Область изобретения
Данное изобретение относится к способу получения клонированных эмбрионов человека путем применения способа межвидовой трансплантации клеточных ядер, более конкретно к способу получения клонированных эмбрионов человека путем применения способа межвидовой трансплантации клеточных ядер, посредством которого ядра соматических клеток, полученных из ткани человека, переносят в зрелые ооциты, происходящие из коровы. Изобретение относится также к клонированным эмбрионам человека, полученным описанным выше способом.
Предпосылки изобретения
В течение продолжительного времени считалось, что производство животных должно проводиться посредством оплодотворения, включающего в себя мужские и женские гаметы. Однако были приложены огромные усилия по получению клонированных животных с идентичным видом и идентичными генетическими характеристиками.
Недавно, различные рекомбинантные растения с желательными признаками ценных культурных растений были успешно получены благодаря разработкам в биотехнологии и генной инженерии (см.: Schweizer et al., Plant Journal, 20: 541-552, 1999). Что касается животных, было много успешных примеров получения клонированных животных, которые включают в себя, например, клонированных ягнят (см. : Wilmut et al., Nature, 385:810-813, 1997), клонированных коров (см.: Wells et al., Reprod. Fertil. and Develop., 10:369-378, 1998) и клонированных мышей (см.: Wakayama et al., Nature, 394:369-374, 1998). Поскольку получение клонированных животных не может быть реализовано без высокой технологии, созданной на основе биотехнологии, считалась нормой оценка технологического развития в соответствующих областях.
Между тем, было известно, что стволовые клетки животных обладают способностью развиваться в любой орган, что побудило к исследованию механизма их дифференцировки в каждый орган путем их получения и культивирования. При проведении этого исследования важно использовать материалы с идентичной тканеспецифичностью для уменьшения вариаций среди многих различных исследований. Однако, очевидно, что подходящие материалы с идентичной тканеспецифичностью не являются все время доступными. Хотя недавно способ клонирования с использованием соматических клеток облегчил получение материалов с идентичной тканеспецифичностью, он был менее удовлетворительным в случае ткани человека.
В этих обстоятельствах существовали убедительные причины для разработки способа получения эмбриональных стволовых клеток человека с идентичной тканеспецифичностью.
Сущность изобретения
Согласно данному изобретению, было обнаружено, что эмбриональные стволовые клетки человека могут быть успешно получены при помощи способа межвидовой трансплантации клеточных ядер, включающего в себя слияние ооцитов коровы и клеток кожи человека и культивирование клонированных эмбрионов человека in vitro до стадии морул/бластоцист.
Первой целью данного изобретения является, следовательно, обеспечение способа получения клонированных эмбрионов человека при помощи способа межвидовой трансплантации ядер.
Другой целью данного изобретения является обеспечение клонированных эмбрионов человека, полученных указанным способом.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Указанные выше и другие цели и признаки данного изобретения станут очевидными из нижеследующего описания, даваемого вместе с сопутствующими чертежами, в которых:
Фигура 1 является фотографией донорных соматических клеток.
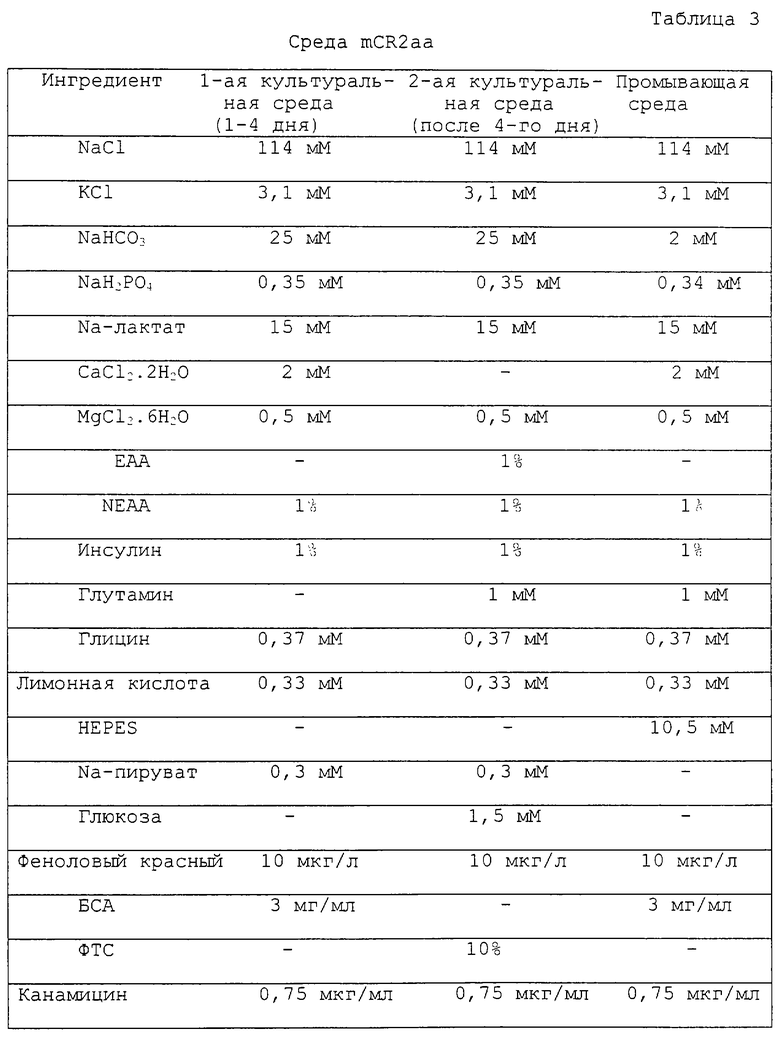
Фигура 2 является фотографией, показывающей процесс разрезания zona pellucida (прозрачной зоны, т.е. блестящей оболочки яйцеклетки) реципиентного ооцита удерживающей пипеткой и разрезающей пипеткой.
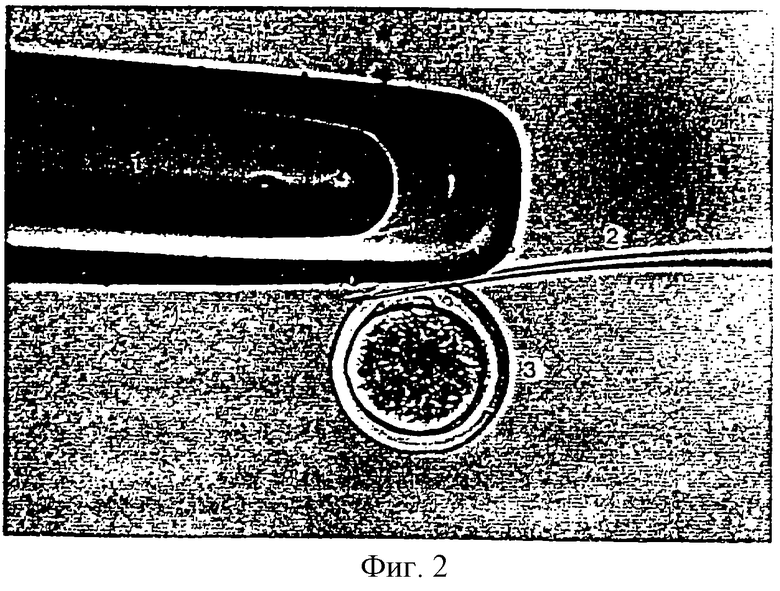
Фигура 3 является фотографией, показывающей процесс энуклеации путем удаления первого полярного тельца и ядра из реципиентного ооцита.
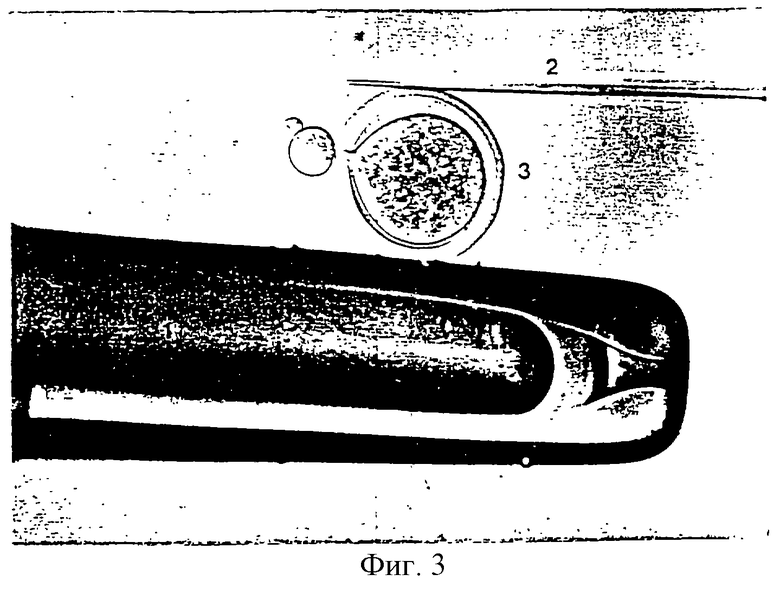
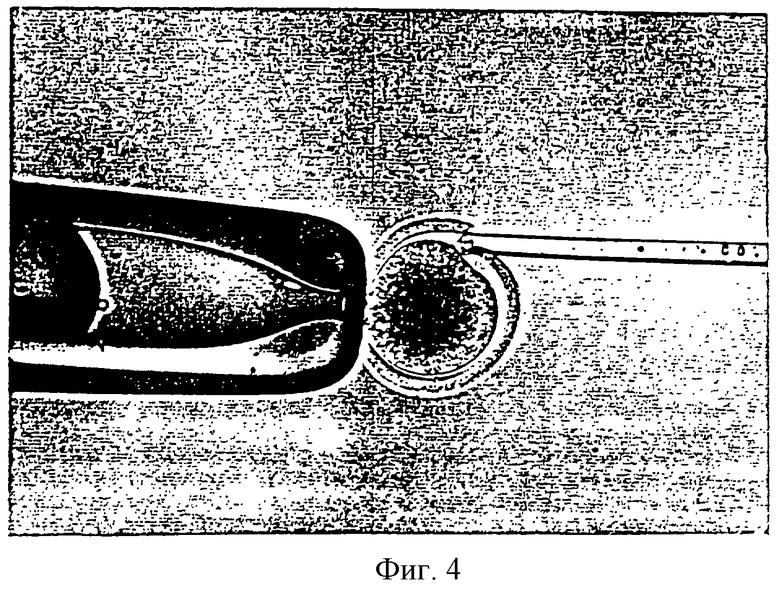
Фигура 4 является фотографией, показывающей процесс переноса соматической клетки в энуклеированный (безъядерный) ооцит удерживающей пипеткой и инъекционной пипеткой.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Способ получения клонированных эмбрионов человека данного изобретения предусматривает стадии получения линий донорных соматических клеток, собранных из человека; созревания ооцитов, собранных из яичника коровы, in vitro; удаления клеток кумулуса (cumulus) (яйцевого холмика на стенке Граафова пузырька, в котором помещается яйцо), окружающих ооциты, разрезания части блестящей зоны созревших ооцитов и выжимания части цитоплазмы, включающей в себя первое полярное тельце, с получением энуклеированных реципиентных ооцитов; перенесения ядра в реципиентный ооцит инъекцией донорных клеток в энуклеированные ооциты с последующими последовательными электрослиянием и активацией электрослитых клеток с получением эмбрионов; постактивации и культивирования эмбрионов in vitro.
Способ получения клонированных эмбрионов данного изобретения дополнительно иллюстрируется следующим образом.
Стадия 1: Получение донорных клеток
Линии соматических клеток, собранных из человека, готовят в качестве донорных клеток: хотя клетки, собранные из человека, не являются ограничением для донорных клеток, предпочтительные клеточные линии включают в себя клетки или фибробласты кожи, собранные из пупочного канатика новорожденных. Более предпочтительной клеточной линией для донорных клеток являются клетки кожи, выделенные из кожной ткани. Указанные клеточные линии получают с использованием общепринятого известного способа (см.: Mather & Barnes, Methods in Cell Biology, vol. 57, Animal Cell Culture Methods, Academic Press, 1998) с некоторыми модификациями.
Например, клетки или фибробласты кожи пупочного канатика новорожденных промывают и измельчают. Затем, эти клетки подвергают обработке трипсином и коллагеназой типа II в условиях 39oС, 5% СО2, с последующим культивированием в DMEM (модифицированной по способу Дульбекко среде Игла), дополненной не являющимися незаменимыми аминокислотами, 10% ФТС (фетальной телячьей сывороткой) и 1% смесью пенициллин-стрептомицин (10000 Е/мл пенициллина, 10 мг/мл стрептомицина) при описанных выше условиях.
Эти соматические клеточные линии хранят с использованием субкультуры, культуры сывороточного голодания или замораживания. Субкультивирование донорных клеточных линий проводят при регулярных интервалах путем замены старой среды на новую среду после трипсинизации. Культивирование с сывороточным голоданием проводят с использованием DMEM, дополненной 0,5% ФТС, и способа Wilmut et al. (см.: Wilmut et al., Nature, 385:810-813, 1997). Хранящиеся таким образом клеточные линии используют для более поздней стадии в качестве донорных клеток.
Стадия 2: Получение реципиентных ооцитов
Незрелые ооциты, собранные из яичника коровы, доводят до созревания in vitro: незрелые ооциты отбирают из яичника в промывающей среде ТСМ199, содержащей 10 мМ HEPES (N-[гидроксиэтил]пиперазин-N'-[2-этансульфоновая кислоту] ) и доводят до созревания культивированием этих клеток в культуральной среде ТСМ199 (содержащей Na-пируват, пенициллин-стрептомицин), дополненной эстрадиолом, FSH (фолликулостимулирующим гормоном) и ФТС, в течение 16-22 часов в условиях 39oС, 5% СО2.
Стадия 3: Энуклеация реципиентных ооцитов
После удаления клеток кумулуса (яйцевого холмика на стенке Граафова пузырька), окружающих зрелые реципиентные ооциты, и разрезания части блестящей зоны ооцитов, часть цитоплазмы, включающую первое полярное тельце, удаляют из ооцитов с получением энуклеированных ооцитов: сначала клетки кумулуса, окружающие зрелые ооциты, удаляют физически денудирующей пипеткой в промывающей среде ТСМ199, содержащей гиалуронидазу. Затем денудированные ооциты промывают промывающей средой ТСМ199 и переносят в раствор цитохалазина В. Для энуклеации денудированных ооцитов часть блестящей зоны денудированных ооцитов пронизывается режущей пипеткой с образованием щели, через которую 10-15% цитоплазмы, включающей в себя первое полярное тельце, может выжиматься из ооцитов. Энуклеированные ооциты промывают и инкубируют в культуральной среде ТСМ199. Указанный раствор цитохалазина В получают разбавлением цитохалазина В, растворенного в ДМСО (диметилсульфоксиде), культуральной средой ТСМ199.
Стадия 4: Электрослияние донорных клеток с реципиентными ооцитами и активация электрослитых клеток
Донорные клетки переносят в реципиентные ооциты с последующим электрослиянием и активацией электрослитых клеток: перед инъекцией донорных клеток в реципиентные ооциты энуклеированные ооциты промывают культуральной средой ТСМ199 и переносят в раствор РНА-Р (фитогемагглютинина). Затем донорные клетки переносят в энуклеированные ооциты инъекцией донорных клеток в щель, сделанную на блестящей зоне ооцитов, в растворе РНА-Р.
Электрослияние проводят с использованием электроманипулятора клеток Electro Cell Manipulator (BTX ECM2001). Реконструированные эмбрионы в растворе маннита, дополненном промывающим раствором ТСМ199, помещают в камеру с двумя электродами, по одному на каждой стороне. Перед помещением эмбрионов с их донорными клетками, обращенными к катоду в камере, эту камеру заполняют раствором маннита. После электрослияния эмбрионов посредством приложения импульса переменного тока 0,75-2,00 кВ/см дважды с интервалом в одну секунду в течение 15 мкс каждый раз, электрослитые эмбрионы промывают раствором маннита и промывающей средой ТСМ199, инкубируют в растворе цитохалазина В и активируют. Электрослияние и активацию проводят одновременно, при условии, что электрослияние проводят в среде с маннитом, содержащей Са2+. В противном случае активацию проводят после электрослияния. При проведении электрослияния в не содержащей Са2+ среде с маннитом стадию активации проводят инкубированием эмбрионов в растворе иономицина в темноте. Затем иономицин удаляют из эмбрионов промыванием их промывающей средой ТСМ199, содержащей ФТС или БСА. Указанный раствор иономицина получают разбавлением иономицина, растворенного в ДМСО, промывающей средой ТСМ199, содержащей БСА.
Стадия 5: Постактивация и культивирование эмбрионов in vitro
Эмбрионы постактивируют и культивируют in vitro: активированные эмбрионы, инкубированные в промывающей среде ТМС199, содержащей ФТС или БСА, постактивируют инкубированием в растворе циклогексимида или DAMP (4-диметиламинопурина) и культивируют in vitro в условиях 5% CO2 или смеси 5% СО2, 7% О2 и 88% N2. Указанный раствор циклогексимида или раствор DAMP получают добавлением циклогексимида, растворенного в этаноле, или DAMP к средам для культивирования in vitro, соответственно. Среды для культивирования in vitro включают в себя среды mTALP (см. таблицу 1) (таблицы 1-11 см. в конце описания), mSOF (см. таблицу 2) и mCR2aa (см. таблицу 3), все из которых содержат NaCl, KCl, NаНСО3, NaH2PO4, CaCl2, Na-лактат, глюкозу, феноловый красный, БСА, канамицин, незаменимые аминокислоты, не являющиеся незаменимыми аминокислоты и L-глутамин.
Необязательно, эмбрионы, культивируемые in vitro, хранят посредством замораживания для более позднего использования и подвергают оттаиванию, когда их предполагают использовать. Для замораживания эмбрионов их промывают ЗФР, содержащим ФТС, помещают в среду для замораживания, содержащую пенициллин-стрептомицин, СаСl2, глюкозу, MgCl2, Na-пируват и ЗФР. Затем эмбрионы в среде для замораживания подвергают медленному замораживанию с последующим быстрым замораживанием в жидком N2. При вынимании замороженных эмбрионов из жидкого N2 и оттаивании их помещают на воздух на приблизительно 5 секунд и затем оттаивают в теплой воде. Для удаления среды для замораживания из оттаявших эмбрионов их помещают последовательно в среды, содержащие глицерин от его высокой концентрации до низкой концентрации.
На основе вышеописанного способа авторы изобретения получили эмбрион, SNU6 (линию соматических клеток человека), с использованием клеток кожи человека в качестве доноров ядер. Этот эмбрион был депонирован Международным депозитарием, КСТС, Корейской Коллекцией Типовых культур (Korean Collection for Type Cultures; KRIBB # 52, Oundong, Yusong-ku, Taejon, 305-333, Republic of Korea) 19 июня 2000 года под номером доступа КСТС 0805ВР.
Данное изобретение иллюстрируется дополнительно в следующих примерах, которые не должны рассматриваться как ограничение объема данного изобретения.
Пример 1: Получение донорных клеток и реципиентных ооцитов
Для получения донорных клеток ткань, собранную из кожи человека, промывали ЗФР (забуферемным фосфатом солевым раствором, GIBCO BRL, Life Technologies, USA) и измельчали до размера 100 меш. Затем эту ткань инкубировали в ЗФР, содержащем 0,25% трипсин, 1 мМ ЭДТА и 1 мг/мл коллагеназы типа II, в течение 1 часа в условиях 39oС, 5% СО2. После расщепления ткани ферментами ее центрифугировали при 1500 об/мин в течение 2 минут и суспендировали в DMEM (модифицированной по способу Дульбекко среде Игла GIBCO BRL, Life Technologies, USA), дополненной 10% ФТС, 1% NEAA (не-незаменимыми аминокислотами) и 1% смесью пенициллин-стрептомицин. Суспензию переносили на чашки для культивирования клеток и инкубировали в условиях 39oС, 5% CO2 с получением линии соматических клеток. После этого клетки трипсинизировали в растворе, содержащем 0,25% трипсин и 1 мМ ЭДТА, и доводили число клеток до 2 х 104 клеток/мл для помещения аликвот клеток в пробирки Эппендорфа.
Фигура 1 изображает соматические клетки, выделенные в виде отдельных клеток для ядерного донора.
С другой стороны, для реципиентных ооцитов, фолликулы, размер которых был 2-6 мм в диаметре, отсасывали из яичников Корейских коров шприцом на 10 мл, имеющим иглу 18G. Затем эту фолликулярную жидкость переносили в чашку 100 мм с сеткой (длина между линиями была 1 см), нарисованной на ее дне, и ооциты с гомогенной цитоплазмой и достаточным числом слоев клеток кумулуса вокруг них подвергали скринингу. Отобранные ооциты промывали три раза 2 мл промывающей среды ТСМ199 (см. таблицу 4) в чашках 35 мм и затем один раз культуральной средой ТСМ199 (см. таблицу 5). Наконец, ооциты культивировали в культуральной среде ТСМ199, содержащей 0,1% раствор эстрадиола (см. таблицу 6), 2,5% раствор фолликулостимулирующего гормона (см. таблицу 7) и 10% ФТС, с получением реципиентных ооцитов.
Пример 2: Перенос ядер соматических клеток
Реципиентные ооциты, полученные в примере 1, промывали один раз промывающей средой ТСМ199 и переносили в 0,1% раствор гиалуронидазы (Sigma Chemical Co. , USA), приготовленный смешиванием 1 мл промывающей среды ТСМ199 с 111 мкл исходного раствора гиалуронидазы (10 мг/мл в промывающей среде ТСМ199). После удаления клеток кумулуса из ооцитов в присутствии 0,1% раствора гиалуронидазы, денудированные ооциты промывали три раза и инкубировали в промывающей среде ТСМ199. Затем эти ооциты переносили в раствор цитохалазина В (Sigma Chemical Co. , USA), приготовленный смешиванием 1 мл промывающей среды ТСМ199, содержащей 10% ФТС, с 1 мкл исходного раствора цитохалазина (7,5 мг/мл в ДМСО), и блестящую зону каждого ооцита разрезали с использованием микроманипулятора для образования щели, через которую можно выжать 10-15% цитоплазмы из ооцита с получением энуклеированного (лишенного ядра) ооцита. Стадия энуклеации более конкретно иллюстрируется следующим образом: рабочую чашку помещали на чашку микроманипулятора и микроманипулятор снабжали удерживающей пипеткой на его левом плече и режущей пипеткой на его правом плече. Затем удерживающую пипетку и режущую пипетку помещали в направлении 9 часов и 3 часов, соответственно, и корректировали для свободного перемещения во всех направлениях путем помещения контроллера (регулятора) пипеток в середину. Эти две пипетки дополнительно корректировали, чтобы не давать им прикасаться к рабочей чашке, и их кончики помещали в середину микрокапельки путем перемещения их вверх и вниз по этой микрокапельке. Затем ооциты переносили из промывающей среды ТСМ199 в раствор цитохалазина В с использованием промывающих пипеток с раструбом (внутренний диаметр >200 мкм). Микроманипулятор сначала фокусировали на ооците с использованием его рукоятки грубой регулировки и рукоятки тонкой регулировки и этот фокус дополнительно корректировали перемещением этих двух пипеток вверх и вниз. Ооцит помещали с его первым полярным тельцем, ориентированным в направлении 12 часов и удерживающую пипетку помещали вблизи этого ооцита в направлении 9 часов ооцита для фиксации ооцита путем применения гидравлического давления. Фигура 2 показывает процесс разрезания блестящей зоны ооцита с использованием удерживающей пипетки и режущей пипетки. Как показано на фигуре 2, ооцит пронизывался режущей пипеткой (2) от направления 1 час к направлению 11 часов с особой осторожностью, чтобы не повредить цитоплазму ооцита. После этого, к удерживающей пипетке (1) прилагали гидравлическое давление для отделения ооцита (3) и удерживающую пипетку приводили в контакт с режущей пипеткой, проникающей через окаймляющую блестящую зону на верхней части первого полярного тельца, для разрезания части блестящей зоны путем соприкосновения этих двух пипеток. Щель на ооците, сделанную, как описано выше, использовали как для энуклеации, так и для инъекции донорной клетки. Фигура 3 показывает процесс энуклеации, удаляющий первое полярное тельце и ядро из ооцита. Как показано на фигуре 3, ооцит (3) помещали таким образом, что его щель была ориентирована вертикально, удерживали удерживающей пипеткой (1) на его нижней части для предотвращения его перемещения и мягко нажимали на его верхнюю часть режущей пипеткой (2) для получения энуклеированного ооцита. Энуклеированный ооцит промывали три раза промывающей средой ТСМ199 и инкубировали в культуральной среде ТСМ199.
После этого донорные клетки, полученные заранее, переносили в энуклеированные ооциты с использованием микроманипулятора. Сначала, инъекционную микрокапельку 4 мкл помещали на середину рабочей чашки с использованием раствора РНА-Р, приготовленного смешиванием 400 мкл промывающего раствора ТСМ199 и 100 мкл исходного раствора РНА-Р (фитогемагглютинина) (0,5 мг/мл в промывающем растворе ТСМ199). Затем делали две микрокапельки для донорных клеток, одну из которых помещали выше, а другую - ниже инъекционной микрокапельки на той же самой рабочей чашке с использованием ЗФР, содержащего 1% ФТС. После распределения этих микрокапелек с минеральным маслом, рабочую чашку помещали на чашку микроманипулятора.
Режущую пипетку, установленную на микроманипуляторе, заменяли инъекционной пипеткой. Энуклеированные ооциты промывали три раза промывающей средой ТСМ199 и переносили в инъекционную микрокапельку. Донорные клетки оттягивали в инъекционную пипетку и переносили в инъекционную микрокпельку. Фигура 4 показывает процесс перенесения соматической клетки в энуклеированный ооцит. Как показано на фигуре 4, энуклеированный ооцит помещали с его щелью, ориентированной в направлении 1 часа, фиксировали при помощи удерживающей пипетки и инъецировали донорной клеткой через щель с использованием инъекционной пипетки и гидравлического давления с получением реконструированного эмбриона. Этот эмбрион промывали три раза промывающей средой ТСМ199 и инкубировали в промывающей среде ТСМ199.
Пример 3: Электрослияние и активация
Реконструированные эмбрионы подвергали электрослиянию с использованием манипулятора клеток Electrocell Manipulator (ЕСМ 2001, ВТХ, USA) с последующей активацией. 15 мкл раствора маннита, содержащего 0,28 М маннита, 0,5 мМ HEPES (рН 7,2), 0,1 мМ MgSO4 и 0,05% БСА, добавляли к культуральной среде ТСМ199, содержащей реконструированные эмбрионы, с использованием пипетки с раструбом для промывания. После 1-минутной инкубации в указанной среде эмбрионы инкубировали в течение 1 минуты в растворе маннита, дополненном промывающим раствором ТСМ199, и наконец переносили в раствор маннита с использованием пипетки с раструбом для промывания. Камеру (камеру 3,2 мм 453) клеточного электроманипулятора заполняли раствором маннита, дополненным промывающей средой ТСМ199, и затем эмбрионы помещали в эту камеру таким образом, что их часть с донорной клеткой была обращена к катоду. После электрослияния эмбрионов путем приложения импульса переменного тока 0,75-2,00 кВ/см дважды с интервалом 1 сек в течение 15 мкс каждый раз, их переносили в промывающую среду ТСМ199 и промывали три раза этой средой от раствора маннита.
Для активации электрослитых эмбрионов их инкубировали в темноте в течение 4 минут в растворе иономицина (Sigma Chemical Co., USA), который был промывающей средой ТСМ199, содержащей 5 мкМ иономицин и 1% БСА. Исходный раствор иономицина получали растворением 1 мг иономицина в 1,34 мл ДМСО. Активированные эмбрионы инкубировали в течение 5 минут в чашке 35 мм, содержащей промывающую среду ТСМ199, дополненную 10% ФТС, для удаления иономицина из этих эмбрионов.
Пример 4: Постактивация и культивирование in vitro электрослитых эмбрионов
Активированные эмбрионы постактивировали в течение 4 часов в 25 мкл раствора циклогексимида (Sigma Chemical Co., USA), приготовленного добавлением исходного раствора циклогексимида (10 мг/мл в этаноле) к среде для культивирования in vitro, mTALP, в конечной концентрации 10 мкг/мл. Затем эмбрионы подвергали скринингу и отобранные эмбрионы инкубировали в течение 7 дней в условиях 39oС, 5% СО2. Во время культивирования развитие эмбрионов подвергали мониторингу по мере прохождения времени (см. таблицу 8).
Как показано в таблице 8, было ясно продемонстрировано, что способ межвидовой трансплантации ядер позволяет получить развитие клонированных эмбрионов человека до стадии морула/бластоциста, что в конечном итоге облегчает получение эмбриональных стволовых клеток человека из развившихся морул/бластоцист.
На основе вышеописанного способа авторы изобретения получили эмбрион, SNU6 (линию соматических клеток человека), с использованием клеток кожи человека в качестве доноров ядер. Этот эмбрион был депонирован международным депозитарием, КСТС (Корейской Коллекцией Типовых Культур; KRIBB #52, Oundong, Yusong-ku, Taejon, 305-333, Republic of Korea) 19 июня 2000 года под номером доступа КСТС 0805ВР.
Пример 5: Замораживание и оттаивание эмбрионов и трансплантация
Эмбрионы замораживали для долгосрочного хранения. Сначала среду для замораживания (см. таблицы 9 и 10) распределяли в чашки 35 мм и морозильник включали для поддержания температуры при -5oС. Эмбрионы, отобранные для замораживания, промывали ЗФР, содержащим 10% ФТС, и инкубировали в среде для замораживания в течение 20 минут. Затем эмбрионы оттягивали во французскую соломинку на 0,25 мл таким образом, что соломинка имела среду для замораживания, содержащую эмбрионы, в середине и два слоя воздуха на обоих концах. После герметизации нагреванием соломинки с использованием нагретого пинцета ее помещали в морозильник, выдерживали при -5oС в течение 5 минут и отделяли пинцетом, предварительно охлажденным жидким N2. После отделения соломинку охлаждали при скорости -0,3oС/мин до -30oС, выдерживали в течение 10 минут, когда температура достигала -30oС. В конце концов эти эмбрионы хранили в резервуаре с жидким N2.
Для оттаивания замороженных эмбрионов среду для оттаивания, содержащую ЗФР, дополненный 20% ФТС, готовили в чашках 35 мм и дополняли глицерином для получения сред для оттаивания, каждая из которых содержала 0%, 3% и 6% глицерина (см. таблицы 9 и 11). Затем замороженную соломинку вынимали из жидкого N2, выдерживали на воздухе в течение 5 секунд и оттаивали в резервуаре (>20 см в диаметре), содержащем теплую воду (30oС). После оттаивания соломинку разрезали в местах слоев воздуха на обоих концах и среду, содержащую эмбрионы, собирали. Эмбрионы исследовали под микроскопом. Для удаления среды для замораживания из эмбрионов их последовательно инкубировали в средах для оттаивания, содержащих 6% глицерин, 3% глицерин и 0% глицерин, каждый раз по 5 минут.
Как ясно показано и объяснено выше, данное изобретение обеспечивает способ получения клонированных эмбрионов человека с использованием способа межвидовой трансплантации ядер, предусматривающего перенесение ядер, происходящих из соматических клеток человека, в ооциты, полученные из коровы, и клонированные эмбрионы человека, полученные указанным способом. Согласно способу данного изобретения, получение клонированных эмбрионов человека может быть использовано для получения эмбриональных стволовых клеток человека, которые могут быть широко применены для различных целей, таких как лечение заболеваний и другие применения в фармацевтике и медицинской науке.
Разнообразные модификации данного изобретения, кроме показанных и описанных здесь, будут очевидными для специалистов в данной области из предыдущего описания. Подразумевается, что подобные модификации находятся в объеме прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОГО ТИГРА ПУТЕМ ПРИМЕНЕНИЯ МЕТОДА МЕЖВИДОВОЙ ТРАНСПЛАНТАЦИИ ЯДЕР | 2000 |
|
RU2205537C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОЙ КОРОВЫ | 2000 |
|
RU2205536C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛОНИРОВАННОГО ПРЕДСТАВИТЕЛЯ СЕМЕЙСТВА ПСОВЫХ | 2006 |
|
RU2391817C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНОЙ СТВОЛОВОЙ КЛЕТКИ ЧЕЛОВЕКА | 2007 |
|
RU2352637C1 |
| ТРАНСГЕННЫЕ КОПЫТНЫЕ ЖИВОТНЫЕ, ИМЕЮЩИЕ ПОНИЖЕННУЮ АКТИВНОСТЬ ПРИОННОГО БЕЛКА, И ИХ ПРИМЕНЕНИЯ | 2003 |
|
RU2384059C2 |
| ПАРТЕНОГЕНЕТИЧЕСКАЯ АКТИВАЦИЯ ЧЕЛОВЕЧЕСКИХ ООЦИТОВ ДЛЯ ПОЛУЧЕНИЯ ЧЕЛОВЕЧЕСКИХ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК | 2006 |
|
RU2469085C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ЭКЗОГЕННОГО БЕЛКА В МОЛОКЕ ТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ И СПОСОБ ОЧИСТКИ БЕЛКОВ ИЗ МОЛОКА | 2004 |
|
RU2360002C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ, КОТОРЫЕ ПРОДУЦИРУЮТ ЭКЗОГЕННЫЕ БЕЛКИ В МОЛОКЕ, И ТРАНСГЕННЫЕ МЛЕКОПИТАЮЩИЕ, ПОЛУЧЕННЫЕ ТАКИМ СПОСОБОМ | 2004 |
|
RU2390562C2 |
| СЛИТЫЙ БЕЛОК С УВЕЛИЧЕННОЙ ЭРИТРОПОЭТИНОВОЙ АКТИВНОСТЬЮ, НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ СЛИТЫЙ БЕЛОК, И СПОСОБ ПОЛУЧЕНИЯ СЛИТОГО БЕЛКА | 2002 |
|
RU2232192C2 |
| ТРАНСГЕНЕЗ У МЛЕКОПИТАЮЩИХ ПУТЕМ ИНТРАЦИТОПЛАЗМАТИЧЕСКОЙ ИНЪЕКЦИИ СПЕРМЫ | 1999 |
|
RU2267270C2 |






Изобретение относится к способам клонирования эмбрионов человека. Способ предусматривает стадии: получения линий донорных соматических клеток, собранных из человека; созревания ооцитов, собранных из яичника коровы, in vitro; удаления клеток кумулуса, окружающих ооциты, разрезания части блестящей зоны созревших ооцитов с получением щели и выжимания через эту щель части цитоплазмы, включающей в себя первое полярное тельце, с получением энуклеированных реципиентных ооцитов; перенесения ядра в реципиентный ооцит инъекцией донорных клеток в безъядерные реципиентные ооциты с последующими последовательными электрослиянием и активацией электрослитых клеток с получением эмбрионов; постактивации и культивирования эмбрионов in vitro. Получение данным способом клонированных эмбрионов человека может быть использовано для получения эмбриональных стволовых клеток человека, которые могут широко применяться в медицине и биологии. 12 з.п. ф-лы, 11 табл., 4 ил.
Приоритет по пунктам и признакам:
30.06.1999 по пп. 1-13;
04.01.2000 установлен по внесенным изменениям в пункты формулы.
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| КЕЧИЯН К.Н | |||
| и др | |||
| Преодоление фактора отсутствия фертильности спермы в процедуре ЭКО с помощью микрохирургии на Zona pellucida | |||
| - Проблемы репродукции | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |

