Изобретение относится к генной инженерии, биотехнологии, микробиологии, иммунологии и может быть использовано для конструирования плазмидной ДНК pD1spGBD, обеспечивающей экспрессию рекомбинантного антигена - домена 1 белка LigA L. interrogans в составе химерного белка D4-GBD с 1,3-β-глюкансвязывающим доменом (GBD), и получения на их основе субъединичной генно-инженерной вакцины против лептоспироза.
В России иммунизация населения против лептоспирозов проводится по эпидемиологическим показаниям, т.е. иммунизации подлежат лица, относящиеся к группам профессионального риска заражения (в любое время года), персонал лабораторий, работающих с патогенными лептоспирами, а также лица, направляемые на строительные и сельскохозяйственные работы в места активно действующих природных и хозяйственных очагов лептоспирозов. В эндемичных по лептоспирозам очагах вакцинируют главным образом детей, а также лиц, подвергающихся по роду деятельности риску заражения (ветеринарные работники, зоотехники, рыболовы, работники очистных канализационных сооружений, складских помещений, рыбоводческих хозяйств, шахтеры, работники субпродуктовых и убойных цехов мясокомбинатов, санитарных боен и убойных пунктов). Ежегодно по данным Федеральной службы по надзору в сфере защиты прав потребителей и благополучия человека РФ вакцинации против лептоспирозов в зависимости от эпидемиологической обстановки подвергаются в среднем от 40 до 70 тыс. человек. Для профилактики заболевания лептоспирозами в нашей стране применяют вакцины, содержащие инактивированные нагреванием или формальдегидом культуры лептоспир основных серологических групп: Icterohaemorrhagiae, Grippotyphosa, Pomona, Sejroe и др. Такие препараты обеспечивают напряженный серовароспецифический иммунитет длительностью 6-12 месяцев.
Инактивированные вакцины против лептоспироза характеризуются высокой реактогенностью и большим риском различных поствакцинальных осложнений вплоть до сывороточной болезни и анафилактического шока. Кроме того, для производства традиционных вакцин требуются дорогие питательные среды для культивирования вакцинных штаммов, сложные условия культивирования, длительное наращивание биомассы лептоспир, а также велика опасность заражения персонала и контаминации окружающей среды.
Известны различные способы производства традиционных противолептоспирозных инактивированных вакцин. В основе этих препаратов лежит выращивание биомассы лептоспир, инактивирование ее формальдегидом и концентрация микробной массы. Такие поливалентные вакцины, содержащие штаммы лептоспир четырех серологических групп (RU 2184775, 10.07.2002) обладают высокими антигенными и протективными свойствами, однако вакцинация этими препаратами обеспечивает лишь серовароспецифический иммунный ответ, а перекрестный иммунитет (невосприимчивость к другим серогруппам лептоспир) отсутствует. Кроме того, длительность иммунного ответа при применении этих препаратов составляет 6-12 месяцев, а затем требуется повторная ревакцинация.
Производство традиционных противолептоспирозных вакцин и наработка специфических антигенов связаны с использованием патогенных штаммов лептоспир, что требует обеспечения особых условий безопасности, для культивирования лептоспир необходимы дорогие многокомпонентные питательные среды. Имеются многочисленные данные о том, что поствакцинальные осложнения часто связаны с недостаточной степенью очистки препаратов от компонентов питательных сред и инактивирующих агентов, используемых в технологическом процессе наращивания патогенных штаммов, а также с использованием различных консервантов, например формальдегида.
Традиционные противолептоспирозные вакцины во многом несовершенны, приводят к большому количеству осложнений, аллергизации, требуют сложных и дорогих условий культивирования вакцинных штаммов, использования многостадийных и сложных схем очистки.
В этой связи особенно актуальной является проблема создания вакцинных препаратов нового поколения, в частности, конструирование безопасной субъединичной вакцины на основе протективных антигенов Leptospira interrogans.
Эти проблемы могут быть решены при использовании генно-инженерных подходов по созданию многокомпонентных субъединичных вакцин, отвечающих современным требованиям.
Субъединичные генно-инженерные вакцины считаются наиболее эффективными и безопасными вакцинными препаратами, т.к. состоят из ограниченного количества высокоочищенных консервативных вакцинно-ценных белков и природных адъювантов, обеспечивающих одновременную защиту от нескольких серогрупп патогенных лептоспир.
Существующие подходы к созданию новых профилактических препаратов основываются главным образом на использовании очищенных иммуногенных компонентов микроорганизмов. К наиболее перспективным вакцинно-ценным антигенам лептоспир относят липополисахариды (ЛПС) и белки наружной оболочки (ОМР - outer membrane proteins). Использование ЛПС в качестве основного компонента противолептоспирозной вакцины проблематично, т.к. ЛПС лептоспир серовароспецифичны и не обеспечивают перекрестного иммунитета при инфицировании другим сероваром возбудителя. В связи с этим в настоящее время интенсивно изучаются различные белки лептоспир, преимущественно белки наружной мембраны (ОМР), на предмет наличия в них консервативных эпитопов для разных сероваров возбудителя.
Одними из кандидатных белков являются специфические консервативные иммуноглобулиноподобные белки Lig (Leptospiral immunoglobulin-like) - липопротеины наружной мембраны лептоспир. Они относятся к надсемейству бактериальных иммуноглобулиноподобных белков, многие из которых являются нефимбрильными адгезинами и белками проникновения. Все эти белки содержат от 10 и более тандемных повторов иммуноглобулиноподобных доменов размером около 90 аминокислотных остатков (Palaniappan R., Chang Yung-Fu, Jusuf S., Artiushin S., Timoney J., McDonough S., Barr S., Divers Т., Simpson K., McDonough P., Mohammed H. Cloning and Molecular Characterization of an Immunogenic LigA Protein of Leptospira interrogans Infect. Immun. Nov. 2002, p.5924-5930).
Гены ligA и ligB обнаружены только у патогенных лептоспир (Matsunaga J., Barocchi M., Croda J., Young Т., Sanchez Y., Siqueira I., Bolin C., Reis M., Riley L., Haake D., Ко A. Pathogenic Leptospira species express surface-exposed proteins belonging to the bacterial immunoglobulin superfamily Mol. Microbiol. 2003, 49(4), 929-945).
Ген белка LigA клонирован и экспрессирован в клетках Е. coli. Однако экспрессии полноразмерного гена LigA добиться не удалось, т.к. продукт был токсичен для Е. coli. Экспрессировать укороченный 90-кДа фрагмент белка (без сигнальной последовательности 31 а.о.) в Е.coli удалось только в виде химерного белка с GST, однако уровень экспрессии был очень низким (Palaniappan R., Chang Yung-Fu, Jusuf S., Artiushin S., Timoney J., McDonough S., Barr S., Divers Т., Simpson K., McDonough P., Mohammed H. Cloning and Molecular Characterization of an Immunogenic LigA Protein of Leptospira interrogans Infect. Immun. Nov. 2002, p.5924-5930).
При культивировании лептоспир in vitro продукты генов не обнаруживаются, экспрессия генов ligA и ligB наблюдается только in vivo, являясь, таким образом, маркером инфекции (Palaniappan R., Chang Yung-Fu, Jusuf S., Artiushin S., Timoney J., McDonough S., Barr S., Divers Т., Simpson K., McDonough P., Mohammed H. Cloning and Molecular Characterization of an Immunogenic LigA Protein of Leptospira interrogans Infect. Immun. Nov. 2002, p.5924-5930). Наличие антител к белкам LigA и LigB позволяет дифференцировать поствакцинальный иммунитет от инфекции. Кроме того, белок LigA представляет интерес в качестве вакцинно-ценного белка и может использоваться в качестве компонента субъединичной генно-инженерной вакцины (Palaniappan R., McDonough S., Divers Т., Chen C.-S., Pan M.-J., Matsumoto M., Chang Y.-F. Immunoprotection of Recombinant Leptospiral Immunoglobulin-Like Protein A against Leptospira interrogans Serovar Pomona Infection Infect Immun. 2006 March; 74(3): 1745-1750).
Несмотря на то, что данные специфические антигены могут использоваться как в диагностических целях, так и в качестве компонента вакцинных препаратов, их сложно получать традиционными способами. Поскольку гены белков LigA и LigB экспрессируются только in vivo, в данном случае клонирование генов этих белков является единственным способом наработать антигены в препаративных количествах.
В процессе производства современных конъюгированных вакцинных препаратов используются различные способы иммобилизации белков, основанные, как правило, на химической пришивке. Эти подходы многостадийны, трудоемки, дорогостоящи и небезопасны с точки зрения обеспечения экологической безопасности производства.
Аналогов иммуностимулирующих препаратов, основанных на использовании протективных антигенов лептоспир, которые иммобилизованы на полисахаридном носителе за счет аффинного взаимодействия, в доступных источниках информации не обнаружено.
В качестве ближайшего по технической сущности аналога принят инъекционный препарат, состоящий из рекомбинантного антигена TUL4 бактерии Francisella tularensis, иммобилизованного на углеводном сорбенте - TUL4spCBD (RU 2270249, кл. МПК С12N 1/21, 20 февраля 2006 г.).
Известный препарат содержит белково-целлюлозный комплекс, который состоит из рекомбинантного белка TUL4spCBD, обладающего антигенными свойствами белка TUL4 - основного иммунодоминантного антигена наружной мембраны Francisella tularensis, иммобилизованного на целлюлозном иммуносорбенте, благодаря наличию целлюлозосвязывающего домена (CBD) гена эндоглюканазы CelD из термофильного микроорганизма Anaerocellum thermophilum.
Препарат рекомбинантного белка TUL4spCBD, иммобилизованного на целлюлозе, предлагается использовать для иммунизации животных с целью получения гипериммунных сывороток к белку TUL4spCBD.
Однако известный препарат имеет недостатки, обусловленные использованием целлюлозы в качестве иммуносорбента. Несмотря на безвредность и отсутствие токсичности, целлюлоза не способна при парентеральной инъекции индуцировать неспецифическую воспалительную реакцию и, соответственно, обеспечивать формирование напряженного иммунного ответа, в частности высокого уровня синтеза антител. Поэтому использование целлюлозного сорбента в качестве носителя рекомбинантных антигенов требует применения дополнительного адъюванта для получения гипериммунной сыворотки, в качестве которого предлагается использовать адъювант Фрейнда, обладающий высокой реактогенностью.
Задачей изобретения является получение препарата белка, обладающего антигенными свойствами LigA.
Технический результат заключается в обеспечении связывания белка, обладающего антигенными свойствами LigA, с зимозаном, что позволяет обеспечивать развитие интенсивного иммунного ответа и высокий уровень синтеза антител без применения дополнительных адъювантов, а также в упрощении способа получения белка при высоком уровне его продукции.
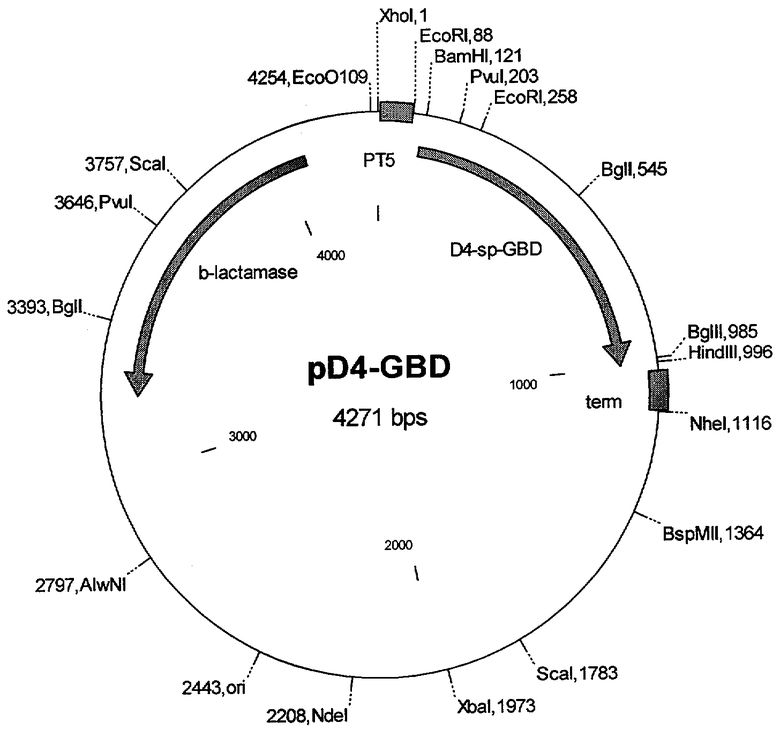
Сущность изобретения состоит в том, что рекомбинантная плазмида pD4spGBD размером 4271 обеспечивает экспрессию бифункционального рекомбинантного белка D4-GBD, состоящего из домена 4 белка LigA L. interrogans, спейсера из остатков глицина и серина и 1,3 β-глюкансвязывающего домена Cl. thermocellum (GBD), и содержит:
- искусственный бактериальный оперон химерного белка, включающий:
промоторную область раннего промотора бактериофага Т5 (7-87 п.н.), обеспечивающего эффективную транскрипцию мРНК,
рекомбинантный ген (115-994 н.п.), кодирующий целевой химерный белок D4-GBD,
нетранслируемую область терминации транскрипции бактериального оперона (1017-1113 н.п.), обеспечивающую эффективное окончание транскрипции;
- бактериальный оперон bla (4066-3206 н.п. комплементарной цепи), кодирующий бета-лактамазу, являющуюся селективным маркером для отбора клонов-трансформантов Е. coli методом контрселекции;
- бактериальный участок инициации репликации типа ColEl (2443 н.п.), обеспечивающий репликацию плазмиды в штаммах Е. coli.
Штамм Escherichia coli M15 [pREP4, pD4spGBD], полученный трансформацией штамма Escherichia coli M15 /pREP4 плазмидой pD4spGBD. Полученный штамм, депонированный во Всероссийской Коллекции Промышленных Микроорганизмов ФГУП ГосНИИгенетика под номером В-9901 от 21 декабря 2007 г., является продуцентом рекомбинантного белка D4-GBD, состоящего из домена 4 белка LigA L. interrogans, спейсера из остатков глицина и серина и 1,3 β-глюкансвязывающего домена Cl. thermocellum (GBD).
Рекомбинантный белок D4-GBD, состоящий из домена 4 белка LigA L. interrogans, спейсера из остатков глицина и серина и 1,3 β-глюкансвязывающего домена Cl. thermocellum (GBD), обладает антигенными свойствами белка LigA L. interrogans и способностью самопроизвольно связываться с 1,3-β-глюкансодержащим сорбентом - зимозаном А.
Способ получения рекомбинантного белка D4-GBD включает выращивание клеток штамма Escherichia coli M15 [pREP4, pD4spGBD], получение супернатанта, содержащего белок, иммобилизацию белка путем добавления к супернатанту суспензии 1,3-β-глюкансодержащего сорбента - зимозана А, инкубирование, отмывание сорбента со связавшимся белком, концентрирование иммобилизованного на зимозане А белка на колонке, снятие белка с сорбента, проведение очистки для получения целевого белка.
Способ исследования связывания белка D4-GBD с антителами сывороток больных включает определение титра специфических антител методом твердофазного непрямого иммуноферментного анализа, для проведения которого в качестве антигена используют рекомбинантный белок D4-GBD, сорбированный на дне лунок микропланшета.
Способ получения специфических антител к белку D4-GBD включает иммунизацию животных рекомбинантным белком D4-GBD, иммобилизованным на зимозане А, путем трехкратного введения препарата нарастающими дозами без применения дополнительных адъювантов, с последующим отбором крови для определения титра специфических антител, приготовление сыворотки и проведение твердофазного непрямого иммуноферментного анализа для определения титра специфических антител.
Таким образом, технический результат достигается за счет того, что создана рекомбинантная плазмида pD4spGBD, кодирующая бифункциональный рекомбинантный белок D4-GBD, с одной стороны содержащий аминокислотную последовательность домена 4 белка LigA, а с другой обладающий способностью самостоятельно связываться с 1,3-β-глюкансодержащим сорбентом, благодаря наличию 1,3-β-глюкансвязывающего домена.
Также указанный технический результат достигается тем, что штамм Escherichia coli M15 [pREP4, pD4spGBD], депонированный во Всероссийской Коллекции Промышленных Микроорганизмов ФГУП ГосНИИгенетика под номером В-9901 от 21 декабря 2007 г., является продуцентом рекомбинантного белка D4-GBD. При этом отсутствие в клетках Е.coli белков, способных связываться с зимозаном А, служит гарантией того, что рекомбинантный белок D4-GBD является единственным белком штамма Escherichia coli M15 [pREP4, pD4spGBD], прочно связывающимся с используемым сорбентом, что обеспечивает возможность одностадийного получения высокоочищенного препарата указанного рекомбинантного белка, иммобилизованного на зимозане А.
Указанный технический результат достигается также тем, что рекомбинантный белок D4-GBD, включающий в себя аминокислотную последовательность, определяющую способность данного белка связываться с 1,3-β-глюканом (зимозаном), позволяет проводить в одну стадию концентрирование, очистку и иммобилизацию белка D4-GBD на 1,3-β-глюкане (зимозане А). Иммобилизация на зимозане обеспечивается за счет присутствия в рекомбинантном белке 1,3-β-глюкансвязывающего домена эндоглюканазы LicA из термофильного микроорганизма Clostridium thermocellum, который обладает высоким сродством к различным 1,3-β-глюканам и обеспечивает прочное связывание с носителем в широком диапазоне значений рН 4-11, температуры 0-75°С и концентраций соли 0-5 М NaCl.
Ключевым моментом разработанной авторами новой технологии является использование зимозана А для аффинной иммобилизации рекомбинантного белка, который обладает выраженными иммуностимулирующими свойствами в отличие от целлюлозного сорбента, обеспечивает развитие интенсивного иммунного ответа и высокий уровень синтеза антител без применения дополнительных адъювантов. При этом сохраняются все достоинства одностадийного выделения и очистки рекомбинантных антигенов за счет аффинной иммобилизации: простота, доступность, экономичность и отсутствие токсичных реагентов.
Препарат зимозан А (1,3-1,6-бета-глюкан дрожжей) разрешен для перорального применения, а также для внутривенного введения как инъекционный препарат (Закенфельд Г.К. Иммунологический механизм действия полисахаридов дрожжевых клеток Saccharomyces cerevisiae. Рига 1990; 152). Зимозан индуцирует неспецифический иммунитет посредством взаимодействия с Toll-like рецепторами TLR2 и TLR6 на поверхности иммунокомпетентных клеток (Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol 1999; 17: 593-623; Dillon S., Agrawal S., Banerjee K. et al. Yeast zymosan, a stimulus for TLR2 and dectin-1, induces regulatory antigenpresenting cells and immunological tolerance. The Journal of Clinical Investigation April 2006, Vol.116 Num. 4: p.916-928; Pasare, C., and Medzhitov, R. 2004. Toll-like receptors and acquired immunity. Semin. Immunol. 16:23-26.).
Основным действующим началом препарата зимозан является 1,3-1,6-β-глюкан, широко распространенный в природе биосовместимый и биодеградируемый полисахарид, который входит в состав клеточной стенки многих растений, водорослей и грибов (Беседнова Н.Н., Иванушко Л.А., Звягинцева Т.Н., Елякова Л.А. Иммунотропные свойства 1,3-1,6-β-D-глюканов). Зимозан способен обеспечивать устойчивость широкого спектра к различным грибным патогенам, содержащим в своем составе 1,3-1,6-β-глюкан (Sato M., Sano Н., Iwaki D. et al. Direct Binding of Toll-Like Receptor 2 to Zymosan, and Zymosan-Induced NF-B Activation and TNF-α Secretion Are Down-Regulated by Lung Collectin Surfactant Protein A. The Journal of Immunology April 25, 2003; p.417-425).
Для преодоления описанных сложностей и получения иммуностимулирующих препаратов, состоящих только из протективных антигенов, иммобилизованных на полисахаридном сорбенте, который может выполнять функции адъюванта, предлагается новая технология, обеспечивающая одновременное выделение, очистку, и концентрирование рекомбинантного белка за счет его аффинной иммобилизации на 1,3-β-глюкансодержащем сорбенте (зимозане А) и позволяющая в результате получать готовый для инъекции препарат.
Предлагаемая авторами технология получения в одну стадию иммуногенной композиции, содержащей иммобилизованные на зимозане А рекомбинантные антигены, заключается в конструировании двухкомпонентного белка, содержащего целевой белок (антиген) и специальный домен, соединенные спейсерной последовательностью. Способность связываться с зимозаном А придает рекомбинантному белку встроенный в него специальный белковый модуль - 1,3-β-глюкансвязывающий домен (GBD). GBD самопроизвольно взаимодействует с зимозаном А с высокой константой связывания. При этом исходные антигенные свойства второго (основного) домена рекомбинантного белка при присоединении GBD не нарушаются. Использование данной технологии позволяет сделать процесс иммобилизации и очистки белковых антигенов одностадийным и практически безинструментальным. Кроме того, предлагаемая технология обеспечивает экологическую безопасность производства, поскольку при этом снимается необходимость использования патогенных штаммов и токсичных химических реагентов.
Достигается указанный технический результат также тем, что способ очистки, концентрирования и получения иммобилизованного рекомбинантного белка D4-GBD включает обработку супернатанта гидролизата клеток Escherichia coli M15 [pREP4, pD4spGBD] суспензией 1,3-β-глюкансодержащего сорбента - зимозана А.
Изобретение поясняется чертежом, на котором приведена схема плазмиды pD4spGBD, нуклеотидная последовательность плазмиды pD4spGBD - последовательность №1, аминокислотная последовательность белка D4 - последовательность №2, глицин-сериновый спейсер - последовательность №3, глюкансвязывающий домен GBD - последовательность №4.
Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам (Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. М.: Мир, 1984; Клонирование ДНК. Методы./ Под ред. Д. Гловера. Пер. с англ. Москва, Мир, 1988; Saiki R.K., Gelfand D.H., Stoffel S., Sharf S.J., Higuchi R., Horn G.T., Mullis K.B., Elich H.A. Science, 1988. V.239, №4839, p.487-491; Sanger F., Nicklen S., Coulson A.R. Proc. Nat. Acad. Sci. USA, 1977, V.74, p.5463-5467).
Примеры
Пример 1. Получение рекомбинантной плазмиды pD4spGBD
а) Химический синтез олигодезоксирибонуклеотидов
Олигодезоксирибонуклеотиды были синтезированы твердофазным амидофосфитным методом и очищены методом электрофореза в 12%-ном полиакриламидном геле.
б) Получение фрагмента гена ligA с последующим его клонированном
Копии фрагмента гена ligA, кодирующего домены 4 белка LigA, получали методом полимеразной цепной реакции, для чего были выбраны праймеры, комплементарные началу и концу интересующей области ДНК. Используя в качестве ДНК-матрицы хромосомную ДНК Leptospira interrogans serovar Copenhageni, методом ПЦР с использованием олигонуклеотидных праймеров состава (подчеркнуты сайты рестриктаз BamHI, XbaI, и BglII)

амплифицировали последовательность нуклеотидов, соответствующую домену 4 гена LigA. Реакцию проводили в 25 мкл реакционной смеси, содержащей 2 мкг ДНК, 10 пкМ каждого праймера A4-For и A4-Rev, 67 мМ трис-HCl (рН 8,8 при 25°С), 15 мМ сульфата аммония, 2,5 мМ хлористого магния, 0,01% Твин-20, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. Taq-ДНК-полимеразы. Реакцию амплификации проводили под вазелиновым маслом по программе: 95°C - 10 мин, 35 циклов (95°C - 30 сек, 67°C - 30 сек, 72°C - 30 сек), затем 72°C - 5 мин с применением активного точного режима регулирования. Синтезированные фрагменты ДНК размером 310 н.п. очищали однократной экстракцией реакционной смеси хлороформом и переосаждением этанолом с добавлением ацетата аммония и затем растворяли в воде.
Для клонирования данного продукта полученные ПЦР-фрагменты, а также ДНК плазмиды pQE16 (Qiagene, USA) гидролизовали рестрикционными эндонуклеазами BamHI и XbaI при 37°C в буфере, содержащем 66 мМ Трис-ацетат (рН 7,9 при 37°C), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Выделенные из геля рестрикционные фрагменты ДНК, соответствующие домену 4 гена ligA размером 310 н.п. и фрагменты плазмиды pQE16 размером 2419 н.п., лигировали с помощью ДНК-лигазы бактериофага Т4. Полученной лигированной смесью трансформировали клетки Е.coli M15[pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с селективными антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI и XbaI, а также рестриктаз, сайты которых находятся внутри клонируемых фрагментов, и отбирали клоны pD4, плазмидные ДНК которых размером 2712 н.п. содержат последовательность домена 4. Первичную структуру полученной плазмиды pD4 подтверждали секвенированием по методу Сэнгера.
в) Получение последовательности, кодирующей Gly-Ser спейсер, с последующим ее клонированном
Для получения последовательности спейсера, состоящего из повторов аминокислотных остатков Gly-Ser, были синтезированы два комплементарных олигонуклеотида следующего состава (подчеркнут сайт эндонуклеазы рестрикции BglII, под нуклеотидной последовательностью первого олигонуклеотида приведена кодируемая последовательность аминокислот):
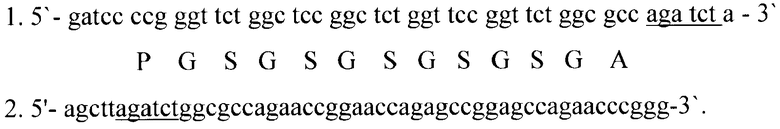
Для получения двуцепочечного фрагмента ДНК смешивали по 20 пкМ каждого олигонуклеотида, прогревали смесь 10 мин при 95°C и охлаждали в течение 4 часов до 25°C. Для клонирования данного продукта ДНК плазмиды pQE13 гидролизовали рестрикционными эндонуклеазами BamHI и HindIII при 37°C в буфере, содержащем 66 мМ Трис-ацетата (рН 7,9 при 37°C), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Выделенный из геля рестрикционный фрагмент плазмиды pQE13 размером 3420 н.п. объединяли с двуцепочечным фрагментом ДНК, кодирующим спейсер, и лигировали с помощью ДНК-лигазы бактериофага Т4. Полученной лигированной смесью трансформировали клетки Е. coli M15[pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI и HindIII, а также рестриктаз, сайты которых находятся внутри клонируемых фрагментов, и отбирали клоны pRsp, плазмидная ДНК которых (размером 3471 н.п.) содержит последовательность Gly-Ser спейсера. Первичную структуру полученной плазмиды подтверждали секвенированием по методу Сэнгера.
г) Получение участка гена licA Clostridium thermocellum, содержащего последовательность 1,3-β-глюкансвязывающего домена, с последующим его клонированном в плазмидную конструкцию pRsp
Копию участка гена licA получали методом полимеразной цепной реакции, для чего были выбраны праймеры, соответствующие началу и концу данной области ДНК. Используя в качестве ДНК-матрицы хромосомную ДНК Clostridium thermocellum, методом ПЦР с использованием олигонуклеотидных праймеров состава (подчеркнуты сайты рестриктаз BamHI и HindIII и BglII):
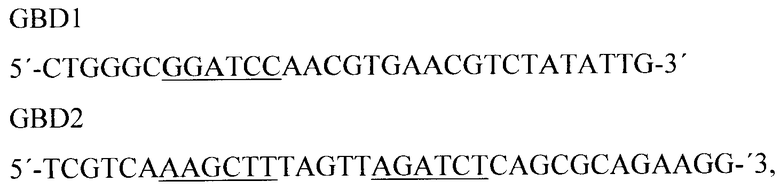
амплифицировали интересующую последовательность нуклеотидов. Реакцию проводили в 25 мкл реакционной смеси, содержащей 2 мкг ДНК, 10 пкМ каждого праймера, 67 мМ трис-HCl (рН 8,8 при 25°C), 15 мМ сульфата аммония, 2,5 мМ хлористого магния, 0,01% Твин-20, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1-2 ед. Taq-ДНК-полимеразы. Реакцию амплификации проводили под вазелиновым маслом: 5 циклов: 93°C - 2 мин, 57°C - 5 мин и 72°C - 30 сек; 30 циклов: 93°C - 1 мин, 63°C - 1 мин и 72°C - 30 сек; затем 63°C - 5 мин и 72°C - 10 мин. Продукт амплификации размером 528 н.п. обрабатывали хлороформом, переосаждали этиловым спиртом и растворяли в воде. Для клонирования данного продукта полученную ДНК PCRGBD гидролизовали рестрикционными эндонуклеазами BamHI и HindIII при 37°C в буфере, содержащем 66 мМ Трис-ацетата (рН 7,9 при 37°C), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Также ДНК плазмиды pRsp гидролизовали рестрикционными эндонуклеазами BglII и HindIII. Выделенные из геля рестрикционные фрагменты: фрагмент ДНК, соответствующий участку гена licA размером 548 н.п., и фрагмент плазмиды pRsp размером 3420 п.н., содержащий селективный маркер, участок инициации репликации, промоторную область и последовательность, кодирующую Gly-Ser спейсер, лигировали с помощью ДНК-лигазы фага Т4. Полученной лигированной смесью трансформировали клетки Е. coli M15 [pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI и HindIII, а также рестриктаз, находящихся внутри клонируемых фрагментов, и отбирали клоны pspGBD (размером 4013 н.п.), плазмидная ДНК которых содержит последовательность участка гена licA со спейсером на N-конце, состоящим из Gly-Ser повторов. Первичную структуру полученной плазмиды подтверждали секвенированием по методу Сэнгера.
д) Получение плазмиды, содержащей последовательности фрагмента гена ligA, Gly-Ser спейсера, и 1,3-β-глюкансвязывающего домена GBD.
Для получения плазмиды, содержащей последовательности фрагмента гена ligA, Gly-Ser спейсера, и 1,3-β-глюкансвязывающего домена GBD, плазмиду pD4 гидролизовали рестрикционными эндонуклеазами XhoI и BglII при 37°C в буфере 66 мМ Трис-ацетат (рН 7,9 при 37°C), 20 мМ ацетат магния, 132 мМ ацетат калия и 0,2 мг/мл BSA в течение 1 часа, а плазмиду pspGBD гидролизовали рестрикционными эндонуклеазами XhoI и BamHI при 37°C в буфере, содержащем 50 мМ Трис-HCl (рН 7,5 при 37°C), 10 мМ хлорида магния, 100 мМ хлорида натрия и 0,1 мг/мл BSA, в течение 1 часа. Выделенные из геля рестрикционные фрагменты: фрагменты плазмидной ДНК pD4, включающие участок гена ligA размером 402 н.п., а также фрагмент ДНК плазмиды pspGBD, включающий Gly-Ser спейсер и последовательность 1,3-β-глюкансвязывающего домена GBD, размером 3869 н.п., лигировали с помощью ДНК-лигазы бактериофага Т4. Полученной лигированной смесью трансформировали клетки Е. coli M15 [pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI, BglII, а также рестриктаз, сайты которых находятся внутри клонируемого фрагмента, и отбирали клоны pD4spGBD, плазмидная ДНК которых содержит последовательность домена 4 гена LigA, Gly-Ser спейсера и 1,3-β-глюкансвязывающего домена GBD размером 4271 н.п. Первичную структуру полученной плазмиды pD4spGBD подтверждали секвенированием по методу Сэнгера.
Пример 2. Получение штамма Е.coli [pREP4, pD4spGBD] - продуцента рекомбинантного белка D4-GBD, состоящего из рекомбинантного домена 4 белка LigA, Gly-Ser спейсера, 1,3-β-глюкансвязывающего домена GBD
Для получения штамма Е.coli [pREP4, pD4spGBD] - продуцента рекомбинантного белка D4-GBD клетки штамма Е.coli M15 [pREP4] трансформировали плазмидной ДНК pD4spGBD.
Полученный штамм Е.coli M15 [pREP4, pD4spGBD] характеризуется следующими признаками.
Культурально-морфологические признаки. Клетки прямые, палочковидные, неподвижные, грамотрицательные. При рассеве на чашке с 1,5% LB-агаром рост в виде отдельных колоний, иногда в R-форме с неровными краями. Хорошо растет на плотных и жидких питательных средах (LB-бульон, LB-агар, МПА, МПБ).
Физиолого-биохимические признаки. Клетки растут в пределах 4-42°C при оптимуме рН 6,8-7,5. Штаммы разлагают глюкозу, маннит с образованием кислоты, не разлагает сахарозу, арабинозу, галактозу, сбраживает мальтозу, ксилозу, сорбит, рамнозу.
Устойчивость к антибиотикам. Существенным при использовании данных штаммов является их чувствительность к налидиксовой кислоте (25 мг/мл), стрептомицину (20 мг/мл) и рифампицину (25 мг/мл). Проявляют устойчивость к ампициллину (до 100 мкг/мл), обусловленную наличием рекомбинантной плазмиды pD4spGBD, и к канамицину (до 25 мкг/мл), обусловленную наличием плазмиды pREP4.
Полученный штамм депонирован во Всероссийской Коллекции Промышленных Микроорганизмов ФГУП ГосНИИгенетика под номером В-9901 от 21 декабря 2007 г. и является продуцентом рекомбинантного белка D4-GBD.
Трансформированные клетки культивировали на селективной агаризованной среде, содержащей штаммовый антибиотик - канамицин и плазмидный антибиотик - ампициллин.
Пример 3. Способ получения препарата белка D4-GBD, иммобилизированного на 1,3-β-глюкане (зимозане), и раствора D4-GBD.
Штамм-продуцент белка D4-GBD Е.coli M15 [pREP4, pD4spGBD] выращивали в 500-1000 мл среды LB с ампициллином и канамицином при 37°C до оптической плотности, соответствующей 1 ед. поглощения при длине волны 600 нм, индуцировали добавлением 100-150 мкл 0,1 М раствора изопропил-бета-D-тиогалактопиранозида и выращивали в течение 3-4 часов. Клетки осаждали центрифугированием при 6000 об/мин в течение 10 мин.
Выделение рекомбинантного белка проводили следующим образом. Биомассу суспендировали в лизатном буфере следующего состава: 50 мМ Трис, рН 8,0; 0,5 М NaCl; 5 мМ MgCl2; 1/1000 объема PMSF; 0,1% Triton Х-100; 0,5 мг/мл лизоцим. Затем лизат разрушали ультразвуком, клеточный дебрис удаляли центрифугированием при 15000 об/мин в течение 20 мин.
Для иммобилизации белка к супернатанту добавляли равный объем 20%-ной суспензии зимозана A (Sigma, США), инкубировали 1 час при перемешивании. Сорбент со связавшимся белком 3 раза отмывали от клеточного лизата в 10-кратном объеме буфера, содержащем 0,5 М NaCl и 0,5% Triton X-100.
Для концентрирования раствора белка D4-GBD зимозан А с иммобилизованным белком вносили в колонку, снимали белок с сорбента раствором 4 М мочевины и проводили очистку методом диализа в течение ночи против раствора PBS (на 1 литр: 8 г NaCl, 0,2 г KCl, 2,7 г Na2HPO4×7H2O, 0,24 г KH2PO4, рН 4,0).
Полученный препарат белка D4-GBD анализировали методом электрофореза по Лэммли в 12% полиакриламидном геле в стандартной системе буферов, с окрашиванием гелей 0,15% раствором Кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и последующим отмыванием в 10% уксусной кислоте.
Концентрация указанного белка составляла 2-7 мг на 1 мл сорбента для иммобилизованного белка D4-GBD, 3-5 мг/мл для раствора белка D4-GBD. Степень чистоты всех препаратов составляла не менее 95%.
Пример 4. Индукция синтеза белка D4-GBD в клетках штамма-продуцента Е.coli M15 [pREP4, pD4spGBD]
Клетки штамма Е.coli [pREP4, pD4spGBD] выращивали в 500 мл среды LB при 37°C до оптической плотности, соответствующей 1 ед. поглощения при длине волны 600 нм, индуцировали 150 мкл 0,1 М раствора изопропил-бета-D-тиогалактопиранозида и выращивали в течение 4 часов.
Для контроля продукции рекомбинантного белка D4-GBD клетками Е. coli M15 [pREP4, pD4spGBD] применяли метод электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Разделение белков проводили в 12% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ Tris-HCl, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). По окончании электрофореза гели окрашивали 0,15% раствором Кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывали в 10% уксусной кислоте. При сравнении спектра белков у штаммов Е.coli M15 [pREP4] и Е.coli M15 [pREP4, pD4spGBD] после добавления индуктора ИПТГ обнаруживали появление дополнительных белковых полос с молекулярной массой 31,8 кДа, что соответствует расчетной молекулярной массе рекомбинантного белка D4-GBD.
Уровень синтеза белка D4-GBD определяли, сравнивая интенсивность окрашивания полосы рекомбинантного белка D4-GBD с полосой соответствующего белка - стандарта молекулярной массы.
Согласно полученным данным, клетки штамма Е.coli M15 [pREP4, pD4spGBD] синтезируют около 50 мг белков D4-GBD на литр культуры клеток при концентрации 109 клеток на мл. Рекомбинантный белок D4-GBD не угнетает рост клеток штамма-продуцента и не является токсичным для них, а присутствие 1,3-β-глюкансвязывающего домена в составе белка D4-GBD защищает его от деградации протеазами клеток Е.coli.
Пример 5. Способ исследования взаимодействия рекомбинантного белка D4-GBD с антигенами сывороток больных лептоспирозом
Для исследования взаимодействия антител сывороток больных с рекомбинантным белком D4-GBD использовали пулы сывороток клинически здоровых и больных лептоспирозом в различные сроки болезни из коллекции лаборатории лептоспирозов ГУ НИИЭМ им. Н.Ф.Гамалеи.
Полистироловую 96-луночную микропанель (Corning) иммобилизовали химерным белком D4-GBD в концентрации 30 мкг/мл на лунку, суспендированным в 30 мМ калий-карбонатном буфере рН 9,2-9,6 в течение 24 часов при +4°C и перемешивании. Лунки дважды промыли дистиллированной водой и трижды фосфатно-буферным раствором (PBS) с 0,05% Tween 20 (PBST). Затем в лунки вносили разведения исследуемых сывороток, приготовленные на PBST, начиная с разведения 1:100. Сыворотки титровали методом двукратных разведений и инкубировали в течение часа на шейкере при +37°C. Затем панель промывали аналогичным образом, и в лунки вносили антивидовой иммунопероксидазный конъюгат (1,0 мг/мл, ООО фирма ИМТЕК) в рабочем разведении 1:10000. Инкубацию проводили при +37°C на шейкере. Затем планшет отмывали, и для проявления реакции в лунки вносили по 100 мкл готовой субстрат-индикаторной смеси, содержащей 0,01% 3,3',5,5'-тетраметилбензидина (ТМБ). Инкубацию с субстратом проводили при комнатной температуре в течение 15 минут. Останавливали реакцию добавлением 50 мкл 8 М H2SO4 в каждую лунку. Оптическое поглощение (ОП) окрашенного продукта пероксидазной реакции определяли на фотометре Multiscan Ascent (ThermoLabsystems) при 450 нм. Титром антител считали обратную величину последнего разведения исследуемого образца, в котором значение оптической плотности (ОП450) в 2 раза превышало ОП450 в отрицательном контроле.
Результаты непрямого ИФА белка D4-GBD с пулами сывороток показали, что указанный рекомбинантный белок взаимодействует с антителами сывороток людей, инфицированных лептоспирами сероваров Icterohaemorrhagiae и Canicola, причем титры сывороток в ИФА совпали с титрами в РМА. Сыворотки, содержащие антитела к сероварам Grippotyphosa, Sejroe, а также сыворотки клинически здоровых людей в непрямом ИФА показали отрицательный результат.
Пример 6. Способ получения специфических антител к рекомбинантному белку D4-GBD
Специфические антитела к рекомбинантному белку D4-GBD получали по специальной схеме. Иммобилизованный на зимозане А рекомбинантный белок D4-GBD и раствор свободного белка D4-GBD использовали для иммунизации кроликов породы шиншилла. Состояние животных в процессе иммунизации было удовлетворительным, в месте инъекций не наблюдалось патологических процессов, изъязвлений кожных покровов, состояние места инъекции не отличалось от неповрежденных кожных покровов животного.
Схема иммунизации включала трехкратное введение препарата возрастающими дозами в количестве 100-300 мкг белка/кг с интервалами 2-3 недели. Отбор крови у иммунизированных кроликов проводили через 10 дней после последней инъекции. Полученные сыворотки исследовали методом непрямого иммуноферментного анализа. Титр специфических антител к белку D4-GBD в полученных сыворотках составил 10000.
Таким образом, конструирование рекомбинантной плазмиды pD4spGBD с использованием технологии создания слитных белков позволяет обеспечить высокий уровень экспрессии домена 4 белка LigA в составе химерного белка D4-GBD с 1,3-β-глюкансвязывающим доменом (GBD) в клетках штамма-продуцента, в то время как отдельный аутентичный домен 4 белка LigA в клетках Е. coli экспрессировать не удается.
Использование штамма M15 Escherichia coli [pREP4, pD4spGBD] обеспечивает высокий уровень синтеза рекомбинантного белка D4-GBD - до 10% от тотального белка клетки - при индукции экспрессии изопропил-тио-галактопиранозидом (ИПТГ).
Предлагаемый способ получения белка позволяет проводить выделение, очистку концентрирование белка и формулирование инъекционного препарата в одну стадию за счет иммобилизации на 1,3-β-глюкансодержащем сорбенте - зимозане А, что значительно сокращает время проведения данных операций и снижает затраты в несколько раз, поскольку является практически безинструментальным процессом и не требует использования дорогостоящих и токсичных реагентов, и в то же время позволяет получить препарат высокой степени чистоты (не ниже 95%), пригодный для иммунизации животных.
Способ исследования связывания белка с антителами позволяет впервые оценить сохранение антигенных свойств рекомбинантного домена 4 белка LigA при его синтезе в составе химерного белка с 1,3-β-глюкансвязывающим доменом (GBD) для обеспечения гарантированного специфического взаимодействия рекомбинантного домена 4 с антителами сывороток больных лептоспирозом.
Использование предлагаемого способа получения специфических антител позволяет иммунизировать животных полученной суспензией частиц зимозана А с иммобилизованным на них белком без применения дополнительных адъювантов, не вызывает побочных эффектов у животных, в отличие, например, от адъюванта Фрейнда, и в то же время обеспечивает высокий уровень синтеза специфических антител при использовании невысоких доз антигена.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD1spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D1-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D1-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D1-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D1-GBD | 2008 |
|
RU2401302C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD5spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D5-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D5-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D5-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D5-GBD | 2008 |
|
RU2401304C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-CFP10-DBD, РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli M15 [pREP4, pESAT6-CFP10-DBD], СПОСОБ ПОЛУЧЕНИЯ, ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ESAT6-CFP10-DBD НА ДЕКСТРАНЕ, РЕКОМБИНАНТНЫЙ БЕЛОК ESAT6-CFP10-DBD И ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК ESAT6-CFP10-DBD | 2013 |
|
RU2539026C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА (ВАРИАНТЫ), ШТАММ ESCHERICHIA COLI (ВАРИАНТЫ) - ПРОДУЦЕНТ ХИМЕРНЫХ БЕЛКОВ, ХИМЕРНЫЙ БЕЛОК (ВАРИАНТЫ), СПОСОБ ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНЫХ БЕЛКОВ НА ЦЕЛЛЮЛОЗЕ, СПОСОБ ИММОБИЛИЗАЦИИ РЕКОМБИНАНТНЫХ БЕЛКОВ НА ПОЛИСТИРОЛЬНЫХ НОСИТЕЛЯХ | 2008 |
|
RU2378371C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА pESAT6-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-DBD, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК ESAT6-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2520737C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА Ag85A-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-DBD, ШТАММ Escherichia coli [pREP4, pAg85A-DBD], ХИМЕРНЫЙ БЕЛОК Ag85A-DBD | 2013 |
|
RU2520078C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422524C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422525C1 |



Изобретение относится к биотехнологии и представляет собой плазмидную ДНК pD4spGBD, обеспечивающую экспрессию рекомбинантного антигена - домена 4 белка LigA L. interrogans в составе химерного белка D4-GBD с 1,3-β-глюкансвязывающим доменом (GBD), и получения на их основе субъединичной генно-инженерной вакцины против лептоспироза. Изобретение позволяет обеспечивать развитие интенсивного иммунного ответа и высокий уровень синтеза антител без применения дополнительных адъювантов. 6 н.п. ф-лы, 1 ил.
1. Рекомбинантная плазмида pD4spGBD размером 4271 н.п., обеспечивающая экспрессию бифункционального рекомбинантного белка D4-GBD, состоящего из домена 4 белка LigA L. interrogans, спейсера из остатков глицина и серина и 1,3 β-глюкансвязывающего домена С1. thermocellum (GBD), содержащая:
искусственный бактериальный оперон химерного белка, включающий: промоторную область раннего промотора бактериофага Т5 (7-87 п.н.), рекомбинантный ген (115 - 994 н.п.), кодирующий целевой химерный белок D4-GBD,
нетранслируемую область терминации транскрипции бактериального оперона (1017-1113 н.п.);
бактериальный оперон bla (4066-3206 н.п. комплементарной цепи), кодирующий бета-лактамазу, являющуюся селективным маркером для отбора клонов-трансформантов Е. coli методом контр-селекции;
бактериальный участок инициации репликации типа ColEl (2443 н.п.).
2. Штамм Escherichia coli M15 [pREP4, pD4spGBD], полученный трансформацией штамма Escherichia coli M15 /pREP4 плазмидой pD4spGBD, депонированный во Всероссийской Коллекции Промышленных Микроорганизмов ФГУП ГосНИИгенетика под номером В-9901 - продуцент рекомбинантного белка D4-GBD, состоящего из домена 4 белка LigA L. Interrogans, спейсера из остатков глицина и серина и 1,3 β-глюкансвязывающего домена Cl. thermocellum (GBD).
3. Способ получения рекомбинантного белка D4-GBD, включающий выращивание клеток штамма Escherichia coli М15 [pREP4, pD4spGBD], получение супернатанта, содержащего белок, иммобилизацию белка путем добавления к супернатанту суспензии 1,3-β-глюкансодержащего сорбента - зимозана А, инкубирование, отмывание сорбента со связавшимся белком, концентрирование иммобилизованного на зимозане А белка на колонке; снятие белка с сорбента, проведение очистки для получения целевого белка.
4. Рекомбинантный белок D4-GBD, состоящий из домена 4 белка LigA L. interrogans, спейсера из остатков глицина и серина, и 1,3 β-глюкансвязывающего домена Cl. thermocellum (GBD), обладающий антигенными свойствами белка LigA L. interrogans, и способностью самопроизвольно связываться с 1,3-β-глюкансодержащим сорбентом - зимозаном А.
5. Способ исследования связывания белка D4-GBD по п.4 с антителами сывороток больных, включающий определение титра специфических антител методом твердофазного непрямого иммуноферментного анализа, для проведения которого в качестве антигена используют рекомбинантный белок D4-GBD по п.4, сорбированный на дне лунок микропланшета.
6. Способ получения специфических антител к белку D4-GBD по п.4, включающий иммобилизацию белка D4-GBD по п.4 на 1,3-β-глюкансодержащем сорбенте - зимозане А, иммунизацию животных иммобилизованным рекомбинантным белком D4-GBD путем трехкратного введения препарата нарастающими дозами без применения дополнительных адъювантов, с последующим отбором крови для определения титра специфических антител, приготовление сыворотки и проведение твердофазного непрямого иммуноферментного анализа для определения титра специфических антител.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| PALANIAPPAN R | |||
| et | |||
| al | |||
| Cloning and molecular characterization of an immunogenic LigA protein of Leptospira interrogans | |||
| Infect Immun | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| MATSUNAGA J | |||
| et | |||
| al | |||
| Pathogenic Leptospira species express surface-exposed proteins belonging to the bacterial immunoglobulin superfamily | |||
| Mol Microbiol | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

