD] с целлюлозным сорбентом за счет аффинного взаимодействия при процедуре инкубации с последующей отмывкой от несвязавшихся бактериальных белков и
выделение целевого продукта.
6. Рекомбинантный белок L1spCBD, включающий фрагмент белка L с последовательностью №3, Gly-Ser спейсер с последовательностью №5, целлюлозосвязывающий домен гена эндоглюканазы CelD из А. thermophilum с последовательностью №6, при этом фрагмент белка L связывается с антителами в сыворотке крови с молекулярной массой, соответствующей (25973±1000) Да (при определении методом электрофореза в ПААГ), рI 8,92.
7. Рекомбинантный белок L2spCBD, включающий фрагмент белка L с последовательностью №4, Gly-Ser спейсер с последовательностью №5, целлюлозосвязывающий домен гена эндоглюканазы CelD из А. thermophilum с последовательностью №6, при этом антигенная специфичность фрагмента белка L связывается с антителами в сыворотке крови с молекулярной массой, соответствующей (25680±1000) Да (при определении методом электрофореза в ПААГ), pI 9,72.
8. Способ иммобилизации рекомбинантных белков L1spCBD и L2spCBD на полистирольных носителях, включающий:
суспендирование чистого рекомбинантного белка по п.6 или 7 и полистирольных носителей в буфере Bead wash buffer;
добавление реакционной смеси и инкубирование компонентов;
отмывку полученного препарата полистирольный носитель+белок и хранение в буфере Bead storage buffer при 4°С в темноте.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| GOLOVLIOV I | |||
| et al | |||
| Adjuvanticity of ISCOMs incorporating a T cell-reactive lipoprotein of the facultative intracellular pathogen Francisella tularensis, Vaccine, 1995, Feb; 13(3) abstract. | |||

