Область техники
[1] Настоящее изобретение относится к клонированному представителю семейства псовых и к способу его получения. Более конкретно, настоящее изобретение относится к способу получения клонированного представителя семейства псовых, включающему энуклеацию зрелого ооцита представителя семейства псовых с получением энуклеированного ооцита-реципиента, проведение переноса ядра в энуклеированный ооцит с использованием соматической клетки представителя семейства псовых в качестве клетки-донора ядра в оптимизированных условиях таким образом, чтобы получить эмбрион с трансплантированным ядром, и перенос эмбриона с трансплантированным ядром в яйцевод суррогатной матери, а также к клонированному представителю семейства псовых, полученному при помощи этого способа.
Предшествующий уровень техники
[2] Благодаря недавним достижениям в развитии технологии трансплантации ядра соматической клетки посредством слияния клеток или внутрицитоплазматической клеточной инъекции получение клонированных животных осуществляют в действительности.
[3] Технология трансплантации ядра соматической клетки, представляющая собой технологию, дающую возможность рождения живых потомков без прохождения мейоза и формирования гаплоидных половых клеток, что обычно происходит в процессе размножения, представляет собой способ формирования новых индивидуумов при помощи трансплантации диплоидных соматических клеток взрослых особей в энуклеированные клетки для формирования эмбрионов, и трансплантации эмбрионов in vivo. Как правило, при применении технологии трансплантации ядра соматической клетки ооциты-реципиенты, которые переносят с ядрами соматических клеток-доноров, используют после их искусственного выращивания in vitro до метафазы II мейоза. Затем, чтобы предотвратить развитие хромосомных аномалий, являющихся результатом трансплантации ядра соматической клетки, из зрелых ооцитов удаляют ядра перед трансплантацией соматических клеток. После инъецирования соматических клеток в перивителлиновое пространство или в цитоплазму зрелых ооцитов энуклеированные ооциты и соматические клетки физически сливают друг с другом с помощью электрической стимуляции. Слитую пару активируют путем электрической стимуляции или при помощи химических веществ и переносят суррогатным матерям для получения живого потомства.
[4]
[5] Такая технология трансплантации ядра соматической клетки может быть широко использована в данной области, например, в целях размножения высших животных, сохранения редких или почти вымерших животных, продукции некоторых питательных веществ, продукции терапевтических биоматериалов, продукции животных для трансплантации органов, продукции животных с заболеваниями или расстройствами, продукции животных, ценных в медицинском отношении для замещения или трансплантации органа, а также восстановления клетки или гена.
[6]
[7] Технология клонирования животных впервые была осуществлена доктором. Вильмутом из Института Рослина, Англия, путем взятия клетки молочной железы от шестилетней овцы, трансплантации данной клетки в энуклеированный ооцит с получением эмбриона с трансплантированным ядром и переноса эмбриона in vivo с получением таким образом клонированного животного, Долли. С тех пор получили клонированных коров, мышей, гусей, свиней и кроликов с помощью трансплантации ядер с использованием соматических клеток, полученных у взрослых животных (WO 9937143A2, ЕР 930009 А1, WO 9934669 A1, WO 9901164 A1 и US 5945577).
[8] Между тем, интерес многих людей привлекало клонирование не только сельскохозяйственных животных, таких как коровы и свиньи, но и клонирование других домашних животных, таких как собаки. Недавно из числа домашних животных впервые клонировали кошку, а также проводили исследования по клонированию собаки.
[9] Тем не менее, до сих пор отсутствуют сообщения об успешном клонировании собак способом трансплантации соматической клетки.
Описание изобретения
Техническая задача
[10] Соответственно, авторы настоящего изобретения предприняли исследования для разработки способа клонирования представителя семейства псовых и, в результате, впервые получили клонированных представителей семейства псовых способом трансплантации соматической клетки в условиях, оптимизированных для электрического слияния, активации эмбриона с трансплантированным ядром и переноса эмбриона суррогатной матери, таким образом, выполнив настоящее изобретение.
[11]
[12] Следовательно, задача настоящего изобретения заключается в создании способа получения эмбриона представителя семейства псовых с трансплантированным ядром с использованием технологии трансплантации ядра соматической клетки.
[13] Другая задача настоящего изобретения заключается в создании эмбриона представителя семейства псовых с трансплантированным ядром, полученного описанным способом.
[14] Еще одна задача настоящего изобретения заключается в создании способа получения клонированного представителя семейства псовых, включающего в себя стадию переноса указанного эмбриона с трансплантированным ядром в суррогатную мать для того, чтобы обеспечить возможность рождения живого потомка.
[15] Еще одна задача настоящего изобретения заключается в создании клонированного представителя семейства псовых, полученного описанным способом.
Техническое решение.
[16] Для выполнения вышеописанных задач в одном из аспектов настоящего изобретения предложен способ получения эмбриона представителя семейства псовых с трансплантированным ядром с использованием технологии трансплантации ядра соматической клетки.
[17] В другом аспекте настоящего изобретения предложен эмбрион собаки с трансплантированным ядром, полученный с помощью описанного способа.
[18] В соответствии с еще одним аспектом настоящего изобретения предложен способ получения клонированного представителя семейства псовых, включающий стадию переноса указанного эмбриона с трансплантированным ядром суррогатной матери для обеспечения возможности рождения живых потомков.
[19] В соответствии с еще одним аспектом настоящего изобретения предложен клонированный представитель семейства псовых, полученный при помощи описанного способа.
[20] Далее настоящее изобретение будет описано подробно.
[21]
[22] Определение терминов
[23] Используемый здесь термин "трансплантация ядра" относится к способу манипуляции с генами для получения идентичной характерной формы и качества, полученных при искусственной комбинации энуклеированной клетки с ядерной ДНК одной клетки.
[24] Используемый здесь термин "эмбрион с трансплантированным ядром" относится к эмбриону, в который ввели или который слили с клеткой-донором ядра.
[25] Использованный здесь термин "клонированный" относится к методике манипуляции с генами для получения нового индивидуума, имеющего идентичный набор генов с другим индивидуумом. Этот термин, в частности в настоящем изобретении, относится к факту, что клетка, эмбриональная клетка, клетка плода и/или клетка животного обладает последовательностью ядерной ДНК, которая по существу близка или идентична последовательности ядерной ДНК другой клетки, эмбриональной клетки, клетки плода и/или клетки животного
[26] Использованный здесь термин "клетка-донор ядра" относится к клетке или ядру из клетки, которое переносят в ооцит-реципиент, используемый в качестве акцептора ядра.
[27] Использованный здесь термин" ооцит-реципиент" относится к ооциту, в который трансплантировали ядро из клетки-донора ядра после того, как собственное ядро было удалено.
[28] Использованный здесь термин "зрелый ооцит" относится к ооциту в метафазе II мейоза.
[29] Использованный здесь термин "энуклеированный ооцит" относится к ооциту, у которого удалили его собственное ядро.
[30] Использованный здесь термин "слияние" относится к комбинации донора ядра и липидной мембраны ооцита-реципиента. Например, липидная мембрана может представлять собой плазматическую мембрану или ядерную мембрану клеток. Слияние может происходить, будучи дополненным электрической стимуляцией донора ядра и ооцита-реципиента, когда они помещены вплотную друг к другу, или когда донор ядра помещен в перивителлиновое пространство ооцита-реципиента.
[31] Использованный здесь термин "активация" относится к стимуляции клеточного деления до, во время или после стадии трансплантации ядра. Предпочтительно, в настоящем изобретении он обозначает стимуляцию клеточного деления после этапа трансплантации ядра.
[32] Использованный здесь термин "живой потомок" обозначает животное, которое выжило, покинув матку. "Живой потомок" может представлять собой животное, которое жило, по меньшей мере, одну секунду, одну минуту, один день, одну неделю, один месяц, шесть месяцев или более чем один год с момента выхода из утробы матери. "Живой потомок" может не требовать системы кровообращения внутриматочной среды для выживания.
[33] Использованный здесь термин "представители семейства псовых" включает собак, волков, лис, шакалов, койотов, корейских волков и енотовидных собак. Предпочтительно он включает собак или волков. Как известно, собаки произошли при одомашнивании диких волков, и, таким образом, они обладают одинаковым хромосомным набором и проявляют сходство в отношении периода беременности и изменений половых гормонов (Seal US et al., Biology Reproduction 1979, 21: 1057-1066).
[34]
[35] Настоящее изобретение характеризуется тем, что клонирование представителя семейства псовых с помощью технологии трансплантации ядра соматической клетки впервые было успешно осуществлено путем получения эмбриона представителя семейства псовых с трансплантированным ядром в условиях, оптимизированных для электрического слияния, и переноса эмбриона с трансплантированным ядром в яйцевод суррогатной матери для получения живого потомка.
[36]
[37] Способ получения эмбриона представителя семейства псовых с трансплантированным ядром согласно настоящему изобретению может включать стадии (а) энуклеации зрелого ооцита представителя семейства псовых с получением энуклеированного ооцита-реципиента; (б) выделения соматической клетки из ткани собаки-донора с получением клетки-донора ядра; (в) микроинъекции клетки-донора ядра со стадии (б) в энуклеированный ооцит со стадии (а) и электрического слияния клетки-донора с энуклеированным ооцитом при напряжении тока 3,0-3,5 кВ/см; и (г) активации слитого ооцита со стадии (в).
[38] Далее описана каждая стадия способа получения эмбриона представителя семейства псовых с трансплантированным ядром согласно настоящему изобретению.
[39]
[40] Стадия 1: Энуклеация ооцитов-реципиентов
[41] Для использования в качестве ооцитов-реципиентов незрелые ооциты, полученные от собак, могут созревать in vitro, или же могут быть собраны ооциты, созревшие in vivo. Как правило, ооциты млекопитающих (например, крупного рогатого скота, свиней и овец) овулируют с образованием зрелых ооцитов, т.е. на стадии II метафазы мейоза, тогда как ооциты представителей семейства псовых овулируют на стадии профазы I мейоза, в отличие от других животных, и созревают, находясь в яйцеводе в течение 48-72 часов. Из-за того, что скорость созревания ядра ооцита представителей семейства псовых является очень низкой и время овуляции и репродуктивная физиология представителей семейства псовых иные, чем у других животных, предпочтительно для использования в качестве ооцитов-реципиентов отбирают ооциты представителей семейства псовых, созревшие in vivo.
[42] Более конкретно, отбор зрелых ооцитов представителей семейства псовых предпочтительно осуществляют через 48-72 часа и, более предпочтительно, через 72 часа после индукции овуляции у представителей семейства псовых. Для этого день овуляции у представителей семейства псовых могут определять с помощью любого способа, известного в области техники. Примеры таких способов определения дня овуляции могут включать тесты вагинальных мазков, измерение уровня половых гормонов в сыворотке и использование ультразвуковых диагностических систем, но не ограничиваются ими. Начало течки у представителей семейства псовых может быть подтверждено набуханием влагалища и выделениями, состоящими из сыворотки и крови.
[43]
[44] В одном из примеров по настоящему изобретению был проведен тест вагинальных мазков и анализ концентрации сывороточного прогестерона; день, когда количество некератинизированных эпителиальных клеток достигает 80% и концентрация сывороточного прогестерона достигает приблизительно 4,0-7,5 нг/мл, рассматривают как день начала овуляции. На основании этого определения ооциты отбирают через 48-72 часа, а предпочтительно через 72 часа после овуляции. Поскольку известно, что время созревания ооцитов, овулирующих у представителей семейства псовых, составляет 48-72 часа после овуляции, авторы настоящего изобретения анализировали ооциты, собранные через 48 часов, 60 часов и 72 часа после овуляции и, в результате, подтвердили, что ооциты, отобранные приблизительно через 72 часа после овуляции, представляют собой зрелые ооциты, соответствующие метафазе II мейоза. Также ооцит, из которого действительно был успешно получен клонированный представитель семейства псовых по настоящему изобретению, представлял собой ооцит, отобранный через 72 часа после овуляции. Это позволяет предполагать, что наиболее предпочтительно отбирать зрелые ооциты у представителей семейства псовых через 72 часа после овуляции.
[45]
[46] В качестве способа отбора ооцитов, созревших in vivo, может быть использован хирургический способ, включающий анестезию животных с последующим вскрытием брюшной полости. Более конкретно, отбор ооцитов, созревших in vivo, могут осуществлять с использованием сальпингэктомии при помощи любого известного способа. Сальпингэктомия представляет собой способ отбора ооцитов после промывки путем промывки яйцевода средой для сбора ооцитов после хирургического удаления яйцевода.
[47] При другом способе ооциты, созревшие in vivo, могут быть отобраны путем внедрения катетера в бахромчатый конец яйцевода и инъекции промывающей жидкости в маточно-трубный контакт с помощью иглы постоянного катетера. Этот способ обладает преимуществом, заключающимся в том, что он не вызывает повреждение яйцевода и, таким образом, позволяет использовать животное-донор ооцитов при следующей течке.
[48] Соответственно, отбор ооцитов, созревших in vivo, предпочтительно осуществляют с использованием способа, включающего применение катетера, который не вызывает повреждения яйцевода. При этом для увеличения скорости отбора ооцитов при помощи способа отбора ооцитов, включающего использование катетера, авторы настоящего изобретения разработали иглу для извлечения ооцитов, которая имеет закругленный конец и, таким образом, легко вставляется во вход яйцевода (см. Фиг.1). Более конкретно, способ отбора ооцитов с использованием иглы, разработанной авторами настоящего изобретения, включает внедрение и лигирование иглы для извлечения ооцитов, имеющей закругленный конец, в яйцевод, с последующей промывкой средой для сбора ооцитов маточно-трубного контакта таким образом, чтобы позволить промывающей среде затечь внутрь иглы для извлечения ооцитов, и наблюдением промывающей среды под микроскопом для отбора зрелых ооцитов.
[49]
[50] После отбора зрелых ооцитов гаплоидные ядра ооцитов удаляют. Энуклеацию ооцитов могут осуществлять при помощи любого способа, известного в области техники (см. патент США No. 4994384; патент США No. 5057420; патент США No. 5945577; Европейский патент No. 0930009 А1; Корейский патент 342437; Kanda et al, J. Vet. Med. Sci., 57(4): 641-646, 1995; Willadsen, Nature, 320:63-65, 1986, Nagashima et al., Mol. Reprod. Dev. 48: 339-343 1997; Nagashima et al., J. Reprod Dev 38: 37-78, 1992; Prather et al., Biol. Reprod 41: 414-418, 1989, Prather et al., J. Exp. Zool. 255:355-358, 1990; Saito et al., Assis Reprod Tech Andro, 259: 257-266, 1992; Terlouw et al., Theriogenology 37:309, 1992).
[51] Предпочтительно, энуклеацию реципиентных ооцитов могут осуществлять при помощи любого из двух следующих способов. Один способ включает удаление клеток кумулюса зрелых ооцитов-реципиентов, рассечение части zona pellucida ооцитов-реципиентов с использованием микроиглы с получением разреза и удаление первого полярного тельца, ядра и прилегающей цитоплазмы (наименьшего возможного количества) через разрез. Другой способ включает удаление клеток кумулюса ооцитов-реципиентов, окраску ооцитов и удаление первого полярного тельца и ядра ооцитов с использованием отсасывающей пипетки. Более предпочтительно для энуклеации ооцитов используют способ аспирации в случае ооцитов с высоким уровнем выживаемости, а способ с формированием разреза используют в случае ооцитов с низким уровнем выживаемости, причем состояние ооцитов-реципиентов оценивают визуально.
[52]
[53] Стадия 2: Приготовление клеток-доноров ядра
[54] В качестве клеток-доноров ядра могут использовать соматические клетки, происходящие от представителей семейства псовых. В частности, соматические клетки, использованные в настоящем изобретении, могут представлять собой эмбриональные клетки представителей семейства псовых, клетки плода, клетки молодых особей или клетки взрослых особей и, предпочтительно, происходят из тканей, таких как кумулюс, кожа, слизистая оболочка ротовой полости, кровь, спинной мозг, печень, легкие, почки, мышцы и репродуктивный тракт и т.д., которые могут быть получены из клеток взрослых особей. Примеры соматических клеток, которые могут быть использованы в настоящем изобретении, включают клетку кумулюса, эпителиальную клетку, фибробласт, нервную клетку, клетку эпидермиса, кератиноцит, гемопоэтическую клетку, меланоцит, хондроцит, эритроцит, макрофаг, моноцит, мышечную клетку, В-лимфоцит, Т-лимфоцит, эмбриональную стволовую клетку, эмбриональную клетку половой линии, но не ограничиваются ими. Более предпочтительно, соматические клетки, которые могут быть использованы в настоящем изобретении, могут включать фибробласт плода, взрослый фибробласт и клетку кумулюса.
[55] Кроме того, клетки-доноры ядра, использованные в настоящем изобретении, могут представлять собой клетки, полученные путем трансформации соматических клеток дикого типа определенными генами с помощью способа переноса генов или способа направленного воздействия на гены. Способ переноса генов или способ направленного воздействия на гены могут быть с легкостью применены любым специалистом в данной области техники, поскольку они известны в области техники.
[56] Соматические клетки, получаемые в качестве клеток-доноров ядра, могут получать при помощи способа, включающего изготовление хирургических образцов или образцов биопсии, и одиночные клетки из этих образцов могут получать при помощи любого способа, известного в области техники. Например, некоторые ткани, из которых клонировали животных, разрезают в асептических условиях с получением хирургического образца или биоптата, образец измельчают, обрабатывают трипсином, и затем выращивают в тканевой культуральной среде. После выращивания в течение 3-4 дней в клеточной культуральной среде рост клеток на культуральном планшете подтверждается. Когда клетки полностью вырастают, некоторое количество ткани замораживают и хранят в жидком азоте для последующего использования, а оставшееся количество пересевают для использования при трансплантации ядра. Клетки, которые непрерывно выращивают для использования при трансплантации ядра, пересевают до 10 раз для того, чтобы предотвратить избыточный рост клеток.
[57]
[58] Культуральная среда, которую используют как описано выше, может представлять собой одну из сред, которые известны в области техники, и примеры такой среды включают ТСМ-199 и DMEM (среда Игла, модифицированная Дульбекко).
[59]
[60] Стадия 3: Микроинъекция и слияние клеток-доноров ядра
[61] Микроинъекцию клеток-доноров ядра в энуклеированные ооциты проводили путем микроинъекции клеток-доноров ядра в область между цитоплазмой и zona pellucida энуклеированных ооцитов с помощью пипетки для переноса.
[62]
[63] Энуклеированные ооциты, в которые с помощью микроинъекции ввели клетки-доноры ядра, электрически сливали с клетками-донорами ядра с использованием клеточного манипулятора.
[64] Электрическое слияние могут осуществлять с помощью постоянного или переменного тока. Предпочтительно, его могут осуществлять при напряжении тока 3,0-3,5 кВ/см, и более конкретно, его могут осуществлять 1-3 раза при напряжении постоянного тока 3,0-3,5 кВ/см для 10-30 □. Наиболее предпочтительно, его могут осуществлять дважды при постоянном токе с напряжением 3,0-3,5 кВ/см для 20 □. Если напряжение тока при слиянии меньше чем 3,0 кВ/см или больше чем 3,5 кВ/см, степень слияния между ооцитами и клетками-донорами ядра будет очень низкой. Описанный выше уровень напряжения при электрическом слиянии характеризуется тем, что он выше, чем уровень напряжения при электрическом слиянии, в общем известном до настоящего времени (1,7-2,0 кВ/см).
[65] В одном из тестовых примеров по настоящему изобретению для определения оптимального уровня напряжения при электрическом слиянии, эмбрионы с трансплантированными ядрами, в которые проводили микроинъекцию клеток-доноров ядер, электрически сливали в условиях с различным напряжением и проверяли степень слияния с помощью микроскопа (см. тестовый пример 2). В результате можно видеть, что степень ядерного слияния была выше при более высоком напряжении, чем при низком напряжении, и самый высокий уровень слияния 75,2% был показан при уровне напряжения 3,0-3,5 кВ/см (см. таблицу 7).
[66]
[67] Слияние клеток-доноров ядер с ооцитами с использованием электрической стимуляции могут производить в среде для слияния. Среда для слияния, используемая по настоящему изобретению, может представлять собой среду, содержащую маннит, MgSO4, Hepes и бычий сывороточный альбумин (БСА).
[68]
[69] Стадия 4: Активация эмбрионов с трансплантированными ядрами
[70] Активация слитых эмбрионов с трансплантированными ядрами представляет собой стадию реактивации временно приостановленного клеточного цикла. Для реактивации клеточного цикла следует снизить активацию клеточных веществ передачи сигнала, выступающих как элементы, приостанавливающие клеточный цикл, таких как MPF, MAP киназа и т.д.
[71] Как правило, способы активации эмбрионов с трансплантированными ядрами включают электрический способ и химический способ. В настоящем изобретении предпочтительна активация эмбрионов с трансплантированными ядрами химическим способом. Химический способ ускоряет активацию эмбрионов с трансплантированными ядрами сильнее, чем электрический способ. Химический способ представляет собой способ обработки эмбрионов с трансплантированными ядрами такими веществами, как этанол, инозитолтрифосфат (IP3), двухвалентные ионы (например, Ca2+ или Sr2+), ингибиторы микротрубочек (например, цитохалазин В), бивалентный ионный ионофор и ингибиторы протеинкиназ, такие как 6-диметиламинопурин, ингибиторы белкового синтеза (например, циклогексимид), форбол 12-миристат-13-ацетат (РМА). Предпочтительно в настоящем изобретении в качестве химического способа активации эмбрионов с трансплантированным ядром могут использовать способ обработки эмбрионов с трансплантированным ядром одновременно или пошагово с помощью кальциевого ионофора и 6-диметиламинопурина. Более предпочтительно, эмбрионы с трансплантированным ядром обрабатывают 5-10 мкМ кальциевым ионофором при 37-39°С в течение 3-6 минут и, затем, 1,5-2,5 мМ 6-диметиламинопурином при 37-39°С в течение 4-5 часов.
[72]
[73] В одном из тестовых примеров по настоящему изобретению, после того как эмбрионы с трансплантированным ядром были активированы электрическим способом и химическим способом, проводили наблюдения за эмбрионами с трансплантированным ядром с целью определения стадии их развития (см. Тестовый Пример 3). В результате оказалось возможным подтвердить, что химическая активация усиливает потенциал к развитию у эмбрионов с трансплантированным ядром, а химическая активация эмбрионов с трансплантированным ядром обеспечивает возможность развития эмбрионов с трансплантированным ядром до стадии морулы (см. Таблицу 8).
[74]
[75] Таким образом, в настоящем изобретении предложены эмбрионы представителей семейства псовых с трансплантированным ядром, полученные с помощью описанного выше способа. Авторы настоящего изобретения назвали один из эмбрионов представителя семейства псовых с трансплантированным ядром, полученный в одном из примеров, "Snuppy" (клонированный эмбрион представителя семейства псовых). "Snuppy" (клонированный эмбрион представителя семейства псовых) был размещен в международном органе депонирования КСТС (Korean Collection for Type Cultures - Корейская коллекция типовых культур; Korean Research Institute of Bioscience and Biotechnology, 52, Oun-dong, Yusong-gu, Daejeon, Korea) 15 июля 2005, под номером КСТС 10831 ВР.
[76] Эмбрионы с трансплантированным ядром хранят в замороженном состоянии и при необходимости их могут использовать после разморозки.
[77] Кроме того, эмбрионы представителей семейства псовых с трансплантированным ядром по настоящему изобретению могут использовать для получения клонированных представителей семейства псовых, помещая их внутрь суррогатных матерей, что обеспечивает возможность рождения живых потомков. Предпочтительно, перенос эмбрионов с трансплантированным ядром по настоящему изобретению суррогатным матерям осуществляют путем переноса в яйцевод суррогатных матерей. Перенос могут осуществлять любым способом, известным в области техники, и, предпочтительно, для переноса клонированных эмбрионов используют катетер.
[78] В одном из примеров по настоящему изобретению клонированных собак "Snuppy" и "NT-2#" впервые получили путем переноса эмбрионов с трансплантированным ядром по настоящему изобретению внутрь яйцеводов суррогатных матерей (см. Пример 6). При этом в одном из тестовых примеров по настоящему изобретению продемонстрировали, что если эмбрионы с трансплантированным ядром по настоящему изобретению переносили в матку суррогатных матерей, то у суррогатных матерей не наступала беременность (см. Тестовый Пример 4). Это позволяет предполагать, что перенос эмбрионов с трансплантированным ядром при получении клонированных собак предпочтительно производить в яйцевод.
[79] Кроме того, при переносе эмбрионов с трансплантированным ядром суррогатным матерям эмбрионы с трансплантированным ядром могут находиться на стадии 1 клетки, 2 клеток или 4 клеток. Кроме того, эмбрионы с трансплантированным ядром могут культивировать в 25 □ микрокаплях mSOF, покрытых минеральным маслом, пока проводится подготовка суррогатных матерей.
[80]
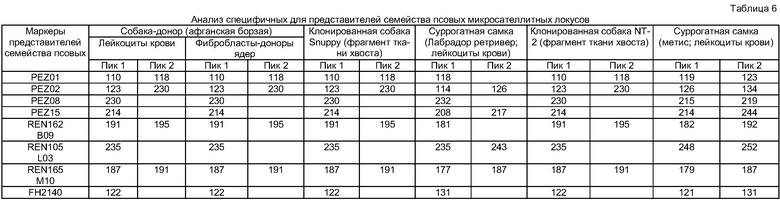
[81] Соответственно, в настоящем изобретении предложены клонированные представители семейства псовых. Клонированные представители семейства псовых обладают точно такими же генетическими характеристиками, что и клетки-доноры ядер или доноры. В одном из примеров по настоящему изобретению клонированных собак получали в соответствии со способом по настоящему изобретению и анализировали их генетические характеристики с использованием микросателлитного анализа (см. Тестовый Пример 1). В результате можно видеть, что клонированные собаки по настоящему изобретению обладают точно такими же генетическими характеристиками, как и клетки-доноры ядер или доноры (см. Таблицу 6).
Благоприятные эффекты
[82] Как было описано выше, в настоящем изобретении предложен способ получения клонированных представителей семейства псовых. Таким образом, настоящее изобретение может внести вклад в развитие исследований в ветеринарной медицине, антропологии и медицинской науке, таких как разведение выдающихся представителей семейства псовых, сохранение редких или почти вымерших представителей семейства псовых, ксенотрансплантацию и модели заболеваний животных.
Краткое описание графических материалов
[83] Фиг.1 представляет собой фотографию, демонстрирующую иглы 15-гейдж и 18-гейдж для извлечения ооцитов, которые использовали для отбора ооцитов у собак в одном из примеров по настоящему изобретению.
[84] Фиг.2 представляет собой фотографию, демонстрирующую клонированную собаку Snuppy, полученную в соответствии со способом по настоящему изобретению, а также собаку-донора (а) и клонированную собаку Snuppy вместе с ее суррогатной матерью (b).
Наилучший способ осуществления изобретения
[85] В дальнейшем, настоящее изобретение описано подробно с помощью примеров. Тем не менее, следует понимать, что эти примеры приведены только в целях иллюстрации, и не подразумевается, что они ограничивают объем настоящего изобретения.
[86]
[87] Пример 1: Отбор реципиентных ооцитов у собак
[88] Собаки, использованные для получения ооцитов-реципиентов, представляли собой 131 самку собак смешанных пород в возрасте 1-3 года, которых содержали в соответствии со стандартами, установленными Национальным Университетом Сеула по содержанию лабораторных животных. Время овуляции определяли с помощью тестов вагинальных мазков и измерения концентрации сывороточного прогестерона у собак в состоянии течки. Зрелые ооциты отбирали через 48-72 часа после овуляции.
[89] Для измерения концентрации сывороточного прогестерона каждый день отбирали 3-5 мл крови и центрифугировали с получением сыворотки, и сыворотку анализировали с использованием DSL-3900 ACTIVE Progesterone Coated-Tube Radioimmunoassay Kit (набор для радиографического иммунного анализа на прогестерон с покрытой трубкой (Diagnostic Systems Laboratories, Inc., TX). День, в который концентрация прогестерона впервые достигала 4,0-7,5 нг/мл, считали днем овуляции (Hase et al., J. Vet. Med. Sci., 62:243-248, 2000).
[90] Для проведения теста вагинального мазка мазки получали каждый день со дня первого проявления проэструса. Мазки получали путем внедрения тампона на стержне в губы влагалища, который затем размазывали по предметному стеклу. После окрашивания Diff-Quik (International chemical Co., Japan) мазки тестировали под микроскопом; время, за которое поверхностные клетки достигали более чем 80% индекса ороговения эпителиальных клеток (Evans J.M. et al., Vet. Rec, 7:598-599, 1970), считали моментом овуляции.
[91] Известно, что время созревания овулировавших ооцитов составляет 48-72 часа после овуляции. Таким образом, авторы настоящего изобретения получали ооциты через 48-72 часа после овуляции следующим образом.
[92] Сначала самкам собак, которые достигли времени получения ооцитов, созревших in vivo, вводили 0,05 мг/кг сульфата атропина и 0,025 мг/кг малеата ацепромазина и подвергали анестезии путем введения 5 мг/кг кетамина. Анестезию поддерживали введением изофлурана.
[93] У подвергнутых анестезии самок собак рассекали брюшную область с помощью 5-10D. Затем в абдоминальную полость яйцевода вводили иглу для извлечения ооцитов с закругленным передним концом (см. Фиг.1) и закрепляли на месте шовной нитью, а затем промывали по направлению вниз средой для сбора ооцитов (см. Таблицу 1), присоединяя катетер 24-гейдж IV в маточно-трубный контакт для того, чтобы промывающая жидкость протекала в иглу 16-гейдж. Промывающую жидкость переносили в стерильную чашку Петри и после этого анализировали под микроскопом для того, чтобы отобрать зрелые ооциты.
[94]
[95]
[96] В результате собрали в среднем по 12 зрелых ооцитов от каждой собаки - в целом 1370 ооцитов.
[97]
[98] Пример 2: энуклеация ооцитов-реципиентов
[99] 0,1% (об./об.) гиалуронидазу (Sigma, USA) добавляли к среде hCR2aa (Таблица 2), приготовленной путем добавления буфера Hepes к среде CR2, не содержащей Са2+ (Charles Rosenkrans 2; Rosenkrans et al., Biol. Reprod. 49, 459-462, 1993). Затем клетки кумулюса ооцитов, полученных в Примере 1, удаляли путем повторного пипетирования описанной выше среды. После этого ооциты окрашивали 5 □/мл бисбензимида (Hoechst 33342) в течение 5 минут и наблюдали под инвертированным микроскопом с эпифлюоресцентным оборудованием при увеличении 200× таким образом, чтобы отобрать только ооциты с первым полярным тельцем. 10% (об./об.) ЭТС и 5 □/мл цитоплазмина В добавляли к среде hCR2aa (Таблица 2), а отобранные ооциты энуклеировали в среде с использованием микроманипулятора (Narishige, Tokyo, Japan). А именно, ооциты удерживали с помощью микропипетки (150 □ внутренний диаметр), и затем удаляли первое полярное тельце, прилегающую цитоплазму (менее чем 5%) и ядра ооцитов с использованием аспирационной пипетки. Энуклеированные ооциты хранили в среде ТСМ-199 (Таблица 3), дополненной 10% (об./об.) ЭТС.
[100]
[101]
[102]
[103]
[104] Пример 3: Приготовление клеток-доноров ядер
[105] В качестве клеток-доноров ядер использовали взрослые фибробласты, отобранные у собак. Для этой цели сначала выделяли биоптат кожи уха у трехлетнего самца афганской борзой. Небольшие кусочки фрагмента ушной ткани промывали 3 раза в DPBS (физиологический раствор Дульбекко, забуференный фосфатом) и измельчали с помощью хирургического лезвия. Измельченную ткань диссоциировали в среде Игла, модифицированной Дульбекко (DMEM; Life Technologies, Rockville, MD), содержащей 0,25% (масс./об.) трипсина и 1 мМ ЭДТА в течение 1 часа при 37°С. Трипсинизированные клетки однократно промывали в DPBS без Са+ и Mg+ путем центрифугирования при 300×g в течение 2 минут и высеивали на 100-мм пластиковые культуральные чашки. Высеянные клетки после этого культивировали в течение 6-8 дней на DMEM, дополненной 10% (об./об.) эмбриональной телячьей сывороткой (ЭТС), 1 мМ глютамина, 25 мМ NaHCO3 и 1% (об./об.) минимальной поддерживающей среды (МПС), вспомогательным аминокислотным раствором (Life Technologies) при 39°С в увлажненной атмосфере с 5% СО и 95% воздуха. После удаления неприкрепившихся кластеров клеток или эксплантатов прикрепившиеся клетки выращивали далее в течение 4-6 дней с трипсинизацией в течение 1 мин с использованием 0,1% трипсина и 0,02% ЭДТК (этилендиамин тетрауксусной кислоты). Затем пересеянные клетки помещали в замораживающую среду и хранили в жидком азоте при -196°С. Замораживающая среда состояла из 80% (об./об.) среды Игла, модифицированной Дульбекко (DMEM), 10% (об./об.) диметилсульфоксида (ДМСО) и 10% (об./об.) эмбриональной телячьей сыворотки (ЭТС).
[106]
[107] Пример 4: Микроинъекция и слияние клеток-доноров ядер с энуклеированными ооцитами
[108] Клетки-доноры ядер, полученные в примере 3, вводили путем микроинъекции в энуклеированные ооциты, полученные в Примере 2. После того, как аспирационную пипетку микроманипулятора в Примере 2 заменяли на пипетку для переноса, фиксированные ооциты обрабатывали 100 мг/мл фитогемагглютинина в среде hCR2aa. Разрез на энуклеированном ооците поддерживали удерживающей пипеткой, и затем туда вводили пипетку для переноса. После этого одиночные клетки, выделенные из фибробластов в Примере 3, инъецировали в пространство между цитоплазмой и zona pellucida энуклеированного ооцита с помощью пипетки для переноса.
[109] Ооциты, в которые инъецировали клетки-доноры ядер, как описано выше, помещали в среду для слияния (содержащую 0,26 М маннит, 0,1 мМ MgSO4, 0,5 мМ Hepes и 0,05% БСА) и переносили в камеру для слияния клеток, оборудованную проволочным электродом из нержавеющей стали (ВТХ 453, 3,2 мм gap; ВТХ, San Diego, CA). После установления равновесия в течение 3 минут, к слившимся парам применяли постоянный электрический ток с напряжением 3,0-3,5 кВ/см в течение 20 секунд с использованием манипулятора ВТХ Electro-cell, таким образом, сливая клетки-доноры с ооцитами. Слияние проводили при низком напряжении (близком к 3,0 кВ/см) если полученные ооциты были слабыми. В случае если ооциты были здоровыми, слияние проводили при высоком напряжении (близком к 3,5 кВ/см). Среднее напряжение, при котором проводили слияние, составляло 3,3 кВ/см.
[110] 1095 слившихся эмбрионов с трансплантированным ядром отбирали с помощью стереомикроскопического тестирования и культивировали в течение 3 часов в модифицированной синтетической жидкости яйцеводов (mSOF), как показано в Таблице 4 (Jang et al., Reprod Fertil Dev, 15, 179-185, 2003).
[111]
[112]
[113] Пример 5: Активация эмбрионов с трансплантированным ядром
[114] Эмрбионы с трансплантированным ядром, полученные в Примере 4, культивировали в mSOF (Таблица 4), содержащей 10 мкМ ионофора, в течение 4 минут при 39°С. Затем эмбрионы промывали и дальше инкубировали в течение 4 часов в mSOF, дополненной 1,9 мМ 6-диметиламинопурином.
[115] Один из эмбрионов представителей семейства псовых с трансплантированным ядром, описанных выше, авторы настоящего изобретения назвали "Snuppy" (клонированный эмбрион представителя семейства псовых), и его разместили в международном органе депонирования КСТС (Корейская коллекция типовых культур; Korean Research Institute of Bioscience and Biotechnology, 52, Oun-dong, Yusong-gu, Daejeon, Korea) 15 июля 2005, под номером КСТС 10831 ВР. Эмбрионы с трансплантированным ядром перед переносом их суррогатным матерям выращивали в 25 □ каплях mSOF, покрытых минеральным маслом.
[116]
[117] Пример 6: Перенос эмбрионов суррогатным матерям и получение клонированных собак
[118] Эмбрионы с трансплантированным ядром из Примера 5 хирургически переносили в яйцевод суррогатных матерей. Перенос производили в зависимости от уровня готовности суррогатной матери после активации эмбрионов с трансплантированным ядром из Примера 5. А именно, когда суррогатные матери были сразу же подготовлены, перенос эмбрионов с трансплантированным ядром проводили немедленно, в противном случае перенос производили на следующий день после активации эмбрионов с трансплантированным ядром (стадия развития эмбриона: 2-клеточная стадия или 4-клеточная стадия). В качестве суррогатных матерей использовали 123 собаки, включающие собак смешанной породы и лабрадоров ретриверов. Выбранные собаки были здоровы, демонстрировали нормально повторяющийся цикл эструса и нормальное состояние матки. 1095 реконструированных эмбрионов из Примера 5 хирургически перенесли суррогатным матерям. Для этой цели суррогатных матерей подвергали анестезии с помощью внутривенной инъекции 0,1 мг/кг ацепромазина и 6 мг/кг пропофола, и состояние анестезии поддерживали с помощью 2% изофлурана. Операционную область анестезированной самки собаки оперировали в стерильных условиях и разрезали по центру брюшины с помощью 5-10D при общем вскрытии брюшной полости таким образом, чтобы обнажить яйцевод. Брюшную полость стимулировали руками, чтобы подтянуть яичник, яйцевод и матку к разрезу. Брыжейку подтянутого яичника осторожно перебирали, чтобы найти открытие яйцевода, и вставляли в яйцевод 3.5F катетер Tom cat (Sherwood, St. Louis, МО), оборудованный 1,0 мл туберкулиновым шприцом (без латекса, Becton Dickinson & СО. Franklin lakes, NJ 07417), чтобы обеспечить достаточное пространство спереди от катетера. Затем эмбрионы с трансплантированным ядром вводили в яйцевод через катетер. В случае если в микроскоп можно было наблюдать успешно инъецированные эмбрионы с трансплантированным ядром, то в брюшную полость инъецировали 500 мл физиологического раствора, содержащего антибиотик. Брюшинный шов накладывали с помощью рассасывающейся нити, а затем накладывали шов на кожу. Для предотвращения постхирургической инфекции в течение трех дней делали инъекции широкого спектра антибиотиков.
[119] Через 22 дня после переноса эмбрионов с трансплантированным ядром суррогатным матерям беременность определяли с использованием ультразвукового сканера с присоединенным линейным зондом 7,0 МГц SONOACE 9900 (Medison Co. LTD, Seoul, Korea). Мониторинг беременности выполняли с помощью ультразвука каждые 2 недели после первого подтверждения. В результате подтвердили, что 3 собаки оказались беременными. Из них одна в последующем была утрачена, а от одного из двух оставшихся животных с помощью кесарева сечения 24 апреля 2005 родилась первая клонированная собака, через 60 дней после переноса эмбрионов с трансплантированным ядром. Вес при рождении составлял 530 г, и клонированный детеныш оказался здоровым. Клонированного щенка назвали "Snuppy" (щенок Национального Университета Сеула - Seoul National University PuDDy). От оставшегося одного животного с помощью кесарева сечения 29 мая 2005 родилась вторая клонированная собака, через 60 дней после переноса эмбрионов с трансплантированным ядром. Вес при рождении составил 550 г, и клонированный щенок оказался здоровым. Второй клонированный щенок был назван "NT-2#".
Способ выполнения изобретения
[120] Тестовый пример 1: Проверка генетической идентичности клонированных собак, полученных по настоящему изобретению
[121]
[122] В соответствии с настоящим изобретением клонированных щенков Snuppy и NT-2#, полученных в Примере 6, тестировали с целью проверки, являются ли клонированные щенки Snuppy и NT-2# генетически идентичными афганской борзой собаке-донору клеток-доноров ядер из Примера 3.
[123] Выделяли геномную ДНК клонированных щенков, собаки-донора, суррогатных реципиентов и фибробластов-доноров ядер. Для этой цели из хвостов щенков получили фрагменты ткани, и собрали образцы крови у собаки-донора и суррогатной матери. Как образцы ткани, так и образцы крови и фибробласты инкубировали в литическом буфере [0,05 М Tris (рН 8.0), 0,05 М ЭДТА (рН 8.0), 0,5% додецилсульфат натрия (SDS)], дополненным 400 □ протеиназы К в течение одной ночи. Затем геномную ДНК из каждого образца экстрагировали фенолом и осаждали этанолом.
[124] Образцы изолированной геномной ДНК растворяли в 50 D ТЕ и использовали для проведения микросателлитного анализа с восемью собачьими маркерами [PEZ01, PEZ02, PEZ08, PEZ15 (см. патент США No. 5874217), REN162B09, REN105L03, REN165M10, FH2140 (см. http://www.fhcre.org/science/dog_ genome/dog.html)] (Francisco, L.V. et al. Mamm. Genome 7, 359-362 1996; Neff, M.W. et al. Genetics. 151, 803-820, 1999; Richman, M. et al. J. Biochem. Biophys. Methods 47, 137-149, 2001; Denise, S. et al. Animal Genetics. 35, 14-17, 2004). Выделенную геномную ДНК в качестве матрицы амплифицировали с помощью ПЦР с использованием флуоресцентно помеченных локус-специфических праймеров (Таблица 5), изготовленных на основании последовательностей известных маркеров. Продукты амплификации анализировали с помощью автоматического анализатора ДНК-последовательностей (ABI 373: Applied Biosystems, Foster City, CA). Реакция ПЦР включала преденатурацию при 94°С в течение 1 мин, последующую денатурацию при 94°С в течение 20 с, отжиг при 58°С в течение 20 с и элонгацию при 74°С в течение 20, длительностью 30 циклов, и затем пост-элонгацию при 74°С в течение 5 мин. Также использовали собственное программное обеспечение (GeneScan и Genotyper; Applied Biosystems) для оценки размера продуктов ПЦР.
[125]
[126]
[127]
[128] В результате обнаружили, что клонированные собаки Snuppy и NT-2#, полученные в соответствии с настоящим изобретением, были полностью генетически идентичными собаке-донору афганской борзой и фибробластам, выделенным из собаки-донора. С другой стороны, клонированные собаки по настоящему изобретению и суррогатные матери (лабрадоры ретриверы и собаки смешанной породы) генетически отличались друг от друга (Таблица 6).
[129]
[130]
[131]
[132] Тестовый пример 2: Оптимизация условий для электрического слияния клеток-доноров ядер с энуклеированными ооцитами
[133] Для оптимизации условия для электрического слияния клеток-доноров ядер с энуклеированными ооцитами клетки-доноры ядер микроинъецировали внутрь энуклеированных ооцитов способом, сходным с примером 4, и клетки-доноры и ооциты сливали друг с другом в условиях варьирующего напряжения 1,7-1,9 кВ/см, 2,1-2,5 кВ/см и 3,0-3,5 кВ/см. Затем реконструированные эмбрионы проверяли на слияние с помощью стереомикроскопа.
[134] В результате можно было видеть, что в условиях напряжения тока 3,0-3,5 кВ/см продемонстрировано, что 203 ооцита с трансплантированным ядром слились (процент слияния 75,2%). Это говорит о том, что эти условия приводят к значительно более высокой эффективности слияния по сравнению с другими условиями (Таблица 7).
[135]
[136]
[137]
[138] Тестовый пример 3: Оптимизация условий для активации эмбрионов с трансплантированным ядром
[139] Эмбрионы с трансплантированным ядром, полученные в Примере 4, активировали с помощью электрического способа и химического способа. Затем над эмбрионами с трансплантированным ядром проводили наблюдения с целью определить стадии их развития. При электрическом способе эмбрионы с трансплантированным ядром из Примера 4 помещали в среду с маннитом (содержащую 0,26 М маннит, 0,1 мМ MgSO4, 0,5 мМ Hepes и 0,05% БСА) и 100 нМ CaCl2 и переносили в камеру для слияния клеток, оборудованную проволочным электродом из нержавеющей стали (ВТХ453, 3.2 мм gap; BTX, San Diego, СА). После достижения равновесия в течение 3 минут к сливающимся парам применяли постоянный ток с напряжением 3,0-3,5 кВ/см в течение 20 секунд с использованием BTX Electro-cell манипулятора, сливая таким образом клетки-доноры с ооцитами.
[140]
[141] При способе химической активации эмбрионы с трансплантированным ядром из Примера 4 помещали в mSOF, содержащую 10 мМ ионофор (Sigma), и выращивали в среде при 39°С в течение 4 минут. Затем культуру промывали и дальше культивировали в mSOF (Таблица 4), дополненной 1,9 мМ 6-диметиламинопурином в течение 4 часов. После завершения выращивания эмбрионы переносили в среду ТСМ199 (Таблица 3).
[142] Каждую стадию развития эмбрионов с трансплантированным ядром, активированных с помощью электрического способа и химического способа, исследовали с использованием стереомикроскопа при увеличении 100х.
[143] В результате подтвердили, что усиление химической активации усиливает потенциал к развитию у эмбрионов с трансплантированным ядром. А именно, было показано, что в случае эмбрионов с трансплантированным ядром, активированных с помощью химического способа, 80% ооцитов достигали 2-клеточной стадии, а в случае эмбрионов с трансплантированным ядром, активированных электрическим способом, только около 53% ооцитов достигали 2-клеточной стадии. Также было обнаружено, что химически активированные эмбрионы продемонстрировали развитие эмбрионов с трансплантированным ядром до стадии морулы, а электрически активированные эмбрионы продемонстрировали развитие только до 16-клеточной стадии (Таблица 8).
[144]
[145]

[146]
[147] Тестовый пример 4: Оптимизация условий для переноса эмбрионов с трансплантированным ядром по настоящему изобретению суррогатным матерям
[148] Эмбрионы с трансплантированным ядром, активированные в Примере 5, культивировали в mSOF (Таблица 4) в инкубаторе при 38-39°С и атмосфере с 5% СО и 5% кислорода. Затем эмбрионы, выращенные до 8-клеточной стадии, погружали в забуференный фосфатом физиологический раствор, содержащий 0,1% эмбриональной телячьей сыворотки, и переносили в рог матки 20 суррогатных матерей (собаки смешанной породы) с помощью соломинки.
[149] Через 22 дня после переноса эмбрионов с трансплантированным ядром беременность детектировали с использованием ультразвукового сканера (Medison Co. LTD, Seoul, Korea) в соответствии со способом, описанным в Примере 6.
[150] В результате было обнаружено, что ни один эмбрион с трансплантированным ядром, перенесенный в матку, не привел к наступлению беременности. Это позволяет предполагать, что предпочтительным является перенос эмбрионов с трансплантированным ядром в яйцевод, как описано в Примере 6.
Промышленная применимость
[151] Как описано выше, в настоящем изобретении предложен способ получения клонированных представителей семейства псовых. Таким образом, настоящее изобретение может внести вклад в развитие исследований в ветеринарной медицине, антропологии и медицинской науке, таких как разведение выдающихся представителей семейства псовых, сохранение редких или почти вымерших видов представителей семейства псовых, ксенотрансплантацию и модели заболеваний на животных.



| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОГО ТИГРА ПУТЕМ ПРИМЕНЕНИЯ МЕТОДА МЕЖВИДОВОЙ ТРАНСПЛАНТАЦИИ ЯДЕР | 2000 |
|
RU2205537C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОЙ КОРОВЫ | 2000 |
|
RU2205536C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННЫХ ЭМБРИОНОВ ЧЕЛОВЕКА ПУТЕМ ПРИМЕНЕНИЯ СПОСОБА МЕЖВИДОВОЙ ТРАНСПЛАНТАЦИИ ЯДЕР | 2000 |
|
RU2216591C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ЭКЗОГЕННОГО БЕЛКА В МОЛОКЕ ТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ И СПОСОБ ОЧИСТКИ БЕЛКОВ ИЗ МОЛОКА | 2004 |
|
RU2360002C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНОВ ЖИВОТНЫХ И СПОСОБ ВЫРАЩИВАНИЯ ЖИВОТНОГО ИЗ ЭМБРИОНОВ | 1998 |
|
RU2216592C2 |
| ЖИВОТНЫЕ, УСТОЙЧИВЫЕ К ВИРУСУ РЕПРОДУКТИВНО-РЕСПИРАТОРНОГО СИНДРОМА СВИНЕЙ | 2012 |
|
RU2644673C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНОЙ СТВОЛОВОЙ КЛЕТКИ ЧЕЛОВЕКА | 2007 |
|
RU2352637C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ, КОТОРЫЕ ПРОДУЦИРУЮТ ЭКЗОГЕННЫЕ БЕЛКИ В МОЛОКЕ, И ТРАНСГЕННЫЕ МЛЕКОПИТАЮЩИЕ, ПОЛУЧЕННЫЕ ТАКИМ СПОСОБОМ | 2004 |
|
RU2390562C2 |
| ПАРТЕНОГЕНЕТИЧЕСКАЯ АКТИВАЦИЯ ЧЕЛОВЕЧЕСКИХ ООЦИТОВ ДЛЯ ПОЛУЧЕНИЯ ЧЕЛОВЕЧЕСКИХ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК | 2006 |
|
RU2469085C2 |
| ТРАНСГЕНЕЗ У МЛЕКОПИТАЮЩИХ ПУТЕМ ИНТРАЦИТОПЛАЗМАТИЧЕСКОЙ ИНЪЕКЦИИ СПЕРМЫ | 1999 |
|
RU2267270C2 |

Изобретение относится к области биотехнологии и генной инженерии. Способ включает стадии энуклеации ооцита представителя семейства псовых с получением энуклеированного ооцита-реципиента, проведение переноса ядра в энуклеированный ооцит с использованием соматической клетки собаки в качестве клетки-донора ядра в оптимизированных условиях таким образом, чтобы получить эмбрион с трансплантированным ядром, и перенос эмбриона с трансплантированным ядром в яйцевод суррогатной матери. Способ может быть использован для разведения выдающихся представителей семейства псовых, сохранения редких или почти вымерших видов представителей семейства псовых, ксенотрансплантации и моделирования заболеваний на животных. 2 н. и 11 з.п. ф-лы, 2 ил., 8 табл.
1. Способ получения эмбриона представителя семейства псовых с трансплантированным ядром, включающий стадии:
(а) энуклеации зрелого ооцита представителя семейства псовых с получением энуклеированного ооцита-реципиента;
(б) выделения соматической клетки из ткани донора - представителя семейства псовых - с получением клетки-донора ядра;
(в) микроинъекции клетки-донора ядра со стадии (б) в энуклеированный ооцит со стадии (а) и электрического слияния клетки-донора с энуклеированным ооцитом при напряжении тока 3,0-3,5 кВ/см и
(г) активации слитого ооцита со стадии (в).
2. Способ по п.1, где зрелый ооцит представляет собой ооцит, созревший in vivo.
3. Способ по п.2, где ооцит, созревший in vivo, получен от представителя семейства псовых через 48-72 ч после овуляции.
4. Способ по п.1, где соматическая клетка на стадии (а) представляет собой клетку, выбранную из группы, состоящей из клетки кумулюса, эпителиальной клетки, фибробласта, нервной клетки, эпидермальной клетки, кератиноцита, гемопоэтической клетки, меланоцита, хондроцита, эритроцита, макрофага, моноцита, мышечной клетки, В-лимфоцита, Т-лимфоцита, эмбриональной стволовой клетки и эмбриональной клетки половой линии, клетки плода, клетки плаценты и клетки эмбриона.
5. Способ по п.1, где соматическая клетка представляет собой фибробласт или клетку кумулюса.
6. Способ по п.1, где электрическое слияние на стадии (в) проводят 1-3 раза при постоянном напряжении тока 3,0-3,5 кВ/см в течение 10-30 с.
7. Способ по п.1, где стадию активации на стадии (г) осуществляют путем обработки слитого ооцита одновременно или последовательно кальциевым ионофором и DMAP (6-диметиламинопурином).
8. Способ по п.7, где способ активации осуществляют путем обработки слитого ооцита 5-10 мкМ кальциевого ионофора при 37-39°С в течение 3-5 мин и затем 1,5 мМ-2,5 мМ DMAP (6-диметиламинопурином) при 37-39°С в течение 4-5 ч.
9. Способ по п.1, где представитель семейства псовых выбран из группы, состоящей из собаки, волка, лисицы, шакала, койота, Корейского волка и енотовидной собаки.
10. Способ по п.1, где представитель семейства псовых выбран из группы, состоящей из собаки, волка и лисицы.
11. Способ получения представителя семейства псовых, включающий стадии:
(а) энуклеации зрелого ооцита представителя семейства псовых с получением энуклеированного ооцита-реципиента;
(б) выделения соматической клетки из ткани донора - представителя семейства псовых - с получением клетки-донора ядра;
(в) микроинъекции клетки-донора ядра со стадии (б) в энуклеированный ооцит со стадии (а) и электрического слияния клетки-донора с энуклеированным ооцитом при напряжении тока 3,0-3,5 кВ/см;
(г) активации слитого ооцита со стадии (в) с получением эмбриона с трансплантированным ядром; и
(д) переноса эмбриона с трансплантированным ядром в яйцевод суррогатной матери для того, чтобы обеспечить возможность рождения живого потомка.
12. Способ по п.11, где представитель семейства псовых выбран из группы, состоящей из собаки, волка, лисицы, шакала, койота, Корейского волка и енотовидной собаки.
13. Способ по п.11, где представитель семейства псовых выбран из группы, состоящей из собаки, лисицы и волка.
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОЙ КОРОВЫ | 2000 |
|
RU2205536C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОГО ТИГРА ПУТЕМ ПРИМЕНЕНИЯ МЕТОДА МЕЖВИДОВОЙ ТРАНСПЛАНТАЦИИ ЯДЕР | 2000 |
|
RU2205537C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННЫХ ЭМБРИОНОВ ЧЕЛОВЕКА ПУТЕМ ПРИМЕНЕНИЯ СПОСОБА МЕЖВИДОВОЙ ТРАНСПЛАНТАЦИИ ЯДЕР | 2000 |
|
RU2216591C2 |

