Настоящее изобретение относится к аденовирусам, к нуклеиновым кислотам, кодирующим эти вирусы, и к их применению, а в частности к получению лекарственного средства для лечения опухолей.
В настоящее время для лечения опухолей используется ряд способов лечения. Помимо хирургии, наиболее часто используются методы химиотерапии и лучевой терапии. Однако применение этих методов ассоциируется с серьезными побочными эффектами. Использование селективных по репликации онколитических вирусов явилось новой идеей, положенной в основу лечения опухолей. Эта идея заключается в инициации селективной внутриопухолевой репликации вирусного агента, которая приводит к репликации вируса, к лизису инфекционной опухолевой клетки и распространению этого вируса на смежные опухолевые клетки. Поскольку способность вируса к репликации ограничена опухолевыми клетками, то в нормальной ткани этот вирус не может реплицироваться, а поэтому эта ткань не подвергается лизису данным вирусом.
В настоящее время были проведены клинические испытания нескольких вирусных систем для оценки лизиса опухоли. Одним из примеров может служить аденовирус d11520 (Onyx-015), который был с успехом использован в фазах клинических испытаний I и II (Khuri F. et al. Nature Medicine 6, 879-885, 2000). Onyx-015 представляет собой аденовирус, у которого был полностью делетирован ген белка Е1В-55кДа. Осуществление полной делеции белка Е1В55кДа этого аденовируса основано на обнаружении того факта, что репликация и, таким образом, лизис клеток могут происходить под действием аденовирусного вектора, дефицитного по р53 (Kirn D. et al., Proc. Am. Soc. Clin. Oncol. 17, 391a, 1998), в то время как нормальные клетки остаются неповрежденными. Более конкретно, генный продукт Е1В-55кДа участвует в ингибировании р53, в транспорте вирусной мРНК и в “отключении” синтеза белка клетки-хозяина. Ингибирование р53 происходит посредством образования комплекса, состоящего из р53 и кодируемого аденовирусом белка Е1В-55кДа, и/или комплекса, состоящего из Е1В-55кДа и Е4orf6. p53, кодируемый TP53, является исходной точкой запуска комплексного регуляторного механизма (Zambetti, G.P. et al., FASEB J. 7, 855-865, 1993), который приводит, помимо прочего, к эффективному ингибированию репликации вирусов, таких как аденовирусы, в клетках. Ген ТР53 является делетированным или мутированным, примерно в 50% всех опухолей человека, что приводит к отсутствию желаемого апоптоза, и стимулируется химиотерапией или лучевой терапией, а поэтому такое лечение опухолей обычно не дает положительного результата.
Другая концепция использования онколитических аденовирусов основана на обнаружении того факта, что если белок Е1А присутствует в специфической делетированной форме или включает одну или несколько мутаций, которые не влияют на связывание с Rb/E2F, и/или с p107/E2F, и/или с p130/E2F, то такой аденовирус не будет индуцировать вступление инфицированных клеток в фазу S и будет способен реплицироваться в опухолевых клетках, не имеющих функционального белка Rb. Кроме того, белок Е1А может быть делетирован у N-конца и включать одну или несколько мутаций в аминокислотной области в положениях 1-76 белков Е1А соответственно, что будет приводить к ингибированию связывания Е1А с р300 и, таким образом, обеспечивать селективную репликацию в опухолевых клетках. Такие методы описаны в качестве примера в европейском патенте ЕР 0931830. Примерами таких вирусов являются AdΔ24, dl922-947, E1Ad/01/07 и СВ016 (Howe J.A. et al., Molecular Therapy 2, 485-495, 2000; Fueyo J. et al., Oncogene 19, 2-12, 2000; Heise C. et al., Nature Medicine 6, 11341139, 2001; Balague C. et al., J. Virol. 75, 7602-7611, 2001). Эти аденовирусные системы для онколизиса, известные специалистам, включают различные делеции в белке Е1А, и такие делеции были сделаны исходя из предположения, что функциональный белок Rb и комплексы, состоящие из неактивного белка Rb и Е2F, соответственно будут блокировать эффективную in vivo репликацию и, тем самым, будут обеспечивать репликацию аденовируса in vivo только в Rb-негативных/мутированных клетках. Эти уже известные аденовирусные системы были получены на основе на Е1А для регуляции in vivo репликации под контролем раннего промотора Е2 (англ., Е2-раннего промотора) и свободного Е2F (Dyson, N.Genes & Development, 12, 2245-2262, 1998).
Другая форма онколитических аденовирусных систем основана на использовании селективных промоторов для специфической экспрессии вирусного онкогена Е1А, который обеспечивает селективную репликацию в опухолевых клетках (Rodriguez, R. et al., Cancer Res. 57, 2559-2563, 1997).
Как описано выше, выбор теневого фенотипа клетки, который является подходящим для реализации соответствующей концепции, лежащей в основе механизма их действия, имеет важное значение для различных концепций действия аденовирусных онколитических вирусов. Другими словами, известные в настоящее время различные аденовирусные системы могут быть использованы лишь в том случае, если выполняются предварительно требуемые молекулярно-биологические условия. В результате этого использование таких систем ограничивается отдельными группами пациентов.
Конкретная проблема, связанная с лечением опухолевых заболеваний, возникла вследствие развития у пациентов резистентности ко многим лекарственным препаратам (обозначаемым МDR), которая представляет собой, в частности, хорошо изученную форму резистентности опухолей к цитостатическим лекарственным средствам (Gottesman and Pastan, Annu. Rev. Biochem. 62, 385-427, 1993). Такая резистентность обусловлена сверхэкспрессией мебрано-ассоциированного транспортного белка, Р-гликопротеина, который принадлежит к так называемым АВС-транспортерам (Stein U. et al., JBC 276, 28562-69, 2001, J. Wijnholds, Novartis Found Symp., 243, 69-79, 2002). Bargou R.C. et al., и Oda Y. et al. (Bargou R.C. et al., Nature Medicine 3, 447-450, 1997; Clin. Cancer Res. 4, 2273-2277, 1998), которые сумели продемонстрировать, что человеческий фактор транскрипции YВ-1, локализованный в ядре, непосредственно участвует в активации экспрессии указанного Р-гликопротеина. Последующие исследования подтвердили, что YВ-1 транспортируется в ядро при различных стрессовых условиях, таких как УФ-излучение, введение цитостатических средств (Koike K. et al., FEBS Lett 17, 390-394, 1997) и гипертермия (Stein U. et al., JBC 276, 28562-69, 2001). Другие исследования подтвердили, что ядерная локализация YВ-1, кроме того, оказывает воздействие на один из АВС-транспортеров. Этот АВС-транспортер называется МRР (англ., белок, ассоциированный с резистентностью ко многим лекарственным препаратам) и участвует в образовании так называемой атипичной независимой от Р-гликопротеина резистентности ко многим лекарственным препаратам (Stein U. et al., JBC 276, 28562-69, 2001).
Проблема, лежащая в основе настоящего изобретения, заключается в разработке технических средств, а в частности способов лечения организма, а более конкретно, организма человека и группы пациентов, соответственно, с использованием специфических онколитических активных агентов. Другая проблема, на решение которой направлено настоящее изобретение, заключается в разработке способа, позволяющего осуществлять лизис опухолей у пациентов с опухолевыми заболеваниями, которые являются резистентными к цитостатическим средствам, а в частности являются резистентными ко многим лекарственным средствам. В конечном счете, проблема, лежащая в основе настоящего изобретения, заключается в получении аденовируса, подходящего для лизиса клеток.
В соответствии с первым аспектом настоящего изобретения, указанная проблема может быть решена путем использования аденовируса, экспрессирующего первый белок, выбранный из группы, включающей белок Е1В и белок Е4, а затем второй белок, выбранный из группы, включающей белок Е1А.
В одном из вариантов осуществления изобретения первым белком является белок Е1В, а предпочтительно Е1В55kD.
В другом варианте осуществления изобретения первым белком является белок Е4, а предпочтительно Е4orf6.
В предпочтительном варианте осуществления изобретения, первый белок представляет собой комбинацию белка Е1В и белка Е4, а предпочтительно, комбинацию белка Е1В55kDa и белка Е4orf6.
В предпочтительном варианте осуществления изобретения, белком Е1А является белок Е1А12S.
Во втором аспекте настоящего изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем использования аденовируса, где указанный аденовирус содержит, по меньшей мере, одну нуклеиновую кислоту, кодирующую белок, выбранный из группы, включающей белки Е1В, белки Е4 и белки Е1А, и где, по меньшей мере, один белок находится под контролем промотора, который отличается от промотора, регулирующего экспрессию белка в аденовирусе дикого типа.
В одном из вариантов второго аспекта изобретения указанным аденовирусом является аденовирус первого аспекта настоящего изобретения.
В одном из вариантов второго аспекта изобретения, по меньшей мере, один белок представляет собой белок Е1В, а предпочтительно белок Е1В55kDa.
В одном из вариантов второго аспекта изобретения, по меньшей мере, один белок представляет собой белок Е4, а предпочтительно белок Е4orf6.
В одном из вариантов второго аспекта изобретения, по меньшей мере, один белок представляет собой белок Е1А, а предпочтительно белок Е1А12S.
В предпочтительном варианте второго аспекта изобретения, по меньшей мере, один белок представляет собой комбинацию белка Е1В и белка Е4, а предпочтительно комбинацию белка Е1В55kD и белка Е4orf6.
В одном из вариантов второго аспекта изобретения, по меньшей мере, один белок представляет собой комбинацию белка Е1В и белка Е1А, а предпочтительно комбинацию белка Е1В55kD и белка Е1А12S.
В предпочтительном варианте второго аспекта изобретения, по меньшей мере, один белок представляет собой комбинацию белка Е4 и белка Е1А, а предпочтительно комбинацию белка Е4orf6 и белка Е1А12S.
В одном из вариантов второго аспекта изобретения, по меньшей мере, один белок представляет собой комбинацию белка Е1В, белка Е4 и белка Е1А, а предпочтительно комбинацию белка Е1В55kD, белка Е4orf6 и белка Е1А12S.
В одном из вариантов второго аспекта изобретения, экспрессия белка Е1В регулируется промотором, где указанный промотор выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е1В.
В одном из вариантов второго аспекта изобретения, экспрессия белка Е4 регулируется промотором, где указанный промотор выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е4.
В предпочтительном варианте второго аспекта изобретения, промотор, регулирующий экспрессию белка Е1А, представляет собой промотор, регулируемый YВ-1, либо промотор, который может регулироваться YВ-1.
В предпочтительном варианте второго аспекта изобретения, промотор, регулирующий экспрессию белка Е1А, представляет собой аденовирусный поздний промотор Е2.
В одном из вариантов первого и второго аспектов изобретения, белок Е4, а предпочтительно белок Е4orf6 и белок Е1В, а предпочтительно белок Е1В55kDa находятся под контролем того же самого или общего промотора.
В третьем аспекте изобретения, проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса, где указанный аденовирус обеспечивает доставку YВ-1 в ядро посредством, по меньшей мере, одного аденовирусного белка, либо такая доставка YВ-1 в ядро опосредуется, по меньшей мере, одним аденовирусным белком, где указанный аденовирусный белок, предпочтительно, отличается от Е1А.
В одном из вариантов третьего аспекта изобретения указанным аденовирусом является аденовирус первого и/или второго аспекта настоящего изобретения.
В четвертом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса, где указанный аденовирус обеспечивает доставку YВ-1 для репликации аденовируса посредством, по меньшей мере, одного аденовирусного белка, либо опосредует такую доставку YВ-1 для репликации аденовируса посредством, по меньшей мере, одного аденовирусного белка, где указанный аденовирусный белок, предпочтительно, отличается от Е1А.
В одном из вариантов четвертого аспекта изобретения указанным аденовирусом является аденовирус первого и/или второго и/или третьего аспекта настоящего изобретения.
В одном из вариантов третьего и четвертого аспектов настоящего изобретения указанным аденовирусным белком является комплекс Е4orf6 и Е1В55kDa.
В пятом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса, где нуклеиновая кислота указанного аденовируса содержит, по меньшей мере, одну функционально неактивную аденовирусную область, где указанная область выбрана из группы, включающей область Е1, область Е3, область Е4 и их комбинации.
В одном из вариантов пятого аспекта изобретения указанной областью является область Е1.
В одном из вариантов пятого аспекта изобретения указанной областью является область Е3.
В одном из вариантов пятого аспекта изобретения указанной областью является область Е4.
В одном из вариантов пятого аспекта изобретения указанная область включает область Е1, область Е3, область Е4.
В шестом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса, где указанный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, включающую, по меньшей мере, один промотор и нуклеиновую кислоту, кодирующую аденовирусный белок, где указанным аденовирусным белком является белок Е1В, а предпочтительно белок Е1В55kDa.
В одном из вариантов шестого аспекта изобретения указанным аденовирусом является аденовирус первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого аспекта настоящего изобретения.
В одном из вариантов шестого аспекта изобретения, указанный промотор отличается от промотора Е1В.
В одном из вариантов шестого аспекта изобретения указанный промотор выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е1В.
В седьмом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса, где указанный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, включающую по меньшей мере, один промотор и нуклеиновую кислоту, кодирующую аденовирусный белок, где указанным аденовирусным белком является белок Е4, а предпочтительно, белок Е4orf6.
В одном из вариантов седьмого аспекта изобретения указанный промотор отличается от промотора Е4.
В одном из вариантов седьмого аспекта изобретения указанный промотор выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е4.
В одном из вариантов седьмого аспекта изобретения указанным промотором является промотор Е1А.
В восьмом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса, где указанный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, включающую, по меньшей мере, один промотор и нуклеиновую кислоту, кодирующую аденовирусный белок, где указанным аденовирусным белком является белок Е1А, а предпочтительно, белок Е1А12S.
В одном из вариантов восьмого аспекта изобретения указанным аденовирусом является аденовирус первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого аспекта, и/или шестого аспекта, и/или седьмого аспекта настоящего изобретения.
В одном из вариантов восьмого аспекта изобретения указанный промотор отличается от промотора Е1А.
В одном из вариантов восьмого аспекта изобретения указанный промотор выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого аспекта, и/или шестого аспекта, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус включает нуклеиновую кислоту, где указанная нуклеиновая кислота кодирует YВ-1.
В предпочтительном варианте восьмого аспекта изобретения указанная нуклеиновая кислота, кодирующая YВ-1, находится под контролем промотора, где указанным промотором является, предпочтительно, Е2-поздний промотор.
В одном из вариантов восьмого аспекта изобретения указанная нуклеиновая кислота, кодирующая YВ-1, находится под контролем промотора, где указанным промотором является YВ-1-зависимый и YВ-1-регулируемый промотор соответственно.
В одном из вариантов восьмого аспекта изобретения указанная нуклеиновая кислота, кодирующая YВ-1, представляет собой часть экспрессионной кассеты, содержащей нуклеиновую кислоту, кодирующую белок Е1А, а предпочтительно нуклеиновую кислоту, кодирующую белок Е1А12S.
В одном из вариантов восьмого аспекта изобретения указанная нуклеиновая кислота, кодирующая белок Е1А, отделена от нуклеиновой кислоты, кодирующей YВ-1, последовательностью IRES.
В одном из вариантов шестого, и/или седьмого, и/или восьмого аспекта изобретения указанная нуклеиновая кислота, кодирующая белок Е1А, а предпочтительно белок Е4orf6, и нуклеиновая кислота, кодирующая белок Е1В, а предпочтительно белок Е1В55kDa, содержится в экспрессионной кассете, где указанные две кодирующие последовательности, предпочтительно, разделены последовательностью IRES.
В предпочтительном варианте восьмого аспекта изобретения указанный промотор экспрессионной кассеты выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанные аденовирусные промоторы отличаются от промотора Е4 и отличаются от промотора Е1В, а предпочтительно, отличаются от промотора Е4 дикого типа и отличаются от промотора Е1В дикого типа.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус содержит экспрессионную кассету, включающую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты выбрана из группы, включающей аптамеры, рибозимы, аптазимы, антисмысловые молекулы и киРНК.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус содержит экспрессионную кассету, включающую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты представляет собой кодирующую нуклеиновую кислоту, и где указанная нуклеиновая кислота кодирует молекулу, выбранную из группы, включающей пептиды, полипептиды, белки, антикалины, антитела и фрагменты антител.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус содержит экспрессионную кассету, включающую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты выбрана из группы, включающей гены, индуцирующие апоптоз, гены пролекарств, ингибиторы протеазы, гены-супрессоры опухоли, цитокины и ингибиторы ангиогенеза.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанным аденовирусом является рекомбинантный аденовирус.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанным аденовирусом является аденовирусный мутант.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус является дефицитным по репликации.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус способен реплицироваться в клетках, содержащих YВ-1 с нарушенной регуляцией или имеющих YВ-1 в ядре.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанные клетки содержат YВ-1 в ядре независимо от клеточного цикла.
В одном из вариантов первого, и/или второго, и/или третьего, и/или четвертого, и/или пятого, и/или шестого, и/или седьмого, и/или восьмого аспекта настоящего изобретения указанный аденовирус не содержит белка Е1А13S, и/или указанный аденовирус не содержит нуклеиновой кислоты, кодирующей белок Е1А13S.
В девятом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения нуклеиновой кислоты, кодирующей аденовирус в соответствии с любым из 1-8 аспектов.
В десятом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения системы репликации, содержащей нуклеиновую кислоту в соответствии с девятым аспектом настоящего изобретения, и нуклеиновую кислоту вируса-помощника, где указанная нуклеиновая кислота вируса-помощника содержит одну или несколько экспрессионных кассет аденовируса в соответствии с любым из 1-8 аспектов.
В одном из вариантов десятого аспекта изобретения указанный аденовирус или нуклеиновая кислота, кодирующая этот аденовирус, не включает экспрессионную кассету, содержащуюся в вирусе-помощнике.
В одиннадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения вектора, содержащего нуклеиновую кислоту в соответствии с девятым аспектом изобретения и/или систему репликации в соответствии с десятым аспектом изобретения.
В одном из вариантов десятого аспекта изобретения указанным вектором является экспрессирующий вектор.
В двенадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем использования клетки, содержащей аденовирус в соответствии с любым из 1-8 аспектов изобретения, и/или нуклеиновую кислоту в соответствии с девятым аспектом изобретения, и/или систему репликации в соответствии с десятым аспектом изобретения, и/или вектор в соответствии с одиннадцатым аспектом изобретения.
В одном из вариантов двенадцатого аспекта изобретения указанной клеткой является эукариотическая клетка, предпочтительно, клетка животного, а более предпочтительно, клетка млекопитающего.
В предпочтительном варианте двенадцатого аспекта изобретения указанной клеткой млекопитающего является клетка, выбранная из группы, включающей клетки мышей, крыс, морских свинок, свиней, овец, коз, крупного рогатого скота, лошадей, собак, кошек и человека.
В тринадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена с использованием организма, предпочтительно, организма млекопитающего, содержащего аденовирус в соответствии с 1-8 аспектами изобретения, нуклеиновую кислоту в соответствии с девятым аспектом изобретения, систему репликации в соответствии с десятым аспектом изобретения, вектор в соответствии с одиннадцатым аспектом изобретения или клетку в соответствии с двенадцатым аспектом изобретения, где указанный организм, предпочтительно, выбран из группы, включающей мышей, крыс, морских свинок, свиней, овец, коз, крупного рогатого скота, лошадей, собак и кошек.
В четырнадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса в соответствии с любыми 1-8 аспектами изобретения, нуклеиновой кислоты в соответствии с девятым аспектом изобретения, системы репликации в соответствии с десятым аспектом изобретения, вектора в соответствии с одиннадцатым аспектом изобретения или клетки в соответствии с двенадцатым аспектом изобретения для репликации аденовируса, предпочтительно, для in vitro репликации аденовируса.
В пятнадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса в соответствии с любыми 1-8 аспектами изобретения, нуклеиновой кислоты в соответствии с девятым аспектом изобретения, системы репликации в соответствии с десятым аспектом изобретения, вектора в соответствии с одиннадцатым аспектом изобретения или клетки в соответствии с двенадцатым аспектом изобретения для получения аденовируса, предпочтительно, для in vitro получения аденовируса.
В шестнадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса в соответствии с любыми 1-8 аспектами изобретения, нуклеиновой кислоты в соответствии с девятым аспектом изобретения, системы репликации в соответствии с десятым аспектом изобретения, вектора в соответствии с одиннадцатым аспектом изобретения или клетки в соответствии с двенадцатым аспектом изобретения для экспрессии генов, а предпочтительно, генов, стимулирующих клеточный лизис, предпочтительно, клеточный лизис в процессе репликации аденовируса, и/или генов, стимулирующих опосредуемый аденовирусом клеточный лизис.
В одном из вариантов шестнадцатого аспекта изобретения экспрессируемыми генами являются трансгены, описанные в настоящей заявке.
В семнадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса в соответствии с любыми 1-8 аспектами изобретения, нуклеиновой кислоты в соответствии с девятым аспектом изобретения, системы репликации в соответствии с десятым аспектом изобретения, вектора в соответствии с одиннадцатым аспектом изобретения или клетки в соответствии с двенадцатым аспектом изобретения для получения лекарственного средства.
В одном из вариантов четырнадцатого-семнадцатого аспектов изобретения указанная клетка, в которой реплицируется аденовирус, содержит в своем ядре YВ-1, а предпочтительно эта клетка содержит YВ-1 в ядре независимо от клеточного цикла.
В одном из вариантов четырнадцатого-семнадцатого аспектов изобретения указанная клетка, в которой реплицируется аденовирус, содержит YВ-1 с нарушенной регуляцией.
В одном из вариантов применения семнадцатого аспекта изобретения указанное лекарственное средство предназначено для лечения опухолевых заболеваний.
В предпочтительном варианте применения семнадцатого аспекта изобретения указанное опухолевое заболевание выбрано из группы, включающей злокачественные заболевания, рак, раковые заболевания и опухоли.
В одном из вариантов применения семнадцатого аспекта изобретения указанные опухоли выбраны из группы, включающей солидные, несолидные, злокачественные и доброкачественные опухоли.
В одном из вариантов применения семнадцатого аспекта изобретения, по меньшей мере, часть клеток, образующих опухоль, содержит в своем ядре YВ-1, а предпочтительно эти клетка содержат YВ-1 в ядре независимо от клеточного цикла.
В одном из вариантов применения семнадцатого аспекта изобретения, по меньшей мере, часть клеток, образующих опухоль, содержит YВ-1 с нарушенной регуляцией.
В одном из вариантов применения семнадцатого аспекта изобретения, по меньшей мере, часть клеток, образующих опухоль, являются Rb-позитивными или Rb-негативными.
В одном из вариантов применения семнадцатого аспекта изобретения, по меньшей мере, часть клеток, образующих опухоль, обладают резистентностью, а предпочтительно, резистентностью к множеству фармацевтически активных агентов.
В предпочтительном варианте применения семнадцатого аспекта изобретения указанной резистентностью является полирезистентность.
В одном из вариантов применения семнадцатого аспекта изобретения указанной резистентностью является резистентность к противоопухолевым средствам, а предпочтительно к цитостатическим средствам, и/или резистентность, индуцируемая излучением.
В одном из вариантов применения семнадцатого аспекта изобретения пациент, для введения которому предназначен данный лекарственный препарат, имеет множество клеток, где указанными клетками являются клетки, описанные в различных вариантах применения семнадцатого аспекта настоящего изобретения.
В одном из вариантов применения семнадцатого аспекта изобретения указанный лекарственный препарат содержит, по меньшей мере, один дополнительный фармацевтически активный агент.
В одном из вариантов применения семнадцатого аспекта изобретения указанный лекарственный препарат вводят вместе с дополнительным фармацевтически активным агентом, либо указанный лекарственный препарат предназначен для такого введения.
В одном из вариантов применения семнадцатого аспекта изобретения указанный дополнительный фармацевтически активный агент выбран из группы, включающей цитокины, ингибиторы металлопротеиназы, ингибиторы ангиогенеза, цитостатические средства, ингибиторы тирозинкиназы и ингибиторы клеточного цикла.
В одном из вариантов применения семнадцатого аспекта изобретения указанный лекарственный препарат вводят до, во время или после облучения.
В предпочтительном варианте применения семнадцатого аспекта изобретения указанное облучение предназначено для лечения опухоли.
В одном из вариантов применения семнадцатого аспекта изобретения клетку или организм, для которых требуется лечение, подвергают лечебному мероприятию, выбранному из группы, включающей облучение, введение цитостатических средств и гипертермию.
В одном из вариантов применения семнадцатого аспекта изобретения указанное лечебное мероприятие является местным или системным.
В одном из вариантов применения семнадцатого аспекта изобретения, указанное облучение проводят с применением излучения высокой энергии, а предпочтительно любого излучения, обычно применяемого для лечения опухолевых заболеваний.
В восемнадцатом аспекте изобретения, проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса в соответствии с любыми 1-8 аспектами изобретения, нуклеиновой кислоты в соответствии с девятым аспектом изобретения, системы репликации в соответствии с десятым аспектом изобретения, вектора в соответствии с одиннадцатым аспектом изобретения или клетки в соответствии с двенадцатым аспектом изобретения в целях получения лекарственного препарата для лечения опухолевых заболеваний, отличающихся тем, что указанное опухолевое заболевание выбрано из группы, включающей опухоли молочной железы, опухоли костей, опухоли желудка, опухоли тонкого кишечника, опухоли желчного пузыря, опухоли поджелудочной железы, опухоли печени, опухоли почек, опухоли головного мозга, опухоли яичника, опухоли кожи, опухоли придатков кожи, опухоли головы и шеи, опухоли матки, опухоли синовиальной оболочки, опухоли гортани, опухоли пищевода, опухоли языка, опухоли предстательной железы, а предпочтительно, одно из вышеупомянутых опухолевых заболеваний, определенных в любом из предыдущих аспектов изобретения.
В девятнадцатом аспекте изобретения проблема, лежащая в основе настоящего изобретения, может быть решена путем применения аденовируса в соответствии с любыми 1-8 аспектами изобретения, нуклеиновой кислоты в соответствии с девятым аспектом изобретения, системы репликации в соответствии с десятым аспектом изобретения, вектора в соответствии с одиннадцатым аспектом изобретения или клетки в соответствии с двенадцатым аспектом изобретения в целях получения лекарственного препарата для лечения опухолевых заболеваний, где указанным опухолеспецифическим промотором является промотор, специфичный к опухоли, для лечения которой предназначен данный лекарственный препарат.
В двадцатом аспекте изобретения, проблема, лежащая в основе настоящего изобретения, может быть решена путем применения фармацевтической композиции, содержащей аденовирус в соответствии с любыми 1-8 аспектами изобретения, нуклеиновую кислоту в соответствии с девятым аспектом изобретения, систему репликации в соответствии с десятым аспектом изобретения, вектор в соответствии с одиннадцатым аспектом изобретения или клетку в соответствии с двенадцатым аспектом изобретения и, необязательно, фармацевтически приемлемый носитель.
Описанный выше аденовирус настоящего изобретения, а в частности аденовирус, описанный в соответствии с 1-8 аспектами настоящего изобретения, также относится к аденовирусам группы I, и к аденовирусам, имеющим трансактивирующий онкогенный белок, такой как, например, Е1А, и/или к аденовирусам, описанным в настоящей заявке, а в частности описанным выше, а также к аденовирусам, используемым в соответствии с настоящим изобретением, а именно к аденовирусам группы II. Все аденовирусы группы I и аденовирусы группы II вместе будут далее называться просто аденовирусами, либо аденовирусами настоящего изобретения, либо вирусами настоящего изобретения.
Настоящее изобретение основано на неожиданном обнаружении того факта, что обратное изменение экспрессионной последовательности аденовирусных генов приводит к эффективной репликации и, необязательно, к лизису клетки, инфицированной аденовирусом. Что касается хронологически меняющейся экспрессии аденовирусных генов, то следует особо подчеркнуть, что в данном случае сначала экспрессируется белок Е1В и белок Е4, называемые далее, отдельно или вместе, первым белком, который экспрессируется перед экспрессией второго белка. Второй белок выбран из группы, включающей белки Е1А. Эта экспрессионная последовательность является обратной по отношению к последовательности аденовируса дикого типа, где сначала экспрессируется белок Е1А, и лишь за ним экспрессируются белок Е1В и белок Е4, что обеспечивает активацию факторов транскрипции, например, их транспорт в ядро инфицированной клетки и влияние на последующую активность репликации или регуляцию такой активности. Кинетика аденовирусных транскриптов в аденовирусах дикого типа описана, например, в работе Glenn G.M. и Ricciardi R.P. Virus Research 1988, 73-91, где сообщается, что в вирусе дикого типа транскрипты Е1А, то есть транскрипт Е1А12S и транскрипт Е1А13S, обычно детектируются до образования транскриптов и продуктов трансляции, Е4orf6 и Е1В55k соответственно. В настоящем изобретении белок Е1В, предпочтительно, представляет собой белок Е1В-55kD, если это не оговорено особо в описании данной заявки. В настоящем изобретении, белок Е4, предпочтительно, представляет собой белок Е4orf6, если это не оговорено особо в описании данной заявки. В настоящем изобретении белок Е1А, предпочтительно, представляет собой белок Е1А12S, если это не оговорено особо в описании настоящей заявки, либо белок Е1А, описанный здесь в связи с Е1А-модифицированными аденовирусами.
В соответствии с настоящим изобретением, белок Е1А, а в частности также белок Е1А12S, может быть в принципе заменен. Такие замененные белки Е1А и белки Е1А12S также относятся к белкам, называемым здесь белком Е1А и белком Е1А12S, соответственно, либо, предположительно, к белкам, охватываемым этим термином, если не указано обратное. Вместо белка Е1А12S может быть также использован белок Е1А, который обладает супрессорным действием на опухоль, и который описан, например, Dickopp A., Esche H., Swart G., Seeber S., Kirch HC, Opalka B. Cancer Gene Ther. 2000, Jul; 7(7):1043-50. Другие производные белков Е1А, а в частности, белок Е1А12S, используемый и/или обозначаемый таким образом в настоящем описании, также, в основном, представляют собой белки, способные высвобождать фактор Е2F из комплекса Rb/Е2F. Такими белками являются, среди прочих, опухолевый антиген обезьяньего вируса 40 (большой Т-антиген SV40), белок папилломавируса Е7 (HPV Е7), описанные Chеllappan S. et al., Proc. Natl. Acad. Sci. USA 1992, 89, 4549-4533.
В соответствии с настоящим изобретением могут быть использованы производные Е4orf6 и Е1В55k, где используемые здесь термины Е4orf6 и Е1В55k включают такие производные. Эти производные описаны, например, Shen Y. et al., J. of Virology 2001, 75, 4297-4307; Querido E. et al., J. of Virology 2001, 75, 699-709.
В соответствии с настоящим изобретением, белок Е1В экспрессируется перед экспрессией белка Е1А, либо белок Е4 экспрессируется перед экспрессией белка Е1А, либо белок Е1В и белок Е4 оба экспрессируются перед экспрессией белка Е1А, где каждый из этих белков описан выше.
Сконструированный таким образом аденовирус способен реплицироваться на очень высоком уровне после инфицирования клетки, которая экспрессирует YВ-1 в ядре, а предпочтительно, экспрессирует YВ-1 в ядре независимо от клеточного цикла, или которая содержит YВ-1 с нарушенной регуляцией, предпочтительно, в цитоплазме. Не имея намерения ограничиваться нижесказанным, авторы настоящего изобретения лишь предполагают, что комплекс, состоящий из белка Е1В и/или белка Е4, и отдельные комплексы, состоящие из этих двух белков, соответственно обладает/обладают способностью транспортировать YВ-1 с нарушенной регуляцией в клеточное ядро, или обладает/обладают способностью инициировать репликацию аденовируса под действием белка Е1В и/или белка Е4, экспрессирующихся перед экспрессией белка Е1А. После транспорта в клеточное ядро или активации, указанный YВ-1 может, как описано в настоящей заявке, обеспечивать эффективную репликацию, в частности, с помощью Е2-позднего промотора. Таким образом, хронологически ранняя экспрессия белка Е1В и/или белка Е4 препятствует прохождению каскада реакций, наблюдаемого для белка дикого типа вместе с начальной экспрессией белка Е1А. В предпочтительном варианте осуществления изобретения, белок Е1А представляет собой белок Е1А, который, в частности, больше не обеспечивает трансактивацию белка Е1В и/или белка Е4, либо он трансактивирует эти белки лишь на очень ограниченном уровне. Предпочтительно, чтобы такая трансактивация была недостаточной для обеспечения эффективной репликации и недостаточной для обеспечения репликации в клетках, которые не содержат YВ-1 в ядре. При этом, предпочтительно, чтобы такая трансактивация не происходила в клетках, которые не содержат YВ-1 в ядре независимо от клеточного цикла, или которые не содержат YВ-1 с нарушенной регуляцией.
Кроме того, настоящее изобретение основано на неожиданном обнаружении того факта, что аденовирус способен реплицироваться на достаточно эффективном уровне, если он содержит, по меньшей мере, нуклеиновую кислоту, которая кодирует белок, где указанный белок выбран из группы, включающей белки Е1В, белки Е4 и белки Е1А, и где, по меньшей мере, один из этих белков находится под контролем промотора, отличающегося от промотора, регулирующего экспрессию соответствующего белка в аденовирусе дикого типа. Такая репликация является особенно эффективной и обычно она приводит к лизису опухоли в том случае, если клетки содержат YВ-1 в ядре, а в частности если они содержат YВ-1 в ядре независимо от клеточного цикла, или в том случае, если такие клетки содержат YВ-1 с нарушенной регуляцией, а в частности, YВ-1 с нарушенной регуляцией, в цитоплазме. Все, что было сказано выше в отношении белков Е1В, относится также к белкам Е4 и к белкам Е1А. В аденовирусах дикого типа белок Е1В регулируется промотором Е1В, белок Е4 регулируется промотором Е4, а белок Е1А регулируется промотором Е1А. Путем выбора промоторов, отличающихся от промоторов, которые регулируют экспрессию вышеупомянутых белков в аденовирусах дикого типа, экспрессия вышеупомянутых белков, а значит и регулирующее взаимодействие отдельных аденовирусных нуклеиновых кислот и белков могут быть изменены. Путем выбора промоторов может быть получен хронологически отличающийся характер экспрессии, который, как предполагается, не ограничивается нижеуказанным механизмом, будет приводить к наблюдаемой репликации в клетках, где указанным механизмом может быть механизм, который был уже описан ранее для хронологически отличающейся экспрессии аденовирусных белков Е1В, Е4 и Е1А. Пример конкретной конструкции для регуляции экспрессии указанных белков промоторами, отличающимися от промоторов, регулирующих экспрессии соответствующих белков в аденовирусе дикого типа, может быть взят из зависимых пунктов формулы изобретения и из экспериментальной части изобретения, а особенно репрезентативными примерами являются вирусы, обозначенные здесь как XVirPSJL1 и XVirPSJL2. Предпочтительным белком Е1В является белок Е1В55kD, предпочтительным белком Е4 является белок Е4orf6, а предпочтительным белком Е1А является белок Е1А12S.
Промоторы, которые, предпочтительно, регулируют белок Е1В, а также белок Е4, выбраны из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, при условии, что указанные используемые аденовирусные промоторы отличаются от промотора Е1В в случае регуляции экспрессии белка Е1В и отличаются от промотора Е4 в случае регуляции экспрессии белка Е4. Применение промотора Е1А для регуляции экспрессии белка Е1В и/или белка Е4 является особенно предпочтительным. Промотор Е1А описан, например, Boulanger P. and Blair E.; Biochem. J. 275, 281-299. Кроме того, может быть также использован каждый из гетерологичных промоторов и любой другой гетерологичный промотор, то есть промотор, который отличается от промотора, регулирующего экспрессию соответствующего белка в аденовирусе дикого типа. Репрезентативным примером такого промотора является промотор CMV, а также другие подходящие промоторы, известные специалистам.
Промотор, используемый для регуляции экспрессии белка Е1А, может быть также выбран из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, при условии, что указанный аденовирусный промотор отличается от промотора Е1А. В настоящем изобретении предусматривается, что один или несколько вышеупомянутых белков, то есть белок Е1В, белок Е4 или белок Е1А, находятся под контролем одного и того же промотора, однако предпочтительно, чтобы под контролем одного и того же промотора находились белок Е1В и белок Е4. Особенно предпочтительно, чтобы экспрессия белка Е1А регулировалась YВ-1-регулируемым промотором или промотором, который может регулироваться YВ-1. Такие промоторы описаны здесь в соответствии с другими аспектами настоящего изобретения. Применение аденовирусного Е2-позднего промотора является особенно предпочтительным для регуляции экспрессии промотора Е1А, поскольку, во-первых, он может регулироваться YВ-1, а во-вторых, при наличии такого промотора, но в отсутствии YВ-1 обнаруживается лишь небольшой уровень транскрипции, который может быть фактически сведен до исключительно низкого уровня, что будет обеспечивать очень хороший уровень регуляции экспрессии нуклеиновой кислоты, которая находится под контролем Е2-позднего промотора. Это будет гарантировать значительное повышение биологической безопасности, а в частности, при применении в медицине.
Кроме того, авторами настоящего изобретения было обнаружено, что аденовирусы могут особенно хорошо реплицироваться в клетках, которые содержат YВ-1 в ядре, а в частности YВ-1 в ядре, независимо от клеточного цикла, и/или которые содержат YВ-1 с нарушенной регуляцией, а предпочтительно, YВ-1 с нарушенной регуляцией в цитоплазме, в том случае, если YВ-1 предназначен для репликации, либо непосредственно, либо опосредованно, в частности, в клеточном ядре, либо в том случае, если доставка YВ-1 прямо или опосредованно обеспечивается аденовирусным белком, где указанный аденовирусный белок отличается от Е1А. Этот аспект настоящего изобретения отличается от другого аспекта, также описанного в настоящем изобретении, тем, что применение трансактивирующих Е1А-модифицированных аденовирусов, а предпочтительно, аденовирусов группы II, позволяет осуществлять репликацию этих вирусов в позитивных по ядерному YВ-1 опухолевых клетках, позитивных по ядерному YВ-1, а в частности, в позитивных по ядерному YВ-1 клетках, которые являются YВ-1-позитивными независимо от клеточного цикла, и в клетках, которые содержат YВ-1 с нарушенной регуляцией, а в частности содержат YВ-1 в цитоплазме, поскольку трансактивирующие свойства белка Е1А, а в частности белка Е1А13S не используются в данном случае, то есть в случае аденовирусов группы I, а используются в предпочтительном варианте, где белок Е1А13S является функционально неактивным, а поэтому он больше неспособен к трансактивации также белков Е4orf6 и Е1В55k, которые участвуют в транспорте и в прямой или опосредованной доставке YВ-1, соответственно, в ядро. Следовательно, в соответствии с этим аспектом настоящего изобретения, эффективная репликация аденовируса невозможна. В соответствии с этим, транспорт YВ-1 в ядро и доставка YВ-1 для репликации аденовируса соответственно больше не могут находиться под контролем прямого или опосредованного действия белка Е1А, но могут происходить благодаря экспрессии белка Е1В, а в частности белка Е1В55kD и/или белка Е4, а в частности белка Е4orf6, который не регулируется Е1А.
Этот вариант аденовируса может быть также получен посредством одного из вышеописанных механизмов, например, путем обеспечения хронологически более ранней экспрессии белка Е1В и/или белка Е4 по сравнению с экспрессией белка Е1А, или путем помещения одного или нескольких белков Е1В, белков Е4 и белков Е1А под контроль промотора, который отличается от промотора, регулирующего экспрессию соответствующего белка в аденовирусе дикого типа.
И, наконец, авторами настоящего изобретения было неожиданно обнаружено, что эффективная репликация аденовируса может также происходить, в частности, в клетках, которые содержат YВ-1 в своем ядре, а особенно, если они содержат YВ-1 в ядре независимо от клеточного цикла, или в клетках, которые содержат YВ-1 с нарушенной регуляцией, предпочтительно, в цитоплазме, в том случае, если, по меньшей мере, один из белков Е1В, белков Е4 и белков Е1А, а в частности, их предпочтительных форм, экспрессируется в экспрессионной кассете под контролем промотора. В одном из своих вариантов, настоящее изобретение, в основном, относится к трем экспрессионным кассетам, каждая из которых содержит один из указанных белков. В альтернативном варианте изобретения экспрессионная кассета может также содержать два или более белков Е1В, Е4 и Е1А и их производные, и возможные замены, соответственно, а особенно в случае Е1А12S. Все, что было сказано выше в отношении аспекта, в котором аденовирусы содержат нуклеиновые кислоты, относящиеся к белкам Е1В, Е4 и Е1А, также применимо для конструирования различных белков и, соответственно, используемых промоторов. При использовании таких экспрессионных кассет, предпочтительно, чтобы белки и кодирующие их нуклеиновые кислоты в геноме аденовируса дикого типа, которые соответствует подходящим белкам экспрессионных кассет, были полностью или частично делетированы, что дает возможность, по меньшей мере, до определенной степени, обеспечивать стабильность вируса и предотвращать рекомбинацию.
В принципе, экспрессионные кассеты могут быть клонированы в каждую область и в каждый сайт аденовируса, соответственно, но при этом, предпочтительно, чтобы одна или несколько кассет были встроены отдельно или в комбинации друг с другом в область Е1, в область Е3 и/или в область Е4 данного вируса. Нуклеиновые кислоты области Е1, Е3 и Е4 могут быть полностью делетированы, частично делетированы либо вообще не делетированы, однако в случае аденовирусов настоящего изобретения предпочтительно, чтобы нуклеиновая кислота, содержащая ген Е1А13S, была инактивирована или делетирована так, чтобы данный вирус не продуцировал какой-либо трансактивирующий белок Е1А. Степень такой делеции в одной или нескольких областях Е1, Е3 и Е4 определяется используемой экспрессионной кассетой и, необязательно, дополнительно введенными чужеродными генами или трансгенами, или другими экспрессионными кассетами, содержащими такие гены, то есть гены, которые отличаются от аденовирусных генов, по меньшей мере, в том смысле, что они не присутствуют в регуляторных последовательностях аденовирусной нуклеиновой кислоты, как это обычно наблюдается в аденовирусе дикого типа, или они не присутствуют в последовательности аденовирусных нуклеиновых кислот аденовирусов дикого типа в каждом сайте. В настоящем изобретении предусматривается, что нуклеиновые кислоты, содержащиеся в одной или нескольких экспрессионных кассетах, кодирующих белок Е1В, белок Е4 и/или белок Е1А, являются частично или полностью делетированными в аденовирусном геноме. В одном из вариантов изобретения, например, в аденовирусе настоящего изобретения, XVirPSJL1 или -2, аденовирусная нуклеиновая кислота, кодирующая Е4orf6, является частично делетированной, однако полная нуклеиновая кислота, кодирующая этот белок, содержится в экспрессионной кассете. Предпочтительно, это может быть также реализовано и для белка Е1В55k (также обозначаемого Е155Kd) и/или белка Е1А12S. В предпочтительных вариантах изобретения, число делеций выбирают так, чтобы достигался максимальный размер упаковки, составляющий примерно 103% от максимального размера упаковки аденовируса дикого типа, хотя этот предел рассматривается лишь как предпочтительный предел. В предпочтительных вариантах изобретения, возможные делеции, которые могут быть сделаны в аденовирусном геноме, также имеют лишь те ограничения, которые гарантируют получение все еще инфекционных и упакованных частиц. Точное число делеций может быть определено любым специалистом, исходя из описания изобретения и в соответствии со стандартными тестами.
В качестве исходного материала для конструирования описанных здесь аденовирусов может быть использован любой аденовирус дикого типа, однако могут быть также использованы и другие аденовирусы, сконструированные в соответствии с техническим описанием, приведенным в настоящем изобретении. Особенно предпочтительно использовать аденовирусы подгруппы С, а в этой подгруппе - аденовирус 2 и аденовирус 5.
Используемые здесь термины белок Е1В и белки Е1В, белок Е4 и белки Е4, а также белок Е1А и белки Е1 являются синонимами, если это не оговорено особо.
Используемый здесь термин “YВ-1 с нарушенной регуляцией” означает описанные здесь молекулу YВ-1 или белок YВ-1, которые присутствуют в форме, количественно и/или качественно отличающейся от YВ-1, обычно присутствующего в клетках, а предпочтительно, в неопухолевых клетках. YВ-1 с нарушенной регуляцией может быть охарактеризован и идентифицирован с помощью конкретных вирусов, способных реплицироваться в присутствии YВ-1 с нарушенной регуляцией, в клетках теневого фенотипа, содержащих такой YВ-1 с нарушенной регуляцией. В соответствии с этим конкретными вирусами являются вирусы, в которых белок Е1А является мутированным и обладает трансактивирующей функцией. Конкретными примерами таких вирусов являются AdΔ24, dl922-947, E1 Ad/01/07 и СВ 016 и/или вирусы, описанные Howe J.A. et al., Molecular Therapy 2, 485-495, 2000; Fueyo J. et al., Oncogene 19, 2-12, 2000; Heise C. et al., Nature Medicine 6, 1134-1139, 2001; Balague C. et al., J. Virol. 75, 7602-7611, 2001; Bautista D.S. et al., Virology 1991, 182, 578-596; Jelsma T.N. et al., Virology 1988, 163, 494-502; Wong H.K. and Ziff E.B., J. of Virology 1994, 68, 4910-4920]. Такая клетка и клетка, имеющая такой клеточный теневой фенотип, соответственно, могут быть использованы для репликации аденовирусов группы I и/или аденовирусов группы II. Кроме того, опухоли, содержащие такие клетки, могут быть подвергнуты лизису под действием аденовирусов настоящего изобретения.
Кроме того, настоящее изобретение основано на неожиданном обнаружении того факта, что репликация ДНК Е1А-модифицированных аденовирусов в опухолевых клетках, позитивных по ядерному YВ-1, обусловлена активацией Е2-позднего промотора. Следует отметить, что Е1А-модифицированные аденовирусы представляют собой аденовирусы, которые (а) обнаруживают снижение уровня репликации, или вообще не реплицируются в клетках, негативных по ядерному YВ-1, по сравнению с соответствующими клетками дикого типа, (b) оказывают трансактивирующее действие, по меньшей мере, на один вирусный ген, где указанный ген, в частности, выбран из группы, включающей Е1В-55kDa, E4orf6, E4orf3 и E3ADP, и/или (с) не транспортируют клеточный YВ-1 в ядро клетки посредством аденовируса. Используемые в настоящем изобретении аденовирусы, кроме того, но необязательно, характеризуются тем, что связывание кодируемого аденовирусом белка Е1А препятствует связыванию Е2F с Rb, и тем, что они обладают способностью растворять соответствующий комплекс, состоящий из Е2F и Rb. Аденовирусы, которые имеют один или несколько вышеупомянутых признаков (а)-(с), а предпочтительно все признаки (а)-(с), являются дефицитными по репликации в клетках, которые не содержат YВ-1 в ядре.
В одном из вариантов осуществления изобретения используемый здесь термин “значительное снижение уровня репликации”, в частности, означает уровень репликации, который в 2 раза, предпочтительно, в 5 раз, более предпочтительно, в 10 раз, а наиболее предпочтительно в 100 раз ниже по сравнению с уровнем репликации в клетках дикого типа. В предпочтительном варианте осуществления изобретения такое сравнение уровней репликации осуществляют с использованием идентичных или аналогичных клеточных линий, идентичных или аналогичных титров инфекционности вируса (множественности заражения, m.o.i., или бляшкообразующих единиц, б.о.е.) и/или идентичных или аналогичных общих экспериментальных условий. Термин “репликация”, в частности, означает образование вирусных частиц. В других вариантах осуществления изобретения термин “степень репликации” может означать уровень синтеза вирусной нуклеиновой кислоты. Методы определения уровней синтеза вирусной нуклеиновой кислоты, а также методы определения образования вирусных частиц известны специалистам.
Полученные данные, методы, применения, или нуклеиновые кислоты, белки, системы репликации и т.п. необязательно ограничены аденовирусами. В принципе, такие системы репликации также имеются и в других вирусах, которые также упоминаются в данном описании.
С использованием вирусов настоящего изобретения или с использованием вирусов, описанных в соответствии с настоящим изобретением, может быть осуществлена репликация, сравнимая с репликацией вируса дикого типа, при уровне инфицирования 1-10 б.о.е./клетку, в отличие от уровня инфицирования в предшествующих работах, составляющего 10-100 б.о.е./клетку.
Термин “клеточный YВ-1” означает любой YВ-1, который кодируется клеткой, а предпочтительно также экспрессируется клеткой, в которой присутствует YВ-1, в частности, перед инфицированием соответствующей клетки аденовирусом, а предпочтительно описанным здесь аденовирусом и/или вирусом-помощником. Однако, в соответствии с настоящим изобретением, “клеточный YВ-1” также означает YВ-1, который вводят в клетку или который продуцируется этой клеткой только после ее обработки экзогенными факторами, например, путем инфицирования вирусом, а предпочтительно, аденовирусом.
Не имея намерения ограничиваться в дальнейшем нижеприведенными соображениями, автор настоящего изобретения лишь высказывает предположение, что при репликации вирусов, используемых в настоящем изобретении, и в соответствии с применением аденовирусов настоящего изобретения, Е2-ранний промотор, т.е. ранний промотор Е2 не “запускается” посредством человеческого клеточного фактора транскрипции Е2F. В этих случаях такой “запуск” репликации не зависит от статуса Rb в клетках, то есть опухолевые клетки, которые подвергаются инфицированию описанными здесь вирусами и которые затем, предпочтительно, подвергаются лизису, могут содержать как функциональный, так и неактивный белки Rb. Кроме того, при применении описанных здесь аденовирусов или описанных здесь условий, для репликации аденовируса не требуется присутствия какого-либо функционального белка р53, однако его присутствие не оказывает негативного влияния на указанную репликацию. Поскольку данная техническая идея выходит за рамки принципа, лежащего в основе применения онколитических или туморолитических аденовирусов типа AdΔ24, dl922-947, E1Ad/01/07, CB016 или аденовирусов, описанных, например, в европейском патенте ЕР 0931830, то в белок Е1А были введены одна или несколько делеций, исходя из предположения, что интактные функциональные белки Rb препятствуют эффективной репликации in vivo, а поэтому репликация аденовируса in vivo происходит лишь в Rb-негативных или мутированных по Rb клетках. Эти известные аденовирусные системы были сконструированы на основе Е1А для регуляции in vivo репликации аденовирусов с помощью раннего Е2-промотора (Е2-раннего промотора) и “свободного Е2F”. Тем не менее, такие уже применявшиеся ранее вирусы могут быть использованы в соответствии с настоящим изобретением для репликации в клетках, которые содержат YВ-1 в ядре независимо от их клеточного цикла, или в клетках, которые содержат YВ-1 с нарушенной регуляцией.
Вирусы, а в частности аденовирусы, описанные в указанном Европейском патенте ЕР 0931830, могут быть использованы в соответствии с настоящим изобретением. Более конкретно, вирусы, описанные в указанном патенте, являются дефицитными по репликации и не содержат экспрессируемого вирусного онкопротеина, способного связываться с функциональным продуктом гена-суппрессора опухоли, Rb. Таким аденовирусом может быть, в частности, любой аденовирус, у которого отсутствует экспрессируемый вирусный онкопротеин Е1А, способный связываться с функциональным продуктом гена-суппрессора опухоли, а более конкретно Rb. Вирусный онкопротеин Е1А может содержать инактивирующую мутацию, например, в домене CR1 в положениях аминокислот 30-85 в аденовирусе Аd 5, который также обозначается здесь Аd5, Аd 5, в положениях нуклеотидов 697-790, и/или в домене CR2 в положениях аминокислот 120-130 в Аd 5 и в положениях нуклеотидов 920-967, которые участвуют в связывании белка р105 Rb, белка р130 и р107. Однако, в соответствии с настоящим изобретением, указанным аденовирусом является аденовирус типа 2 dl312 или аденовирус типа 5 NТ dl1010.
При применении аденовирусов в соответствии с настоящим изобретением в целях приготовления лекарственного препарата, а в частности в целях приготовления лекарственного препарата для лечения опухолевых заболеваний и других описанных здесь заболеваний, и при применении аденовирусов в соответствии с настоящим изобретением, а также при применении аденовирусов настоящего изобретения для репликации в клетках, которые содержат YВ-1 в ядре, а предпочтительно содержат YВ-1 в ядре независимо от клеточного цикла, или которые содержат YВ-1 с нарушенной регуляцией, предпочтительно, в цитоплазме, репликация, в конечном счете, происходит в тех клетках, которые имеют YВ-1 в ядре, предпочтительно, независимо от клеточного цикла, то есть другими словами в клетках, позитивных по ядерному YВ-1 или в клетках, которые содержат YВ-1 с нарушенной регуляцией. Следует особо подчеркнуть, что сами по себе аденовирусы не реплицируются или реплицируются лишь на значительно более низком уровне в клетках, содержащих YВ-1 не в ядре, а лишь в цитоплазме, или в клетках, которые не содержат YВ-1 с нарушенной регуляцией. Следовательно, для успешной репликации этих вирусов необходимо, чтобы YВ-1 присутствовал в ядре клетки, предпочтительно, независимо от клеточного цикла, либо чтобы присутствовал YВ-1 с нарушенной регуляцией. Как будет более подробно описано ниже, это может быть осуществлено, например, путем создания в клетках таких условий, которые будут обеспечивать экспрессию или присутствие YВ-1, предпочтительно, независимо от клеточного цикла, или присутствие YВ-1 с нарушенной регуляцией в ядре, или экспрессию YВ-1 с нарушенной регуляцией. Подходящими условиями могут быть, например, кодирование и экспрессия, соответственно, YВ-1 аденовирусами, которые либо применяются в соответствии с настоящим изобретением, либо являются объектом настоящего изобретения, и которые, помимо аденовирусных генов, также содержат генетическую информацию, кодирующую YВ-1, а в частности обеспечивающую его экспрессию. Другими способами обеспечения транспорта, индуцирования или экспрессии YВ-1 в ядре клетки является создание в клетке или в организме, содержащем такую клетку, стрессовых условий, таких как введение цитостатических средств, облучение, гипертермия и т.п. В предпочтительном варианте осуществления изобретения указанным облучением является любое облучение, которое применяется, например, для лечения опухолевых заболеваний.
Аденовирусы, используемые в настоящем изобретении, а в частности, для лизиса опухоли, а также аденовирусы, применяемые в предпочтительных вариантах настоящего изобретения, отличаются тем, что они не реплицируются в клетках, не содержащих YВ-1 в своем ядре независимо от клеточного цикла, и которые, таким образом являются негативными по ядерному YВ-1, или которые не содержат какого-либо YВ-1 с нарушенной регуляцией.
Другим отличительным признаком части аденовирусов, применяемых в соответствии с настоящим изобретением и отличающихся от аденовирусов настоящего изобретения, является то, что они кодируют вирусный онкоген, который также называется здесь онкогенным белком, где указанным онкогенным белком, предпочтительно, является Е1А, и где указанный онкогенный белок способен активировать, по меньшей мере, один вирусный ген, который влияет на репликацию вируса и/или на лизис клеток, инфицированных указанным вирусом. При этом его влияние на репликацию должно быть, предпочтительно, таким, чтобы данный вирус реплицировался лучше в присутствии онкогенного белка, чем в его отсутствие. Этот процесс также называется здесь трансактивацией, а в частности Е1А-трансактивацией, в случае, когда указанная трансактивация опосредуется Е1А. Термины “трансактивировать” или “трансактивация” описывают, предпочтительно, процесс, в котором соответствующий вирусный онкопротеин влияет на экспрессию и/или транскрипцию одного или нескольких других генов, отличающихся от самого гена, кодирующего указанный вирусный онкогенный белок, то есть регулирует его/их экспрессию и/или трансляцию, а в частности активирует эту экспрессию/трансляцию. Такими вирусными генами являются, предпочтительно, Е1В55kDa, E4orf6, E4orf3 и E3ADP, а также любые комбинации вышеупомянутых генов и генных продуктов соответственно.
Другим, хотя и необязательным отличительным признаком аденовирусов, применяемых в соответствии с настоящим изобретением, а также аденовирусов настоящего изобретения, является их способность связываться и способность конкретно кодируемых ими белков связываться, соответственно, с супрессором опухоли Rb. В принципе, в настоящем изобретении аденовирусы, применяемые в соответствии с настоящим изобретением, могут связываться, а могут и не связываться с Rb. Применение любых двух альтернативных вариантов аденовирусов не зависит от статуса Rb в обрабатываемых клетках или в клетках, предназначенных для обработки.
Для придания онкопротеину Е1А неспособности связываться с Rb в него могут быть введены нижеследующие делеции, например делеция в области CR1 (положения аминокислот 30-85 в Аd5) и делеция в области CR2 (положения аминокислот 120-139 в Аd5). При этом область CR3 сохраняется и может оказывать свое трансактивирующее действие на другие ранние вирусные гены.
Для придания онкопротеину Е1А способности к связыванию с Rb в данном онкопротеине Е1А, в принципе, могут быть созданы следующие делеции: делеция области CR3 (положения аминокислот 140-185); делеция N-конца (положения аминокислот 1-29); делеция аминокислот в положениях 85-119; и делеция С-конца (положения аминокислот 186-289). Указанные выше области не препятствуют связыванию E2F с Rb. При этом трансактивирующая функция сохраняется, хотя и является несколько ослабленной по сравнению с функцией Аd5 дикого типа.
Кроме того, в соответствии с настоящим изобретением, особенно, если это касается аденовирусов настоящего изобретения, белок Е1А, а в частности, белок Е1А12S, конструируют так, чтобы в одном из вариантов изобретения он обладал способностью связываться с Rb, а в другом варианте изобретения он не обладал способностью связываться с Rb, где указанным белком Е1А12S является белок Е1А, а в частности белок Е1А12S, который определен в настоящем изобретении, и который, тем не менее, в предшествующих работах иногда называется модифицированным Е1А12S. Соответствующее конструирование белка Е1А12S может быть осуществлено любым специалистом, а в частности в отношении вышеупомянутых делеций белка Е1А, который также называется здесь просто Е1А.
Такие аденовирусы, которые, в основном, уже известны специалистам и которые не обнаруживают какой-либо трансактивации, в основном рассматриваются как дефицитные по репликации. Однако заслуга автора настоящего изобретения состоит в том, что им было установлено, что указанные вирусы, тем не менее, способны реплицироваться в подходящей среде, а в частности в подходящем клеточном теневом фенотипе. Такой подходящий клеточный теневой фенотип индуцируется или обеспечивается присутствием YВ-1 в ядре, а предпочтительно, присутствием YВ-1 в ядре независимо от клеточного цикла, или присутствием YВ-1 с нарушенной регуляцией. Термины “клетки” или “клеточные системы”, используемые здесь в связи с каждым и любым другим аспектом настоящего изобретения, означают фрагменты клеток или фракции клеточных экстрактов, а также клетки, которые присутствуют in vitro, in vivo или in situ. Кроме того, термины “клеточные системы” или “клетки” также включают клетки, присутствующие в клеточной культуре, в культуре тканей, в культуре органов или в какой-либо другой ткани или в другом органе in vivo и in situ соответственно; выделенные клетки; клетки, присутствующие в группах или составляющие часть тканей, органов или организмов, и которые могут также присутствовать, предпочтительно, в живом организме. Указанным организмом, предпочтительно, является организм любых позвоночных, а более предпочтительно, организм млекопитающего. Более предпочтительным организмом является организм человека. Другие предпочтительные организмы описаны в соответствии с различными аспектами настоящего изобретения.
Кроме того, в настоящем изобретении, которое основано на описанной здесь технической идее, были продуцированы новые вирусы, которые подвергались репликации, характерной для описанных здесь аденовирусов, и аденовирусов, описанных в предшествующих работах, в клетках, которые являются позитивными по ядерному YВ-1, предпочтительно, позитивными по ядерному YВ-1 независимо от клеточного цикла, или которые содержат YВ-1 с нарушенной регуляцией. Другими словами, а в частности исходя, предпочтительно, из уже известных аденовирусов, могут быть сконструированы другие вирусы, которые имеют описанные здесь признаки, необходимые для их использования в настоящем изобретении.
В соответствии с настоящим изобретением, модифицированный онкопротеин Е1А различных аденовирусов, применяемых в соответствии с настоящим изобретением, в отличие от вирусов настоящего изобретения, обладает способностью трансактивировать ранние вирусные гены, такие как Е1В55К, Е4orf3, E4orf6, E3ADP, в клетках, позитивных по ядерному YB-1, или в клетках, содержащих YВ-1 с нарушенной регуляцией. При этом предпочтительно, чтобы в вирусном геноме отсутствовали какие-либо другие замены, а соответствующий аденовирус мог, так или иначе, соответствовать аденовирусу дикого типа или его производному.
Описанные здесь вирусы, которые кодируют трансактивирующий онкогенный белок, определенный в настоящем изобретении, включают, например, аденовирусы AdΔ24, dl922-947, E1Ad/01/07, CB106 и/или аденовирусы, описанные в европейском патенте ЕР 0931830, каждый из которых способен трансактивировать ранние гены, такие как Е1В, Е2, Е3 и/или Е4, и каждый из которых сравним с аденовирусами дикого типа, а в частности с Ad5 дикого типа. В этих случаях конкретная область белка Е1А ответственна за трансактивацию. В белке Е1А аденовирусов различных серотипов присутствуют три в высокой степени консервативных области. Область CR1 простирается от аминокислоты в положении 41 до аминокислоты в положении 80, область CR2 простирается от аминокислоты в положении 120 до аминокислоты в положении 139, а область CR3 простирается от аминокислоты в положении 140 до аминокислоты в положении 188. Трансактивирующая функция обусловлена главным образом присутствием области CR3 в белке Е1А. Аминокислотная последовательность CR3, присутствующая в вышеупомянутых аденовирусах, остается неизмененной. Это приводит к трансактивации ранних генов Е1В, Е2, Е3 и Е4 независимо от того, присутствует ли YВ-1 в ядре или в цитоплазме.
В противоположность этому, в рекомбинантном аденовирусе dl520 область CR3 была делетирована. Таким образом, dl520 экспрессирует так называемый белок Е1А12S, который не содержит аминокислотную последовательность области CR3. Таким образом, dl520 может обладать лишь очень слабым трансактивирующим действием, в частности, на область Е2, а поэтому он не реплицируется в клетках, негативных по ядерному YВ-1. В клетках, позитивных по ядерному YВ-1, YВ-1 является ответственным за трансактивацию области Е2 и тем самым обеспечивает эффективную репликацию dl520. Этот факт был положен в основу применения таких систем, как dl520, или систем на основе dl520 в описанных здесь целях. Более важное отличие между двумя ранее описанными группами аденовирусов, таких как, например, дельта 24 (также называемого здесь АdΔ24) и, например, dl520, заключается в том, что ранние гены Е1В, Е3 и Е4 являются в более высокой степени трансактивированными в клетках, позитивных по ядерному YВ-1 независимо от клеточного цикла, или в клетках, содержащих YВ-1 с нарушенной регуляцией, чем в клетках, негативных по ядерному YВ-1, или в клетках, которые не содержат YВ-1 с нарушенной регуляцией. В противоположность этому, в случае дельта 24 такого отличия в трансактивации в указанных клетках либо не наблюдалось вообще, либо эти отличия были лишь очень незначительными. Однако трансактивация dl520, а более конкретно, белка Е1А12S, была значительно слабее, чем действие аденовируса дикого типа. Тем не менее, такая трансактивация является достаточной для эффективной репликации вируса в клетках, позитивных по ядерному YВ-1, как показано также в примере 10. Описанная в настоящей заявке, а в частности, описанная в этой связи конструкция белка Е1А и нуклеиновой кислоты, кодирующей описанный белок, где указанный белок Е1А имеет, по сравнению с онкогенным белком Е1А дикого типа, одну или несколько делеций и/или мутаций включая, особенно предпочтительно, конструкции белка Е1А, описанные для аденовирусов dl520 или АdΔ24, dl922-947, Е1Аd/01/07, СВ106 и/или аденовирусов, описанных в европейском патенте ЕР0931830, представляют собой варианты вирусов, а в частности аденовирусов, репликация которых регулируется, а предпочтительно, преимущественно, регулируется активацией Е2-позднего промотора. При этом предпочтительно, чтобы указанная делеция была выбрана из группы, включающей делеции области CR3, делеции N-конца и делеции С-конца. Другие варианты белка Е1А, которые обеспечивают такой тип репликации аденовирусов, могут быть генерированы специалистом исходя из описания настоящей заявки. Вариантом белка Е1А, описанным ранее, является вариант, который может быть использован в связи с использованием аденовирусов настоящего изобретения, также называемых здесь аденовирусами настоящего изобретения или аденовирусами группы I.
Аденовирусы настоящего изобретения, а в частности аденовирусы группы I, которые также называются здесь производными и которые могут быть использованы в соответствии с настоящим изобретением, обычно имеют делецию Е1, делецию Е1/Е3 и/или делецию Е4, то есть соответствующие аденовирусы не способны генерировать функционально активные продукты экспрессии Е1, и/или Е3, и/или Е4 и соответствующие им продукты. Либо, другими словами, эти аденовирусы способны генерировать лишь функционально неактивные продукты экспрессии Е1, Е3 и/или Е4, где функционально неактивный продукт экспрессии Е1, Е3 и/или Е4 представляет собой продукт экспрессии, который либо вообще не присутствует в качестве продукта экспрессии, либо он присутствует на уровне транскрипции и/или трансляции, либо он присутствует в форме, которая не обладает, по меньшей мере, одной из функций, присущей аденовирусу дикого типа. Эта(и) функция(и), присущие продукту экспрессии аденовируса дикого типа, известна(ы) специалистам и описана(ы) у Russell W.C. Journal of Virology, 81, 2573-2604, 2000. В работе Russell (см. выше) описаны также принципы конструирования аденовирусов и аденовирусных векторов, которые вводятся в настоящее описание посредством ссылки. В соответствии с настоящим изобретением, модифицированный онкопротеин Е1А, то есть онкопротеин, который больше не является трансактивирующим белком Е1А, и другие белки, такие как Е1А12S, Е1В-55К, E4orf6 и/или E3ADP (белок гибели аденовируса, ADP)) (Tollefson A. et al., J. Virology, 70, 2296-2306, 1996), экспрессируются в таком векторе отдельно или в любой комбинации. Эти отдельно названные гены, а также описанные здесь трансгены могут быть независимо друг от друга клонированы в область Е1, и/или Е3, и/или Е4 и могут экспрессироваться с помощью подходящего промотора или под контролем подходящего промотора. В основном каждая из областей Е1, Е3 и Е4 также является подходящей в качестве сайта клонирования в нуклеиновой кислоте аденовируса. В некоторых вариантах осуществления изобретения подходящими промоторами являются промоторы, описанные здесь в связи с регуляцией и экспрессией, соответственно, Е1А, а предпочтительно, модифицированного Е1А.
И наконец, в одном из вариантов осуществления изобретения, аденовирусы группы II, используемые в соответствии с настоящим изобретением, являются дефицитными по Е1В, а в частности, дефицитными по Е1В19кДа. Используемый здесь термин “дефицитный” относится, в основном, к состоянию, при котором Е1В не обладает всеми свойствами белка Е1В дикого типа, и не обладает, по меньшей мере, одним из этих свойств.
По меньшей мере, некоторые варианты аденовирусов группы II, применяемые в соответствии с настоящим изобретением, известны специалистам. Аденовирусами, применяемыми в соответствии с настоящим изобретением, предпочтительно, являются рекомбинантные аденовирусы, а в частности, аденовирусы которые, в отличие от аденовирусов дикого типа, имеют модификации, внесенные в соответствии с описанными здесь методами. Делеции и мутации в последовательностях нуклеиновой кислоты аденовируса, которые не играют важной роли с точки зрения настоящего изобретения, могут быть соответственно внесены самим специалистом. Такие делеции могут быть, например, сделаны в части нуклеиновой кислоты, кодирующей Е3 и Е4, как описано в настоящей заявке. Делеция Е4 является особенно предпочтительной, при условии, что такая делеция не распространяется на белок Е4orf6, или, другими словами, предпочтительно, чтобы аденовирус, применяемый в соответствии с настоящим изобретением, кодировал Е4orf6. В предпочтительных вариантах осуществления изобретения такие аденовирусные нуклеиновые кислоты могут быть, кроме того, упакованы в вирусные капсиды и, таким образом, могут образовывать инфекционные частицы. Этот принцип может быть применен и в отношении нуклеиновых кислот настоящего изобретения. При этом следует отметить, что, в основном, аденовирусные системы могут быть дефицитными по одному или нескольким продуктам экспрессии. В связи с этим следует принять во внимание, что в случае аденовирусов группы I и аденовирусов группы II такие дефицитные аденовирусы могут быть получены путем мутации или делеции нуклеиновой кислоты, кодирующей указанный продукт экспрессии, где такие мутации и делеции соответственно могут быть осуществлены либо полностью, либо до определенной степени, при которой этот продукт экспрессии больше не будет продуцироваться, либо эти дефицитные аденовирусы могут быть получены с помощью регуляторных элементов и элементов, регулирующих экспрессию, таких как промоторы и факторы транскрипции которые отсутствуют или активируются по механизму, отличающемуся от механизма активации дикого типа, либо на уровне нуклеиновых кислот (отсутствие промотора; цис-активные элементы), либо на уровне системы трансляции и системы транскрипции, соответственно (транс-активные элементы). В частности, последний аспект может зависеть от соответствующего теневого фенотипа клетки.
Помимо использования уже известных аденовирусов в соответствии с настоящим изобретением, могут быть также использованы новые аденовирусы, такие как аденовирусы группы II, в целях, которые были уже описаны здесь для других аденовирусов. Эти новые аденовирусы настоящего изобретения могут быть получены исходя из описанной здесь технической идеи. Особенно предпочтительными и репрезентативными являются, например, вирусы Xvir03 и Xvir03/01, показанные на фиг.16 и 17, а принцип их конструирования также проиллюстрирован в примерах 11 и 12.
В случае вектора Xvir03, промотор CMV клонируют в область Е1, под контролем которой находятся нуклеиновые кислоты для Е1В55К и Е4orf6, разделенные последовательностью IRES. Благодаря клонированию этих двух генов в вирус и благодаря продуцированию из них генных продуктов, соответственно достигается эффективная репликация, которая фактически соответствует репликации вирусов дикого типа, при этом селективная репликация сохраняется в клетках, а предпочтительно, в опухолевых клетках, поскольку такая репликация происходит, в частности, в клетках, позитивных по ядерному YВ-1, а более конкретно, в клетках, которые содержат YВ-1 с нарушенной регуляцией в соответствии с настоящим описанием. В одном из вариантов изобретения клетками, в которых присутствует YВ-1 с нарушенной регуляцией, являются такие клетки, которые обнаруживают повышенную экспрессию YВ-1, а предпочтительно, компартментнезависимую экспрессию YВ-1 по сравнению с нормальными или неопухолевыми клетками.
Другим вариантом вируса Xvir03 является вирус Xvir03/01, в который, в предпочтительном варианте изобретения, клонируют терапевтические гены или трансгены, находящиеся под контролем специфического промотора, а в частности опухолеспецифического или тканеспецифического промотора. Что касается такого вируса, то его область Е4 также является функционально неактивной, а предпочтительно делетированной. Описанные здесь трансгены могут быть также клонированы в область Е4, и такое клонирование может быть осуществлено либо альтернативно, либо в дополнение к клонированию трансгенов в область Е3.
Описанные здесь трансгены, а в частности трансгены, описанные ниже, могут быть также экспрессированы в сочетании с аденовирусами или с помощью аденовирусов настоящего изобретения, то есть аденовирусов группы I и их нуклеиновых кислот соответственно, или систем репликации настоящего изобретения и, таким образом, они входят в состав экспрессионной кассеты, содержащей промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты кодирует один или несколько указанных трансгенов. Области Е1, Е3 и/или Е4 являются особенно подходящими клонирующими сайтами в аденовирусном геноме, однако клонирующие сайты не ограничиваются указанными областями.
Указанными терапевтическими генами могут быть гены пролекарства, гены цитокинов, апоптоз-индуцирующие гены, гены-супрессоры опухоли, гены ингибиторов металлопротеиназ, и/или гены ингибиторов ангиогенеза и ингибиторов тирозинкиназы. Кроме того, могут быть экспрессированы киРНК, аптамеры, антисмысловые молекулы и рибозимы, которые, предпочтительно, направлены против ассоциированных с раком молекул-мишеней. Предпочтительно, указанные отдельные или множество молекул-мишеней выбраны из группы, включающей релевантные факторы резистентности, антиапоптозные факторы, онкогены, факторы ангиогенеза, ферменты ДНК-синтеза, ферменты репарации ДНК, факторы роста и их рецепторы, факторы транскрипции, металлопротеиназы, а в частности металлопротеиназы межклеточного матрикса и активатор плазминогена урокиназного типа. Предпочтительные их варианты уже описаны в настоящей заявке в соответствии с другими аспектами изобретения.
Подходящими генами пролекарства, которые могут быть использованы в предпочтительных вариантах осуществления изобретения, являются, например, гены цитозиндезаминазы, тимидинкиназы, карбоксипептидазы, урацилфосфорибозил-трансферазы или пурин-нуклеозид-фосфорилазы (PNP); [Kirn et al, Trends in Molecular Medicine, volume 8, no. 4 (suppl), 2002; Wybranietz W.A. et al., Gene Therapy, 8, 1654-1664, 2001; Niculescu-Duvaz et al., Curr. Opin. Mol. Therapy, 1, 480.486, 1999; Koyama et al., Cancer Gene Therapy, 7, 1015-1022, 2000; Rogers et al., Human Gene Therapy, 7, 2235-2245, 1996; Lockett et al., Clinical Cancer Res., 3, 2075-2080, 1997; Vijayakrishna et al., J. Pharmacol. And Exp. Therapeutics, 304, 1280-1284, 2003].
Подходящими цитокинами, которые могут быть использованы в предпочтительных вариантах осуществления изобретения, являются, например, GM-CSF, TNF-альфа, Il-12, Il-2, Il-6, CSF или интерферон-гамма; [Gene Therapy, Advances in Pharmacology, volume 40, editor: J. Thomas August, Academic Press; Zhang and Degroot, Endocrinology, 144, 1393-1398, 2003; Descamps et al., J. Mol. Med., 74, 183-189, 1996; Majumdar et al., Cancer Gene Therapy, 7, 1086-1099, 2000].
Подходящими апоптоз-индуцирующими генами, которые могут быть использованы в предпочтительных вариантах осуществления изобретения, являются, например, гены декорина [Tralhao et al., FASEB J., 17, 464-466, 2003]; ретинобластомы 94 [Zhang et al., Cancer Res. 63, 760-765, 2003]; Bax и Bad [Zhang et al., Hum. Gene Ther., 20, 2051-2064, 2002]; апоптин [Noteborn & Pietersen, Adv. Exp. Med. Biol., 465, 153-161, 2000]; ADP [Toth et al., Cancer Gene Therapy, 10, 193-200, 2003]; bcl-xs [Sumantran et al., Cancer Res, 55, 2507-2512, 1995]; Е4orf4 [Braithwaite & Russell, Apoptosis, 6, 359-370, 2001]; FasL, Apo-1 и Trail [Boehringer Manheim, Guide to Apoptotic Pathways, Arai et al., PNAC, 94, 13862-13867, 1997]; Bims [Yamaguchi et al., Gene Therapy, 10, 375-385, 2003]; GNR163 [Oncology News, 17 Junе, 2000].
Подходящими генами-супрессорами опухоли, которые могут быть использованы в предпочтительных вариантах осуществления изобретения, являются, например, E1A, p53, p16, p21, p27 или MDA-7 [Opalka et al., Cell Tissues Organs, 172, 126-132, 2002, Ji et al., Cancer Res., 59, 3333-3339, 1999, Su et al., Oncogene, 22, 1164-1180, 2003].
Подходящими ингибиторами ангиогенеза, которые могут быть использованы в предпочтительных вариантах осуществления изобретения, являются, например, эндостатин или ангиостатин [Hajitou et al., FASEB J. 16, 1802-1804, 2002], и антитела против VEGF [Ferrara N., Semin Oncol. 2002 Dec; 29 (6 Suppl 16): 10-4].
Подходящими ингибиторами металлопротеиназы, которые могут быть использованы в предпочтительных вариантах осуществления изобретения, являются, например, Timp-3 [Ahonen et al., Mol. Therapy, 5, 705-715, 2002]; PAI-1 [Soff et al., J. Clin. Invest., 96, 2593-2600, 1995]; Timp-1 [Brandt K. Curr. Gene Therapy, 2, 255-271, 2002].
Другими трансгенами настоящего изобретения, которые могут экспрессироваться аденовирусами группы I и аденовирусами группы II, также являются ингибиторы тирозинкиназы. Примерами тирозинкиназ являются EGFR (рецептор эпидермального фактора роста) [Onkologie, Entstehung und Progression maligner Tumoren; author: Christoph Wagner, Georg Thieme Verlag, Stuttgart, 1999]. Предпочтительным ингибитором тирозинкиназы является герцептин [Zhang H et al., Cancer Biol Ther., 2003, Jul-Aug; 2 (4 suppl 1): S122-6].
киРНК (короткая интерферирующая РНК) состоит из двух, а предпочтительно, из двух отдельных цепей РНК, которые гибридизуются друг с другом вследствие комплементарности оснований, то есть, это означает, что они состоят, в основном, из спаренных оснований и имеют длину, предпочтительно, до 50 нуклеотидов, более предпочтительно, от 18 до 30 нуклеотидов, а еще более предпочтительно, менее чем 25 нуклеотидов, и наиболее предпочтительно 21, 22 или 23 нуклеотида, где указанные числа относятся к одной цепи киРНК, а в частности означают длину фрагмента указанной одной цепи, которая гибридизуется с одной, а более конкретно, со второй одной цепью, и образует с ней пару оснований. киРНК специфически индуцирует или опосредует деградацию мРНК. Требуемая для этого специфичность обеспечивается последовательностью киРНК, а следовательно, ее сайтом связывания. Подвергаемая деградации последовательность-мишень в основном комплементарна первой или второй из указанных цепей, образующих киРНК. Хотя точный механизм ее действия пока неясен, однако предполагается, что киРНК участвует в биологическом механизме ингибирования отдельных аллелей в клетках в процессе их развития и самозащиты от вирусов. киРНК-опосредованная интерференция РНК используется как средство для специфической супрессии или даже полного подавления экспрессии белка путем введения геноспецифической двухцепочечной РНК. Для высших организмов киРНК, имеющая длину от 19 до 23 нуклеотидов, является особенно подходящей, поскольку она не приводит к активации неспецифической защитной реакции, такой как интерлейкиновый ответ. Непосредственная трансфекция двухцепочечной РНК, состоящей из 21 нуклеотидов и имеющей симметричные 2-нуклеотидные выступающие 3'-концы, может опосредовать интерференцию РНК в клетках млекопитающих и является в высокой степени эффективной по сравнению с молекулами, используемыми в других технологиях, например, с рибозимами и антисмысловыми молекулами (Elbashir S. Harborth J. Lendeckel W. Yalvcin A. Weber K. Tuschl T: Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411:494-498). Для ингибирования экспрессии нужного гена необходимы лишь очень небольшие молекулы киРНК. В предшествующих работах, во избежание ограничений для экзогенно вводимой киРНК, которые, в частности, накладываются временной природой феномена интерференции и специфической доставкой молекул киРНК, использовались векторы, которые обеспечивают экспрессию эндогенной киРНК. Для этого, например, олигонуклеотиды, имеющие длину в 64 нуклеотида, встраивают в вектор, содержащий нацеливающие последовательности длиной в 19 нуклеотидов, как в смысловой, так и в антисмысловой ориентации, разделенные, например, спейсерной последовательностью длиной в 9 нуклеотидов. Полученный транскрипт имел укладку в виде шпилечной структуры, включающей структуру типа “стебель”, состоящую, например, из 19 пар нуклеотидов. Петля этой структуры быстро деградирует в клетке, в результате чего образуется функциональная молекула киРНК (Brummelkamp et al., Science, 296, 550-553, 2002).
Нуклеиновая кислота, кодирующая YВ-1, которая может быть частью аденовирусов одного из вариантов аденовирусов, используемых в настоящем изобретении, а в частности аденовирусов группы II, а также аденовирусов настоящего изобретения, то есть аденовирусов группы I, может содержать последовательность нуклеиновой кислоты, которая опосредует транспорт YВ-1 в ядро. Нуклеиновые кислоты, аденовирусы и аденовирусные системы настоящего изобретения, а также уже известные аденовирусы, такие как, например, Onyx-015, AdΔ24, dl922-947, E1Ad/01/07, CB016, dl520 и аденовирусы, описанные в патенте ЕР 0931830, могут быть использованы как аденовирусы и аденовирусные системы соответственно и соответствующие нуклеиновые кислоты в комбинации с этими нуклеиновыми кислотами в соответствии с настоящим изобретением. Подходящие последовательности нуклеиновых кислот, опосредующих транспорт в ядро, известны специалистам и описаны, например, в работах Whittaker G.R. et al., Virology; 246, 1-23, 1998; Friedberg E.C., TIBS 17, 347, 1992; Jans D.A. et al., Bioassays 2000 Jun; 22(6): 532-44; Yoneda Y., J. Biochem (Tokyo) 1997 May; 121(5): 811-7; Boulikas, T., Crit. Rev. Eukariot. Gene Expr. 1993; 3(3): 193-227; Lyons R.H., Mol. Cell. Biol., 7, 2451-2456, 1987. Последовательности нуклеиновой кислоты, опосредующие транспорт в ядро, могут быть использованы для реализации различных принципов. Один из таких принципов заключается, например, в том, что YВ-1 образует гибридный белок вместе с сигнальным пептидом, или он доставляется таким сигнальным пептидом и транспортируется в клеточное ядро благодаря этому сигнальному пептиду, после чего происходит репликация аденовирусов настоящего изобретения.
Другой принцип, который может быть реализован при конструировании аденовирусов, используемых в соответствии с настоящим изобретением, а в частности аденовирусов группы II, а также аденовирусов настоящего изобретения, то есть аденовирусов группы I, заключается в том, что YВ-1 может быть доставлен транспортной последовательностью, которая после синтеза, предпочтительно, в цитоплазме, переносит или транспортирует YВ-1 в клеточное ядро и стимулирует репликацию вируса. Примером особенно эффективной последовательности нуклеиновой кислоты, опосредующей транспорт в ядро, является последовательность ТАТ ВИЧ, которая, среди прочих других подходящих последовательностей нуклеиновой кислоты такого типа, описана в работе Efthymiadis A. Briggs, L.J, Jans, D.A., JBC 273, 1623-1628, 1998. В настоящем изобретении описаны аденовирусы, используемые в соответствии с настоящим изобретением, а предпочтительно, аденовирусы группы II, а также аденовирусы настоящего изобретения, то есть аденовирус группы I, и содержащие последовательности нуклеиновой кислоты, кодирующие пептиды, опосредующие транспорт в ядро.
В настоящем изобретении предусматривается, что YВ-1 присутствует в своей полноразмерной форме, а в частности в форме, которая соответствует YВ-1 дикого типа. Кроме того, в настоящем изобретении предусматривается, что YВ-1 используется или присутствует в виде производного, например, в укороченной или в усеченной форме. В соответствии с настоящим изобретением указанное используемое или присутствующее производное YВ-1 представляет собой YВ-1, способный связываться с Е2-поздним промотором, и таким образом активировать экспрессию гена в области Е2 аденовируса. Такие производные, в частности, включают описанные здесь производные YВ-1. Другие производные могут быть получены путем делеции одной или нескольких аминокислот у N-конца, у С-конца или внутри аминокислотной последовательности. В настоящем изобретении также предусматривается, что фрагменты YВ-1 могут быть также использованы в качестве белков YВ-1, определенных в настоящем изобретении. В работе Jrchott [JBC 2003, 278, 27988-27996] описаны различные фрагменты YВ-1, характеризующиеся делециями у С- и N-конца. Распределение различных фрагментов YВ-1 показало, что домен холодового шока (CSD), а также С-конец являются подходящими для регулируемого клеточным циклом транспорта YВ-1 в клеточное ядро. Таким образом, в настоящем изобретении предусматривается, что укороченный YВ-1 (также называемый далее белком YВ-1), в случае экспрессии Е1В55k и Е4orf6, осуществляемой в соответствии с настоящим изобретением, лучше мигрирует в ядро, а поэтому индуцирует более сильный СРЕ, для которого не требуется лучшего связывания с Е2-поздним промотором, по сравнению с нативным YВ-1, а поэтому также нельзя исключить, что укороченный YВ-1 будет лучше мигрировать в ядро и стимулировать оба эффекта, то есть индуцирование СРЕ и связывание с Е2-поздним промотором. И, наконец, такие укороченные фрагменты YВ-1 могут также лучше мигрировать в ядро и более эффективно связываться с Е2-поздним промотором, не индуцируя при этом более сильного СРЕ. В настоящем изобретении также предусматривается, что укороченные белки YВ-1 и их фрагменты, соответственно, содержат другие последовательности, описанные здесь для полноразмерного YВ-1, а в частности сигнальные последовательности, ответственные за клеточную локализацию (NLS) и т.п.
Что касается вышеупомянутых других различных генов и генных продуктов, кодируемых и экспрессируемых аденовирусами соответственно, то в принципе они могут также кодироваться и экспрессироваться соответственно в любой комбинации.
В описании настоящего изобретения термины “аденовирус” и “аденовирусная система” имеют, в основном, одинаковый смысл. Термин “аденовирус”, в частности, означает полную вирусную частицу, содержащую капсид и нуклеиновую кислоту. Термин “аденовирусная система” имеет, в частности, тот смысл, что указанная нуклеиновая кислота, в отличие от нуклеиновой кислоты дикого типа, является модифицированной. Такие модификации, предпочтительно, включают изменения в структуре генома аденовируса, которые могут возникать в результате делеции, и/или добавления, и/или мутации промоторов, регуляторных последовательностей и/или кодирующих последовательностей, таких как рамки считывания. Кроме того, более предпочтительно термин “аденовирусная система” используется в сочетании с вектором, который может применяться, например, в генотерапии.
Приведенные выше комментарии, включая любое использование, а также конструирование аденовирусов и аденовирусных систем соответственно, могут также относиться и к кодирующим их нуклеиновым кислотам, и наоборот.
В соответствии с настоящим изобретением предусматривается, что аденовирусы, используемые в настоящем изобретении, а в частности аденовирусы группы II, а также аденовирусы группы I и кодирующие их нуклеиновые кислоты, представляют собой любую соответствующую аденовирусную нуклеиновую кислоту, которая, взятая отдельно или в комбинации с другими последовательностями нуклеиновой кислоты, инициирует событие репликации. Как указывается в настоящем описании, для репликации описанных последовательностей и/или генных продуктов может потребоваться вирус-помощник. Это до некоторой степени относится и к кодирующим последовательностям нуклеиновой кислоты, и такими последовательностями нуклеиновой кислоты являются известные последовательности нуклеиновой кислоты, а поэтому в настоящем изобретении предусматривается использование не только идентичных последовательностей, но также и отличающихся от них последовательностей. Используемый здесь термин “производные последовательности”, в частности, означает любые последовательности, которые продуцируют генный продукт, либо нуклеиновую кислоту, либо полипептид, обладающие функцией, соответствующей одной из функций непроизводной последовательности. Это может быть определено путем проведения рутинных тестов, известных специалистам. Примерами таких производных последовательностей нуклеиновой кислоты являются последовательности нуклеиновой кислоты, которые кодируют тот же самый генный продукт, а в частности, ту же самую аминокислотную последовательность, но которые имеют при этом отличающиеся последовательности оснований, что обусловлено вырожденностью генетического кода.
Что касается аденовирусов, применяемых в соответствии с настоящим изобретением, то есть аденовирусов группы II, и/или соответствующих систем репликации аденовирусов настоящего изобретения и их применения в соответствии с настоящим изобретением соответственно, то в одном из вариантов осуществления изобретения указанная аденовирусная нуклеиновая кислота является дефицитной по экспрессии онкогенного белка, а в частности дефицитной по белку Е1А, что означает, что она либо не кодирует белок 12S Е1А (также обозначаемый здесь Е1А12S), либо не кодирует белок 13S Е1А (также обозначаемый здесь Е1А13S), либо не кодирует ни белок 12S Е1А, ни белок 13S Е1А, либо она является модифицированной, как определено в настоящем описании, если не указано обратное; а указанная система репликации аденовируса, кроме того, содержит нуклеиновую кислоту вируса-помощника, где указанная нуклеиновая кислота вируса-помощника содержит последовательность нуклеиновой кислоты, кодирующую онкогенный белок, а в частности белок Е1А, который имеет нижеуказанные свойства и наделяет этими свойствами указанный аденовирус соответственно, а именно он, предпочтительно, не реплицируется в клетках, негативных по ядерному YВ-1, но реплицируется в позитивных по ядерному YВ-1 клетках, независимо от их клеточного цикла, или в клетках, содержащих YВ-1 с нарушенной регуляцией; он трансактивирует, по меньшей мере, один вирусный ген, а в частности Е1В55kDa, E4orf6, E4orf3 и/или E3ADP, в позитивных по ядерному YВ-1 клетках, и/или не транспортирует клеточный YВ-1 в ядро. В настоящем изобретении предусматривается, что описанные здесь трансгены кодируются отдельно или в комбинации друг с другом указанным вирусом-помощником и/или экспрессируются в нем. Это относится к вирусам-помощникам как для аденовирусов группы I, так и для аденовирусов группы II.
Кроме того, в одном из вариантов такой аденовирусной системы репликации настоящего изобретения указанная аденовирусная нуклеиновая кислота и/или нуклеиновая кислота вируса-помощника присутствует(ют) в качестве векторов, способных к репликации.
В настоящем изобретении также предусматривается, что нуклеиновая(ые) кислота(ы), кодирующая(ие) аденовирусы группы I и/или аденовирусы группы II, предпочтительно присутствует(ют) в экспрессирующем векторе, и что этот экспрессирующий вектор используется в настоящем изобретении.
В другом своем аспекте настоящее изобретение также относится к группе векторов, включающей, по меньшей мере, два вектора, где указанная группа векторов содержит, в целом, описанную здесь аденовирусную систему репликации для аденовирусов группы I, и/или для аденовирусов группы II, описанных в настоящей заявке, и где указанная группа векторов используется в соответствии с настоящим изобретением. В одном из вариантов изобретения каждый компонент аденовирусной системы репликации расположен на отдельном векторе, а предпочтительно, на экспрессирующем векторе.
И, наконец, в своем другом аспекте настоящее изобретение относится к применению клетки, которая содержит одну или несколько нуклеиновых кислот, кодирующих описанные здесь аденовирусы группы I и/или аденовирусы группы II, которые, предпочтительно, используются в настоящем изобретении и которые предназначены для использования в соответствии с настоящим изобретением, и/или соответствующую аденовирусную систему репликации, и/или соответствующий вектор, и/или группу векторов настоящего изобретения, где указанное применение предназначено почти для таких же целей, как и описанные здесь различные аденовирусы.
Описанные выше конструкции аденовирусов, а в частности их нуклеиновых кислот и нуклеиновых кислот, кодирующих эти аденовирусы, могут быть также введены в клетку, а в частности в опухолевую клетку по частям в виде множества компонентов, где благодаря присутствию этих различных отдельных компонентов они могут действовать аддитивно, так, как действовали бы отдельные компоненты, происходящие от одной нуклеиновой кислоты и одного или нескольких аденовирусов соответственно.
Нуклеиновые кислоты, которые используются в соответствии с настоящим изобретением и которые кодируют аденовирусы группы I и/или аденовирусы группы II, а также соответствующие аденовирусные системы или их части, могут также присутствовать в качестве векторов. При этом предпочтительными являются вирусные векторы. В случае, если нуклеиновые кислоты включают аденовирусные нуклеиновые кислоты, то вирусной частицей является вектор. Однако в настоящем изобретении указанные нуклеиновые кислоты присутствуют в плазмидном векторе. В любом случае вектор содержит элементы, обеспечивающие и регулирующие размножение встроенной нуклеиновой кислоты, то есть репликацию и, необязательно, экспрессию встроенной нуклеиновой кислоты. Подходящие векторы, а предпочтительно, экспрессирующие векторы и их соответствующие элементы известны специалистам и описаны, например, в работе Grunhaus A., Horwitz, M.S., 1994, Adenoviruses as cloning vectors. In Rice C., editor, Seminars in Virology, London: Saunders Scientific Publications.
В одном из своих аспектов настоящее изобретение относится к группе векторов, описанных в вышеуказанном варианте изобретения, где указанные различные элементы нуклеиновой кислоты необязательно находятся только на одном векторе. В соответствии с этим, указанная группа векторов состоит, по меньшей мере, из двух векторов. Так или иначе, вся информация, касающаяся указанных векторов, может быть также применена как к векторам, так и к группе векторов соответственно.
Аденовирусы группы I и/или аденовирус группы II характеризуются описанными здесь нуклеиновыми кислотами и генными продуктами соответственно и могут, так или иначе, содержать все указанные элементы, известные специалистам, которые присутствуют в аденовирусах дикого типа (Shenk T.: Adenoviridae: The virus and their replication. Fields Virology, 3rd edition, edit. Fields B.N., Knipe, D.M., Howley, P.M. et al., Lippincott-Raven Publishers, Philadelphia, 1996, chapter 67).
Краткое обсуждение репликации аденовирусов приводится ниже лишь в целях иллюстрации, но не в целях ограничения настоящего изобретения.
Репликация аденовирусов представляет собой очень сложный процесс и обычно в нем участвует человеческий фактор транскрипции Е2F. Во время инфицирования вирусом сначала экспрессируются “ранние гены” Е1, Е2, Е3 и Е4. Группа “поздних генов” ответственна за синтез вирусных структурных белков. Для активации как ранних, так и поздних генов, область Е1, состоящая из двух транскрипционных единиц Е1А и Е1В, которые кодируют различные белки Е1А и Е1В, играет решающую роль, поскольку указанные транскрипционные единицы индуцируют транскрипцию генов Е2, Е3 и Е4 (Nevins J.R. Cell 26, 213-220, 1981). Кроме того, белки Е1А могут инициировать синтез ДНК в покоящихся клетках, и, тем самым, стимулировать их вступление в фазу S (см. Boulanger and Blair, 1991). Кроме того, они взаимодействуют с супрессорами опухолей класса Rb (Whyte P. et al., Nature 334, 124-127, 1988). В этом случае высвобождается клеточный фактор транскрипции Е2F. Затем факторы Е2F могут связываться с соответствующими промоторными областями как клеточных, так и вирусных генов (в частности, с аденовирусным ранним промотором Е2) и тем самым инициировать транскрипцию и репликацию (Nevins J.R. Science 258, 424-429, 1992). Активность рRb и Е2F регулируется посредством фосфорилирования. Гипофосфорилированная форма рRb, в частности, присутствует в фазах G1 и М. В противоположность этому гиперфосфорилированная форма рRb присутствует в фазах S и G2. Е2F высвобождается из комплекса, состоящего из Е2F и гипофосфорилированного рRb, путем фосфорилирования рRb. Высвобождение Е2F из комплекса, состоящего из Е2F и гипофосфорилированного рRb, приводит к транскрипции Е2F-зависимых генов. Белок Е1А связывается только с гипофосфорилированной формой рRb, но при этом связывание Е1А с рRb происходит, главным образом, через область CR2 белка Е1А. Кроме того, он также связывается с областью CR1, хотя и с более низкой аффинностью (Ben-Israel and Kleiberger, Frontiers in Bioscience, 7, 1369-1395, 2002; Helt and Galloway, Carcinogenesis, 24, 159-169, 2003).
Генные продукты области Е2 особенно необходимы для инициации и осуществления репликации, поскольку они кодируют три главных белка. Транскрипция белков Е2 регулируется двумя промоторами, “Е2-ранним Е2F-зависимым промотором”, который также называется здесь “Е2-ранним промотором” или “ранним промотором Е2”, и “Е2-поздним промотором” (Swaminathan and Thimmapaya, The Molecular Repertoire of Adenoviruses III: Current Topics in Microbiology and Immunology, Vol.199, 177-194, Springer Verlag 1995). Кроме того, продукты области Е4 вместе с белком Е1А и Е1В-55kDa играют решающую роль для активности Е2F и стабильности р53. Так, например, указанный промотор Е2 даже в большей степени трансактивируется посредством прямого взаимодействия белка E4orf6/7, кодируемого областью Е4, с гетеродимером, состоящим из Е2F и DР1 (Swaminathan and Thimmapaya, JBC 258, 736-746, 1996). Кроме того, комплекс, состоящий из Е1В-55kDa и Е4orf6, инактивируется р53 (Steegenga W.T. et al., Oncogene 16, 349-357, 1998), в результате чего литический инфекционный цикл успешно завершается. Более того, белок Е1В-55kDa, поскольку он стимулируется путем взаимодействия с белком Е4orf6, обладает другой важной функцией, а именно экспортом вирусной РНК из ядра клетки, при этом клеточные РНК остаются в ядре (Bridge and Ketner, Virology 174, 345-353, 1990). Другим важным наблюдением явилось то, что указанный белковый комплекс, состоящий из Е1В-55kDa/Е4orf6, локализован в так называемых “вирусных тельцах включения”. Было высказано предположение, что эти структуры представляют собой сайты репликации и транскрипции (Ornelles and Shenk, J. Virology 65, 424-429, 1991).
Другой областью, которая играет важную роль в репликации, а в частности в высвобождении аденовирусов, является область Е3. Более конкретно, область Е3 содержит генетическую информацию для различных сравнительно небольших белков, которые не имеют важного значения для инфекционного цикла аденовируса in vitro, то есть они не играют важной роли в клеточной культуре. Однако эти белки имеют решающее значение для выживания вируса во время острой и/или латентной инфекции in vivo, поскольку они обладают, среди прочих, иммунорегуляторными и апоптотическими функциями (Marshall S. Horwitz, Virololgie, 279, 1-8, 2001; Russell, см. выше). Было показано, что белок, имеющий размер примерно 11,6 кДа, индуцирует клеточную гибель. Такой белок, благодаря своей функции, называется АDP то есть английским термином “белок гибели аденовируса” (Tollefson J. Virology, 70, 2296-2306, 1996). Этот белок образуется преимущественно в поздней фазе инфекционного цикла. Кроме того, сверхэкспрессия указанного белка приводит к более эффективному лизису инфекционных клеток (Doronin et al., J. Virology, 74, 6147-6155, 2000).
Кроме того, автору настоящего изобретения известно, что Е1А-делетированные вирусы, а в частности вирусы, которые не содержат белок 12S Е1А, а также не экспрессируют белок 13S Е1А, могут весьма эффективно реплицироваться при более высоких m.o.i. (Nevins J.R., Cell 26, 213-220, 1981), однако, эти вирусы не могут быть использованы в каких-либо клинических целях. В литературе такой феномен известен как “Е1А-подобная активность”. Также известно, что из 5 белков, кодируемых Е1А, два белка, а именно белки 12S и 13S, регулируют и индуцируют соответственно, экспрессию других аденовирусных генов (Nevins, J.R. Cell 26, 213-220, 1981; Boulanger P. and Blair E.; Biochem. J. 275, 281-299, 1991). В соответствии с этим было показано, что трансактивирующая функция преимущественно обеспечивается CR3-областью белка 13S (Wong H.K. and Ziff E.B., J. Virol., 68, 4910-20, 1994). Аденовирусы, которые имеют значительные делеции в CR1-области и/или CR2 и/или в CR3-области белка 13S, являются, в основном, дефектными по репликации, однако, они все же трансактивируют вирусные гены и промоторы в других клеточных линиях, а в частности в области Е2 (Wong H.K. and Ziff E.B. J. Virol. 68, 4910-20, 1994; Mymryk J.S. and Bayley S.T., Virus Research 33, 89-97, 1994).
После инфицирования клетки, обычно опухолевой клетки, аденовирусом дикого типа происходит индуцирование транспорта YВ-1 в ядро, которое опосредуется Е1А, Е1В-55kDa и Е4orf6, и YВ-1 локализуется вместе с Е1В-55kDa в вирусных тельцах включения в ядре, что позволяет обеспечивать эффективную репликацию вируса в клеточном ядре как in vitro, так и in vivo. Ранее было уже обнаружено, что Е4orf6 связывается с Е1В-55К (Wiegel S. and Dobblestein, M.J. Virology, 74, 764-772, 2000; Keith N. Leppard, Seminars in Virology, 8, 301-307, 1998) и таким образом опосредует транспорт и распределение Е1В-55kDa в ядре, что обеспечивает оптимальное продуцирование вируса и репликацию аденовируса соответственно. Эффективная репликация вируса в соответствии с настоящем изобретением возможна благодаря взаимодействию Е1А, Е1В-55К и YВ-1 и комплекса, состоящего из Е1В-55К/Е4orf6, с YВ-1 соответственно и совместной локализации YВ-1 и Е1В-55kDa в ядре в так называемых вирусных тельцах включения, а поэтому описанные здесь вирусы могут быть использованы для репликации в клетках, позитивных по ядерному YВ-1, а предпочтительно, в клетках, которые содержат YВ-1 в ядре независимо от клеточного цикла, и/или в клетках, которые содержат или обнаруживают YВ-1 с нарушенной регуляцией, и/или они могут быть использованы в целях изготовления лекарственного препарата соответственно для лечения заболеваний, в которых участвуют клетки, позитивные по ядерному YВ-1, предпочтительно, клетки, которые содержат YВ-1 в ядре независимо от клеточного цикла, и/или клетки, которые содержат или обнаруживают YВ-1 с нарушенной регуляцией. Таким образом, репликация, которая может быть осуществлена в данном клеточном теневом фенотипе, приводит к лизису клеток, к высвобождению вируса и к инфицированию и лизису соседних клеток, а в случае инфицирования опухолевой клетки и опухоли соответственно, такая репликация, в конечном счете, приводит к лизису опухоли, т.е. к онколизису.
YВ-1 принадлежит к группе в высокой степени консервативных факторов, которые связываются с инвертированной последовательностью СААТ, то есть с так называемым Y-боксом. Они могут быть активными в регуляции как на уровне транскрипции, так и на уровне трансляции (Wolffe A.P. Trends in Cell Biology 8, 318-323, 1998). Растущее число Y-бокс-зависимых путей регуляции было обнаружено при активации, а также при ингибировании генов, ассоциированных с ростом и апоптозом клеток (Swamynathan S.K. et al., FASEB J. 12, 515-522, 1998). Так, например, YВ-1 непосредственно взаимодействует с р53 (Okamoto T. et al., Oncogene 19, 6194-6202, 2000) и играет важную роль в экспрессии гена Fas (Lasham A. et al., Gene 252, 1-13, 2000), в экспрессии генов MDR и MRP (Stein U. et al., JBC 276, 28562-69, 2001; Bargou R.C. et al., Nature Medicine 3, 447-450, 1997) и в активации топоизомераз и металлопротеиназ (Mertens P.R. et al., JBC 272, 22905-22912, 1997; Shibao K. et al., Int. J. Cancer 83, 732-737, 1999). Кроме того, YВ-1 участвует в регуляции стабильности мРНК (Chen C.Y. et al., Genes and Development 14, 1236-1248, 2000) и в процессах репарации (Ohga T. et al., Cancer Res. 56, 4224-4228, 1996; Izumi H. et al., Nucleic Acid Research 2001, 29, 1200-1207; Ise T. et al., Cancer Res., 1999, 59, 342-346).
Ядерная локализация YВ-1 в опухолевых клетках, либо присутствие YВ-1, в ядре независимо от клеточного цикла, либо транслокация YВ-1 с нарушенной регуляцией, присутствующего в цитоплазме, в ядро под действием аденовирусов группы I и/или аденовирусами группы II, приводит к Е1А-независимой репликации вируса, где, в частности, ни белок 12S Е1А, ни белок 13S Е1А не присутствуют и не используются в экспрессированной форме соответственно (Holm P.S. et al., JBC 277, 10427-10434, 2002), а в случае сверхэкспрессии белка YВ-1 она приводит к резистентности ко многим лекарственными препаратам. Кроме того, известно, что аденовирусные белки, такие как, например, Е4orf6 и Е1В-55kDa оказывают положительное действие на репликацию вируса (Goodrum F.D. and Ornelles D.A. J. Virology 73, 7474-7488, 1999), а функциональный белок Е1А ответственен за активацию других продуктов вирусных генов (таких как E4orf6, E3ADP и E1B-55K)(Nevins J.R., Cell 26, 213-220, 1981). Однако этого не происходит в случае применения Е1А-минус-аденовирусов, известных специалистам, в которых белок 13S Е1А отсутствует. Ядерная локализация YВ-1 в полирезистентных клетках, которые содержат YВ-1 в своем ядре, обеспечивает репликацию и образование частиц таких Е1А-минус-вирусов соответственно. Однако в данном случае эффективность репликации вируса и образования частиц снижается в несколько раз по сравнению с вирусами дикого типа Аd5. В отличие от этого комбинация YВ-1 обеспечивает в высокой степени эффективную YВ-1-опосредуемую репликацию и образование частиц вируса, и, тем самым, онколизис, где YВ-1 либо уже содержится в ядре опухолевой клетки, что может быть результатом локализации YВ-1 в ядре независимо от клеточного цикла, или где YВ-1 с нарушенной регуляцией, присутствующий в цитоплазме, транспортируется в ядро аденовирусами группы I и/или аденовирусами группы II, или его транспорт в клеточное ядро индуцируется экзогенными факторами (например, применением цитостатических средств, облучением или гипертермией), то есть факторами, индуцирующими его присутствие в ядре, в частности независимо от клеточного цикла, или где YВ-1 вводится в качестве трансгена с помощью вектора вместе с системой, предпочтительно, с аденовирусной системой, которая “запускает” аденовирусные гены, но при этом не наблюдается репликации вируса. Это может быть применено также и к аденовирусам настоящего изобретения, аденовирусам группы I, которые способны эффективно реплицироваться благодаря их специфической конструкции и действию, которое оказывает белок Е1В, предпочтительно, белок Е1В55К, и/или белок Е4, а предпочтительно, белок Е4orf6, на эффективную мобилизацию YВ-1, предпочтительно, в ядро. Подходящими цитостатическими средствами, которые могут быть использованы вместе с описанными здесь аденовирусами в соответствии с различными аспектами настоящего изобретения, являются цитостатические средства, принадлежащие к следующим группам, а именно: антрациклины, такие как, например, дауномицин и адриамицин; алкилирующие агенты, такие как, например, циклофосфамид; алкалоиды, такие как этопозид; виналкалоиды, такие как, например, винкристин и винбластин; антиметаболиты, такие как, например, 5-фторурацил и метотрексат; производные платины, такие как, например, цисплатин; ингибиторы топоизомеразы, такие как, например, камфотецин, СРТ-11; таксаны, такие как, например, таксол, паклитаксел, ингибиторы гистон-деацетилазы, такие как, например, FR901228, MS-27-275, трихостатин А, модуляторы MDR, такие как, например, MS-209, VX-710 и производные гелданамицина, такие как, например, 17-ААG. Описанные здесь аденовирусы, а в частности рекомбинантные аденовирусы, которые способны реплицироваться лишь в клетках, позитивных по ядерному YВ-1, и в клетках, которые содержат регулируемый YВ-1, предпочтительно, в цитоплазме, ограничены в отношении своей способности к трансактивации вирусных генов Е1В55kDa, E4orf6, E4orf3 и E3ADP, по сравнению с соответствующей трансактивирующей способностью аденовирусов дикого типа, а в частности Аd5 дикого типа. Автором настоящего изобретения было неожиданно обнаружено, что такая ограниченная трансактивирующая способность может быть компенсирована за счет экспрессии соответствующих генов, а в частности Е1В55К и Е4orf6 в сочетании с ядерной локализацией YВ-1. Как будет показано в нижеследующих примерах, в таких условиях степень репликации вируса и образования частиц соответственно возрастают до уровня, сравнимого с уровнем репликативной активности и активности в образовании частиц аденовирусов дикого типа соответственно.
Лекарственный препарат, в комбинации с которым или для изготовления которого применяются описанные здесь аденовирусы настоящего изобретения, обычно вводят системно, хотя в настоящем изобретении предусматривается и местное применение или локальная доставка указанного лекарственного препарата. Такое введение осуществляют в целях инфицирования клеток аденовирусом и в целях стимуляции репликации аденовируса именно в тех клетках, которые индуцируют, предпочтительно этиологически, развитие данного состояния, обычно заболевания, где для его диагностики и/или предупреждения и/или лечения применяется лекарственный препарат настоящего изобретения.
Такой лекарственный препарат, предпочтительно, используют для лечения злокачественных заболеваний, опухолевых заболеваний, раковых заболеваний, рака и опухолей, где используемые здесь термины являются синонимами, если это не оговорено особо. Опухолевыми заболеваниями, предпочтительно, являются заболевания, при которых YВ-1 уже присутствует в клеточном ядре в соответствии с механизмом, лежащим в основе данного опухолевого заболевания, а в частности, патологическим механизмом, предпочтительно, независимым от клеточного цикла; либо заболевания, при которых присутствие YВ-1 в клеточном ядре обусловлено экзогенными факторами, где указанные факторы благоприятствуют переносу YВ-1 в клеточное ядро или его индуцированию или экспрессии в ядре. Используемые здесь термины “опухоль” или “опухолевое заболевание” означают как злокачественные, так и доброкачественные опухоли, а также солидные и диффузные опухоли и ассоциированные с ними заболевания. В одном из вариантов изобретения указанный лекарственный препарат содержит, по меньшей мере, одно дополнительное фармацевтически активное соединение. Природа и количество такого дополнительного фармацевтически активного соединения будет зависеть от показаний к его применению. В случае, когда данный лекарственный препарат применяется для лечения и/или предупреждения опухолевых заболеваний, обычно используются цитостатические средства, такие как, например, цисплатин, таксол, даунобластин, даунорубицин, адриамицин и/или митоксантрон, или другие цитостатические средства или группы цитостатических средств, описанные в настоящей заявке, а предпочтительно, описанные в связи с опосредуемой цитостатическими средствами ядерной локализацией YВ-1.
Лекарственный препарат настоящего изобретения может присутствовать в виде различных композиций, а предпочтительно, в жидкой форме. Кроме того, указанный лекарственный препарат может содержать стабилизаторы, буферы, консерванты и т.п., которые обычно применяются в фармацевтической практике.
Автором настоящего изобретения было неожиданно обнаружено, что описанные в настоящем изобретении вирусы, а предпочтительно аденовирусы группы I и/или аденовирусы группы II могут с большим успехом применяться для лечения указанных опухолей, и они могут быть также использованы в целях изготовления лекарственных препаратов для лечения тех опухолей, которые содержат YВ-1 в клеточном ядре независимо от клеточного цикла. Обычно YВ-1 находится в цитоплазме, а в частности в перинуклеарной плазме. Во время G1/S-фазы клеточного цикла YВ-1 может присутствовать в ядре как нормальных, так и опухолевых клеток, где часть YВ-1 остается в цитоплазме [Jürchott K et al., JBC 2003, 278, 27988-27996]. Однако этого недостаточно, чтобы обеспечить вирусный онколизис с применением модифицированных таким образом аденовирусов. Сравнительно низкая эффективность таких аттенюированных аденовирусов, описанных в предшествующих работах, в конечном счете, обусловлена их неправильным применением. Другими словами, такие аденовирусные системы могут быть использованы, в частности, с более высокой эффективностью в том случае, если при введении описанных здесь аттенюированных или модифицированных вирусов, а предпочтительно, аденовирусов группы I и/или аденовирусов группы II, создаются молекулярно-биологические условия, необходимые для вирусного онколизиса. В случае описанных здесь аденовирусов, применяемых в соответствии с настоящим изобретением, таких как AdΔ24, dl922-947, E1Ad/01/07, CB016, dl520, и рекомбинантных аденовирусов, описанных в Европейском патенте ЕР 0931830, такие необходимые условия создаются в таких опухолях, клетки которых содержат локализованный в ядре YВ-1 независимо от клеточного цикла. Такой тип ядерной локализации может быть обусловлен самой природой опухоли, либо факторами или агентами, описанными в настоящем изобретении. Таким образом, в настоящем изобретении определена новая группа опухолей и опухолевых заболеваний, а также пациенты, которые могут быть подвергнуты успешному лечению с применением вирусов настоящего изобретения, а предпочтительно, аденовирусов группы I и/или аденовирусов группы II, а также аттенюированных или модифицированных аденовирусов, уже описанных в литературе.
Другой группой пациентов, которые могут быть подвергнуты лечению в соответствии с настоящим изобретением с применением аденовирусов группы I и/или аденовирусов группы II или аденовирусов, применяемых в соответствии с настоящим изобретением, которые уже были описаны в литературе, или с применением аденовирусов, описанных здесь впервые, а предпочтительно, с применением таких аденовирусов, которые имеют мутации и делеции соответственно в белке Е1А, не препятствующие связыванию Rb/Е2f, но которые не реплицируются в негативных по ядерному YВ-1 клетках, или которые имеют очень низкий уровень репликации, как определено в данном описании, и/или которые имеют и/или обнаруживают делетированный онкопротеин, а в частности Е1А, например, в случае вирусов AdΔ24, dl922-947, E1Ad/01/07, CB016 и аденовирусов, описанных в европейском патенте ЕР 0931830, являются такие пациенты, у которых при создании и реализации определенных условий будет осуществляться миграция YВ-1 в клеточное ядро или его индукция в клеточном ядре или его перенос в клеточное ядро, либо присутствие YВ-1 с нарушенной регуляцией. Применение аденовирусов группы I и/или аденовирусов группы II для пациентов этой группы основано на обнаружении того факта, что индуцирование репликации вируса основано на ядерной локализации YВ-1 с последующим связыванием YВ-1 с Е2-поздним промотором. На основании описанных здесь данных был сделан вывод, что описанные здесь аденовирусы, такие как AdΔ24, dl922-947, E1Ad/01/07, CB016 и/или аденовирусы, описанные в европейском патенте ЕР 0931830, могут также реплицироваться в тех клетках, которые являются позитивными по ядерному YВ-1, и/или в клетках, которые содержат YВ-1 с нарушенной регуляцией, как определено в настоящем изобретении. Таким образом, эти аденовирусы могут быть использованы для лечения заболеваний и групп пациентов в соответствии с настоящим изобретением, имеющих клетки с указанными признаками, а в частности, если эти клетки участвуют в развитии соответствующего заболевания, подвергаемого лечению. Наличие указанных признаков является основой для успешного применения аденовирусов AdΔ24, dl922-947, E1Ad/01/07, CB016 и аденовирусов, описанных в европейском патенте ЕР 0931830, и аденовирусов группы I и/или аденовирусов группы II для лечения, в соответствии с настоящим изобретением, тех опухолей, которые содержат YВ-1 в ядре независимо от их клеточного цикла, или которые содержат YВ-1 с нарушенной регуляцией, как определено в настоящем описании. Другой группой пациентов, которые могут быть подвергнуты лечению в соответствии с настоящим изобретением с применением вирусов, которые описаны здесь как подходящие для применения в настоящем изобретении, и с применением вирусов, которые описаны здесь впервые, а в частности аденовирусов группы I и/или аденовирусов группы II соответственно являются пациенты, имеющие клетки, позитивные по ядерному YВ-1, и/или клетки, которые стали позитивными по ядерному YВ-1 в результате лечения, описанного ниже; и/или пациенты, которые были подвергнуты такому лечению, а именно лечению, проводимому до введения аденовирусов, одновременно с введением соответствующих вирусов или после введения аденовирусов. В настоящем изобретении предусматривается, что пациентами, имеющими клетки, позитивные по ядерному YВ-1, являются пациенты, которые, в частности, содержат различные опухолеобразующие клетки, имеющие YВ-1 в ядре независимо от их клеточного цикла, и/или пациенты, у которых в этих клетках имеется YВ-1 с нарушенной регуляцией. Одним из таких методов лечения является введение цитостатических средств, описанных в общих чертах в настоящей заявке и/или применяемых в комбинации с противоопухолевой терапией. Кроме того, к такой группе методов лечения принадлежит облучение, а предпочтительно, облучение, используемое в противоопухолевой терапии. Термин “облучение” означает, в частности, облучение лучами высокой энергии, а предпочтительно, радиоактивное облучение, предпочтительно, используемое в противоопухолевой терапии. Другими методами лечения являются гипертермия и применение гипертермии соответственно, а предпочтительно, гипертермии, используемой в противоопухолевой терапии. В особенно предпочтительном варианте осуществления изобретения применяют местную гипертермию. И, наконец, другим методом лечения является гормонотерапия, а в частности гормонотерапия, используемая в противоопухолевой терапии. Для такой гормонотерапии обычно используются антиэстрогены и/или антиандрогены. В соответствии с этим, антиэстрогены, такие как тамоксифен, применяются, в частности, для лечения рака молочной железы, а антиандрогены, такие как, например, флутамид или ацетат ципротерона применяются, в частности, для лечения рака предстательной железы.
Описанные здесь аденовирусы могут быть также использованы для лечения опухолей, где указанная опухоль выбрана из группы, включающей первичные опухоли, вторичные опухоли, третичные опухоли и метастатические опухоли. В соответствии с этим, предпочтительно, чтобы указанные опухоли обладали, по меньшей мере, одним из нижеследующих признаков, а именно, чтобы они имели YВ-1 в клеточном ядре независимо от клеточного цикла и независимо от причины локализации YВ-1 в ядре, и/или чтобы они содержали YВ-1 с нарушенной регуляцией.
В соответствии с настоящим изобретением указанными клетками и опухолями соответственно, содержащими клетки, в которых реплицируются или способны реплицироваться аденовирусы настоящего изобретения, являются клетки и опухоли, обладающие одним или несколькими описанными здесь признаками, а в частности признаками, заключающимися в том, что они имеют YВ-1 в клеточном ядре независимо от клеточного цикла и независимо от причины локализации YВ-1 в ядре, и/или они имеют YВ-1 с нарушенной регуляцией, и в том, что эти клетки и опухоли, соответственно могут быть обработаны аденовирусами группы I и/или аденовирусами группы II в соответствии с настоящим изобретением, и что указанные аденовирусы могут быть использованы в целях изготовления лекарственного препарата для лечения заболеваний, где указанные аденовирусы экспрессируют нуклеиновую кислоту, кодирующую YВ-1. Следовательно, существует, предпочтительно, три категории клеток и соответствующих опухолей, в которых аденовирусы группы I и/или аденовирусы группы II настоящего изобретения могут реплицироваться, и которые могут быть подвергнуты лечению, а предпочтительно, лизису с применением этих аденовирусов, соответственно:
Группа А: Клетки, которые имеют YВ-1 в ядре независимо от клеточного цикла;
Группа В: Клетки, которые не имеют YВ-1 в ядре, а в частности независимо от клеточного цикла, но которые содержат YВ-1 с нарушенной регуляцией; и
Группа С: Клетки, которые не имеют YВ-1 в ядре, а в частности независимо от клеточного цикла, и которые не содержат YВ-1 с нарушенной регуляцией.
Что касается клеток группы А, то аденовирусы настоящего изобретения, а в частности, аденовирусы группы I, которые не экспрессируют дополнительный YВ-1, могут быть использованы для репликации или лизиса. Однако такие аденовирусы настоящего изобретения, а в частности аденовирусы группы I, которые экспрессируют дополнительный YВ-1, также могут быть использованы для репликации и лизиса. Это также относится к группе В. Не ограничиваясь нижесказанным, автор имеет основание предположить, что под действием белка Е1В, а в частности белка Е1В55К, и/или белка Е4, а в частности белка Е4orf6, эффективная репликация обеспечивается локализацией YВ-1 в ядре и переносом YВ-1 в ядро, соответственно. Этот процесс поддерживается YВ-1, дополнительно экспрессируемым аденовирусами.
В случае группы С, предпочтительно, аденовирусы настоящего изобретения, а в частности аденовирусы группы I, которые дополнительно экспрессируют YВ-1, могут быть использованы для репликации или лизиса. И в данном случае имеются основания предполагать, без каких-либо ограничений, что вышеуказанные механизмы репликации вируса являются не такими активными в конкретном клеточном теневом фенотипе, чтобы могла происходить эффективная репликация. Эффективная репликация может происходить только благодаря доставке YВ-1 и экспрессии YВ-1, соответственно, где лежащий в ее основе механизм, предположительно, заключается в том, что сверхэкспрессия YВ-1 приводит к ядерной локализации YВ-1, на что также указывается в работах Bargout [Bargout R.C. et al., Nature Medicine 1997, 3, 447-450] и Jurchott [Jurchott K et al., JBC 2003, 278, 27988-27996].
В соответствии с настоящим изобретением некоторые клетки, образующие опухоль, содержат в своем ядре либо эндогенный YВ-1, либо YВ-1, локализованный в ядре после индукции и активного транспорта в ядро, либо YВ-1, который имеет нарушенную регуляцию, как определено в настоящем изобретении. При этом предпочтительно, чтобы примерно 5% или выше, то есть 6%, 7%, 8% и т.п. опухоль-образующих клеток представляли собой клетки, позитивные по ядерному YВ-1, или клетки, в которых присутствует YВ-1 с нарушенной регуляцией. Что касается других опухолей, таких как опухоли молочной железы, остеосаркома, карцинома яичника, карцинома синовиальной оболочки или карцинома легкого, то процент опухолевых клеток, которые содержат YВ-1 с нарушенной регуляцией, или которые обнаруживают ядерную локализацию YВ-1 независимо от клеточного цикла, может составлять примерно от 30 до 50% [Kohno K. et al., BioEssays 2003, 25, 691-698]. Такие опухоли могут быть предпочтительно подвергнуты лечению аденовирусами в соответствии с настоящим изобретением. Ядерная локализация YВ-1 может быть индуцирована путем создания внешних и местных стрессовых условий соответственно. Такая индукция может быть, например, достигнута посредством облучения, а в частности УФ-облучения, введения уже описанных здесь цитостатических средств и гипертермии. Что касается гипертермии, то в этой связи важно отметить, что она может быть реализована очень специфическим способом, а более конкретно, локальным способом, так, чтобы мог обеспечиваться также специфический ядерный транспорт YВ-1 в ядро клетки, в результате чего будут выполняться условия, требуемые для репликации аденовируса, а следовательно, будет осуществляться лизис клетки и опухоли, которые имеют, предпочтительно, ограниченную локализацию (Stein U. Jurchott K. Walther W., Bergmann S. Schlag P.M, Royer H.D. J. Biol. Chem. 2001, 276(30):28562-9; Hu Z., Jin S, Scotto K.W, J. Biol. Chem. 2000 Jan 28; 275(4):2979-85; Ohga T., Uchiumi T. Makino Y. Koike K, Wada M, Kuwano M. Kohno K.J. Biol. Chem. 1998, 273(11):5997-6000).
Лекарственный препарат настоящего изобретения может быть также введен пациентам или группам пациентов, либо он может быть предназначен для такого введения пациентам, у которых соответствующее предварительное, последующее или одновременное лечение этим препаратом будет индуцировать транспорт YВ-1, в частности, в соответствующих опухолевых клетках и продуцирование в этих клетках YВ-1 с нарушенной регуляцией, соответственно.
На основании изложенной здесь технической идеи, каждый специалист, в целях генерирования различных вариантов аденовирусов, которые могут быть использованы в настоящем изобретении, может внести соответствующие модификации, в частности в Е1А, которыми могут быть, например, делеции или точковые мутации.
Как уже объяснялось выше, аденовирусы группы I и/или группы II способны реплицироваться в указанных клетках и в клеточных системах, соответственно, которые имеют YВ-1 в своем ядре. Для ответа на вопрос, могут ли аденовирусы, используемые в настоящем изобретении, также реплицироваться и таким образом лизировать опухоль, статус клеток, с точки зрения присутствия или отсутствия Rb, то есть продукта-супрессора опухоли ретинобластомы, не имеет решающего значения. Кроме того, что касается применения указанных аденовирусов в соответствии с настоящим изобретением, то не следует принимать в расчет р53-статус инфицированных клеток, то есть клеток, инфицируемых или обрабатываемых описанными здесь аденовирусными системами, которые применяются к клеткам, позитивным по ядерному YВ-1, то есть к клеткам, содержащим YВ-1 в ядре независимо от их клеточного статуса, где этот р53-статус, а также Rb-статус не влияют на репликацию аденовируса, применяемого для реализации описанной здесь технической идеи.
Трансактивирующий онкоген и онкогенный белок соответственно, а в частности Е1А, предпочтительно, аденовирусов группы II, могут находиться под контролем собственных природных аденовирусных промоторов и/или под контролем опухолеспецифического или тканеспецифического промотора. Подходящие неаденовирусные промоторы могут быть выбраны из группы, включающей промотор цитомегаловируса, промотор RSV (вируса саркомы Рауса), аденовирусный промотор Vа I и невирусный промотор YВ-1 (Makino Y. et al., Nucleic Acids Res. 1996, 15, 1873-1878). Другими промоторами, которые могут быть использованы в соответствии с любым из описанных здесь аспектов настоящего изобретения, могут быть промотор теломеразы, промотор альфа-фетопротеина (AFP), промотор карциноэмбрионального антигена (СЕА) (Cao G., Kuriyama, S., Gao, J. Mitoro, A., Cui, L., Nakatani, T., Zhang X., Kikukawa, M., Pan, X. Fukui, H., Qi, Z. Int. J. Cancer, 78, 242-247, 1998), промотор L-пластина (Chung I. Schwartz, P.E., Crystal R.C., Pizzorno, G., Leavitt, J. Deisseroth, AB. Cancer Gene Therapy, 6, 99-106, 1999), промотор аргинина-вазопрессина (Coulson J.M. Staley, J. Woll, P.J. British J. Cancer 80, 1935-1944, 1999), промотор E2f (Tsukada et al. Cancer Res., 62, 3428-3477), промотор уроплакина II (Zhang et al., Cancer Res., 62, 3743-3750, 2002), промотор PSA (Hallenbeck P.L., Chang Y.N., Hay, C, Golightly, D., Stewart, D., Lin, J. Phipps, S., Chiang Y.L. Human Gene Therapy, 10, 1721-1733, 1999), промотор тирозиназы (Nettelbeck D.M. Anti-Cancer Drugs, 14, 577-584, 2003), промотор циклооксигеназы 2 (Nettelbeck D.M., Rivera, A.A., Davydova J. Dieckmann, D., Yamamoto M., Curiel D.T. Melanoma Res., 13, 287-292, 2003) и индуцирующие системы, такие как тетрациклин (Xu X.L., Mizuguchi H., Mayumi T., Hayakawa, T. Gene, 309, 145-151, 2003). Кроме того, YB-1-зависимым Е2-поздним промотором аденовирусов, описанным в заявке на патент Германии DE 10150984.7, является промотор, который может быть использован в настоящем изобретении.
Известно, что промотор теломеразы играет очень важную роль в клетках человека. В соответствии с этим, активность теломеразы регулируется посредством транскрипционного контроля гена обратной транскриптазы теломеразы (hTERT), которая является каталитической субъединицей данного фермента. Активность экспрессии теломеразы в опухолевых клетках человека составляет 85%. В противоположность этому, она является неактивной в большинстве нормальных клеток. Исключением являются лишь зародышевые клетки и эмбриональная ткань. (Braunstein I. et al., Cancer Research, 61, 5529-5536, 2001; Majumdar A.S. et al., Gene Therapy 8, 568-578, 2001). Более детальное исследование промотора hTERT выявило, что фрагменты 283 п.н. и 82 п.н., соответственно расположенные на значительном расстоянии от инициирующего кодона, являются достаточными для специфической экспрессии в опухолевых клетках (Braunstein I. et al., Majumdar A.S. et al., см. выше). Поэтому этот промотор и специфические фрагменты соответственно являются подходящими для специфической экспрессии в опухолевых клетках гена, в частности трансгена, а предпочтительно, одного из описанных здесь трансгенов. Такой промотор будет обеспечивать экспрессию модифицированного онкогена, а предпочтительно, онкогенного белка Е1А лишь в опухолевых клетках. Кроме того, в одном из вариантов осуществления изобретения рассматривается экспрессия трансгена в аденовирусном векторе, находящемся под контролем этих промоторов, а предпочтительно, трансгена, выбранного из группы, включающей Е4orf6, Е1В55kD, АDP и YВ-1. В настоящем изобретении также предусматривается, что открытая рамка считывания трансактивирующего онкогенного белка, а в частности белка Е1А, соответствует рамке считывания одного или нескольких генных продуктов аденовирусной системы. Однако эта открытая рамка считывания трансактивирующего белка Е1А может быть также независимой от них.
В настоящем изобретении предусматривается, что различные промоторы, описанные выше, также применяются в соответствии с различными вариантами аденовирусов настоящего изобретения, предпочтительно, аденовирусов группы I, а в частности, в том случае, если используемый промотор отличается от промотора, регулирующего экспрессию соответствующего белка или продукта экспрессии в аденовирусах дикого типа. Таким образом, вышеупомянутые промоторы являются подходящими гетерологичными промоторами, определенными в настоящем изобретении. В предпочтительных вариантах аденовирусов настоящего изобретения, а в частности аденовирусов группы I, предусматривается, что при применении аденовирусов для клеток группы А и В, определенных выше, экспрессия белка Е1В и/или белка Е4 начинается с таких гетерологичных промоторов, где предпочтительно, но не исключительно, экспрессия белка Е1А регулируется YВ-1. Экспрессия белка Е1А в этом и других вариантах изобретения находится под контролем YВ-1-регулируемого промотора, такого как, например, аденовирусный Е2-поздний промотор. Это также справедливо для случая, где белок Е1В и/или белок Е4 экспрессируется/экспрессируются в экспрессионной кассете.
В предпочтительных вариантах аденовирусов настоящего изобретения, а в частности аденовирусов группы I, предусматривается, что при применении аденовирусов в сочетании с клетками группы С, каждый и независимо взятый промотор представляет собой опухолеспецифический, органоспецифический или тканеспецифический промотор. В соответствии с этим достаточно, чтобы, по меньшей мере, одним из таких промоторов, регулирующих экспрессию белка Е1В, белка Е4 и/или белка Е1А, был специфический промотор. Такая специфичность по отношению к опухоли, органу и ткани, является гарантией того, что репликация аденовирусов настоящего изобретения будет происходить только в клетках соответствующих опухолей, органов или тканей, и что помимо этого никакая другая ткань не будет повреждена в результате репликации аденовирусов, то есть, например, она не будет подвергаться лизису. При этом, предпочтительно, чтобы второй, а более предпочтительно все три белка находились под контролем таких опухолеспецифических, органоспецифических или тканеспецифических промоторов. С использованием таких аденовирусов могут также подвергаться лизису те клетки, которые не образуют опухоли, или которые не могут образовывать такую опухоль, но которые по тем или иным причинам, то есть по медицинским показаниям, должны быть разрушены или удалены из организма, предпочтительно, из организма млекопитающего, а более предпочтительно, из организма человека, по той причине, например, что эти клетки продуцируют нежелательный фактор, либо продуцируют такой фактор на слишком высоком уровне.
В одном из вариантов настоящего изобретения предусматривается, что клетки, для лизиса которых применяются аденовирусы настоящего изобретения, являются резистентными, а предпочтительно, они являются полирезистентными.
Резистентностью, описанной в настоящем изобретении и характерной для опухолей и пациентов, подвергаемых лечению, является резистентность, опосредуемая нижеследующими генами, которыми являются, но не ограничиваются ими: гены MDR, MRP, топоизомеразы, ВСL2, глутатион-2-трансферазы (GST), протеинкиназы С (PKC). Поскольку действие цитостатических средств основано, среди прочих, на индуцировании апопотоза, то экспрессия генов, ответственных за апоптоз, играет важную роль в развитии любой резистентности, которая ассоциируется с нижеследующими факторами, а именно, с Fas, фактором семейства ВСL2, HSP 70 и EGFR [Kim et al., Cancer Chemther. Pharmacol. 2002, 50, 343-352].
В работе Levenson et al. [Levenson V.V. et al., Cancer Res., 2000, 60, 5027-5030] указывается, что по сравнению с нерезистентными опухолевыми клетками в резистентных опухолевых клетках уровень экспрессии YВ-1 значительно повышается.
Используемый здесь термин “резистентность” означает, предпочтительно, резистентность против описанных здесь цитостатических средств. Такая резистентность ко многим лекарственным препаратам, предпочтительно, ассоциируется с экспрессией, а более предпочтительно, со сверхэкспрессией мембраноcвязанного транспортного белка Р-гликопротеина, который может быть использован в качестве маркера для выявления соответствующих клеток, а также опухолей, имеющих такой маркер, и соответствующих групп пациентов. Используемый здесь термин “резистентность” также означает резистентность, опосредуемую Р-гликопротеином, которая также называется классической резистентностью, и атипичную резистентность, которая представляет собой МRP-опосредуемую резистентность, или резистентность другого типа, не опосредуемую Р-гликопротеином. Другим маркером, который коррелирует с экспрессией YВ-1, является топоизомераза II-альфа. Таким образом, для того чтобы решить, будет ли для данного пациента успешным лечение с использованием аденовирусов настоящего изобретения, в методе скрининга, вместо или помимо определения уровня YВ-1 в ядре, может быть использована топоизомераза II альфа. Другим маркером, который, в принципе, может быть использован аналогично Р-гликопротеину, является MRP. Еще одним маркером, по меньшей мере, до той степени, в которой это относится к клеткам колоректальной карциномы или к пациентам с колоректальной карциномой, является РCNA (англ., ядерный антиген пролиферирующих клеток) (Hasan S. et al., Nature 15, 387-391, 2001), как, например, описано у Shibao K. et al. (Shibao K. et al., Int. Cancer, 83, 732-737, 1999). И наконец, исходя из описанного выше, экспрессия MDR (резистентность ко многим лекарственным препаратам) является маркером (Oda Y. et al., Clin. Cancer Res., 4, 2273-2277, 1998), по меньшей мере, для клеток рака молочной железы и клеток остеосаркомы. Другим возможным маркером, который может быть использован в настоящем изобретении, является р73 (Kamiya M., Nakazatp, Y., J. Neurooncology 59, 143-149 (2002); Stiewe et al., J. Biol. Chem. 278, 14230-14236, 2003).
И наконец, следует также отметить, что YВ-1 может быть использован в настоящем изобретении в качестве прогностического маркера в клетках рака молочной железы. В соответствии с этим, рецидив после хирургической операции и химиотерапии наблюдается лишь у пациентов, в первичной опухоли которых наблюдается повышенный уровень экспрессии YВ-1 [Janz M. et al., Int. J. Cancer 2002, 97, 278-282].
Важное преимущество настоящего изобретения состоит в том, что лечению с использованием в настоящем изобретении описанных здесь аденовирусов могут быть подвергнуты также пациенты, которые по тем или иным причинам не могут дальше продолжать обычное консервативное лечение, и где последующее лечение опухолевого заболевания уже известными методами не может дать ожидаемого успеха, а в частности, где использование цитостатических средств и облучения больше не представляется целесообразным и больше не может давать положительные результаты с точки зрения их воздействия на опухоль или снижения размеров опухоли. Используемый здесь термин “опухоль”, в общих чертах, означает любое опухолевое или раковое заболевание, при котором в ядре опухолевой клетки содержится, предпочтительно, независимо от ее клеточного цикла, эндогенный YВ-1, или YВ-1, индуцированный воздействием описанных здесь экзогенных факторов, и/или YВ-1 с нарушенной регуляцией.
Кроме того, описанные здесь вирусы могут быть использованы, в основном, для лечения опухолей. Такие опухоли, предпочтительно, выбраны из группы, включающей рак молочной железы, карциному яичника, рак предстательной железы, остеосаркому, глиобластому, меланому, мелкоклеточную карциному легких и рак толстой и прямой кишки. Другими опухолями являются опухоли, которые обладают описанной здесь резистентностью, а предпочтительно, опухоли, которые обладают полирезистентностью, а в частности также опухоли, относящиеся к вышеописанной группе. Особенно предпочтительными опухолями являются опухоли, выбранные из группы, включающей опухоли молочной железы, опухоли костей, опухоли желудка, опухоли тонкого кишечника, опухоли желчного пузыря, опухоли поджелудочной железы, опухоли печени, опухоли почек, опухоли головного мозга, опухоли яичника, опухоли кожи, опухоли придатков кожи, опухоли головы и шеи, опухоли матки, опухоли синовиальной оболочки, опухоли гортани, опухоли слюнных желез, опухоли пищевода, опухоли языка и опухоли предстательной железы. В соответствии с настоящим изобретением, предпочтительно, чтобы манифестации этих опухолей полностью соответствовали их описанию в настоящем изобретении.
Аденовирусами настоящего изобретения, предпочтительно, являются аденовирусы группы I, а аденовирусами, применяемыми в соответствии с настоящим изобретением, предпочтительно, являются аденовирусы группы II.
Применение описанных здесь аденовирусов, а в частности аденовирусов группы I и/или аденовирусов группы II, в качестве лекарственных средств, а в частности для системного введения, может быть улучшено путем подходящей доставки аденовирусов. Инфицирование опухолевых клеток аденовирусами до некоторой степени зависит, среди прочих, от присутствия рецептора коксакивируса-аденовируса САR и различных интегринов. По мере увеличения уровня их экспрессии в клетках, титры их инфекции могут становиться очень низкими (б.о.е./клетку). До настоящего времени были разработаны различные стратегии для достижения так называемой повторной доставки рекомбинантных аденовирусов, например, путем встраивания гетерологичных последовательностей в область фибриллярного утолщения с использованием биспецифических антител; покрытия аденовирусов полимерами, введения лигандов в фибриллу Аd; замены утолщения аденовируса серотипа 5 и фибриллярного стержня и утолщения аденовируса серотипа 5 на утолщение аденовируса серотипа 3 и фибриллярный стержень и утолщение Аd35; и модификации основания пентонов, соответственно (Nicklin S.A., et al., Molecular Therapy 2001, 4, 534-542; Magnusson M.K. et al., J. of Virology 2001, 75, 7280-7289; Barnett B.G. et al., Biochimica et Biophysica Acta 2002, 1575, 1-14). Что касается различных аспектов настоящего изобретения, то реализация таких дополнительных вариантов и отличительных признаков, соответственно, в аденовирусах настоящего изобретения и аденовирусах, применяемых в соответствии с настоящим изобретением, а в частности в аденовирусах группы I и в аденовирусах группы II, входит в объем настоящего изобретения.
В другом своем аспекте, настоящее изобретение относится к способу отбора пациентов, которые могут быть подвергнуты лечению с использованием модифицированного аденовируса, то есть аденовируса, применяемого в соответствии с настоящим изобретением, такого как, например, AdΔ24, dl922-947, E1Ad/01/07, CB016, или вирусов, описанных в Европейском патенте ЕР 0931830, и/или аденовируса группы I и/или аденовируса группы II, где указанный способ включает следующие стадии:
- анализ образца опухолевой ткани и
- определение локализации YВ-1 в ядре клетки, независимо от ее клеточного цикла, или наличия в клетках YВ-1 с нарушенной регуляцией.
Вместо детекции или помимо детекции присутствия YВ-1 может быть осуществлена детекция вышеупомянутых маркеров.
В случае, если опухолевая ткань или ее часть содержит YВ-1 в клеточном ядре, а предпочтительно, независимо от клеточного цикла, или если она содержит YВ-1 с нарушенной регуляцией, то описанные здесь аденовирусы, а в частности аденовирусы группы I и/или аденовирусы группы II, могут быть применены в соответствии с настоящим изобретением.
В одном из вариантов осуществления способа настоящего изобретения предусматривается, что анализ опухолевой ткани проводят с использованием агента, выбранного из группы, включающей антитела против YВ-1, аптамеры против YВ-1, энантиомеры (оптические изомеры) против YВ-1, а также антикалины против YВ-1. В общих чертах, указанные агенты могут быть продуцированы и использованы, соответственно, в качестве соответствующих маркеров. Методы получения антител, а в частности моноклональных антител, известны специалистам. Другими агентами для специфической детекции YВ-1 или маркеров, являются пептиды, которые с высокой аффинностью связываются со структурами-мишенями, а в данном случае с YВ-1 или с указанными маркерами. Для генерирования таких пептидов существуют уже описанные методы, такие как метод фагового представления. Для этих целей в качестве исходного материала используют пептидную библиотеку, где отдельные пептиды имеют длину примерно 8-20 аминокислот, а размер библиотеки составляет примерно от 102 до 1018, а предпочтительно, от 108 до 1015 различных пептидов. Конкретными формами полипептидов, связывающихся с молекулой-мишенью, являются так называемые антикалины, которые, например, описаны в заявке на патент Германии DЕ 19742706.
Дополнительными агентами для специфического связывания с YВ-1 или с описанными здесь соответствующими маркерами и таким образом, для детекции независимой от клеточного цикла локализации YВ-1 в клеточном ядре являются так называемые аптамеры, то есть D-нуклеиновые кислоты, которые присутствуют либо в форме РНК или ДНК, либо в форме одной цепи или двойной цепи, и которые специфически связываются с молекулой мишенью. Получение аптамеров описано, например, в Европейском патенте ЕР 0533838. Конкретными вариантами аптамеров являются так называемые аптазимы, описанные, например, у Piganeau N. et al. (2000), Angew. Chem. Int. Ed., 39, no. 29, pages 4369-4373. Эти аптазимы представляют собой конкретные варианты аптамеров, поскольку, помимо аптамерной части, они содержат рибозимную часть, и после связывания с молекулой-мишенью или высвобождения молекулы-мишени, связывающейся с аптамерной частью, рибозимная часть становится каталитически активной и расщепляет субстрат нуклеиновой кислоты с продуцированием сигнала.
Другой формой аптамеров являются так называемые энантиомеры, то есть нуклеиновые кислоты, связывающиеся с молекулой-мишенью и состоящие из L-нуклеиновых кислот. Метод получения таких энантиомеров описан, например, в WO 98/08856.
Образец опухолевой ткани может быть взят путем пункции или при хирургической операции. Для того чтобы определить, присутствует ли YВ-1 в ядре клеток независимо от его клеточного цикла, часто проводят микроскопические анализы и/или иммуногистологические анализы, а предпочтительно, анализы с использованием антител или любых других описанных выше агентов. Дополнительными методами детекции YВ-1 в ядре и его локализации в ядре независимо от клеточного цикла являются методы, известные специалистам. Так, например, локализация YВ-1 может быть легко детектирована путем сканирования срезов ткани, окрашенных на YВ-1. Частота встречаемости YВ-1 в ядре уже указывает на то, что его локализация в ядре не зависит от клеточного цикла. Другим возможным способом детекции YВ-1 в ядре независимо от клеточного цикла является окрашивание, свидетельствующее о присутствии YВ-1, а также определение локализации YВ-1 в ядре и фазы клеточного цикла. Однако такой способ, а также детекция YВ-1, соответственно, могут быть также осуществлены с использованием вышеупомянутых агентов против YВ-1. Детекцию этих агентов осуществляют в соответствии с процедурами, известными специалистам. Поскольку указанные агенты специфически связываются с YВ-1, но не с какими-либо другими структурами в анализируемом образце, а в частности с другими структурами клеток, то локализация указанных агентов, а следовательно, и их специфическое связывание с YВ-1, а также локализация YВ-1 могут быть соответственно детектированы и оценены с помощью соответствующего мечения этих агентов. Методы мечения указанных агентов известны специалистам. Те же самые методы могут быть также применены для определения наличия и числа клеток в образце, содержащих YВ-1 с нарушенной регуляцией. Поскольку YВ-1 с нарушенной регуляцией также указывает на сверхэкспрессию, в отличие от YВ-1 с ненарушенной регуляцией, то относительная экспрессия YВ-1 по сравнению со стандартным образцом может быть использована для установления нарушения регуляции YВ-1 в анализируемой клетке.
Ниже приводится подробное описание настоящего изобретения со ссылками на графический материал и примеры, которые представлены для иллюстрации новых признаков, вариантов и преимуществ настоящего изобретения.
На фиг.1 показаны структурные конструкции аденовирусных векторов, называемых здесь АdЕ1/Е3-минус, которые представляют собой Е1/Е3-делетированные аденовирусы, аденовирусов дикого типа и аденовируса dl520.
На фиг.2 представлены домены белка Е1А, связывающиеся с р300, р107 и р105.
На фиг.3 представлены клетки U2ОS, которые не содержат YВ-1 в своем ядре после инфицирования Е1/Е3-делетированным аденовирусом Аd5, называемым здесь Е1/Е3-минус Аd5, и аденовирусом dl520.
На фиг.4 представлены клетки 257RDB, которые не содержат YВ-1 в своем ядре после инфицирования Е1/Е3-делетированным аденовирусом Аd5, называемым здесь Е1/Е3-минус Аd5, и аденовирусом dl520.
На фиг.5 представлены клетки 257RDB и клетки U2ОS после инцифирования аденовирусом dl1119/1131.
На фиг.6 представлен результат анализа ЕМSA, который подтверждает, что YВ-1 присутствует в полирезистентных клетках и в клеточных линиях 257RDB, 181RDB, МСF-7Ad соответственно, тогда как в ядрах клеток U2ОS и НеLa YВ-1 не присутствует.
На фиг.7 представлены структурные конструкции белка Е1А аденовируса дикого типа, аденовируса dl520 и аденовируса dl1119/1131.
На фиг.8 представлена гистограмма, иллюстрирующая эффективную репликацию аденовирусов в присутствии дополнительных экспрессируемых вирусных белков в абсолютных величинах.
На фиг.9 представлена гистограмма, иллюстрирующая увеличение эффективности репликации аденовирусов в присутствии дополнительных экспрессируемых вирусных белков.
На фиг.10 показаны лунки с культивируемыми клетками U2ОS после их окрашивания кристаллическим фиолетовым и после инфицирования аденовирусом dl520 при 10 и 30 б.о.е./клетку, соответственно, и лунки с контрольными клетками (К), в которые не был введен даунорубицин, и лунки с клетками, в которые было введено 40 нг даунорубицина на 1 мл, соответственно.
На фиг.11 показаны лунки с культивируемыми клетками НеLa после их окрашивания кристаллическим фиолетовым и после инфицирования аденовирусом dl520 при 10 и 30 б.о.е./клетку, соответственно, и лунки с контрольными клетками (К), в которые не был введен даунорубицин, и лунки с клетками, в которые было введено 40 нг даунорубицина на мл, соответственно.
На фиг.12 представлен график зависимости объема опухолей, имеющих различное происхождение (RDB257 и НеLa), от времени после обработки PBS и dl520 соответственно.
На фиг.13 представлены фотографии умерщвленных мышей с развивающимися опухолями RDB257 после их обработки PBS и 5×108 dl520 соответственно.
На фиг.14 представлены результаты Вестерн-блот-анализа клеточного экстракта (подкожных растущих опухолей) клеток RDB257 и клеток НеLa после инфицирования dl520.
На фиг.15 представлена гистограмма, иллюстрирующая эффективность репликации и образование частиц, соответственно, dl520 и аденовируса дикого типа в опухолевых клетках, позитивных по ядерному YВ-1 (257RDB и 181RDB), и в опухолевых клетках, негативных по ядерному YВ-1 (НеLa, U2ОS).
На фиг.16 представлена структурная конструкция аденовируса дикого типа и аденовирусного вектора АdXvir03.
На фиг.17 представлена структурная конструкция аденовирусного вектора АdXvir03/01.
На фиг.18А/В показаны лунки с культивируемыми клетками 181RDB (фиг.18А) и клетками 272RDB (фиг.18В) после их окрашивания кристаллическим фиолетовым и после инфицирования аденовирусом Аd312 (20 б.о.е./клетку), Xvir03 (5 б.о.е./клетку), и лунки с контрольными клетками (неинфицированными), где окрашивание кристаллическим фиолетовым осуществляли через пять дней после инфицирования.
На фиг.19 представлены результаты Нозерн-блот-анализа экспрессии гена Е2 в клетках А549 и в клетках U2OS после инфицирования аденовирусом дикого типа Аd5 и аденовирусом Аd312.
На фиг.20 представлены результаты Нозерн-блот-анализа экспрессии гена Е2 в клетках U2OS после инфицирования аденовирусом дикого типа и аденовирусом дельта-24 через 12 часов и 24 часа.
На фиг.21 представлена структурная конструкция аденовирусного вектора XVirPSJL1.
На фиг.22 представлена структурная конструкция аденовирусного вектора XVirPSJL2.
На фиг.23 показаны лунки с культивируемыми клетками НеLa после их окрашивания кристаллическим фиолетовым и после инфицирования аденовирусом dl520 с использованием различных б.о.е./клетку.
На фиг.24 представлена гистограмма, иллюстрирующая активность люциферазы в клетках U2OS, клетках НеLa и клетках 257RDB при использовании различных промоторных фрагментов аденовирусного Е2-позднего промотора.
На фиг.25 представлена гистограмма, иллюстрирующая число вирусных частиц через два дня и пять дней после инфицирования клеток U2OS YВ-1-экспрессирующим аденовирусом и вирусом Аd312, где показано различие между внутриклеточными оставшимися вирусными частицами (закрашенные черным) и высвобожденными внеклеточными вирусными частицами (горизонтальные штрихи).
Пример 1: Типы модификаций Е1А, которые могут содержать аденовирусы, используемые в настоящем изобретении
На фиг.1 представлена структурная конструкция аденовирусных векторов АdЕ1/Е3-минус, то есть Е1/Е3-делетированных аденовирусов, аденовируса дикого типа и аденовируса dl520.
Аденовирус АdЕ1/Е3-минус не содержит область, кодирующую функциональный Е1А или функциональные Е1В или Е3, и используется в экспериментах настоящего изобретения в качестве контроля токсичности.
Ген Е1А дикого типа кодирует все 5 белков, которые продуцируются посредством альтернативного сплайсинга РНК Е1А. Среди этих продуцируемых белков имеются два различных белка, а именно белок, состоящий из 289 аминокислот, и белок, состоящий из 243 аминокислот. dl520 не кодирует белок в 289 аминокислот, поскольку он имеет делецию во фрагменте CR3 гена Е1А, а поэтому продукт гена 13S отсутствует. Аденовирус dl520, который может быть использован в настоящем изобретении, называется вирусом 12S Е1А и известен специалистам. Аденовирус dl347 (Wong H.K. & Ziff E.B. J. Virol. 68, 4910-4920, 1994), известный из предшествующих работ, представляет собой вирус 12S Е1А, который также может быть использован в настоящем изобретении.
В белке, состоящем из 289 аминокислот и кодируемом мРНК 13S Е1А, имеются 3 области, которые являются консервативными для различных подтипов аденовирусов. Эти области называются CR1, CR2 и CR3. Области CR1 и CR2 присутствуют в обоих белках Е1А (Е1А 12S и Е1А 13S), то есть в белке, состоящем из 289 аминокислот, и в белке, состоящем из 243 аминокислот, а область CR3 присутствует только в более крупном из двух вышеупомянутых белков.
Область CR3 необходима для активации вирусных генов, а в частности Е1В, Е2, Е3 и Е4. Вирусы, которые содержат лишь меньший по размеру белок, то есть белок, состоящий из 243 аминокислот, обладают лишь очень слабым трансактивирующим действием на вирусные гены и не стимулируют репликацию аденовирусов в тех клетках, которые не содержат YВ-1 в своем ядре. Поскольку YВ-1 присутствует в ядре только опухолевых клеток и может быть детектирован только в этих клетках, то указанный вектор является подходящим для индуцирования опухолеспецифической репликации.
Из-за делеции области CR3 в dl520 этот аденовирус не может переносить клеточный YВ-1 в клеточное ядро, то есть не может осуществлять так называемую транслокацию, и таким образом YВ-1 отсутствует в положении, позволяющем этому аденовирусу реплицироваться в клетках, негативных по ядерному YВ-1, а поэтому этот вирус может быть использован в настоящем изобретении, поскольку он обладает трансактивирующим действием, необходимым для целей настоящего изобретения.
Пример 2: Механизм действия аденовирусов в зависимости от статуса Rb в клетках
На фиг.2 представлены домены белка Е1А, связывающиеся с р300, р107 и р105. Р300, а также р107 представляют собой клеточный связывающий белок. Связывание белка ретинобластомы (рRb), белка-супрессора опухоли, опосредуется CR1 и CR2. Исследования показали, что рRb и р107/р300 находятся в комбинации с клеточным фактором транскрипции Е2F, эффективным регулятором транскрипции. Белок Е1А дикого типа препятствует связыванию Е2F с Rb. Высвобождаемый таким образом Е2F связывается с ранним промотором Е2 и индуцирует репликацию аденовирусов.
Из предшествующих работ известно, что некоторые делеции в онкопротеине Е1А могут приводить к образованию рекомбинантных аденовирусных векторов, таких как упомянутые ниже векторы, которые обладают способностью реплицироваться преимущественно в Rb-негативных клетках и могут быть использованы в настоящем изобретении. Так, например, аденовирусный вектор dl922-947 имеет делецию в области CR2 (в положениях аминокислот 122-129), а вектор СВ016 имеет делеции в области CR1 (в положениях аминокислот 27-80) и в области CR2 (в положениях аминокислот 122-129). Вектор Е1Аdl01/07 имеет делецию в области CR2 (в положениях аминокислот 111-123). Кроме того, из-за дополнительной делеции у N-конца (в положениях аминокислот 4-25) он не связывается с белком р300. Аденовирусный вектор AdΔ24 имеет делецию в области CR2 (в положениях аминокислот 120-127). Аденовирусный вектор, описанный в патенте ЕР 0931830, имеет делеции в области CR1 и в области CR2.
Механизм связывания Е2F/RВ и механизм высвобождения Е2F, опосредуемые Е1А, фундаментально отличаются от механизма, лежащего в основе настоящего изобретения. В отличие от предположений, высказываемых в предшествующих работах, авторы настоящего изобретения считают, что решающую роль в репликации вируса играет не отделение Е2F от белка Rb, хотя оно и является важным, а ядерная локализация человеческого фактора транскрипции YВ-1. В нормальных клетках этот фактор транскрипции присутствует только в цитоплазме на протяжении всего клеточного цикла. После инфицирования аденовирусом этот фактор при определенных условиях направляется в ядро или уже присутствует в ядре в некоторых клеточных системах, таких как клетки при некоторых опухолевых заболеваниях, включая, например, но не ограничиваясь ими, рак молочной железы, карциному яичника, карциному предстательной железы, остеосаркому, глиобластому, меланому, мелкоклеточную карциному легких и колоректальную карциному.
Пример 3: Инфицирование клеток U2ОS
Высевали 100000 клеток U2ОS на лунку. На следующий день клетки инфицировали различными аденовирусами, представленными на фиг.3. Инфицирование проводили в 500 мкл бессывороточной среды DMEM при 37°С в течение 1 ч. Затем инфекционную среду удаляли и заменяли 2 мл полной среды (10% FCS/DMEM). Анализ проводили через 3 дня путем окрашивания кристаллическим фиолетовым.
Как видно на фиг.3, клетки U2ОS, которые не имели YВ-1 в ядре, не обнаруживали лизиса, на что указывало окрашивание кристаллическим фиолетовым, после инфицирования двумя различными аденовирусами, а именно Е1/Е3-делетированным аденовирусом, называемым Е1/Е3-минус, и аденовирусом dl520, который может быть использован в настоящем изобретении. Для этого сначала среду удаляли. Затем клетки покрывали кристаллическим фиолетовым (50% ЕТОН, 3% формальдегид, 5% уксусная кислота, 1% кристаллический фиолетовый) и инкубировали при комнатной температуре в течение 5-10 минут. После этого 6-луночные планшеты тщательно ополаскивали водой и сушили при комнатной температуре.
Это подтверждает вывод, лежащий в основе настоящего изобретения, который заключается в том, что для индуцирования лизиса инфицированных клеток вирусами, используемыми в настоящем изобретении, необходимо присутствие YВ-1.
Пример 4: Инфицирование клеток 257RDB
Высевали 100000 клеток 257RDB на лунку. На следующий день клетки инфицировали различными аденовирусами, представленными на фиг.4. Инфицирование проводили в 500 мкл бессывороточной среды DMEM при 37°С в течение 1 ч. Затем инфекционную среду удаляли и заменяли 2 мл полной среды (10% FCS/DMEM). Анализ проводили через три дня путем окрашивания кристаллическим фиолетовым.
Результат этого эксперимента проиллюстрирован на фиг.4. Аденовирус, называемый Е1/Е3-минус-Аd5, который является Е1/Е3-делетированным, не обнаруживал какого-либо лизирующего действия при низких m.o.i. (б.о.е./клетку) после инфицирования клеток 257RDB, имеющих YВ-1 в своем ядре. В противоположность этому, dl520, который как показано в примере 3, не реплицировался в негативных по ядерному YВ-1 клетках, но при этом кодировал трансактивирующий онкогенный белок Е1А в соответствии с настоящим изобретением, обеспечивал фактически полный лизис при m.o.i. (множественности заражения) 40 б.о.е. на клетку, и преобладающий лизис при m.o.i. 10 б.о.е. на клетку. Отсюда можно сделать вывод, что для dl520 и аналогичных вирусов, таких как описанные здесь dl1119/1131 или АdXvir03, требуется m.o.i., которая примерно на один порядок меньше (в десять раз) по сравнению с m.o.i. для Е1-делетированного или Е1/Е3-делетированного аденовирусов, что подтверждает их пригодность для клинического применения.
Как показано на фиг.7, белок Е1А вируса dl520 отличается тем, что в нем область CR3 является делетированной, что приводит к трансактивации, необходимой для использования в настоящем изобретении и для репликации в клетках, позитивных по ядерному YВ-1.
Пример 5: Инфицирование клеток 257RDB и U2ОS вирусом dl1119/1131
Как показано на фиг.5, лизис клеток U2ОS, негативных по ядерному YВ-1, не наблюдается после их инфицирования при m.o.i.= 20 б.о.е. на клетку аденовирусом dl1119/1131, который имел делецию в положении аминокислот 4-138 белка Е1А и в нуклеиновой кислоте, кодирующей этот белок, и кроме того, содержал стоп-кодон после аминокислоты 218, при этом экспрессируемый усеченный белок Е1А содержал область CR3 полного белка Е1А. В качестве негативного контроля использовали слой неинфицированных клеток.
В противоположность этому, при m.o.i. = 20 б.о.е. на клетку наблюдался фактически полный лизис указанного клеточного слоя, осуществляемый аденовирусом dl1119/1131 в клеточной системе, такой как клетки 257RDB, содержащие YВ-1 в своем ядре, то есть клетки, позитивные по ядерному YВ-1. Таким образом, этот пример является дополнительным подтверждением того, что модифицированный онкогенный белок Е1А, который, как показано на фиг.7, содержит, например, только область CR3 и не содержит области CR1 и области CR2, обеспечивает требуемую трансактивацию в клетках, позитивных по ядерному YВ-1, которая необходима для репликации аденовирусов в соответствии с настоящим изобретением и которая приводит к репликации вируса. Поэтому аденовирус dl1119/1131 является дополнительным аденовирусом, который может быть использован в настоящем изобретении. В настоящем изобретении также показано, что могут быть использованы вирусы, сконструированные аналогично вирусу dl1119/1131, которые имеют такую же область CR3, но в отличие от него они также содержат область CR1 и/или область CR2.
Пример 6: Детекция ядерного YВ-1 в клетках, резистентных ко многим лекарственным препаратам
Этот пример основан на предположении, что ядерный YВ-1, в качестве фактора транскрипции, должен связываться с Y-боксом (последовательностью СААТ) в промоторе mdr1 (промоторе гена резистентности ко многим лекарственным препаратам). Для детекции этого ядерного YВ-1 проводили так называемый анализ ЕМSA (анализ на сдвиг электрофоретической подвижности). Для этого ядерный белок выделяли, а затем 1-10 мкг белка инкубировали вместе с коротким ДНК-фрагментом (олигонуклеотидом) при 37°С. Для определения ядерного YВ-1 использовали следующий олигонуклеотид: промотор mdr1 для U2ОS (в положениях от -86 до -67): TGAGGCTGATTGGCTGGGCA (Х-бокс подчеркнут).
Перед проведением этой процедуры, ДНК-фрагмент у его 5'-конца подвергали радиоактивному мечению 32Р. Затем проводили разделение в нативном полиакриламидном геле. В случае, если белок YВ-1 связывался с последовательностью в указанном олигонуклеотиде, то он мог быть детектирован, поскольку любой несвязанный олигонуклеотид мигрировал в геле быстрее, чем связанный олигонуклеотид (Holm P.S. et al., JBC 277, 10427-10434; Bargou R.C. et al., Nature Medicine 3, 447-450, 1997).
Как можно видеть на фиг.6, анализ ЕМSA показал, что в отличие от клеточных линий U2ОS и клеток НеLa, YВ-1 присутствует в ядре полирезистентных клеток 257RDB, 181RDB и МСF-7Ad.
Результаты, представленные в примерах 4 и 5, подтвердили, что в отличие от клеток U2ОS аденовирусы dl520 и dl1119/1131 реплицируются в клетках, позитивных по ядерному YВ-1, таких как, например, 257RDB, и индуцируют их лизис. Эти результаты подтверждают выводы о том, что указанные аденовирусы могут быть использованы в настоящем изобретении. Кроме того, эти результаты подтвердили, что по сравнению с аденовирусом дикого типа даже слабая трансактивация вирусных генов в клетках, позитивных по ядерному YВ-1, опосредуемая модифицированными или делетированными продуктами гена Е1А, приводит к успешной репликации и лизису таких клеток в присутствии YВ-1 в их ядре, включая, например, полирезистентные клетки, и что описанные здесь аденовирусы могут быть также использованы для лизиса таких опухолей.
Пример 7: Повышение эффективности репликации аденовирусов Е1-минус
В этом примере показано, что ранние вирусные гены Е1В-55К и Е4orf6 могут быть заменены посредством трансфекции плазмидой рЕ4orf6 и посредством инфицирования Е1/Е3-делетированным аденовирусом Аd-55К. Аd-55К представляет собой Е1/Е3-делетированный вирус, где ген Е1В-55К клонирован в Е1 и находится под контролем CMV. Эта замена была необходима для того, чтобы АdYВ-1, то есть аденовирус, экспрессирующий YВ-1, не экспрессировал эти ранние гены, и чтобы автор настоящего изобретения мог удостовериться, что замена этих ранних генов в системе репликации, содержащей ядерный YВ-1, может повышать эффективность репликации и эффективность образования вирусных частиц соответственно до уровня, сравнимого с уровнем эффективности аденовирусов дикого типа, таких как Аd5.
Для этого осуществляли следующую процедуру:
Все 105 клеток U2ОS трансфицировали плазмидой рЕ4orf6 с использованием липофектамина. Плазмида рЕ4orf6 несет ДНК-последовательность, кодирующую ранний вирусный ген Е4orf6, находящийся под контролем CMV.
Через 24 часа после трансфекции плазмидой рЕ4orf6 клетки инфицировали YВ-1-экспрессирующим Е1/Е3-делетированным аденовирусом АdYВ-1 (50 б.о.е./клетку) и Е1/Е3-делетированным аденовирусом Аd-55К, содержащим ген Е1В-55kDa, (50 б.о.е./клетку). Вирус Аd-55К представляет собой Е1/Е3-делетированный вирус, несущий в качестве трансгена вирусный ген Е1В-55К, находящийся под контролем CMV.
Затем, через 5 дней после инфицирования (=post infectionem), клетки удаляли из среды (2 мл). Высвобождение вирусных частиц из выделенных клеток проводили путем трехкратного поочередного замораживания и оттаивания (цикл оттаивания/замораживания). После этого клетки 293 подвергали анализу методом бляшек в целях выявления образовавшихся инфекционных частиц (бляшкообразующих единиц на мл (б.о.е./мл)). Результаты представлены на фиг. 8 и 9. На фиг.8 представлен результат анализа методом бляшек, выраженный в абсолютных величинах. Наиболее значимое различие по сравнению с инфицированием, проводимым лишь одним АdYВ-1, обнаруживалось при трансфекции плазмидой рЕ4orf6 и при ко-инфицировании двумя вирусами АdYВ-1 и Аd-55К. На фиг.9 представлен результат фиг.8, где показано, во сколько раз эффективность репликации превышает эффективность репликации, определенной для АdYВ-1. Клетки, инфицированные плазмидой рЕ4orf6, а затем АdYВ-1 и Е1В-55К (Аd-55К), продуцировали в 25 раз большее количество б.о.е./мл.
На основании этих результатов можно сделать вывод, что замена на Е1В-55К и Е4orf6 приводит к 25-кратному увеличению числа образующихся вирусов (б.о.е./мл) после инфицирования Е1/Е3-делетированным аденовирусом АdYВ-1. Аддитивное действие Е1В-55К и Е4orf6 на продуцирование бляшкообразующих единиц (б.о.е.) значительно превышало действие каждого из указанных двух генных продуктов.
Контрольные эксперименты, проводимые для одной плазмиды, экспрессирующей ЕGFP, ясно указывали на то, что в этом выбранном экспериментальном методе только 10% клеток были успешно трансфецированы плазмидой рЕ4orf6. Число частиц, образующихся в клетках, которые экспрессировали как Е1В-55К, так и Е4orf6, было сравнимо с числом частиц человеческого аденовируса типа 5 (дикого типа). Это подтверждает открытие, лежащее в основе настоящего изобретения, а именно, что экспрессия Е4orf6 и Е1В-55К, в сочетании с ядерной локализацией YВ-1, обеспечивает репликацию аденовируса и образование вирусных частиц, а в частности Е1А-делетированных аденовирусов, сравнимые с репликацией и образованием вирусных частиц одного из аденовирусов дикого типа, Аd5.
Пример 8: Повышенная репликации аденовирусов, которые не реплицируются в клетках, негативных по ядерному YВ-1, в клетках, позитивных по ядерному YВ-1, после введения цитостатических средств
Как известно из предшествующих работ, введение различных цитостатических средств индуцирует ядерную локализацию человеческого фактора транскрипции YВ-1. Как было обнаружено автором настоящего изобретения, фактор YВ-1, локализованный в ядре, регулирует репликацию аденовируса посредством активации аденовирусного Е2-позднего промотора. Комбинация обоих эффектов может быть использована для обеспечения специфического лизиса опухоли.
Для осуществления анализов на онколизис проводили следующую процедуру: 200000 клеток (HeLa и U2OS, соответственно) высевали в каждую лунку 6-луночного планшета. На следующий день добавляли 40 нг/мл (конечная концентрация) даунорубицина. После инкубирования в течение 3 часов клетки инфицировали 10 и 30 б.о.е. dl520/клетку соответственно. Затем клетки инкубировали в среде, не содержащей цитостатического средства. Через 3-5 дней клетки окрашивали кристаллическим фиолетовым.
Как видно на фиг. 10 и 11, введение даунорубицина индуцирует репликацию dl520 посредством ядерной локализации YВ-1. Таким образом, dl520, взятый в комбинации с цитостатическим средством и с даунорубицином, обладает значительно более сильным онколитическим действием по сравнению с одним даунорбицином.
Пример 9: In vivo-лизис опухоли посредством dl520
Клетки НеLa (негативные по ядерному YВ-1) и клетки 257RDB (позитивные по ядерному YВ-1), используемые в данном in vivo исследовании, размножали в стерильных условиях для культивирования клеток. Перед инъекцией этих клеток мышам (штамм CD1NuNu) для образования подкожной опухоли клетки собирали путем трипсинизации, растворяли в среде DMEM (10% FCS), подсчитывали и один раз промывали PBS. Затем клетки центрифугировали, PBS отсасывали и эти клетки распределяли в свежем PBS в нужном количестве. В данном исследовании число подкожно инъецированных клеток составляло 5×106 для обеих клеточных линий. Инъецирование проводили подкожно в один бок животных, причем для лучшей идентификации клетки НеLa инъецировали в правый бок, а клетки 257RDB инъецировали в левый бок. Рост опухолей контролировали два раза в неделю и при этом измеряли длину и ширину опухоли с использованием штангенциркуля с нониусом. Исходя из результатов этих измерений объем опухоли вычисляли по следующей математической формуле:
3/4π·а/2·(b/2)2,
где а=длина, b=ширина.
После того как опухоль достигала объема 200-520 мм3, в эту опухоль вводили вирус и PBS в качестве негативного контроля соответственно. Инъецируемые объемы были идентичными и каждый из них составлял по 50 мкл. Эту процедуру повторяли в течение 3 последующих дней. Общая доза вводимых вирусов составляла 5×108 б.о.е. Рост опухоли регистрировали два раза в неделю и вычисляли ее объем. По окончании исследования мышей умерщвляли, и опухоли удаляли для последующего анализа.
Результаты представлены на фиг. 12 и 13.
На фиг.12 представлен график, иллюстрирующий объем опухоли в зависимости от времени и различных схем обработки. В случае образования опухоли клетками RDB257, после инъекции PBS наблюдался значительный рост опухоли примерно от 438 мм3 до 1466 мм3. Под влиянием вектора dl520, используемого в настоящем изобретении, рост опухоли обнаруживал значительное снижение. Исходя из среднего размера опухолей, составляющего 344 мм3, размер опухоли увеличился лишь на 21%, то есть составлял всего 543 мм3.
В примере настоящего изобретения опухоль, состоящая из клеток НеLa, была использована в качестве контроля, и после введения PBS вела себя так же, как и опухоль, образованная клетками RDB257, после введения PBS. Однако опухоли, образованные клетками НеLa и обработанные dl520, еще обнаруживали значительный рост, и их объем, исходя из начального объема 311 мм3, достигал 1954 мм3.
На фиг.13 представлена фотография умерщвленных мышей с растущей опухолью, образованной RDB257. На этой фотографии хорошо видно, что после введения аденовируса dl520 в соответствии с настоящим изобретением наблюдается значительное снижение роста опухоли. В данном случае наблюдалось даже снижение объема опухоли (на 1-й день после введения вируса dl520 этот объем составлял 515 мм3, а на 30-й после введения вируса dl520 объем опухоли составлял 350 мм3).
Пример 10: Саузерн-блот-анализ опухолевой ДНК
ДНК экстрагировали из образца опухоли, который был взят из средней части опухоли, продуцированной как описано в примере 9. Для выделения образца использовали набор, предназначенный для выделения тканей (Dneasy Tissue Kit of Qiagen). ДНК выделяли в соответствии с инструкциями производителей. В соответствии с этим, ДНК выделяли из клеток путем щелочного лизиса. После этого выделенную ДНК очищали на колонке. Затем концентрацию выделенной ДНК определяли путем фотометрии при 260 нм. Анализ проводили с использованием 2 мкг образцов ДНК, которые гидролизовали 10 единицами рестриктирующего фермента Kpn I. Полученные образцы подвергали электрофоретическому разделению в 0,8% агарозном геле. После этого ДНК блоттировали на найлоновой мембране (по системе, описанной Schleicher & Schuell). ДНК, блоттированную на мембране, гибридизовали со специфическим 1501 п.н.-ДНК-зондом. Этот 1501 п.н.-ДНК-зонд специфически связывался с 3369 п.н.-Kpn I-фрагментом в Е2А-кодирующей последовательности Ad5. Зонд получали заранее с помощью ПЦР (праймер: 5'- GTC GGA GAT CAG ATC CGC GT (SEQ ID NO:2), 5'-GAT CCT CGT CGT CTT CGC TT (SEQ ID NO:3)) и радиоактивно метили 32Р. Затем мембрану промывали и экспонировали с пленкой.
Результат Саузерн-блот-анализа опухолевой ДНК представлен на Фиг.14. Этот анализ подтвердил, что в резистентных клетках RDB257 in vitro реплицировался только dl250, как показано на дорожках 3, 4 и 5. Дорожка 1 относится к позитивному контролю Аd-5d, а дорожки 6, 7 и 8 относятся к ДНК клеток НеLa, которые были инфицированы dl520. Поскольку клетки НеLa не являются позитивными по ядерному YВ-1, то вирус dl520 не реплицировался, а поэтому последовательность Е2А не детектировалась.
Другой результат, полученный для dl520, представлен на фиг.15. С помощью анализа методом бляшек проводили исследование на образование вирусных частиц (б.о.е./мл) после инфицирования аденовирусом dl520 и аденовирусом дикого типа. Различные опухолевые клетки, позитивные по ядерному YВ-1 (257RDB и 181RDB), и опухолевые клетки, негативные по ядерному YВ-1, были инфицированы аденовирусом dl520 и аденовирусом дикого типа.
Для этого проводили следующую процедуру:
100000-200000 клеток высевали в так называемые планшеты, имеющие 6 лунок (6-луночные планшеты) в среде L15 (резистентные клетки) и среде DMEM (нерезистентные клетки), содержащей 10% FCS. Через 24 часа проводили инфицирование аденовирусом dl520 и аденовирусом дикого типа (10 б.о.е./клетку). Через 3 дня после инфицирования (post infectionem) вирусные частицы выделяли из клеточной суспензии (3 мл) путем проведения трехкратного поочередного замораживания и оттаивания. После этого клетки 293 подвергали анализу методом бляшек в целях выявления образовавшихся инфекционных частиц (бляшкообразующих единиц на мл (б.о.е./мл). Результаты представлены на фиг.15. Результат анализа методом бляшек показал, что dl520 реплицировался в позитивных по ядерному YВ-1 клетках (257RDB и 181RDB) аналогично аденовирусу дикого типа. Таким образом, с использованием описанных здесь аденовирусов в настоящем изобретении, может быть достигнута эффективная репликация, аналогичная репликации одного из аденовирусов дикого типа.
Пример 11: Структурная конструкция аденовирусного вектора Xvir03
На фиг.16 представлена структурная конструкция аденовирусного вектора Xvir03. Аденовирус Xvir03 представляет собой так называемый Е1/Е3-делетированный аденовирус. Это означает, что в нем отсутствовали белки Е1А, Е1В и Е3, которые являются функциональными в отношении репликации аденовируса. Делеция области Е1 простиралась от аминокислоты 342 до аминокислоты 3528, а делеция области Е3 простиралась от аминокислоты 27865 до аминокислоты 30995. Используемый здесь термин “Е1-делетированный вирус” означает вирус, в котором Е1 больше не является функционально активным. Это может быть достигнуто путем инактивации с использованием какой-либо другой наиболее интактной последовательности нуклеиновой кислоты и аминокислотной последовательности, однако это также может быть достигнуто путем делеции области Е1, кодирующей белки, имеющие разные размеры. Из-за отсутствия белков Е1А и Е1В и нуклеиновых кислот, кодирующих эти белки, область Е4, такая как Е4orf6, либо очень слабо экспрессируется (примерно на 1-5% по сравнению с аденовирусами дикого типа), либо вообще не экспрессируется. Вирусные гены Е1В-55К и Е4orf6 экспрессируются в области Е1 посредством гетерологичного промотора CMV (Clontech: Plasmid pShuttle), встроенного в Xvir03. Вместо промотора CMV может быть использован любой из промоторов, описанных в настоящей заявке в связи с экспрессией Е1А. Открытые рамки считывания обоих генов присоединены друг к другу посредством так называемой последовательности IRES (внутреннего сайта связывания с рибосомой) (Pelletier J. and Sonenberg, N. Nature, 1988, 334, 320-325). Этот элемент (Novagen: рCITE) обеспечивает экспрессию 2 белков из одной мРНК.
Конструирование вектора
Плазмиду Е1В55k-pShuttle получали путем клонирования открытой рамки считывания Е1В55k из рСGNE1B от M. Dobelstein (University of Marburg) с помощью ферментов ХbaI и BfrI в вектор pShuttle (Clontech). Затем Е1В55k в pShuttle линеаризовали ферментом ApaI, затупляли по концам и разрезали ферментом NheI.
Во втором векторе плазмиду рсDNA3.1(+)(Invitrogen), за которой следовал элемент IRES, используемый в качестве ПЦР-продукта, клонировали с использованием рСIТЕ-4а(+) (от компании Novagen) в качестве матрицы путем ТА-клонирования в рестрикционный EcoRV-сайт, а Е4orf6, взятый из плазмиды рCMV-Е4orf6 (М.Dobelstein, University of Marburg), клонировали с помощью BamHI, в результате чего получали IRES-Е4orf6-рсDNA3.1(+). IRES-Е4orf6 в рсDNA3.1(+) линеаризовали ферментом NotI, затупляли по концам, а затем фрагмент IRES-Е4orf6 разрезали ферментом NheI. Фрагмент IRES-Е4orf6 присоединяли к открытому вектору Е1В55k-pShuttle (затупленному, NheI). Затем эту кассету клонировали из Е1В55k-IRES-Е4orf6-pShuttle вместе с промотором CMV и конъюгатом “бычий гормон роста-(ВGH)-polyA” в плазмиду ΔЕ1, ΔЕ3-Adeno-X (Clontech) с использованием ферментов I-Ceu I и PI-SceI, и полученную конструкцию обозначали AdcmvE1B/IRES/E4orf6. Затем конструировали аденовирус в соответствии с инструкциями производителя (Clontech). Адено-плазмиду, линеаризованную ферментом РасI и имеющую экспрессионный элемент CMV-Е1В55k-IRES-Е4orf6-ВGH-polyA, трансфицировали в клетки НЕК293 и через 11 дней после трансфекции абляционные клетки удаляли вместе со средой для высвобождения аденовирусов путем проведения нескольких циклов замораживания-оттаивания.
Вышеописанный вектор, как и другие описанные здесь вирусы, в принципе, являются подходящими для использования в настоящем изобретении. В частности, вышеописанный вектор является подходящим для репликации и стимуляции лизиса, например, в клетках, позитивных по ядерному YВ-1, а также в клетках, где нарушена регуляция YВ-1, то есть в клетках, где по сравнению с нормальными клетками и неопухолевыми клетками наблюдается сверхэкспрессия YВ-1 соответственно. В частности, этот вектор может быть использован для лечения таких же заболеваний или таких же групп пациентов или групп пациентов с одним и тем же заболеванием, которые были упомянуты выше в связи с применением других аденовирусов, используемых в настоящем изобретении, и других аденовирусов, описанных в настоящем изобретении.
Пример 12: Структурная конструкция аденовирусного вектора Xvir03/01
Как можно видеть на фиг.17, Xvir03/01 представляет собой другое производное Xvir03. Терапевтические гены, такие как, например, описанные здесь гены и трансген могут быть клонированы в область Е3. Кроме того, во избежание гомологичной рекомбинации с Е4orf6 из экспрессионной кассеты Xvir03, была введена делеция в область Е4. Это позволяет клонировать в данную конструкции более крупные трансгены. Делетированная область Е3 содержит рестрикционные SacI-, NdeI и NheI-сайты для введения кассеты, в которую могут быть, например, клонированы терапевтические трансгены.
Получение плазмиды для клонирования терапевтических генов в область Е3, а также для создания делеций в области Е4
рAdeno-X-плазмида от Clontech имеет рестрикционный сайт для фермента SfuI, расположенный за 3'ITR-областью, которая отсутствует в аденовирусе дикого типа. Е3-Е4-область была вырезана из рAdenoX (Clontech) с помощью фермента SрeI (положение 23644) и фермента SfuI и перенесена в pcDNA3.1(+) (Invitrogen) с получением pcDNA3.1-E3Δ27865-30995-Е4. Большую часть Е4ORF6, а именно 33241-33875, удаляли с использованием PstI и получали pcDNA3.1-E3Δ27865-30995, Е4Δ33241-33875. Для проведения последующей модификации Xvir03, Е3/Е4-делетированную область, вырезанную из pcDNA3.1-E3Δ27865-30995, Е4Δ33241-33875, клонировали с помощью ферментов SfuI и SpeI в плазмиду pAdenoX и получали pAdenoXE3Δ27865-30995, Е4Δ33241-33875.
Затем экспрессионную кассету так же, как было описано для Xvir03, клонировали с помощью I-Ceu I и PI-SceI из E1B55k-IRES-E4orf6-pShuttle вместе с промотором CMV и конъюгатом “бычий гормон роста-(ВGH)-polyA” в плазмиду pAdenoX-E3Δ27865-30995, Е4Δ33241-33875, и полученную конструкцию обозначали AdcmvE1B/IRES/E4orf6-ΔЕ4. Затем был сконструирован аденовирус в соответствии с инструкциями производителей (Clontech).
Вышеописанный вектор, как и другие описанные здесь вирусы, в принципе, может быть использован в настоящем изобретении. В частности, вышеописанный вектор является подходящим для репликации в клетках, позитивных по ядерному YВ-1, а также в клетках, в которых нарушена регуляция YВ-1, то есть в клетках, где, по сравнению с нормальными клетками и неопухолевыми клетками, наблюдается сверхэкспрессия YВ-1, а поэтому он является подходящим для лизиса клеток. В частности, этот вектор может быть использован для лечения таких заболеваний или таких групп пациентов или групп пациентов с тем же самым заболеванием пациентов, которые были упомянуты выше в связи с применением других аденовирусов, используемых в настоящем изобретении, и других аденовирусов, описанных в настоящем изобретении.
Пример 13: Онколитический эффект Xvir03 в клетках 257RDB и 181RDB
Высевали 100000 клеток (257RDB и 181RDB) на лунки планшета, имеющего шесть лунок (6-луночного планшета). На следующий день клетки, как показано на фиг.18, инфицировали Аd312 (20 б.о.е./клетку) и Xvir03 (5 б.о.е./клетку). Инфицирование проводили в 500 мкл бессывороточной среды DMEM при 37°С в течение 1 ч. Затем инфекционную среду удаляли и заменяли 2 мл полной среды (10% FCS/DMEM). Анализ проводили через 5 дней путем окрашивания кристаллическим фиолетовым. Результат представлен на фиг. 18А и 18В.
Как видно на фиг. 18А и 18В, полирезистентные клетки, имеющие YВ-1 в ядре, подвергались лизису после инфицирования Аd312 и Xvir03 только в случае Xvir03, на что указывало окрашивание клеток кристаллическим фиолетовым. Для этого сначала среду удаляли. Затем клетки покрывали кристаллическим фиолетовым (50% ЕТОН, 3% формальдегид, 5% уксусная кислота, 1% кристаллический фиолетовый) и инкубировали при комнатной температуре в течение 5-10 минут. После этого 6-луночные планшеты тщательно ополаскивали водой и сушили при комнатной температуре.
Автору настоящего изобретения известно, что Е1А-делетированные вирусы (например, Аd312), которые не являются трансактивирующими аденовирусами в соответствии с настоящим изобретением, могут, однако, с высокой эффективностью реплицироваться при более высоких m.o.i. (Nevins J.R., Cell 26, 213-220, 1981), но они тем не менее не могут быть использованы в клинических целях. В литературе такой феномен известен как “Е1А-подобная активность”. Используемым здесь аденовирусом Аd312 является Е1А-делетированный вирус. При используемом титре (20 б.о.е./клетку), который был еще выше, чем клинически предпочтительный титр, ранние аденовирусные гены, такие как Е1В55k и Е4orf6, либо вообще не экспрессировались, либо экспрессировались лишь на очень незначительном уровне (Nevins J.R., Cell 26, 213-220, 1981). Как уже было описано выше, эти гены и белки играют важную роль в репликации вируса. В противоположность этому, указанные гены и белки соответственно экспрессировались аденовирусом Xvir03 (фиг.16). Как можно видеть на фиг. 18А и 18В, экспрессия генов Е1В-55К и Е4orf6 будет приводить к эффективной репликации вируса и к лизису клеток, и к тому же при более низком требуемом титре инфекционности (выраженном в б.о.е./клетку). Это подтверждает утверждение, лежащее в основе настоящего изобретения и заключающееся в том, что экспрессия Е4orf6 и Е1В-55К (в отсутствии Е1А) в сочетании с ядерной локализацией YВ-1, способна индуцировать в высокой степени эффективную репликацию аденовируса. Необходимый титр аденовируса, который позволяет использовать его в клинических целях, составляет всего лишь 1-5 б.о.е./клетку.
Таким образом, подтверждается вывод, лежащий в основе настоящего изобретения и заключающийся в том, что для получения вирусов, которые могут быть использованы в настоящем изобретении, и для лизиса инфицированных клеток необходимо присутствие в их ядре YВ-1, а в частности, такое присутствие должно быть независимым от клеточного цикла.
Пример 14: Нозерн-блот-анализ экспрессии гена Е2 аденовируса Аd312
В каждом случае 1 миллион клеток А549 и U2OS высевали в 10 см-чашки Петри. На следующий день клетки инфицировали Аd312 (50 б.о.е./клетку) и Adwt (которыл служил в качестве контроля, 5 б.о.е./клетку). Высокий титр используемого вируса Аd312 давал Е1-независимую репликацию в опухолевых клетках. Инфицирование проводили в 1-2 мл бессывороточной среды DMEM в течение 1 часа при 37°С. Затем инфекционную среду удаляли и заменяли 10 мл полной среды (10% FCS/DMEM). Через 3 дня РНК выделяли. Затем концентрацию выделенной РНК определяли на фотометре при 260 нм. После этого образцы РНК подвергали электрофоретическому разделению в 0,8% формальдегидном агарозном геле. Затем РНК блоттировали на найлоновой мембране (по системе, описанной Schleicher & Schuell). РНК, блоттированную на мембране, гибридизовали с “ранним зондом” Е2 и “поздним зондом” Е2. Этот 1501 п.н.-”поздний зонд” специфически связывался на участке за Е2-поздним промотором. Зонд получали заранее с помощью ПЦР (праймер: 5'- GTC GGA GAT CAG ATC CGC GT (SEQ ID NO:4), 5'-GAT CCT CGT CGT CTT CGC TT (SEQ ID NO:5)) и радиоактивно метили 32Р. В противоположность этому, ранний зонд связывался на участке между Е2-ранним промотором и Е2-поздним промотором (положения: 226791-227002) и также был получен с помощью ПЦР (праймер: 5'- AGCTGATCTTCGCTTTTG (SEQ ID NO:6), 5'-GGATAGCAAGACTCTGAC AAAG (SEQ ID NO:7)) Затем мембрану промывали и экспонировали с пленкой.
Результат анализа представлен на фиг.19. Как ранний, так и поздний зонды продуцировали специфические сигналы при контрольном инфицировании аденовирусом дикого типа, тогда как опухолевые клетки, инфицированные Аd312, продуцировали специфический сигнал только при использовании позднего зонда. Этот анализ подтвердил открытие, лежащее в основе настоящего изобретения, и заключающееся в том, что экспрессия Е4orf6 и Е1В55К и отсутствие Е1А обеспечивают транспорт сверхэкспрессируемого YВ-1 и YВ-1 с нарушенной регуляцией соответственно в клеточное ядро, и тем самым индуцируют экспрессию гена Е2, которая является необходимым условием эффективной репликации аденовируса.
Пример 15: Нозерн-блот-анализ экспрессии гена Е2 аденовируса Аd-delta24
В каждом случае 1 миллион клеток U2OS высевали в 10 см-чашки Петри. На следующий день клетки инфицировали аденовирусом delta24 (Аddelta24) (10 б.о.е./клетку) и аденовирусом дикого типа (Adwt) (который служил в качестве контроля, 10 б.о.е./клетку). Используемый рекомбинантный аденовирус Аddelta24 (Fueyo, J. et al., Oncogene, 19, 2-12, 2000) имел специфическую делецию в области CR2 белка Е1А, а поэтому он мог реплицироваться только в Rb-негативных опухолях. Кроме того, этот вирус экспрессировал гены Е1В55k и Е4orf6 на уровне, сравнимом с аденовирусом дикого типа. Инфицирование проводили в 1-2 мл бессывороточной среды DMEM в течение 1 часа при 37°С. Затем инфекционную среду удаляли и заменяли 10 мл полной среды (10% FCS/DMEM). Через 12 ч и 24 ч РНК выделяли. Затем концентрацию выделенной РНК определяли на фотометре при 260 нм. После этого образцы РНК подвергали электрофоретическому разделению в 0,8% формальдегидном агарозном геле. Затем РНК блоттировали на найлоновой мембране (по системе, описанной Schleicher & Schuell). РНК, блоттированную на мембране, гибридизировали с “ранним зондом” и “поздним зондом”. Этот “поздний зонд”, содержащий 1501 п.н., специфически связывался на участке за Е2-поздним промотором. Зонд получали заранее с помощью ПЦР (праймер: 5'- GTC GGA GAT CAG ATC CGC GT (SEQ ID NO:4), 5'-GAT CCT CGT CGT CTT CGC TT (SEQ ID NO:5)) и радиоактивно метили 32Р. Однако ранний зонд связывался на участке между Е2-ранним промотором и Е2-поздним промотором и также был получен с помощью ПЦР (праймер: 5'- AGCTGATCTTCGCTTTTG (SEQ ID NO:6), 5'-GGATAGCAAGACTCTGACAAAG (SEQ ID NO:7)). Затем мембрану промывали и экспонировали с пленкой.
Результат анализа представлен на фиг.20.
Через 12 часов только поздний зонд продуцировал специфический сигнал. А уже через 24 часа ранний зонд также продуцировал сигнал в клетках, инфицированных Аddelta24. Однако по сравнению с аденовирусами дикого типа такой сигнал был значительно слабее. Этот результат также подтверждает открытие, лежащее в основе настоящего изобретения и заключающееся в том, что экспрессия Е4orf6 и Е1В-55К обеспечивает транспорт в клеточное ядро сверхэкспрессируемого YВ-1 и YВ-1 с нарушенной регуляцией соответственно, которые затем связываются с Е2-поздним промотором и индуцируют экспрессию гена Е2.
Пример 16: Структурное конструирование аденовирусных векторов XvirPSJL1 и XvirPSJL2
Описание векторов: Векторы группы XvirPSJL, которые представляют собой варианты вирусов, именуемых здесь аденовирусами группы I, и которые представлены здесь векторами и аденовирусами XvirPSJL1 и XvirPSJL2 соответственно, способны реплицироваться, подобно аденовирусу dl520, не только в позитивных по ядерному YВ-1 клетках, а, в частности, в опухолевых клетках, но также и в опухолевых клетках, в которых присутствует сверхэкспрессируемый YВ-1 и YВ-1 с нарушенной регуляцией, соответственно. Вирусные гены Е1В55k и Е4orf6 экспрессируются только в dl520-инфицированных клетках, позитивных по ядерному YВ-1, под контролем промотора Е1В и промотора Е4, соответственно, а экспрессия Е1В55k и Е4orf6 в XvirPSJL происходит под контролем промотора цитомегаловируса (CMV). Однако вместо промотора CMV могут быть использованы и другие промоторы, а в частности опухолеспецифические, тканеспецифические и органоспецифические промоторы. Благодаря экспрессии Е1В55k и Е4orf6, сверхэкспрессируемый YВ-1 и YВ-1 с нарушенной регуляцией соответственно, транспортируются в ядро и инициируют репликацию аденовируса. Аденовирусные векторы группы XvirPSJL, описанные в данной заявке, объединяют различные элементы и функции аденовирусных векторов dl520, Xvir03 и АdYВ-1 в одном векторе. Подобно вектору dl520 вирусы XvirPSJL содержат ген Е1А12S. Этот ген и соответствующий генный продукт, соответственно, ответственны за индуцирование фазы S инфицированной клетки, а также стимулируют репликацию вируса и улучшают эффект от проведения химиотерапии и облучения. Аналогично Xvir03, вирусы XvirPSJL содержат экспрессионную кассету CMV-Е1В55k/IRES/Е4orf6, которая необходима для эффективной репликации и опосредованного или прямого транспорта YВ-1 с нарушенной регуляцией в ядро, который, предпочтительно, содержится в опухолевых клетках. Таким образом, репликация возможна только в клетках, а предпочтительно, в опухолевых клетках, в которых сверхэкспрессируется YВ-1 или присутствует YВ-1 с нарушенной регуляцией. Кроме того, Р53 подвергали деградации с помощью комплекса Е1В55k/Е4orf6. Последовательность, кодирующую человеческий фактор транскрипции YВ-1 получали из вируса АdYВ-1. Эндогенный, то есть уже присутствующий в клетке YВ-1 усиливает репликацию вируса. Экспрессия как Е1А12S, так и YВ-1 регулируется YВ-1-зависимым аденовирусным Е2-поздним промотором. В соответствии с этим, могут быть также использованы специфические промоторы, а в частности опухолеспецифические, тканеспецифические или органоспецифические промоторы. Другим отличительным признаком этих вирусов является то, что в этих вирусах область Е4 делетирована. Этот вектор содержит рестрикционные сайты, посредством которых, в случае аденосирусных векторов XvirPSJL1 и XvirPSJL2, могут экспрессироваться различные трансгены, описанные в данной заявке, такие как рибозимы, антисмысловые молекулы, киРНК, апоптоз-индуцирующие гены, цитокины и гены пролекарств. Их экспрессия может также регулироваться опухолеспецифическими, тканеспецифическими или органоспецифическими промоторами, описанными в настоящей заявке. Локализация экспрессионных кассет не является фиксированной, а особенно, если это не относится к областям Е1, Е3 и Е4 или к областям, находящихся внутри них, и они могут иметь любое расположение. Эти векторы реплицируются независимо от р53- или Rb-статуса опухолевых клеток.
Структурные конструкции рекомбинантных аденовирусов XvirPSJL1 и XvirPSJL2 представлены на фиг. 21 и 22.
Генерирование кассеты Е2-late-YB1IRES/12S
рAdeno-X-плазмида от Clontech/BD Biosciences, которая была использована в качестве описанного здесь исходного материала, содержала геномную нуклеиновую кислоту аденовируса Аd5 и имела рестрикционный SfuI-сайт, расположенный за 3'ITR-областью, которая отсутствует в аденовирусе дикого типа. Е3-Е4-область переносили с помощью фермента SрeI (положение 23644) и фермента SfuI из рAdenoX (Clontech) в pcDNA3.1(+) (Invitrogen) и обозначали pcDNA3.1-E3Δ27865-30995-Е4. Большую часть Е4ORF6, а именно нуклеотиды 33241-33875, удаляли с использованием PstI. Полученный таким образом фрагмент обозначали pcDNA3.1-E3Δ27865-30995, Е4Δ33241-33875.
Е2-поздний промотор вырезали из рGL3-EGFР (Holm P.S. et al., JBC 2002, 277, 10427-10434) ферментами SacI и NheI и клонировали в pcDNA3.1-E3Δ27865-30995, Е4Δ33241-33875. При этом область Е3 была дополнительно делетирована на участке нуклеотидов Δ27593-31509. Полученный таким образом фрагмент обозначали Е2-late- pcDNA3.1-E3Δ27593-31509, Е4Δ33241-33875.
кДНК для продукта Е1А-243АА генерировали с помощью ОТ-ПЦР, выделяли и последовательность секвенировали и клонировали в pcDNA3.1(+) (Invitrogen) с использованием ферментов BamHI и EcoRI. Е1А-12S-pcDNA3.1+ линеаризовали NheI и BamHI, затупляли по концам полимеразой Т4 и создавали выступающие Т-концы с помощью полимеразы Taq и dТТР. Элемент IRES клонировали в виде ПЦР-продукта (матрица=рCITE, Novagen) в вектор Е1А-12S-pcDNA3.1+ (стратегия ТА-клонирования).
YВ-1-EcoRI-фрагмент выделяли из вектора pHVad2c (Holm et al., JBC 2002, 277, 10427-10434) и затупляли по концам. Вектор pShuttle (коммерчески доступный, поставляемый фирмой BD Biosciences) линеаризовали ферментом XbaI, концы затупляли, а затем дефосфорилировали и лигировали с предварительно продуцированной YВ-1-кодирующей нуклеиновой кислотой. Полученный таким образом вектор обозначали YВ-1-рShuttle. Клонирование в вектор pShuttle приводило к получению нуклеиновой кислоты, кодирующей фрагмент YВ-1 со стоп-кодоном, находящимся внутри рамки считывания. YВ-1-кодирующую нуклеиновую кислоту клонировали из YВ-1-pShuttle с помощью ферментов NheI и BfrI в вектор IRES-E1A-12S в pcDNA3.1(+). Полученный таким образом фрагмент обозначали YВ-1 (EcoRI-EcoRI со стоп-кодоном)-IRES-E1A-12S-pcDNA3.1(+).
Затем кассету YВ-IRES-E1A12S вырезали ферментом PmeI и клонировали в NheI-линеаризованный затупленный по концам и дефосфорилированный вектор Е2late-pcDNA3.1-E3Δ27593-31509, Е4Δ33241-33875. Таким образом, вторая кассета находилась в делетированном участке области Е3.
Трансгенную кассету, содержащую конструкцию нуклеиновой кислоты Е2late-YВ-IRES-E1A12S, клонировали вместе с оставшимися аденовирусными последовательностями E3Δ27593-31509, Е4Δ33241-33875 с помощью ферментов SfuI и SpeI в вектор pAdenoX Clontech (=pAdenoX/Е2late-YВ-1-IRES-E1A12S/Е3Δ27593-31509, Е4Δ33241-33875).
Кассету CMV-Е1В55k/IRES/Е4orf6 вырезали ферментами I-CeuI и PI-Scel из вектора pShuttle, описанного выше для Xvir03, и встраивали в вектор AdenoX/Е2late-YВ-1-IRES-E1A12S/Е3Δ27593-31509, Е4Δ33241-33875.
Затем этот вектор линеаризовали ферментом Рас I, трансфецировали в клетки 293, и выделяли рекомбинантные аденовирусы XvirPSJL1 и XvirPSJL2 соответственно без трансгенов, как показано на чертеже, в соответствии с инструкциями производителя.
Пример 17: Инфицирование клеток НеLa аденовирусом dl520
100000 клеток НеLa высевали на чашку. На следующий день клетки инфицировали различными титрами (б.о.е./мл) аденовируса dl520. Инфицирование проводили в 500 мкл бессывороточной среды DMEM в течение 1 часа при 370 , затем инфекционную среду удаляли и заменяли 2 мл полной среды (10% FCS/DMEM). Через 3-5 дней проводили анализ путем окрашивания кристаллическим фиолетовым.
Результат этого эксперимента представлен на фиг.23. Аденовирус dl520 не обнаруживал какого-либо лизиса при низких m.o.i. (5-10 б.о.е./мл) после инфицирования клеток НеLa, не содержащих YВ-1 в ядре. В противоположность этому, dl520 обнаруживал фактически полный лизис при m.o.i. (множественности заражения) 100-200 б.о.е. на клетку и все еще преобладающий лизис при m.o.i. 50 б.о.е. на клетку. На основании этого можно сделать вывод, что dl520 и аналогичные вирусы, способные “запускать” экспрессию аденовирусных генов Е1В55k и Е4orf6 при более высоких m.o.i., являются подходящими для прямого или опосредованного транспорта сверхэкспрессируемого YВ-1 или YВ-1 с нарушенной регуляцией в ядро и тем самым для индуцирования клеточного лизиса.
Пример 18: Люциферазный анализ для определения активности Е2-позднего промотора
Известно, что YВ-1 связывается с аденовирусным Е2-поздним промотором в ядре (Holm et al., JBC 2002, 277, 10427-10434), и что этот промотор также является наиболее подходящим для экспрессии нуклеиновых кислот. Использование аденовирусного Е2-позднего промотора продиктовано, в частности, тем, что он может регулироваться YВ-1, причем YВ-1 действует как позитивный эффектор, то есть указанный промотор является активным лишь в присутствии YВ-1 в ядре. При этом указанный аденовирусный Е2-поздний промотор может быть в высокой степени селективно регулируемым, а поэтому он может быть использован в системах, в которых YВ-1 присутствует в ядре и фактически предотвращает любую экспрессию нуклеиновой кислоты, которая находится под контролем аденовирусного Е2-позднего промотора, в том случае, если YВ-1 не присутствует в ядре в качестве эффектора и регулятора соответственно. Е2-поздний промотор содержит 3 Y-бокса (ССААТ), которые являются подходящими для активации гена Е2. Были получены различные конструкции Е2-позднего промотора и эти конструкции были протестированы на их специфичность и активность. Анализ проводили как описано ниже.
Клеточные линии EPG-257 RDB (эпителиальной карциномы желудка), которые имеют YВ-1 в ядре, клеточные линии HeLa (эпителиальной карциномы шейки матки) и клеточные линии U2OS (остеосаркомы) высевали в 6-луночные планшеты при трех различных концентрациях клеток. На следующий день лунки, которые обнаруживали 70%-ную конфлюэнтность, использовали для трансфекции. Для каждой лунки 500 нг очищенной плазмидной ДНК различных конструкций Е2-позднего промотора (Spin Miniprep, Qiagen) в люциферазных векторах (коммерчески доступных и поставляемых фирмой Promega, исходная плазмида: pGL3-энхансер) добавляли в 500 мкл среды OptiMEM в реакционный 1,5 мл-сосуд с плотно закрывающейся крышкой и 5 мкл DOTAP добавляли в 500 мкл среды в другой реакционный сосуд с плотно закрывающейся крышкой. Оба раствора объединяли и смешивали. Смесь инкубировали в течение 30 минут при комнатной температуре для образования комплекса. Затем клетки три раза ополаскивали PBS и покрывали слоем смеси для трансфекции. Планшеты инкубировали при 37°С в течение 5 часов, а затем снова три раза ополаскивали PBS и загружали полную среду.
Через 48 часов после инфицирования клетки обрабатывали с помощью набора для люциферазного анализа, Luciferase Assay System Kit of Promega (Cat. № Е1500): каждую лунку покрывали слоем из 500 мкл буфера для лизиса, а затем клетки смывали с луночного планшета 1 мл-пипеткой через 10 минут при комнатной температуре, и переносили в реакционный сосуд с плотно закрывающейся крышкой емкостью 1,5 мл. Затем клеточный лизат центрифугировали при 4°С в течение 15 минут при 14000 об/мин. В каждые 50 мкл супернатанта добавляли 100 мкл люциферазного субстрата и проводили измерение на сцинтилляционном и люминисцентном счетчике TopCount (Canberra-Packard GmbH, 63303 Dreieich) Microplate Scintillation & Luminescence counter, в черных планшетах с 96 лунками при длине волны 945 нм.
Белки оценивали с помощью набора для анализа белков, ВСА Protein Assay Reagent Kit, каталожный номер 23227 (PIERCE, Rockford, Illinois, USA) при 570 нм в планшет-ридере для считывания показаний кинетической флуоресценции/оптической плотности (Molecular Dynamics) в биолюминометре (BioluminTM 960). Соответствующие световые сигналы образцов транслировали в соответствующее количество белка (RLU/мкг белка).
Были использованы следующие плазмиды: плазмида “pGL3-энхансер” (Promega), из которой был удален энхансер с помощью ферментов BamHI (2250 п.н.) и BsaBI (2003 п.н.) и которая служила для контрольного считывания. Различные конструкции промотора Е2 клонировали в MCS в векторе рGL3, не содержащем энхансера, с помощью рестрикционных ApaI- и SacI-сайтов. Промотор hCMV клонировали с помощью ферментов BglII и HindIII в энхансер рGL3, и этот промотор служил в качестве позитивного контроля. Такой позитивный контроль позволял оценивать эффективность трансфекции и также служил эталонной величиной люциферазной активности. Для каждой клеточной линии CMV-контроль принимали за 100%, а ферментативную активность, продуцируемую конструкциями промотора Е2, определяли по отношению к этой величине и представляли в виде гистограммы на фиг.24.
Различные конструкции обозначали следующим образом:
1) содержит Y-бокс I, II и III, соответствующий последовательности нуклеотидов 25932-26179 п.н. (называемой последовательностью аденовируса дикого типа, см. также часть приведенной далее области Е2);
2) содержит Y-бокс II и III, соответствующий последовательности нуклеотидов 25932-26127 п.н. (называемой последовательностью аденовируса дикого типа, см. также часть приведенной далее области Е2 аденовируса);
3) содержит Y-бокс III, соответствующий последовательности нуклеотидов 25932-26004 п.н. (называемой последовательностью аденовируса дикого типа, см. также часть приведенной далее области Е2 аденовируса);
4) не содержит Y-бокса и служит в качестве контроля.
Часть области Е2 аденовируса (взята из Virology 1992, 186, 280-285) (сайты связывания с YВ-1 показаны жирным шрифтом):



Результаты, представленные на фиг.24, убедительно подтверждают, что отдельные промоторные фрагменты, которые содержат различные Е2-late/Y-боксы, являются подходящими для экспрессии терапевтических трансгенов в опухолевых клетках, позитивных по ядерному YВ-1, а поэтому они могут быть использованы в качестве промоторов, определенных в настоящем изобретении.
Пример 19: Влияние YВ-1, экспрессируемого аденовирусом, на высвобождение частиц
Клетки человеческой остеосаркомы (U2OS) инфицировали Е1/Е3-делетированным аденовирусным вектором АdYВ-1, и Аd312, имеющим только делетированный Е1А, при m.o.i. 50 б.о.е./клетку. АdYВ-1 содержит в своем геноме последовательность, кодирующую клеточный фактор транскрипции YВ-1, а поэтому он экспрессирует Y-бокс-связывающий белок 1 (YВ-1). Для оценки уровня высвобождения вирусных частиц после инфекции в виде бляшкообразующих единиц (б.о.е.) супернатант культуральной среды или оставшийся клеточный слой выделяли через 2 дня и 5 дней после инфицирования соответственно. Высвобождение внутриклеточных частиц проводили за 3 цикла замораживания/оттаивания. Число частиц определяли с помощью анализа методом бляшек, проводимого на клетках 293.
Результаты представлены на фиг.25, где сплошные гистограммы обозначают число оставшихся внутриклеточных вирусных частиц, а косо заштрихованные гистограммы обозначают высвобожденные внеклеточные вирусные частицы.
Результаты, представленные на фиг.25, подтверждают, что АdYВ-1 в целом продуцирует больше б.о.е., чем Аd312, и высвобождает больше частиц. Через 5 дней АdYВ-1-инфицированные клетки явно обнаруживали цитопатический эффект (СРЕ) в отличие от Аd312-инфицированных клеток.
Отличительные признаки настоящего изобретения, описанные в данной заявке, формула изобретения, а также графический материал, взятые отдельно или в любой их комбинации, имеют важное значение для реализации различных вариантов настоящего изобретения.
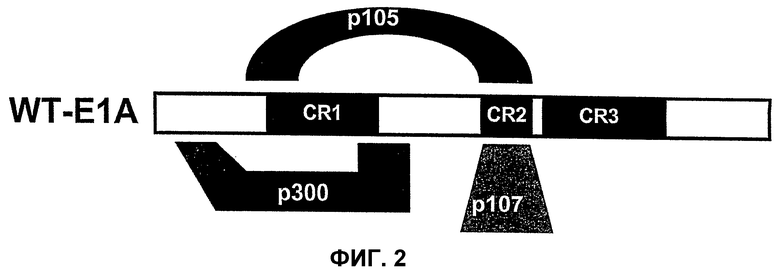
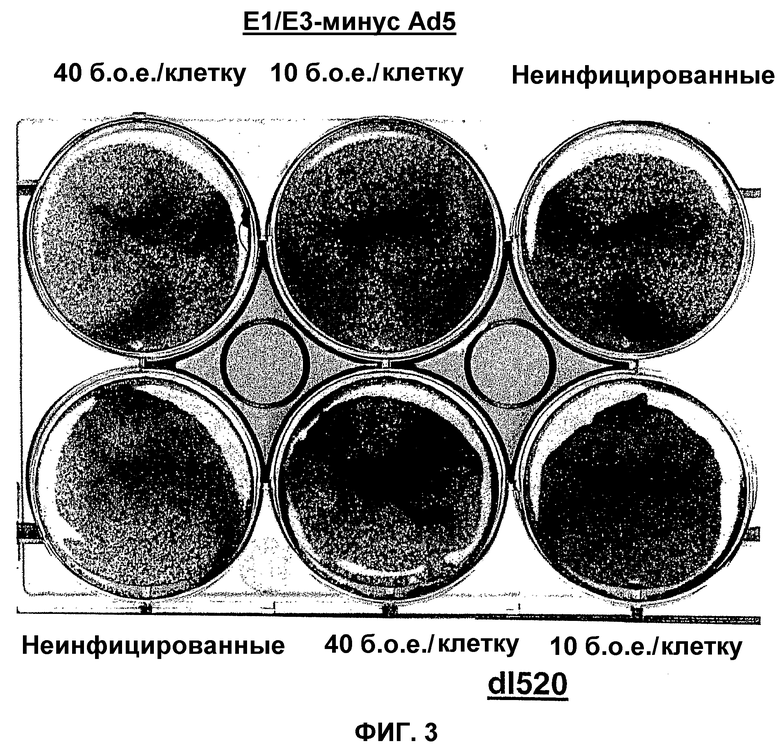
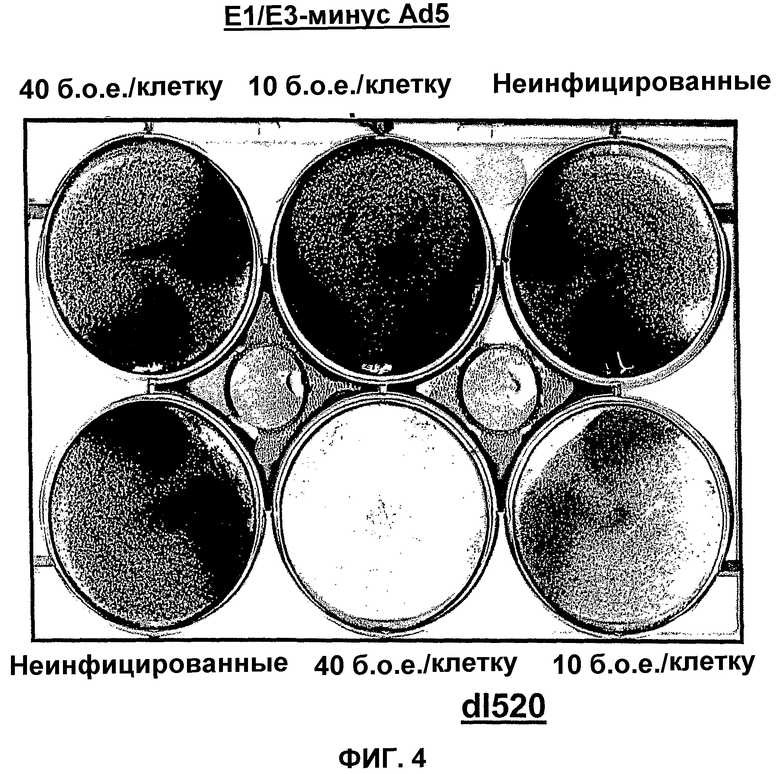
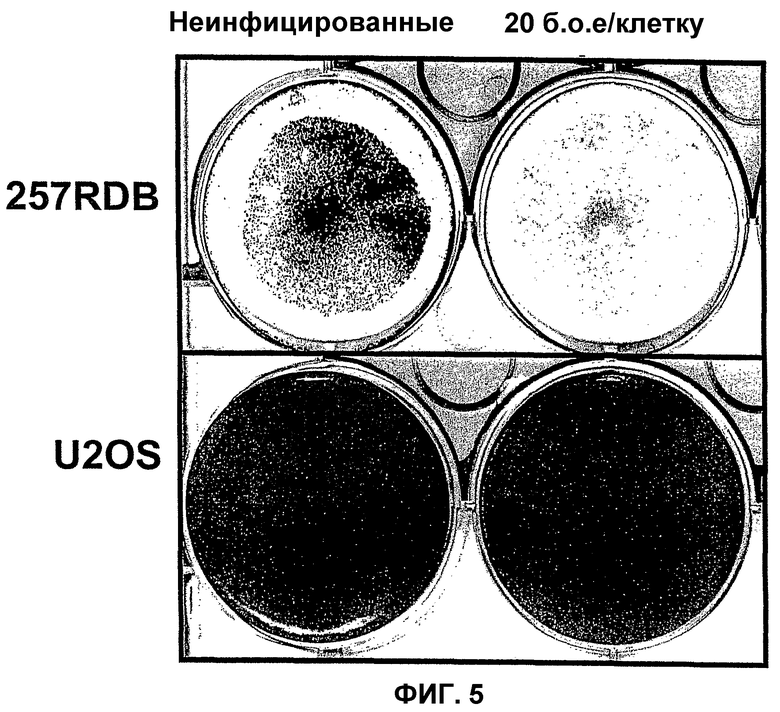
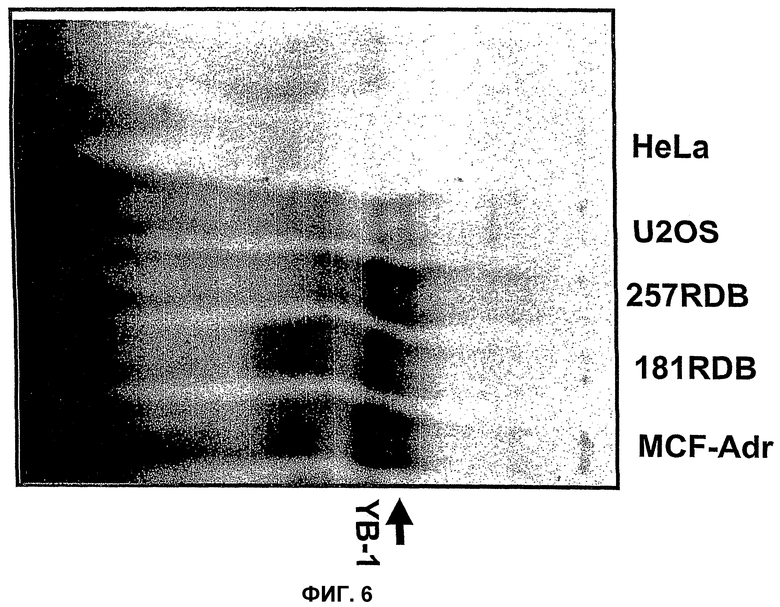
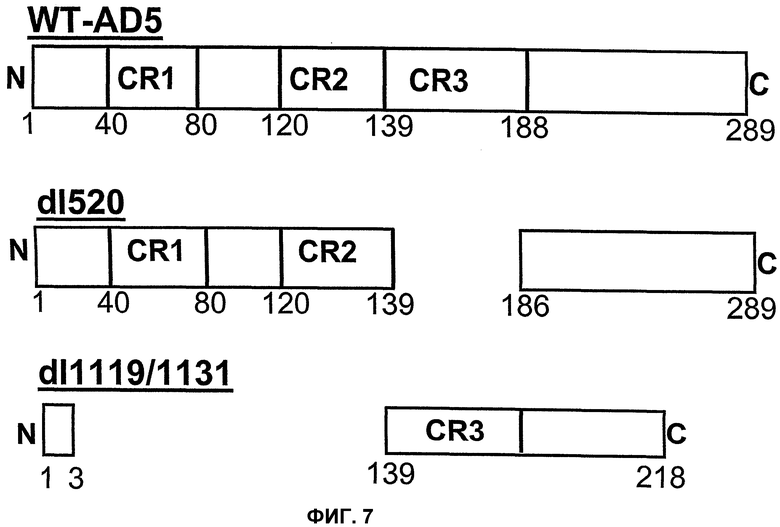
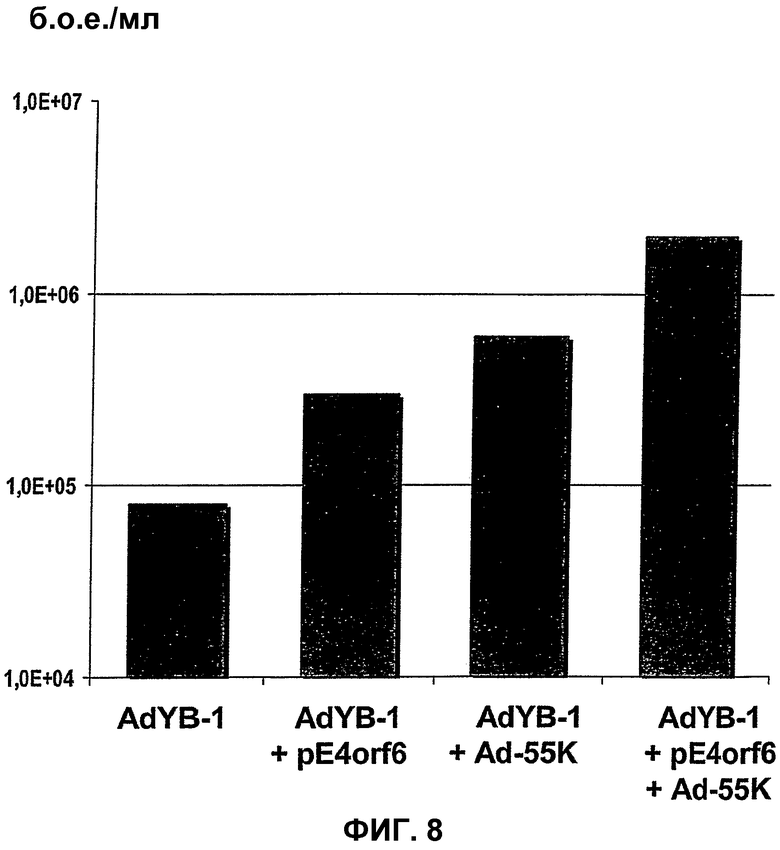
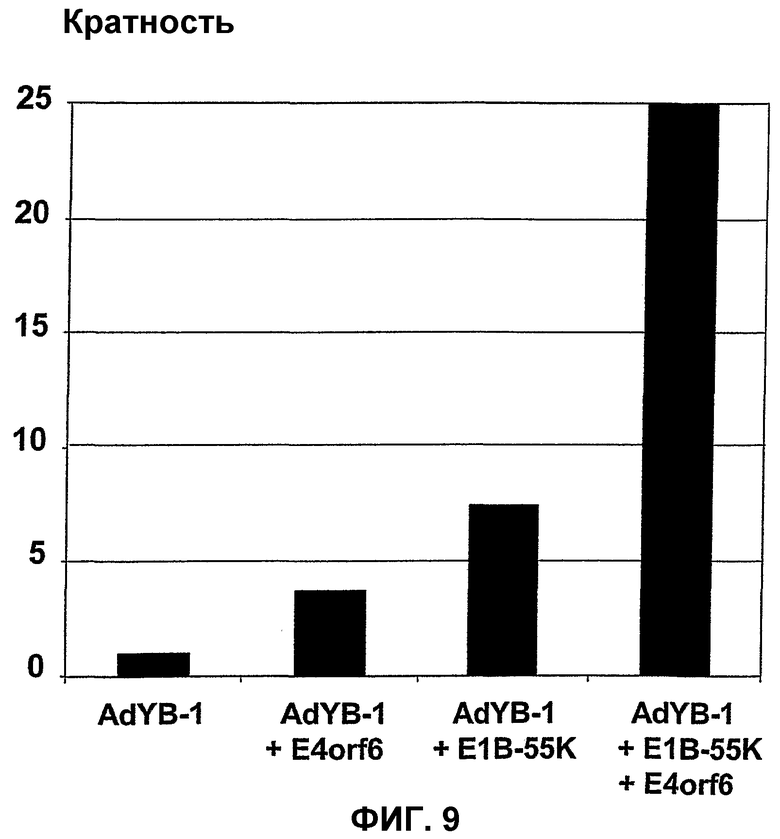
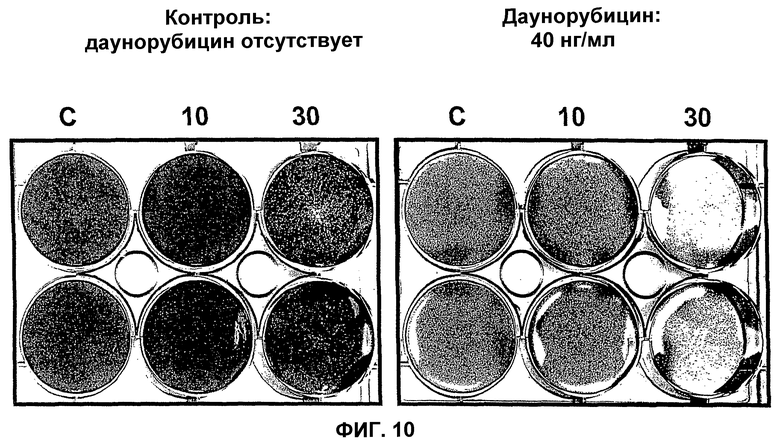
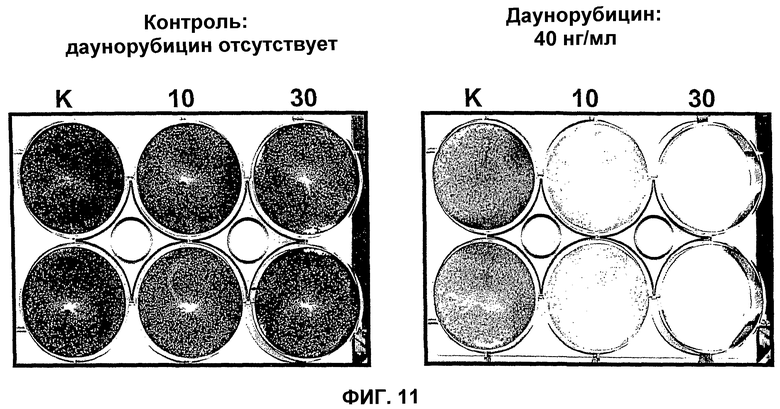
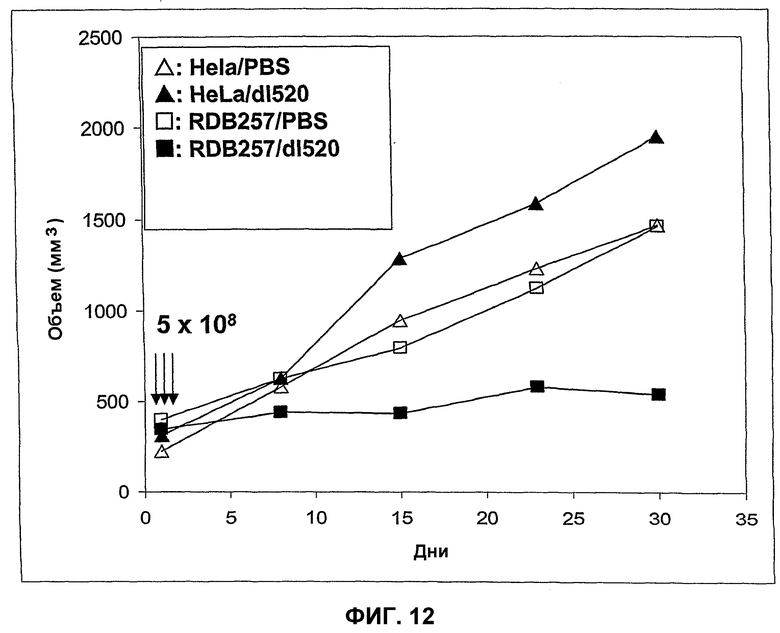
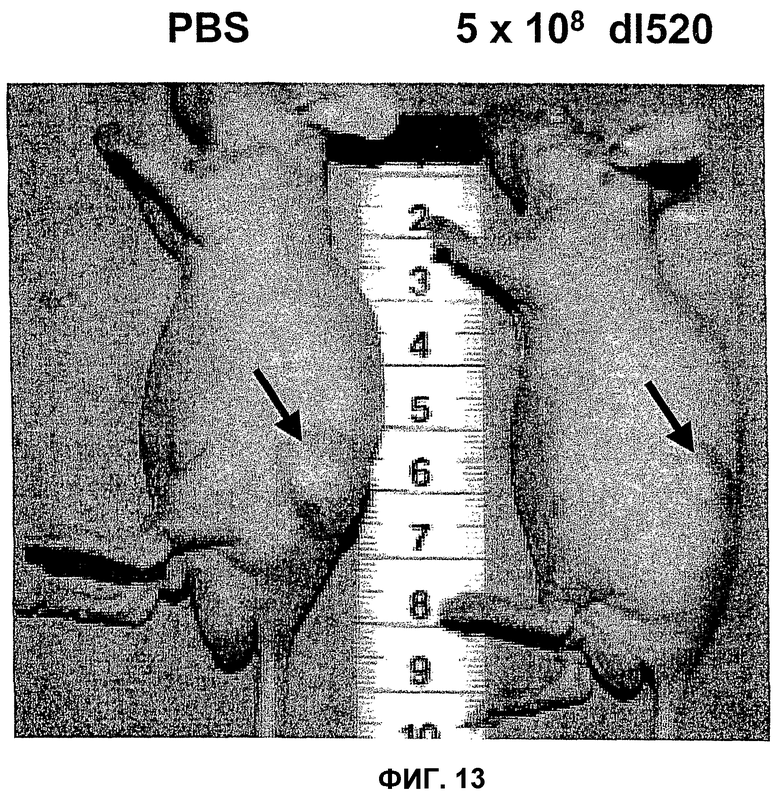
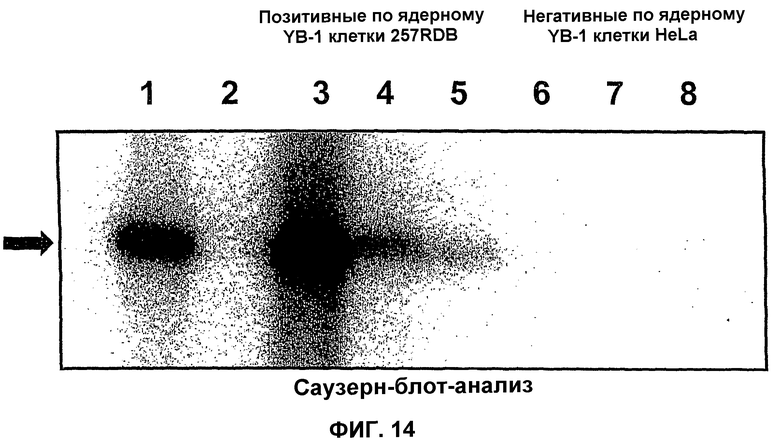
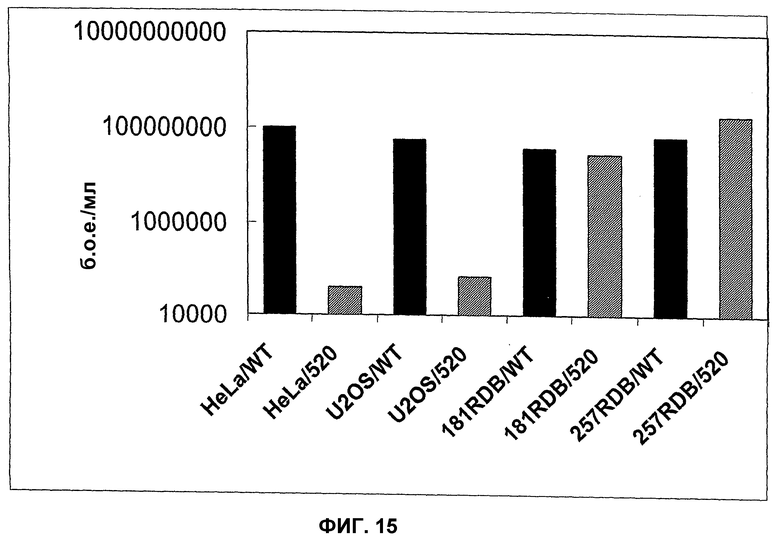
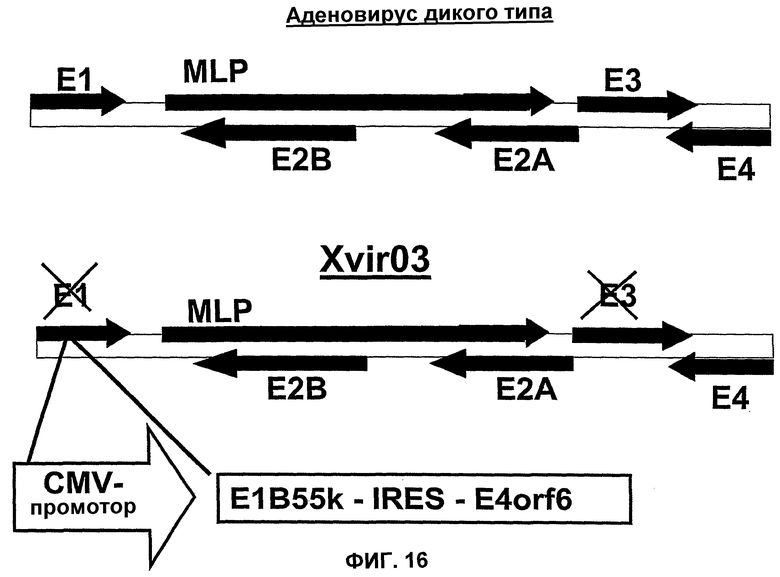
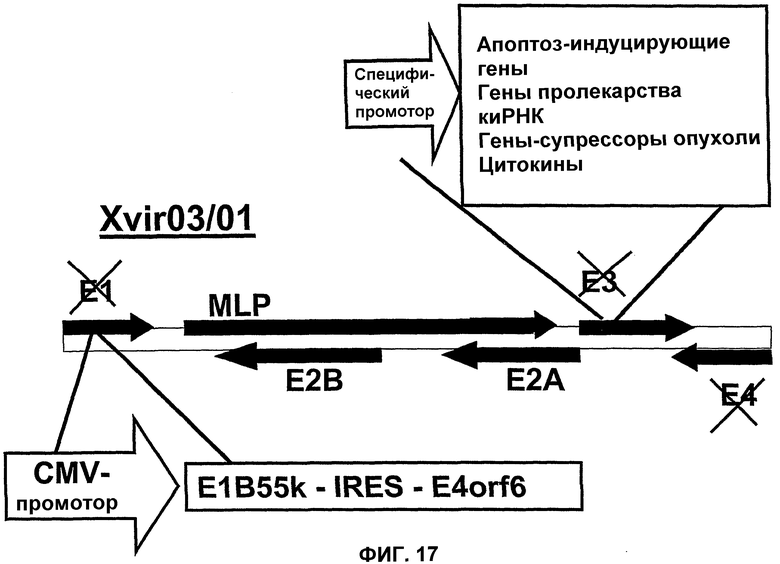
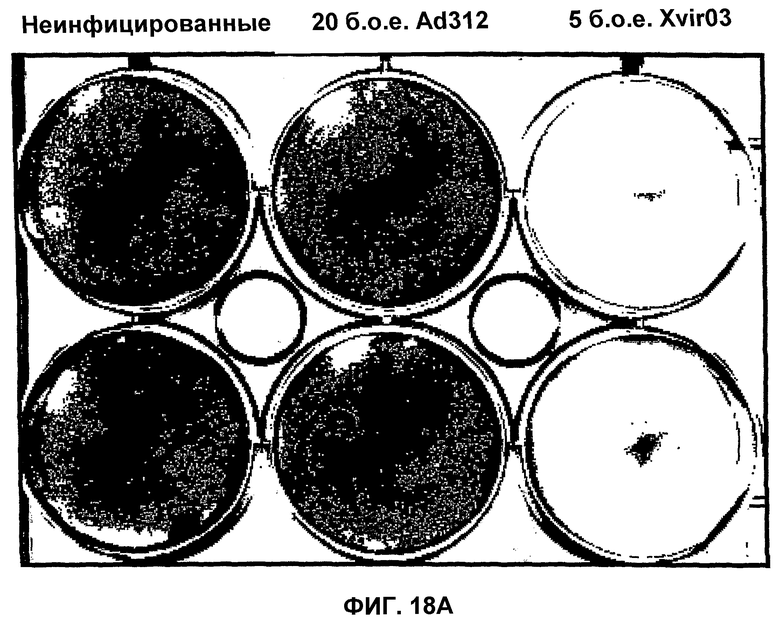
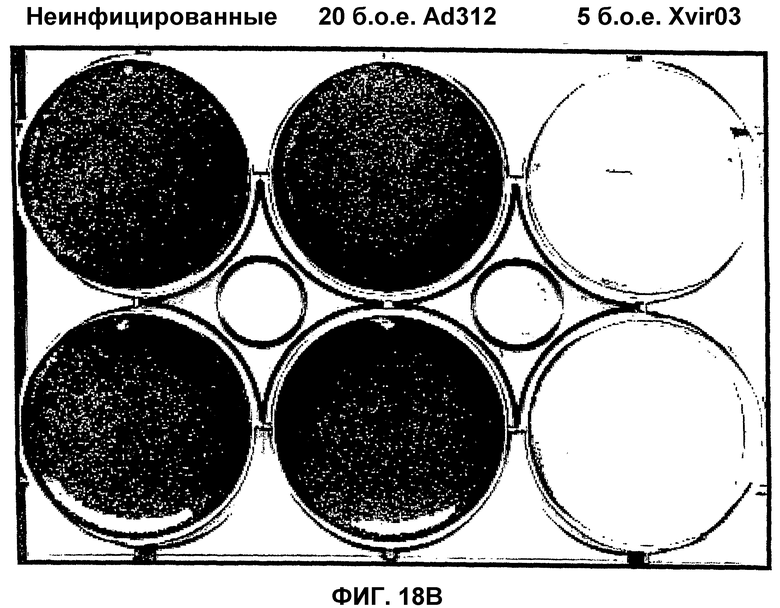
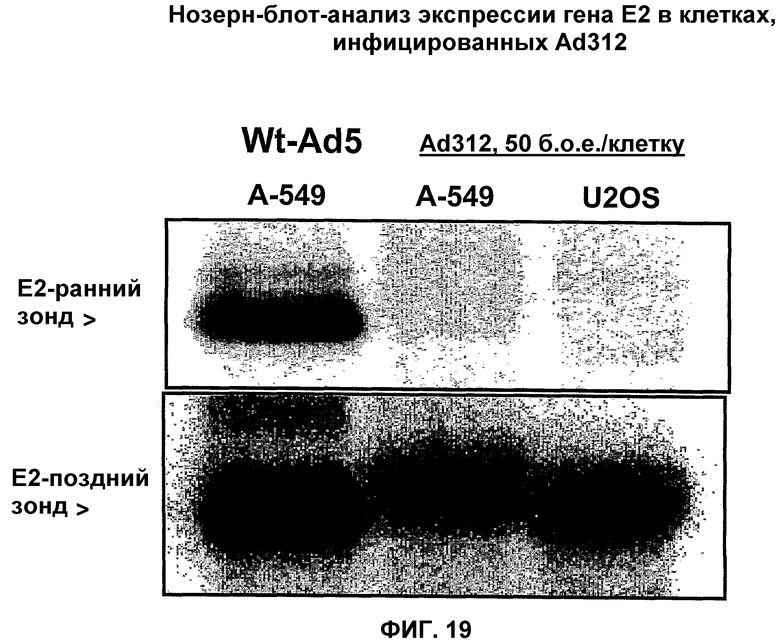
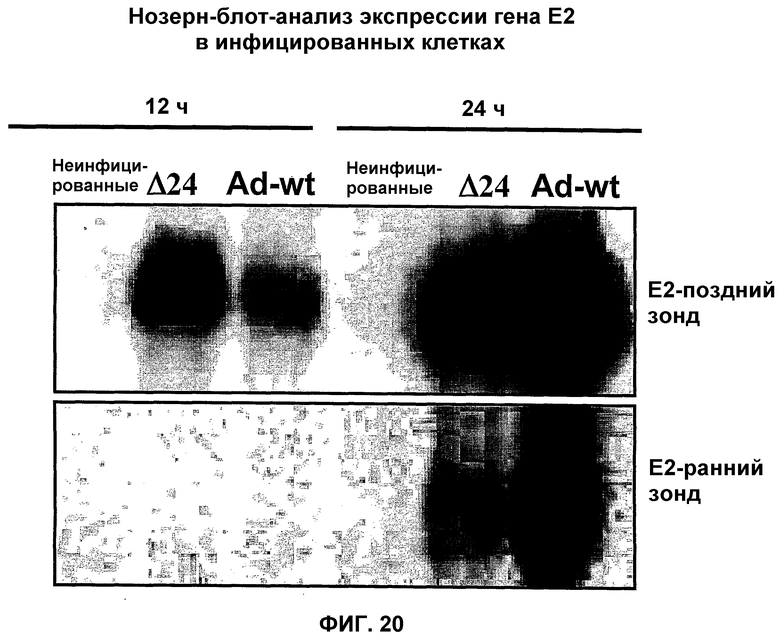
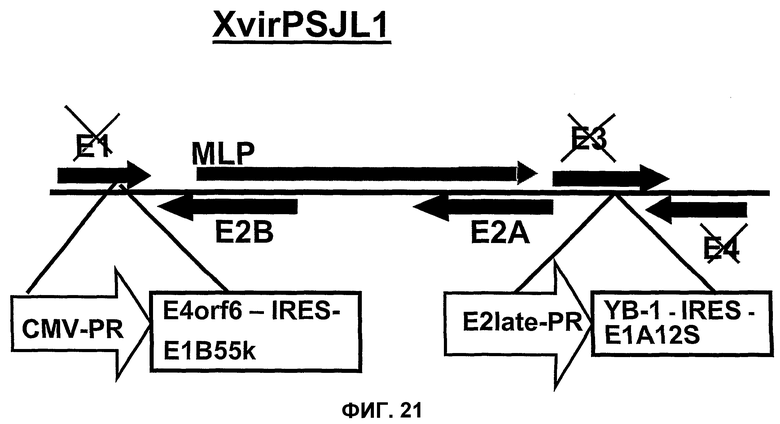
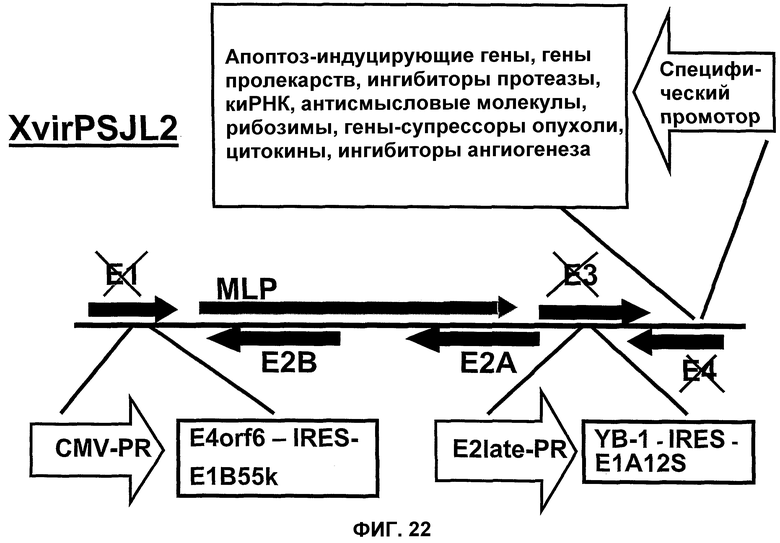
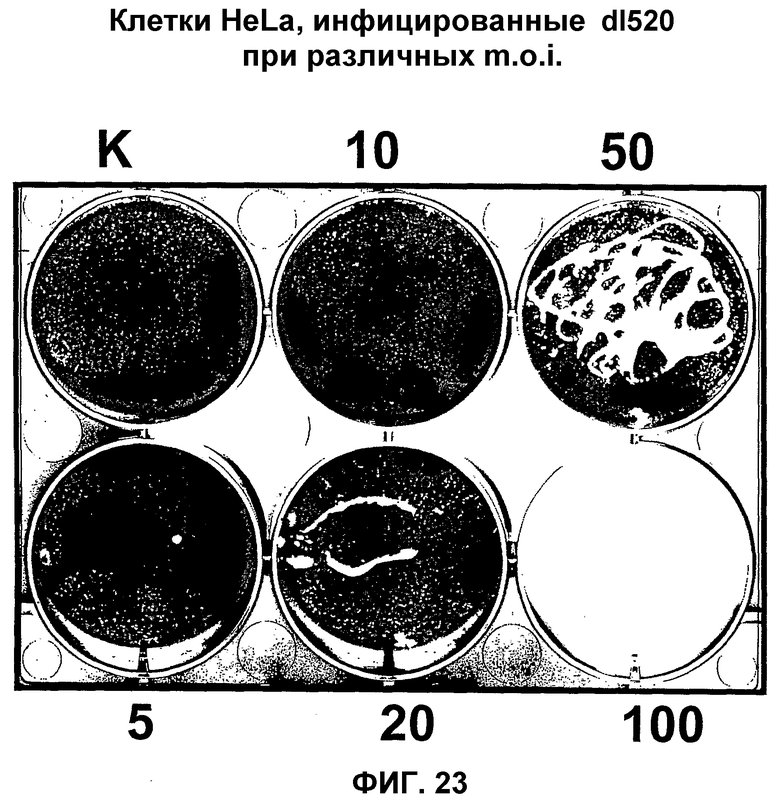
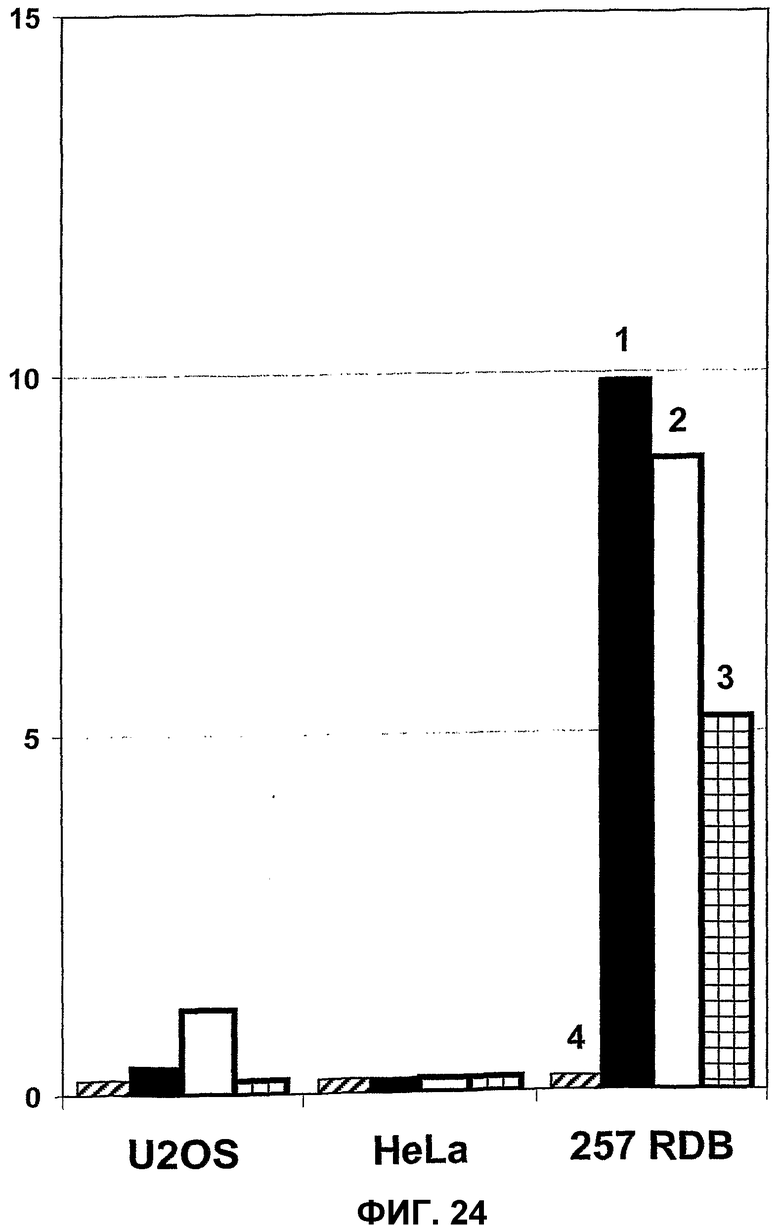

Изобретение относится к области биотехнологии. Рекомбинантный аденовирус экспрессирует белок, выбранный из группы, включающей в себя белок Е1В и Е4 и их комбинацию, а затем второй белок, представляющий собой белок Е1А. Рекомбинантный аденовирус состоит из экспрессионной кассеты, кодирующей белок Е1В и белок Е4, под контролем промотора, отличного от промотора, регулирующего экспрессию в аденовирусе дикого типа. Также, рекомбинантный аденовирус необязательно содержит нуклеиновую кислоту, кодирующую YB-1, экспрессионные кассеты, содержащие промоторы и нуклеиновые кислоты, кодирующие различные гены. Предложены также нуклеиновая кислота, кодирующая такой рекомбинантный аденовирус и их применение. Рекомбинантный аденовирус может быть использован в медицине для лечения опухолевых заболеваний. 4 н. и 26 з.п. ф-лы, 26 ил.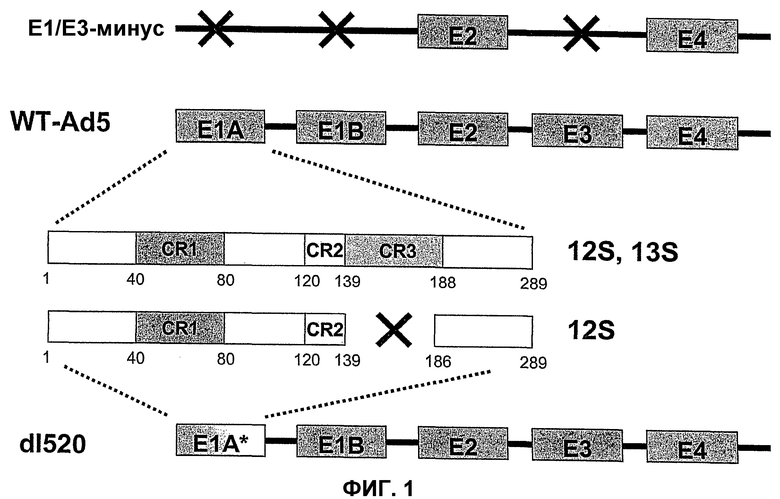
1. Рекомбинантный аденовирус, предназначенный для лечения опухолевых заболеваний, экспрессирующий первый белок, выбранный из группы, включающей в себя белок Е1В и белок Е4 и их комбинации, а затем второй белок, представляющий собой белок Е1А,
где рекомбинантный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, содержащую промотор и нуклеиновую кислоту, кодирующую белок, выбранный из группы, состоящей из указанного первого белка и указанного второго белка, где промотор отличается от промотора, регулирующего экспрессию белка в аденовирусе дикого типа, где нуклеиновая кислота аденовируса необязательно содержит, по меньшей мере, одну функционально неактивную аденовирусную область, где указанная область выбрана из группы, включающей область Е1, область Е3, область Е4 и их комбинации,
где рекомбинантный аденовирус необязательно содержит нуклеиновую кислоту, где указанная нуклеиновая кислота кодирует YB-1, где рекомбинантный аденовирус необязательно содержит экспрессионную кассету, содержащую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты выбрана из группы, включающей в себя аптамеры, рибозимы, аптазимы, антисмысловые молекулы и киPHK,
где рекомбинантный аденовирус необязательно содержит экспрессионную кассету, содержащую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты представляет собой кодирующую последовательность нуклеиновой кислоты, где указанная нуклеиновая кислота кодирует молекулу, выбранную из группы, включающей в себя пептиды, полипептиды, белки, антикалины, антитела и фрагменты антител,
где рекомбинантный аденовирус необязательно содержит экспрессионную кассету, содержащую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты выбрана из группы, включающей в себя апоптозиндуцирующие гены, гены пролекарств, гены ингибиторов протеаз, гены-супрессоры опухоли, гены цитокинов и гены ингибиторов ангиогенеза, и где дополнительно необязательно рекомбинантный аденовирус способен реплицироваться в клетках, содержащих YB-1 с нарушенной регуляцией или содержащих YB-1 в ядре.
2. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный первый белок представляет собой комбинацию белка Е1В и белка Е4, предпочтительно комбинацию белка E1B55kD и белка E4orf6.
3. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанным белком Е1А является белок E1A12S.
4. Рекомбинантный аденовирус по п.1, отличающийся тем, что экспрессия белка Е1В регулируется промотором, выбранным из группы, включающей в себя опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е1В.
5. Рекомбинантный аденовирус по п.1, отличающийся тем, что экспрессия белка Е4 регулируется промотором, выбранным из группы, включающей в себя опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е4.
6. Рекомбинантный аденовирус по п.4 или 5, где указанным аденовирусным промотором является промотор Е1А.
7. Рекомбинантный аденовирус по п.1, отличающийся тем, что экспрессия белка Е1А регулируется промотором, выбранным из группы, включающей опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный аденовирусный промотор отличается от промотора Е1А.
8. Рекомбинантный аденовирус по п.7, отличающийся тем, что указанный промотор, регулирующий экспрессию белка Е1А, представляет собой промотор, регулируемый YB-1, либо промотор, который может регулироваться YB-1.
9. Рекомбинантный аденовирус по п.8, отличающийся тем, что указанный промотор, регулирующий экспрессию белка Е1А, представляет собой аденовирусный поздний промотор Е2.
10. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус обеспечивает доставку YB-1 в ядро посредством, по меньшей мере, одного аденовирусного белка или такая доставка YB-1 в ядро опосредуется, по меньшей мере, одним аденовирусным белком, где указанный аденовирусный белок предпочтительно отличается от Е1А.
11. Рекомбинантный аденовирус по п.10, отличающийся тем, что указанным аденовирусным белком является комплекс E4orf6 и Е1В55kD.
12. Рекомбинантный аденовирус по п.1, отличающийся тем, что нуклеиновая кислота указанного аденовируса содержит, по меньшей мере, одну функционально неактивную аденовирусную область, где указанная область выбрана из группы, включающей область Е1, область Е3, область Е4 и их комбинации.
13. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, содержащую, по меньшей мере, один промотор и нуклеиновую кислоту, кодирующую аденовирусный белок, где указанным аденовирусным белком является белок Е1В, предпочтительно белок E1B55kD.
14. Рекомбинантный аденовирус по п.13, отличающийся тем, что указанный промотор выбран из группы, включающей в себя опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы, где указанный промотор отличается от промотора Е1В.
15. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, содержащую, по меньшей мере, один промотор и нуклеиновую кислоту, кодирующую аденовирусный белок, где указанным аденовирусным белком является белок Е4, предпочтительно белок E4orf6.
16. Рекомбинантный аденовирус по п.13 или 14, отличающийся тем, что указанным промотором является промотор Е1А.
17. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит, по меньшей мере, одну экспрессионную кассету, содержащую, по меньшей мере, один промотор и нуклеиновую кислоту, кодирующую аденовирусный белок, где указанным аденовирусным белком является белок Е1А, предпочтительно белок E1A12S.
18. Рекомбинантный аденовирус по п.17, отличающийся тем, что указанный промотор выбран из группы, включающей в себя опухолеспецифические промоторы, органоспецифические промоторы, тканеспецифические промоторы, гетерологичные промоторы и аденовирусные промоторы.
19. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит нуклеиновую кислоту, где указанная нуклеиновая кислота кодирует YB-1.
20. Рекомбинантный аденовирус по п.19, отличающийся тем, что указанная нуклеиновая кислота, кодирующая YB-1, находится под контролем промотора, где указанным промотором являются YB-1-зависимый и YB-1-регулируемый промоторы соответственно.
21. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит экспрессионную кассету, содержащую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты выбрана из группы, включающей в себя аптамеры, рибозимы, аптазимы, антисмысловые молекулы и киРНК.
22. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит экспрессионную кассету, содержащую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты представляет собой кодирующую нуклеиновую кислоту и где указанная нуклеиновая кислота кодирует молекулу, выбранную из группы, включающей в себя пептиды, полипептиды, белки, антикалины, антитела и фрагменты антител.
23. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус содержит экспрессионную кассету, содержащую промотор и последовательность нуклеиновой кислоты, где указанная последовательность нуклеиновой кислоты выбрана из группы, включающей в себя апоптозиндуцирующие гены, гены пролекарств, гены ингибиторов протеаз, гены-супрессоры опухоли, гены цитокинов и гены ингибиторов ангиогенеза.
24. Рекомбинантный аденовирус по п.1, отличающийся тем, что указанный аденовирус способен реплицироваться в клетках, содержащих YB-1 с нарушенной регуляцией или содержащих YB-1 в ядре.
25. Нуклеиновая кислота, кодирующая рекомбинантный аденовирус по любому из пп.1-24.
26. Применение рекомбинантного аденовируса по любому из пп.1-24 для лизиса опухолевых клеток.
27. Применение нуклеиновой кислоты по п.25 для лизиса опухолевых клеток.
28. Применение по п.26 или 27, отличающееся тем, что, по меньшей мере, часть клеток, образующих опухоль, содержит в ядре YB-1, предпочтительно YB-1 в ядре независимо от клеточного цикла.
29. Применение по п.26 или 27, отличающееся тем, что, по меньшей мере, часть клеток, образующих опухоль, обладает резистентностью, предпочтительно резистентностью к множеству фармацевтически активных агентов.
30. Применение по п.26 или 27, отличающееся тем, что опухолевые клетки подвергают лечебному мероприятию, выбранному из группы, включающей в себя облучение, введение цитотоксических средств и гипертермию.
| WO 9846779 A1, 22.10.1998 | |||
| СПОСОБ ПОЛУЧЕНИЯ со-ОКСИКИСЛОТ | 0 |
|
SU170951A1 |
| WONG H.K | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| GANLY I | |||
| et al., "Replication and cytolysis of an E1B-attenuated adenovirus in drug-resistant ovarian | |||

