Область техники, к которой относится изобретение
Изобретение относится к области лечения заболеваний, и конкретнее, к рекомбинантному аденовирусу для генной терапии или виротерапии, содержащему альбумин-связывающий участок на наружной поверхности аденовирусного гексонного белка, в частности, к онколитическому аденовирусу, содержащему альбумин-связывающий участок, и его применению для профилактики и/или лечения рака. Указанные аденовирусы «защищены» от нейтрализующих антител, находящихся в кровяном русле, и таким образом особенно подходят для системного введения.
Уровень техники
Аденовирусы широко использовались в качестве векторов для доставки генов в генной терапии, а также в качестве онколитических агентов для лечения рака. Они обладают рядом характерных свойств, которые делают их пригодными для этих применений. То есть, их структура и биология много изучались, что позволяет легко модифицировать их геном, они способны инфицировать реплицирующиеся и нереплицирующиеся клетки, и они могут быть легко получены в высоких титрах для их применения в клинике. С точки зрения безопасности, они не вызывают угрожающих жизни заболеваний у людей, и их геном не является интегративным, что предупреждает инсерционный мутагенез. Результаты клинических испытаний с аденовирусными векторами свидетельствуют о низкой токсичности и хорошем профиле безопасности, хотя эффективность все еще нуждается в улучшении, особенно, когда вирус вводится системно.
В области генной терапии системное введение, то есть, инъекции в кровоток эндовенозно или внутриартериально, могут быть необходимы для достижения многих различных органов или диссеминированных клеток. Например, в терапии рака с использованием аденовирусных векторов и онколитических аденовирусов необходимо системное введение для лечения диссеминированных опухолей, на поздней или метастатической стадии. Тем не менее, у аденовирусов имеются существенные ограничения при введении в кровоток, что ухудшает эффективность терапии. Аденовирус типа 5 (Ad5) подвергается многочисленным нейтрализующим взаимодействиям в кровотоке, что резко снижает биодоступность вируса. Депонирование в печени представляет собой серьезное препятствие для терапии, поскольку > 90% введенной дозы удерживается этим органом, главным образом, макрофагами печени, называемыми купферовскими клетками, а также эндотелиальными синусоидальными клетками (LSECs) и гепатоцитами. Прямое взаимодействие с клетками крови и белками также представляет собой важный барьер. Ad5 может непосредственно связываться с клетками крови, такими как эритроциты, посредством рецептора CAR и с тромбоцитами через интегрины. Антитела могут не только непосредственно нейтрализовать вирус, но также могут «запускать» врожденный иммунный ответ посредством активации комплемента и «стыковки» вирусных частиц с Fc-рецепторами моноцитов и нейтрофилов. Кроме того, повторное введение вектора повышает уровни нейтрализующих анти-Ad-антител (NAb) и, следовательно, усиливает нейтрализацию вируса. Опсонизация аденовируса антителами и комплементом также увеличивает клиренс купферовскими клетками. В целом, эти взаимодействия приводят к очень короткому периоду полураспада Ad в крови, составляющему примерно нескольких минут у мышей и людей.
Предпринимались интенсивные усилия для того, чтобы избежать нейтрализации антителами и иммунными клетками, когда аденовирус вводится системно.
Тестировалась химическая модификация капсида аденовируса полимерами (полиэтиленгликолем (ПЭГ) или N-(2-гидроксипропил)метакриламидом (НРМА)). Конъюгация полимера на поверхности вируса позволяет вирусу избежать нейтрализации антителами и иммунными клетками, и также отменяет связывание с CAR, интегрином и FX. Тем не менее, полимеры, конъюгированные с капсидом, не переходят вирусному потомству и увеличивают сложность крупномасштабного производства по GMP для клинического применения.
В WO 2011/129468 A9 раскрыт химерный аденовирус, способный избегать иммунного распознавания нейтрализующими антителами. Указанный аденовирус был получен посредством генетической модификации капсида аденовируса человека серотипа 5, где ген, который кодирует гексонный белок, был заменен геном гексона из обезьяньего аденовируса серотипа 19. Полученный химерный аденовирус также проявлял более высокую противоопухолевую активность по сравнению с тем же самым аденовирусом без генетической модификации.
Предпринималось несколько попыток получить аденовирус, защищенный белком альбумином (см. WO 2007/050128 А2). Однако экспериментальные данные показали, что аденовирус, имеющий капсид, модифицированный альбумин-связывающим доменом, не защищен от нейтрализующих антител (Hedley S.J. et al., 2009. The Open Gene Therapy Journal, 2:1-11).
Таким образом, по-прежнему существует необходимость в дополнительно генетически модифицированном аденовирусе, подходящем для системного введения и способного избежать действия нейтрализующих антител.
Сущность изобретения
В первом аспекте изобретение относится к аденовирусному геному, отличающемуся тем, что он содержит последовательность, кодирующую альбумин-связывающий участок, вставленную в кодирующую область гипервариабельного участка 1 (HVR1) гексонного белка, что приводит к экспрессии слитого белка, содержащего гексонный белок и альбумин-связывающий участок, и где альбумин-связывающий участок располагается на наружной поверхности гексонного белка, когда сборка гексонного белка имеет место в аденовирусном капсиде.
Во втором аспекте настоящее изобретение относится к рекомбинантному аденовирусу, имеющему аденовирусный геном по изобретению.
В третьем аспекте настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество рекомбинантного аденовируса по настоящему изобретению вместе с фармацевтически приемлемым носителем.
В четвертом аспекте настоящее изобретение относится к рекомбинантному аденовирусу или фармацевтической композиции по настоящему изобретению для применения в медицине.
В еще одном аспекте настоящее изобретение относится к рекомбинантному аденовирусу или фармацевтической композиции по настоящему изобретению для применения в профилактике и/или лечении рака у млекопитающего, где аденовирус представляет собой онколитический аденовирус или аденовирус, содержащий ген, используемый в терапии рака, вставленный в его геном.
Краткое описание чертежей
Фиг. 1. Схема инсерции альбумин-связывающего домена (ABD) в ICOVIR15-ABD. ABD 3 из стрептококкового белка G (SEQ ID NO:1) фланкирован двумя линкерами GSGS (SEQ ID NO:2) и инсерцирован в середину гипервариабельного участка 1 (HVR1) гексона онколитического вируса ICOVIR15 с получением ICOVIR15-ABD. LITR/RITR, правый и левый инвертированные концевые повторы; MLP, главный поздний промотор; E1Ap, модифицированный промотор E1A; E1A-Δ24, мутантный вариант белка E1A, где аминокислоты 121-129 полипептидной цепи делецированы; L1-L5, поздние гены; волокно RGD, модифицированное RGD волокно инсерцией пептида RGD в H1-петлю волокна.
Фиг. 2. Изображение аденовируса, содержащего альбумин-связывающий домен (ABD), вставленный в гексон. По сравнению с немодифицированным аденовирусом ICOVIR15 (слева), ABD-модифицированный вирус ICOVIR-15-ABD (справа) покрыт альбумином, присутствующим в крови, защищая тем самым вирус от нейтрализующих антител.
Фиг. 3. Кинетика продукции вирусов ICOVIR15-ABD и ICOVIR15. Конфлюентные клетки А549 заражали 800 вирусными частицами (vp) на клетку. Через четыре часа (ч) после заражения вирус извлекали, клетки трижды промывали PBS и инкубировали с несодержащей вирус средой. Клеточные экстракты собирали через 4, 24, 48 и 72 ч после заражения и титровали методом окрашивания с использованием антитела к гексону. Образцы оценивали в трех повторностях. Среднее значение±стандартное отклонение приведено в виде столбцов (хотя их трудно различить, с учетом низких значений). TU/мл, трансдуцирующие единицы на мл. *Статистическая значимость по сравнению с группой ICOVIR15 (p≤0,05).
Фиг. 4. Сравнительная цитотоксичность in vitro ICOVIR15 и ICOVIR15-ABD в присутствии или отсутствии человеческого сывороточного альбумина (HSA). Клетки A549, Sk-mel28, HEK293 и MCF-7 инфицировали указанными вирусами из расчета от 10000 до 0,0001 вирусных частиц (vp) на клетку. Показаны значения IC50 (количество vp на клетку, необходимое для того, чтобы вызвать снижение жизнеспособности клеточной культуры на 50%) на 7 сутки после заражения. Три различных повторности количественно определяли для каждой клеточной линии. Среднее значение±стандартное отклонение приведено в виде столбцов. MOI, множественность инфекции.
Фиг. 5. ICOVIR15-ABD связывает человеческий и мышиный альбумин по данным ELISA. А) Лунки покрывали человеческим или бычьим сывороточным альбумином (HSA или BSA, который связывается или не связывается с ABD, соответственно). Определение связывания проводили при трех различных количествах вирусного белка (0,25, 2,5 и 25 нг). Было детектировано связывание аденовирусов ICOVIR15 и ICOVIR15-ABD с покрытыми альбумином лунками после инкубации с антителом к гексону и меченным пероксидазой вторичным антителом с использованием колориметрического анализа. Образцы оценивали в трех повторностях. Среднее значение±стандартное отклонение приведено в виде столбцов. OD, оптическая плотность. *Статистическая значимость по сравнению с другими группами (p≤0,05). B) Лунки покрывали бычьим, человеческим или мышиным сывороточным альбумином (BSA, HSA или MSA). Количество тестированного вирусного белка составляло 25 нг. Было детектировано связывание аденовируса с покрытыми альбумином лунками после инкубации с антителом к гексону и меченным пероксидазой вторичным антителом с использованием колориметрического анализа. В опыт включали контрольную группу без аденовируса. Образцы оценивали в трех повторностях. Среднее значение±стандартное отклонение приведено в виде столбцов. OD, оптическая плотность. *Статистическая значимость по сравнению с другими группами (p≤0,05).
Фиг. 6. Связывание с альбумином защищает аденовирус от нейтрализующих антител in vitro. Аденовирусы AdGL и AdGL-ABD с покрытием человеческим сывороточным альбумином (HSA) или без, инкубировали с серийными разведениями нейтрализующего антитела Ab6982 в течение 1 ч при температуре 37°С. Затем добавляли клетки НЕК293 для получения множественности инфекции, равной 0,5 трансдуцирующих единиц (TU) на клетку. Через 24 ч после заражения трансдукцию клеток анализировали по экспрессии люциферазы. В опыт включали контроль без антител (Ab) (контроль «без Ab»), чтобы получить значение инфекции, равное 100%. Образцы оценивали в трех повторностях. Среднее значение±стандартное отклонение приведено в виде столбцов е.
Фиг. 7. ICOVIR15-ABD показывает повышенную in vitro цитотоксичность в присутствии нейтрализующих антител при защите сывороточным альбумином человека (HSA). ICOVIR15 и ICOVIR15-ABD инкубировали с серийными разведениями нейтрализующего антитела Ab6982 (Nab, промышленно доступные поликлональные анти-HAd5-антитела) в течение 1 ч в присутствии или отсутствии человеческого сывороточного альбумина (HSA). Клетки А549 добавляли для получения множественности инфекции, равной 600 вирусных частиц (vp) на клетку. Процент выживших клеток (содержание белка в лунках) измеряли на 4 сутки после заражения. Образцы оценивали в трех повторностях. Среднее значение±стандартное отклонение приведено в виде столбцов.
Фиг. 8. Инсерция ABD увеличивает период полураспада аденовируса в плазме крови. Мышам nude вводили смесь ICOVIR15 и ICOVIR15-ABD в соотношении 1:1 в суммарной дозе 5×1010 вирусных частиц (vp) на мышь (n=5). Образцы крови собирали через 5, 15 мин, 1, 4 и 24 ч после введения и центрифугировали для получения сыворотки. Проводили ПЦР-амплификацию гипервариабельного участка 1 (HVR1) гексона аденовируса и образцы анализировали с помощью электрофореза. Инсерция ABD увеличивает размер HVR1 с 299 до 361 п.н. Показаны дорожки в геле со стандартом с несколькими соотношениями геномов ICOVIR15-ABD:ICOVIR15 (0,2, 1, 5, 10 и 50), контролем до введения (t0), водой, используемой в качестве отрицательного контроля для ПЦР (H2O) и ПЦР проб сыворотки (от #1 до #5).
Фиг. 9. Противоопухолевая активность ICOVIR15-ABD после системного введения in vivo. Мышам nude, несущим подкожные ксенотрансплантаты меланомы (Sk-mel28), вводили однократно внутривенно дозу забуференного фосфатом физиологического раствора (PBS), ICOVIR15 или ICOVIR15-ABD (5×1010 вирусных частиц (vp) на мышь). Приведены графики объем опухолей±SEM (n=10-12). *Статистическая значимость по сравнению с группой PBS (p≤0,05).
Фиг. 10. Трансдукция печени и опухоли in vivo аденовирусным вектором, модифицированным альбумин-связывающим доменом в HVR1 гексона, сохраняется у предварительно иммунизированных аденовирусом мышей. Мышей C57BL/6, несущих подкожные ксенотрансплантаты меланомы (B16-CAR), иммунизировали внутрибрюшинной инъекцией hAd5wt (2×1010 вирусных частиц (vp) на мышь) или растворителем, и через 7 суток внутривенно вводили AdGL (вектор GFP-люцифераза) или AdGL-ABD (3×1010 вирусных частиц на мышь). Через 3 суток анализировали активность люциферазы в печени и опухоли биолюминесцентной томографией (IVIS). Среднее значение±стандартное отклонение приведено в виде столбцов (печень n=4-6, опухоли n=8-12). с: секунды; sr: стерадиан.
Фиг. 11. Инсерция ABD в гипервариабельный участок 5 не влияет на жизнеспособность вируса. Клетки НЕК293 трансфектировали плазмидой pAdZGL-Н5-ABD для получения вируса AdGL-Н5-ABD. Через одну неделю собирали клетки и супернатант и подвергали лизису с помощью трех циклов замораживания-оттаивания. Клеточный экстракт, содержащий вирус, титровали на клетках НЕК293 с помощью метода бляшек. Показаны лунки, соответствующие разведениям 1E6, 1E7 и 1E8, где видны бляшки, свидетельствующие о размножении вируса.
Фиг. 12. Альбумин-связывающий домен, инсерцированный в HVR5, не защищает аденовирус от нейтрализующих антител, в отличие от того же домена, инсерцированного в HVR1. Опыт с нейтрализацией in vitro проводили на клетках НЕК293 и Sk-mel28, сравнивая AdGL, AdGL-H1-ABD и AdGL-H5-ABD. Аденовирусы инкубировали в течение 1 ч с серийными разведениями нейтрализующего антитела Ab6982 в присутствии или отсутствии человеческого сывороточного альбумина (HSA). Клетки последовательно добавляли для получения множественности инфекции, равной 10 вирусных частиц (vp) на клетку (НЕК293) и 40 vp на клетку (Sk-mel28). Через двадцать четыре часа после заражения анализировали трансдукцию клеток по экспрессии люциферазы. В опыт включали контроль без антител (Ab) (контроль «без Ab»), чтобы получить значение инфекции, равное 100%. Образцы оценивали в трех повторностях. Среднее значение±стандартное отклонение приведено в виде столбцов.
Подробное описание изобретения
Авторы настоящего изобретения установили, что аденовирус, генетически модифицированный альбумин-связывающим участком, на наружной поверхности капсида, в частности, на наружной поверхности аденовирусного гексонного белка, способен приобрести «альбуминовый щит», позволяющий вирусу избежать действия нейтрализующих антител и повышающий его персистентность в крови после системного введения. Этот результат является неожиданным, поскольку предшествующие попытки модифицировать аденовирус альбумин-связывающим доменом закончились неудачей в отношении усиления защиты аденовируса от нейтрализующих антител (Hedley S.J. et al., 2009. The Open Gene Therapy Journal, 2:1-11).
Кроме того, когда рекомбинантный аденовирус является онколитическим аденовирусом, то инсерция альбумин-связывающего участка повышает его противоопухолевую активность. В этом смысле, указанные генетически модифицированные аденовирусы имеют потенциальную ценность для преодоления ограничений системного введения, в частности, для лечения рака.
Результаты, приведенные в примерах настоящего изобретения, ясно показывают, что селективный по репликации онколитический аденовирус (ICOVIR15-ABD), содержащий последовательность, кодирующую альбумин-связывающий домен (ABD) из стрептококкового белка G, инсерцированный в гипервариабельный участок 1 (HVR1) кодирующей последовательности гексонного белка, экспонирует данный домен на его капсиде, способствуя связыванию альбумина, защищая аденовирус от нейтрализующих антител, повышая его период полураспада в плазме крови и усиливая его противоопухолевую эффективность. Экспериментальные примеры, приведенные в настоящем изобретении, также показывают, что инсерция ABD из стрептококкового белка G в HVR1 гексонного белка дефицитного по репликации аденовируса (AdGL-ABD) защищает аденовирус от нейтрализующих антител (фиг. 6). Таким образом, эти результаты показывают, что альбуминовое покрытие аденовируса функционирует в качестве щита, чтобы «спрятать» вирусные белки и избежать множественных нежелательных взаимодействий в крови (нейтрализующие антитела, поглощение клетками крови и его захват печенью), улучшая его фармакокинетику. Это особенно важно, когда аденовирусный вектор для генной терапии, вакцины или онколитический аденовирус вводится повторно. Таким образом, генетически модифицированные аденовирусы по изобретению пригодны для системного введения.
Аденовирусы по изобретению
Результаты, полученные в настоящем изобретении, показывают, что аденовирус, содержащий альбумин-связывающий участок, на наружной поверхности аденовирусного гексонного белка, может быть покрыт альбумином, тем самым защищая его от нейтрализующих антител, присутствующих в кровотоке. Данный защитный эффект наблюдается для репликативных (ICOVIR15-ABD) и нерепликативных (AdGL-ABD) аденовирусов.
В одном аспекте настоящее изобретение относится к рекомбинантному аденовирусу, имеющему аденовирусный геном, отличающийся тем, что он содержит последовательность, кодирующую альбумин-связывающий участок, вставленный в кодирующую область гипервариабельного участка 1 (HVR1) гексонного белка, что приводит к экспрессии слитого белка, содержащего гексонный белок и альбумин-связывающий участок, и где альбумин-связывающий участок располагается на наружной поверхности гексонного белка, когда сборка гексонного белка имеет место в аденовирусном капсиде.
В рамках изобретения, термин «аденовирус» относится к любому вирусу, который может быть квалифицирован как аденовирус, т.е. любой вирус, относящийся к семейству Adenoviridae, характеризующийся тем, что он является вирусом без оболочки с икосаэдрическим нуклеокапсидом, содержащим двухцепочечную ДНК генома. Этот термин включает любой аденовирус, способный инфицировать человека или животного, включая все группы, подгруппы и серотипы, которые используют CAR в качестве рецептора для инфицирования клеток-мишеней. Аденовирусы по настоящему изобретению включают, не ограничиваясь этим, аденовирусы птицы, собаки, лошади, коровы, овцы, свиньи, человека или лягушки. В предпочтительном варианте осуществления аденовирус по изобретению представляет собой аденовирус человека, т.е. аденовирус, способный инфицировать людей. Согласно изобретению «серотип» представляет собой любой из иммунологически различных типов аденовирусов. Существует по меньшей мере 57 серотипов аденовируса человека, которые подразделяются на несколько подгрупп (от А до G). Изобретение предполагает применение любого аденовирусного серотипа, известного в данной области, включая, не ограничиваясь этим, любой из серотипов, указанных в таблице 1.
Таблица 1. Несколько примеров аденовирусных подгрупп и серотипов, подходящих для применения в настоящем изобретении
В предпочтительном варианте осуществления изобретения аденовирус человека выбран из группы, состоящей из аденовируса человека серотипов 1 до 57.
В другом предпочтительном варианте осуществления изобретения аденовирус относится к подгруппе С, более предпочтительно представляет серотип 5.
Человеческий аденовирус серотипа 5 (Ad5) ассоциирован со слабыми респираторными инфекциями. Последовательность гена человеческого аденовируса серотипа 5 можно найти в GenBank: AY339865.1 (версия от 13 августа 2007 г.).
Аденовирус по изобретению представляет рекомбинантный аденовирус. В рамках изобретения, термин «рекомбинантный» относится к аденовирусу, который отсутствует в природе. Этот рекомбинантный аденовирус содержит одну или более модификаций по сравнению с диким типом. Такие модификации включают, не ограничиваясь этим, модификации в геноме аденовируса, который упакован в частицы, для того, чтобы получить инфекционный вирус. Другие модификации позволяют получить дефицитный по репликации вирус (т.е. вирус, который не может репродуцироваться) удалением гена из генома вируса, который имеет критическое значение для репликации. Примеры модификаций включают делеции, известные в данной области, такие как делеции в одной или более кодирующих областей Е1a, E1b, E2a, E2b, Е3 или Е4. Другие возможные модификации включают делеции всех кодирующих областей генома аденовируса. Такие аденовирусы известны как «лишенные существенного внутреннего содержимого» - «gutless» аденовирусы. Химерные аденовирусы, образованные комбинацией элементов различных серотипов, также включаются.
Термин «рекомбинантный» также включает условные по репликации аденовирусы, представляющие собой вирусы, которые предпочтительно реплицируются в определенных типах клеток или тканей, но в меньшей степени или вообще не реплицируются в других типах. Например, среди аденовирусов по изобретению имеются аденовирусы, которые реплицируются в аномально пролиферирующей ткани, такой как солидные опухоли и другие новообразования. Они включают вирусы, раскрытые в патенте США № 5998205 и в патенте США № 5801029. Такие вирусы иногда называют «цитолитическими» или «цитопатическими» вирусами (или векторами), и, если они имеют такой эффект на опухолевые клетки, то называются «онколитическими» вирусами (или векторами).
В одном варианте осуществления аденовирус является репликативным аденовирусом, в частности, онколитическим аденовирусом.
В еще одном варианте осуществления изобретения аденовирус является нерепликативным аденовирусом или дефицитным по репликации аденовирусом. Дефицитный по репликации аденовирус или нерепликативный аденовирус представляет собой аденовирус, неспособный реплицироваться в клетке-мишени, он используется в генной терапии в качестве носителя генов в клетки-мишени, поскольку цель состоит в том, чтобы экспрессировать терапевтический ген внутри клетки, но не лизировать клетку.
Рекомбинантный аденовирус по настоящему изобретению модифицирован посредством инсерции гетерологичной последовательности на наружной поверхности аденовирусного гексонного белка. В частности, гетерологичная последовательность кодирует альбумин-связывающий участок.
Частица аденовируса состоит из капсида, который включает вирусную ДНК. В рамках изобретения, термин «капсид» относится к белковой оболочке вируса, образованной субъединицами, называемыми капсомерами, которые могут быть пятиугольными или шестиугольными. Аденовирусный капсид имеет форму икосаэдра, который имеет 20 равносторонних треугольных граней. Большая часть капсида образована гексонным белком, и каждая вершина имеет комплекс, образованный пентоновым основанием и белком волокна.
В рамках изобретения, термин «аденовирусный гексонный белок» или «гексонный белок» (ранее известный как «белок II») относится к основному структурному белку капсида, найденного в аденовирусах, который самоассоциируется с образованием тримеров, каждый в форме шестиугольника. 240 гексонных тримеров собираются с обеспечением аденовирусного капсида. Гексонный белок имеет важное значение для сборки капсида вируса, определения икосаэдрической симметрии капсида и целостности капсида. Основные структурные свойства гексонного белка одинаковы у аденовирусов различных серотипов, но гексонный белок отличается по размеру и иммунологическим свойствам между серотипами. В настоящем изобретении термин «гексонный белок» охватывает гексонный белок любого аденовируса, включая, не ограничиваясь этим, белок, определяемый последовательностью из базы данных UniProt с инвентарным номером P04133 от 19 февраля 2014 года, который соответствует гексонному белку человека аденовируса C серотипа 5; белок, определяемый последовательностью из базы данных UniProt с инвентарным номером P03277 от 19 февраля 2014 года, который соответствует гексонному белку аденовируса человека C серотипа 2; белок, определяемый последовательностью из базы данных UniProt с инвентарным номером P42671 от 19 февраля 2014 года, который соответствует гексонному белку аденовируса птиц gal1 (штамм Phelps); и белок, определяемый последовательностью из базы данных UniProt с инвентарным номером P11819 от 19 февраля 2014 года, который соответствует гексонному белку аденовируса человека F серотипа 40. Выражение включает все природные варианты гексонного белка, которые естественным путем возникают в других подгруппах или серотипах.
В настоящем изобретении выражение «наружная поверхность гексонного белка» относится к областям гексонного белка, которые экспонированы на поверхности капсида. Для того, чтобы определить: вставлен альбумин-связывающий участок по настоящему изобретению во внутреннюю часть или в наружную поверхность аденовирусного гексонного белка, можно провести анализ детектирования связывания с сывороточным альбумином человека, как описано в экспериментальном разделе данной заявки (например, анализ ELISA) или анализ нейтрализации in vitro. Если человеческий сывороточный альбумин может связаться с аденовирусом, то тогда альбумин-связывающий участок, был введен в наружную поверхность аденовирусного гексонного белка.
Сообщалось, что петля 1 (L1) и петля 2 (L2) гексонного белка экспонированы снаружи структуры вирусного капсомера. L1 содержит шесть гипервариабельных участков (HVRs), т.е. от HVR1 до HVR6, и L2 содержит седьмой гипервариабельный участок (HVR7).
В рамках изобретения, термин «гипервариабельный участок или «HVR» относится к области, варьирующей по длине и последовательности между аденовирусными серотипами, образующей часть поверхностных экспонированных петлей. Существует семь гипервариабельных участков аденовирусного гексона для каждой субъединицы тримера (Biere B. and Schweiger B., J. Clin. Virol., 2010; 47 (4):366-371). В контексте настоящего изобретения номенклатура, используемая для HVRs, представляет номенклатуру, описанную Crawford-Miksza и Schnurr (Crawford-Miksza and Schnurr, 1996, Virology, 224 (2):357-367). В предпочтительном варианте осуществления настоящего изобретения HVR является HVR1. Инсерция определенного остатка в область HVR приводит к 240 раз × 3 или 720 вставкам в целом в аденовирусный вектор. В предпочтительном варианте осуществления последовательность, кодирующую альбумин-связывающий участок, вставляют таким образом, что полученный слитый белок содержит альбумин-связывающий участок после аминокислоты D150 гексонного белка согласно нумерации для гексонного белка, имеющего инвентарный номер в GenBank BAG48782.1 от 14 июня 2008, соответствующий гексонному белку человеческого аденовирусного серотипа 5.
В более предпочтительном варианте осуществления изобретения нуклеотидная последовательность полного модифицированного аденовирусного гексона, имеющего ABD, инсерцированный в HVR1 (ABD-HVR1), представляет SEQ ID NO:3. Заявители настоящего изобретения показали, что инсерция альбумин-связывающего домена в другой HVR (в частности, HVR5) дает жизнеспособный вирус, но не защищает аденовирус от нейтрализующих антител. Примеры, представленные в настоящей заявке, показывают, что последовательность, кодирующая альбумин-связывающий участок, инсерцирована в HVR5 таким образом, что полученный в результате слитый белок содержит альбумин-связывающий участок после аминокислоты A274 гексонного белка согласно нумерации гексонного белка, имеющего инвентарный номер BAG48782.1 в GenBank, от 14 июня 2008, соответствующий гексонному белку человеческого аденовируса серотипа 5. Нуклеотидная последовательность полного модифицированного аденовирусного гексона, содержащего ABD, инсерцированный в HVR5 (ABD-HVR5), представляет SEQ ID NO:4. На фиг. 12 показано, что альбумин-связывающий домен является функциональным при вставке в HVR1, но не в HVR5.
Альбумин-связывающий участок может быть непосредственно присоединен к гексонному белку, т.е. N- и С-конец альбумин-связывающего участка непосредственно связан с гексонным белком. Однако также возможно, чтобы альбумин-связывающий участок был связан с гексонным белком посредством линкерной последовательности. Таким образом, в еще одном варианте осуществления N- и/или С-конец альбумин-связывающего участка связан с гексонным белком посредством линкерной последовательности.
В рамках изобретения, термин «линкерная последовательность» относится к аминокислотной последовательности, которая функционирует в качестве шарнирной области между гексонным белком и альбумин-связывающим участком, обеспечивая пространство между обоими элементами и гарантируя, что вторичная структура гексонного белка не нарушается за счет присутствия участка ABD, и наоборот. Линкерная последовательность может быть любой длины, которая позволяет обоим элементам перемещаться независимо друг от друга, сохраняя при этом трехмерную форму отдельных элементов. В предпочтительном варианте осуществления линкерная последовательность представляет собой гибкий пептидный линкер длиной из 31 аминокислоты или менее. Более предпочтительно линкерная последовательность содержит менее 10 аминокислот, менее 5 аминокислот, менее 4 аминокислот или 2 аминокислот. В одном варианте осуществления линкерная последовательность содержит 2 или более аминокислот, выбранных из группы, состоящей из глицина, серина, аланина и треонина. В еще одном варианте осуществления указанный линкер представляет собой полиглициновый линкер. Показательные, неограничивающие примеры линкерных последовательностей включают SGGTSGSTSGTGST (SEQ ID NO: 5), AGSSTGSSTGPGSTT (SEQ ID NO: 6), GGSGGAP (SEQ ID NO: 7) и GGGVEGGG (SEQ ID NO: 8). Данные последовательности были использованы для связывания сконструированных биспиралей с другими белковыми доменами (Muller K.M. et al., Meth. Enzymology, 2000, 328:261-281). Предпочтительно, чтобы линкерная последовательность включала последовательность GSGS (SEQ ID NO: 2). Другие линкеры, известные в данной области, могут быть использованы в качестве альтернативы (Reddy Chichili V. P., Kumar V. and Sivaraman J. (2013). Linkers in the structural biology of protein-protein interactions. Protein Science, 22 (2):153 -67).
Следовательно, аденовирус по настоящему изобретению имеет альбумин-связывающий участок, на наружной поверхности гексонного белка, таким образом, способствуя покрытию аденовирусного капсида альбумином.
В рамках изобретения, термин «альбумин» относится к члену семейства белков альбуминов, которые представляют собой водорастворимые глобулярные белки, умеренно растворимые в концентрированных растворах солей и подвергающиеся тепловой денатурации. Альбумины обычно обнаруживаются в плазме крови. Сывороточный альбумин, продуцируемый печенью, растворяется в плазме крови и является наиболее распространенным белком в крови у млекопитающих. В частности, термин «сывороточный альбумин» относится к глобулярному белку, который в организме человека кодируется геном ALB (UniGene Hs. 418167). Человеческий сывороточный белок альбумин представляет белок, определяемый последовательностью из базы данных Uniprot с инвентарным номером P02768 от 19 марта 2014 года.
В рамках изобретения, термин «альбумин-связывающий участок» относится к любой аминокислотной последовательности, способной связываться с альбумином, то есть обладающей аффинностью связывания с альбумином. Предпочтительно, он способен связываться с сывороточным альбумином, более предпочтительно, с человеческим сывороточным альбумином. Термин «альбумин-связывающий участок» включает, не ограничиваясь этим, встречающиеся в природе альбумин-связывающие домены (ABD) (например, ABD, присутствующие в бактериальных белках), и альбумин-связывающие последовательности из синтетических пептидов. В предпочтительном варианте осуществления альбумин-связывающий участок выбран из альбумин-связывающего домена из стрептококкового белка G, альбумин-связывающего домена из белка PAB Peptostreptococcus magnus, альбумин-связывающего пептида, имеющего коровую последовательность DICLPRWGCLW (SEQ ID NO:9) и их функционально эквивалентных вариантов. В более предпочтительном варианте осуществления изобретения альбумин-связывающий домен происходит из стрептококкового белка G.
Термин «альбумин-связывающий домен» относится к любой области из встречающегося в природе белка, который способен связываться с альбумином с достаточной специфичностью, с тем, чтобы обеспечить защиту от нейтрализующих антител.
В рамках изобретения, термин «альбумин-связывающий домен из стрептококкового белка G» или «ABD из стрептококкового белка G» относится к домену, который состоит из 46 аминокислотных остатков, образующих трехспиральный пучок (Kraulis P.J. et al., FEBS Lett, 1996; 378:190-4), и связывается с высокой аффинностью с человеческим и мышиным альбумином, но не с бычьим альбумином (Konig Т. and Skerra A., J. Immunol. Methods, 1998; 218: 73-83). Существует несколько альбумин-связывающих доменов в стрептококковом белке G. В предпочтительном домене альбумин-связывающий участок представляет альбумин-связывающий домен 3 из стрептококкового белка G. Предпочтительно, чтобы последовательность альбумин-связывающего домена 3 из стрептококкового белка G представляла последовательность SEQ ID NO: 1.
В рамках изобретения, термин «альбумин-связывающий домен из белка Peptostreptococcus magnus PAB» относится к альбумин-связывающему домену из белка PAB из Finegoldia magna (ранее известному как Peptostreptococcus magnus), известному как «модуль GA», который способен связывать альбумин (Lejon S., 2004. J. Biol. Chem., 279: 42924-42928). Белок PAB из Finegoldia magna представляет белок, определяемый последовательностью из базы данных в GenBank с инвентарным номером CAA54857.1 от 9 сентября 2004 года.
В рамках изобретения, термин «альбумин-связывающий пептид, имеющий коровую последовательность DICLPRWGCLW (SEQ ID NO:9)» относится к пептидам, которые связывают альбумин, полученный из фаговых клонов RA и SA, как описано Dennis M.S. et al. (J. Biol. Chem., 2002 277:35035-35043).
Изобретение также охватывает функционально эквивалентные варианты таких альбумин-связывающих участков. В рамках изобретения, термин «функционально эквивалентный вариант» относится к любому полипептиду, полученному из альбумин-связывающего участка посредством инсерции, делеции или замены одного или более остатков, и который по существу сохраняет способность взаимодействовать с альбумином, как определено выше. В предпочтительном варианте осуществления полипептид рассматривается в качестве функционально эквивалентного варианта альбумин-связывающего участка, если он проявляет способность в связывании с альбумином по меньшей мере на уровне 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% от способности в связывании с альбумином альбумин-связывающего домена SEQ ID NO: 1. Предпочтительно полипептид рассматривается в качестве функционально эквивалентного варианта альбумин-связывающего участка, если он способен нейтрализовать антитела с эффективностью по меньшей мере на уровне 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% от эффективности альбумин-связывающего домена SEQ ID NO:1.
Подходящими функциональными вариантами являются варианты, которые показывают степень идентичности по отношению к альбумин-связывающим доменам или альбумин-связывающим последовательностям, раскрытым в настоящем изобретении, по меньшей мере 25% идентичность аминокислотной последовательности, например, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99%. Степень идентичности между двумя полипептидами определяется с использованием компьютерных алгоритмов и методов, которые широко известны специалистам в данной области техники. Идентичность между двумя аминокислотными последовательностями предпочтительно определяют с использованием алгоритма BLASTP [BLAST Manual, Altschul S. et al., NCBI NIH Bethesda, MD. 20894, Altschul S. et al., J. Mol. Biol. 215: 403-410 (1990)], хотя, также можно использовать другие подобные алгоритмы. BLAST и BLAST 2.0 используются с параметрами, описанными в настоящем описании, для определения процента идентичности последовательностей. Программное обеспечение для осуществления анализов BLAST является общедоступным через Национальный центр биотехнологической информации.
Функционально эквивалентные варианты альбумин-связывающих участков могут представлять собой производные альбумин-связывающих доменов и альбумин-связывающих последовательностей. Термин «производные» включает, не ограничиваясь этим, альбумин-связывающие домены из бактерий, модифицированные с целью повышения их аффинности к альбумину, например, описанные Johansson M.U. et al. (J. Biol. Chem., 2002, 277:8114-8120), Jonsson A. et al. (Protein Eng. Des. Sel., 2008, 21:515-527) и Linhult М. et al. (Protein Sci., 2002, 11:206-213). Например, производное может представлять модифицированный стрептококковый G ABD ABD035, раскрытый Jonsson A. et al. (Protein Eng. Des. Sel., 2008, 21:515-527).
Рекомбинантные аденовирусы могут быть получены с помощью стандартных методов молекулярной биологии, известных в данной области техники (Chillon and Bosch. Adenovirus. Methods and Protocols. 3rd edition. Methods in Molecular Biology, 1089. Springer Protocols. Humana Press. (2014)).
Аденовирус, который содержит альбумин-связывающий участок по настоящему изобретению, размножается и амплифицируется согласно стандартным методам в области аденовирусных векторов, как описано Chillon and Bosch. Adenovirus. Methods and Protocols. 3rd edition. Methods in Molecular Biology, 1089. Springer Protocols. Humana Press. (2014); и Alemany R., Zhang W. Oncolytic adenoviral vectors. Totowa, NJ.:Humana Press, 1999. Клеточные линии, обычно используемые в области генной терапии и виротерапии, представляют клеточные линии НЕК-293 и А549. Предпочтительным способом размножения является инфицирование клеточной линии, которая обеспечивает репликацию аденовируса. Линия клеток аденокарциномы легкого А549 является примером такой клеточной линии. Размножение осуществляется, например, следующим образом: клетки А549 высевают в пластиковые планшеты для культивирования клеток и инфицируют из расчета 100 вирусных частиц на клетку. Через двое суток наличие цитопатического эффекта свидетельствует о продукции вируса, когда клетки отделяются, образуя «виноградоподобные» кластеры. Клетки собирают в пробирки. После центрифугирования при 1000 g в течение 5 мин клеточный осадок замораживают и оттаивают три раза, чтобы разрушить клетки. Полученный клеточный экстракт центрифугируют при 1000 g в течение 5 мин и супернатант, содержащий вирус, наслаивают на градиент хлорида цезия и центрифугируют в течение 1 ч при 35000 g. Собирают полосу вируса, полученную из градиента, и снова наслаивают на другой градиент хлорида цезия и центрифугируют в течение 16 ч при 35000 g. Собирают полосу вируса и подвергают диализу против смеси PBS-10% глицерин. Диализованный вирус распределяют на аликвотные порции и хранят при -80°С. Количественное определение числа вирусных частиц и бляшкообразующих единиц проводится согласно стандартным протоколам. Забуференный фосфатом физиологический раствор (PBS) с 5% глицерина является стандартным составом, используемым для хранения аденовируса. Тем не менее, были описаны другие составы, которые повышают стабильность вируса.
Методы очистки аденовирусов, содержащих альбумин-связывающий участок, для их применения в профилактике или лечении рака, являются такими же, как методы, описанные для других аденовирусов и аденовирусных векторов, используемых в виротерапии и генной терапии рака.
Аденовирус может быть использован для нацеливания на аномальные клетки, например, любые клетки, которые являются вредными или иным образом нежелательными in vivo. Широкий ряд примеров включает клетки, вызывающие аутоиммунное заболевание, рестеноз и образование рубцовой ткани.
Аденовирусы по изобретению могут быть селективно распределены in vivo в определенной ткани, что позволяет значительно сократить экспрессию в немишеневой или неопухолевой ткани.
Репликативный аденовирус по изобретению может иметь модификации в его геномной последовательности, придающие избирательную репликацию в клетке. Для того чтобы направить экспрессию аденовируса в ткань, где такая экспрессия необходима, или в опухолевую ткань, подлежащую лечению, аденовирус по изобретению может включать тканеспецифический промотор или опухолеспецифический промотор. Таким образом, в одном варианте осуществления аденовирус дополнительно содержит тканеспецифический промотор или опухолеспецифический промотор.
В предпочтительном варианте осуществления тканеспецифический промотор или опухолеспецифический промотор представляет собой промоторные последовательности для регуляции экспрессии одного или более генов, выбранных из группы, состоящей из Е1a, Е1b, Е2 и Е4. Предпочтительно промотор регулирует экспрессию Е1a.
В рамках изобретения, термин «промотор» используется согласно его общепризнанному в данной области значению. Он означает область ДНК, как правило, выше кодирующей последовательности гена, которая связывает РНК-полимеразу и направляет фермент в нужный сайт старта транскрипции. Указанный промотор контролирует вирусные гены, которые начинают репликацию.
Термин «тканеспецифический» означает, что промотор, с которым операбельно связан ген, существенный для репликации, функционирует специфически в этой ткани таким образом, что репликация протекает в этой ткани. Это может произойти в результате наличия в этой ткани, и отсутствия в немишеневых тканях, факторов транскрипции с позитивной регуляцией, которые активируют промотор. Это также может иметь место в результате отсутствия факторов, ингибирующих транскрипцию, которые обычно встречаются в немишеневых тканях и предупреждают транскрипцию за счет промотора. Таким образом, когда происходит транскрипция, то она протекает в ген, существенный для репликации, такой как в ткани-мишене, то происходит репликация вектора и сопутствующие функции.
Тканевая специфичность особенно важна в отношении нацеленности на аномальный аналог определенного типа ткани, одновременно избегая воздействия на нормальный аналог ткани или избегая воздействия на окружающие ткани другого типа, оказывая лечебный эффект на патологическую ткань. В конкретном варианте осуществления промотор представляет собой «опухольспецифический» промотор, означая, что промотор специфически функционирует в опухолевых тканях. Например, рекомбинантные аденовирусы по изобретению пригодны для лечения метастазов в печени. Одним конкретным примером является рак толстой кишки, который часто метастазирует в печень. Было установлено, что даже тогда, когда рак толстой кишки метастазирует в печень, то промотор СЕА активен в клетках метастазов, но не в нормальных клетках печени. Следовательно, печень здорового взрослого человека не будет поддерживать репликацию вируса, который имеет вирусные гены, необходимые для репликации, связанные с СЕА-специфическим промотором при раке толстой кишки. Репликация будет иметь место в первичных раковых клетках. Другим примером может служить промотор альфа-фетопротеина, который активен только в гепатоцеллюлярной карциноме. Еще одним примером является промотор тирозиназы, который активен только в меланоме, и не в нормальной коже. В каждом случае, как ожидается, репликация происходит в аномальных, но не в нормальных клетках.
Примерами тканеспецифичных промоторов являются, не ограничиваясь этим, промотор альфа-фетопротеина, промотор DE3, промотор тирозиназы, промотор карциноэмбрионального антигена (СЕА), промотор сурфактантного белка, промотор E2F, промотор теломеразы hTERT, промотор простат-специфического антигена, промотор СОХ-2, промотор гена альбумина, коровый промотор вируса гепатита, промотор глобулин-связывающего белка, который связывается с тироксином, и промотор ErbB2.
В предпочтительном варианте осуществления промотор выбран из группы, состоящей из промотора E2F, промотора теломеразы hTERT, промотора тирозиназы, промотора простат-специфического антигена, промотора альфа-фетопротеина и промотора СОХ-2.
Аденовирусы по изобретению особенно пригодны для лечения рака. Все опухоли потенциально поддаются лечению аденовирусом по изобретению. Типы опухолей включают, не ограничиваясь этим, гематопоэтические опухоли, опухоли поджелудочной железы, опухоли нервной системы, опухоли печени, желудочно-кишечного тракта, эндокринной системы, желчных путей, синопульмонарные опухоли, опухоли головы и шеи, саркому и карциному мягких тканей, опухоли кожи, опухоли репродуктивного тракта и тому подобное. Предпочтительные опухоли для лечения представляют собой опухоли с высоким митотическим индексом по сравнению с нормальной тканью, предпочтительно солидные опухоли.
В предпочтительном варианте осуществления аденовирус по изобретению представляет собой онколитический аденовирус.
В рамках изобретения, термин «онколитический аденовирус» относится к любому аденовирусу, который способен реплицироваться или который является компетентным по репликации в опухолевых клетках, даже без селективности. Терапевтическое действие онколитических аденовирусов основано на их способности реплицироваться и лизировать опухолевые клетки, которые должны быть элиминированы. Гибель опухолевых клеток можно детектировать с помощью любого метода из уровня техники, например, определением количества жизнеспособных клеток, цитопатического эффекта, апоптоза опухолевых клеток, синтеза вирусных белков в опухолевых клетках (например, метаболическое мечение, вестерн-блот вирусных белков или ПЦР с обратной транскрипцией вирусных генов, необходимых для репликации) или определением уменьшения размеров опухоли.
Другой стратегией для достижения селективной репликации в опухолях является элиминация вирусных функций, которые необходимы для репликации в нормальных клетках, но которые не нужны в опухолевых клетках. Это включает, например, элиминацию ранних функций Е1А, который блокируют путь ретинобластомы (PRB). Селективная репликация таких мутантов была показана в нескольких документах предшествующего уровня техники. Другие вирусные гены, которые непосредственно взаимодействуют с pRB, такими как E4 и E4orf6/7, являются кандидатами для делеции с целью достижения селективной репликации в опухолевых клетках.
Еще одной модификацией, описанной для достижения селективной репликации в опухолях, является делеция аденовирусных генов, кодирующих вирус-ассоциированные РНК (VA-РНК). Эти РНК блокируют противовирусную активность интерферона, и их делеция приводит к получению аденовирусов, которые чувствительны к ингибированию интерфероном. За счет характерного усечения на пути интерферона в опухолевых клетках, такие аденовирусы обычно реплицируются в опухолях.
Таким образом, в еще одном варианте осуществления аденовирус по изобретению дополнительно содержит мутации в одном или более генах, выбранных из группы, состоящей из Е1а, Е1b, Е4 и VA-РНК, для достижения селективной репликации в опухолях. Предпочтительно мутации находятся в Е1а. В предпочтительном варианте осуществления мутация в Е1а представляет делецию некоторых аминокислот белка Е1А, влияющих на взаимодействие Е1А с pRB, предпочтительно представляет делецию аминокислот 121-129 полипептидной цепи (делеция Δ24).
В рамках изобретения, выражение «селективная репликация» означает, что аденовирус обладает более высокой эффективностью репликации в опухолевых клетках, чем в нормальных клетках (например, в 1000 раз выше, чем в нормальных клетках).
В рамках изобретения, термин «репликация» относится к удвоению количества аденовирусных векторов, которое происходит на уровне нуклеиновой кислоты или на уровне инфекционной вирусной частицы. В случае ДНК-содержащих вирусов, репликация на уровне нуклеиновой кислоты представляет репликацию ДНК. Однако репликация также включает образование инфекционных ДНК-содержащих вирусных частиц.
Репликацию аденовируса можно анализировать с помощью хорошо известных методик. Анализы репликации аденовирусного вектора в клетке обычно включают детектирование полинуклеотида, вирионов или инфекционного вируса. Разнообразие хорошо известных методов, которые могут быть использованы для этой цели, включает определение количества меченого субстрата, включенного в полинуклеотид, в течение определенного периода в клетке.
Когда репликация включает ДНК-полинуклеотид, то часто в качестве меченого субстрата используется 3H-тимидин. В этом случае уровень репликации определяется отделением ДНК вектора от массы клеточной ДНК и измерением количества трития, специфически включенного в ДНК вектор.
Репликацию полинуклеотидного вектора также можно детектировать посредством лизиса или пермеабилизации клеток для высвобождения полинуклеотида, затем выделением полинуклеотида и количественным определением непосредственно ДНК или РНК, которая выделена. Репликация полинуклеотида также может быть детектирована с помощью количественной ПЦР с использованием праймеров, специфичных для анализируемого полинуклеотида.
Вирионы можно анализировать с помощью методов подсчета на основе электронной микроскопии, хорошо известных в данной области техники, и выделением вирионов и определением содержания белка и нуклеиновых кислот, и мечением вирусных геномных полинуклеотидов или белков вириона и определением количества вириона по количеству полинуклеотида или белка.
Другая стратегия для достижения селективности аденовируса для опухолевой клетки заключается в модификации белков капсида вируса, принимающих участие в инфицировании клетки-хозяина, для нацеленности аденовируса к рецептору, находящемуся в опухолевой клетке. Модификация белков капсида, которые используются вирусом для инфицирования клетки, также может быть использована для повышения инфекционной активности аденовируса (т.е. повышения проникновения вируса в клетку). Нацеленность аденовируса на опухоль также может быть достигнута с помощью бифункциональных лигандов, которые связаны с вирусом в одном конце и с рецептором опухоли в другом.
Таким образом, в еще одном варианте осуществления аденовирус по изобретению дополнительно включает модификации капсида для повышения его инфекционной активности или его нацеленности к рецептору, находящемуся в опухолевой клетке. В более предпочтительном варианте осуществления модификация капсида представляет инсерцию мотива RGD (мотива аргинин-глицин-аспарагин) в петлю H1 аденовирусного белка волокна. Данная инсерция позволяет аденовирусу использовать интегрины для «прикрепления» в клетке, и не только для интернализации, как это имеет место с аденовирусом дикого типа. Использование интегринов в качестве клеточных рецепторов вируса повышает его инфекционную активность и онколитическую эффективность. В еще одном варианте осуществления онколитический аденовирус имеет капсид, модифицированный посредством замены гепаринсульфат-связывающего домена KKTK (SEQ ID NO: 10) в аденовирусном волокне на домен RGDK (SEQ ID NO: 11) (N. Bayo et al., Human Gene Therapy, 2009, 20:1214-21). Другой стратегией для повышения инфекционной активности аденовирусов для клеток-мишеней является замена фрагмента волокна гомологичным фрагментом из другого серотипа. Обычно shaft-домен и knob-домен волокна человеческих аденовирусов, полученных из серотипа 5, замещаются на shaft-домен и knob-домен волокна человеческих аденовирусов серотипа 3 или 35. Полученные рекомбинантные аденовирусы с геномами, происходящими из других серотипов, известны в данной области техники как химерные аденовирусы. В еще одном варианте осуществления аденовирус по изобретению дополнительно содержит химерный капсид, полученный из аденовирусов различных серотипов. В еще одном предпочтительном варианте осуществления модификация капсида представляет замену фрагмента гена волокна гомологичным фрагментом из аденовируса другого серотипа с образованием химерного аденовируса.
В предпочтительном варианте осуществления онколитический аденовирус представляет опухолеселективный реплицирующийся аденовирус, отличающийся содержанием мутантного варианта белка Е1a, где аминокислоты 121-129 полипептидной цепи делецированы (делеция Δ24), что оказывает влияние на взаимодействие Е1а с pRB, инсерцией четырех сайтов связывания E2F и одного сайта связывания Sp1 в эндогенный промотор Е1А для регуляции экспрессии Е1a, и, наконец, инсерцией пептида RGD в аденовирусное волокно для повышения инфекционной активности вируса. ICOVIR15-ABD представляет предпочтительный вариант осуществления настоящего изобретения. Указанные модификации могут присутствовать в комбинации в одном и том же аденовирусе или по отдельности.
В предпочтительном варианте осуществления онколитический аденовирус представляет опухолеселективный реплицирующийся аденовирус, отличающийся наличием делеции некоторых аминокислот белка Е1a, влияющих на взаимодействие Е1a с pRB, предпочтительно делецию аминокислот 121-129 полипептидной цепи (делеция Δ24).
В еще одном предпочтительном варианте осуществления онколитический аденовирус представляет собой опухолеселективный реплицирующийся аденовирус, отличающийся наличием инсерции четырех сайтов связывания E2F и одного сайта связывания Sp1 в эндогенный промотор Е1a для регуляции экспрессии Е1a.
В еще одном предпочтительном варианте осуществления онколитический аденовирус представляет опухолеселективный реплицирующийся аденовирус, отличающийся наличием инсерции пептида RGD в аденовирусное волокно для повышения инфекционной активности вируса.
Геном аденовируса также может содержать гетерологичный ген, который кодирует терапевтический белок, таким образом, что гетерологичный ген экспрессируется в инфицированной клетке. В рамках изобретения, термин «терапевтический белок» относится к белку, который, как можно ожидать, обеспечивает определенный лечебный эффект при экспрессии в данной клетке. Указанные гетерологичные генные продукты могут иметь место в реплицирующемся или нереплицирующемся аденовирусе. Вставленный терапевтический ген может представлять любой ген, который используется в генной терапии или в вакцинации. Предпочтительно гетерологичный ген используется в генной терапии рака. Инсерция терапевтического гена в геном онколитического аденовируса дает «вооруженный онколитический аденовирус», что повышает цитотоксичность онколитического аденовируса по отношению к опухолевым клеткам. Например, указанный гетерологичный ген может, среди прочего, привести к гибели опухолевой клетки, активировать иммунную систему против опухоли, ингибировать ангиогенез, элиминировать внеклеточный матрикс, индуцировать апоптоз. В этих случаях, каким образом и когда будет осуществляться экспрессия терапевтического гена, будет иметь решающее значение для конечного результата терапевтического подхода.
Следовательно, в одном варианте осуществления изобретения аденовирус содержит один или более неаденовирусных генов, вставленных в геном указанного аденовируса. В предпочтительном варианте осуществления гены представляют гены, используемые в генной терапии или в вакцинации. В более предпочтительном варианте осуществления гены представляют гены, используемые в генной терапии рака. Предпочтительно гены, используемые в генной терапии рака, по меньшей мере, представляют ген, выбранный из группы, состоящей из пролекарство-активирующих генов, генов-супрессоров опухолей, генов, кодирующих противоопухолевые интерферирующие РНК, и иммуностимулирующих генов.
В рамках изобретения, термин «неаденовирусный ген» относится к гетерологичному гену, отсутствующему в геноме аденовируса дикого типа.
В рамках изобретения, термин «ген, используемый в генной терапии» относится к гену, который может быть использован в качестве лекарственного средства для профилактики или лечения наследственного или приобретенного заболевания или состояния посредством доставки указанной терапевтической ДНК в клетки пациента. Специалисту в данной области техники должно быть понятно, что термин «генная терапия» включает использование ДНК, которая кодирует функциональный, терапевтический ген, для замены мутантного гена, или использование ДНК, которая кодирует терапевтический белок. Например, ДНК может кодировать фермент, гормон, рецептор или полипептид, которые имеют терапевтическую ценность. Любой ген, который может быть использован для лечения заболевания, которое подходит для лечения генной терапией, может быть вставлен в геном аденовируса по изобретению. Гены, используемые в генной терапии, могут представлять собой, без ограничения, гены, кодирующие ферменты, производные крови, гормоны, интерлейкины, интерфероны, TNF, факторы роста, нейротрансмиттеры или их предшественники, или синтетические ферменты, трофические факторы, например, BDNF, CNTF, NGF, IGF, GMF, aFGF, bFGF, NT3, NT5 и тому подобное; аполипопротеины, например, ApoAI, ApoAIV, ApoE, и тому подобное; дистрофин или минидистрофин; гены-супрессоры опухолей, например, р53, Rb, RaplA, DCC, к-rev; гены, кодирующие факторы, участвующие в свертывании крови, например, факторы VII, VIII, IX; пролекарство-активирующие гены, например, тимидинкиназа, цитозиндезаминаза; весь или фрагмент природного или искусственного иммуноглобулина (Fa, ScFv и тому подобное). Терапевтический ген также может представлять собой антисмысловой ген или последовательность, экспрессия которой в клетке-мишени обеспечивает экспрессию генов или транскрипцию клеточных мРНК для регуляции.
В рамках изобретения, термин «ген, используемый в вакцинации» относится к гену, кодирующему антигенный пептид, способный генерировать иммунный ответ у человека или животных, с целью получения профилактической или терапевтической вакцины. Такие антигенные пептиды могут представлять собой, без ограничения, специфические для вируса Эпштейна-Барра, вируса ВИЧ, вируса гепатита В, вируса болезни Ауески и опухолеспецифические пептиды.
В рамках изобретения, термин «пролекарство-активирующие гены» относится к генам, кодирующим продукт, который действует на нетоксичное пролекарство, превращая нетоксичное пролекарство в форму, которая является токсичной для ткани-мишени. Предпочтительно, чтобы токсин обладал противоопухолевой активностью или элиминировал клеточную пролиферацию.
Примеры пролекарство-активирующих генов включают, не ограничиваясь этим, ген тимидинкиназы. Тимидинкиназа вируса герпеса простого катализирует фосфорилирование ганцикловира с образованием токсичного ганцикловира нуклеотидфосфата. Данное соединение функционирует в качестве терминатора транскриции и ингибитора ДНК-полимеразы, предупреждает синтез ДНК и, таким образом, является цитотоксическим. В одном варианте осуществления пролекарство-активирующим геном является ген тимидинкиназы, предпочтительно вирусной тимидинкиназы, выбранной из группы, состоящей из тимидинкиназы вируса герпеса простого, тимидинкиназы цитомегаловируса и тимидинкиназы вируса ветряной оспы. Когда используются вирусные тимидинкиназы, то взаимодействующий агент или химиотерапевтическое средство предпочтительно представляет аналог нуклеозидов, например, выбранный из группы, состоящей из ганцикловира, ацикловира и 1-2-дезокси-2-фтор-D-арабинофуранозил-5-иодурацила (FIAU). Такие взаимодействующие агенты эффективно используются вирусными тимидинкиназами в качестве субстратов, и такие взаимодействующие агенты, таким образом, включаются летально в ДНК опухолевых клеток, экспрессирующих вирусные тимидинкиназы, что приводит к гибели клеток-мишеней. В еще одном варианте осуществления пролекарство-активирующий ген является цитозиндезаминазой. Цитозиндеаминаза катализирует превращение 5'-фторцитозина в противораковый препарат, 5'-фторурацил, который является высоко цитотоксическим. Таким образом, клетки-мишени, которые экспрессируют ген цитозиндеаминазы, превращают 5-фторцитозин в 5-фторурацил и погибают. Обсуждение таких «суицидных» генов, смотри в публикации Blaese Р.М. et al., Eur. J. Cancer 30A:1190-1193 (1994).
В рамках изобретения, термин «гены-супрессоры опухолей» относится к антионкогенам или генам, которые защищают клетки от одного шага на пути к раку. Когда один из этих генов мутирует, вызывая потерю или снижение его функции, то клетка может прогрессировать к раку, как правило, в сочетании с другими генетическими изменениями.
Примерами генов-супрессоров опухолей являются, не ограничиваясь этим, белок-супрессор опухолей р53, кодируемый геном ТР53, PTEN, pVHL, APC, CD95, ST5, YPEL3, ST7 и ST14.
В рамках изобретения, термин «гены, кодирующие противоопухолевые интерферирующие РНК» относится к генам, кодирующим терапевтически полезные молекулы РНК для лечения опухолей, т.е. siРНК (Dorsett and Tuschl (2004) Nature Rev. Drug Disc., 3:318-329). В некоторых случаях гены могут быть включены в рекомбинантный аденовирус по изобретению для дополнительного повышения способности аденовируса вызывать гибель клетки линии дифференцировки моноцитов/макрофагов, однако, не оказывая какого-либо прямого воздействия на саму клетку. Они включают гены, кодирующие siРНК, способные ингибировать активность факторов, негативно влияющих на презентацию молекулами MHC класса I, блокировать комплемент, ингибировать IFN и IFN-индуцированные механизмы, хемокины и цитокины, гибель, опосредованную NK-клетками, отрицательную регуляцию иммунного ответа (например, IL-10, TGF-бета) и металлопротеиназы, которые могут разрушить внеклеточный матрикс и усилить распространение вируса в опухоли.
В рамках изобретения, термин «иммуностимулирующие гены» относится к генам, которые активируют иммунную систему против опухоли. Другие примеры гетерологичных генов или их фрагментов, включают таковые, которые кодируют иммуномодулирующие белки, такие как цитокины или хемокины. Примеры включают интерлейкин-2, патенты США № 4738927 или 5641665; интерлейкин-7, патенты США № 4965195 или 5328988; и интерлейкин-12, патент США № 5457038; фактор некроза опухолей альфа, патенты США № 4677063 или 5773582; интерферон-гамма, патенты США № 4727138 или 4762791; или GM CSF, патенты США № 5393870 или 5391485, Mackensen et al. (1997) Cytokine Growth Factor Rev. 8:119-128).
Эти модификации в геноме аденовируса не исключают друг друга.
Аденовирусные геномы по изобретению
Настоящее изобретение также относится к геному аденовируса. В одном аспекте настоящее изобретение относится к аденовирусному геному, отличающемуся тем, что он содержит последовательность, кодирующую альбумин-связывающий участок, вставленный в кодирующую область гипервариабельного участка 1 (HVR1) гексонного белка, что приводит к экспрессии слитого белка, включающего гексонный белок и альбумин-связывающий участок, и где альбумин-связывающий участок располагается на наружной поверхности гексонного белка, когда сборка гексонного белка имеет место в аденовирусном капсиде.
В рамках изобретения, выражение «аденовирусный геном» относится к последовательности двухцепочечной ДНК, которая в присутствии соответствующих белков, может быть упакована, приводя к образованию полной аденовирусной частицы. Для того, чтобы имела место такая упаковка, последовательность должна соответствовать некоторым условиям, которые могут быть обобщены следующим образом:
- иметь отдельный аденовирусный ITR, по одному на каждом из ее концов;
- включать сигнал упаковки Psi между обоими ITR, расположенными таким образом, чтобы расстояние между 5'-концом сигнала упаковки Psi и 3'-концом ближайшего к нему ITR не превышало расстояние, которое будет препятствовать упаковке природного аденовируса, расстояние, которое составляет 200 п.н. в случае человеческого аденовируса серотипа 5, и которое, как предполагается, по аналогии, должно быть примерно равным для других серотипов, поскольку как было видно, введение последовательностей между ITR и сигналом упаковки в последовательности, которая обычно разъединяет их, уменьшает способность аденовируса к упаковке, вызывая уменьшение общего числа полученных аденовирусных частиц, несмотря на отсутствие существенных изменений во времени, необходимом для их упаковки;
- расстояние между концами обоих ITR не должно составлять более чем 105% от размера генома аденовируса, присутствующего в природе, к которому относятся белки, образующие капсид.
Аденовирусный геном предпочтительно является дефицитным, по меньшей мере, по одной функции гена, существенного для репликации вируса, что приводит к «дефицитному по репликации» аденовирусному вектору. Под термином «дефицитный по репликации» подразумевается, что аденовирусный вектор содержит аденовирусный геном, в котором отсутствует по меньшей мере одна функция существенного для репликации гена (т.е. так, что аденовирусный вектор не реплицируется в типичных клетках-хозяевах, в частности, в клетках пациента-человека, которые могли бы быть заражены аденовирусным вектором в процессе лечения согласно настоящему изобретению).
Более предпочтительно, дефицитный по репликации аденовирусный вектор содержит аденовирусный геном, дефицитный по меньшей мере по одной функции гена, существенного для репликации, одной или более областей генома аденовируса. В связи с этим, аденовирусный вектор является дефицитным по меньшей мере по одной функции существенного гена области Е4 или области E1 аденовируса, необходимых для репликации вируса. В дополнение к дефициту в области Е1 рекомбинантный аденовирус также может содержать мутацию в главном позднем промоторе (MLP). Более предпочтительно, аденовирусный вектор является дефицитным по меньшей мере по одной функции существенного гена области E1 и по меньшей мере части области E3 (например, делеция Xba I области Е3). В отношение области E1, то аденовирусный вектор может быть дефицитным (например, в результате делеции) по меньшей мере по части области Е1a и по меньшей мере части области Е1b. Например, аденовирусный вектор может содержать делецию всей области Е1 и части области E3 аденовируса (т.е., нуклеотиды 355-3511 и 28593-30470). В однократно дефицитном аденовирусном векторе может быть делецировано примерно от 356 до 3329 и от 28594 до 30469 нуклеотидов (основываясь на геноме аденовируса серотипа 5). Альтернативно в геноме аденовирусного вектора может быть делецировано примерно от 356 до 3510 и от 28593 до 30470 нуклеотидов (основываясь на геноме аденовируса серотипа 5), в результате чего аденовирусный вектор имеет делеции в областях E1, Е3 и Е4 аденовирусного генома.
В рамках изобретения, термин «дефицит в гене, функции гена или гене или геномной области» определяется как делеция достаточного количества генетического материала вирусного генома с ухудшением или отменой функции гена, нуклеиновокислотная последовательность которого была делецирована полностью или частично. Делеции целой области гена часто не требуется для разрушения функции гена, существенного для репликации. Однако с целью обеспечения достаточного пространства в аденовирусном геноме для одного или более трансгенов, может быть желательной делеция большей части области гена. Несмотря на то, что делеция генетического материала является предпочтительной, мутация генетического материала посредством добавления или замены также подходит для нарушения функции гена. Функции гена, существенного для репликации, представляют такие функции генов, которые необходимы для репликации (например, размножения) и кодированы, например, аденовирусными ранними областями (например, области El, E2 и E4), поздними областями (например, области L1-L5), генами, участвующими в вирусной упаковке (например, ген IVa2) и ассоциированные с вирусом РНК (например, NA-РНК-1 и/или NA-РНК-2).
Аденовирусный вектор также может иметь делецированный по существу весь аденовирусный геном, за исключением ITR и последовательности упаковки. Такие векторы известны в данной области техники как «лишенные существенного внутреннего содержимого» - «gutless» или хелпер-зависимые аденовирусные векторы. В этом случае последовательность гексона модифицирована таким образом, чтобы было обеспечено содержание альбумин-связывающего участка хелперного аденовируса. 5'- или 3'-области аденовирусного генома, содержащего ITR и последовательность упаковки, не должны происходить из аденовируса одного и того же серотипа, что и остальная часть вирусного генома. Например, 5'-область аденовирусного генома серотипа 5 (то есть, область генома 5' к аденовирусной области E1) может быть заменена соответствующей областью аденовирусного генома серотипа 2 (например, область генома Ad5 5' к области генома Е1 аденовируса замещается нуклеотидами 1-456 генома Ad2). Однако дефицит аденовирусного генома аденовирусного вектора согласно способу по настоящему изобретению предпочтительно ограничивается функциями гена, существенного для репликации, кодируемого ранними областями генома аденовируса.
Согласно изобретению инвертированный концевой повтор или ITR понимается как последовательности примерно из 100 п.н., которые находятся на обеих сторонах линейного генома аденовируса и которые имеют существенное значение для репликации аденовирусного генома (Stow N.D., 1982, Nucl. Acid Res, 10:5105-5109).
Согласно изобретению аденовирусный сигнал упаковки ψ понимается как последовательность примерно из 160 п.н., которая, в случае аденовируса серотипов 2 и 5, простирается между положениями 190 и 350 генома. Элиминация последовательности генома аденовируса не дает молекулам ДНК, которые образуются в процессе размножения вируса, эффективно включаться в недавно образовавшийся капсид (Hearing P. et al., 1987, J. Virol., 61:2555-2558), но она не предупреждает репликацию генома в «упаковочной» клетке, в отличие от элиминации ITR.
Все варианты осуществления, раскрытые в контексте аденовирусов по изобретению, применимы к аденовирусным геномам по изобретению.
В частности, в варианте осуществления аденовирусный геном происходит из аденовируса человека, предпочтительно выбранного из группы, состоящей из аденовируса человека серотипов 1-57, более предпочтительно серотипа 5.
В еще одном варианте осуществления альбумин-связывающий участок выбран из альбумин-связывающего домена из стрептококкового белка G, альбумин-связывающего домена из белка PAB Peptostreptococcus magnus, альбумин-связывающего пептида, имеющего коровую последовательность DICLPRWGCLW (SEQ ID NO:9), и их функционально эквивалентных вариантов. В более предпочтительном варианте осуществления изобретения альбумин-связывающий участок представляет альбумин-связывающий домен 3 из стрептококкового белка G, предпочтительно имеющий последовательность (SEQ ID NO:1).
В еще одном варианте осуществления последовательность, кодирующую альбумин-связывающий участок, вставляют таким образом, что полученный слитый белок содержит альбумин-связывающий участок после аминокислоты D150 гексонного белка согласно нумерации гексонного белка, имеющего инвентарный номер в GenBank BAG48782.1. В предпочтительном варианте осуществления изобретения нуклеотидная последовательность полного модифицированного аденовирусного гексона, имеющего ABD, вставленный в HVR1 (ABD-HVR1), представляет SEQ ID NO:3.
В еще одном варианте осуществления N- и/или С-конец альбумин-связывающего участка связан с гексонным белком линкерной последовательностью, где предпочтительно линкерная последовательность содержит последовательность GSGS (SEQ ID NO:2).
В еще одном варианте осуществления аденовирусный геном дополнительно содержит тканеспецифический промотор или опухолеспецифичный промотор. В предпочтительном варианте тканеспецифический промотор или опухолеспецифический промотор представляют промоторные последовательности для регуляции экспрессии одного или более генов, выбранных из группы, состоящей из E1a, E1b, E2 и E4; более предпочтительно промотор выбран из группы, состоящей из промотора E2F, промотора теломеразы hTERT, промотора тирозиназы, промотора простат-специфического антигена, промотора альфа-фетопротеина и промотора СОХ-2.
В еще одном варианте осуществления изобретения аденовирус представляет онколитический аденовирус, предпочтительно аденовирус, где его аденовирусный геном дополнительно содержит мутации в одном или более генов, выбранных из группы, состоящей из Е1а, Е1В, Е4 и VA-РНК, для достижения селективной репликации в опухолях.
В еще одном варианте осуществления аденовирусный геном дополнительно содержит модификации капсида для повышения инфекционной активности аденовируса или его нацеленности на рецептор, присутствующий в опухолевой клетке. Предпочтительно модификация капсида представляет инсерцию мотива RGD в петлю H1 аденовирусного белка волокна. В еще одном варианте осуществления аденовирусный геном представляет химерный геном аденовируса, полученный из одного данного серотипа, который содержит фрагмент или часть его генома, замещенные гомологичным фрагментом генома другого серотипа. Предпочтительно указанный химерный аденовирус представляет аденовирус человека серотипа 5, который содержит фрагмент гена волокна, замещенный гомологичным фрагментом другого серотипа, предпочтительно аденовируса человека 3 или аденовируса человека 35. В предпочтительном варианте осуществления модификация капсида представляет замещение фрагмента гена волокна гомологичным фрагментом аденовируса другого серотипа с образованием химерного аденовируса.
В еще одном варианте осуществления аденовирусный геном содержит дополнительные гены, вставленные в указанный геном. В варианте осуществления указанные гены используются в генной терапии или в вакцинации. Предпочтительно указанные гены представляют гены, используемые в генной терапии рака, более предпочтительно, по меньшей мере, ген, выбранный из группы, состоящей из пролекарство-активирующих генов, генов-супрессоров опухолей, генов, кодирующих противоопухолевые интерферирующие РНК и иммуностимулирующих генов.
Композиции по изобретению
Рекомбинантные аденовирусы по изобретению можно использовать для получения фармацевтической композиции. Таким образом, еще один аспект настоящего изобретения представляет фармацевтическую композицию, содержащую терапевтически эффективное количество рекомбинантного аденовируса по настоящему изобретению вместе с фармацевтически приемлемым носителем.
В рамках изобретения, термин «фармацевтическая композиция» относится к композиции, которая адаптирована для введения заранее определенной дозы одного или более терапевтических полезных агентов в клетку, группу клеток, орган, ткань или организм.
Рекомбинантные аденовирусы вводятся в эффективных количествах. Термин «терапевтически эффективное количество» понимается как количество, способное обеспечить терапевтический эффект, и которое может быть определено специалистом в данной области с использованием обычно используемых средств. Эффективное количество будет варьировать в зависимости от конкретного состояния, подлежащего лечению, возраста и физического состояния пациента, который подвергается лечению, тяжести состояния, продолжительности лечения, характера сопутствующей или комбинированной терапии (если таковые имеются), конкретного пути введения и других подобных факторов, находящихся в пределах знаний и опыта практикующего врача. Предпочтительно, в общем, чтобы использовалась максимальная доза, то есть, самая высокая безопасная доза с точки зрения врача. Например, если пациент имеет опухоль, то эффективное количество может представлять такое количество, которое уменьшает объем опухоли или нагрузку (что, например, определяется посредством визуализации опухоли). Эффективные количества также можно оценить по наличию и/или количеству раковых клеток в крови или другой жидкости или ткани организма (например, с помощью биопсии). Если опухоль оказывает отрицательное влияние на нормальное функционирование ткани или органа, то эффективное количество может быть определено оценкой нормального функционирования ткани или органа. Специалистам в данной области техники должно понятно, что дозы могут быть также определены с использованием руководства Goodman and Goldman's The Pharmacological Basis of Therapeutics, Ninth Edition (1996), Appendix II, pp. 1707-1711 и руководства Goodman and Goldman's The Pharmacological Basis of Therapeutics, Tenth Edition (2001), Appendix II, pp. 475-493.
В рамках изобретения, термин «фармацевтически приемлемый носитель» означает нетоксичный, инертный, твердый, полутвердый или жидкий наполнитель, разбавитель, инкапсулирующий материал или вспомогательное вещество любого типа, приемлемые для пациента с фармакологической/токсикологической точки зрения и для производства фармацевтической химии с физической/химической точки зрения относительно композиции, формуляции, стабильности, переносимости пациентом и биодоступности. В монографии Remington's Pharmaceutical Sciences. Ed. by Gennaro, Mack Publishing, Easton, Pa., 1995 раскрываются различные носители, используемые в получении фармацевтических композиций, и известные способы их получения. Некоторые примеры материалов, которые могут служить в качестве фармацевтически приемлемых носителей, включают, не ограничиваясь этим, сахара, такие как лактоза, глюкоза и сахароза; крахмалы, такие как кукурузный крахмал и картофельный крахмал; целлюлоза и ее производные, такие как натриевая соль карбоксиметилцеллюлоза, этилцеллюлоза и ацетат целлюлозы; порошкообразный трагакант; солод; желатин; тальк; наполнители, такие как какао-масло и воска для суппозиториев; масла, такие как арахисовое масло, хлопковое масло; сафлоровое масло; кунжутное масло; оливковое масло; кукурузное масло и соевое масло; гликоли, такие как пропиленгликоль; сложные эфиры, такие как этилолеат и этиллаурат; агар; поверхностно-активные вещества, такие как твин™ 80; буферные агенты, такие как гидроксид магния и гидроксид алюминия; альгиновую кислоту; не содержащую пирогенов воду; изотонический солевой раствор; раствор Рингера; этиловый спирт; и фосфатные буферные растворы, а также другие нетоксичные совместимые смазывающие вещества, такие как лаурилсульфат натрия и стеарат магния, а также красители, высвобождающие агенты, агенты для покрытий, подсластители, вкусовые и ароматизирующие агенты, консерванты и антиоксиданты также могут присутствовать в композиции, согласно мнению составителя. Если фильтрация или другие методы конечной стерилизации не применимы, то композиции могут быть получены в асептических условиях.
Фармацевтические композиции по изобретению можно вводить пациенту любыми способами, известными в данной области техники, включая пероральный и парентеральный пути. Согласно таким вариантам осуществления композиции по настоящему изобретению можно вводить инъекцией (например, внутривенной, подкожной или внутримышечной, внутрибрюшинной инъекцией). В конкретном варианте осуществления рекомбинантные аденовирусы по настоящему изобретению вводят пациенту системно, например, внутривенной инфузией или инъекцией. Инъекционные препараты, например стерильные инъекционные водные или масляные суспензии, могут быть приготовлены в соответствии с известными в данной области техники способами с использованием подходящих диспергирующих или смачивающих агентов и суспендирующих агентов. Стерильный инъекционный препарат также может представлять собой стерильный инъекционный раствор, суспензию или эмульсию в нетоксичном, приемлемом для парентерального введения разбавителе или растворителе, например, в виде раствора в 1,3-бутандиоле. Среди приемлемых носителей и растворителей, которые можно использовать, можно указать воду, раствор Рингера, фармакопея США, и изотонический раствор хлорида натрия. Кроме того, стерильные нелетучие масла обычно используются в качестве растворителя или суспендирующей среды. С этой целью может быть использовано любое мягкое нелетучее масло, включая синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, используются в приготовлении инъекционных препаратов. В одном варианте осуществления рекомбинантный аденовирус суспендируют в жидком носителе, содержащем 1% (мас./ об.) натриевой соли карбоксиметилцеллюлозы и 0,1% (об./об.) твина™ 80. Инъекционные составы можно стерилизовать, например, фильтрацией через фильтр, задерживающий бактерии, или включением стерилизующих агентов в форме стерильных твердых композиций, которые могут быть растворены или диспергированы в стерильной воде или другой стерильной среде для инъекций непосредственно перед использованием.
Композиции могут включать рекомбинантный аденовирус в качестве единственного активного агента, или в комбинации с другим терапевтическим агентом.
В предпочтительном варианте осуществления композиция содержит онколитический аденовирус или аденовирус, содержащий один или более генов, используемых в генной терапии рака, вставленных в геном аденовируса. В данном конкретном случае композиции могут включать этот аденовирус в качестве единственного активного агента для лечения опухоли, или в комбинации с другим терапевтическим агентом, таким как химиотерапевтический препарат или вектор со вставленным терапевтическим геном.
Терапевтические применения аденовируса по изобретению
Имеется большой опыт в применении дефектных по репликации и компетентных по репликации аденовирусов в области генной терапии, и в области вакцинации.
В еще одном аспекте настоящее изобретение относится к рекомбинантному аденовирусу по изобретению или фармацевтической композиции по изобретению для применения в медицине. Аденовирусы по изобретению могут быть предназначены для лечения любого вида заболевания, для которого требуется введение такого аденовируса в кровоток.
Авторы настоящего изобретения также показали, что аденовирусы по изобретению особенно пригодны для лечения рака.
В еще одном аспекте настоящее изобретение относится к рекомбинантному аденовирусу по изобретению или фармацевтической композиции по изобретению для применения в профилактике и/или лечении рака у млекопитающего, где аденовирус представляет онколитический аденовирус или аденовирус, содержащий один или более генов, используемых в генной терапии рака, вставленных в геном аденовируса.
Альтернативно изобретение относится к применению рекомбинантного аденовируса по изобретению или фармацевтической композиции по изобретению для производства лекарственного средства для профилактики и/или лечения рака у млекопитающего, где аденовирус является онколитическим аденовирусом или аденовирусом, содержащим один или более генов, используемых в генной терапии рака, вставленных в геном аденовируса.
Альтернативно изобретение относится к способу профилактики и/или лечения рака у млекопитающего, включающему введение указанному млекопитающему рекомбинантного аденовируса по изобретению или фармацевтической композиции по изобретению, где аденовирус является онколитическим аденовирусом или аденовирусом, содержащим один или более генов, используемых в генной терапии рака, вставленных в геном аденовируса.
В рамках изобретения, термин «профилактика» относится к профилактическому или предупреждающему способу, где аденовирус вводят на начальной или ранней стадии болезни (то есть, предзлокачественной стадии развития опухоли), или также для предупреждения ее начала.
Аденовирусные векторы обычно использовались для приготовления вакцин, которые индуцируют иммунитет против белков патогенных микроорганизмов. Например, были широко описаны рекомбинантные аденовирусные вакцины против ВИЧ, вируса бешенства, вируса денге, вируса Эбола, коронавируса или возбудителя ТОРС, вируса папилломы человека, вируса гепатита С, вируса гепатита В, ротавируса, вируса кори, респираторно-синцитиального вируса, цитомегаловируса, вируса герпеса простого 2 типа, вируса Эпштейна-Барра, вируса гриппа, Trypanosoma cruzi и Plasmodium falciparum и других.
В еще одном аспекте настоящее изобретение относится к рекомбинантному аденовирусу по изобретению или фармацевтической композиции по изобретению для применения в профилактике инфекционного заболевания у млекопитающего.
Альтернативно изобретение относится к применению рекомбинантного аденовируса по изобретению или фармацевтической композиции по изобретению для производства лекарственного средства, предпочтительно вакцины, для профилактики инфекционного заболевания у млекопитающего.
Альтернативно изобретение относится к способу профилактики инфекционного заболевания у млекопитающего, включающему введение указанному млекопитающему рекомбинантного аденовируса по изобретению или фармацевтической композиции по изобретению.
В рамках изобретения, выражение «инфекционное заболевание» или «инфекция» относится к заболеванию, вызванному проникновением в организм хозяина инфекционного или патогенного агента, такого как вирусы, вироиды и прионы; микроорганизмы, такие как бактерии; паразиты, такие как нематоды (в том числе круглые черви и острицы), членистоногие, такие как клещи, клещики, блохи и вши; грибы и простейшие.
Рекомбинантные аденовирусы по изобретению пригодны для производства вакцин для профилактики любого вида инфекционного заболевания.
В предпочтительном варианте осуществления инфекционное заболевание вызвано патогенным агентом, выбранным из группы, состоящей из ВИЧ, вируса бешенства, вируса денге, вируса Эбола, коронавируса или возбудителя ТОРС, вируса папилломы человека, вируса гепатита С, вируса гепатита В, ротавируса, вируса кори, респираторно-синцитиального вируса, цитомегаловируса, вируса герпеса простого 2 типа, вируса Эпштейна-Барра, вируса гриппа, Trypanosoma cruzi и Plasmodium falciparum.
Термины «лечить» или «лечение» относятся к терапевтическому лечению, цель которого состоит в том, чтобы контролировать прогрессирование заболевания до или после того, как появились клинические симптомы. Контроль прогрессирования заболевания понимается как положительные или желаемые клинические результаты, которые включают, не ограничиваясь этим, ослабление симптомов, уменьшение продолжительности течения заболевания, стабилизацию патологических состояний (в частности, избежание дополнительных функциональных расстройств), задержку прогрессирования заболевания, улучшение патологического состояния и ремиссию (частичную и полную). Контроль прогрессирования заболевания также включает пролонгирование выживаемости по сравнению с ожидаемой выживаемостью, когда лечение не применялось.
Например, в случае лечения рака, ответ может контролироваться наблюдением за одним или более следующих эффектов: (1) ингибирование, до некоторой степени, роста опухоли, включая (i) замедление (ii), ингибирование ангиогенеза и (iii) полное торможение роста; (2) снижение количества опухолевых клеток; (3) поддержание на одном уровне размера опухоли; (4) уменьшение размера опухоли; (5) ингибирование, включая (i) снижение, (ii) замедление или (iii) полное предотвращение инфильтрации опухолевых клеток в периферические органы; (6) ингибирование, включая (i) снижение, (ii) замедление или (iii) полное предотвращение метастазирования; (7) усиление противоопухолевого иммунного ответа, что может привести к (i) поддержанию на одном уровне размера опухоли, (ii) уменьшению размера опухоли, (iii) замедлению роста опухоли, (iv) уменьшению, замедлению или предотвращению инвазии и/или (8) облегчение до некоторой степени тяжести или количества одного или нескольких симптомов, связанных с расстройством.
Термин «рак» относится к заболеванию, которое характеризуется бесконтрольным делением клеток (или увеличением выживаемости клеток или устойчивости к апоптозу), способностью указанных клеток проникать в другие соседние ткани (инвазия) или распространяться в другие области тела, где клетки обычно не находятся (метастазы), через лимфатические и кровеносные сосуды. В рамках изобретения, термин «рак» включает, не ограничиваясь этим, следующие виды рака: рак молочной железы; рак желчных путей; рак мочевого пузыря; рак головного мозга, включая глиобластому и медуллобластому; рак шейки матки; хориокарциному; рак толстой кишки; рак эндометрия; рак пищевода; рак желудка; гематологические новообразования, включая острый лимфоцитарный лейкоз и миелогенный лейкоз; Т-клеточный острый лимфобластный лейкоз/лимфому; волосатоклеточный лейкоз; хронический миелогенный лейкоз, множественную миелому; СПИД-ассоциированные лейкозы и Т-клеточный лейкоз/лимфому; интраэпителиальные неоплазии, включая болезнь Боуэна и болезнь Педжета; рак печени; рак легких; лимфомы, включая болезнь Ходжкина и лимфоцитарные лимфомы; нейробластомы; рак ротовой полости, включая плоскоклеточную карциному; рак яичников, включая виды рака, развивающиеся из эпителиальных клеток, стромальных клеток, зародышевых клеток и мезенхимальных клеток; рак поджелудочной железы; рак предстательной железы; рак прямой кишки; саркомы, включая лейомиосаркому, рабдомиосаркому, липосаркому, фибросаркому и остеосаркому; рак кожи, включая меланому, рак клеток Меркеля, саркому Капоши, базально-клеточную карциному и плоскоклеточный рак; рак яичка, включая герминогенные опухоли, такие как семинома, несеминомные опухоли (тератома, хориокарцинома), стромальные опухоли и герминтативноклеточные опухоли; рак щитовидной железы, включая аденокарциному и медуллярную карциному щитовидной железы; и рак почек, включая аденокарциному и опухоль Вильмса. В одном варианте осуществления рак представляет собой рак молочной железы. В другом варианте осуществления рак представляет меланому. Другие виды рака известны специалисту с обычной квалификацией в данной области техники.
Аденовирус по настоящему изобретению можно вводить млекопитающему. В рамках изобретения, термин «млекопитающее» относится к любому виду млекопитающих, в том числе, не ограничиваясь этим, домашних и сельскохозяйственных животных (коров, лошадей, свиней, овец, коз, собак, кошек или грызунов), приматов и человека. Предпочтительно млекопитающим является человек. В контексте настоящего изобретения, млекопитающее страдает раком или имеет риск страдать раком.
Для лечения опухолей на животных моделях или у пациентов аденовирусы могут быть доставлены местным или региональным введением посредством интратуморальной или внутриполостной инъекцией или системно посредством инъекции в кровоток. Вирус можно также вводить в сосудистую сеть опухоли. Поскольку рекомбинантные аденовирусы по изобретению защищены от нейтрализующих антител, находящихся в крови, то они особенно хорошо подходят для системного введения. Следовательно, в предпочтительном варианте осуществления изобретения аденовирус по изобретению вводится системно. В рамках изобретения, выражение «системное введение» относится к пути введения в кровеносную систему, таким образом, что воздействие приходится на весь организм. Предпочтительно введение является парентеральным (как правило, внутриартериальной или эндовенозной инъекцией, инфузией или имплантацией).
Аденовирус по изобретению идеально вводить до того, как он свяжется с сывороточным альбумином, и он свяжется с альбумином в крови подвергаемого лечению пациента. Однако для повышения персистентности аденовирусов в крови с целью повышения возможного достижения диссеминированных опухолевых узелков, капсид может быть покрыт альбумином до введения аденовируса.
Лечение опухолей аденовирусами по изобретению можно использовать в комбинации с другими терапевтическими методами, такими как химиотерапия или лучевая терапия, как было описано ранее в области онколитической аденовируса.
Протоколы применения аденовирусов, описанных в настоящем изобретении для лечения рака, представляют такие же процедуры, которые используются в областях виротерапии и генной терапии с использованием аденовирусов.
Настоящее изобретение также относится к следующему:
[1]. Аденовирусный геном, отличающийся тем, что он содержит последовательность, кодирующую альбумин-связывающий участок, вставленную в кодирующую область гипервариабельного участка 1 (HVR1) гексонного белка, что приводит к экспрессии слитого белка, содержащего гексонный белок и альбумин-связывающий участок, и где альбумин-связывающий участок располагается на наружной поверхности гексонного белка, когда сборка гексонного белка имеет место в аденовирусном капсиде.
[2]. Аденовирусный геном по п.[1], где геном происходит из аденовируса человека.
[3]. Аденовирусный геном по п.[2], где аденовирус человека выбран из группы, состоящей из аденовируса человека серотипов 1-57.
[4]. Аденовирусный геном по п.[3], где аденовирус человека представляет серотип 5.
[5]. Аденовирусный геном по п.п. [1], [2], [3] или [4], где альбумин-связывающий участок выбран из альбумин-связывающего домена из стрептококкового белка G, альбумин-связывающего домена из белка PAB Peptostreptococcus magnus, альбумин-связывающего пептида, имеющего коровую последовательность DICLPRWGCLW (SEQ ID NO:9), и их функционально эквивалентных вариантов.
[6]. Аденовирусный геном по п.[5], где альбумин-связывающий участок представляет альбумин-связывающий домен 3 из стрептококкового белка G.
[7]. Аденовирусный геном по п.[6], где последовательность альбумин-связывающего домена 3 из стрептококкового белка G представляет SEQ ID NO:1.
[8]. Аденовирусный геном по п.п. [1], [2], [3], [4], [5], [6] или [7], где HVR гексонного белка выбран из группы, состоящей из HVR1, HVR2, HVR3, HVR4, HVR5, HVR6 и HVR7.
[9]. Аденовирусный геном по п.[8], где HVR гексонного белка представляет HVR1.
[10]. Аденовирусный геном по п.[9], где последовательность, кодирующая альбумин-связывающий участок, вставлена таким образом, что полученный слитый белок содержит альбумин-связывающий участок после аминокислоты D150 гексонного белка согласно нумерации гексонного белка, имеющего инвентарный номер в GenBank BAG48782.1.
[11]. Аденовирусный геном по п.[8], где HVR гексонного белка представляет HVR5.
[12]. Аденовирусный геном по п.[11], где последовательность, кодирующая альбумин-связывающий участок, вставлена в HVR5 таким образом, что полученный в результате слитый белок содержит альбумин-связывающий участок после аминокислоты A274 гексонного белка согласно нумерации гексонного белка, имеющего инвентарный номер BAG48782.1 в GenBank.
[13]. Аденовирусный геном по одному из пунктов [1]-[12], где N- и/или С-конец альбумин-связывающего участка связан с гексонным белком линкерной последовательностью.
[14]. Аденовирусный геном по п.[13], где указанная линкерная последовательность содержит последовательность GSGS (SEQ ID NO:2).
[15]. Аденовирусный геном по одному из пунктов [1]-[14], где аденовирусный геном дополнительно содержит тканеспецифический промотор или опухолеспецифичный промотор.
[16]. Аденовирусный геном по п.[15], где тканеспецифический промотор или опухолеспецифический промотор представляют промоторные последовательности для регуляции экспрессии одного или более генов, выбранных из группы, состоящей из E1a, E1b, E2 и E4.
[17]. Аденовирусный геном по п.[16], где промотор выбран из группы, состоящей из промотора E2F, промотора теломеразы hTERT, промотора тирозиназы, промотора простат-специфического антигена, промотора альфа-фетопротеина и промотора СОХ-2.
[18]. Аденовирусный геном по одному из пунктов [1]-[17], где аденовирус является онколитическим аденовирусом.
[19]. Аденовирусный геном по п.[18], где указанный аденовирусный геном дополнительно содержит мутации в одном или более генов, выбранных из группы, состоящей из Е1а, Е1b, Е4 и VA-РНК, для достижения селективной репликации в опухолях.
[20]. Аденовирусный геном по одному из пунктов [1]-[19], где аденовирусный геном дополнительно содержит модификации капсида для повышения инфекционной активности аденовируса или его нацеленности на рецептор, присутствующий в опухолевой клетке.
[21]. Аденовирусный геном по п.[20], где модификация капсида представляет инсерцию мотива RGD в петлю H1 аденовирусного белка волокна.
[22]. Аденовирусный геном по одному из пунктов [1]-[21], где аденовирусный геном содержит один или более дополнительных генов, вставленных в геном аденовируса.
[23]. Аденовирусный геном по п.[22], где дополнительные гены представляют один или более неаденовирусных генов, используемых в генной терапии или в вакцинации.
[24]. Аденовирусный геном по п.[23], где указанные гены используются в генной терапии рака.
[25]. Аденовирусный геном по п.[24], где указанные гены, используемые в генной терапии рака, представляют собой по меньшей мере ген, выбранный из группы, состоящей из пролекарство-активирующих генов, генов-супрессоров опухолей, генов, кодирующих противоопухолевые интерферирующие РНК, и иммуностимулирующих генов.
[26]. Рекомбинантный аденовирус, имеющий аденовирусный геном по одному из пунктов [1]-[25].
[27]. Фармацевтическая композиция, содержащая терапевтически эффективное количество рекомбинантного аденовируса по п.[26] вместе с фармацевтически приемлемым носителем.
[28]. Рекомбинантный аденовирус по п.[26] или фармацевтическая композиция по п.[27] для применения в медицине.
[29]. Рекомбинантный аденовирус по п.[26] или фармацевтическая композиция по п.[27] для применения в профилактике и/или лечении рака у млекопитающего, где аденовирус является онколитическим аденовирусом или аденовирусом, имеющим аденовирусный геном по п.п. [24]-[25].
[30]. Рекомбинантный аденовирус для применения по п.п. [28] или [29], где млекопитающее является человеком.
[31]. Рекомбинантный аденовирус для применения по п.п. [28], [29] или [30], где аденовирус вводится системно.
Следующие примеры приведены только как иллюстративные и не должны быть истолкованы как ограничивающие объем настоящего изобретения.
Примеры
Материалы и методы
Клеточные линии. Клетки НЕК293 (почки эмбриона человека), А549 (аденокарцинома легкого человека), Sk-mel28 (меланома) и MCF7 (аденокарциномы молочной железы человека) были получены из Американской коллекции типовых культур (АТСС, Manassas, VA). Все линии опухолевых клеток, за исключением MCF7, поддерживали на модифицированной по способу Дульбекко среде Игла, с добавлением 5% фетальной бычьей сыворотки при температуре 37°С, в атмосфере 5% СО2. Клетки MCF7 поддерживали на среде RPMI 1640 с добавлением 10% фетальной бычьей сыворотки. Все клеточные линии регулярно тестировались на наличие микоплазм.
Конструирование вирусов. Онколитический аденовирус ICOVIR15 был описан ранее (Rojas J.J. et al. Minimal RB-responsive E1A promoter modification to attain potency, selectivity and transgen-arming capacity oncolytic adenoviruses. Mol. Ther., 2010; 18 (11):1960-71). AdGL представляет вектор первого поколения с делецированной областью E1, экспрессирующий кассету слитого белка EGFP-люцифераза из pEGFPLuc (Clontech). Проводили инсерцию кассеты промотор CMV-EGFPLuc-полиА, заменяющей область E1, следуя протоколу рекомбинации, представляющему адаптированный протокол Stanton et al. (Stanton R.J. et al. Reengineering adenovirus vector systems to enable high-throughput analyses of gene function. Biotechniques, 2008; 45(6):659-62, 664-8), основанному на гомологичной рекомбинации в бактериях, используя позитивную-негативную селекцию. Кассету CMV-GFPLuc, фланкированную гомологичными областями E1, использовали для замены маркеров позитивной-негативной селекции pAd5-CV5-Е3+, обычно используемых для конструирования аденовирусных векторов с делецией Е1. ICOVIR15 размножали в клетках А549 и дефицитный по репликации AdGL размножали в клетках HEK293.
ICOVIR15-ABD и AdGL-ABD конструировали инсерцией альбумин-связывающего домена (ABD), фланкированного двумя линкерами (GSGS) (SEQ ID NO: 2), в гипервариабельный участок 1 (HVR1) аденовирусного гексона после D150 аминокислоты. Нуклеотидная последовательность полного модифицированного гексона, содержащего ABD, инсерцированного в HVR1 (ABD-HVR1), представляет SEQ ID NO:3. Все модификации выполняли, следуя протоколу рекомбинации, представляющему адаптированный протокол Stanton et al. (Stanton R.J. et al. Reengineering adenovirus vector systems to enable high-throughput analyses of gene function. Biotechniques, 2008; 45(6):659-62, 664-8), основанному на гомологичной рекомбинации в бактериях, используя позитивную-негативную селекцию с кассетой RpsL-Neo.
Во-первых, кассету rpsLNeo амплифицировали с помощью ПЦР из pJetRpsLNeo, плазмиды, содержащей маркеры позитивной-негативной селекции rpsLneo, клонированные в pJet1.2/blunt (GenScript, Wheelock House, Гонконг), с использованием олигонуклеотидов HVR1rpsLF 5'- GCCCTGGCTCCCAAGGGTGCCCCAAATCCTTGCGAATGGGGCCTGGTGATGATGGC-3' (SEQ ID NO: 12) и HVR1rpsLR 5'- GTAATATTTATACCAGAATAAGGCGCCTGCCCAAATACGTGAGTTCAGAAGAACTCGTCAAGAAG-3'(SEQ ID NO: 13). Кассету вставляли в HVR1 плазмид pAdZICOVIR15 и pAdZGL с получением pAdZICOVIR15-H1-rpsLNeo и pAdZGL-H1-rpsLNeo. Во-вторых, получали фрагмент линкер-ABD-линкер с помощью ПЦР с использованием следующих перекрывающихся олигонуклеотидов:
CCG-3'
GCG-3'
TC-3'
Данный фрагмент использовали для замены кассеты rpsLNeo в обеих плазмидах с получением pAdZICOVIR15-H1-ABD и pAdZGL-H1-ABD.
Также ABD вставляли в гипервариабельный участок 5 AdGL после аминокислоты A274. Нуклеотидная последовательность полного модифицированного гексона, содержащего ABD, вставленный в HVR5 (ABD-HVR5), представляет SEQ ID NO: 4. Для этой цели кассету rpsLNeo амплифицировали с помощью ПЦР из pJetRpsLNeo с использованием следующих олигонуклеотидов H5rpsLF и H5rpsLR
gaactcgtcaagaag-3'
и вставляли в плазмиду pAdZGL с получением pAdZGL-H5-rpsLNeo. На второй стадии рекомбинации фрагмент линкер-ABD-линкер амплифицировали с помощью ПЦР из pAdZICOVIR15-H1-ABD с использованием следующих олигонуклеотидов ABDH5F и ABDH5R:
tgtttctgattattacaagaatttgattaacaatgccaaaaccgtcgagg-
gcgtaaaggctctgatcgacgaaatacttgcggccctaccc-3'
tgtttctgattattacaagaatttgattaacaatgccaaaaccgtcgaggg-cgtaaaggctctgatcgacgaaatacttgcggccctaccc-3'
Данный фрагмент использовали для замены rpsLNeo из pAdZGL-H5-rpsLNeo с получением pAdZGL-H5-ABD.
Кроме того, домен также можно инсерцировать в любую из гипервариабельных участков гексонного белка. Для этого rpsLNeo должен быть вставлен в требуемую область и затем заменен фрагментом ABD, полученным с помощью ПЦР, с плечами гомологии для рекомбинации в конкретную гипервариабельную петлю.
ICOVIR15-ABD (ABD в HVR1), AdGL-ABD (ABD в HVR1) и AdGL-Н5-ABD (ABD в HVR5) получали трансфекцией полученных плазмид с использованием стандартного кальций-фосфатного протокола в клетках HEK293. Онколитической аденовирус ICOVIR15-ABD очищали методом бляшек и затем амплифицировали в клетках А549. Аденовирусный вектор AdGL-ABD очищали методом бляшек и затем амплифицировали в клетках HEK293. Оба вируса очищали с использованием двойного градиента с хлоридом цезия, следуя стандартным методикам.
Инкубацию вирусов с сывороточным альбумином человека (HSA) проводили в течение одного часа при комнатной температуре со средой, содержащей 1 мг/мл HSA.
Анализ продукции вируса. Клетки A549 инфицировали 800 вирусными частицами (vp) каждого вируса на клетку с получением 80-100% заражения. Клетки трижды промывали PBS через 4 ч после заражения и инкубировали со свежей средой. В указанные временные точки клетки собирали и три раза замораживали-оттаивали с получением клеточного экстракта. Вирусные титры определяли методом окрашивания, основанным на использовании антитела к гексону (Cascallo М., et al. Systemic toxicity-efficacy profile of ICOVIR-5, a potent and selective oncolytic adenovirus based on the pRB pathway. Mol. Ther., 2007; 15:1607-15) в клетках НЕК293.
Анализ цитотоксичности in vitro. Анализ цитотоксичности проводили посевом 10000 клеток HEK293, 30000 клеток А549, 10000 клеток Sk-mel28 и 20000 клеток MCF-7 на лунку в 96-луночных планшетах. Перед инфицированием вирусы инкубировали в течение одного часа при комнатной температуре со средой или средой, содержащей 1 мг/мл сывороточного альбумина человека (HSA). Клетки заражали в нормальной среде или в альбумин-содержащей среде, серийными разведениями (1/3 для клеток HEK293, Sk-mel28 и MCF-7 и 1/5 для клеток А549), начиная с 10000 вирусных частиц/ клетку. На 7 сутки после заражения планшеты промывали PBS и окрашивали на общее содержание белка (проба с бицинхониновой кислотой; Pierce Biotechnology, Rockford, IL) и определяли оптическую плотность. Количество вирусных частиц на клетку, необходимое для торможения роста на 50% (IC50), определяли по кривым доза-реакция стандартной нелинейной регрессией (GraFit; Erithacus Software, Horley, Великобритания) с использованием адаптированного уравнения Хилла.
Детектирование связывания с человеческим и мышиным сывороточным альбумином. Детектирование связывания с человеческим сывороточным альбумином (HSA) и мышиным сывороточным альбумином (MSA) проводили, следуя протоколу ELISA, предсавляющему адаптированный протокол Konig и Skerra (Konig T., Skerra A. Use of an albumin-binding domain for the selective immobilization of recombinant capture antibody fragments on ELISA plates. J. Immunol. Methods, 1998; 218:73-83). Инкубацию проводили в течение 1 ч при комнатной температуре с последующими тремя стадиями отмывки 200 мкл PBS, содержащим 0,1% твина 20, который также использовали в качестве буфера для разведения вирусов и антител.
96-луночный планшет покрывали 200 мкл HSA или BSA (Sigma) в концентрации 2 мг/мл, разведенных в PBS. Оставшиеся сайты связывания на пластиковой поверхности блокировали 2 мг/мл бычьего сывороточного альбумина, разведенного PBS, содержащего 0,5% твина 20. На следующей стадии добавляли очищенные вирусы в объеме 50 мкл. Для определения связывания были испытаны три разных количества вирусного белка (25, 2,5 и 0,25 нг вирусного белка) (фиг. 5A). Концентрацию вирусного белка в очищенных образцах вируса определяли количественно с помощью анализа белка Bio-Rad. Детектирование вирусов проводили с использованием антитела против гексона из супернатанта гибридомы 2Hx-2 (ATCC® HB-8117™) (50 мкл на лунку при разведении 1/5) и поликлонального козьего антимышиного антитела, конъюгированного с пероксидазой хрена (50 мкл на лунку при разведении 1/2000). Лунки окрашивали добавлением 100 мкл на лунку свежеприготовленного смешанного раствора субстрата пероксидазы - 3,3',5,5'-тетраметилбензидина и инкубировали в течение 15 мин при встряхивании. Реакцию останавливали добавлением 100 мкл 2 Н серной кислоты и измеряли оптическую плотность при длине волны 450 нм.
Тот же самый эксперимент проводили с покрытием 96-луночного планшета 200 мкл BSA, HSA или MSA (Sigma) в концентрации 2 мг/мл, разведенных в PBS, и тестировали 25 нг вирусного белка для определения связывания. В опыт включали контрольную группу без вируса (фиг. 5В).
Анализ антитело-опосредуемой нейтрализации. Антитело-опосредуемую нейтрализацию анализировали на уровне инфекции (трансдукция аденовирусами репортером люцифераза-GFP) и репликации (цитотоксичность, опосредованная аденовирусами, компетентными по репликации). Промышленно доступное антитело Ab6982 (анти-Ad5, кроличье поликлональное антитело, Abcam) использовали в качестве нейтрализующего антитела. Для анализа инфекционной активности, начиная с разведения 1/100 стокового раствора антитела, готовили серийные разведения 1/6 антитела в среде, содержащей различные аденовирусы (AdGL или AdGL-ABD) в 96-луночных планшетах.
После инкубации в течение 1 ч при комнатной температуре добавляли клетки 1E5 HEK293 или 3E4 Sk-mel28 на лунку с получением требуемой множественности инфекции (0,5 TU/клетку или 10 вирусных частиц/клетку для клеток НЕК293 и 40 вирусных частиц/ клетку для клеток Sk-mel28). Через двадцать четыре часа после инфицирования среду удаляли и клетки лизировали добавлением 50 мкл реагента для лизиса клеток (Promega, Madison, Wi) и один раз замораживали-оттаивали. Лизаты центрифугировали при 13000 g в течение 5 мин при 4°С и измеряли активность фермента люциферазы в супернатанте с помощью реагента для анализа люциферазы (Promega) с использованием люминометра (Berthold Junior, Berthold GmbH&Co KG, Германия).
Для анализа репликации готовили серийные разведения 1/2 антитела, начиная с разведения 1/100, стокового раствора антитела. Такое последовательное разведение проводили с использованием среды, содержащей ICOVIR15 или ICOVIR15-ABD. Через 1 ч после инкубации с антителом при комнатной температуре, добавляли клетки 3E5 А549 на лунку для получения множественности инфекции 600 вирусных частиц/клетку. На 4 сутки после инфицирования анализировали жизнеспособность по окрашиванию на общее содержание белка, как описано выше в подразделе «Анализ цитотоксичности in vitro» в разделе «Материалы и методы».
Клиренс из крови in vivo. Исследования in vivo проводили в лаборатории ICO-IDIBELL (Барселона, Испания) AAALAC, блоке 1155, и их проведение было одобрено Этическим комитетом по проведению экспериментов на животных IDIBELL. Мышам-самкам BALB/C nu/nu вводили внутривенно смесь ICOVIR15 и ICOVIR15-ABD (соотношение 1:1) в общей дозе 5×1010 вирусных частиц в объеме 10 мл/кг в PBS (n=5). Через 5 мин, 15 мин, 1 ч, 4 ч и 24 ч после введения, отбирали образцы крови из хвостовой вены. Образцы крови центрифугировали при 5000 g в течение 5 мин при 4°С для отделения клеточной фракции и сбора сыворотки крови. Образцы сыворотки обрабатывали протеиназой К и SDS в течение 45 мин при 54°C и затем в течение 10 мин при 90°С для протеолиза капсида и высвобождения вирусной ДНК. Переваренные образцы амплифицировали с помощью ПЦР с использованием гексон HVR1-фланкирующих олигонуклеотидов Ad19121F 5'-CTGGACATGGCTTCCACGTA-3' (SEQ ID NO: 25) и Ad19300R 5'-GCTCGTCTACTTCGTCTTCG-3' (SEQ ID NO: 26), и анализировали электрофорезом на 1% агарозном геле. Поскольку ICOVIR15-ABD имеет инсерцию ABD в середине HVR1, то размер ПЦР-продукта будет больше: ожидается, что ПЦР-продукт из ICOVIR15-ABD составляет 361 п.н. по сравнению с 199 п.н. из ICOVIR15.
Противоопухолевая эффективность in vivo. Опухолевые ксенотрансплантаты подкожной меланомы получали инъекцией 1×107 клеток Sk-mel28 в бок 6-недельных мышей-самок Balb/C nu/nu. Когда опухоли достигали размера 150 мм3 (день эксперимента 0), мышей рандомизировали (n=10-12 животных в группе) и однократно внутривенно вводили PBS или 5×1010 вирусных частиц ICOVIR15 или ICOVIR15-ABD в объеме 10 мл/кг в PBS в хвостовую вену. Размер опухолей и состояние мышей оценивали трижды в неделю. Объем опухолей измеряли с помощью цифрового штангенциркуля и определяли с использованием уравнения V (мм3)=π/6×W2×L, где W и L представляют ширину и длину опухоли, соответственно. Статистическую значимость различий в размере опухолей между группами лечения оценивали с помощью двухсторонего t-теста Стьюдента.
Трансдукция in vivo. Опухоли меланомы воспроизводили имплантацией 1×106 клеток B16-CAR подкожно в оба бока 6-недельных мышей-самок C57BL6 (n=4-6 животных в каждой группе). Когда опухоли достигали размера 100 мм3, мышам вводили внутрибрюшинно PBS (группа интактных мышей) или 2×1010 вирусных частиц hAd5wt (предварительно иммунизированная группа). Через семь суток после иммунизации животным однократно внутривенно вводили PBS, AdGL или AdGL-ABD в дозе 3×1010 вирусных частиц на мышь. Через 3 суток после того, как мышам вводили вектор, они получали внутрибрюшинно инъекцию 250 мкл D-люциферина (15 мг/мл; Biosynth, Staad, Швейцария) и мышей подвергали эвтаназии для отбора образцов печени и опухоли для биолюминесцентной томографии (IVIS). Органы исследовали на приборе IVIS Lumina XR (Caliper Life Sciences, Hopkinton, MA), и программное обеспечение Living Image v4.0 использовали для количественной оценки эмиссии света.
Результаты
Получение и характеристика ICOVIR15-ABD. Для получения альбумин-связывающего аденовируса, альбумин-связывающий домен 3 из белка G стрептококковых бактерий (SEQ ID NO: 1) инсерцировали в HVR1 гексона ICOVIR15, с получением онколитического аденовируса ICOVIR15-ABD (фиг. 1 и 2). Домен был вставлен, фланкированный двумя линкерами (GSGS) (SEQ ID NO: 2), в середину HVR1 после аминокислоты D150 без делеции какой-либо последовательности гексона (фиг. 1).
Для анализа влияния инсерции ABD на репликацию вируса, заявители сравнивали кинетику репликации ICOVIR15 и ICOVIR15-ABD в клетках А549. Как показано на фиг. 3, несмотря на то, что кинетика продукции обоих вирусов была сходной, наблюдали снижение общего выхода продуцированного ICOVIR15-ABD. Также анализировали способность вируса вызывать гибель раковых клеток in vitro. Эксперименты по оценке цитотоксичности проводили в клетках HEK293, А549, Sk-mel28 и MCF-7, в присутствии или отсутствии HSA. Значения IC50 не указывали на наличие существенных различий между вирусами в трех из четырех испытанных клеточных линий, и трехкратное повышение IC50 для ICOVIR15-ABD по сравнению с ICOVIR15 в клетках MCF-7 указывает на некоторое снижение цитотоксичности в этой клеточной линии (фиг. 4). Во всех случаях добавление HSA не влияло на цитотоксичность.
Эксперимент с ELISA проводили для демонстрации связывания ICOVIR15-ABD c HSA и MSA. Лунки покрывали MSA, HSA или BSA (следует отметить, что ABD связывается с MSA и HSA, но не с BSA) и анализировали связывание обоих вирусов ICOVIR15 или ICOVIR15-ABD. Положительный сигнал получали при добавлении ICOVIR15-ABD как в покрытые HSA лунки (фиг. 5А), так и покрытые MSA лунки (фиг. 5В), и интенсивность сигнала возрастала с увеличением количества используемого вируса (фиг. 5А). Когда использовали BSA вместо HSA или MSA, то никакого сигнала не наблюдали, независимо от количества добавленного вируса, указывая на то, что вирус может связываться с человеческим и мышиным альбумином, но не с бычьим альбумином. Как и следовало ожидать, ни в одном случае не наблюдали связывания с вирусом ICOVIR15.
Связывание с альбумином защищает аденовирус от нейтрализующих антител in vitro. Продемонстрировав связывание ICOVIR15-ABD с человеческим альбумином, заявители тестировали, насколько такое связывание может защитить вирус от нейтрализующих антител (NABS) in vitro. Для этого конструировали аденовирусный вектор, экспрессирующий слитый белок GFP-люцифераза, модифицированный ABD в гексоне, названный AdGL-ABD. Исследовали эффективность трансдукции AdGL-ABD в клетках НЕК293 после инкубации с серийными разведениями промышленно доступного нейтрализующего антитела Ab6982 (кроличье поликлональное антитело к Ad5) в присутствии и отсутствие HSA. Как показано на фиг. 6, были достигнуты одинаковые уровни трансдукции с немодифицированным вектором AdGL независимо от инкубации с альбумином. В отличие от этого HSA защищал AdGL-ABD от нейтрализации. Интересно отметить, что AdGL-ABD нейтрализовался в меньшей степени, чем немодифицированный вектор AdGL, даже если его не инкубировали с альбумином, указывая, что простая модификация HVR1 с ABD уже препятствует связыванию некоторых NAbs.
Кроме того, также анализировали способность вирусов приводить к гибели раковых клеток в присутствии нейтрализующих антител. Для этой цели клетки A549 инфицировали ICOVIR15 или ICOVIR15-ABD, предварительно инкубированные с серийными разведениями нейтрализующего антитела Ab6982, в присутствии и отсутствии HSA, и выживаемость клеток анализировали через 4 суток после заражения. В отсутствии человеческого альбумина оба вируса показывали одинаковую способность вызывать гибель опухолевых клеток (фиг. 7), и лишь небольшое повышение цитотоксичности наблюдали с ICOVIR15-ABD, вероятно, за счет некоторого уклонения от действия нейтрализующих антител, наблюдаемое в трансдукции (фиг. 6). Важно отметить, что, когда человеческий альбумин добавляли в среду, то цитотоксичность ICOVIR15-ABD была значительно выше по сравнению с ICOVIR15, для которого она оставалась без изменений.
ICOVIR15-ABD показывает повышенный период полураспада в плазме крови. Для исследования того, что связывание с альбумином может уменьшить быстрый клиренс аденовируса из крови, исследовали фармакокинетику ICOVIR15-ABD после системного введения in vivo. Мышам вводили смесь ICOVIR15 и ICOVIR15-ABD в соотношении 1:1 в суммарной дозе 5×1010 вирусных частиц на мышь, и отбирали образцы крови на различные временные точки. Амплификацию HVR1 гексона проводили с помощью ПЦР в образцах сыворотки. За счет инсерции ABD получали полосу 361 п.н. с ICOVIR15-ABD, в то время как размер полосы для ICOVIR15 составлял всего 199 п.н. Следовательно, сравнивая относительную интенсивность полос на каждый временной точке, можно было определить, какой вирус сохраняется дольше в кровотоке. На фиг. 8 показаны результаты электрофореза ПЦР-продуктов всех образцов, включая стандарт с несколькими соотношениями ICOVIR15-ABD: ICOVIR15 (0,2, 1, 5, 10 и 50), контроль до введения и воду, используемую в качестве отрицательного контроля. Полосы с одинаковой интенсивностью получали в контроле до введения и через 5 мин после инъекции. Затем отмечали изменение интенсивности полос, поскольку полоса, соответствующая ICOVIR15-ABD, становилась более интенсивной, чем полоса ICOVIR15. Через 1 ч после инъекции четко видно различное сохранение обоих вирусов в пользу ICOVIR15-ABD. Эти данные указывают на то, что через 5 мин после инъекции ICOVIR15 подвергается клиренсу из крови гораздо быстрее, чем ICOVIR15-ABD, демонстрируя лучшую фармакокинетику вируса, модифицированного ABD.
Противоопухолевая активность ICOVIR15-ABD после системного введения in vivo. После того, как был показан повышенный период полураспада ICOVIR15-ABD в плазме крови, вирус был испытан в отношении того, насколько это переносится на повышение противоопухолевой эффективности после системного введения. Мышам, несущим опухолевые ксенотрансплантаты опухоли Sk-mel28 (меланома), вводили однократно внутривенно дозу забуференного фосфатом физиологического раствора (PBS), ICOVIR15 или ICOVIR15-ABD из расчета 5×1010 вирусных частиц на мышь. На сутки 38 после обработки животных вынужденно подвергали эвтаназии за счет большого размера PBS-обработанных опухолей. Оба вируса были способны значительно снижать рост опухолей по сравнению с PBS (фиг. 9). Однако лечение ICOVIR15-ABD показало статистическое значимое снижение роста опухолей, начиная с суток 21 до окончания лечения, в то время как ICOVIR15 не мог статистически значимо контролировать рост опухоли до суток 35. На сутки 38, когда животных подвергали эвтаназии, ICOVIR15 вызывал снижение роста опухолей в 1,4 раза по сравнению с 2-кратным снижением с ICOVIR15-ABD.
Связывание с альбумином защищает аденовирус от предварительной иммунизации анти-HAd5 in vivo. Иммунокомпетентных мышей C57BL6, несущих опухоли меланомы B16-CAR, иммунизировали внутрибрюшинно введением 2×1010 вирусных частиц hAd5wt или PBS (предварительно иммунизированные или нативные группы). Через семь суток мышам вводили однократно внутривенно дозу PBS, AdGL или AdGL-ABD из расчета 3×1010 вирусных частиц на мышь. Через 3 суток после введения вектора мышей подвергали эвтаназии и отбирали образцы печени и опухолей для биолюминесцентной томографии in vivo (IVIS). Не наблюдали существенных различий между векторами в трансдукции печени и опухолей у нативных животных (фиг. 10). Следует отметить, что, когда животные были предварительно иммунизированы немодифицированным вектором AdGL, то отмечали полную нейтрализацию, поскольку трансдукция печени и опухолей полностью отсутствовала. В противоположность, AdGL-ABD был способен поддерживать тот же уровень трансдукции в печени и опухолях, что указывает на защиту от предварительной иммунизации анти-HAd5.
Инсерция ABD в гипервариабельный участок 5. Для проверки того, можно ли провести такую инсерцию в другие гипервариабельные участки гексона, заявители конструировали вектор AdGL-H5-ABD. Клетки НЕК293 трансфекцировали плазмидой pAdZGL-Н5-ABD для получения вируса AdGL-Н5-ABD. Через одну неделю после трансфекции собирали клетки и супернатант и подвергали лизису с помощью трех циклов замораживания-оттаивания. Клеточный экстракт, содержащий вирус, титровали в клетках НЕК293, с помощью метода бляшек. Лунки, соответствующие разведениям 1E6, 1E7 и 1E8, показаны на фиг. 11, где четко видны бляшки с размножением вируса. Инсерцию ABD в HVR5 подтверждали секвенированием генома вируса. Это показывает возможность инсерции ABD в другие гипервариабельные участки, не оказывая влияния на жизнеспособность вируса.
Инсерция ABD в гипервариабельный участок 5 не защищает аденовирус от нейтрализующих антител. Для проверки того, насколько инсерция ABD в гипервариабельный участок 5, также может защитить аденовирус от нейтрализующих антител, заявители сравнивали эффективность трансдукции векторов AdGL, AdGL-H1-ABD и AdGL-H5-ABD после инкубации с серийными разведениями Ab6982 NAb с или без HSA, в клетках HEK293 и Sk-mel28. Как показано на фиг. 6, инкубация с HSA обеспечивала явное преимущество трансдукции с AdGL-H1-ABD в обеих клеточных линиях, в то время как она не оказывала существенного влияния на немодифицированный вектор AdGL (фиг. 12). Удивительно, но добавление человеческого альбумина не увеличивало уровни трансдукции AdGL-H5-ABD. Это указывает на то, что ABD является функциональным при инсерции в HVR1, но не в HVR5.








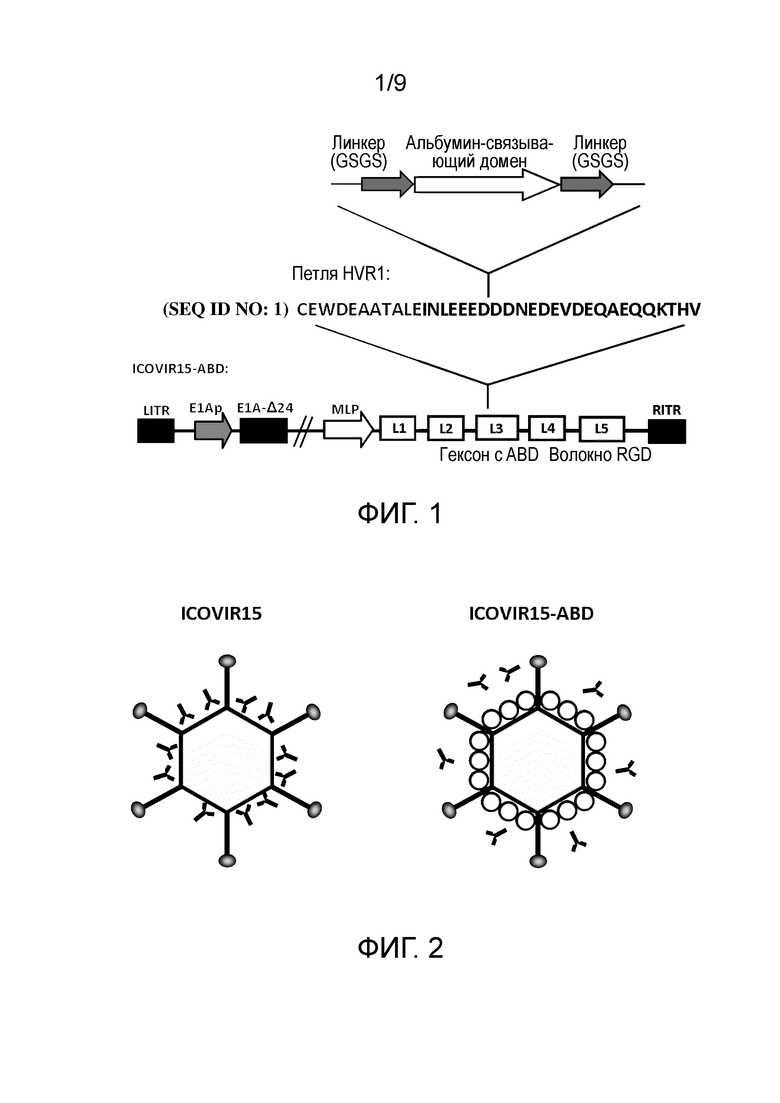
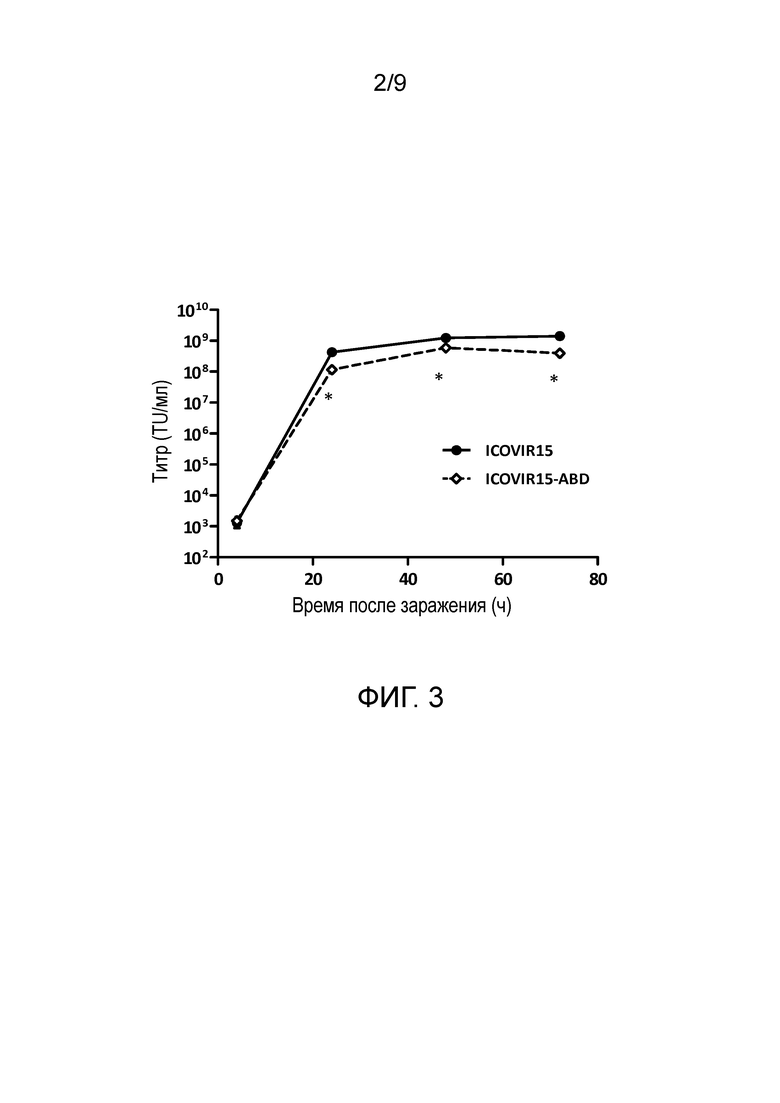
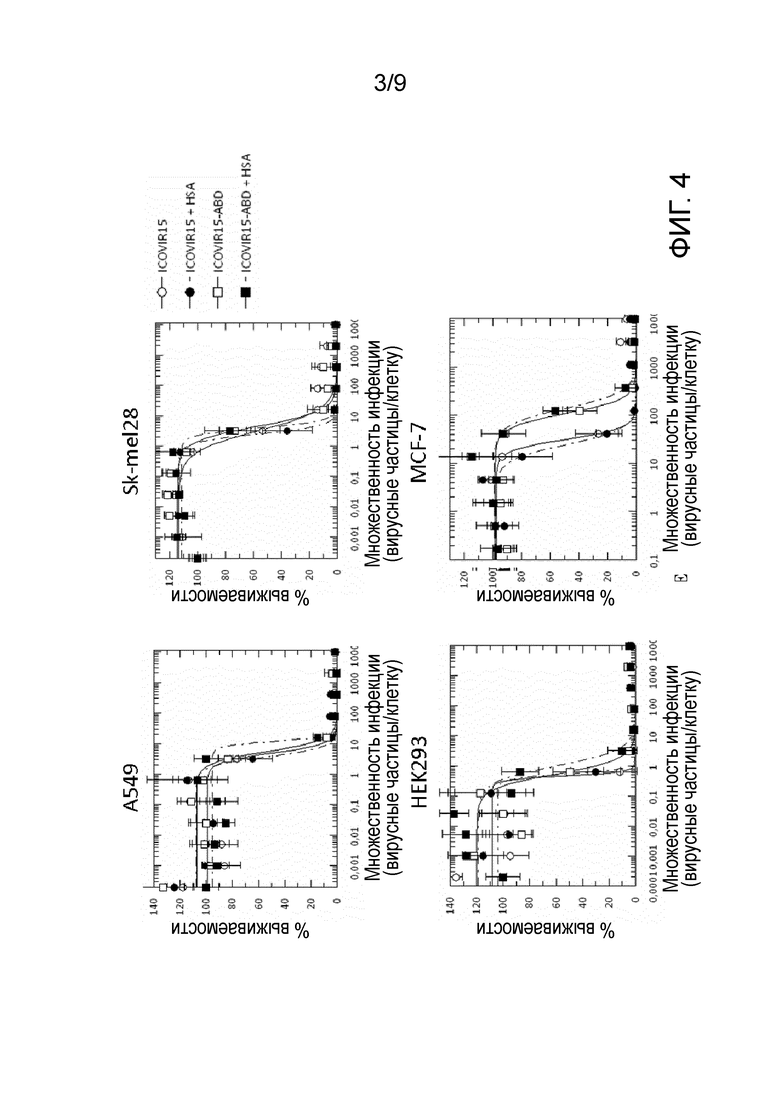
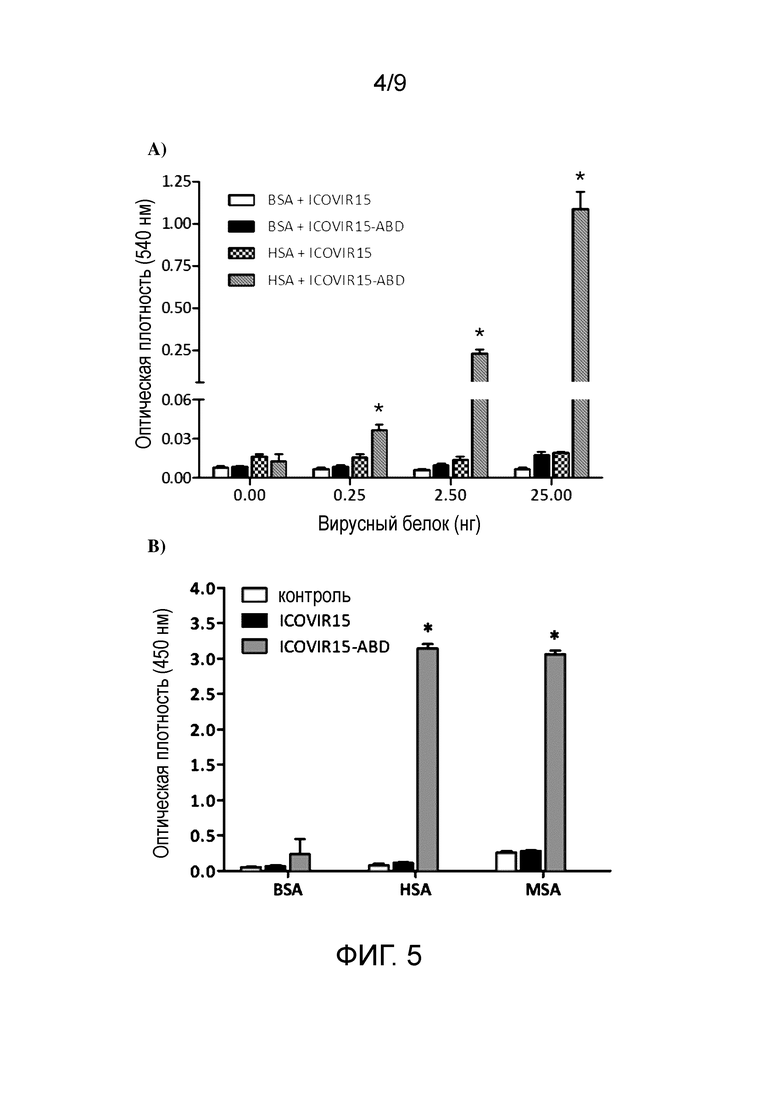
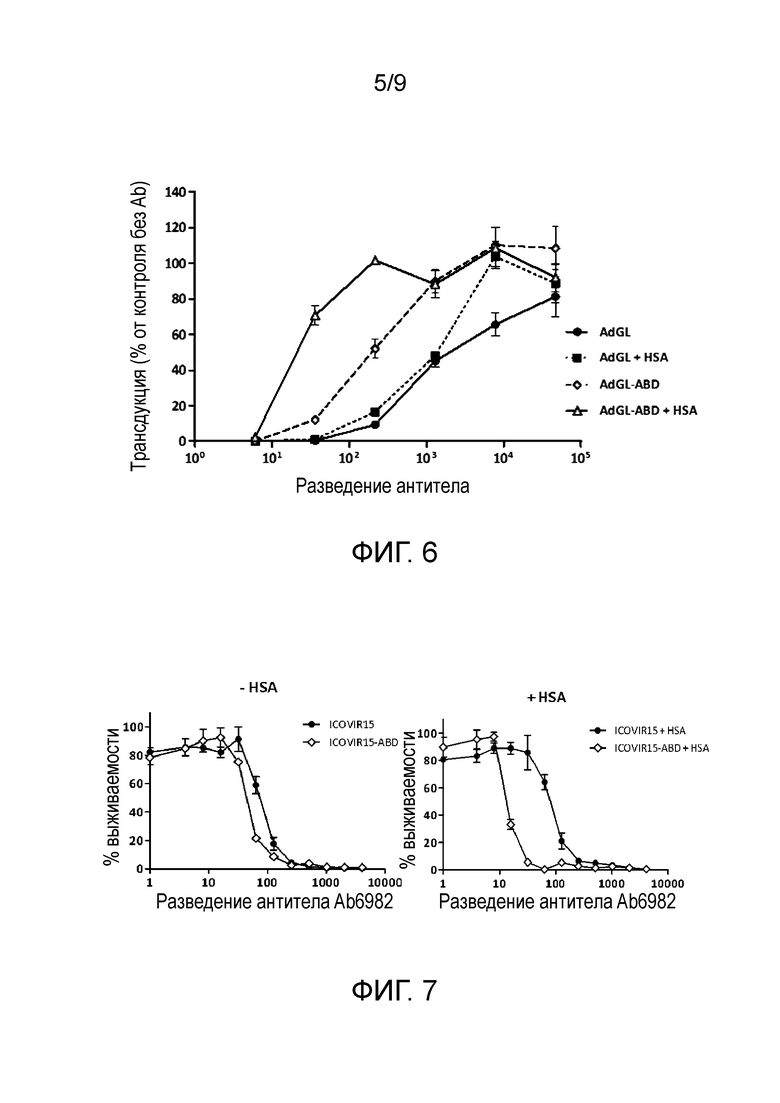
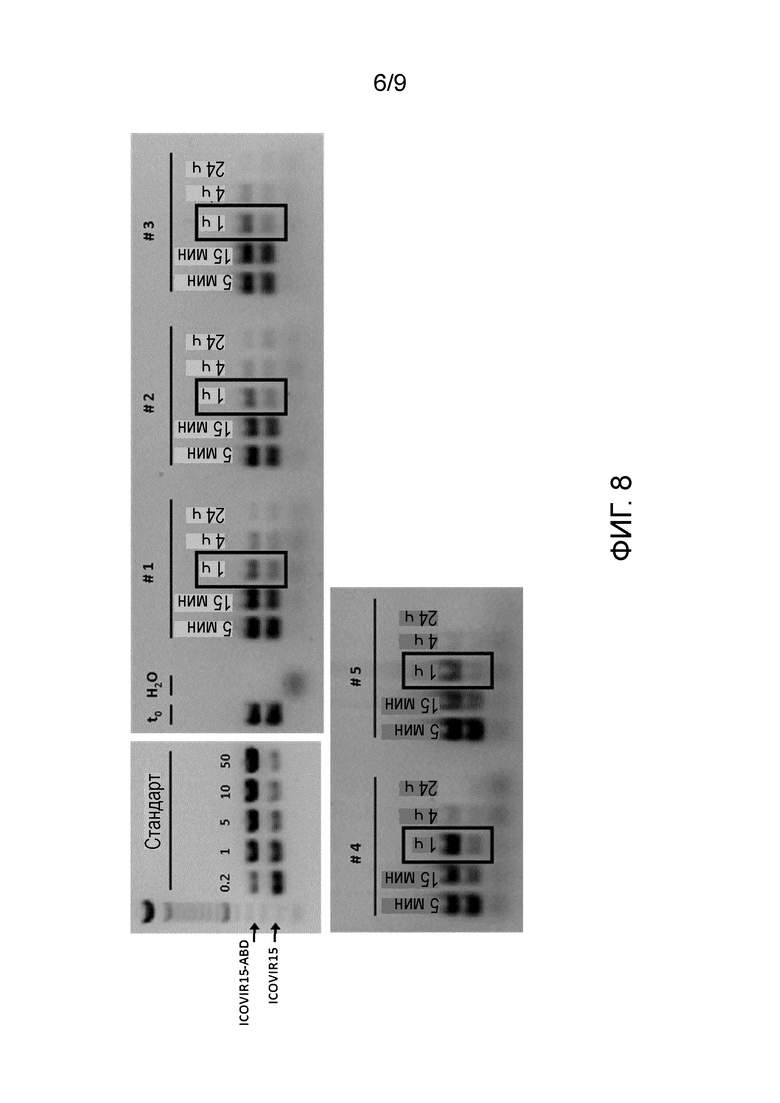
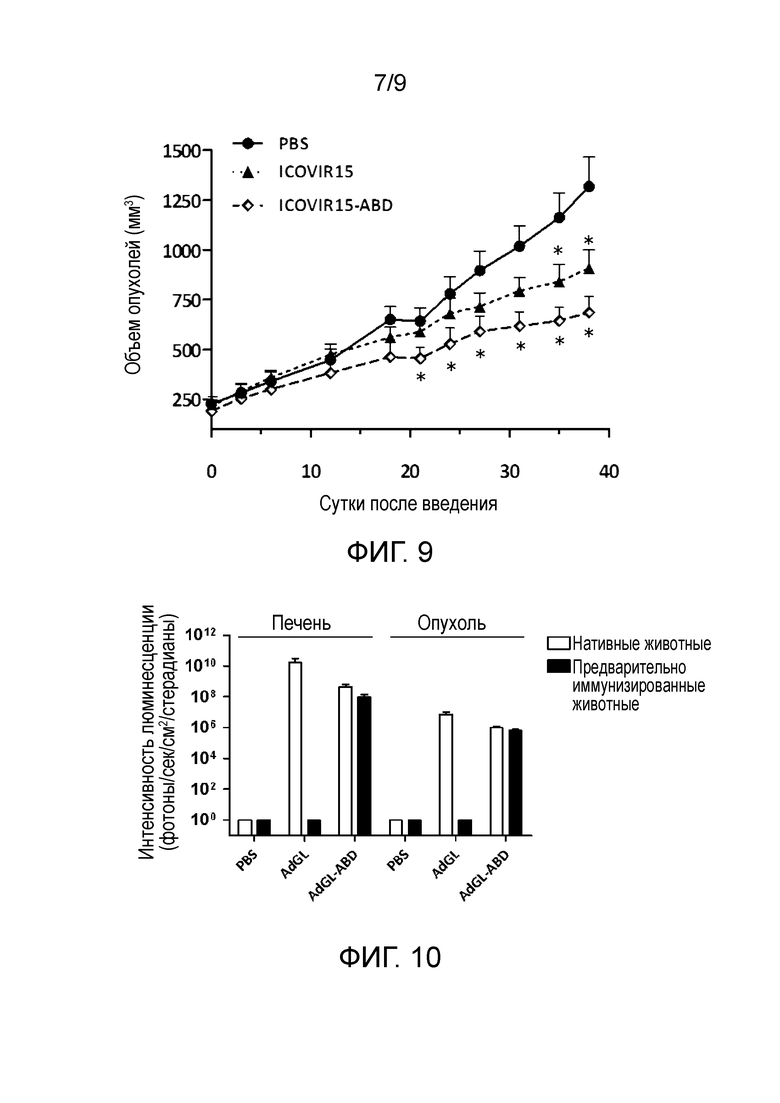
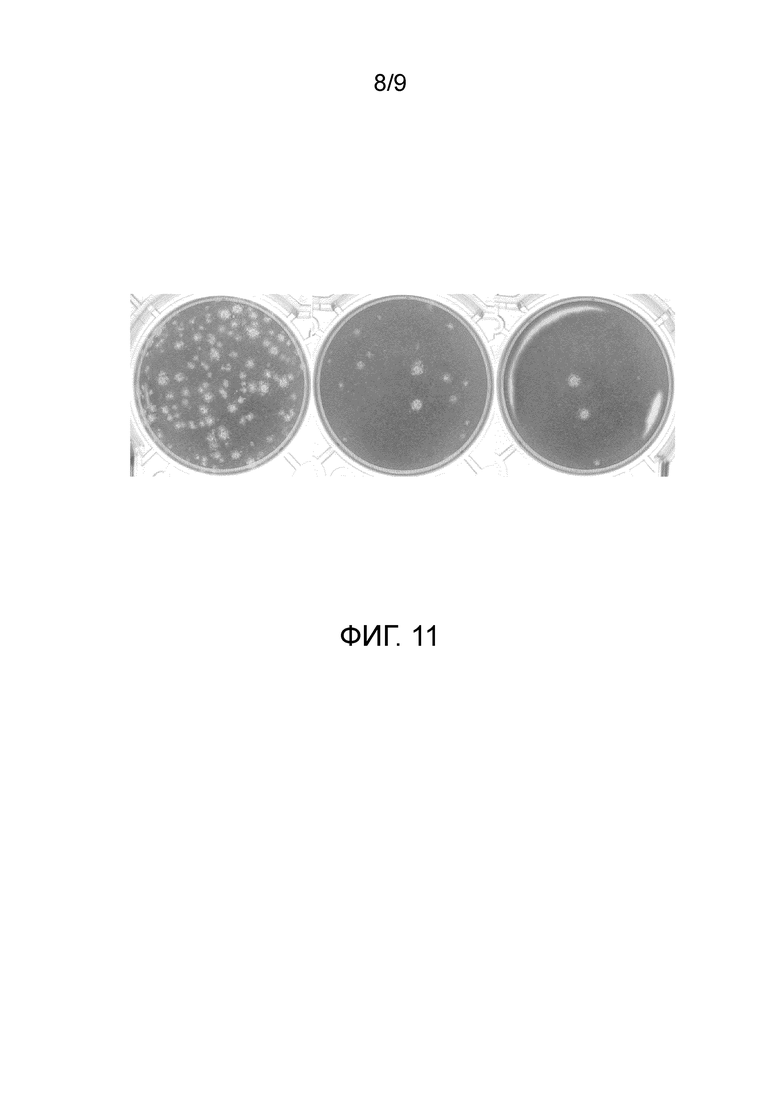
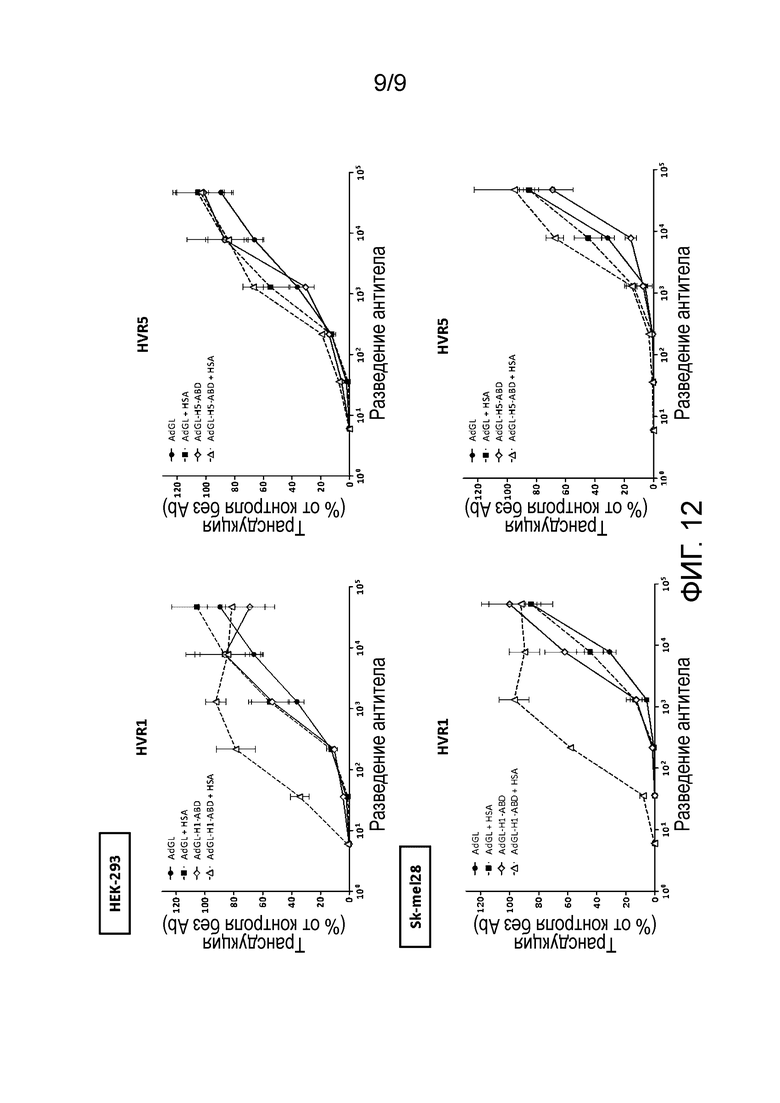
Изобретение относится к области биотехнологии и молекулярной биологии. Предложен аденовирусный геном для защиты аденовируса от нейтрализующих антител, отличающийся тем, что он содержит последовательность, кодирующую альбумин-связывающий участок, вставленную в кодирующую область гипервариабельного участка 1 (HVR1) гексонного белка, что приводит к экспрессии слитого белка, содержащего гексонный белок и альбумин-связывающий участок, и где альбумин-связывающий участок располагается на наружной поверхности гексонного белка, когда сборка гексонного белка имеет место в аденовирусном капсиде. Изобретение может быть применено в медицине в профилактике и/или лечении рака. 7 н. и 26 з.п. ф-лы, 12 ил., 1 табл., 1 пр.
1. Аденовирусный геном для защиты аденовируса от нейтрализующих антител, отличающийся тем, что он содержит последовательность, кодирующую альбумин-связывающий участок, вставленную в кодирующую область гипервариабельного участка 1 (HVR1) гексонного белка, что приводит к экспрессии слитого белка, содержащего гексонный белок и альбумин-связывающий участок, и где альбумин-связывающий участок располагается на наружной поверхности гексонного белка, когда сборка гексонного белка имеет место в аденовирусном капсиде.
2. Аденовирусный геном по п.1, где аденовирус представляет аденовирус человека.
3. Аденовирусный геном по п.2, где аденовирус человека выбран из группы, состоящей из аденовируса человека серотипов 1-57.
4. Аденовирусный геном по п.3, где аденовирус человека представляет аденовирус серотипа 5.
5. Аденовирусный геном по п.1, где альбумин-связывающий участок выбран из альбумин-связывающего домена из стрептококкового белка G, альбумин-связывающего домена из белка PAB Peptostreptococcus magnus, альбумин-связывающего пептида, имеющего коровую последовательность DICLPRWGCLW (SEQ ID NO:9), и их функционально эквивалентных вариантов.
6. Аденовирусный геном по п.5, где альбумин-связывающий участок представляет альбумин-связывающий домен 3 из стрептококкового белка G.
7. Аденовирусный геном по п.6, где последовательность альбумин-связывающего домена 3 из стрептококкового белка G представляет SEQ ID NO:1.
8. Аденовирусный геном по п.1, где последовательность, кодирующая альбумин-связывающий участок, вставлена таким образом, что полученный слитый белок содержит альбумин-связывающий участок после аминокислоты D150 гексонного белка согласно нумерации гексонного белка, имеющего инвентарный номер в GenBank BAG48782.1.
9. Аденовирусный геном по п.1, где N- и/или С-конец альбумин-связывающего участка связан с гексонным белком линкерной последовательностью.
10. Аденовирусный геном по п.9, где указанная линкерная последовательность содержит последовательность GSGS (SEQ ID NO:2).
11. Аденовирусный геном по п.1, где указанный аденовирусный геном дополнительно содержит тканеспецифический промотор или опухолеспецифичный промотор.
12. Аденовирусный геном по п.11, где тканеспецифический промотор или опухолеспецифический промотор представляют промоторные последовательности для регуляции экспрессии одного или более генов, выбранных из группы, состоящей из E1a, E1b, E2 и E4.
13. Аденовирусный геном по п.12, где промотор выбран из группы, состоящей из промотора E2F, промотора теломеразы hTERT, промотора тирозиназы, промотора простат-специфического антигена, промотора альфа-фетопротеина и промотора СОХ-2.
14. Аденовирусный геном по п.1, где аденовирус является онколитическим аденовирусом.
15. Аденовирусный геном по п.14, где указанный аденовирусный геном дополнительно содержит мутации в одном или более генов, выбранных из группы, состоящей из Е1а, Е1b, Е4 и VA-РНК, для достижения селективной репликации в опухолях.
16. Аденовирусный геном по п.1, где аденовирусный геном дополнительно содержит модификации капсида для повышения инфекционной активности аденовируса или его нацеленности на рецептор, присутствующий в опухолевой клетке.
17. Аденовирусный геном по п.16, где модификация капсида представляет инсерцию мотива RGD в петлю H1 аденовирусного белка волокна.
18. Аденовирусный геном по п.16, где модификация капсида представляет замену участка гена волокна гомологичным участком из аденовируса другого серотипа с получением химерного аденовируса.
19. Аденовирусный геном по п.1, где аденовирусный геном содержит один или более неаденовирусных генов, вставленных в указанный геном, и где указанные гены представляют гены, используемые в генной терапии или в вакцинации.
20. Аденовирусный геном по п.19, где указанные гены используются в генной терапии рака.
21. Аденовирусный геном по п.20, где указанные гены, используемые в генной терапии рака, представляют собой по меньшей мере ген, выбранный из группы, состоящей из пролекарство-активирующих генов, генов-супрессоров опухолей, генов, кодирующих противоопухолевые интерферирующие РНК, и иммуностимулирующих генов.
22. Рекомбинантный аденовирус, имеющий аденовирусный геном по одному из пп. 1-21, для защиты аденовируса от нейтрализующих антител.
23. Фармацевтическая композиция для лечения рака, содержащая терапевтически эффективное количество рекомбинантного аденовируса по п.22 вместе с фармацевтически приемлемым носителем.
24. Применение рекомбинантного аденовируса по п.22 или фармацевтической композиции по п.23 в лечении рака у млекопитающего, где аденовирус является онколитическим аденовирусом или аденовирусом, имеющим аденовирусный геном, содержащий один или более неаденовирусных генов, используемых в генной терапии рака, вставленных в геном аденовируса.
25. Применение по п.24, где млекопитающее является человеком.
26. Применение по п.24, где аденовирус вводят системно.
27. Применение рекомбинантного аденовируса по п.22 или фармацевтической композиции по п.23 для получения лекарственного средства для лечения рака у млекопитающего, где аденовирус является онколитическим аденовирусом или аденовирусом, имеющим аденовирусный геном, содержащий один или более неаденовирусных генов, используемых в генной терапии рака, вставленных в геном аденовируса.
28. Применение рекомбинантного аденовируса по п.22 или фармацевтической композиции по п.23 для получения вакцины для профилактики инфекционного заболевания у млекопитающего.
29. Применение по одному из пп. 27 или 28, где млекопитающим является человек.
30. Применение по п.27 или 28, где аденовирус вводят системно.
31. Способ лечения рака у млекопитающего, включающий введение указанному млекопитающему рекомбинантного аденовируса по п.22 или фармацевтической композиции по п.23, отличающийся тем, что аденовирус является онколитическим аденовирусом или аденовирусом, имеющим аденовирусный геном, содержащий один или более неаденовирусных генов, используемых в генной терапии рака, вставленных в геном аденовируса.
32. Способ по п.31, где млекопитающим является человек.
33. Способ по п.31, отличающийся тем, что аденовирус вводят системно.
| US 2008112929 A1, 15.05.2008 | |||
| HONGJU WU et all., "Identification of Sites in Adenovirus Hexon for Foreign Peptide Incorporation", Journal of virology, vol.79, no.6, 01.03.2005, pp.3382-3390 | |||
| HEDLEY S.J | |||
| et all., "Assesment of Genetic Shielding for Adenovirus Vectors", The open Gene Theerapy Journal, vol.2, 2009, pp.1-11 | |||
| ТЕРНОВОЙ В.А | |||
| и др., |

